
Ecofysiologisch onderzoek naar het effect van...
102
Faculteit Bio-ingenieurswetenschappen Academiejaar 2011-2012 Ecofysiologisch onderzoek naar het effect van droogtestress op de cavitatie- en hydraulische geleidbaarheidsdynamiek bij druif (Vitis vinifera L. cv. Chardonnay) Lies Kips Promotor: Prof. dr. ir. Kathy Steppe Tutor: ir. Annelies Baert Masterproef voorgedragen tot het behalen van de graad van Master in de bio-ingenieurswetenschappen: Milieu-technologie
Transcript of Ecofysiologisch onderzoek naar het effect van...

Faculteit Bio-ingenieurswetenschappen
Academiejaar 2011-2012
Ecofysiologisch onderzoek naar het effect van droogtestress op de
cavitatie- en hydraulische geleidbaarheidsdynamiek bij druif (Vitis
vinifera L. cv. Chardonnay)
Lies Kips
Promotor: Prof. dr. ir. Kathy Steppe
Tutor: ir. Annelies Baert
Masterproef voorgedragen tot het behalen van de graad van
Master in de bio-ingenieurswetenschappen: Milieu-technologie


Faculteit Bio-ingenieurswetenschappen
Academiejaar 2011-2012
Ecofysiologisch onderzoek naar het effect van droogtestress op de
cavitatie- en hydraulische geleidbaarheidsdynamiek bij druif (Vitis
vinifera L. cv. Chardonnay)
Lies Kips
Promotor: Prof. dr. ir. Kathy Steppe
Tutor: ir. Annelies Baert
Masterproef voorgedragen tot het behalen van de graad van
Master in de bio-ingenieurswetenschappen: Milieu-technologie


De auteur en de promotor geven de toelating deze masterproef voor consultatie beschikbaar te
stellen en delen van de masterproef te kopieren voor persoonlijk gebruik. Elk ander gebruik valt
onder de beperkingen van het auteursrecht, in het bijzonder met betrekking tot de verplichting
de bron uitdrukkelijk te vermelden bij het aanhalen van resultaten uit deze masterproef.
The author and the promoter give the permission to use this thesis for consultation and to copy
parts of it for personal use. Every other use is subject to the copyright laws, more specifically the
source must be extensively specified when using results from this thesis.
Gent, juni 2012
De promotor De tutor De auteur
Prof. dr. ir. Kathy Steppe ir. Annelies Baert Lies Kips


Dankwoord
De totstandkoming van deze thesis kan vergeleken worden met een Chardonnay druif, de basis
van een kwaliteitsvol glas wijn. Zo vormen Prof. dr. ir. Kathy Steppe en ir. Annelies Baert het
vruchtvlees van de druif. Met hun kennis, enthousiasme en talloze suggesties werd gezorgd voor
voldoende sappigheid en volume als basis van smaakvol glas wijn. Wanneer dit vruchtvlees van
slechte kwaliteit is, heeft het plukken van de druiven weinig zin. Bedankt voor de aanmoedigingen
en de opbouwende kritiek!
De andere thesisstudenten waarmee ik talloze uren samen in het lokaaltje heb gewerkt, waren een
perfecte afspiegeling van de pitjes en de schil. Deze taaie en bittere elementen lijken op het eerste
zicht eerder een noodzakelijk kwaad, maar niets is minder waar! Ze bevatten tannines die instaan
voor een belangrijk aandeel van de smaak en dus net zoals jullie, kleur en aroma geven aan de
wijn.
Een glas wijn met teveel alcohol bestaat niet. Daarom stellen Lidewei, Erik, Philip en Geert de
extra toegevoegde suiker voor die na fermentatie voor een stevige hoeveelheid alcohol zorgt. Door
kenners ook wel ’amelioration’ van de wijn genoemd wordt. Zonder jullie hulp zou ik nergens
geraakt zijn, bedankt!
De avonden die we gevuld hebben met thesispraat zijn niet meer op een hand te tellen en waren
broodnodig voor het relativeren van de tegenslagen. Daarom zijn Laura, Jarinda, Sofie, Lien,
Julie, Nele, Valerie, Patrijs, Toon, Tom, Stijn, Tracy, Hanne, Jessie, mama, papa, Roger, Lut,
Julie & Julie, Marie, Ward en nog zoveel anderen de zuren: een bijzonder belangrijk onderdeel
voor de conservatie van de wijn, waardoor deze met de jaren beter wordt!
Tot slot zijn er de klimatologische omstandigheden, zoals zeer mooi uitgebeeld door Jan. Uitermate
flexibel, onvoorspelbaar en broodnodig. De Chardonnay druif kan zich verrassend goed aanpassen
aan diverse omstandigheden en wordt dus onder kenners niet voor niets ”de hoer van de wijngaard”
genoemd.
Het was een plezier samen te werken met jullie!
v


Samenvatting
Cavitatie in planten is het ontstaan van luchtbellen in watergevulde xyleemvaten. Dergelijke
luchtgevulde vaten verbreken het watercontinuum en zorgen voor een verhoogde hydraulische
weerstand van de opwaartse sapstroom met nefaste gevolgen voor de groei en overleving van de
plant. Cavitatie wordt in de hand gewerkt door droge omgevingscondities die in het kader van de
huidige klimaatverandering steeds meer zullen voorkomen. Een goede kennis inzake cavitatie en
de bijhorende gevolgen voor de plant-waterbalans zijn bijgevolg van cruciaal belang.
Tegenwoordig wordt de cavitatiegevoeligheid van een plant gekarakteriseerd aan de hand van een
destructieve vulnerability curve (VC) waarin het verband tussen de waterpotentiaal en de hy-
draulische geleidbaarheid wordt weergegeven. De diverse methoden die hiertoe gebruikt worden,
genereren echter uiteenlopende resultaten en worden hevig bekritiseerd. Bovendien vereisen beide
parameters destructieve staalname wat de praktische uitvoering bemoeilijkt. De doelstelling van
deze thesis was het opstellen van een niet-destructieve VC waarin de cavitatiedynamiek recht-
streeks wordt weergegeven in functie van de xyleeminkrimping.
Experimenten werden uitgevoerd op afgeknipte takken van druivelaars (Vitis vinifera L. cv. Char-
donnay). Deze werden toegelaten verregaand uit te drogen waarbij ondertussen de (i) diameter,
(ii) cavitatie, (iii) hydraulische geleidbaarheid en (iv) waterpotentialen werden opgemeten. Hiertoe
werd respectievelijk gebruik gemaakt van een ’Linear Variable Displacement Transducer’ (LVDT-
sensor), een ultrasone cavitatiesensor, een XYL’EM of gravitatiekolom en een drukbom. Tot slot
werd ook het gewichtsverlies van de tak opgevolgd doorheen de dehydratatieperiode.
Uit de resultaten bleek dat het gelijktijdig opmeten van de schors- en xyleemdiameter in combi-
natie met het cavitatiepatroon en het waterverlies cruciale informatie bood over de plant tijdens
verregaande dehydratatie. Diverse stadia van uitdroging werden gekarakteriseerd waarbinnen ge-
differentieerd werd tussen de verschillende achterliggende mechanismen en een verband gelegd werd
tussen de verschillende parameters. Bovendien werd een nauwe overeenkomst gevonden tussen het
voorkomen van cavitatie en diameterdalingen onder de voorheen laagst behaalde waarde. In een
tweede luik van deze thesis werden zowel destructieve als niet-destructieve VC opgesteld. De
constructie van destructieve VC werd op verschillende punten bemoeilijkt (e.g. verstopping van
de vaten) waardoor de resultaten moeilijk te interpreteren waren. Op basis van de experimenteel
verworven kennis werden een aantal sleutelpunten aangehaald die in de toekomst gerespecteerd
dienen te worden om de natuurlijke situatie in de plant niet te beınvloeden tijdens het meetproces.
vii

viii
In vergelijking met deze gecompliceerde, destructieve methode bieden niet-destructieve VC een
belangrijke meerwaarde in het onderzoek naar de waterrelaties in druivelaars. De combinatie van
cavitatiesignalen en xyleeminkrimping in een niet-destructieve VC werd nog nooit eerder opge-
meten tijdens dergelijke sterke uitdroging waardoor de curve uniek is in zijn soort. De verkregen
sigmoıdale curves bevestigen dat beide parameters (cavitatie en xyleeminkrimping) eenduidig ge-
relateerd zijn en een belangrijke informatiebron vormen voor de cavitatie- en waterstatus van de
plant. Aan de hand van beide parameters kan de threshold bepaald worden vanaf wanneer cavita-
tie optreedt en vanaf wanneer schade wordt toegericht aan het watergeleidingssysteem. De studie
wijst uit dat het van cruciaal belang is de tak volledig te laten uitdrogen vooraleer kwantitatieve
schattingen gemaakt kunnen worden op basis van deze curve. Bovendien vertonen de resultaten
een seizoenale trend in de VC: naarmate het seizoen vorderde, ging eenzelfde hoeveelheid cavitatie
gepaard met een grotere inkrimping. Dit suggereert een seizoenale variatie in de elasticiteit, het
aantal vaten en de gevoeligheid van de vaten. Deze bevinding is van essentieel belang voor het
potentieel toekomstig gebruik van niet-destructieve VC in de plant-gebaseerde irrigatiecontrole.

Abstract
Cavitation in plants is the development of air bubbles in water filled xylem vessels. Those air
filled vessels break the water continuum and cause an elevated hydraulic resistance in the upward
sap flow with detrimental consequences for the growth and survival of the plant. Cavitation is
amplified in dry environmental conditions which are expected to occur more frequently as a con-
sequence of climate change. A sound understanding of cavitation and of the corresponding effect
on the water balance of the plant are therefore of the utmost importance.
Currently the plant’s sensitivity to cavitation is demonstrated by means of a destructive vulnerabi-
lity curve (VC) where the relationship between the water potential and the hydraulic conductivity
is shown. Various methods which are used to construct this VC, generate diverging results and
are under critical debate. Furthermore, destructive sampling is required for both parameters
which complicates the practical procedure. The goal of this thesis was the construction of a non-
destructive VC where the cavitation events are presented as a function of the shrinkage of the
xylem tissue.
Experiments were performed on cut branches of grapevine (Vitis vinifera L. cv. Chardonnay).
The samples were allowed to dehydrate under constant monitoring of the diameter and cavitation
and measurements were made of the hydraulic conductance and the water potential. Those para-
meters were measured respectively with (i) a ’Linear Variable Displacement Transducer’ (LVDT),
(ii) an ultrasonic cavitation sensor, (iii) a XYL’EM or a gravity column and (iv) a pressure bomb.
Finally, the weight loss was monitored through the dehydration period.
The results indicate that measurements of the total diameter and the xylem diameter combined
with the cavitation pattern, generate crucial information on the plant during dehydration. From
the relationship between those parameters, we were able to distinguish various dehydration stages,
characterised by different underlying mechanisms. Furthermore, we found a distinct correspon-
dence between cavitation events and shrinkage of the xylem diameter. As a second part of this
thesis, we developed both destructive and non-destructive VC. In the construction of destructive
VC, we have encountered a number of problems (e.g. clogging of the xylem vessels) which have
impeded the interpretation of the results. On the basis of experimentally gathered knowledge, a
few key points were emphasized to make the measuring process as minimally invasive as possible.
Compared to the complicated destructive method, non-destructive VC offer an important value in
the research of water relations in grapevine. Using cavitation events in combination with shrinkage
ix

x
of the xylem diameter in a non-destructive VC has never been used on cut branch segments, which
makes this curve unique. The resulting sigmoid curves confirmed that both parameters (cavitation
and xylem diameter) are related and thus constitute an important source of information on the
water status of the plant. Using the non-destructive VC, the cavitation threshold can be deter-
mined, as well as the point where damage in the water conducting system starts to occur. The
thesis revealed that it is of utmost importance that the branch is allowed to dehydrate completely
before quantitative estimations are based on the curve. Finally, results indicated a seasonal trend
in the VC: as the season proceeded, the same amount of cavitation events corresponded with a
larger shrinkage. This suggests a seasonal variation in the elasticity, the amount of vessels and
the vulnerability to cavitation of the vessels. This conclusion is essential for the potential future
use of non-destructive VC in plant-based irrigation.

Inhoudsopgave
Lijst van figuren xv
Inleiding xvii
1 Literatuurstudie 3
1.1 Watertransport van bodem tot atmosfeer . . . . . . . . . . . . . . . . . . . . . . . 3
1.1.1 Watertransport doorheen wortel en stengel . . . . . . . . . . . . . . . . . . 3
1.1.2 Verdamping van water naar de atmosfeer . . . . . . . . . . . . . . . . . . . 5
1.1.3 De waterpotentiaal als drijvende kracht . . . . . . . . . . . . . . . . . . . . 6
1.2 Hydraulische geleidbaarheid . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
1.2.1 Hydraulische geleidbaarheid op plantniveau . . . . . . . . . . . . . . . . . . 9
1.2.2 Hydraulische geleidbaarheid op xyleemniveau . . . . . . . . . . . . . . . . . 9
1.2.3 Opmeten van hydraulische geleidbaarheid . . . . . . . . . . . . . . . . . . . 10
1.3 Stamdiametervariaties . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
1.4 Cavitatie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
1.4.1 Fysische oorzaken van cavitatie . . . . . . . . . . . . . . . . . . . . . . . . . 13
1.4.2 Ontstaan en verspreiding van cavitatie binnen de plant . . . . . . . . . . . . 13
1.4.3 Gevolgen van cavitatie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
1.4.4 Herstelmechanismen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 16
1.4.5 Opmeten van cavitatie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
1.4.6 Cavitatie: trade-off veiligheid versus hydraulische geleidbaarheid . . . . . . 19
1.5 De vulnerability curve . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
2 Materialen en Methode 23
2.1 Experimentele proefopzet . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
2.2 Microklimaat . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
2.3 Waterpotentiaal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 26
2.3.1 Werkingsprincipe van de drukbom . . . . . . . . . . . . . . . . . . . . . . . 27
2.3.2 De drukbom in de praktijk . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
2.4 Hydraulische geleidbaarheid . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29
xi

xii Inhoudsopgave
2.4.1 Opmeten van de hydraulische geleidbaarheid: XYL’EM . . . . . . . . . . . 29
2.4.2 De XYL’EM in de praktijk . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
2.4.3 Opmeten van de hydraulische geleidbaarheid: gravitatiekolom . . . . . . . . 31
2.5 Diametervariaties . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
2.5.1 Werkingsprincipe van de LVDT-sensor . . . . . . . . . . . . . . . . . . . . . 31
2.5.2 De LVDT-sensor in de praktijk . . . . . . . . . . . . . . . . . . . . . . . . . 33
2.6 Cavitatie . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33
2.6.1 Opmeten van cavitatie: ultrasone akoestische emissies . . . . . . . . . . . . 33
2.6.2 Werkingsprincipe van de cavitatiesensor . . . . . . . . . . . . . . . . . . . . 34
2.6.3 Cavitatiesensor in de praktijk . . . . . . . . . . . . . . . . . . . . . . . . . . 34
2.6.4 Gevoeligheid van de cavitatiesensor . . . . . . . . . . . . . . . . . . . . . . . 34
3 Resultaten 39
3.1 Microklimaat . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
3.2 Gevoeligheid van de cavitatiesensor . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
3.3 Benchdrying experimenten . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
3.3.1 Algemeen verloop . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
3.3.2 Cavitatiedynamiek . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43
3.3.3 Diameterverloop in relatie tot cavitatie tijdens beperkte uitdroging . . . . . 43
3.3.4 Diameterverloop in relatie tot cavitatie tijdens sterke uitdroging . . . . . . 46
3.3.5 Diameterverloop in relatie tot waterverlies tijdens sterke uitdroging . . . . . 47
3.4 Destructieve vulnerability curve . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 48
3.5 Niet-destructieve vulnerability curve . . . . . . . . . . . . . . . . . . . . . . . . . . 49
4 Discussie 55
4.1 Cavitatiedynamiek tijdens periodes van zwakke en sterke uitdroging . . . . . . . . 55
4.2 Diameterverloop doorheen verregaande dehydratatie . . . . . . . . . . . . . . . . . 57
4.2.1 Verklaring diameterverloop gedurende periode 1 . . . . . . . . . . . . . . . 57
4.2.2 Verklaring diameterverloop gedurende periode 2 en 3 . . . . . . . . . . . . . 58
4.2.3 Verklaring diameterverloop gedurende periode 4 . . . . . . . . . . . . . . . 59
4.3 Destructieve vulnerability curve . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
4.3.1 Verklaring van het verschil in destructieve VC . . . . . . . . . . . . . . . . . 60
4.3.2 Kwantitatieve interpretatie van de destructieve VC . . . . . . . . . . . . . . 62
4.4 Niet-destructieve vulnerability curve . . . . . . . . . . . . . . . . . . . . . . . . . . 63
4.4.1 Opstellen van een niet-destructieve VC . . . . . . . . . . . . . . . . . . . . . 63
4.4.2 Praktisch gebruik en toepassingen van de niet-destructieve VC . . . . . . . 66
5 Besluit en toekomstperspectief 71
Bibliografie 73

Lijst van afkortingen en symbolen
Tabel 1: Lijst van afkortingen.
Afkorting Betekenis
APK ’Acoustic Emissions Peak Amplitude’
ASCO-P ’Acoustic Signal Conditioner with Peak Detector’
ASL ’Average Signal Level’
dB Decibel
DOY Dag van het jaar
LVDT ’Linear Variable Displacement Transducer’
PAR ’Photosynthetic Active Radiation’
PLC ’Percentage Loss of Hydraulic Conductivity’
RH ’Relative Humidity’
SPAC ’Soil Plant Atmosphere Continuum’
UAE ’Ultrasonic Acoustic Emissions’
VC ’Vulnerability Curve’
VD Verzadigingsdeficiet
xiii

Tabel 2: Lijst van symbolen.
Symbool Betekenis Eenheid
cUAE50 Inkrimping corresponderend met 50 % cavitatie mm
d Binnendiameter van het ide kanaal m
e0v Verzadigde dampdruk kPa
ev Actuele dampdruk kPa
F Steady-state waterflux kg.s−1
Kh Maximale hydraulische geleidbaarheid kg.s−1.MPa−1
K′h Actuele hydraulische geleidbaarheid kg.s−1.MPa−1
kh Specifieke hydraulische geleidbaarheid kg.m.s−1.Pa−1
Pv Absolute druk van lucht Pa
Px Absolute druk in het xyleem Pa
P50 Waterpotentiaal bij 50 % verlies aan hydraulische geleidbaarheid MPa
R Hydraulische weerstand MPa.s.kg−1
rp Kromtestraal van de pitporie m
T Temperatuur ◦C
σ Oppervlaktespanning van water N.m−1
ρw Dichtheid van water kg.m−3
ηw Dynamische viscositeit van water Pa.s
∆D Inkrimping mm
∆W Gewichtsverlies g
∆Ψ Potentiaalverschil Pa
Ψx Xyleemwaterpotentiaal Pa
Ψs Waterpotentiaal in het opslagweefsel Pa
Ψw Waterpotentiaal Pa
Ψg Gravitationele potentiaal Pa
Ψm Matrixpotentiaal Pa
Ψp Hydrostatische potentiaal Pa
Ψπ Osmotische potentiaal Pa
xiv

Lijst van figuren
1.1 Illustratie van de wtransportroute van water van bodem tot atmosfeer . . . . . . . 4
1.2 Transpiratiedynamiek en stamdiametervariaties . . . . . . . . . . . . . . . . . . . . 8
1.3 Schematische weergave van de waterstroming doorheen het SPAC . . . . . . . . . . 10
1.4 Illustratie van de ’air-seeding’ hypothese . . . . . . . . . . . . . . . . . . . . . . . . 15
1.5 Model van het herstelmechanisme van embolismen in xyleemvaten van druivelaar. . 17
2.1 Opstelling van de druivelaars in de serre . . . . . . . . . . . . . . . . . . . . . . . . 23
2.2 Schematische weergave van de volledige proefopstelling . . . . . . . . . . . . . . . . 24
2.3 Gedetailleerd overzicht van het benchdrying experiment . . . . . . . . . . . . . . . 25
2.4 De Scholander-drukbom . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
2.5 Voorstelling van de diverse componenten van de waterpotentiaal . . . . . . . . . . 28
2.6 Opstelling van de verticale gravitatiekolom . . . . . . . . . . . . . . . . . . . . . . 32
2.7 Vereenvoudigde weergave van de werking van de LVDT-sensor . . . . . . . . . . . . 33
2.8 Detailweergave van de geınstalleerde cavitatiesensor . . . . . . . . . . . . . . . . . 35
3.1 Microklimaat . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
3.2 Reikwijdte van de cavitatiesensor . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41
3.3 Detectie threshold van de cavitatiesensor . . . . . . . . . . . . . . . . . . . . . . . . 42
3.4 Cavitatiedynamiek en diameterverloop van een afgesneden druiventak . . . . . . . 42
3.5 Xyleem- en schorsdiameterverloop doorheen experiment 5 . . . . . . . . . . . . . . 44
3.6 Differentiatie tussen inkrimpend en uitzettend xyleemweefsel in combinatie met het
voorkomen van cavitatiesignalen . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
3.7 Verloop van het volume en het gewicht van de tak doorheen experiment 6 . . . . . 47
3.8 Destructieve vulnerability curve . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 49
3.9 Niet-destructieve vulnerability curve . . . . . . . . . . . . . . . . . . . . . . . . . . 51
3.10 Detail van een niet-destructieve vulnerability curve . . . . . . . . . . . . . . . . . . 52
xv


Inleiding
Watertransport in bomen is een intrigerend fenomeen. Planten transporteren dagelijks water over
afstanden die onze menselijke capaciteiten ruim overschrijden. Desondanks is het precieze mecha-
nisme van deze sapstroom nog niet volledig opgehelderd.
De cohesietheorie stelt dat de opwaartse sapstroom ontstaat door een gradient van zuigspanningen
opgebouwd doorheen het watercontinuum onder invloed van transpiratie. Droogtestress en/of een
toenemende mate van transpiratie veroorzaakt verhoogde zuigspanningen in het vasculair weef-
sel, wat uiteindelijk cavitatie kan induceren. Cavitatie is het breken van het watercontinuum in
xyleemvaten waardoor de vaten gevuld worden met lucht in plaats van met water. Dit leidt tot
verminderde hydraulische geleidbaarheid en resulteert dus in een substantiele vermindering van
het watertransport. Zo vormt het een van de belangrijkste beperkingen in het overleven van de
plant in een droge omgeving. Als reactie op deze condities kan de plant zijn stomatale geleid-
baarheid verlagen zodat de transpiratie afneemt en zuigspanningen gereduceerd worden tot op
een niveau waarop het water zijn metastabiele, vloeibare staat kan behouden. Op die manier kan
de sapstroom onderhouden blijven (e.g. Dixon & Joly, 1895; Tyree & Sperry, 1988; Holtta et al.,
2002). De stomata staan echter ook in voor de gasuitwisseling met de atmosfeer en houden dus
nauw verband met de fotosyntheseprocessen in de plant. Zo heeft stomatale sluiting ook invloed op
de netto assimilatie, het competitief vermogen en uiteindelijk de groei en overleving van de plant
(e.g. Choat et al., 2010). Om een goede waterbalans te behouden is efficient watertransport dus
van cruciaal belang. De globale klimaatverandering gaat gepaard met een stijgende gemiddelde
luchttemperatuur, wat een vergrootte evapotranspiratie tot gevolg kan hebben. Daarenboven
wordt een stijging in zowel frequentie als duur van acute hittegolven voorspeld. Deze verhogen
zelfs in gematigde gebieden de kans op erg droge periodes. Voornamelijk deze laatste kunnen een
groot effect uitoefenen op het overleven van planten, gezien veel soorten nu reeds dicht bij hun
droogtelimiet leven. Het overleven van de planten zal dus meer en meer afhankelijk zijn van een
goede waterbalans en het behoud van de waterkolom (e.g. Schultz, 2000; Martınez-Vilalta et al.,
2002).
In dit onderzoek werden kwantitatieve gegevens verzameld van de waterrelaties in druivelaar Vitis
vinifera L. cv. Chardonnay. Druivelaars groeien vaak in gebieden waar kans is op intensieve droog-
testress en waar cavitatie dus een veel voorkomend fenomeen is (e.g. Schultz, 2003; Lovisolo et al.,
2008). Anatomisch wordt de druivelaar gekenmerkt door grote xyleemvaten, die een goede basis
xvii

bieden om de cavitatiedynamiek te onderzoeken (Sperry et al., 1987; Schultz & Matthews, 1988;
Chaves et al., 2010). Daarnaast is het een gewas met hoge economische waarde, wat een verbeterd
inzicht in de waterrelaties relevant maakt. Zo kan een gestuurde irrigatiecontrole een belangrijke
meerwaarde bieden in de druifkwaliteit en bijhorende wijnkwaliteit. Cavitatie is een interdiscipli-
nair onderwerp dat een brug slaat tussen biologie en fysica. Gezien veel factoren omtrent cavitatie
en de droogterespons van planten nog niet volledig opgehelderd zijn, is het onderhevig aan een
levendig debat. Dat maakt cavitatie tot een ’hot topic’ dat constant aan nieuwe inzichten wint.
Het doel van deze thesis is tweevoudig. Enerzijds wordt getracht een beter inzicht te krijgen in
de cavitatiedynamiek door het zoeken naar ecofysiologische verbanden tussen het verloop van (i)
cavitatie, (ii) hydraulische geleidbaarheid, (iii) diametervariaties en (iv) waterpotentialen. An-
derzijds beogen we een nieuwe maat te vinden om de gevoeligheid van de plant aan cavitatie
te karakteriseren. Standaard wordt hiervoor gebruik gemaakt van een destructieve vulnerability
curve (VC), waarin de hydraulische geleidbaarheid wordt onderzocht in functie van de waterpo-
tentiaal. Beide parameters vereisen echter destructieve staalname en nieuwe variabelen zijn dan
ook gewenst om de waterstatus in de toekomst op een niet-invasieve manier in kaart te brengen.
Door middel van een niet-destructieve VC (bijvoorbeeld op basis van het verband tussen cavitatie
en diametervariaties) zou men in de toekomst vlotter kunnen anticiperen op de nefaste gevolgen
van cavitatie met betrekking tot een verbeterde irrigatiecontrole. Daarom werd in deze thesis
onderzocht of er een consistente relatie bestaat tussen het cavitatiepatroon en diametervariaties
doorheen verregaande uitdroging van een druiventak.
In de literatuurstudie (hoofdstuk 1) worden de relevante fysiologische processen besproken met
betrekking tot cavitatie. Hierin komt het watertransport aan bod, gevolgd door diverse aspecten
van de hydraulische geleidbaarheid, stamdiametervariaties en cavitatie en tot slot wordt finaal de
destructieve VC behandeld. De algemene proefopzet van de experimenten wordt besproken in
hoofdstuk 2 (materialen en methode). Daarnaast worden de gehanteerde technieken uiteengezet
die toegepast werden om het microklimaat en de vier variabelen op te meten (waterpotentiaal,
hydraulische geleidbaarheid, diametervariaties en cavitatie). In hoofdstuk 3 (resultaten) worden
de relevante data gepresenteerd en beschreven. In de discussie (hoofdstuk 4) wordt vervolgens kri-
tisch gereflecteerd en worden de resultaten in perspectief gezet. Er wordt onder andere nagegaan
hoe de plant zich gedraagt tijdens verregaande uitdroging. Welke cavitatiedynamiek vertoont de
plant onder diverse omstandigheden? Hoe verloopt het verlies aan hydraulische geleidbaarheid en
is dit verlies reversibel? Gaat het hervullen van de vaten ook gepaard met cavitatie? Hoe uit zich
dit in de diameter van de tak? Kunnen we differentieren tussen diameterinkrimping ten gevolge
van stijgende zuigspanningen enerzijds en het legen van de waterreserves anderzijds? Zijn deze
trends gerelateerd aan het cavitatiepatroon? Welk verloop vertoont de niet-destructieve VC? Is
dit verloop constant of is er een seizoenale trend te detecteren? Finaal worden de belangrijkste
inzichten kernachtig samengevat en wordt een toekomstperspectief geboden in het laatste deel van
deze thesis, hoofdstuk 5 (besluit en toekomstperspectieven).
xviii

LITERATUURSTUDIE


Hoofdstuk 1
Literatuurstudie
1.1 Watertransport van bodem tot atmosfeer
Om alle vitale functies te kunnen vervullen, transporteert de plant grote hoeveelheden water.
Onder vochtige omstandigheden kan 5 − 10 kg water m−2 dag−1 verdampt worden door stomata
ter hoogte van de bladeren, die zich op meer dan 100 m van de bodem kunnen bevinden (Cruiziat
& Tyree, 1990; Cochard, 2006). Massale hoeveelheden water moeten dus beduidende afstanden
afleggen, wat een efficient watertransport van cruciaal belang maakt.
Voordat er wordt ingegaan op de drijvende kracht en de bijhorende mechanismen, wordt eerst het
structurele verloop van het watertransport en de anatomische eigenschappen van het vasculair
weefsel toegelicht. Figuur 1.1 illustreert de weg die water aflegt vanuit de bodem doorheen de
plant tot in de atmosfeer.
1.1.1 Watertransport doorheen wortel en stengel
Wateropname start ter hoogte van de wortel. Uit de rhizosfeer wordt het water opgenomen via de
wortelhaartjes, waarna het beweegt doorheen de epidermis tot in de cortex. Na het kruisen van
de endodermis en de pericyclus migreert het water richting vasculair weefsel, wat zich hogerop
ook doortrekt in stengel en blad (Cochard, 2006). Dit vasculair weefsel bestaat uit xyleem, floeem
en cambiumweefsel. Zowel xyleem als floeem worden jaarlijks gesynthetiseerd door het vasculair
cambium, een dunne laag die zich tussen beide situeert. Het floeem bevindt zich aan de buitenkant
van de cambiumlaag en bestaat uit een dunne laag cellen die zorgt voor het transport en de
distributie van fotosyntheseproducten vanuit de bladeren naar diverse organen. In het xyleem
stroomt het sap in tegenovergestelde richting; het vervoert het opgenomen water vanuit de wortel
in de richting van de bladeren. Secundair xyleem wordt jaarlijks naar het centrum van de stam
afgescheiden waardoor de stamdiameter toeneemt en groeiringen ontstaan (Tyree & Ewers, 1991).
In deze thesis ligt de focus op het opwaarts watertransport doorheen de xyleemvaten. Daarom zal
3
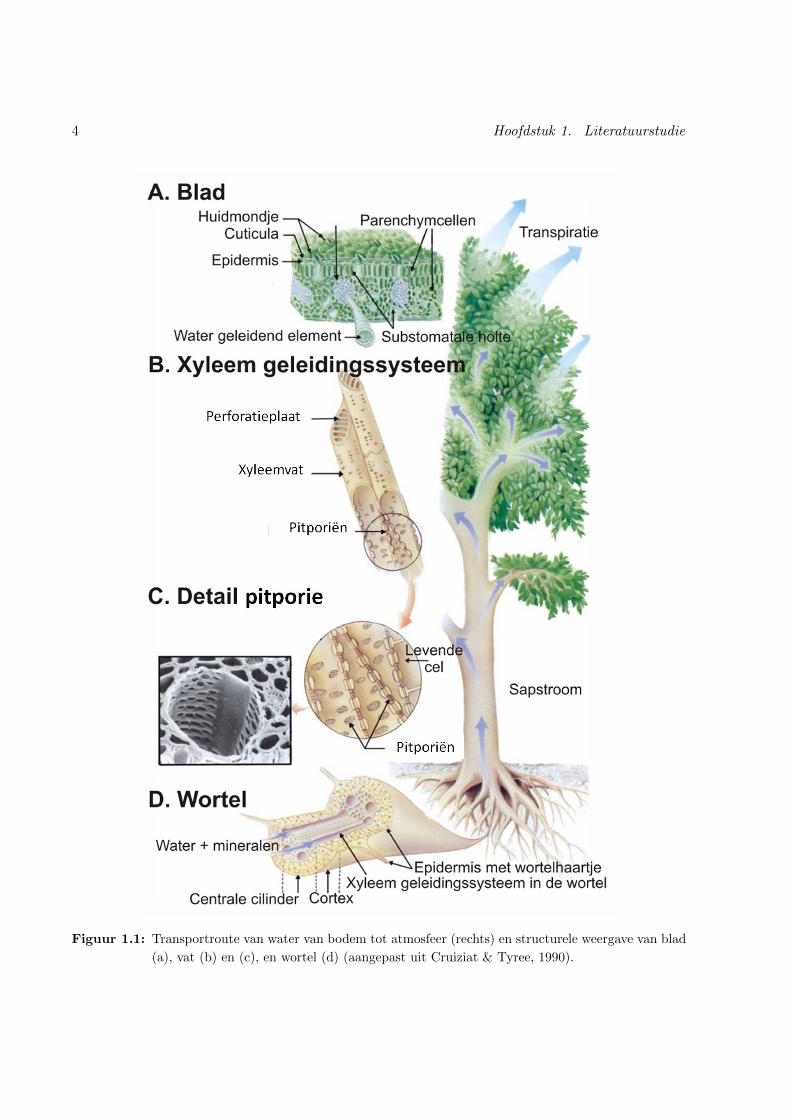
4 Hoofdstuk 1. Literatuurstudie
Figuur 1.1: Transportroute van water van bodem tot atmosfeer (rechts) en structurele weergave van blad
(a), vat (b) en (c), en wortel (d) (aangepast uit Cruiziat & Tyree, 1990).

Hoofdstuk 1. Literatuurstudie 5
in het vervolg sapstroom gebruikt worden als synoniem voor opwaarts watertransport
Anatomisch bestaat het xyleem uit twee celtypes: tracheeen en tracheıden. Tracheıden komen
voor in alle vasculaire planten en varieren in lengte van 2 tot 6 mm, terwijl tracheeen of vatelemen-
ten hoofdzakelijk voorkomen in angiospermen (figuur 1.1 B). Deze vatelementen zijn aan elkaar
verbonden via perforatieplaten en vormen zo lange houtvaten van uiteenlopende afmetingen (1 mm
tot meer dan 1 m) (Cruiziat & Tyree, 1990; Tyree & Ewers, 1991). Watertransport over lange
afstand grijpt plaats in de lumina van deze xyleemelementen. Ze zijn enkel functioneel in dode
toestand wat een laagweerstandige axiale sapstroom mogelijk maakt. Aangezien de houtvaten zich
niet oneindig uitstrekken moet water ook lateraal tussen vaten kunnen bewegen. De aanwezigheid
van pitporien op de laterale zijwanden maken dergelijke radiale stroming mogelijk (figuur 1.1 C).
Zo een pitporie bestaat uit een netwerk van cellulose microfibrillen waarin tortueuze kanalen de
microporien vormen (Cruiziat & Tyree, 1990; Choat et al., 2008). Deze bepalen de pitporositeit
en zullen erg belangrijk blijken in de vatconnectiviteit en gevoeligheid aan cavitatie (sectie 1.4)
(Cruiziat & Tyree, 1990; Tyree & Zimmermann, 2001).
Naast de watergeleidende elementen bestaat het xyleemweefsel ook uit vezels en parenchymcellen.
Vezels hebben een verstevigde secundaire celwand voor structurele steun. Parenchymcellen zijn
de enige levende celtypes en vervullen een opslagfunctie voor water en koolhydraten. Zo kunnen
ze instaan voor lateraal transport van water en opgeloste stoffen en bovendien actief reageren op
pathogenen en verwondingen (Tyree & Ewers, 1991; Steppe, 2004).
1.1.2 Verdamping van water naar de atmosfeer
Vanuit de stam wordt het water getransporteerd via de xyleemvaten tot in de bladeren, waar de
tracheeen zich transformeren tot nauwe nerven. Hier komt het water in de celwand van de mesofiel-
cellen terecht, vanwaar de verdamping plaatsgrijpt. Deze verdamping is in sterke mate afhankelijk
van het microklimaat via een combinatie van drie factoren: zonlicht, temperatuur (T) en relatieve
vochtigheid (RH, ’relative humidity’) van de atmosfeer. Onder invloed van fotosynthetisch actieve
straling (PAR, ’photosynthetically active radiation’) worden de huidmondjes of stomata geopend
en kan de plant latente energie gebruiken om water te verdampen in de substomatale ruimte
(figuur 1.1 A). Hier schommelt de RH rond de 100%, terwijl de atmosferische luchtvochtigheid
meestal veel lager ligt. Dit betekent een drijvende kracht voor diffusie en resulteert in het verlies
van waterdamp doorheen de huidmondjes (Cruiziat & Tyree, 1990; Jones, 1992; Cochard, 2006).
Het effect van RH en T wordt gecombineerd in een parameter en uitgedrukt als het verzadigings-
deficiet (VD). Dit VD is het verschil tussen de werkelijke en de verzadigde hoeveelheid vocht in
de lucht zodat een grotere waarde een sterkere drijvende kracht voor transpiratie betekent. Onder
invloed van deze concentratiegradient zal gasvormig water tenslotte vanuit de huidmondjes tot in
de atmosfeer diffunderen (e.g. Cruiziat & Tyree, 1990; Tyree, 1997). Deze huidmondjes verkeren
echter niet altijd in geopende toestand. Naast een uitgang voor gasvormig water verzorgen ze

6 Hoofdstuk 1. Literatuurstudie
immers gelijktijdig toegang tot CO2 uit de lucht. Dit is nodig voor fotosyntheseprocessen in de
plant. Een optimale balans tussen waterverlies en CO2-opname wordt bewerkstelligd door regeling
van de openingsgraad van de huidmondjes, ook wel de stomatale weerstand genoemd. Daardoor
bestaat er een verband tussen fotosynthese en de waterstatus van de plant en kan een verminderde
sapstroom verstrekkende gevolgen hebben voor de fotosynthese en uiteindelijk de overleving van
de plant (Cruiziat & Tyree, 1990).
1.1.3 De waterpotentiaal als drijvende kracht
De waterpotentiaal is een maat voor de beschikbaarheid van water. De grootte van de water-
potentiaal duidt de kracht aan waarmee water wordt aangezogen (ψ < 0) of afgestoten (ψ > 0).
Doorheen het hydraulisch netwerk van de plant wordt een gradient van negatieve waterpotentialen
(zuigspanningen) opgebouwd. In de bodem heerst de minst negatieve waterpotentiaal, terwijl de
waarden steeds negatiever worden naar de bladeren toe. Water stroomt spontaan in de richting
van dalende (meer negatieve) waterpotentialen, in overeenstemming met de wet van de minimale
energie en wordt zo doorheen het xyleem ’omhooggetrokken’ tot in de bladeren (sectie 1.2). Sap-
stroom is bijgevolg een passief proces met als drijvende kracht het verschil in waterpotentiaal
(Tyree & Zimmermann, 2001; Jury & Horton, 2004).
Cohesie theorie
In 1895 beschreven Dixon en Joly de ’Cohesion-Tension theory’, een verklarende theorie omtrent
het watertransport in planten, die tot op vandaag grotendeels wordt geaccepteerd. De theorie stelt
dat de waterpotentiaal in de plant bijna steeds negatief is, waardoor het volledige hydraulische
netwerk van de plant onder zuigspanning staat. Deze zuigspanning wordt veroorzaakt door trans-
piratie ter hoogte van de huidmondjes en zet zich voort over de gehele lengte van de plant. De
waterkolom houdt stand als gevolg van de heersende cohesiekrachten tussen watermoleculen on-
derling. Het hydraulisch netwerk kan dus als een continuum doorheen de gehele plant worden
beschouwd en wordt dikwijls benoemd als een bodem-plant-atmosfeer-continuum (SPAC, ’soil
plant atmosphere continuum’)(Cruiziat & Tyree, 1990; Tyree & Zimmermann, 2001).
Dagelijks verloop van de waterpotentiaal en de diameter
Aangezien transpiratie een diurnaal, zongebaseerd proces is, vertoont ook de waterpotentiaal van
de plant een dagelijkse cyclus (Irvine en Grace, 1997). ’s Morgens is de boom in verzadigde toe-
stand. Er bestaat geen waterpotentiaalgradient tussen de wortels en de bladeren waardoor het
water in evenwicht is en de sapstroom stilstaat. Onder invloed van zonlicht gaan de stomata open
en start de transpiratie. De waterpotentiaal ter hoogte van de huidmondjes daalt. Als gevolg van
deze daling ontstaat een potentiaalgradient in het xyleem waardoor waterstroming op gang komt.

Hoofdstuk 1. Literatuurstudie 7
Deze zuigspanning wordt sequentieel doorgegeven aan de diverse organen van de kruin naar de
wortels toe (Tyree & Zimmermann, 2001; Steppe et al., 2006).
Ten gevolge van de hydraulische weerstand doorheen de plant, is de zuigspanning niet meteen
voelbaar ter hoogte van de wortels (Peramaki et al., 2001; Zweifel & Item, 2000; Sevanto et al.,
2011). Door deze vertraging start de wateropname in de wortels niet gelijktijdig met de trans-
piratie waardoor de boom initieel beroep moet doen op interne reserves. Deze bevinden zich in
elk waterhoudend orgaan: bladeren, vruchten, takken, stam en wortels (Ueda & Shibata, 2001).
Het aanspreken van dergelijke reserves resulteert in een radiale waterstroming. Op die manier
is de plant tijdelijk in staat om snel te reageren op varierende transpiratie, onafhankelijk van de
worteltoevoer. Zo worden grote schommelingen in de xyleem waterpotentiaal in beperkte mate
opgevangen. Ondertussen zet de zuigspanning zich verder door in de richting van de wortels en
onder invloed van cohesiekrachten en komt er een verticale wateropname op gang. De radiale
watertoevoer houdt aan tot wanneer beide potentialen eenzelfde grootte hebben bereikt. De drij-
vende kracht is dan gelijk aan nul. De waterreserves worden dus niet enkel aangesproken in tijden
van droogte, maar zijn intrinsiek verbonden met de dagelijkse sapstroom (Zweifel & Item, 2000).
Rond de middag bereikt de boom een steady-state-toestand waarin evenveel water wordt opgeno-
men als er getranspireerd wordt. Op dit moment is de waterpotentiaal het meest negatief en is
de diameter het kleinst. ’s Avonds vindt het omgekeerde proces plaats: de wateropname door de
wortels overschrijdt de transpiratie waardoor interne reserves opnieuw aangevuld worden (Irvine &
Grace, 1997; Peramaki et al., 2001; Sevanto et al., 2005; Steppe et al., 2006). Op deze manier ont-
staat een sterke hydraulische koppeling tussen de opslagpools en de transpiratiestroom en worden
extreme variaties in waterpotentiaal gebufferd (Steppe et al., 2006). Dergelijke hydratatiecycli
vertalen zich in een diurnaal diameterpatroon, corresponderend met het dagelijks verloop van
de waterpotentiaal. Naarmate de opslagpools overdag uitgeput raken, krimpt de stam terwijl
deze ’s avonds en ’s nachts zal toenemen door het opnieuw aanvullen van de reserves (Irvine &
Grace, 1997; Cochard et al., 2001; Intrigliolo & Castel, 2007; Almeras, 2008). Het verloop van de
transpiratie en de diametervariaties zijn schematisch weergegeven in figuur 1.2.
Opmeten van de waterpotentiaal: controversieel debat omtrent de validiteit van de
cohesietheorie en de drukbom
Ondanks het ontstaan van de cohesietheorie in 1894 werd het opmeten van dergelijke nega-
tieve waterpotentialen pas mogelijk gemaakt in de jaren 1950-1960 met de ontwikkeling van de
thermokoppel-psychrometer (Spanner, 1951) en de drukbom (Scholander et al., 1964). Bladwa-
terpotentialen tot -10 MPa werden opgemeten en deze ontwikkeling werd beschouwd als bevesti-
ging van de cohesietheorie (Kolb & Davis, 1994). Omwille van zijn eenvoudige hanteerbaarheid
werd de drukbom uitvoerig gebruikt door plantfysiologen en vond hij zijn praktische intrede in
irrigatiecontrole. In 1994 lanceerde Zimmerman de xyleemdrukprobe, een toestel dat toeliet de
xyleemwaterpotentiaal ’in situ’ te meten. De opgemeten zuigspanningen lagen veel lager dan de

8 Hoofdstuk 1. Literatuurstudie
Figuur 1.2: (A)Illustratie van de dagelijkse dynamiek van transpiratie en stamdiametervariaties en (B)
de overeenkomstige uitputting en bijvulling van het opslagweefsel. Verder wordt de richting
en grootte van de opwaartse en radiale waterstroming aangegeven op diverse tijdstippen
van de dag met Ψx en Ψs de waterpotentiaal van respectievelijk het xyleemweefsel en het
opslagweefsel (aangepast uit Fernandez & Cuevas, 2010).
metingen van de drukbom (0 tot -1 MPa), waardoor de accuraatheid van de drukbom in twijfel
werd getrokken (Steudle, 2001). Daardoor werd het debat omtrent de basis van de opwaartse
sapstroom opnieuw aangewakkerd, wat doorgaat tot op vandaag.
Zo bekritiseren Westhoff et al. (2009) en Zimmermann et al. (2004) het voorkomen van een groot
watercontinuum doorheen de plant, alsook de grootteorde van de waterpotentialen. Ze opperen
dat de opwaartse sapstroom niet zomaar als een geısoleerd mechanisme kan beschouwd worden
waarin xyleemvaten niet meer zijn dan kwetsbare kanalen. Volgens hen bewerkt een samenspel
van diverse krachten (e.g. osmotische, capillaire, elektrische en transpiratiekrachten) over korte
afstanden de sapstroom zodat het water in verschillende stappen omhooggetrokken wordt. In
het licht van deze theorie, ook wel de ’Watergate’ of ’Multi-Force’ theorie genoemd, stellen ze de
werking van de Scholander-drukbom in vraag, die volgens hen de grootte van de zuigspanning
overschat. Ze baseren zich hiervoor op resultaten van de xyleemdrukprobe en cryo-scanning
elektronenmicroscopie (Wei et al., 1999).

Hoofdstuk 1. Literatuurstudie 9
Onderzoek naar het exacte mechanisme achter de opwaartse sapstroom in planten blijft dus een
’hot topic’ en is onderhevig aan een levendig debat (Cochard et al., 2001; Steudle, 2001; Wei
et al., 2001). Desondanks is de cohesietheorie tot op vandaag nog grotendeels aanvaard en blijft
de drukbom de standaard methode voor het opmeten van waterpotentialen (Holbrook et al., 1995;
Cochard et al., 2001; Wei et al., 2001; Choat et al., 2010; Ruger et al., 2010).
1.2 Hydraulische geleidbaarheid
1.2.1 Hydraulische geleidbaarheid op plantniveau
Opwaartse sapstroom doorheen het SPAC wordt bewerkstelligd onder invloed van een constante
afname in waterpotentiaal (meer negatief) in de richting van de bladeren. Naar analogie met
elektrische circuits beschreef van den Honert (1948) elk medium (e.g. bodem, wortels, takken) als
een afzonderlijke hydraulische weerstand waarover de sapstroom stroomt. Hierin is hydraulische
weerstand het inverse van hydraulische geleidbaarheid (figuur 1.3). Dit concept stelde hem in staat
de wet van Ohm te introduceren om de steady-state-waterflux (F [kg.s−1]) doorheen de plant te
kunnen kwantificeren:
F =∆ψ
R= ∆ψKh (1.1)
Hierin is het potentiaalverschil ∆ψ [MPa] de drijvende kracht, R ondervonden weerstand [MPa.s.kg−1] en Kh de hydraulische geleidbaarheid [kg.s−1.MPa−1]. Vergelijking 1.1 biedt slechts een ver-
eenvoudigde kwantificatie van het watertransport, gezien capacitieve aspecten (e.g. wateropslag)
niet in rekening worden gebracht (Boyer, 1985; Tyree & Ewers, 1991).
1.2.2 Hydraulische geleidbaarheid op xyleemniveau
Puur fysisch gezien kan sapstroom doorheen xyleemvaten vergeleken worden met waterstroming
doorheen capillaire kanalen. In de hydrodynamica wordt dergelijke stroming gedefinieerd door
middel van de wet van Hagen-Poiseuille:
F =πρw
∑ni=1 d
4
128ηw
∆ψ
l= kh
∆ψ
l(1.2)
Hierin is ρw de dichtheid van water (998.2 kg.m−3 bij 20◦C), ηw de dynamische viscositeit van
water (1.002 10−3 Pa.s bij 20◦C), d de binnendiameter van het ide kanaal (m), ∆ψl de drukpoten-
tiaalgradient (Pa.m−1) en kh de specifieke hydraulische geleidbaarheid (kg.m.s−1.Pa−1). Hieruit
blijkt dat de specifieke hydraulische geleidbaarheid evenredig is aan de som van de vatdiameters
tot de vierde macht. Kleine variaties van deze diameters oefenen dus een significante invloed uit
op de waterstroming (Tyree & Ewers, 1991; Jury & Horton, 2004). Capillaire kanalen zijn ech-
ter een sterke vereenvoudiging van de werkelijke xyleemvaten (e.g. aanwezigheid van pitporien)
waardoor de maximale kh in de meeste soorten varieert tussen 20-100% van deze theoretische
waarde (Tyree & Ewers, 1991). De hydraulische geleidbaarheid van het xyleem (Kh) is een maat

10 Hoofdstuk 1. Literatuurstudie
Figuur 1.3: Illustratie van het pad dat water aflegt doorheen het bodem-plant-atmosfeer continuum
(SPAC, ’soil-plant-atmosphere continuum’)(rechts) en de corresponderende weergave van het
elektrisch analogon (links). Watertransport grijpt plaats onder invloed van een potentiaal-
gradient in de richting van dalende waterpotentialen (Ψ). Elk medium (wortel, stam, blad,
stomata en grenslaag (GL)) vertegenwoordigt een hydraulische weerstand (R). De totale ma-
croscopische weerstand wordt gezien als een serieschakeling van deze individuele weerstanden
(aangepast uit Cruiziat & Tyree (1990)).
voor het vermogen van het vasculair systeem om water en opgeloste stoffen te transporteren onder
optimale omstandigheden (Espino & Schenk, 2011). Hoe groter de hydraulische geleidbaarheid,
des te kleiner de transportweerstand die het sap ondervindt en des te meer water naar de kruin
kan stromen onder een gegeven waterpotentiaal. De maximale geleidbaarheid wordt bepaald door
de vatdiameter, het aantal xyleemvaten en de ruimtelijke verdeling van deze vaten (Durante et al.,
2011). Het xyleem opereert echter niet altijd onder optimale condities onder andere door obstruc-
ties (e.g. cavitaties, tyloses) in de xyleemvaten en afname van de stomatale geleiding (Tyree &
Sperry, 1989; Sevanto et al., 2005). De werkelijke hydraulische geleidbaarheid ligt dus meestal
lager dan de maximale waarde (Tyree & Sperry, 1988).
1.2.3 Opmeten van hydraulische geleidbaarheid
De standaard methode om de hydraulische geleidbaarheid te kwantificeren bestaat erin om een
ontgaste, gedestilleerde oplossing door het taksegment te sturen onder lage druk. De hydraulische

Hoofdstuk 1. Literatuurstudie 11
geleidbaarheid wordt vervolgens gedefinieerd als de ratio van het debiet en de drukgradient over de
sectie (formule 1.1) (Sperry & Tyree, 1988). Wanneer dergelijke meting wordt uitgevoerd onder
toenemende dehydratatie van de plant (benchdrying) zal de embolisatiegraad toenemen en de
hydraulische geleidbaarheid systematisch afnemen (Choat et al., 2010). Op die manier kan het
uitdrogingsprofiel aan de hand van een percentage verlies aan geleidbaarheid (percentage loss of
conductivity, PLC) uitgedrukt worden :
PLC = 100(1 −K ′hKh
) (1.3)
waarin K ′h en Kh respectievelijk de actuele en de maximale hydraulische geleidbaarheid voorstellen
[kg.s−1.MPa−1] (Kolb et al., 1996; Rosner et al., 2006; Choat et al., 2010).
Dergelijke methode kan uitgevoerd worden met een eenvoudige waterkolom waarin een gravitaire
gradient gecreeerd wordt doorheen de tak. Alternatief kan gebruik gemaakt worden van de
XYL’EM, gebaseerd op de methode van Sperry & Tyree (1988) en toegepast door onder meer
Cochard et al. (2000). Het werkingsprincipe wordt uiteengezet in sectie 2.4.
1.3 Stamdiametervariaties
Variaties in de stamdiameter van de plant resulteren uit een irreversibele groeigebaseerde compo-
nent enerzijds en een reversibele component ten gevolge van variaties in de plant-waterbalans
anderzijds (Irvine & Grace, 1997; Steppe et al., 2006; Perez-Diez et al., 2010). Reversibele
diametervariaties vloeien voort uit het aanvullen of uitputten van waterreserves in de plant.
Daarom worden de diametervariaties in combinatie met de wateropslagmechanismen besproken.
Er zijn drie opslagmechanismen te onderscheiden en elk van deze grijpt plaats bij verschillende
waterpotentialen in verschillende weefsels.
Elastische inkrimping correspondeert met het dagelijkse verloop van de waterpotentiaal en doet
zich voor ter hoogte van het floeemweefsel, de schors en de levende cellen in het xyleem (Figuur
1.2). Als reactie op een daling in de xyleemwaterpotentiaal in de voormiddag zal een radiale
sapstroom optreden vanuit deze weefsels om de waterbalans in evenwicht te brengen. Naarmate
de opslagpool overdag verder uitgeput raakt, krimpt de stam terwijl deze ’s avonds en ’s nachts
zal toenemen door het opnieuw aanvullen van de reserves. Dergelijke hydratatiecycli resulteren
in een diurnaal diameterpatroon dat in grootte bepaald wordt door de elasticiteit van de weefsels
(Irvine & Grace, 1997; Cochard et al., 2001; Intrigliolo & Castel, 2007; Almeras, 2008). Naast
levende weefsels ondergaat ook het dode xyleemweefsel dagelijks elastische variaties in omvang
(Irvine & Grace, 1997; Zweifel & Item, 2000; Cochard et al., 2001; Peramaki et al., 2001; Ueda
& Shibata, 2001; Zweifel et al., 2001; Rosner et al., 2009; Perez-Diez et al., 2010). Onder in-
vloed van toenemende zuigspanningen ondergaan xyleemvaten een inkrimping. Deze zogenaamde
’zuigspanningsinkrimping’ is een rechtstreeks gevolg van de cohesietheorie waarbij de adhesie-
krachten tussen water en xyleemwanden zorgen voor een inkrimping bij grote zuigspanningen.

12 Hoofdstuk 1. Literatuurstudie
Veranderingen in de waterpotentiaal worden dus op een directe wijze gereflecteerd in xyleemdia-
metervariaties (Irvine & Grace, 1997; Peramaki et al., 2001). Naast de elastische opslagcapaciteit
van de plant, kan ook capillariteit bijdragen tot de waterbalans, zij het onder erg kleine zuig-
spanningen. Deze reserve wordt reeds uitgeput rond -0.5 MPa, waardoor het belang voor de
transpiratiestroom gering is (Tyree et al., 1984).
Tenslotte kan cavitatie een rol spelen in de wateropslagcapaciteit van de plant. Wanneer een
xyleemvat caviteert, wordt het gevuld met lucht en wordt de volledige waterinhoud vrijgesteld in
de transpiratiestroom (Holtta et al., 2002, 2009). De mate waarin dit mechanisme een belangrijke
rol speelt in het bufferen van de waterbalans van de plant varieert van soort tot soort. Een
aantal auteurs oppert dat caviterende vaten de stamdiameterafname kortstondig tegengaan als
gevolg van het ontspannen van de zuigspanning (Irvine & Grace, 1997; Rosner et al., 2009, 2010;
Rosner & Karlsson, 2011). Holtta et al. (2009) stelt echter dat de capacitieve effecten als gevolg
van cavitatie in angiospermen erg weinig bijdragen tot diametervariaties door hun rigiditeit. Dit
is in overeenstemming met bevindingen van diverse andere auteurs (Schultz & Matthews, 1988;
Peramaki et al., 2001; Steppe et al., 2006; Intrigliolo & Castel, 2007; Holtta et al., 2009).
Sapstroom en diametervariaties zijn dus sterk gerelateerd aan elkaar en geven een indicatie van
de waterstatus van de plant. Onder condities van droogtestress zal het water sterker gebon-
den zijn aan de bodem, waardoor een grotere zuigspanning nodig is om eenzelfde sapstroom te
bewerkstelligen. De waterpotentiaal ter hoogte van het xyleem vertoont dus een sterkere daling,
waardoor de waterreserves dieper worden aangesproken. Toenemende stadia van uitdroging ver-
oorzaken op die manier een toenemende afname van de diameter, bovenop het diurnaal patroon.
De schorsdiameter (het weefsel buiten de cambiale zone) zal voornamelijk dalen als gevolg van
uitputting van de waterreserves terwijl het xyleem onder invloed van toenemende zuigspanning
zal krimpen (Sevanto et al., 2005; Holtta et al., 2007; Fernandez & Cuevas, 2010).
1.4 Cavitatie
Zoals blijkt uit de cohesietheorie schuilt de moeilijkheid van het watertransport niet in het over-
bruggen van het hoogteverschil tot aan de kruin, maar wel in het onderhouden van de cohesie-
krachten in de waterkolom (Cruiziat & Tyree, 1990). De heersende zuigspanningen waaraan het
water in de plant langdurig onderworpen wordt, zorgen er namelijk voor dat het water in een
metastabiele toestand verkeert. Dit betekent dat de aanwezigheid van een luchtbel voldoende
kan zijn om de cohesiekrachten tussen watermoleculen te verbreken. Dit heet het caviteren van
xyleemvaten en leidt tot verstoppingen of embolismen in het vasculair weefsel. Zo wordt het water-
continuum doorheen de plant verbroken en wordt de sapstroom gehinderd (Tyree & Zimmermann,
2001; Steppe, 2004; Jia et al., 2006; Choat et al., 2008).

Hoofdstuk 1. Literatuurstudie 13
1.4.1 Fysische oorzaken van cavitatie
Planten ondergaan dagelijks zuigspanningen die cavitatie kunnen induceren. Droogte, sterke
transpiratie, vries-en-dooicycli of pathogenen verhogen het risico op cavitatie en kunnen in extreme
mate leiden tot de dood van de plant (Tyree & Zimmermann, 2001; Steppe, 2004; Jia et al., 2006;
Choat et al., 2008). De voornaamste trigger voor cavitatie is droogtestress, gekenmerkt door
periodes van hoge transpiratie en/of bodemwatertekort. In deze thesis wordt dan ook gefocust
op cavitatie als gevolg van droogte. Vooraleer dieper ingegaan wordt op deze vorm van cavitatie
worden de twee andere mechanismen kort toegelicht.
De essentie van vriesgeınduceerde cavitatie is dat gas heel zwak oplosbaar is in bevroren water.
Bijgevolg zullen er tijdens het vriesproces gasbelletjes ontstaan uit het luchtverzadigde xyleemsap.
Die zullen doorgaans groter zijn in brede vaten omdat deze meer opgeloste lucht per eenheid lengte
bevatten (Wheeler et al., 2005). Wanneer het dooiproces aanvangt zullen de meeste gasbellen
opnieuw oplossen in de vloeistoffase. Gasbelletjes in grote vaten kunnen gemakkelijk samensmelten
en zo uitgroeien tot grote bellen. Grote gasbellen lossen echter trager op, waardoor de kans groter
is dat er nog lucht aanwezig is wanneer de kritieke zuigkracht bereikt wordt. Wanneer dit het
geval is, expandeert de gasbel en wordt het vat gecaviteerd. De gevoeligheid aan vriesgeınduceerde
cavitatie kan dus rechtstreeks gelinkt worden aan de vatdiameter (Tyree & Zimmermann, 2001;
Pittermann & Sperry, 2003).
Tenslotte kunnen ook pathogenen cavitatie veroorzaken. Deze scheiden toxines af waartegen de
plant zich beschermt door middel van de vorming van gezwellen die de xyleemvaten verstoppen.
Pathogenen kunnen ook zelf de vaten verstoppen door hun fysieke aanwezigheid of ze kunnen de
samenstelling van het xyleemsap wijzigen. Al deze factoren houden een groeiend risico op cavitatie
in (Tyree & Zimmermann, 2001; McElrone et al., 2003).
1.4.2 Ontstaan en verspreiding van cavitatie binnen de plant
Er zijn verschillende mechanismen die het voorkomen van cavitatie verklaren. Een eerste mecha-
nisme is het homogeen nucleatieprincipe. Dit principe berust op het spontaan ontstaan van een
luchtbel in een functionerend vat. Hiervoor zijn echter heel grote zuigspanningen nodig (boven
100 MPa) waardoor dit mechanisme minder relevant blijkt onder biologische omstandigheden (Ty-
ree & Zimmermann, 2001; Holtta et al., 2002; Cochard, 2006). Het heterogene nucleatieprincipe
kan reeds onder veel kleinere zuigspanningen plaatsgrijpen en cavitatie induceren. Hierbij worden
adhesiekrachten tussen watermoleculen en de vatwand verbroken ter hoogte van een onregelma-
tigheid of pitporie.
Het meest aanvaarde mechanisme voor de verspreiding van droogtegeınduceerde cavitatie is de ’air-
seeding’ hypothese (Sperry & Tyree, 1988; Cochard, 1992; Salleo et al., 1996; Tyree, 1997; Holtta
et al., 2002, 2005). Deze verklaart de verspreiding van cavitatie in aangrenzende xyleemvaten aan

14 Hoofdstuk 1. Literatuurstudie
de hand van de structuur van pitmembranen en wordt toegelicht aan de hand van figuur 1.4. In
figuur 1.4 A staan twee aangrenzende xyleemvaten in verbinding met elkaar via de pitporien op hun
laterale wanden. Omdat het ene gecaviteerde vat (V1) met lucht gevuld is en het andere vat (V2)
met water vormen zich lucht-water-menisci over hun gemeenschappelijke pitporien (Figuur 1.4 B).
In het functionele vat heerst een zuigkracht, terwijl het gecaviteerde vat zich op atmosfeerdruk
bevindt (Figuur 1.4 B en C). Hierdoor ontstaat een drukverschil over de menisci. Het maximale
drukverschil dat onderhouden kan worden zonder dat de meniscus doorslaat, wordt gegeven door
de wet van Laplace:
Pv − Px = 2σ/rp (1.4)
met Pv de absolute druk van de lucht [Pa], Px de absolute druk in het xyleem [Pa], σ opper-
vlaktespanning van water (0.0728 N.m−1 bij 20◦C) en rp kromtestraal van de pitporie. Onder
invloed van toenemende droogte of sterke transpiratie vergroot de zuigkracht in het functionele
vat en stijgt het drukverschil over de meniscus (Figuur 1.4 B en C). Indien dit drukverschil de
threshold (bepaald op basis van vergelijking 1.4) overschrijdt, slaat de meniscus door en wordt de
gasbel in het lumen van V2 gezogen (Figuur 1.4 D). Volgens de wet van Laplace kan een grotere
pitporie slechts een kleiner drukverschil onderhouden. Het maximaal te onderhouden drukver-
schil wordt dus bepaald door de grootste pitporie (grootste rp). Onder invloed van de heersende
zuigspanning expandeert de luchtbel instantaan en vult het volledige vat zich met waterdamp.
V2 is nu ook gecaviteerd en de druk is er gelijk aan de verzadigde dampdruk (2.3 kPa). Enkele
milliseconden later wordt de waterdamp vervangen door lucht op atmosfeerdruk (Cruiziat & Ty-
ree, 1990; Tyree & Zimmermann, 2001). Deze kan aangevoerd worden uit intercellulaire ruimten,
andere geemboliseerde vaten (V1), of diffunderen uit naburige geleidende vaten (Jackson & Grace,
1996). Op dat moment is het vat V2 geemboliseerd en disfunctioneel (Figuur 1.4 E en F) (Tyree
& Zimmermann, 2001; Jia et al., 2006). Een geemboliseerd vat kan dus naburige vaten aanzetten
tot cavitatie en zich zo voortzetten doorheen het volledige vasculaire systeem. De aanwezigheid
van pitporien remt deze verspreiding van lucht doorheen het vasculair systeem af en functioneert
zo als veiligheidsklep voor de plant (Cruiziat & Tyree, 1990; Jackson & Grace, 1996; Choat et al.,
2008). Dit is een eerder paradoxaal gegeven, gezien het net deze pitporien zijn die zorgen voor
een geıntegreerd hydraulisch netwerk.
1.4.3 Gevolgen van cavitatie
Cavitatie kan in extreme mate leiden tot ’runaway’ cavitatie (Milburn, 1973). Luchtgevulde va-
ten kunnen geen zuigspanningen doorgeven, waardoor de sapstroom niet meer kan passeren via
geemboliseerde vaten. Hierdoor ontstaat een daling in hydraulische geleidbaarheid, waardoor gro-
tere zuigspanningen nodig zijn om aan een constante transpiratievraag te voldoen. Dit induceert
op zijn beurt steilere drukgradienten en vergroot in een vicieuze cirkel het risico op cavitatie.
Het voorkomen van deze extensieve vorm van cavitatie is echter nog niet experimenteel bewezen
en kan een zeldzaam fenomeen zijn. Cavitatie is immers in veel planten een dagelijks fenomeen
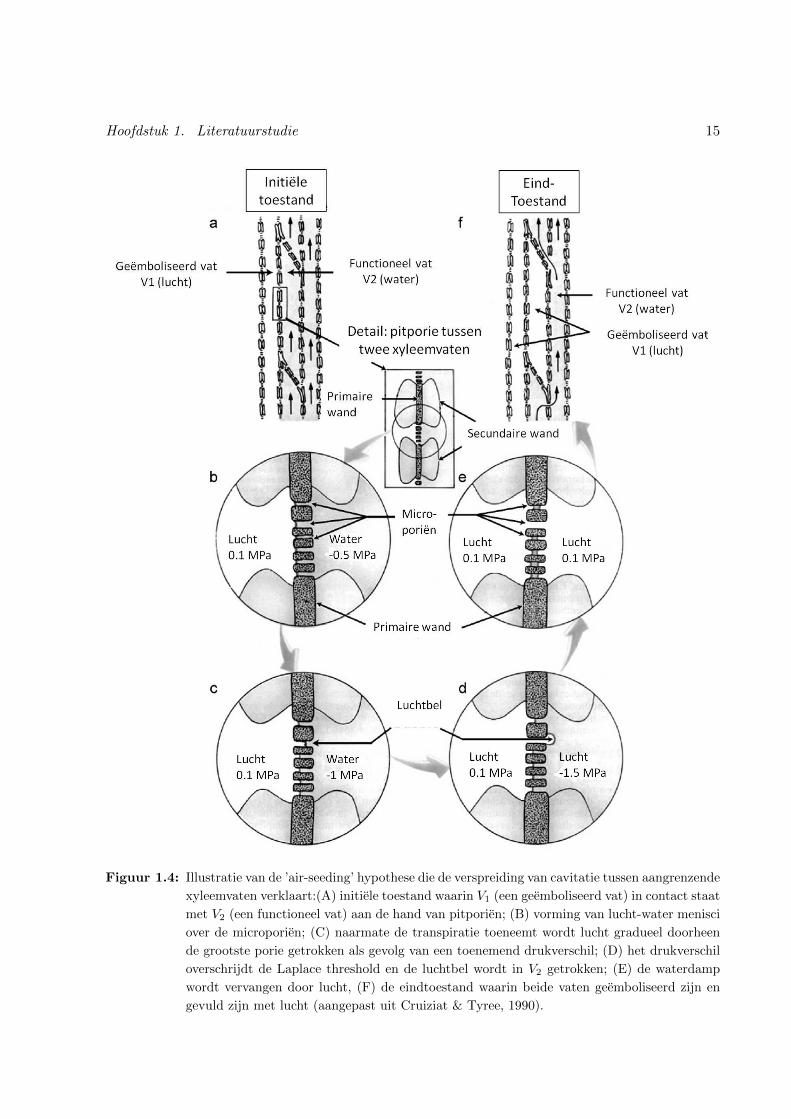
Hoofdstuk 1. Literatuurstudie 15
Figuur 1.4: Illustratie van de ’air-seeding’ hypothese die de verspreiding van cavitatie tussen aangrenzende
xyleemvaten verklaart:(A) initiele toestand waarin V1 (een geemboliseerd vat) in contact staat
met V2 (een functioneel vat) aan de hand van pitporien; (B) vorming van lucht-water menisci
over de microporien; (C) naarmate de transpiratie toeneemt wordt lucht gradueel doorheen
de grootste porie getrokken als gevolg van een toenemend drukverschil; (D) het drukverschil
overschrijdt de Laplace threshold en de luchtbel wordt in V2 getrokken; (E) de waterdamp
wordt vervangen door lucht, (F) de eindtoestand waarin beide vaten geemboliseerd zijn en
gevuld zijn met lucht (aangepast uit Cruiziat & Tyree, 1990).

16 Hoofdstuk 1. Literatuurstudie
waaraan de plant is aangepast. Ten gevolge van intensieve transpiratie rond de middag ondervindt
de waterpotentiaal een dip. Hierdoor stijgt de zuigspanning, wat een aantal vaten doet caviteren.
Deze dagelijkse embolisaties kunnen zich in de loop van de avond en nacht herstellen waardoor
een stabiele embolisatiecyclus doorlopen wordt (Cruiziat & Tyree, 1990; Cochard, 1992; Zufferey
et al., 2011). De meeste bomen kunnen niveaus van 5 - 20% verlies aan hydraulische geleidbaar-
heid verdragen vooraleer een onstabiele ’runaway’ toestand bereikt wordt (Tyree & Sperry, 1988;
Tyree & Ewers, 1991).
Naast het herstellen van gecaviteerde vaten trachten diverse planten ’runaway’ cavitatie en ver-
regaande dehydratatie te vermijden door middel van stomatale sluiting. Hierdoor vermindert de
transpiratie en kan de waterpotentiaal boven de threshold gehandhaafd worden (e.g. Kolb et al.,
1996; Salleo et al., 2001; Holtta et al., 2002, 2011; Zufferey et al., 2011). Deze stomatale respons
impliceert echter een reductie van de gasuitwisseling met de atmosfeer wat zich vertaalt in een
verlaagde fotosynthese. Op lange termijn kan dit leiden tot een koolstoftekort met nefaste ge-
volgen voor de groei en overleving van de plant. Diverse auteurs stellen dat het voordelig kan
zijn voor de plant om cavitatie in beperkte mate te tolereren, op voorwaarde dat de vaten tijdig
hervuld kunnen worden. Op die manier kan de stomatale sluiting uitgesteld worden, waardoor
meer koolstof kan worden opgenomen (Tyree & Zimmermann, 2001; Holtta et al., 2009; Choat
et al., 2010; Zufferey et al., 2011).
1.4.4 Herstelmechanismen
Gezien het dagelijks voorkomen van cavitatie, moeten planten een mechanisme hebben ontwik-
keld voor het hervullen van gecaviteerde xyleemvaten. Op die manier kan de sapstroom terug
hersteld worden en wordt de plant vrijwaard van systematisch verlies aan hydraulische geleid-
baarheid (Brodersen et al., 2010). Ondanks talrijk bewijs van dergelijk proces bleven de details
ervan gedurende vele jaren een raadsel (e.g. Salleo et al., 1996).
Recente experimenten, waaronder in vivo waarneming door middel van rontgenstraling in Vitis
vinifera L., hebben een beter inzicht gegeven in dit herstelmechanisme (Zwieniecki et al., 2000;
Brodersen et al., 2010). Brodersen et al. (2010) stelden een model op om het herstel van cavitatie
in druivelaars te beschrijven (Figuur 1.5). Hierin wordt het herstelmechanisme toegeschreven aan
tijdelijk opgebouwde drukgradienten tussen xyleem en omringende weefsels, die tot stand komen
door een samenspel van (i) de instroom van water en opgeloste stoffen, (ii) heersende potentialen
en (iii) de bevochtigbaarheid van xyleemwanden (Bucci et al., 2003; Brodersen et al., 2010). Ca-
vitatieherstel blijkt een dynamisch proces waarbij de plant, via actieve participatie van het floeem
en parenchymcellen, een opmerkelijke controle heeft over de stroming en distributie van water
(Lovisolo et al., 2008). Brodersen et al. (2010) stellen verder dat het herstelmechanisme in de
praktijk gelijktijdig en gecoordineerd plaatsgrijpt binnen een groepje xyleeemvaten. Deze onder-
gaan een hydraulische isolatie, waardoor een lokale overdruk gecreeerd kan worden. Het transport
van opgeloste stoffen gaat door totdat alle kanalen binnen deze groep van xyleemvaten gevuld zijn.

Hoofdstuk 1. Literatuurstudie 17
Op die manier worden de pas hervulde vaten niet meteen aangesloten op de sapstroom, waardoor
lokaal aangevoerde stoffen niet meteen verloren gaan in naburige vaten. Dit maakt cavitatieher-
stel mogelijk tijdens de transpiratie (Tyree, 1997; Zwieniecki et al., 2000; Brodersen et al., 2010;
Zufferey et al., 2011). Deze gecoordineerde vorm van herstel zou gestimuleerd worden door diffuse
signalen zoals vibraties en planthormonen (Salleo et al., 1996; Bucci et al., 2003; Lovisolo et al.,
2008). Geemboliseerde vaten worden over een tijdspanne van een aantal uur opgevuld, afhankelijk
Figuur 1.5: Model van het herstelmechanisme van embolismen in de xyleemvaten van druivelaars.(A1,
B1) Onder invloed van embolismen scheidt het floeem lokaal opgeloste stoffen af die via pa-
renchymcellen in het xyleemkanaal terechtkomen. (A2, B2) Hierdoor wordt een osmotische
gradient gecreeerd waardoor water spontaan toestroomt vanuit naburige vezels en parenchym-
cellen en het kanaal binnenkomt via talrijke porien. (A3-6, B3-6) Initieel worden waterdrup-
pels gevormd op de binnenwand van het kanaal. (C1-3)Deze smelten samen en expanderen
waardoor het aanwezige gas gevangen en samengedrukt wordt in progressief kleinere bellen
(4a-4b) tot ze oplossen of ontsnappen via hydrofobe microkanalen in de vatwand. S: opgeloste
stoffen, W: water. (Brodersen et al., 2010).
van hun diameter en voorafgaande waterstatus. Hoe groter de diameter, hoe langer het opvullen
duurt, waardoor de kans stijgt dat het vat nog niet is opgevuld tegen het begin van de volgende
transpiratiecyclus. In dit geval zal het herstelmechanisme falen (Salleo et al., 1996; Brodersen
et al., 2010; Zufferey et al., 2011).

18 Hoofdstuk 1. Literatuurstudie
1.4.5 Opmeten van cavitatie
In een caviterend xyleemvat wordt het sap dat onder zuigspanning staat op een aantal microse-
conden verdreven en vervangen door waterdamp (Tyree & Sperry, 1988; Jia et al., 2006; Rosner
et al., 2006). Deze plotse drukverandering zorgt voor de productie van energie onder de vorm van
ultrasone akoestische signalen (’Ultrasonic Acoustic Emissions’, UAE). Met behulp van deze be-
vindingen legden Milburn & Johnson (1966) de beginselen vast voor een non-destructieve methode
om cavitatie op te meten. Met behulp van een ultrasone cavitatiesensor kunnen de cavitatiesigna-
len gedetecteerd worden en kan de stresstoestand van de plant in beeld gebracht worden (Cruiziat
& Tyree, 1990; Jackson & Grace, 1996; Cochard, 2006; Rosner et al., 2006).
Interpretatie van cavitatiesignalen
De interpretatie van de UAE-signalen is echter niet eenvoudig. Diverse factoren dienen in acht
te worden genomen voordat conclusies kunnen getrokken worden. Zo wordt de signaalsterkte en
-voortplanting beınvloed door de weglengte die het signaal moet afleggen, de dichtheid van het
hout, de orientatie van de vezels en de hydratatietoestand van het staal (Rosner et al., 2006; Mayr
& Rosner, 2011). Verder kunnen diverse andere processen in levende planten ook UAE uitstu-
ren, waardoor vaak geen 1:1 relatie voorkomt tussen het aantal UAE en het aantal gecaviteerde
vaten (Rosner et al., 2006). In principe kan elke (dode) watergevulde cel die voldoende sterke
wanden heeft om negatieve drukken te weerstaan, caviteren. In angiospermen kunnen dus ook
parenchymcellen en tracheıden bijdragen tot UAE-signalen. Verder kunnen reflecties van golven
of droogtescheuren aan de basis liggen van deze niet-lineaire relatie. Vaten kunnen ten slotte ook
hervuld worden zodat ze meermaals UAE produceren. Deze fenomenen bemoeilijken de interpre-
tatie van de UAE resultaten. Veel auteurs nemen aan dat UAE-signalen hoofdzakelijk afkomstig
zijn van cavitatie, maar deze kunnen niet met zekerheid als enige oorzaak worden aangeduid
(Sperry & Tyree, 1988; Jackson & Grace, 1996; Kikuta & Salleo, 1997; Rosner et al., 2010).
Bij de interpretatie van de UAE moet ook rekening gehouden worden met de vatdiameter. Zoals
voorgeschreven door de wet van Poiseuille (formule 1.2) is de sapstroom evenredig met de diameter
tot de vierde macht. Cavitatie in een groter vat is dus hydraulisch significanter en zal resulteren
in een sterkere daling in hydraulische geleidbaarheid. Corresponderend wordt verondersteld dat
grotere vaten onder zuigspanning meer elastische energie opslaan waardoor ze energetisch hogere
UAE waarden produceren. Gezien het aantal UAE als parameter niet differentieert tussen energe-
tische signaalsterkten, is dit onderscheid tussen vatdiameters moeilijk te maken. De relatie tussen
het cumulatief aantal UAE en het verlies aan geleidbaarheid zal bijgevolg niet lineair zijn, aange-
zien niet alle cellen even groot zijn en dus niet gelijkmatig bijdragen aan de totale hydraulische
conductiviteit (Rosner et al., 2006).
Het is duidelijk dat de interpretatie van het aantal UAE enige voorzichtigheid vereist (Jackson
& Grace, 1996). Rosner et al. (2006) is van mening dat een meer gesofisticeerde analyse van de

Hoofdstuk 1. Literatuurstudie 19
UAE-signalen noodzakelijk is om de hydraulische gevoeligheid te beschrijven (Mayr & Rosner,
2011).
1.4.6 Cavitatie: trade-off veiligheid versus hydraulische geleidbaarheid
Uit de ’air-seeding’ hypothese (sectie 1.4.2) blijkt dat kleine pitporien een grote barriere betekenen
voor de verspreiding van cavitatie en als veiligheidklep dienen in de verspreiding van cavitatie.
Kleine pitporien gaan echter gepaard met een functionele kost. De sapstroom ondervindt namelijk
een weerstand in de xyleemvaten als gevolg van (i) een lumenweerstand en (ii) een porieweerstand.
Wijde porien zijn dus cruciaal voor efficient watertransport, maar ze impliceren gelijktijdig een
risico op cavitatie, wat hen inherent gevoeliger maakt aan droogtegeınduceerde cavitatie. In
houtachtige planten bestaat dus een trade-off tussen xyleemefficientie en xyleemresistentie voor
cavitatie (Kolb & Davis, 1994; Rosner et al., 2006; Loepfe et al., 2007; Choat et al., 2008; Rosner
et al., 2008; Christman et al., 2009; Rosner et al., 2009; Cai & Tyree, 2010; Holtta et al., 2011).
Initieel werd verondersteld dat de pitporositeit aan de basis lag van deze trade-off en ’getuned’
moest worden door de plant om de hydraulische structuur te optimaliseren. Empirische data
onderschrijven deze stelling echter niet altijd (Hacke et al., 2006).
Verschillende auteurs hebben dit fenomeen onderzocht. Zo verklaren Choat et al. (2008) deze
afwijking door middel van het principe van de ’air-seeding’ theorie. De cavitatiethreshold wordt
namelijk bepaald door de maximale poriediameter in plaats van de porositeit waarmee de ge-
middelde poriediameter wordt uitgedrukt. De hydraulische geleidbaarheid op zijn beurt wordt
in mindere mate beınvloed door de aanwezigheid van een paar grote porien en is bijgevolg wel
gerelateerd aan de porositeit. De hydraulische trade-off is volgens hen dus toe te schrijven aan
een verband tussen porositeit, relevant voor de hydraulische geleidbaarheid en maximale porie-
grootte, gerelateerd aan cavitatie (Hacke et al., 2006). In zijn pitoppervlaktehypothese relateert
Wheeler et al. (2005) de maximale poriegrootte aan de pitmembraanoppervlakte per vat. Een
grotere oppervlakte betekent immers meer pitporien en op basis van de probabiliteitstheorie dus
meer kans op een grote porie. De gemiddelde porositeit van de pitmembranen verandert daar-
entegen niet noodzakelijk mee met de pitoppervlakte. Planten zouden dus beter beschermd zijn
tegen cavitatie wanneer ze minder pitoppervlakte per kanaal bezitten. Dit heeft ook consequen-
ties naar geleidbaarheid toe gezien minder pitoppervlakte een grotere hydraulische weerstand
betekent. Aangezien de pitmembraanoppervlakte in directe relatie staat met de vatdiameter en
-lengte (Wheeler et al., 2005; Hacke et al., 2006), komt een kleinere diameter overeen met minder
pitmembraan en dus een grotere veiligheid voor cavitatie. Anderzijds heeft een kleinere diameter
een nefaste invloed op de hydraulische geleidbaarheid, gekwantificeerd door de wet van Poiseuille
(formule 1.2). Loepfe et al. (2007) trekt de analyse ten slotte open naar systeemniveau. Een
belangrijk aandeel van de sapstroomweerstand is namelijk toe te schrijven aan de connectiviteit
tussen de vaten, naast de porie- en lumenweerstand. Hierdoor schrijven de auteurs de trade-off
toe aan een combinatie van zowel pitporiegrootte als vatlengte en connectiviteit (Loepfe et al.,

20 Hoofdstuk 1. Literatuurstudie
2007; Cai & Tyree, 2010; Durante et al., 2011).
1.5 De vulnerability curve
De gevoeligheid van het xyleem voor cavitatie kan grafisch afgebeeld worden aan de hand van een
vulnerability curve (VC). Hierop wordt het verlies aan hydraulische geleidbaarheid weergegeven
in functie van de waterpotentiaal nodig om dit verlies te induceren. De relatie tussen beide para-
meters verloopt sigmoıodaal en kan gefit worden aan een Chapman curve (y0 +a (1−exp(−bx))c).
Gevoeligere vaten caviteren bij minder negatieve waterpotentialen. Hoe negatiever de waterpo-
tentiaal wordt, hoe meer vaten disfunctioneel worden door cavitatie en hoe lager de hydraulische
geleidbaarheid wordt. De VC geeft dus een beeld van de droogtegevoeligheid van een plant. Om
VC eenvoudig te kunnen interpreteren maakt men vaak gebruik van de P50. Hiermee wordt de
waterpotentiaal aangeduid die overeenkomt met een 50% verlies aan geleidbaarheid (e.g. Cai &
Tyree, 2010; Choat et al., 2010).
Een VC is geldig voor een soort. De spatiale verdeling van de pitporositeit tussen de verschillende
xyleemvaten van een plant verklaart de sigmoıdale vorm. Niet alle vaten hebben immers dezelfde
poriegroottedistributie en caviteren dus niet allemaal bij eenzelfde waterpotentiaal (Choat et al.,
2008). De procedure om een standaard VC op te stellen, is tijdrovend en vereist destructieve
staalname. Dit dient bovendien steeds in labo’s te worden uitgevoerd wat langetermijnopvol-
ging bemoeilijkt. Daarom wordt in deze studie nagegaan of niet-destructieve VC hier een uitweg
kunnen bieden.

MATERIAAL EN METHODE


Hoofdstuk 2
Materialen en Methode
2.1 Experimentele proefopzet
De experimenten werden uitgevoerd op eenjarige druivelaars (Vitis vinifera cv. Chardonnay),
opgekweekt op de faculteit bio-ingenieurswetenschappen in Gent (figuur 2.1). In het najaar werden
ze toegelaten in de buitenlucht te groeien. Op 12 maart 2012 werden ze verhuisd naar een serre,
waar ze vanaf dan permanent verbleven en tweemaal per week van water voorzien werden.
Figuur 2.1: Opstelling van de druivelaars in de serre (links), aluminium zakjes om de transpiratie stop te
zetten (rechts boven) en doorsnede van een tak (rechts onder).
23

24 Hoofdstuk 2. Materialen en Methode
Per experiment werden voor zonsopgang twee taksegmenten verzameld (50-90 cm in lengte en 0.5-
0.9 cm in diameter) op basis van vergelijkbare eigenschappen (lengte, gewicht, bladeren). Deze
werden zo snel mogelijk een tweede keer afgeknipt onder water om luchtintrede in de xyleemvaten
te vermijden. Vervolgens werden ze getransporteerd naar het labo in een vochtige, zwarte plas-
tiekzak. Op die manier werd transpiratie en waterverlies verhinderd (Schultz & Matthews, 1988).
In het labo werden ze gedurende ten minste 1 uur in een donkere ruimte geplaatst om de tak in
evenwicht te laten komen (Cochard et al., 2001).
De proefopstelling bestond uit drie verschillende onderdelen en dus drie aparte taksegmenten (fi-
guur 2.2). De eerste tak werd op een balans bevestigd om het gewichtsverlies bij uitdroging te
kunnen opvolgen. De tweede tak werd in twee stukken geknipt. Van een deel werden de blaadjes
gebruikt om de stamwaterpotentialen op te meten. Het andere deel werd periodiek aan het wa-
terreservoir gekoppeld en hiertoe vastgezet met klemmen. Op deze tak werden additioneel vier
sensoren bevestigd: (i) een cavitatiesensor, (ii) twee LVDT-sensoren (’Linear Variable Displace-
ment Transducer’) en (iii) een temperatuursensor (figuur 2.3).
Figuur 2.2: Schematische weergave van de voorbereiding van de proefopstelling. Tak 1 wordt op een ba-
lans geplaatst om het waterverlies bij toenemende dehydratatie op te meten. Tak 2 wordt in
twee delen verdeeld: blaadjes van deel a worden gebruikt in de drukbom om waterpotentiaal-
metingen uit te voeren. Op het resterende deel b van tak 2 worden 4 sensoren geplaatst: (i)
een cavitatiesensor (zwart cirkeltje), (ii) twee LVDT-sensoren (’Linear Variable Displacement
Transducer’, pijltjes) en (iii) een temperatuursensor (streepje).
De voorbereiding duurde maximaal 1.5 uur en in die tussentijd werden alle takuiteinden in para-
film gewikkeld om waterverlies tegen te gaan (Hietz et al., 2008). De vier geınstalleerde sensoren
verzamelden data en schreven per seconde een gemiddelde waarde weg naar de datalogger (NI
USB-6009, National Instruments,Zaventem, Belgie) (figuur 2.3). Om geleidbaarheidsdata te be-
komen werd de tak gemiddeld om het uur, vijf minuten aan een externe watertoevoer gehangen.
Tegelijkertijd werden het gewicht en de waterpotentialen opgemeten. Om eenzelfde uitdrogings-
profiel te bekomen werden ook de andere twee takken telkens vijf minuten aan water gekoppeld.
Naarmate toenemende dehydratatiestadia doorlopen werden, daalde de hydraulische geleidbaar-

Hoofdstuk 2. Materialen en Methode 25
heid. Praktisch betekende dit dat er minder water doorheen de tak kon stromen onder eenzelfde
drukverschil. Daarom werden de takken naar het einde toe langer aangesloten aan externe water-
toevoer om een meetbare hoeveelheid water te kunnen opvangen (tot 30 minuten). Hiervoor werd
uiteraard gecorrigeerd in de berekening van de hydraulische geleidbaarheid.
Figuur 2.3: Gedetailleerd overzicht van het benchdrying experiment: een druiventak waarop diverse sen-
soren bevestigd zijn (pijltjes) wordt horizontaal vastgeklemd tussen twee statieven. Verder
wordt het microklimaat opgemeten met behulp van een relatieve-vochtigheidssensor en een
temperatuursensor, die rechts gepositioneerd zijn, net buiten het beeld van de foto
Verschillende experimenten werden uitgevoerd in de periode tussen 20 september 2011 (DOY
263) tot 28 mei 2012 (DOY 148). Bepaalde aspecten van de experimentele procedure werden
licht aangepast en geoptimaliseerd bij opeenvolgende experimenten in het kader van praktische
bevindingen. Deze worden weergegeven in tabel 2.1. Relevante verschillen in de procedure tussen
diverse experimenten zullen daarenboven duidelijk aangegeven worden in de betreffende delen van
hoofdstuk 3 en 4.
2.2 Microklimaat
De heersende T en RH bepalen de mate en snelheid waarmee de tak zal uitdrogen. Het zijn
bijgevolg sleutelvariabelen bij het correct interpreteren van de resultaten. Om deze klimatologische
variabelen op te meten werd gebruik gemaakt van:

26 Hoofdstuk 2. Materialen en Methode
� een thermokoppel (type PCS Schaevitz 10/94 LBB-312-PA-020)
� een RH-sensor (Rotronic, USA)
De relatieve vochtigheid geeft aan hoeveel waterdamp een luchtmengsel bevat en wordt gedefini-
eerd als de ratio van de werkelijke dampdruk en de verzadigde dampdruk van het luchtmengsel
bij omgevingstemperatuur. Uit de combinatie van T en RH kan het VD bepaald worden (sectie
1.1.2). Het is een maat voor de drogingskracht van lucht en staat rechtstreeks in verband met de
transpiratie. Mathematisch kan het VD worden weergegeven als het verschil tussen de verzadigde
dampdruk (e0v [kPa]) en de werkelijke dampdruk bij omgevingstemperatuur (ev [kPa]):
V D = e0v − ev (2.1)
waarin de verzadigingsdampdruk bepaald wordt door de luchttemperatuur (T [◦C]):
e0v = 0.6108 exp(
17.27 T
T + 237.3) (2.2)
en werkelijke dampdruk een fractie uitmaakt van de verzadigingsdampdruk (Goldstein et al., 1998;
Steppe et al., 2004):
ev = e0v
RH
100(2.3)
Tabel 2.1: Overzicht van de verschillende gehanteerde technieken voor (i) opmeten van de hydraulische
geleidbaarheid en (ii) het opmeten van de waterpotentiaal bij vier opeenvolgende experimen-
ten. De andere variabelen werden op identieke wijze opgevolgd. Alle experimenten werden
uitgevoerd op druivelaars (Vitis vinifera L. cv. Chardonnay).
Naam DOY Gehanteerde techniek
Hydraulische geleidbaarheid Waterpotentiaal
Experiment 1 335-350 (2011) XYL’EM Tak uiteindje
Experiment 2 16-20 (2012) XYL’EM Tak uiteindje
Experiment 3 79-85 (2012) XYL’EM Tak uiteindje
Experiment 4 85-97 (2012) Horizontale gravitatiekolom Blad
Experiment 5 100-107 (2012) Verticale gravitatiekolom Blad
Experiment 6 115-122 (2012) Verticale gravitatiekolom Blad
Experiment 7 136-148 (2012) - -
DOY, dag van het jaar
2.3 Waterpotentiaal
De diversiteit aan fysische of chemische krachten die op het water werken, zorgen ervoor dat de
waterpotentiaal (Ψw [MPa]) ontleed kan worden in vier componenten:
Ψw = Ψg + Ψp + Ψm + Ψπ (2.4)

Hoofdstuk 2. Materialen en Methode 27
met Ψg de gravitationele potentiaal, Ψp de hydrostatische potentiaal, Ψm de matrixpotentiaal en
Ψπ de osmotische potentiaal (e.g. Tyree & Zimmermann, 2001). De gravitationele component
houdt rekening met het verschil in hoogte t.o.v. het referentieniveau. Deze component is enkel
significant bij verticale waterkolommen groter dan 1 m en bijgevolg verwaarloosbaar in taksegmen-
ten. De osmotische potentiaal is een maat voor de chemische aantrekking van water door opgeloste
stoffen. De matrixpotentiaal drukt de capillaire krachten uit die gepaard gaan met lucht-water-
menisci. Deze wordt vaak gecombineerd met de hydrostatische potentiaal en voorgesteld als de
drukcomponent van de totale waterpotentiaal (Jones, 1992; Tyree & Zimmermann, 2001; Jury &
Horton, 2004).
2.3.1 Werkingsprincipe van de drukbom
De Scholander-drukbom (PMS Instrument Company, Albany, USA) kwantificeert de drukcom-
ponent van de totale waterpotentiaal op een afgesneden blaadje of taksegment. De osmotische
component is meestal verwaarloosbaar, waardoor men bovendien een indicatie krijgt van de totale
waterpotentiaal (Turner, 1988).
Figuur 2.4: (A) Schematische voorstelling van de Scholander-drukbom om de waterpotentiaal van plan-
tensegmenten op te meten en (B) opstelling van de drukbom met bijhorende gasfles waarmee
de druk kan worden opgebouwd in de drukkamer tot wanneer de waterkolom ter hoogte van
het snijvlak verschijnt.
Water in een intact blad kan zich in twee compartimenten bevinden: in de levende celinhoud
(protoplast, Ψpw) of in celwanden en xyleemvaten (de apoplast, Ψa
w). Wanneer transpirerende
bladeren een evenwichtstoestand hebben bereikt (waterverlies = wateropname), staan beide com-
partimenten op eenzelfde totale waterpotentiaal (Ψpw=Ψa
w), maar de bijdrage van de verschillende
componenten is verschillend (figuur 2.5 A). Het celsap in levende cellen bevat een hoge concentra-
tie opgeloste stoffen en heeft dus een negatieve osmotische potentiaal (Ψpπ). Deze opgeloste stoffen
oefenen een aantrekkingskracht uit op het water en genereren op die manier een turgordruk, die
een positieve drukpotentiaal (Ψpp) genereert. Water in de apoplast bevat een lagere concentratie

28 Hoofdstuk 2. Materialen en Methode
opgeloste stoffen waardoor de osmotische potentiaal er veel kleiner is (Ψaπ). De cellulose mi-
crofibrillen ter hoogte van de celwanden genereren echter capillaire krachten die leiden tot een
matrixpotentiaal (Ψam). Daarnaast staan de xyleemvaten onder een zuigspanning ten gevolge van
de vertraagde wateropname door de wortels in vergelijking met het waterverlies. Aangezien het
onmogelijk is om het onderscheid te maken tussen capillaire krachten in celwanden en zuigspan-
ning in xyleemvaten, worden beide termen gesubstitueerd in een totale negatieve drukcomponent
(Ψap). Het samenspel van deze componenten wordt weergegeven in figuur 2.5 A.
Figuur 2.5: Diagram van de verschillende componenten van de waterpotentiaal in (A) een intact blad
en (B) een afgesneden blad in de drukbom. Ψm,Ψp en Ψπ duiden respectievelijk op de
matrixpotentiaal, de drukpotentiaal en de osmotische potentiaal waarbij superscript a voor
de apoplast staat en superscript b voor de protoplast. Pgas is de aangelegde druk in de
drukkamer.
2.3.2 De drukbom in de praktijk
De avond voor de start van het benchdrying experiment werden de intacte blaadjes in aluminium
zakjes verpakt om ervoor te zorgen dat ze in evenwicht kwamen met de stam. Zo boden de
potentiaalmetingen op de bladeren een indicatie van de stamwaterpotentiaal (e.g. Choat et al.,
2010). Wanneer het blad werd afgeknipt, werd het snijvlak blootgesteld aan atmosfeerdruk.
Onder invloed van de heersende negatieve waterpotentiaal in de apoplast trok het xyleemsap
zich weg van het snijvlak, in de omringende cellen. De xyleemvaten werden gevuld met lucht en
de drukcomponent Ψap in de apoplast viel weg (figuur 2.5 B). Het staal werd na afsnijden zo snel

Hoofdstuk 2. Materialen en Methode 29
mogelijk omgekeerd in de drukbom geplaatst, zodat het snijvlak doorheen het rubberen luchtdicht
afsluitelement stak (figuur 2.4 B). Op die manier werd de transpiratie stopgezet en verdeelden de
drukgradienten zich in het staal. Vervolgens werd de druk binnenin de kamer opgebouwd door
middel van gecomprimeerd gas. Onder invloed van deze uitwendige druk steeg de drukcomponent
in de protoplast en werd het water terug in de apoplast gedwongen. De waterkolom werd terug
omhoog gedrukt in de xyleemvaten en op het moment dat de meniscus van de waterkolom ter
hoogte van het snijvlak opdook, compenseerde de uitwendige druk de oorspronkelijke zuigkracht in
het xyleem exact in grootte en kon deze laatste dus worden afgelezen op de naald van de drukbom
(figuur 2.5 B) (Turner, 1988; Wei et al., 1999). De stamwaterpotentiaalmetingen werden om het
uur uitgevoerd vanaf de start van het benchdrying experiment tot wanneer de potentiaal een
waarde van 5 MPa overschreed en onbetrouwbaar werd. Aangezien er in de winter geen blaadjes
voorhanden waren, werd het uiteinde van de intacte tak ten minste 12 uur voor het experiment
artificieel afgedicht met silicone (TEC 7, Novatech N.V., Olen, Belgie). Nadat het water in de tak
in evenwicht was gekomen in de donkere ruimte, werd het uiteindje afgeknipt (5 cm) onder water.
Op die manier ontstond een taksegment waarop stamwaterpotentiaalmetingen uitgevoerd konden
worden.
2.4 Hydraulische geleidbaarheid
Voor het opmeten van de hydraulische geleidbaarheid werden twee methoden gehanteerd. In de
experimenten tot en met 26 maart 2012 (DOY 85) werd de XYL’EM gebruikt. Ten gevolge van
technische problemen werd vanaf dan overgeschakeld op de gravitatiekolom. Beide technieken
berusten echter op hetzelfde principe gepostuleerd door Sperry & Tyree (1988) (sectie 1.2.3).
2.4.1 Opmeten van de hydraulische geleidbaarheid: XYL’EM
De hydraulische geleidbaarheid van een taksegment (50 - 90 cm) werd gekwantificeerd aan de hand
van de XYL’EM (Bronkhorst, Clermont-Ferrand-Theix, France). Dit bestaat uit een hogedrukvat
(typisch 150 kPa), een lagedrukvat (typisch 3-10 kPa) en een debietmeter (liquiflow, 0.1-5 g.h−1).
Lage- en hogedrukmetingen werden uitgevoerd met gefilterd (Whatman filter papers, 41) Volvic
water waaraan 10 mM KCl en 1 mM CaCl2 werden toegevoegd (van Ieperen et al., 2000; van Doorn
et al., 2011). Dit werd op voorhand ontgast onder laboratoriumvacuum met een magnetische
roerder gedurende ten minste 2 uur en vervolgens door een membraanfilter gestuurd (Liqui-cel mini
module, Membrana GmbH, Wuppertal, Duitsland) (Espino & Schenk, 2011). Tenslotte passeerde
het water nogmaals twee filters (Nalgene, 0.45 MIC, 25 mm) en werd finaal bij kamertemperatuur
(15-22 ◦C) doorheen de tak gestuurd. Na 1-5 minuten werd het debiet afgelezen wanneer de waarde
min of meer constant bleef (van Doorn et al., 2011). De hydraulische geleidbaarheid werd hieruit
afgeleid aan de hand van formule 1.1.

30 Hoofdstuk 2. Materialen en Methode
2.4.2 De XYL’EM in de praktijk
Het basale uiteinde van de tak werd ontdaan van schors- en floeemweefsel en herhaaldelijk her-
sneden onder water. De groene onderdelen van de tak (blaadjes, twijgen,...) werden ingepakt
in aluminiumfolie om versnelde transpiratie tegen te gaan. Vervolgens werd dit uiteinde aan de
hand van plastieken buisjes verbonden aan het XYL’EM lagedrukwaterreservoir. Lagedrukme-
tingen verzekerden dat embolismen niet op een artificiele manier verwijderd werden en gaven dus
de heersende hydraulische geleidbaarheid weer. Hogedrukmetingen (flushen) hadden net tot doel
deze embolismen op te lossen om zo de maximale hydraulische geleidbaarheid te kwantificeren.
Het flushen van lange stalen veroorzaakte echter problemen waarvan de oorsprong nog niet hele-
maal duidelijk is (o.a. verstoppen van de vaten, zwelling van de pitporien). Hierdoor ontstond er
een plotse afname in de geleidbaarheid na het flushen en waren deze resultaten onbetrouwbaar.
De hogedrukmetingen werden bijgevolg niet gebruikt om de maximale hydraulische geleidbaar-
heid te bepalen. Er werd echter aangenomen dat de initiele Kh-waarde, de maximale hydrauli-
sche geleidbaarheid voorstelde, aangezien (i) de tak voor zonsopgang is afgeknipt onder water en
nadien niet meer toegelaten werd om te transpireren en (ii) omdat de plant voldoende werd be-
waterd. Tussen twee metingen door werd het takje toegelaten uit te drogen in de omgevingslucht
(benchdrying), zodat een brede range aan waterpotentialen bereikt werd. Dit zorgde ervoor dat
de embolisatiegraad en de hydraulische geleidbaarheid systematisch respectievelijk toe- en afna-
men. De gemeten hydraulische geleidbaarheid in dergelijke stadia werd dan uitgedrukt als een
percentage van de initiele maximale waarde (formule 1.3). Tenslotte werd de verkregen waarde
nog gecorrigeerd voor veranderingen in viscositeit ten gevolge van temperatuurschommelingen
(K20 = KT 3.49399.3252+√
54.2176−TT+32.7425 )(Cochard, 2008).
De metingen werden uitgevoerd op takken met een lengte die de maximale vatlengte overschreed
(50 - 90 cm). Wanneer dit niet het geval is, riskeert men dat vaten aan beide kanten opengesne-
den worden. Op die manier fungeert de tak als open buis, waar het water rechtstreeks doorheen
kan stromen, onafhankelijk van de aanwezigheid van gecaviteerde vaten en wordt de hydraulische
geleidbaarheid overschat. Deze maximale vatlengte werd opgemeten door luchtinjectie in een on-
dergedompelde tak. De tak werd stelselmatig korter afgeknipt tot wanneer een continue stroom
luchtbelletjes tevoorschijn kwam. De maximale vatlengte werd dan gedefinieerd als de lengte van
de overblijvende tak (e.g. Skene & Balodis, 1968; Sperry et al., 1987). Resultaten varieerden tus-
sen 41 en 67 cm in overeenstemming met bevindingen van Choat et al. (2010) waarin de lengte
varieerde tussen 50 - 90 cm. Andere auteurs melden waarden die varieren tussen 11.6 cm (Jacob-
sen & Pratt, 2012) en 12.8 cm (Wheeler et al., 2005). Aangezien deze nog kleiner zijn dan onze
waarden, is de gehanteerde taklengte een conservatieve waarde en bijgevolg een goede aanname.
Gezien problemen werden ondervonden bij zowel hoge- als lagedrukmetingen werd de XYL’EM
aan een aantal technische testen onderworpen. Hierin werd vastgesteld dat (i) de debietmeter een
dalende stroming registreerde, ondanks een constante weerstand en drukverschil en (ii) dat de
debietmeter een stroming aangaf die in grootte niet overeenkwam met de werkelijke waterstroom.

Hoofdstuk 2. Materialen en Methode 31
Omdat vermoed werd dat deze problemen deels aardden in verstopping van de buisjes door algen
en particulair materiaal, werden de buisjes uit het toestel geautoclaveerd en onderworpen aan een
javelbehandeling. Bij verder gebruik werd het zeer omzichtig gehanteerd om nieuwe onzuiverhe-
den zoveel mogelijk te voorkomen. De testresultaten bleken echter nog niet consistent, waardoor
besloten werd over te schakelen op een tweede techniek om de hydraulische geleidbaarheid op te
meten.
2.4.3 Opmeten van de hydraulische geleidbaarheid: gravitatiekolom
Het principe van de gravitatiekolom berust, net zoals de XYL’EM, op het induceren van een
waterstroming doorheen de tak, die in dit geval wordt opgewekt met behulp van een constante
gravitaire drukhoogte (figuur 2.6). Een reservoir met gedestilleerd water werd op 1.5 m boven de
tak geplaatst en verbonden via een waterkolom op de tak. Aan de hand van een overloopme-
chanisme en een constante druppelsgewijze watertoevoer werd een constante drukhoogte van 1 m
opgebouwd (9.79 kPa). Per meting werd het gedestilleerd water gedurende een bepaald tijdsinter-
val opgevangen onderaan de tak en afgewogen op een nauwkeurige balans (Radwag, 0.01g). Uit
de gewichtstoename per tijdsinterval kon het massadebiet doorheen de tak berekend worden en in
combinatie met de drukhoogte werd finaal de hydraulische geleidbaarheid bekomen (formule 1.1).
2.5 Diametervariaties
Stamdiametervariaties worden opgenomen met behulp van een ’Linear Variable Displacement
Transducer’ (LVDT) sensor. Dit is een elektrische transducer waarmee een mechanische verplaat-
sing vertaald wordt in een elektrisch signaal [Volt] dat naar een datalogger wordt gestuurd. Het
gemeten voltage kan dan aan de hand van kalibratie terug omgerekend worden tot een fysische
diametervariatie in millimeter.
2.5.1 Werkingsprincipe van de LVDT-sensor
Een LVDT-sensor bestaat uit een staaf die tegen de tak wordt geplaatst aan een zijde en vrij kan
bewegen naar de andere kant. Op het einde van deze staaf hangt een beweegbare ferromagne-
tische, cilindervormige kern. Rond deze kern zijn drie spoelen bevestigd: een primaire spoel en
twee secundaire spoelen (figuur 2.7 A). Het geheel wordt op zijn plaats gehouden door middel van
een op maat gemaakte houder die aan de stam wordt bevestigd met elastiekjes. Een veer zorgt
voor nauw contact zodat de staaf de stamdiametervariaties mooi kan opvolgen.
Er wordt een wisselstroom aangelegd over de primaire spoel die een voltage in beide secundaire
spoelen induceert. Deze laatste zijn serieel verbonden, maar in omgekeerde volgorde, waardoor
hun voltages een tegengestelde polariteit bezitten. Het outputsignaal van de LVDT-sensor is het

32 Hoofdstuk 2. Materialen en Methode
Figuur 2.6: (A)Opstelling van de gravitatiekolom waarin de tak in verticale positie aan (B) het waterre-
servoir gekoppeld wordt met behulp van (C) een druppelmechanisme. De pijltjes duiden op
(a) de aansluiting van de waterkolom aan de tak , (b) de overloopbuis voor overtollig water,
(c) het schaaltje om het water op te vangen, (d) het waterreservoir op 1 m drukhoogte, (e)
de wateroverloop (f) en het druppelmechanisme.
verschil tussen deze secundaire voltages. Wanneer de kern beweegt, varieren de geınduceerde vol-
tages in de secundaire spoelen en dus ook het spanningsverschil. Wanneer de kern zich in het
centrum bevindt, dus symmetrisch t.o.v. beide spoelen, wordt in deze secundaire spoelen een
spanning geınduceerd, gelijk in grootte en tegengesteld in teken (figuur 2.7 A). Het spannings-
verschil is bijgevolg gelijk aan nul (figuur 2.7 B). Wanneer de stam vervolgens uitzet of inkrimpt,
beweegt de magnetische kern in de richting van een van de twee spoelen. Het spanningsverschil
stijgt van nul tot een maximale waarde (binnen het bereik van de sensor). Een positie van de
kern volledig rechts of links komt overeen met de respectievelijke positie 100% (+) en 100% (-)
(figuur 2.7 B). De grootte van dit spanningsverschil is proportioneel met de afstand afgelegd door
de kern: hoe verder de kern zich verplaatst, hoe groter het outputsignaal. De term ’linear’ verwijst
naar dit fenomeen. De fase van het outputvoltage wordt bepaald door de richting waarin de kern
beweegt. Op deze manier biedt de LVDT-sensor een rechtstreekse maat voor de kwantificatie van
diametervariaties.

Hoofdstuk 2. Materialen en Methode 33
Figuur 2.7: Vereenvoudigde weergave van het werkingsprincipe van de LVDT-sensor (aangepast uit
Steppe et al., 2004). (A) De LVDT-sensor bestaat uit een ferromagnetische kern, omringd
door drie spoelen waarover een wisselspanning wordt gezet. (B) Bij uitzetten of inkrimpen
van de stam beweegt de kern waardoor een spanningsverschil ontstaat. Verschillende posities
van de kern resulteren in een verschillend spanningsverschil.
2.5.2 De LVDT-sensor in de praktijk
Per tak werden twee LVDT-sensoren bevestigd, waarvan een op de schors en een rechtstreeks op
het xyleem. Ter hoogte van de sensor op het xyleem werden de schors, het floeem en het cambium
zorgvuldig weggesneden zodat de effecten van deze levende weefsels geelimineerd werden. Vanaf
DOY 85 werd het xyleem onder de sensor ingewreven met vaseline zodat er geen lokaal versnelde
uitdroging plaatsgreep.
2.6 Cavitatie
2.6.1 Opmeten van cavitatie: ultrasone akoestische emissies
Caviterende vaten stoten ultrasone akoestische emissies (UAE [dB]) uit die te detecteren zijn
met een cavitatiesensor (Milburn & Johnson, 1966; Tyree et al., 1984). Deze bestaat uit een
’AE-signal conditioner with peak detector’ (ASCO-P, Vallen Systeme GmbH, Duitsland) en een
piezo-elektrische sensor (VS150-M, Vallen Systeme, GmbH, Duitsland). De sensor meet signalen
op binnen een frequentierange van 90 - 295 kHz. Deze range is in overeenstemming met de hui-
dige literatuur, waar gesteld wordt dat de hoogste amplitude van de cavitatiesignalen een sterke
frequentiecomponent heeft in de range van 100 - 300 kHz (e.g. Tyree & Sperry, 1989).

34 Hoofdstuk 2. Materialen en Methode
2.6.2 Werkingsprincipe van de cavitatiesensor
De ruwe signalen die de piezo-elektrische sensor opving, werden versterkt en ongewenste frequen-
ties werden weggefilterd. Het resterende signaal werd omgezet naar een logaritmische vorm en
werden door een ’peak stretcher’ gestuurd. Deze hield de piekwaarden bij en verlengde ze over 50
milliseconden (’APK’,’Acoustic Emission Peak Amplitude’). Dit APK-signaal geeft de piekampli-
tude weer over een vooropgesteld tijdsinterval en wordt uitgedrukt in Volt. Het is in staat snelle,
momentane signalen op te vangen zoals deze geproduceerd door caviterende vaten. Per seconde
werd op die manier een gemiddelde van 1000 APK-signalen bijgehouden. Om de signalen in dB
om te rekenen werd gebruik gemaakt van volgende formule:
dBAE =VAE 100
4(2.5)
met dBAE het UAE-signaal uitgedrukt in dB en VAE het UAE-signaal uitgedrukt in Volt.
2.6.3 Cavitatiesensor in de praktijk
Omdat de cavitatiesensor gevoelig is aan drukverschillen, werd deze op het volledig verzadigde
houtstaal geplaatst met behulp van een klemmechanisme zoals voorgeschreven in Rosner et al.
(2006) (figuur 2.8). Met behulp van veren en een draaischroef werd op die manier een constante
druk bekomen op de piezo-elektrische sensor en werden de signalen niet beınvloed door een ver-
minderde druk naarmate de tak inkromp. Daarnaast werd een dunne laag vaseline aangebracht
tussen de sensor en de tak om het akoestische contact te verbeteren. Het geheel werd tenslotte
afgeschermd van achtergrondlawaai met behulp van mastiek.
2.6.4 Gevoeligheid van de cavitatiesensor
Gezien diverse processen in de plant geluid kunnen veroorzaken, diende onderscheid gemaakt te
worden tussen caviterende vaten en achtergrondruis. Hiertoe werd een minimale signaalintensiteit
vooropgesteld. Het cumulatieve aantal UAE met een intensiteit groter dan deze cavitatiethreshold,
biedt een indicatie van de mate van cavitatie in de plant. In de literatuur worden varierende
waarden gebruikt tussen 25 en 35 dB (Zeugin, 2004; Jia et al., 2006; Mayr & Rosner, 2011). In
deze thesis werd de grenswaarde experimenteel vastgesteld door de sensor vrij in de lucht te hangen
zodat geen verstoring optrad. De intensiteit van de omgevingsruis schommelde tussen 27 en 32 dB.
De detectiethreshold werd gelijkgesteld aan de ondergrens plus 8 dB (Rosner et al., 2006). Zo werd
een waarde van 35 dB bekomen waarmee gediscrimineerd kon worden tussen achtergrondsignalen
en signalen die voortkwamen uit caviterende vaten.
Tot slot diende ook de verspreiding van de signalen doorheen het hout getest te worden, aangezien
die beınvloed wordt door eigenschappen als waterinhoud en orientatie van de houtvezels. Dit werd
onderzocht door middel van een potloodtest (e.g. Jackson & Grace, 1996). Hierin werd de sensor
op een houten tak bevestigd. Vervolgens werd de stift van een vulpotlood op toenemende afstand

Hoofdstuk 2. Materialen en Methode 35
Figuur 2.8: Detail van de cavitatiesensor, geınstalleerd op een tak met behulp van mastiek, vaseline en
een klemmechanisme om constante druk uit te oefenen.
van de sensor geplaatst onder een hoek van ongeveer 30° en met zachte druk afgebroken op het
hout. Dit veroorzaakte een kunstmatig UAE-signaal dat door de sensor gedetecteerd kon worden.
Naarmate de afstand vergrootte, nam de intensiteit van het signaal af tot op het moment dat dit
niet meer te onderscheiden viel van de achtergrondruis (Mayr & Rosner, 2011).


RESULTATEN


Hoofdstuk 3
Resultaten
In de volgende secties wordt eerst de algemene dynamiek besproken van het cavitatie- en diame-
terpatroon opgemeten tijdens het ’benchdrying’ experiment. De parameters worden geanalyseerd
in het licht van de heersende omgevingsfactoren en de algemene trend wordt besproken (sectie
3.1 - 3.3). Vervolgens wordt vanuit de destructieve VC de brug geslagen tussen ultrasone cavita-
tiesignalen en xyleemfluctuaties om tot een niet-destructieve VC te komen (sectie 3.4 - 3.5). Er
wordt gewerkt met data uit zes verschillende experimenten, die analoog verliepen tenzij anders
vermeld. Daarom zullen de resultaten als een geheel besproken worden.
3.1 Microklimaat
Het verloop van de ecofysiologische parameters wordt in belangrijke mate gecontroleerd door het
microklimaat. Daarom is het van cruciaal belang om een aantal microklimatologische variabelen
op te meten. In figuur 3.1 A en C wordt de T [◦C] van de tak (grijs) en de omgeving (zwart)
afgebeeld voor twee verschillende experimenten. In figuur 3.1 B en D worden de RH ([%], grijs,
rechter y-as) en het VD ([kPa], zwart, linker y-as) weergegeven.
De T en VD-data uit experiment 3 (figuur 3.1 C en D) lagen doorheen het gehele experiment
ongeveer 20% hoger dan deze uit experiment 2 (figuur 3.1 A en B). De T schommelde op dagelijkse
basis met maxima gedurende de dag en waarden van de tak die enigszins hoger lagen dan de
omgeving. Het VD vertoonde een verloop analoog aan de T met pieken overdag en dalen gedurende
de avond en nacht. Het VD staat in nauwe relatie tot de transpiratie waarbij een hoge VD waarde
(dus een lage RH) een sterke drijvende kracht betekent (e.g. Leonardi et al., 2000; Steppe et al.,
2004). Op die manier beınvloedt het VD de waterrelaties in de plant met sterke implicaties
voor het verloop van de waterpotentiaal, het diameter- en cavitatiepatroon en de hydraulische
geleidbaarheid. Het VD is dan ook een sleutelvariabele in deze analyse.
39
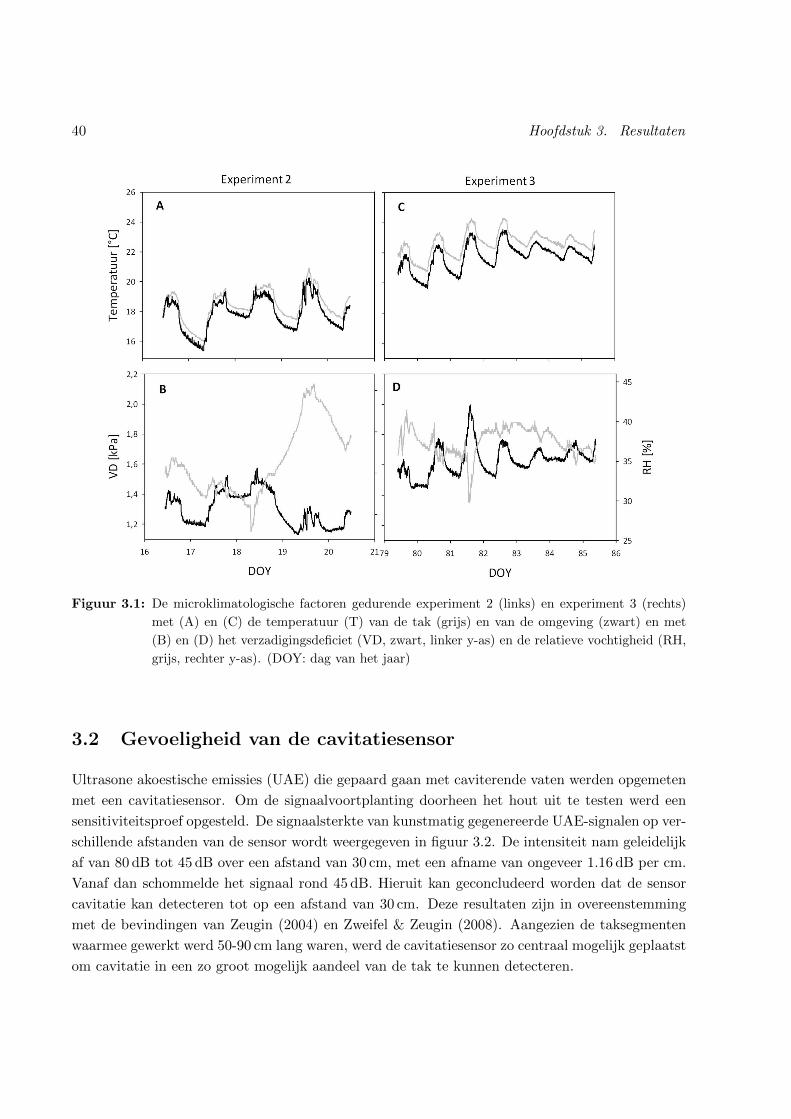
40 Hoofdstuk 3. Resultaten
Figuur 3.1: De microklimatologische factoren gedurende experiment 2 (links) en experiment 3 (rechts)
met (A) en (C) de temperatuur (T) van de tak (grijs) en van de omgeving (zwart) en met
(B) en (D) het verzadigingsdeficiet (VD, zwart, linker y-as) en de relatieve vochtigheid (RH,
grijs, rechter y-as). (DOY: dag van het jaar)
3.2 Gevoeligheid van de cavitatiesensor
Ultrasone akoestische emissies (UAE) die gepaard gaan met caviterende vaten werden opgemeten
met een cavitatiesensor. Om de signaalvoortplanting doorheen het hout uit te testen werd een
sensitiviteitsproef opgesteld. De signaalsterkte van kunstmatig gegenereerde UAE-signalen op ver-
schillende afstanden van de sensor wordt weergegeven in figuur 3.2. De intensiteit nam geleidelijk
af van 80 dB tot 45 dB over een afstand van 30 cm, met een afname van ongeveer 1.16 dB per cm.
Vanaf dan schommelde het signaal rond 45 dB. Hieruit kan geconcludeerd worden dat de sensor
cavitatie kan detecteren tot op een afstand van 30 cm. Deze resultaten zijn in overeenstemming
met de bevindingen van Zeugin (2004) en Zweifel & Zeugin (2008). Aangezien de taksegmenten
waarmee gewerkt werd 50-90 cm lang waren, werd de cavitatiesensor zo centraal mogelijk geplaatst
om cavitatie in een zo groot mogelijk aandeel van de tak te kunnen detecteren.
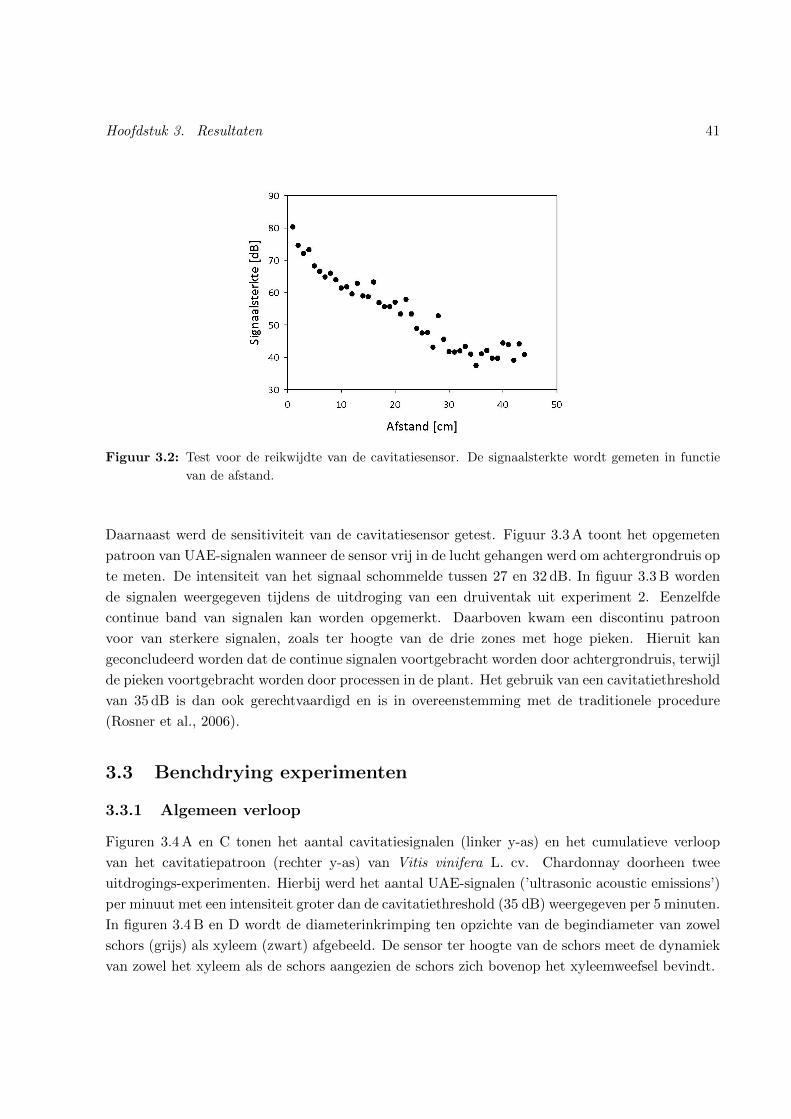
Hoofdstuk 3. Resultaten 41
Figuur 3.2: Test voor de reikwijdte van de cavitatiesensor. De signaalsterkte wordt gemeten in functie
van de afstand.
Daarnaast werd de sensitiviteit van de cavitatiesensor getest. Figuur 3.3 A toont het opgemeten
patroon van UAE-signalen wanneer de sensor vrij in de lucht gehangen werd om achtergrondruis op
te meten. De intensiteit van het signaal schommelde tussen 27 en 32 dB. In figuur 3.3 B worden
de signalen weergegeven tijdens de uitdroging van een druiventak uit experiment 2. Eenzelfde
continue band van signalen kan worden opgemerkt. Daarboven kwam een discontinu patroon
voor van sterkere signalen, zoals ter hoogte van de drie zones met hoge pieken. Hieruit kan
geconcludeerd worden dat de continue signalen voortgebracht worden door achtergrondruis, terwijl
de pieken voortgebracht worden door processen in de plant. Het gebruik van een cavitatiethreshold
van 35 dB is dan ook gerechtvaardigd en is in overeenstemming met de traditionele procedure
(Rosner et al., 2006).
3.3 Benchdrying experimenten
3.3.1 Algemeen verloop
Figuren 3.4 A en C tonen het aantal cavitatiesignalen (linker y-as) en het cumulatieve verloop
van het cavitatiepatroon (rechter y-as) van Vitis vinifera L. cv. Chardonnay doorheen twee
uitdrogings-experimenten. Hierbij werd het aantal UAE-signalen (’ultrasonic acoustic emissions’)
per minuut met een intensiteit groter dan de cavitatiethreshold (35 dB) weergegeven per 5 minuten.
In figuren 3.4 B en D wordt de diameterinkrimping ten opzichte van de begindiameter van zowel
schors (grijs) als xyleem (zwart) afgebeeld. De sensor ter hoogte van de schors meet de dynamiek
van zowel het xyleem als de schors aangezien de schors zich bovenop het xyleemweefsel bevindt.
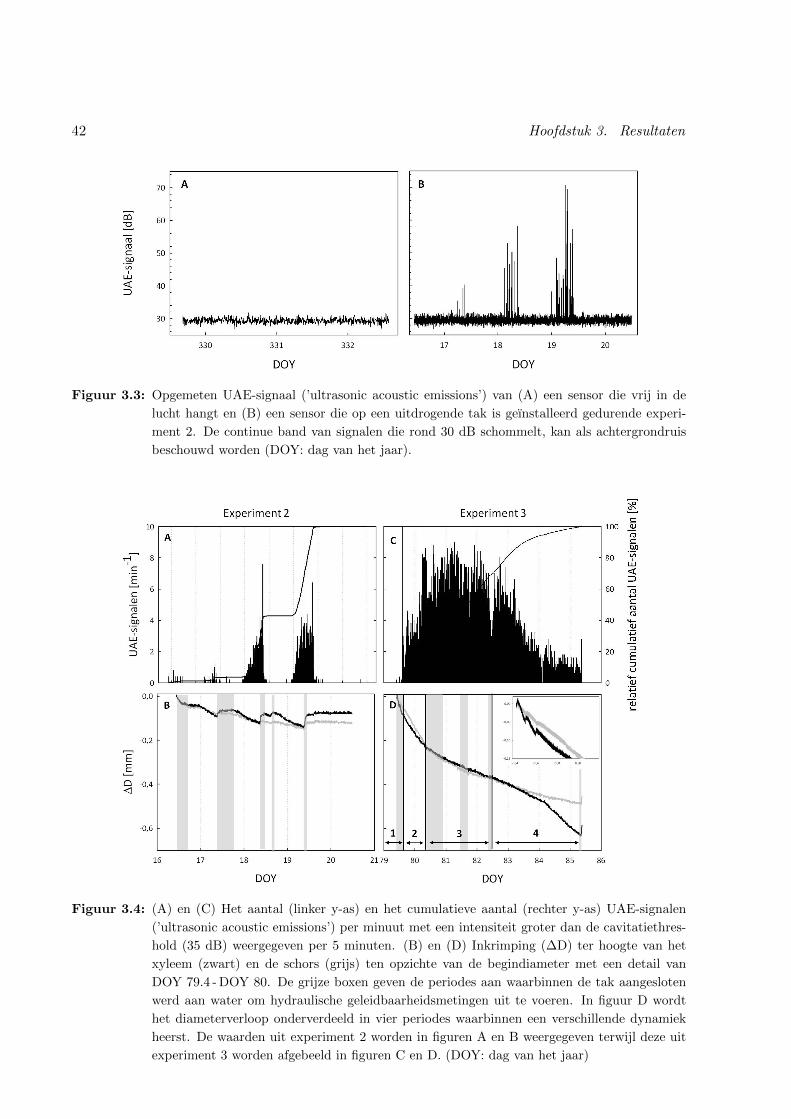
42 Hoofdstuk 3. Resultaten
Figuur 3.3: Opgemeten UAE-signaal (’ultrasonic acoustic emissions’) van (A) een sensor die vrij in de
lucht hangt en (B) een sensor die op een uitdrogende tak is geınstalleerd gedurende experi-
ment 2. De continue band van signalen die rond 30 dB schommelt, kan als achtergrondruis
beschouwd worden (DOY: dag van het jaar).
Figuur 3.4: (A) en (C) Het aantal (linker y-as) en het cumulatieve aantal (rechter y-as) UAE-signalen
(’ultrasonic acoustic emissions’) per minuut met een intensiteit groter dan de cavitatiethres-
hold (35 dB) weergegeven per 5 minuten. (B) en (D) Inkrimping (∆D) ter hoogte van het
xyleem (zwart) en de schors (grijs) ten opzichte van de begindiameter met een detail van
DOY 79.4 - DOY 80. De grijze boxen geven de periodes aan waarbinnen de tak aangesloten
werd aan water om hydraulische geleidbaarheidsmetingen uit te voeren. In figuur D wordt
het diameterverloop onderverdeeld in vier periodes waarbinnen een verschillende dynamiek
heerst. De waarden uit experiment 2 worden in figuren A en B weergegeven terwijl deze uit
experiment 3 worden afgebeeld in figuren C en D. (DOY: dag van het jaar)

Hoofdstuk 3. Resultaten 43
3.3.2 Cavitatiedynamiek
De cavitatiedynamiek in de twee experimenten verschilt in termen van patroon en grootteorde.
Daar waar cavitatie bij experiment 2 in twee goed te onderscheiden periodes optrad (figuur 3.4 A),
kwam cavitatie continu voor tijdens experiment 3 (figuur 3.4 C). Hier namen de signalen in een
eerste fase snel toe tot wanneer een maximale hoeveelheid van 8 signalen per minuut bereikt werd.
Dit hoge aantal hield 3 dagen aan en nam in een tweede fase geleidelijk af naar een uitgesproken
lagere waarde van ongeveer 2 signalen per minuut.
Daarnaast was de hoeveelheid cavitatiesignalen geproduceerd in experiment 3 aanzienlijk groter
dan deze in experiment 2. Daar waar zich in totaal 21 268 signalen voordeden in experiment
3, traden er slechts 1 993 op in experiment 2, wat een aandeel uitmaakt van 9%. De duur van
experiment 2 bedroeg maar 80% van die van experiment 3, maar zelfs na correctie voor deze ver-
schillende termijn bleef de cavitatieintensiteit beduidend lager in experiment 2 (9.6%). Tenslotte
manifesteerde cavitatie zich sneller in experiment 3 (na 4.5 h in vergelijking met 18 h in experiment
2). Tijdens de beginfase van de uitdroging (in experiment 2 tot DOY 17.2 en in experiment 3 tot
DOY 79.6) waren de cavitatiesignalen gering. Uit analyse van de data werd een nauwe overeen-
komst ontdekt tussen het tijdstip van aan- en afkoppeling van de watertoevoer en het voorkomen
van deze kleine hoeveelheid UAE-signalen. Het genereren van een waterstroom doorheen de tak
zorgde namelijk voor een lichte verstoring van de opstelling, waardoor dergelijke schokken wellicht
als cavitatiesignalen opgevangen werden door de cavitatiesensor. Daarom worden de weinige sig-
nalen die toch voorkwamen tijdens de beginfase als een artefact van de experimentele procedure
beschouwd en werd er aangenomen dat er weinig of geen cavitaties zijn opgetreden.
3.3.3 Diameterverloop in relatie tot cavitatie tijdens beperkte uitdroging
Het verloop in figuur 3.4 B en D vertoont een algemeen dalend verloop met een aantal duidelijk
stijgende periodes (e.g. DOY 17.4 in figuur 3.4 B). Deze dynamiek was ook aanwezig in het begin
van het experiment 3 (zoals weergegeven door het detail) en in experiment 5 (figuur 3.5). Deze
uitzettingen waren het gevolg van een geınduceerde waterstroming doorheen de tak door middel
van de XYL’EM. De tijdstippen waarop dergelijke stroming werd aangelegd, worden per dag omvat
door een grijze rechthoek. Zoals blijkt uit figuren 3.4 D en 3.5 resulteerde watercontact niet langer
in een diameteruitzetting wanneer een inkrimping van meer dan 0.15 mm had plaatsgevonden
(e.g. grijze rechthoek ter hoogte van DOY 80.4). Daarnaast verschilde de onderlinge respons van
schors- en xyleemweefsel tussen verschillende experimenten. Daar waar de diameteruitzetting zich
het sterkst manifesteerde ter hoogte van het xyleemweefsel in experiment 2 en 3 (figuur 3.4 B en D)
was het omgekeerde waar te nemen in experiment 5 (figuur 3.5). Hierop wordt dieper ingegaan in
sectie 3.3.4. Verschillen tussen xyleem- en schorsweefsel worden hier tijdelijk buiten beschouwing
gelaten en er wordt gefocust op de xyleemdiameter gezien deze, overeenkomstig de huidige kennis,
duidelijk in relatie staat tot de waterpotentiaal en dus tot het cavitatiepatroon (Irvine & Grace,
1997; Offenthaler et al., 2001; Peramaki et al., 2001; Holtta et al., 2005).
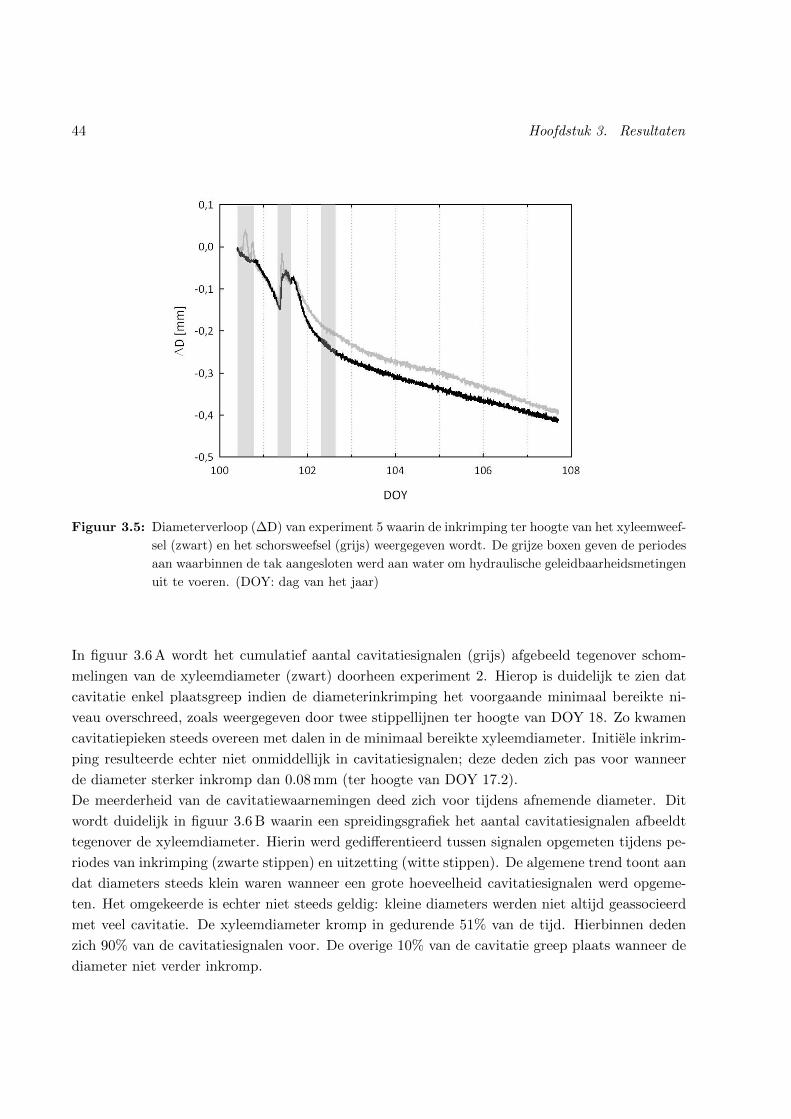
44 Hoofdstuk 3. Resultaten
Figuur 3.5: Diameterverloop (∆D) van experiment 5 waarin de inkrimping ter hoogte van het xyleemweef-
sel (zwart) en het schorsweefsel (grijs) weergegeven wordt. De grijze boxen geven de periodes
aan waarbinnen de tak aangesloten werd aan water om hydraulische geleidbaarheidsmetingen
uit te voeren. (DOY: dag van het jaar)
In figuur 3.6 A wordt het cumulatief aantal cavitatiesignalen (grijs) afgebeeld tegenover schom-
melingen van de xyleemdiameter (zwart) doorheen experiment 2. Hierop is duidelijk te zien dat
cavitatie enkel plaatsgreep indien de diameterinkrimping het voorgaande minimaal bereikte ni-
veau overschreed, zoals weergegeven door twee stippellijnen ter hoogte van DOY 18. Zo kwamen
cavitatiepieken steeds overeen met dalen in de minimaal bereikte xyleemdiameter. Initiele inkrim-
ping resulteerde echter niet onmiddellijk in cavitatiesignalen; deze deden zich pas voor wanneer
de diameter sterker inkromp dan 0.08 mm (ter hoogte van DOY 17.2).
De meerderheid van de cavitatiewaarnemingen deed zich voor tijdens afnemende diameter. Dit
wordt duidelijk in figuur 3.6 B waarin een spreidingsgrafiek het aantal cavitatiesignalen afbeeldt
tegenover de xyleemdiameter. Hierin werd gedifferentieerd tussen signalen opgemeten tijdens pe-
riodes van inkrimping (zwarte stippen) en uitzetting (witte stippen). De algemene trend toont aan
dat diameters steeds klein waren wanneer een grote hoeveelheid cavitatiesignalen werd opgeme-
ten. Het omgekeerde is echter niet steeds geldig: kleine diameters werden niet altijd geassocieerd
met veel cavitatie. De xyleemdiameter kromp in gedurende 51% van de tijd. Hierbinnen deden
zich 90% van de cavitatiesignalen voor. De overige 10% van de cavitatie greep plaats wanneer de
diameter niet verder inkromp.
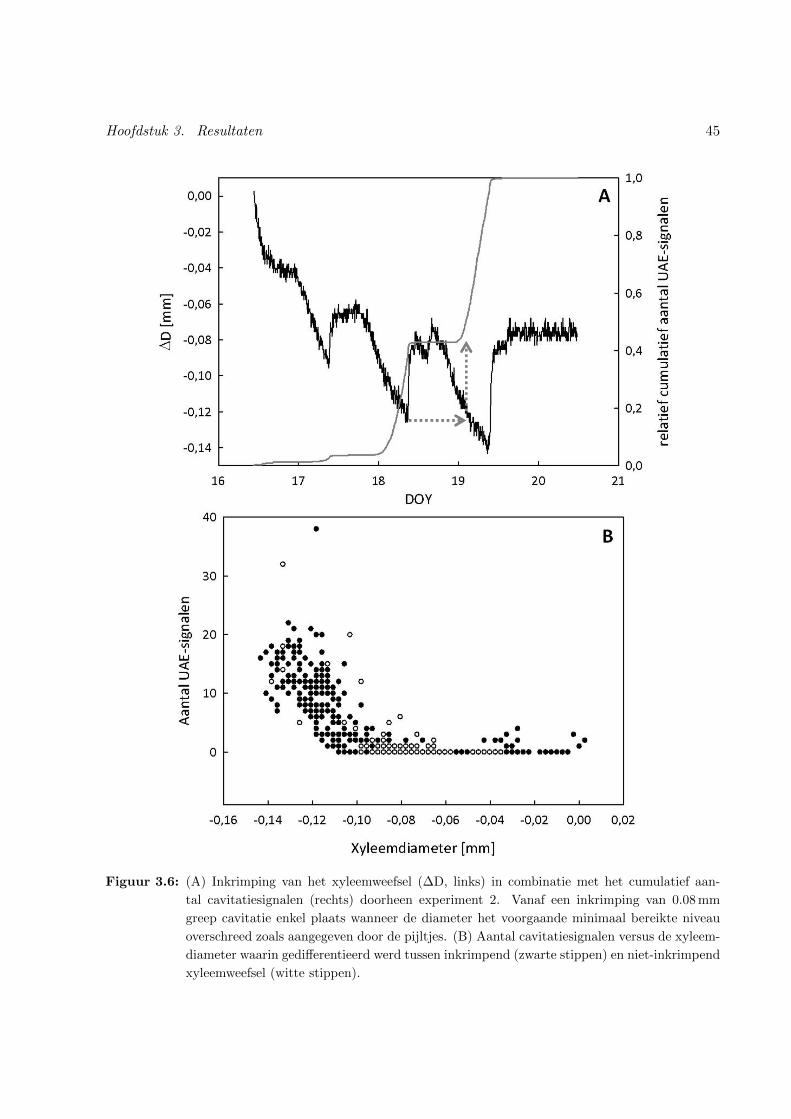
Hoofdstuk 3. Resultaten 45
Figuur 3.6: (A) Inkrimping van het xyleemweefsel (∆D, links) in combinatie met het cumulatief aan-
tal cavitatiesignalen (rechts) doorheen experiment 2. Vanaf een inkrimping van 0.08 mm
greep cavitatie enkel plaats wanneer de diameter het voorgaande minimaal bereikte niveau
overschreed zoals aangegeven door de pijltjes. (B) Aantal cavitatiesignalen versus de xyleem-
diameter waarin gedifferentieerd werd tussen inkrimpend (zwarte stippen) en niet-inkrimpend
xyleemweefsel (witte stippen).

46 Hoofdstuk 3. Resultaten
3.3.4 Diameterverloop in relatie tot cavitatie tijdens sterke uitdroging
Wanneer de diameter doorheen een lange, intense uitdrogingsperiode beschouwd werd, kon het
verloop in vier periodes onderverdeeld worden, zoals weergegeven in figuur 3.4 D. Binnen elke
periode dreven andere mechanismen de inkrimping, mechanismen die telkens op een andere manier
aan de waterrelaties gekoppeld konden worden.
Tijdens de eerste periode trad nog geen cavitatie op. Zowel schors- als xyleemweefsel fluctueerden
met de periodiek aangelegde waterstroming zoals ook waar te nemen in experiment 2 en 5 ( figuur
3.4 B en 3.5). Het xyleem vertoonde echter een sterkere uitzettingsdynamiek in vergelijking met
de schors in experiment 2 en 3. Dit is in strijd met de huidige literatuur waar gesteld wordt
dat het xyleemweefsel meer rigide is en dus minder varieert in diameter (Schultz & Matthews,
1988; Irvine & Grace, 1997; van Ieperen et al., 2000; Steppe, 2004). In de hypothese dat het
blootgestelde xyleemweefsel ter hoogte van de LVDT-sensor artificieel sneller zou uitdrogen, werd
vanaf experiment 4 vaseline aangebracht om deze versnelde uitdroging tegen te gaan (Alsina
et al., 2007). Vanaf dan waren de resultaten conform de verwachtingen, waarbij uitzettingen
in het schorsweefsel zich sterker manifesteerden, zoals te zien ter hoogte van de grijze boxen
(DOY 100 en DOY 101) in experiment 5 (figuur 3.5). Ondanks de beperkte afmetingen van
het blootgestelde xyleemoppervlak, werd dus proefondervindelijk vastgesteld dat voldoende zorg
moet besteed worden aan het tegengaan van versnelde dehydratatie. Het diameterverloop tijdens
de eerste periode wordt daarom verder geanalyseerd met behulp van figuur 3.5. Watercontact
doorheen de eerste dag van de uitdroging resulteerde niet in een toename van de xyleemdiameter.
Na dehydratatie doorheen de nacht bewerkstelligde het water echter wel een sterke uitzetting van
het xyleem. Tijdens de tweede dag van de uitdroging (DOY 101) waren een paar uur tussen
twee metingen reeds voldoende om in een zichtbare uitzetting te resulteren (DOY 101.7). De
schorsdiameter zette reeds vanaf het begin van de uitdroging uit telkens wanneer water werd
toevoerd. De schorsschommelingen worden afgebeeld bovenop de variaties in xyleemdiameter
doordat het schorsweefsel bovenop xyleemweefsel ligt.
Vanaf periode 2, dus wanneer de diameter kromp onder 0.1 mm in experiment 3 (figuur 3.4 D)
trad cavitatie op. Naar analogie met Cochard et al. (2001) kan de cavitatiethreshold uitgedrukt
worden als een mate van inkrimping. Uit de resultaten van experimenten 1 tot 7 volgt dat de
druivelaars caviteerden vanaf een inkrimping van 0.1 tot 0.2 mm.
Aan het begin van periode 3 vielen xyleem- en schorsdiameter samen op een waarde van -0.2 mm
en vertoonden vanaf dan een parallel verloop (periode 3 van figuur 3.4 D en figuur 3.5). De
inkrimping verliep aan een trager tempo in vergelijking met de voorgaande periodes. Bovendien
resulteerde het aanleggen van een waterstroming doorheen de tak hier niet meer in een toename
van de diameter (e.g. grijze rechthoek ter hoogte van DOY 80.4 in figuur 3.4 D). Analyse van
de andere experimenten wees uit dat vanaf een inkrimping van 0.15 - 0.2 mm, in geen van beide
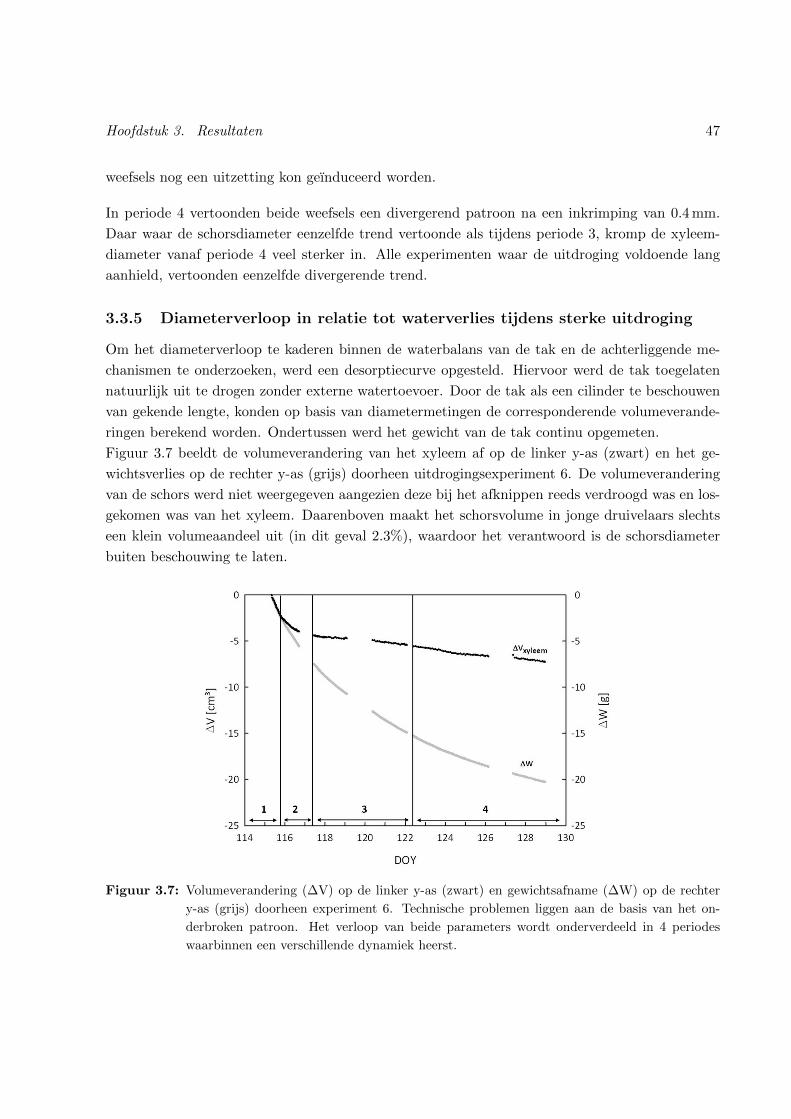
Hoofdstuk 3. Resultaten 47
weefsels nog een uitzetting kon geınduceerd worden.
In periode 4 vertoonden beide weefsels een divergerend patroon na een inkrimping van 0.4 mm.
Daar waar de schorsdiameter eenzelfde trend vertoonde als tijdens periode 3, kromp de xyleem-
diameter vanaf periode 4 veel sterker in. Alle experimenten waar de uitdroging voldoende lang
aanhield, vertoonden eenzelfde divergerende trend.
3.3.5 Diameterverloop in relatie tot waterverlies tijdens sterke uitdroging
Om het diameterverloop te kaderen binnen de waterbalans van de tak en de achterliggende me-
chanismen te onderzoeken, werd een desorptiecurve opgesteld. Hiervoor werd de tak toegelaten
natuurlijk uit te drogen zonder externe watertoevoer. Door de tak als een cilinder te beschouwen
van gekende lengte, konden op basis van diametermetingen de corresponderende volumeverande-
ringen berekend worden. Ondertussen werd het gewicht van de tak continu opgemeten.
Figuur 3.7 beeldt de volumeverandering van het xyleem af op de linker y-as (zwart) en het ge-
wichtsverlies op de rechter y-as (grijs) doorheen uitdrogingsexperiment 6. De volumeverandering
van de schors werd niet weergegeven aangezien deze bij het afknippen reeds verdroogd was en los-
gekomen was van het xyleem. Daarenboven maakt het schorsvolume in jonge druivelaars slechts
een klein volumeaandeel uit (in dit geval 2.3%), waardoor het verantwoord is de schorsdiameter
buiten beschouwing te laten.
Figuur 3.7: Volumeverandering (∆V) op de linker y-as (zwart) en gewichtsafname (∆W) op de rechter
y-as (grijs) doorheen experiment 6. Technische problemen liggen aan de basis van het on-
derbroken patroon. Het verloop van beide parameters wordt onderverdeeld in 4 periodes
waarbinnen een verschillende dynamiek heerst.

48 Hoofdstuk 3. Resultaten
Analoog aan figuur 3.4 D kunnen ook hier dezelfde 4 periodes onderscheiden worden. Gedurende
periode 1 onderging de tak geen tot een heel beperkte hoeveelheid cavitatie (resultaten niet weer-
gegeven). De volumeverandering en de gewichtsverandering waren proportioneel (∆W/∆V=1) en
namen snel af. Vanaf periode 2 was de relatie tussen beide parameters niet meer lineair (∆W/∆V=
1.9) als gevolg van een zwakkere volumeafname. Dit betekent dat de hoeveelheid verdampt water
niet meer uitgedrukt kon worden als een volumeverandering. Ondertussen bereikte de cavitatie
een gemiddelde waarde van 2.5 signalen per minuut (resultaten niet weergegeven). Doorheen pe-
riode 3 onderging het xyleem weinig extra inkrimping. Het waterverlies bleef echter toenemen
waardoor proportioneel veel meer water verloren ging per volumetrische inkrimping (∆W/∆V =
7).
Tenslotte stegen de cavitatiesignalen tot een maximale hoeveelheid van 5 signalen per minuut,
een niveau dat werd aangehouden tot aan het begin van periode 4. In deze laatste periode nam
het xyleemvolume terug sterker toe en bereikte finaal eenzelfde snelheid van afname als het wa-
terverlies (∆W/∆V=1.2). Op dat moment waren ook de cavitatiesignalen afgenomen tot een
gemiddelde waarde van 1 signaal per minuut. In experiment 7 werd een analoog patroon opgeme-
ten (resultaten niet weergegeven).
3.4 Destructieve vulnerability curve
Toenemende zuigspanning in het xyleem veroorzaakt cavitatie. Dergelijke luchtgevulde vaten ver-
oorzaken een obstructie in het xyleemweefsel met een daling van de hydraulische geleidbaarheid
tot gevolg. Deze relatie wordt standaard weergegeven in een VC waarin de afname in hydraulische
geleidbaarheid (toenemende PLC [%]) uitgezet wordt in functie van de variatie in de waterpoten-
tiaal [MPa]. In figuur 3.8 worden de resultaten uit experimenten 1, 2, 3, 5 en 6 weergegeven.
De eerste drie experimenten werden uitgevoerd aan de hand van de XYL’EM methode, terwijl
experimenten 5 en 6 gegenereerd werden aan de hand van de gravitatiekolom. De VC uit expe-
riment 1, 2 en 3 vertonen een analoge trend en kunnen elk gefit worden aan een Chapman curve
(R2 respectievelijk 0.97, 0.96 en 0.99). Het gemiddelde van deze curves wordt voorgesteld door
de zwarte lijn waarbij de grijze zone de standaardafwijking voorstelt. De bijhorende P50-waarde
bedraagt -1.99 MPa. Experimenten 5 en 6 vertonen weinig gelijkenis met de voorgaande experi-
menten zowel wat betreft grootte-orde als wat betreft vorm.
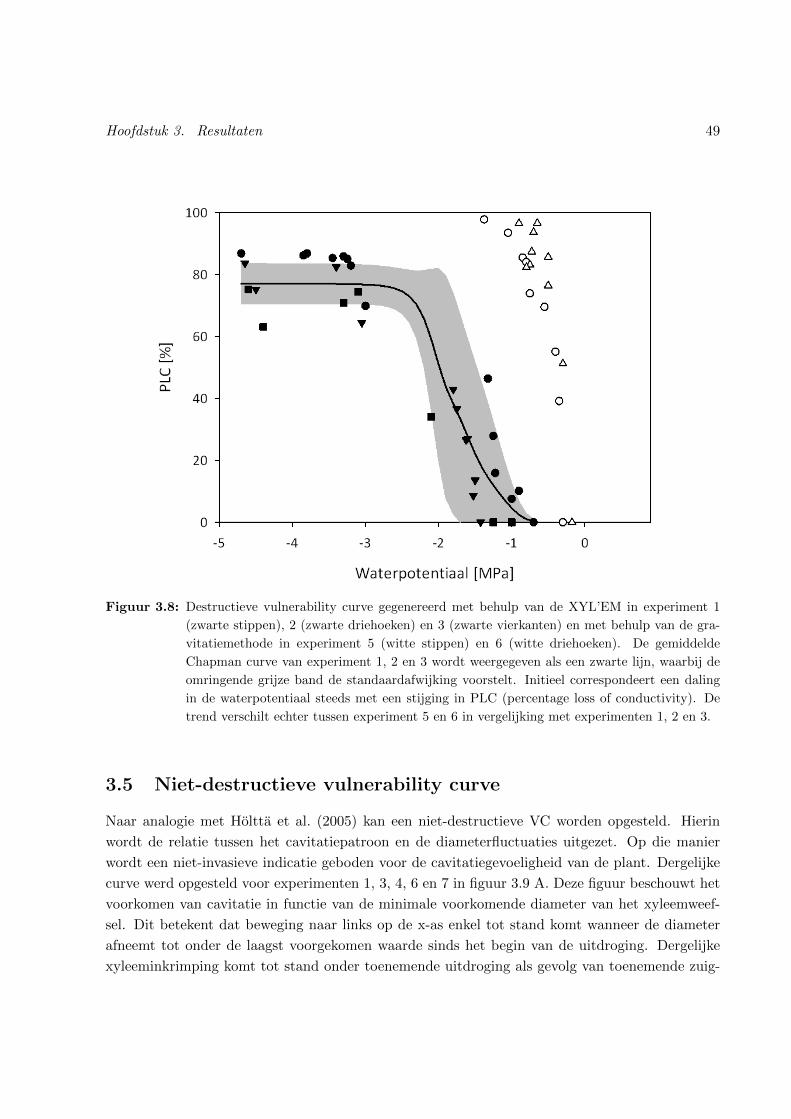
Hoofdstuk 3. Resultaten 49
Figuur 3.8: Destructieve vulnerability curve gegenereerd met behulp van de XYL’EM in experiment 1
(zwarte stippen), 2 (zwarte driehoeken) en 3 (zwarte vierkanten) en met behulp van de gra-
vitatiemethode in experiment 5 (witte stippen) en 6 (witte driehoeken). De gemiddelde
Chapman curve van experiment 1, 2 en 3 wordt weergegeven als een zwarte lijn, waarbij de
omringende grijze band de standaardafwijking voorstelt. Initieel correspondeert een daling
in de waterpotentiaal steeds met een stijging in PLC (percentage loss of conductivity). De
trend verschilt echter tussen experiment 5 en 6 in vergelijking met experimenten 1, 2 en 3.
3.5 Niet-destructieve vulnerability curve
Naar analogie met Holtta et al. (2005) kan een niet-destructieve VC worden opgesteld. Hierin
wordt de relatie tussen het cavitatiepatroon en de diameterfluctuaties uitgezet. Op die manier
wordt een niet-invasieve indicatie geboden voor de cavitatiegevoeligheid van de plant. Dergelijke
curve werd opgesteld voor experimenten 1, 3, 4, 6 en 7 in figuur 3.9 A. Deze figuur beschouwt het
voorkomen van cavitatie in functie van de minimale voorkomende diameter van het xyleemweef-
sel. Dit betekent dat beweging naar links op de x-as enkel tot stand komt wanneer de diameter
afneemt tot onder de laagst voorgekomen waarde sinds het begin van de uitdroging. Dergelijke
xyleeminkrimping komt tot stand onder toenemende uitdroging als gevolg van toenemende zuig-

50 Hoofdstuk 3. Resultaten
spanning (Irvine & Grace, 1997). Op die manier kan het verloop van de minimaal voorkomende
diameter geınterpreteerd worden als de zuigspanning, die toeneemt naarmate de xyleemdiameter
verder afneemt. In tegenstelling tot de andere experimenten stellen de waarden uit experiment 7
de inkrimping van de volledige tak voor (xyleemdiameter + schorsweefsel). Dit is het gevolg van
praktische problemen met de sensor die de xyleemdiametervariaties opmat. Zoals boven gesteld,
maakt het schorsweefsel bij druivelaars echter slechts een klein volumetrisch aandeel uit van de
tak, waardoor aangenomen wordt dat de curve een analoog verloop vertoont als de overeenkom-
stige xyleem-gebaseerde curve. Dit werd bevestigd door de andere experimenten (resultaten niet
weergegeven) waardoor de curve van experiment 7 analoog als de andere curves zal behandeld
worden.
De VC van de vijf experimenten vertoonden een sigmoıdaal verloop met een R2 tussen 0.991 en
0.999. Analoog aan P50 -waarde van de destructieve VC kan hier een cUAE50-waarde bepaald
worden. Dit is de xyleeminkrimping die correspondeert met 50 % van de cavitatiesignalen. Deze
cUAE50-waarden bedroegen respectievelijk -0.27, -0.34, -0.30, -0.55 en -1.06 mm voor de expe-
rimenten 1, 3, 4, 6 en 7. Experiment 6 vertoonde een discontinu patroon met een aantal open
ruimtes (e.g. ter hoogte van een inkrimping van 0.4 mm). Deze werden veroorzaakt door techni-
sche problemen met de computer. Er wordt bijgevolg gesuggereerd dat de werkelijke curve een
iets steiler patroon vertoonde met een kleinere cUAE50-waarde.
In figuur 3.9 B worden dezelfde VC weergegeven met additioneel de curve uit experiment 2 (ter
hoogte van -0.1 mm). In deze figuur wordt de y-as in absoluut aantal UAE-signalen uitgedrukt.
Zowel de diameterinkrimping als het aantal UAE-signalen van experiment 2 waren veel kleiner
dan deze van de andere experimenten, waaruit blijkt dat deze curve slechts de beginfase van een
volledige VC afbeeldt. Ook tussen de andere experimenten heerst een grote variabiliteit. De hoe-
veelheid cavitatiesignalen nam af van december (experiment 1) tot maart (experiment 3 en 4) en
steeg spectaculair in mei (experimenten 6 en 7): daar waar eind mei ruim 54 000 cavitatiesignalen
werden opgemeten, werd een totale hoeveelheid van 5 821 signalen geregistreerd in maart: een
aandeel van slechts 11%.

Hoofdstuk 3. Resultaten 51
Figuur 3.9: Niet-destructieve vulnerability curve waarbij het aantal cavitatiesignalen (A) relatief en (B)
absoluut wordt uitgedrukt ten opzichte van de minimaal behaalde xyleemdiameter. Het cu-
mulatieve aantal gemeten UAE (’ultrasonic acoustic emissions’) is een maat voor het verlies
aan geleidbaarheid terwijl de minimaal bereikte xyleemdiameter de maximaal bereikte zuig-
spanning voorstelt. Kleurcode: experiment 1 (zwart), experiment 2 (groen), experiment 3
(donkergrijs), experiment 4 (lichtgrijs), experiment 6 (donkerblauw) en experiment 7 (licht-
blauw). De zwarte curves stellen telkens de overeenkomstige sigmoıdale curves voor.
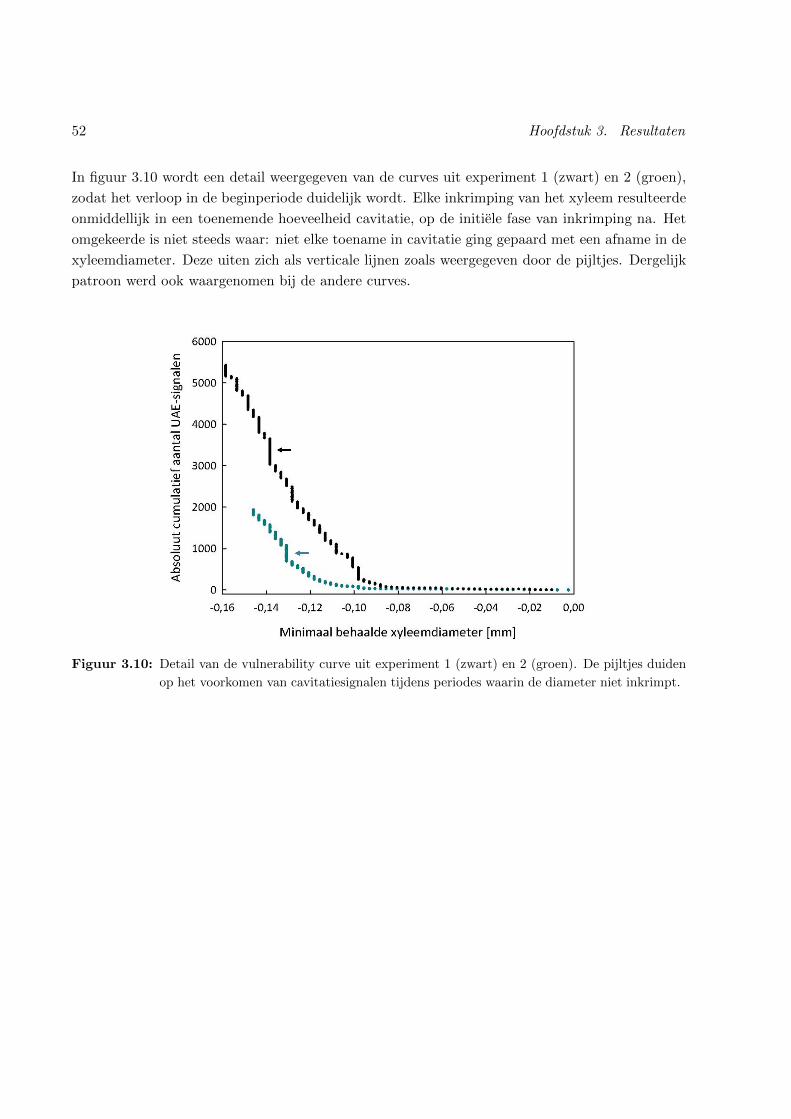
52 Hoofdstuk 3. Resultaten
In figuur 3.10 wordt een detail weergegeven van de curves uit experiment 1 (zwart) en 2 (groen),
zodat het verloop in de beginperiode duidelijk wordt. Elke inkrimping van het xyleem resulteerde
onmiddellijk in een toenemende hoeveelheid cavitatie, op de initiele fase van inkrimping na. Het
omgekeerde is niet steeds waar: niet elke toename in cavitatie ging gepaard met een afname in de
xyleemdiameter. Deze uiten zich als verticale lijnen zoals weergegeven door de pijltjes. Dergelijk
patroon werd ook waargenomen bij de andere curves.
Figuur 3.10: Detail van de vulnerability curve uit experiment 1 (zwart) en 2 (groen). De pijltjes duiden
op het voorkomen van cavitatiesignalen tijdens periodes waarin de diameter niet inkrimpt.

DISCUSSIE


Hoofdstuk 4
Discussie
4.1 Cavitatiedynamiek tijdens periodes van zwakke en sterke uit-
droging
De uitgesproken verschillen in cavitatiepatroon tussen experimenten 2 en 3 (figuur 3.4) kunnen in
belangrijke mate toegeschreven worden aan het microklimaat. Een hoger VD tijdens experiment
3 betekende immers een grotere drijvende kracht voor de transpiratie en dus een snellere en meer
intense uitdroging van de tak (Leonardi et al., 2000). Dit zorgde voor een snellere afname van de
waterpotentiaal waardoor de cavitatiethreshold, i.e. het maximale drukverschil dat de pitporien
konden weerstaan, sneller overschreden werd en cavitatie in experiment 3 sneller aanving.
Na 1 dag uitdroging in experiment 3 kon de periodiek aangelegde waterstroom tijdens hydraulische
geleidbaarheidsmetingen geen diametertoename meer bewerkstelligen. Een mogelijke verklaring
voor dit fenomeen is de aanzienlijke hoeveelheid cavitatie die reeds had plaatsgevonden op dat
moment. Hierdoor ontstond een afname in de hydraulische geleidbaarheid (Tyree & Sperry, 1989;
Sevanto et al., 2005), waardoor de watertoevoer doorheen de tak geblokkeerd werd en het water de
vaten niet voldoende diep kon penetreren. Op die manier kan de waterinput onvoldoende geweest
zijn om het waterverlies door uitdroging te compenseren, waardoor geen tijdelijke relaxatie van de
zuigspanning kon optreden. Anderzijds kunnen dergelijke verstoppingen ook artificieel gegenereerd
zijn door particulair materiaal, luchtbellen of spontane gelvorming in de tak (Espino & Schenk,
2011; van Doorn et al., 2011; Jacobsen & Pratt, 2012). Onafhankelijk van de oorzaak kan dit
fenomeen ook de verschillende cavitatiepatronen verklaren. Omdat de zuigspanning een continu
toenemende trend vertoonde, werd de cavitatiethreshold van steeds meer vaten overschreden en
kon cavitatie onverstoord doorgaan. Tijdens experiment 2 was de cavitatie minder ver gevorderd
waardoor periodieke wateraanleg wel uitgesproken diameterstijgingen kon blijven bewerkstelligen,
die bovendien correspondeerden met een pauze in de cavitatiesignalen: uitzettende diameters be-
tekenden immers geen extra daling in de waterpotentiaal waardoor weinig extra cavitatie optrad
55

56 Hoofdstuk 4. Discussie
(figuur 3.6). Enkel wanneer de diameter daalde onder het voorheen minimaal bereikte niveau,
werden nieuwe cavitatiethresholds overschreden wat resulteerde in bijkomende cavitatie (Holtta
et al., 2005). Op die manier kan het verschil in cavitatiepatroon tussen beide experimenten ver-
klaard worden.
Het lage VD in combinatie met de korte dehydratatieperiode in experiment 2 kan aan de basis
liggen van de lage hoeveelheid UAE-signalen: de tak onderging geen volledige uitdroging waardoor
slechts een beperkt aandeel van de vaten gecaviteerd waren. In experiment 3 onderging de tak een
meer intense uitdroging, zodat deze zich na 5 dagen reeds in een verregaand dehydratatiestadium
bevond en het grootste aandeel van de vaten gecaviteerd waren. Dit uit zich bovendien in de
beperkte hoeveelheid UAE-signalen die nadien voorkwamen. Analyse van de signaalintensiteit uit
experiment 3 bevestigt een analoge trend als de hoeveelheid UAE-signalen in figuur 3.4 C (resul-
taten niet weergegeven): een initiele snelle toename met nadien een afname in de signaalsterkte.
Tyree & Sperry (1989) stellen dat toenemende zuigspanningen resulteren in een steeds grotere
energievrijgave, wat de toenemende signaalsterkte kan verklaren. De oorzaak van de beduidende
daling in aantal en sterkte van de cavitatiesignalen is nog grotendeels ongekend, gezien de be-
perkte literatuur omtrent cavitatie in angiospermen tijdens verregaande uitdroging. Ondanks
nog geen eenduidige verklaring voor de laag-energetische signalen is gevonden, wordt algemeen
aangenomen dat deze emissies niet voortkomen uit cavitatiewaarnemingen van watergeleidende
xyleemvaten (Wolkerstorfer et al., 2012). Desalniettemin werd uitgebreid onderzoek gedaan naar
coniferen, die tijdens verregaande dehydratatiestadia grote hoeveelheden laag-energetische signa-
len voortbrachten (e.g. Mayr & Rosner, 2011; Wolkerstorfer et al., 2012). Daar worden dergelijke
laag-energetische signalen deels toegeschreven aan cavitatie van kleine erg resistente tracheıden,
die ondanks de hoge zuigspanning slechts een beperkte energie uitstoten door hun kleine volume
(Mayr & Rosner, 2011). Daarnaast kunnen andere houtelementen zoals niet-geleidende vezels en
levende parenchymcellen verantwoordelijk zijn voor de laag-energetische UAE-signalen, zo stel-
len Sandford & Grace (1985). Tenslotte kunnen ook andere plantaardige processen aan de basis
liggen zoals microscheuren in celwanden als gevolg van uitdroging (Kowalski & Smockzkiewicz,
2004; Rosner, 2004; Wolkerstorfer et al., 2012).
Het cavitatiepatroon uit experiment 3 komt echter niet volledig overeen met de dynamiek waarge-
nomen in coniferen: terwijl zowel de intensiteit als het aantal signalen afnam bij druivelaars steeg
de hoeveelheid signalen van lage intensiteit in coniferen (Wolkerstorfer et al., 2012). Dit verschil
kan toegeschreven worden aan de verschillende anatomische structuur waardoor een andere cavi-
tatiedynamiek gegenereerd kan worden. Coniferen bezitten immers enkel tracheıden die kleiner
zijn in afmeting dan de lange houtvaten in druivelaars (Tyree & Zimmermann, 2001). Zo hebben
Holtta et al. (2005) in totaal ruim 50 000 UAE-signalen opgemeten over een dehydratatieperiode
van 3 dagen in Pinus sylvestris, terwijl in druivelaars slechts 20 000 emissies werden opgemeten
over een periode van 7 dagen.

Hoofdstuk 4. Discussie 57
Een aantal auteurs oppert dat het grote aantal UAE-signalen die massaal voorkomen, een indicatie
bieden voor het verlies aan hydraulische geleidbaarheid en dus voor het totaal aantal watergelei-
dende vaten in de tak (Tyree et al., 1984; Cochard, 1992; Holtta et al., 2005). Er heerst echter
onenigheid omtrent deze hypothese. Zo opperen Sandford & Grace (1985) dat het opgemeten
aantal UAE-signalen slechts een aandeel van 16% van de vaten representeert. Ze schrijven dit
toe aan het beperkte bereik van de cavitatiesensor en het tegelijk voorkomen van meerdere UAE-
signalen die als 1 signaal worden opgemeten. Onderzoek van Rosner et al. (2006) wijst dan weer
uit dat diverse andere plantaardige processen, dus niet afkomstig van geleidende xyleemvaten, aan
de basis kunnen liggen van het grotere aantal emissies. Daarnaast wordt de interpretatie van het
aantal UAE-signalen in termen van aantal vaten bemoeilijkt bij het ’in situ’ gebruik van de sensor
doordat de vaten kunnen hervullen en dus meermaals UAE-signalen kunnen produceren (Jackson
& Grace, 1996). Hoewel de precieze relatie nog onzeker is, wordt algemeen aangenomen dat het
totaal aantal UAE-signalen sterk gecorreleerd is met het aantal xyleemvaten (Rosner et al., 2006;
Mayr & Rosner, 2011; Wolkerstorfer et al., 2012).
4.2 Diameterverloop doorheen verregaande dehydratatie
Toenemende uitdroging gaat gepaard met een diameterinkrimping waartoe verschillende mechanis-
men bijdragen. Aan de hand van figuur 3.4 wordt onderscheid gemaakt tussen deze verschillende
mechanismen doorheen de dehydratatieperiode. Figuur 3.7 biedt additionele informatie bij het dif-
ferentieren tussen de verschillende mechanismen aan de hand van de relatie tussen volumeafname
en waterverlies. Op die manier kan cavitatie ook fysisch gelinkt worden aan waterverlies.
4.2.1 Verklaring diameterverloop gedurende periode 1
Bij een inkrimping kleiner dan 0.1 mm overschreed de waterpotentiaal de cavitatiethreshold nog
niet, waardoor dergelijke afname niet resulteerde in cavitatie (figuur 3.4C en D). Bovendien waren
xyleemdiametervariaties hier proportioneel gerelateerd aan het waterverlies (∆W/∆V=1, figuur
3.7). De diameterinkrimping kan bijgevolg toegeschreven worden aan het verlies van capillair wa-
ter en waterverlies uit levende cellen (elastische cellen, floeem, waterreserves tussen xyleemvaten
en in de schors, levende xyleemparenchymcellen,...) (Tyree et al., 1984; Zweifel & Item, 2000;
Almeras, 2008). Deze laatste zijn zeer flexibel waardoor de inkrimping zich aan snel tempo kan
voltrekken.
Zoals inkrimpende diameters gerelateerd zijn aan de waterrelaties van de plant, zijn ook uitzet-
tende weefsels afhankelijk van de waterbalans (figuur 3.4 B en 3.5). In de beginfase veroorzaakte
periodiek contact met een externe waterbron een onmiddellijke toename in de totale diameter.
Deze kan te wijten zijn aan het hervullen van de waterreserves (e.g. Irvine & Grace, 1997; Co-
chard et al., 2001; Intrigliolo & Castel, 2007; Almeras, 2008).

58 Hoofdstuk 4. Discussie
Los van het bovenliggend weefsel onderging ook de xyleemdiameter een uitzetting bij watercontact,
die toegeschreven kan worden aan de tijdelijke relaxatie van de negatieve zuigspanning (e.g. Irvine
& Grace, 1997; Offenthaler et al., 2001). In tegenstelling tot de totale diameter genereerde water-
contact hier pas een uitzetting na een initiele periode van uitdroging, wanneer de waterpotentiaal
reeds voldoende gedaald was. De mate waarin watertoevoer de xyleemdiameter beınvloedt, hangt
dus af van de dehydratatietoestand van de tak en bewijst de rigiditeit van het xyleemweefsel.
4.2.2 Verklaring diameterverloop gedurende periode 2 en 3
Het diameterverloop doorheen periode 2 en 3 kan toegeschreven worden aan de combinatie van
twee factoren. Enerzijds wordt verondersteld dat de dimensies van de levende cellen verder bleven
afnemen. Daarnaast werd de cavitatiethreshold overschreden aan het begin van periode 2 zodat
vanaf dat moment ook de inelastische xyleemvaten ineenkrompen. Toenemende zuigspanning
zorgt immers voor het samentrekken van de vaten als gevolg van de adhesiekrachten tussen het
vrije water en de vatwand (Irvine & Grace, 1997; Peramaki et al., 2001). Ten gevolge van hun
rigiditeit kunnen deze slechts een beperkte inkrimping bewerkstelligen (Schultz & Matthews, 1988;
Peramaki et al., 2001; Steppe et al., 2006), waardoor proportioneel meer water verloren ging per
volumeverandering. Dit vertaalde zich in een toenemende ∆W/∆V ratio (figuur 3.7) (Zweifel &
Item, 2000).
De sterkere diameterdaling gedurende periode 2 kan te wijten zijn aan een grotere bijdrage van
de levende cellen, terwijl cavitatie de bovenhand kreeg in periode 3. In die vaten die nog vrij
water bezaten, ontwikkelden zich immers steeds sterkere zuigspanningen, waardoor steeds meer
vaten gecaviteerd werden, terwijl de levende cellen uitgeput geraakten en hun aandeel afnam.
Deze hypothese wordt bevestigd door de toenemende ∆W/∆V ratio enerzijds en het toenemende
aantal cavitatiesignalen anderzijds.
De verschillende inkrimpingssnelheid tussen periode 2 en 3 kan ook verklaard worden aan de hand
van de trade-off tussen gevoeligheid en hydraulische geleidbaarheid. Hierin wordt gesteld dat cavi-
tatieresistentie binnen 1 soort toeneemt naarmate de vatwand steviger is en de diameter afneemt
(uitgedrukt als een hoge vatwand/vatlumen-ratio) (e.g. Rosner et al., 2010). Aangezien de gevoe-
ligste vaten het eerst caviteren, zou periode 2 theoretisch gepaard gaan met enerzijds een sterker
waterverlies en anderzijds een grotere diameterinkrimping. Brede vaten houden immers meer wa-
ter op en worden gekenmerkt door een lagere vatwand/vatlumen-ratio, waardoor ze minder goed
bestand zijn tegen inkrimping onder invloed van zuigspanning. Zo kan de snellere diameterdaling
doorheen periode 2 in figuur 3.4 D het gevolg zijn van de cavitatie van een aantal gevoelige brede
vaten, terwijl de tragere inkrimping in periode 3 kan voortkomen uit meerdere kleine caviterende
vaten. Deze hypothese wordt bevestigd door het grotere aantal cavitatiesignalen dat optrad vanaf
de grens tussen periode 2 en 3.

Hoofdstuk 4. Discussie 59
Wanneer de diameters sterker zijn afgenomen dan 0.15 - 0.2 mm, reageerden ze niet meer op de
aangelegde waterstroming in tegenstelling tot de respons tijdens periode 1 (sectie 4.2.1). Zoals
uiteengezet in sectie 4.1 kan dit ter hoogte van het xyleemweefsel te wijten zijn aan de afname
van de hydraulische geleidbaarheid als gevolg van verregaande cavitatie of artificieel geınduceerde
verstopping, waardoor het water niet meer diep in de tak kon doordringen. De verklaring voor
de continue diameterdaling ter hoogte van het schorsweefsel kan gevonden worden in de afname
van activiteit van de levende cellen. Door verregaande dehydratatie zijn de waterreserves immers
uitgeput. Hierdoor kan de turgordruk niet meer hersteld worden (Holtta et al., 2006), waardoor
het schorsweefsel afsterft en bijgevolg niet meer kan reageren op watertoevoer. Dit impliceert dat
de levende cellen geen onafhankelijke diameterveranderingen meer kunnen ondergaan, zodat de
totale diameter vanaf dan enkel variaties in het onderliggende xyleemweefsel reflecteert en beide
diameters een parallel verloop vertonen. Huidige kennis omtrent het hervullen van vaten tijdens
verregaande dehydratatie is schaars, gezien onderzoek zich meestal toespitst op het hervullen van
de vaten over een korte tijdspanne of ’in situ’ (e.g. Sperry et al., 1987). Desalniettemin wordt
hiermee het belang benadrukt van een intact en onverstoord geleidingsweefsel om watertransport
te kunnen blijven onderhouden.
4.2.3 Verklaring diameterverloop gedurende periode 4
Wanneer een inkrimping van 0.4 mm werd bereikt, onderging het xyleemweefsel plots een sterkere
diameterafname, die divergeerde van de totale diameter. Het minder expressieve verloop van de
schorsdiameter kan voortkomen uit het loslaten van schors- en xyleemweefsel waardoor een in-
krimping van het xyleemweefsel niet noodzakelijk resulteert in een even grote daling in de totale
diameter. Bovendien vertegenwoordigt het schorsweefsel een zeer beperkt volume aandeel in de
tak (2-5% in deze experimenten) waardoor het xyleemweefsel procentueel gezien veel sterker kan
inkrimpen (Almeras, 2008).
De sterkere mate van xyleemdaling op zijn beurt kan toegeschreven worden aan twee factoren.
Vooreerst kan de grote hoeveelheid cavitatie die ervoor optrad, een belangrijke rol spelen (figuur
3.4 C). Vanaf het moment dat de diameter sterker inkromp bij het begin van periode 4, vielen
de opgemeten cavitatiesignalen immers terug van een hoog niveau van ongeveer 8 signalen per
minuut naar een niveau van gemiddeld 2 signalen per minuut. Hieruit kan dus worden afgeleid
dat de meerderheid van de cavitatiewaarnemingen reeds had plaatsgevonden en een aanzienlijk
aandeel van de vaten reeds gecaviteerd was. Het vasculair weefsel, dat initieel een stevig geheel
vormde, opgebouwd uit functionele vaten onder zuigspanning, was op dat moment voor een be-
langrijk aandeel met lucht gevuld. De individuele vaten werden minder in vorm gehouden door
een geheel van omringende functionele vaten zodat het xyleemweefsel een sterkere inkrimping kon
ondergaan. Daarnaast wordt verondersteld dat ook het verlies aan gebonden water een rol kan
spelen in deze finale fase. Wanneer een xyleemvat caviteert, wordt het vrij water in de xyleem-

60 Hoofdstuk 4. Discussie
lumina immers vervangen door lucht, zodat enkel gebonden water overblijft ter hoogte van de
vatwand. Dit wordt het verzadigingspunt van het hout genoemd (’fiber saturation point’). Ver-
damping van dit gebonden water zorgt voor een contractie van de microfibrillen in de vatwand
(Rosner et al., 2010) en kan op die manier bijdragen tot een xyleeminkrimping evenredig met het
waterverlies, zoals waargenomen in figuur 3.7 (∆W/∆V=1.2). Rosner et al. (2010) stelt bovendien
dat dergelijke contracties gepaard gaan met de uitstoot van laag energetische UAE-signalen, wat
in overeenstemming is met het patroon in signaalintensiteit (resultaten niet weergegeven).
4.3 Destructieve vulnerability curve
Het opmeten van de hydraulische geleidbaarheid (Kh) is niet eenvoudig. In diverse andere onder-
zoeken werden meermaals problemen gerapporteerd, zoals onstabiele of continu en snel afnemende
Kh-waarden tijdens het flushen (Sperry & Tyree, 1988; Espino & Schenk, 2011; van Doorn et al.,
2011; Jacobsen & Pratt, 2012). Hoewel in deze thesis niet gewerkt werd met flushen onder hoge-
druk, leden de XYL’EM metingen onder analoge problemen. Bijkomende experimenten (resultaten
niet weergegeven), waarbij de tak niet toegelaten werd om verregaand uit te drogen, resulteer-
den daarenboven ook in een continue en verregaande afname van de geleidbaarheid. Door deze
problemen werd de initieel gehanteerde XYL’EM methode in vraag gesteld en werd besloten de
volgende experimenten uit te voeren aan de hand van de gravitatiekolom. Het resultaat wordt
weergegeven in figuur 3.8, waarbij experimenten 1, 2 en 3 opgemeten werden met de XYL’EM,
terwijl de verticale gravitatiekolom gebruikt werd in experimenten 5 en 6.
4.3.1 Verklaring van het verschil in destructieve VC
De VC gegenereerd door de twee methoden vertoonden weinig overeenkomst. Verschillende feno-
menen kunnen hiervoor aan de basis liggen. Er wordt gesuggereerd dat niet zozeer de verschillende
gehanteerde techniek, maar eerder de verschillende samenstelling van het water aan de basis ligt
van deze afwijkende resultaten. In de gravitatiemethode werd immers zuiver, niet-ontgast ge-
destilleerd water gebruikt als medium in tegenstelling tot het ontgast, gefilterd Volvic water met
toegevoegde kationen in de XYL’EM methode. Zo opperen Espino & Schenk (2011) dat lucht-
verzadigd water kunstmatige belvorming kan induceren, waardoor vaten geblokkeerd worden en
de hydraulische geleidbaarheid sneller afneemt als deze die veroorzaakt wordt door natuurlijke
dehydratatie. Daarnaast kan particulair materiaal in onvoldoende gefilterd water de pitporien
verstoppen, waardoor de doorstroming van water verhinderd wordt en te hoge PLC-waarden op-
gemeten worden (Espino & Schenk, 2011). Van Doorn et al. (2011) stellen bovendien dat kationen
nodig zijn om aan de hand van elektrostatische interacties met het pitmembraan, een daling in de
waterstroming te kunnen tegengaan. Bijgevolg wordt geconcludeerd dat bovenstaande problemen
meer uitgesproken tot uiting kwamen bij de gravitaire methode en op die manier resulteerden in
een snellere afname in hydraulische geleidbaarheid. Desondanks worden analoge verschijnselen

Hoofdstuk 4. Discussie 61
ook in de XYL’EM experimenten (1, 2 en 3) niet uitgesloten. Het geleidingsweefsel in planten
heeft een complexe structuur. Dit maakt hen kwetsbaar voor cavitatie of verstoppingen zodat de
aanwezigheid van het kleinste partikeltje of luchtbelletje nauwkeurig verhinderd moet worden. Bij
afknippen en artificieel doorstromen met water wordt een natuurlijke situatie gecreeerd en kan dit
mechanisme aangetast worden, zoals blijkt uit deze resultaten.
Hoe langer de taksegmenten (50 - 90 cm), hoe groter bovenstaande effecten. In lange takken dient
namelijk meer water doorheen de tak te stromen waardoor er meer kans is op een artificiele
daling van de hydraulische geleidbaarheid (modificatie van de pitporie structuur, verstoppen van
de vaten,...) (Ayup et al., 2012). Het gebruik van kortere segmenten was echter niet mogelijk
aangezien druivelaars lange vaten bezitten. Bijgevolg dient omzichtig omgesprongen te worden
met de voorbereiding van het water bij metingen van de hydraulische geleidbaarheid van soort
met lange vaten.
Een verklaring voor de afname in geleidbaarheid bij continue hydratatie van takken, kan gevon-
den worden in recent onderzoek van Jacobsen & Pratt (2012). Zij ondervonden een daling in
de geflushte Kh-waarde. Deze waarde zou echter constant moeten blijven, onafhankelijk van het
dehydratatiestadium, gezien de aanwezige embolismen verwijderd worden aan de hand van hoge
druk en de tak op die manier terug in verzadigde toestand wordt gebracht. De auteurs schrijven
deze daling toe aan plantaardige gelvorming die een verstopping veroorzaakt in de xyleemvaten
(Jacobsen & Pratt, 2012). Sun et al. (2008) stellen dat deze gelvorming in druivelaars als natuur-
lijke reactie kan optreden bij zowel verwondingen als bij embolisaties. Hieruit kan dus niet bepaald
worden of gelvorming in afgeknipte takken artificieel geınduceerd werd door het afknippen van de
takken, dan wel natuurlijk geınduceerd werd door cavitatie. De belangrijkste consequentie van
deze gelhypothese voor de constructie van destructieve VC is dat het opmeten van het verloop van
de hydraulische geleidbaarheid niet relatief ten opzichte van de geflushte waarde mag gebeuren.
Jacobsen & Pratt (2012) stellen immers dat de gevormde gels niet verwijderd kunnen worden door
een hoge druk, waardoor de maximale Kh onderschat wordt en de PLC systematisch overschat
wordt (formule 1.3). De Kh-waarden dienen dus in absolute termen als een ’afname in geleid-
baarheid’ geınterpreteerd te worden. Op die manier verkregen Jacobsen & Pratt (2012) VC die
wijzen op een veel grotere gevoeligheid in vergelijking met de bevindingen van Choat et al. (2010).
In deze thesis werd daarom gewerkt met de absolute waarden voor daling in geleidbaarheid. De
initiele Kh-waarde werd hier immers als de maximale geleidbaarheid beschouwd, aangezien de tak
voor zonsopgang werd afgesneden van een goed bewaterde plant en dus aangenomen werd dat de
tak volledig verzadigd was bij de initiele meting. Alhoewel het voorkomen van gelvorming plau-
sibel is en als verklaring kan dienen voor de afname in geleidbaarheid bij continue watertoevoer,
kan deze niet de oorzaak zijn van de beperkte overeenkomst tussen de VC.
Een laatste mogelijke verklaring voor die afwijking kan geboden worden door de stamwaterpoten-
tiaal. De wijze waarop deze werd opgemeten, verschilt immers tussen beide proefopzetten. De

62 Hoofdstuk 4. Discussie
XYL’EM experimenten grepen plaats tijdens de herfst en winter, wanneer geen blaadjes voor-
handen waren. Daarom werd de waterpotentiaal tijdens experimenten 1, 2 en 3 gemeten op een
kort afgesneden uiteinde van de tak (sectie 2.3). In experimenten 5 en 6 werden blaadjes ge-
bruikt, gewikkeld in aluminiumfolie. De data bevestigen dat de waterpotentiaal sneller en dieper
daalde in het taksegmentje in vergelijking met de bladeren van een langere tak (resultaten niet
weergegeven). Ten gevolge van zijn beperkte afmetingen kon dit immers sneller uitdrogen. Daar-
enboven werd het niet sequentieel aangelegd aan een lagedruk-waterstroming, corresponderend
met de hydraulische geleidbaarheidsmetingen in de grote tak. Het beschouwde verschil tussen de
VC kan dus verklaard worden door een samenspel van de watersamenstelling en de methode van
waterpotentiaalmetingen.
4.3.2 Kwantitatieve interpretatie van de destructieve VC
De VC opgemeten met de XYL’EM vlakten af ter hoogte van PLC-waarden van 80% (figuur 3.8).
Theoretisch wordt echter verwacht dat het eindpunt van dergelijke curve zich ter hoogte van 100%
zou bevinden. Dit kan voortkomen uit het feit dat de drukbommetingen op een takuiteinde werden
uitgevoerd. Onder -5 MPa was dit takuiteinde reeds in zo’n uitgedroogde toestand, dat de meting
niet meer accuraat was en de correcte waterpotentiaal bijgevolg niet meer te bepalen was. Hogere
PLC-waarden werden wel opgemeten (tot 92% zoals gemeten in experiment 2), maar correspon-
deerden met waterpotentialen die niet meer konden opgemeten worden op het taksegment. In
experimenten 5 en 6 daalden de waterpotentialen aan minder snel tempo waardoor de 100% hier
wel bereikt werd. Daarenboven werd getwijfeld aan de accuraatheid van de hoogste PLC-waarden,
gezien deze dicht rond de grens van nauwkeurigheid van het XYL’EM-toestel schommelden.
De P50-waarde berekend op basis van de XYL’EM experimenten bedroeg -1.99 MPa, waaruit volgt
dat reeds 50% cavitatie optreedt bij waterpotentialen van -2 MPa. Gerapporteerde P50-waarden
lopen sterk uiteen. Zo rapporteerden Choat et al. (2010) een P50-waarde van -2.97 MPa en toonden
aan dat PLC-waarden in Vitis vinifera L. cv. Chardonnay niet stegen boven 35% bij waterpoten-
tialen van -2 MPa. De data uit experimenten 1, 2 en 3 suggereren dus een grotere gevoeligheid
aan droogtestress in vergelijking met Choat’s bevindingen. Alsina et al. (2007) rapporteren een
P50-waarde van -2.27 MPa, die dichter aanleunt bij de resultaten uit deze thesis. Resultaten van
Jacobsen & Pratt (2012) op Vitis vinifera L. cv. Glenora tonen dan weer een analoge trend als deze
bekomen met de gravitatiekolom (experimenten 5 en 6), waarin de hydraulische geleidbaarheid
zeer snel afneemt en de P50-waarde -0.51 MPa bedraagt. Ook Wheeler et al. (2005) rapporteren
een erg lage P50-waarde van -0.76 MPa.
Uit bovenstaande analyse kan geconcludeerd worden dat zowel de PLC-waarden als de waterpo-
tentialen sterk afhankelijk zijn van de wijze waarop de proef wordt uitgevoerd en bijgevolg moeilijk
te verzamelen zijn op een consistente manier. Daarenboven circuleren veel uiteenlopende hydrau-

Hoofdstuk 4. Discussie 63
lische VC in de literatuur, waardoor hevig gedebatteerd wordt over de betrouwbaarheid van de
hydraulische meetmethoden. Het opstellen van een niet-destructieve VC die minder afhankelijk is
van de wijze van dataverzameling, zou dan ook een belangrijke meerwaarde kunnen bieden in het
onderzoek naar de waterrelaties in druivelaars.
4.4 Niet-destructieve vulnerability curve
Een niet-destructieve VC biedt de mogelijkheid om continu en niet-destructief de waterstatus van
de plant na te gaan. Holtta et al. (2005) stelt een curve voor waarin het cavitatiepatroon wordt uit-
gezet ten opzichte van de xyleemdiameter. Gecaviteerde vaten betekenen immers een obstructie in
het vasculair weefsel en zorgen zo voor het verlies aan hydraulische geleidbaarheid in de plant. Op
basis van dit verband wordt gesuggereerd dat bij toenemende uitdroging UAE-signalen, uitgesto-
ten door caviterende vaten, eenzelfde trend vertonen als het verlies in hydraulische geleidbaarheid.
Dit werd experimenteel bevestigd door diverse auteurs (e.g. Cochard, 1992; Hacke & Sauter, 1996;
Barnard et al., 2011). Op basis van deze bevindingen wordt aangenomen dat cavitatiesignalen
als indicator kunnen dienen voor de gevoeligheid aan droogte in druivelaars (e.g. Rosner, 2004;
Holtta et al., 2005).
In overeenstemming met bevindingen van diverse auteurs kan de xyleemdiameter in een niet-
destructieve VC geınterpreteerd worden als de maximaal bereikte zuigspanning in het vasculair
weefsel (Irvine & Grace, 1997; Zweifel & Item, 2000; Cochard et al., 2001; Peramaki et al., 2001).
De zuigspanning in het xyleem vergroot immers door toenemende uitdroging, waardoor de tak
inkrimpt (Irvine & Grace, 1997; Peramaki et al., 2001; Sevanto et al., 2005). Ook de totale
takinkrimping (van zowel de schors als het onderliggende xyleem) kan gerelateerd worden aan
de waterpotentiaal, maar deze relatie is minder rechtlijnig te interpreteren. In de schors treedt
namelijk een waaier aan andere processen op die een effect kunnen uitoefenen op de schorsdiameter
(e.g. legen van de waterreserves, levende cellen, radiale groei, osmotische aanpassing)(Offenthaler
et al., 2001). Variaties in de xyleemdiameter staan dus in directer verband met de waterbalans
van de plant en met de cavitatiesignalen en kunnen als niet-invasieve kwantificatie dienen voor de
waterpotentiaal (Holtta et al., 2005). Bij druivelaars is deze schorsdiameter echter zo beperkt dat
ook de totale diameter gebruikt kan worden (experiment 7) .
4.4.1 Opstellen van een niet-destructieve VC
De niet-destructieve VC in figuur 3.9 A toonden een sigmoıdaal profiel waarin cavitatie steeg
met toenemende zuigspanning. Dit is in overeenstemming met de deterministische ’air-seeding’
hypothese (Holtta et al., 2005). Volgens deze hypothese ontstaat cavitatie in een vat na het
overschrijden van een welbepaalde cavitatiethreshold. Ten gevolge van de distributie in pitpo-
riegrootte binnen de diverse xyleemvaten, heerst er een range aan deze thresholds binnen het

64 Hoofdstuk 4. Discussie
vasculair weefsel. Naarmate de zuigspanning toeneemt, wordt de threshold stelselmatig in meer
vaten overschreden, met een toenemend aantal caviterende vaten tot gevolg. Uit figuur 3.9 A blijkt
dat de initiele diameterdaling niet gepaard ging met caviterende vaten. Gedurende deze periode
overschreed de waterpotentiaal die overeenkwam met dergelijke diameters de cavitatiethreshold
niet, waardoor de waterkolom behouden bleef en geen cavitatie optrad (e.g. Tyree et al., 1984;
Kikuta et al., 2003).
De VC waren gelijkaardig qua vorm, maar vertoonden een verschuiving doorheen het seizoen
(figuur 3.9 A). Op het einde van het seizoen (december, experiment 1) vertoonde het xyleem een
zwakke inkrimping. Naarmate het nieuwe seizoen vorderde, vertoonde de tak een veel sterkere
inkrimping voor eenzelfde hoeveelheid cavitatie en caviteerde dus nog niet bij een inkrimping
die wel reeds cavitatie induceerde op het einde van het seizoen (maart tot mei, experimenten
3 - 7). Omdat cavitatiethresholds bepaald worden door pitporiekarakteristieken, welke op hun
beurt grotendeels anatomisch bepaald zijn en dus vastliggen per cultivar (e.g. Choat et al., 2008),
wordt aangenomen dat de takken eenzelfde range aan waterpotentialen ondergingen. Eenzelfde
waterpotentiaalverschil (∆Ψ) vertaalde zich dus in een kleinere xyleeminkrimping (∆VV ) naarmate
het seizoen vorderde.
Er wordt geopperd dat dit fenomeen verklaard kan worden door de xyleemdiameter als maat voor
de waterpotentiaal. Beide parameters zijn gerelateerd aan de hand van de elasticiteitsmodulus (ε
[MPa]) van de xyleemwanden zoals uitgedrukt door formule 4.1:
∆Ψ = ε∆V
V(4.1)
Hierin wordt een grotere elasticiteit van de celwanden uitgedrukt als een kleinere elasticiteitsmodu-
lus, die de potentiaalverandering weergeeft die overeenkomt met een bepaalde volumeverandering
(Lambers et al., 2008). Intrigliolo & Castel (2007) stellen dat de elasticiteit van de xyleemvaten
varieert doorheen het seizoen, met een grotere elasticiteit (kleinere ε) aan het begin van het sei-
zoen. Zo komt eenzelfde potentiaalverschil in het begin van het seizoen overeen met een grotere
volumeverandering. Dit uit zich in figuur 3.9 A door een verschuiving naar links op de x-as naar-
mate het nieuwe seizoen vordert en de vaten meer elastisch worden. De uiterst rechtse curves
komen immers overeen met druivelaars uit december en maart terwijl de uiterst linkse curves in
mei werden opgemeten. Op basis van figuur 3.9 A kan een keerpunt verwacht worden, waarbij de
elasticiteit terug zal afnemen en curves terug naar rechts op de x-as zullen opschuiven.
De xyleeminkrimping is bijgevolg een maat voor de waterpotentiaal, maar de kwantitatieve relatie
varieert doorheen het seizoen als gevolg van een variabele elasticiteit. Sperry et al. (1987) stellen
dat druivelaars overgaan van een dormant stadium naar een fase van actieve groei en bladexpansie
eind mei. Omdat de druivelaars vanaf 12 maart 2012 in de serres waren opgekweekt, kan deze
overgang zich vroeger hebben voorgedaan, waardoor de elasticiteit reeds begin mei (experiment
6) beduidend groter was. Ook Alsina et al. (2007) rapporteren een toename in de elasticiteit
(afnemende elasticiteitsmodulus) van 10-jarige Chardonnay druivelaars doorheen een periode van

Hoofdstuk 4. Discussie 65
eind mei - eind september. Dit past in onze resultaten en suggereert dat het keerpunt niet voor
september optreedt. Verder onderzoek is vereist om deze seizoenale trend beter in kaart te bren-
gen.
In figuur 3.9 B worden de destructieve VC in cumulatief absoluut aantal signalen weergegeven.
Hierin blijkt duidelijk dat naast de inkrimping ook het aantal cavitatiesignalen sterk varieerde
doorheen het seizoen. Het totaal aantal gecaviteerde vaten vertoonde een dalende trend vanaf
december (experiment 1 en 3), met een minimum eind maart (experiment 4), en opnieuw een
spectaculaire stijging begin mei (experimenten 6 en 7). Dit kan een uiting zijn van de overgang van
dormante toestand naar actieve groei. Zo stellen Sperry et al. (1987) dat xyleemvaten gedurende
de winter grotendeels met lucht gevuld zijn door vries-geınduceerde cavitatie. Dergelijke vaten
waren bijgevolg reeds gecaviteerd bij de aanvang van het dehydratatie-experiment en konden
op die manier geen UAE-signalen meer genereren. Voor de bladzetting worden de vaten terug
gevuld met sap om de transpiratiestroom te kunnen onderhouden. Het herstel van deze oude
xyleemvaten is cruciaal, gezien nieuwe xyleemvaten slechts gevormd worden na het ontluiken van
de bladeren. Dit hervullen is zichtbaar onder de vorm van ’bleeding’, waarbij xyleemsap uit de
druiventakken druppelt. Het wordt veroorzaakt door worteldruk (tot 500 kPa), waardoor het
sap uit de vaten naar boven wordt gestuwd en gasbellen verwijderd worden (e.g. Sperry et al.,
1987). Op die manier kan de trend in de VC in figuur 3.9 B geınterpreteerd worden: in maart
werd een druiventak onderzocht die nog grotendeels vrij was van blaadjes maar reeds worteldruk
vertoonde. De opgemeten cavitatie was dus enkel het gevolg van de oude xyleemvaten die nog
geleidend waren of reeds opnieuw met sap gevuld waren onder invloed van worteldruk. In mei
bevonden de druivelaars zich in een stadium van actieve groei, waarin de bladeren reeds ontloken
waren en nieuwe xyleemvaten gevormd waren, met een hoger aantal UAE-signalen tot gevolg. In
overeenstemming met deze resultaten rapporteren ook Tibbetts & Ewers (2000) een seizoenale
trend in het aantal cavitaties. Gezien de seizoenale trend een belangrijke invloed uitoefent op het
verloop en de cUAE50-waarde van de curve, zal deze in rekening gebracht moeten worden bij de
praktische toepassing van de VC.
De niet-destructieve VC uit experiment 2 (figuur 3.9 B en 3.10) week af van de andere VC wat
betreft vorm en grootteorde. De tak werd immers niet toegelaten om volledig uit te drogen. Uit
figuur 3.9 B blijkt dat de curve geen volledig sigmoıdaal profiel vertoonde, maar slechts een de-
tail voorstelde van de initiele fase van een VC doorheen een volledige dehydratatie. Een analoge
vorm werd gerapporteerd door Holtta et al. (2005), die een niet-destructieve VC ’in situ’ opstel-
den van een Schotse den. Als gevolg van neerslag en bodemvocht onderging de den ook geen
volledige dehydratatie, vandaar de analogie met de resultaten uit experiment 2. Daarenboven
bedroeg de cumulatieve hoeveelheid cavitatiesignalen in experiment 2 slechts een fractie van het
totaal aantal vaten (figuur 3.9 B). Hieruit kan worden afgeleid dat het opstellen van een volledige
niet-destructieve VC enkel destructief kan gebeuren door een verregaande uitdroging te bewerk-

66 Hoofdstuk 4. Discussie
stelligen. Een onvolledige uitdroging bij ’in situ’ experimenten zorgt er bovendien voor dat de
P50-waarden bekomen uit dergelijke experimenten niet kwantitatief beschouwd kunnen worden.
Eens de curve is opgesteld voor het betreffende soort, kan de opvolging van beide parameters wel
op een niet-destructieve manier plaatsvinden.
Figuur 3.10 toont het precieze verloop van een VC: daar waar afnemende diameters steeds ge-
paard gingen met een toenemend aantal cavitatiesignalen, was het omgekeerde niet steeds waar.
Dit wordt ook duidelijk in figuur 3.6 B, waar een klein aandeel (10%) van de cavitatiesignalen
optrad wanneer de diameter niet inkromp. Deze resultaten zijn in overeenstemming met de bevin-
dingen van Holtta et al. (2005). Een aantal oorzaken kunnen aan de basis van dit fenomeen liggen.
Enerzijds kunnen dergelijke signalen een uiting zijn van een zogenaamd stochastisch embolisatie-
patroon. Deze theorie berust op homogene of heterogene nucleatie waarin spontaan luchtbellen
ontstaan onder invloed van thermodynamische fluctuaties. Deze veroorzaken cavitatie die niet kan
voorspeld worden door thresholds. Hoe langer een zekere zuigspanning aanhoudt, des te meer kans
op dergelijke spontante embolisatie. Anderzijds kan ook het deterministische mechanisme aan de
basis van dergelijk patroon liggen, waarin de vaten niet kunnen caviteren bij hun vooropgestelde
threshold ten gevolge van een tijdelijke ongunstige toestand (e.g. belemmering van luchtintrede)
(Kikuta et al., 2003; Holtta et al., 2005). Tenslotte kunnen deze pieken ook het gevolg zijn van
schokken, veroorzaakt door het uitvoeren van de metingen. Bijkomende experimenten moeten
hier uitsluitsel bieden.
4.4.2 Praktisch gebruik en toepassingen van de niet-destructieve VC
Aan de hand van de relatie tussen xyleeminkrimping (∆D) en waterpotentiaal (Ψ) kan een verband
gelegd worden tussen de niet-destructieve en de destructieve VC. Volgende relatie werd opgesteld
tussen beide parameters op basis van experimenten 4, 5 en 6: ∆D=0.0373 + 0.1331 Ψ (R2=0.74;
resultaten niet weergegeven). Aan de hand van deze vergelijking kon de waterpotentiaal berekend
worden, die overeenkomt met een welbepaalde diameterinkrimping. Zo werd de waterpotentiaal
berekend voor de aanvang van cavitatie, alsook deze waarbij de cUAE-50-waarde bereikt werd.
De inkrimping die overeenkwam met de start van de cavitatie voor experimenten 1 tot 7 lag tus-
sen 0.1 en 0.2 mm. Gebaseerd op bovenstaande relatie werd de overeenkomstige waterpotentiaal
berekend (tussen -0.47 en -1.22 MPa), die dus een indicatie biedt van de cavitatiethreshold. Op
basis van de wet van Laplace (formule 1.4) corresponderen dergelijke cavitatiethresholds met pit-
poriedimensies van 0.10 - 0.30 µm. Sperry et al. (1987) bevestigt dat dergelijke waarden binnen
de poriedimensies liggen van verschillende hardhoutspecies.
Daarnaast werden de cUAE50-waarden van experimenten 1, 3, 4, 6 en 7 omgerekend tot wa-
terpotentialen van respectievelijk -2.31, -2.83, -2.53, -4.41 en -8.24 MPa. Deze waterpotentialen
kunnen op hun beurt vergeleken worden met de corresponderende waarden uit de destructieve
VC. In tegenstelling tot de geobserveerde P50-waarde van de destructieve VC (-1.99 MPa, figuur

Hoofdstuk 4. Discussie 67
3.8), stemmen de waarden van de niet-destructieve VC wel overeen met de P50-waarden voor
Chardonnay druivelaars zoals gerapporteerd door Choat et al. (2010) (-2.97 MPa ) en Alsina
et al. (2007) (-2.27 MPa). De afwijkende waarden van experimenten 6 en 7 kunnen veroor-
zaakt zijn door de variabele elasticiteit van de xyleemvaten. De relatie tussen ∆D en Ψ werd
immers collectief opgesteld waardoor een gemiddelde elasticiteit wordt aangenomen voor de vol-
ledige meetperiode. Wanneer dit verband echter enkel op basis van experiment 6 wordt opgesteld
(∆D=0.0402 + 0.1481 Ψ; R2=0.76), zijn de cUAE50-waarden (respectievelijk -3.99 en -7.4 MPa)
nog steeds meer negatief dan de andere waarden. Hieruit blijkt dat nog andere oorzaken aan de
basis liggen van dit gegeven en wordt een seizoenale trend gesuggereerd in de gevoeligheid van
de vaten. Melcher et al. (2003) stellen een toenemende gevoeligheid aan cavitatie vast op jaar-
basis in suikeresdoorn en suggereren dat oudere xyleemvaten een verzwakte weerstand bezitten
door (i) voorgaande cavitatiewaarnemingen, (ii) schade gedurende de wintermaanden of (iii) door
verloop van tijd. Op basis van deze bevindingen kan vooropgesteld worden dat de druivelaars
van december tot maart (experimenten 1-4) een verzwakte weerstand voor cavitatie ontwikkeld
hebben, resulterend in een minder negatieve cUAE50-waarde, waarna de ontwikkeling van nieuwe
vaten in mei (figuur 3.9 B) voor een versterkte weerstand tegen cavitatie gezorgd kan hebben.
Het dient benadrukt te worden dat ouderdom een factor kan zijn die meespeelt, maar niet met
zekerheid als de enige correcte oorzaak van de discrepantie aangeduid kan worden. De opgestelde
relatie is immers slechts een grove benadering en biedt louter een indicatie van de relatie tussen
diameterinkinkrimping en waterpotentiaal. Om meer inzicht te krijgen in de situatie dienen de
niet-destructieve VC in de toekomst gerelateerd te worden aan de heersende waterpotentialen om
zo meer duidelijkheid te creeeren omtrent het seizoenale verloop van zowel de elasticiteit als de
gevoeligheid van de vaten.
Daar waar een destructieve VC een directe maat geeft van een fysische eigenschap van de plant die
gelinkt kan worden aan de overleving, is cavitatie minder eenvoudig te relateren aan een fysische
toestand. In hoeverre de P50-waarde een interpretatie geeft van de hydraulische geleidbaarheid
en zo een beeld geeft van de mate waarin een plant leidt onder droogtestress, is immers nog onder
debat. Alhoewel diverse studies een nauwe overeenkomst hebben uitgewezen tussen P50-waarden
en cUAE50-waarden (e.g. Cochard, 1992; Hacke & Sauter, 1996; Rosner, 2004; Barnard et al.,
2011), stellen Kikuta et al. (2003) dat het verlies aan hydraulische geleidbaarheid meestal groter
is dan het percentage gecaviteerde vaten, wat wordt bevestigd in onze data. Reeds voordat de
laatste UAE-signalen worden opgemeten, zo opperen zij, heeft de plant immers zijn geleidbaar-
heid verloren omdat de waterstroming slechts tot stand komt in een aaneenschakeling van niet
gecaviteerde vaten. Zo draagt een niet gecaviteerd vat dat geısoleerd voorkomt in de plant, niet
bij tot de geleidbaarheid, maar kan wel nog een UAE-signaal genereren. De VC mag bijgevolg niet
als een louter fysische eigenschap van een individueel vat beschouwd worden, maar dient eerder
als eigenschap van de volledige weefselstructuur geınterpreteerd te worden (Kikuta et al., 2003;
Loepfe et al., 2007). Daarnaast kan ook de gelhypothese van Jacobsen & Pratt (2012) hier een
invloed uitoefenen: als gevolg van gelvorming liggen gecaviteerde vaten niet als enige aan de ba-

68 Hoofdstuk 4. Discussie
sis van afnemende hydraulische geleidbaarheid, wat de discrepantie tussen de hydraulische en de
akoestische methodes kan verklaren. Jackson et al. (1995) en Manoharan & Pammenter (2005) on-
dervonden dan weer een omgekeerde relatie. Ze stelden vast dat de hydraulische P50-waarde meer
negatief is dan de cUAE50-waarde. De brede waaier aan verschillende resultaten bewijst dat akoes-
tische data niet eenduidig gebruikt kunnen worden om het verlies aan hydraulische geleidbaarheid
kwantitatief te beoordelen. Zoals Manoharan & Pammenter (2005) het gepast verwoorden: ”De
invloed van cavitatiewaarnemingen op de hydraulische geleidbaarheid wordt niet op een directe
manier gemeten door akoestische methoden en daarom is de interpretatie van gevoeligheid op
basis van deze methoden meer subjectief”. Een mogelijke oorzaak in de moeilijke interpretatie
van cavitatiesignalen in termen van hydraulische geleidbaarheid is de eenheid ’aantal cavitatie-
signalen’. Deze differentieert niet voldoende tussen de hydraulische significantie van verschillende
vaten, zoals Rosner et al. (2006) stellen. De onduidelijke herkomst van de signalen in combinatie
met de verspreiding van de signalen doorheen het hout, evenals overlappende cavitatiesignalen
en signalen buiten het opgemeten frequentiegebied dragen bij tot deze onzekerheid (Sandford &
Grace, 1985; Jackson & Grace, 1996).
Desalniettemin is het een uiterst gebruiksvriendelijke methode om verschillende soorten te verge-
lijken qua droogtegevoeligheid (e.g. Alsina et al., 2007). Aan de hand van de cavitatiesensor kan
de waterpotentiaalthreshold bepaald worden vanaf het ogenblik waarop schade wordt toegebracht
aan het watergeleidingssysteem. Daarenboven kunnen continue UAE-signalen bijdragen tot het
onderzoek naar het effect van microklimatologische variabelen op de cavitatietrend (Jackson &
Grace, 1996). Tenslotte kan de cavitatiesensor op verschillende plantenorganen geplaatst worden,
waardoor de vergelijking tussen verschillende plantenorganen toegelaten wordt (Jackson & Grace,
1996) om de spatiale variatie in het voorkomen van cavitatie te onderzoeken.

BESLUIT EN TOEKOMSTPERSPECTIEF


Hoofdstuk 5
Besluit en toekomstperspectief
Het doel van deze thesis was tweevoudig. Ten eerste werd er gezocht naar verbanden tussen het
cavitatiepatroon in relatie tot diametervariaties onder diverse omstandigheden om tot een beter
inzicht in de uitdrogingsdynamiek te komen. Onder condities van beperkte uitdroging, zoals le-
vende planten dagelijks ondergaan, bleek het diameterverloop een goede indicator te zijn voor
het optreden van cavitatie: afnemende diameters onder het voorheen laagst voorgekomen niveau
gingen vrijwel steeds gepaard met cavitatie. Daarentegen bood het gelijktijdig opmeten van de
schors- en xyleemdiameter in combinatie met het cavitatiepatroon en het waterverlies cruciale in-
formatie over de plant tijdens verregaande dehydratatie. Door de combinatie van deze parameters
konden diverse stadia van uitdroging in de plant gekarakteriseerd worden en kon gedifferentieerd
worden tussen de verschillende mechanismen die sequentieel plaatsgrepen. Op die manier kan de
huidige beperkte kennis omtrent verregaande dehydratatie in angiospermen aangevuld worden en
kan de methode bijdragen tot het interpreteren van de niet-destructieve vulnerability curve (VC),
het tweede objectief van deze thesis.
Vooraleer werd overgegaan op de niet-destructieve VC werden destructieve VC opgesteld, zoals
tot op heden standaard toegepast. Zo werd getracht de niet-destructieve VC te kaderen en te
staven met huidige kennis. Het opstellen van dergelijke destructieve VC bleek echter geen si-
necure bij druivelaars. Er werden verschillende aandachtspunten aangehaald die gerespecteerd
dienen te worden om de natuurlijke situatie in de plant niet te sterk te modificeren tijdens het
meetproces. Zo dient voldoende zorg besteed te worden aan de samenstelling van het water waar-
mee de tak doorstroomd wordt, om verstopping, belvorming en zwelling van pitporien tegen te
gaan. Daarnaast moet de lengte van de tak beperkt worden, rekening houdend met de maximale
vatlengte van het species. Tenslotte dient de duur van hydraulische metingen beperkt te worden
om gelvorming en dus artificiele verstopping te minimaliseren (<1 dag (Jacobsen & Pratt, 2012)).
Desalniettemin blijft het een gecompliceerde procedure, die veel ervaring vergt en een destructieve
staalname vereist. Ook in de literatuur vertonen de destructieve VC sterk uiteenlopende resul-
71

72 Hoofdstuk 5. Besluit en toekomstperspectief
taten en blijken ze erg afhankelijk van de omstandigheden en de wijze waarop de experimenten
werden uitgevoerd. Er werd dan ook geconcludeerd dat het opstellen van een niet-destructieve
VC, die minder afhankelijk is van de wijze van data verzameling, een belangrijke meerwaarde kan
bieden in het onderzoek naar de waterrelaties in druivelaars.
De voorgestelde niet-destructieve VC toonden de relatie tussen de xyleemdiameter en de UAE.
De combinatie van beide parameters werd nog nooit opgemeten tijdens verregaande dehydrata-
tie, waardoor de curves uniek zijn in hun soort. De resultaten bevestigden dat beide parameters
eenduidig gerelateerd zijn en een belangrijke informatiebron vormen voor de cavitatiestatus van
de plant. Aan de hand van de niet-destructieve VC kon de waterpotentiaalthreshold bepaald
worden vanaf wanneer cavitatie optrad en vanaf wanneer grote schade werd toegericht aan het
watergeleidingssysteem. Onze studie toont aan dat het van cruciaal belang is de tak volledig te
laten uitdrogen om een juiste en volledige inschatting te maken van de droogtegevoeligheid van
een species. Enkel op die manier caviteert de meerderheid van de vaten en kunnen de waarden
relatief uitgezet worden. Tenslotte was er een duidelijke seizoenale trend te beschouwen tussen
de diverse niet-destructieve VC die werden opgesteld van december tot mei. Hieruit werd gesug-
gereerd dat de seizoenale ontwikkeling van de plant opgevolgd kan worden aan de hand van een
niet-destructieve VC, waarbij een indicatie wordt gekregen van de ontwikkeling van nieuwe vaten
en de trend in de elasticiteit.
Toekomstig onderzoek dient te focussen op een meer gesofisticeerde analyse van de cavitatiesigna-
len, zodat gedifferentieerd kan worden tussen de relevantie van de diverse cavitatiesignalen om hun
effect op de waterbalans te kunnen inschatten. Op die manier kan tot een betere overeenkomst
gekomen worden tussen de hydraulische en de akoestische methoden. Recente, nieuwe software
waarmee diverse eigenschappen van de UAE-signalen opgemeten kunnen worden, zijn veelbelo-
vend en voorspellen een belangrijke rol van UAE in het monitoren van de waterbalans. Deze
toekomstige methoden zouden daarenboven kunnen bijdragen tot het onderscheid in de verschil-
lende bronnen van cavitatiesignalen (e.g. levende cellen, microscheuren), wat tot dusver nog louter
op interpretatie berustte. Daarnaast dient de relatie tussen het verloop van niet-destructieve VC
en de corresponderende waterpotentialen verder onderzocht te worden om op die manier tot een
beter zicht te komen van het seizoenale verloop van de elasticiteit en de gevoeligheid van de vaten.
Tot slot dient het gebruik van de niet-destructieve VC praktisch uitgewerkt en geımplementeerd
te worden, zodat ze in de toekomst kan bijdragen tot het modelleren en opbouwen van irrigatie-
strategieen, specifiek afgestemd op de plant.

Bibliografie
Almeras, T. (2008). Mechanical analysis of the strains generated by water tension in plant stems.
Part II: strains in wood and bark and apparent compliance. Tree Physiology, 28 (10), 1513–1523.
Alsina, M. M., de Herralde, F., Aranda, X., Save, R., & Biel, C. (2007). Water relations and
vulnerability to embolism are not related: Experiments with eight grapevine cultivars. Vitis,
46 (1), 1–6.
Ayup, M., Hao, X., Chen, Y., Li, W., & Su, R. (2012). Changes of xylem hydraulic efficiency
and native embolism of Tamarix ramosissima Ledeb. seedlings under different drought stress
conditions and after rewatering. South African Journal of Botany, 78, 75–82.
Barnard, D. M., Meinzer, F. C., Lachenbruch, B., McCulloh, K. A., Johnson, D. M., & Woodruff,
D. R. (2011). Climate-related trends in sapwood biophysical properties in two conifers: avoi-
dance of hydraulic dysfunction through coordinated adjustments in xylem efficiency, safety and
capacitance. Plant, Cell and Environment, 34 (4), 643–654.
Boyer, J. S. (1985). Water transport. Annual Review of Plant Physiology, 36, 473–516.
Brodersen, C. R., McElrone, A. J., Choat, B., Matthews, M. A., & Shackel, K. A. (2010). The
dynamics of embolism repair in xylem: in vivo visualizations using high-resolution computed
tomography. Plant physiology, 154 (3), 1088–95.
Bucci, S. J., Scholz, F. G., Goldstein, G., Meinzer, F. C., & Sternberg, L. D. S. (2003). Dynamic
changes in hydraulic conductivity in petioles of two savanna tree species: factors and mecha-
nisms contributing to the refilling of embolized vessels. Plant, Cell and Environment, 26 (10),
1633–1645.
Cai, J. & Tyree, M. T. (2010). The impact of vessel size on vulnerability curves: data and models
for within-species variability in saplings of aspen, Populus tremuloides Michx. Plant, Cell and
Environment, 33 (7), 1059–69.
Chaves, M. M., Zarrouk, O., Francisco, R., Costa, J. M., Santos, T., Regalado, A. P., Rodrigues,
M. L., & Lopes, C. M. (2010). Grapevine under deficit irrigation: hints from physiological and
molecular data. Annals of Botany, 105 (5), 661 –676.
73

74 Bibliografie
Choat, B., Cobb, A. R., & Jansen, S. (2008). Structure and function of bordered pits: new
discoveries and impacts on whole-plant hydraulic function. New phytologist, 177 (3), 608–625.
Choat, B., Drayton, W. M., Brodersen, C. R., Matthews, M. A., Shackel, K. A., Wada, H., &
McElrone, A. J. (2010). Measurement of vulnerability to water stress-induced cavitation in
grapevine: a comparison of four techniques applied to a long-vesseled species. Plant, Cell and
Environment, 33 (9), 1502–12.
Christman, M. A., Sperry, J. S., & Adler, F. R. (2009). Testing the ’rare pit’ hypothesis for xylem
cavitation resistance in three species of Acer. New phytologist, 182 (3), 664–674.
Cochard, H. (1992). Vulnerability of several conifers to air embolism. Tree physiology, 11 (1),
73–83.
Cochard, H. (2006). Cavitation in trees. Comptes Rendus Physique, 7 (9-10), 1018–1026.
Cochard, H. (2008). Xylem instruction manual and tutorial for xylem embolism measurements.
Clermont-Ferrand-Theix, Frankrijk: Bronkhorst.
Cochard, H., Bodet, C., Ameglio, T., & Cruiziat, P. (2000). Cryo-Scanning Electron Microscopy
Observations of Vessel Content during Transpiration in Walnut Petioles. Facts or Artifacts?
Plant physiology, 124 (3), 1191–1202.
Cochard, H., Forestier, S., & Ameglio, T. (2001). A new validation of the Scholander pres-
sure chamber technique based on stem diameter variations. Journal of Experimental Botany,
52 (359), 1361–1365.
Cruiziat, P. & Tyree, M. T. (1990). La montee de la seve dans les arbres. La recherche, 21 (220),
406–414.
Dixon, J. H. & Joly, J. (1895). On the ascent of sap. Philosophical transactions of the Royal
Society of London, 186, 563–576.
Durante, M., Maseda, P. H., & Fernandez, R. J. (2011). Xylem efficiency vs. safety: Acclima-
tion to drought of seedling root anatomy for six Patagonian shrub species. Journal of Arid
Environments, 75 (5), 397–402.
Espino, S. & Schenk, H. J. (2011). Mind the bubbles: achieving stable measurements of maximum
hydraulic conductivity through woody plant samples. Journal of Experimental Botany, 62 (3),
1119–1132.
Fernandez, R. & Cuevas, M. V. (2010). Agricultural and Forest Meteorology Irrigation scheduling
from stem diameter variations: A review. Agricultural and Forest Meteorology, 150 (2), 135–151.

Bibliografie 75
Goldstein, G., Andrade, J. L., Meinzer, F. C., Holbrook, N. M., Cavelier, J., & Jackson, P. (1998).
Stem water storage and diurnal patterns of water use in tropical forest canopy trees. Plant,
Cell and Environment, 21 (4), 397–406.
Hacke, U. G. & Sauter, J. J. (1996). Drought-lnduced Xylem Dysfunction in Petioles, Branches,
and Roots of Populus balsamifera L . and Alnus glutinosa (L.) Gaertn. Plant physiology, 111 (2),
413–417.
Hacke, U. G., Sperry, J. S., Wheeler, J. K., & Castro, L. (2006). Scaling of angiosperm xylem
structure with safety and efficiency. Tree physiology, 26 (6), 689–701.
Hietz, P., Rosner, S., Sorz, J., & Mayr, S. (2008). Comparison of methods to quantify loss of
hydraulic conductivity in Norway spruce. Annals of Forest Science, 65 (5), 502–509.
Holbrook, N. M., Burns, M. J., & Field, C. B. (1995). Negative xylem pressures in plant-a test of
the balancing pressure technique. Science, 270 (5239), 1193–1194.
Holtta, T., Cochard, H., Nikinmaa, E., & Mencuccini, M. (2009). Capacitive effect of cavitation
in xylem conduits: results from a dynamic model. Plant, Cell and Environment, 32 (1), 10–21.
Holtta, T., Juurola, E., Lindfors, L., & Porcar-Castell, A. (2011). Cavitation induced by a
surfactant leads to a transient release of water stress and subsequent run away embolism in
Scots pine (Pinus sylvestris) seedlingsf. Journal of Experimental Botany, 63 (2), 1057–1067.
Holtta, T., Vesala, T., & Nikinmaa, E. (2007). A model of bubble growth leading to xylem conduit
embolism. Journal of Theoretical Biology, 249 (1), 111–23.
Holtta, T., Vesala, T., Nikinmaa, E., Peramaki, M., Siivola, E., & Mencuccini, M. (2005). Field
measurements of ultrasonic acoustic emissions and stem diameter variations. New insight into
the relationship between xylem tensions and embolism. Tree physiology, 25 (2), 237–43.
Holtta, T., Vesala, T., Peramaki, M., & Nikinmaa, E. (2002). Relationships between Embolism,
Stem Water Tension, and Diameter Changes. Journal of Theoretical Biology, 215 (1), 23–38.
Holtta, T., Vesala, T., Sevanto, S., Peramaki, M., & Nikinmaa, E. (2006). Modeling xylem and
phloem water flows in trees according to cohesion theory and Munch hypothesis. Trees, 20,
67–78.
Intrigliolo, J. R. & Castel, J. R. (2007). Evaluation of grapevine water status from trunk diameter
variations. Irrigation Science, 26 (1), 49–59.
Irvine, J. & Grace, J. (1997). Continuous measurements of water tensions in the xylem of trees
based on the elastic properties of wood. Planta, 202 (4), 455–461.

76 Bibliografie
Jackson, G. E. & Grace, J. (1996). Field measurements of xylem cavitation: are acoustic emissions
useful? Journal of Experimental Botany, 47 (304), 1643–1650.
Jackson, G. E., Irvine, J., & Grace, J. (1995). Xylem cavitation in Scots pine and Sitka spruce
saplings during water stress. Tree Physiology, 15, 783–790.
Jacobsen, A. L. & Pratt, R. B. (2012). No evidence for an open vessel effect in centrifuge-based
vulnerability curves of a long-vesselled liana (Vitis vinifera ). New phytologist, 194 (4), 982–990.
Jia, X., Zhang, L., Ma, R., Wang, Z., Zhang, Q., & Yao, Y. (2006). Ultrasonic Acoustic Emissions
from Leaf Xylem of Potted Wheat Subject to a Soil Drought and Rewatering Cycle. Agricultural
sciences in China, 5 (5), 346–355.
Jones, H. G. (1992). Plants and microclimate. A quantitative approach to environmental plant
physiology (2nd ed.). Cambridge, Verenigd Koninkrijk: Cambridge University Press.
Jury, W. A. & Horton, R. (2004). Soil physics (6th ed.). Hoboken, Belgie: John Wiley & Sons
Inc.
Kikuta, S. B., Hietz, P., & Richter, H. (2003). Vulnerability curves from conifer sapwood sections
exposed over solutions with known water potentials. Experimental Biology, 54 (390), 2149–2155.
Kikuta, S. B. & Salleo, S. (1997). Ultrasound acoustic emissions from dehydrating leaves of
deciduous and evergreen trees. Plant, Cell and Environment, 20 (11), 1381–1390.
Kolb, K. J. & Davis, S. D. (1994). Drought Tolerance and Xylem Embolism in Co-Occurring
Species of Coastal Sage and Chaparral. Ecology, 75 (3), 648–659.
Kolb, K. J., Sperry, J. S., & Lamont, B. B. (1996). A method for measuring xylem hydraulic
conductance and embolism in entire root and shoot systems. Journal of Experimental Botany,
47 (304), 1805–1810.
Kowalski, S. J. & Smockzkiewicz, A. (2004). Identification of wood destruction during drying.
Maderas. Ciencia y tecnologıa, (2nd), 133–143.
Lambers, H., Stuart Chapin III, F., & Pons, T. L. (2008). Plant Physiological Ecology (2nd ed.).
New York, USA.
Leonardi, C., Guichard, S., & Bertin, N. (2000). High vapour pressure deficit influences growth,
transpiration and quality of tomato fruits. Scientia Horticulturae, 84 (3-4), 285–296.
Loepfe, L., Martınez-Vilalta, J., Pinol, J., & Mencuccini, M. (2007). The relevance of xylem
network structure for plant hydraulic efficiency and safety. Journal of Theoretical Biology,
247 (4), 788–803.

Bibliografie 77
Lovisolo, C., Perrone, I., Hartung, W., & Schubert, A. (2008). An abscisic acid-related reduced
transpiration promotes gradual embolism repair when grapevines are rehydrated after drought.
New Phytologist, 180 (3), 642–651.
Manoharan, P. & Pammenter, N. W. (2005). Comparison between ultrasonic acoustic emission
(UAE) and hydraulically measured loss of hydraulic conductance in Eucalyptus spp. Clone
GU210. Journal of science of the University of Kelaniya Sri Lanka, 2, 85–104.
Martınez-Vilalta, J., Pinol, J., & Beven, K. (2002). A hydraulic model to predict drought-induced
mortality in woody plants: an application to climate change in the Mediterranean. Ecological
Modelling, 155 (2-3), 127–147.
Mayr, S. & Rosner, S. (2011). Cavitation in dehydrating xylem of Picea abies: energy properties
of ultrasonic emissions reflect tracheid dimensions. Tree Physiology, 31 (1), 59–67.
McElrone, A. J., Sherald, J. L., & Forseth, I. N. (2003). Interactive effects of water stress and
xylem-limited bacterial infection on the water relations of a host vine. Journal of Experimental
Biology, 54 (381), 419–430.
Melcher, P. J., Zwieniecki, M. A., & Holbrook, N. M. (2003). Vulnerability of Xylem Vessels to Ca-
vitation in Sugar Maple. Scaling from Individual Vessels to Whole Branches. Plant Physiology,
131, 1775–1780.
Milburn, J. A. (1973). Cavitation studies on whole Ricinus plants by acoustic detection. Planta,
112, 333–342.
Milburn, J. A. & Johnson, R. P. C. (1966). The conduction of sap. Detection of vibrations
produced by sap cavitation in Rhicins xylem. Planta, 69, 43–52.
Offenthaler, I., Hietz, P., & Richter, H. (2001). Wood diameter indicates diurnal and long-term
patterns of xylem water potential in Norway spruce. Trees, 15 (4), 215–221.
Peramaki, M., Nikinmaa, E., Sevanto, S., Ilvesniemi, H., Siivola, E., Hari, P., & Vesala, T. (2001).
Tree stem diameter variations and transpiration in Scots pine : an analysis using a dynamic
sap flow model. Tree Physiology, 21 (12-13), 889–897.
Perez-Diez, J., Garcia-Prada, J., Romera-Juarez, F., & Diez-Jimenez, E. (2010). Mechanical
behaviour analyses of sap ascent. Journal of Biological Physics, 36 (4), 355–363.
Pittermann, J. & Sperry, J. S. (2003). Tracheid diameter is the key trait determining the extent
of freezing- induced embolism in conifers. Tree Physiology, 23, 907–914.
Rosner, S. (2004). Acoustic detection of cavitation events in water conducting elements of Norway
spruce sapwood. Journal of acoustic emission, 22, 110–118.

78 Bibliografie
Rosner, S. & Karlsson, B. (2011). Hydraulic efficiency compromises compression strength perpen-
dicular to the grain in Norway spruce trunkwood. Trees, 25 (2), 289–299.
Rosner, S., Karlsson, B., Konnerth, J., & Hansmann, C. (2009). Shrinkage processes in standard-
size Norway spruce wood specimens with different vulnerability to cavitation. Tree physiology,
29 (11), 1419–1431.
Rosner, S., Klein, A., Muller, U., & Karlsson, B. (2008). Tradeoffs between hydraulic and mecha-
nical stress responses of mature Norway spruce trunk wood. Tree Physiology, 28 (8), 1179–1188.
Rosner, S., Klein, A., Wimmer, R., & Karlsson, B. (2006). Extraction of features from ultra-
sound acoustic emissions: a tool to assess the hydraulic vulnerability of Norway spruce. New
Phytologist, 171 (1), 105–116.
Rosner, S., Konnerth, J., Plank, B., Salaberger, D., & Hansmann, C. (2010). Radial shrinkage and
ultrasound acoustic emissions of fresh versus pre-dried Norway spruce sapwood. Trees, 24 (5),
931–940.
Ruger, S., Ehrenberger, W., Arend, M., Geßner, P., Zimmermann, G., Zimmermann, D., Bentrup,
F., Nadler, A., Raveh, E., Sukhorukov, V. L., & Zimmermann, U. (2010). Comparative mo-
nitoring of temporal and spatial changes in tree water status using the non-invasive leaf patch
clamp pressure probe and the pressure bomb. Agricultural Water Management, 98 (2), 283–290.
Salleo, S., Lo Gullo, M. A., De Paoli, D., & Zippo, M. (1996). Xylem recovery from cavitation-
induced embolism in young plants of Laurus nobilis a possible mechanism. New Phytologist,
132 (1), 47–56.
Salleo, S., Lo Gullo, M. A., Raimondo, F., & Nardini, A. (2001). Vulnerability to cavitation of leaf
minor veins: any impact on leaf gas exchange? Plant, Cell and Environment, 24 (8), 851–859.
Sandford, A. P. & Grace, J. (1985). The measurement and interpretation of ultrasound from
woody stems. Journal of Experimental Botany, 36 (163), 298–311.
Scholander, P. F., Hammel, H. T., Hemmingsen, E. A., & Bradstreet, E. D. (1964). Hydrostatic
pressure and osmotic potential in leaves of mangroves and some other plants. Botany, 52,
119–125.
Schultz, H. R. (2000). Climate change and viticulture: A European perspective on climatology,
carbon dioxide and UV-B effects. Australian Journal of Grape and Wine Research, 6 (1), 2–12.
Schultz, H. R. (2003). Differences in hydraulic architecture account for near-isohydric and aniso-
hydric behaviour of two field-grown Vitis vinifera L. cultivars during drought. Plant, Cell and
Environment, 26 (8), 1393–1405.

Bibliografie 79
Schultz, H. R. & Matthews, M. A. (1988). Resistance to Water Transport in Shoots of Vitis
vinifera L. : Relation to Growth at Low Water Potential. Plant physiology, 88 (3), 718–24.
Sevanto, S., Holtta, T., & Holbrook, N. M. (2011). Effects of the hydraulic coupling between
xylem and phloem on diurnal phloem diameter variation. Plant, Cell and Environment, 34 (4),
690–703.
Sevanto, S., Holtta, T., Markkanen, T., & Peramaki, M. (2005). Relationships between diurnal
xylem diameter variation and environmental factors in Scots pine. Boreal environment, 10 (5),
447–458.
Skene, D. S. & Balodis, V. (1968). A study of vessel length in Eucalyptus obliqua L’Herit. Journal
of Experimental Botany, 19 (4), 825–830.
Spanner, D. C. (1951). The Peltier effect and its use in the measurement of suction pressure.
Journal of experimental botany, 2 (2), 145–168.
Sperry, J. S., Holbrook, N. M., Zimmermann, M. H., & Tyree, M. T. (1987). Spring Filling of
Xylem Vessels in Wild Grapevine. Plant physiology, 83 (2), 414–417.
Sperry, J. S. & Tyree, M. T. (1988). Mechanism of Water Stress-Induced Xylem Embolism. Plant
Physiology, 88 (3), 581–587.
Steppe, K. (2004). Diurnal dynamics of water flow through trees: design and validation of a
mathematical flow and storage model. PhD thesis.
Steppe, K., Cnudde, V., Girard, C., Lemeur, R., Cnudde, J., & Jacobs, P. (2004). Use of X-ray
computed microtomography for non-invasive determination of wood anatomical characteristics.
Journal of Structural Biology, 148 (1), 11–21.
Steppe, K., De Pauw, D. J. W., Lemeur, R., & Vanrolleghem, P. A. (2006). A mathematical model
linking tree sap flow dynamics to daily stem diameter fluctuations and radial stem growth. Tree
physiology, 26 (3), 257–73.
Steudle, E. (2001). The cohesion-tension mechanism and the acquisition of water by plant roots.
Annual Review of Plant Physiology and Plant Molecular Biology, 52 (1), 847–875.
Sun, Q., Rost, T. L., & Matthews, M. A. (2008). Wound-induced vascular occlusions in Vitis
vinifera (Vitaceae): Tyloses in summer and gels in winter. American Journal of Botany, 95 (12),
1498–1505.
Tibbetts, T. J. & Ewers, F. W. (2000). Root pressure and specific conductivity in temperate lia-
nas: exotic Celastrus orbiculatus (Celastraceae) vs. native Vitis riparioa (Vitaceae). American
Journal of Botany, 87 (9), 1272–1278.

80 Bibliografie
Turner, N. C. (1988). Measurement of plant water status by the pressure chamber technique.
Irrigation Science, 9 (4), 289–308.
Tyree, M. T. (1997). The Cohesion-Tension theory of sap ascent: current controversies. Journal
of Experimental Botany, 48 (10), 1753–1765.
Tyree, M. T., Dixon, M. A., Tyree, E. L., & Johnson, R. (1984). Ultrasonic Acoustic Emissions
from the Sapwood of Cedar and Hemlock . Plant physiology, 75 (4), 988–992.
Tyree, M. T. & Ewers, F. W. (1991). The hydraulic architecture of trees and other woody plants.
New phytologist, 119 (3), 345–360.
Tyree, M. T. & Sperry, J. S. (1988). Do woody plants operate near the point of catastrophic
xylem dysfunction caused by dynamic water stress? Answers from a model. Plant physiology,
88 (3), 574–80.
Tyree, M. T. & Sperry, J. S. (1989). Characterization and propagation of acoustic emission signals
in woody plants: towards an improved acoustic emission counter. Plant, Cell and Environment,
12 (4), 371–382.
Tyree, M. T. & Zimmermann, M. H. (2001). Xylem structure and the ascent of sap (2nd ed.).
Springer-Verlag.
Ueda, M. & Shibata, E. (2001). Diurnal changes in branch diameter as indicator of water status
of Hinoki cypress Chamaecyparis obtusa. Trees, 15 (6), 315–318.
van den Honert, T. H. (1948). Water transport in plants as a caternary process (3th ed.). Faraday
Society Discussions.
van Doorn, W. G., Hiemstra, T., & Fanourakis, D. (2011). Hydrogel Regulation of Xylem Water
Flow : An Alternative Hypothesis. Plant physiology, 157 (4), 1642–1649.
van Ieperen, W., van Meeteren, U., & van Gelder, H. (2000). Fluid ionic composition influences
hydraulic conductance of xylem conduits. Journal of Experimental Botany, 51 (345), 769–776.
Wei, C., Steudle, E., Tyree, M. T., & Lintilhac, P. M. (2001). The essentials of direct xylem
pressure measurement. Plant, Cell and Environment, 24 (5), 549–555.
Wei, C., Tyree, M. T., & Steudle, E. (1999). Direct measurement of xylem pressure in leaves
of intact maize plants. A test of the cohesion-tension theory taking hydraulic architecture into
consideration. Plant physiology, 121 (4), 1191–206.
Westhoff, M., Zimmermann, D., Schneider, H., Wegner, L. H., Geßner, P., Jakob, P., & Bamberg,
E. (2009). Evidence for discontinuous water columns in the xylem conduit of tall birch trees.
Plant Biology, 11 (3), 307–327.

Wheeler, J. K., Sperry, J. S., Hacke, U. G., & Hoang, N. (2005). Inter-vessel pitting and cavitation
in woody Rosaceae and other vesselled plants: a basis for a safety versus efficiency trade-off in
xylem transport. Plant, Cell and Environment, 28 (6), 800–812.
Wolkerstorfer, S. V., Rosner, S., & Hietz, P. (2012). An improved method and data analysis for
ultrasound acoustic emissions and xylem vulnerability in conifer wood. Physiologia Plantarum.
Zeugin, F. (2004). Trockenstress-Untersuchung an Flaumeiche und Waldfohre im Wallis mittels
der Ultrasonic Acoustic Emission-Technik. PhD thesis.
Zimmermann, U., Schneider, H., Wegner, L. H., & Haase, A. (2004). Water ascent in tall trees:
does evolution of land plants rely on a highly metastable state? New Phytologist, 162 (3),
575–615.
Zufferey, V., Cochard, H., Ameglio, T., Spring, J.-L., & Viret, O. (2011). Diurnal cycles of
embolism formation and repair in petioles of grapevine (Vitis vinifera cv. Chasselas). Journal
of experimental botany, 62 (11), 3885–94.
Zweifel, R. & Item, H. (2000). Stem radius changes and their relation to stored water in stems of
young Norway spruce trees. Trees, 15 (1), 50–57.
Zweifel, R., Item, H., & Hasler, R. (2001). Link between diurnal stem radius changes and tree
water relations. Tree Physiology, 21 (12-13), 869–877.
Zweifel, R. & Zeugin, F. (2008). Ultrasonic acoustic emissions in drought-stressed trees: more
than signals from cavitation? The New phytologist, 179 (4), 1070–9.
Zwieniecki, M. A., Hutyra, L., Thompson, M. V., & Holbrook, N. M. (2000). Dynamic changes in
petiole specific conductivity in red maple (Acer rubrum L .), tulip tree (Liriodendron tulipifera
L .) and northern fox grape (Vitis labrusca L .). Plant, Cell and Environment, 23 (4), 407–414.



















