
Literatuurstudie Steffen De Vreese - Functionele en...
27
Transcript of Literatuurstudie Steffen De Vreese - Functionele en...


Vrijwaringsclausule Universiteit Gent, haar werknemers of studenten bieden geen enkele garantie met betrekking tot de juistheid of volledigheid van de gegevens vervat in deze masterproef, noch dat de inhoud van deze masterproef geen inbreuk uitmaakt op of aanleiding kan geven tot inbreuken op de rechten van derden. Universiteit Gent, haar werknemers of studenten aanvaarden geen aansprakelijkheid of verantwoordelijkheid voor enig gebruik dat door iemand anders wordt gemaakt van de inhoud van de masterproef, noch voor enig vertrouwen dat wordt gesteld in een advies of informatie vervat in de masterproef.

UNIVERSITEIT GENT
FACULTEIT DIERGENEESKUNDE
Academiejaar 2012- 2013
Functionele en beschrijvende anatomie van het gehoorapparaat van Cetacea
door
Steffen De Vreese
Promotoren: Drs. Marjan Doom Literatuurstudie in het kader Prof. dr. Paul Simoens van de Masterproef
© 2013 Steffen De Vreese

VOORWOORD Dank aan prof. dr. Paul Simoens om het mij mogelijk te maken over dit onderwerp een studie
te maken. Veel dank aan mijn promotor, drs. Marjan Doom, voor de hulp, de aanwijzingen en
de snelle verbeteringen van mijn proefversies. Daarnaast dank aan Martin Schellinck,
bibliothecaris op de afdeling Dierkunde (Ledeganck). Zonder hem was er essentiële,
interessante literatuur niet beschikbaar geweest voor de student. Dank aan Mathilde De Moor
voor de vertaling van de Latijnse zin uit Plinius’ Naturalis Historia.
Al is de wetenschap en haar bijhorende kennis de laatste jaren snel ontwikkeld en uitgebreid,
uiteraard met behulp van allerhande nieuwe en innovatieve technieken, toch blijft er een
mysterie aanwezig rond alles wat zich in de oceaan afspeelt en heeft afgespeeld. De evolutie
die walvissen hebben ondergaan is zo uniek dat het, me dunkt, wel moet fascineren. De
levenswijze van elke soort op zich is uniek en bovendien verbazingwekkend. Hun
communicatieve vaardigheden zijn soms zo inventief en hun gedrag zo menselijk dat hun
blijkbare intelligentie sowieso ontzag opwekt bij de aanschouwer. En dan is er nog niet
gesproken over hun anatomie. Alleen al de omvang van sommige soorten is bijzonder
indrukwekkend. Dit zijn slechts enkele voorbeelden van de fascinerende facetten van
walvissen en hoe meer men zich hierin verdiept, hoe groter de verbazing telkens blijkt. Het
gehoorapparaat is één van die onderdelen die toont hoe ingenieus de natuur is. Het is een
puzzelstukje dat zelf is opgebouwd uit nog kleinere stukjes die naadloos aansluiten om zo
een prachtig geheel te vormen, met name de majestueuze dieren die onze verbeelding
prikkelen, de walvissen. Hopelijk geniet u evenveel van deze literatuurstudie als ik had bij het
ontdekken van allerlei fascinerende feiten, maar ook van nog onopgeloste vragen. Het
mysterie blijft bestaan, maar hier is al een glimp…

INHOUDSOPGAVE
SAMENVATTING ............................................................................................. 1
INLEIDING ....................................................................................................... 2
1. Taxonomie ................................................................................................ 3
2. Invloed van omgeving op het gehoor ....................................................... 3
2.1 Fysica .................................................................................................. 3
2.2 Functionele classificatie ...................................................................... 5
3. Anatomie .................................................................................................. 6
3.1 Het uitwendig oor ................................................................................ 6
3.2 Het tympano-periotisch complex ......................................................... 8
3.3 Het middenoor .................................................................................... 9
3.4 Het binnenoor ................................................................................... 11
BESPREKING ................................................................................................ 16
BIJLAGEN ...................................................................................................... 18
REFERENTIELIJST ....................................................................................... 20

1
SAMENVATTING Deze literatuurstudie levert een glimp in de anatomie van het gehoorapparaat van walvissen.
De studie brengt een beeld van de feiten, de anatomische bevindingen en hun functionaliteit,
maar ook van de tegenstrijdigheden binnen de literatuur. Er wordt ingegaan op de anatomie,
zowel macroscopisch als microscopisch, en de bijhorende functie van verschillende
onderdelen van het gehoorapparaat van walvissen. Inleidend wordt een beknopte
voorstelling van de evolutie weergegeven van de voorouders van walvisachtigen die als
landzoogdier hun weg naar zee terugvonden. Hierdoor ondergingen deze dieren, die volledig
aangepast waren aan het leven op land, enkele noodzakelijke aanpassingen. Deze evolutie
impliceerde onder andere dat oren, die voorheen geluid opvingen dat zich voortplantte in
lucht, nu onder water moesten functioneren. Om enigszins inzicht te krijgen in de impact van
deze verandering, wordt een kleine inleiding gegeven in de fysische aspecten van het horen
onder water. Geluid gedraagt zich namelijk anders onder water dan in lucht, met andere
snelheden en intensiteiten waar het oor zich aan heeft aangepast.
Wegens de enorme variatie van de verschillende soorten binnen de groep van walvissen
wordt in deze literatuurstudie een classificatie gehanteerd die eerder functioneel is en zo een
overzicht behoudt.
Nadien wordt dieper ingegaan op de unieke bouw van het gehoorapparaat van het uitwendig
‘zichtbare’ deel naar de diepere structuren die in de schedel verborgen liggen. De uitwendige
opening, de onderkaken en hun verbindingen met het middenoor wordt eerst besproken. Dan
volgt een beschrijving van een deel van de schedel, namelijk het tympano-periotisch
complex, dat het midden- en binnenoor omgeeft. Verder wordt ingegaan op het middenoor
met zijn typische aanpassingen en de anatomie van de gehoorbeentjes en tuba auditiva.
Tenslotte volgt het binnenoor met specifieke bijzonderheden van de cochlea en het
vestibulum.

2
INLEIDING
"Delphinum audire manifestum est; nam et cantu mulcentur et capiuntur attoniti sono;
quanam audiant, mirum."
Plinius, Naturalis Historia
Vertaling: 'Het is duidelijk dat dolfijnen kunnen horen. Want ze worden zowel gesust door gezang als
verbijsterd verleid door geluid. Hoe ze dan horen, is me een raadsel.'
De kennis over de anatomie van het gehoorapparaat van walvissen is erbarmelijk in
vergelijking met die van onze huisdieren. Toch is deze kennis cruciaal om te begrijpen welke
invloed de mens heeft (vb. door scheepvaart) op de walvispopulatie. Om dit te begrijpen,
moet er gestart worden aan de basis en dat is de anatomie. Hier in het bijzonder de anatomie
van het gehoorstelsel.
Doordat walvissen geëvolueerd zijn uit landzoogdieren, heeft het gehoorapparaat zich
evolutionair aangepast aan het leven onder water. Het uitwendig oor heeft mogelijks zijn
functie verloren en bij tandwalvissen zijn bijvoorbeeld de onderkaken een belangrijke rol gaan
spelen in het mechanisme van echolocatie (Norris, 1968)
Over de functie van het middenoor en elk van zijn onderdelen bestaat er nog steeds geen
consensus. Structuren zoals de gehoorbeentjes zijn wel aanwezig, maar is er is geen sluitend
bewijs over de functionaliteit ervan (Ketten, 1997).
Het binnenoor vertoont specifieke eigenschappen die gepaard gaan met het leven onder
water. De reductie van het vestibulum en geassocieerde zenuwvezels is significant, maar de
reden erachter nog onbekend. De cochlea vertoont specifieke verschillen tussen de
verschillende soorten, wat onder andere te zien is in de bouw van de basilaire membraan
(Wever et al., 1971b).
Horen is essentieel voor walvissen om te overleven. Enkele specifieke kenmerken van het
gehoor van walvissen zijn onder andere de echolocatie (van belang bij Odontoceti) waarbij
ultrasone klanken moeten waargenomen worden en bij Mysticeti de lange-
afstandscommunicatie (waarover nog veel discussies bestaan) waarbij specifieke infrasone
klanken worden geproduceerd. Aangezien men uit de anatomie niet kan afleiden welke
frequenties (ultra- en infrasoon) walvisachtigen (Cetacea) kunnen horen, gebruikt men als
maatstaf de frequenties van geproduceerde geluiden die bijgevolg slechts een schatting
geven (Ketten, 1997).
Een betere kijk op de anatomie van het gehoorapparaat leidt tot een betere interpretatie van
de functie ervan. Daarnaast is het essentiële kennis voor verder onderzoek in deze nog
mysterieuze tak van het dierenrijk.

3
1. Taxonomie
De naam ‘Cetacea’ stamt af van het Griekse woord Κῆτος, dat ‘zeemonster’ of ‘reusachtige
vis’ betekent (Aristoteles, Historia Animalium IV 241).
Omwille van het feit dat sommige walvisachtigen of Cetacea op levende prooien jagen en dat
men fossielen van hun voorouders heeft teruggevonden waarvan het gebit duidelijk carnivoor
is, werd reeds vroeg aanvaard dat walvissen waarschijnlijk afstammen van creodonte
(primitieve rovers) zoogdieren (Kellogg, 1936). Vermoed wordt dat de overgang van
evenhoevige landdieren naar volledig aquatische Cetacea ruwweg 13 miljoen jaar in beslag
heeft genomen, namelijk tussen ongeveer 50 en 37 miljoen jaar geleden (Bajpai et al., 2007).
De vroege Cetacea, die men Archaeoceti noemt, leefden in het Eoceen (45-30 miljoen jaar
geleden). Men heeft zelfs fossiele restanten gevonden die ongeveer 52 miljoen jaar oud
waren (Fordyce, 1994). Deze eerste zeezoogdieren zijn verdwenen rond het einde van het
Oligoceen (30-19 miljoen jaar geleden). De twee huidige suborden ontstonden in het vroege
Oligoceen, de tandwalvissen (Odontoceti) rond 32 miljoen jaar geleden en de baleinwalvissen
(Mysticeti) rond 34,5 miljoen jaar geleden. Andere wetenschappers (Thewissen en Williams,
2002) beweren dat de onderverdeling in die twee suborden plaatsvond in het late Eoceen
(ongeveer 37 miljoen jaar geleden). Los van de discussie over het exacte tijdstip bestaat er
momenteel wel een consensus dat Odontoceti en Mysticeti afstammen van een
gemeenschappelijke voorouder (Fordyce, 2001) en dat die voorouder ontsprong uit de
Archaeoceti (Fordyce, 2001 en 2008). Dit is in tegenstelling tot vroegere wetenschappelijke
strekkingen, toen algemeen werd aanvaard dat elke genoemde suborde een afzonderlijke
terrestrische oorsprong had (Reysenbach De Haan, 1936).
Taxonomie: Zie bijlage I
2. Invloed van omgeving op gehoor 2.1 Fysica
Vooraleer er kan ingegaan worden op de typische anatomie van het gehoorapparaat, dient
men eerst de fysische achtergrond te begrijpen, namelijk hoe geluid zich onder water anders
gedraagt dan in lucht. Hierin spelen drie fysische principes een belangrijke rol. Allereest zijn
de factoren die de geluidssnelheid beïnvloeden bepalend.
Geluidssnelheid: c = λ.f
Hierin is c geluidssnelheid (m/s)
λ golflengte van het geluid (m)
f frequentie van het geluid (Hz)
De geluidssnelheid is direct afhankelijk van de lokale densiteit van een medium (Ketten,
1998). Deze densiteit wordt in zeewater voornamelijk beïnvloed door de zoutconcentratie,
temperatuur en druk. In water bedraagt de geluidssnelheid gemiddeld 1530 m/s (in lucht:

4
ongeveer 340 m/s). Voor een gegeven frequentie plant het geluid zich onder water sneller
voort dan in de lucht en is dus de golflengte langer. Hierdoor zal het geluid zich tevens verder
voortplanten aangezien lagere golflengtes minder attenueren (Nummela et al., 2007).
Elk medium heeft een akoestische impedantie die voortplanting van geluid karakteriseert. De
impedantie is een maat voor de hinder die een geluidsgolf ondervindt wanneer deze overgaat
naar een ander medium (Rienstra & Hirschberg, 2004). Bij de overgang van een geluidsgolf
van één medium (vb. lucht in het middenoor) naar een ander (vb. vloeistof in het binnenoor)
geeft de ratio van de akoestische impedanties van beide media de hoeveel energie weer die
weerkaatst wordt. Hoe groter de ratio, hoe groter de weerkaatsing (Nummela et al., 2007).
De akoestische impedantie wordt voorgesteld door: Z = p/v = c.ρ
Hierin is Z de akoestisch impedantie (Pa.s/m)
p de geluidsdruk (Pa)
v de partikelsnelheid (m/s)
ρ de densiteit van het medium (kg/m3)
luchtdensiteit: 1,293 kg/m3 (op zeeniveau) waterdensiteit: 1000 kg/m3
c de geluidssnelheid (m/s)
De akoestische impedantie van lucht bedraagt Zlucht= 400 Pa.s/m en deze van water
Zwater=1500 Pa.s/m.
Hemilä et al. (2001) hebben vastgesteld dat de impedantie van de cochlea van tandwalvissen
vrij goed overeenstemt met deze van water. Hieruit volgt dat een geluidsgolf die het
middenoor binnentreedt weinig zal reflecteren.
Een derde belangrijke wet: I = p.v = p.[p/(c.ρ)] = p2/(c.ρ)
Hierin is I de geluidsintensiteit in een medium
p de effectieve geluidsdruk (Pa) van een medium
c de geluidssnelheid (m/s) in een medium
(c.ρ) de impedantie (cfr. supra) van dat medium
Om het geluid onder water even goed te horen als in lucht, moet Iwater = Ilucht. Indien
bovenstaande formule wordt omgezet, wordt pwater = 59,7 . plucht . Dit betekent dat voor een
gelijke geluidsintensiteit en dus eenzelfde prikkeling van het oor de geluidsdruk onder water
ongeveer 60 maal groter moet zijn (Ketten, 1997, 1998; Nummela, 2007).
Deze informatie indachtig, is het logisch dat er grote verschillen bestaan tussen de anatomie
van het gehoorapparaat bij de verschillende species afhankelijk van welke frequenties er
gedetecteerd moeten kunnen worden. Hierdoor stuiten we onmiddellijk op het probleem dat
het praktisch niet mogelijk is om te weten welke frequenties kunnen gedetecteerd worden
door alle verschillende soorten Cetacea.

5
Sales en Pye (1974) vermelden verscheidene experimentele studies, uitgevoerd om de
reikwijdte van het gehoorstelsel bij Odontoceti aan te tonen; hierbij rekende men altijd op een
fysieke reactie van een getraind dier op een gegeven geluidssignaal. De testresultaten bleken
consequent en Johnson (1966) concludeerde zo dat dolfijnen tot 150kHz kunnen horen.
De specifieke reikwijdte is voor vele Cetacea niet geweten, daarom gebruikt men piekspectra
van uitgezonden vocalisaties om walvissen verder in te delen in verschillende groepen.
Ketten (1997) vermeldt: “Mammalian vocalizations generally have peak spectra at or near the
frequency of best sensitivity for that species; therefore, spectral analyses of underwater
recordings of emitted sounds provide reasonable indirect estimates of cetacean hearing.”
Dit is toepasbaar voor studies rond communicatie binnen dezelfde species. Maar
daarenboven kunnen ze buiten die reikwijdte ook geluiden detecteren, essentieel voor
detectie van prooien, roofdieren en andere significante omgevingsgeluiden (Ketten, 2009).
2.2 Functionele classificatie
Allereerst worden Cetacea onderverdeeld in Odontoceti (tandwalvissen), die specifieke
geluiden produceren met een hoge frequentie (t.b.v. echolocatie), en Mysticeti
(baleinwalvissen), die laagfrequente geluiden kunnen produceren. Volgens Ketten (1984) kan
men Cetacea op basis van de geproduceerde piekspectra nog verder onderverdelen in
functionele groepen.
Er zijn twee ultrasone Odontocete groepen: Type I met piekcentra boven 100 kHz en type II
met piekspectra lager dan 80 kHz, elk met hun specifieke habitat en sociale interactie. Type I
vindt men terug aan kusten en in rivieren. Deze dieren leiden vaak een solitair leven of in
kleine groepen. Hun gehoorapparaat is aangepast aan hoge frequenties die gebruikt worden
bij echolocatie (Ketten 1998). Type II vindt men terug aan kusten maar ook verder in zee.
Deze vertoeven in grotere groepen en gebruiken geluid ter communicatie met dieren van
dezelfde soort en om relatief grote objecten op afstand te detecteren door gebruik te maken
van lagere ultrasone frequenties (40-70 kHz) (Ketten, 1998; Ketten en Wartzok, 1990).
Alle Mysticeti worden gegroepeerd onder het type M (Ketten, 1998) al zijn er mogelijks
verdere onderverdelingen te maken (Würsig en Clarck, 1993). Frequenties van vocalisaties
van baleinwalvissen bevinden in het sonische (20 Hz - 20 kHz) tot infrasone (<20 Hz),
afhankelijk van het soort vocalisatie (Ketten, 1997, 1998). Al is er heel weinig over geweten,
toch veronderstelt men dat Mysticeti geen gebruik maken van echolocatie (Ketten, 1992)

6
3. Anatomie
Het gehoorapparaat van zoogdieren kan worden ingedeeld in drie entiteiten.
Het uitwendig oor, dat geluid opvangt en geleidt naar het trommelvlies; het met lucht gevulde
middenoor, dat via gehoorbeentjes en membranen de trillingen versterkt en deze doorgeeft
aan het binnenoor; Dit laatste is met vocht gevuld en laat als bandfilter een bepaalde
frequentieband door (de daarbuiten gelegen frequenties worden geblokkeerd of niet
gecapteerd) om dan de mechanische trillingen te converteren naar elektrische signalen, die
naar de grote hersenen worden doorgestuurd (Ketten, 1998).
De anatomie van het gehoorapparaat van Cetacea is uniek omdat het eigenschappen bezit
die gelijkaardig zijn aan die van landzoogdieren, zoals bijvoorbeeld een met lucht gevuld
middenoor en een spiraalvormige cochlea, maar ook specifieke aanpassingen aan het leven
onder water heeft ondergaan (zie verder). Deze adaptaties zijn duidelijk verschillend bij de
drie functionele groepen (I, II, M) (Ketten, 1998).
3.1 Het uitwendig oor
Het uitwendig oor wordt onderverdeeld in de pinna of auricula (oorschelp), de concha
auriculae (het tegen de schedel aanliggende trechtervormige deel van de oorschelp) en de
meatus acusticus externus (de uitwendige gehoorgang) (Ketten, 2000). Pinnae (oorschelpen)
zijn afwezig bij alle Cetacea (Reysenbach de Haan, 1956; Ketten, 1998, 2000). De porus
acusticus externus (de uitwendige gehooropening) uit zich als een klein kuiltje in de huid, met
een diameter die zelfs bij de grootste Mysticeti slechts 3mm groot is (Ketten, 2000). Bij
Odontoceti is de uitwendige gehoorgang zeer smal en wordt wijder naar mediaal toe, waar
het blind eindigt (Reysenbach de Haan, 1956). Het bevat geen aantoonbare verbinding met
het trommelvlies of middenoor (Ketten, 1998, 2000). Bij Mysticeti is de diameter in het
algemeen minder dan drie mm (Ketten, 2000) en versmalt verder naar mediaal tot het lumen
verdwijnt en overgaat in een bindweefselstreng (Reysenbach de Haan, 1956). Verder naar
het middenoor toe verschijnt er opnieuw een lumen dat is opgevuld met een compacte
cerumen-plug (oorsmeer). Deze plug bestaat uit verschillende samengepakte,
gedesquameerde epitheellagen (Reysenbach de Haan, 1956; Ketten, 1998). Ze is trouwens
al aanwezig voor de geboorte en wordt groter naarmate het gehoorkanaal groeit
(Reysenbach de Haan, 1956). Er is onduidelijkheid over de functionele betekenis van enig
segment in dit gehele kanaal (Ketten, 1992, 1998, 2000). Aangezien de
geluidsdoorlaatbaarheid van het lichaam van walvisachtigen in de buurt komt van deze van
zeewater, hoeft geluid niet noodzakelijk via het gehoorkanaal binnen te dringen, maar zou het
langs alle kanten het binnenoor kunnen bereiken (Reysenbach de Haan, 1956). Het midden-
en binnenoor zijn evenwel omgeven door poreus bot dat luchtsinussen bevat (zie verder:
Tympano-periotisch complex) en zorgt voor akoestische isolatie (Reysenbach de Haan, 1960;
Branstetter, 2006; Nummela et al., 2007).

7
Geluidsperceptie van ultrasone geluiden (dus bij Odontoceti) verloopt via de onderkaak. De
mandibula bevat een mandibulair kanaal waarin onder andere vet, zenuwen en bloedvaten te
vinden zijn. Dit kanaal wordt snel heel smal naar rostraal toe, maar is prominent naar caudaal
toe. Daarenboven heeft het caudale deel van de onderkaak van Odontoceti enkele unieke
eigenschappen. De laterale zijde van de mandibula bevat lokaal een ovaalvormige regio die
zeer dunwandig is. Dit wordt het ‘pan bone’ genoemd. Net lateraal van die regio liggen zachte
weefsels (vnl. vet) die een ‘akoestisch raam’ vormen. Dit beduidt dat op die plaats
geluidssignalen de onderkaak kunnen binnendringen tot in het mandibulair kanaal. Verder
naar caudaal worden die signalen doorgegeven aan het tympano-periotisch complex en het
middenoor (Norris, 1968). Deze beschrijving werd later door Ketten (1997) geamendeerd.
Deze auteur vermeldt dat het mandibulair vetkanaal dienst doet als een kanaal met lage
weerstand waarin geluidsignalen zich voortplanten en dat het ‘pan bone’ dient als een
akoestisch raam dat de signalen doorgeeft aan het middenoor. De externe geluiden worden
bij deze theorie opgevangen door het rostrale deel van de onderkaak. Bovendien werd er via
MRI (Magnetic Resonance Imaging) een tweede trompetvormig vetkanaal opgemerkt dat zich
lateraal van het ‘pan bone’ bevindt. Dit duidt mogelijks op een gesegmenteerde
geluidscaptatie waarbij er afhankelijk van de frequentie een andere absorptieroute wordt
gevolgd (zie Fig.1). Dit zou het oriëntatievermogen ten goede komen.
Verscheidene auteurs beschrijven dat bruinvissen met het hoofd draaiende bewegingen
maken tijdens echolocatie (Norris, 1968). Dit zou een logisch gevolg kunnen zijn van
bovenstaande theorie. Bij Mysticeti vindt men geen gelijkaardige structuren terug in de
onderkaken (Ketten, 1997). Recent MRI-onderzoek van een dwergvinvis gaf aan dat een
groot, welgevormd vetlichaam zich lateraal, dorsaal en caudaal van de ramus mandibulae
bevindt, en bovendien ook contact maakt met het tympano-periotisch complex (Yamato et al,
2012). Er is vooralsnog geen bewijs dat dit een functionele rol heeft in het gehoorapparaat
van Mysticeti. Het caudomediale deel van de mandibula van Odontoceti is niet afgelijnd door
bot (Norris, 1968). Een andere (eventueel additieve) hypothese van geluidsperceptie stelt dat
geluid via de mondholte en de keel vervolgens via vet aan de mediale zijde van de mandibula
dringt en dan aan het middenoor wordt doorgegeven (Barroso, 2010; Barroso et al., 2012).

8
Fig. 1 Model van paden die geluid kan afleggen bij de dolfijn (uit Ketten, 1997). (Es: uitgaande
geluidssignalen; Is: binnenkomende geluidsignalen; vs: divertikel van de vestibulaire zak; ts: divertikel
van de tubulaire nasale zak; ps: divertikel van de premaxillaire nasale zak). Let hier voornamelijk op
binnenkomende signalen (Is). Frontaal invallende geluiden (mogelijks voor echolocatie) dringen via de
onderkaak door het mandibulair kanaal. Lateraal vormen de trompetvormige vetkwabben (lateral
channel), die eenzelfde densiteit hebben als het vet in het mandibulair kanaal, een toegang voor
invallende geluiden langs de achter- of zijkant van het dier (mogelijks voor lagere frequenties). Dit duidt
op een eventueel gesegmenteerd gehoor.
3.2 Het tympano-periotisch complex
Het tympano-periotisch complex (T-P complex) is een beenderige structuur, samengesteld uit
het os temporale (slaapbeen) en het os petrosum (rotsbeen), die het midden- en binnenoor
omgeeft (Ketten, 1997; Nummela et al., 1999)(Fig.2). Het tympanisch gedeelte van het os
temporale, dat zich het meest lateraal bevindt, heeft contact met de omliggende zachte
(vet)weefsels waarin geluidsgolven worden opgevangen en zou aldus mee vibreren
(Nummela et al., 1999a). Dit deel van het bot, dat zo dun is dat het geluidsignalen zeer
efficiënt kan overdragen, noemt men de tympanische plaat en is het functionele equivalent
van het trommelvlies (= tympanische membraan) van landzoogdieren (Fleischer, 1978;
Nummela et al., 1999a, 1999b). De volumeverhouding tussen het tympanisch en periotisch
bot is bij Odontoceti ongeveer gelijk, terwijl bij Mysticeti het eerstgenoemde bijna het dubbele
volume inneemt (Ketten, 1993). Bij de Odontoceti is de bulla hol en wordt afgelijnd door
compact bot van het tympanische gedeelte van het os temporale. Het is volledig geïsoleerd
van de rest van schedel in een peribullaire holte en wordt ter plaatse gehouden door middel
van enkele ophangbanden (Reysenbach de Haan, 1956; Ketten en Wartzok, 1990; Ketten,
1998). Bij Mysticeti heeft het T-P complex wel contact met de rest van de schedel via de

9
processus mastoideus (Nummela et al., 1999). Bij alle Cetacea is het T-P complex dorsaal en
mediaal omgeven door lucht bevattende (i.e. schuim bevattende) sinussen die zorgen voor
akoestische isolatie (Reysenbach de Haan, 1960; Ketten en Wartzok, 1990; Ketten 1997;
Nummela et al., 2007). Deze sinussen zijn overigens verbonden met de luchtwegen waardoor
ze eveneens een functie zouden hebben in de drukregeling rond het complex (Reysenbach
de Haan, 1960). Doordat het T-P complex zich in een uitgebreide peribullaire holte bevindt
krijgt de walvisschedel een typische paddenstoelvorm (Ketten, 1997).
3.3 Het middenoor
Over de verschillende functies en de specifieke werking van het middenoor van Cetacea
bestaat nog geen consensus (Reysenbach de Haan, 1956; Ketten, 1997). Er is
overeenstemming over het feit dat de gehoorbeentjes (malleus (hamer), incus (aambeeld),
stapes (stijgbeugel)) een osseuze ketting vormen die trillingen, veroorzaakt door
binnenkomende geluidsgolven, overbrengen van de tympanische plaat van de mandibula
naar het fenestra vestibuli (= ovale venster) van het binnenoor (Nummela et al., 1999a,
1999b; Kinkel et al., 2001; Nummela et al., 2007). Door deze overdracht worden de trillingen
versneld zonder dat de druk verhoogt (Nummela et al., 1999a). Dit proces gaat omgekeerd bij
landzoogdieren waar de snelheid wordt gereduceerd en de druk verhoogd (Nummela et al.,
1999a). Gehoorbeentjes van alle Odontoceti zijn gemiddeld zwaar en solide (Reysenbach de
Haan, 1956; Norris, 1968). Hun grootte, hardheid en vorm vertonen wel veel variatie tussen
verschillende species (Reysenbach de Haan, 1956). Er is een synostose tussen de malleus
en de tympanic plate via een beenderige kam, de processus gracilis (Reysenbach de Haan,
1956, 1960; Nummela et al., 1999a, 1999b, 2007).
Het middenoor van Cetacea vertoont aanpassingen aan de snelle en hevige
drukveranderingen die optreden bij duiken (Ketten, 1997) en aan het waarnemen van geluid
onder water (Reysenbach de Haan, 1960). Voor eenzelfde geluidsintensiteit moet onder
water de geluidsdruk ongeveer 60 maal groter zijn (cfr. Fysica 2.1). Het middenoor is hieraan
aangepast en is dus zwaar en compact, net zoals het beenweefsel van het hele
tympanopetromastoid is (= os petrotympanicum (Reysenbach de Haan, 1956)) (Reysenbach
de Haan, 1960). Het tympanisch bot heeft lateraal een dunne wand, zoals bij alle zoogdieren,
maar bevat bij Cetacea mediaal een dikke, uitpuilende structuur, het involucrum (Nummela et
al., 2007).
Ook de tuba auditiva van Cetacea heeft typische aanpassingen ondergaan. Zoals bij alle
zoogdieren vormt deze buis een verbinding tussen de farynx en het cavum tympani van het
middenoor. Bij Mysticeti is de tuba auditiva verbonden met de sinus pterygoideus
(Reysenbach de Haan, 1956). De buis is breed en haar wand is stevig, waardoor ze onder
drukveranderingen open blijft en het oor zo beschermt tegen barotrauma (Ketten, 1997). Ze
bevat bovendien mucosale kleppen die openen naar het cavum tympani toe (Reysenbach de
Haan, 1956).
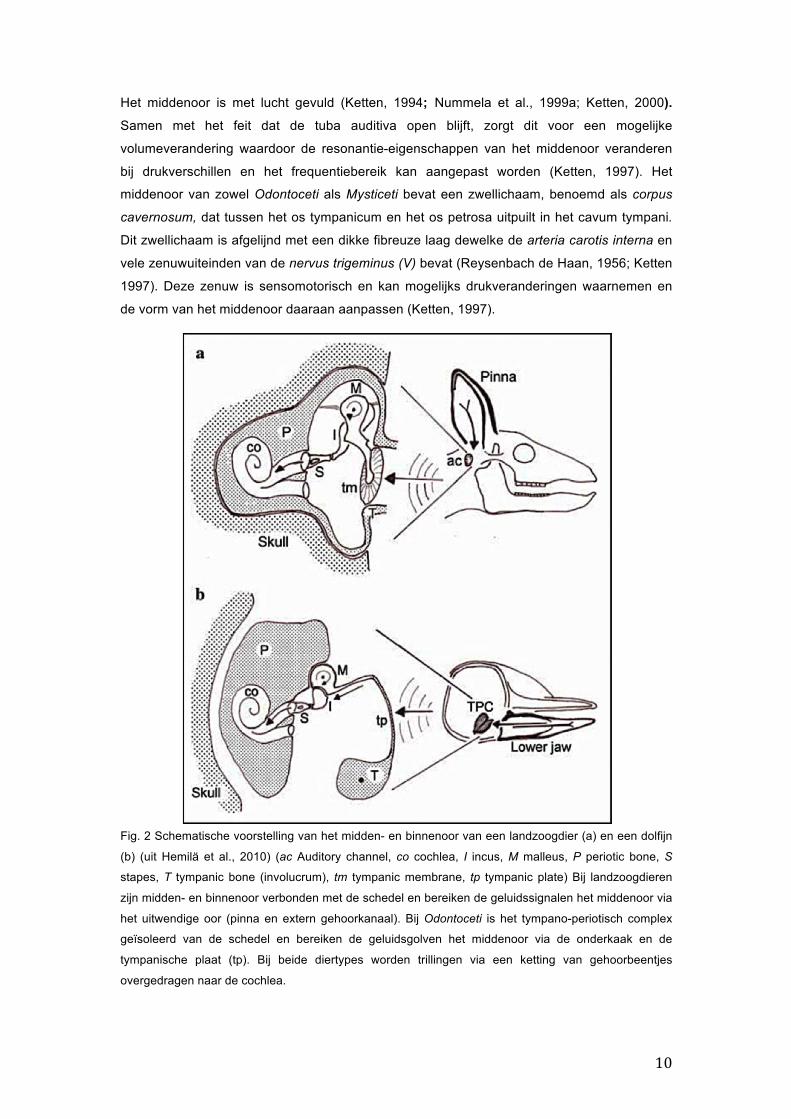
10
Het middenoor is met lucht gevuld (Ketten, 1994; Nummela et al., 1999a; Ketten, 2000).
Samen met het feit dat de tuba auditiva open blijft, zorgt dit voor een mogelijke
volumeverandering waardoor de resonantie-eigenschappen van het middenoor veranderen
bij drukverschillen en het frequentiebereik kan aangepast worden (Ketten, 1997). Het
middenoor van zowel Odontoceti als Mysticeti bevat een zwellichaam, benoemd als corpus
cavernosum, dat tussen het os tympanicum en het os petrosa uitpuilt in het cavum tympani.
Dit zwellichaam is afgelijnd met een dikke fibreuze laag dewelke de arteria carotis interna en
vele zenuwuiteinden van de nervus trigeminus (V) bevat (Reysenbach de Haan, 1956; Ketten
1997). Deze zenuw is sensomotorisch en kan mogelijks drukveranderingen waarnemen en
de vorm van het middenoor daaraan aanpassen (Ketten, 1997).
Fig. 2 Schematische voorstelling van het midden- en binnenoor van een landzoogdier (a) en een dolfijn
(b) (uit Hemilä et al., 2010) (ac Auditory channel, co cochlea, I incus, M malleus, P periotic bone, S
stapes, T tympanic bone (involucrum), tm tympanic membrane, tp tympanic plate) Bij landzoogdieren
zijn midden- en binnenoor verbonden met de schedel en bereiken de geluidssignalen het middenoor via
het uitwendige oor (pinna en extern gehoorkanaal). Bij Odontoceti is het tympano-periotisch complex
geïsoleerd van de schedel en bereiken de geluidsgolven het middenoor via de onderkaak en de
tympanische plaat (tp). Bij beide diertypes worden trillingen via een ketting van gehoorbeentjes
overgedragen naar de cochlea.
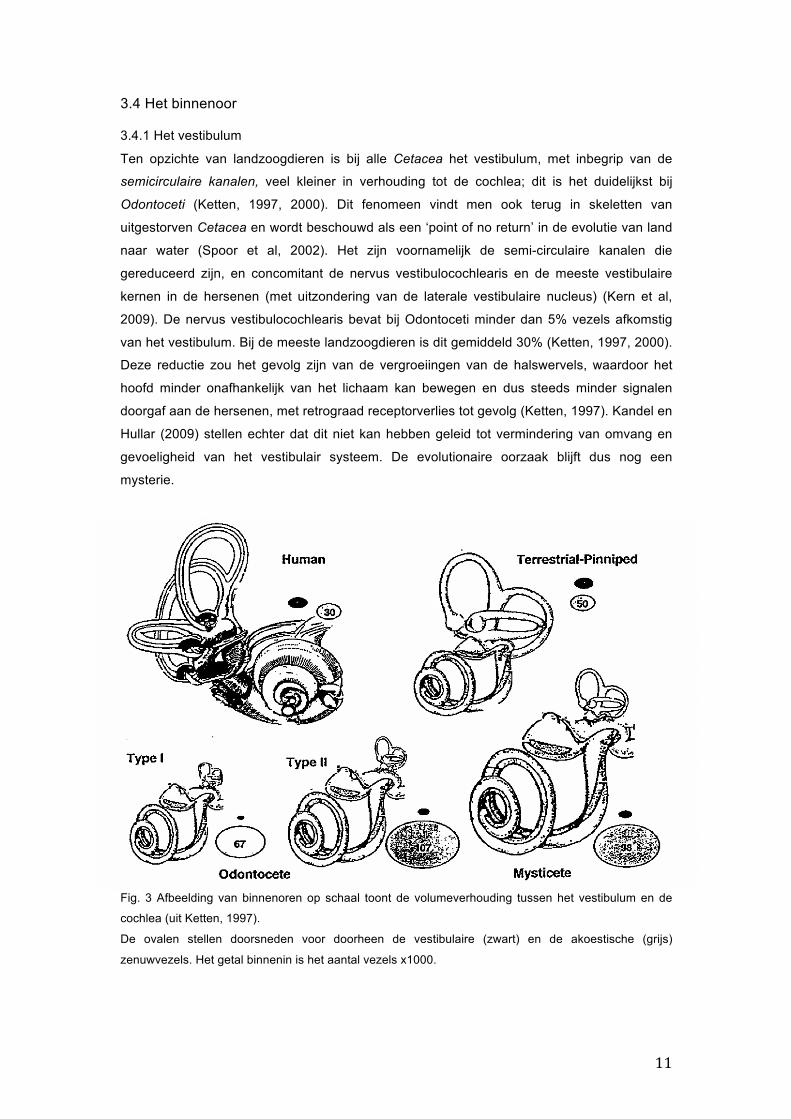
11
3.4 Het binnenoor
3.4.1 Het vestibulum
Ten opzichte van landzoogdieren is bij alle Cetacea het vestibulum, met inbegrip van de
semicirculaire kanalen, veel kleiner in verhouding tot de cochlea; dit is het duidelijkst bij
Odontoceti (Ketten, 1997, 2000). Dit fenomeen vindt men ook terug in skeletten van
uitgestorven Cetacea en wordt beschouwd als een ‘point of no return’ in de evolutie van land
naar water (Spoor et al, 2002). Het zijn voornamelijk de semi-circulaire kanalen die
gereduceerd zijn, en concomitant de nervus vestibulocochlearis en de meeste vestibulaire
kernen in de hersenen (met uitzondering van de laterale vestibulaire nucleus) (Kern et al,
2009). De nervus vestibulocochlearis bevat bij Odontoceti minder dan 5% vezels afkomstig
van het vestibulum. Bij de meeste landzoogdieren is dit gemiddeld 30% (Ketten, 1997, 2000).
Deze reductie zou het gevolg zijn van de vergroeiingen van de halswervels, waardoor het
hoofd minder onafhankelijk van het lichaam kan bewegen en dus steeds minder signalen
doorgaf aan de hersenen, met retrograad receptorverlies tot gevolg (Ketten, 1997). Kandel en
Hullar (2009) stellen echter dat dit niet kan hebben geleid tot vermindering van omvang en
gevoeligheid van het vestibulair systeem. De evolutionaire oorzaak blijft dus nog een
mysterie.
Fig. 3 Afbeelding van binnenoren op schaal toont de volumeverhouding tussen het vestibulum en de
cochlea (uit Ketten, 1997).
De ovalen stellen doorsneden voor doorheen de vestibulaire (zwart) en de akoestische (grijs)
zenuwvezels. Het getal binnenin is het aantal vezels x1000.

12
3.4.2 De cochlea
Er is weinig informatie beschikbaar over de anatomie van de cochlea bij Mysticeti en de
informatie die voorhanden is, toont weinig specifieke eigenschappen aan (Ketten, 1992).
Verder wordt in deze literatuurstudie enkel gesproken over de cochlea bij Odontoceti, tenzij er
expliciet vermeld wordt dat het over Mysticeti gaat. Het slakkenhuis van Cetacea is
macroscopisch gelijkaardig aan dat van andere zoogdieren (Ketten, 1992). De grootte van de
cochlea is gecorreleerd met de grootte van het dier (Ketten, 1992) De cochlea wordt inwendig
onderverdeeld in de scala tympani (ventraal), de scala media of ductus cochlearis en de
scala vestibuli (dorsaal). Deze drie kanalen lopen spiraalvormig rond de modiolus, de
beenderige kam waarin de nervus cochlearis (als vertakking van de nervus
vestibulocochlearis, de VIIIe kopzenuw) loopt (Ketten, 1997; Ketten en Wartzok, 1990). Deze
zenuw zendt verdere vertakkingen naar ganglia, die ofwel gedeeltelijk omgeven zijn door het
botweefsel van de modiolus, ofwel bijna volledig vrij liggen in de scala tympani (Wever et al.,
1971a). De diameter van het canalis spiralis cochleae neemt af naar de apex cochleae toe
(Ketten en Wartzok, 1990).
Fig 4. Tekening van een doorsnede doorheen het canalis spiralis cochleae (uit Dellman en Brown,
1981)
Er zijn twee opvallende anatomische kenmerken die invloed hebben op de
resonantiekarakteristieken en die ervoor zorgen dat hoge frequenties kunnen waargenomen
worden, namelijk de lamina spiralis ossea (een binnenste en buitenste) en de membraneuze
basilaire membraan (Ketten, 1992). Tussen de verschillende functionele types Odontoceti
(type I en II) zien we hierin verschillen die passen bij de levensstijl en de bijhorende meer
voorkomende geluidsfrequenties (Ketten, 1992). De binnenste lamina spiralis ossea is een
beenderige kam, uitgaande van de modiolus tot ongeveer halverwege de ductus cochlearis.

13
De basilaire membraan hecht hierop vast en vormt de scheiding tussen de scala tympani en
de scala media. Hierop rust het organum spirale (orgaan van Corti), waarin geluidstrillingen
worden omgezet naar zenuwsignalen (Ketten, 1997). Naar de buitenzijde van de cochlea toe
gaat de basilaire membraan over in een buitenste lamina spiralis ossea, die een verlenging
van het beenderig cochleaire kapsel is (Wever et al., 1971b; Ketten en Wartzok, 1990). De
verbinding tussen de basilaire membraan en buitenste lamina spiralis ossea vindt men in de
eerste 8 tot 10 mm van de cochlea, verder naar de apex toe verdwijnt die buitenste lamina
spiralis ossea en hecht de basilaire membraan rechtstreeks op het spiraal ligament (crista
spiralis)(Wever et al., 1971b; Ketten en Wartzok, 1990). De basilaire membraan verschilt in
dikte en breedte naargelang de positie van basis tot apex (zie fig. 4). In de smalle basale
regio is ze kort en dik, wat ideaal is voor hoge geluidsfrequenties, en naar de apex toe rekt ze
uit, wordt dunner en is dus goed om lage frequenties te detecteren (Ketten en Wartzok,
1990). Een studie bij 4 tuimelaars (Tursiops truncatus) toont dat de breedte van de basilaire
membraan aan de basis van de cochlea ongeveer 25 µm meet en in het apicale einde van de
cochlea ongeveer 350 µm breed is, en dus 14 keer breder wordt dan aan de basis. Bij de
mens is dit ongeveer 6,25 keer, waar de membraan aan de basis minstens 80 µm meet.
(Wever et al., 1971b). Bij de Mysticeti varieert de basilaire membraanbreedte van 100 µm aan
de basis tot meer dan 1500 µm in de apex (Ketten, 1997). Ook de binnenste lamina spiralis
ossea is breed aan de basis en wordt smal naar de apex toe (Wever et al., 1971b).
Fig. 5 Schematische voorstelling van de afmetingen van basilaire membraan van basis tot apex (uit
Ketten, 1997)
Op histologische preparaten van dolfijnen toont het orgaan van Corti zich als een stevige
structuur met relatief grote en dicht opeengepakte steuncellen. Deze steuncellen zijn veel
groter dan bij andere zoogdieren en zijn typisch voor Odontoceti (Wever et al, 1971a). De

14
stria vascularis is uitermate dens en het ligamentum spirale is opgebouwd uit strak geweven
collageenvezels (Ketten, 1997) Deze verschillende onderdelen van de cochlea bij Odontoceti
geven het geheel een robuust karakter dat aangepast is aan ultrasone geluidsreceptie.
(Wever et al, 1971a).
Gegevens over de cochlea van Mysticeti zijn schaars en komen hoofdzakelijk uit de dissectie
van aangespoelde dieren met redelijk lange post mortem-tijden. De cochlea vertoonde in
deze studies niet de cellulaire ontwikkeling zoals bij Odontoceti. Het spiraal ligament, de stria
vascularis en de steuncellen vertoonden hier meer gelijkenissen met die van de mens. Deze
informatie blijkt toch relevant, aangezien het binnenoor van Odontoceti, na gelijkaardige post
mortem-tijd, zijn cellulaire kenmerken blijft bewaren en zelfs bij heftig verval niet op het oor
van Mysticeti gelijkt (Ketten, 1992, 1997).
Berekende schattingen van het aantal (binnenste en buitenste) haarcellen in de cochlea van
de dolfijn (Tursiops truncatus) geven een gelijkaardig aantal (+/- 15.000) als bij de mens
(Wever et al., 1971c). Men vindt wel een significant verschil in het totaal aantal
ganglioncellen. Bij de dolfijn telt men er ongeveer drie maal zoveel als bij de mens
(gemiddeld 30.500 bij mens) (Wever et al., 1971c). Wanneer men de ratio’s van
ganglioncellen ten opzichte van haarcellen vergelijkt, is dit bij de mens ongeveer 2 op 1; bij
de dolfijn is dit meer dan 5 op 1 (Wever et al., 1971c). Recentere data geven specifiekere
ratio’s weer: 7,3 op 1 bij type I Odontoceti, 6,5 op 1 bij type II Odontoceti en 5,7 op 1 voor
Mysticeti (Ketten, 1997). Uit deze cellulaire verdeling kan men twee mogelijke redeneringen
afleiden: hoge geluidsfrequenties vereisen meer zenuwgeleiding of de dolfijn ontvangt in het
algemeen meer informatie van de cochlea dan de mens (Wever et al., 1971c). Dit laatste past
in het plaatje van het gesofisticeerde mechanisme van echolocatie, maar verklaart niet de
gelijkaardige innervatie bij Mysticeti. Men veronderstelt dus dat ofwel de fysica van ‘horen
onder water’ deze verdeling kan verklaren ofwel dat Mysticeti even complexe informatie halen
uit lage en infrasone geluiden en eventueel zelfs gebruik maken van infrasone echolocatie
(Ketten, 1997).

15
Fig. 6 Detailtekening van een doorsnede door de ductus cochlearis van een tuimelaar (Tursiops truncatus) (uit Wever et al., 1971a)

16
BESPREKING
Het gehoor van Cetacea blijft in heel veel aspecten een mysterie. Algemeen kan gesteld
worden dat de anatomie van het gehoorapparaat van Odontoceti door een weinig aantal
onderzoekers vrij goed is beschreven, maar dat over de specifieke functies van de
verschillende onderdelen nog veel twijfel en discussie bestaat. Het gehoorapparaat van
Mysticeti is veel minder onderzocht, en roept nog veel meer vragen op.
In de literatuur is men voorzichtig wat betreft de functionaliteit van vele structuren. Daarom
beperken veel studies zich voornamelijk tot het beschrijven van anatomische bevindingen
zonder er functies aan te koppelen. De porus acusticus externus bijvoorbeeld is bij alle
Cetacea aanwezig, maar over diens functie en die van het aansluitende gehoorkanaal
bestaat nog discussie (Ketten, 1992, 1998, 2000). Het is ook mogelijk dat deze structuur
slechts een rol speelt bij enkele soorten. Hierover is weinig geweten. Maar met de
ontwikkeling van moderne beeldvormingstechnieken zou men hier inzicht in kunnen
verwerven.
Over andere structuren en bijhorende functies heeft men wel een consensus bereikt. Zo is
men het er over eens dat de mandibulae van Odontoceti wel degelijk een rol spelen bij het
opvangen van ultrasone geluiden (>20kHz) (Norris, 1968; Ketten, 1997; Barroso 2010, 2012).
Over het exacte mechanisme is er echter ook hier discussie, die al meer dan 50 jaar aan de
gang is. Meerdere theorieën zijn voorgesteld. Ketten (1994, 1997) vermeldt de mogelijkheid
van een gesegmenteerd gehoor, waarbij de weerkaatste echolocatie-geluiden via de
onderkaken worden opgevangen, maar andere geluiden (met een lagere frequentie) via
laterale vetkanalen zouden worden waargenomen. Barroso (2010, 2012) opteert een andere
theorie waarbij geluiden via de mondholte zouden worden opgenomen. Er is nog geen
sluitend bewijs gevonden hoe het geheel precies zou functioneren.
Ook over het tympano-periotisch complex bestaat nog veel onduidelijkheid. In deze
literatuurstudie werden drie bronnen aangehaald waarin wordt beschreven dat de bullae los
van de schedel staan, omgeven door peribullaire holtes waarin ze ophangen met
weefselbanden. Specifiekere details over hoe dit precies past in het grotere plaatje, en dus
vooral het waarom achter dit unieke kenmerk wordt in deze bronnen slechts vaag
beschreven. Er zijn vermeldingen dat deze holtes bestaan uit schuim bevattende sinussen
(Reysenbach de Haan, 1960; Ketten en Wartzok, 1990; Ketten 1997; Nummela et al., 2007),
maar hierover is ook meer onderzoek gewenst.
Hetzelfde geldt voor het middenoor. Over de verschillende functies en de specifieke werking
van het middenoor van Cetacea bestaat nog geen consensus (Reysenbach de Haan; Ketten,
1997).

17
De anatomie van het binnenoor is vrij goed beschreven bij Odontoceti. De anatomische
verschillen bij de verschillende soorten zijn duidelijk aanwezig en bovendien in correlatie met
de geluiden waarmee die soorten voornamelijk te maken krijgen (Wever et al., 1971b; Ketten
en Wartzok, 1990). Bij Mysticeti zijn de gegevens dan weer schaars.
Aangezien geluid een essentiële rol speelt bij onder andere de communicatie, de oriëntering,
het detecteren van prooien, de voortplanting en vele andere aspecten van een walvisleven
waarvan we mogelijks geen idee hebben, is het belangrijk om te begrijpen hoe dit precies
werkt opdat we meer inzicht zouden krijgen in hun leefwereld om zo te streven naar een
harmonischere relatie tussen walvis en mens.

18
BIJLAGEN Bijlage I: Taxonomie (Mead en Brownell, 2005): Klasse Mammalia
Infaklasse Placentalia
Superorde Laurasiotheria
Cetartiodactyla
Orde Cetacea: Walvissen
Suborde Archaeoceti: uitgestorven walvisachtigen
Suborde Mysticeti: Baleinwalvissen
Familie Balaenidae (Echte walvissen)
Balaenae (één species: Groenlandse walvis)
en Eubalaena (Noordkapers: drie species)
Familie Balaenopterdidae (Vinvissen)
Balaenoptera (Echte vinvissen: zes species)
Megaoptera (één species: Bultrug)
Familie Eschrichtiidae (Grijze walvissen)
(één species: Grijze walvis)
Familie Neobalaenidae
hier vermeld als aparte familie, maar wordt soms
geklasseerd als geslacht binnen de familie Balaenidae
(één species: Dwergwalvis)
Suborde Odontoceti: Tandwalvissen
Familie Delphinidae (Dolfijnachtigen)
Omvat o.a. Gewone dolfijn, Langsnuit gewone dolfijn,
Dwerggriend, Griend, Grijze dolfijn, Sarawakdolfijn, Orka of
Zwaardwalvis, Irawaddydolfijn, Witlipdolfijn en Tuimelaar
Familie Monodontidae (Gordeldolfijnen)
Omvat Beluga en Narwal
Familie Phocoenidae (Bruinvissen)
Omvat o.a. Gewone bruinvis, Californische bruinvis,
Brilbruinvis, Bruinvis van Burmeister, Brilbruinvis, Chinese
bruinvis en Dallbruinvis
Familie Physeteridae (Potvissen)
Omvat Potvis, Dwergpotvis en Kleinste potvis
Familie Platanistidae (Indische rivierdolfijnen)
Omvat Gangesdolfijn en Indusdolfijn
Familie Iniidae (Rivierdolfijnen)

19
Omvat o.a. Orinocodolfijn, Chinese vlagdolfijn en La
Platadolfijn
Familie Ziphiidae (Spitssnuitdolfijnen)
Omvat o.a. Gewone spitssnuitdolfijn, Spitssnuitdolfijn van
Gervais, Spitssnuitdolfijn van True, Butskop, Zwarte Dolfijn,
Dolfijn van Cuvier, Pacifische spitssnuitdolfijn, Japanse
spitssnuitdolfijn en vele andere.
(bron Nederlandse namen: Shirihai en Jarret, 2008)

20
REFERENTIELIJST 1. Aristoteles, Historia Animalium IV 241. Bron: The Works of Aristotle, vertaald naar het
Engels door D’Arcy Wentworth Thompson, 1910
2. Bajpai, S., Nummela, S., Thewissen J.G.M., Hussain, T. and Kumar K. (2007). Sound
transmission in archaic and modern whales: Anatomical adaptations for underwater
hearing. Anatomical Record, 290, 716–733
3. Branstetter B.K. & Mercado E. (2006). Sound Localization by Cetaceans. International
Journal of Comparative Psychology, 19, 26-61
4. Barroso C. (2010). Shape Analysis of Odontocete Mandibles: Functional and Evolutionary
Implications. Thesis Master of Science in Biology, San Diego State University, San Diego,
p. 28-38
5. Barroso C., Cranford T.W. & Berta A. (2012). Shape Analysis of Odontocete Mandibles:
Functional and Evolutionary Implications. Journal of Morphology, 273, 1021-1030
6. Dellmann H.D. & Brown E.M. (1981). Handbook of Veterinary Histology, 2nd edition,
Philadelphia, p. 436
7. Fleischer G. (1978). Evolutionary Principles of the Mammalian Middle Ear. Adv. Anat.
Embryol. Cell Biol., 55, 1-70
8. Fordyce R.E. & Barnes L.G. (1994). The Evolutionary History of Whales and Dolphins,
Annual Review of Earth and Planetary Sciences, 22, 419-455
9. Fordyce R.E., de Muizon C. (2001). Evolutionary history of the cetaceans: a review. In:
Mazin & de Buffrénil 2001, Secondary Adaptation of Tetrapods to Life in Water, p. 169-
234
10. Fordyce R.E. (2008). Cetacean Evolution, Cetacean Fossil Record, in Perrin W.F.,
Würsig B., Thewissen J.G.M. (Editors) Encyclopedia of Marine Mammals, second edition,
Academic Press, p. 201-215
11. Hemilä S., Nummela S. & Reuter T. (2001). Modeling whale audiograms: effects of bone
mass on high-frequency hearing. Hearing Research, 151, 221-226
12. Hemilä S., Nummela S. & Reuter T. (2010). Anatomy and physics of the exceptional
sensitivity of dolphin hearing (Odontoceti: Cetacea), Journal of Comparative Physiology
A, 196, 165-179
13. Johnson C.S. (1966). Auditory tresholds of bottlenose poroise (Tursiops truncatus,
Montagu), U.S. Naval Ordnance Test Station Technical Publication No. 4178, p.1-28.
Bron: Sales en Pye (1974)
14. Kandel B.M. & Hullar T.E. (2009). The relationship of head movements to semicircular
canal size in cetaceans. Journal of Experimental Biology, 213, 1175-1181
15. Kellogg R. (1936). A review of the Archaeoceti. Carnegie Institution of Washington,
482,1–366

21
16. Kern A., Seidel K. & Oelschläger H.H.A. (2009). The Central Vestibular Complex in
Dolphins and Humans: Functional Implications of Deiters’ Nucleus. Brain Behavior Evol,
73, 102-110
17. Ketten D.R. (1984). Correlations of morphology with frequency for odontocete cochlea:
Systematics and topology. Ph.D. dissertation, The Johns Hopkins University, Baltimore,
Md, 335pp. Vermeld in: Ketten D.R. (1997), Structure and Function in Whale Ears
18. Ketten D.R. & Wartzok D. (1990). Three-Dimensional Reconstructions of the Dolphin Ear.
In: Thomas J. & Kastelein R. (Editors) Sensory Abilities of Cetaceans, New York, p. 81-
105
19. Ketten D.R. (1992). The Marine Mammal Ear: Specializations for Aquatic Audition and
Echolocation. In: The Evolutionary Biology of Hearing, Webster D.B., Fay R.R., and
Popper A. N. (editors), Springer-Verslag, p. 717-750.
20. Ketten D.R. (1994). Functional Analyses of Whale Ears: Adaptations for Underwater
Hearing. I.E.E.E Proceedings in Underwater Acoustics, 1, 264-270
21. Ketten D.R. (1997). Structure and Function in Whale Ears. The International Journal of
Animal Sound and its Recording, 8, 103-135
22. Ketten D.R. (1998). Marine Mammal Auditory Systems: a Summary of Audiometric and
Marine Mammal Anatomical Data and It’s Implications for Underwater Acoustic Impacts,
Boston, 75 pp.
23. Ketten D.R. (2000). Cetacean Ears. In: Au W.W.L., Popper A.N. & Fay R.R. (editors)
Hearing by Whales and Dolphins, Springer-Verslag, New York, p.44-108
24. Ketten D.R. (2009). Marine Mammal Auditory Systems: a Summary of Audiometric and
Marine Mammal Anatomical Data and its Implications for Underwater Acoustic Impacts,
Boston, 23 pp.
25. Kinkel, M.D., Thewissen J.G.M. & Oelschläger H.A. (2001). Rotation of Middle Ear
Ossicles During Cetacean Development. Journal of Morphology, 249, 126–131
26. Mead J.G. and Brownell, Robert L. Jr. (2005). Order Cetacea. In: Wilson, Don E. and
Reeder, D. M., Mammal Species of the World, Third Edition. Baltimore, MD, Johns
Hopkins University Press, p. 723-743
27. Nummela S., Wägar T., Hemilä S. & Reuter T. (1999a). Scaling of the cetacean middle
ear. Hearing Research, 133, 71-81
28. Nummela S., Wägar T., Hemilä S. & Reuter T. (1999b). The anatomy of the killer whale
middle ear (Orcinus Orca). Hearing Research, 133, 61-70
29. Nummela S., Thewissen J.G.M., Bajpaj S., Hussain T. & Kumar K. (2007). Sound
Transmission in Archaic and Modern Whales: Anatomical Adaptations for Underwater
Hearing. The Anatomical Record, 290, 716-733
30. Norris, K. S. (1968). The Evolution of Acoustic Mechanisms in Odontocete Cetaceans. In:
Evolution and Environment (E. T. Drake, ed). Yale University Press; New Haven. pp. 297-
324

22
31. Reysenbach de Haan F.W. (1956). De Ceti Auditu (Hearing in Whales), Acta Oto-
laryngologica: Supplementum, 134, 1-114
32. Reysenbach de Haan F.W. (1960). Some Aspects of Mammalian Hearing under Water,
Proc. R. Soc., Londen, 152, 54-62
33. Rienstra S.W. & Hirschberg A. (2012). An introduction to Acoustics, Eindhoven, p. 36
34. Sales, G. & Pye, D. (1974). Ultrasonic Communication by Animals. John Wiley and Sons,
New York, p. 202-223
35. Shirihai H. en Jarret B. (2008). Gids van alle zeezoogdieren. Tirion Natuur, Baarn, pp. 21-
246
36. Spoor F., Bajpai S., Hussain S.T., Kumar K. & Thewissen J.G.M. (2002). Vestibular
Evidence for the Evolution of Aquatic Behaviour in Early Cetaceans. Nature, 417, 163-
166
37. Thewissen J.G.M. & Williams E. M. (2002). The Early Radiations of Cetacea (Mammalia):
Evolutionary Pattern and Developmental Correlations, Annual Review of Ecology and
Systematics, 33, 73-90
38. Wever E.G., McCormick J.G., Palin J. & Ridgway S.H. (1971a). The Cochlea of the
Dolphin, Tursiops truncatus: General Morphology. Proc. Nat. Acad. Sci. USA, 68, n°10,
2381-2385
39. Wever E.G., McCormick J.G., Palin J. & Ridgway S.H. (1971b). The Cochlea of the
Dolphin, Tursiops truncatus: The Basilar Membrane. Proc. Nat. Acad. Sci. USA, 68, n°11,
2706-2711
40. Wever E.G., McCormick J.G., Palin J. & Ridgway S.H. (1971c). The Cochlea of the
Dolphin, Tursiops truncatus: Hair Cells and Ganglion Cells. Proc. Nat. Acad. Sci. USA,
68, n°12, 2908-2912
41. Würsig B. & Clark. C. (1993). Behavior in The Bowhead. Society for Marine Mammalogy,
spec. publ., 2, 157-200. Vermeld in: Ketten D.R. (1997) Structure and Function in Whale
Ears. The International Journal of Animal Sound and its Recording, 8, 103-135
42. Yamato M., Ketten D.R., Arruda J., Cramer S. & Moore K. (2012). The Auditory Anatomy
of the Minke Whale (Balaenoptera acutorostrata): A Potential Fatty Sound Reception
Pathway in a Baleen Whale. The Anatomical Record, 295, 991–998


















