
De Moleculaire Diagnose Van Brugada Syndroom.pdf;Jsessionid=D4FDE564B1661307E395F66E278A55F6
86
De moleculaire diagnose van Brugada syndroom Peirsman Liszl Academiejaar 2009-2010 Dr. Sonia Van Dooren Afstudeerwerk ingediend voor het behalen van het diploma Bachelor in de Biomedische laboratoriumtechnologie – afstudeerrichting Farmaceutische en biologische laboratoriumtechnologie
-
Upload
stef-straf -
Category
Documents
-
view
139 -
download
6
Transcript of De Moleculaire Diagnose Van Brugada Syndroom.pdf;Jsessionid=D4FDE564B1661307E395F66E278A55F6

De moleculaire diagnose van Brugada
syndroom
Peirsman Liszl
Academiejaar 2009-2010
Dr. Sonia Van Dooren
Afstudeerwerk ingediend voor het behalen van het diploma Bachelor
in de Biomedische laboratoriumtechnologie – afstudeerrichting
Farmaceutische en biologische laboratoriumtechnologie


De moleculaire diagnose van Brugada
syndroom
Peirsman Liszl
Academiejaar 2009-2010
Dr. Sonia Van Dooren
UZ Brussel – Centrum voor Medische GeneticaCampus JetteLaarbeeklaan 1011090 Brussel

Dankwoord
Dit afstudeerwerk is er alleen maar gekomen dankzij de steun en hulp van vele mensen!
Hieronder een paar woordjes om iedereen te bedanken.
Het was een hele ervaring om gedurende vier maanden mee te draaien in een diagnostisch
labo. Het opstarten van het onderzoek, het onder de knie krijgen van de technieken en de
theoretische begeleiding waren niet mogelijk geweest zonder de samenwerking en sfeer
die heerst in het labo. Daarom wil ik ook alle laboranten bedanken die steeds paraat waren
om mij te helpen deze stage tot een goed einde te brengen. In het bijzonder wil ik Kristof
Endels bedanken. Een betere stagementor had ik mij niet kunnen voorstellen. Ik wens hem
te bedanken voor zijn hulp, steun en de grappige momenten tijdens mijn stage. Ben Caljon
wens ik te bedanken voor zijn ondersteuning op vlak van informatica en theoretische
achtergrond van allerlei zaken.
Uiteraard wil ik ook Dr. Sonia Van Dooren bedanken. Als stagepromotor heeft zij er steeds
op toegezien dat alles vlot verliep en er mee voor gezorgd dat dit afstudeerwerkvorm kreeg.
Zij was steeds bereid uitleg te geven op al mijn vragen. Bedankt voor de vele suggesties en
tips en mij te doen doorzetten op momenten dat het even moeilijk ging.
Ook mijn ouders, broers, grootouders en toekomstige schoonfamilie mogen niet ontbreken.
Hen wil ik bedanken voor hun steun en luisterend oor. Ook mijn medestudenten Sarah,
Phebe en Noortje ben ik erg dankbaar. Samen een hapje eten of plezier maken om er
nadien weer helemaal ontspannen in kunnen vliegen.
Als laatste wil ik mijn verloofde Jan bedanken om mij te helpen bij de vormgeving van dit
afstudeerwerk, maar vooral voor de steun en de peptalks gedurende deze stageperiode.
Bedankt dat je er bent voor mij.
Dankzij iedereen hierboven vernoemd en alle anderen die ik vergeten ben, heb ik een
geweldige stageperiode gehad waar ik met een glimlach aan ga blijven terugdenken!

INHOUDSTAFEL
1. Inleiding.................................................................................................................... 111.1. Het hart...................................................................................................................... 121.2. Opbouw van het hart................................................................................................. 131.2.1. De hartwand .......................................................................................................... 131.2.2. Twee helften.......................................................................................................... 141.3. Prikkelgeleiding......................................................................................................... 141.4. Het elektrocardiogram (ECG) ................................................................................... 151.5. Hartritmestoornissen en hun ECG............................................................................ 161.5.1. Normaal ECG........................................................................................................ 161.5.2. De QT-syndromen................................................................................................. 171.5.3. Brugada syndroom (BrS) ...................................................................................... 191.5.4. Catecholaminerge polymorfe ventriculaire tachycardie (CPVT)........................... 222. Materiaal en methoden........................................................................................... 222.1. Primer design ............................................................................................................ 222.1.1. Principe.................................................................................................................. 222.2. DNA extractie ............................................................................................................ 242.2.1. Automatische DNA extractie ................................................................................. 242.2.2. Manuele DNA extractie ( proteïnase-digestie en fenol-chloroform extractie) ...... 262.3. Polymerase Chain Reaction (PCR) .......................................................................... 272.3.1. Principe.................................................................................................................. 272.3.2. Reagentia: AmpliTaq® 360 DNA-polymerase (Applied Biosystems Kit nr: 4398828)
292.3.3. Protocol ................................................................................................................. 302.4. Gelelektroforese........................................................................................................ 302.4.1. Principe.................................................................................................................. 302.4.2. Reagentia .............................................................................................................. 322.4.3. Protocol ................................................................................................................. 322.5. Enzym zuivering........................................................................................................ 322.5.1. Principe.................................................................................................................. 322.5.2. Reagentia (New England Biolabs) ........................................................................ 332.5.3. Protocol ................................................................................................................. 332.6. BigDye Cycle Sequencing ........................................................................................ 342.6.1. Principe.................................................................................................................. 342.6.2. Reagentia .............................................................................................................. 352.6.3. Protocol ................................................................................................................. 362.6.4. Analyse.................................................................................................................. 372.7. High Resolution Melting ............................................................................................ 382.7.1. Principe.................................................................................................................. 382.7.2. Reagentia .............................................................................................................. 422.7.3. Protocol ................................................................................................................. 422.7.4. Analyse.................................................................................................................. 443. Resultaten................................................................................................................ 463.1. Sequencing van SCN5A voor vier indexpatiënten ................................................... 463.2. SCN5A RT-PCR oppuntstelling & HRM pre-validatie .............................................. 503.2.1. SCN5A RT-PCR oppuntstelling ............................................................................ 503.2.2. SCN5A HRM pre-validatie exon scanning............................................................ 563.2.3. SCN5A HRM pre-validatie: gen-scanning ............................................................ 614. Besluit ...................................................................................................................... 705. Literatuurlijst ........................................................................................................... 726. Lijst met figuren ...................................................................................................... 737. Lijst met tabellen..................................................................................................... 75

Lijst met afkortingen
A: Adenine
ARVD/C: aritmogene rechter ventrikel dysplasie/ cardiomyopathie
AV-klep: atrioventriculaire-klep
BLAST: Basic Local Alignment Search Tool
Bp: basenparen
BrS: Brugada syndroom
C: Cytosine
C: hartritme
CCC: covalently closed conformation
CO2: koolzuur
Cp: cross point
CPVT: catecholaminerge polymorfe ventriculaire tachycardie
CRL: Contiguous Read Length
DCM: gedilateerde cardiomyopathie
ddNTP: dideoxynucleotide trifosfaat
DNA: deoxynucleotide acid
dNDP: deoxynucleotide difosfaat
dNMP: deoxynucleotide monofofaat
dNTP: deoxynucleotide trifosfaat
ds: double stranded
ECG: elektrocardiogram
EDTA: ethyleendiaminetetra-azijnzuur
F-primer: forward primer
G: Guanine
GPD1-L: glycerol-3-phospfate dehydrogenase 1-like
H-brug: waterstofbrug
HCM: hypertrofische cardiomyopathie
HRM: High Resolution Melting
LQTS: long QT-syndroom
NaAc: NatriumAcetaat
NCBI : The National Center for Biotechnology Information
NTC: no template control, negatieve controle
O2: zuurstof
OC: open nicked conformation
OD: optische densiteit
PCR: Polymerase Chain Reaction
RNA: ribonucleotide acid

Rpm: rounds per minute
R-primer: reverse primer
RT: real-time
RyR2: cardiale ryanodine receptor type 2
SA-knoop: sino-atriale knoop
SCD: sudden cardiac death
SNP: single nucleotide polymorphism
SQTS: short QT-syndroom
ss: single stranded
SSS: sick sinus syndroom
T: thymine
T: Thymine
Ta: annealingstemperatuur
TBE-buffer: Tris-Boorzuur-EDTA buffer
TE-buffer: Tris-EDTA buffer
Tm: smelttemperatuur
TRL: Trimmed Read Length
TRRM: Terminator Ready Reaction Mix
UV-licht: ultraviolet-licht

Samenvatting
Deze studie focust zich op de moleculaire diagnose van Brugada syndroom. Brugada
syndroom is een aangeboren hartritmestoornis in een structureel normaal hart, te wijten
aan een onevenwicht tussen in- en uitwaartse ionenstromen. Brugada syndroom wordt tot
op heden voornamelijk gelinkt met het SCN5A gen, dat codeert voor een Na+-kanaal.
Het doel van dit onderzoek was het ontwikkelen en uitvoeren van de moleculaire diagnose
van BrS op het SCN5A gen. Deze studie werd voorafgegaan door de oppunstelling van de
directe nucleotide cycle sequencing van SCN5A. Gezien de grootte van dit gen (28 exonen)
werd daarnaast ook de voorkeur gegeven aan de ontwikkeling van een diagnostische high-
throughput assay via een intermediair mutatiedetectie platform aan de hand van High-
Resolution-Melting (HRM) analyse. In een vorig werk werd deze oppunstelling opgestart en
in dit werk werd deze gefinaliseerd en geprevalideerd.
In een eerste deel van dit eindwerk werd de bidirectionele cycle sequencing van SCN5A
uitgevoerd voor vier indexpatiënten, die allen een typisch Brs ECG hebben. Bij deze
indexpatiënten werden in totaal 7 single nucleotide polymorphisms (SNPs) gevonden. Bij 1
indexpatiënt werd een splice-acceptorsite mutatie in intron 24 ontdekt, die, na verder
onderzoek in de familie, kan worden beschouwd als de familie-specifieke causale mutatie
van Brugada syndroom.
In het tweede deel werd de verdere oppuntstelling en prevalidatie van de SCN5A HRM
analyse uitgevoerd. Dit vereiste in eerste instantie de oppuntstelling van de real-time PCRs
voorafgaand aan de HRM analyse. De 14 nieuw ontwikkelde RT-PCRs werden steeds
geëvalueerd op sensitiviteit en specificiteit. Finaal kon de HRM analyse oppuntgesteld
worden voor 24 van de 28 SCN5A exonen. Voor de HRM-prevalidatie werd enerzijds een
exon-gerichte scanning uitgevoerd op de geïdentificeerde SNPs in de stalen van de
indexpatiënten uit deel 1. Alle gevonden SNPs konden ook via HRM-analyse opgepikt
worden, met uitzondering van de diep-intronische mutaties die met de HRM-primersets niet
gecoverd worden. Anderzijds werd een SCN5A gen-scanning uitgevoerd op stalen van 6
nieuwe BrS indexpatiënten en van de indexpatiënt met een causale BrS mutatie uit deel 1.
Hierbij werden een aantal abberante smeltpatronen gedetecteerd die via cycle sequencing
als 5 verschillende SNPs en 2 causale BrS mutaties in respectievelijk exon 21 en intron 21
geïdentificeerd werden.
Bijgevolg werd aangetoond dat de HRM analyse kan gebruikt worden als primaire
genscanningmethode voor SCN5A in het kader van de moleculaire diagnose van Brugada
syndroom. Dankzij de oppuntstelling zullen in de toekomst sneller mutaties en SNPs
kunnen worden gedetecteerd bij patiënten met dit syndroom.

Summary
This study is focused on the molecular diagnosis of Brugada syndrome. Brugada syndrome
is a congenital cardiac arrhythmia in a structurally normal hearth, due to an inbalance
between inward and outward ionic currents. Until now, the SCN5A gene, coding for a Na+
channel, is mainly linked with Brugada syndrome.
The aim of the research project was to develop and to perform the molecular diagnosis of
Brugada syndrome on the SCN5A gene. The study has been preceded by the optimization
of direct nucleotide cycle sequencing of SCN5A. Given the size of this gene (28 exons),
preference was given to the development of a diagnostic high-throughput assay through an
intermediary mutation detection platform, using High-Resolution-Melting (HRM) analysis.
This optimization had already started before this study. With this study it has been finalized
and prevalidated.
In a first part of the research project, the bidirectional cycle sequencing of SCN5A was
performed on four index patients, all showing a typical BrS ECG. Within these index
patients, 7 single nucleotide polymorphisms (SNPs) in total were found. Within one index
patient, a splice-acceptorsite mutation in intron 24 was discovered, that can be considered
to be a family specific causal mutation of Brugada syndrome.
The second part of the research project was focused on the further optimization and
prevalidation of the SCN5A HRM analysis. For this, it was first required to have the real
time PCRs optimized prior to the HRM analyses. The 14 newly designed RT-PCRs were
always assessed in function of their sensitivity and their specificity. The end result was that
the HRM analysis could be optimized for 24 of the 28 SCN5A exons. For the HRM
prevalidation, SCN5A exon-focused scanning was performed on the one side, and gene
scanning on the other side. The exon-focused scanning was executed on the identified
SNPs in the samples of the index patients of the first part of the research project. All of the
discovered SNPs could also be detected through HRM-analyses, with the exception of
deep intronic mutations that were not covered with the HRM primer sets. The SCN5A gene
scanning was executed on samples of 6 new BrS index patients and of the index patient
with the causal BrS mutation, discovered during the first part of the research project. During
this scanning, a number of aberrant melting patterns were detected, that could be identified
as 5 different SNPs and 2 causal mutations in exon 21 and intron 21, respectively.
As a result, it was shown that the HRM analysis can be used as a primary gene scanning
method within the framework of the molecular diagnosis of Brugada syndrome. Thanks to
the optimization of this method it might become easier in the future to detect SNPs and

mutations within patients having Brugada syndrome.
11
1. Inleiding
Genetisch onderzoek heeft uitgewezen dat aangeboren, primaire hartritmestoornissen
kunnen te wijten zijn aan ionenkanalen die niet optimaal functioneren. Deze ionenkanalen
staan in voor de initiatie en propagatie van de actiepotentiaal (het elektrisch signaal dat de
hartspier nodig heeft om samen te trekken). Momenteel zijn er heel wat cardiogenen
beschreven die elk coderen voor of betrokken zijn in het correct functioneren van een
ionenkanaal of een (subeenheid) (tabel 1). Wanneer dergelijk ionenkanaal wordt up- of
downgereguleerd, kan dit een effect hebben op het vermogen van de actiepotentiaal,
waardoor er een hartritmestoornis kan opgewekt worden. Het zijn vooral de K+-, Na+- en
Ca2+-kanalen die aangetast zijn. [1]
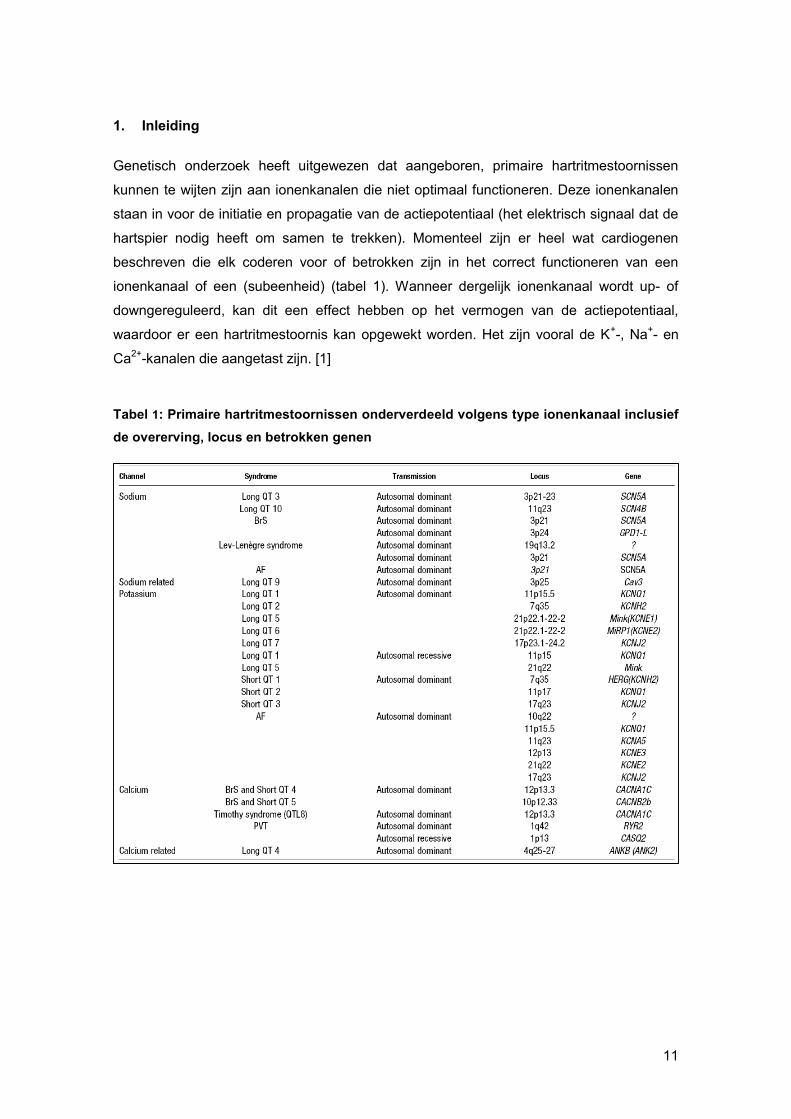
Tabel 1: Primaire hartritmestoornissen onderverdeeld volgens type ionenkanaal inclusief
de overerving, locus en betrokken genen

12
Primaire hartritmestoornissen als long QT-syndroom (LQTS); short QT-syndroom (SQTS),
Brugada syndroom (BrS) en catecholaminerge polymorfe ventriculaire tachycardie (CPVT)
zijn elektrische hartstoornissen die voorkomen in een structureel normaal hart. [2] (Een
tachycardie wordt gekenmerkt door een hartfrequentie hoger dan 100 slagen per minuut)
Deze syndromen worden voornamelijk via een autosomaal dominant patroon overgeërfd.
Uiteraard bestaan er ook secundaire hartritmestoornissen, te wijten aan een structureel
abnormaal hart. Dit zijn hypertrofische cardiomyopathie (HCM), gedilateerde
cardiomyopathie (DCM) en aritmogene rechter ventrikel dysplasie/ cardiomyopathie
(ARVD/C). Deze aandoeningen worden gekenmerkt door afwijkingen aan de sarcomeren
of het cytoskelet. [3]
Daarnaast kan het sick sinus syndroom (SSS) ook hartritmestoornissen veroorzaken. Deze
aandoening wordt gekenmerkt door degeneratieve fibrose of ionenkanaal stoornissen in de
sinusknoop, wat leidt tot hartritmestoornissen. [4]
Deze aangeboren hartritmestoornissen kunnen levensbedreigend zijn en kennen een hoge
mortaliteit. Het is daarom belangrijk dat de moleculaire ‘pathways’ ontrafeld worden om een
zo snel mogelijke diagnose te stellen, een goede therapie te starten en nieuwe targets te
vinden voor de ontwikkeling van nieuwe geneesmiddelen. [1]
Dit eindwerk spitst zich vooral toe op de moleculaire diagnose van 1 van de primaier
hartritmestoornissen, het Brugada syndroom aan de hand van enerzijds een direct
mutatiedetectie platform via directe nucleotide sequencing en anderzijds een intermediair
mutatiedetectie platform via High-Resolution-Melting (HRM) analyse.
1.1. Het hart
Het hart ligt in het midden van de borstholte en loopt uit naar de linkerkant. Het vormt een
stomp kegelvorming orgaan, ter grootte van een vuist, dat met de punt naar links beneden
is gericht. Deze punt wordt ook wel de apex genoemd. [3] Het is een spier die op een
consistente en gecontroleerde manier samentrekt. Deze regelmatige contracties zorgen
ervoor dat het bloed wordt rondgepompt doorheen het volledige menselijke lichaam. Op
deze manier voorziet de hartspier erin dat de in het bloed aanwezige zuurstof en
voedingsstoffen de organen en weefsels bereiken. De hartspier zorgt er ook voor dat
koolzuur (CO2) wordt afgevoerd naar de longen en daar, in de longblaasjes, wordt
uitgewisseld voor zuurstof (O2).
Bepaalde chemische stoffen die in de bloedbaan worden vrijgezet, waaronder adrenaline,
bepalen samen met de nervus vagus de snelheid waarmee de pacemakercellen een
elektrisch signaal genereren. Deze signalen zorgen voor het samentrekken en ontspannen
van de hartspier. [3]
Normaal trekt het hart 60 tot 100 keer samen per minuut. Hartritmestoornissen ontstaan
wanneer het hart onregelmatig samentrekt. Dit kan leiden tot doorbloedingsstoornissen in
allerlei organen, waardoor deze te weinig zuurstof en voedingsstoffen krijgen. Dit is
problematisch aangezien tekort aan O2 kan leiden tot orgaanfalen.
Hartritmestoornissen kunnen ook een oorzaak zijn van ‘sudden cardiac death’ (SCD). Dit is
een plots hartfalen en komt vaak voor bij personen die leiden aan genetische
hartaandoeningen en geleidingsstoornissen. Primair en secundaire hartritmestoornissen
hebben een ander werkingsmechanisme maar hebben vaak SCD als gevolg. [5]
In deze studie leggen we de nadruk op de eerste categorie van hartritmestoornissen, met
name diegene die te wijten zijn aan defecten in ionenkanalen of proteïnen die helpen in de
correcte expressie of functie van de ionenkanalen.
1.2. Opbouw van het hart
1.2.1. De hartwand
Het hart is omgeven door een stug, niet-elastisch bindweefselvlies. Dit wordt het pericard of
hartzakje genoemd en is een beschermingslaag voor het hart.
H
o
13
et hart heeft een hartwand die bestaat uit 3 lagen. Deze zijn van binnen naar buiten
pgebouwd uit: i) het endocard, ii) het myocard en iii) het epicard. Het endocard,
Figuur 1: Opbouw hartwand (epicard/pericard)
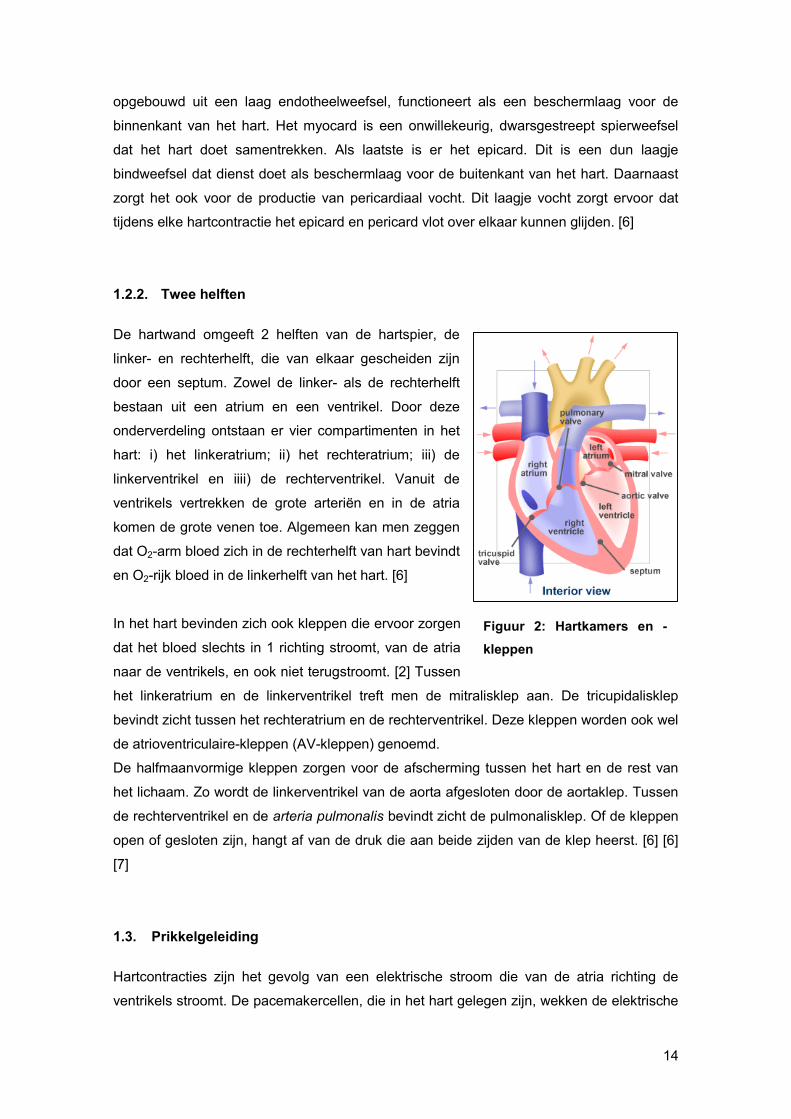
opgebouwd uit een laag endotheelweefsel, functioneert als een beschermlaag voor de
binnenkant van het hart. Het myocard is een onwillekeurig, dwarsgestreept spierweefsel
dat het hart doet samentrekken. Als laatste is er het epicard. Dit is een dun laagje
bindweefsel dat dienst doet als beschermlaag voor de buitenkant van het hart. Daarnaast
zorgt het ook voor de productie van pericardiaal vocht. Dit laagje vocht zorgt ervoor dat
tijdens elke hartcontractie het epicard en pericard vlot over elkaar kunnen glijden. [6]
1.2.2. Twee helften
De hartwand omgeeft 2 helften van de hartspier, de
linker- en rechterhelft, die van elkaar gescheiden zijn
door een septum. Zowel de linker- als de rechterhelft
bestaan uit een atrium en een ventrikel. Door deze
onderverdeling ontstaan er vier compartimenten in het
hart: i) het linkeratrium; ii) het rechteratrium; iii) de
linkerventrikel en iiii) de rechterventrikel. Vanuit de
ventrikels vertrekken de grote arteriën en in de atria
komen de grote venen toe. Algemeen kan men zeggen
dat O2-arm bloed zich in de rechterhelft van hart bevindt
en O2-rijk bloed in de linkerhelft van het hart. [6]
In het hart bevinden zich ook kleppen die ervoor zorgen
dat het bloed slechts in 1 richting stroomt, van de atria
naar de ventrikels, en ook niet terugstroomt. [2] Tussen
het linkeratrium en de linkerventrikel treft men de mitra
bevindt zicht tussen het rechteratrium en de rechterventrike
de atrioventriculaire-kleppen (AV-kleppen) genoemd.
De halfmaanvormige kleppen zorgen voor de afscherming
het lichaam. Zo wordt de linkerventrikel van de aorta afges
de rechterventrikel en de arteria pulmonalis bevindt zicht d
open of gesloten zijn, hangt af van de druk die aan beide
[7]
1.3. Prikkelgeleiding
Hartcontracties zijn het gevolg van een elektrische stroo
ventrikels stroomt. De pacemakercellen, die in het hart gel
Figuur 2: Hartkamers en -
kleppen
14
lisklep aan. De tricupidalisklep
l. Deze kleppen worden ook wel
tussen het hart en de rest van
loten door de aortaklep. Tussen
e pulmonalisklep. Of de kleppen
zijden van de klep heerst. [6] [6]
m die van de atria richting de
egen zijn, wekken de elektrische
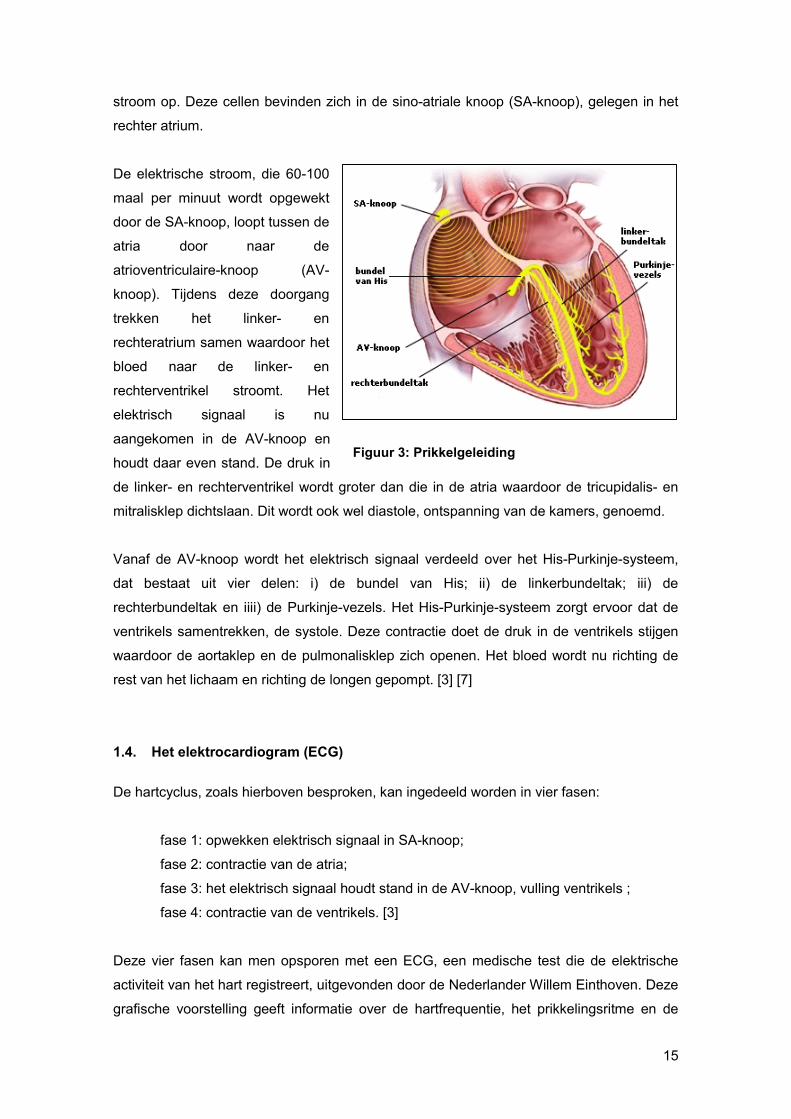
15
stroom op. Deze cellen bevinden zich in de sino-atriale knoop (SA-knoop), gelegen in het
rechter atrium.
De elektrische stroom, die 60-100
maal per minuut wordt opgewekt
door de SA-knoop, loopt tussen de
atria door naar de
atrioventriculaire-knoop (AV-
knoop). Tijdens deze doorgang
trekken het linker- en
rechteratrium samen waardoor het
bloed naar de linker- en
rechterventrikel stroomt. Het
elektrisch signaal is nu
aangekomen in de AV-knoop en
houdt daar even stand. De druk in
de linker- en rechterventrikel wordt groter dan die in de atria waardoor de tricupidalis- en
mitralisklep dichtslaan. Dit wordt ook wel diastole, ontspanning van de kamers, genoemd.
Vanaf de AV-knoop wordt het elektrisch signaal verdeeld over het His-Purkinje-systeem,
dat bestaat uit vier delen: i) de bundel van His; ii) de linkerbundeltak; iii) de
rechterbundeltak en iiii) de Purkinje-vezels. Het His-Purkinje-systeem zorgt ervoor dat de
ventrikels samentrekken, de systole. Deze contractie doet de druk in de ventrikels stijgen
waardoor de aortaklep en de pulmonalisklep zich openen. Het bloed wordt nu richting de
rest van het lichaam en richting de longen gepompt. [3] [7]
1.4. Het elektrocardiogram (ECG)
De hartcyclus, zoals hierboven besproken, kan ingedeeld worden in vier fasen:
fase 1: opwekken elektrisch signaal in SA-knoop;
fase 2: contractie van de atria;
fase 3: het elektrisch signaal houdt stand in de AV-knoop, vulling ventrikels ;
fase 4: contractie van de ventrikels. [3]
Deze vier fasen kan men opsporen met een ECG, een medische test die de elektrische
activiteit van het hart registreert, uitgevonden door de Nederlander Willem Einthoven. Deze
grafische voorstelling geeft informatie over de hartfrequentie, het prikkelingsritme en de
Figuur 3: Prikkelgeleiding

16
impulsuitbreiding. [3]
Het ECG wordt onderverdeeld in 6
golven van P to en met U.Voor het
begin van de P-top vindt fase 1
plaats: de opwekking van het
elektrisch signaal in de SA-knoop.
De P-top geeft de depolarisatie
van de atria weer, wat
overeenkomt met fase 2. De
repolarisatie van de atria is op zich
niet waar te nemen als een
grafische verandering omdat die
golf overgaat in de volgende golf,
het QRS-complex. Juist voor dit QRS-complex vindt fase 3 plaats: het stand houden van
het elektrisch signaal in de AV-knoop.
Het QRS-complex geeft de depolarisatie van de ventrikels weer, fase 4 van de hartcyclus.
De T-golf weerspiegelt uiteindelijk de repolarisatie van de ventrikels. De U-golf geeft de
ontspanning van het hart weer, juist voor de volgende samentrekking. [3] [8]
Het PR-interval loopt gelijk met de AV-geleidingstijd. Het QT-interval geeft de totale
tijdsduur van de- en repolarisatie van de ventrikels weer. Deze is afhankelijk van de
hartfrequentie. [8]
1.5. Hartritmestoornissen en hun ECG
1.5.1. Normaal ECG
Figuur 5 geeft de link weer
tussen de verschillende
ionenstromen en het ECG.
Wanneer tijdens fase 0 grote
hoeveelheden Na+ het hart
binnenstromen, krijgt men de
vorming van het QRS-complex,
de ventrikels trekken samen.
Fase 1 en 2 worden gekenmerkt
door een trage K+-uitstroom.
Gedurende fase 1 wordt de
Figuur 4: Het ECG
Figuur 5: Ionenstroom gelinked aan ECG

uitwaartse K+-stroom in evenwicht gehouden door een inwaartse Na+-stroom en Ca2+-
stroom. Een plateau-fase wordt gevormd. Tijdens fase 3 vindt een snelle K+-uitstroom
plaats. Deze 3 fasen geven de repolarisatie van de ventrikels weer. Fase 4, wat niet wordt
weergegeven in figuur 5, is de rustfase. Na deze fase is het hart klaar voor een nieuwe
cyclus. [1]
1.5.2. De QT-syndromen
1.5.2.1. Long QT-syndroom (LQTS)
LQTS is een overerfbare hartritme aandoening die wordt gekenmerkt door een verlenging
van het QTc-interval. Deze aandoening komt ongeveer voor bij 1 op de 10.000 individuen.
[7] Het QTc-interval is het QT-interval gecorrigeerd voor het hartritme (c) en wordt gemeten
tijdens een ECG aan de hand van de formule van Bazett:
Een nor
langer d
interval
hartritme
60-70%
vrouwen
mortalite
Naarma
verwaar
Onderst
het func
penetrat
17
maal QTc-interval bedraagt ongeveer tussen de 350-400 ms. Een QTc-interval
an 440 ms wordt vaak als abnormaal beschouwd. [6] Patiënten met een QTc-
groter dan 500 ms worden aanschouwd als patiënten met een hoog risico op
stoornissen. [10]
van de patiënten met LQTS zijn vrouwen. Men vermoedt dat het QTc, dat bij
van nature iets langer is dan bij mannen, de oorzaak hiervan is. Ook de hogere
it bij jonge, mannelijke LQTS-patiënten kan hiertoe bijdragen.
te de leeftijd ouder stijgt, is het verschil in incidentie tussen mannen en vrouwen te
lozen. [8]
aande tabel geeft voor de 3 belangrijkste LQTS, het aangetasten type ionenkanaal,
tioneel effect, de frequentie van voorkomen, het typische ECG, de ‘triggers’ en de
ie (fenotypische uiting van de mutatie) weer.
QTc = QT/√RR

18
Tabel 2: Correlatie tussen genotype en fenotype voor de drie belangrijkste LQT-genen
LQT1 of KCNQ1 codeert voor het IKs-ionenkanaal. Dit ionenkanaal staat in voor een trage
uitwaartse K+-stroom. K+-efflux zorgt voor repolarisatie van de ventrikels. Wanneer dit gen
gemuteerd is, resulteert dit in een verminderde IKs-ionenstroom door een defect in het
transport van de kanaalprotëinen naar het plasmamembraan, een falen in de kanaal
assemblage of niet functionele IKs-ionenkanalen. Dit heeft een tragere repolarisatie van de
ventrikels tot gevolg. [1] [10] [11]
LQT2 of KCNH2 codeert voor het IKr ionenkanaal. Dit ionenkanaal zorgt voor de snelle
uitwaartse K+-stroom. Een mutatie in dit gen zorgt opnieuw voor een verminderde K+-efflux
wat leidt tot tragere repolarisatie van de ventrikels waardoor de actiepotentiaal wordt
verlengd. [1] [10] [11]
Het INa ionenkanaal wordt gecodeerd door LQT3 of SCN5A. De Na+-instroom door dit
ionenkanaal zorgt voor de depolarisatie van de ventrikels. ‘Gain of function’ mutaties in dit
gen vergroten de Na+-instroom door een langer aanhoudende instroom, een vertraagde
inactivatiesnelheid, een sneller inactivatieherstel, een verschuiving in activiteit van
ionenkanaal of een abnormale interactie met de β-subeenheid. Hierdoor wordt de
depolarisatie van de ventrikels verlengd. [1] [10] [11]
1.5.2.2. Short QT-syndroom (SQTS)
SQTS is een zeldzame hartritmestoornis waarbij de patiënten een QTc-interval hebben dat
korter is dan 300-330 ms. Door ventriculaire fibrillaties hebben ze ook een hoog risico op
SCD. [4] [8] [11] Op het ECG worden vaak puntige T-golven zonder vlak ST-segment
waargenomen. De diagnose wordt gesteld aan de hand van verscheidene factoren: verkort
QTc-interval, syncope, ventriculaire fibrillatie of polymorfe ventriculaire tachycardie. [5] [9]

19
In de literatuur zijn er 5 genen beschreven die aanleiding kunnen geven tot SQTS. Bij
SQT1 veroorzaken mutaties in het KCNH2-gen een verhoogde IKr-uitstroom. Dit leidt tot
een verkorting van de repolarisatie van de ventrikels. ‘Triggers’ voor dit syndroom zijn idem
aan deze van LQT2: sport, lawaai en rust. [5] [9] [13]
Bij SQT2 ligt de oorzaak in mutaties in het KCNQ1-gen wat de IKs-uitstroom verhoogt en zo
het QT-interval verkort. [5] [9] [13]
Mutaties in het KCNJ2-gen, het IK1-kanaal, veroorzaken SQT3. De IK1-uitstroom wordt
verhoogd waardoor de laatste repolarisatie fase versnelt. SQT3 kan aanleiding geven tot
onverwacht wakker worden ’s nachts of hartkloppingen. [5] [7] [13]
Als laatste worden SQT4 en SQT5 geassocieerd met mutaties in het CACNA1C-gen en het
CACNB2b-gen. Deze 2 genen coderen respectievelijk voor de α1- en β1-subeenheid van
het L-type Ca2+-kanaal. Deze mutaties verlagen de ICa,L–instroom wat eveneens een
verkorting van de repolarisatie-fase geeft. [9]
1.5.3. Brugada syndroom (BrS)
In 1992 werd het BrS voor het eerst beschreven, gekenmerkt door een typisch ECG met
een verlenging van het ST-segment in de rechter precordiale geleider. Dit syndroom, met
een prevalentie van 5/10.000 personen, kenmerkt zich door een rechter bundel blok,
ventriculaire hartritmestoornissen en SCD zonder enige structurele hartafwijkingen. De
aandoening valt onder de ionenkanaal stoornissen en wordt autosomaal dominant
overgeërfd. Patiënten kunnen dit syndroom elk op een andere manier ervaren, gaande van
hartkloppingen en duizeligheid tot syncope en SCD. Deze symptomen komen vaak tot
uiting wanneer de patiënt rustig is, zoals ’s nachts. [2] [5] [14] [16]
Figuur 6: ECG Short QT versus normaal ECG
20
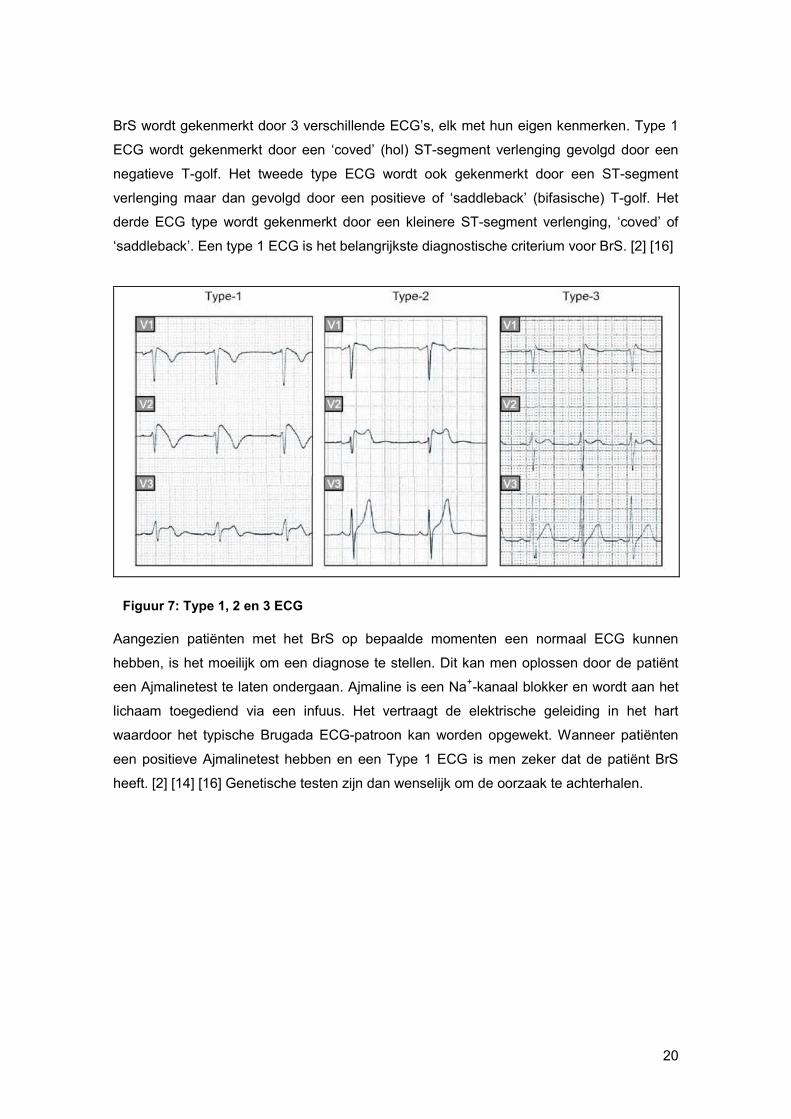
BrS wordt gekenmerkt door 3 verschillende ECG’s, elk met hun eigen kenmerken. Type 1
ECG wordt gekenmerkt door een ‘coved’ (hol) ST-segment verlenging gevolgd door een
negatieve T-golf. Het tweede type ECG wordt ook gekenmerkt door een ST-segment
verlenging maar dan gevolgd door een positieve of ‘saddleback’ (bifasische) T-golf. Het
derde ECG type wordt gekenmerkt door een kleinere ST-segment verlenging, ‘coved’ of
‘saddleback’. Een type 1 ECG is het belangrijkste diagnostische criterium voor BrS. [2] [16]
Aangezien patiënten met het BrS op bepaalde momenten een normaal ECG kunnen
hebben, is het moeilijk om een diagnose te stellen. Dit kan men oplossen door de patiënt
een Ajmalinetest te laten ondergaan. Ajmaline is een Na+-kanaal blokker en wordt aan het
lichaam toegediend via een infuus. Het vertraagt de elektrische geleiding in het hart
waardoor het typische Brugada ECG-patroon kan worden opgewekt. Wanneer patiënten
een positieve Ajmalinetest hebben en een Type 1 ECG is men zeker dat de patiënt BrS
heeft. [2] [14] [16] Genetische testen zijn dan wenselijk om de oorzaak te achterhalen.
Figuur 7: Type 1, 2 en 3 ECG

21
SCN5A, de α-subeenheid van het Na+-kanaal, is lang het enige gen geweest dat aan het
BrS werd gelinkt. Er zijn in dit gen al honderden mutaties gekend die een functieverlies
geven van de INa-instroom. Dysfunctie van het Na+-kanaal enerzijds maar ook een
verminderde vorming van Na+-kanalen anderzijds zijn oorzaak van deze verlaagde INa-
stroom. [14] [16]
Door een verminderde Na+-instroom tijdens fase 0 en een normale K+-uitstroom tijdens
fase 1 ontstaat er een onevenwicht tussen deze ionenstromen wat de hartritmestoornissen
veroorzaakt.
Hoewel mutaties in het SCN5A-gen
sterk gelinkt worden aan het BrS, wordt
er slechts bij 18-30% van de patiënten
ook een mutatie gevonden. In familiale
gevallen van BrS is het percentage aan
mutaties dan weer hoger in vergelijking
met sporadische gevallen. Deze zaken
wijzen er allemaal op dat het BrS een
syndroom is met een grote genetische
heterogeniteit.
Recent werden er naast SCN5A 4
nieuwe andere genen geassocieerd met
BrS. Mutaties in het gen, glycerol-3-
phosphate dehydrogenase 1-like
(GPD1-L) leiden indirect tot een
functieverlies van het Na+-kanaal. Mutaties in GPD1-L verminderen de aanvoer van Na+-
kanalen naar het celmembraan. [2] [16]
Figuur 8: Verschil in actiepotentiaal tussen normaal ECG (a), type 1 ECG (c) en type 2 ECG
Figuur 9: Verstoorde ionenstroom bij BrS

22
Mutaties in de 2 genen, CACNA1c en CACNB2b, die coderen voor 2 Ca2+-kanalen, kunnen
op hun beurt ook zorgen voor een verstoorde ionenstroom met BrS in combinatie met
SQTS tot gevolg. Daarnaast zijn mutaties in het KCNE3-gen ook geassocieerd met het BrS.
Dit gen codeert voor de β-subeenheid van een eiwit dat verantwoordelijk is voor de
uitwaartse K+-stroom. [16]
Door de ontdekking van deze nieuwe genen wordt BrS niet meer omschreven als een
louter Na+-kanaal stoornis, maar meer als een syndroom te wijten aan een onevenwicht
tussen de in- en uitwaartse ionenstromen. [16]
1.5.4. Catecholaminerge polymorfe ventriculaire tachycardie (CPVT)
Patiënten met CPVT hebben polymorfe tachycardiën uitgelokt door stress situaties. Bij
deze aandoening zorgen ‘gain-of-function’ mutaties in het cardiale ryanodine receptor type
2 (RyR2) gen voor een ongecontroleerde Ca2+ vrijzetting wat leidt tot hartritmestoornissen.
[2]
2. Materiaal en methoden
2.1. Primer design
2.1.1. Principe
Amplificatie van een specifiek ‘deoxynucleotide acid’ (DNA) fragment verkrijgt men alleen
wanneer men specifieke primersets gebruikt. Deze primersets zijn synthetische
oligonucleotiden die complementair en anti-parallel zijn aan de te amplificeren DNA
sequentie.
Voor de ontwikkeling van primersets voor HRM-analyse maakt men gebruik van het
gespecialiseerde computerprogramma ‘LigtScanner Primer Design Sofware 1.0’. Bij het
ontwikkelen van primers zijn er enkele punten waarop men moet letten.
2.1.1.1. Primer selectie: lengte en regio
Om een specifieke amplificatie te krijgen, kunnen primers best een lengte van 18-24
basenparen (bp) hebben. Primers met een lager aantal bp hebben echter een grotere kans
om aspecifiek in het genoom te binden.
De parameters van het programma worden zodanig ingesteld dat primers gegenereerd

23
worden die maximaal fragmenten van 350-400 bp overspannen. Fragmenten groter dan
400 bp zullen de mutatie detectie gevoeligheid verlagen, waardoor de kans op vals
negatieven groter wordt. De regio waarin primers worden ontwikkeld, ligt meestal een 50-
tot 100-tal basen verwijderd van het begin en eind van het exon, om een maximale mutatie
detetectie gevoeligheid aan de intron-exon boundaries te garanderen.
2.1.1.2. Primer specificiteit
De specificiteit van een primerset gaat men na met het computerprogramma Primer BLAST
‘Basic Local Alignment Search Tool‘ van ‘The National Center for Biotechnology’ (NCBI).
Dit programma gaat na hoe groot de kans is dat een primer ergens aspecifiek in het
humaan genoom kan binden.
Daarnaast vermijdt men beter eventueel aanwezige single nucleotide polymorphisms (SNP)
binnen primers, aangezien ze, afhankelijk van hun locatie, de oorzaak kunnen zijn van
allel-dropout of een verlaagde opbrengst van de polymerase-chain-reaction (PCR)-reactie.
De aanwezigheid van SNPs onder de ontworpen primers wordt nagegaan met het
softwarepakket ‘Alamut 1.5’. Dit databaseprogramma met een zeer gebruiksvriendelijke
interfase registreert en update continu gekende en beschreven SNPs en mutaties zodat
deze gemakkelijk op te sporen zijn in de sequentie van een bepaald gen. [16] [17] [18] [19]
[20]
2.1.1.3. De annealingstemperatuur
De smelttemperatuur (Tm), de temperatuur waarbij 50% van de primer-matrijs structuur
volledig gedenatureerd is, kan ondermeer bepaald worden met de formule van Thein en
Wallace en is afhankelijk van het Guanine (G)/ Cytosine (C)-gehalte:
Tm = 4°C x (GC) + 2°C x (AT)
Primers met een GC-gehalte van 40-60% zullen een stabiele binding geven met het
complementaire DNA fragment, aangezien GC-bindingen sterker zijn dan Adenine (A)/
Thymine (T)-bindingen (3 versus 2 waterstofbruggen (H-bruggen)). Ontwikkeling van
primers in een regio waar het percentage GC te hoog is, bemoeilijkt de amplificatie. [18] [20]
[21]
In functie van de Tm bepaalt men de annealingstemperatuur (Ta) van de primer. Deze Ta ligt
meestal 1°-5°C lager dan het primerpaar met de laagste Tm. Toch moet de optimale Ta van

24
elke primerset experimenteel bepaald worden.
Bij primer design is het belangrijk primerparen te ontwikkelen waarvan de Tm’s ongeveer
dezelfde zijn. Primerparen met sterk verschillende Tm’s resulteren doorgaan in inefficiënte
amplificatie.
Wanneer men een te lage Ta instelt, zullen de primers aspecifiek binden met een lagere
opbrengst van het gewenste DNA fragment tot gevolg. Bij een te hoge Ta zal de primerset
niet tot niet goed aan de complementaire DNA sequentie kunnen binden met geen tot lage
opbrengst als gevolg.
2.1.1.4. Zelfcomplementariteit van de primer
Primers met aan het 3’-uiteinde een G of een C helpen bij de correcte binding aan en
verlenging van het matrijs DNA. Een opeenvolging van >3 verschillende G of C’s aan het
3’-uiteinde is best te vermijden. Deze G- of C-stretch kan, door complementariteit, ‘hairpin’-
structuren (intramoleculaire hybridisatie) vormen, waardoor de primer niet meer kan binden
en niet meer beschikbaar is voor verlenging aan het 3’-OH-uiteinde. Een thymine (T) of T-
stretch aan het 3’-uiteinde wordt beter vermeden omdat deze aanleiding geeft tot een minst
stabiele binding aan het matrijs DNA.
Voor een primerpaar is het ook belangrijk dat de 3’-uiteinden geen complementaire
sequenties bevatten. Dit kan zelf- of primerset-dimerisatie bevorderen waardoor de
primer(s) evengoed niet meer beschikbaar zijn. Met het freeware programma ‘Auto Dimer’
kan men de kans op dimerisatie en hairpin structuren nagaan. Wanneer men in dit
programma de score op zelfcomplementariteit laag instelt (1 voor een perfecte ‘match’, -1
een ‘mismatch’, -0,25 voor een base (of N) met een N en -2 voor een 'gap') kan deze
maximaal gedetecteerd worden. Het is dan aan de gebruiker om uit te maken of de
gevonden zelf-complementariteit en hairpins significant de PCR-efficiëntie zullen
beïnvloeden. [20][21]
2.2. DNA extractie
2.2.1. Automatische DNA extractie
2.2.1.1. Principe
Met behulp van de ‘Multiprobe II Plus EX + Gripper’ robot en de Chemagen ‘Chemagic
Magnetic separation Module I’ kan men op een automatische manier DNA extraheren uit 5-
7 ml EDTA-bloed (EDTA wordt gebruikt als een coagulans).
Het EDTA-bloed wordt gemengd met een hoeveelheid lysebuffer en protease. De

25
lysebuffer zorgt ervoor dat de cellen opzwellen en openbarsten en het protease gaat de
aanwezige eiwitten afbreken. Hierdoor komt het DNA vrij, waarna het in aanwezigheid van
de bindingbuffer wordt gebonden aan ‘gecoate beads’. Wanneer men de magneet aanzet,
zullen de magnetisch ‘gecoate beads’, waaraan het DNA is gebonden, aangetrokken
worden. Op die manier kan, na een aantal wasstappen, het supernatans met cellulaire
resten verwijderd worden. Tenslotte elueert men het geëxtraheerde DNA in bijhorende
elutiebuffer. [22]
Met de Nanodrop® ND-1000 bepaalt men de concentratie en de
zuiverheid van het geëxtraheerde DNA. De concentratie van het
DNA-staal wordt bepaald aan de hand van de optische densiteit
(OD) bij 260 nm. De ratio 260/280, die de contaminatie door
protëinen weergeeft, moet rond de 1,8 liggen. De ratio 260/230
geeft de zuiverheid van het DNA-staal weer in verhouding met de
aanwezige hoeveelheid ribonucleotide acid (RNA). Een ratio van
1,8-2,2 is zeer goed en toont aan dat er geen contaminatie
aanwezig is. [22]
2.2.1.2. Reagentia (GC Biotech)
- Lysebuffer;
- Protease;
- Bindingbuffer;
- Magnetische beads;
- 5 wasbuffers;
- Elutiebuffer
2.2.1.3. Protocol
DNA-extractie:
- Zet het toestel en het programma klaar;
- Breng maximum 7 ml bloed aan in de 50 ml falcon tubes;
- Plaatsing stalen, tubes, tips;
- Vul de nodige vloeistoffen aan (magnetische beads, protease, elutiebuffer, buffer);
- Start de run (per run kan er uit twaalf bloestalen DNA geëxtraheerd worden);
Figuur 10:
Nanodrop® ND-1000

26
Nanodrop:
- Schakel de software in;
- Kalibreren (blanco instellen);
- Concentratie stalen meten.
2.2.2. Manuele DNA extractie ( proteïnase-digestie en fenol-chloroform extractie)
2.2.2.1. Principe
De proteïnase K digestie en fenol-chloroform extractie is een techniek die wordt toegepast
voor DNA-extractie uit andere stalen dan bloedstalen, bv. prenatale chorion villi of
amniocyten, biopten, urine, enz. [23]
Een lysebuffer let detergent en proteïnase K worden toegevoegd aan cellen om de
celmembraan te lyseren en het DNA vrij te maken van proteïnen.
Tijdens de fenol-chloroform extractie zal het vrijgekomen DNA oplossen in de waterige fase
(bovenaan) terwijl alle andere celcomponenten oplossen in de organische fase (onderaan).
Dit opgezuiverd DNA kan men gebruiken voor moleculaire testen. [23]
2.2.2.2. Reagentia
- 1x Tris-EDTA buffer (TE-buffer) pH 8 (Promega);
- 4mg/ml Proteïnase-K: 20mg gelyofiliseerd oplossen in 5ml water (Merck);
- Fenol (Sigma);
- Chloroform/isoamylalcohol 24:1 (VWR);
- Absolute Ethanol (apotheek);
- 3M NatriumAcetaat (24.61 g NaAc aanlengen tot 100 ml met gesteriliseerd water);
- Zuiver, steriel en pyrogeenvrij water (Kela);
- 10% SDS (Sigma).
waterige fase
organische fase

27
2.2.2.3. Protocol
- Voeg 25 μl Proteïnase-K, 425 μl TE-buffer en 50 μl 10% SDS toe aan CVS-, spier-,
hart-, of leverstalen;
- Voeg 5 μl Proteïnase-K, 445 μl TE-buffer en 50 μl 10% SDS toe aan AC-, fibro-,
leuco-, EBV-, NS-, of CVSLTstalen;
- Voeg 500 μl van de Proteïnase-K/TE-buffer/10%SDS oplossing toe aan het te
bereiden staal en incubeer het 2u in een warmwaterbad van 45°C;
- Vortex het nadien 1 maal en voeg 500 μl fenol toe;
- Bereid 70% ethanol-oplossing (300 μl aqua ad injectabilia en 700 μl absolute
ethanol);
- Bereid Ethanol/NaAcetaat-oplossing (1 ml absolute ethanol en 45 µl 3 M
Natriumacetaatoplossing);
- Meng de fenol- en waterfase homogeen door de eppendorftube enkele malen om
te zwenken;
- Centrifugeer gedurende 5 min. aan 8000 ‘rounds per minute’ (rpm) (6726g);
- Pipetteer de bovenste fase over naar een nieuw eppendorftube en voeg 500 µl
chloroform/isoamyethanol 24:1 (96 ml chloroform en 4 ml isoamylalcohol ) toe;
- Centrifugeer na mengen 5 min. aan 8000rpm (6726g);
- Pipetteer de bovenste fase over naar een nieuwe eppendorftube en voeg opnieuw
500 µl chloroform/isoamyethanol 24:1 toe;
- Centrifugeer na mengen opnieuw 5 min aan 8000 rpm (6726g);
- Pipetteer de bovenste fase over naar een hemolysebuisje;
- Na vortexen wordt een slijmerige vlok, DNA, gevormd;
- Pik de DNA-vlok op met een toegeschroeide pasteurpipet;
- Spoel de DNA-vlok met 70% ethanol en laat drogen aan de lucht;
- Los de vlok op in de gewenste hoeveelheid TE-buffer;
- Draai parafilm rond het dekseltje van tube en laat het overnacht op een rad in de
koelkamer (+4°C) oplossen;
- Bepaal de concentratie aan DNA met de Nanodrop;
- Bewaar het DNA op 4°C.
2.3. Polymerase Chain Reaction (PCR)
2.3.1. Principe
PCR is een methode waarmee men, vertrekkende van een kleine hoeveelheid DNA, een
28
specifieke DNA-sequentie exponentieel kan amplificeren. Het is een methode die in vitro
wordt uitgevoerd en die wordt gekenmerkt door een cyclische afwisselende
temperatuursstijging en –daling.
Om een specifieke DNA-sequentie te amplificeren maakt men gebruik van primers (± 20
bp), die complementair en anti-parallel zijn aan de matrijs, en van een thermostabiel DNA
polymerase, Taq polymerase. Dit Taq polymerase wordt geïsoleerd uit Thermus Aquaticus.
Het is actief tot een temperatuur van 96°C en heeft volgende eigenschappen:
- 5’ → 3’ polymerase activiteit;
- 5’ → 3’ exonuclease activiteit;
- Geen 3’ → 5’ exonuclease activiteit (geen proofreading);
- Verlengt 3’ uiteinde met 1 Adenine (A);
- Heeft matrijs en deoxynucleotide trifosfaten (dNTPs) nodig om zijn polymerase
activiteit uit te voeren;
- Heeft Mg2+-ionen nodig als cofactor. [24]
Een PCR-reactie bestaat uit volgende stappen:
- Initiële denaturatie bij 95°C voor 1’;
- Denaturatie bij 95°C voor 30’’;
- Annealing bij Ta van de primerset voor 30’’;
- Extensie bij 72°C;
- Finale extensie bij 72°C;
- Koeling bij 10°C.
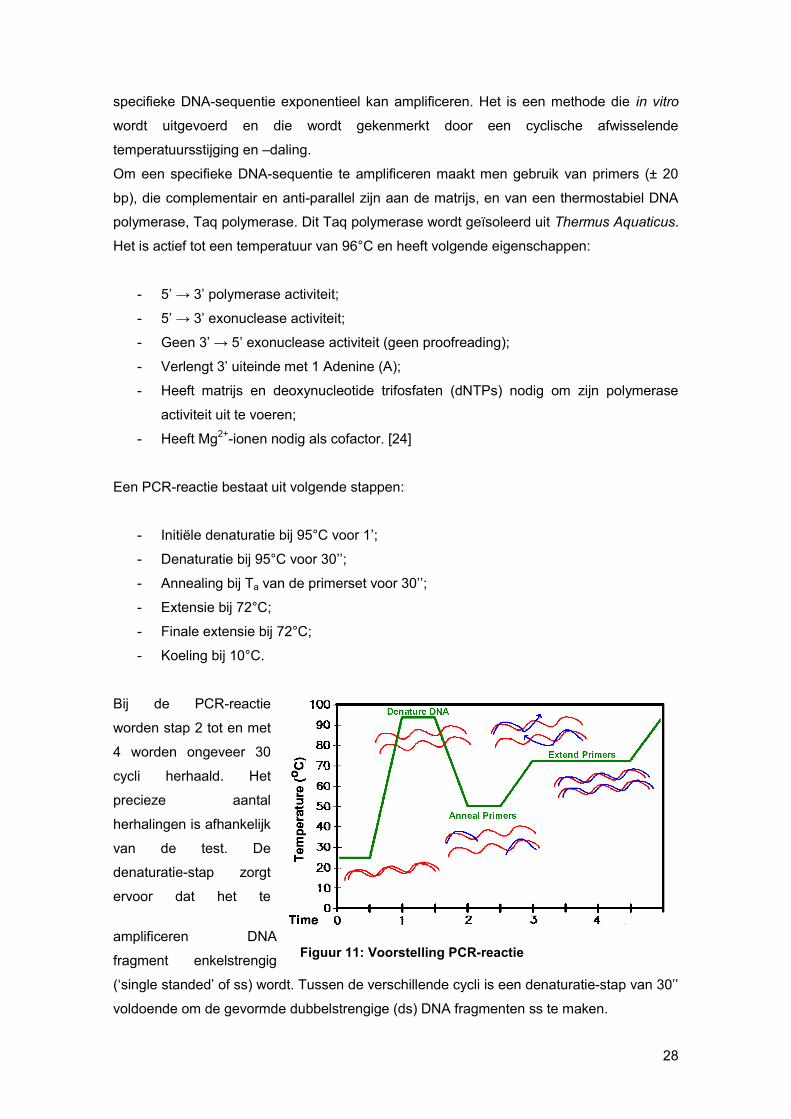
Bij de PCR-reactie
worden stap 2 tot en met
4 worden ongeveer 30
cycli herhaald. Het
precieze aantal
herhalingen is afhankelijk
van de test. De
denaturatie-stap zorgt
ervoor dat het te
amplificeren DNA
fragment enkelstrengig
(‘single standed’ of ss) wordt. Tussen de verschillende cycli is een denaturatie-stap van 30’’
voldoende om de gevormde dubbelstrengige (ds) DNA fragmenten ss te maken.
Figuur 11: Voorstelling PCR-reactie

29
De temperatuur van de annealing-stap – de temperatuur waarop de primers binden op het
ss DNA- ligt ideaal net onder de Tm van de primers. De extensie-stap gebeurt op 72°C, de
temperatuur waarop het Taq pol het 3’ OH-uiteinde van de primers verlengt. Nadat het
ingegeven aantal cycli doorlopen is, vindt de finale extensie plaats. Na deze stap zijn alle
gevormde DNA fragmenten in principe volledig ds. Nadien wordt het PCR-product gekoeld
op 10°C, een temperatuur waarop het zeer stabiel is.
Na ongeveer 30 cycli zal er geen exponentiële stijging meer zijn. De PCR reactie zal op dat
moment zijn plateau-fase bereiken. Reagentia (primermix, dNTPs) die zijn opgebruikt, het
Taq pol dat niet meer optimaal werkt of de opstapeling van PPi, dat inhiberend werkt op de
PCR-reactie, zijn mogelijke oorzaken van de plateau-fase. [25] [26]
Wanneer men verschillende primersets met eenzelfde in silico bepaalde Ta range wil
samen testen, kan men opteren voor een ‘touchdown’ PCR. Bij een ‘touchdown’ PCR wordt
de Ta over een range van maximaal 10°C stapsgewijs per cyclus met 0,5°C verlaagd. Op
deze manier zullen de verschillende primersets elk bij hun eigen ideale temperatuur
‘annealen’.
Om de downstream directe nucleotide cycle sequencing-reactie te standaardiseren,
worden aan de PCR-primers M13-tags gekoppeld. Deze M13-tags zullen niet aspecifiek
binden binnen het genoom maar worden wel mee geamplificeerd. Voor meer uitleg zie 2.6.
Om na te gaan of er tijdens de PCR-reactie geen contaminatie is opgetreden, neemt men
steeds een ‘no template control of NTC’ mee. Deze NTC bevat alle reagentia behalve DNA.
De NTC wordt ook steeds meegenomen tijdens de gelcontrole.
2.3.2. Reagentia: AmpliTaq® 360 DNA-polymerase (Applied Biosystems Kit nr:4398828)
- AmpliTaq 360 10X PCR buffer (stockconcentratie) → 1X (werkconcentratie);
- 25mM MgCl2-oplossing (stockconcentratie) → ± 2mM (werkconcentratie);
- 200 μM dNTPs (GE Healthcare);
- AmpliTaq 360 5U/μl → 1U;
- primer mix forwared (F)/reverse (R) ( finaal elk 2 μM) (IDT);
- DNA verdunning (finaal 1,70 μg/ml);
- Water tot 50 μl (Kela);

30
- Veriti® Thermal Cycler (Applied Biosystems).
2.3.3. Protocol
- Nummer de nodige PCR tubes;
- Maak reactie mix, steeds in overmaat;
- Voeg het Taq pol op het einde toe (zie tabel 3);
- Verdeel 30 μl mix uit in elke PCR tube;
- Voeg 5 μl primermix toe (bevat forward en reverse primers) toe aan elke PCR tube;
- Maak DNA verdunning onder de laminaire flow om DNA-contaminatie te vermijden,
steeds in overmaat;
- Voeg 15 μl van de DNA-verdunning toe aan de genummerde PCR tubes, steeds
onder de laminaire flow om DNA-contaminatie te vermijden;
- Plaats de PCR-tubes in de Veriti® Thermal Cycler (applied Biosystems) en stel het
PCR-programma in;
Tabel 3: Samenstelling reactiemix PCR
Volumes in µl
# react.
8/ 12,75 102
1x 5 40
2 mM 4 32
200 µM 8 64
1 U 0,25 2
30 240
AmpliTaq (5U/µl)
totaal
Water
GeneAmp 10x PCR Buffer II
25 mM MgCl2 solution
dNTPs
Lotnummer (+ firma) Finale
conc.
Samenstelling 1 reactie
2.4. Gelelektroforese
2.4.1. Principe
De migratie van moleculen, in een dragermateriaal, doorheen een elektrisch veld noemt
men elektroforese. Deze moleculen kunnen gescheiden worden op basis van:
- grootte;
- conformatie;
- ladingsdensiteit.

31
De grootte komt voor een DNA fragment overeen met de lengte. DNA fragmenten met een
groot aantal bp zullen trager migreren dan DNA fragmenten met een kleiner aantal bp.
‘Covalently closed conformation’ (CCC), ‘open nicked conformation’ (OC) en lineair zijn de
3 conformatie vormen waarin DNA kan voorkomen. PCR-producten zijn lineaire DNA
fragmenten en migreren sneller dan OC en CCC DNA.
Bij neutrale pH is de lading van elk DNA-molecule hetzelfde. De fosfaatgroepen van de
fosfaat-pentose ruggengraat zijn op die pH geïoniseerd, wat het DNA een netto negatieve
lading geeft. De DNA moleculen zullen hierdoor allemaal naar de positieve pool (anode)
van het elektrisch veld migreren. [26]
Als dragermateriaal gebruikt men het lineair polysaccharide agarose dat geëxtraheerd
wordt uit zeewier en is opgebouwd uit galactose en een galactose derivaat. Het wordt
opgelost in Tris-Boorzuur-EDTA-buffer (TBE-buffer) en is vloeibaar bij 95°C. Bij afkoelen,
zullen er zich H-bruggen vormen tussen de bouwstenen waardoor men gel-vorming krijgt.
Om de DNA fragmenten na scheiding te visualiseren onder ultraviolet licht (U.V-licht) voegt
men aan de TBE-buffer, opgelost in H20, 100 μl Gelred toe. Gelred is een fluorescente dye
die, in tegenstelling tot ethidiumbromide, veiliger en veel gevoeliger is. Het intercaleert
tussen de baseparen en absorbeert U.V.-licht van 300-360 nm, waarna het een fluorescent
signaal uitzendt. De sterkte van een gekleurd bandje op gel komt overeen met de
hoeveelheid geamplificeerd DNA. [27]
Wanneer men een PCR van een bepaald exon op
punt stelt, is het belangrijk na te gaan of men het
juiste fragment heeft geamplificeerd. Dit kan men
doen door een moleculaire gewichtsladder mee op
gel te zetten. Deze moleculaire gewichtsladder
bevat x-aantal fragmenten waarvan de lengte gekend
is. Aan de hand hiervan kan men de lengte van het
geamplificeerde DNA fragment afleiden.
Voor een controle van PCR-fragmenten voor sequencing, met een lengte tussen 200 à
500bp, maakt men een gel van 2% agarose. Voor real-time (RT) PCR-controle tussen 100
en 350bp maakt men een gel van 3% agarose, om een betere scheiding van fragmenten te
bekomen.
Figuur 12: Foto gelelektroforese

32
2.4.2. Reagentia
- UltraPure Agarose Electrophoresis (invitrogen);
- 10X TBE-buffer (stockconcentratie) → 1X (werkconcentratie) (Invitrogen);
- Gelred 10μl in 100 μl (Biotium);
- Eventueel moleculaire gewichtsmerkers (Roche en New England Bio-Labs);
- Draagbakje en kammetjes (Eurogentec Mupid-ex);
- Elektroforesekamer (Eurogentec Mupid-ex);
- UV-transilluminator + camera (TFP-M/WL Vilber Lourmat).
2.4.3. Protocol
- Meet 70 ml TBE-buffer 1X af in een maatcilinder en breng over in een fles (idem
voor RT-PCR-controle);
- Weeg 1,4 g agarose (2,1 g voor RT-PCR-controle) af in een weegschuitje en voeg
toe aan de TBE-buffer 1X;
- Meng de agarose en verwarm in een microgolfoven tot
alle agarose is opgelost;
- Laat 10 min afkoelen en giet oplossing in een
draagbakje + kammetjes;
- Laat de gel stollen;
- Haal de kammetjes uit gel;
- Leg de gel in een elektroforesekamer (gevuld met TBE-
buffer);
- Laad 10 μl PCR-product (5 μl voor RT-PCR-controle);
- Stel de timer in op 20 minuten (60 minuten voor RT-
PCR-controle);
- Stel de spanning in op 100V;
- Visualiseer na de aangegeven tijd het DNA in de gel onder de U.V.-transilluminator
en maak een foto.
2.5. Enzym zuivering
2.5.1. Principe
Na een PCR-reactie bevindt er zich in de PCR-tube nog onverbruikte primersets en vrije
dNTPs die moeten verwijderd worden vooraleer men de sequencing kan verderzetten. De
aanwezigheid van zowel een F- als een R-primer kunnen het elektroferogram, dat aan het
Figuur 13:
Gelektroforesekamer +
kammetjes

33
einde van de sequencing bekomen wordt, verstoren. Bovendien kan een te hoge
concentratie aan vrije dNTPs de incorporatie van dideoxynucleotiden trifosfaten (ddNTPs)
bemoeilijken. [28]
Voor de enzymzuivering gebruikt men 2 enzymen: Exonuclease I en Antartic Phosphatase.
Exonuclease I breekt ssDNA en dus de primers in een 3’ →5’ richting af en vormt zo
deoxynucleotide monofosfaten (dNMPs). Antartic Phosphatase breekt de vrije dNTPs af tot
deoxynucleotide difosfaten (dNDPs), dNMPs en zo tot nucleosides. Deze componenten
kunnen niet meer verbruikt worden en dus ook niet meer interfereren in verdere reacties.
[28]
nucleosidedNMPotideoligonucle ePhosphatasAntarcticIeExonucleas
nucleosidedNMPdNDPdNTP ePhosphatasAntarcticePhosphatasAntarcticePhosphatasAntarctic
2.5.2. Reagentia (New England Biolabs)
- Exonuclease I 20 U/μl (stockconcentratie) → 1 U/μl;
- Antartic Phosphatase 5 U/μl (stockconcentratie) → 1 U/μl;
- Beide opgelost in 100 μl H2O
2.5.3. Protocol
- Nummer de nodige PCR-tubes;
- Voeg aan elke PCR-tube 1 μl enzym-mix (5 μl Exonuclease I, 20 μl Antartic
Phosphatase en 75 μl H20 voor 100 μl enzym-mix) toe;
- Voeg 5 μl PCR-product toe;
- Zet PCR-tubes in Veriti® Thermal Cycler en stel volgend programma in:
Tijd Temperatuur
Incubatie 15' 37°C
Denaturatie 20' 80°C
Koeling ∞ 10°C
De stalen zijn nu zuiver om een BigDye Cycle Sequencing reactie uit te voeren.

34
2.6. BigDye Cycle Sequencing
2.6.1. Principe
Met sequencing bepaalt men de opeenvolging van basen in een DNA-keten. De techniek,
gebaseerd op de keten-terminatie methode van Sanger, is geautomatiseerd en maakt
gebruik van 4 fluorescent gelabelde ddNTPs: ddATP; ddGTP; ddTTP en ddCTP. De
ddNTPs hebben aan hun 3’-uiteinde een H-groep in plaats van een OH-groep en worden in
ondermaat toegevoegd aan het reactiemengsel. De H-groep die het ddNTP draagt,
voorkomt verlenging van de DNA-keten aan het 3’-Huiteinde. [28]
In plaats van, zoals bij een
PCR-reactie, de F-en R-primer
toe te voegen, voegt men hier
enkel de F- of R-primer toe. Het
geamplificeerde DNA fragment
zal nu slechts langs 1 zijde
verlengd worden met een
lineaire amplificatie tot gevolg.
Door de ddNTPs in ondermaat
toe te voegen, zullen ze door
het Taq pol op willekeurige
posities ingebouwd worden wat
resulteert in fluorescent gelabelde ssDNA fragmenten van verschillende lengte. [25] [28]
Voor de BigDye Cycle Sequencing-reactie heeft men - juist zoals bij een PCR-reactie -
Figuur 14: Verschil tussen
dNTP en ddNTP
Figuur 15: BigDye Cycle sequencing

35
buffer, Taq-pol en dNTPs nodig. De ddNTPs zijn specifiek voor de BigDye Cycle
Sequencing-reactie.
Het Taq pol, de dNTPs en de ddNTPs zitten samen in een mastermix, Terminator Ready
Reaction Mix (TRRM). De buffer, het water, de M13 F- of R-primers en het gezuiverde
PCR-product moeten nog apart toegevoegd worden. [28]
Om na de BigDye Cycle Sequencing de niet verbruikte M13 primers, dNTPs en ddNTPs te
verwijderen, gebruikt men de BigDye® X-TerminatorTM Purification Kit. Deze kit bevat een
buffer, SAM-oplossing en hars, X-Terminator. De X-terminator hars zal onder constant
schudden alle vrije componenten binden en capteren. Tijdens en na het afdraaien, zakt de
X-Terminator uit naar de bodem. De bovenstaande SAM-buffer bevat de ssDNA
fragmenten en is vrij van interfererende deeltjes. [28]
De ssDNA fragmenten worden gescheiden en geanalyseerd op de ABI 3130 xl Genetic
Analyzer. In dit toestel worden de ssDNA fragmenten gescheiden via capillaire
elektroforese. De uitgezonden fluorescentie wordt in bepaalde golflengtegebieden (virtuele
filters) opgevangen en gecollecteerd door een CCD camera. De opgevangen
lichtintensiteiten worden opgeslagen als digitale signalen die de computer dan met behulp
van het Sequence Analysis software v5.2 verwerkt tot een elektroferogram die de
opeenvolgende basen weergeven.
2.6.2. Reagentia
BigDye Cycle Sequencing (Applied Biosystems):
- 1 μl Gezuiverd PCR product;
- 1.5 μl BigDye Sequencing buffer 5X (stockconcentratie) → 0.75x
(werkconcentratie);
- 1 μl TRRM (bevat Taq pol, dNTPs en ddNTPs) 2.5x (stockconcentratie) → 0.25x
(werkconcentratie);
- 1 μl M13 primer (F en R) 0.16 μM (werkconcentratie);
- 5,5 μl water.
Plaatzuivering (Applied Biosystems):
- 35 μl SAM-buffer;
- 7.5 μl X-Terminator (deze moet voor gebruik zeer goed gemixt worden);

36
- Schudapparaat (IKA® MS3 Digital).
2.6.3. Protocol
BigDye Cycle Sequencing:
- 9 μl mix (buffer, TRRM, M13 primer en water) in elke PCR tube en 1 μl enzym
gezuiverd PCR product;
- Stel programma in en run op de Veriti® Thermal Cycler:
Tijd Temperatuur
Denaturatie 1' 96°C
Deanturatie 10' 96°C
Annealing 5' 50°C
Extentie 4' 60°C
Koeling ∞ 10°C
Plaatzuivering:
- Meng voor 1 staal 35 μl SAM en 7,5 μl X-Terminator;
- Voeg per well 42.5 μl van de oplossing toe aan de 96-well plaat;
- Voeg10 μl Big Dye product toe;
- Laat 30 min schudden op 3000 rmp en daarna 2 min afdraaien 1000 g;
- Plaat laden in de ABI 3130 xl Genetic AnalyzerSequencing inzetten
=> namen ingeven;
=> oven aanzetten;
=> programma instellen (standaard, kort of lange injectie, lange runtijd,…).
25 x

37
2.6.4. Analyse
De ABI 3130xl Genetic Sequencer geeft, na de run, de resultaten weer in
elektroferogrammen. Deze elektroferogrammen zijn een grafische weergave van de
nucleotidensequenties. Ze worden manueel gelezen, aan de hand van een
referentiesequentie. Nadien worden de sequenties nagekeken in het software programma
‘Variant Reporter v1.1’. Om een analyse in Variant Reporter v1.1 uit te voeren, worden
eerst enkele analyseparameters ingesteld waaronder ‘basecalling’ en ‘trimming. Tijdens de
‘basecalling’ gaat het programma na of het aan elke piek, gegenereerd in het
elektroferogram, een base kan toeschrijven. Nadien wordt de sequentie ‘getrimmed’.
Tijdens deze stap worden 5’ en 3’ uiteinden van de sequentie verwijderd volgens vooraf
aangegeven criteria. Na het instellen kunnen de referentiesequentie en de te analyseren
sequenties in het programma geïmporteerd en geanalyseerd worden. Na deze stap wordt
eerst aan de hand van de kwaliteitsparameters nagegaan welke sequenties al dan niet
voldoen aan de analyseparameters. De onderzochte kwaliteitsparameters zijn ‘Average
Trace Score’, ‘Median PUP score ’, ‘Contiguous Read Lenght’ (CRL), ‘Trimmed Read
Lenght’ (TRL) en ‘Signal Strength’. De belangrijkst sequentie kwaliteitsparameter is
‘Average Trace Score’ en geeft een kwaliteitswaarde aan de sequenties die overblijven na
‘trimming’. Een trace score van 30 stemt overeen met een 99.9% zeker basecalling. De
‘Median PUP score’ geeft de zuiverheid van de sequenties weer: hoe hoger de PUP score,
hoe minder achtergrondsequentie er aanwezig is. De CRL zegt iets over de continue lengte
van de sequentie waarin de basecalling vlot verloopt. De TRL laat zien of de sequentie nog
aan de analyseparameters voldoet na ‘trimming’. Als laatste is er de ‘Signal Strength’, die
weergeeft welke sequenties een piekintensiteit hebben hoger dan de minimum ingestelde
piekintensiteit van 200.
Of een sequentie wel of niet voldoet aan elk van deze kwaliteitsparameters wordt
weergegeven aan de hand van een kleurencode. Sequenties die groen kleuren zijn OK.
Sequenties die oranje kleuren zijn twijfelachtig. Sequenties met een rode kleur, worden niet
goedgekeurd.
Bidirectionele veranderingen ten opzichte van de referentiesequentie worden nagekeken in
Alamut v1.5 en gerapporteerd. [28]

38
2.7. High Resolution Melting
2.7.1. Principe
HRM is een sensitieve gen-scanning methode, gebaseerd op heteroduplex analyse, die
wordt gebruikt om de aanwezigheid van mutaties en SNPs op te sporen. In eerste instantie
amplificeert men een specifiek PCR-fragment via RT-PCR-reactie. Na de RT-PCR-reactie
wordt het amplicon onderworpen aan de HRM-analyse.
De HRM-methode is een methode met verscheidene voordelen:
- Er zijn slechts kleine hoeveelheden reagentia nodig (20 μl reactievolume);
- Er is slechts een kleine hoeveelheid staal nodig (HRM-analyse direct na PCR);
- Het is een simpele en snelle techniek om de aanwezigheid van mutaties op te
sporen;
- Een snelle optimalisatie is mogelijk;
- Hoge reproduceerbaarheid;
- Het heeft een hoge nauwkeurigheid en sensitiviteit;
- De HRM-analyse wordt uitgevoerd in een gesloten systeem (minder kans op cross-
contaminatie);
- Het PCR-product van de HRM-analyse kan gebruikt worden om een BigDye Cycle
Sequencing-reactie op uit te voeren. [29] [30]
Een RT-PCR-reactie is identiek aan een gewone PCR-reactie met als extra voordeel dat
men de reactie kan volgen in de tijd. Om de PCR-reactie in ‘real time’ te kunnen volgen,
wordt er aan de Master Mix een fluorescerende dye – Resolight – toegevoegd. Resolight
heeft enkel affiniteit voor ds DNA en satureert deze ook volledig wanneer het in overmaat
wordt toegevoegd. Tijdens de amplificatie nestelt de fluorescente dye zich in de ds DNA
streng. Naarmate er meer kopijen van het te amplificeren DNA fragment gevormd worden,
stijgt de fluorescentie.
Figuur 16: Voorstelling elektroferogram
39
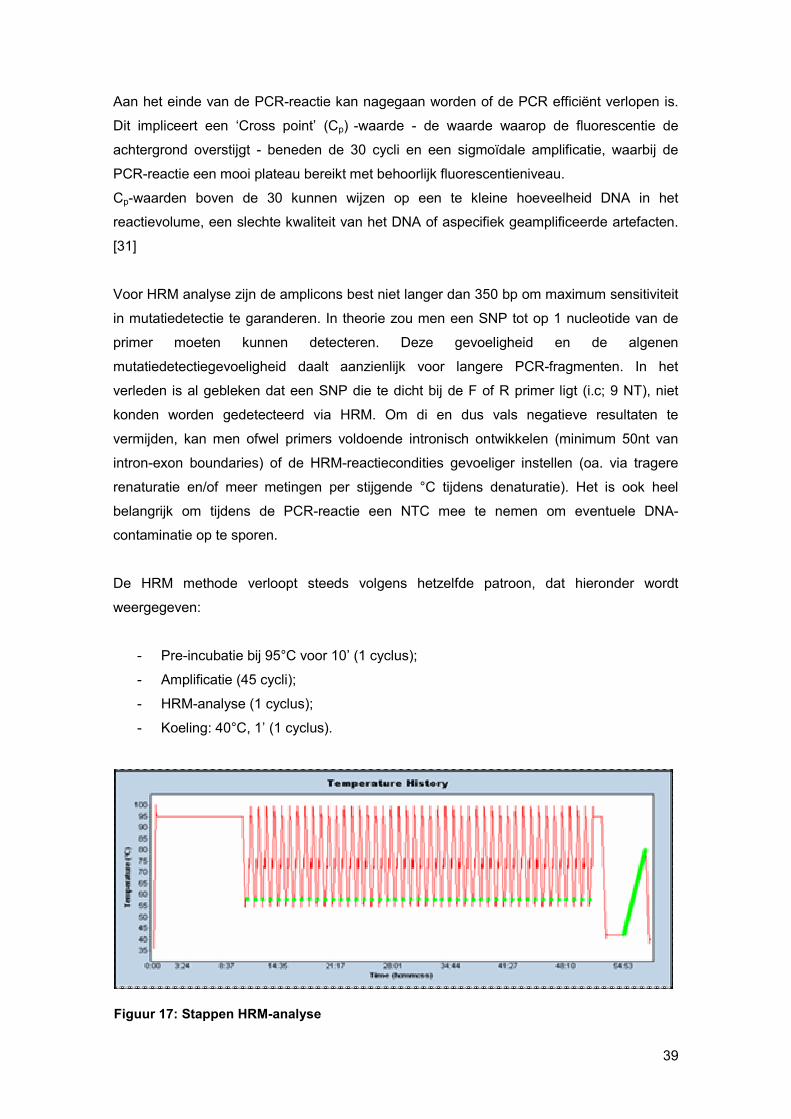
Aan het einde van de PCR-reactie kan nagegaan worden of de PCR efficiënt verlopen is.
Dit impliceert een ‘Cross point’ (Cp) -waarde - de waarde waarop de fluorescentie de
achtergrond overstijgt - beneden de 30 cycli en een sigmoïdale amplificatie, waarbij de
PCR-reactie een mooi plateau bereikt met behoorlijk fluorescentieniveau.
Cp-waarden boven de 30 kunnen wijzen op een te kleine hoeveelheid DNA in het
reactievolume, een slechte kwaliteit van het DNA of aspecifiek geamplificeerde artefacten.
[31]
Voor HRM analyse zijn de amplicons best niet langer dan 350 bp om maximum sensitiviteit
in mutatiedetectie te garanderen. In theorie zou men een SNP tot op 1 nucleotide van de
primer moeten kunnen detecteren. Deze gevoeligheid en de algenen
mutatiedetectiegevoeligheid daalt aanzienlijk voor langere PCR-fragmenten. In het
verleden is al gebleken dat een SNP die te dicht bij de F of R primer ligt (i.c; 9 NT), niet
konden worden gedetecteerd via HRM. Om di en dus vals negatieve resultaten te
vermijden, kan men ofwel primers voldoende intronisch ontwikkelen (minimum 50nt van
intron-exon boundaries) of de HRM-reactiecondities gevoeliger instellen (oa. via tragere
renaturatie en/of meer metingen per stijgende °C tijdens denaturatie). Het is ook heel
belangrijk om tijdens de PCR-reactie een NTC mee te nemen om eventuele DNA-
contaminatie op te sporen.
De HRM methode verloopt steeds volgens hetzelfde patroon, dat hieronder wordt
weergegeven:
- Pre-incubatie bij 95°C voor 10’ (1 cyclus);
- Amplificatie (45 cycli);
- HRM-analyse (1 cyclus);
- Koeling: 40°C, 1’ (1 cyclus).
Figuur 17: Stappen HRM-analyse

40
Tijdens de pre-incubatie op 95°C wordt het Taq pol geactiveerd en denatureert het ds DNA
fragment. In de 2de stap van het proces vindt de amplificatie plaats, die uit 3 stappen
bestaat. De denaturatie-stap van 10’’ op 96°C zorgt ervoor dat zowel het DNA fragment als
de primers ss zijn. Tijdens de annealing-stap van 15’’ zullen de primers binden aan de
complementaire sequenties in het ss DNA fragment. Als laatste vindt de extensie plaats op
72°C voor 15’’. De primers zullen nu verlengd worden aan het 3’-uiteinde.
De 3de stap in het proces is de HRM-analyse. Eerst is er een denaturatie-stap van 1’ op
95°C waarbij de gevormde ds DNA fragmenten denatureren. Bij daling van de temperatuur
tot 40°C worden de homo- en heteroduplexen gevormd (zie verder). Nadien stijgt de
temperatuur snel naar 65°C in 28.4’’ waarna de temperatuur zeer geleidelijk aan klimt tot
96°C. Tijdens deze stap, welke ongeveer 15’ in beslag neemt, wordt de fluorescentie
gemeten. [32]
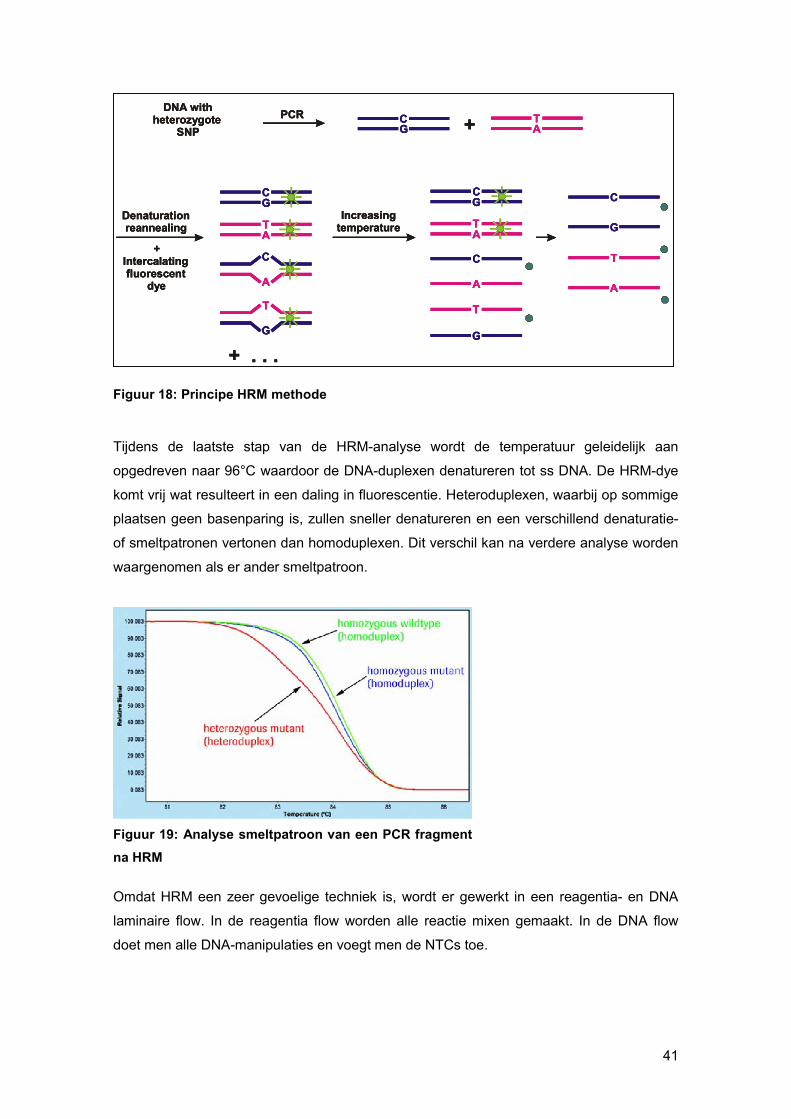
Om minieme verschillen in DNA-sequenties waar te nemen moet men ‘spiken’ met een
controle staal, waarvan de DNA-sequentie gekend is. Van dit controlestaal wordt 5 ng DNA
(1μl van 5 ng/μl) toegevoegd aan de reactie-mix ten opzichte van 20 ng patiënt DNA (4 μl
van 5ng/ μl). Tijdens de denaturatie-stap van de HRM-analyse zullen zowel het ds DNA
van het patiënt staal als van het controle staal denatureren. Tijdens de afkoeling naar 40°C
zullen de ss DNA fragmenten van het patiënten staal en het controle staal ‘ad random’ met
elkaar basenparen. Op die manier worden DNA-duplexen gevormd. Als het patiënt-
amplicon niet verschilt van het controle-amplicon krijgt men enkel de vorming van
homoduplexen. Van zodra patiënt- en controle-amplicon van elkaar verschillen worden
heteroduplexen gevormd.
In de heteroduplexen is complementaire basenparing niet mogelijk op de mutatie of SNP
posities, waardoor de HRM-dye niet kan intercaleren op die plaats.
DNA withheterozygote
SNP
PCR+
+ . . .
T
C
T
T T
T
T
C
C CC
C
G
A
A
A A
AA
G
G G
G
G
Denaturationreannealing
+Intercalatingfluorescent
dye
Increasingtemperature
DNA withheterozygote
SNP
PCR+
+ . . .
T
C
T
T T
T
T
C
C CC
C
G
A
A
A A
AA
G
G G
G
G
Denaturationreannealing
+Intercalatingfluorescent
dye
Increasingtemperature
DNA withheterozygote
SNP
PCR+
+ . . .
T
C
T
T T
T
T
C
C CC
C
G
A
A
A A
AA
G
G G
G
G
Denaturationreannealing
+Intercalatingfluorescent
dye
Increasingtemperature
Tijdens de laatste stap van de HRM-analyse wordt de temperatuur geleidelijk aan
opgedreven naar 96°C waardoor de DNA-duplexen denatureren tot ss DNA. De HRM-dye
komt vrij wat resulteert in een daling in fluorescentie. Heteroduplexen, waarbij op sommige
plaatsen geen basenparing is, zullen sneller denatureren en een verschillend denaturatie-
of smeltpatronen vertonen dan homoduplexen. Dit verschil kan na verdere analyse worden
waargenomen als er ander smeltpatroon.
Figuur 18: Principe HRM methode
Figuur 19: Analyse smeltpatroon van een PCR fragment
41
Omdat HRM een zeer gevoelige techniek is, wordt er gewerkt in een reagentia- en DNA
laminaire flow. In de reagentia flow worden alle reactie mixen gemaakt. In de DNA flow
doet men alle DNA-manipulaties en voegt men de NTCs toe.
na HRM

42
2.7.2. Reagentia
Master Mix 2x (stockconcentratie) → 1x (werkconcentratie) (Roche Diagnostics) bestaande
uit:
- FastStart Taq DNA polymerase (gebruikt bij hot start PCR), i.e., een chemisch
gemodificeerde vorm van het thermostabiele-recombinante Taq DNA polymerase,
die wordt geactiveerd bij 95°C tijdens de pre-incubatie en inactief is bij
temperaturen tot 75°C. Hierdoor worden minder aspecifieke producten gevormd en
stijgt de specificiteit en sensitiviteit van de RT-PCR-reactie bijgevolg;
- 2x reactiebuffer (samenstelling onbekend);
- dNTP mix (dATP, dUTP, dCTP en dGTP);
- High Resolution Melting Dye (Resolight);
- De Master Mix wordt best bij -15°C - 25°C en uit het licht bewaard;
- Vortex de Master Mix zeer goed voor gebruik
MgCl2 25mM (stockconcentratie) → 1,0mM – 3,5mM (werkconcentratie) (De concentratie
MgCl2 in de reactiemix is afhankelijk van exon tot exon. De optimale concentratie moet
experimenteel onderzocht worden);
H2O (een bepaalde hoeveelheid water is nodig om het eindvolume op 20 μl te krijgen);
Primers (Voor deze techniek is specifieke amplificatie zeer belangrijk. Primerconcentratie is
dus best tussen de 0,1-0,3 μM elk. Meestal worden concentraties van 0,2 μM gebruikt).
2.7.3. Protocol
2.7.3.1. Oppuntstelling RT-PCR-reactie
- Maak test-DNA verdunning van 5ng/μl;
- Maak MgCl2-verdunningen;
- Maak reactiemix (steeds overmaat) (zie tabel 4);
- Voeg per rij telkens een andere MgCl2-concentratie toe;
- Verdeel 12.2 μl reactie mix uit in de welletjes voor de NTCs en 5 μL H2O voor de
helft van de NTCS;
- Voeg de juiste hoeveelheid test-DNA toe aan de reactiemix;
- Verdeel 17.2 μl reactiemix uit in welletjes voor het test-DNA;
- Voeg aan de overige NTCs 5 μl H2O toe (in de DNA-laminaire flow);
43
- Sluit de plaat af met de LightCycler® 480 Sealing Foil;
- Draai de plaat kort af (short spin: 10 sec aan 2000 g);
- Zet plaat in LightCycler® 480 (Roche) en start het RT-PCR programma;
- Analyseer de resultaten aan de hand van de LightCycler® 480 Gene Scanning
Software.
Tabel 4: Samenstelling reactiemix HRM
reagentia. stockconc. userconc. 1x
mastermix(x) 2 1 10
25 mM MgCl2 25 2,5 2
H20 1,4
totaal reactiemix 13,4
2,5 μM primermix 2,5 μM 200 nM 1,6
spike DNA 5 ng/µl 1
test DNA 5 ng/µl 4
totaal 20
2.7.3.2. Scanning patiënten
- Maak DNA verdunningen;
- Maak reactiemix (steeds overmaat);
- Verdeel 15 μL reactiemix uit in de welletjes voor de NTCs en 5 μL H2O voor de helft
van de NTCs;
- Voeg juiste hoeveelheid spike DNA toe aan reactiemix;
- Verdeel 16 μL reagentia mix met spike DNA uit in welletjes voor het test-DNA;
- Voeg aan elke well 4 μL patiënt-DNA toe;
- Voeg aan de overige NTCs 5 μl H2O toe (in de DNA-laminarie flow);
- Sluit de plaat af met de LightCycler® 480 Sealing Foil;
- Draai de plaat kort af (short spin: 10 sec aan 2000 g);
- Zet plaat in LightCycler® 480 (Roche) en start het RT-PCR + HRM programma;
- Analyseer de resultaten aan de hand van de LightCycler® 480 Gene Scanning
Software.
2.7.4. Analyse
Nadat het volledige programma doorlopen is, worden de resultaten geanalyseerd met de
LightCycler® 480 Gene Scanning Software.
Bij een oppuntstelling wordt er eerst gekeken bij welke MgCl2-concentratie de beste PCR-
reactie curve gevormd wordt. Elke PCR-reactie heeft namelijk een ideale concentratie aan
MgCl2 nodig aangezien enkel nucleotiden, gecomplexeerd met Mg2+-ionen, kunnen
gebruikt worden als substraat voor de groeiende DNA-keten. Ook het FastStart Taq DNA
pol heeft een bepaalde hoeveelheid MgCl2 nodig om zijn productiviteit en betrouwbaarheid
te vrijwaren. [33]
Nadien wordt via gelelektroforese nagegaan of de amplificatie specifiek was. Wanneer men
positieve NTCs heeft, worden deze enderzijds mee op gel geladen om mogelijke PCR
contaminatie te onderscheiden van mogelijke primer-dimeren.
44
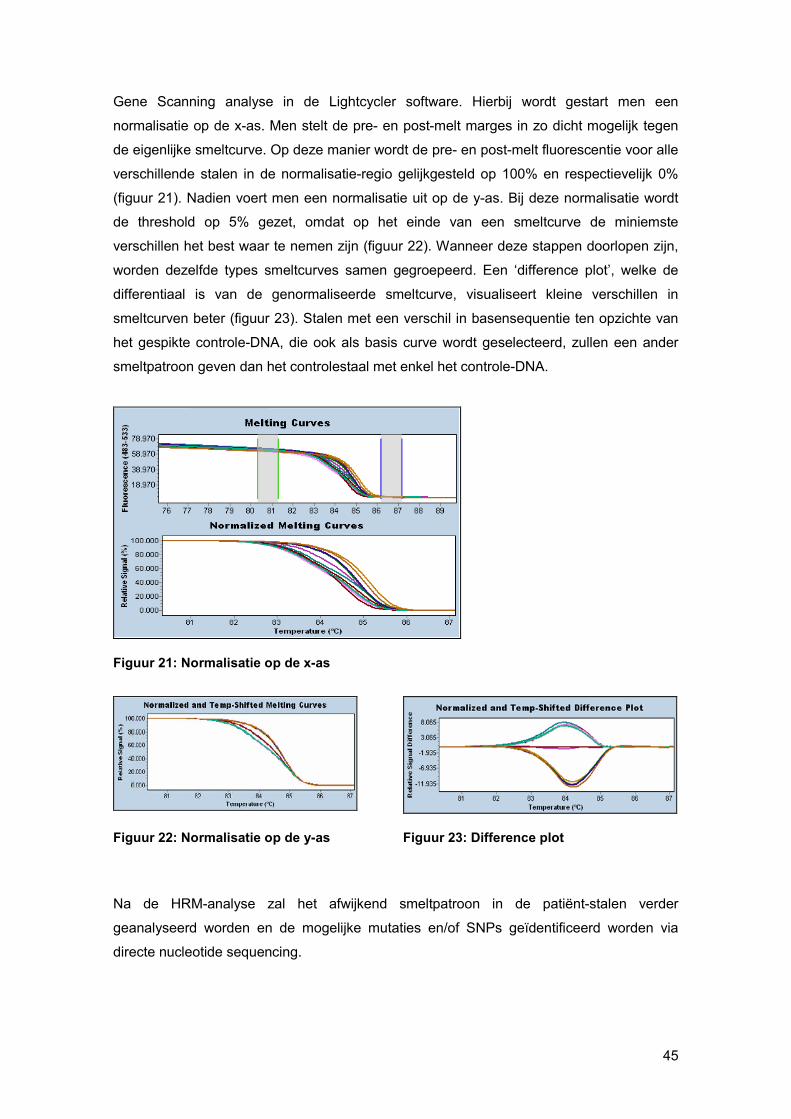
Anderzijds kan men de specificiteit van de RT-PCR-reactie ook controleren via een RT-
PCR smeltcurve met behulp van Tm-calling. Deze Tm-calling is een grafische voorstelling
van de fluorescente veranderingen wanneer ds DNA overgaat naar ss-vorm. Bij deze Tm-
calling kan men nakijken of NTCs, die fluorescent signaal geven, het resultaat zijn van
PCR-contaminatie dan wel primer-dimeren. Figuur 20 geeft een voorbeeld van een Tm-
calling. De grote piek stelt het geamplificeerde DNA-fragment voor. De klein piek geeft de
smeltcurve van primer-dimeren weer. [34]
Wanneer men op de patiënten-stalen een HRM-analyse wil uitvoeren, slecteert men een
Figuur 20: Tm calling
45
Gene Scanning analyse in de Lightcycler software. Hierbij wordt gestart men een
normalisatie op de x-as. Men stelt de pre- en post-melt marges in zo dicht mogelijk tegen
de eigenlijke smeltcurve. Op deze manier wordt de pre- en post-melt fluorescentie voor alle
verschillende stalen in de normalisatie-regio gelijkgesteld op 100% en respectievelijk 0%
(figuur 21). Nadien voert men een normalisatie uit op de y-as. Bij deze normalisatie wordt
de threshold op 5% gezet, omdat op het einde van een smeltcurve de miniemste
verschillen het best waar te nemen zijn (figuur 22). Wanneer deze stappen doorlopen zijn,
worden dezelfde types smeltcurves samen gegroepeerd. Een ‘difference plot’, welke de
differentiaal is van de genormaliseerde smeltcurve, visualiseert kleine verschillen in
smeltcurven beter (figuur 23). Stalen met een verschil in basensequentie ten opzichte van
het gespikte controle-DNA, die ook als basis curve wordt geselecteerd, zullen een ander
smeltpatroon geven dan het controlestaal met enkel het controle-DNA.
Na de HRM-analyse zal het afwijkend smeltpatroon in de patiënt-stalen verder
geanalyseerd worden en de mogelijke mutaties en/of SNPs geïdentificeerd worden via
directe nucleotide sequencing.
Figuur 21: Normalisatie op de x-as
Figuur 22: Normalisatie op de y-as Figuur 23: Difference plot
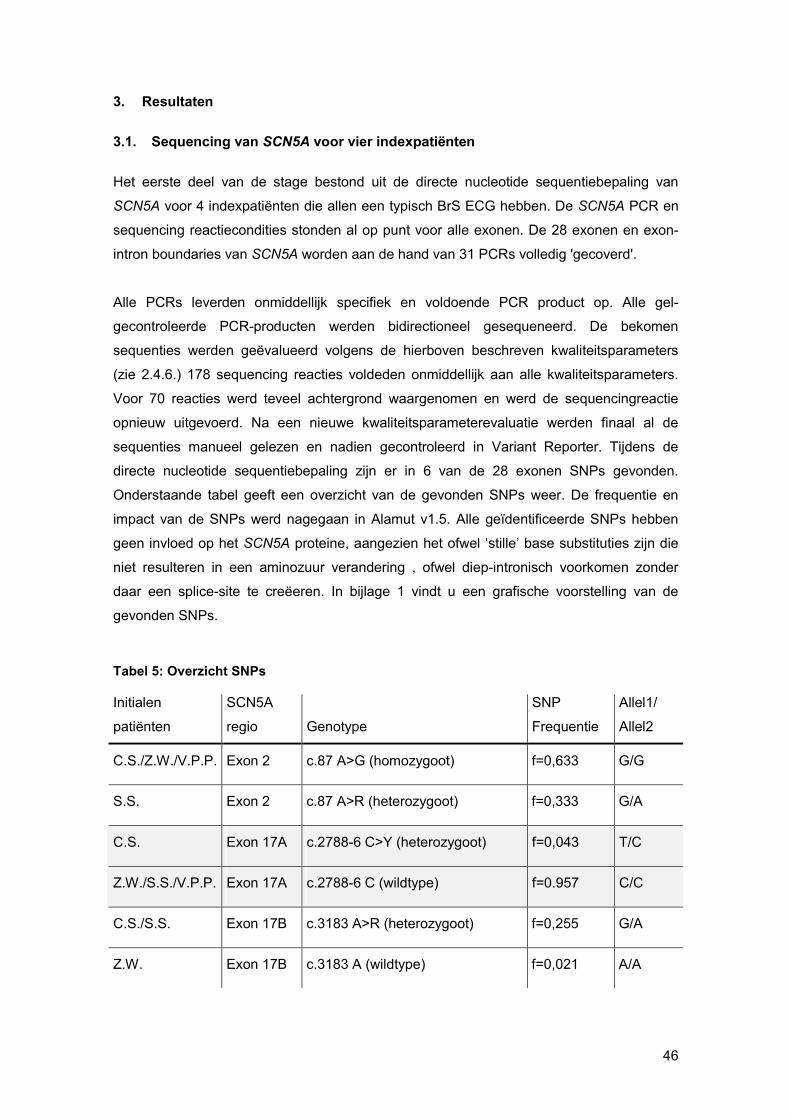
46
3. Resultaten
3.1. Sequencing van SCN5A voor vier indexpatiënten
Het eerste deel van de stage bestond uit de directe nucleotide sequentiebepaling van
SCN5A voor 4 indexpatiënten die allen een typisch BrS ECG hebben. De SCN5A PCR en
sequencing reactiecondities stonden al op punt voor alle exonen. De 28 exonen en exon-
intron boundaries van SCN5A worden aan de hand van 31 PCRs volledig 'gecoverd'.
Alle PCRs leverden onmiddellijk specifiek en voldoende PCR product op. Alle gel-
gecontroleerde PCR-producten werden bidirectioneel gesequeneerd. De bekomen
sequenties werden geëvalueerd volgens de hierboven beschreven kwaliteitsparameters
(zie 2.4.6.) 178 sequencing reacties voldeden onmiddellijk aan alle kwaliteitsparameters.
Voor 70 reacties werd teveel achtergrond waargenomen en werd de sequencingreactie
opnieuw uitgevoerd. Na een nieuwe kwaliteitsparameterevaluatie werden finaal al de
sequenties manueel gelezen en nadien gecontroleerd in Variant Reporter. Tijdens de
directe nucleotide sequentiebepaling zijn er in 6 van de 28 exonen SNPs gevonden.
Onderstaande tabel geeft een overzicht van de gevonden SNPs weer. De frequentie en
impact van de SNPs werd nagegaan in Alamut v1.5. Alle geïdentificeerde SNPs hebben
geen invloed op het SCN5A proteine, aangezien het ofwel ‘stille’ base substituties zijn die
niet resulteren in een aminozuur verandering , ofwel diep-intronisch voorkomen zonder
daar een splice-site te creëeren. In bijlage 1 vindt u een grafische voorstelling van de
gevonden SNPs.
Tabel 5: Overzicht SNPs
Initialen
patiënten
SCN5A
regio Genotype
SNP
Frequentie
Allel1/
Allel2
C.S./Z.W./V.P.P. Exon 2 c.87 A>G (homozygoot) f=0,633 G/G
S.S. Exon 2 c.87 A>R (heterozygoot) f=0,333 G/A
C.S. Exon 17A c.2788-6 C>Y (heterozygoot) f=0,043 T/C
Z.W./S.S./V.P.P. Exon 17A c.2788-6 C (wildtype) f=0.957 C/C
C.S./S.S. Exon 17B c.3183 A>R (heterozygoot) f=0,255 G/A
Z.W. Exon 17B c.3183 A (wildtype) f=0,021 A/A
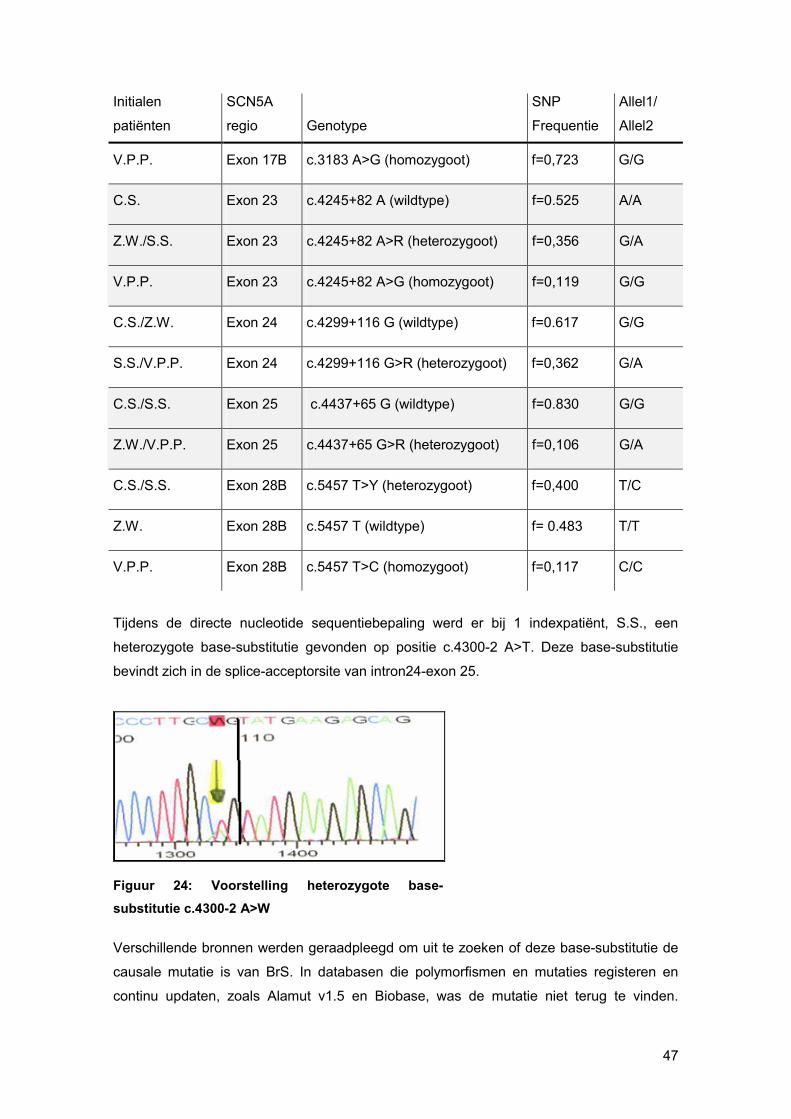
47
Initialen
patiënten
SCN5A
regio Genotype
SNP
Frequentie
Allel1/
Allel2
V.P.P. Exon 17B c.3183 A>G (homozygoot) f=0,723 G/G
C.S. Exon 23 c.4245+82 A (wildtype) f=0.525 A/A
Z.W./S.S. Exon 23 c.4245+82 A>R (heterozygoot) f=0,356 G/A
V.P.P. Exon 23 c.4245+82 A>G (homozygoot) f=0,119 G/G
C.S./Z.W. Exon 24 c.4299+116 G (wildtype) f=0.617 G/G
S.S./V.P.P. Exon 24 c.4299+116 G>R (heterozygoot) f=0,362 G/A
C.S./S.S. Exon 25 c.4437+65 G (wildtype) f=0.830 G/G
Z.W./V.P.P. Exon 25 c.4437+65 G>R (heterozygoot) f=0,106 G/A
C.S./S.S. Exon 28B c.5457 T>Y (heterozygoot) f=0,400 T/C
Z.W. Exon 28B c.5457 T (wildtype) f= 0.483 T/T
V.P.P. Exon 28B c.5457 T>C (homozygoot) f=0,117 C/C
Tijdens de directe nucleotide sequentiebepaling werd er bij 1 indexpatiënt, S.S., een
heterozygote base-substitutie gevonden op positie c.4300-2 A>T. Deze base-substitutie
bevindt zich in de splice-acceptorsite van intron24-exon 25.
Verschillende bronnen werden geraadpleegd om uit te zoeken of deze base-substitutie de
causale mutatie is van BrS. In databasen die polymorfismen en mutaties registeren en
continu updaten, zoals Alamut v1.5 en Biobase, was de mutatie niet terug te vinden.
Figuur 24: Voorstelling heterozygote base-
substitutie c.4300-2 A>W
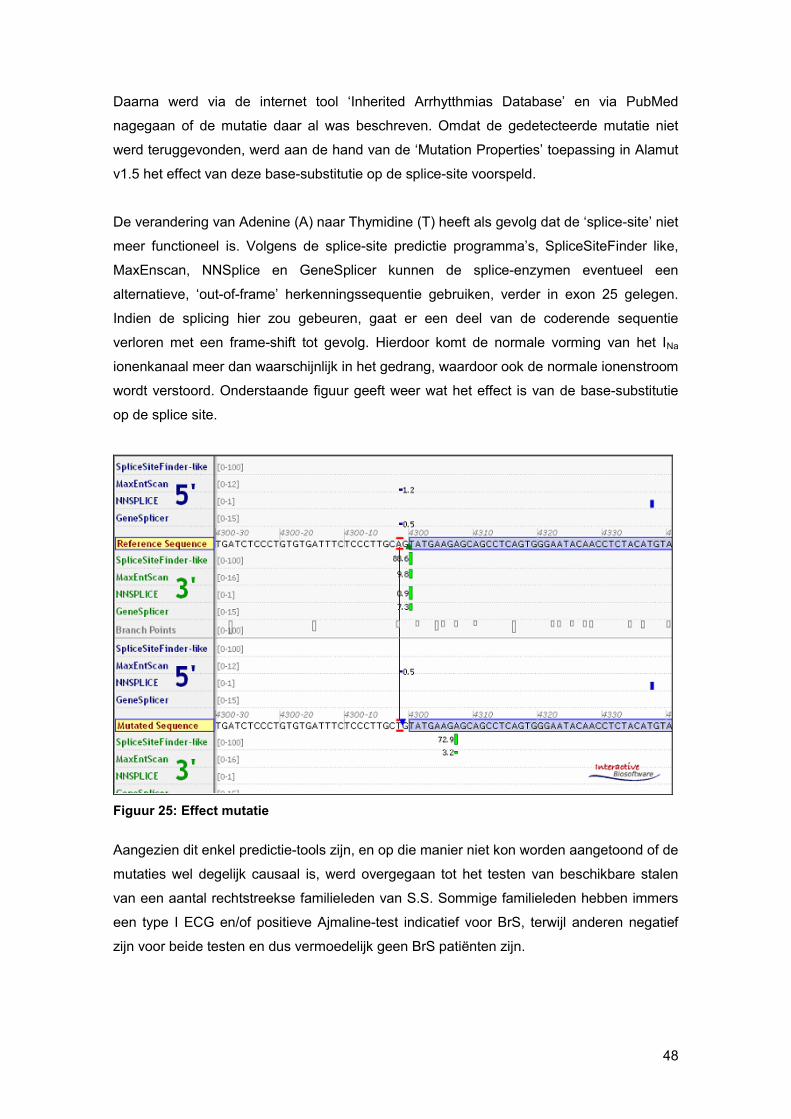
Daarna werd via de internet tool ‘Inherited Arrhytthmias Database’ en via PubMed
nagegaan of de mutatie daar al was beschreven. Omdat de gedetecteerde mutatie niet
werd teruggevonden, werd aan de hand van de ‘Mutation Properties’ toepassing in Alamut
v1.5 het effect van deze base-substitutie op de splice-site voorspeld.
De verandering van Adenine (A) naar Thymidine (T) heeft als gevolg dat de ‘splice-site’ niet
meer functioneel is. Volgens de splice-site predictie programma’s, SpliceSiteFinder like,
MaxEnscan, NNSplice en GeneSplicer kunnen de splice-enzymen eventueel een
alternatieve, ‘out-of-frame’ herkenningssequentie gebruiken, verder in exon 25 gelegen.
Indien de splicing hier zou gebeuren, gaat er een deel van de coderende sequentie
verloren met een frame-shift tot gevolg. Hierdoor komt de normale vorming van het INa
ionenkanaal meer dan waarschijnlijk in het gedrang, waardoor ook de normale ionenstroom
wordt verstoord. Onderstaande figuur geeft weer wat het effect is van de base-substitutie
op de splice site.
48
Aangezien dit enkel predictie-tools zijn, en op die manier niet kon worden aangetoond of de
mutaties wel degelijk causaal is, werd overgegaan tot het testen van beschikbare stalen
van een aantal rechtstreekse familieleden van S.S. Sommige familieleden hebben immers
een type I ECG en/of positieve Ajmaline-test indicatief voor BrS, terwijl anderen negatief
zijn voor beide testen en dus vermoedelijk geen BrS patiënten zijn.
Figuur 25: Effect mutatie
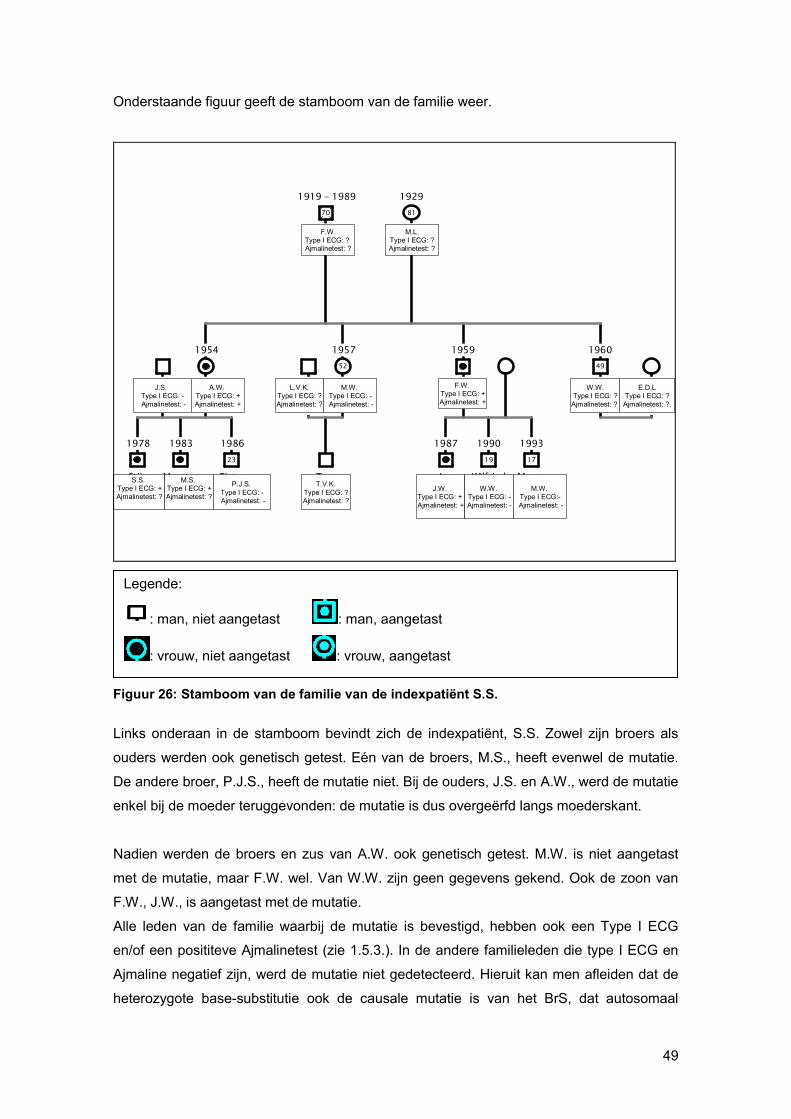
Onderstaande figuur geeft de stamboom van de familie weer.
1919 - 1989
Ferdinand
Weyns
70
1929
Maria
Leppens
81
1954
Anita
Weyns
1957
Martine
Weyns
52
1959
Fernand
Weyns
1960
Werner
Weyns
49
Jef
Sterckx
1978
Stijn
Sterckx
1983
Maarten
Sterckx
1986
Pieter
Jan
Sterckx
23
Luk
Van
Kets
Tom
Van
Kets
1987
Jan
Weyns
1990
Wilfried
Weyns
19
1993
Marco
Weyns
17
Erna
De
Loose
F.W.Type I ECG: ?Ajmalinetest: ?
M.L.Type I ECG: ?Ajmalinetest: ?
J.S.Type I ECG: -Ajmalinetest: -
A.W.Type I ECG: +Ajmalinetest: +
S.S.Type I ECG: +Ajmalinetest: ?
M.S.Type I ECG: +Ajmalinetest: ?
P.J.S.Type I ECG: -Ajmalinetest: -
L.V.K.Type I ECG: ?Ajmalinetest: ?
M.W.Type I ECG: -Ajmalinetest: -
F.W.Type I ECG: +Ajmalinetest: +
J.W.Type I ECG: +Ajmalinetest: +
T.V.K.Type I ECG: ?Ajmalinetest: ?
W.W.Type I ECG: -Ajmalinetest: -
M.W.Type I ECG:-Ajmalinetest: -
W.W.Type I ECG: ?Ajmalinetest: ?
E.D.LType I ECG: ?Ajmalinetest: ?.
Legende:
: man, niet aangetast : man, aangetast
: vrouw, niet aangetast : vrouw, aangetast
49
Links onderaan in de stamboom bevindt zich de indexpatiënt, S.S. Zowel zijn broers als
ouders werden ook genetisch getest. Eén van de broers, M.S., heeft evenwel de mutatie.
De andere broer, P.J.S., heeft de mutatie niet. Bij de ouders, J.S. en A.W., werd de mutatie
enkel bij de moeder teruggevonden: de mutatie is dus overgeërfd langs moederskant.
Nadien werden de broers en zus van A.W. ook genetisch getest. M.W. is niet aangetast
met de mutatie, maar F.W. wel. Van W.W. zijn geen gegevens gekend. Ook de zoon van
F.W., J.W., is aangetast met de mutatie.
Alle leden van de familie waarbij de mutatie is bevestigd, hebben ook een Type I ECG
en/of een posititeve Ajmalinetest (zie 1.5.3.). In de andere familieleden die type I ECG en
Ajmaline negatief zijn, werd de mutatie niet gedetecteerd. Hieruit kan men afleiden dat de
heterozygote base-substitutie ook de causale mutatie is van het BrS, dat autosomaal
Figuur 26: Stamboom van de familie van de indexpatiënt S.S.

50
dominant wordt overgeërfd.
3.2. SCN5A RT-PCR oppuntstelling & HRM pre-validatie
3.2.1. SCN5A RT-PCR oppuntstelling
Een HRM analyse kan maar efficiënt verlopen als een optimaal RT-PCR product wordt
gegenereerd. Hierbij wordt gestreefd naar sensitieve en specifieke RT-PCRs. Sensitiviteit
wordt geëvalueerd op basis van bekomen Cp-waarden < 30 en fluorescentie. De
specificiteit wordt beoordeeld op basis van de smeltcurve (Tm-calling) en agarose
gelelektroforese.
De HRM analyse voor 28 exonen van SCN5A werd in eerste instantie op punt gesteld door
een vorige student met primers in-house ontwikkeld via de ‘Primer Blast’ software. Onder
verschillende uitgeteste condities, bleek op basis van de best bekomen sensitiviteit en
specificiteit, dat voor 13 exonen beter nieuwe primers werden ontwikkeld.
In dit werk werden de nieuwe oppuntstellingen uitgevoerd voor de RT-PCRs op 13 SCN5A
exonen, wat overeenkomt met 14 RT-PCR-reacties (nl. exon 7, 9, 12b, 14, 16a, 17a, 17b,
19, 20, 21 22, 25, 27, 28e).
Aangezien intussen een publicatie over SCN5A HRM analyse was verschenen in de
literatuur [35], werden de primersequenties voor de oppuntstelling van de overige exonen,
na controle op correctheid, overgenomen uit het artikel.
Voor de oppuntstelling van de RT-PCR-reactie voorafgaand aan de HRM-analyse van
SCN5A werkt men steeds via een bepaald patroon.
De eerste stap in de oppuntstelling is het uitvoeren van een MgCl2-titratie van 1,5mM tot
3,5mM met een interval van 0,5mM voor elke RT-PCR-reactie. De MgCl2-concentratie is
een belangrijke component in een RT-PCR-reactie. Enkel dNTPs, gecomplexeerd met
MgCl2 worden ingebouwd in de groeiende matrijs. Ook het Taq pol heeft een welbepaalde
concentratie aan MgCl2 nodig om zijn polymerase activiteit ten opzichte van de DNA-matrijs
ideaal uit te voeren.
Er werd gestart met een ‘touchdown’ Ta gaande van 70°C→60°C met een daling van
0,5°C/cyclus. Afhankelijk van het verloop en de specificiteit van de RT-PCR-reactie werden
er verdere stappen ondernomen. Bijlage 2 bevat een overzicht van de verschillende
stappen in de oppuntstelling van de RT-PCR-reactie voor SCN5A.
Het te doorlopen stappenplan wordt aan de hand van enkele PCRs voor exonen 16 (16a),
17 (17a en 17b) en 19 hieronder nader verklaard.
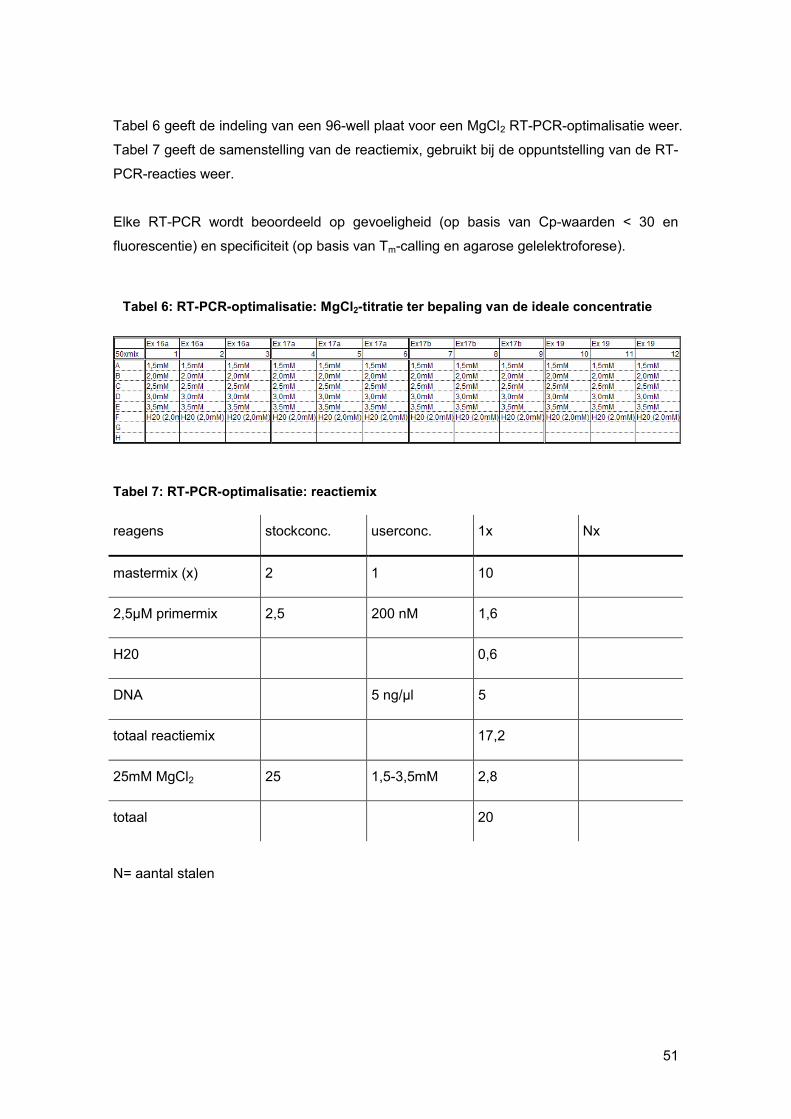
Tabel 6 geeft de indeling van een 96-well plaat voor een MgCl2 RT-PCR-optimalisatie weer.
Tabel 7 geeft de samenstelling van de reactiemix, gebruikt bij de oppuntstelling van de RT-
PCR-reacties weer.
Elke RT-PCR wordt beoordeeld op gevoeligheid (op basis van Cp-waarden < 30 en
fluorescentie) en specificiteit (op basis van Tm-calling en agarose gelelektroforese).
51
Tabel 7: RT-PCR-optimalisatie: reactiemix
reagens stockconc. userconc. 1x Nx
mastermix (x) 2 1 10
2,5μM primermix 2,5 200 nM 1,6
H20 0,6
DNA 5 ng/μl 5
totaal reactiemix 17,2
25mM MgCl2 25 1,5-3,5mM 2,8
totaal 20
N= aantal stalen
Tabel 6: RT-PCR-optimalisatie: MgCl2-titratie ter bepaling van de ideale concentratie
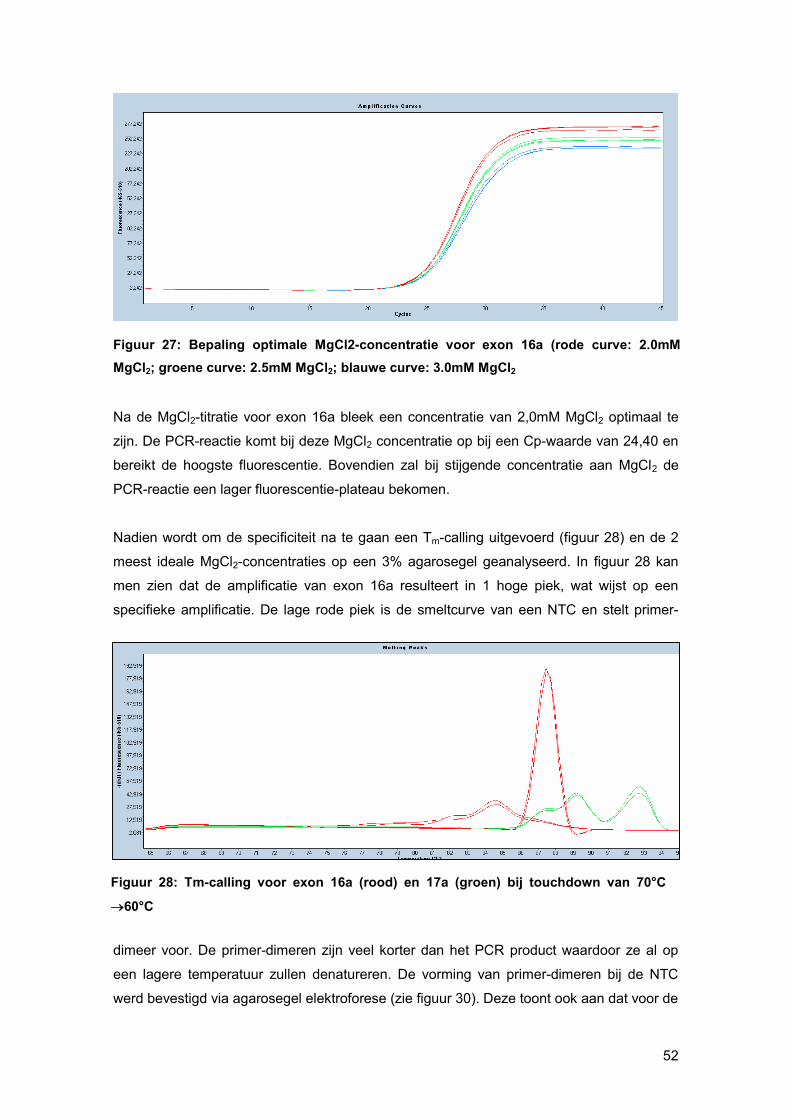
F
52
Na de MgCl2-titratie voor exon 16a bleek een concentratie van 2,0mM MgCl2 optimaal te
zijn. De PCR-reactie komt bij deze MgCl2 concentratie op bij een Cp-waarde van 24,40 en
bereikt de hoogste fluorescentie. Bovendien zal bij stijgende concentratie aan MgCl2 de
PCR-reactie een lager fluorescentie-plateau bekomen.
Nadien wordt om de specificiteit na te gaan een Tm-calling uitgevoerd (figuur 28) en de 2
meest ideale MgCl2-concentraties op een 3% agarosegel geanalyseerd. In figuur 28 kan
men zien dat de amplificatie van exon 16a resulteert in 1 hoge piek, wat wijst op een
specifieke amplificatie. De lage rode piek is de smeltcurve van een NTC en stelt primer-
dimeer voor. De primer-dimeren zijn veel korter dan het PCR product waardoor ze al op
een lagere temperatuur zullen denatureren. De vorming van primer-dimeren bij de NTC
werd bevestigd via agarosegel elektroforese (zie figuur 30). Deze toont ook aan dat voor de
Figuur 27: Bepaling optimale MgCl2-concentratie voor exon 16a (rode curve: 2.0mM
MgCl2; groene curve: 2.5mM MgCl2; blauwe curve: 3.0mM MgCl2
iguur 28: Tm-calling voor exon 16a (rood) en 17a (groen) bij touchdown van 70°C
60°C
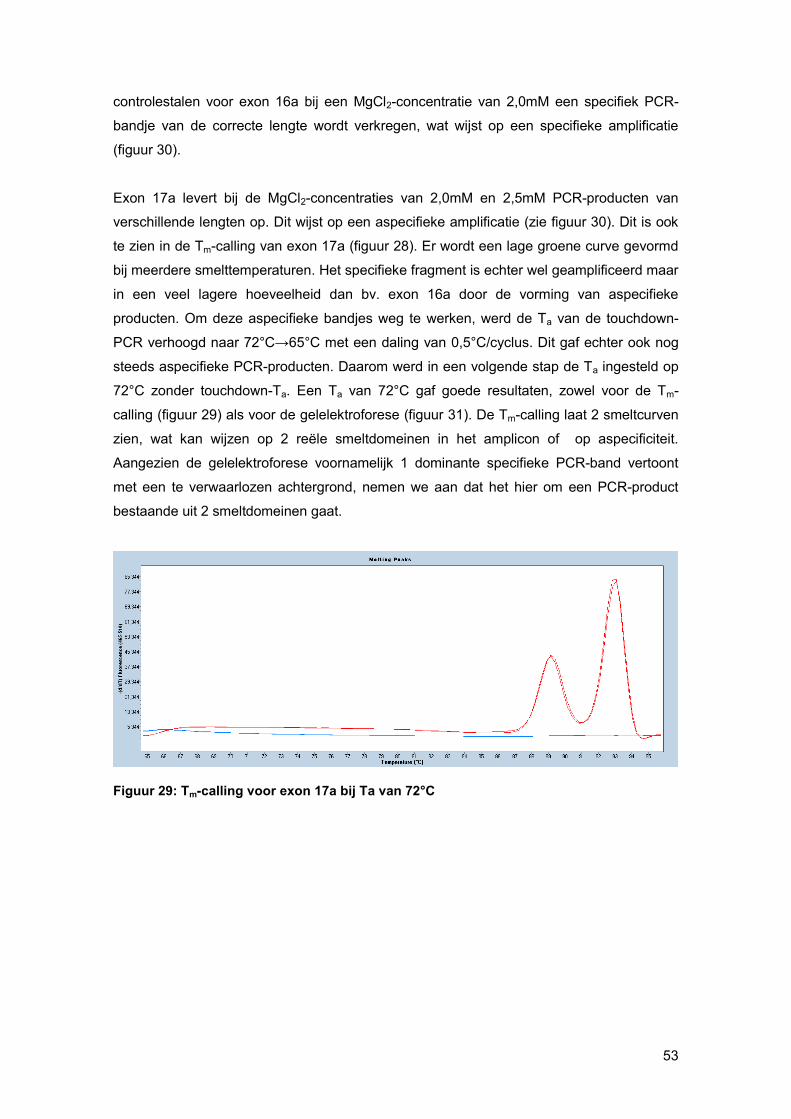
53
controlestalen voor exon 16a bij een MgCl2-concentratie van 2,0mM een specifiek PCR-
bandje van de correcte lengte wordt verkregen, wat wijst op een specifieke amplificatie
(figuur 30).
Exon 17a levert bij de MgCl2-concentraties van 2,0mM en 2,5mM PCR-producten van
verschillende lengten op. Dit wijst op een aspecifieke amplificatie (zie figuur 30). Dit is ook
te zien in de Tm-calling van exon 17a (figuur 28). Er wordt een lage groene curve gevormd
bij meerdere smelttemperaturen. Het specifieke fragment is echter wel geamplificeerd maar
in een veel lagere hoeveelheid dan bv. exon 16a door de vorming van aspecifieke
producten. Om deze aspecifieke bandjes weg te werken, werd de Ta van de touchdown-
PCR verhoogd naar 72°C→65°C met een daling van 0,5°C/cyclus. Dit gaf echter ook nog
steeds aspecifieke PCR-producten. Daarom werd in een volgende stap de Ta ingesteld op
72°C zonder touchdown-Ta. Een Ta van 72°C gaf goede resultaten, zowel voor de Tm-
calling (figuur 29) als voor de gelelektroforese (figuur 31). De Tm-calling laat 2 smeltcurven
zien, wat kan wijzen op 2 reële smeltdomeinen in het amplicon of op aspecificiteit.
Aangezien de gelelektroforese voornamelijk 1 dominante specifieke PCR-band vertoont
met een te verwaarlozen achtergrond, nemen we aan dat het hier om een PCR-product
bestaande uit 2 smeltdomeinen gaat.
Figuur 29: Tm-calling voor exon 17a bij Ta van 72°C

54
Lo
g2
ladd
er
Ex16
a2
.0m
M
Ex16
a2
.5m
M
Ex16
aN
TC
Ex17
a2
.0m
M
Ex17
a2
.5m
M
MW
5la
dd
er
Ex17
a2
.0m
M
Ex17
a2
.5m
M
De RT-PCR-reactie van de 2 laatste RT-PCRs op de 96-well plaat, exon 17b en exon 19
kwam veel te laat op. Hierdoor werd de PCR plateau fase niet meer bereikt, wat cruciaal is
voor een betrouwbare HRM-analyse (zie figuur 31). Dit kan eventueel te wijten zijn aan het
feit dat de touchdown Ta 70°C→60°C te hoog is voor de primers om hun complementaire
sequentie te vinden. Vervolgens wordt de touchdown Ta verlaagd naar 65°C→55°C met
een daling van 0,5°C/cyclus.
Figuur 30: Agarosegel-elektroforese van
touchdown PCR Ta 70°C 60°C
Figuur 31: Agarosegel-elektroforese van
touchdown PCR Ta 72°C
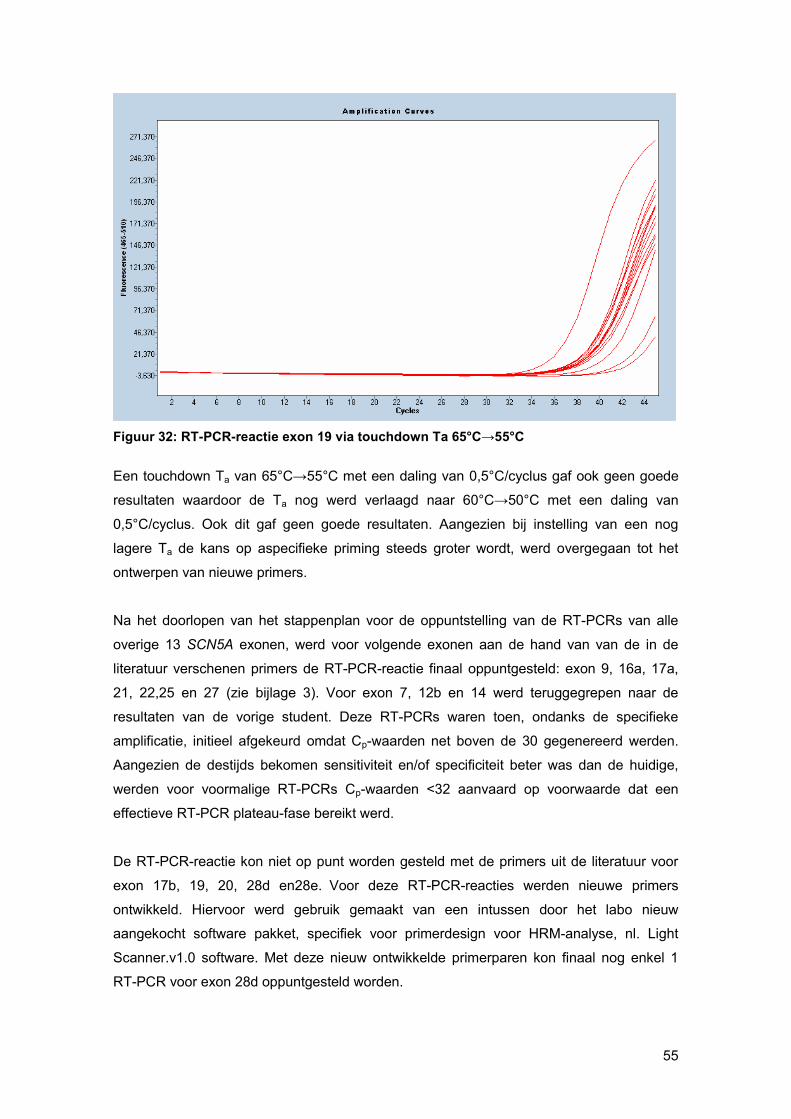
55
Een touchdown Ta van 65°C→55°C met een daling van 0,5°C/cyclus gaf ook geen goede
resultaten waardoor de Ta nog werd verlaagd naar 60°C→50°C met een daling van
0,5°C/cyclus. Ook dit gaf geen goede resultaten. Aangezien bij instelling van een nog
lagere Ta de kans op aspecifieke priming steeds groter wordt, werd overgegaan tot het
ontwerpen van nieuwe primers.
Na het doorlopen van het stappenplan voor de oppuntstelling van de RT-PCRs van alle
overige 13 SCN5A exonen, werd voor volgende exonen aan de hand van van de in de
literatuur verschenen primers de RT-PCR-reactie finaal oppuntgesteld: exon 9, 16a, 17a,
21, 22,25 en 27 (zie bijlage 3). Voor exon 7, 12b en 14 werd teruggegrepen naar de
resultaten van de vorige student. Deze RT-PCRs waren toen, ondanks de specifieke
amplificatie, initieel afgekeurd omdat Cp-waarden net boven de 30 gegenereerd werden.
Aangezien de destijds bekomen sensitiviteit en/of specificiteit beter was dan de huidige,
werden voor voormalige RT-PCRs Cp-waarden <32 aanvaard op voorwaarde dat een
effectieve RT-PCR plateau-fase bereikt werd.
De RT-PCR-reactie kon niet op punt worden gesteld met de primers uit de literatuur voor
exon 17b, 19, 20, 28d en28e. Voor deze RT-PCR-reacties werden nieuwe primers
ontwikkeld. Hiervoor werd gebruik gemaakt van een intussen door het labo nieuw
aangekocht software pakket, specifiek voor primerdesign voor HRM-analyse, nl. Light
Scanner.v1.0 software. Met deze nieuw ontwikkelde primerparen kon finaal nog enkel 1
RT-PCR voor exon 28d oppuntgesteld worden.
Figuur 32: RT-PCR-reactie exon 19 via touchdown Ta 65°C→55°C

56
In bijlage 3 vindt men een overzicht van de ideale condities voor elk exon. 25 RT-PCR-
reacties waren al oppuntgesteld door een vorige student. 7 nieuwe RT-PCR-reacties
werden in dit werk oppuntgesteld met de in de literatuur beschreven primers. Voor 6 RT-
PCR-reacties (exon 17b, 19, 19bis, 20, 28d en 28e) werden nieuwe primers ontwikkeld,
waarbij enkel voor RT-PCR 28d aanvaardbare resultaten bekomen werden. Voor de
overige 4 RT-PCRs werd na 3 tot 4 pogingen van primerdesign en RT-PCR optimalisatie
geopteerd om geen gen-scanning via HRM-analyse uit te voeren voorafgaand aan directe
nucleotide sequencing.
3.2.2. SCN5A HRM pre-validatie exon scanning
Tijdens de directe nucleotidesequencing van SCN5A op de vier indexpatiënten uit 3.1.,
werden 7 SNPs gevonden in 6 exonen. Om de gevoeligheid van de SCN5A HRM-analyse
te testen, werd nagegaan of deze SNPs ook wel degelijk konden gedetecteerd worden met
de oppuntgestelde RT-PCRs en HRM-analyse. Aldus werden de DNA-stalen van deze 4
indexpatiënten onderworpen aan een exon-specifieke HRM-analyse, met name enkel voor
de 6 hoger beschreven exonen (zie tabel 5). De DNA-stalen van de patiënten en een
controlestal werden steeds in duplo getest en gespiked met controlestaal.
3.2.2.1. Exon 2: SNP c. 87 A>G
Figuur 33 toont de 2 vormen van de SNP. De rode curves stellen de heterozygote SNP
detectie voor. De blauwe curves, waaronder ook het controle DNA, vertegenwoordigen de
homozygote SNP. Hieruit kan men besluiten dat de HRM-analyse deze SNP kan
detecteren. De aanwezigheid van deze SNP heeft ook geen invloed op de correcte
eiwitvorming, aangezien het een synonieme basesubstitutie is, die niet resulteert in een
aminozuurverandering volgens Alamut en de internet tool ‘Inherited Arrhytthmias
Database’.
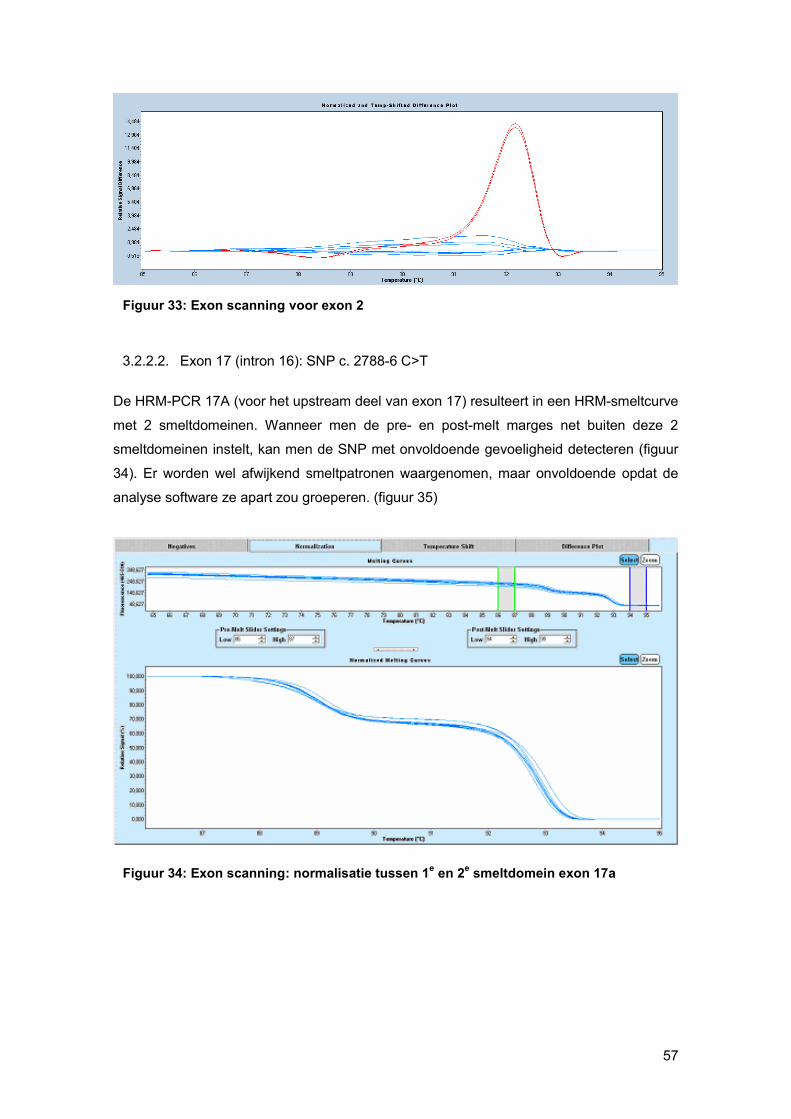
57
3.2.2.2. Exon 17 (intron 16): SNP c. 2788-6 C>T
De HRM-PCR 17A (voor het upstream deel van exon 17) resulteert in een HRM-smeltcurve
met 2 smeltdomeinen. Wanneer men de pre- en post-melt marges net buiten deze 2
smeltdomeinen instelt, kan men de SNP met onvoldoende gevoeligheid detecteren (figuur
34). Er worden wel afwijkend smeltpatronen waargenomen, maar onvoldoende opdat de
analyse software ze apart zou groeperen. (figuur 35)
Figuur 34: Exon scanning: normalisatie tussen 1e
en 2e
smeltdomein exon 17a
Figuur 33: Exon scanning voor exon 2
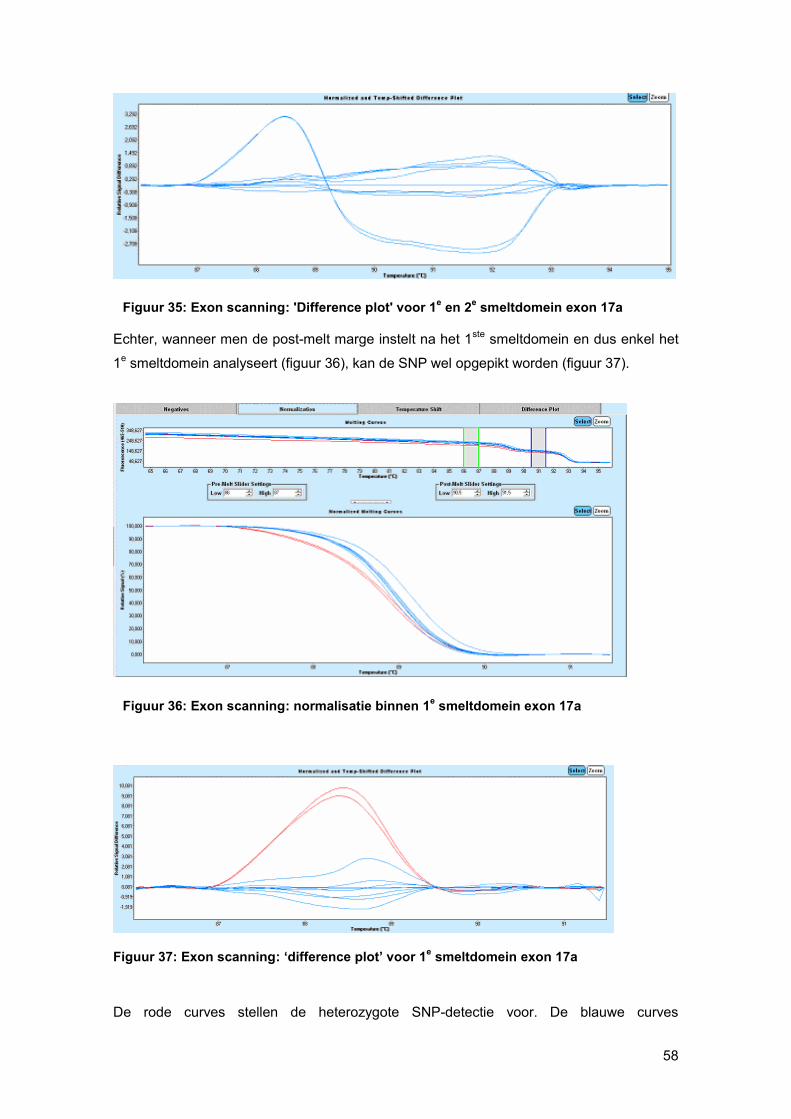
58
Echter, wanneer men de post-melt marge instelt na het 1ste smeltdomein en dus enkel het
1e smeltdomein analyseert (figuur 36), kan de SNP wel opgepikt worden (figuur 37).
Figuur 37: Exon scanning: ‘difference plot’ voor 1e
smeltdomein exon 17a
De rode curves stellen de heterozygote SNP-detectie voor. De blauwe curves
Figuur 35: Exon scanning: 'Difference plot' voor 1e
en 2e
smeltdomein exon 17a
Figuur 36: Exon scanning: normalisatie binnen 1e
smeltdomein exon 17a

59
vertegenwoordigen de wildtypes.
Deze SNP staat niet beschreven in de ‘Inherited Arrhytthmias Database’. Volgens Alamut
is het een gekende SNP die geen invloed heeft op de splicing.
3.2.2.3. Exon 17: SNP c. 3183 A>G
Voor het B-deel van exon 17 kon geen HRM-analyse uitgevoerd worden, aangezien er
geen optimaal primerpaar gevonden werd, waardoor de RT-PCR-reactie niet
geoptimaliseerd werd. Voor deel B moet men direct overgaan tot een directe nucleotide
sequencing. Volgens de internet tool ‘Inherited Arrhytthmias Database’ heeft deze
synonieme SNP geen effect op de correcte vorming van het eiwit.
3.2.2.4. Exon 23 (intron 23): SNP c. 4245+82 A>G
Tijdens de directe nucleotidesequencing van exon 23 werd bij de 4 indexpatiënten de
homozygote en heterozygote vorm van de SNP alsook wildtype op positie c.4245+82
teruggevonden. Tijdens de HRM-analyse konden deze vormen niet van elkaar gescheiden
worden wanneer men de pre- en post melt marge instelde voor en na de 2 smeltdomeinen.
Wanneer de pre melt marge werd ingesteld tussen de 2 smeltdomeinen en dus na het 1e
smeltdomein, slaagde men er wel in de verschillende vormen van de SNP op te sporen
(figuur 38 ). De groene en rode curves stellen respectievelijk de homozygote en
heterozygote SNP voor. Het controle DNA werd als basislijn geselecteerd en vormt samen
met de wildtypen de blauwe curven (figuur 39).
Volgens Alamut 1.5 en de ‘Inherited Arrhytthmias Database’ heeft deze SNP geen effect
op de correcte vorming van het eiwit.
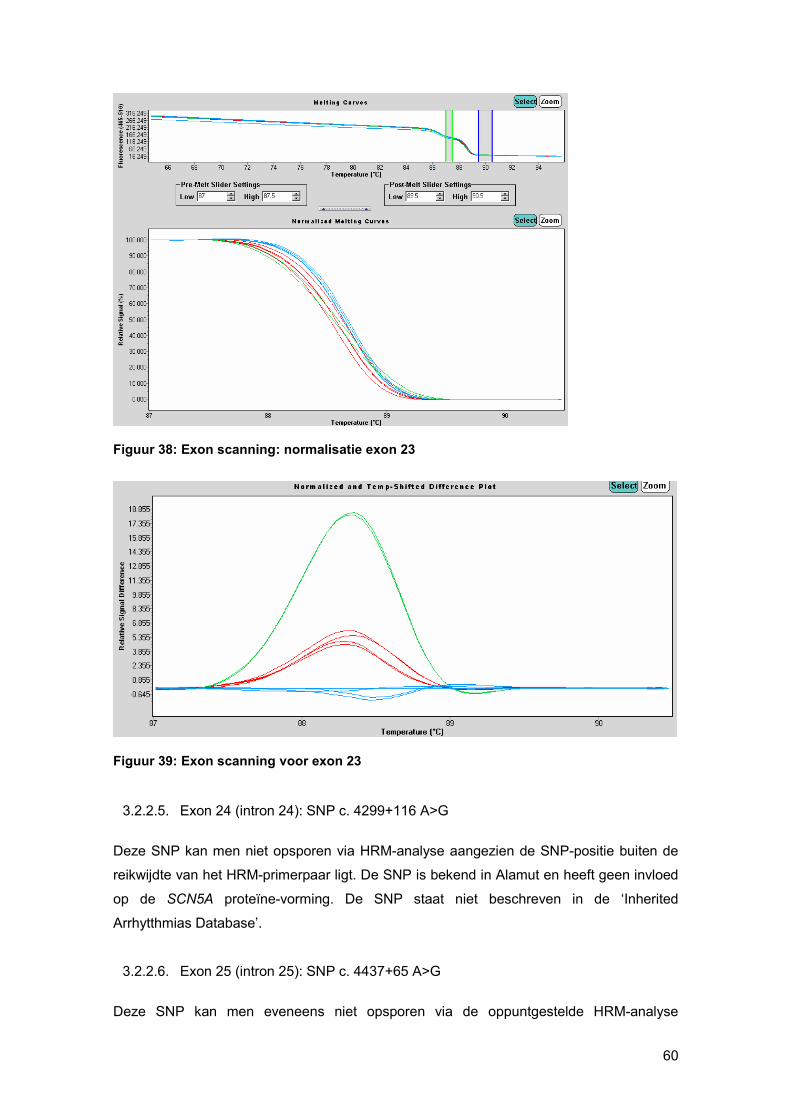
Figuur 38: Exon scanning: normalisatie exon 23
60
3.2.2.5. Exon 24 (intron 24): SNP c. 4299+116 A>G
Deze SNP kan men niet opsporen via HRM-analyse aangezien de SNP-positie buiten de
reikwijdte van het HRM-primerpaar ligt. De SNP is bekend in Alamut en heeft geen invloed
op de SCN5A proteïne-vorming. De SNP staat niet beschreven in de ‘Inherited
Arrhytthmias Database’.
3.2.2.6. Exon 25 (intron 25): SNP c. 4437+65 A>G
Deze SNP kan men eveneens niet opsporen via de oppuntgestelde HRM-analyse
Figuur 39: Exon scanning voor exon 23
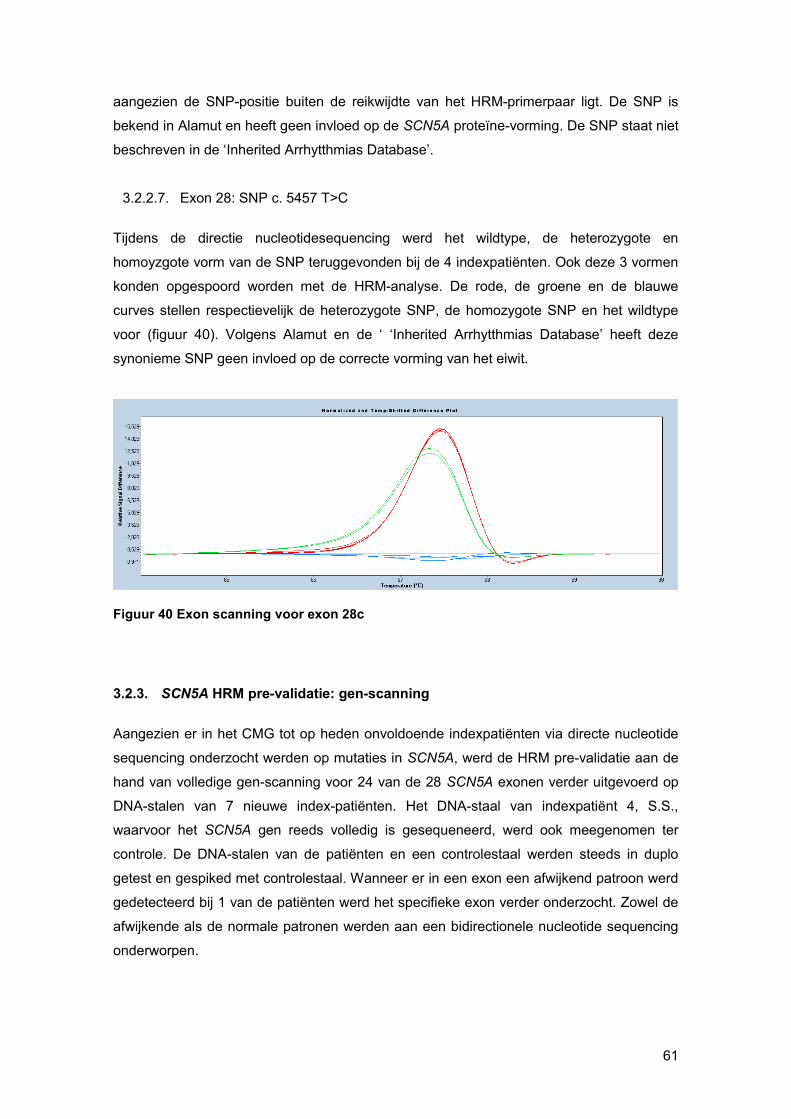
61
aangezien de SNP-positie buiten de reikwijdte van het HRM-primerpaar ligt. De SNP is
bekend in Alamut en heeft geen invloed op de SCN5A proteïne-vorming. De SNP staat niet
beschreven in de ‘Inherited Arrhytthmias Database’.
3.2.2.7. Exon 28: SNP c. 5457 T>C
Tijdens de directie nucleotidesequencing werd het wildtype, de heterozygote en
homoyzgote vorm van de SNP teruggevonden bij de 4 indexpatiënten. Ook deze 3 vormen
konden opgespoord worden met de HRM-analyse. De rode, de groene en de blauwe
curves stellen respectievelijk de heterozygote SNP, de homozygote SNP en het wildtype
voor (figuur 40). Volgens Alamut en de ‘ ‘Inherited Arrhytthmias Database’ heeft deze
synonieme SNP geen invloed op de correcte vorming van het eiwit.
Figuur 40 Exon scanning voor exon 28c
3.2.3. SCN5A HRM pre-validatie: gen-scanning
Aangezien er in het CMG tot op heden onvoldoende indexpatiënten via directe nucleotide
sequencing onderzocht werden op mutaties in SCN5A, werd de HRM pre-validatie aan de
hand van volledige gen-scanning voor 24 van de 28 SCN5A exonen verder uitgevoerd op
DNA-stalen van 7 nieuwe index-patiënten. Het DNA-staal van indexpatiënt 4, S.S.,
waarvoor het SCN5A gen reeds volledig is gesequeneerd, werd ook meegenomen ter
controle. De DNA-stalen van de patiënten en een controlestaal werden steeds in duplo
getest en gespiked met controlestaal. Wanneer er in een exon een afwijkend patroon werd
gedetecteerd bij 1 van de patiënten werd het specifieke exon verder onderzocht. Zowel de
afwijkende als de normale patronen werden aan een bidirectionele nucleotide sequencing
onderworpen.
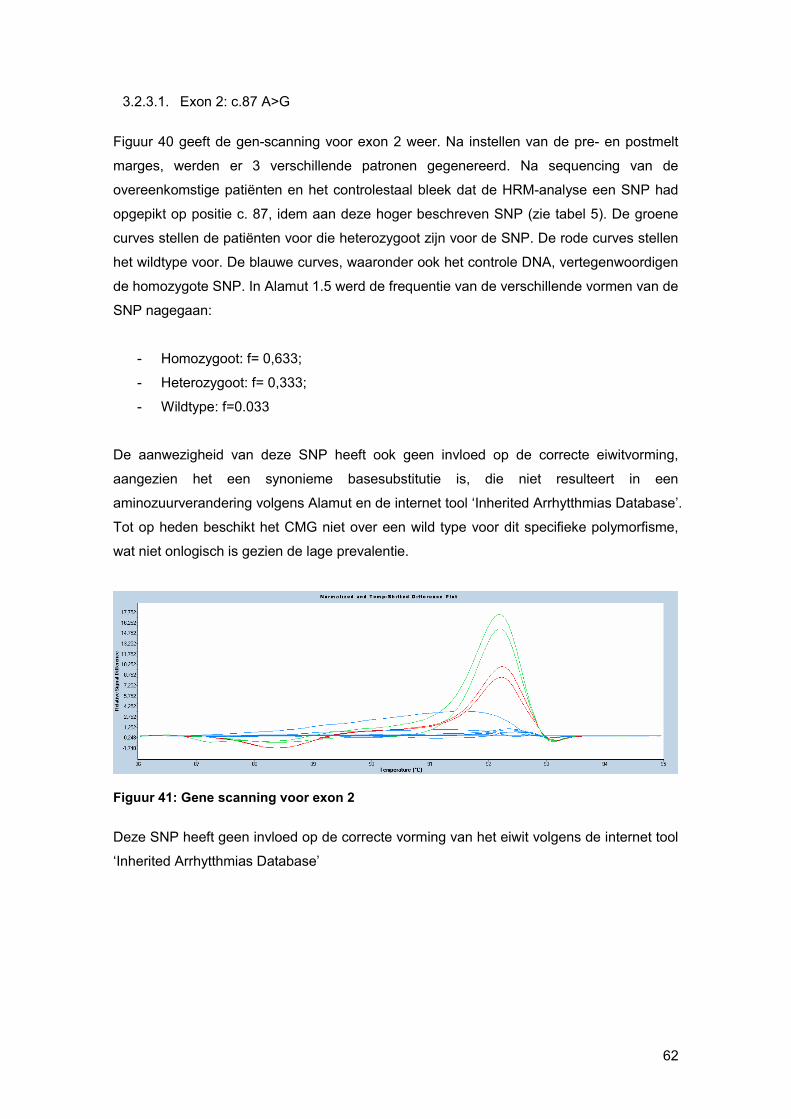
62
3.2.3.1. Exon 2: c.87 A>G
Figuur 40 geeft de gen-scanning voor exon 2 weer. Na instellen van de pre- en postmelt
marges, werden er 3 verschillende patronen gegenereerd. Na sequencing van de
overeenkomstige patiënten en het controlestaal bleek dat de HRM-analyse een SNP had
opgepikt op positie c. 87, idem aan deze hoger beschreven SNP (zie tabel 5). De groene
curves stellen de patiënten voor die heterozygoot zijn voor de SNP. De rode curves stellen
het wildtype voor. De blauwe curves, waaronder ook het controle DNA, vertegenwoordigen
de homozygote SNP. In Alamut 1.5 werd de frequentie van de verschillende vormen van de
SNP nagegaan:
- Homozygoot: f= 0,633;
- Heterozygoot: f= 0,333;
- Wildtype: f=0.033
De aanwezigheid van deze SNP heeft ook geen invloed op de correcte eiwitvorming,
aangezien het een synonieme basesubstitutie is, die niet resulteert in een
aminozuurverandering volgens Alamut en de internet tool ‘Inherited Arrhytthmias Database’.
Tot op heden beschikt het CMG niet over een wild type voor dit specifieke polymorfisme,
wat niet onlogisch is gezien de lage prevalentie.
Deze SNP heeft geen invloed op de correcte vorming van het eiwit volgens de internet tool
‘Inherited Arrhytthmias Database’
Figuur 41: Gene scanning voor exon 2
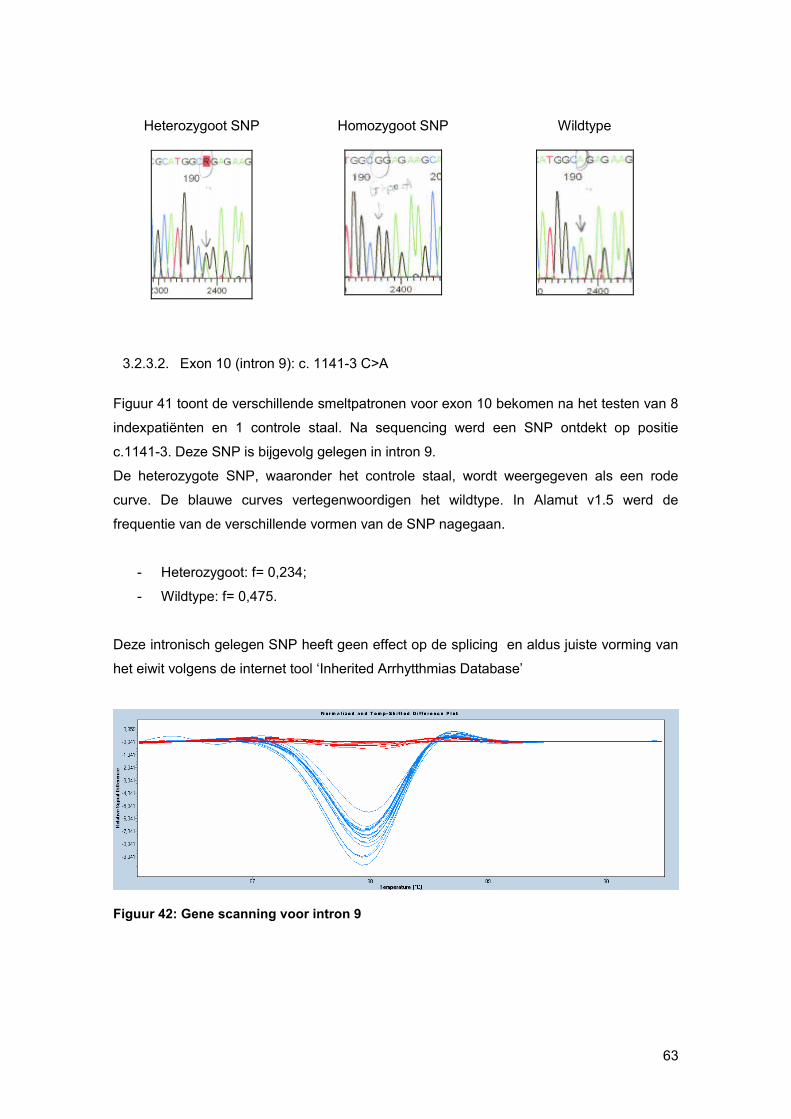
63
Heterozygoot SNP Homozygoot SNP Wildtype
3.2.3.2. Exon 10 (intron 9): c. 1141-3 C>A
Figuur 41 toont de verschillende smeltpatronen voor exon 10 bekomen na het testen van 8
indexpatiënten en 1 controle staal. Na sequencing werd een SNP ontdekt op positie
c.1141-3. Deze SNP is bijgevolg gelegen in intron 9.
De heterozygote SNP, waaronder het controle staal, wordt weergegeven als een rode
curve. De blauwe curves vertegenwoordigen het wildtype. In Alamut v1.5 werd de
frequentie van de verschillende vormen van de SNP nagegaan.
- Heterozygoot: f= 0,234;
- Wildtype: f= 0,475.
Deze intronisch gelegen SNP heeft geen effect op de splicing en aldus juiste vorming van
het eiwit volgens de internet tool ‘Inherited Arrhytthmias Database’
Figuur 42: Gene scanning voor intron 9

64
Heterozygoot SNP Wildtype
3.2.3.3. Exon 12: c. 1673 A>G
Na het testen van 8 indexpatiënten en 1 controlestaal, gaf de smeltcurve voor exon 12 3
patronen. Na sequencing bleken deze patronen de verschillende vormen van 1 SNP op
positie c.1673 voor te stellen. De patiënten die homozygoot zijn voor de SNP worden
gegroepeerd onder de groene curves. De patiënten die heterozygoot zijn voor de SNP
worden verzameld onder de rode curve. De wildtypes worden weergegeven als blauwe
curves. Het controle staal is wederom heterozygoot voor de SNP.
In Alamut v1.5 werd de frequentie van de SNP nagegaan:
- Homozygoot: f= 0.033;
- Heterozygoot: f= 0.300;
- Wildtype: f=0.667.
Deze base-substitutie heeft als gevolg dat er een Arginine in plaats van een Histidine wordt
gevormd. Beide aminozuren zijn allebei basische aminozuren. Het effect zal bijgevolg
miniem zijn. Deze positie is gekend als een cSNP in de Inherited Arrytthmias Database.
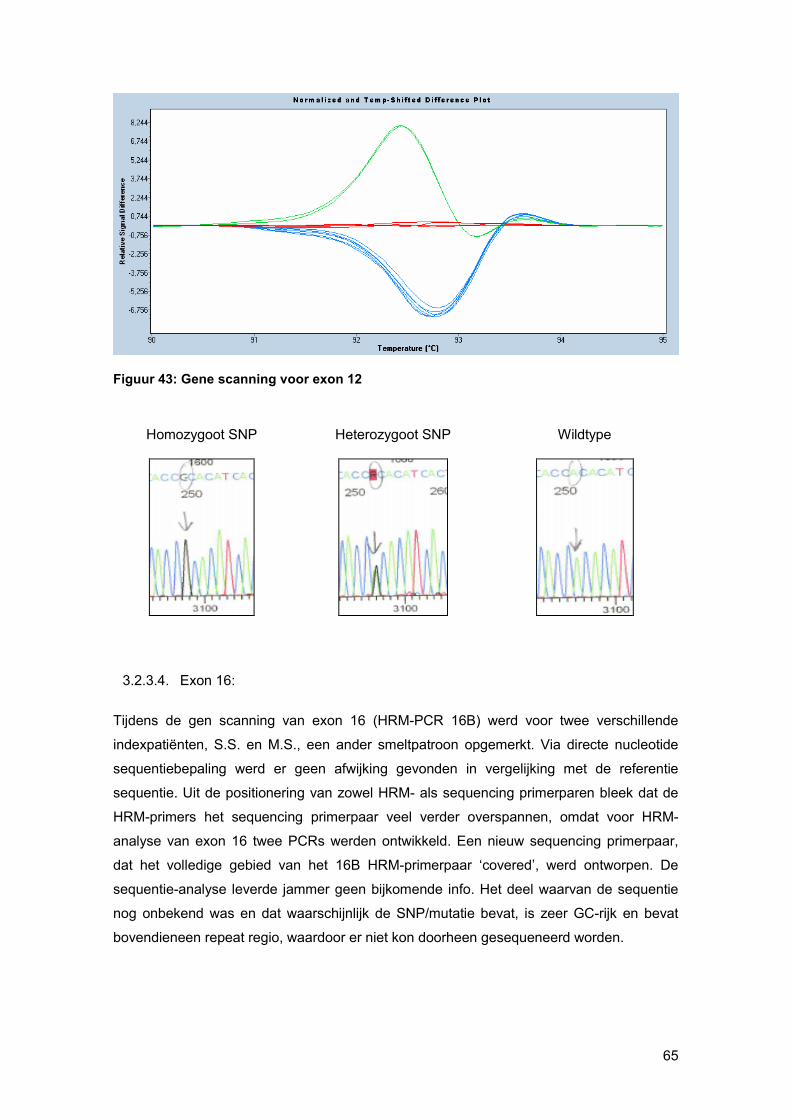
65
Figuur 43: Gene scanning voor exon 12
Homozygoot SNP Heterozygoot SNP Wildtype
3.2.3.4. Exon 16:
Tijdens de gen scanning van exon 16 (HRM-PCR 16B) werd voor twee verschillende
indexpatiënten, S.S. en M.S., een ander smeltpatroon opgemerkt. Via directe nucleotide
sequentiebepaling werd er geen afwijking gevonden in vergelijking met de referentie
sequentie. Uit de positionering van zowel HRM- als sequencing primerparen bleek dat de
HRM-primers het sequencing primerpaar veel verder overspannen, omdat voor HRM-
analyse van exon 16 twee PCRs werden ontwikkeld. Een nieuw sequencing primerpaar,
dat het volledige gebied van het 16B HRM-primerpaar ‘covered’, werd ontworpen. De
sequentie-analyse leverde jammer geen bijkomende info. Het deel waarvan de sequentie
nog onbekend was en dat waarschijnlijk de SNP/mutatie bevat, is zeer GC-rijk en bevat
bovendieneen repeat regio, waardoor er niet kon doorheen gesequeneerd worden.
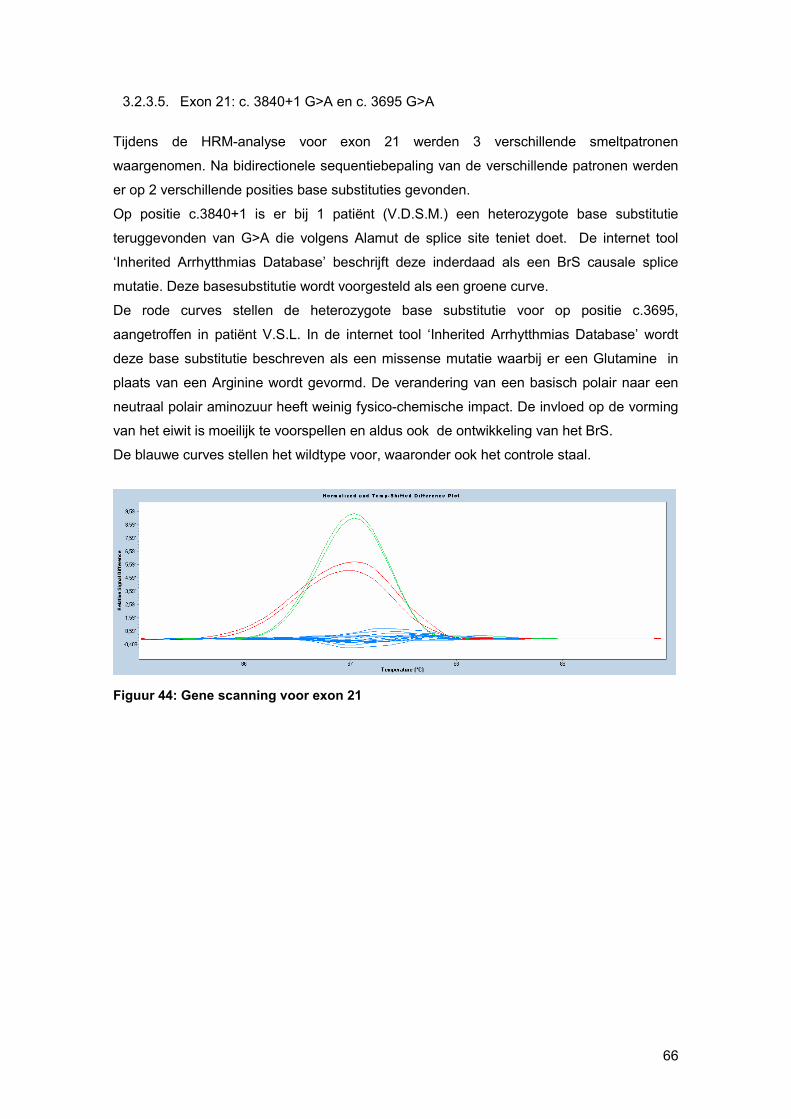
66
3.2.3.5. Exon 21: c. 3840+1 G>A en c. 3695 G>A
Tijdens de HRM-analyse voor exon 21 werden 3 verschillende smeltpatronen
waargenomen. Na bidirectionele sequentiebepaling van de verschillende patronen werden
er op 2 verschillende posities base substituties gevonden.
Op positie c.3840+1 is er bij 1 patiënt (V.D.S.M.) een heterozygote base substitutie
teruggevonden van G>A die volgens Alamut de splice site teniet doet. De internet tool
‘Inherited Arrhytthmias Database’ beschrijft deze inderdaad als een BrS causale splice
mutatie. Deze basesubstitutie wordt voorgesteld als een groene curve.
De rode curves stellen de heterozygote base substitutie voor op positie c.3695,
aangetroffen in patiënt V.S.L. In de internet tool ‘Inherited Arrhytthmias Database’ wordt
deze base substitutie beschreven als een missense mutatie waarbij er een Glutamine in
plaats van een Arginine wordt gevormd. De verandering van een basisch polair naar een
neutraal polair aminozuur heeft weinig fysico-chemische impact. De invloed op de vorming
van het eiwit is moeilijk te voorspellen en aldus ook de ontwikkeling van het BrS.
De blauwe curves stellen het wildtype voor, waaronder ook het controle staal.
Figuur 44: Gene scanning voor exon 21

67
Heterozygote splice mutatie
c. 3840+1 G>A
Heterozygoot mutatie:
c. 3695 G>A
3.2.3.6. Exon 23 (intron 23): 4245+82 A>G
Figuur 44 is een voorstelling de verschillende smeltpatronen die na sequencing analyse
een SNP in intron 23 blijken te vertegenwoordigen. De rode curve stelt de HRM-analyse
voor van de patiënten die homozygoot zijn voor de SNP. De blauwe curve stelt de HRM-
analyse voor van de patiënten die heterozygoot zijn voor de SNP, waaronder het controle
staal. Een staal dat het wildtype voorstelt, is, werd tot op heden nog niet gevonden in de
reeds geteste patiënten populatie. Binnen de blauwe curves dient wel een aanzienlijke
afwijking ten opzichte van de basiscurve te worden opgemerkt. In de toekomst moet via
sequencing worden onderzocht of deze blauwe curves misschien toch het smeltpatroon
van een (andere) SNP weergeven, dan wel dat de omliggende G-stretch verantwoordelijk
is voor de grotere afwijking binnen de wild types.
In Alamut v1.5 is de frequentie van de SNP nagegaan:
- Homozygoot: f= 0,119;
- Heterozygoot: f=0,356.
Via Alamut en de internet tool ‘‘Inherited Arrhytthmias Database’ kon nagegaan worden dat
deze SNP geen invloed heeft op de juiste vorming van het eiwit.
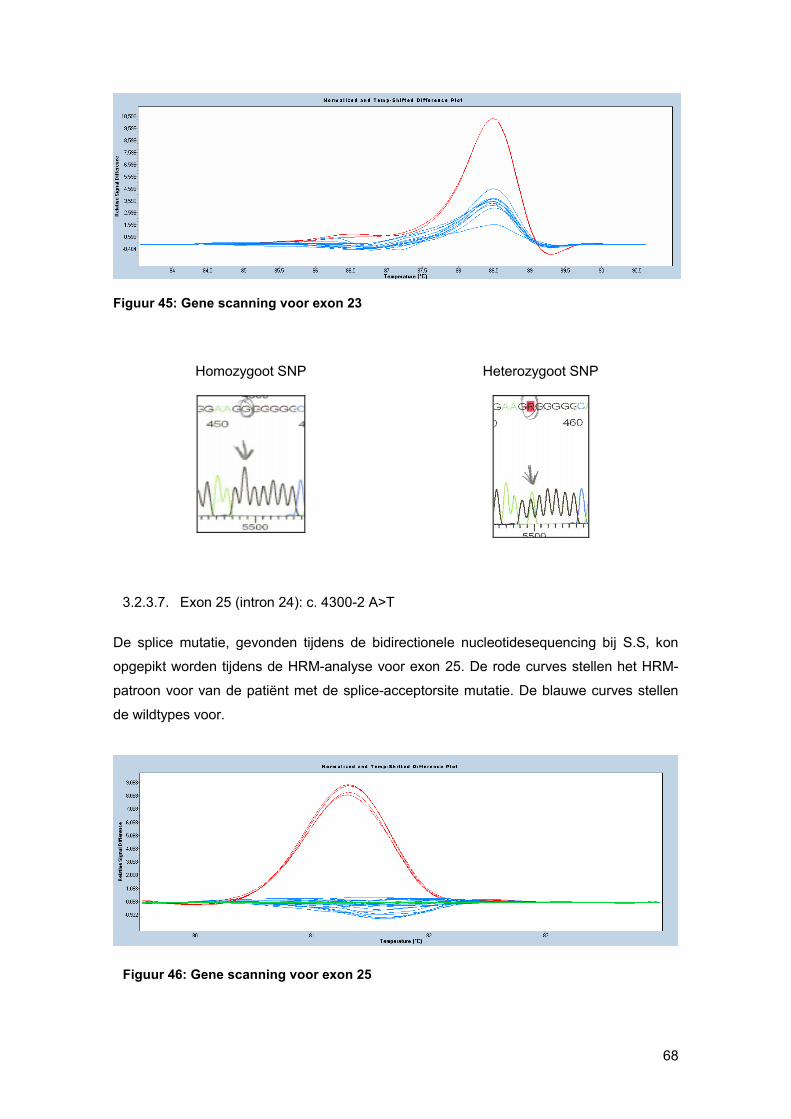
Homozygoot SNP Heterozygoot SNP
3.2.3.7. Exon 25 (intron 24): c. 4300-2 A>T
De splice mutatie, gevonden tijdens de bidirectionele nucleotidesequencing bij S.S, kon
opgepikt worden tijdens de HRM-analyse voor exon 25. De rode curves stellen het HRM-
patroon voor van de patiënt met de splice-acceptorsite mutatie. De blauwe curves stellen
de wildtypes voor.
Figuur 45: Gene scanning voor exon 23
68
Figuur 46: Gene scanning voor exon 25
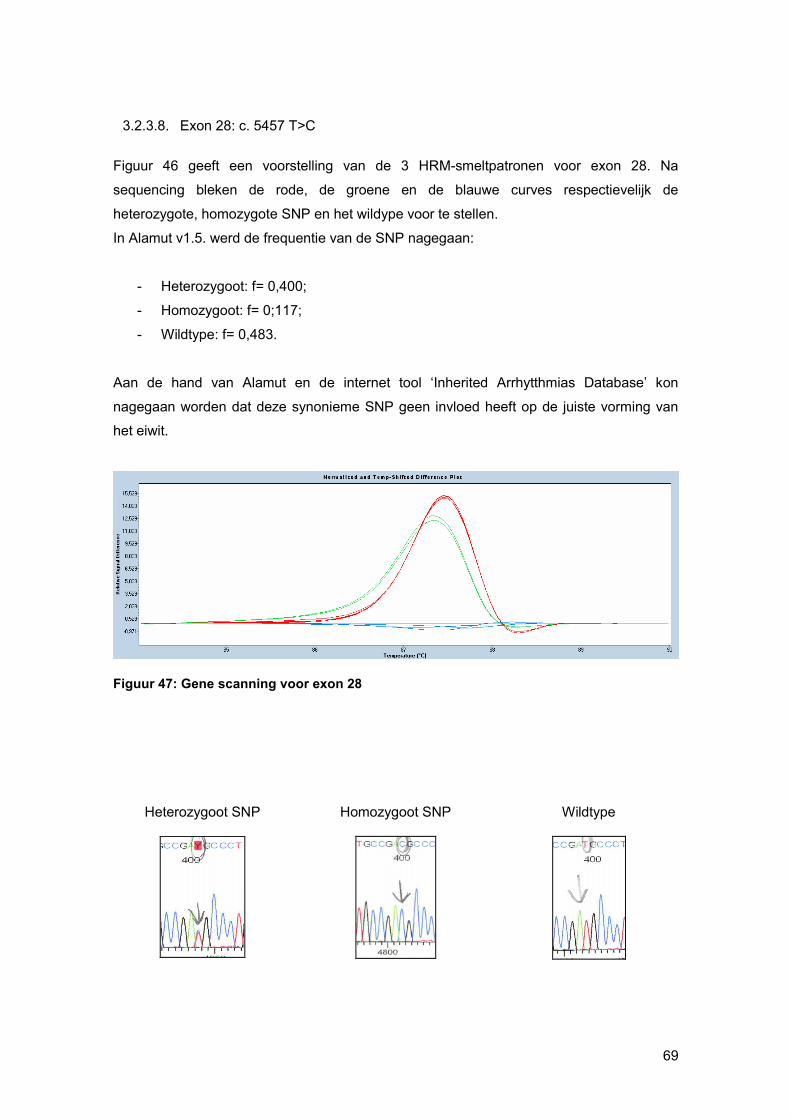
69
3.2.3.8. Exon 28: c. 5457 T>C
Figuur 46 geeft een voorstelling van de 3 HRM-smeltpatronen voor exon 28. Na
sequencing bleken de rode, de groene en de blauwe curves respectievelijk de
heterozygote, homozygote SNP en het wildype voor te stellen.
In Alamut v1.5. werd de frequentie van de SNP nagegaan:
- Heterozygoot: f= 0,400;
- Homozygoot: f= 0;117;
- Wildtype: f= 0,483.
Aan de hand van Alamut en de internet tool ‘Inherited Arrhytthmias Database’ kon
nagegaan worden dat deze synonieme SNP geen invloed heeft op de juiste vorming van
het eiwit.
Heterozygoot SNP Homozygoot SNP Wildtype
Figuur 47: Gene scanning voor exon 28

70
4. Besluit
Deze studie had tot doel de moleculaire diagnose van BrS op termijn mogelijk te maken
aan de hand van een high-througput mutatiedetectie platform, nl. HRM analyse. Het
gebruik van de HRM analyse zou namelijk toelaten om de diagnose van BrS veel sneller te
laten verlopen in vergelijking met de traditionele techniek van de bidirectionele nucleotide
sequencing.
Om HRM analyse mogelijk te maken, dienden allereerst de 37 RT-PCR-reacties van het
SCN5A gen op punt gesteld te worden. Pas nadat de RT-PCR optimalisatie voltooid was,
kon overgegaan worden op het testen van de detectiegevoeligheid van de HRM-analyse.
Dit moest in de eerste plaats uitgevoerd worden op DNA-stalen van indexpatiënten,
waarvan .de volledige SCN5A exon en flankerende intron nucleotidesequentie gekend is,
en dus alle mogelijke polymorfismen en/of mutaties geïdentificeerd zijn.
Met betrekking tot de eerste vereiste – de oppuntstelling van de 37 RT-PCR-reacties – kon
beroep worden gedaan op een eerdere poging tot oppuntstelling. Bij deze eerste poging
was men er reeds in geslaagd 21 van de 37 RT-PCR-reacties op punt te stellen. Bij de
aanvang van het huidige onderzoek werd opnieuw gestart voor de 16 overige RT-PCR- en
HRM-analyses. Op basis van primers, beschreven in de literatuur, werd er in geslaagd om
7 van de 16 RT-PCR-reacties verder op punt te stellen. Voor de overblijvende 9 RT-PCR-
reacties, werden de resultaten van de eerste poging tot oppuntstelling vergeleken met die
van de tweede poging met primers uit de literatuur. Uit deze vergelijking bleek dat bij 4 RT-
PCR-reacties de resultaten van de eerste poging tot oppuntstelling beter en alsnog
bruikbaar waren. Voor de overige 5 RT-PCR-reacties moesten nieuwe primers ontwikkeld
worden. Hiervoor werd het computerprogramma Light Scanner gebruikt. Met deze nieuwe
primers werd in een derde poging één bijkomende RT-PCR-reactie oppuntgesteld. De 4
overblijvende RT-PCR-reacties konden niet op punt worden gesteld tijdens dit onderzoek.
Bijgevolg zal men de exonen gelinkt aan deze RT-PCR-reacties niet kunnen onderwerpen
aan een HRM-analyse. Voor deze exonen (m.n. 17, 19, 20 en 28) zal men toevlucht
moeten blijven nemen in directe nucleotide sequencing.
Voor de tweede vereiste – beschikken over stalen van indexpatiënten met geïdenticeerde
SNPs en/of mutaties – werd het DNA van 4 indexpatiënten eerst onderzocht via directe
nucleotide sequencing. Bij dit onderzoek werden in totaal 8 basesubstituties gevonden.
Zeven bleken gekende SNPs te zijn, terwijl één nog niet beschreven mutatie werd
gevonden. Aan de hand van de familiale segregatie van de mutatie, enkel in familieleden
die positief zijn voor de klinische diagnostische criteria voor BrS, kon worden aangetoond,
dat de gevonden mutatie causaal is voor BrS.

71
Deze aanwezige SNPs en mutatie werden gebruikt als basis voor de HRM-prevalidatie in
het kader van detectiegevoeligheid. Voor 4 exonen met SNPs (m.n. 2, 17a, 23, 28b)
werden aberrante smeltpatronen opgepikt. Voor 2 van deze 4 exonen bekwam men echter
enkel de gewenste gevoeligheid, indien hun tweeledige smeltdomeinen apart geanalyseerd
werden. Voor 2 exonen (24, 25) konden de SNPs niet worden gedetecteerd, aangezien
deze zich in een gebied bevinden dat niet wordt gecoverd door de HRM-primers. Voor
exon 17b stond de RT-PCR-reactie niet op punt en kon dus niet worden overgaan tot HRM-
analyse.
Uit deze resultaten is gebleken dat de HRM analyse althans voor de onderzochte exonen
voldoende gevoelig is. Ingeval de PCR uit meerdere smeltdomeinen bestaat, dienen deze
ook apart geanalyseerd te worden .
Achteraf werd de HRM-detectiegevoeligheid verder onderzocht aan de hand van gen-
scanning voor alle ontwikkelde SCN5A HRM-PCRs dmv van 6 nieuwe indexpatiënten. Bij
deze 6 patiënten werden via HRM analyse in totaal 8 afwijkende smeltpatronen
gedetecteerd, welke via sequencing werden geïdentificeerd als 6 SNPs en twee mutaties.
Deze totale analyse (HRM SCN5A gen scanning + exon-specifieke sequencing) werd
uitgevoerd op twee weken tijd, terwijl een sequenicng analyse van alle exonen voor deze 6
patiënten waarschijnlijk meer dan een maand in beslag had genomen.
Dankzij het gebruik van HRM als screeningmethode zal het dan ook mogelijk worden om in
de toekomst sneller tot een moleculaire diagnose van Brugada syndroom te komen. In de
komende maanden zullen beide platforms echter in parallel getest worden om de HRM-
detectiegevoeligheid nog verder te onderzoeken.

72
5. Literatuurlijst
1. Wolf C.M., Berul C.I. Molecular Mechanisms of Inherited Arrhytmias. Current
Genomics 2008, 9, p. 160-168;
2. http://cardio.nettools.be/default.aspx?url=p_119.htm geraadpleegd op 26 april 2010;
3. Campuzano O., Sarquella-Brugada G., Bugada R., Brugada P., Brugada J. The
Genetic Basis of Malignant Arrhythmias and Cardiomyopathies Rev Esp Cardiol. 2009,
62(4), p. 422-436;
4. L.L. Kirchmann Anatomie en fysiologie van de mens. 2005, Elsevier gezondheidzorg,
Maarssen;
5. S. Silbernagl, A. Despopoulos Atlas van de fysiologie. 2008, SESAM/Hbuitgevers,
Baarn;
6. Sovari A.A., Kocheril A.G., Assadi R., Baas A.S., Zareba W., Rosero S. Long QT
Syndrome 2009, emedicine.medscape.com;
7. Morita H., Wu J., Zipers D.P. The QT syndromes: long and short Lancet, 2008, 372, p.
750-763;
8. Schwartz P.J. The congenital long QT syndromes from genotype to phenotype: clinical
implications Journal of Internal Medicine, 2006, 259, p. 39-47;
9. Medeiros-Domingo A., Iturralde-Torres P., Ackerman M.J. Clinical and Genetics
Characteristics of Long QT Syndrome Rev Esp Cardiol, 2007, 60, p. 739-752;
10. Reinig M.G., Engel T.R. The Shortage of Short QTq Chest, 2007, DOI
10.1378/chest.06-2133;
11. Brugada R., Hong K., Cordeiro J.M., Dumaine R. Short QT Syndrome Canadian
Medical Association Journal, 2005, 173(11), p. 1349-1354;
12. Antzelevitch C. The Brugada Syndrome: diagnostic criteria and cellular mechanisms
European Heart Journal, 2001, 22, p. 356-363;
13. Brugada P., Benito B., Brugada R., Brugada J. Brugada Syndrome: Update 2009
Hellenic J Cardiol, 2009, 50, p. 352-372;
14. Farwell D., Gollob M.H. Electrical heart disease: Genetic and molecula basis of cardiac
arrhytmias in normal strucural hearts Can. J. Cardiol., 2007, 23, p. 16A-22A;
15. Benito B., Brugada J., Brugada R., Brugada P. Brugada Syndrome Rev. Esp. Cardiol.,
2009, 62(11), p. 1297-1315;
16. http://www.premierbiosoft.com/tech_notes/PCR_Primer_Design.html geraadpleegd op
8 mei 2010;
17. http://www.premierbiosoft.com/tech_notes/PCR_Primer_Design.html geraadpleegd op
8 mei 2010;
18. https://www.roche-applied-science.com/sis/amplification/index.jsp?id=AM050300
geraadpleegd op 8 mei 2010;
19. http://blast.ncbi.nlm.nih.gov/Blast.cgi geraadpleegd op 1 mei 2010;

73
20. Real-Time PCR: From Theory to Practice, invitrogen;
21. http://www.nfstc.org/pdi/Subject04/pdi_s04_m01_02_f.htm geraadpleegd op 2 mei
2010;
22. SOP DNA bereiding UZ Brussel;
23. SOP fenol/chloroform extractie UZ Brussel;
24. http://www.invitrogen.com/site/us/en/home/Products-and-
Services/Applications/Nucleic-Acid-Amplification-and-Expression-Profiling/PCR/PCR-
Misc/Taq-DNA-Polymerase.html geraadpleegd op 2 mei 2010;
25. Watson J.D., Baker T.A., Bell S.P., Gann A., Levine M., Losick R. Molecular Biology of
the Gene. 2008, Pearson Education, San Fransisco;
26. Koolman J., Röhm K.-H. Atlas van de biochemie. 2004, SESAM/HBuitgevers, Baarn
27. http://www.biotium.com/product/price_and_info.asp?item=41001&Sub_Section=09A
geraadpleegd op 2 mei 2010;
28. SOP DNA sequencing UZ Brussel;
29. http://www3.appliedbiosystems.com/AB_Home/applicationstechnologies/High-
Resolution-Melting/index.htm geraadpleegd op 11/03/2010
30. https://www.roche-applied-science.com/sis/rtpcr/htc/index.jsp?&id=htc_000000
geraadpleegd op 9 mei 2010;
31. LightCycler® High Resolution Melting Master versie maart 2007;
32. Bijsluiter Light Cycle® 480 High Resolution Melting Master, versie juli 2009;
33. http://www.primerdesign.co.uk/Download%20material/Beginners%20guide%20to%20r
eal-time%20PCR.pdf geraadpleegd op 11 maart 2010;
34. Real-Time PCR Reaction: From Theory to Practice, Invitrogen;
35. Millat G., Chanavat V., Rodriguez-Lafrasse C., Rousson R., Rapid, sensitive and
inexpensive detection of SCN5A genetic variations by high resolution melting analysis
Clinical Biochemistry, 2008, 42, p. 491-499.
6. Lijst met figuren
1. http://www.10voorbiologie.nl/index.php?cat=9&id=573&par=586&sub=589
geraadpleegd op 13 mei 2010;
2. https://www.healthforums.com/print/articlePrint?brand=HealthForums.com&pkg=hf2&c
ontentID=9770 geraadpleegd op 13 mei 2010;
3. http://cardio.nettools.be/default.aspx?url=p_99.htm geraadpleegd op 13 mei 2010;
4. http://www.virtualmedicalcentre.com/healthinvestigations.asp?sid=28 geraadpleegd op
15 april 2010;
5. http://drsvenkatesan.wordpress.com/2009/08/26/why-ecg-is-greatly-influensed-by-
serum-potasium-levels-and-least-affected-by-serum-sodium-levels/ geraadpleegd op

74
13 mei 2010;
6. Brugada R., Hong K., Cordeiro J.M., Dumaine R. Short QT Syndrome Canadian
Medical Association Journal, 2005, 173(11), 1349-1354;
7. Brugada P., Benito B., Brugada R., Brugada J. Brugada Syndrome: Update 2009
Hellenic J Cardiol, 2009, 50, p. 352-372;
8. Antzelevitch C. The Brugada Syndrome: diagnostic criteria and cellular mechanisms
European Heart Journal, 2001, 22, p. 356-363;
9. Brugada P., Benito B., Brugada R., Brugada J. Brugada Syndrome: Update 2009
Hellenic J Cardiol, 2009, 50, p. 352-372;
10. http://www.genehk.com/news/pr/pr_s03.html geraadpleegd op 8 mei 2010;
11. http://www.mun.ca/biology/scarr/PCR_simplified.html geraadpleegd op 8 mei 2010;
12. https://sites.google.com/a/luther.edu/genetics/students/adam-ragheb/identification-of-
taster-and-nontaster-genotypes-by-restriction-enzyme-digestion-and-gel-
electrophoresis geraadpleegd op 8 mei 2010;
13. http://kenickbiochem09.wikispaces.com/Western+Blotting+--By+Ongori,+David?f=print
geraadpleegd op 8 mei 2010;
14. http://home.scarlet.be/~tsk05520/biomolespa/Enzimas/enzima-01.jpg geraadpleegd op
8 mei 2010;
15. http://www3.appliedbiosystems.com/cms/groups/portal/documents/web_content/cms_0
51951.gif geraadpleegd op 8 mei 2010;
16. http://dnasequencing.wordpress.com/2007/10/26/chain-termination-methods/
graadpleegd op 8 mei 2010;
17. Introducing High-Resolution Melting on the LightCycler® 480;
18. Introducing High-Resolution Melting on the LightCycler® 480;
19. Introducing High Resolution Melting on the LightCycler® 480 System;
20. Tm-calling;
21. Introducing High Resolution Melting on the LightCycler® 480 System;
22. Introducing High Resolution Melting on the LightCycler® 480 System;
23. Introducing High Resolution Melting on the LightCycler® 480 System.
24. Voorstelling heterozygote base-substitutie;
25. Effect mutatie;
26. Stamboom familie indexpatiënt;
27. Bepaling optimalisatie MgCl2 concentratie voor exon 16a;
28. Tm-calling voor exon 16a en 17a;
29. Tm-calling voor exon 17a;
30. Agarosegel-elektroforese 1;
31. Agarosegel-elektroforese 2;
32. RT-PCR-reactie exon 19;

75
33. Exon scanning voor exon 2;
34. Exon scanning : normalisatie exon 17a;
35. Exon scanning: ‘Differende plot’ exon 17a;
36. Exon scanning: normalisatie exon 17a;
37. Exon scanning: ‘Difference plot’ exon 17a;
38. Exon scanning: normalisatie voor exon 23
39. Exon scanning voor exon 23
40. Exon scanning voor 28c;
41. Gene scanning voor exon 2;
42. Gene scanning voor inron 9;
43. Gene scannning voor exon 12;
44. Gene scanning voor exon 21;
45. Gene scanning voor exon 23;
46. Gene scanning voor exon 25;
47. Gene scanning voor exon 28
7. Lijst met tabellen
1. The Genetic Basis of Malignant Arrhythmias and Cardiomyopathies
2. Clinical and Genetic Characteristics of Long QT Syndrome; Rev Esp Cardiol.
2007;60:739-52;
3. Samenstelling reactiemix PCR;
4. Samenstelling reactiemix HRM;
5. Overzicht SNPs;
6. RT-PCR-optimalisatie: MgCl2-titratie ter bepaling van de ideale concentratie;
7. RT-PCR-optimalisatie: reactiemix.
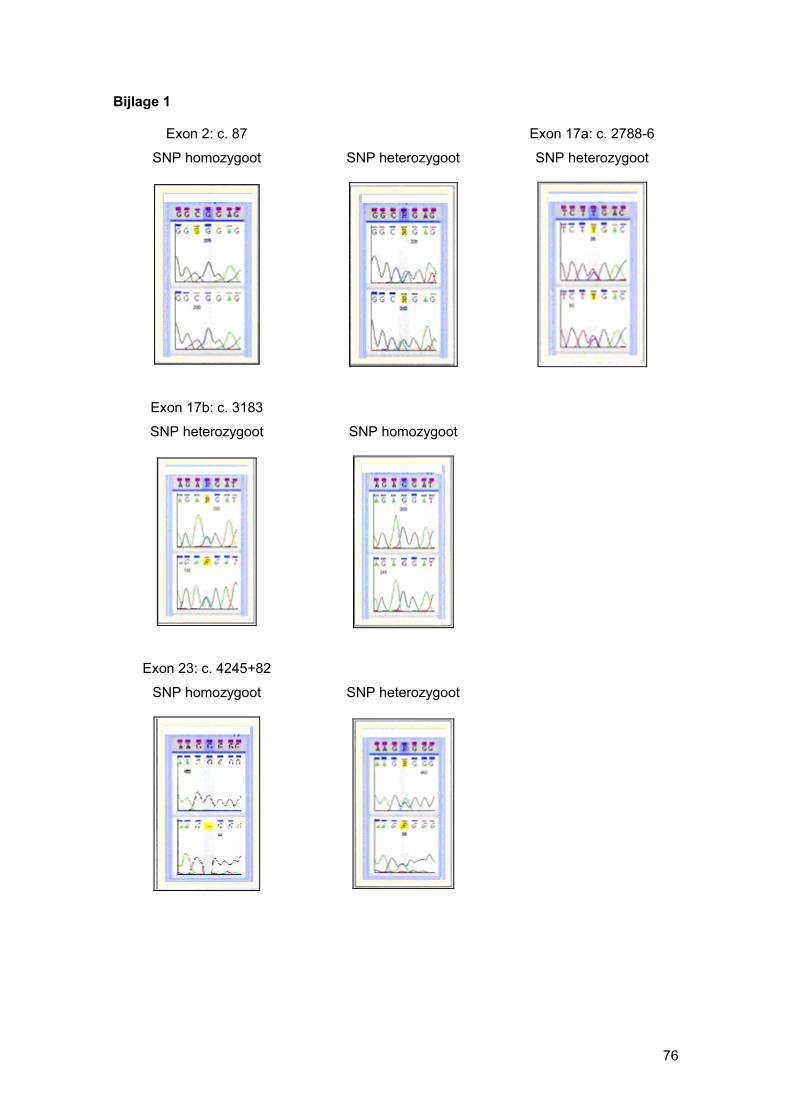
76
Bijlage 1
Exon 2: c. 87 Exon 17a: c. 2788-6
SNP homozygoot SNP heterozygoot SNP heterozygoot
Exon 17b: c. 3183
SNP heterozygoot SNP homozygoot
Exon 23: c. 4245+82
SNP homozygoot SNP heterozygoot
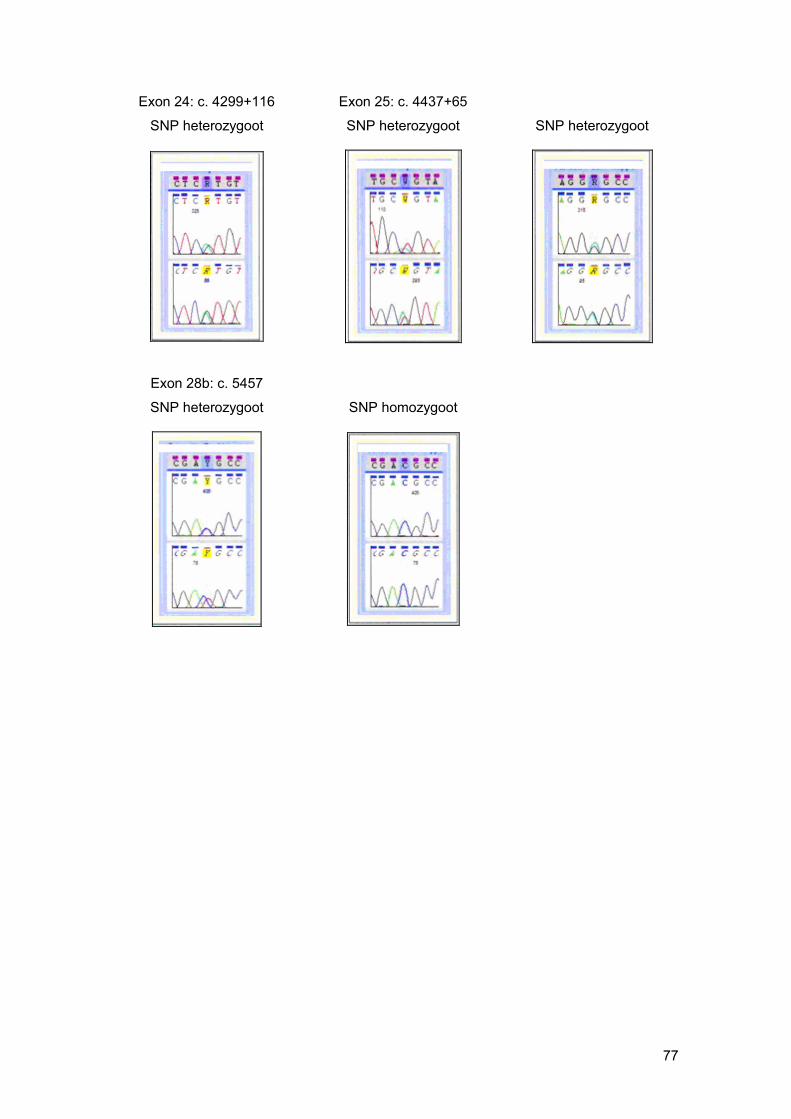
77
Exon 24: c. 4299+116 Exon 25: c. 4437+65
SNP heterozygoot SNP heterozygoot SNP heterozygoot
Exon 28b: c. 5457
SNP heterozygoot SNP homozygoot
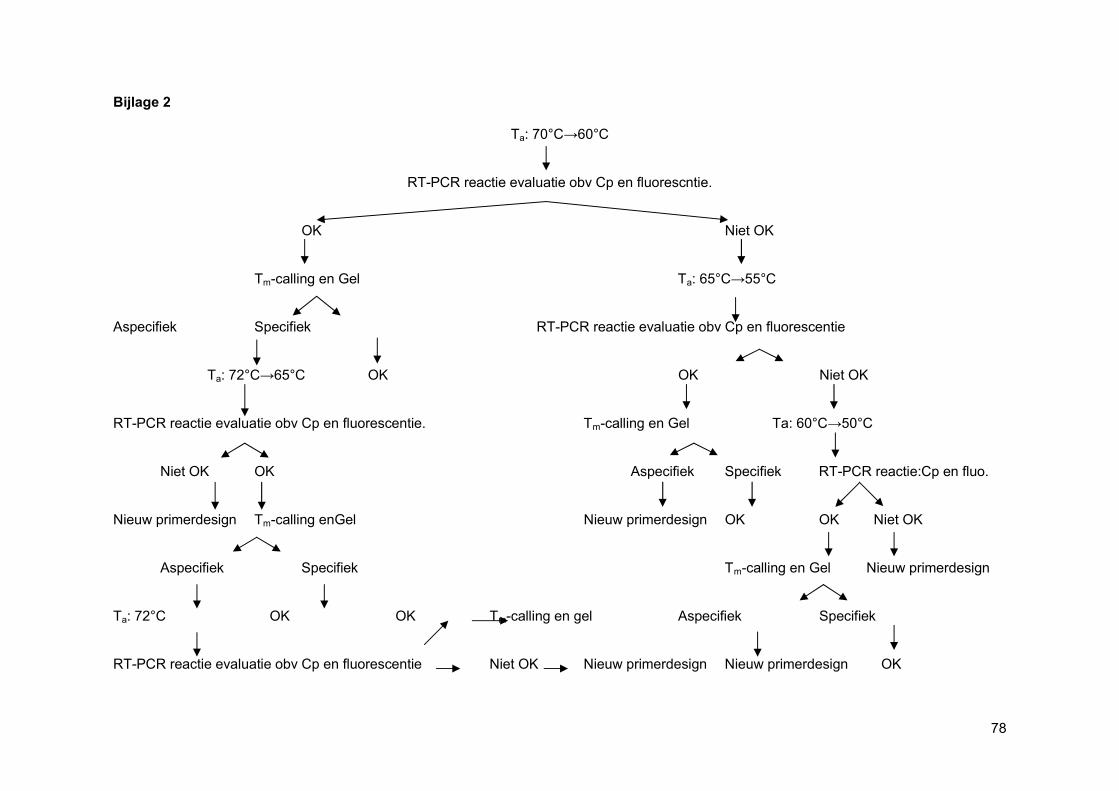
78
Bijlage 2
Ta: 70°C→60°C
RT-PCR reactie evaluatie obv Cp en fluorescntie.
OK Niet OK
Tm-calling en Gel Ta: 65°C→55°C
Aspecifiek Specifiek RT-PCR reactie evaluatie obv Cp en fluorescentie
Ta: 72°C→65°C OK OK Niet OK
RT-PCR reactie evaluatie obv Cp en fluorescentie. Tm-calling en Gel Ta: 60°C→50°C
Niet OK OK Aspecifiek Specifiek RT-PCR reactie:Cp en fluo.
Nieuw primerdesign Tm-calling enGel Nieuw primerdesign OK OK Niet OK
Aspecifiek Specifiek Tm-calling en Gel Nieuw primerdesign
Ta: 72°C OK OK Tm-calling en gel Aspecifiek Specifiek
RT-PCR reactie evaluatie obv Cp en fluorescentie Niet OK Nieuw primerdesign Nieuw primerdesign OK
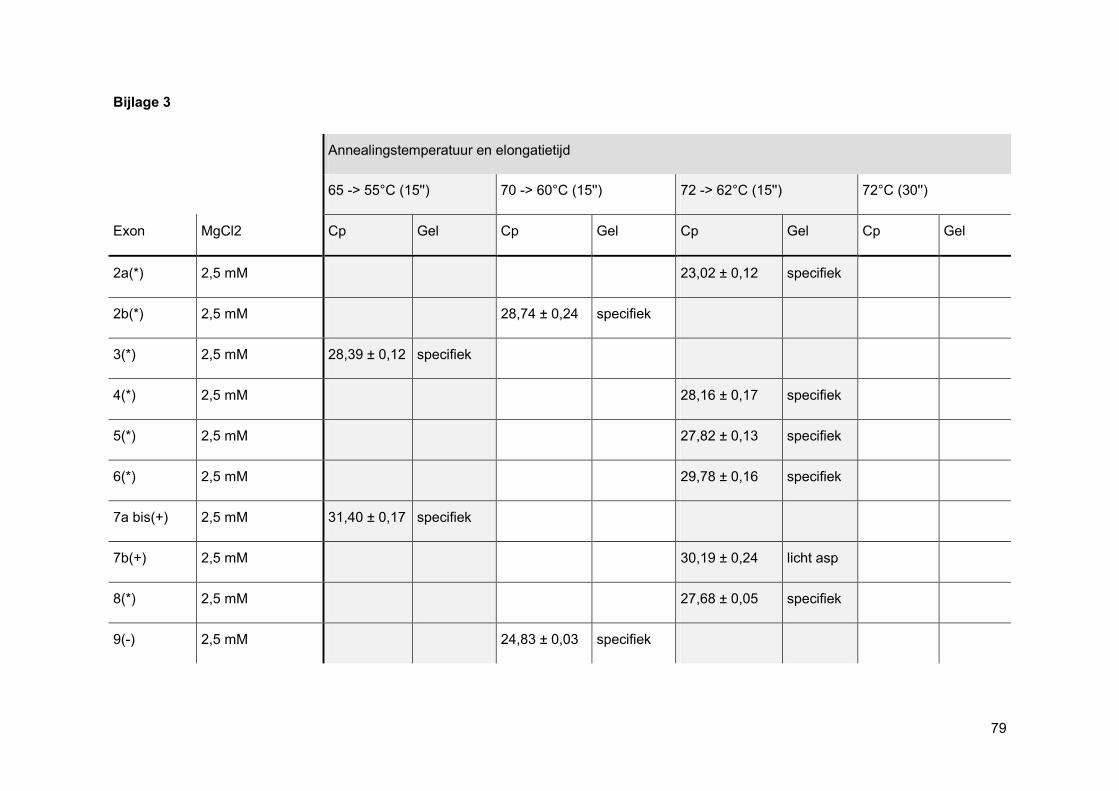
79
Bijlage 3
Annealingstemperatuur en elongatietijd
65 -> 55°C (15'') 70 -> 60°C (15'') 72 -> 62°C (15'') 72°C (30'')
Exon MgCl2 Cp Gel Cp Gel Cp Gel Cp Gel
2a(*) 2,5 mM 23,02 ± 0,12 specifiek
2b(*) 2,5 mM 28,74 ± 0,24 specifiek
3(*) 2,5 mM 28,39 ± 0,12 specifiek
4(*) 2,5 mM 28,16 ± 0,17 specifiek
5(*) 2,5 mM 27,82 ± 0,13 specifiek
6(*) 2,5 mM 29,78 ± 0,16 specifiek
7a bis(+) 2,5 mM 31,40 ± 0,17 specifiek
7b(+) 2,5 mM 30,19 ± 0,24 licht asp
8(*) 2,5 mM 27,68 ± 0,05 specifiek
9(-) 2,5 mM 24,83 ± 0,03 specifiek
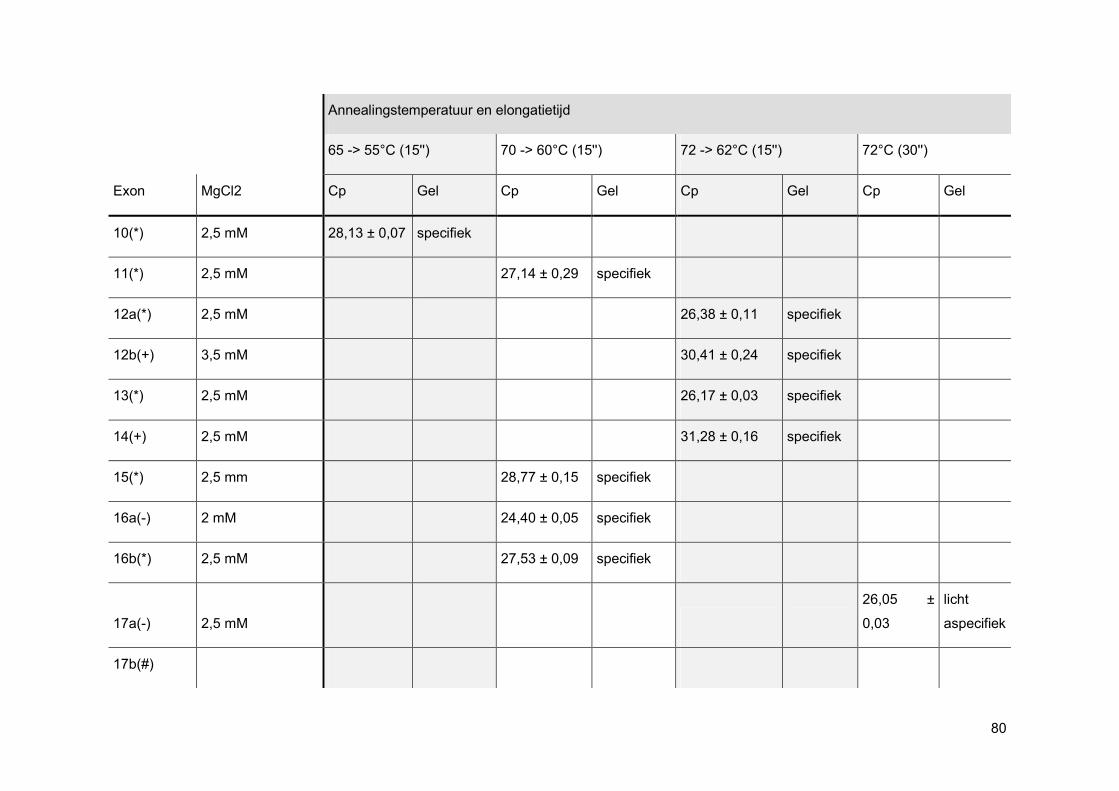
80
Annealingstemperatuur en elongatietijd
65 -> 55°C (15'') 70 -> 60°C (15'') 72 -> 62°C (15'') 72°C (30'')
Exon MgCl2 Cp Gel Cp Gel Cp Gel Cp Gel
10(*) 2,5 mM 28,13 ± 0,07 specifiek
11(*) 2,5 mM 27,14 ± 0,29 specifiek
12a(*) 2,5 mM 26,38 ± 0,11 specifiek
12b(+) 3,5 mM 30,41 ± 0,24 specifiek
13(*) 2,5 mM 26,17 ± 0,03 specifiek
14(+) 2,5 mM 31,28 ± 0,16 specifiek
15(*) 2,5 mm 28,77 ± 0,15 specifiek
16a(-) 2 mM 24,40 ± 0,05 specifiek
16b(*) 2,5 mM 27,53 ± 0,09 specifiek
17a(-) 2,5 mM
26,05 ±
0,03
licht
aspecifiek
17b(#)
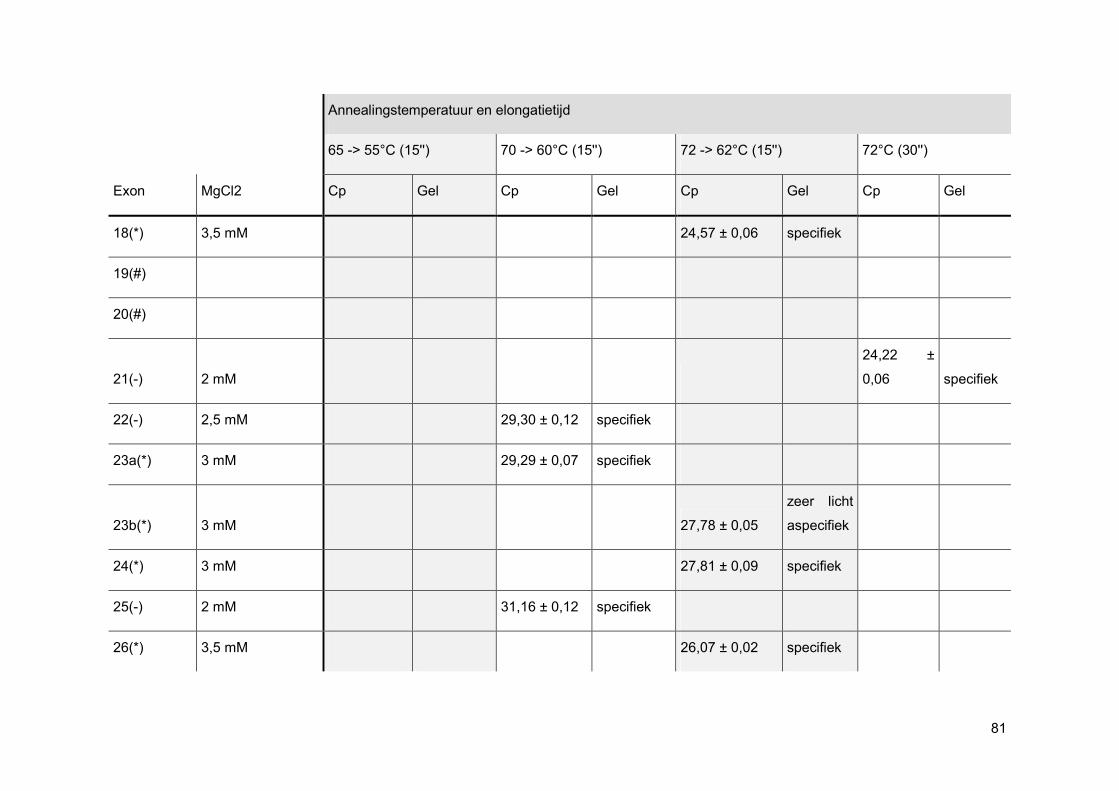
81
Annealingstemperatuur en elongatietijd
65 -> 55°C (15'') 70 -> 60°C (15'') 72 -> 62°C (15'') 72°C (30'')
Exon MgCl2 Cp Gel Cp Gel Cp Gel Cp Gel
18(*) 3,5 mM 24,57 ± 0,06 specifiek
19(#)
20(#)
21(-) 2 mM
24,22 ±
0,06 specifiek
22(-) 2,5 mM 29,30 ± 0,12 specifiek
23a(*) 3 mM 29,29 ± 0,07 specifiek
23b(*) 3 mM 27,78 ± 0,05
zeer licht
aspecifiek
24(*) 3 mM 27,81 ± 0,09 specifiek
25(-) 2 mM 31,16 ± 0,12 specifiek
26(*) 3,5 mM 26,07 ± 0,02 specifiek
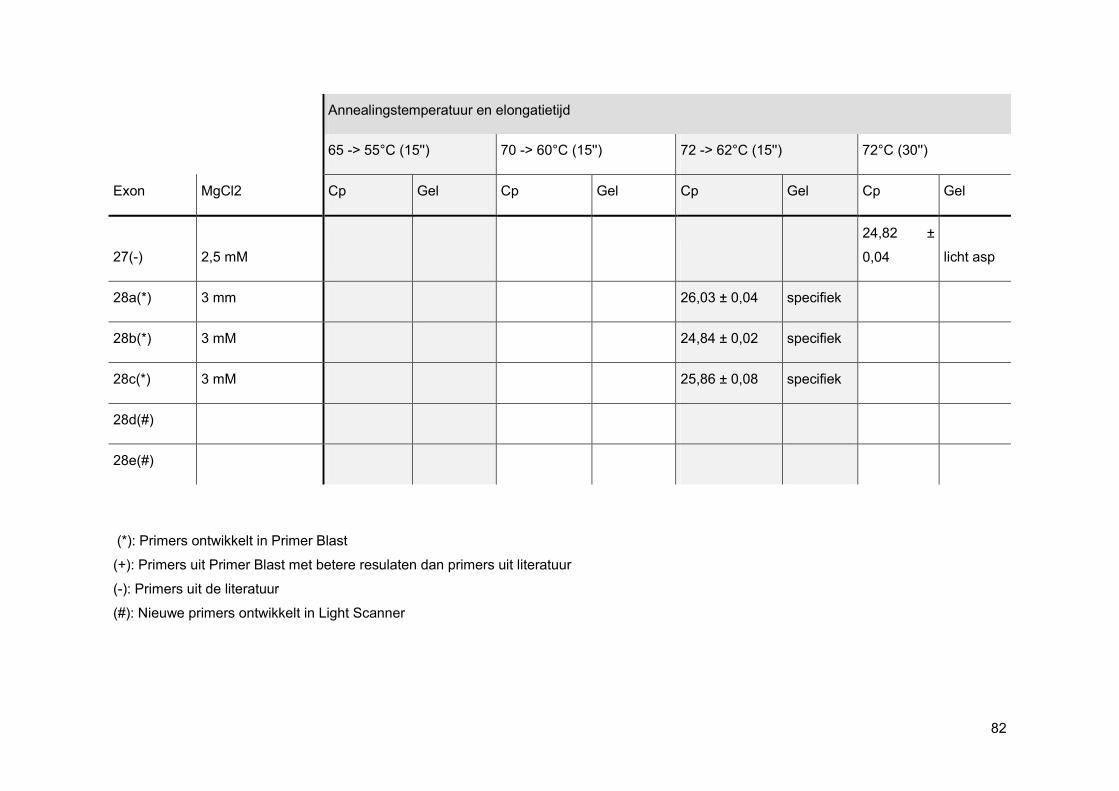
82
Annealingstemperatuur en elongatietijd
65 -> 55°C (15'') 70 -> 60°C (15'') 72 -> 62°C (15'') 72°C (30'')
Exon MgCl2 Cp Gel Cp Gel Cp Gel Cp Gel
27(-) 2,5 mM
24,82 ±
0,04 licht asp
28a(*) 3 mm 26,03 ± 0,04 specifiek
28b(*) 3 mM 24,84 ± 0,02 specifiek
28c(*) 3 mM 25,86 ± 0,08 specifiek
28d(#)
28e(#)
(*): Primers ontwikkelt in Primer Blast
(+): Primers uit Primer Blast met betere resulaten dan primers uit literatuur
(-): Primers uit de literatuur
(#): Nieuwe primers ontwikkelt in Light Scanner
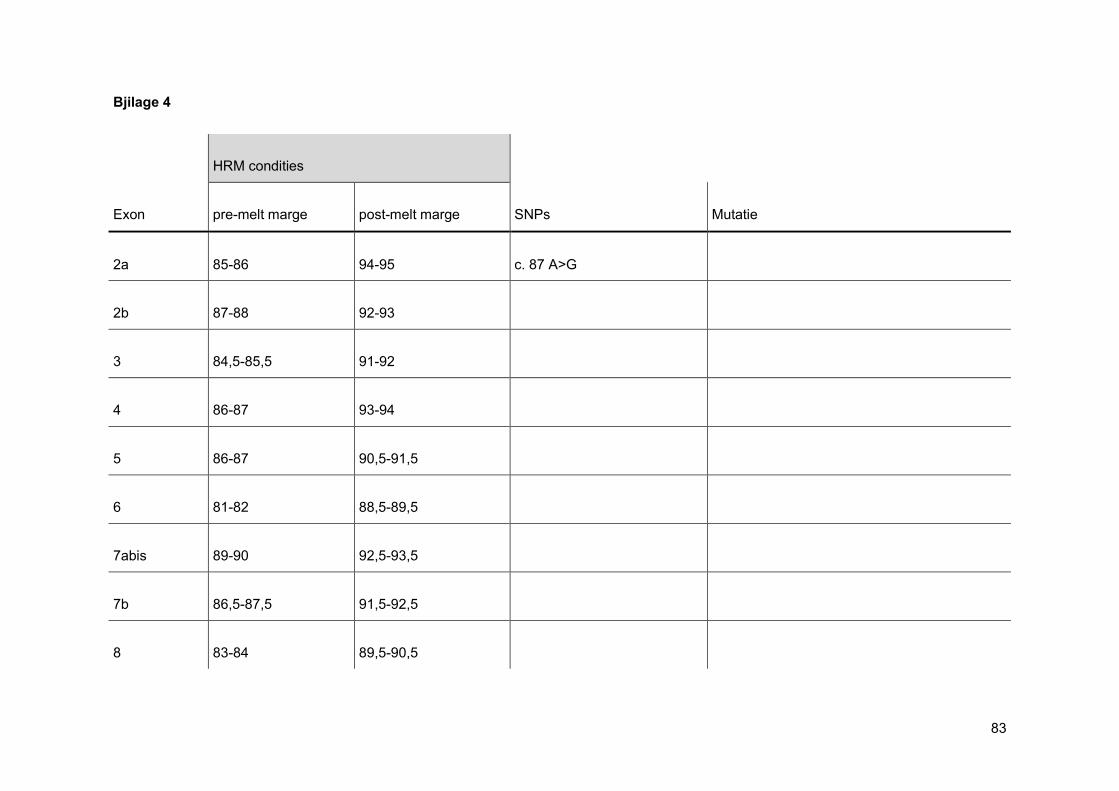
83
Bjilage 4
HRM condities
Exon pre-melt marge post-melt marge SNPs Mutatie
2a 85-86 94-95 c. 87 A>G
2b 87-88 92-93
3 84,5-85,5 91-92
4 86-87 93-94
5 86-87 90,5-91,5
6 81-82 88,5-89,5
7abis 89-90 92,5-93,5
7b 86,5-87,5 91,5-92,5
8 83-84 89,5-90,5
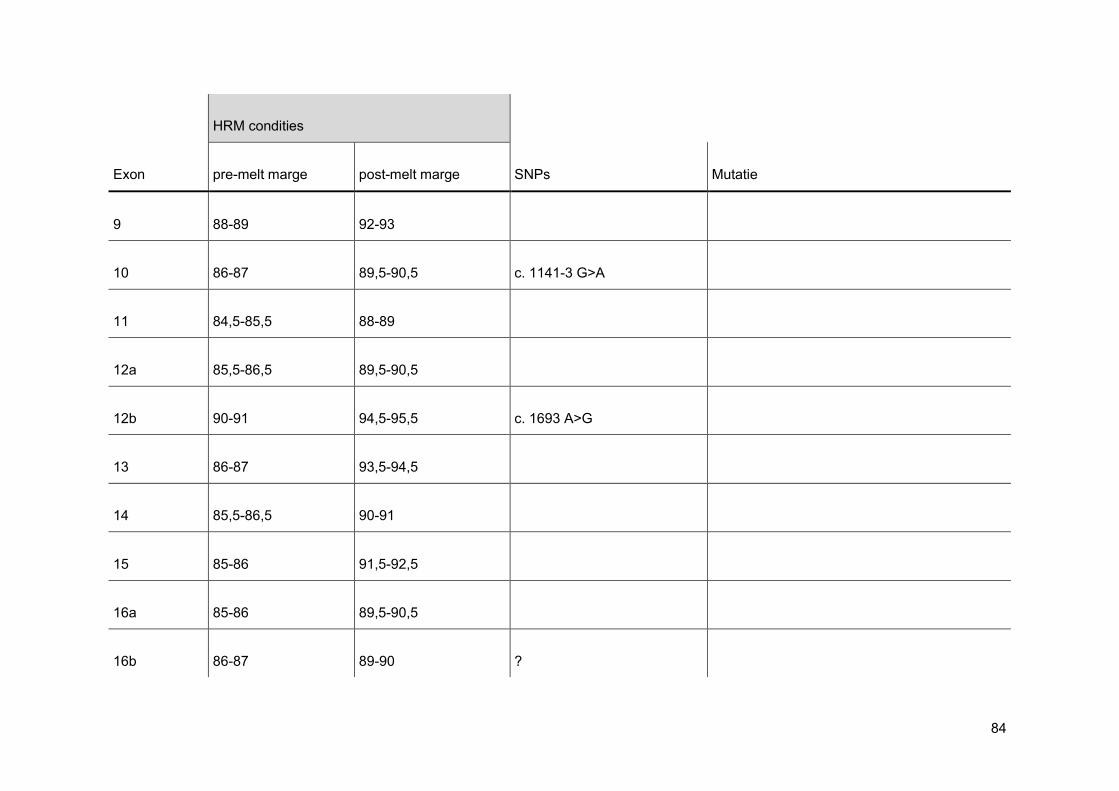
84
HRM condities
Exon pre-melt marge post-melt marge SNPs Mutatie
9 88-89 92-93
10 86-87 89,5-90,5 c. 1141-3 G>A
11 84,5-85,5 88-89
12a 85,5-86,5 89,5-90,5
12b 90-91 94,5-95,5 c. 1693 A>G
13 86-87 93,5-94,5
14 85,5-86,5 90-91
15 85-86 91,5-92,5
16a 85-86 89,5-90,5
16b 86-87 89-90 ?
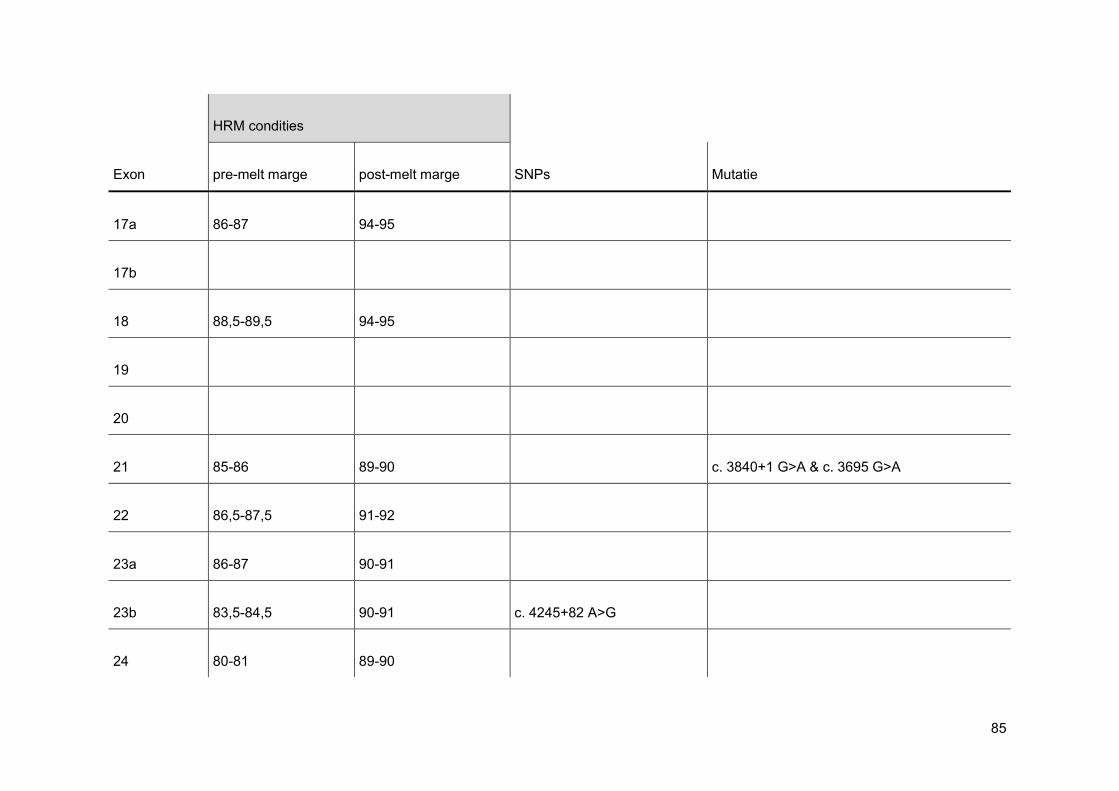
85
HRM condities
Exon pre-melt marge post-melt marge SNPs Mutatie
17a 86-87 94-95
17b
18 88,5-89,5 94-95
19
20
21 85-86 89-90 c. 3840+1 G>A & c. 3695 G>A
22 86,5-87,5 91-92
23a 86-87 90-91
23b 83,5-84,5 90-91 c. 4245+82 A>G
24 80-81 89-90
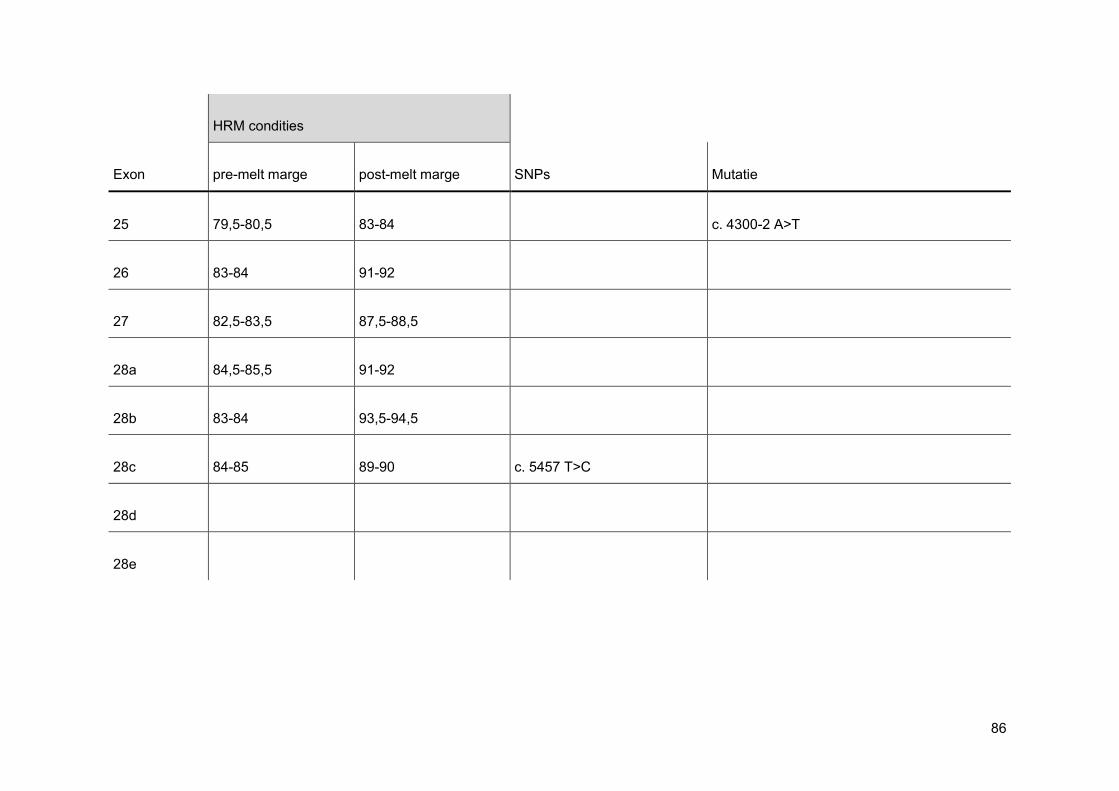
86
HRM condities
Exon pre-melt marge post-melt marge SNPs Mutatie
25 79,5-80,5 83-84 c. 4300-2 A>T
26 83-84 91-92
27 82,5-83,5 87,5-88,5
28a 84,5-85,5 91-92
28b 83-84 93,5-94,5
28c 84-85 89-90 c. 5457 T>C
28d
28e


















