
Bladluizen (Sitobion avenae) als vector voor de...
75
Faculteit Bio-ingenieurswetenschappen Vakgroep Gewasbescherming Academiejaar 2010-2011 Bladluizen (Sitobion avenae) als vector voor de transmissie van plantpathogene schimmels (Fusarium graminearum) Nathalie DE ZUTTER Promotor: Prof. dr. ir. G. SMAGGHE Copromotor: Prof. dr. ir. G. HAESAERT Masterproef voorgedragen tot het behalen van de graad van Master in de Bio-ingenieurswetenschappen: Landbouwkunde
Transcript of Bladluizen (Sitobion avenae) als vector voor de...

Faculteit Bio-ingenieurswetenschappen
Vakgroep Gewasbescherming
Academiejaar 2010-2011
Bladluizen (Sitobion avenae) als vector
voor de transmissie van plantpathogene schimmels
(Fusarium graminearum)
Nathalie DE ZUTTER
Promotor: Prof. dr. ir. G. SMAGGHE
Copromotor: Prof. dr. ir. G. HAESAERT
Masterproef voorgedragen tot het behalen van de graad van
Master in de Bio-ingenieurswetenschappen: Landbouwkunde

ii
De auteur en de promotor geven de toelating deze scriptie voor consultatie beschikbaar te
stellen en delen ervan te kopiëren voor persoonlijk gebruik. Elk ander gebruik valt onder de
beperkingen van het auteursrecht, in het bijzonder met betrekking tot de verplichting
uitdrukkelijk de bron te vermelden bij het aanhalen van de resultaten uit deze scriptie.
The author and the promotor give the permission to use this thesis for consultation and to
copy parts of it for personal use. Every other use is subject to the copyright laws, more
specifically the source must be extensively specified when using results from this thesis.
Gent
Juni 2011
De promotor, De copromotor,
Prof. dr. ir. Guy Smagghe Prof. dr. ir. Geert Haesaert
De auteur,
Nathalie De Zutter

iii
Woord vooraf
Langs deze weg wil ik iedereen bedanken die me geholpen heeft bij het verwezenlijken van
mijn thesis.
In de eerste plaats wil ik mijn promotor Prof. dr. ir. Guy Smagghe (Universiteit Gent,
vakgroep Gewasbescherming) bedanken voor het opvolgen van mijn thesis en het mogelijk
maken dat ik rond dit boeiende thema kon werken.
Deze thesis werd uitgevoerd in samenwerking met de Hogeschool Gent (Associatie
Universiteit Gent, campus Schoonmeersen, departement Biowetenschappen en
Landschapsarchitectuur). Daar verdienen vele mensen een woord van dank.
Prof. dr. ir. Geert Haesaert, mijn copromotor, bedank ik voor zijn raad tijdens de
vergaderingen. Ook bedankt voor het naleeswerk en de informatie die u mij gegeven hebt.
Een woord van dank gaat uit naar dr. ir. Kris Audenaert. Bedankt voor alles zo goed na te
lezen, voor alle uitleg omtrent statistische verwerking, om mij steeds met raad en daad bij te
staan,… Uw opbouwende kritiek is een belangrijke schakel geweest in het schrijven van dit
eindwerk.
Tot slot bedank ik nog vele andere mensen met de gedachte „alle beetjes helpen‟:
Dr. ir. Isabelle Vantornhout verdient een dankwoord voor haar uitleg omtrent de Jackknife
methode. Ik bedank ook Ellen Ydens voor haar informatie en tips, Nick Deschuyffeleer om
mij wegwijs te maken in het „schimmelkot‟ en Sofie Landschoot voor haar opzoekingswerk in
de database. I also thanks Mohamad Hamshou for helping me with the assay of intrinsic
growth of aphids. Ook bedankt aan Annelies De Roissart voor haar inzet voor mijn thesis en
voor alle informatie die ze me gegeven heeft. Jammer dat onze samenwerking niet langer kon
duren.
Gent, juni 2011,
Nathalie De Zutter

iv
Inhoudsopgave
1. Inleiding ............................................................................................................................. 1
2. Literatuurstudie ................................................................................................................ 3
2.1. Bladluizenproblematiek bij tarwe ........................................................................................... 3
2.1.1. Inleiding ......................................................................................................................... 3
2.1.2. Schadebeeld .................................................................................................................... 3
2.1.3. Epidemiologie................................................................................................................. 4
2.1.4. Algemene levenscyclus .................................................................................................. 4
2.1.5. Belangrijkste bladluissoorten op graangewassen ............................................................ 5
2.1.5.1. Sitobion avenae, de grote graanluis......................................................................... 6
2.1.5.2. Metopolophium dirhodum, de roos-grasluis ............................................................ 7
2.1.5.3. Rhopalosiphum padi, de vogelkersluis .................................................................... 7
2.2. Fusariumziekte-complex bij tarwe ......................................................................................... 8
2.2.1. Inleiding ......................................................................................................................... 8
2.2.2. Opbrengst- en kwaliteitsverliezen ................................................................................... 9
2.2.3. Mycotoxineproductie ...................................................................................................... 9
2.2.4. Taxonomie en levenscyclus van Fusarium graminearum ............................................. 10
2.2.5. Resistentie van tarwe tegen FHB .................................................................................. 12
2.3. Signaaltransductie in plant-pathogeen- en plant-insect- interactie ........................................ 13
2.3.1. Constitutieve en induceerbare afweer ........................................................................... 13
2.3.2. Herkenning van schimmel of insect .............................................................................. 13
2.3.2.1. Inleiding ................................................................................................................ 13
2.3.2.2. Herkenning schimmel ........................................................................................... 13
2.3.2.3. Herkenning insecten ............................................................................................. 15
2.3.3. Complexiteit van plantafweer t.a.v. schimmel- of insectenaanval ................................ 15
2.3.3.1. Plantafweer t.a.v. schimmels................................................................................. 15
2.3.3.2. Plantafweer t.a.v. insecten .................................................................................... 17
2.4. Interactie tussen insecten en pathogenen .............................................................................. 20
2.4.1. Insecten als vector van pathogenen ............................................................................... 20
2.4.1.1. Virussen ................................................................................................................ 20
2.4.1.2. Bacteriën ............................................................................................................... 21
2.4.1.3. Schimmels ............................................................................................................ 21
3. Materiaal en methoden ................................................................................................... 23
3.1. Studie-organismen ................................................................................................................ 23

v
3.1.1. Tarwezaden en hun oorsprong ...................................................................................... 23
3.1.2. Kweek van bladluissoorten S. avenae en M. dirhodum ................................................. 23
3.1.3. Schimmel F. graminearum (GFP transformant) ........................................................... 23
3.1.3.1. Inoculatie van F. graminearum op PDA medium ................................................. 23
3.1.3.2. Sporenoplossing van F. graminearum maken ....................................................... 24
3.1.3.3. Controle van groei van GFP transformant (F. graminearum) m.b.v.
fluorescentiemicroscoop ........................................................................................................... 24
3.2. Preferentietesten ................................................................................................................... 24
3.2.1. Aanmaken van steriele tarwekiemlingen ...................................................................... 24
3.2.2. Uitzaaien van tarwe en productie van aren ................................................................... 25
3.2.3. Ontwikkeling van preferentietesten .............................................................................. 25
3.2.3.1. Met kiemlingen ..................................................................................................... 25
3.2.3.2. Met aren ................................................................................................................ 25
3.2.3.3. Met behandelde aren ............................................................................................. 26
3.2.3.4. Met artificieel dieet aangelengd met DON ............................................................ 26
3.3. Verspreiding F. graminearum door S. avenae ...................................................................... 27
3.3.1. Orale opname van macroconidia door S. avenae .......................................................... 27
3.3.1.1. Medium met antibiotica aanmaken ....................................................................... 28
3.3.2. Uitwendige acquisitie en verspreiding F. graminearum door S. avenae ....................... 28
3.3.2.1. Wateragar aanmaken............................................................................................. 29
3.4. Overleving A. pisum en S. avenae op artificieel dieet met DON .......................................... 29
3.4.1. Acyrthosiphon pisum, de erwtenbladluis ....................................................................... 29
3.4.2. Proefopzet met bladluissystemen .................................................................................. 30
3.5. Reproductie S. avenae op artificieel dieet met DON ............................................................ 30
3.6. Statistische dataverwerking .................................................................................................. 31
4. Resultaten ........................................................................................................................ 32
4.1. Preferentietesten tussen tarwerassen ..................................................................................... 32
4.1.1. Met kiemlingen ............................................................................................................. 32
4.1.1.1. Preferentie ............................................................................................................. 32
4.1.1.2. Snelheid van keuze ............................................................................................... 33
4.1.2. Met aren ........................................................................................................................ 34
4.1.2.1. Preferentie ............................................................................................................. 34
4.1.2.2. Snelheid van keuze ............................................................................................... 34
4.1.3. Vergelijking voor S. avenae tussen aren en kiemlingen ................................................ 34
4.2. Aantrekking S. avenae tot DON ........................................................................................... 35
4.2.1. Preferentietesten met behandelde aren .......................................................................... 35

vi
4.2.1.1. Preferentie op korte termijn .................................................................................. 35
4.2.1.2. Snelheid van keuze ............................................................................................... 36
4.2.1.3. Preferentie op lange termijn .................................................................................. 37
4.2.2. Preferentietesten met artificieel dieet met DON ........................................................... 37
4.2.3. Overleving A. pisum en S. avenae op artificieel dieet met DON ................................... 39
4.2.4. Reproductie S. avenae op artificieel dieet met DON..................................................... 40
4.3. Verspreiding F. graminearum door S. avenae ...................................................................... 44
4.3.1. Orale opname van macroconidia door S. avenae .......................................................... 44
4.3.2. Metingen van stiletten S. avenae en M. dirhodum ........................................................ 44
4.3.3. Uitwendige acquisitie en verspreiding F. graminearum door S. avenae ....................... 44
5. Discussie ........................................................................................................................... 47
5.1. Preferentietesten tussen tarwerassen ..................................................................................... 47
5.1.1. Preferentie .................................................................................................................... 47
5.1.2. Snelheid van keuze ....................................................................................................... 48
5.1.3. Link preferentie – gevoeligheid voor aarfusarium ........................................................ 49
5.2. Aantrekking van S. avenae tot DON ..................................................................................... 50
5.2.1. Preferentietesten met behandelde aren .......................................................................... 50
5.2.2. Preferentietesten met artificieel dieet met DON ........................................................... 51
5.2.3. Overleving A. pisum en S. avenae op artificieel dieet met DON ................................... 52
5.2.4. Reproductie S. avenae op artificieel dieet met DON..................................................... 53
5.3. Verspreiding F. graminearum door S. avenae ...................................................................... 55
5.3.1. Orale opname van macroconidia door S. avenae .......................................................... 55
5.3.2. Uitwendige acquisitie en verspreiding F. graminearum door S. avenae ....................... 55
6. Conclusie .......................................................................................................................... 57
7. Samenvatting en toekomstperspectieven ...................................................................... 59
8. Summary .......................................................................................................................... 61
9. Referenties ....................................................................................................................... 62

vii
Lijst met afkortingen
A adult
ABA abscisinezuur
Avr avirulentiegenen
BYDV Barley Yellow Dwarf Virus
C artificieel dieet aangelengd met 0 mg/l DON
CAD Cadenza
CON aren geïnfecteerd met gedestilleerd water
CYDV Cereal Yellow Dwarf Virus
Da artificieel dieet aangelengd met 0,25 mg/l DON
DAS diacetoxyscirpenol
Db artificieel dieet aangelengd met 1 mg/l DON
DG18 dichloram glycerol agar
DON deoxynivalenol
DON aren geïnfecteerd met het mycotoxine DON
DT verdubbelingstijd
EAG elektroantennogram
EBF (E)-β-farnesene
ET ethyleen
ETI Effector Triggered Immunity
ETS Effector Triggered Susceptibility
FHB Fusarium Head Blight
FUS aren geïnfecteerd met sporen van F. graminearum (GFP)
FX fusarenone-X
GA giberellinezuur
GFP Green Fluorescent Protein
HOM Homeros
HR Hypersensitieve Respons
JA jasmonijnzuur
LEX Lexus
lx leeftijdspecifieke overleving
MeJA methyljasmonaat
mx leeftijdsspecifieke vruchtbaarheid

viii
N1 N4 1e 4
e nimfale stadium
NIV nivalenol
NCBI National Center for Biotechnology Information
NO stikstofmonoxide
PAL Fenylalanine ammonia-lyase
PAMP Pathogen-Associated Molecular Pattern
PDA Potato Dextrose Agar
PI proteïnase-inhibitors
PLRV potato leafroll virus
POD peroxidasen
PPO polyfenoloxidasen
PR pathogenese-gerelateerd
PRR Pathogen Responsive Receptors
PTI PAMP Triggered Immunity
PVY potato virus Y
R resistentiegenen
R0 netto vermenigvuldigingssnelheid
rm intrinsieke groeisnelheid
ROS Reactive Oxygen Species
SA salicylzuur
SAH Sahara
SAR Systemic Acquired Resistance
SPSS Statistical Package for Social Sciences
T gemiddelde generatietijd
TUL Tulsa
VBA Visual Basic for Applications
λ finale groeisnelheid

1
1. Inleiding
De voorspelling dat het wereldwijde bevolkingsaantal snel groeit, impliceert dat de
voedselproductie aanzienlijk zal moeten stijgen om aan de vraag naar voedsel te kunnen
voldoen (Zhu-Salzman et al., 2005). Tarwe (Triticum spp.) is voor de mens één van de
belangrijkste voedselbronnen. Het levert meer dan 20 procent van de calorieën en proteïnen in
de menselijke voeding. Dit gewas is in meer dan 40 landen en voor meer dan 35 procent van
de wereldbevolking het hoofdvoedsel. Het belang van tarwe steunt op de brede genetische
basis waardoor het een goede adaptatie bezit t.a.v. verschillende hoogtes, klimaatscondities en
bodemcondities. Dit laat toe om tarwe te telen onder diverse omgevingsomstandigheden.
Tevens is tarwe makkelijk te transporteren en kan voor lange tijd opgeslagen worden wanneer
de bewaarcondities goed zijn. Het graan wordt verwerkt tot brood en andere producten. Zo is
T. aestivum geschikt als broodgraan terwijl durumtarwe zeer geschikt is voor de productie van
pasta‟s. Tarwe is daarenboven een belangrijke grondstof die in industriële processen verwerkt
wordt tot zetmeel, gluten en alcohol. Het gewas is eveneens van groot belang in de
veevoederindustrie (Bushuk, 1998). Om een stijging in de voedselproductie te bereiken, wordt
er een efficiëntere agrarische productie geëist. Deze gaat gepaard met een optimale
teelttechniek en gewasbescherming. De impact van onkruidconcurrentie, schimmel- en
insectenschade moet zo laag mogelijk gehouden worden en hierbij komt de maatschappelijke
druk om het gebruik van pesticiden te beperken (El-Wakeil et al., 2010).
Bladluizen kunnen schade veroorzaken bij wintertarwe (Triticum aestivum L.). De drie
belangrijkste bladluissoorten op tarwe zijn de grote graanluis (Sitobion avenae F.), de roos-
grasluis (Metopolophium dirhodum Wlk.) en de vogelkersluis (Rhopalosiphum padi L.) (van
Emden & Harrington, 2007). Zoals vele bladluizen brengen deze aan de tarwebladeren
zuigschade aan, waardoor gele vlekken ontstaan. Ze scheiden honingdauw uit waarin
roetdauwschimmels zich ontwikkelen die de fotosynthese van de plant kunnen hinderen.
Tevens kunnen ze door te zuigen aan de groeiende korrels de korrelvulling negatief
beïnvloeden (Stijger, 2005).
Tarwe is tevens gevoelig voor talrijke schimmelziekten (roesten, meeldauw,
bladvlekkenziekte, aarziekte, etc.). Een belangrijke ziekte op granen is aarfusarium (of
Fusarium head blight (FHB)). Verscheidene schimmelsoorten van het geslacht Fusarium
vormen een complex (Goswami & Kistler, 2004) dat zowel kiemplantenziekte, voetziekte als
aaraantastingen kan veroorzaken (Kema et al., 2002). In eenzelfde aar kunnen meerdere
Fusariumsooren worden teruggevonden en de samenstelling van de populatie kan verschillen
naargelang graansoort, teeltsysteem en omgevingsomstandigheden. Problemen met Fusarium
spp. verschillen per plaats, jaar en teeltsysteem (Kema et al., 2002). Ook
populatieverschuivingen in het Fusariumziekte-complex zijn mogelijk (Audenaert et al.,
2009). Fusarium graminearum is één van de belangrijkste soorten uit het complex binnen
Europa. Deze pathogeen zorgt ervoor dat de graankorrels verschrompelen en kan in een paar
weken tijd, bij voor de schimmel gunstige weersomstandigheden en een gevoelig
tarwegenotype, een volledige oogst vernielen. De economische graanverliezen kunnen hoog
oplopen en zijn afhankelijk van de Fusariumsoort die de aantasting veroorzaakt, het genotype
van de tarwe en de omgevingscondities (McMullen et al., 1997). Naast rechtstreekse
economische schade produceren Fusarium spp. ook verschillende mycotoxines. Dit zijn

2
verdedigingsstoffen of virulentiefactoren die door schimmels worden geproduceerd (Van
Outryve, 2007a). Vaak heeft elke soort zijn eigen mycotoxinepatroon (Vogelgsang et al.,
2008). Deoxynivalenol, een mycotoxine geproduceerd door onder andere F. graminearum,
kan nadelige gevolgen hebben voor de gezondheid van mens en dier (McMullen et al., 1997).
Door het gezondheidsrisico dat met mycotoxines gepaard gaat en de economische verliezen
wordt er veel aandacht besteedt aan FHB resistentie in tarwe (McMullen et al., 2008).
Schimmels, maar ook virussen en bacteriën, kunnen verspreid worden door insecten en deze
hebben vaak een verwoestend effect op de plant (El-Hamalawi, 2007). Zo kunnen
oevervliegen Fusarium oxysporum f.sp. basilici verspreiden op veldsla en Fusarium
avenaceum verspreiden op Lisianthus (El-Hamalawi & Stanghellini, 2005).
De transmissie van plantpathogene schimmels door insecten is belangrijk bij de
schimmelverspreiding (El-Hamalawi, 2007). In deze thesis wordt de tritrofe interactie tussen
de tarweplant, de bladluissoort S. avenae en de schimmel F. graminearum onderzocht.
Terwijl de wintertarwe zich ontwikkelt, wordt deze aangevallen door bladluizen. In het
aarstadium kan de plant geïnfecteerd worden door de schimmel. Maar wat is de interactie
tussen de bladluis en deze schimmel? Concreet wordt nagegaan of S. avenae de verspreiding
van F. graminearum in de hand werkt. Er wordt in deze thesis getracht om een antwoord te
formuleren op volgende vragen: voelen de bladluizen zich aangetrokken tot zieke planten en
prefereren ze tarwecultivars die vaak geïnfecteerd worden met Fusarium spp.? Kan deze
bladluissoort de schimmel uitwendig aan zijn lichaam meedragen en zijn deze bladluizen in
staat om schimmelsporen op te zuigen en dus deze inwendig op te nemen? Kunnen ze de
pathogeen overbrengen naar gezonde tarweplanten? Met behulp van een F. graminearum
isolaat die getransformeerd is met het Green Fluorescent Protein (GFP) kan de verspreiding
van deze schimmel door bladluizen gevolgd worden.

3
2. Literatuurstudie
2.1. Bladluizenproblematiek bij tarwe
2.1.1. Inleiding
Bladluizen van de familie Aphididae (Fig. 1), of de echte bladluizen, zijn kleine insecten van
2 tot 4 mm lang met een variërende kleur. Determinatie gebeurt aan de hand van verschillende
morfologische eigenschappen. Deze insecten hebben twee complexogen, 3 paar poten en een
rostrum, ook een zuigsnavel genoemd, die varieert in lengte. In het rostrum zitten stiletten die
dienst doen als zuigbuis en speekselbuis. Ze hebben lange antennes en helder doorzichtige
vleugels. Een bladluis beschikt ook over corniculi of siphunculi. Dit zijn structuren die zich
op het abdomen bevinden en zijn de uitmondingen van wasklieren. De klieren scheiden
wasexcreten uit die alarmferomonen bevatten. De cauda is het staartje aan het einde van het
abdomen (Tirry, 2009).
Fig. 1: Morfologie van een bladluis (Tirry, 2009)
2.1.2. Schadebeeld
Bladluizen kunnen tarwegewassen op verschillende manieren beschadigen. Meestal komen ze
voor op de groeipunten van een gewas, zoals de toppen van groeiende bladeren, omdat op die
plaatsen veel voedingsstoffen in de sapstroom zitten die nodig zijn voor de groei van de plant.
Bladluizen voeden zich met hun zuigende en stekende monddelen op het floëem. Dit zorgt
voor groeiremmingen bij de plant alsook voor omgekrulde bladeren met gele vlekken. Door
zich op de sapstroom te voeden, nemen de bladluizen teveel suikers op. Dit teveel aan suikers
scheiden ze uit via de anus onder de vorm van honingdauw. Deze vloeistof maakt het
gewasoppervlak kleverig. Hierin kunnen roetdauwschimmels groeien die met hun zwart
mycelium de bladeren van de plant bedekken en de fotosynthese hinderen. Hierdoor kan de
opbrengst van het gewas dalen (Stijger, 2005). Een aantal bladluissoorten komen ook voor op
de graankorrels. Niet alleen de graanopbrengsten dalen door bladluisbeschadiging, ook de
bakkwaliteit van het graan gaat erop achteruit door veranderingen in de proteïnesamenstelling (Poehling et al., 2007; El-Wakeil et al., 2010). Een andere, belangrijke, schadefactor van
bladluizen is virusoverdracht (zie 2.4.1.1.) (Katis et al., 2007; Peters et al., 2009).

4
2.1.3. Epidemiologie
Er bestaan zowel gevleugelde als ongevleugelde bladluizen (dimorfisme) (Braendle et al.,
2006). De vleugelloze individuen zijn de apterae, de bladluizen met vleugels zijn de alatae
(Müller et al., 2001). De verplaatsing van bladluizen kan van zowel intrinsieke als extrinsieke
oorsprong zijn. Intrinsieke krachten, die genetisch voorgeprogrammeerd zijn, zorgen ervoor
dat de bladluis zich verplaatst, bijvoorbeeld wegvluchten van een vijand of op zoek gaan naar
voedsel bij honger. Met extrinsieke krachten wordt bedoeld dat de bladluis zich verplaatst
door stimuli uit de omgeving, bijvoorbeeld alarmferomonen uitgescheiden door andere
soortgenoten (Irwin et al., 2007). Als het dimorfisme door de omgeving wordt gestimuleerd,
wordt dit vleugelpolyfenisme genoemd (Braendle et al., 2006).
Gevleugelde bladluizen ontstaan meestal door te hoge populatiedensiteiten, door een gebrek
aan voedsel op de waardplant of om migratie van een primaire naar een secundaire
waardplant mogelijk te maken (Braendle et al., 2006). In de herfst speelt bij sommige
bladluissoorten ook de fotoperiode, langere nachten en kortere dagen, een rol (Williams &
Dixon, 2007). In mindere mate zet een lager wordende temperatuur ook aan tot de vorming
van gevleugelde bladluizen. Bij te hoge populatiedensiteiten is het vooral het fysieke contact
die de overgang stimuleert. Ook door het verschijnen van natuurlijke vijanden of de
aanwezigheid van pathogenen in de plant kan de overgang ontstaan (Müller et al., 2001). Al
deze factoren verschillen uiteraard van soort tot soort en zijn van belang voor
voorspellingsmodellen van bladluispopulaties (Williams & Dixon, 2007).
Algemeen wordt vleugelpolyfenisme beschouwd als een aanpassing aan veranderende
omgevingsomstandigheden, wat de verspreidingscapaciteit van het insect doet stijgen en hen
toelaat om te ontsnappen aan ongunstige omgevingen (Castañeda et al., 2010).
Een andere verplaatsing is de niet-intentionele verplaatsing. De bladluis wordt, zonder dat het
zijn bedoeling is, verplaatst naar de sink door de zwaartekracht, luchtstromingen,
voorbijkomende dieren, landbouwwerktuigen, etc. (Irwin et al., 2007).
2.1.4. Algemene levenscyclus
De voortplanting bij bladluizen kan complex zijn. Bladluizen kunnen zich ongeslachtelijk
voortplanten via parthenogenese, maar een geslachtelijke voortplanting bestaat ook (Dedryver
et al., 1998). Bij een ongeslachtelijke voortplanting (anholocyclisch) zullen de bladluizen
continue vivipaar zijn en de nimfen levend baren. Een holocyclische voortplanting houdt in
dat de bladluizen zich vermenigvuldigen door afwisselend geslachtelijke bevruchting
(oviparie, eileggend) en parthenogenese (Simon et al., 2002; McLean et al., 2009). Er bestaan
ook nog tussenliggende vormen van voortplanting. Hierbij zullen pathenogenetische wijfjes
verschillende aantallen seksueel voortplantende bladluizen produceren (Dedryver et al.,
1998). Niet alleen hun voortplanting kan variëren, bladluizen hebben ook een
waardplantwissel. Bladluizen kunnen een éénhuizige (mono-oecisch) of een tweehuizige (di-
oecisch) levenscyclus hebben. Hierbij wordt verwezen naar het al dan niet hebben van een
wissel tussen hout- en kruidachtige waardplanten (Müller et al., 2001).
De tweehuizige levenscyclus (Fig. 2b) van de familie Aphididae begint in de herfst wanneer
er aan seksuele reproductie wordt gedaan op de primaire, meestal houtige, waardplant. De
eitjes die gelegd worden, moeten de winter overleven. In de lente komt uit een winterei de

5
fundatrix (stammoeder). Zij produceert op parthenogenetische wijze de eerste generatie(s)
vleugelloze wijfjes, de fundatrigenia. Deze produceren nieuwe bladluizen die naar de
secundaire, meestal kruidachtige, waardplant migreren en zich daar de hele zomer
pathenogenetisch voortplanten. Ze zijn meestal vivipaar waardoor er een snelle
vermenigvuldiging is. Later worden de bladluizen gevleugeld. De mannetjes en gynoparae
migreren terug naar de primaire waardplant. De gynoparae produceren seksuele vrouwtjes
(oviparae) die met de mannetjes paren en wintereitjes produceren (Williams & Dixon, 2007).
In een éénhuizige levenscyclus (Fig. 2a) zorgt de stammoeder voor de nieuwe vrouwelijke
generaties, die meestal ongevleugeld zijn en zich op pathenogenetische wijze voortplanten.
Deze blijven zich voeden op dezelfde soort waardplant of een zeer nauwe verwant van de
waardplant. In de herfst ontstaan sexuparae die zowel mannetjes als wijfjes produceren.
Dankzij geslachtelijke bevruchting van de oviparae met de mannetjes, kunnen er voor
overwintering wintereitjes geproduceerd worden (Williams & Dixon, 2007).
a b Fig. 2: Eénhuizige (a) en tweehuizige levenscyclus (b) (van Emden & Harrington, 2007)
2.1.5. Belangrijkste bladluissoorten op graangewassen
S. avenae, M. dirhodum en R. padi zijn de belangrijkste bladluizen die tarwe kunnen
aantasten. Ze zijn allen ondergebracht in de subfamilie Aphidinae (Fig. 3).
Fylum: Arthropoda
Klasse: Insecta
Orde: Hemiptera
Suborde: Sternorrhyncha
Superfamilie: Aphidoidea
Familie: Aphididae
Subfamilie: Aphidinae
Stam: Macrosiphini
Geslacht: Sitobion
Soort: avenae
Geslacht: Metopolophium
Soort: dirhodum
Stam: Aphidini
Geslacht: Rhopalosiphum
Soort: padi Fig. 3: Taxonomie van de belangrijkste bladluissoorten op tarwe (Blackman & Eastop, 2007; PaDIL, 2010)

6
In Europa koloniseren deze drie bladluissoorten vooral wintertarwe. Hierop kunnen ze
simultaan voorkomen, toch prefereert S. avenae de tarwearen, terwijl R. padi en M. dirhodum
vooral de bladeren verkiezen. Bladluizen kunnen eind april te zien zijn op wintertarwe.
Zelden ontstaan er hoge bladluispopulaties vòòr de aarvorming eind mei. De opbouw van de
bladluispopulaties is gecorreleerd met de rijping van de waardplant en de reproductie van de
bladluizen daalt aanzienlijk na het deegrijpstadium van de aren. Daarom ligt de kritische
periode vooral in de lente en het begin van de zomer en duurt deze niet langer dan zes weken.
De populatiedensiteit blijft laag tot aan de bloei, waarna een exponentiële stijging volgt. De
hoge densiteit aan individuen op dat moment blijft slechts korte tijd behouden door de
verrotting van de plant en de komst van natuurlijke vijanden. Daarna verschijnen de
bladluizen terug in de herfst op nieuw gezaaide wintertarwe (Poehling et al., 2007).
2.1.5.1. Sitobion avenae, de grote graanluis
Het geslacht Sitobion bevat Europees-Aziatische en Afrikaanse bladluissoorten waarvan het
grootste deel zich voedt op grassen. De grote graanluis, Sitobion avenae Fabricius (Fig. 4a),
heeft geen waardplantwissel en is dus éénhuizig (Blackman & Eastop, 2007). S. avenae,
waarschijnlijk Europees van oorsprong, komt voor in Noordwest-Europa en in het
mediterrane gebied, het oosten van Indië en Nepal, het noorden en zuiden van Afrika en in
Noord- en Zuid-Amerika (Blackman & Eastop, 2007). Volgens PaDIL (2010) komt de soort
zelden voor in tropische klimaten en helemaal niet in Australië. Deze bladluizen hebben vier
nimfale stadia met ongeveer dezelfde kleur als de adulten maar ze hebben nog niet de
morfologische kenmerken ontwikkeld om precies gedetermineerd te worden. De eerste twee
stadia beschikken over antennen met vijf segmenten, de laatste twee stadia beschikken over
antennen met zes segmenten. In sommige gevallen kunnen de mannelijke nimfen herkend
worden aan hun bruinachtige kleur die ook nog te zien is als ze adult zijn (PaDIL, 2010).
De apterae op grassen en granen zijn spoelvormig. De kleur varieert van geel-groen naar
roodbruin, soms blinkend. Ze hebben zwarte antennen en siphunculi met een bleke cauda. De
apterae koloniseren alle soorten Poaceae, alle granen en weidegrassen in gematigde gebieden
en andere monocotyle planten. Ze kunnen zich over grote afstanden verspreiden (Blackman &
Eastop, 2007). De derde en vierde nimfale stadia en de apterae van S. avenae zijn mobiel en
verplaatsen zich frequent. Ze blijven gemiddeld 1,9 dagen op eenzelfde aar en verplaatsen
zich gemiddeld twee meter per dag. De gevleugelde vormen bewegen ook doorheen het
gewas in hun late nimfale stadia‟s en worden door de luchtbeweging verspreid als ze adulten
zijn (PaDIL, 2010).
De soort legt wintereitjes maar bij milde winters kunnen ze overleven via pathenogenese
(Blackman & Eastop, 2007). Zeer koude winters kunnen een hoge of zelfs volledige
mortaliteit teweegbrengen bij pathenogenetische graanluispopulaties (Helden & Dixon, 2002).
Wintereitjes daarentegen zijn kouderesistent en hun mortaliteit is onafhankelijk van de
winterkoude (Helden & Dixon, 2002; Simon et al., 2002). Zo kon aangetoond worden dat in
het Verenigd Koninkrijk de frequentie van anholocyclische voortplanting daalt en de
frequentie van holocyclische voortplanting stijgt als richting het noorden wordt gegaan. Dit is
te verklaren doordat een holocyclische voortplanting wintereitjes produceert die beter tegen
koude bestand zijn dan actieve bladluisvormen (Loxdale & Lushai, 2007).

7
Directe schade wordt veroorzaakt door bladluizen die zich voeden op de aren van
graangewassen (Blackman & Eastop, 2007). Populaties die groot genoeg zijn, kunnen
nutriënten uit de plant verwijderen wat kan leiden tot minder aren, minder graankorrels per
aar en een gereduceerd korrelgewicht. Behalve een geelverkleuring van de bovenste bladeren
en aren, wat een symptoom is bij vele bladluissoorten en plantpathogenen, zijn er geen
duidelijke symptomen zichtbaar (PaDIL, 2010). De indirecte schade houdt het overbrengen
van virussen in zoals het dwergvergelingsvirus op diverse graansoorten (barley yellow dwarf
virus, BYDV), zowel binnen als tussen de verschillende graansoorten (Blackman & Eastop,
2007) en het dwergmozaïekvirus op maïs (maize dwarf mozaic virus) (PaDIL, 2010).
2.1.5.2. Metopolophium dirhodum, de roos-grasluis
De roos-grasluis, Metopolophium dirhodum Walker (Fig. 4b) werd het eerst gedocumenteerd
in 1981 in Nieuw-Zeeland, waarschijnlijk geïntroduceerd door menselijke handel via schepen
en vliegtuigen of via het transport van commerciële gewassen en stockages van plantaardig
materiaal zoals aardappelpootgoed (Loxdale & Lushai, 2007). Deze soort is wijd verspreid
maar komt niet voor in de tropen (Poehling et al., 2007). De bladluis is lichtgroen met
lichtgekleurde poten en komt vrijwel uitsluitend voor op bladeren en stengels (Wittouck et al.,
2006). Ze hebben een waardplantwissel van Rosaceae naar grassen (Loxdale & Lushai, 2007).
Op de primaire waardplanten gebeurt overwintering onder de vorm van eitjes (Wittouck et al.,
2006). De soort kan BYDV verspreiden (Hermans et al., 2009). M. dirhodum kan voor grote
schade zorgen doordat deze bladluis toxische stoffen injecteert in maïsplanten. Dit wordt
toxemiase genoemd. Als gevolg hiervan hebben de planten een groeivertraging en een minder
ontwikkeld wortelgestel. Er ontstaan wit-gele strepen op de bladeren en misvormde bladeren
die later ook kunnen scheuren. Vooral laat gezaaide maïspercelen lopen gevaar (Verhiest,
2009).
2.1.5.3. Rhopalosiphum padi, de vogelkersluis
De vogelkersluis, Rhopalosiphum padi Linnaeus (Fig. 4c), is een belangrijke plaag die
wereldwijd terug te vinden op alle belangrijke graangewassen en weidegrassen. Deze bladluis
komt voor op verscheidene soorten Poaceae en andere monocotyle planten. Zijn primaire
waardplant is de vogelkers (Prunus padus). De apterae zijn ovaal en hebben verscheidene
tinten groen als kleur, vaak met roestkleurige vlekken (patches) rond de siphunculi. Het is een
vector van vele virussen. Het kan onder andere het gele dwergvirus op graan (cereal yellow
dwarf virus, CYDV) en het bladvlekkenvirus op maïs (maize leaf fleck virus) overdragen
(Blackman & Eastop, 2007).
a b c Fig. 4: Grote graanluis (Sitobion avenae) (a), de roos-grasluis (Metopolophium dirhodum) (b) en de
vogelkersluis (Rhopalosiphum padi) (van Emden & Harrington, 2007) (c)

8
2.2. Fusariumziekte-complex bij tarwe
2.2.1. Inleiding
Fusarium head blight (FHB) is in vele gebieden van de wereld een economisch belangrijke
ziekte bij granen. Deze ziekte wordt veroorzaakt door een complex van schimmels waarvan in
Europa F. avenaceum, F. culmorum, F. graminearum, F. poae en Microdochium nivale
(vroeger F. nivale) de voornaamste zijn (Waalwijk et al., 2003).
In eenzelfde aar kunnen dus meerdere Fusariumsoorten worden teruggevonden. De
samenstelling van de fusariumpopulatie kan verschillen door externe factoren (Kema et al.,
2002). Een gecombineerd effect van geografie, weersomstandigheden, fungicide-
behandelingen, gewasrotatie, waardplantresistentie en grondbewerking bepalen het resultaat
en de samenstelling van de FHB populatie. Zo prefereert F. graminearum vooral hogere
temperaturen, terwijl F. culmorum, F. poae, F. avenaceum en M. nivale de neiging hebben om
te domineren in koudere regionen (Doohan et al., 2003).
In Nederland trad een verschuiving op van F. culmorum in de vroege jaren van 1990 naar F.
graminearum in 2000 en 2001 (Waalwijk et al., 2003). Een analoog fenomeen trad op in
Vlaanderen. In 2007 en 2008 kwam F. culmorum duidelijk minder frequent voor dan begin de
jaren 2000, terwijl F. poae het meest dominant was, gevolgd door F. avenaceum en M. nivale
in 2007 en gevolgd door F. graminearum in 2008 (Audenaert et al., 2009).
Symptomen van Fusarium spp. zijn afhankelijk van de Fusariumsoorten en het moment van
infectie. Afhankelijk van de soort kunnen ze kiemplantenziekte, voetziekte en aarfusarium
(kafjesrood) veroorzaken. Zo kan F. poae enkel aren infecteren (Kema et al., 2002) en
veroorzaken F. graminearum, F. pseudograminearum en F. culmorum onder meer voetziekte
(Backhouse & Burgess, 2002). Ook een Fusariumbesmetting van zaaizaad is mogelijk maar
draagt weinig bij tot het ontstaan van aarfusarium. Het kan de opkomst wel sterk negatief
beïnvloeden (Kema et al., 2002; Groot, 2003).
De meest bestudeerde en belangrijkste Fusariumsoort in Europa is F. graminearum. Vooral
graangewassen zoals tarwe zijn tijdens hun bloei gevoelig aan deze pathogeen. De schimmel
ontwikkelt zich in gematigde en semitropische gebieden bij frequente regenval en bij hoge
relatieve vochtigheden. Op tarwe verschijnen donkerbruine/zwarte vlekken op de kafjes van
de granen, waarnaar verwezen wordt als schurft (scab). Aarpakjes kleuren bleek na
aantasting, dit wordt blight genoemd (Fig. 5). Wit mycelium, dat later roos kleurt, koloniseert
de volledige aar (Goswami & Kistler, 2004). De eerste symptomen van FHB zijn te zien in
het midden van de aar omdat daar de bloei begint en de vochtigheid groter is (Walter et al.,
2010). Bij een aarinfectie ontstaan verkleurde en verschrompelde graankorrels (tombstones).
De korrels rijpen voortijdig af en de hele aar sterft. Deze schimmel kan in een paar weken tijd
een volledige oogst vernielen (McMullen et al., 1997).

9
Fig. 5: Aren met een verschillende mate van aantasting (Derycke et al., 2007)
2.2.2. Opbrengst- en kwaliteitsverliezen
Een infectie van enkele graankorrels kan leiden tot het afsterven van de gehele aar (Wang et
al., 2005). FHB kan graanverliezen doen oplopen van 30 tot 70 procent (Waalwijk et al.,
2003). Naast deze ernstige opbrengstverliezen, en de daarmee gepaard gaande economische
verliezen, is er ook een verminderde graankwaliteit. Een Fusariuminfectie kan zetmeelkorrels,
opslagproteïnen en celwanden in graankorrels aantasten en zorgt voor significante
veranderingen in koolhydraten, lipiden en de proteïnesamenstelling. De kwaliteit van de
opslagproteïnen en de verwerkingseigenschappen van het deeg worden hierdoor beïnvloed.
Door de degradatie van het glutengehalte, vertonen de geïnfecteerde tarwegranen een
verminderde bakkwaliteit (Wang et al., 2005). Naast een verlies aan bakkwaliteit treedt ook
een verminderd broodvolume op (Prangle et al., 2005).
Hoewel FHB symptomen een klassieke impact hebben op het duizend-korrelgewicht van de
granen en op de bak- en brouwkwaliteit (Prangle et al., 2005; Wang et al., 2005; Wang et al.,
2008), is de grootste zorg rond FHB de productie van mycotoxines.
2.2.3. Mycotoxineproductie
Mycotoxines zijn secundaire metabolieten die door schimmels worden geproduceerd. Tabel 1
geeft de mycotoxines weer die de verschillende Fusariumsoorten produceren.
Tabel 1: Mycotoxines van Fusariumsoorten op tarwe in Europa (Bottalico & Perrone, 2002)

10
Trichothecenen, een mycotoxinegroep geproduceerd door Fusarium spp., zijn tricyclische
sesquiterpenen (Doohan et al., 2003). Ze worden onderverdeeld in type-A trichothecenen,
zoals diacetoxyscirpenol (DAS), T-2 en HT-2, en type-B trichothecenen zoals deoxynivalenol
(DON), nivalenol (NIV) en fusarenone-X (FX). Deze categorieën zijn gebaseerd op de
afwezigheid (type-A) of de aanwezigheid (type-B) van een keto groep op C-8 van het
trichothecene skelet. F. sporotrichioides, F. poae en F. equiseti zijn type-A producenten,
terwijl F. graminearum en F. culmorum type-B trichothecenen produceren.
Studies tonen aan dat verschillende isolaten van dezelfde Fusariumsoort variërende
concentraties en types toxineproductie kunnen hebben. Verschillen in type toxineproductie
worden gebruikt om F. graminearum te groeperen in twee verschillende chemotypes die
ofwel primair DON en geacetyleerde vormen van DON produceren, ofwel primair NIV en/of
FX (Vogelgsang et al., 2008).
Toxiciteit van trichothecenen is te wijten aan hun vermogen om de eiwitsynthese te inhiberen.
Ze binden op de 60S subunit van eukaryotische ribosomen en tasten het peptidyltransferase
aan. Deze mycotoxinegroep inhibeert enerzijds de initiatie of de elongatie van de
peptideketen. DON is een voorbeeld van een elongatie-inhibitor (Goyarts et al., 2006).
Hierdoor veroorzaakt DON, ook vomitoxine genoemd, bij blootstelling aan hoge dosissen
braakneigingen en voedselweigering bij niet-herkauwers, maar ook bij andere dieren en de
mens vormt het een bedreiging voor de gezondheid (McMullen et al., 1997). Omdat toxines
hitte- en soms zuurbestendig zijn, breken ze niet af bij de verwerking van granen (Van
Outryve, 2007a). De Europese Commissie introduceerde daarom in 2006 een wetgeving
omtrent de maximaal toelaatbare gehaltes aan mycotoxines in granen en graanproducten voor
voedingsdoeleinden. Het maximumgehalte aan DON in onbewerkte granen bedraagt 1250
µg/kg. Granen bestemd voor menselijke consumptie, zoals meel en bloem, hebben een norm
van 750 µg/kg. In brood mag er niet meer dan 500 µg/kg DON zitten. Bij varkensvoeder is de
richtlijn 0,9 mg/kg. Bij herkauwers is dit 5 mg/kg (Van Outryve, 2007b).
2.2.4. Taxonomie en levenscyclus van Fusarium graminearum
F. graminearum Schwabe (teleomorf: Gibberella zeae (Schwein.) Petch.) (Fig. 6) produceert
DON, wat in Europa het voornaamste mycotoxine in graan is (Desjardins et al., 2004).
Superrijk: Eukaryota
Rijk: Fungi
Fylum: Ascomycota
Subfylum: Pezizomycotina
Klasse: Sordariomycetidae
Subklasse: Hypocreomycetidae
Orde: Hypocreales
Familie: Nectriaceae Geslacht: Gibberella
Soort: zeae Fig. 6: Taxonomie van F. graminearum gebaseerd op de geslachtelijk fase Gibberella zeae
(Goswami & Kistler, 2004)

11
Deze hemibiotrofe schimmel maakt zowel geslachtelijke als ongeslachtelijke structuren
tijdens zijn levenscyclus (Fig. 7). De geslachtelijke sporen (ascosporen) worden verspreid via
de wind. De ongeslachtelijke sporen (conidia) worden via regenspatten verspreid. De
banaanvormige ascosporen zijn verzameld in buisvormige zakjes of asci in het perithecium.
Ze vormen het primaire inoculum (Trail, 2009). F. graminearum wordt als een monocyclische
ziekte beschouwd, wat wil zeggen dat het primaire inoculum voor de ziekte zorgt, maar dat er
geen nieuw inoculum wordt aangemaakt in datzelfde groeiseizoen (Trail, 2009). Onder
condities die echter bevorderlijk zijn voor F. graminearum zullen de ascosporen bijdragen tot
een verhoogde inoculumdruk en zullen er twee of zelfs meerdere cycli per groeiseizoen
optreden (Obst et al., 2002).
De ascosporen worden via de wind verspreid (Trail, 2009) en bij landing op een bloeiende aar
van tarwe is adhesie van cruciaal belang. De sporen mogen dan niet meer verplaatst worden
door wind, regen of andere krachten en de plakkerige celwanden van de ascosporen spelen
hierin een rol. Daarna volgt de kieming van de sporen met de vorming van hyfen. De
schimmel dringt de plant binnen via natuurlijke openingen zoals de stomata of door directe
penetratie met behulp van zijn hyfen. Bij aarkafjes dringen ze onder andere binnen langs
spleten tussen de aan elkaar grenzende bovenste (palea) en onderste (lemma) kroonkafjes
(Walter et al., 2010). Vlak na infectie verspreidt de schimmel zich intercellulair in de plant en
begint ook de genexpressie voor het aanmaken van DON. DON veroorzaakt necrose van de
weefsels en laat de schimmel toe om zich ook in de aarspil te verspreiden. Later verspreidt de
schimmel zich intracellulair (Trail, 2009). Hij wordt necrotroof en doodt het plantenweefsel
op het moment dat de assimilatenstroom afneemt (Goswami & Kistler, 2004).
Ongeslachtelijke sporen (conidia) ontwikkelen zich op de oppervlakte van de geïnfecteerde
plantendelen. Deze worden door regenspatten verspreid of komen in het gewasresidu op het
veld terecht, waar ze overleven (saprofytisch) en de winter doorbrengen (Trail, 2009). In de
lente zorgen warme, vochtige weersomstandigheden voor de ontwikkeling van de conidia en
de vorming van perithecia waarin nieuwe asci en ascosporen geproduceerd worden (Goswami
& Kistler, 2004). Fusariumschimmels zijn ook in staat om overlevingsstructuren te
produceren (chlamydosporen) die lange tijd kunnen achterblijven in de bodem en als nieuwe
infectiebron dienen (Kema et al., 2002).
Fig. 7: Levenscyclus van F. graminearum (Trail, 2009)

12
2.2.5. Resistentie van tarwe tegen FHB
Tarwerassen kunnen variëren van zeer gevoelig tot bijna ongevoelig voor aarfusarium.
Hoewel een variatie van resistentie tegen FHB gekend is, is geen enkele tarwe- of
gerstvariëteit absoluut resistent (Walter et al., 2010). Er bestaan vijf types van resistentie. De
eerste twee worden algemeen herkend. Type I resistentie vermindert het aantal initiële
infecties (Bai & Shaner, 2004; McMullen et al., 2008; Walter et al., 2010). Eigenschappen
van de plant betreffende ontwikkeling, fysiologie, morfologie alsook biochemische
eigenschappen helpen mee om de aanhechting en infectie van de schimmel te voorkomen.
Planthoogte, tijdstip en duur van de bloei, aanwezigheid van kafnaaldjes en de compactheid
van aren zijn slechts enkele van die eigenschappen. Oppervlakten van de aren bedekt met was
zorgen ervoor dat er minder water beschikbaar is voor de kieming van conidia en dragen
eveneens bij tot de type I resistentie (Walter et al., 2010). Type II resistentie beperkt de
verspreiding van de schimmel vanuit de infectieplaats (Bai & Shaner, 2004; McMullen et al.,
2008; Walter et al., 2010). Deze resistentie houdt in dat de plant anatomische eigenschappen
heeft zoals vaatbundels met een kleinere diameter, compactere vaatbundels in de aarspil,
dikkere celwanden, etc. De plant biedt ook chemische weerstand door nieuw gesynthetiseerde
pathogeen-gerelateerde proteïnes en flavonoïdes en de biotransformatie van DON in het
minder giftige DON-3-glucoside (Walter et al., 2010). Type III resistentie houdt accumulatie
van DON in graankorrels tegen. Dit kan enerzijds te wijten zijn aan degradatie van DON door
plantenzymen gedurende de korrelontwikkeling. Anderzijds kunnen er hoge concentraties van
DON aanwezig zijn in de aarweefsels die geen graankorrels zijn, maar is DON niet in staat
om naar de korrels te migreren (Bai & Shaner, 2004). Type IV resistentie biedt weerstand
tegen graaninfectie en bevordert het behoud van graankorrelaantal en –grootte. Type V is
tolerantie (Bai & Shaner, 2004). Tolerante planten kunnen de opbrengst handhaven ondanks
de aanwezigheid van FHB en DON (McMullen et al., 2008).
Verbetering van FHB resistentie is een hoge prioriteit in veredelingsprogramma‟s van tarwe
en gerst en rassen met verschillende niveaus van resistentie tegen FHB zijn ontwikkeld
(McMullen et al., 2008). Tarweveredelingsprogramma‟s baseren zich sterk op resistentie
afgeleid van de genetische bronnen zoals Sumai 3, een zomertarweras van Aziatische
afkomst. Deze resistentiebron wordt wereldwijd gebruikt in tarwe (Bai & Shaner, 2004) en is
ontwikkeld uit Funo en een Taiwanese tarwe. Opmerkelijk is dat beide ouders matig resistent
zijn, waaruit geconcludeerd kan worden dat de resistentie van Sumai 3 berust op transgressie
(Kema et al., 2002). Sumai 3 heeft de hoogste graad van type II resistentie (Bai & Shaner,
2004) maar bezit ook type I resistentie (Kema et al., 2002). Het ras heeft een snellere
accumulatie van glucanases en chitinases dan gevoeligere rassen. Onderzoekers vinden dat
deze resistentiebron meer overerfbaar, stabiel en consistent is over verschillende omgevingen
dan de meeste andere bronnen (Bai & Shaner, 2004).

13
2.3. Signaaltransductie in plant-pathogeen- en plant-insect-
interactie
2.3.1. Constitutieve en induceerbare afweer
Planten hebben zowel constitutieve (passieve) als induceerbare (actieve) afweermechanismen.
In vergelijking met constitutieve afweermechanismen springen geïnduceerde mechanismen
efficiënter om met energie omdat constitutieve afweer continu aanwezig is en energie vergt.
Daarentegen komt de geïnduceerde afweer (wondrespons) enkel tot stand als de plant wordt
aangevallen door vijanden. Een afweer kan zowel structureel zijn, zoals fysische barrières die
plagen en pathogenen kunnen tegenhouden, als chemisch, zoals secundaire metabolieten die
vaak toxisch zijn voor plagen en pathogenen. Secundaire metabolieten zijn componenten die
niet vereist zijn voor normale plantengroei of -reproductie en zijn vaak uniek voor specifieke
plantensoorten (Howe & Jander, 2008; Zhao et al., 2009).
2.3.2. Herkenning van schimmel of insect
2.3.2.1. Inleiding
Planten zijn onderworpen aan aanvallen van een wijde waaier aan pathogenen en insecten
(Glazebrook, 2005). Er bestaan 120 schimmelgenera, 30 types virussen en 8 bacteriegenera
die verantwoordelijk zijn voor 11 000 ziekten die beschreven zijn bij planten (Ferreira et al.,
2006). Omdat voor de verschillende pathogenen complexe afweersystemen ontwikkelend zijn
door planten, wordt er dieper ingegaan op de reacties van een plant ten aanzien van
schimmels en insecten. Alvorens de plant een afweerrespons op gang kan brengen, moet deze
eerst zijn aanvaller herkennen.
2.3.2.2. Herkenning schimmel
Als een schimmel een plant tracht binnen te dringen, ontstaat er een strijd. Het doel van de
schimmel is om de plant ziek te maken. Het doel van de plant is om de schimmel buiten te
sluiten en ervoor te zorgen dat hij niet kan overgaan van extracellulaire groei naar invasieve
groei met parasitisme en ziekte tot gevolg. De afweermechanismen van planten tegen
schimmels kunnen pas efficiënt werken als de plant de schimmel snel kan herkennen.
Elicitoren spelen hierin een cruciale rol. Een elicitor is een term die gebruikt wordt om eender
welke molecule aan te duiden die een plantafweerreactie kan uitlokken. Deze kunnen zowel
van de schimmel komen (exogene elicitoren) maar kunnen ook componenten van de
waardplant zelf zijn, die gemodificeerd zijn door de schimmel (endogene elicitoren)
Afhankelijk van hun werkingsmechanisme kunnen elicitoren opgedeeld worden in aspecifiek
en specifiek (Ferreira et al., 2006).
Aspecifieke elicitoren kunnen onder andere glycoproteïnen, peptiden, koolhydraten en lipiden
zijn. Deze elicitoren zijn betrokken bij de eerste lijn van plantafweer (zie 2.3.3.1.). Hun
aanwezigheid is vaak constitutief. Ze worden vrijgesteld bij een aanval van schimmels en
signaleren de aanwezigheid van een potentiële pathogeen aan zowel een waard- als een niet-
waardplant, op een niet-cultivar-specifieke manier. Een aantal van hen kunnen beschouwd
worden als moleculaire handtekeningen van de schimmel (PAMP, Pathogen-Associated
Molecular Pattern). Dit zijn pathogeen-specifieke moleculaire sequenties of structuren die
14
vaak onmisbaar zijn voor de levensstijl van de pathogeen. Ze lokken bij de plant een receptor-
gemedieerde immuunrespons uit (Niks & Marcel, 2009). Een voorbeeld hiervan zijn de
elicitoren die vrijgesteld worden bij de enzymatische degradatie van de celwandpolymeren
van de schimmel wanneer de schimmel zijn weg in de waardplant maakt. Zulke elicitoren,
zoals oligomeren van chitine of glucanen, zijn onmisbaar voor de schimmel (Ferreira et al.,
2006). Chitine kan gedetecteerd worden door de chitine-bindende proteïnen en chitinases van
de plant. Glucaan door glucanases en thaumatine gelijkende proteïnen (Walter et al., 2010).
Specifieke elicitoren zijn uniek voor een bepaalde schimmel en lokken een zeer cultivar-
specifieke respons uit. Waar de fytopathologie het vroeger hield op de productie van
elicitoren (gecodeerd door avirulentiegenen (Avr) van de pathogeen) in een plant-pathogeen
interactie, en de daaropvolgende herkenning door receptoren (producten van de resistentie (R)
genen van een plant) in het zogenaamde gen-om-gen complex (Avr-R), heeft deze perceptie
een zeer grote evolutie ondergaan in de laatste jaren.
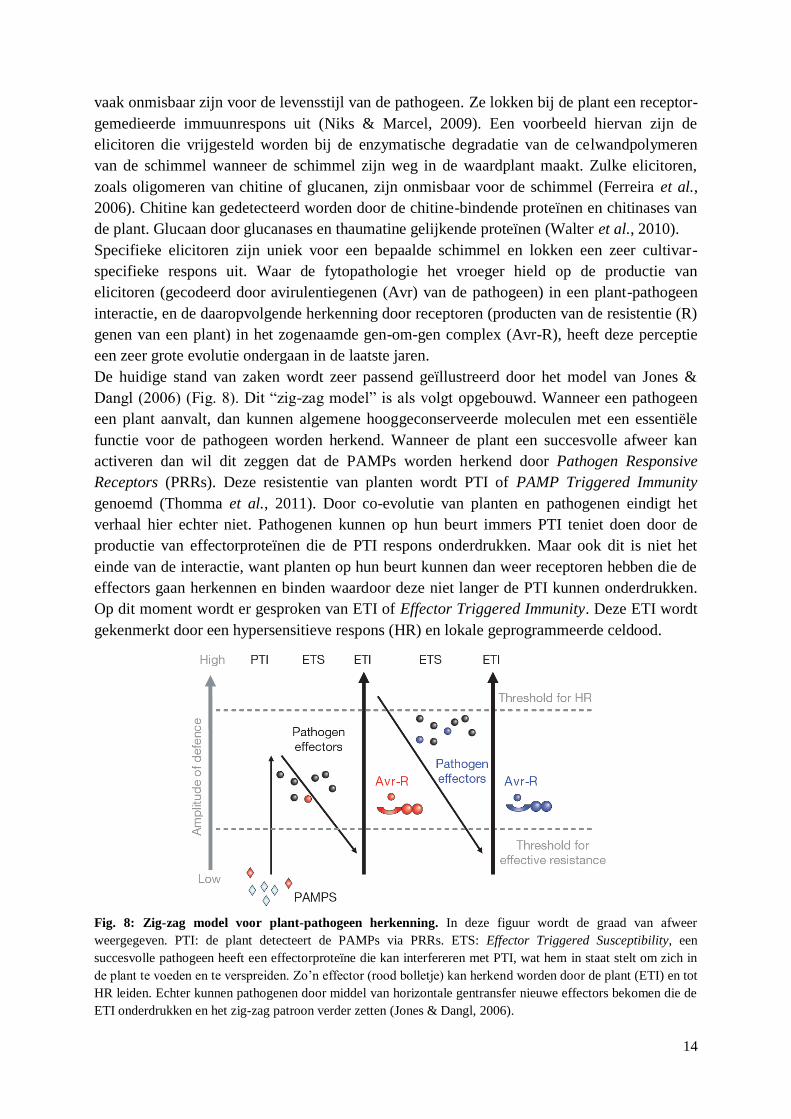
De huidige stand van zaken wordt zeer passend geïllustreerd door het model van Jones &
Dangl (2006) (Fig. 8). Dit “zig-zag model” is als volgt opgebouwd. Wanneer een pathogeen
een plant aanvalt, dan kunnen algemene hooggeconserveerde moleculen met een essentiële
functie voor de pathogeen worden herkend. Wanneer de plant een succesvolle afweer kan
activeren dan wil dit zeggen dat de PAMPs worden herkend door Pathogen Responsive
Receptors (PRRs). Deze resistentie van planten wordt PTI of PAMP Triggered Immunity
genoemd (Thomma et al., 2011). Door co-evolutie van planten en pathogenen eindigt het
verhaal hier echter niet. Pathogenen kunnen op hun beurt immers PTI teniet doen door de
productie van effectorproteïnen die de PTI respons onderdrukken. Maar ook dit is niet het
einde van de interactie, want planten op hun beurt kunnen dan weer receptoren hebben die de
effectors gaan herkennen en binden waardoor deze niet langer de PTI kunnen onderdrukken.
Op dit moment wordt er gesproken van ETI of Effector Triggered Immunity. Deze ETI wordt
gekenmerkt door een hypersensitieve respons (HR) en lokale geprogrammeerde celdood.
Fig. 8: Zig-zag model voor plant-pathogeen herkenning. In deze figuur wordt de graad van afweer
weergegeven. PTI: de plant detecteert de PAMPs via PRRs. ETS: Effector Triggered Susceptibility, een
succesvolle pathogeen heeft een effectorproteïne die kan interfereren met PTI, wat hem in staat stelt om zich in
de plant te voeden en te verspreiden. Zo‟n effector (rood bolletje) kan herkend worden door de plant (ETI) en tot
HR leiden. Echter kunnen pathogenen door middel van horizontale gentransfer nieuwe effectors bekomen die de
ETI onderdrukken en het zig-zag patroon verder zetten (Jones & Dangl, 2006).

15
2.3.2.3. Herkenning insecten
De activering van plantrespons wordt voorafgegaan door de snelle herkenning van
insectenschade door de plant. Enerzijds is deze herkenning gebaseerd op de gen-om-gen
hypothese en anderzijds op de herkenning van de door insecten gemodificeerde
plantcomponenten. Net zoals de herkenning van schimmels, is de herkenning van
insectenschade door planten gebaseerd op de gen-om-gen interactie tussen de elicitoren van
insecten en de resistentiegenen van de plant (Howe & Jander, 2008).
Elicitoren laten de plant toe om een onderscheid te maken tussen mechanische schade en
schade aangebracht door herbivoren. Weefselbeschadiging is niet altijd een betrouwbare
indicator voor een aanval door insecten. Om te vermijden dat de plant onnodig
afweermechanismen op gang brengt, die energie vergen, moet de plant een onderscheid
kunnen maken tussen schade aangebracht door insecten en mechanische schade door
bijvoorbeeld hagel of wind. Eerst en vooral herkennen de planten schade door insecten aan de
orale secreties van de insecten (Howe & Jander, 2008). Speeksel van insecten bevat namelijk
verscheidene hydrolytische enzymen die dienst doen als elicitoren. Voorbeelden van deze
enzymen zijn peroxidase en pectinase (Zhu-Salzman et al., 2005). Planten kunnen herbivoren
ook herkennen aan de elicitoren in de ovipositiesecretie die uitgescheiden wordt tijdens
ovipositie van herbivoren (Dicke et al., 2003; Steidle et al., 2005). Hoewel het grote aantal
insectensoorten dat afweerresponsen kunnen uitlokken, zijn er slechts een klein aantal plant-
insect combinaties bestudeerd. Er moet dus nog veel onderzoek gedaan worden naar orale
insectensecreties die waardplantafweerresponsen kunnen uitlokken (Howe & Jander, 2008).
De plant kan zijn beschadigd weefsel ook herkennen aan het feit dat herbivoren de
componenten in het plantenweefsel modificeren (Smith & Boyko, 2006). De gewonde
plantencellen zenden dan signaalmoleculen uit die een afweerrespons op gang brengen (Howe
& Jander, 2008). In tegenstelling tot de eerste manier van herkenning (gen-om-gen), die
specifiek van toepassing is op herbivoorresistente planten, komt deze tweede manier van
herkenning zowel voor bij herbivoorresistente als -gevoelige planten (Smith & Boyko, 2006).
2.3.3. Complexiteit van plantafweer t.a.v. schimmel- of insectenaanval
2.3.3.1. Plantafweer t.a.v. schimmels
Planten worden continu blootgesteld aan potentiële schimmels waarvan ze ziek kunnen
worden. Ziekte kan zich echter enkel voordoen als de waardplant gevoelig is voor een
bepaalde pathogeen, de pathogeen zelf virulent is en de omgevingsomstandigheden optimaal
zijn voor ziekteontwikkeling. Met andere woorden, niet alle schimmels kunnen elke plant ziek
maken en een plant kan ook niet gevoelig zijn voor alle soorten schimmels. Slechts een klein
deel van de schimmels zijn in staat om een plant te invaderen en ziek te maken. Tevens moet
opgemerkt worden dat schimmelpopulaties en waardplanten veranderen wanneer ze hun
ontwikkelingstadia doorlopen, wat kan bijdragen tot veranderingen in pathogeenvirulentie en
de waardplantgevoeligheid (Ferreira et al., 2006).
Schimmels zijn dus niet altijd in staat om de plant ziek te maken. Dit komt omdat planten
reeds miljoenen jaren samen evolueren met schimmels en omdat ze beide succesvol
chemische wapens creëren om elkaar aan te vallen. Elke keer dat de plant een nieuwe
defensieve innovatie ontwikkelt tegen een schimmel, vindt deze een manier om de afweer te
omzeilen. Deze complexe co-evolutie verklaart niet alleen de specificiteit tussen de schimmel

16
en zijn waardplant maar tevens ook de inefficiëntie of overtolligheid van sommige genen die
vaak coderen voor enzymen die overlappende activiteiten vertonen. Zo creëerden planten
complexe series van chemische en structurele barrières die op meerdere niveaus (multi-level)
werken en zowel constitutief als induceerbaar van aard kunnen zijn (Ferreira et al., 2006).
De constitutieve afweer van een plant is zowel structureel als chemisch. Deze afweer
beschermt tegen een niet-specifieke brede waaier aan schimmels.
De wasachtige cuticula en de celwanden van planten zijn fysische barrières (structureel)
waarnaar verwezen wordt als de eerst lijn van plantafweer. Ze beschermen namelijk tegen het
binnendringen van de schimmel (Lazniewska et al., 2010). Tarweresistentie tegen infectie van
Fusarium spp. is vaak gecorreleerd met de verdikking van de celwand. Celwandversterking
en een verhoogde lignine-inhoud worden verondersteld de instroom van Fusariumtoxines en
de uitstroom van plantnutriënten te beperken. Celwandversterkingen en lignificatie als reactie
op Fusarium spp. kunnen bekomen worden door polyfenoloxidasen (PPO) die de oxidatie van
polyfenolen tot quinonen katalyseren, en door peroxidasen (POD) die de oxidatie van fenolen
en lignificatie bevorderen. Infectie door F. graminearum en DON kan respectievelijk
resulteren in een verhoogde activiteit van POD in tarwezaailingen en POD genexpressie in
tarwearen. Deze enzymen zijn enerzijds constitutief aanwezig maar vertonen anderzijds een
verhoogde activiteit als de plant aangevallen wordt (geïnduceerde afweer) (Walter et al.,
2009).
Fenolen (chemisch) treden op als celwandversterkers en antischimmelcomponenten. Het
fenolische p-coumarinezuur is een precursor van vele afweercomponenten, zoals flavonoïden
en ligninecomponenten. Fenylalanine ammonia-lyase (PAL) katalyseert de eerste stap in de
fenylpropanoïdpathway. Deze pathway speelt eveneens een rol in de resistentie van tarwe
voor F. graminearum en DON (Walter et al., 2009). De plant beschikt tevens over vele andere
constitutieve secundaire metabolieten met een antischimmelwerking of metabolieten die de
binnendringende schimmel op een chemische manier afweren (Ferreira et al., 2006).
Naast een constitutieve afweer bestaat er ook een geïnduceerde afweer die opnieuw
structureel en chemisch kan zijn. Deze afweer is vooral geldig tegen specifieke schimmels.
Een structurele, geïnduceerde afweer is de hypersensitieve respons (HR),
celwandverstevigingen door oxidatieve cross-linking van celwandcomponenten (gefaciliteerd
door waterstofperoxide en POD) , afzetting van callose en lignine, etc. (Glazebrook, 2005;
Ferreira et al., 2006).
HR houdt in dat de plant ervoor zorgt dat er een versnelde afsterving is van de plantcellen
rond de aanvalplaats. Hierdoor ontwikkelen zich necrotische laesies (geprogrammeerde
celdood) (Heath, 2000; Lazniewska et al., 2010). Deze respons ontneemt de pathogeen
nutriënten en produceert toxische moleculen die de groei van de pathogeen beperken tot een
kleine regio in de plant. De HR verleent resistentie aan een groot deel van potentiële
pathogenen en is tegen biotrofe schimmels actief (Heath, 2000). Dit is niet het geval bij
necrotrofen aangezien deze schimmels voordeel hebben bij de dode cellen van de plant
(Glazebrook, 2005).
Voorbeelden van chemische, induceerbare afweer zijn veranderingen in de permeabiliteit van
plasmamembranen of een snelle oxidatieve burst die leidt tot de productie van ROS (Reactive

17
Oxygen Species) zoals superoxide, waterstofperoxide en hydroxylradicalen die een
antimicrobiële werking hebben (Ferreira et al., 2006). De oxidatieve burst is één van de eerste
plantreacties op necrotrofe pathogenen. In het geval van F. graminearum zal DON echter zelf
de waterstofperoxideproductie in tarwe induceren en de geprogrammeerde celdood activeren.
Tarwe kan zichzelf dus tegen Fusarium spp. verdedigen door oxidatieve stress tegen te gaan,
omdat DONsynthese geïnitieerd wordt als de schimmel oxidatieve stress ondervindt (Walter
et al., 2009). Naast een oxidatieve burst is er een verhoogde productie van secundaire
metabolieten zoals fytoalexinen. De inductie van fosfolipasen, die inwerken op de lipide-
gebonden onverzadigde vetzuren in het membraan, zorgt namelijk voor de vrijstelling van
jasmonaat (JA, jasmijnzuur), methyljasmonaat (MeJA) en andere gerelateerde moleculen.
Hierdoor ontstaat er een transcriptionele activering van afweergerelateerde genen die coderen
voor pathogenese-gerelateerde (PR-) proteïnen zoals chitinasen, glucanasen en proteasen
(Ferreira et al., 2006).
Activering van de lokale respons (HR) op de plaats van infectie kan gevolgd worden door
secundaire immuniteit van de plant, namelijk de systemisch verworven resistentie (systemic
acquired resistance, SAR). SAR is een langdurige resistentie die werkt tegen schimmels die
weefselnecrose kunnen veroorzaken en kan weken tot maanden aanhouden. Het fenomeen
kan na de HR optreden omdat hierbij een signaal geproduceerd wordt dat via het floëem naar
andere plantendelen wordt vervoerd. In de systemische bladeren (bladeren die ook resistentie
verworven hebben maar niet aangevallen zijn, bevinden zich in een primed state) worden
SAR-genen geïnduceerd die coderen voor PR-proteïnen die microbiële celwandcomponenten
hydrolyseren (Conrath, 2006).
Lange tijd werd verondersteld dat in Arabidopsis thaliana en andere dicotyle planten de SA-
afhankelijke respons werd gebruikt tegen biotrofe schimmels (schimmels die nutriënten
opnemen uit levende plantencellen). Tegen necrotrofen (schimmels die plantenweefsel
afdoden) werd verondersteld dat de respons afhankelijk was van ethyleen (ET) en JA
(Glazebrook, 2005; Lazniewska et al., 2010). Deze responsen vertonen een aanzienlijke
overlap met de respons ten aanzien van wonden, die ook onder de controle van JA en ET
staan (Glazebrook, 2005). Omdat levensstijlen van schimmels niet steeds even gemakkelijk
kunnen worden onderverdeeld in necrotroof of biotroof, hangt de positieve of negatieve
interactie tussen SA (salicylzuur) en JA/ET af van de specifieke pathogeen. In natuurlijke
omstandigheden moeten planten zich verdedigen tegen meerdere soorten pathogenen. Daarom
induceren ze complexe verdedigingsmechanismen waarbij het nog onduidelijk is waarom ze
de ene respons boven de andere verkiezen (Bari & Jones, 2009). Hoe dan ook, de respons
tegen pathogenen is steeds afhankelijk van het type pathogeen maar ook van het type
waardplant (Bostock, 2005). Voor monocotyle planten is de plantrespons tegen pathogenen
namelijk meer afhankelijk van JA dan van SA, terwijl dit bij dicotylen net omgekeerd is. Er
moet dus voorzichtig worden omgesprongen met de extrapolatie van data afkomstig van A.
thaliana (dicotyl) naar graangewassen (monocotyl) (Walter et al., 2009).
2.3.3.2. Plantafweer t.a.v. insecten
Planten reageren op herbivorie door de productie van toxines en defensieve proteïnen om
fysiologische processen in insecten te belemmeren. De enorme diversiteit aan
gespecialiseerde plantcomponenten en secundaire metabolieten, vloeit voort uit een co-

18
evolutionaire strijd tussen herbivoren en planten om te eten of niet gegeten te worden. Als
insecten de plant aanvallen, zorgen de constitutieve en de geïnduceerde afweer voor de
bescherming van de plant. Beide afweren kunnen zowel direct als indirect zijn. Het
gecombineerde effect van directe en indirecte afweer levert een duurzame resistentie op tegen
een breed spectrum aan arthropoden (Howe & Jander, 2008).
Fysische barrières (directe constitutieve afweer), ook wel de eerste lijn van afweer genoemd,
hinderen vraat door herbivoren. Voorbeelden van dergelijke barrières zijn celwanden,
cuticula, trichomen, etc. De constitutieve afweer van de plant zorgt er tevens voor dat er
toxische metabolieten worden aangemaakt (indirecte afweer). Belangrijke secundaire
metabolieten zijn hydroxamzuur, alkaloïden, glucosinolaten, terpenoïden en C-6 aldehyden
(Goggin, 2007). Han et al. (2009) onderzocht bij tarwe verschillende constitutieve enzymen.
Activiteit van PAL, PPO en POD werd teruggevonden in de vlagblad- en aarstadia van tarwe
nadat deze door S. avenae werden aangetast. PAL is een enzym uit de fenylpropanoïd
pathway die instaat voor de productie van componenten (zoals coumarine) die toxisch zijn
voor zowel insecten als pathogenen, bovendien zal dit enzym finaal leiden tot productie van
onder andere SA, wat een sleutelregulator is in plantafweer. PPO en POD zijn oxidasen die
instaan voor de productie van quinonen van verscheidene fenolische precursoren. De graad
van activiteit van deze enzymen was afhankelijk van de gevoeligheid van het tarweras voor
bladluizen en het stadium waarin de tarwe zich bevindt. Hoewel deze enzymen constitutief
aanwezig zijn, vertonen ze een verhoogde activiteit als de plant door insecten wordt
aangevallen. Deze kunnen dus ook als een onderdeel van de geïnduceerde resistentie
beschouwd worden (Han et al., 2009). Plantcomponenten zoals fenolen en hydroxaminezuren
dragen bij tot resistentie bij verschillende groeistadia van granen. Ze schrikken bladluizen af
door hun antixenotische en antibiotische werking (Eleftherianos et al., 2006; Zhao et al.,
2009). Antixenose duidt op het verhinderen van bladluizen om zich te gaan settelen op granen
en deze koloniseren. Antibiose duidt op het reduceren van de populatiegroei van bladluizen
(Eleftherianos et al., 2006; Goggin, 2007). Vele van deze metabolieten zijn potentieel toxisch
voor de plant zelf. Daarom worden niet-toxische precursoren gestockeerd die geactiveerd
worden bij insectenaanvallen (Howe & Jander, 2008).
De geïnduceerde verdediging komt op gang bij de herkenning van insecten. Signaalmoleculen
alarmeren de plant zodat deze verschillende pathways kan activeren die tot resistentie kunnen
leiden. Relatief weinig is geweten over het doorgeven van signalen die een link vormen tussen
de insectspecifieke elicitoren en de plantafweerresponsen die ze uitlokken. Onder andere het
calciumion (Ca2+
) is betrokken als boodschapper in vele plantsignaalpathways, ook tegen
herbivorie. Cytosolische Ca2+
-inhoud is onder normale omstandigheden veel lager dan in
organellen of in apoplastische vloeistof. Kortstondige verhoging van de cytosolische Ca2+
-
levels activeren calmoduline en andere calcium-gewaarwordende proteïnen die als reactie
voor signaaltransducties zorgen waaronder transcriptionele responsen (Howe & Jander, 2008).
ROS, die ook activators zijn van afweersignaalpathways, ontstaan in planten die door
bladluizen aangevallen worden. De betrokkenheid van ROS bij pathogeenresistentie is goed
gekend maar deze componenten kunnen ook een direct nadelig effect hebben op het
darmweefsel van arthropoden (Smith & Boyko, 2006).

19
Wanneer de plant signalen krijgt dat deze aangevallen wordt door insecten worden twee
belangrijke signaalpathways geactiveerd als reactie op verwonding, namelijk de JA-
(octadecanoïd) pathway en de SA-pathway (van Dam, 2009; Zhao et al., 2009). Vooral de
jasmonaatpathway speelt een centrale rol in het bevorderen van resistentie tegen een breed
spectrum aan insecten (Howe & Jander, 2008). De signaalpathways kunnen geïnduceerd
worden door vraat en speeksel van herbivoren. Kauwende insecten induceren de JA-pathway
terwijl de insecten met stekende en zuigende monddelen de SA-pathway ook kunnen
activeren (Zhao et al., 2009). Insecten met een verschillend voedingsgedrag initiëren dus
verschillende maar toch overlappende patronen van genexpressie, terwijl aanvallers met
hetzelfde voedingsgedrag zeer gelijkaardige responsen uitlokken (Howe & Jander, 2008). Er
zijn ook nog andere signaalpathways zoals ET, abscisinezuur (ABA), giberellinezuur (GA),
ROS en stikstofmonoxide (NO). Als gevolg van de pathways worden metabolieten
geproduceerd die aan arthropoden zoals bladluizen weerstand moeten bieden (Smith &
Boyko, 2006).
Deze verschillende pathways leiden tot geïnduceerde plantresistentie (direct of indirect). De
directe geïnduceerde verdediging als reactie op herbivoren houdt in dat er via verschillende
pathways een snelle accumulatie is van verdedigingsproteïnen zoals proteïnase-inhibitors
(PIs) en PPO (Zhao et al., 2009) die het plantenweefsel voor het insect moeilijker verteerbaar
maken (Piesik et al., 2010). PIs tasten verteringsproteasen in darmen van de insecten aan. Dit
resulteert in aminozuurdeficiënties die de groei en ontwikkeling van de herbivoor negatief
beïnvloeden. Dit wordt vaak door het insect zelf gedwarsboomd doordat deze digestieve
proteasen kan aanmaken die ongevoelig zijn voor de PIs van de waardplant of deze PIs
inactiveren (Howe & Jander, 2008).
De indirecte geïnduceerde verdediging ontstaat op de plaats waar de schade is aangebracht en
verspreidt zich systemisch naar andere weefsels. In de niet beschadigde bladeren van de door
herbivoren aangevallen planten komt de afweer dus ook op gang. Deze systemische resistentie
zorgt voor een efficiënte resistentie tegen aanvallen die zich in de toekomst kunnen voordoen
(Howe & Jander, 2008). Tevens zorgt deze indirecte afweer ervoor dat er volatiele
componenten gemaakt worden die bij naburige planten ook een systemische resistentie
teweeg brengen (Zhao et al., 2009; Piesik et al., 2010). Zo is MeJA, waarvan JA de precursor
is, een volatiele signaalmolecule die communicatie tussen planten teweeg brengt. Ook kunnen
volatielen natuurlijke vijanden van de herbivoren aantrekken (Zhu-Salzman et al., 2005; Zhao
et al., 2009; Piesik et al., 2010). Chemische signalen die zowel voordelig zijn voor de
ontvangers (natuurlijke vijanden) als voor de verzenders (aangevallen planten) worden
synomonen genoemd (Steidle et al., 2005). Bladeren produceren continu kleine hoeveelheden
van deze volatielen maar extra volatielen worden uitgescheiden als de plant beschadigd wordt
door herbivoren. De chemische componenten in deze stoffen variëren tussen planten en
herbivoorsoorten (Dicke et al., 2003). De verschillende combinaties in volatielen kunnen
parasiterende en predaterende insecten op een selectieve manier aantrekken (Zhu-Salzman et
al., 2005; El-Wakeil et al., 2010). Ook extraflorale nectarklieren worden gestimuleerd om
carnivore arthropoden te lokken (Goggin, 2007).

20
2.4. Interactie tussen insecten en pathogenen
2.4.1. Insecten als vector van pathogenen
2.4.1.1. Virussen
Insecten die infectieuze pathogenen met zich meedragen, worden vectoren genoemd. Tussen
vectoren en virussen bestaat er een specifieke relatie. In vele gevallen kunnen virussen slechts
door één type insect worden verspreid. Zo kan een virus dat bladluizen als vector heeft niet
verspreid worden door tripsen en vice versa (Peters et al., 2009). Verschillende soorten
insecten zijn vectoren: bladluizen, dwergcicaden, spoorcicaden, witte vlieg, schildluizen,
tripsen, etc. (Höfte, 2009). Om een virusepidemiologie te begrijpen is het van belang om de
biologie en ecologie van zijn vector te bestuderen (Davis & Radcliffe, 2008a).
Virussen zoals het dwergvergelingsvirus op graan (barley/cereal yellow dwarf virus,
BYDV/CYDV) en het bladrolvirus op aardappelen (potato leafroll virus, PLRV) behoren tot
de familie Luteoviridae. Ze worden op persistente wijze verspreid door bladluizen (Powell et
al., 1997; Yang et al., 2008). Dit wil zeggen dat het virus in het bladluislichaam kan
circuleren gedurende een paar dagen en pas na passage doorheen de insectenweefsels via de
monddelen kan worden overgedragen. De bladluis kan levenslang infectieus blijven (Höfte,
2009; Peters et al., 2009). Bladluizen verwerven het virus door zich met hun stekend-
zuigende monddelen te voeden op geïnfecteerd floëem. Virions worden via het stilet
opgenomen in het bladluislichaam waar ze circuleren in het hemolymfe. Ze worden terug via
de speekselbuis in het bladluisstilet doorgegeven aan andere planten (Yang et al., 2008). Meer
dan 20 verschillende bladluissoorten, waaronder R. padi en M. dirhodum, kunnen het BYDV
doorgeven. Dit virus kan wintergerst en -tarwe aantasten. In de winter zullen de bladtoppen
van aangetaste wintertarwe geel verkleuren. Het virus veroorzaakt dwerggroei en een
verminderde wortelgroei. Bij een voorjaarsinfectie treedt er een roodverkleuring op van het
blad, te beginnen bij de bladtop. De beste manier om het virus te bestrijden, is door zijn
vectoren, in dit geval bladluizen, te bestrijden (Hermans et al., 2009). Bladluizen zijn tevens
een vector van het dwergmozaïekvirus op maïs (maize dwarf mozaic virus). Dit virus wordt
op een niet-persistente wijze verspreid (PaDIL, 2010). Dit wil zeggen dat het virus voorkomt
op het stilet van de zuigende bladluis, reeds nadat deze zich enkele seconden tot enkele
minuten op de geïnfecteerde plant heeft gevoed. Het virus blijft slechts enkele uren op de
bladluis aanwezig. De verspreiding van niet-persistente virussen gaat dus veel sneller dan de
verspreiding van persistente virussen (Höfte, 2009; Peters et al., 2009).
De groene perzikluis (Myzus persicae) is een polyfage bladluissoort die minstens 860
plantensoorten als waardplant heeft en een vector is van minstens 100 plantenvirussen. Het is
een efficiënte vector van het aardappelvirus (potato virus Y, PVY) en PLRV (Davis &
Radcliffe, 2008a). Het PLRV wordt op persistente wijze doorgegeven en zorgt voor een
verlies aan aardappelopbrengsten, chlorose en het donker worden van vasculaire bundels. Dit
leidt tot een lagere kwaliteit en vermarktbaarheid. Het virus komt voor in het floëem van de
plant en wordt niet alleen verspreid door de groene perzikluis maar ook door verscheidene
andere bladluissoorten zoals de aardappelbladluis (Macrosiphum euphorbiae) en de
katoenbladluis (Aphis gossypii) (Davis & Radcliffe, 2008b). Ook de sojaboonbladluizen
(Aphis glycines) zijn in staat om PLRV door te geven aan gevoelige aardappelrassen. Deze

21
exotische bladluissoort kan ook nog andere persistente en niet-persistente virussen doorgeven.
Hun hoge reproductie, korte generatieperiode en vermogen om alatae te vormen, verhogen de
kans dat het PLRV wordt verspreid. Daarbij komt dat aardappelen gevoelig zijn voor het virus
in hun vegetatieve fase. De virustransmissie door deze bladluis wordt verhoogd omdat deze
vooral in het begin van het groeiseizoen in het gewas arriveert (Davis & Radcliffe, 2008a).
2.4.1.2. Bacteriën
Bladluizen worden beschouwd als strikte vectoren van fytopathogene virussen, ondanks hun
frequent fysiek contact met bacteriële epifyten. Als bladluizen landen op een waardplant,
maken ze met hun stilet proefboringen in het weefsel om de geschiktheid van de plant na te
gaan. Ze prikken in de epidermale lagen om de zeefvaten van het floëem te lokaliseren.
Sommige bladluissoorten spuiten speeksel in het weefsel die het plantmateriaal voorverteert
waarna ze het terug opzuigen. Het speeksel kan in contact komen met verschillende bacteriële
epifyten zoals de alomtegenwoordige fytopathogene Pantoea agglomerans en Pseudomonas
syringae. De erwtenbladluis (Acyrthosiphon pisum) kan P. syringae opnemen en deze bacterie
kan zich in het bladluislichaam vermenigvuldigen. Daarna wordt ze uitgescheiden samen met
de honingdauw en komt op het plantoppervlak van al dan niet waardplanten terecht
(Stavrinides et al., 2009).
De Aziatische citrusbladvlo (Diaphorina citri) is meestal een minder belangrijke plaag op
citrusplanten. Echter als deze een vector is van de bacteriën die vergroening veroorzaken op
citrus, wordt het insect als een belangrijke plaag beschouwd. De pathogeen veroorzaakt gele
scheuten, vlekziekte, chlorose, etc. Het fruit kleurt niet naar behoren, vandaar de naam
vergroeningsziekte. De twee floëemgelimiteerde bacteriesoorten die deze ziekte veroorzaken
zijn Liberibacter africanus en asiaticus (Halbert & Manjunath, 2004).
Insecten zoals cicaden voeden zich op xyleemsap en zijn vectoren van Xylella fastidiosa. Dit
is een bacterie die onder andere een invloed heeft op de structurele polysacchariden van
planten. Deze bacteriën koloniseren het nauwe kanaal in de chitineuze monddelen van de
vectoren (Killiny et al., 2010). Andere xyleemzuigende insecten zoals scherpschutters
(Cicadellinae) en schuimbeestjes (Cercopidae) zijn eveneens vectoren van Xylella fastidiosa.
Deze bacterie veroorzaakt de Pierceziekte (Californische wijnziekte) op wijndruiven. Het
verhindert de sapstroom waardoor bladeren verdrogen en afsterven (Hopkins & Purcell,
2002).
2.4.1.3. Schimmels
Plantenziekten veroorzaakt door schimmels, met insecten als vectoren, kunnen verwoestende
effecten hebben op planten. Zo kan Verticillium albo-atrum, een verwelkingsziekte op
luzerne, verspreid worden door de erwtenbladluis (Acyrthosiphon pisum), de bladsnijderbij
(Megachile rotanata) en de champignonmug (Bradysia impatiens). Phytium wortelrot wordt
verspreid door oevervliegen (Scatella stagnalis) en Fusarium oxysporum door
champignonmuggen (Bradysia spp.) (El-Hamalawi, 2007).
Thielaviopsis basicola is een schimmel die zich in de bodem verspreidt en onder andere de
wortels van o.a. veldsla (Valerianella locusta) infecteert. Veldsla kent ook een andere vijand,
namelijk oevervliegen (Scatella stagnalis). Onderzoek toont aan dat oevervliegen deze
schimmel kunnen verspreiden. De chlamydosporen van T. basicola overleven de passage
doorheen het spijsverteringsstelsel van de vliegen en levende propagules komen in de

22
uitwerpselen van de insecten terecht. De leefbaarheid van de propagules wordt niet beïnvloed
door de acquisitietijd. Oevervliegen, zowel de adulten als de larven, dragen dus bij tot de
verspreiding van deze schimmel op veldsla (Stanghellini et al., 1999). Niet alleen T. basicola
maar ook Verticillium dahliae en F. oxysporum f.sp. basilici worden door oevervliegen
verspreid. Dit zijn eveneens bodemschimmels. De verspreiding van sporen die aan de
buitenkant van de lichamen van de vliegen hangt, zoals aan de poten en vleugels, is ook van
belang (El-Hamalawi, 2007).
Fusarium avenaceum, die ook in het Fusarium ziektecomplex terug te vinden is (Kema et al.,
2002), veroorzaakt stengelrot bij lisianthus (Eustoma grandiflorum). Oevervliegen,
champignonmuggen en motmuggen (Psychodidae spp.) kunnen de bovengrondse
levensvormen (macroconidia) van deze schimmel extern opnemen. Zowel solitaire als
geclusterde macroconidia worden verspreid. Oevervliegen zijn tevens in staat om
macroconidia intern op te nemen en leefbare propagules uit te scheiden. De hoeveelheid
inoculum verplaatst door deze insecten blijkt genoeg te zijn om de lisianthusplanten te
infecteren en ziek te maken (El-Hamalawi & Stanghellini, 2005).
Oevervliegen kunnen ook Pythium aphanidermatum verspreiden wanneer ze zich voeden op
de geïnfecteerde komkommerwortels (Cucumis sativus). Alle larvale stadia en ook de adulten
bevatten oösporen van deze schimmel in hun spijsverteringsstelsel. Daar larven in direct
contact komen met de komkommerwortels en dus ook met P. aphanidermatum konden ze de
sporen opnemen. Adulten komen echter niet in direct contact met de geïnfecteerde wortels.
Sporen in hun spijsverteringsstelsel zijn dus opgenomen in de larvale stadia maar zijn na
verpopping nog inwendig achtergebleven. Na uitscheiding hebben deze sporen nog steeds
voldoende kiemkracht om gezonde komkommerplanten te infecteren (Stanghellini &
Goldberg, 1990).
Phytophthora citricola veroorzaakt kanker bij de stam van avocadobomen (Persea
americana). Meestal treedt deze pathogeen de boom binnen via wonden die dicht bij de
bodem gelegen zijn, met als gevolg dat de boom suikerhoudende exudaten lekt. Tot deze
stoffen voelen mieren (Iridomyrmex humilis) zich aangetrokken. Door ermee in contact te
komen, kunnen ze zowel oösporen als myceliumfragmenten van deze pathogeen verspreiden
naar de bodem of naar ander wonden (El-Hamalawi & Menge, 1996).
Onderzoek toont aan dat er een interactie is tussen de stengelboorder Apion onopordi en de
roestschimmel Puccinia punctiformis op akkerdistel (Cirsium arvense). De wijfjes van de
kever zorgen tijdens de ovipositie ervoor dat akkerdistel wordt geïnfecteerd met roest. De
adulte insecten die ontstaan zijn uit eitjes gelegd op geïnfecteerde akkerdistel zijn significant
zwaarder dan adulten op gezonde planten. De reden hiervoor is onbekend maar de zwaardere
kevers produceren meer en grotere eitjes, waardoor zwaarder zijn dus een fitnessvoordeel
betekent (Friedli & Bacher, 2001). Sporen van P. punctiformis kunnen ook door andere
herbivore insecten verspreid worden. Zo kunnen de zwarte bonenluis (Aphis fabae), de grote
distelluis (Uroleucon cirsii) en de schildpadtor (Cassida rubiginosa), als ze zich gevoed
hebben met geïnfecteerde akkerdistelplanten, gezonde planten besmetten met sporen die aan
hun lichaam en poten hangen (Kluth et al., 2002).

23
3. Materiaal en methoden
3.1. Studie-organismen
3.1.1. Tarwezaden en hun oorsprong
Er werd gewerkt met vijf tarwerassen (Triticum aestivum): Lexus (LEX), Homeros (HOM),
Tulsa (TUL), Sahara (SAH) en Cadenza (CAD). Hun mandatarissen waren Clovis Matton
(LEX en HOM), Ets. L. Rigaux (TUL), Aveve (SAH) en KWS Benelux (CAD).
3.1.2. Kweek van bladluissoorten S. avenae en M. dirhodum
Vleugelloze S. avenae en M. dirhodum bevonden zich elk apart in plexiglazen bakken die aan
de zijkanten voorzien zijn van gaas. In de plexiglazen bakken werden tarweplanten (SAH)
van ongeveer een week oud geplaatst waarop de bladluizen zich konden voeden. De
bladluizen werden gestimuleerd om zich parthenogenetisch voort te planten in een
klimaatskamer met een temperatuur van 21 ± 1°C, een relatieve vochtigheid van 60% en een
lichtregime van 16u licht/8u duisternis.
De mengcultuur van S. avenae werd aangekocht als een Banker-System-E bij Biobest
Belgium N.V. uit Westerlo. De bladluizen, zowel nimfen als adulten, bevonden zich in een
plastieken doos. Transport hiervan gebeurde in een piepschuimen doos voorzien van een
koelelement. Bij aankomst werd de plastieken doos in de plexiglazen bak geplaatst. De
mengcultuur van M. dirhodum was afkomstig van veldbemonsteringen in Bottelare
(Proefhoeve, Hogent).
3.1.3. Schimmel F. graminearum (GFP transformant)
In de proeven werd gebruik gemaakt van een GFP transformant van F. graminearum stam
8/1. De klasse I transformant die in staat is om DON te produceren werd gebruikt (Jansen et
al., 2005). Deze bevond zich in de collectie van schimmels in de Hogeschool Gent (campus
Schoonmeersen). Eveneens werd DON zelf ingeschakeld in de proeven. Het gebruikte DON
was een supernatans van F. graminearum die opgekweekt was op een DON inducerend
medium (Gardinez et al., 2009).
3.1.3.1. Inoculatie van F. graminearum op PDA medium
Een GFP transformant van F. graminearum stam 8/1 (klasse I) werd geïnoculeerd door een
klein stukje mycelium op een petriplaat met potato dextrose agar (PDA) medium (OXIOD,
Erembodegem, België) te inoculeren. Deze procedure gebeurde onder een verticale laminaire
flow om zo steriel mogelijk te kunnen werken en zodoende contaminatie te vermijden. Hierna
werd deze petrischaal onder een lichtregime van UV/duisternis (12u 365 nm 10W/12u)
geplaatst. Na 7 tot 10 dagen bij 20°C trad sporulatie op (Fig. 9). De schimmel werd bij 4 °C
bijgehouden tot wanneer de proeven startten.

24
a b
Fig. 9: Petriplaat (diameter = 9 cm) met sporuleerde F. graminearum (GFP) (a: bovenkant, b: onderkant).
Goede sporulatie kan te zien zijn aan de rooskleuring van het medium, groei van de schimmel in concentrische
cirkels en weinig luchtmycelium.
3.1.3.2. Sporenoplossing van F. graminearum maken
Er werd gewerkt met steriel water waarin 0,01% detergent werd aangebracht. Als detergent
werd polyoxyethylensorbitanmonoleat (Tween20) gebruikt dat ervoor zorgt dat de sporen niet
samenklonteren. Deze oplossing werd op de petriplaat, waarin de sporulerende schimmel
groeit (zie 3.1.3.1.), gegoten. Door met een steriele spatel, verhit in een bunsenbrander, over
het conidia-houdende mycelium te wrijven, werden de sporen geoogst. Deze sporenoplossing
werd in een falconbuisje van 50 ml opgevangen. 5 µl van deze sporenoplossing werd met een
pipet in het groefje van de Bürkertelkamer gebracht. Via capillaire krachten werd de
oplossing doorheen het raster gezogen. Hierna werd er een dekglaasje overgelegd. De
telkamer werd bekeken onder een microscoop (Olympus XC 50) met fasecontrast. Er werd een
verdunning van 1018
sporen per ml oplossing gemaakt.
3.1.3.3. Controle van groei van GFP transformant (F. graminearum) m.b.v.
fluorescentiemicroscoop
Uit de 4 kwadranten van de petriplaat werd een klein stukje mycelium genomen. Dit werd op
een draagglaasje gelegd, waarna er een druppel steriel water werd overgegoten met een pipet.
Daarna werd er een dekglaasje overgelegd. Dit preparaat werd onder de
fluorescentiemicroscoop (Olympus XC 50, X-Cite Series 120 PC Q, Cell*Family software)
bekeken. Er werd gezocht naar fluorescerende myceliumdraden en sporen die duidelijk te
onderscheiden waren van autofluorescentie. Deze waren afkomstig van F. graminearum
(GFP).
3.2. Preferentietesten
3.2.1. Aanmaken van steriele tarwekiemlingen
In falconbuisjes van 15 ml werd 4 g steriel wit zand en 0,5 ml steriel water gedaan. Hierin
werd een steriel zaadje van een bepaald tarweras geplant. Het zaadje werd steriel gemaakt
door het 1 minuut in water met 1% NaOCl onder te dompelen, waarna het in steriel water
werd afgespoeld. De buisjes werden in een groeikamer geplaatst met een lichtregime van 16u
licht/8u duisternis en een temperatuur van 22°C.

25
3.2.2. Uitzaaien van tarwe en productie van aren
Tarwe werd uitgezaaid in potten met een diameter van 14 cm voorzien van potgrond.
Gedurende de groei werden ze in een groeikamer gehouden van 22°C, 60% relatieve
vochtigheid en een lichtregime van 16u licht/8u duisternis. Ze kregen voldoende water en
werden twee keer bemest met 1,5 g/l Alkristal 1 (oplosmeststof, N-P-K 17-6-10, chloorarm).
Na circa vijf maanden produceerden de tarweplanten aren.
3.2.3. Ontwikkeling van preferentietesten
Om te achterhalen of bladluizen bepaalde tarwerassen prefereren, werden preferentietesten
ontwikkeld. Dit werd gedaan met behulp van een t-stukje die twee falconbuizen met elkaar
verbond. Via het t-stukje werden 10 adulte bladluizen in het systeem gebracht met behulp van
een penseel en de opening werd met parafilm afgesloten.
3.2.3.1. Met kiemlingen
Er werden preferentietesten met steriele tarwekiemlingen uitgevoerd. 7 dagen oude, steriele
tarwezaailingen (zie 3.2.1. voor de aanmaak van steriele kiemlingen) werden twee aan twee
gecombineerd door 2 falconbuisjes (15 ml) tegenover elkaar te plaatsen met behulp van een t-
stukje (Fig. 10). De preferentie van bladluizen voor tarwerassen LEX, TUL, SAH en HOM
werd onderzocht. De tarwecultivars werden zowel onderling als met zichzelf gecombineerd.
Elk uur werd nagegaan hoeveel bladluizen voor het ene of het andere tarweras kozen. Dit
werd 7u lang gedaan. Na 24u, en indien nodig na 48u, werd het opnieuw geobserveerd. De
procedure werd zowel met M. dirhodum als S. avenae doorlopen. Elke combinatie werd 5
keer herhaald.
Fig. 10: Systeem voor preferentietesten met tarwekiemlingen
3.2.3.2. Met aren
Bij preferentietesten met aren werden tarwearen gebruikt zoals bekomen in 3.2.2. De
rassencombinaties LEX-LEX en HOM-HOM werden gemaakt. Niet bloeiende aren werden in
falconbuisjes (50 ml) gebracht waarvan de punt was afgesneden. Deze buisjes werden twee
aan twee verbonden met een t-stukje (Fig. 11). De uiteinden van de falconbuizen werden
afgesloten met behulp van parafilm. 10 adulte bladluizen van S. avenae werden langs de derde
opening van het t-stukje toegevoegd met behulp van een penseel. Daarna werd de opening
afgesloten met parafilm. Elk uur werd geobserveerd hoeveel bladluizen er uit het t-stukje
verdwenen waren. Dit gaf een idee over het aantal bladluizen die reeds een keuze gemaakt
hadden. Later werden de bladluizen op de aren geteld. Dit gebeurde op destructieve wijze.
Elke combinatie werd 5 keer herhaald.
Fig. 11: Systeem voor preferentietesten met tarwearen

26
3.2.3.3. Met behandelde aren
1. Op korte termijn
Aren (CAD) werden geïnfecteerd met F. graminearum (GFP) (FUS), met het mycotoxine
DON of met gedestilleerd water (+ 0,01% detergent, Tween20) (CON). Er werd gebruik
gemaakt van boeiende planten (groeistadium 60 volgens de Zaddokschaal) omdat aren besmet
worden door F. graminearum met behulp van sporen die via de meeldraden de plant
binnendringen.
Elke dag, gedurende 4 dagen, werden er 12 onafhankelijke (van een verschillende plant) aren
behandelend. Vier aren ondergingen met behulp van een pipet een puntinoculatie met sporen
van F. graminearum (GFP). Per aar werden er twee aarpakjes geïnoculeerd met elk 20 µl
sporenoplossing bestaande uit 106
sporen per ml water waarin 0,01% detergent was opgelost
(zie 3.1.3.2. voor het aanmaken van een sporenoplossing). Deze korrels werden gemerkt met
een stift (Audenaert et al., 2010). Bij vier andere onafhankelijke aren werden er twee
aarpakjes bedruppeld met elk 20 µl water waarin 0,01% detergent was toegevoegd. Deze
deden dienst als controle. Bij de overblijvende 4 planten werd er telkens per aar twee
aarpakjes met 20 µl DON (10 mg/l water) behandelend. Elke plant werd gelabeld.
Na de infectie bevonden de planten zich 24u in 100% relatieve vochtigheid zodat de sporen
konden kiemen en de plant binnendringen. Daarna kregen de planten 24u om te drogen bij een
lichtregime van 16u licht/8u duisternis. 48u na de infectie werden de aren in falconbuisjes (50
ml) gebracht waarvan de punt was afgesneden. Deze buisjes werden twee aan twee verbonden
met een t-stukje (Fig. 11). De uiteinden van de falconbuizen werden afgesloten met behulp
van parafilm. 10 adulte bladluizen van S. avenae werden langs de derde opening van het t-
stukje toegevoegd met behulp van een penseel. Daarna werd de opening afgesloten met
parafilm. De bladluizen kregen 48u om één van de twee aren te kiezen en er zich op te
voeden. Daarna werden de aren afgeknipt van de plant en samen met de bladluizen die erop
zaten één dag in de diepvries gelegd. Later werden de bladluizen op de aren geteld. Dit
gebeurde op destructieve wijze. In deze proef werden zowel dezelfde als verschillende
behandelingen twee aan twee gecombineerd.
Als controle werd bij elke plant een extra aar geïnoculeerd met dezelfde behandeling. 14
dagen na inoculatie van deze aren werd de infectie beoordeeld op basis van het oppervlak van
de aren waarop Fusariumsymptomen te zien waren: 1 = gezond, 2 = tot 25%, 3 = 25 tot 50%,
4 = 50 tot 75%, 5 = 75 tot 100% van de aar bedekt met symptomen (zie Fig. 5).
2. Op lange termijn
Er werd tevens eenzelfde proef opgesteld maar hierbij werd pas na 14 dagen geobserveerd
hoeveel bladluizen zich op de aren met de verschillende behandelingen bevonden.
3.2.3.4. Met artificieel dieet aangelengd met DON
Hebben bladluizen (S. avenae) een voorkeur voor DON zelf, losgekoppeld van de
afweerreactie dat DON veroorzaakt bij planten?
Voor deze proef werden falconbuisjes (15 ml) doorgesneden en op een t-stukje geplaatst (Fig.
12). Langs beide kanten kwam een parafilmsachet (2 uitgerekte lagen parafilm over elkaar
met daartussen een vloeistof) voor met daarin 70 µl van een behandeling. Er waren 3
behandelingen: 0,25 mg/l DON (Da) (gemaakt uit 500 µl artificieel dieet zoals beschreven in

27
Febvay et al., (1988), 25 µl DON (10 mg/l) en 475 µl steriel water), 1 mg/l DON (Db)
(gemaakt uit 500 µl artificieel dieet, 100 µl DON (10 mg/l) en 400 µl steriel water) en de
controle (C) (gemaakt uit 500 µl artificieel dieet en 500 µl steriel water). Deze oplossingen
werden in de vortex geschud alvorens de sachets gemaakt werden. De 3 behandelingen
werden onderling gecombineerd. Per systeem werden er 10 adulte bladluizen toegevoegd met
een penseel via het t-stukje en de opening werd afgesloten met parafilm. Na 1u, 2u, 3u, 4u,
24u en 48u werd nagegaan welke keuze de bladluizen gemaakt hadden. Elke combinatie werd
6 keer herhaald.
Om na te gaan of bladluizen zich aangetrokken voelden tot mogelijke volatiele componenten
in DON werd in 20 systemen, zoals hierboven beschreven, één bladluis toegevoegd. Aan de
ene kant bevond zich een C-behandeling, aan de andere een Db-behandeling. Na 1u, 2u, 3u,
4u, 5u, 24u, 48u en 72u werd nagegaan welke keuze de bladluizen gemaakt hadden.
Fig. 12: T-stukje met aan beide uiteinden een stukje falconbuis (15 ml)
3.3. Verspreiding F. graminearum door S. avenae
3.3.1. Orale opname van macroconidia door S. avenae
In deze proef werd er nagegaan of bladluizen de sporen van F. graminearum (GFP) kunnen
opnemen en uitscheiden.
Falconbuisjes (50 ml) werden doorgesneden. Langs één kant werd er een parafilmsachet
gemaakt. Daarin werd met een pipet 125 µl artificieel dieet (Febvay et al., 1988) voor
bladluizen gedaan dat aangelengd was met 125 µl steriel water met 1018
sporen per ml (zie
3.1.3.2. om een sporenoplossing te maken). Deze oplossing werd in de vortex geschud
alvorens de sachets gemaakt werden. Langs de andere kant werden er 25 adulte bladluizen in
het systeem toegevoegd en afgesloten met parafilm. Elke behandeling (artificieel dieet met
sporen van F. graminearum en zonder sporen ter controle) werd één keer uitgevoerd. De
bladluizen kregen 2u tijd om zich op de parafilmsachet te voeden. Hierbij werd verondersteld
dat de sporen na 2u nog niet waren overgegaan tot kieming (Audenaert et al., 2010). Om
bezinking van de sporen te verhinderen, werden de systemen elke 15 minuten omgedraaid. Na
2u werden alle zuigende bladluizen in petrischalen op medium met antibiotica (zie 3.3.1.1.)
geplaatst. De bladluizen konden vrij rondlopen in de petrischaal. De platen werden na
minimum 4 dagen incubatie bij 25°C gecontroleerd op fluorescentie onder de
fluorescentiemicroscoop (zie 3.1.3.3.). Met deze proefopzet kon nagegaan worden of
bladluizen eventueel opgenomen sporen konden uitscheiden.
Om na te gaan of bladluizen sporen inwendig kunnen opnemen, moesten deze gegrind
(gemaald) worden. Hierbij werden 50 adulte bladluizen in een bovenstaand beschreven
systeem toegevoegd. Na 2u werden de zuigende bladluizen gegrind in een mortier. Deze

28
massa werd in een petriplaat met medium en antibiotica uitgeplaat. Na minimum 4 dagen
werd deze plaat onder de fluorescentiemicroscoop gecheckt. Dit werd 1 keer uitgevoerd.
Om na te gaan of het grinden de sporen echter niet beschadigde en kieming hierdoor
verhinderde, werden 30 bladluizen samen met een sporenoplossing van 100 sporen per ml in
een mortier gegrind, uitgeplaat en na minimum 4 dagen gecheckt onder de
fluorescentiemicroscoop. Dit werd ook 1 keer uitgevoerd.
3.3.1.1. Medium met antibiotica aanmaken
In een fles van 1 liter werd 31,8 g dichloram glycerol agar (DG18) gedaan. Dit werd
aangevuld met 2,5 mg malachiet groen en 300 mg chloramfenicol om bacteriegroei tegen te
gaan. Het poedermengsel werd aangelegd met gedestilleerd water tot 1 liter. Na goed
schudden, werd het in de autoclaaf steriel (15 minuten aan een temperatuur van 120°C (1
atm)) gemaakt, later afgekoeld en uitgegoten in petrischalen met een diameter van 9 cm onder
een verticale laminaire flow. Het deksel werd pas op de petrischaal teruggeplaatst wanneer het
medium gedroogd was. Dit voorkwam condensvorming.
3.3.2. Uitwendige acquisitie en verspreiding F. graminearum door S. avenae
Er werd nagegaan of de bladluizensoort S. avenae in staat is om F. graminaerum (GFP)
uitwendig op het lichaam te dragen en te verplaatsen. Deze proef werd op verschillende
wijzen uitgevoerd.
Allereerst werd nagegaan hoe groot de proportie aan adulte bladluizen was die in staat waren
om F. graminearum (GFP) met zich mee te dragen als ze uit de sporulerende schimmel
kwamen. In 5 petriplaten met sporulerende F. graminearum (zie 3.1.3.1.) werden 10
bladluizen gevoegd. Deze kregen 4u de tijd om hierin rond te lopen en daarna werd het deksel
van de petriplaat verwijderd. Wanneer de bladluizen uit de petriplaat kropen, werden ze met
een penseel opgenomen en in een petriplaat met medium en antibiotica geplaatst (zie 3.3.1.1.).
Na minimum 4 dagen incubatie bij 25°C werden de platen onder de fluorescentiemicroscoop
gecheckt op F. graminearum (GFP) (zie 3.1.3.3.). Deze proef werd 2 keer herhaald.
Een proefopzet (Fig. 13) met 15 petrischalen (diameter = 14 cm) werd aangewend om na te
gaan of S. avenae als vector van F. graminearum (GFP) kan optreden. Daarin werd wateragar
(zie 3.3.2.1. voor de aanmaak van wateragar) gegoten. In elke petrischaal werd een steriel
zaadje van de tarwecultivar Homeros gelegd (zaadje steriel gemaakt zoals in 3.2.1.). Na een
week incubatie van de platen bij een lichtregime van 16u licht/8u donker en 22°C was de
kiemling in het eerste bladstadium. In elke petriplaat werden met behulp van een penseel 5
adulte bladluizen geplaatst, tegenover de kiemling, die 4u in sporulerende F. graminearum
rondgelopen hadden. Deze platen werden omgekeerd gelegd, daar de bladluizen de neiging
hadden om naar het licht te lopen en het contact tussen bladluizen en het wateragar verzekerd
was. Na 15 minuten, 1,5u en 24u werden van 5 platen de bladluizen weggenomen en elk apart
in petrischalen op medium met antibiotica geplaatst, waarin ze vrij konden rondlopen. De
kiemling werd in stukjes geknipt en eveneens op medium met antibiotica gelegd (Fig. 14).
Deze proef werd in de tijd herhaald. Ter controle werden er 3 platen met wateragar en een
steriele kiemling gemaakt waarbij geen bladluizen werden toegevoegd.

29
Fig. 13: Petriplaat (diameter = 14 cm) met wateragar en daarin 5 bladluizen en een steriele kiemling
(P: plaats waar kiemling zich bevindt, BL: startpunt van de bladluizen)
Fig. 14: Petriplaat (diameter = 9 cm) met in stukken gesneden kiemling
3.3.2.1. Wateragar aanmaken
15 g bacteriologisch agar werd aangelengd met gedestilleerd water tot 1 liter. Na goed
schudden, werd het in de autoclaaf steriel gemaakt, later afgekoeld en uitgegoten onder een
verticale laminaire flow in petrischalen met een diameter van 14 cm.
3.4. Overleving A.pisum en S. avenae op artificieel dieet met DON
3.4.1. Acyrthosiphon pisum, de erwtenbladluis
De erwtenbladluis, Acyrthosiphon pisum (Hemiptera: Aphididae), is een groene bladluis die
leeft op peulvruchten. Meestal behoren zijn waardplanten tot de familie van de Fabaceae.
Deze bladluissoort komt wereldwijd voor. In koud-gematigde gebieden is A. pisum
holocyclisch en produceert oviparae en mannetjes. Ze hebben geen echte waardplantwissel. In
Europa produceren ze in de herfst alate sexuparae en mannetjes. Hun geslachtelijke fase
maken ze door op Vicia (Blackman & Eastop, 2007). Deze bladluissoort doorloopt vier
nimfale stadia alvorens adult te worden. Zijn genoom is gekend (The International Aphid
Genomics Consortium, 2010).
Een stockcultuur van A. pisum bevindt zich in het laboratorium onder gestandaardiseerde
condities: 23-25°C, een relatieve vochtigheid van 60-70% en een lichtregime van 16u licht/8u
duisternis. Deze bladluizen worden gekweekt op tuinbonen (Vicia faba) (Hamshou et al.,
2010).

30
3.4.2. Proefopzet met bladluissystemen
Om na te gaan wat de invloed van DON is op de overleving van S. avenae en A. pisum, werd
volgend experiment opgezet. Artificieel dieet voor bladluizen (zie Febvay et al. (1988) voor
de samenstelling) werd aangelengd met verschillende concentraties van DON, namelijk 0,1
mg/l, 0,5 mg/l en 1 mg/l DON. Er werden 3 bladluissystemen (Fig. 15) per concentratie DON
gemaakt met elk 10 nimfen. Een controle (0 mg/l DON) werd eveneens opgezet. 3 dagen lang
werden de overlevende nimfen geteld. Gedurende dit experimenten kregen de bladluizen om
de 2 dagen nieuwe voeding. Per systeem bestond deze voeding uit 200 µl oplossing van
artificieel dieet voor bladluizen aangelengd met verschillende concentraties van DON of
steriel water voor de controle.
Fig. 15: Aanmaak van bladluissystemen. De 2e foto is een parafilmsachet (zie 3.2.3.4.). De bladluizen zitten
tussen deze sachet en het deksel (4e foto). Daarna worden deze systemen omgekeerd geplaatst zodat de
bladluizen ondersteboven aan het dieet kunnen zuigen.
3.5. Reproductie S. avenae op artificieel dieet met DON
De ontwikkeling en reproductie van S. avenae werd voor verschillende concentraties van
DON (0,25 mg/l en 1 mg/l) nagegaan. At random geselecteerde adulte bladluizen van S.
avenae uit de stockcultuur werden op tarwebladeren (SAH) gezet. Neonaten geproduceerd na
24u werden gebruikt om de proef te starten. De leeftijd van deze neonaten was 0 tot 24u. Die
dag werd als dag 0 beschouwd. De nimfale ontwikkeling en mortaliteit werden dagelijks
gescoord. De aanwezigheid van exuvia werden gebruikt voor het determineren van een
vervelling. Voor de controle (0 mg/l DON) en de behandelingen werden 5 systemen (Fig. 15)
opgezet. In elk systeem zaten er 2 neonaten. Wanneer ze het adulte stadium bereikten, werden
ze elk apart in een systeem gezet. De nimfen die ze produceerden, werden dagelijks geteld en
met een fijn penseel voorzichtig weggenomen zodat de adult zo weinig mogelijk gestoord
werd. Gedurende dit experiment kregen de bladluizen om de 2 dagen nieuwe voeding. Per
systeem bestond deze voeding uit 200 µl oplossing van artificieel dieet voor bladluizen
aangelengd met verschillende concentraties van DON of steriel water voor de controle.
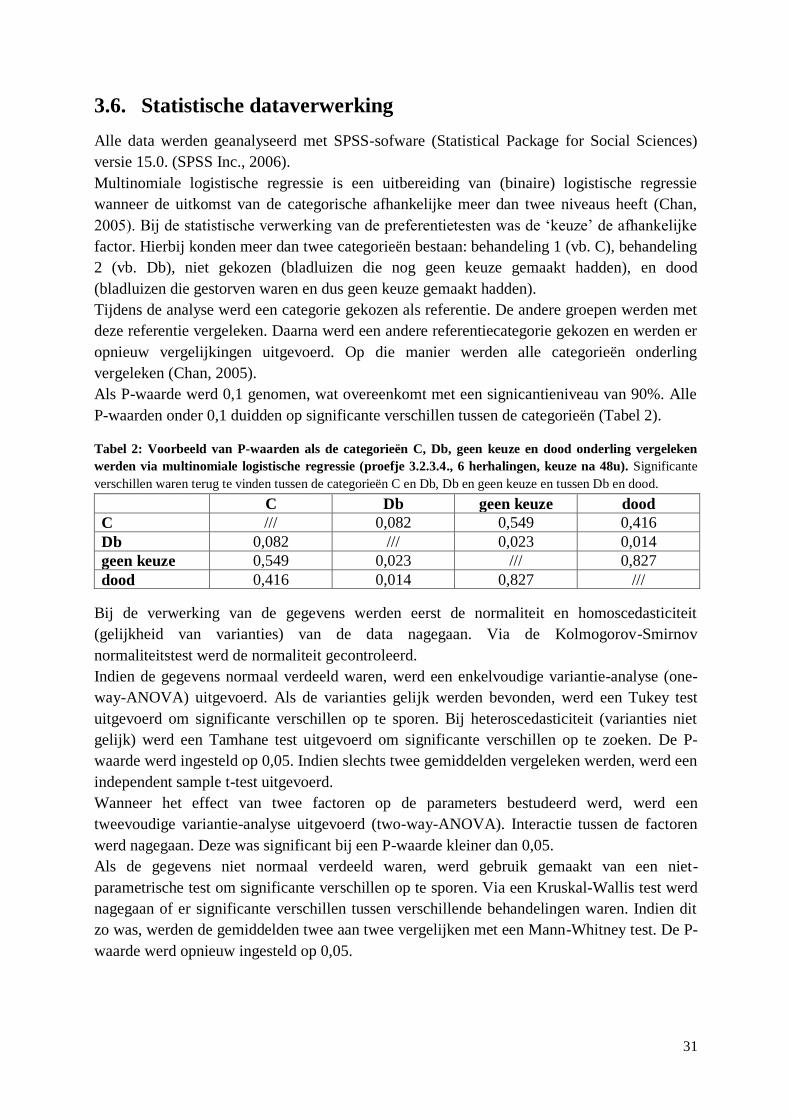
31
3.6. Statistische dataverwerking
Alle data werden geanalyseerd met SPSS-sofware (Statistical Package for Social Sciences)
versie 15.0. (SPSS Inc., 2006).
Multinomiale logistische regressie is een uitbereiding van (binaire) logistische regressie
wanneer de uitkomst van de categorische afhankelijke meer dan twee niveaus heeft (Chan,
2005). Bij de statistische verwerking van de preferentietesten was de „keuze‟ de afhankelijke
factor. Hierbij konden meer dan twee categorieën bestaan: behandeling 1 (vb. C), behandeling
2 (vb. Db), niet gekozen (bladluizen die nog geen keuze gemaakt hadden), en dood
(bladluizen die gestorven waren en dus geen keuze gemaakt hadden).
Tijdens de analyse werd een categorie gekozen als referentie. De andere groepen werden met
deze referentie vergeleken. Daarna werd een andere referentiecategorie gekozen en werden er
opnieuw vergelijkingen uitgevoerd. Op die manier werden alle categorieën onderling
vergeleken (Chan, 2005).
Als P-waarde werd 0,1 genomen, wat overeenkomt met een signicantieniveau van 90%. Alle
P-waarden onder 0,1 duidden op significante verschillen tussen de categorieën (Tabel 2).
Tabel 2: Voorbeeld van P-waarden als de categorieën C, Db, geen keuze en dood onderling vergeleken
werden via multinomiale logistische regressie (proefje 3.2.3.4., 6 herhalingen, keuze na 48u). Significante
verschillen waren terug te vinden tussen de categorieën C en Db, Db en geen keuze en tussen Db en dood.
C Db geen keuze dood
C /// 0,082 0,549 0,416
Db 0,082 /// 0,023 0,014
geen keuze 0,549 0,023 /// 0,827
dood 0,416 0,014 0,827 ///
Bij de verwerking van de gegevens werden eerst de normaliteit en homoscedasticiteit
(gelijkheid van varianties) van de data nagegaan. Via de Kolmogorov-Smirnov
normaliteitstest werd de normaliteit gecontroleerd.
Indien de gegevens normaal verdeeld waren, werd een enkelvoudige variantie-analyse (one-
way-ANOVA) uitgevoerd. Als de varianties gelijk werden bevonden, werd een Tukey test
uitgevoerd om significante verschillen op te sporen. Bij heteroscedasticiteit (varianties niet
gelijk) werd een Tamhane test uitgevoerd om significante verschillen op te zoeken. De P-
waarde werd ingesteld op 0,05. Indien slechts twee gemiddelden vergeleken werden, werd een
independent sample t-test uitgevoerd.
Wanneer het effect van twee factoren op de parameters bestudeerd werd, werd een
tweevoudige variantie-analyse uitgevoerd (two-way-ANOVA). Interactie tussen de factoren
werd nagegaan. Deze was significant bij een P-waarde kleiner dan 0,05.
Als de gegevens niet normaal verdeeld waren, werd gebruik gemaakt van een niet-
parametrische test om significante verschillen op te sporen. Via een Kruskal-Wallis test werd
nagegaan of er significante verschillen tussen verschillende behandelingen waren. Indien dit
zo was, werden de gemiddelden twee aan twee vergelijken met een Mann-Whitney test. De P-
waarde werd opnieuw ingesteld op 0,05.
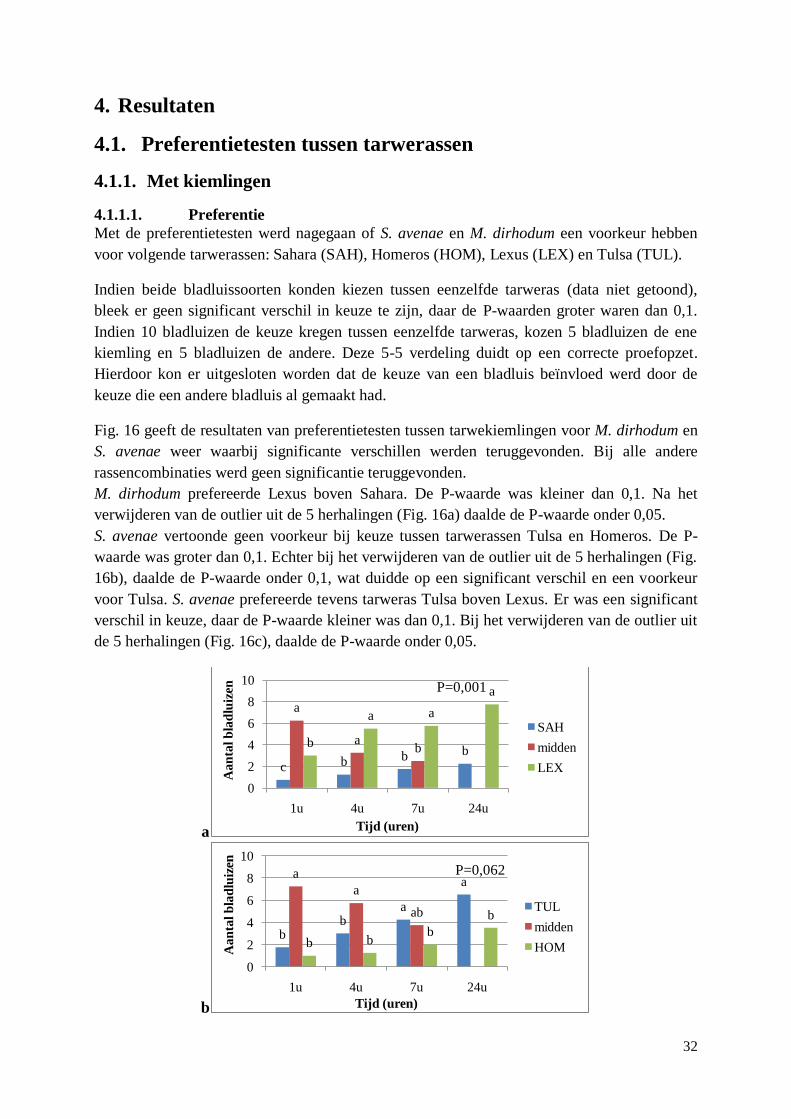
32
4. Resultaten
4.1. Preferentietesten tussen tarwerassen
4.1.1. Met kiemlingen
4.1.1.1. Preferentie
Met de preferentietesten werd nagegaan of S. avenae en M. dirhodum een voorkeur hebben
voor volgende tarwerassen: Sahara (SAH), Homeros (HOM), Lexus (LEX) en Tulsa (TUL).
Indien beide bladluissoorten konden kiezen tussen eenzelfde tarweras (data niet getoond),
bleek er geen significant verschil in keuze te zijn, daar de P-waarden groter waren dan 0,1.
Indien 10 bladluizen de keuze kregen tussen eenzelfde tarweras, kozen 5 bladluizen de ene
kiemling en 5 bladluizen de andere. Deze 5-5 verdeling duidt op een correcte proefopzet.
Hierdoor kon er uitgesloten worden dat de keuze van een bladluis beïnvloed werd door de
keuze die een andere bladluis al gemaakt had.
Fig. 16 geeft de resultaten van preferentietesten tussen tarwekiemlingen voor M. dirhodum en
S. avenae weer waarbij significante verschillen werden teruggevonden. Bij alle andere
rassencombinaties werd geen significantie teruggevonden.
M. dirhodum prefereerde Lexus boven Sahara. De P-waarde was kleiner dan 0,1. Na het
verwijderen van de outlier uit de 5 herhalingen (Fig. 16a) daalde de P-waarde onder 0,05.
S. avenae vertoonde geen voorkeur bij keuze tussen tarwerassen Tulsa en Homeros. De P-
waarde was groter dan 0,1. Echter bij het verwijderen van de outlier uit de 5 herhalingen (Fig.
16b), daalde de P-waarde onder 0,1, wat duidde op een significant verschil en een voorkeur
voor Tulsa. S. avenae prefereerde tevens tarweras Tulsa boven Lexus. Er was een significant
verschil in keuze, daar de P-waarde kleiner was dan 0,1. Bij het verwijderen van de outlier uit
de 5 herhalingen (Fig. 16c), daalde de P-waarde onder 0,05.
a
b
c b b b
a
abb
a a
a
0
2
4
6
8
10
1u 4u 7u 24u
Aan
tal b
lad
luiz
en
Tijd (uren)
SAH
midden
LEX
P=0,001
bb
a
aa
a
ab
b bb
b
0
2
4
6
8
10
1u 4u 7u 24u
Aan
tal b
lad
luiz
en
Tijd (uren)
TUL
midden
HOM
P=0,062
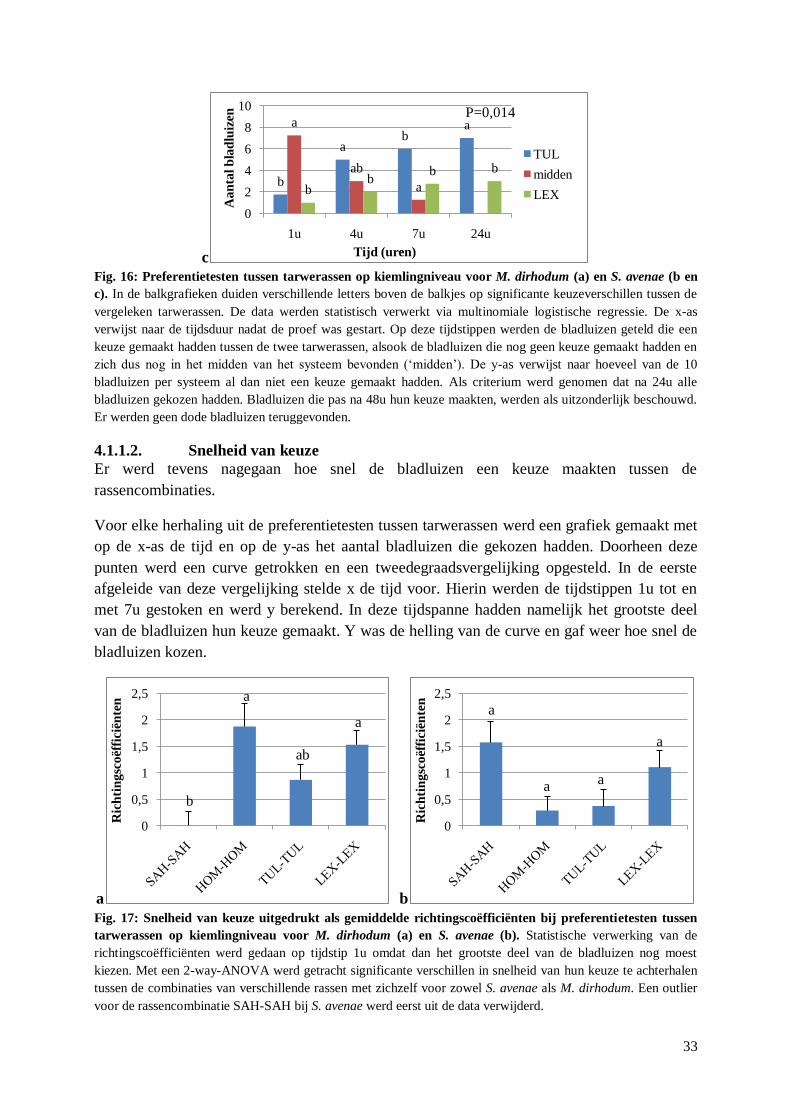
33
c
Fig. 16: Preferentietesten tussen tarwerassen op kiemlingniveau voor M. dirhodum (a) en S. avenae (b en
c). In de balkgrafieken duiden verschillende letters boven de balkjes op significante keuzeverschillen tussen de
vergeleken tarwerassen. De data werden statistisch verwerkt via multinomiale logistische regressie. De x-as
verwijst naar de tijdsduur nadat de proef was gestart. Op deze tijdstippen werden de bladluizen geteld die een
keuze gemaakt hadden tussen de twee tarwerassen, alsook de bladluizen die nog geen keuze gemaakt hadden en
zich dus nog in het midden van het systeem bevonden („midden‟). De y-as verwijst naar hoeveel van de 10
bladluizen per systeem al dan niet een keuze gemaakt hadden. Als criterium werd genomen dat na 24u alle
bladluizen gekozen hadden. Bladluizen die pas na 48u hun keuze maakten, werden als uitzonderlijk beschouwd.
Er werden geen dode bladluizen teruggevonden.
4.1.1.2. Snelheid van keuze
Er werd tevens nagegaan hoe snel de bladluizen een keuze maakten tussen de
rassencombinaties.
Voor elke herhaling uit de preferentietesten tussen tarwerassen werd een grafiek gemaakt met
op de x-as de tijd en op de y-as het aantal bladluizen die gekozen hadden. Doorheen deze
punten werd een curve getrokken en een tweedegraadsvergelijking opgesteld. In de eerste
afgeleide van deze vergelijking stelde x de tijd voor. Hierin werden de tijdstippen 1u tot en
met 7u gestoken en werd y berekend. In deze tijdspanne hadden namelijk het grootste deel
van de bladluizen hun keuze gemaakt. Y was de helling van de curve en gaf weer hoe snel de
bladluizen kozen.
a b
Fig. 17: Snelheid van keuze uitgedrukt als gemiddelde richtingscoëfficiënten bij preferentietesten tussen
tarwerassen op kiemlingniveau voor M. dirhodum (a) en S. avenae (b). Statistische verwerking van de
richtingscoëfficiënten werd gedaan op tijdstip 1u omdat dan het grootste deel van de bladluizen nog moest
kiezen. Met een 2-way-ANOVA werd getracht significante verschillen in snelheid van hun keuze te achterhalen
tussen de combinaties van verschillende rassen met zichzelf voor zowel S. avenae als M. dirhodum. Een outlier
voor de rassencombinatie SAH-SAH bij S. avenae werd eerst uit de data verwijderd.
b
ab
aa
ab
abb
b b
0
2
4
6
8
10
1u 4u 7u 24uA
an
tal
bla
dlu
izen
Tijd (uren)
TUL
midden
LEX
P=0,014
0
0,5
1
1,5
2
2,5
Ric
hti
ngsc
oëf
fici
ënte
n
b
a
ab
a
0
0,5
1
1,5
2
2,5
Ric
hti
ngsc
oëf
fici
ënte
n
a
aa
a

34
Het gemiddelde van de richtingscoëfficiënten voor M. dirhodum na 1u bij keuze tussen
dezelfde tarwerassen (kiemlingen) bedroeg 1,07 ± 0,41. Bij S. avenae was dit 0,75 ± 0,31.
Significante verschillen in richtingscoëfficiënten werden voor M. dirhodum teruggevonden
tussen HOM-HOM en SAH-SAH en tussen LEX-LEX en SAH-SAH (Fig. 17a). Er werden
geen significante verschillen voor S. avenae teruggevonden (Fig. 17b).
Eveneens met een 2-way-ANOVA werd er tussen beide bladluissoorten na 1u een significant
verschil in richtingscoëfficiënten teruggevonden voor SAH-SAH (P-waarde = 0,0264) en
HOM-HOM (P-waarde = 0,0140). Voor TUL-TUL (P-waarde = 0,9332) en LEX-LEX (P-
waarde = 0,9720) werden geen significante verschillen tussen beide bladluissoorten
gevonden. Bij de combinatie SAH-SAH koos S. avenae sneller dan M. dirhodum. Bij de
combinatie HOM-HOM was dit omgekeerd.
4.1.2. Met aren
4.1.2.1. Preferentie
Een gelijkaardige procedure werd uitgevoerd bij de preferentietesten op aarniveau waarbij de
bladluissoort S. avenae kon kiezen tussen dezelfde rassencombinaties (LEX en HOM). Beide
combinaties hadden 5 herhalingen. Eerst werd de preferentie nagegaan. Er werden geen
significante verschillen in preferentie teruggevonden met een multinomiale logistische
regressie wanneer de bladluizen konden kiezen tussen hetzelfde ras (LEX of HOM). Dit wijst
dus op een betrouwbare proefopzet.
4.1.2.2. Snelheid van keuze
Daarna werd de snelheid van keuze op aarniveau nagegaan. De gemiddelde
richtingcoëfficiënt voor LEX-LEX na 1u was 3,72 ± 0,14. Voor HOM-HOM bedroeg dit 3,66
± 0,17. Hiertussen werden geen significante verschillen in richtingscoëfficiënten na 1u
teruggevonden met een independent sample t-test.
4.1.3. Vergelijking voor S. avenae tussen aren en kiemlingen
Voor de kiemlingen bedroegen de richtingscoëfficiënten na 1u voor LEX-LEX 1,11 ± 0,31 en
voor HOM-HOM 0,29 ± 0,27. De richtingcoëfficiënt voor aren na 1u voor LEX-LEX was
3,72 ± 0,14. Voor HOM-HOM bedroeg dit 3,66 ± 0,17.
Met een independent sample t-test werd er een significant verschil teruggevonden (P-waarde
< 0,001) voor HOM-HOM tussen kiemlingen en aren. Voor LEX-LEX werd ook een
significant verschil teruggevonden (P-waarde < 0,001).
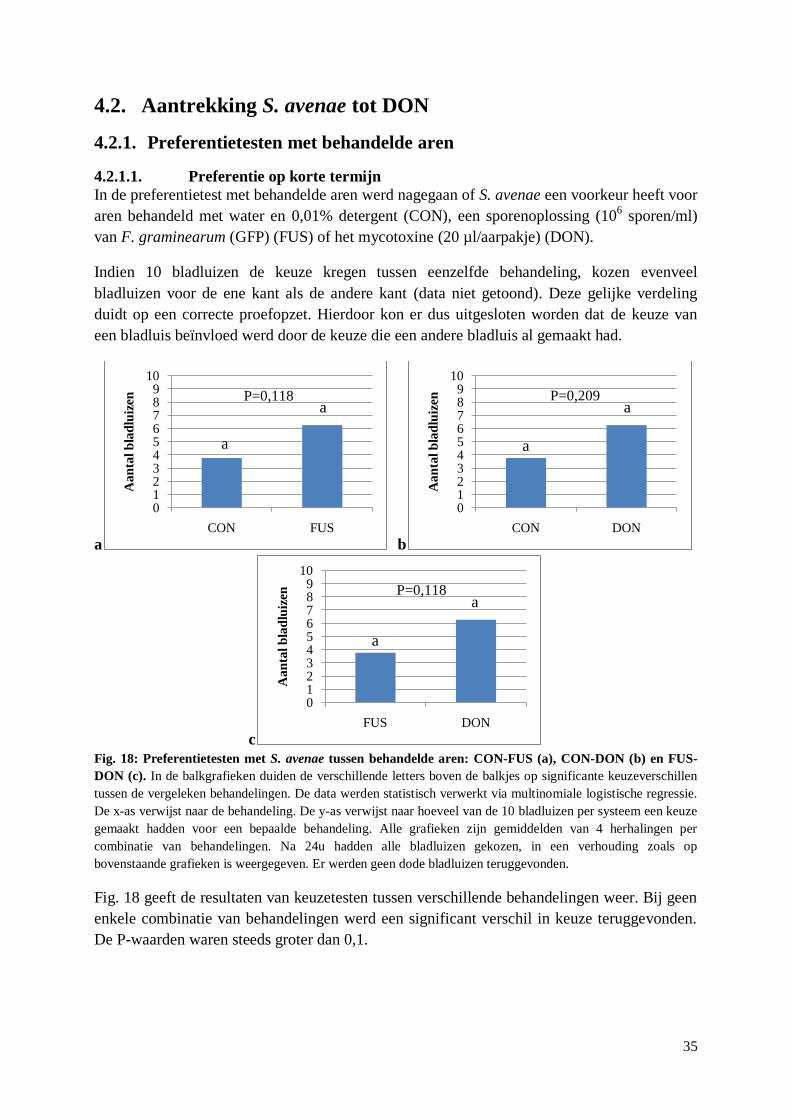
35
4.2. Aantrekking S. avenae tot DON
4.2.1. Preferentietesten met behandelde aren
4.2.1.1. Preferentie op korte termijn
In de preferentietest met behandelde aren werd nagegaan of S. avenae een voorkeur heeft voor
aren behandeld met water en 0,01% detergent (CON), een sporenoplossing (106 sporen/ml)
van F. graminearum (GFP) (FUS) of het mycotoxine (20 µl/aarpakje) (DON).
Indien 10 bladluizen de keuze kregen tussen eenzelfde behandeling, kozen evenveel
bladluizen voor de ene kant als de andere kant (data niet getoond). Deze gelijke verdeling
duidt op een correcte proefopzet. Hierdoor kon er dus uitgesloten worden dat de keuze van
een bladluis beïnvloed werd door de keuze die een andere bladluis al gemaakt had.
a b
c
Fig. 18: Preferentietesten met S. avenae tussen behandelde aren: CON-FUS (a), CON-DON (b) en FUS-
DON (c). In de balkgrafieken duiden de verschillende letters boven de balkjes op significante keuzeverschillen
tussen de vergeleken behandelingen. De data werden statistisch verwerkt via multinomiale logistische regressie.
De x-as verwijst naar de behandeling. De y-as verwijst naar hoeveel van de 10 bladluizen per systeem een keuze
gemaakt hadden voor een bepaalde behandeling. Alle grafieken zijn gemiddelden van 4 herhalingen per
combinatie van behandelingen. Na 24u hadden alle bladluizen gekozen, in een verhouding zoals op
bovenstaande grafieken is weergegeven. Er werden geen dode bladluizen teruggevonden.
Fig. 18 geeft de resultaten van keuzetesten tussen verschillende behandelingen weer. Bij geen
enkele combinatie van behandelingen werd een significant verschil in keuze teruggevonden.
De P-waarden waren steeds groter dan 0,1.
a
a
0123456789
10
CON FUS
Aan
tal b
lad
luiz
en P=0,118
a
a
0123456789
10
CON DON
Aan
tal b
lad
luiz
en P=0,209
a
a
0123456789
10
FUS DON
Aan
tal b
lad
luiz
en P=0,118
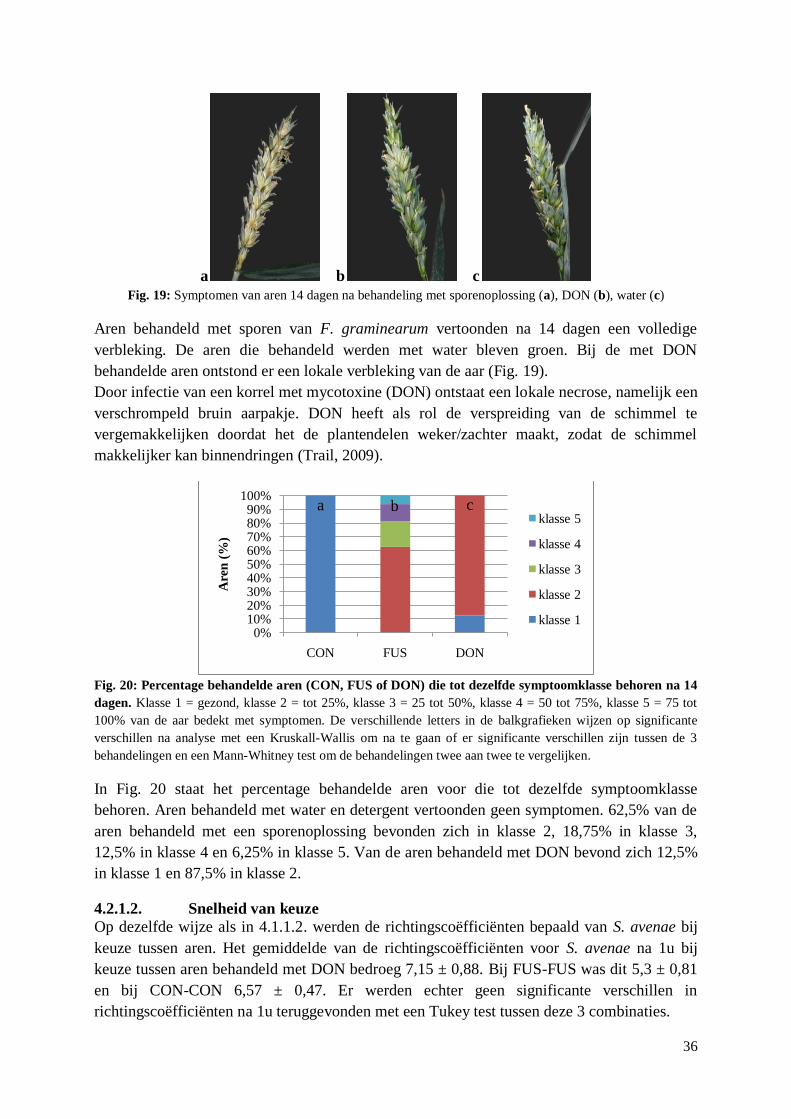
36
a b c
Fig. 19: Symptomen van aren 14 dagen na behandeling met sporenoplossing (a), DON (b), water (c)
Aren behandeld met sporen van F. graminearum vertoonden na 14 dagen een volledige
verbleking. De aren die behandeld werden met water bleven groen. Bij de met DON
behandelde aren ontstond er een lokale verbleking van de aar (Fig. 19).
Door infectie van een korrel met mycotoxine (DON) ontstaat een lokale necrose, namelijk een
verschrompeld bruin aarpakje. DON heeft als rol de verspreiding van de schimmel te
vergemakkelijken doordat het de plantendelen weker/zachter maakt, zodat de schimmel
makkelijker kan binnendringen (Trail, 2009).
Fig. 20: Percentage behandelde aren (CON, FUS of DON) die tot dezelfde symptoomklasse behoren na 14
dagen. Klasse 1 = gezond, klasse 2 = tot 25%, klasse 3 = 25 tot 50%, klasse 4 = 50 tot 75%, klasse 5 = 75 tot
100% van de aar bedekt met symptomen. De verschillende letters in de balkgrafieken wijzen op significante
verschillen na analyse met een Kruskall-Wallis om na te gaan of er significante verschillen zijn tussen de 3
behandelingen en een Mann-Whitney test om de behandelingen twee aan twee te vergelijken.
In Fig. 20 staat het percentage behandelde aren voor die tot dezelfde symptoomklasse
behoren. Aren behandeld met water en detergent vertoonden geen symptomen. 62,5% van de
aren behandeld met een sporenoplossing bevonden zich in klasse 2, 18,75% in klasse 3,
12,5% in klasse 4 en 6,25% in klasse 5. Van de aren behandeld met DON bevond zich 12,5%
in klasse 1 en 87,5% in klasse 2.
4.2.1.2. Snelheid van keuze
Op dezelfde wijze als in 4.1.1.2. werden de richtingscoëfficiënten bepaald van S. avenae bij
keuze tussen aren. Het gemiddelde van de richtingscoëfficiënten voor S. avenae na 1u bij
keuze tussen aren behandeld met DON bedroeg 7,15 ± 0,88. Bij FUS-FUS was dit 5,3 ± 0,81
en bij CON-CON 6,57 ± 0,47. Er werden echter geen significante verschillen in
richtingscoëfficiënten na 1u teruggevonden met een Tukey test tussen deze 3 combinaties.
0%10%20%30%40%50%60%70%80%90%
100%
CON FUS DON
Are
n (
%)
klasse 5
klasse 4
klasse 3
klasse 2
klasse 1
a b c
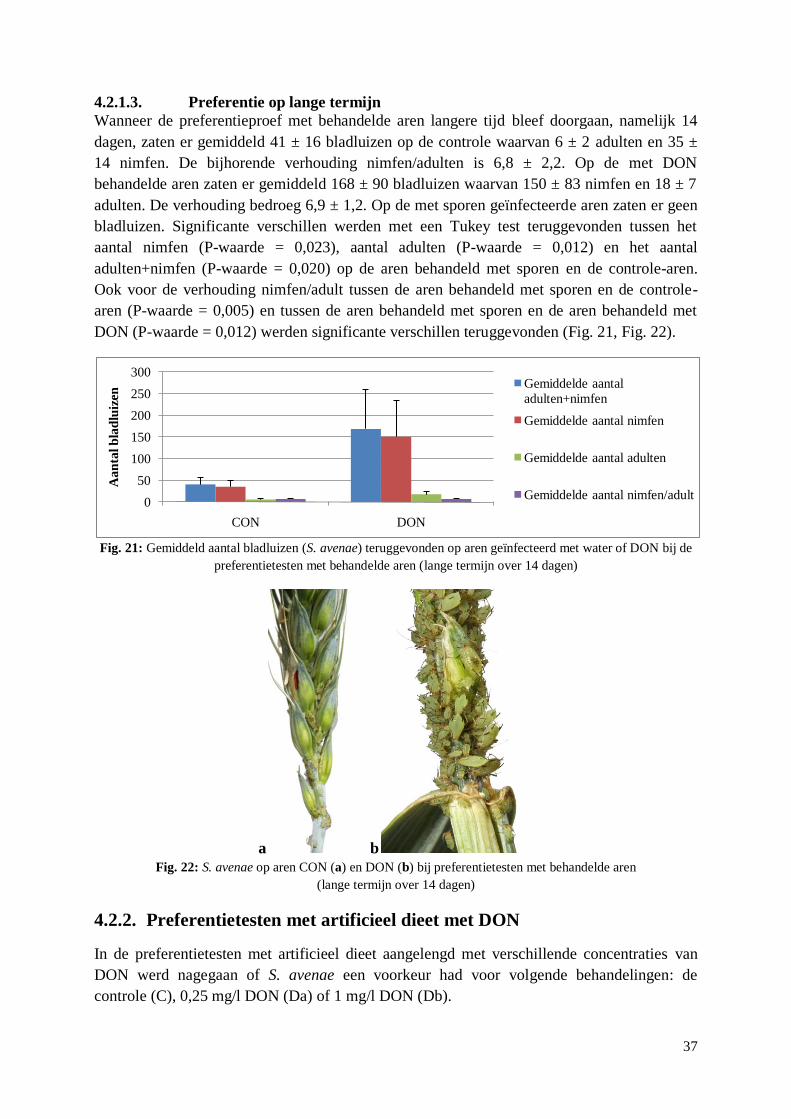
37
4.2.1.3. Preferentie op lange termijn
Wanneer de preferentieproef met behandelde aren langere tijd bleef doorgaan, namelijk 14
dagen, zaten er gemiddeld 41 ± 16 bladluizen op de controle waarvan 6 ± 2 adulten en 35 ±
14 nimfen. De bijhorende verhouding nimfen/adulten is 6,8 ± 2,2. Op de met DON
behandelde aren zaten er gemiddeld 168 ± 90 bladluizen waarvan 150 ± 83 nimfen en 18 ± 7
adulten. De verhouding bedroeg 6,9 ± 1,2. Op de met sporen geïnfecteerde aren zaten er geen
bladluizen. Significante verschillen werden met een Tukey test teruggevonden tussen het
aantal nimfen (P-waarde = 0,023), aantal adulten (P-waarde = 0,012) en het aantal
adulten+nimfen (P-waarde = 0,020) op de aren behandeld met sporen en de controle-aren.
Ook voor de verhouding nimfen/adult tussen de aren behandeld met sporen en de controle-
aren (P-waarde = 0,005) en tussen de aren behandeld met sporen en de aren behandeld met
DON (P-waarde = 0,012) werden significante verschillen teruggevonden (Fig. 21, Fig. 22).
Fig. 21: Gemiddeld aantal bladluizen (S. avenae) teruggevonden op aren geïnfecteerd met water of DON bij de
preferentietesten met behandelde aren (lange termijn over 14 dagen)
a b Fig. 22: S. avenae op aren CON (a) en DON (b) bij preferentietesten met behandelde aren
(lange termijn over 14 dagen)
4.2.2. Preferentietesten met artificieel dieet met DON
In de preferentietesten met artificieel dieet aangelengd met verschillende concentraties van
DON werd nagegaan of S. avenae een voorkeur had voor volgende behandelingen: de
controle (C), 0,25 mg/l DON (Da) of 1 mg/l DON (Db).
0
50
100
150
200
250
300
CON DON
Aan
tal b
lad
luiz
en
Gemiddelde aantal
adulten+nimfen
Gemiddelde aantal nimfen
Gemiddelde aantal adulten
Gemiddelde aantal nimfen/adult
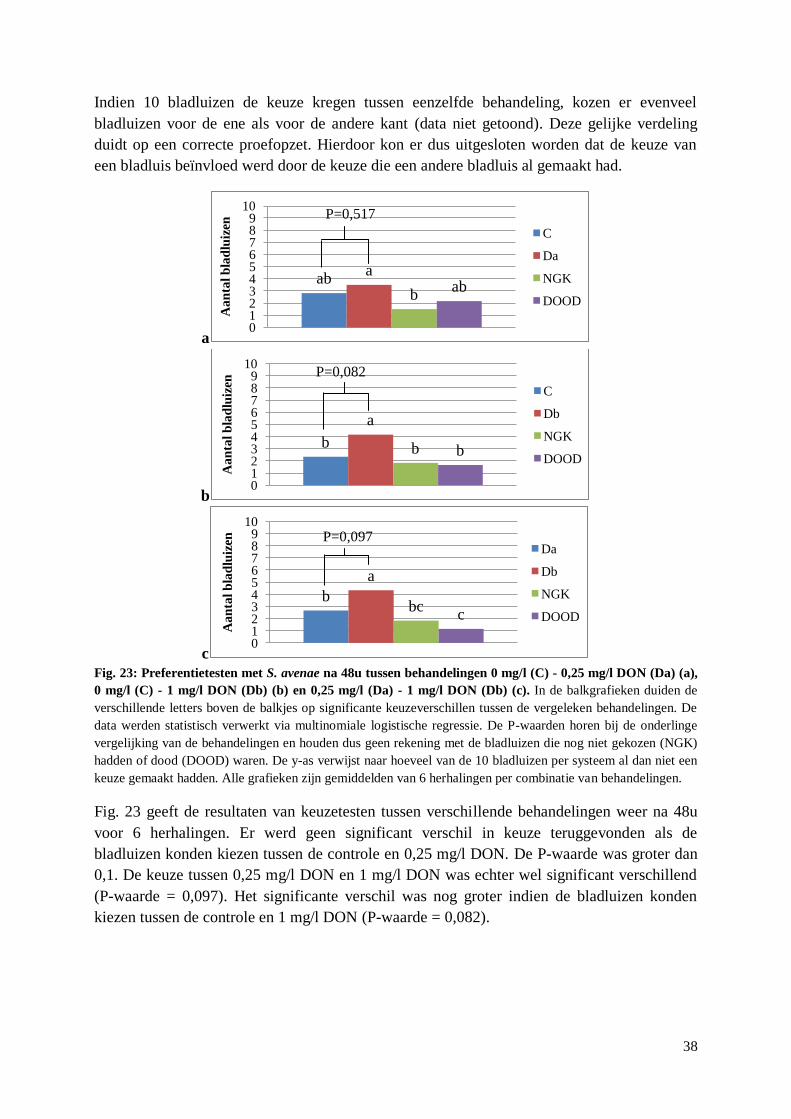
38
Indien 10 bladluizen de keuze kregen tussen eenzelfde behandeling, kozen er evenveel
bladluizen voor de ene als voor de andere kant (data niet getoond). Deze gelijke verdeling
duidt op een correcte proefopzet. Hierdoor kon er dus uitgesloten worden dat de keuze van
een bladluis beïnvloed werd door de keuze die een andere bladluis al gemaakt had.
a
b
c
Fig. 23: Preferentietesten met S. avenae na 48u tussen behandelingen 0 mg/l (C) - 0,25 mg/l DON (Da) (a),
0 mg/l (C) - 1 mg/l DON (Db) (b) en 0,25 mg/l (Da) - 1 mg/l DON (Db) (c). In de balkgrafieken duiden de
verschillende letters boven de balkjes op significante keuzeverschillen tussen de vergeleken behandelingen. De
data werden statistisch verwerkt via multinomiale logistische regressie. De P-waarden horen bij de onderlinge
vergelijking van de behandelingen en houden dus geen rekening met de bladluizen die nog niet gekozen (NGK)
hadden of dood (DOOD) waren. De y-as verwijst naar hoeveel van de 10 bladluizen per systeem al dan niet een
keuze gemaakt hadden. Alle grafieken zijn gemiddelden van 6 herhalingen per combinatie van behandelingen.
Fig. 23 geeft de resultaten van keuzetesten tussen verschillende behandelingen weer na 48u
voor 6 herhalingen. Er werd geen significant verschil in keuze teruggevonden als de
bladluizen konden kiezen tussen de controle en 0,25 mg/l DON. De P-waarde was groter dan
0,1. De keuze tussen 0,25 mg/l DON en 1 mg/l DON was echter wel significant verschillend
(P-waarde = 0,097). Het significante verschil was nog groter indien de bladluizen konden
kiezen tussen de controle en 1 mg/l DON (P-waarde = 0,082).
aba
bab
0123456789
10
Aan
tal
bla
dlu
izen
C
Da
NGK
DOOD
P=0,517
b
a
b b
0123456789
10
Aan
tal
bla
dlu
izen
C
Db
NGK
DOOD
P=0,082
b
a
bcc
0123456789
10
Aan
tal b
lad
luiz
en
Da
Db
NGK
DOOD
P=0,097
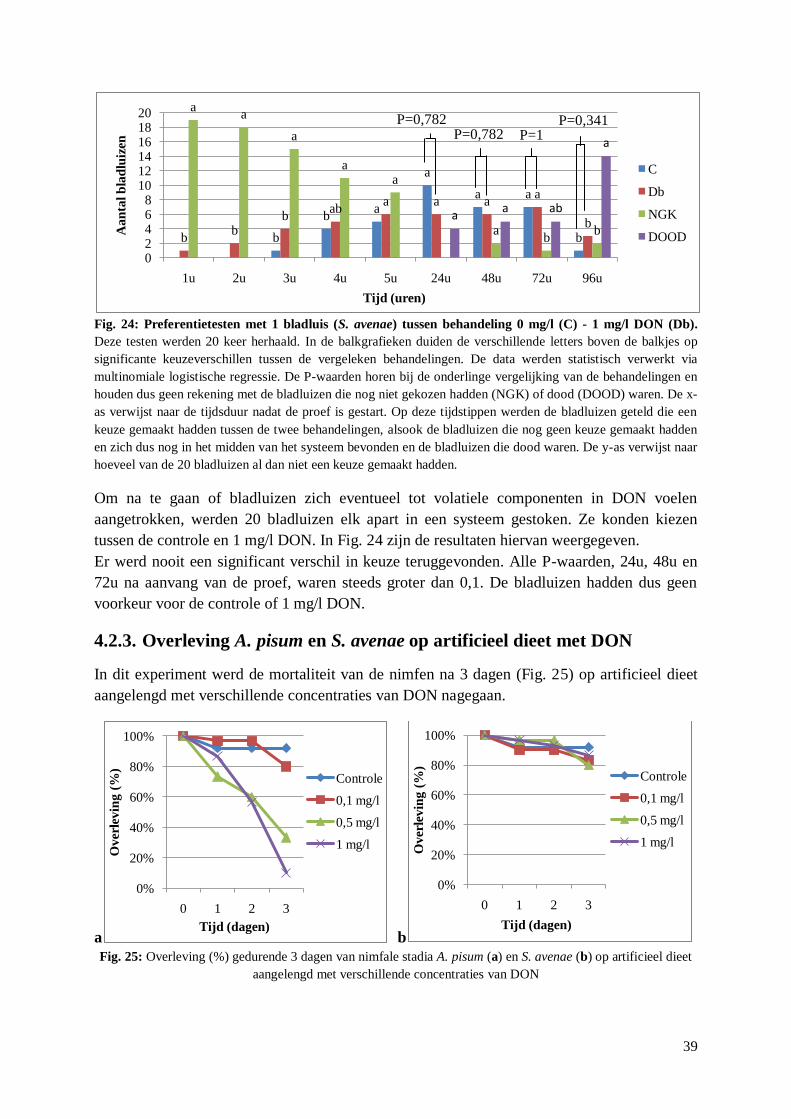
39
Fig. 24: Preferentietesten met 1 bladluis (S. avenae) tussen behandeling 0 mg/l (C) - 1 mg/l DON (Db).
Deze testen werden 20 keer herhaald. In de balkgrafieken duiden de verschillende letters boven de balkjes op
significante keuzeverschillen tussen de vergeleken behandelingen. De data werden statistisch verwerkt via
multinomiale logistische regressie. De P-waarden horen bij de onderlinge vergelijking van de behandelingen en
houden dus geen rekening met de bladluizen die nog niet gekozen hadden (NGK) of dood (DOOD) waren. De x-
as verwijst naar de tijdsduur nadat de proef is gestart. Op deze tijdstippen werden de bladluizen geteld die een
keuze gemaakt hadden tussen de twee behandelingen, alsook de bladluizen die nog geen keuze gemaakt hadden
en zich dus nog in het midden van het systeem bevonden en de bladluizen die dood waren. De y-as verwijst naar
hoeveel van de 20 bladluizen al dan niet een keuze gemaakt hadden.
Om na te gaan of bladluizen zich eventueel tot volatiele componenten in DON voelen
aangetrokken, werden 20 bladluizen elk apart in een systeem gestoken. Ze konden kiezen
tussen de controle en 1 mg/l DON. In Fig. 24 zijn de resultaten hiervan weergegeven.
Er werd nooit een significant verschil in keuze teruggevonden. Alle P-waarden, 24u, 48u en
72u na aanvang van de proef, waren steeds groter dan 0,1. De bladluizen hadden dus geen
voorkeur voor de controle of 1 mg/l DON.
4.2.3. Overleving A. pisum en S. avenae op artificieel dieet met DON
In dit experiment werd de mortaliteit van de nimfen na 3 dagen (Fig. 25) op artificieel dieet
aangelengd met verschillende concentraties van DON nagegaan.
a b
Fig. 25: Overleving (%) gedurende 3 dagen van nimfale stadia A. pisum (a) en S. avenae (b) op artificieel dieet
aangelengd met verschillende concentraties van DON
b
ba
a
a a
bbb
bab
a a aa
b
aa
a
aa
ab
b
aa ab
a
02468
101214161820
1u 2u 3u 4u 5u 24u 48u 72u 96u
Aan
tal
bla
dlu
izen
Tijd (uren)
C
Db
NGK
DOOD
P=0,782P=0,782 P=1
P=0,341
0%
20%
40%
60%
80%
100%
0 1 2 3
Over
levin
g (
%)
Tijd (dagen)
Controle
0,1 mg/l
0,5 mg/l
1 mg/l
0%
20%
40%
60%
80%
100%
0 1 2 3
Over
levin
g (
%)
Tijd (dagen)
Controle
0,1 mg/l
0,5 mg/l
1 mg/l

40
Tabel 3: Overleving nimfale stadia van A. pisum en S. avenae na 3 dagen op artificieel dieet aangelengd
met verschillende concentraties van DON. De kleine letters duiden op significante verschillen tussen
concentraties van DON, de grote letters duiden op significante verschillen tussen de bladluissoorten, na
statistische verwerking van de data met een 2-way-ANOVA.
Controle 0,1 mg/l DON 0,5 mg/l DON 1 mg/l DON
A. pisum 92% ± 0,8% aA 80% ± 4,7% aA 33,3% ± 7,2% bB 10% ± 4,7% bB
S. avenae 83,3% ± 2,7% aA 83,3% ± 7,2% aA 80% ± 0% aA 86,7% ± 7,2% aA
De overleving van A. pisum na 3 dagen (Tabel 3) op de controle en op 0,1 mg/l DON was
significant verschillend van de overleving op 0,5 en 1 mg/l DON. Voor S. avenae werden
geen significante verschillen teruggevonden tussen de verschillende concentraties van DON.
Eveneens met een 2-way-ANOVA werd er tussen de bladluissoorten een significant verschil
in overleving op DON teruggevonden na 3 dagen voor 1 mg/l (P-waarde < 0,001) en voor 0,5
mg/l (P-waarde = 0,0034) maar geen significant verschil voor 0,1 mg/l (P-waarde = 1) en 0
mg/l (P-waarde = 0,9628) (Tabel 3).
4.2.4. Reproductie S. avenae op artificieel dieet met DON
In het experiment omtrent reproductie was het de bedoeling om de overlevings- en
reproductieparameters te achterhalen van S. avenae als deze zich voedde op artificieel dieet
met DON (0,25 mg/l en 1 mg/l).
Tabel 4: Ontwikkelingsduur (in dagen) voor de 4 nimfale stadia van S. avenae. De verschillende letters
duiden op significante verschillen tussen de behandelingen na statistische verwerking met een independent
sample t-test.
Ontwikkelingsduur (in dagen) Controle 0,25 mg/l DON
N1 1,1 ± 0,3 a 1,0 ± 0,5 a
N2 1,4 ± 0,5 a 1,8 ± 0,7 a
N3 1,4 ± 1,0 a 1,3 ± 0,5 a
N4 1,9 ± 0,4 a 1,9 ± 0,4 a
Totaal 5,9 ± 1,1 a 5,9 ± 0,7 a
Er werden geen significante verschillen tussen de controle en 0,25 mg/l DON teruggevonden
voor alle nimfale stadia en de totale nimfale duur. De concentratie 1 mg/l DON werd niet in
beschouwing genomen omdat hierbij de mortaliteit te hoog lag (Tabel 4).
In Fig. 26 is de overleving van de nimfen en het verloop van hun ontwikkeling weergegeven
in functie van hun leeftijd.
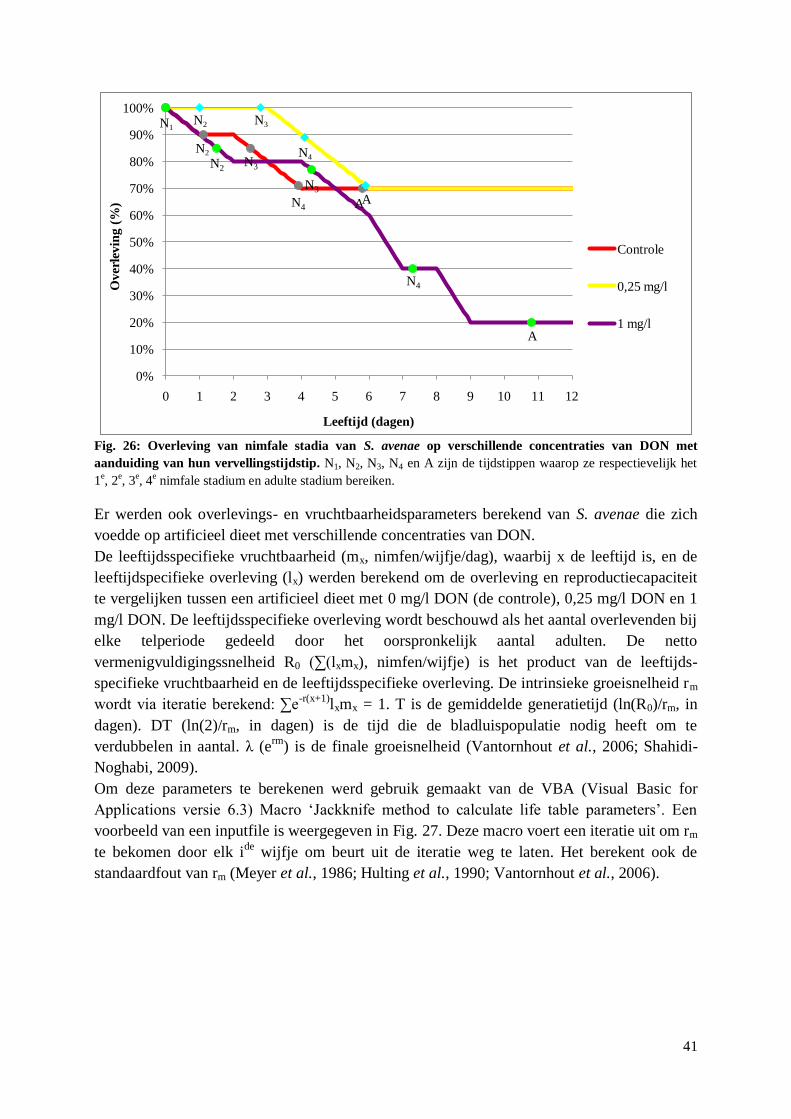
41
Fig. 26: Overleving van nimfale stadia van S. avenae op verschillende concentraties van DON met
aanduiding van hun vervellingstijdstip. N1, N2, N3, N4 en A zijn de tijdstippen waarop ze respectievelijk het
1e, 2
e, 3
e, 4
e nimfale stadium en adulte stadium bereiken.
Er werden ook overlevings- en vruchtbaarheidsparameters berekend van S. avenae die zich
voedde op artificieel dieet met verschillende concentraties van DON.
De leeftijdsspecifieke vruchtbaarheid (mx, nimfen/wijfje/dag), waarbij x de leeftijd is, en de
leeftijdspecifieke overleving (lx) werden berekend om de overleving en reproductiecapaciteit
te vergelijken tussen een artificieel dieet met 0 mg/l DON (de controle), 0,25 mg/l DON en 1
mg/l DON. De leeftijdsspecifieke overleving wordt beschouwd als het aantal overlevenden bij
elke telperiode gedeeld door het oorspronkelijk aantal adulten. De netto
vermenigvuldigingssnelheid R0 (∑(lxmx), nimfen/wijfje) is het product van de leeftijds-
specifieke vruchtbaarheid en de leeftijdsspecifieke overleving. De intrinsieke groeisnelheid rm
wordt via iteratie berekend: ∑e-r(x+1)
lxmx = 1. T is de gemiddelde generatietijd (ln(R0)/rm, in
dagen). DT (ln(2)/rm, in dagen) is de tijd die de bladluispopulatie nodig heeft om te
verdubbelen in aantal. λ (erm
) is de finale groeisnelheid (Vantornhout et al., 2006; Shahidi-
Noghabi, 2009).
Om deze parameters te berekenen werd gebruik gemaakt van de VBA (Visual Basic for
Applications versie 6.3) Macro „Jackknife method to calculate life table parameters‟. Een
voorbeeld van een inputfile is weergegeven in Fig. 27. Deze macro voert een iteratie uit om rm
te bekomen door elk ide
wijfje om beurt uit de iteratie weg te laten. Het berekent ook de
standaardfout van rm (Meyer et al., 1986; Hulting et al., 1990; Vantornhout et al., 2006).
N2
N3
N4 A
N2 N3
N4
A
N1
N2
N3
N4
A
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 1 2 3 4 5 6 7 8 9 10 11 12
Over
levin
g (
%)
Leeftijd (dagen)
Controle
0,25 mg/l
1 mg/l
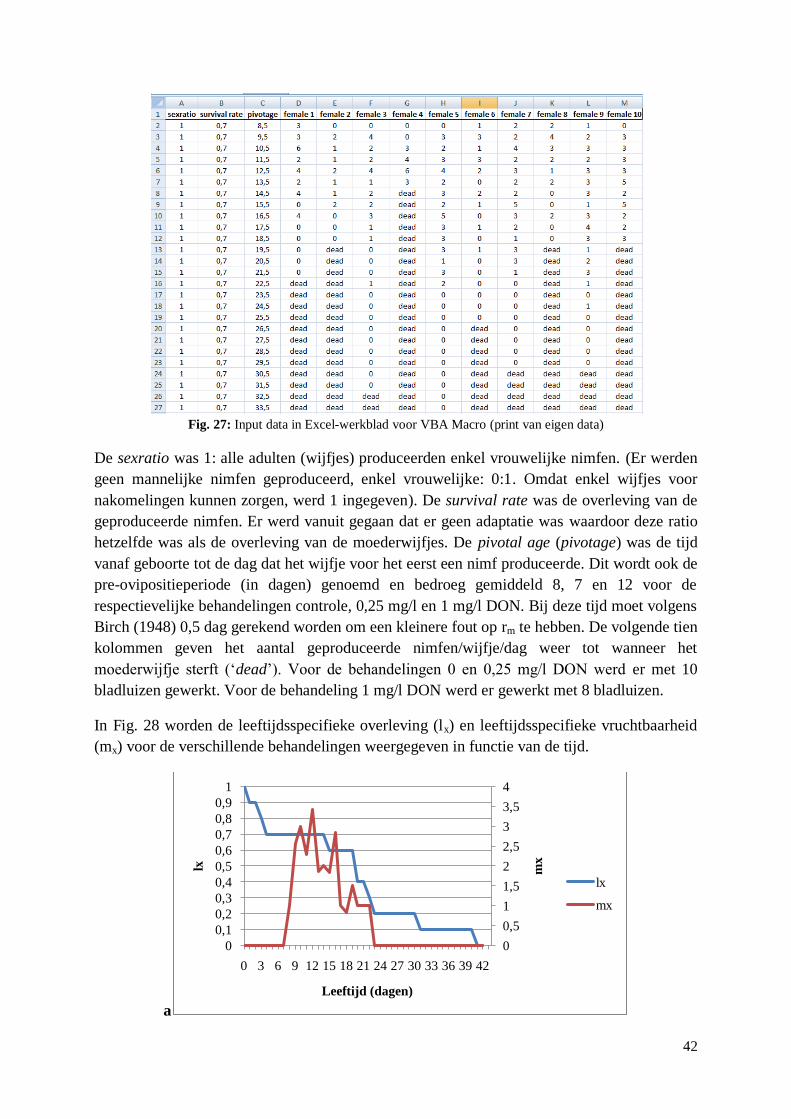
42
Fig. 27: Input data in Excel-werkblad voor VBA Macro (print van eigen data)
De sexratio was 1: alle adulten (wijfjes) produceerden enkel vrouwelijke nimfen. (Er werden
geen mannelijke nimfen geproduceerd, enkel vrouwelijke: 0:1. Omdat enkel wijfjes voor
nakomelingen kunnen zorgen, werd 1 ingegeven). De survival rate was de overleving van de
geproduceerde nimfen. Er werd vanuit gegaan dat er geen adaptatie was waardoor deze ratio
hetzelfde was als de overleving van de moederwijfjes. De pivotal age (pivotage) was de tijd
vanaf geboorte tot de dag dat het wijfje voor het eerst een nimf produceerde. Dit wordt ook de
pre-ovipositieperiode (in dagen) genoemd en bedroeg gemiddeld 8, 7 en 12 voor de
respectievelijke behandelingen controle, 0,25 mg/l en 1 mg/l DON. Bij deze tijd moet volgens
Birch (1948) 0,5 dag gerekend worden om een kleinere fout op rm te hebben. De volgende tien
kolommen geven het aantal geproduceerde nimfen/wijfje/dag weer tot wanneer het
moederwijfje sterft („dead‟). Voor de behandelingen 0 en 0,25 mg/l DON werd er met 10
bladluizen gewerkt. Voor de behandeling 1 mg/l DON werd er gewerkt met 8 bladluizen.
In Fig. 28 worden de leeftijdsspecifieke overleving (lx) en leeftijdsspecifieke vruchtbaarheid
(mx) voor de verschillende behandelingen weergegeven in functie van de tijd.
a
0
0,5
1
1,5
2
2,5
3
3,5
4
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42
mx
lx
Leeftijd (dagen)
lx
mx
43
b
c
Fig. 28: Leeftijdsspecifieke overleving (lx) en leeftijdsspecifieke vruchtbaarheid (mx) op artificieel dieet
aangelengd met 0 mg/l (a), 0,25 mg/l (b) en 1 mg/l DON (c) voor 10 bladluizen (S. avenae) waarvan bij de
overlevende adulten ook de reproductie werd nagegaan.
De gemiddelde adulte levensduur van bladluizen waarbij reproductie werd nagegaan in de
controlegroep was 16,2 ± 8,1 dagen. Voor de behandeling 0,25 mg/l DON was dit 19,7 ± 4,9
dagen en voor 1 mg/l DON 14,9 ± 9,5 dagen. Hiertussen werden met een Tukey test geen
significante verschillen teruggevonden.
Tabel 5 geeft de reproductieparameters voor de verschillende behandelingen weer.
Tabel 5: Overleving- en vruchtbaarheidsparameters van S. avenae voor verschillende concentraties van
DON toegevoegd aan het artificieel dieet. De verschillende letters duiden op significante verschillen binnenin
een rij. Deze werden met Tukey of Tamhane testen (afhankelijk van al dan niet gelijke varianties)
teruggevonden, waarbij de P-waarden steeds kleiner dan 0,001 bedroegen.
Parameters Controle 0,25 mg/l DON 1 mg/l DON
R0 17,43 a 13,65 b 2,08 c
rm 0,225 ± 0,008 b 0,231 ± 0,007 a 0,046 ± 0,011 c
T 12,71 b 11,33 c 16,16 a
DT 3,38 b 3,29 c 15,72 a
λ 1,228 b 1,235 a 1,045 c
0
0,5
1
1,5
2
2,5
3
3,5
4
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42
mx
lx
Leeftijd (dagen)
lx
mx
0
0,5
1
1,5
2
2,5
3
3,5
4
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42m
x
lx
Leeftijd (dagen)
lx
mx

44
4.3. Verspreiding F. graminearum door S. avenae
4.3.1. Orale opname van macroconidia door S. avenae
Om na te gaan of adulte S. avenae de sporen van F. graminearum (GFP) kunnen opnemen,
werden de bladluizen die aan de sporenoplossing zogen, uitgeplaat op medium met
antibiotica. Hierbij werden de petrischalen na minimum 4 dagen incubatie bij 25°C gecheckt
onder de fluorescentiemicroscoop. Hierop werd geen fluorescentie waargenomen. In de
uitscheidingen van de bladluizen zaten dus geen sporen.
Om na te gaan of de bladluizen de sporen konden opnemen maar niet uitscheiden, werden 30
zuigende bladluizen gegrind en uitgeplaat. Ook hier werd geen fluorescentie waargenomen op
de petriplaten. Bij het grinden van bladluizen en sporen samen, werd er wel fluorescentie
teruggevonden. De sporen konden ook na het grinden op met medium verder leven.
4.3.2. Metingen van stiletten S. avenae en M. dirhodum
Onder de binoculair werden de stiletafmetingen van adulte S. avenae en adulte M. dirhodum
bekeken (Fig. 29). Van elke bladluizensoort werden 20 bladluizen bekeken.
Bij S. avenae bedroeg de gemiddelde lengte van het stilet 357 µm ± 58 µm en de breedte 63
µm ± 13 µm. Bij M. dirhodum was dit 316 µm ± 51 µm en 44 µm ± 10 µm.
a b
Fig. 29: Uitwendige afmetingen van het stilet van S. avenae (a) en M. dirhodum (b)
4.3.3. Uitwendige acquisitie en verspreiding F. graminearum door S. avenae
Allereerst werd nagegaan hoe groot de proportie adulte bladluizen was die inoculum droeg op
hun lichamen na 4u in sporulerende F. graminearum (GFP) te lopen. Deze werden op
medium met antibiotica uitgeplaat. In de eerste herhaling droegen er 2, 1, 1, 9 en 2 van de 10
bladluizen per sporulerende plaat inoculum. In de tweede herhaling was dit 0, 6, 2, 2, 1 van de
10 bladluizen. Gemiddeld gezien droegen 26% ± 4% van de bladluizen inoculum.
Er werden ook enkele adulte bladluizen die uit de sporulerende schimmel kwamen, bekeken
onder de fluorescentiemicroscoop. Zowel aan de poten als aan de antennes van de bladluizen
werd inoculum teruggevonden. Het mycelium maar ook de sporen kunnen op medium groeien
en gedetecteerd worden via fluorescentie (Fig. 30).
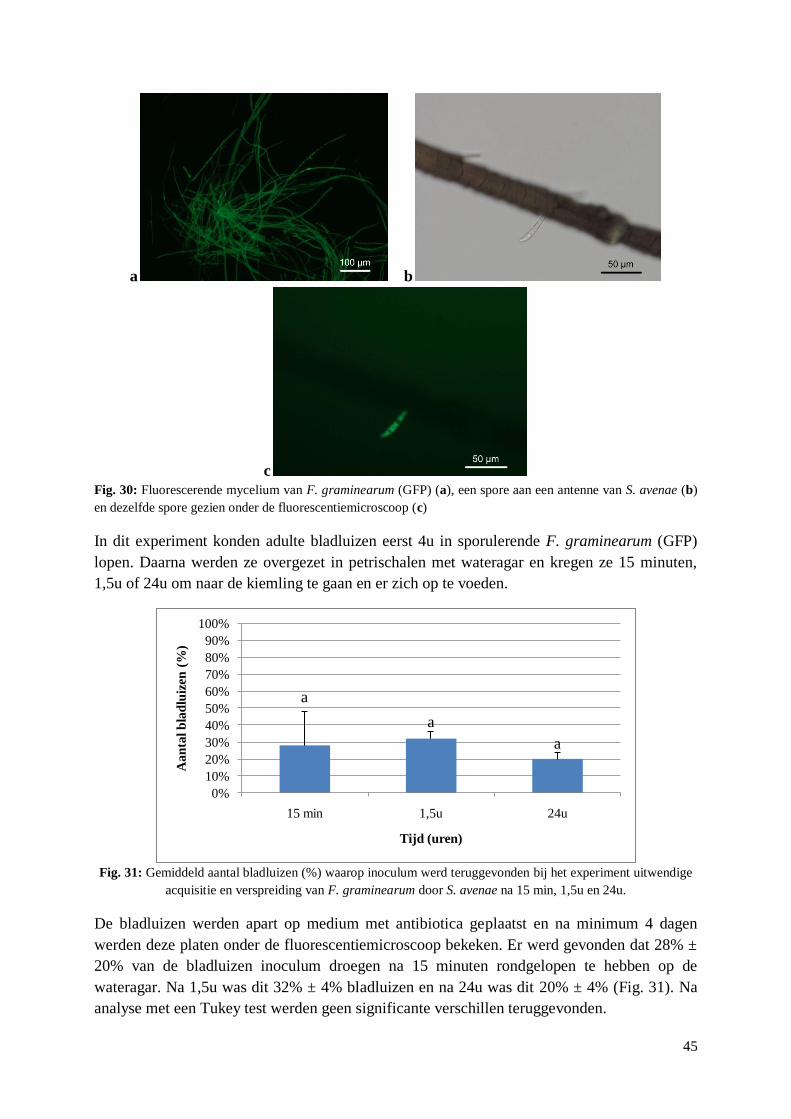
45
a b
c
Fig. 30: Fluorescerende mycelium van F. graminearum (GFP) (a), een spore aan een antenne van S. avenae (b)
en dezelfde spore gezien onder de fluorescentiemicroscoop (c)
In dit experiment konden adulte bladluizen eerst 4u in sporulerende F. graminearum (GFP)
lopen. Daarna werden ze overgezet in petrischalen met wateragar en kregen ze 15 minuten,
1,5u of 24u om naar de kiemling te gaan en er zich op te voeden.
Fig. 31: Gemiddeld aantal bladluizen (%) waarop inoculum werd teruggevonden bij het experiment uitwendige
acquisitie en verspreiding van F. graminearum door S. avenae na 15 min, 1,5u en 24u.
De bladluizen werden apart op medium met antibiotica geplaatst en na minimum 4 dagen
werden deze platen onder de fluorescentiemicroscoop bekeken. Er werd gevonden dat 28% ±
20% van de bladluizen inoculum droegen na 15 minuten rondgelopen te hebben op de
wateragar. Na 1,5u was dit 32% ± 4% bladluizen en na 24u was dit 20% ± 4% (Fig. 31). Na
analyse met een Tukey test werden geen significante verschillen teruggevonden.
a
a
a
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
15 min 1,5u 24u
Aan
tal b
lad
luiz
en (
%)
Tijd (uren)
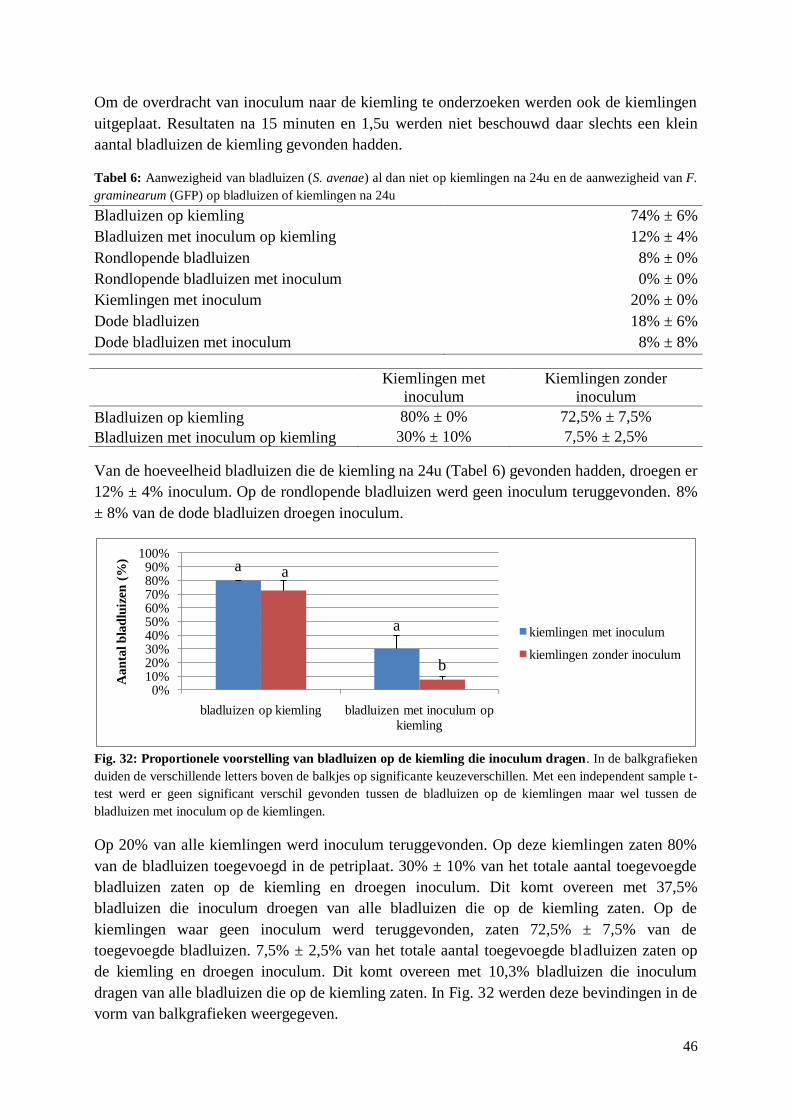
46
Om de overdracht van inoculum naar de kiemling te onderzoeken werden ook de kiemlingen
uitgeplaat. Resultaten na 15 minuten en 1,5u werden niet beschouwd daar slechts een klein
aantal bladluizen de kiemling gevonden hadden.
Tabel 6: Aanwezigheid van bladluizen (S. avenae) al dan niet op kiemlingen na 24u en de aanwezigheid van F.
graminearum (GFP) op bladluizen of kiemlingen na 24u
Bladluizen op kiemling 74% ± 6%
Bladluizen met inoculum op kiemling 12% ± 4%
Rondlopende bladluizen 8% ± 0%
Rondlopende bladluizen met inoculum 0% ± 0%
Kiemlingen met inoculum 20% ± 0%
Dode bladluizen 18% ± 6%
Dode bladluizen met inoculum 8% ± 8%
Kiemlingen met
inoculum
Kiemlingen zonder
inoculum
Bladluizen op kiemling 80% ± 0% 72,5% ± 7,5%
Bladluizen met inoculum op kiemling 30% ± 10% 7,5% ± 2,5%
Van de hoeveelheid bladluizen die de kiemling na 24u (Tabel 6) gevonden hadden, droegen er
12% ± 4% inoculum. Op de rondlopende bladluizen werd geen inoculum teruggevonden. 8%
± 8% van de dode bladluizen droegen inoculum.
Fig. 32: Proportionele voorstelling van bladluizen op de kiemling die inoculum dragen. In de balkgrafieken
duiden de verschillende letters boven de balkjes op significante keuzeverschillen. Met een independent sample t-
test werd er geen significant verschil gevonden tussen de bladluizen op de kiemlingen maar wel tussen de
bladluizen met inoculum op de kiemlingen.
Op 20% van alle kiemlingen werd inoculum teruggevonden. Op deze kiemlingen zaten 80%
van de bladluizen toegevoegd in de petriplaat. 30% ± 10% van het totale aantal toegevoegde
bladluizen zaten op de kiemling en droegen inoculum. Dit komt overeen met 37,5%
bladluizen die inoculum droegen van alle bladluizen die op de kiemling zaten. Op de
kiemlingen waar geen inoculum werd teruggevonden, zaten 72,5% ± 7,5% van de
toegevoegde bladluizen. 7,5% ± 2,5% van het totale aantal toegevoegde bladluizen zaten op
de kiemling en droegen inoculum. Dit komt overeen met 10,3% bladluizen die inoculum
dragen van alle bladluizen die op de kiemling zaten. In Fig. 32 werden deze bevindingen in de
vorm van balkgrafieken weergegeven.
a
a
a
b
0%10%20%30%40%50%60%70%80%90%
100%
bladluizen op kiemling bladluizen met inoculum op
kiemling
Aan
tal b
lad
luiz
en (
%)
kiemlingen met inoculum
kiemlingen zonder inoculum

47
5. Discussie
5.1. Preferentietesten tussen tarwerassen
5.1.1. Preferentie
Onderzoek toonde aan dat M. dirhodum het tarweras Lexus boven Sahara prefereerde.
Daarentegen had S. avenae geen voorkeur voor Lexus of Sahara, maar prefereerde deze Tulsa
boven Homeros en Lexus. Deze preferentie was op zijn beurt niet bij M. dirhodum terug te
vinden. In Fig. 33 wordt hiervan een schematisch voorstelling gegeven. Er kon dus
geconcludeerd worden dat de preferentie voor een bepaald tarweras afhankelijk was van de
soort bladluis. Dit wijst op een intrinsiek verschil in preferentie tussen de soorten S. avenae en
M. dirhodum.
a b Fig. 33: Schematische voorstelling van preferentie tussen tarwerassen
voor M. dirhodum (a) en S. avenae (b)
Deze bevindingen zijn uniek, daar er voor de eerste keer onderzoek werd gedaan naar de
preferentie van vleugelloze S. avenae en M. dirhodum voor verschillende tarwerassen. In het
verleden werd dit nog nooit met M. dirhodum uitgevoerd. Er werden door Nicol et al. (1992)
wel reeds preferentietesten gedaan tussen verschillende tarwerassen met S. avenae, echter
werd toen met de gevleugelde vormen gewerkt. Preferentietesten met bladluizen tussen
verschillende waardplanten, en de zoektocht naar de reden waarom de ene waardplant
geprefereerd wordt boven de andere, komen wel frequenter aan bod in de literatuur.
Vaak speelt een visuele respons een rol bij het kiezen van een geschikte waardplant
(Pettersson et al., 2007). Bladluizen zullen preferentieel landen op geel gekleurde
oppervlakten omdat deze bepaalde golflengtes van licht reflecteren. Onderzoek toont echter
aan dat niet alleen lichtgolflengtes maar vooral plantvolatielen een rol spelen bij de keuze van
een plant. Bladluizen reageren namelijk op de „geur‟ van een plant en dat speelt een rol in het
vinden van waardplanten en het discrimineren van andere plantensoorten. Hoe dit juist in zijn
werk gaat, is nog niet geweten. Bladluizen bezitten een waaier van olfactorische organen op
hun antennes en met elektrofysiologische methoden kon aangetoond worden dat bladluizen
reageren op een variëteit aan plantvolatielen (Pettersson et al., 2007). Zo ontdekten Visser &
Piron (1997) met behulp van een elektroantennogram (EAG) een significant verschil in
respons ten aanzien van 35 plantvolatielen tussen bladluissoorten Brevicoryne brassicae en
Myzus persicae waaronder hexylacetaat, butylisothiocyanaat, methylsalicylaat, etc. Dit kon
zowel om aantrekking als afstoting gaan. Zo is methylsalicylaat repellent voor R. padi en
wordt het geproduceerd door vogelkers om ervoor te zorgen dat de bladluizen in de lente deze

48
waardplant verlaten en op zoek gaan naar een zomerwaardplant. Methylsalicylaat draagt dus
bij tot discriminatie van een waardplant (Pickett & Glinwood, 2007). R. padi bleek ook
gevoelig te zijn voor benzaldehyde, wat een belangrijk volatiel is van zijn winterwaardplant
vogelkers (Park et al., 2000). Voor S. avenae werden de hoogste EAGs gevonden bij hexanal
en ook bij benzaldehyde, linalool, etc. vertoonde deze bladluissoort significante responsen. M.
dirhodum had, net zoals S. avenae, de hoogste EAG bij hexanal. Gelijkenissen in responsen
ten aanzien van plantvolatielen hebben mogelijks te maken met het feit dat beide
bladluissoorten tot dezelfde familie (Aphididae) en stam (Macrosiphini) behoren (Visser &
Fu-shun, 1995). Kloonpopulaties van alate S. avenae prefereren hun landbouwkundige
waardplant tarwe boven hun natuurlijke waardplant kropaar (Dactylis glomerata L.). De
specifieke waardplantkarakteristieken die deze preferentie veroorzaken bleven onbekend.
Echter bleek uit voorgaand onderzoek dat de proportie alate S. avenae die tarwerassen
koloniseren negatief gecorreleerd was met hydroxaamzuur die deze rassen bevatten (Lushai et
al., 1997). Lushai et al. (1997) konden ook opmerken dat niet alleen de genetische
achtergrond van de bladluizen bijdraagt tot de preferentie maar ook het feit dat indien
bladluizen verschillende generaties op een alternatieve waardplant worden opgegroeid dat dit
kan bijdragen tot een andere preferentie van de alatae. De preferentie van bladluizen kan dus
beïnvloed worden door hun voorgaande waardplanten.
De reden waarom in dit onderzoek M. dirhodum Lexus verkoos boven Sahara en ook waarom
S. avenae Tulsa verkoos boven Homeros en Lexus is niet geweten. Ogenschijnlijk zien alle
kiemlingen van de verschillende tarwerassen er morfologisch hetzelfde uit.
Veronderstellingen waarom deze bladluizen zich meer of minder aangetrokken voelen tot
bepaalde tarwerassen blijven dus hypothetisch maar steunen gezien de proefopzet wellicht op
volatielen.
5.1.2. Snelheid van keuze
Om na te gaan of er een invloed was van tarwerassen op de snelheid waarmee bladluizen
kiezen, werden er richtingscoëfficiënten berekend na 1u voor dezelfde tarwerassen-
combinaties. Dit werd niet gedaan voor verschillende rassencombinaties omdat hier de
preferentie mogelijks een invloed op de snelheid zou hebben. Voor S. avenae werd gevonden
dat er geen invloed was van de tarwerassen (kiemlingen) op de snelheid van hun keuze. M.
dirhodum koos echter significant sneller voor kiemlingen van Homeros en Lexus dan voor
Sahara. Hier hadden deze tarwerassen wel een invloed op de snelheid van hun keuze.
Gemiddeld gezien kiest M. dirhodum 1,4 keer sneller dan S. avenae bij de keuze tussen
dezelfde tarwekiemling. Er is dus een intrinsiek verschil tussen bladluissoorten in snelheid
van hun keuze bij kiemlingen. Wanneer het experiment van de preferentie tussen tarwerassen
op kiemlingen vergeleken werd met het experiment met de aren, koos S. avenae significant
sneller (8x) voor aren dan voor kiemlingen (zelfde tarwerassen met elkaar gecombineerd,
namelijk Lexus en Homeros). Volgens Poehling et al. (2007) komt S. avenae vooral voor op
tarwearen, terwijl M. dirhodum bladeren verkiest. Dit is echter louter preferentieel gezien,
maar hiermee zou misschien een link kunnen gelegd worden met de hogere snelheid van
keuze van S. avenae voor aren.

49
5.1.3. Link preferentie – gevoeligheid voor aarfusarium
Er werd tevens getracht een link te leggen tussen de voorkeur voor bepaalde tarwerassen bij
beide bladluissoorten en de gevoeligheid van die tarwerassen voor aarfusarium. Volgens
Haesaert et al. (2009) is het tarweras Lexus gevoelig, Homeros matig gevoelig, Tulsa matig
tolerant en Sahara tolerant. Hoewel M. dirhodum het gevoelige tarweras Lexus verkoos en dus
zijn kans om een vector van de schimmel te zijn groter was, zou dit in realiteit waarschijnlijk
zelden voorkomen. M. dirhodum verkiest namelijk de bladeren van tarwe boven de aren
(Poehling et al., 2007). Deze laatste worden door F. graminearum geïnfecteerd. S. avenae
verkoos het matig tolerante tarweras Tulsa. Ook hier kon geen link gelegd worden tussen de
voorkeur van de bladluissoort en de gevoeligheid van Tulsa voor F. graminearum, daar Tulsa
matig tolerant is. Echter moet opgemerkt worden dat hoewel in veldonderzoek van Audenaert
et al. (2009) Tulsa zeer weinig symptomen vertoont en dus resistent lijkt, er toch hoge
concentraties van DON in dit tarweras werden teruggevonden. Dit veldonderzoek werd
uitgevoerd op 7 verschillende veldproeflocaties waarbij in 3 daarvan Tulsa behoorde tot de
top drie van de tarwerassen met de hoogste DONconcentraties. Hoewel in deze proefopzet
niet gewerkt werd met DON, blijkt S. avenae zich aangetrokken te voelen tot een tarweras die
hoge concentraties van DON kan bevatten.

50
5.2. Aantrekking van S. avenae tot DON
5.2.1. Preferentietesten met behandelde aren
Reeds bij de preferentietesten met behandelde aren kwam de preferentie van S. avenae voor
aren geïnfecteerd met DON naar voor. Hierbij werden er geen significante verschillen tussen
verschillende behandelingen teruggevonden, maar dit kon te wijten zijn aan het beperkt aantal
herhalingen. Bij de keuze tussen CON-DON en FUS-DON kozen er meer bladluizen voor
DON dan voor de andere behandelingen. Door het beperkt aantal herhalingen werden er
echter geen significante verschillen gevonden. Bladluizen kozen tevens 1,3 keer sneller voor
de combinatie DON-DON dan voor FUS-FUS, echter zonder significant verschil. Indien de
proef 14 dagen duurde, werden er 4 keer meer bladluizen teruggevonden op de aren
behandeld met DON dan op de controle-aren. Opnieuw bleken bladluizen zich aangetrokken
te voelen tot DON.
Kleurvariatie tussen aren met verschillende behandelingen als reden voor deze aantrekking
kon uitgesloten worden daar de infectie op het moment van de keuze nog niet groot genoeg
was. Een mogelijke verklaring voor de aantrekking van S. avenae tot DON geïnfecteerde aren
zijn aggregatieferomonen. Hierbij kan echter de opmerking gemaakt worden dat een groot
deel van de bladluizen (70% bij CON-DON en 73% bij FUS-DON) reeds gekozen had na 1u.
Hoewel bladluizen in deze tijdspanne mogelijks wel al aggregatieferomonen produceren, kan
in vraag gesteld worden of de hoeveelheid hiervan voldoende is om anderen aan te trekken.
Het deel dat echter pas na 1u koos, kon zich eventueel wel aangetrokken voelen tot
aggregatieferomonen.
Een andere verklaring kan zijn dat de plant volatielen zoals MeJA en ET produceert tijdens
zijn afweermechanisme tegen DON en de bladluizen zich door deze volatielen aangetrokken
voelen. Nog een andere verklaring voor de aantrekking van S. avenae tot DON zijn de
volatiele componenten in DON. Mogelijks konden deze volatiele componenten in de
preferentietesten met artificieel dieet niet voldoende doorheen de parafilm dringen en door de
bladluizen gedetecteerd worden.
DON behoort tot de B-groep van de trichothecenen. Dit zijn tricyclische sesquiterpenen
(Doohan et al., 2003). Sesquiterpenen komen ook voor bij de afweerrespons van planten
tegen herbivoren. Vraat aan bladeren stimuleert namelijk de emissie van volatiele
componenten. Deze kunnen herbivoren afstoten maar ook natuurlijke vijanden van de
herbivoren aantrekken. De twee meest voorkomende bestanddelen in deze volatiele mengsels
zijn terpenen (C10 monoterpenen en C15 sesquiterpenen) en green leaf volatielen (o.a. C6
aldehyden). In de groep van de terpenen bestaat het sesquiterpeen (E)-β-farnesene (EBF).
Deze component is ook een algemeen voorkomend alarmferomoon bij bladluizen. Het wordt
vrijgesteld door bladluizen wanneer ze door natuurlijke vijanden worden aangevallen en zorgt
ervoor dat andere bladluizen in de omgeving stoppen met zich te voeden en wegvluchten
(Unsicker et al., 2009). A. thaliana, gemodificeerd om EBF te produceren, had een repellente
werking op de bladluissoort Myzus persicae in vergelijking met de niet-emitterende
controlelijnen. Het gedrag van de bladluizen werd hierbij 15 minuten lang gevolgd (Beale et
al., 2006). Kunert et al. (2010) vonden echter geen direct effect van EBF op deze

51
bladluissoort. EBF kon de (niet) gevleugelde bladluizen er niet significant van weerhouden de
planten te koloniseren. Dit werd verklaard doordat A. thaliana EBF continu vrijstelde en
bladluizen hieraan gewend raakten. Ook de Vos et al. (2010) onderzochten het gedrag van de
bladluizen indien ze continu werden blootgesteld aan EBF. Na 3 generaties trad er gewenning
op. Ze voelden zich niet meer afgestoten door EBF. Wanneer diezelfde bladluizen niet meer
blootgesteld werden, trad na 3 generaties terug gevoeligheid op. Deze gedragsverandering
werd niet veroorzaakt door een genetische mutatie maar door desensibilisatie in de
transcriptionele respons op EBF (de Vos et al., 2010). In volatiele mengels zijn EBF en
andere sesquiterpenen niet steeds even repellent. Zo bevat de olie van Hemizygia petiolata
Ashby (Lamiaceae) in minder mate ook nog germacreen. Hiertoe voelt S. avenae zich zeer
sterk afgestoten, terwijl deze zich slechts matig afgestoten voelt door EBF (Bruce et al.,
2005).
Hoewel in dit onderzoek S. avenae zich aangetrokken voelt tot het sesquiterpeen DON, blijkt
uit andere studies dat sesquiterpenen die vrijgesteld worden bij de afweerrespons van planten
een (matig) repellente werking hebben.
5.2.2. Preferentietesten met artificieel dieet met DON
Bij de preferentietesten met artificieel dieet trad een gelijkaardig fenomeen op. Ook hier
hadden de bladluizen een voorkeur voor het medium aangelengd met DON. Significante
verschillen werden teruggevonden tussen de controle en 1 mg/l DON en tussen 0,25 mg/l en 1
mg/l DON. De P-waarde was kleiner voor de eerste combinatie dan voor de tweede, wat erop
duidde dat het significant verschil groter werd naarmate het verschil in concentratie groter
werd. De keuze van de bladluizen bleek dus dosisafhankelijk te zijn. De reden waarom in dit
experiment de bladluizen voor DON kozen was onduidelijk. Indien 20 bladluizen in aparte
systemen konden kiezen tussen de controle en 1 mg/l DON, werd er geen significant verschil
teruggevonden. Hieruit kon dus geconcludeerd worden dat ze zich niet aangetrokken voelden
tot eventuele volatiele componenten in DON. Deze conclusie was echter enkel geldig als met
zekerheid kon worden aangetoond dat volatiele componenten doorheen de parafilm konden
dringen. In Fig. 24 was te zien hoe er na 24u meer bladluizen aan de controle zogen dan aan 1
mg/l DON (statistisch gezien was er echter geen significant verschil). Na 72u was de
verhouding 50-50 geworden. Enkele bladluizen die na 24u de controle verkozen, werden na
72u op het artificieel dieet met DON terug gevonden. Hoewel er eerst gedacht werd dat dit te
maken kon hebben met het feit dat hun overleving of reproductie op artificieel medium
aangelengd met DON bevoordeeld kon zijn, weerlegde het experiment rond de reproductie
van S. avenae op artificieel dieet deze hypothese: de mortaliteit van de nimfen op 1 mg/l DON
was 80% en er werd ook geen reproductievoordeel gevonden op 1 mg/l DON ten opzichte van
de controle.
Ook in ander onderzoek wordt de preferentie van insecten voor schimmels onderzocht. El-
Hamalawi (2007) onderzocht de aantrekking van oevervliegen tot verschillende substraten
zoals verschillende pathogene schimmels (Verticillium dahliae, Fusarium oxysporum f.sp.
basilici en Thielaviopsis basicola) opgegroeid op PDA medium, op algen en op een
nutriëntenmedium van PDA en V8 broth. De insecten vlogen bij aanvang van het experiment

52
doelloos rond. Het duurde minstens twee uur alvorens ze in een bepaalde richting vlogen en
zich settelden op een bepaald substraat. De beste attractant voor oevervliegen waren de algen
(waarschijnlijk omdat dit hun primaire voedselbron was), gevolgd door de schimmelculturen
op medium (waar de insecten ook konden van eten en beschouwden als een individuele bron
van nutriënten). Echter tussen deze schimmelculturen werden er geen significante verschillen
in aantrekking teruggevonden.
Vrouwtjes van de stengelboorder Apion onopordi herkenden geïnfecteerde akkerdistels en
konden ook de verschillende stadia van de roest Puccinia punctiformis onderscheiden.
Hierdoor prefereerden ze de jongere stadia met de pycnia (puisten waarin ongeslachtelijke
sporen zitten en die volatielen verspreiden om insecten te lokken zodat deze de sporen naar
het monokaryotische ontvangstmycelium kunnen verplaatsen) en vermeden ze de oudere
stadia met de urediosporen. Dit kon te verklaren zijn doordat planten geïnfecteerd met deze
oudere stadia binnen een maand afsterven en dus niet lang genoeg overleven om de volledige
ontwikkeling van de kever te ondersteunen (Friedli & Bacher, 2001).
Net zoals de stengelboorder A. onopordi waardplanten geïnfecteerd met P. punctiformis
herkent, lijkt S. avenae in staat te zijn om de schimmel F. graminearum, of toch het
mycotoxine DON dat deze produceert, te herkennen. Hoewel S. avenae zich aangetrokken
voelt tot DON, kan niet met zekerheid gezegd worden dat dit net zoals bij de stengelboorder
te maken heeft met volatiele componenten van de schimmel. Zoals eerder vermeld was de
voorkeur van S. avenae voor DON dosisafhankelijk. Indien de aantrekking van S. avenae tot
volatiele componenten in DON uitgesloten kon worden, zouden mogelijks
aggregatieferomonen van de bladluizen een reden zijn waarom er meer bladluizen voor
hogere concentraties van DON kozen. Echter de reden hiervoor is opnieuw onbekend.
5.2.3. Overleving A. pisum en S. avenae op artificieel dieet met DON
Uit de resultaten bleek dat de mortaliteit van nimfale A. pisum na 3 dagen steeg naarmate de
concentraties van DON in hun artificieel dieet stegen. 0,5 mg/l en 1 mg/l DON zorgden voor
significante verschillen in overleving in vergelijking met lagere concentraties van DON. Bij
nimfale S. avenae was dit niet het geval. Deze had een significante betere overleving bij de
twee hoogste concentraties van DON in vergelijking met A. pisum. Er kan dus gesteld worden
dat A. pisum veel gevoeliger was aan het mycotoxine dan S. avenae.
Toxiciteit van trichothecenen is te wijten aan hun vermogen om de eiwitsynthese te inhiberen.
Ze binden op de 60S subunit van eukaryotische ribosomen en tasten het peptidyltransferase
aan. Het mycotoxine DON inhibeert de elongatie van de peptideketen (Goyarts et al., 2006).
Een hypothese voor de lagere gevoeligheid van S. avenae voor DON was dat er door co-
evolutie van deze bladluissoort met F. graminearum enige gewenning aan DON is ontstaan.
Het zou kunnen dat gedurende deze evolutie het peptidyltransferase gemodificeerd is
waardoor het minder gevoelig is aan aantasting door DON. Uiteraard blijft dit puur
hypothetisch. Nucleotidensequenties van dit enzym blijken na gebruik te maken van NCBI
(National Center for Biotechnology Information) enkel gekend te zijn voor de bladluissoort A.
pisum. Er kon dus geen vergelijking gemaakt worden tussen verschillende bladluissoorten.

53
5.2.4. Reproductie S. avenae op artificieel dieet met DON
Wanneer de overleving van S. avenae op concentraties 0 mg/l, 0,25 mg/l en 1 mg/l DON voor
langere tijd onderzocht werd, overleefden slechts 20% van de nimfen lang genoeg om adult te
worden op 1 mg/l DON. Bij de controle en 0,25 mg/l was dit 70%. Er werden geen
significante verschillen in nimfale levensduur op de controle en 0,25 mg/l DON gevonden,
wat er op duidt dat ze geen voordeel konden halen uit het al dan niet voeden op DON. In
beide gevallen ontwikkelden ze even snel. Ook tussen de levensduur van de adulten werd er
wegens grote variatie geen significante verschillen tussen de 3 behandelingen teruggevonden.
Wanneer de reproductie werd geobserveerd op de verschillende behandelingen werd
gevonden dat de intrinsieke groeisnelheid rm beduidend lager lag voor 1 mg/l DON dan voor
de lagere concentraties. De netto vermenigvuldigingssnelheid R0 lag eveneens veel lager. Ook
de finale groeisnelheid λ lag dus lager voor 1 mg/l DON. De gemiddelde generatietijd T lag
significant hoger omdat de pre-ovipositieperiode hoger lag, daar de moederbladluizen een
zeer lange nimfale periode doormaakten. Uiteraard lag de verdubbelingstijd DT ook hoger
voor 1 mg/l DON omdat de bladluizen minder reproduceerden.
Er moet echter opgemerkt worden dat in de natuur de bladluizen die zich voeden op
geïnfecteerde aren nooit aan zo‟n hoge concentratie DON (1 mg/l) blootgesteld worden, tenzij
bij zeer hevige infecties. Er blijkt namelijk dat gedurende 2002-2010 slechts 0,59% tarwearen
geïnfecteerd waren met 0,9 tot 1 mg/l DON (S. Landschoot, persoonlijke communicatie). De
lagere overlevings- en reproductieparameters konden dus toxicologisch te verklaren zijn.
36,87% van de aren bevatten 0,1 tot 0,3 mg/l DON. Deze concentraties komen in realiteit dus
veel frequenter voor. rm lag iets hoger voor 0,25 mg/l dan voor de controle (echter significant
verschillend). Dit duidde op een heel klein reproductievoordeel. In de preferentietesten met
behandelde aren op lange termijn zaten er na 14 dagen 6,9 ± 1,2 nimfen per adult op de aren
behandeld met DON en 6,8 ± 2,2 nimfen per adult op de controle-aren. Ook hier kwam een
heel klein reproductievoordeel terug, echter zonder significant verschil. λ lag iets hoger, T en
DT iets lager, allen met significante verschillen, wat dus op iets betere overlevings- en
vruchtbaarheidsparameters duidde voor 0,25 mg/l DON dan voor de controle.
Zoals eerder vermeld werd M. persicae die leefde op EBF-producerende A. thaliana na 3
generaties ongevoelig voor EBF. Deze EBF-ongevoelige bladluizen produceerden meer
nakomelingen in vergelijking met de bladluizen die gevoelig waren voor EBF. Echter hadden
EBF-gevoelige bladluizen een hogere overlevingskans in de nabijheid van predatoren daar ze
zich afgestoten voelden door de planten die predatoren aantrokken (de Vos et al., 2010). In
het experiment van Kunert et al. (2010) werd geen significant verschil teruggevonden in de
reproductie van M. persicae tussen EBF-producerende A. thaliana en de controleplanten.
Uit dit onderzoek kon dus geconcludeerd worden dat S. avenae op 0,25 mg/l DON een klein
maar toch significant reproductievoordeel heeft. In de literatuur werd gevonden dat EBF-
ongevoelige M. persicae meer nakomelingen konden produceren. Echter kan de link met
gewenning van S. avenae aan DON, zoals M. persicae gewend kon worden aan EBF, niet met
zekerheid gelegd worden, daar slechts één generatie op DON werd opgegroeid en dit
mogelijks te weinig was om gewenning te laten optreden. De reden waarom er een
reproductievoordeel optrad bij bladluizen op 0,25 mg/l DON bleef dus onduidelijk.

54
Overlevings- en vruchtbaarheidsparameters zijn goede indicatoren voor populatiegroei onder
een gegeven set van condities. Er kon aangetoond worden dat de concentratie van DON in het
artificiële dieet waaraan de bladluizen zich konden voedden, een invloed hadden op deze
parameters. Hoewel de bladluizen op elke behandeling nimfen konden produceren, werden er
toch significante verschillen in reproductie teruggevonden.
Ook andere auteurs maken van deze parameters gebruik. Obopile & Ositile (2010) vonden
aan de hand van deze parameters verschillen in overleving en reproductie van de bladluissoort
Aphis craccivora op vijf verschillende variëteiten cowpea (Vigna unguiculata). Deze stelden
dat een kortere pre-ovipositieperiode en een hogere reproductiecapaciteit van een insect op
een waardplant de geschiktheid van een waardplant reflecteert. Davis & Radcliffe (2008a)
vonden dat de bladluissoort M. persicae de hoogste reproductiecapaciteit had op wintertarwe,
gevolgd door rogge, gerst en haver. Dus ook hier werden significante verschillen tussen de
parameters teruggevonden. Guldemond et al. (1998) stelden dat het ontwikkelingsstadium van
de plant een effect heeft op de reproductiecapaciteit van bepaalde graanbladluis-plant
interacties. Afhankelijk van het graangewas, presteerde de vogelkersluis R. padi het beste op
zaailingen van gerst, op de stadia van stengelverlenging tot aarvorming bij tarwe en
uitstoeling tot aarvorming bij haver.
De fitness van insecten op planten geïnfecteerd met schimmels wordt eveneens onderzocht.
Friedli & Bacher (2001) onderzochten de prestaties van de stengelboorder A. onopordi op
akkerdistel geïnfecteerd met de roest P. punctiformis. De adulte insecten die ontstaan waren
uit eitjes afgelegd op geïnfecteerde akkerdistel waren significant zwaarder dan adulten op
gezonde planten. De reden hiervoor bleef onbekend maar de zwaardere kevers produceerden
meer en grotere eitjes, waardoor zwaarder zijn dus een fitnessvoordeel betekende.

55
5.3. Verspreiding F. graminearum door S. avenae
5.3.1. Orale opname van macroconidia door S. avenae
Zoals in de resultaten reeds vermeld was er geen orale opname van macroconidia als de
bladluizen zich voedden op artificieel dieet met sporen in. De reden hiervoor was dat de
sporen van F. graminearum mogelijks te groot waren om doorheen de opening van het stilet
te kunnen. Macroconidia hebben een grootte van 41-60 x 4-5,5 µm. Microconidia zijn
afwezig (Samson et al., 2004). De stiletafmetingen gegeven in de resultaten zijn uitwendig
gemeten. Stiletten hebben echter inwendig nog een zuigbuis zitten waarlangs voedsel en
eventuele sporen worden opgezogen. Van deze buis is de grootte van de opening bij S. avenae
niet gekend. Dit is echter wel gekend bij de bladluissoort M. persicae. De opening van de
zuigbuis bij deze soort is 0,7 µm (Katis et al., 2007) en dus niet groot genoeg om sporen van
F. graminearum op te nemen.
In tegenstelling tot de bladluissoort S. avenae, zijn oevervliegen wel in staat om propagules
van schimmels inwendig op te nemen en terug uit te scheiden. El-Hamalawi (2007) beweerde
dat er minstens 10, 20 en 20 minuten blootstelling van oevervliegen aan de schimmelculturen
nodig was om leefbare propagules van respectievelijk V. dahliae, F. oxysporum f.sp. basilici
en T. basicola terug te vinden in de uitscheidingen van de insecten. Gemiddeld 8,3% (na 10
minuten blootstelling) en 34,3% (na 20 minuten) van de uitscheidingen bevatten propagules
van V. dahliae. Voor F. oxysporum f.sp. basilici en T. basicola was dit respectievelijk 8,1 en
8% na 20 minuten blootstelling. De leefbaarheid van de propagules werd niet beïnvloed door
de acquisitietijd. El-Hamalawi & Stanghellini (2005) beweerden dat oevervliegen ook in staat
zijn om sporen van F. avenaceum op te nemen daar de schimmelkolonies teruggevonden op
het Fusarium-selectieve medium (Komada‟s medium) afkomstig waren van insect-
uitscheidingen die macroconidia bevatten. In een ander onderzoek werd de inwendige opname
van P. aphanidermatum door oevervliegen onderzocht op komkommerwortels. Bij 97% van
de eerste en tweede larvale stadia, 20% van het derde larvale stadium en 10% van de adulten
werden oösporen in hun spijsverteringsstelsel teruggevonden. In adulte oevervliegen konden 1
tot 25 oösporen geteld worden, bij de larven was dit 25 tot 680 oösporen. Of de kiemkracht
van deze sporen veranderde gedurende de inwendige opname was niet geweten, echter was
30% van de sporen uit het spijsverteringsstelsel van de oevervliegen kiemkrachtig
(Stanghellini & Goldberg, 1990).
5.3.2. Uitwendige acquisitie en verspreiding F. graminearum door S. avenae
Er werd nagegaan wat de proportie bladluizen was die inoculum (sporen en mycelium) droeg
op hun lichamen. Slechts 26% van de bladluizen was drager van de schimmel als ze uit een
sporulerende plaat kwamen. Zo‟n sporulerende plaat kon echter beschouwd worden als een
worst-case scenario in vergelijking met een besmette aar. Dit laag percentage kon te wijten
zijn aan het feit dat S. avenae weinig beharing heeft, wat de acquisitie van inoculum negatief
kan beïnvloeden en omdat de bladluizen meestal de niet-besmette zones in de petriplaat met
sporulerende schimmel opzochten. De acquisitie van inoculum door de bladluis was ook zeer
variabel. Deze grote variatie kon te wijten zijn aan het feit dat niet alle bladluizen evenveel

56
rondliepen en dus niet evenveel kans hadden om inoculum op te nemen en omdat de opname
van inoculum afhankelijk was van de sporulatieverdeling en -intensiteit in een plaat. Over het
algemeen kan gesteld worden dat S. avenae slechts in beperkte mate een vector is van de
schimmel.
Na 15 minuten rondlopen op de wateragar droegen 28% ± 20% bladluizen inoculum. Na 1,5u
was dit 32% ± 4% en na 24u was dit 20% ± 4% (Fig. 31). Het gemiddelde over alle
tijdstippen heen is 27%. Ook hier werd de proportie bladluizen die inoculum droeg nadat ze
uit een sporulerende plaat kropen (26% ± 4%) teruggevonden. Over de 3 tijdstippen heen
bleven de bladluizen drager van inoculum. Er droegen proportioneel gezien ook meer
bladluizen inoculum op een kiemling met inoculum dan op een kiemling zonder inoculum.
Bladluizen raakten echter het inoculum willekeurig kwijt en er was niet geweten hoeveel
inoculum de bladluizen droegen.
Naast de interne opname en uitscheiding, werden volgens El-Hamalawi (2007) propagules
van V. dahliae, F. oxysporum f.sp. basilici en T. basicola ook uitwendig aan het lichaam van
oevervliegen aangetroffen, vooral aan de haren op de poten en aan de vleugels. El-Hamalawi
& Stanghellini (2005) beweerden dat oevervliegen in staat zijn om sporen van F. avenaceum
uitwendig aan het lichaam mee te dragen daar de schimmelkolonies teruggevonden op het
Fusarium selectieve medium niet alleen afkomstig waren van insectuitscheidingen die
macroconidia bevatten, maar ook van solitaire of geclusterde macroconidia die uitwendig aan
de insectenlichamen hingen. Bij champignonmuggen en motmuggen was dit ook het geval.
Deze drie insecten konden in een tijdsperiode van vier dagen de macroconidia transporteren
naar minstens 45% van de lisianthusplanten. De hoeveelheid inoculum verplaatst door
insecten bleek genoeg te zijn om deze planten te infecteren en ziek te maken (El-Hamalawi &
Stanghellini, 2005). De wijfjes van stengelboorder A. onopordi zorgden tijdens de ovipositie
ervoor dat akkerdistel werd geïnfecteerd met de roest P. punctiformis Friedli & Bacher,
2001). Ook de zwarte bonenluis, de grote distelluis en de schildpadtor konden, als ze zich
gevoed hadden met geïnfecteerde akkerdistelplanten, gezonde planten besmetten met sporen
die aan hun lichaam en poten hingen (Kluth et al., 2002).

57
6. Conclusie
In deze thesis werd de tritrofe interactie tussen de tarweplant, de bladluissoort S. avenae en de
schimmel F. graminearum onderzocht.
De groei van een tarweplant is een dynamisch fenomeen waar S. avenae zich aan aanpast.
Tijdens de vegetatieve stadia van tarwe bevindt deze bladluissoort zich op de bladeren. Eens
de aarvorming begint, migreren de bladluizen zich naar de aren. In het aarstadium kan de
plant tevens geïnfecteerd worden door F. graminearum. Deze schimmel produceert gedurende
zijn infectie van de aren het mycotoxine DON. S. avenae voelt zich tot deze geïnfecteerde
aren aangetrokken en maakt hiervoor ook het snelst zijn keuze (Fig. 34). Wat het mechanisme
achter deze aantrekkingskracht is, is niet geweten. Maar op de vraag „waarom‟ deze
bladluissoort de geïnfecteerde aren prefereert, kunnen wel enkele hypotheses voorgesteld
worden.
Fig. 34: Schematische weergave van de snelheid van keuze voor S. avenae. De snelheid van keuze is
weergegeven door de dikte van de lijnen. Er kan duidelijk opgemerkt worden dat er voor de aren steeds sneller
gekozen wordt dan voor de kiemlingen. Indien deze bladluissoort tussen aren moet kiezen, koos deze sneller
voor de met DON geïnfecteerde aren dan voor de gezonde aren (CON). Voor de gezonde aren werd sneller
gekozen dan voor de aren geïnfecteerd met sporen (FUS) en waarbij myceliumgroei al matig waarneembaar was.
Op het moment dat de bladluizen hun keuze maakten, was de symptoomontwikkeling nog maar net gestart.
Mogelijks is initiële infectie van aren een reden waarom deze aren geprefereerd worden door
S. avenae, daar deze weten dat op dat moment de aren in bloei staan en dus een verhoogde
sapstroom van de plant naar deze plaats gaat. Op die plaats kunnen ze dus voordeel hebben
om zich te voeden. Eveneens zorgt DON er in het begin van een aarinfectie ervoor dat het
weefsel zachter wordt gemaakt (zodat de schimmel zich makkelijk in de aar kan verspreiden).
Ook dan hebben de bladluizen een voordeel. Het is voor hen dan makkelijker om zich op de
aar te voeden en ze verkiezen deze met DON geïnfecteerde aren dan ook boven andere.

58
Deze voordelen zijn echter van korte duur. Wanneer de infectie erger wordt en de schimmel
de aar verder koloniseert, verandert dat voordeel in een nadeel. Waar de bladluizen op korte
termijn een voordeel van DON ondervonden, voelen ze op langere termijn een negatieve
invloed van de infectie op hun levensprocessen. Hun mortaliteit stijgt door te hoge
concentraties van DON. Ook hun reproductie gaat door teveel DON erop achteruit. Vooral
hoge concentraties van 1 mg/l DON zijn toxisch. Echter komen zo‟n hoge concentraties in de
realiteit zelden voor.
Omwille van dit nadelig effect, dat ontstaat naarmate de infectie erger wordt, is de migratie
van een bladluis een dynamisch fenomeen. Omdat een zwaar geïnfecteerde aar niet meer
voldoet aan de eisen van een bladluis (de concentraties van DON liggen zo hoog dat het
toxisch wordt, de sapstroom in de aar ligt stil en het mycelium werkt de migratie van de
bladluizen tegen) zullen de bladluizen, nog voor deze nadelige situatie zich voordoet, weg
migreren naar minder aangetaste aren of waardplanten buiten het veld. Waarschijnlijk
ontstaan dan gevleugelde bladluizen.
Waar concentraties van 1 mg/l DON in realiteit zelden voorkomen, komt de concentratie 0,25
mg/l DON wel frequenter voor. Hiermee zullen de bladluizen wel in contact komen, als ze
zich voeden op een geïnfecteerde aar. Bij deze concentratie werd in vergelijking met
concentratie 0 mg/l DON een klein reproductievoordeel teruggevonden. Dit was, in tegen-
stelling tot 1 mg/l DON, niet toxicologisch te verklaren. Een hypothese hiervoor is co-evolutie
van S. avenae en F. graminearum die tot enige gewenning aan DON leidde. Deze stelling kan
gestaafd worden daar DON voor de bladluissoort A. pisum veel toxischer is dan voor S.
avenae. Aangezien A. pisum op peulvruchten leeft, kwam deze gedurende zijn evolutie niet in
contact met aren geïnfecteerd met F. graminearum en dus ook niet met DON. Door de co-
evolutie van S. avenae samen met F. graminearum hebben deze bladluizen leren omgaan met
de toxiciteit van DON en hebben ze de voordelen van DON in aren (meer sapstroom door
bloei en zachtere weefsels) achterhaald.
Wanneer getracht wordt een link te leggen tussen de voorkeur van S. avenae voor met DON
geïnfecteerde aren en zijn rol als vector van F. graminearum, kan volgende conclusie
getrokken worden: wanneer de infectie van een aar zover gevorderd is dat er myceliumgroei
ontstaat, migreren de bladluizen naar de gezonde aren. Hierbij zijn ze in staat om mycelium te
verplaatsen. Echter zijn slechts 26% van alle bladluizen een vector (en dit in een worst-case
scenario), daar hun lichaam weinig behaard is. Ze zijn dus niet zo‟n goede drager van
mycelium of sporen van de schimmel. Toch mogen bladluizen als vector van F. graminearum
niet onderschat worden. Eén leefbare spore of één myceliumstuk kan aanleiding geven tot
infectie. Hoewel er dus een laag percentage bladluizen drager is, zullen deze zeker voor
infectie van gezonde aren zorgen en mogen deze bladluispopulaties in monitoringssystemen
van F. graminearum niet vergeten worden. Verder onderzoek moet echter meer duidelijkheid
brengen omtrent hun impact in de verspreiding van plantpathogene schimmels.

59
7. Samenvatting en toekomstperspectieven
Deze thesis onderzoekt de tritrofe interactie tussen de schimmel, Fusarium graminearum, en
de grote graanluis, Sitobion avenae, die beide op wintertarwearen voorkomen en deze
beschadigen. Afzonderlijk veroorzaken ze grote opbrengst- en kwaliteitsverliezen van tarwe
maar hoe deze beschadigers van tarwe met elkaar interageren, is nog niet geweten. Concreet
werd er nagegaan of S. avenae een vector kon zijn van F. graminearum.
Allereerst werd onderzocht welke tarwerassen S. avenae prefereert. In deze proeven werd ook
een vergelijking gemaakt met een andere schadelijke bladluissoort, de roosgrasluis,
Metopolophium dirhodum. Er werd gevonden dat deze twee bladluissoorten een intrinsiek
verschil in preferentie van tarwerassen hebben en ook de snelheid waarmee ze een keuze
maken, is verschillend. Zo prefereert M. dirhodum het tarweras Lexus boven Sahara maar
prefereert S. avenae Tulsa boven Lexus en Homeros. Daarnaast is M. dirhodum ook sneller
dan S. avenae bij de keuze tussen kiemlingen, terwijl S. avenae sneller een beslissing maakt
als deze kan kiezen tussen aren, dan als deze moet kiezen tussen kiemlingen. Een link tussen
geprefereerde tarwevariëteiten en gevoeligheid van deze variëteiten voor een infectie van de
schimmel kon niet aangetoond worden.
Er werd ook nagegaan of S. avenae zich aangetrokken voelt tot de schimmel. Uit de resultaten
bleek dat deze bladluizen zich niet aangetrokken voelden tot de schimmel zelf, maar wel tot
het mycotoxine dat deze produceerde. Zowel op aarniveau als op artificieel niveau werd de
stelling dat deze bladluissoort zich aangetrokken voelt tot deoxynivalenol (DON), bevestigd.
S. avenae voelde zich aangetrokken tot aren geïnfecteerd met DON. Ook hun
beslissingssnelheid was groter als ze konden kiezen tussen met DON geïnfecteerde aren dan
tussen aren geïnfecteerd met sporen. Op lange termijn kwamen ook meer bladluizen voor op
de aren geïnfecteerd met DON dan op de controle-aren. Op artificieel niveau prefereerden ze
kunstmatig vloeibare voeding aangelengd met DON. De stelling dat ze zich tot DON
aangetrokken voelen, werd nogmaals bevestigd doordat gevonden werd dat aantrekking
afhankelijk was van de concentratie van DON in deze voeding. Hiervoor werd een klein
voordeel gevonden in reproductie op een lage concentratie DON, gebruik makende van
overlevings- en reproductieparameters volgens de Jackknifemethode. Een hoge concentratie
DON benadeelde echter de reproductie. Overleving van S. avenae op hoge concentraties van
DON was wel beduidend hoger in vergelijking met de bladluissoort Acyrthosiphon pisum.
Tot slot was er nog het onderzoek naar S. avenae als vector van F. graminearum. Dit bestond
uit twee delen. In een eerste deel werd nagegaan of deze bladluissoort in staat was om sporen
van de schimmel op te zuigen en eventueel uit te scheiden. Hierbij werd teruggevonden dat
deze bladluis niet in staat is om de sporen op te zuigen, mogelijks doordat de sporen te groot
zijn om doorheen het stilet van de bladluis te raken. Een tweede deel van dit onderzoek ging
na of S. avenae de schimmel uitwendig aan zijn lichaam kon dragen en verplaatsen. Deze
bladluissoort is in staat om zowel sporen als mycelium te dragen en te verplaatsen maar
slechts een beperkt percentage is een vector. Er werd gevonden dat in een worst-case scenario
slechts een kwart van de bladluizen een vector is van F. graminearum. Toch is dit een niet te
onderschatten hoeveelheid, daar slechts 1 leefbare spore of stuk mycelium kan aanleiding
geven tot aarinfectie.

60
De stelling, „Aantrekking van S. avenae tot DON‟, biedt mogelijkheid voor verder onderzoek.
Om deze stelling kracht bij te zetten, kan gebruik gemaakt worden van de DON Knock-Out
variant van F. graminearum om hiermee preferentietesten op kiemling- en aarniveau op te
zetten. Agressiviteitstesten met F. graminearum en zijn DON Knock-Out variant zijn
eveneens gewenst, daar de agressiviteit van een pathogeen een invloed kan hebben op de
aantrekking van bladluizen tot tarweplanten.
Om wat meer fundamenteel onderzoek te doen, kan er op moleculair niveau nagegaan worden
wat er in een aar gebeurt als deze geïnfecteerd raakt met de schimmel of met DON. Misschien
zijn volatiele componenten, geproduceerd door de tarweaar als reactie op de schimmel of zijn
mycotoxine, een mogelijke reden waarom S. avenae zich aangetrokken voelt tot DON.
Verder onderzoek naar interactie tussen S. avenae en F. graminearum kan gedaan worden op
aarniveau. Er kan nagegaan worden of aren geïnfecteerd met de schimmel of besmet met
DON een invloed hebben op de groei, ontwikkeling en reproductie van bladluizen.
Omgekeerd kan onderzocht worden of aren beschadigd door bladluizen het infectieproces van
F. graminearum beïnvloeden. Ook zou het nuttig zijn om het gewicht van de bladluizen op
verschillende concentraties van DON in artificieel medium na te gaan. Eveneens is het
interessant om de reden te achterhalen waarom S. avenae op lage concentraties van DON kan
overleven, terwijl A. pisum dat niet kan.
Enkele andere interessante pistes zijn de preferentie van S. avenae voor verschillende
tarwerassen op aarniveau nagaan en vergelijken met hun preferentie op kiemlingniveau. Ook
preferentietestjes tussen combinaties van aren, kiemlingen en topbladen kunnen opgezet
worden voor zowel S. avenae en M. dirhodum.
Om het luik S. avenae als vector van de schimmel verder te onderzoeken, kunnen
experimenten worden opgezet die nagaan in hoeverre het verplaatste inoculum voor infectie
van aren zorgt. Dit kan gedaan worden door bladluizen in de sporulerende schimmel te laten
lopen en daarna manueel over te zetten op aren. In een later experiment kan onderzocht
worden of bladluizen het inoculum van aar naar aar kunnen verplaatsen en of dit inoculum
dan nog steeds voor infectie kan zorgen. Een ander onderzoekluik omtrent bladluizen als
vector is werken met gevleugelde bladluizen. Hoe snel worden bladluizen gevleugeld op
geïnfecteerde aren en dragen deze gevleugelde vormen ook bij tot de verspreiding van
Fusarium spp.?

61
8. Summary
This thesis explores the tritrophic interaction between the fungus Fusarium graminearum and
the aphid species Sitobion avenae. Both occur on winter wheat spikes and damage
it. Individually they cause major yield and quality losses of wheat but how they interact is not
yet known. Specifically, this thesis examined whether S. avenae could be a vector of
F. graminearum.
First, the wheat varieties preferred by S. avenae and Metopolophium dirhodum were
examined. It was found that different aphid species differ in the intrinsic preference of wheat
varieties and also the speed with which they choose, is different. M. dirhodum prefers the
wheat cultivar Lexus over Sahara but prefers S. avenae Tulsa over Lexus and Homeros. In
addition, M. dirhodum chooses faster than S. avenae in the choice between seedlings, while
S. avenae makes a faster decision when it can choose between ears, than when it must choose
between seedlings. A link between the preferred wheat cultivars and sensitivity of this species
to infection with F. graminearum could not be demonstrated.
It was also examined whether S. avenae is attracted to fungus. The results showed that the
aphids are not attracted by the fungus itself, but to the mycotoxin produced by the
fungus. Both on ear level as on artificial liquid food, the argument that this aphid species is
attracted to the mycotoxin deoxynivalenol (DON) is confirmed. Their decision speed was
greater if they could choose between DON infected ears than between ears infected with
spores. In the long term there were more aphids on the ears infected with DON than on the
control ears. At artificial level they preferred the diet diluted with DON. The claim that they
are attracted to DON, was again confirmed because it was found that attraction was dependent
on the concentration of DON in their diet. There was a small benefit found in reproduction on
low concentrations of DON, using survival and reproductive parameters in the Jackknife
method. On high concentrations there was a significant lower reproduction. Survival of
S. avenae on high concentrations of DON was significantly higher compared with the aphid
species Acyrthosiphon pisum.
Finally there was the investigation of S. avenae as a vector of F. graminearum. This consisted
of two parts. The first part examined whether this aphid species was able to take up spores of
the fungus and excrete them. It was found that this aphid is unable to suck up the spores,
possibly because the spores are too large to go through the stylet of the aphids. A second part
of this study examined whether S. avenae could carry the fungus on the outside of his body
and spread it. This aphid species is capable of carrying and spreading both spores and
mycelium, but only a small percentage is a vector. It was found that in a worst-case scenario,
only one quarter of the aphids is a vector of F. graminearum. Yet this is not an inconsiderable
amount, since only one viable spore or one piece of mycelium can lead to ear infection.

62
9. Referenties
Athanassiou C., Palyvos N., Eliopoulos P., Papadoulis G. (2001). Distribution and migration of insects and mites
in flat storage containing wheat. Phytoparasitica, 29, 379-392.
Audenaert K., Callewaert E., Höfte M., De Saeger S., Haesaert G. (2010). Hydrogen peroxide induced by the
fungicide prothioconazole triggers deoxynivalenol (DON) production by Fusarium graminearum. BMC
Microbiology, 10, 112.
Audenaert K., Van Broeck R., Bekaert B., De Witte F., Heremans B., Messens K., Höfte M., Haesaert G. (2009).
Fusarium head blight (FHB) in Flanders: population diversity, inter-species associations and DON
contamination in commercial winter wheat varieties. European Journal of Plant Pathology, 125, 445-458.
Backhouse D., Burgess L. (2002).Climatic analysis of the distribution of Fusarium graminearum, F.
pseudograminearum and F. culmorum on cereals in Australia. Australasian Plant Pathology, 31, 321-327.
Bai G., Shaner G. (2004). Managment and resistance in wheat and barley to Fusarium Head Blight. Annual
Review of Phytopathology, 42, 135-161.
Bari R., Jones J. (2009). Role of plant hormones in plant defense responses. Plant Molecular Biology, 69, 473-
488.
Beale M., Birkett M., Bruce T., Chamberlain K., Field L., Huttly A., Martin J., Parker R., Phillips A., Pickett A.,
Prosser I., Shewry P., Smart L., Wadhams L., Woodcock C., Zhang Y. (2006). Aphid alarm pheromone
produced by transgenic plants affects aphid and parasitoid behavior. Proceedings of the National Academy of
Sciences, 27, 10509-10513.
Birch L. (1948). The intrinsic rate of natural increase of an insect population source. Journal of Animal Ecology,
17, 15-26.
Blackman R., Eastop V. (2007). Aphids as crop pests: taxonomic issues (Hfst 1, 30blz.). Boek van CAB
international.
Bostock R. (2005). Signal crosstalk and induced resistance: straddling the line between cost and benefit. Annual
Review of Phytopathology, 43, 545-580.
Bottalico A., Perrone G. (2002). Toxigenic Fusarium species and mycotoxins associated with head blight in
small-grain cereals in Europe. European Journal of Plant Pathology, 108, 611-624.
Braendle C., Davis G., Brisson J., Stern D. (2006). Wing dimorphism in aphids. Heredity, 97, 192-199.
Bruce T., Birkett M., Blande J., Hooper A., Martin J., Khambay B., Prosser I., Smart L., Wadhams L. (2005).
Response of economically important aphids to components of Hemizygia petiolata essential oil. Pest
Management Science, 61, 1115-1121.
Bushuk W. (1998). Wheat breeding for end-product use. Euphytica, 100, 137-145.
Castañeda L., Figueroa C., Bacigalupe R., Nespolo R. (2010). Effects of wing polymorphism, aphid genotype
and host plant chemistry on energy metabolism of the grain aphid, Sitobion avenae. Journal of Insect
Physiology, 56, 1920-1924.
Chan Y. 2005. Multinomial logistic regression. Singapore Medical Journal, 46, 259-269.
Conrath U. (2006). Systemic acquired resistance. Plant Signaling&Behavior, 1, 179-184.
Davis J., Radcliffe E. (2008a). Reproduction and feeding behavior of Mycus persicae on four cereals. Journal of
Economic Entomology, 101, 9-16.

63
Davis J., Radcliffe E. (2008b). Importance of an aphid species in vectoring a persistently transmitted potato
virus: Aphis glycines is a vector of potato leafroll virus. Plant Disease, 92, 1515-1523.
Dedryver C., Le Gallic J., Gauthier J., Simon J. (1998). Life cycle of the cereal aphid Sitobion avenae:
polymorphism and comparison of life history traits associated with sexuality. Ecological Entomology, 23, 123-
132.
Derycke V., Audenaert K., Haesaert G. (2007). Kunstmatige infectie om rassen te vergelijken.
Landbouw&Techniek, 18, 40.
Desjardins A., Jarosz A., Plattner R., Alexander N., Brown D., Jurgenson J. (2004). Patterns of trichothecene
production, genetic variability, and virulence to wheat of Fusarium graminearum from smallholder farms in
Nepal. Journal of Agricultural and Food Chemistry, 52, 6341-6346.
de Vos M., Cheng W., Summers H., Raguso R., Jander G. (2010). Alarm pheromone habituation in Myzus
persicae has fitness consequences and causes extensive gene expression changes. Proceedings of the National
Academy of Sciences, 107, 14673-14678.
Dicke M., van Poecke R., de Boer J. (2003). Inducible indirect defense of plants: from mechanisms to ecological
functions. Basic and Applied Ecology, 4, 27-42.
Doohan F., Brennan J., Cooke B. (2003). Influence of climatic factors on Fusarium species pathogenic to
cereals. European Journal of Plant Pathology, 109, 755-768.
El-Hamalawi Z. (2007). Attraction, acquisition, retention and spatiotemporal distribution of soilborne plant
pathogenic fungi by shore flies. Annals of Applied Biology, 152, 169-177.
El-Hamalawi Z., Menge J. (1996). The role of snails and ants in transmitting the avocado stem cancer pathogen,
Phytophthora citricola. Journal of the American Society for Horticultural Science, 121, 973-977.
El-Hamalawi Z., Stanghellini M. (2005). Disease development on lisianthus following aerial transmission of
Fusarium avenaceum by adult shore flies, fungus gnats, and moth flies. Plant Disease, 89, 619-623.
El-Wakeil N., Volkmar C., Sallam A. (2010). Jasmonic acid induces resistance to economically important insect
pests in winter wheat. Pest Managment Science, 66, 549-554.
Eleftherianos I., Vamvatsikos P., Ward D., Gravanis F. (2006). Changes in the levels of plant total phenols and
free amino acids induced by two cereal aphids and effects on aphid fecundity. Journal of Applied Entomology,
130, 15-19.
Febvay G., Delobel B., Rahbe Y. (1988). Influence of the amino acid balance on the improvement of an artificial
diet for a biotype of Acyrthosiphon pisum (Homoptera: Aphididae). Canadian Journal of Zoology, 66, 2449-
2453.
Ferreira R., Monteiro S., Freitas R., Santos C., Chen Z., Batista L., Duart J., Borges A., Teixeira A. (2006).
Fungal pathogens: the battle for plant infection. Critical Reviews in Plant Sciences, 25, 505-524.
Friedli J., Bacher S. (2001). Mutualistic interaction between a weevil and a rust fungus, two parasites of the
weed Cirsium arvense. Oecologia, 129, 571-576.
Gardiner
D., Kazan
K., Manners
J. (2009). Nutrient profiling reveals potent inducers of trichothecene
biosynthesis in Fusarium graminearum. Fungal Genetics and Biology, 46, 604-613.
Glazebrook J. (2005). Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annual
Review of Phytopathology, 43, 205-227.
Goggin F. (2007). Plant-aphid interactions: molecular and ecological perspectives. Current Opinion in Plant
Biology, 10, 399-408.

64
Goswami R., Kistler H. (2004). Heading for disaster: Fusarium graminearum on cereal crops. Molecular Plant
Pathology, 5, 515-525.
Goyarts T., Grove N., Dänicke S. (2006). Effects of the Fusarium toxin deoxynivalenol from naturally
contaminated wheat given subchronically or as one single dose on the in vivo protein synthesis of peripheral
blood lymphocytes and plasma proteins in the pig. Food and Chemical Toxicology, 44, 1953-1965.
Groot S. (2003). Fusarium in zaaizaad tarwe: oplossing voor fusarium-probleem pas op termijn beschikbaar.
Ekoland, 2, 28-29.
Guldemond J., van den Brink W., den Belder E. (1998). Methods of assessing population increase in aphids and
the effect of growth stage of the host plant on population growth rates. Entomologia Experimentalis et Applicata,
86, 163-173.
Haesaert G., Derycke V., Heremans B., Audenaert K., Wittouck D., Willaert L. (2009). Granen oogst 2009:
ziektebestrijding wintertarwe (Hfst 5, deel 2). Boek van Landbouwcentrum Granen Vlaanderen vzw. 194blz.
Halbert S., Manjunath K. (2004). Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of
citrus: a literature review and assessment of risk in Florida. Florida Entomologist, 87, 330-353.
Hamshou M., Smagghe G., Shahidi-Noghabi S., De Geyter E., Lannoo N., Van Damme E. (2010). Insecticidal
properties of Sclerotinia sclerotiorum agglutinin and its interaction with insect tissues and cells. Insect
Biochemistry and Molecular Biology, 40, 883-890.
Han Y., Wang Y., Bi J., Yang X., Huang Y., Zhao X., Hu Y., Cai Q. (2006). Constitutive and induced activities
of defense-related enzymes in aphid-resistant and aphid-susceptible cultivars of wheat. Journal of Chemical
Ecology, 35, 176-182.
Heath M. (2000). Hypersensitive response-related death. Plant Molecular Biology, 44, 321-334.
Helden A., Dixon A. (2002). Life-cycle variation in the aphid Sitobion avenae: costs and benefits of male
production. Ecological Entomology, 27, 692-701.
Hermans I., Haesaert G., Derycke V., Wittouck D. (2009). Bestrijding van bladluizen in wintergranen.
Landbouw&Techniek, 18, 19-21.
Höfte M. (2009). Gewasbeschadigers, partim: plantenziekten. Cursus, Gent, Faculteit Bio-
ingenieurswetenschappen. 132blz.
Hopkins D., Purcell A. (2002). Xulella fastidiosa: cause of Pierce‟s disease of grapevine and other emergent
diseases. Plant Disease, 86, 1056-1066.
Howe G., Jander G. (2008). Plant immunity to insect herbivores. Annual Review of Plant Physiology, 59, 41-66.
Hulting F., Orr D., Obrycki J. (1990). A computer program for calculation and statistical comparison of intrinsic
rates of increase and associated life table parameters. Florida Entomologist, 73, 601-612.
Irwin M., Kampmeier G., Weisser W. (2007). Aphids as crop pests: aphid movement: process and consequences
(Hfst 7, 34blz.). Boek van CAB international.
Jansen C., von Wettstein D., Schafer W., Kogel K-H., Felk A., Maier F. (2005). Infection patterns in barley and
wheat spikes inoculated with wild-type and trichodiene synthase gene disrupted Fusarium graminearum.
Proceedings of the National Academy of Sciences of the United States of America, 102, 16892-16897.
Jones J., Dangl J. (2006). The plant immune system. Nature, 444, 323-329.
Katis N., Tsitsipis J., Stevens M., Powell G. (2007). Aphids as crop pests: transmission of plant viruses (Hfst 14,
38blz.). Boek van CAB international.

65
Kema G., Köhl J., Kroon L., Löffler H., Meekes E., Schepers H., Scholten O., van Harmelen M., Waalwijk C.
(2002). Fusarium in granen. Identificatie van cruciale lacunes in kennis omtrent een internationaal probleem.
Plant research international B.V, Wageningen. 60blz.
Killiny N., Prado S., Almeida R. (2010). Chitin utilization by the insect-transmitted bacterium Xylella fastidiosa.
Applied and Environmental Microbiology, 76, 6134-6140.
Kluth S., Kruess A., Tscharntke T. (2002). Insects as vectors of plant pathogens: mutualistic and antagonistic
interactions. Oecologia, 133, 193-199.
Krupinsky J., Bailey K., McMullen M., Gossen B., Turkington T. (2002). Managing plant disease risk in
diversified cropping systems. Agronomy Journal, 94, 198-209.
Kunert G., Reinhold C., Gershenzon J. (2010). Constitutive emission of the aphid alarm pheromone, (E)-β-
farnesene, from plants does not serve as a direct defense against aphids. BMC Ecology, 10, 23-35.
Lazniewska J., Macioszek V., Lawrence C., Kononowicz A. (2010). Fight to the death: Arabidopsis thaliana
defense response to fungal necrotrophic pathogens. Acta Physiologiae Plantarum, 32, 1-10.
Loxdale H., Lushai G. (2007). Aphids as crop pests: population genetic issues: the unfolding story using
molecular markers (Hfst 2, 38blz.). Boek van CAB international.
Lushai G., Sherratt T., David O., De Barro P., Maclean N. (1997). Host selection by winged summer females of
the aphid Sitobion avenae. Entomologia Experimentalis et Applicata, 85, 199-209.
McLean A., Ferrari J., Godfray H. (2009). Effects of the maternal and pre-adult host plant on adult performance
and preference in the pea aphid, Acyrthosiphon pisum. Ecological Entomology, 34, 330-338.
McMullen M., Jones R., Gallenberg D. (1997). Scab of wheat and barley: a re-emerging disease of devastating
impact. Plant Disease, 81, 1340-1348.
McMullen M., Zhong S., Neate S. (2008). Fusarium Head Blight (scab) of small grains. Plant Disease
Management, NDSU Extention Service. 6blz.
Meyer J., Ingersoll C., McDonald L., Boyce M. (1986). Estimating uncertainty in population growth rates:
jackknife vs. bootstrap techniques. Ecology, 67, 1156-1166.
Müller C., Williams I., Hardie J. (2001). The role of nutrition, crowding and interspecific interactions in the
development of winged aphids. Ecological Entomology, 26, 330-340.
Nicol D., Copaja S., Wratte S., Niemeyer H. (1992). A screen of worldwide wheat cultivars for hydroxamic acid
levels and aphid antixenosis. Annals of Applied Biology, 121, 11-18.
Niks R., Marcel T. (2009). Nonhost and basal resistance: how to explain specificity? New Phytologist, 182, 817-
828.
Obopile M., Ositile B. (2010). Life table parameters of cowpea aphid, Aphis craccivora Koch (Homoptera:
Aphididae) on five cowpea Vigna unguiculata (L. Walp.) varieties. Journal of Pest Science, 83, 9-14.
Obst, A., Günther, B., Beck, R., Lepschy, J., Tischner, H. (2002). Weather conditions conducive to Giberella
zeae and Fusarium graminearum head blight of wheat. Journal of Applied Genetics, 43, 185-192.
PaDIL (2010). Diagnostic methods for wheat aphid Sitobion avenae. Cooperative Research Centre for National
Plant Biosecurity. 22blz.
Park K., Elias D., Donato B., Hardie J. (2000). Electroantennogram and behavioural responses of different forms
of the bird cherry-oat aphid, Rhopalosiphum padi, to sex pheromone and a plant volatile. Journal of Insect
Physiology, 46, 597-604.

66
Peters D., Vermunt A., Arkesteijn M. (2009). Vooral bladluizen en tripsen brengen virussen over. Onder glas, 1,
15-17.
Pettersson J., Tjallingii W., Hardie J. (2007). Aphids as crop pests: hostplant selection and feeding (Hfst 4,
26blz.). Boek van CAB international.
Piesik D., Lyszczarz A., Tabaka P., Lamparski R., Bocianowski J., Delaney K. (2010). Volatile induction of
three cereals: influence of mechanical injury and insect herbivory on injured plants and neighbouring uninjured
plants. Annals of Applied Biology, 157, 425-434.
Pickett J., Glinwood R. (2007). Aphids as crop pests: chemical ecology (Hfst 9, 24blz.). Boek van CAB
international.
Poehling H-M., Freier B., Klüken M. (2007). Aphids as crop pests: IPM case studies: grain (Hfst 25, 16blz.).
Boek van CAB international.
Powell G., Hardie J., Pickett J. (1997). Laboratory evaluation of antifeedant compounds for inhibiting settling by
cereal aphids. Entomologia Experimentalis et Applicata, 84, 189-193.
Prangle A., Birzele B., Kramer J., Meier A., Modrow H., Kohler P. (2005). Fusarium-inoculated wheat:
deoxynivalenol contents and baking quality in relation to infection time. Food Control, 16, 739-745.
Sadras V., Fereres A., Ratcliffe R. (1999). Wheat growth, yield and quality as affected by insect herbivores (Hfst
9, 46blz.). Boek ‘Wheat, ecology and physiology of yield determination’ van Satorre E., Slafer G. 456blz.
Samson R., Hoekstra E., Frisvad J. (2004). Introduction to food- and airborne fungi. Boek van CBS
(Centraalbureau voor schimmelcultures, Utrecht). 398 blz.
Shahidi-Noghabi S., Van Damme E., Smagghe G. (2009). Expression of Sambucus nigra agglutinin (SNA-I‟)
from elderberry bark in transgenic tobacco plants results in enhanced resistance to different insect species.
Transgenic Research, 18, 249-259.
Simon J-C., Rispe C., Sunnucks P. (2002). Ecology and evolution of sex aphids. Ecology&Evolution, 17, 34-39.
Smith C., Boyko E. (2006). The molecular bases of plant resistance and defense responses to aphid feeding:
current status. Entomologia Experimentalis et Applicata, 122, 1-16.
SPSS Inc. (2006). SPSS Base 15.0 User‟s Guide. 610blz.
Stanghellini M., Goldberg N. (1990). Ingestion-egestion and aerial transmission of Pythium aphanidermatum by
shore flies (Ephydrinae: Scatella stagnalis). Phytopathology, 80, 1244-1246.
Stanghellini M., Rasmussen S., Kim D. (1999). Aerial transmission of Thielaviopsis basicola, a pathogen of
corn-salad, by adult shore flies. Phytopathology, 89, 476-479.
Stavrinides J., McCloskey J., Ochman H. (2009). Pea aphid as both host and vector for the phytopathogenic
bacterium Pseudomonas syringae. Applied and Environmental Microbiology, 75, 2230-2235.
Steidle J., Fischer A., Gantert C. (2005). Do grains whisper for help? Evidence for herbivore induced synomones
in wheat grains. Entomologia Experimentalis et Applicata, 115, 239-245.
Stijger, H. (2005). Bladluizen veruit de snelst groeiende insectenplaag. Onder glas, 11, 46-49.
The International Aphid Genomics Consortium. (2010). PLos Biology, 8, 24blz.
Thomma B., Nürnberger T., Joosten M. (2011). Perspective of PAMPs and effectors: the blurred PTI-ETI
dichotomy. The Plant Cell, 23, 4-15.
Tirry L. (2009). Gewasbeschadigers, partim: dierlijke plantenbeschadigers. Cursus, Gent, Faculteit Bio-
ingenieurswetenschappen. 182blz.

67
Trail F. (2009). For blighted waves of grain: Fusarium graminearum in the postgenomics era. Plant Physiology,
149, 103-110.
Unsicker S., Kunert G., Gershenzon J. (2009). Protective perfumes: the role of vegetative volatiles in plant
defense against herbivores. Current Opinion in Plant Biology, 12, 479-485.
van Dam N. (2009). How plants cope with biotic interactions. Plant Biology, 11, 1-5.
van Emden H., Harrington R. (2007). Aphids as crop pests. Boek van CAB international. 752blz.
Van Outryve J. (2007a). Natuur als tegenstrever. Landbouw&Techniek, 18, 34-36.
Van Outryve J. (2007b). Europa stelt de norm. Landbouw&Techniek, 18, 28-31.
Vantornhout I., Minnaert H., Tirry L., De Clercq P. (2006). Influence of diet on life table parameters of Iphiseius
degenerans. Experimental and Applied Acarology, 35, 183-195.
Verhiest G. (2009). Bladluizen koloniseren massaal. Boer&Tuinder, 27, 10.
Visser J., Fu-shun Y. (1995). Electroantennogram responses of the grain aphids Sitobion avenae (F.) and
Metopolophium dirhodum (Walk.) (Hom., Aphididae) to plant odour components. Journal of Applied
Entomology, 119, 539-542.
Visser J., Piron P. (1997). Odour response profiles in aphids differentiating for species, clone, form and food.
Proceedings of the Section Experimental and Applied Entomology of the Netherlands Entomological Society, 8,
115-120.
Vogelgsang S., Sulyok M., Hecker A., Jenny E., Krska R., Schuhmacher R., Forrer H-R. (2008). Toxigenicity
and pathogenicity of Fusarium poae and Fusarium avenaceum on wheat. European Journal of Plant Pathology,
122, 265-276.
Waalwijk C., Kastelein P., de Vries I., Kerényi Z., van der Lee T., Hesselink T., Köhl J., Kema G. (2003). Major
changes in Fusarium species in the Netherlands. European Journal of Plant Pathology, 109, 743-754.
Wang J., Pawelzik E., Weinert J., Zhao Q., Wolf G. (2008). Factors influencing falling number in winter wheat.
European Food Research and Technology, 226, 1365-1371.
Wang J. Wieser H., Pawelzik E., Weinert J., Keutgen A., Wolf G. (2005). Impact of the fungal protease
produced by Fusarium culmorum on the protein quality and breadmaking properties of winter wheat. European
Food Research and Technology, 226, 552-559.
Walter S., Nicholson P., Doohan F. (2010). Action and reaction of host and pathogen during Fusarium head
blight disease. New Phytologist, 185, 54-66.
Williams I., Dixon A. (2007). Aphids as crop pests: life cycles and polymorphism (Hfst 3, 18blz.). Boek van
CAB international.
Wittouck D., Haesaert G., Ver Elst P. (2006). Schadelijke insecten, hoe raak je ervanaf? Landbouw&Techniek,
11, 22-23.
Yang X., Thannhauser T., Burrows M., Cox-Foster D., Gildow F., Gray S. (2008). Coupling genetics and
proteomics to identify aphid proteins associated with vector-specific transmission of polerovirus (Luteoviridae).
Journal of Virology, 82, 291-299.
Zhao L., Chen J., Cheng D., Sun J., Liu Y., Tian Z. (2009). Biochemical and molecular characterizations of
Sitobion avenae-induced wheat defense responses. Crop Protection, 28, 435-442.
Zhu-Salzman K., Bi J-L., Liu T-X. (2005). Molecular strategies of plant defense and insect counter-defense.
Insect Science, 12, 3-15.


















