biosdocumenten.pbworks.combiosdocumenten.pbworks.com/w/file/fetch/73056386/Ge… · Web viewDit...
42
Genetica: samenvatting Hoofdstuk 11 Voortplanting Geslachtelijk voortplanting is mogelijk dankzij meiose. Bij meiose krijgen de nakomelingen het genetisch materiaal van de 2 ouders. De cellen die in de meiose meedoen noemt men de gameten en deze zijn haploïd (=n). De andere niet-reproductieve cellen noemt men somatische cellen en zijn diploid (=2n). De gameten zullen zich tijdens de voortplanting samenvoegen en een zygote vormen, dit noemt de bevruchting of syngamy. De gameten hebben maar n-chromosomen omdat anders het chromosomen aantal bij elke bevruchting zou verdubbelen. Tijdens de ontwikkeling van de diploïde zygote worden de cellen die de gameten zullen worden apart gezet. Dit noemt men de germ-line cellen; deze zullen later meiose onder gaan om n-chromosomen te krijgen. Kenmerken van de Meiose De meiose is onderverdeeld in 2 processen: meiose 1 en meiose 2. Elk proces heeft een profase, metafase, anafase en telofase. Meiose 1 Profase Voor deze fase is het DNA al verdubbeld. Tijdens deze fase zullen de homologe chromosomen (die elk bestaan uit 2 zusterchromatiden) associëren met elkaar, dit proces wordt synapsis genoemd. Tussen de 2 homologe chromosomen zal een synaptonemaal complex ontstaan: proteïnebruggen die de 2 aan elkaar binden. De structuur die hierbij ontstaat noemt men een tetrade of bivalent. Er zal ook crossing-over of genetische recombinatie gebeuren: de homologen zullen chromosoommateriaal met elkaar uitwisselen en zo het genetisch materiaal dus recombineren. Deze wordt geïnduceerd door de recombination nodules, enzymen die de chromatiden zullen breken en terug aan elkaar zetten. De plaats waar de crossing-over gebeurt noemt men de chiasmata (sing.: chiasma). Na de crossing-over zal het synaptonemaal complex af gebroken worden en worden de homologen bij elkaar gehouden door de chiasmata + elk zusterchromatide
Transcript of biosdocumenten.pbworks.combiosdocumenten.pbworks.com/w/file/fetch/73056386/Ge… · Web viewDit...

Genetica: samenvatting
Hoofdstuk 11
VoortplantingGeslachtelijk voortplanting is mogelijk dankzij meiose. Bij meiose krijgen de nakomelingen het genetisch materiaal van de 2 ouders. De cellen die in de meiose meedoen noemt men de gameten en deze zijn haploïd (=n). De andere niet-reproductieve cellen noemt men somatische cellen en zijn diploid (=2n).De gameten zullen zich tijdens de voortplanting samenvoegen en een zygote vormen, dit noemt de bevruchting of syngamy. De gameten hebben maar n-chromosomen omdat anders het chromosomen aantal bij elke bevruchting zou verdubbelen. Tijdens de ontwikkeling van de diploïde zygote worden de cellen die de gameten zullen worden apart gezet. Dit noemt men de germ-line cellen; deze zullen later meiose onder gaan om n-chromosomen te krijgen.
Kenmerken van de MeioseDe meiose is onderverdeeld in 2 processen: meiose 1 en meiose 2. Elk proces heeft een profase, metafase, anafase en telofase.
Meiose 1 Profase
Voor deze fase is het DNA al verdubbeld. Tijdens deze fase zullen de homologe chromosomen (die elk bestaan uit 2 zusterchromatiden) associëren met elkaar, dit proces wordt synapsis genoemd. Tussen de 2 homologe chromosomen zal een synaptonemaal complex ontstaan: proteïnebruggen die de 2 aan elkaar binden. De structuur die hierbij ontstaat noemt men een tetrade of bivalent. Er zal ook crossing-over of genetische recombinatie gebeuren: de homologen zullen chromosoommateriaal met elkaar uitwisselen en zo het genetisch materiaal dus recombineren. Deze wordt geïnduceerd door de recombination nodules, enzymen die de chromatiden zullen breken en terug aan elkaar zetten. De plaats waar de crossing-over gebeurt noemt men de chiasmata (sing.: chiasma). Na de crossing-over zal het synaptonemaal complex af gebroken worden en worden de homologen bij elkaar gehouden door de chiasmata + elk zusterchromatide blijft bij elkaar dankzij het gemeenschappelijke centromeer. Hierop zullen tijdens de metafase de microtubuli binden.
MetafaseIn de metafase zullen de chromosomen zich oplijnen aan de evenaar van de cel of de metaplaat. De orientatie van elk chromosoom is willekeurig, zowel de moeder als de vaderlijke homoloog kan worden georiënteerd in de richting van een bepaalde pool. Hierdoor zeggen we dat meiosis 1 zorgt vooronafhankelijke verscheidenheid van de chromosomen. Aan de centromeren zullen vanuit de polen de kinetochore micotubuli binden.
AnafaseNu gaan de kinetochore microtubuli verkorten en zal er een spanning ontstaan. Deze spanning zal de homologen uit elkaar trekken. Elke pool zal 1 van de homologen krijgen. De zusterchromatiden blijven aan elkaar omdat ze fungeren als 1 eenheid. Op de tekening is ook

zichtbaar dat de ruimtelijke liggen de microtubuli alleen kunnen binden aan de centromeren van de homologen en niet van de zusterchromatiden.
TelofaseDe homologe chromosomen zijn nu afgescheiden van elkaar en beginnen met de vorming van een cluster aan elke pool. Ook het nucleair membraan zal ook terug vormen. Elke kern bevat nu 2 zusterchromatiden, deze zijn verschillend van elkaar dankzij de crossing-over in profase 1.
Bij sommige dieren gebeurt de meiosis zonder recombinatie, men spreekt dan van achiasmatische scheiding. Dit geeft dan gameten met genetische informatie van 1 ouder, er zal dus geen verschil zijn tussen ouder en nakomeling.De eerste meiose wordt ook de reductiedeling genoemd omdat het aantal chromosomen vermindert tot n.Tussen de 2 meiotische delingen is er geen verdubbeling van het DNA.Meiose 2de 2de meiotische deling is gelijk aan een mitotische deling zonder de voorafgaande DNA-verdubbeling. Bij deze deling zullen de zustercromatide scheiden maar blijft het aantal chromosomen hetzelfde. Als deze deling gedaan is heeft men 4 haploïde cellen. Bij dieren zullen deze zich direct omvormen tot gameten. Bij planten en fungi delen ze zich mitotisch met als gevolg dat er meer gameten zijn maar ook adulten met verschillende chromosomenparen in de gameten.
ProfaseHet nuclaire membraan zal terug oplossen en er wordt een nieuwe spoelfiguur gevormd.
MetafaseDe kinetochore microtubuli binden aan de centromeren van de zusterchromatiden. Deze zullen de chromosomen naar de meta-plaat brengen als een gevolg van de spanning.
AnafaseDe microtubuli zullen zich verkorten en de zusterchromatiden uit elkaar trekken. De zusterchromatiden migreren nu elk naar een pool van de cel.
TelofaseDe nucleaire enveloppe wordt terug gevormd. Hierna volgt cytokinesis: de cel wordt in gesnoerd en er worden 2 dochtercellen gecreeërd.

Fouten in de meiose Non-disjunctie: het falen van de chromosomen om naar de tegenliggende polen te migreren
gedurende elk van de meiotische delingen. Aneupoloïde gameten: gameten met ontbrekende of extra chromosomen. Die is bij de mens
de grootste oorzaak van spontane abortus.
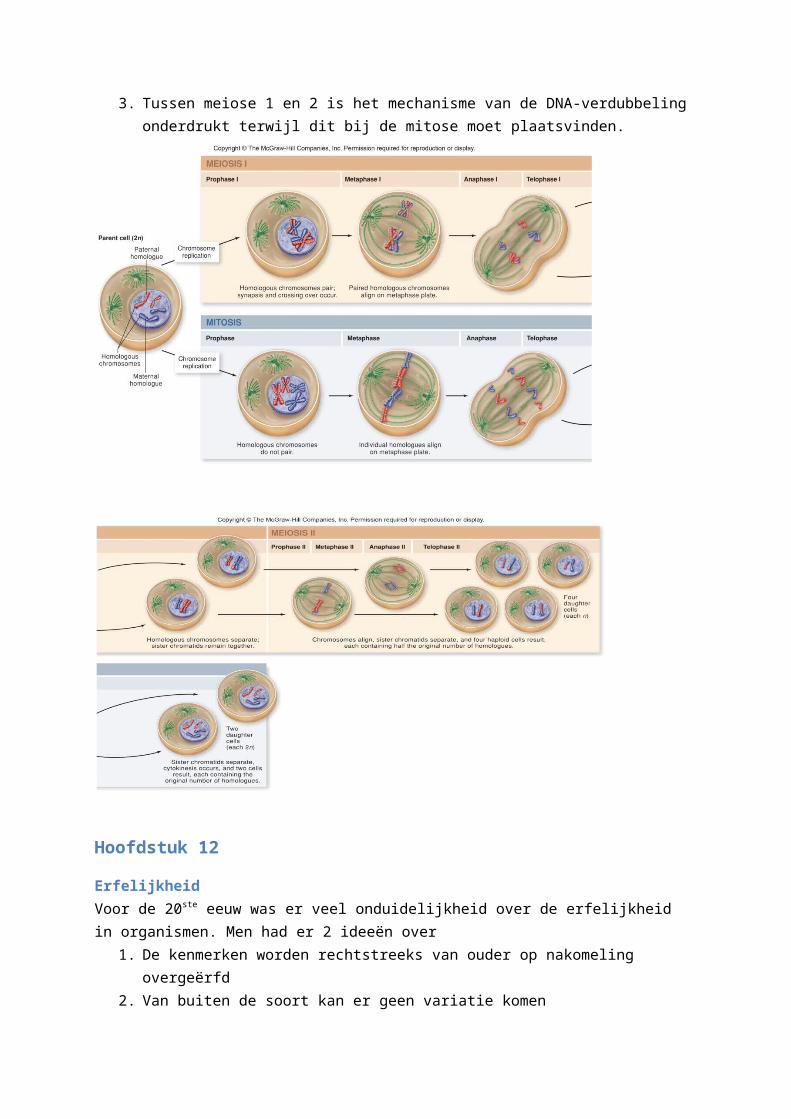
De verschillen met mitose1. Bij de mitose zullen de homologe chromosomen in de metafase zich onafhankelijk van elkaar
gedragen.2. In de anafase zijn het niet de homologen die splitsen maar de zusterchromatiden.3. Tussen meiose 1 en 2 is het mechanisme van de DNA-verdubbeling onderdrukt terwijl dit bij
de mitose moet plaatsvinden.
Hoofdstuk 12
ErfelijkheidVoor de 20ste eeuw was er veel onduidelijkheid over de erfelijkheid in organismen. Men had er 2 ideeën over
1. De kenmerken worden rechtstreeks van ouder op nakomeling overgeërfd2. Van buiten de soort kan er geen variatie komen
Dit leidde tot een paradox, want als men deze 2 ideeën aanneemt dan zou na een bepaalde tijd de leden van 1 soort allemaal hetzelfde uiterlijk hebben.
Geschiedenis Josef Kölreuter
1760 heeft hij verschillende tabaksplanten kunnen kruisen. Hij heeft dus de eerste hybriden gemaakt. De hybride nakomelingen verschilden van hun ouders. In de 2de generatie werd er nog meer variatie ontdekt.Dit leidde tot de conclusie dat kenmerken niet rechtstreeks van ouder op nakomeling worden overgeërfd.
T.A. KnightDe eerste persoon die 2 raszuivere lijnen van de tuinboon kruiste. Met een raszuivere lijn bedoeld dat de nakomelingen geproduceerd uit zelfbevruchting uniform bleven van de ene generatie naar de volgende. Knight ontdekte dat de F1-generatie leek op 1 ouderlijke lijn maar dat in de 2de generatie de beide kenmerken waren te vinden.
Gregor MendelGregor mendel (1822-1884) deed ook kruisproeven met de tuinboon. Er zijn een aantal redenen waarom de tuinboon zo vaak word gebruikt:
1. De plant kan hybride nakomelingen verkrijgen (onderzoek Knight)2. Er zijn verschillende variëteiten van de tuinboon3. Ze zijn gemakkelijk te kweken en hebben een korte generatietijd. Dit maakt het
mogelijk om erfelijk op te onderzoeken.4. Zowel de vrouwelijke als mannelijke geslachtsorganen zijn ingesloten in elke bloem.
De gameten van 1 bloem kunnen samensmelten tot een levensvatbare nakomeling, dit noemt men zelfbevruchting. men kan de bevruchting controleren door het mannelijke geslachtsorgaan te verwijderen en pollen in te brengen van een andere plant.
Tijdens Mendel zijn experimenten had hij 3 stadia:I. Hij zorgde ervoor dat de variëteiten raszuiver waren door er zelfkruising mee uit te
voerenII. Deze zuivere lijnen liet hij een kruisbestuiving of reciproke kruising ondergaan voor
een bepaald kenmerk (pollen van een witte bloem op een paarse en omgekeerd)III. De hybride nakomelingen werden onderworpen aan zelfbevruchting zodat de
erfelijkheid van de kenmerken kon gevolgd worden en de nakomelingen met kenmerk konden geteld worden.

De eerste kruising die Mendel deed was een monohybride kruising: hierbij heeft men maar 2 variaties op 1 kenmerk. Mendel deed dit voor 7 kenmerken. Bij de F1 generatie (nakomelingen van de parentale planten) leken de planten maar op 1 ouder. Dit kenmerk men dominant, het andere kenmerk noemt men recessief.De F2 generatie (nakomelingen van de F1 generatie) had wel planten met het recessieve kenmerk. Na een telling kwam Mendel uit op een 3:1 verhouding.Deze 3:1 verhouding is eigenlijk 1:2:1 verhouding, ¼ van de planten was dominant raszuiver, ¼ was recessief raszuiver en 2/4 was dominant niet zuiver (kenmerkende verdeling voor een monohybride kruising).Hiermee bewees hij dat het erfelijkheidsmodel van toen fout was en gaf een nieuw:
1) Ouders geven discrete informatie van de kenmerken door aan hun nakomelingen. Deze informatie noemt men de genen.
2) Elke nakomeling ontvangt 1 kopie van een gen van elke ouder3) Niet elk gen is identiek, de alternatieve vorm van een gen noemt men een allel. Als
men 2 gameten heeft met hetzelfde allel, dan is de nakomeling homozygoot. Is het allel verschillend dan is de nakomeling heterozygoot.
4) Onder de allelen is er geen menging of verandering van elkaar. Daardoor als de nakomeling gameten produceert, de allelen willekeurig scheiden in de gameten.
5) Alleen het dominante allel zal voor dat kenmerk geuit worden, het andere wordt onderdrukt.
Het fenotype van een organisme is de uiterlijke verschijning ervan, het genotype zijn al de allelen die dat organismen heeft. Zo is de 3:1 verhouding phenotypische verhouding en de 1:2:1 de genotypische verhouding.Dit leidde ook tot het Principle of Segregation: de twee allelen voor een gen scheiden tijdens de gameetvorming en worden weer willekeurig verenigd, een van elke ouder, tijdens de bevruchting.Deze scheiding van de allelen gebeurt in de meiose.
Het Punnett vierkant maakt het mogelijk om deze erfelijkheidstesten symbolisch op te lossen, zonder dat men honderden planten moet kweken.Ook menselijke kenmerken zijn dominant en recessief. Omdat men mensen niet kan kweken zoals planten, onderzoekt men deze kenmerken via stamboomanalyse.
Juveniel glaucoom:De ziekte zorgt voor de degeneratie van de oogziekte wat uiteindelijk tot blindheid leidt. Uit een stamboom heeft men kunnen afleiden dat het om een dominant allel gaat, het komt in elke generatie van de familie voor. Dit is nagenoeg onmogelijk voor een recessief allel, aangezien men daarvoor grote getallen van ondirecte individuen met het allel dan nodig heeft
Albinisme:Hierbij wordt er geen pigment aangemaakt in het haar/huid/ogen. Het is een recessief kenmerk en verspreidt zich gelijkmatig over mannen en vrouwen. Meestal wordt het veroorzaakt doordat men een niet-functioneel allel van het enzyme tyrosinase heeft.

Het volgende onderzoek dat Mendel ondernam was dihybride kruisingen, dit zijn kruisingen waarbij de overerving van 2 kenmerken van de ouders worden gevolgd. De heterozygoten zijn nu ook dubbel heterozygoot. Na de 3 stadia van zijn experiment kwam Mendel in de F2 generatie aan een 9:3:3:1 verhouding van de onafhankelijke kenmerken. Hieruit volgt zijn 2de wet van erfelijkheid of the Principle of Independent Assortement: In een dihybride kruising, sorteren de allelen van elk gen onafhankelijk of met andere woorden de scheiding van de verschillende allel paren is onafhankelijk. De oorzaak hiervan ligt in de onafhankelijke op lijning van de chromosomenparen aan de metaplaat in de 1ste metafase van de meiose.Om de waarschijnlijkheid van een monohybride kruising te voorspellen zijn er een aantal regels:
Optelregel: de waarschijnlijkheid dat 2 wederzijds uitsluitende gebeurtenissen tegelijk gebeuren is de som van hun individuele waarschijnlijkheid.
Vermenigvuldigingsregel: de waarschijnlijkheid dat 2 onafhankelijke gebeurtenissen tegelijk gebeuren is het product van hun individuele waarschijnlijkheid.
de vermenigvuldigingsregel kunnen we ook toepassen op een dihybride kruising. Dit komt omdat we een dihybride kruising kunnen als 2 onafhankelijke monohybride kruisingen. En omdat ze onafhankelijk zijn mogen we de vermenigvuldigingsregel toepassen.Mendel heeft ook de testkruising uitgevonden. Bij een testkruising wilt men het onbekende genotype achterhalen, dit doet men door de onbekende te laten kruisen met de recessieve homozygoot van dat kenmerk.
Als men na de kruising dan recessieve kenmerken ziet weet men dat de onbekende deze ook in zijn genotype heeft. Indien niet dan is de onbekende homozygoot voor het dominante kenmerk.
Uitbreiding op MendelMendel’s erfelijkheidsmodel gaat uit van een paar foute vereenvoudigingen:
elke kenmerk wordt bepaald door een enkel gen met twee alternatieve allelen het milieu heeft geen effect op het fenotype van het organisme er zijn maar 2 allelen voor elk gen de producten van genen gedragen zich onafhankelijk er is een dominante-recessieve relatie tussen allelen

In de moderne genetica heeft men ontdekt dat men deze vereenvoudigingen niet zomaar mag weg laten want ze hebben wel degelijk invloed of zijn fout.
Zo kan het zijn dat een kenmerk wordt beïnvloed door meer als 1 gen, dit noemt men dan polygene overerving. Deze kenmerken hebben een continue variatie en als men ze sorteert
bekomt men een klokhistogram. Deze kenmerken noemt men quantitatieve kenmerken: hoe groter het aantal genen beïnvloeden van een kenmerk, meer continu de verwachte distributie van de versie van dat kenmerk.
Genen kunnen meer als 2 allelen hebben. Een voorbeeld hiervan is het menselijke bloedtype. Hiervoor heeft men 3 allelen die het bloedtype bepalen.
1 allel kan effect hebben op meerdere fenotypes, men noemt dit dan pleiotropie. Zo een allel kan dan dominant zijn voor het ene kenmerk en recessief voor het andere. Deze allelen liggen aan de basis van menselijke ziekten als mucovisidose en sikkelcelanemie.
Bepaalde kattensoorten hebben een allel dat alleen de productie toelaat van pigment onder een bepaalde temperatuur. De extremiteiten (oren, benen, etc.) zullen dus alleen pigment produceren. Hierbij heeft het milieu dus een grote invloed.
Sommige genproducten reageren wel met elkaar. Dit zorgt ervoor dat de verhouding die men verwacht bij onafhankelijke sortering (1:2:1, …) veranderd. Dit gebeurt zelf bij genen die op verschillende chromosomen liggen met onafhankelijke sortering;
De relatie dominant- recessief verbergt een heleboel biochemische reacties. Men heeft verschillende vormen van dominantie die elk een ander effect hebben op de reacties
Onvolledige dominantie: hierbij zal de heterozygote nakomeling lijken op beide ouders en het kenmerk zal tussenin die van de ouders liggen (vb. rood+wit= roos)
Codominantie: hierbij zal het fenotype van de nakomeling enkele kenmerken tonen van de fenotypes van de homozygoten. Dit is het geval bij bloedtype.
Hoofdstuk 13
Genen op de geslachtschromosomenThomas H. Morgan was de eerste die kon bewijzen dat genen op chromosomen lagen (1910). Hiermee bewijs hij de chromosomale theorie van overerving voorgesteld door Walter Sutton in 1902.
Na proeven met de fruitvlieg ontdekte Morgan dat het gen voor de oogkleur op het X-chromosoom ligt. Zo een kenmerk noemt men geslachtsgebonden kenmerken. Het geslacht wordt bepaald door geslachtschromosoom, een vrouw heeft 2 X-chromosomen en een man 1 X- en Y-chromosoom (bij mensen). Bij de mens is de standaardembryo vrouwelijk. Het zal pas veranderen als het SRY-gen op het Y-chromosoom wordt geactiveerd.

Uitzonderingen hierop bevestigen dit mechanisme, zo kan er een deel van Y-chromosoom over gaan naar het X-chromosoom en een vrouwelijk individu (XX) zal toch ontwikkelen tot man. Een andere genetische aandoening zorgt ervoor dat de mannelijke hormonen geen effect hebben en een XY-individu zich vrouwelijk ontwikkelt. Dit kan ook gebeuren na een mutatie van het SRY-gen.Op het Y-chromosoom zitten er maar weinig allelen dit heeft als gevolg dat de recessieve allelen op het X-chromosoom vaak tot uiting komen omdat deze geen actieve tegenhanger hebben. Bij andere soorten heeft het milieu soms invloed op het geslachtsbepalende gen. Ook deze chromosomen kunnen ziektes op zich dragen, een ervan is hemofilie. Het een recessief allel op het X-chromosoom. Daardoor zullen de mannen het zeker vertonen en zijn heterozygote vrouwen drager. Het allel beïnvloed een proteïne in een reeks van eiwitten die zorgen voor het vormen van bloedklonters. Bij mensen die de ziekte hebben zullen wondjes niet vanzelf stoppen met bloeden. Aangezien vrouwelijke individu’s 2 X-chromosomen hebben moet er 1 worden gedeactiveerd om gelijke genexpressie te hebben als een man. Dit noemt men doseringscompensatie. Als het geslacht van het individu bepaald is zal het bij vrouwen als in de embryonale fase X-chromosomen deactiveren. Het X-chromosoom dat gedeactiveerd wordt verschilt van cel tot cel is willekeurig. Het X-chromosoom zal dan condenseren tot een Barr-lichaampje en zich hechten aan het nucleair membraan. De deactivatie heeft als gevolg dat als een vrouw heterozygoot is voor een bepaald allel op het X-chromosoom het in sommige cellen wordt geuit en in andere niet vb. lapjeskat. Men noemt dit genetisch mozaïek.
Uitzonderingen op de chromosoomtheorieMitochondrieën en chloroplasten bevatten ook DNA dat niet word doorgegeven via Mendeliaanse overerving. Al s een zygote gevormd wordt zal deze van elke ouder evenveel genetisch materiaal ontvangen maar al het mitochondriaal en chloroplastisch DNA komt van een enkele ouder meestal de moeder. Dit noemt men maternale overerving.
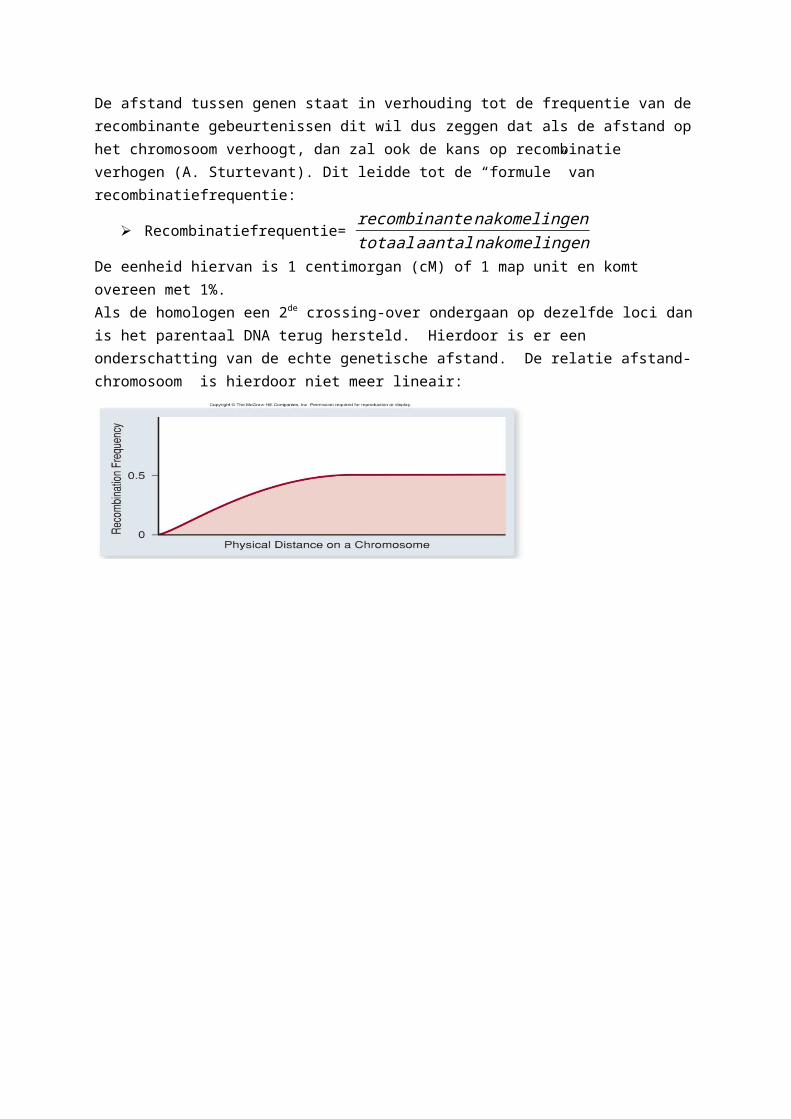
Gene mappingMen kan genen in kaart brengen dankzij crossing-over. Want als er een crossing-over gebeurt worden de allelen gerecombineerd en vormen ze recombinante gameten. De afstand tussen genen staat in verhouding tot de frequentie van de recombinante gebeurtenissen dit wil dus zeggen dat als de afstand op het chromosoom verhoogt, dan zal ook de kans op recombinatie verhogen (A. Sturtevant). Dit leidde tot de “formule” van recombinatiefrequentie:
Recombinatiefrequentie= recombinantenakomelingentotaal aantal nakomelingen
De eenheid hiervan is 1 centimorgan (cM) of 1 map unit en komt overeen met 1%.Als de homologen een 2de crossing-over ondergaan op dezelfde loci dan is het parentaal DNA terug hersteld. Hierdoor is er een onderschatting van de echte genetische afstand. De relatie afstand-chromosoom is hierdoor niet meer lineair:

Op groter afstanden hebben meerdere recombinaties plaats (gevolg uit A.Sturtevant) dit zorgt ervoor dat bij oneven crossing-overs recombinante gameten worden gevormd, en bij even de parentale gameten. Als men dan nog verder gaat zullen het aantal recombinante gameten gelijk worden aan de parentale gameten en de loci zullen onafhankelijke verscheidenheid tonen.Om te weten of 2 genen op een grotere afstand crossing-over hebben gedaan voert men een driepuntstestkruising uit. Zo is het probleem van even crossing-overs opgelost omdat men aan het derde gen kan zien of er een crossing-over is gebeurt.
Het menselijk genoom: Tot in 1980 was men afhankelijk van historische stambomen om menselijke genen in kaart te brengen. Er konden ook geen merkers worden gebruikt omdat het menselijk genoom zo groot is. Dit veranderde met de anonieme merkers, genetische merkers die opgespoord kunnen worden aan de hand van moleculaire technieken maar die geen fenotype veroorzaken. Van deze merkers heeft men er duizenden kunnen vinden en heeft men een map kunnen maken van het menselijk genoom.Met deze informatie kan men nu nog verder gaan. Met behulp van SNP’s (= sinle-nucleotide polymorphims) kan men enkelvoudige basen identificeren en in kaart brengen. Zo kan men personen vergelijken, de genetische kaart van een persoon een hogere “resolutie” geven waardoor er in de forensische analyse minder fouten in kunnen gemaakt worden.
Genetische aandoeningen1. Sikkelcelanemie
Het is eerste ziekte waarvan men heeft kunnen aantonen dat het wordt veroorzaakt door een mutatie in een proteïne. De ziekte tast de hemoglobine aan zodat er een verstoorde zuurstoftoevoer is naar de weefsels. De hemoglobine gaat door de mutatie aan elkaar beginnen te plakken en karakteristieke vorm aannemen van een sikkel.Homozygoten voor deze ziekten hebben een verminderde levensduur en onderbroken ziekteprofiel. De heterozygoten lijken normaal maar hun bloedcellen hebben een verminderde eigenschap om zuurstof te dragen. Het blijkt wel dat de heterozygoten een verhoogde weerstand hebben tegen de parasiet die malaria veroorzaakt.
2. NondisjunctieDit is het falen van homologe of zuster chromatiden om te scheiden tijdens de meiose.dit leidt tot verschillende genetische aandoeningen:

Aneuploïdie, hierbij verliest of krijgt men een extra chromosoom. De individu dat een chromosoom verliest heeft monosomie. Deze individu’s overleven de embryonale fase niet. Personen die een extra chromosoom verkrijgen hebben trisomie. Dit is alleen levensvatbaar voor de chromosomen (menselijk) 13,15,18,21,22. Bij de eerste 3 zullen de personen zware lichamelijk defecten ontwikkelen en sterven binnen een paar maanden. Bij trisomie 21 en 22 zal de ontwikkeling van het skelet vertraagd zijn en verkrijgt men korte, slechte spierspanning. Trisomie 21 is dan ook nog eens gekenmerkt met een mentale handicap. Trisomie werd het eerst beschreven in 1866 door J.langdon Down en word daarom ook Down syndrome genoemd. De ziekte wordt beïnvloed door de leeftijd van de moeder. Dit komt omdat de eicellen van een vrouw worden gemaakt in de embryonale fase. Als de vrouw kinderen krijgt zijn de eicellen even oud als de vrouw. Hierdoor is er een grotere kans dat celdelings problemen zich hebben verzamelt in de vrouwelijke gameten.
3. Nondisjunctie van de geslachtschromosomenDeze non-disjunctie veroorzaakt minder problemen, de personen bereiken vaak volwassenheid en zijn vruchtbaar.
XXX zygote: hierbij is er zygote gevormd met een X gameet en een XX gameet. De persoon zal een vrouw worden met 1 actieve X-chromosoom en 2 Barr-lichaampjes
XXY zygote: de persoon zal mannelijke zijn van geslacht maar met veel vrouwelijke kenmerken met soms mentale achterstand. Dit noemt het Klinefelter syndroom.
YO: er wordt een zygote gevormd tussen een Y gameet en een gameet zonder geslachtschromosoom, het individu is niet levensvatbaar.
XO: de persoon zal ontwikkelen tot een vrouw met lichamelijke aandoeningen. Ze zal steriel zijn, de voortplantingsorganen zullen zich nooit volledig ontwikkelen en een ingekorte nek. De mentale capaciteiten van deze persoon zullen laag tot normaal zijn. Dit noemt het Turner-syndroom.
XYY: de persoon zal ontwikkelen tot een normale man, dit noemt het Jacob syndroom;
4. Genomische inprentingHierbij is het fenotype dat vertoond wordt bij de nakomeling afhankelijk van welke ouder de nakomeling het allel heeft gekregen. Sommige genen zijn geïnactiveerd in de vaderlijke kiembaan en komen dus niet tot uitdrukking in de zygote. Andere genen worden geïnactiveerd in de moederlijke kiembaan met dezelfde resultaat. Dit maakt de zygote haploïde voor een ingedrukt gen. Prader-Willi syndroom en Angelman syndroom worden beiden veroorzaak door een deletie op chromosoom 15. Als chromosoom 15 met de deletie geërfd is van de vader dan zal de nakomeling PWS vertonen. Komt het van de moeder zal het kind AS vertonen.
5. Opsporing van genetische aandoeningenMen heeft verschillende manieren om genetische aandoening op te sporen:
1) Stamboomanalyse:door de stamboom van een persoon te analyseren kan met soms de waarschijnlijkheid berekenen of die persoon een drager is van een bepaalde genetische aandoening.

2) Amniocentesis of vruchtwaterpunctie: hierbij gaat men met een naald amniotische vloeistof opzuigen. In de vloeistof zitten cellen van foetus die men kan laten groeien in cultuur en analyseren.
3) Chorionische vlokkentest: hierbij worden er cellen verwijderd van het chorion. Men doet hetzelfde als bij de vruchtwaterpunctie alleen heeft men sneller resultaat.
Hoofdstuk 14
Aard van het genetische materiaal Frederick Griffith:1928
Griffith deed experimenten met de bacterie die longontsteking veroorzaakt. Hij werkte met de normale vorm (de ziekteverwekker) enen mutatie ervan (veroorzaakt geen ziekte). Nadat hij muizen injecteerde met de normale vorm, de mutatie en een mengeling waarbij de normale was afgedood waren dit de resultaten:
De normale vorm leidde tot ziekte en uiteindelijk de dood De mutatie zorgde voor geen ongemakken Het mengsel leidde terug tot ziekte en de dood.
Griffith besloot dat de informatie die de virulentie (=het ziekteverwekkend vermogen) bepaalde overgedragen was van de dode cellen naar de ongevaarlijke gemuteerde cellen, hij noemde dit tranformatie.In de moderne genetica weten we dat DNA werd overgedragen tussen de verschillende cellen
Avery, Macleod and McCarty:1944Ze herhaalden het experiment van Griffith maar verwijderden van de bacteriën bijna alle proteïnen. Ze merkten dat er geen reductie was in de transformatie activiteit, meer nog het afgezonderde product had de karakteristieken van DNA:
1) De elementaire compositie kwam overeen met DNA2) Na centrifugatie bleek het dezelfde dichtheid te hebben als DNA3) De vetten en proteïnen verwijderen leverde geen verminderde activiteit op4) Alleen DNA-verterende enzymen zorgden ervoor dat de activiteit
verminderde/stopte.Deze experimenten steunde de theorie dat DNA het genetisch materiaal is.
Hershey and ChaseZij deden experimenten met bacteriofagen, virussen die alleen bacteriën infecteerden en bestonden uit DNA en proteïnen. Een deel van de fagen werd gemerkt met radioactief zwavel, dit kwam dan in de proteïnen terecht van de fagen werd gemerkt met radioactief fosfor wat in het DNA terecht kwam. Ze lieten de fagen bacteriën infecteren en zorgden ervoor dat ze na de infectie de fagen op het oppervlak va de bacterie verwijderden. Hiermee verzekerden dat ze alleen het genetisch materiaal detecteerden.Ze concludeerden dat het DNA het genetische materiaal was.

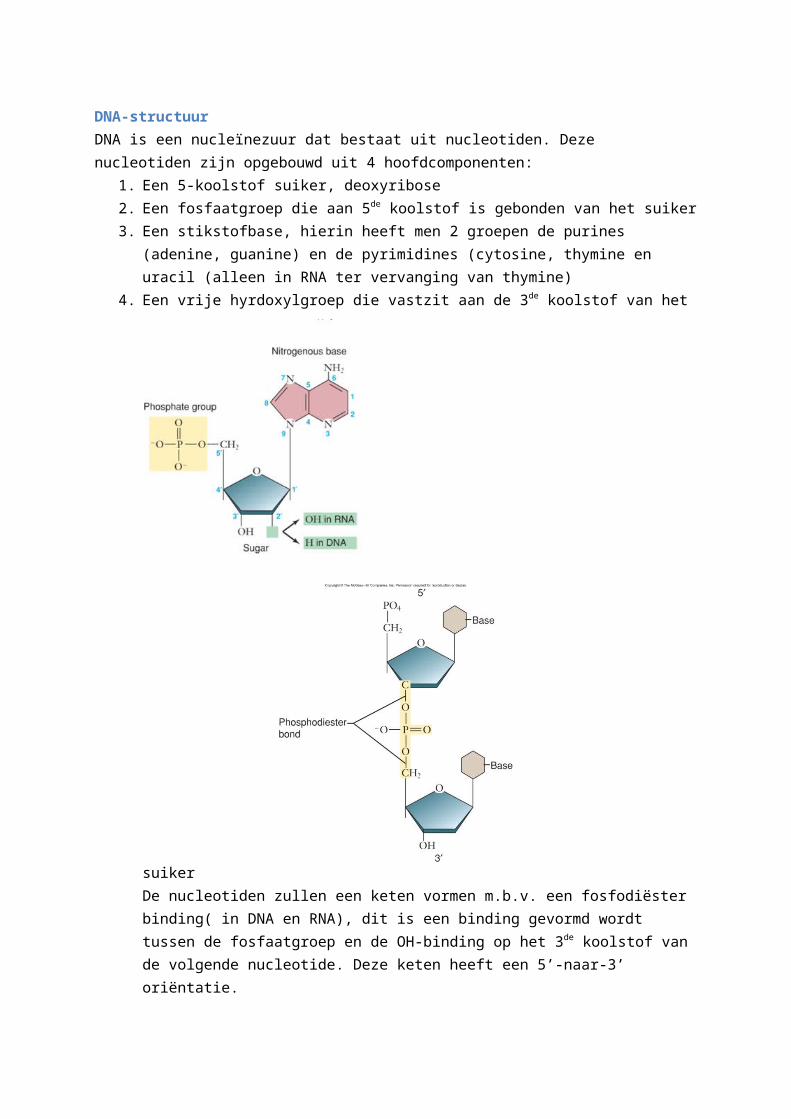
DNA-structuurDNA is een nucleïnezuur dat bestaat uit nucleotiden. Deze nucleotiden zijn opgebouwd uit 4 hoofdcomponenten:
1. Een 5-koolstof suiker, deoxyribose2. Een fosfaatgroep die aan 5de koolstof is gebonden van het suiker3. Een stikstofbase, hierin heeft men 2 groepen de purines (adenine, guanine) en de
pyrimidines (cytosine, thymine en uracil (alleen in RNA ter vervanging van thymine)4. Een vrije hyrdoxylgroep die vastzit aan de 3de koolstof van het suiker
De nucleotiden zullen een keten vormen m.b.v. een fosfodiëster binding( in DNA en RNA), dit is een binding gevormd wordt tussen de fosfaatgroep en de OH-binding op het 3de koolstof
van de volgende nucleotide. Deze keten heeft een 5’-naar-3’ oriëntatie.
Erwin Chargaff ontdekte dat DNA ondanks de weinige bouwstenen geen simpel herhalend polymeer is. Hij ontdekte wel een aantal regels inzake de stikstofbasen:
I. De hoeveelheid adenine is altijd gelijk aan de hoeveel thymineII. De hoeveelheid guanine is altijd gelijk aan de hoeveelheid cytosine
III. Hieruit volgt dat er altijd een gelijke verhouding van purines en pyrimidines is.

Rosalind Franklin voerde een X-straal diffractie om DNA te analyseren. Bij X-straal diffractie gaat men een molecule bombarderen met X-stralen, deze stralen gaan dan afbuigen door de moleculen die ze tegen komen waarna er een patroon kan worden gevonden op een fotografische film. Zo ontdekte ze dat DNA de vorm had van een helix, deze een diameter heeft van 2nm en elke 3,4 nm een volledige draai maakt.
In 1953 leidde Watson en Crick een structuur af voor DNA uit de vorige bevindingen. Ze komen tot een dubbele helix: de strengen zijn polymeren van nucleotiden die worden ondersteund door de fosfodiëster ruggegraat (herhalende suiker en fosfaatgroepen gebonden door de fosfodiëster binding). De strengen zijn antiparallel (1 keten van 5’ naar 3’ en de andere omgekeerd) en slingeren rond 1 as. Ze worden bij elkaar gehouden door de complementaire base ( A&T, G&C) wat zorgt voor een consistente diameter. Dit hebben ze kunnen afleiden zonder zelf 1 experiment te doen.
DNA replicatieBij replicatie moeten de dochterstrengen volledig gelijk zijn aan de parentale strengen. Hiervoor zijn drie modellen mogelijk die dit toelaten:
1. Conservatief: de parentale dubbele helix blijft intact en de nieuwe copies bestaan uit volledige nieuwe moleculen.
2. Semi-conervatief: hierbij worden de parentale strengen uit elkaar getrokken. Hierop wordt dan een complementaire streng gebouwd bestaande uit nieuwe moleculen. De dochtercellen bestaan dus uit 1 nieuwe streng en 1 parentale streng
3. Dispersief: kopieën van het DNA zou bestaan uit mengsels van parentale en nieuw gesynthetiseerde strengen
In 1958 voerde M. Meselson en F. Stahl een experiment uit om te achterhalen welk model klopte. Ze zouden DNA voor 2 replicaties volgen adhv zware stikstof (stikstofbasen). De eerste replicatie deden ze op een medium met de zware stof, de 2de replicatie gebeurde op een medium met normale stikstof. Ze extraheerden het DNA en centrifugeerde dit. Na centrifugatie bleek het semi-conservatieve model te kloppen want:
I. 2 dichtheden werden niet waargenomen na de eerste ronde m.a.w. het conservatieve model klopt niet
II. Na de 2 ronde had men wel 2 banden van verschillende dichtheid met de 2 dichtheid tussen die van zware en normale stikstof in
Om replicatie uit te voeren heeft men 3 dingen nodig: iets om te kopiëren (=het ouderlijk DNA), iets wat het kopiëren uitvoert (=enzymen) en bouwstenen om het kopiëren mogelijk te maken (=nucleotide fosfaten). Replicatie bestaat uit 3 opeenvolgende processen: initiatie: het begin van de replicatie, elongatie: de nieuwe DNA strengen worden aangemaakt, terminatie: de replicatie wordt beëindigt.De elongatie wordt mogelijk gemaakt door DNA polymerase, dit enzym paart de bestaande DNA-basen met de complementaire nucleotiden. DNA polymerase synthetiseert alleen in 5’-naar 3’ richting en om te beginnen moet er eerst een primer aangemaakt worden dat alleen kan door RNA polymerase.
Prokaryote replicatieEukaryoten hebben een enkel circulair DNA-streng. De replicatie begint aan de origin en stopt aan de de termination. De replicatie word uigevoerd door 2 polymerasen die in tegengestelde richting werken.
In E.coli heeft men 3 verschillende DNA polymerasen kunnen onderscheiden. Elk hebben ze een verschillende functie:
DNA polymerase 1: werkt op de achterliggende streng of lagging strand, het verwijderd de primers om de lagging strand en vervangt ze door DNA.
DNA polymerase 2: deze polymerase zorgt niet voor de aanmaak van een nieuwe streng maar staat in voor het herstel van DNA.
DNA polymerase 3: het is deze polymerase dat fungeert als hoofd replicatieënzyme. Het zorgt voor de aanmaak van het DNA op de parentale strengen.
Naast deze functie’s hebben de polymerasen ook bijkomende enzymactiviteiten, met name de eigenschap om fosfodiësterbindingen tussen nucleotiden te breken. Als dit gebeurt binnen het DNA zelf dan spreekt men van endonucleases, gebeurt dit op het einde van de DNA streng dan noemt men ze exonucleases. Pol 1, Pol 2 en Pol 3 hebben allemaal een exonuclease activiteit: dit gaat in de 3’-naar-5’ richting (omgekeerde richting van de synthese), hierbij gaan ze de streng dan proeflezen en de foute basen verwijderen. Pol 1 heeft ook nog een 5‘-naar-3’ exonuclease activiteit.
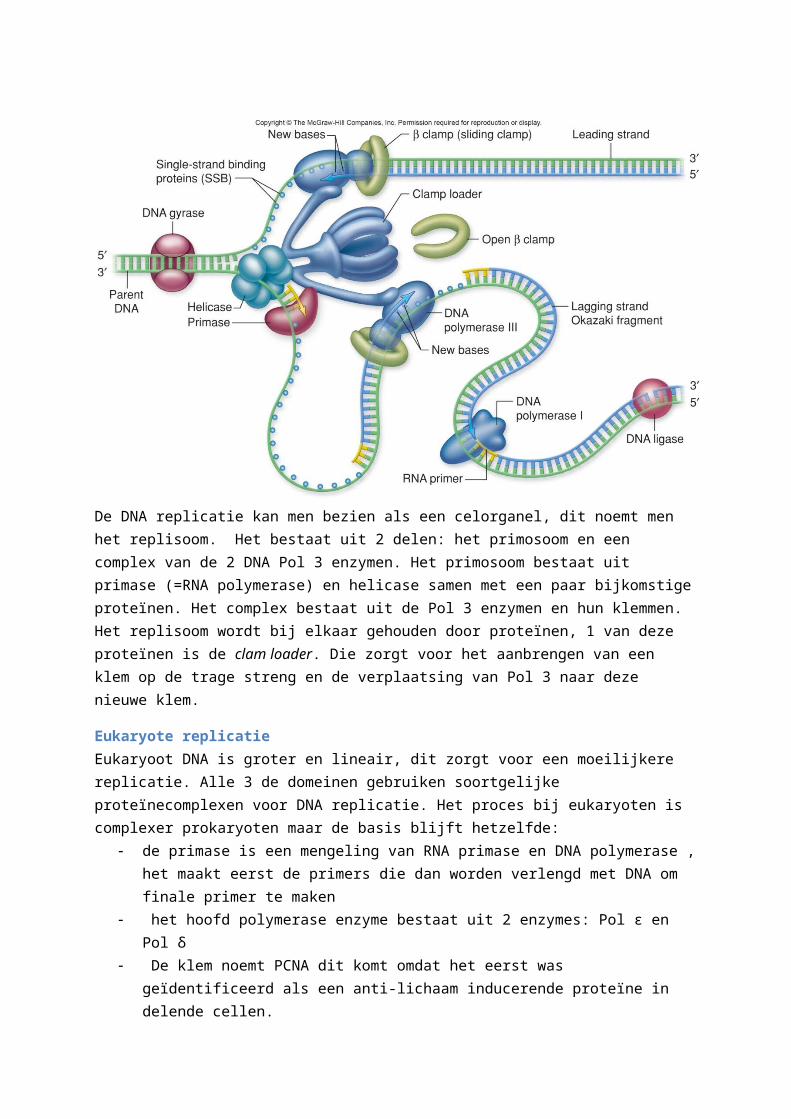
Om het DNA te repliceren moet het eerst ontwonden worden, dit word gedaan door de Helicasen die daarvoor ATP gebruiken. De enkele strengen van DNA die ontstaan door de ontwinding zijn onstabiel,dit komt omdat de hydrofobe basen worden blootgesteld aan het waterig milieu. Dit wordt opgelost doordat de cel het proteïne SSB (single-strand-binding) gebruikt om de strengen te bedekken en zorgen dat ze niet terug aan elkaar binden.De ontwinding van het DNA zorgt ook voor torsiespanning verderop in het DNA molecule. DNA is van nature al opgewonden en zal door de ontwinding verder opwinden, dit zorgt voor een effect genaamd supercoiling, dit noemt men ook de topologische toestand van DNA. Deze wordt tegen gegaan door het enzym topoisomerase. DNA gyrase zal dan verder helpen bij de replicatie.Door de antiparallelle bouw van DNA heeft bij de ontwinding een leading strand of leidende streng en lagging strand of trage streng. Bij de trage streng moet de DNA replicatie gebeuren in omgekeerde syntheserichting van polymerase (3’-naar-5’). Om de replicatie te beginnen wordt de helix gedeeltelijk geopend en vormt een replicatievork. DNA primase op RNA polymerase zal een primer aanmaken voor DNA polymerase, deze wordt later verwijderd en vervangen door DNA.
Synthese aan de leidende streng: op de primer zal DNA Pol 3 binden en beginnen aan de replicatie. Omdat Pol 3 gedurende de hele synthese blijft zitten heeft deze een speciale eigenschap: het vermogen van een polymerase om vast te blijven zitten aan de matrijs wordt processivity genoemd. Pol 3 blijft

op de streng zitten dankzij de β-subeenheid. Deze eenheid bestaat uit 2 identieke proteïnen die samen een cirkel vormen, het is deze cirkel dat zal fungeren als een klem om Pol 3 vast te houden. De structuur wordt de slinding clamp genoemd. Men noemt deze synthese continu.
Synthese op de trage streng:De synthese van DNA gebeurt hier in stukjes, deze stukjes DNA worden Okazaki fragmenten genoemd. Pol 2 zorgt voor de aanmaak van de Okazaki fragmenten. Voor elk Okazaki fragment word er een nieuwe primer gemaakt, dit gebeurt door Pol 1. Dit komt omdat het zijn 5’-naar-3’ exonuclease activiteit gebruikt om de primers te verwijderen om dan volgens normale synthese te vervangen door DNA. DNA ligase zorgt er dan voor dat de opening tussen de Okazaki fragmenten word gesloten.
De DNA replicatie kan men bezien als een celorganel, dit noemt men het replisoom. Het bestaat uit 2 delen: het primosoom en een complex van de 2 DNA Pol 3 enzymen. Het primosoom bestaat uit primase (=RNA polymerase) en helicase samen met een paar bijkomstige proteïnen. Het complex bestaat uit de Pol 3 enzymen en hun klemmen. Het replisoom wordt bij elkaar gehouden door proteïnen, 1 van deze proteïnen is de clam loader. Die zorgt voor het aanbrengen van een klem op de trage streng en de verplaatsing van Pol 3 naar deze nieuwe klem.
Eukaryote replicatieEukaryoot DNA is groter en lineair, dit zorgt voor een moeilijkere replicatie. Alle 3 de domeinen gebruiken soortgelijke proteïnecomplexen voor DNA replicatie. Het proces bij eukaryoten is complexer prokaryoten maar de basis blijft hetzelfde:
de primase is een mengeling van RNA primase en DNA polymerase , het maakt eerst de primers die dan worden verlengd met DNA om finale primer te maken

het hoofd polymerase enzyme bestaat uit 2 enzymes: Pol ε en Pol δ De klem noemt PCNA dit komt omdat het eerst was geïdentificeerd als een anti-lichaam
inducerende proteïne in delende cellen. Door de enorme hoeveel van data zou het een hele tijd duren eren de replicatie voltooid is,
dit lost de cel op doordat er meerdere origins zijn op de chromosomen. Een gevolg daarvan is dat er dus ook meerdere replicons zijn. De origins zijn niet zo speciefiek als bij eukaryoten, hun herkenning gebeurt dankzij chromatine structuur en volgorde van de aminozuren.
Op het einde van eukaryote chromosomen vindt men telomeren. Deze structuren beschermen de uiteinden tegen nucleasen en behouden de integriteit. Ze bestaan uit specifiek DNA gemaakt door telomerase. Dankzij de specifieke synthese richting van de polymerase, lineaire chromosomen en de primer worden de chromosomen elke deling korter. Dit komt omdat bij de trage streng aan het 3’ uiteinde een gat blijft nadat de primer verwijderd is. Telomerase gebruikt een eigen RNA-template om telomeren te maken. Dit heeft als voordeel dat het enzyme kleine stukjes enkelstrengig DNA kan maken, het replisoom maakt dan de andere streng. Dit complete stuk wordt dan aan het chromosoom gehangen en nu kan polymerase verder aanbouwen.

DNA herstelFouten in het DNA kunnen gebeuren in de replicatie, ze kunnen ook ontstaan door mutagenen: een stof die het DNA beschadigt en mutaties veroorzaakt (vb UV-licht, X-stralen,…). Ze worden tegen gegaan doordat de polymerasen kunnen proeflezen en doordat er herstelmechanismen zijn aangemaakt. Deze vallen in 2 categorieën:
Specifiek herstelHierbij gaan de systemen zich richten op een 1 mutatie en alleen deze herstellen.
1. Fotoherstel: dit is eenbeschadiging die wordt veroorzaakt door UV-licht. Het UV-licht zorgt ervoor dat dat 2 naburige thymines gaan binden met elkaar en een thymine dimeer vormen. Hetstel hiervan gebeurt door het enzym fotolyase. Dit enzym gaat boven het dimeer zitten en gebruikt dan licht uit het zichtbaarspectrum om energie te verzamelen. Deze energie word dan gebruikt om de thymines te breken en het DNA in zijn oorspronkelijke vorm te krijgen.
Niet-speciefiek herstelHierbij gebruikt de cel 1 systeem dat vele beschadigingen kan herstellen.
1. Excisieherstel: het beschadigde gebied wordt herstelt en verwijderd. Na verwijdering zal Pol 1 of Pol 2 zorgen voor vervanging van het verwijderde gebied.













Hoofdstuk 15
Hoofdstuk 16