
Radiogenomics voor predictie van radiotoxiciteit bij...
77
Radiogenomics voor predictie van radiotoxiciteit bij patiënten behandeld met radiotherapie voor H&N tumoren. Iris Rossaert Verhandeling ingediend tot het verkrijgen van de graad van Master in de Biomedische Wetenschappen Promotor: Prof. Dr. H. Thierens Vakgroep Medische Basiswetenschappen Academiejaar 2009-2010
Transcript of Radiogenomics voor predictie van radiotoxiciteit bij...

Radiogenomics voor predictie van
radiotoxiciteit bij patiënten behandeld met
radiotherapie voor H&N tumoren.
Iris Rossaert
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Prof. Dr. H. Thierens
Vakgroep Medische Basiswetenschappen
Academiejaar 2009-2010

“De auteur en de promotor geven de toelating deze scriptie voor consultatie beschikbaar te
stellen en delen ervan te kopiëren voor persoonlijk gebruik. Elk ander gebruik valt onder de
beperkingen van het auteursrecht, in het bijzonder met betrekking tot de verplichting
uitdrukkelijk de bron te vermelden bij het aanhalen van resultaten uit deze masterproef.”
Datum
HANDTEKENING STUDENT HANDTEKENING PROMOTOR Iris Rossaert Prof. Dr. H Thierens

Voorwoord Hierbij wil ik graag iedereen bedanken die mij bijgestaan heeft tijdens de realisatie van deze
thesis.
In de eerste plaats wil ik mijn promotor Prof. Dr. H. Thierens en Dr. Ir. Kim De Ruyck
bedanken voor de mogelijkheid die ze mij gaven om dit onderzoek uit te voeren.
In het bijzonder wil ik mijn begeleidster Joke Werbrouck bedanken voor alle hulp en steun
die ze mij bood bij het aanleren van de verschillende technieken en methoden. Bedankt ook
voor al de hulp bij de statistische verwerking en bij het schrijven van deze thesis. Naast Joke
wil ik ook Kim De Ruyck bedanken voor het aanleren van de nodige technieken gedurende
mijn stageperiode.
Verder wil ik ook Virginie De Gelder, Laurence Beels en Sofie De Langhe bedanken omdat
ook zij telkens klaar stonden voor praktische hulp in het labo.
Graag wil ik ook het volledige dienstpersoneel speciaal bedanken voor de aangename
werkomgeving. In het bijzonder bedank ik hierbij mijn mede-thesisstudenten Ans, Marlies,
Charlot en Siska. Bedankt voor al jullie hulp en steun en al de leuke momenten waardoor de
aangename sfeer, ook in meer stressvolle periodes nooit echt verdween.
Ten slotte wil ik al mijn vrienden en familie bedanken. Een extra dankwoord gaat uit naar
mijn ouders en vriend Daan waarbij ik altijd terecht kon, niet alleen dit jaar maar gedurende
mijn hele opleiding.
Bedankt, deze thesis was niet mogelijk zonder jullie hulp en steun!

Inhoudstafel Afkortingen ................................................................................................................................ 5
Samenvatting .............................................................................................................................. 1
Inleiding ..................................................................................................................................... 2
1. Hoofd- en halskanker ..................................................................................................... 2
1.1 Types Tumoren ........................................................................................................ 2
1.2 Behandeling ............................................................................................................. 3
1.2.1 Chirurgie, chemotherapie en cetuximab ........................................................... 3
1.2.2 Radiotherapie .................................................................................................... 3
2. Weefselreacties bij H&N kankerpatiënten na radiotherapie .......................................... 4
2.1 Introductie ................................................................................................................ 4
2.2 Types weefselreacties .............................................................................................. 5
2.2.1 Mucositis .......................................................................................................... 5
2.2.2 Dermatitis ......................................................................................................... 6
2.2.3 Xerostomie........................................................................................................ 6
2.2.4 Dysfagie ............................................................................................................ 6
2.2.5 Fibrose .............................................................................................................. 7
2.3 Het herstel van RT geïnduceerde DNA DSBs ......................................................... 7
A. Homologe recombinatie ................................................................................... 8
B. Niet homologe end-joining ............................................................................... 8
2.4 TGFβ1 ...................................................................................................................... 9
2.4.1 Inleiding ............................................................................................................ 9
2.4.2 Signaalpathway ................................................................................................. 9
2.5 SNPs en de ontwikkeling van normale weefselcomplicaties ................................. 10
2.5.1 SNPs ............................................................................................................... 10
2.5.2 TGFβ1 c.-509 C>T en TGFβ1 c.-800 G>A .................................................... 11
2.5.3 TGFβ1 c.74 G>C ............................................................................................ 11

2.5.4 Ku70 c.-1310 C>G .......................................................................................... 12
2.5.5 XRCC3 c.722 C>T .......................................................................................... 12
3. Doelstelling .................................................................................................................. 12
Materialen en Methoden ........................................................................................................... 14
1. Introductie .................................................................................................................... 14
2. Studiepopulatie ............................................................................................................. 14
3. Dosimetrische gegevens ............................................................................................... 15
4. DNA purificatie ............................................................................................................ 16
5. Concentratie -en kwaliteitbepaling van het DNA ........................................................ 17
6. Polymerase chain reaction (PCR) ................................................................................ 18
6.1 Principe .................................................................................................................. 18
6.2 PCR mix ................................................................................................................. 19
6.3 Primers ................................................................................................................... 19
6.4 PCR Controle ......................................................................................................... 20
7. SNP bepaling ................................................................................................................ 21
7.1 Restriction fragment length polymorphism (RFLP) .............................................. 21
7.2 High resolution melting (HRM) ............................................................................. 22
7.2.1 Primer bepaling............................................................................................... 24
7.2.2 Optimalisatie HRM protocols ......................................................................... 24
8. Kwaliteitscontroles ....................................................................................................... 25
9. Statistische Analyse ...................................................................................................... 25
Resultaten ................................................................................................................................. 26
1. Reeds geoptimaliseerde protocols ................................................................................ 26
2. Optimalisatie HRM Protocols ...................................................................................... 26
2.1 XRCC3 c.722C>T .................................................................................................. 27
2.2 Ku70 c.-1310C>G .................................................................................................. 28
2.3 Controlestalen ........................................................................................................ 31

3. Associatiestudie naar chronische radiosensitiviteit ...................................................... 31
3.1 Analyse genotype-onafhankelijke factoren ............................................................ 32
3.1.1. Therapiegerelateerde parameters .................................................................... 32
3.1.2. Patiëntgerelateerde parameters ....................................................................... 32
3.2 Analyse naar de associatie tussen SNPs en chronische radiosensitiviteit .............. 32
A. Chronische dysfagie ....................................................................................... 33
B. Xerostomie...................................................................................................... 33
C. Fibrose ............................................................................................................ 34
4. Associatiestudie naar acute radiosensitiviteit ............................................................... 35
4.1 Analyse genotype-onafhankelijke factoren ............................................................ 35
4.1.1 Therapiegerelateerde parameters .................................................................... 35
A. Acute dysfagie ................................................................................................ 35
B. Mucositis ........................................................................................................ 37
C. Dermatitis ....................................................................................................... 37
4.1.2 Patiëntgerelateerde parameters ....................................................................... 38
4.2 Analyse van de associatie van SNPs en acute radiosensitiviteit ............................ 38
A. Acute dysfagie ................................................................................................ 39
B. Mucositis ........................................................................................................ 39
C. Dermatitis ....................................................................................................... 40
Discussie ................................................................................................................................... 41
1. Optimalisatie HRM protocols ...................................................................................... 41
1.1 XRCC3 c.722 C>T ................................................................................................. 41
1.2 Ku70 c.-1310 C>G ................................................................................................. 41
2. Associatiestudie naar chronische en acute radiosensitiviteit ........................................ 41
2.1 Chronische normale weefselcomplicaties .............................................................. 42
2.1.1 XRCC3 c.722 C>T .......................................................................................... 42
2.1.2 Ku70 c.-1310 C>G .......................................................................................... 42

2.1.3 TGFβ1 c.-509 C>T ......................................................................................... 43
2.1.4 TGFβ1 c.-800 G>A......................................................................................... 43
2.1.5 TGFβ1 c.74G>C ............................................................................................. 44
2.2 Acute normale weefselcomplicaties ...................................................................... 44
2.2.1 XRCC3 c.722C>T ........................................................................................... 45
2.2.2 Ku70 c.-1310 C>G .......................................................................................... 46
2.2.3 TGFβ1 c.-509 C>T ......................................................................................... 46
2.2.4 TGFβ1 c.-800 G>A......................................................................................... 46
2.2.5 TGFβ1 c.74G>C ............................................................................................. 47
Besluit ....................................................................................................................................... 47
Referentielijst ........................................................................................................................... 48
Bijlage A: Algemene patiëntgegevens ........................................................................................ i
1. Patiëntdata voor analyse naar chronische toxiciteit ........................................................ i
2. Patiëntdata voor analyse naar acute toxiciteit ............................................................... iii
Bijlage B: CTC-scores ............................................................................................................... v
Bijlage C: Gebruikte RFLP protocols ...................................................................................... vii
Bijlage D: Resultaten van de genotypering ............................................................................. viii
Bijlage E: Geoptimaliseerde HRM protocols ........................................................................... xii
1. XRCC3 c.722C>T......................................................................................................... xii
2. Ku70 c.-1310C>G ....................................................................................................... xiii
Bijlage F: Onderzochte dosisparameters ................................................................................. xiv
1. Chronische weefselcomplicaties ................................................................................. xiv
2. Acute weefselcomplicaties ........................................................................................... xv
Bijlage G: SNP associatie ....................................................................................................... xvi
1. Chronische weefselcomplicaties ................................................................................. xvi
2. Acute weefselcomplicaties ......................................................................................... xvii

Afkortingen
BSA Bovine Serum Albumine
cAMP Cyclic Adenosine Monophosphate
CRE cAMP Responsive Elements
CREB cAMP Responsive Element-Binding
CTC Common Toxicity Criteria
CTCAE Common Terminology Criteria for Adverse Events
DD Dosis-de-escalatie
dH2O Dnase en Rnase vrij gedestilleerd water
DMSO DiMethylSulfOxide
DNA DeoxyRiboNucleicAcid
DNA-PK(cs) DNA-afhankelijke Proteïne Kinase katalytische subunit
dNTP DeoxyNucleotideTrisPhosphate
DrW Drinks/Week
ds DNA Dubbel streng DNA
DSB Dubbel Streng Breuken
DVH Dosis Volume Histogram
ECM ExtraCellulaire Matrix
EDTA EthyleenDiamineTetraAcetaat
EGFR Epidermal Growth Factor Receptor
EtBr EthydiumBromide
H&N Hoofd- en Hals (Head and Neck)
HE Heterozygoot
HK Heelkunde
HN Homozygoot Normaal
HNSCC Head and Neck Squamous Cell Carcinoma
HR Homologe Recombinatie
HRM High Resolution Melting
HV Homozygoot Variant
HWE Hardy-Weinberg Equilibrium
IMRT Intenstiteit gemoduleerde Radiotherapie (Intensity Modulated Radiation Therapy)
IR Infrarood
KT Kamertemperatuur
LAP Latency Associated Peptide
MgCl2 MagnesiumdiChloride
MLC Multileaf Collimator

NCBI National Center for Biotechnology Information
NHEJ Niet Homologe End-Joining
nt Nucleotide
Oligo Calc Oligonucleotide properties Calculator
OR Odds Ratio
PBS Phosphate Buffered Saline
PC Faryngeale constrictorspieren (Pharyngeal Constrictor muscles)
PCR Polymerase Chain Reaction
PEG Percutane Endoscopische Gastrostomie
PY Pack Years
QOL Levenskwaliteit (Quality Of Life)
RBC Rode Bloed Cellen
RE Restrictie-Enzym
RFLP Restriction Fragment Length Polymorphism
RNS Reactive Nitrogen Species
ROS Reactive Oxygen Species
rpm Rounds per minut
RT Radiotherapie
SNP Single Nucleotide Polymorfisme
SPSS Statistical Package for the Social Sciences
ss DNA Enkelstrengig DNA (Single Stranded DNA)
Ta Annealings Temperatuur
Taq Thermus aquaticus
TBE Tris Boraat EDTA2NA
TE Tris EDTA
TF TranscriptieFactor
TGFβ Transforming Growth Factor Beta
Tm Smelttemperatuur (melting Temperature)
Tris Tris-hydroxymethyl-aminomethaan
U Units
UES Upper Esophageal Sphincter
UV UltraViolet
UZG Universitair Ziekenhuis Gent
V Volt

1
Samenvatting
Radiotherapie (RT) is een belangrijk onderdeel van de behandeling van hoofd- en hals (H&N)
tumoren. Eén van de grote nadelen is dat ook een zeker volume gezond weefsel aanwezig is in
het bestralingsveld. De ontwikkeling van dermatitis, dysfagie en mucositis (acuut) en
xerostomie, fibrose en dysfagie (chronisch) vormt het grootste probleem bij de behandeling
van H&N tumoren met RT. In deze thesis werd onderzocht of bepaalde single nucleotide
polymorfisment (SNPs) geassocieerd zijn met de ontwikkeling van acute en chronische
weefselcomplicaties in een groep van hoofd- en hals kankerpatiënten behandeld met
intensiteit gemoduleerde radiotherapie (IMRT). Er werden SNPs onderzocht in DNA dubbel
streng breuk (DSB) herstelgenen (XRCC3 c.722C>T en Ku70 c.-1310C>G) en in het TGFβ1
gen (c.-509C>T, c.-800G>A, c.74G>C) wegens hun belangrijke rol in de weefselrespons na
de blootstelling aan ioniserende straling.
Van elk van de geïncludeerde patiënten werd DNA verkregen na isolatie uit lymfocyten of uit
vol bloed. De regio rond de te onderzoeken SNP werd via Polymerase Chain Reaction (PCR)
geamplificeerd. De uiteindelijke SNP bepaling werd uitgevoerd via Restriction Fragment
Length Polymorphism (RFLP) voor de SNPs in de TGFβ1 genen of via High Resolution
Melting (HRM) analyse voor de SNPs in de DNA DSB herstel genen. Gedurende deze thesis
werden de HRM protocols voor XRCC3 c.722C>T en Ku70 c.-1310C>G geoptimaliseerd.
Voor het onderzoek naar chronische weefselcomplicaties (fibrose, xerostomie en dysfagie)
werd een groep van 61 H&N kankerpatiënten geïncludeerd. Binnen deze groep bleek het
TGFβ1 c.-800 GA genotype significant geassocieerd te zijn met een verhoogd risico op de
ontwikkeling van ernstige fibrose.
Met betrekking tot acute weefselcomplicaties (dermatitis, mucositis en dysfagie) werd een
groep van 79 nieuw verzamelde H&N kankerpatiënten geanalyseerd. De TGFβ1 c.74G>C
SNP werd geassocieerd met een verhoogd risico bij de ontwikkeling van ernstige mucositis.
Deze bevinding was bij de aanwezigheid van 1 of 2 variante allelen significant. In
tegenstelling tot de chronische complicaties, verklaren de dosisparameters een deel van de
acute stralingstoxiciteiten. De gemiddelde dosis (Dmean) op de skin5mm, bovenste faryngeale
constrictorspier (SPC), mondholte en mucosa spelen een rol in de ontwikkeling van
dermatitis, dysfagie en mucositis respectievelijk. Concomitante chemotherapie verhoogt
significant het risico op de ontwikkeling van ernstige acute dysfagie en blijkt een risicofactor
voor de ontwikkeling van ernstige mucositis.

Inleiding
2
Inleiding
1. Hoofd- en halskanker
Hoofd- en halskanker (H&N kanker) is met een wereldwijde incidentie van 650000 gevallen
per jaar, een relatief veel voorkomende kanker (6% van alle kankers wereldwijd) [1]. De
incidentie van H&N kanker stijgt met de leeftijd en bereikt een maximum rond de leeftijd van
50 jaar [2]. Dit type kanker komt frequenter voor bij mannen dan bij vrouwen (man:vrouw
ratio van 4-5:1) [3].
1.1 Types Tumoren
H&N kanker is een verzamelnaam voor een heterogene groep tumoren die voornamelijk
ontstaan in de mondholte, farynx (nasofarynx, orofarynx, hypofarynx) en larynx (supraglottis,
glottis, subglottis) (figuur 1.1). De identificatie van de anatomische locatie is belangrijk voor
de klinische classificatie van de tumoren en bijgevolg voor de gebruikte behandelingsmethode
[4].
Figuur 1.1: Anatomie van de H&N regio. Bron: Vokes et al. [4]
De tumoren kunnen verder gekarakteriseerd worden door hun verschillende histologische
oorsprong. Het grootste deel van de tumoren (85%) zijn de epidermoid epitheliomas of
squameuze carcinomas (HNSCC). Deze ontstaan in de cellen die de muceuze oppervlaktes
aflijnen ter hoogte van het respiratoir en digestieve stelsel, namelijk de plaveiselcellen. Eerder
zeldzame histologische varianten zijn de verruceuze, basaloide, papillaire, adenoid, spoelcel -
en adenocarcinoma [5].

Inleiding
3
1.2 Behandeling
1.2.1 Chirurgie, chemotherapie en cetuximab
Chirurgie is vooral belangrijk bij de behandeling van resecteerbare tumoren ter hoogte van de
larynx, hypopharynx en oropharynx. De resectie van tumorweefsel gaat echter dikwijls
gepaard met een verlies aan gezond weefsel en/of functionaliteit van bepaalde weefsels. Na
chirurgie kunnen de patiënten o.a. praat- en slikproblemen ontwikkelen zoals het geval is bij
een volledige laryngectomie. Na chirurgie zal meestal een bijkomende adjuvante therapie
(radiotherapie, RT en/of chemotherapie) volgen om eventueel niet-waarneembare uitzaaiingen
te bestrijden en het risico op herval te vermijden [7].
Het standaard chemotherapeuticum dat gebruikt wordt bij de behandeling van H&N kanker is
het platinumderivaat cisplatinum [1]. Cisplatinum interageert met het DNA van sneldelende
(kanker)cellen. Indien de DNA schade niet hersteld wordt, zal dit aanleiding geven tot
geprogrammeerde celdood (apoptose) van deze cellen [8]. Door de systemische werking van
chemotherapie, kan deze therapie gepaard gaan met complicaties zoals haarverlies,
misselijkheid en anemie. Meestal wordt bij de behandeling van HNSCC chemotherapie in
combinatie met RT (concomitant) toegediend. De combinatie van beide cytotoxische agentia
leidt tot een meer efficiënte behandeling H&N tumoren [9].
Sinds kort werd bij de behandeling van H&N kanker ook target gerichte therapie
geïntroduceerd. Het standaard moleculair target agent dat hierbij wordt gebruikt is cetuximab.
Cetuximab is een monoclonaal antilichaam gericht tegen de epidermale groei factor receptor
(EGFR). Dit antilichaam wordt ook meestal in combinatie gebruikt met RT en zal het
cytotoxisch effect van de bestraling bevorderen wat leidt tot een verhoogde overleving en
tumorcontrole [10]. Een belangrijk nadeel van deze therapie is het optreden van dermatitis dat
in sommige gevallen zeer ernstige vormen kan aannemen. Giro et al. toonden een ernstige
huidtoxiciteit aan in 50% van de behandelde patiënten [11].
1.2.2 Radiotherapie
RT is een algemeen gebruikte techniek bij de behandeling van H&N kanker. Een standaard
RT protocol bestaat uit een gefractioneerde dosistoediening van 2Gy/fractie 5dagen/week
gedurende 7 weken [7]. RT is, in tegenstelling tot chirurgie, geen invasieve techniek. In
vergelijking met chemotherapie heeft RT het grote voordeel dat het een eerder lokale
behandeling is die geen systemische bijwerkingen teweegbrengt. Bij RT worden ioniserende
stralen (IR) gebruikt om de tumorcel zodanig te beschadigen dat ze afsterft. Met de

Inleiding
4
vooruitgang in de gebruikte technologieën wordt de stralingsbundel voornamelijk gericht op
het carcinogeen weefsel waarbij het gezond weefsel een zo laag mogelijke stralingsdosis
krijgt. Naast het tumorweefsel is ook een zeker volume van het omliggende gezonde weefsel
onvermijdelijk in de stralingsbundel aanwezig, waardoor lokaal ongewenste complicaties van
het normale weefsel kunnen optreden [12]. Deze complicaties waren de basis voor het
onderzoek naar een alternatieve techniek waarbij de dosis ter hoogte van het normale weefsel
werd geminimaliseerd: de intensiteit gemoduleerde RT (IMRT). Bij deze techniek worden
meerdere stralingsbundels gebruikt waarbij de intensiteit van de bundels met behulp van
multileaf collimatoren (MLCs) zodanig gemoduleerd wordt dat er een reductie optreedt van
de dosis ter hoogte van het gezonde weefsel, terwijl het tumorweefsel de vereiste hogere
homogene dosis krijgt [13].
2. Weefselreacties bij H&N kankerpatiënten na radiotherapie
2.1 Introductie
Zoals reeds vermeld in sectie 1.3.1 kan bij de behandeling van H&N tumoren met IMRT ook
het gezond weefsel ernstig beschadigd worden waardoor ongewenste weefselreacties kunnen
ontstaan [12,14]. Omdat tumoren behorend tot de groep van H&N kanker, gelegen zijn in een
functioneel kritische regio, kunnen weefselcomplicaties in deze omgeving een sterke daling
van de levenskwaliteit (QOL) veroorzaken. Zo kunnen na bestraling bepaalde fysiologische
functies, zoals de eet- en spraakmogelijkheid, en het uitzicht van de patiënt beïnvloed worden
[7,15].
Bij de behandelde patiënten wordt onderling een grote variabiliteit gezien in de ernst van de
bijwerkingen, gaande van bijna afwezig tot zeer ernstig. Dit wordt zelfs gezien bij patiënten
die een gelijkaardig behandelingsschema ondergingen. De patiënten krijgen voor de
verschillende weefselreacties een score, gaande van 1 tot 5, afhankelijk van de ernst van de
toxiciteit. Hierbij wordt gebruik gemaakt van de Common Terminology Criteria for Adverse
Events v3.0 (CTCAEv3.0) schaal [16].
De variabiliteit in ernst van bijwerkingen tussen de patiënten wordt deels (±30%) verklaard
door therapiegerelateerde factoren (de gebruikte dosis, het bestraalde weefselvolume, de
fractionatie,…) en patiëntspecifieke factoren (rookgedrag1, geslacht, leeftijd en
alcoholgebruik2). Uit verschillende studies betreffende zeldzame genetische syndromen zoals
1 Het rookgedrag wordt uitgedrukt in pack-years (PY): (aantal sigaretten per dag/20) x aantal jaren gerookt. 2 Het alcoholgebruik wordt uitgedrukt in drinks/week (DrW) ongeacht het soort drank (bier, wijn, sterke drank).

Inleiding
5
Ataxia Telangiectasia en het Nijmengen Breakage Syndroom werd reeds gesuggereerd dat de
genetische achtergrond ook aan de basis kan liggen van de inter-individuele verschillen in
stralingsgevoeligheid. Deze syndromen worden enerzijds gekenmerkt door mutaties in genen
betrokken in de detectie van DNA schade en herstel en anderzijds door een extreem
verhoogde stralingsgevoeligheid van de patiënten [17,18]. Voor de grootste met groep RT-
behandelde patiënten (±70%) wordt dus aangenomen dat ook de genetische achtergrond een
belangrijke rol speelt voor de variabiliteit in ernst van de optredende weefselreacties [14]. Het
onderwerp van deze thesis kadert in deze problematiek.
Wegens het optreden van ernstige bijwerkingen bij een niet te verwaarlozen aantal patiënten,
wordt bij het bepalen van de tumordosis rekening gehouden met de tolerantiedosis van de
normale weefsels aanwezig in het bestralingsveld. Hierbij wordt een maximale vernietiging
van de tumorcellen met een aanvaardbaar risico op complicaties nagestreefd [12,19].
De weefselcomplicaties kunnen onderverdeeld worden in 3 groepen. Enerzijds heeft men de
acute weefselcomplicaties, deze treden typisch op tijdens of enkele weken na RT en zijn
voornamelijk het resultaat van depletie van parenchymale cellen (hypoplasie) en beschadiging
van stamcellen waardoor de normale turn-over van het weefsel verstoord is [20]. Indien de
complicaties zich maanden (≥6maanden) tot jaren na de therapie ontwikkelen spreekt men
over late (chronische) weefselcomplicaties [21]. Als derde groep onderscheidt men ook nog
de consequentiële late effecten. Dit zijn ‘late’ weefselreacties die ontstaan uit voormalige,
persisterende acute complicaties [22]. Voor deze scriptie werden enkel de acute en late
toxiciteiten beschouwd.
2.2 Types weefselreacties
2.2.1 Mucositis
Mucositis is een inflammatiereactie van de slijmvliezen die ontstaat tijdens of na bestraling
door een beschadiging en verlies van de mucosale membranen en het onderliggende zachte
weefsel [23]. Bijna alle H&N kankerpatiënten die behandeld worden met RT (98%)
ontwikkelen mucositis. Hierbij ontwikkelt 28% van deze patiënten een zeer ernstige vorm
(CTC 3-4) [24]. Het klinisch beeld wordt gekenmerkt door het optreden van erytheem,
ulceraties en pijn. Deze kunnen enerzijds leiden tot eet-, slik- en praatmoeilijkheden wat de
QOL van de patiënten ondermijnt. Anderzijds is mucositis de meest significante
dosislimiterende weefselcomplicatie. De symptomen zijn dikwijls zo ernstig dat de

Inleiding
6
stralingsdosis moet gereduceerd worden of de therapie (tijdelijk) moet worden stopgezet.
Hierdoor heeft mucositis ook een indirect effect op de tumor respons [25].
2.2.2 Dermatitis
Dermatitis is een inflammatie van de huid. Bij bestraling kunnen de radiosensitieve
progenitorcellen in de basale laag van de epidermis beschadigd worden en afsterven. Hierdoor
worden de gedesquameerde cellen ter hoogte van het epitheel niet meer vervangen, waardoor
de epidermis geleidelijk aan ‘verdwijnt’ en de dermis bloot komt te liggen. Typische
kenmerken van dermatitis zijn huiderytheem, hyperpigmentatie, haarverlies, droge en
vochtige desquamatie [23].
2.2.3 Xerostomie
Xerostomie of verminderde en meer visceuze speekselproductie ontstaat door bestraling van
de speekselklieren. De speekselproductie kan hierbij dalen tot meer dan 50% na 1 week, en
tot 90% tijdens de 2e week van de behandeling [23]. Het is een frequent voorkomende
toxiciteit die voornamelijk als chronisch beschreven wordt. Belangrijke gevolgen van
xerostomie zijn een verminderde orale hygiëne, periodieke microbiële infecties,
moeilijkheden bij het slikken en kauwen met wijziging van het dieet als gevolg, cariës en een
verminderde smaak wegens aantasting van de tong [26]. Alhoewel het gebruik van IMRT al
geresulteerd heeft in een betere sparing van de parotisklieren blijft xerostomie nog steeds een
belangrijke complicatie [23].
2.2.4 Dysfagie
De meest kritische weefselreactie na RT bij H&N kankerpatiënten is dysfagie
(slikmoeilijkheden) waarbij het normale eetpatroon van de patiënt in het gedrang kan komen.
Dysfagie kan zowel acuut als laat optreden. Kritische structuren bij het ontstaan van dysfagie
zijn de circulaire faryngeale constrictorspieren (PCs). Deze zijn belangrijk in de de faryngeale
fase van het slikproces (propulsie van voedsel). Dysfunctie van de PCs na RT voor H&N
kankerpatiënten werd reeds beschreven als de belangrijkste oorzaak voor het optreden van
dysfagie [27]. Degeneratie en verlies van elasticiteit van de PCs kan leiden tot nutritionele
deficiëntie met gewichtsverlies en langdurige sondevoeding (percutane endoscopische
gastrostomie, PEG). Bij het ontstaan van aspiratiepneumonie3 kan dysfagie zelfs
3 Aspiratie van bepaalde stoffen zoals voedsel waardoor het longweefsel ontsteekt.

Inleiding
7
levensbedreigend zijn [28]. Bij de meeste patiënten keert de normale slikfunctie ongeveer 3
maanden na de therapie terug. Een bepaalde groep patiënten ontwikkelen de slikproblemen
pas jaren na de behandeling. De voortdurende cascade van cytokines, hypoxie en chronische
oxidatieve stress resulteert in fibrotisch weefsel en stramheid. Samen met oedeem, atrofie en
neuronale schade kan dit leiden tot functieverlies en chronische dysfagie als gevolg [29].
2.2.5 Fibrose
Fibrose is een belangrijke late weefselcomplicatie na concomitante RT en wordt beschouwd
als een nooit genezende wonde. Tijdens een normale wondheling worden fibroblasten
transiënt geactiveerd tot myofibroblasten, wat leidt tot proliferatie en collageenafzetting. Na
wondheling zullen deze myofibroblasten in apoptose gaan. Tijdens een fibrotische reactie is
deze feedback controle afwezig wat resulteert in chronisch geactiveerde myofibroblasten,
overmatige collageenafzetting en extracellulaire matrix (ECM) productie. Hierdoor wordt het
normale weefsel atrofisch en vervangen door littekenweefsel. Een mogelijke oorzaak van
deze chronische cellulaire activatie is een stralingsgeïnduceerde verandering in de expressie
van bepaalde cytokines, chemokines en groeifactoren door macrofagen, endotheelcellen en
fibroblasten [30].
2.3 Het herstel van RT geïnduceerde DNA DSBs
IR zoals gebruikt bij RT, kunnen verschillende types van DNA schade veroorzaken
waaronder enkelstrengbreuken, dubbel streng breuken (DSB) en beschadiging van basen.
Hiervan zijn de DSBs voornamelijk verantwoordelijk voor het optreden van de
radiobiologische effecten. Niet herstelde DSBs zijn namelijk kritisch voor het optreden van
stralingsgeïnduceerde celdood. Een haperende of volledig deficiënte respons op de inductie
van dit type breuken kan de stralingsgevoeligheid van de cellen en als gevolg de gevoeligheid
voor ernstige normale weefselcomplicaties bepalen [31].
De twee belangrijkste pathways voor het herstel van DSBs zijn de homologe recombinatie
(HR) en de niet-homologe end Joining (NHEJ). Terwijl in mammalia de NHEJ voornamelijk
tussenkomt bij het DNA herstel in de G1 en M fase van de celcyclus, is de HR de
belangrijkste pathway voor DSB herstel tijdens de S en G2 fase. Een schematisch overzicht
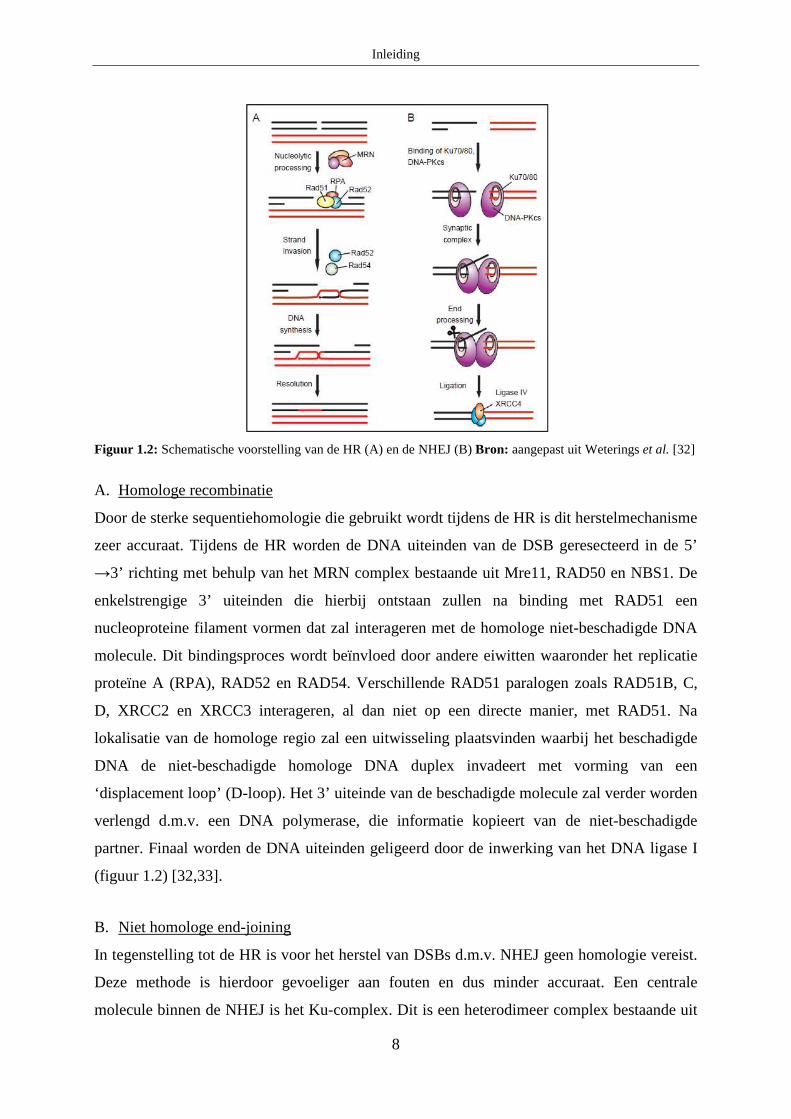
van de HR en NHEJ is weergegeven in figuur 1.2 [32].
Inleiding
8
Figuur 1.2: Schematische voorstelling van de HR (A) en de NHEJ (B) Bron: aangepast uit Weterings et al. [32]
A. Homologe recombinatie
Door de sterke sequentiehomologie die gebruikt wordt tijdens de HR is dit herstelmechanisme
zeer accuraat. Tijdens de HR worden de DNA uiteinden van de DSB geresecteerd in de 5’
→3’ richting met behulp van het MRN complex bestaande uit Mre11, RAD50 en NBS1. De
enkelstrengige 3’ uiteinden die hierbij ontstaan zullen na binding met RAD51 een
nucleoproteine filament vormen dat zal interageren met de homologe niet-beschadigde DNA
molecule. Dit bindingsproces wordt beïnvloed door andere eiwitten waaronder het replicatie
proteïne A (RPA), RAD52 en RAD54. Verschillende RAD51 paralogen zoals RAD51B, C,
D, XRCC2 en XRCC3 interageren, al dan niet op een directe manier, met RAD51. Na
lokalisatie van de homologe regio zal een uitwisseling plaatsvinden waarbij het beschadigde
DNA de niet-beschadigde homologe DNA duplex invadeert met vorming van een
‘displacement loop’ (D-loop). Het 3’ uiteinde van de beschadigde molecule zal verder worden
verlengd d.m.v. een DNA polymerase, die informatie kopieert van de niet-beschadigde
partner. Finaal worden de DNA uiteinden geligeerd door de inwerking van het DNA ligase I
(figuur 1.2) [32,33].
B. Niet homologe end-joining
In tegenstelling tot de HR is voor het herstel van DSBs d.m.v. NHEJ geen homologie vereist.
Deze methode is hierdoor gevoeliger aan fouten en dus minder accuraat. Een centrale
molecule binnen de NHEJ is het Ku-complex. Dit is een heterodimeer complex bestaande uit

Inleiding
9
twee subunits: Ku70 (XRCC6) en Ku80 (XRCC5). Dit heterodimeer kan d.m.v. zijn open
ringstructuur binden aan de beide DNA-uiteinden die ontstaan bij de DSB inductie. De
binding van het heterodimeer beschermt enerzijds de uiteinden tegen verdere degradatie en
zorgt er anderzijds voor dat andere componenten van de NHEJ de DSB kunnen bereiken.
Door binding van het Ku-complex wordt het DNA-afhankelijke proteïne kinase katalytische
subunit (DNA-PKcs) aangetrokken en vormt dit samen met het heterodimeer het DNA-
afhankelijk proteïne kinase (DNA-PK; XRCC7). Dit kinase is eveneens een sleutelmolecule
in de NHEJ en zal enkel functioneel zijn in associatie met het DNA gebonden Ku-eiwit. Naast
het samenbrengen van de DNA uiteinden zal dit kinase ook instaan voor de fosforylatie van
bepaalde targetmoleculen waaronder XRCC4. XRCC4 vormt een complex met Ligase IV dat
verantwoordelijk is voor de ligatie van de DNA uiteinden. Vooraleer deze ligatie kan
gebeuren, worden de DNA uiteinden bewerkt tot ligeerbare uiteinden. In dit proces zijn
verschillende nucleasen en polymerasen betrokken [32,33].
2.4 TGFβ1
2.4.1 Inleiding
Zoals eerder vermeld is fibrose één van de voornaamste late weefselcomplicaties na RT. Het
ontstaan van fibrotisch weefsel ter hoogte van de H&N regio kan aanleiding geven tot de
ontwikkeling van late slikproblemen. Een sleutelmolecule bij het ontstaan van fibrose is
transforming growth factor β (TGFβ). TGFβ is een multifunctionele cytokine dat betrokken is
bij drie belangrijke biologische functies. Naast zijn functie bij de aanmaak van ECM en
collageen, speelt de molecule ook een rol in de regulatie van celgroei-inhibitie en oefent hij
een immuunsuppressieve activiteit uit [30]. Van de 3 gekende isovormen (TGFβ1-3) is
TGFβ1 de dominant voorkomende in humaan plasma (>95%) [34].
2.4.2 Signaalpathway
TGFβ wordt gesecreteerd als een inactief, latent complex. Pas na klieving van het ‘latency
associated peptide’ (LAP) kan TGFβ binden aan zijn functionele receptor (TGFβR1-
TGFβR2). IR is één van de weinige exogene factoren die een activatie van TGFβ kan
induceren (figuur 1.3). IR kan enerzijds zorgen voor een directe activering van TGFβ door
dissociatie van het LAP. Anderzijds leidt blootstelling aan IR tot beschadiging van
endotheliale cellen wat op zich leidt tot een vrijstelling van pro-fibrotische cytokines
waaronder TGFβ. Ook reactieve zuurstof species (ROS) en reactieve stikstof radicalen (RNS),
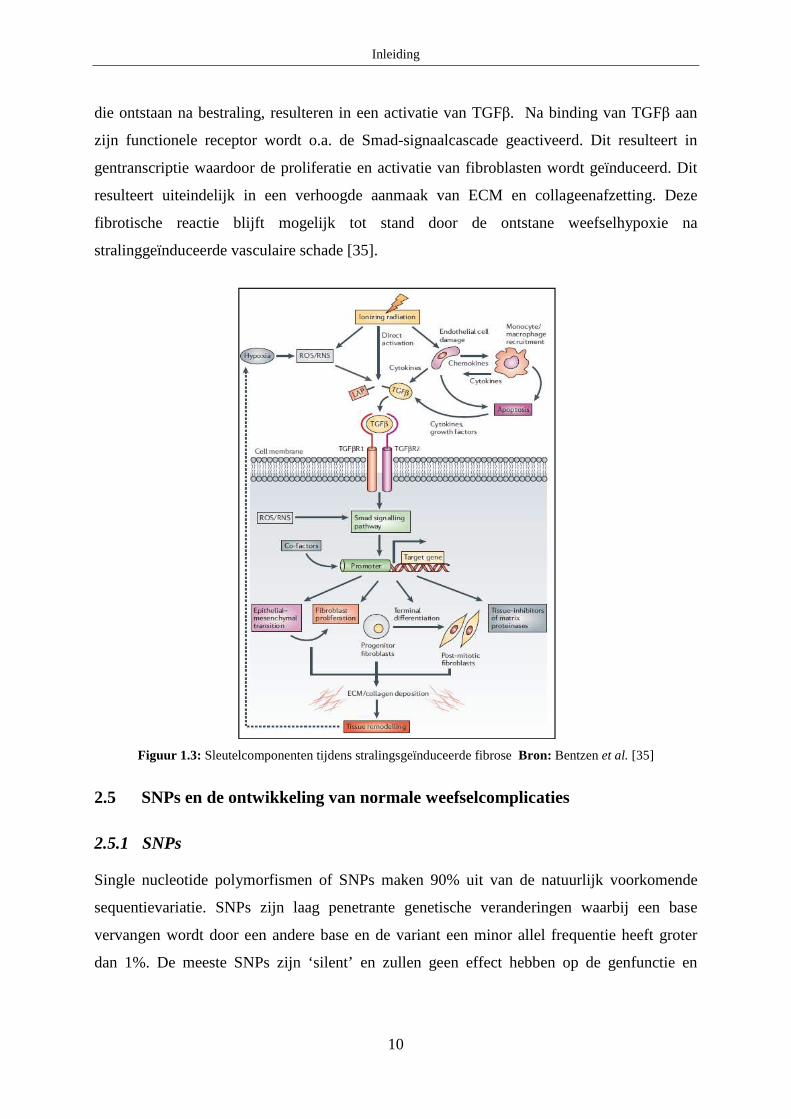
Inleiding
10
die ontstaan na bestraling, resulteren in een activatie van TGFβ. Na binding van TGFβ aan
zijn functionele receptor wordt o.a. de Smad-signaalcascade geactiveerd. Dit resulteert in
gentranscriptie waardoor de proliferatie en activatie van fibroblasten wordt geïnduceerd. Dit
resulteert uiteindelijk in een verhoogde aanmaak van ECM en collageenafzetting. Deze
fibrotische reactie blijft mogelijk tot stand door de ontstane weefselhypoxie na
stralinggeïnduceerde vasculaire schade [35].
Figuur 1.3: Sleutelcomponenten tijdens stralingsgeïnduceerde fibrose Bron: Bentzen et al. [35]
2.5 SNPs en de ontwikkeling van normale weefselcomplicaties
2.5.1 SNPs
Single nucleotide polymorfismen of SNPs maken 90% uit van de natuurlijk voorkomende
sequentievariatie. SNPs zijn laag penetrante genetische veranderingen waarbij een base
vervangen wordt door een andere base en de variant een minor allel frequentie heeft groter
dan 1%. De meeste SNPs zijn ‘silent’ en zullen geen effect hebben op de genfunctie en

Inleiding
11
expressie. Een klein deel van de SNPs echter zijn gelegen in (exon) en rondom (o.a.
promotor) gensequenties en kunnen zo de expressie en functie van deze beïnvloeden [36,37].
SNPs in kritische regio’s van genen betrokken in het herstel van DSBs en in TGFβ1 kunnen
dus de werking van de resulterende proteïnen en cytokine beïnvloeden. Hierdoor kunnen
SNPs aan de basis liggen van het optreden van variaties in de ernst van normale
weefselcomplicaties na RT. Er zijn heel wat proteïnen betrokken bij DSB herstel en in de
TGFβ pathway. Er werden 2 SNPs in DSB herstelgenen (Ku70 c.-1310 en XRCC3 c.722) en 3
SNPs in het TGFβ1 gen geselecteerd.
2.5.2 TGFβ1 c.-509 C>T en TGFβ1 c.-800 G>A
TGFβ1 c.-509C>T (of c.-1347 C>T) en TGFβ1 c.-800G>A (of c.-1638 G>A) zijn SNPs
gelegen in de promotor regio van het TGFβ1 gen. SNPs in de promotor regio van een gen
kunnen de expressie van dit gen veranderen en hierdoor de concentratie van het circulerend
eiwit beïnvloeden. Dit werd aangetoond voor het TGFβ1 c.-509 HV genotype (TT) in de
studie van Grainger et al. waarin dit genotype significant geassocieerd werd met een
verhoging van de circulerende TGFβ1 concentratie [38]. Reeds meerdere studies met
betrekking tot verschillende tumoren beschreven de TGFβ1 c.-509 SNP als een risicofactor
bij de ontwikkeling van late weefselcomplicaties na behandeling met RT [20,39-41]. Ondanks
deze resultaten bestaat er geen consensus over de invloed van de SNP op de functie van het
resulterende proteïne [42-44].
De TGFβ1 c.-800G>A SNP is gelegen in een consensus gebied van de CREB4 (cAMP
responsive element-binding) familie van transcriptiefactoren (TF) waardoor deze SNP
mogelijks een invloed heeft op de transcriptionele functionaliteit van deze TF [38]. In de
literatuur verschenen slechts een beperkt aantal artikels die de associatie van deze SNP en de
ontwikkeling van weefselcomplicaties na RT onderzochten [39,41].
2.5.3 TGFβ1 c.74 G>C
De TGFβ1 c.74 G>C SNP is een SNP gelegen in exon 1 van het TGFβ1 gen. Dit exon bevindt
zich in de signaal peptide van het TGFβ1 eiwit waardoor deze SNP het transport van het eiwit
doorheen het endoplasmatisch reticulum zou kunnen beïnvloeden [39]. Uit de in vitro studie
van Awad et al. bleek dat homozygositeit voor het normale allel van deze SNP gepaard gaat
met een hogere productie van TGFβ1 [45]. Onderzoek van deze SNP en de ontwikkeling van 4 CREB is een transcriptiefactor die bindt aan specifieke DNA sequenties, meerbepaald de cAMP responsive elements (CRE) .

Inleiding
12
weefselcomplicaties na RT bij H&N kankerpatiënten werd tot nog toe niet onderzocht. Ook
bij andere kankertypes werd geen associatie gevonden [39-41].
2.5.4 Ku70 c.-1310 C>G
Zoals blijkt uit sectie 2.3.B is Ku70 een centrale molecule binnen de NHEJ. Samen met het
Ku80 eiwit vormt hij een heterodimeer complex dat vroeg in NHEJ-signaalcascade bindt aan
de DNA uiteinden die ontstaan na de inductie van een DSB. De Ku70 c.-1310 C>G SNP is
een SNP gelegen in de promotorregio van het Ku70 gen. Reeds verschillende studies toonden
aan dat de afwezigheid of verminderde aanwezigheid van het Ku70 eiwit in cellijnen de
stralingsgevoeligheid van deze cellen verhoogt [46,47]. Een SNP in de promotorregio van dit
gen zou de concentratie van Ku70 en aldus de stralingsgevoeligheid kunnen beïnvloeden. In
de literatuur werd nog maar weinig onderzoek verricht met betrekking tot deze SNPs en
radiosensitiviteit [48 19].
2.5.5 XRCC3 c.722 C>T
XRCC3 is één van de 5 RAD51 paralogen dat interageert met RAD51 bij zijn functie in de
HR (sectie 2.3.A) [33]. De XRCC3 c.722 C>T SNP is gelegen in codon 241 van het gen. Het
is een SNP die een aminozuurwijziging (Thr-Met) in het XRCC3 eiwit teweegbrengt. Deze
SNP bevindt zich in de nabijheid van het Phe249 residu. Dit residu is een essentieel
aminozuur ter hoogte van de RAD51-bindingsplaats. De aanwezigheid van de SNP zou de
interactie tussen XRCC3 en RAD51kunnen beïnvloeden [49].
De associatie van de XRCC3 c.722 SNP en de ontwikkeling van acute weefselcomplicaties
werd net als de voornoemde Ku70 SNP binnen de eigen onderzoeksgroep onderzocht in een
populatie H&N kankerpatiënten [19]. Met betrekking tot andere types tumoren werd ook in
andere studies onderzoek gedaan naar de associatie van deze SNP en de ontwikkeling van
acute en late weefselcomplicaties na RT. Met betrekking tot de resultaten van deze
onderzoeken werd echter geen consensus gevonden [40,43, 50-52].
3. Doelstelling
De ernstige acute en late weefselcomplicaties die optreden bij een bepaalde subgroep RT-
behandelde H&N kankerpatiënten behandeld met RT vormen een groot probleem. Deze
weefseltoxiciteiten kunnen resulteren in een verhoogde morbiditeit en mortaliteit en een
verminderde QOL [23]. Bij het optreden van weefselcomplicaties bij de behandelde H&N

Inleiding
13
kankerpatiënten ziet men een variabiliteit in de ernst van deze reacties. Voor de grootste groep
patiënten (±70%) wordt hierbij aangenomen dat de genetische achtergrond een belangrijke rol
speelt [14].
In deze scriptie zal de associatie onderzocht worden tussen de hierboven aangehaalde SNPs in
DSB herstelgenen en TGFβ1 en de ontwikkeling van ernstige acute en late normale
weefselcomplicaties in een groep van RT behandelde H&N kankerpatiënten. Het doel is om
genetische variaties te identificeren die de ontwikkeling van ernstige acute en late
weefselcomplicaties kunnen voorspellen voor de start van de therapie. Hierdoor zouden de
meest radiosensitieve patiënten kunnen geïdentificeerd worden met een in de toekomst
mogelijke individualisering van het behandelingsschema.
In een eerste luik van deze thesis wordt de associatie onderzocht tussen SNPs in DNA
herstelgenen en TGFβ1 en het optreden van chronische weefselcomplicaties (chronische
dysfagie, fibrose en xerostomie). De SNPs die hierbij zullen worden onderzocht zijn XRCC3
c.722C>T, Ku70 c.-1310C>G, TGFβ1c.-509C>T, TGFβ1 c.-800G>A en TGFβ1 c.74G>C.
In een tweede luik wordt in een nieuw verzamelde groep H&N kankerpatiënten de relatie
onderzocht tussen de eerder vermelde SNPs en het optreden van acute weefselcomplicaties
(dysfagie, mucositis en dermatitis). Deze groep omvat naast patiënten die een standaard
IMRT behandeling kregen (standaardgroep) ook patiënten uit de dosis de-escalatie (DD)
studie, die op de vakgroep “RT en experimenteel kankeronderzoek” loopt [53]. In deze thesis
zullen gegevens van patiënten uit de DD studie ook afzonderlijk vergeleken worden met de
data van patiënten die een standaard IMRT therapie ondergingen.

Materialen en Methoden
14
Materialen en Methoden 1. Introductie
In deze thesis werd de associatie nagegaan tussen 5 SNPs en het optreden van acute en late
normale weefselcomplicaties. Hiervoor werden bloedstalen verzameld van patiënten met een
primaire H&N tumor, behandeld met IMRT in het Universitair Ziekenhuis Gent (UZG). Uit
deze stalen werd het DNA uit lymfocyten of direct uit vol bloed geïsoleerd. Door gebruik te
maken van specifieke primerparen werd het DNA fragment dat de te onderzoeken polymorfe
plaats bevat geamplificeerd via Polymerase Chain Reaction (PCR). De uiteindelijke SNP
bepaling werd uitgevoerd d.m.v. Restriction Fragment Length Polymorphism (RFLP) of High
Resolution Melting (HRM) analyse.
2. Studiepopulatie
Voor deze scriptie werden bloedstalen verzameld van 79 H&N kankerpatiënten die behandeld
werden met IMRT op de dienst RT van het UZ Gent. Hiervan werden er 52 gedurende het
jaar van deze scriptie gecollecteerd, de overige stalen werden reeds verzameld binnen de
onderzoeksgroep. De studiepopulatie voor de analyses naar acute weefseltoxiciteiten
(dermatitis, mucositis en dysfagie) bestond uit 67 mannen (85%) en 12 vrouwen (15%). De
gemiddelde leeftijd van de patiënten op het tijdstip van de therapie bedraagt 60.9 jaar
(spreiding: 43-86 jaar). Deze groep bevat 46 patiënten uit de DD groep en 33 patiënten uit de
standaardgroep.
Voor de analyses naar late weefseltoxiciteiten (fibrose, xerostomie en dysfagie) werden 61
patiënten geïncludeerd, waarvan 51 mannen (84%) en 10 vrouwen (16%). Deze patiënten
vormen een subgroep van de populatie geïncludeerd in een reeds gepubliceerd onderzoek naar
acute radiosensitiviteit[19]. De gemiddelde leeftijd van deze populatie was 59.7 jaar
(spreiding: 40-80). De patiënten waren gemiddeld 4.1 jaar in follow-up (spreiding: 2.7-5.3)
Naast RT onderging een deel van de patiënten ook chemotherapie en/of chirurgie (heelkunde,
HK). Een overzicht van de verkregen behandelingen van de patiënten geïncludeerd in de
analyses naar acute en late toxiciteiten worden weergegeven in tabel 2.1.
Van elke patiënt die deelneemt aan de studie werd gevraagd een lijst in te vullen voor het
verkrijgen van informatie betreffende hun rook- en drinkgedrag. Deze gegevens, inclusief
leeftijd en geslacht staan beschreven in bijlage A.

Materialen en Methoden
15
Tabel 2.1: Overzicht behandelingen
Behandeling Acute weefseltoxiciteiten Late weefseltoxiciteiten
n (%) n (%)
Exclusief RT 25 (31.6) 21 (34.5)
Adjuvante RT na heelkundige ingreep 13 (16.5) 15 (24.5)
Heelkunde gevolgd door radiochemotherapie 4 (5.1) 16 (26.2)
Chemoradiotherapie* 37 (46.8) 9 (14.8) Afkortingen: RT = radiotherapie *2 patiënten kregen bijkomend een behandeling met cetuximab
De acute stralingsgeïnduceerde weefselcomplicaties namelijk faryngeale en oesofageale
dysfagie, mucositis en dermatitis werden wekelijks gescoord gebruik makend van de
CTCAEv3.0 schaal [16]. Voor de late weefselcomplicaties meer bepaald fibrose, xerostomie,
late faryngeale en oesofageale dysfagie gebeurde de scoring gedurende het jaar na de therapie
om de 3 maanden, gedurende het 2e jaar om de 4 maanden en gedurende het 3e, 4e en 5e jaar
om de 6 maanden. Men beschouwt de complicaties die optreden vanaf 6 maanden na de
therapie als late weefselcomplicaties. De CTC-scores voor de verschillende
weefselcomplicaties staan beschreven in bijlage B.
Vóór bloedafname werd aan de patiënten gevraagd een ‘informed consent’ te ondertekenen.
Deze studie werd goedgekeurd door de Ethische Comité van het UZG.
3. Dosimetrische gegevens
De stralingsdosis die de normale weefsels ontvangen is een eerste bepalende factor voor het
ontstaan van stralingstoxiciteiten. De dosis-volume histogrammen (DVHs) van de weefsels
die een kritische rol spelen in de ontwikkeling van de acute en late weefselcomplicaties
werden bestudeerd. De DVHs van deze kritische structuren werden bekomen van de dienst
RT van het UZG. Op basis van deze informatie werd de gemiddelde dosis (Dmean) op elk van
de structuren berekend.
De kritische structuren die werden onderzocht bij het ontstaan van dysfagie (acuut/laat) zijn
de PCs (UPC, MPC en IPC), de supraglottis (larynx), het bovenste deel van de slokdarm
(upper eosofagus) en de sfinkter aldaar (upper oesophageal sphincter, UES).Voor deze
structuren werd naast de Dmean ook de minimale dosis op de omtrek5 (Dmin_circumferentie) in
5 Dmin_circumferentie wordt gedefinieerd als de laagste isodose die de twee naburige craniocaudale faryngeale constrictorspieren omvat.

Materialen en Methoden
16
rekening gebracht. Skin5mm6 is de structuur die gelinkt wordt aan optreden van dermatitis. De
Dmean ter hoogte van de mondholte en de mucosa werd onderzocht voor een mogelijke
associatie met het ontstaan van acute mucositis. Met betrekking tot xerostomie is de Dmean op
de rechter en linker parotisklier een bepalende factor. Voor fibrose werd de Dmean ter hoogte
van de subcutis onderzocht [19,27].
4. DNA purificatie
De isolatie van het genomisch DNA gebruikt tijdens deze scriptie gebeurde direct uit humaan
bloed (EDTA buis) of uit humane lymfocyten die reeds geïsoleerd werden uit heparine buisjes
en ingevroren in vloeibare stikstof (-196°C).
Voor de DNA purificatie uit vol bloed worden als eerste de rode bloedcellen (RBC)
gelyseerd. Hiervoor wordt 3 ml vol bloed toegevoegd aan 9 ml RBC-lyse oplossing (Qiagen).
De buisjes worden gedurende 5 min op kamertemperatuur (KT) gehouden en tussentijds
enkele malen omgekeerd. Na de incubatie worden de buisjes 2 min aan 2000xg (4600 rpm)
centrifugeerd (Centrifuge 5415 R, Eppendorf) en wordt het supernatans bijna volledig
verwijderd. Om de verkregen celpellet op te lossen in de overgebleven vloeistof worden de
buisjes hevig gevortext (Genius3, IKA). Vervolgens wordt de cellyse oplossing toegevoegd (3
ml). Na deze stap kan het protocol ongeveer 2 jaar onderbroken worden. Voor de isolatie van
DNA uit lymfocyten worden de cellen na ontdooiing overgebracht in 1.5 ml epjes waarna ze
gedurende 5 min gecentrifugeerd worden aan 300xg (1800 rpm). Vervolgens wordt, na
verwijderen van het supernatans, de verkregen celpellet geresuspendeerd in phosphate
buffered saline (PBS) (Invitrogen) tot een totaalvolume van 200 µl. Wederom worden de
epjes gecentrifugeerd gedurende 10 sec aan 16000xg (13100 rpm). Het supernatans wordt
verwijderd met behoud van 40 µl vloeistof. Om de cellen in de overgebleven vloeistof op te
lossen, wordt hevig gevortext. Na deze stap wordt de cellyse oplossing (Qiagen) toegevoegd
(600 µl) en kan ook hier het protocol ongeveer 2 jaar onderbroken worden.Voor de verdere
DNA isolatie wordt voor zowel humaan bloed (●) als voor humane lymfocyten (♦) een
gelijkaardig protocol gebruikt. Na de cellyse worden de aanwezige eiwitten geprecipiteerd.
Hiervoor wordt proteïne precipitatie buffer toegevoegd aan het cellysaat (●: 1 ml; ♦: 200 µl)
waarna gedurende 20 sec aan hoge snelheid gevortext wordt. Vervolgens worden de
buisjes/epjes preferentieel 5 min op ijs of 10 min in de ijskast geplaatst om de eiwitten beter
6 Skinn5mm wordt gedefineerd als een laagje van 5 mm dat een groot deel van de epidermis en dermis omvat. Dit laagje wordt craniaal afgelijnd t.h.v. de Meatus Akousticus Externus en caudaal t.h.v. wervel C6.

Materialen en Methoden
17
te laten neerslaan. De buisjes/epjes worden hierna gecentrifugeerd (●: 5 min aan 2000xg: ♦: 2
min aan 16000xg) zodat de eiwitten neerslaan en een pellet vormen. Bij de verdere
opzuivering van het DNA wordt het DNA eerst geprecipiteerd. Hierbij wordt het supernatans
overgebracht in een buisje/epje met 100% isopropanol (VWR) (●: 3 ml; ♦: 600 µl). Nadien
wordt de oplossing ongeveer 50 keer omgekeerd totdat de witte DNA draden zichtbaar
worden. Vervolgens worden de buisjes/epjes gecentrifugeerd (●: 3 min aan 2000xg; ♦: 1 min
aan 16000xg) waarbij het DNA als een witte pellet zichtbaar wordt. Het supernatans wordt
afgepipetteerd en de DNA pellet wordt gewassen met 70% ethanol (Sigma) (●:3 ml; ♦:600
µl). Daarna worden de buisjes/epjes opnieuw gecentrifugeerd (●: 1 min aan 2000xg; ♦: 1 min
aan 16000xg), wordt het supernatans verwijderd en laat men de buisjes/epjes gedurende 10
min drogen aan de lucht. De laatste stap van de DNA purificatie is de DNA hydratie.
Hiervoor wordt DNA hydratie oplossing (Qiagen) toegevoegd (●: 500 µl; ♦: 150 µl) en
worden de buisjes/epjes gedurende 1 uur op 65°C (Polystat 24, Fisher Scientific) geïncubeerd.
In de laatste worden de buisjes/epjes na een overnachtperiode op KT kort gecentrifugeerd,
overgebracht in 1.5 ml buisjes en worden ze bewaard op -20°C.
5. Concentratie -en kwaliteitbepaling van het DNA
Voor verdere analyses wordt de DNA oplossing van alle stalen op een concentratie gebracht
van 25ng/µl. Voor de concentratiebepaling van het geïsoleerde DNA wordt gebruik gemaakt
van de NanoDrop ND-1000 V3.5 spectrometer. Deze is beschikbaar op de afdeling Medische
Genetica van het UZG. Aan de hand van absorbantiemetingen kan de concentratie en de
zuiverheid van het staal gemeten worden. De verhouding van de absorbantie bij 260 en 280
nm van 1.8 wordt beschouwd als een zuiver staal, een contaminatie met eiwitten zal deze
waarde doen dalen. De eventuele verdunning gebeurt door toevoeging van TE buffer (10mM).
Voor het maken van een 100mM stockoplossing wordt 6.05 g Tris en 1.86 g EDTA opgelost
in 500ml Dnase en Rnase vrij gedestilleerd water (dH2O, Sigma). De oplossing wordt
aangezuurd met 1M HCl tot een pH van 8.5 wordt bekomen. Voor het maken van de
werkoplossing wordt de stockoplossing 10 x verdund met dH2O.

Materialen en Methoden
18
6. Polymerase chain reaction (PCR)
6.1 Principe
PCR is een in vitro techniek die typisch gebruikt wordt voor de multiplicatie van een
welbepaalde DNA fragment (amplicon). Door gebruik te maken van een specifiek primerpaar
kan de plaats van de te onderzoeken polymorfe regio geamplificeerd worden.
Het algemene PCR proces bestaat uit 3 stappen: denaturatie, aanhechting of annealing van de
primers en primerextensie. In de eerste stap wordt het DNA gedenatureerd. Deze stap gebeurt
bij een temperatuur van 95°C waardoor het dubbelstrengig DNA (double stranded DNA,
dsDNA) enkelstrengig (single stranded DNA, ssDNA) wordt. Tijdens de tweede stap (de
annealing) wordt de temperatuur verlaagd tot de annealingstemperatuur (Ta) van de primers,
wat hen toelaat om te binden aan hun complementaire sequenties op het ssDNA. De Ta is
afhankelijk van de base samenstelling van de gekozen primers en wordt als volgt bepaald: Ta
= [(#T/A × 2) + (#G/C × 4)] − 2. De laatste stap van de PCR reactie is de extentie van de
primers. Bij deze stap wordt de temperatuur verhoogd tot 72°C waarbij het gebruikte hot start
Taq polymerase zijn functie kan uitvoeren. Dit polymerase bindt deoxyribonucleotide
trifosfaten (dNTPs) aan de gebonden primers en synthetiseert de volledige complementaire
DNA streng. Deze 3 stappen worden verschillende malen herhaald wat uiteindelijk resulteert
in een zeer groot aantal kopijen van het DNA fragment [54]. Het algemene PCR programma
dat in deze thesis wordt gebruikt is weergegeven in tabel 2.2.
Tabel 2.2: Algemeen PCR programma
95°C 5 min
95°C 30 sec
Ta 30 sec 35 cycli
72°C 30 sec
72°C 10 min
15°C 10 min
De gebruikte PCR protocols worden beschreven in tabel 2.3 deze werden reeds
geoptimaliseerd binnen de onderzoeksgroep. Er werd gewerkt met de Bio-rad c1000 Thermal
Cycler PCR toestel.
Materialen en Methoden
19
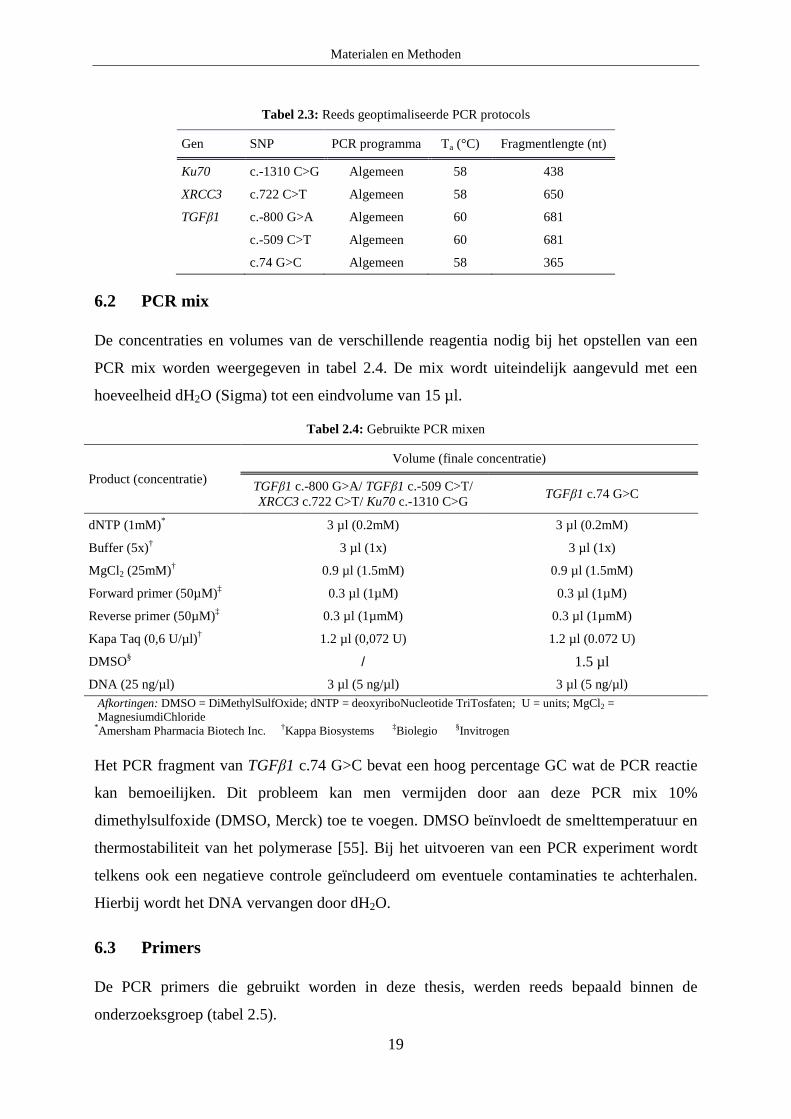
Tabel 2.3: Reeds geoptimaliseerde PCR protocols
Gen SNP PCR programma Ta (°C) Fragmentlengte (nt)
Ku70 c.-1310 C>G Algemeen 58 438
XRCC3 c.722 C>T Algemeen 58 650
TGFβ1 c.-800 G>A Algemeen 60 681
c.-509 C>T Algemeen 60 681
c.74 G>C Algemeen 58 365
6.2 PCR mix
De concentraties en volumes van de verschillende reagentia nodig bij het opstellen van een
PCR mix worden weergegeven in tabel 2.4. De mix wordt uiteindelijk aangevuld met een
hoeveelheid dH2O (Sigma) tot een eindvolume van 15 µl.
Tabel 2.4: Gebruikte PCR mixen
Product (concentratie)
Volume (finale concentratie)
TGFβ1 c.-800 G>A/ TGFβ1 c.-509 C>T/ XRCC3 c.722 C>T/ Ku70 c.-1310 C>G
TGFβ1 c.74 G>C
dNTP (1mM)* 3 µl (0.2mM) 3 µl (0.2mM)
Buffer (5x)† 3 µl (1x) 3 µl (1x)
MgCl2 (25mM)† 0.9 µl (1.5mM) 0.9 µl (1.5mM)
Forward primer (50µM)‡ 0.3 µl (1µM) 0.3 µl (1µM)
Reverse primer (50µM)‡ 0.3 µl (1µmM) 0.3 µl (1µmM)
Kapa Taq (0,6 U/µl)† 1.2 µl (0,072 U) 1.2 µl (0.072 U)
DMSO§ / 1.5 µl DNA (25 ng/µl) 3 µl (5 ng/µl) 3 µl (5 ng/µl)
Afkortingen: DMSO = DiMethylSulfOxide; dNTP = deoxyriboNucleotide TriTosfaten; U = units; MgCl2 = MagnesiumdiChloride
*Amersham Pharmacia Biotech Inc. †Kappa Biosystems ‡Biolegio §Invitrogen
Het PCR fragment van TGFβ1 c.74 G>C bevat een hoog percentage GC wat de PCR reactie
kan bemoeilijken. Dit probleem kan men vermijden door aan deze PCR mix 10%
dimethylsulfoxide (DMSO, Merck) toe te voegen. DMSO beïnvloedt de smelttemperatuur en
thermostabiliteit van het polymerase [55]. Bij het uitvoeren van een PCR experiment wordt
telkens ook een negatieve controle geïncludeerd om eventuele contaminaties te achterhalen.
Hierbij wordt het DNA vervangen door dH2O.
6.3 Primers
De PCR primers die gebruikt worden in deze thesis, werden reeds bepaald binnen de
onderzoeksgroep (tabel 2.5).

Materialen en Methoden
20
Tabel 2.5: Gebruikte PCR primers
Gen Nr. Naam Sequentie Lengte
(nt) Ta
(°C)
Ku70 c.-1310 C>G P1a Ku70/1233F 5'-CTTCAGACCACTCTCTTCTC-3' 20 58
P2a Ku70/1670R 5'-TCACCTCACAGTAGTCGTTG-3' 20 58
XRCC3 c.722 C>T 27 XRCC3/17.630F 5'-GACACCTCTACAGAGGACG-3' 19 58
28 XRCC3/18.279R 5'-TTCTCGATGGTTAGGCACAG-3' 20 58
TGFβ1 c.-800 G>A / TGFβ1 c.-509 C>T*
39 TGFb1/142F 5'-GCAGTTGGCGAGAACAGTTG-3' 20 60
40 TGFb1/722R 5'-TGGGTCACCAGAGAAAGAGG-3' 20 60
TGFβ1 c.74 G>C 41 TGFb1/1.936F 5'-TGTTCGCGCTCTCGGCAG-3' 18 58
42 TGFb1/2.300R 5'-GACCTCCTTGGCGTAGTAG-'3 19 58 *Beide SNPs liggen in hetzelfde PCR fragment
Voor de keuze van de primerparen wordt rekening gehouden met de lengte van de primers
welke ideaal 20 nucleotiden (nt) is, de Ta is best gelegen tussen 58°C en 62°C en identiek
voor het PCR primerpaar. Optimaal eindigt de forward primer met een G of C, terwijl de
reverse primer begint met een G of C dit wegens de sterkere interactie met de doelsequentie
van deze nt. Men vermijdt ook best herhalingen van 3 of meer gelijke nt om aspecifieke
binding te voorkomen.
6.4 PCR Controle
De controle van de PCR reacties gebeurt d.m.v. gelektroforese. Hierbij wordt een 1.5%
agarosegel gemaakt door opkoken, tot een heldere vloeistof van 1.5 g agarose (Invitrogen) in
100 ml TBE buffer (0.5x). Deze oplossing wordt in een daarvoor voorziene houder gegoten
waarin kammetjes geplaatst worden voor het vormen van de laantjes waarin het PCR product
kan worden geladen. Voor de aanmaak van een stockoplossing TBE buffer (10x) wordt 54 g
Tris, 27.5 g en 4.65 g EDTA opgemengd in 500 ml dH2O. Deze stockoplossing wordt 20 keer
verdund met dH2O om de werkoplossing (0.5x) te krijgen.
Na het afkoelen en opstijven van de agarose gel (na ± 20 min) kan de gel geladen worden.
Hiervoor wordt aan 1 µl ladingsbuffer7 2 µl PCR product toegevoegd. Samen met een extra 6
of 16 µl dH2O, afhankelijk van de grootte van de gebruikte laantjes, wordt het PCR product
op gel gebracht. Naast de PCR stalen wordt ook 1 µl DNA ladder (Invitrogen) in één van de
7 Voor de aanmaak van ladingsbuffer wordt 500 µl glycerol (om het DNA te verzwaren) en 250 µl broomfenolblauw (voor de visualisatie van het DNA in de gel) opgemengd met 250 µl dH2O.

Materialen en Methoden
21
laantjes gepipetteerd. Deze wordt zichtbaar op gel als meerdere bandjes van gekende grootte.
Aan de hand van deze ladder kan nagegaan worden of het PCR bandje op de juiste hoogte
voorkomt en dus het gewenste amplicon verkregen werd. Na het laden van de gel stelt men
het elektroforese toestel (Cosmo bio, Westburg) in op 135 Volt voor ongeveer 30 min. De
DNA ladder en de PCR bandjes worden gevisualiseerd door het gel gedurende 40 à 45 min in
een ethidiumbromide (EtBr , Gibco) bad (0.1%) te leggen en daarna op een UV plaat (UV
Transilluminator, UPV) te plaatsten.
7. SNP bepaling
In deze thesis wordt voor de bepaling van de genetische variaties in de patiëntstalen gebruik
gemaakt van 2 technieken: RFLP, een post-PCR techniek en HRM waarbij PCR en SNP
detectie gebeuren in één reactie.
7.1 Restriction fragment length polymorphism (RFLP)
Bij RFLP zal na PCR van de te onderzoek sequentie het verkregen amplicon door
welbepaalde restrictie-enzymen (RE) geknipt worden. Bij de keuze van het RE wordt gewerkt
met de restriction mapper software [56]. Met deze software kan het PCR fragment virtueel
door verschillende enzymen geknipt worden. Het ideale RE knipt op de plaats van de
polymorfe site waardoor de aan- of afwezigheid van de te onderzoeken SNP een ander
knippatroon geeft en fragmentanalyse de SNP bepaling toelaat. De gevormde fragmenten
worden d.m.v. gelelektroferese op basis van hun lengte gescheiden. Voor een RFLP reactie
wordt gewerkt in een totaalvolume van 30 µl. Naast het RE wordt ook een enzym specifieke
buffer voorzien. De gebruikte hoeveelheid bedraagt steeds 10% (3 µl) van het eindvolume. De
hoeveelheid PCR product en het aantal U van het RE werden voor de geselecteerde SNPs via
optimalisatie reeds bepaald binnen de onderzoeksgroep. Afhankelijk van het gebruikte RE
wordt ook bovine serum albumine (BSA) toegevoegd aan de digest mix. Met behulp van
dH2O wordt er uiteindelijk aangevuld tot een totaalvolume van 30 µl per staal. Na het
toevoegen van de digest mix aan het DNA worden de gebruikte epjes goed gevortext en
gecentrifugeerd. Daarna worden ze gedurende een geoptimaliseerde periode geïncubeerd bij
een specifieke temperatuur in een warmwaterbad (Polystat 24, Fisher Scientific). Hierna
worden de epjes kort gecentrifugeerd en worden ze voor minimaal 30 min in de ijskast
bewaard. De in deze thesis gebruikte RFLP protocols staan beschreven in bijlage C . Deze
werden reeds binnen de onderzoeksgroep geoptimaliseerd.

Materialen en Methoden
22
Voor de controle van het digest wordt aan het digestproduct 3 µl (10% van het eindvolume)
ladingsbuffer toegevoegd en wordt het product geladen op een 2% agarosegel8. Om na te gaan
of het PCR product effectief geknipt werd, wordt er eveneens in één van de laantjes niet-
geknipt PCR product geladen. Na het laden van de gel stelt men het elektroforese toestel in op
135 Volt gedurende 30 à 40 min. De digestproducten worden net zoals de PCR producten
gevisualiseerd op een UV plaat na incubatie in een EtBr bad (0.1%).
7.2 High resolution melting (HRM)
HRM is een high-throughput techniek waarbij de genotypering gebeurt op basis van het
smeltpatroon van het PCR amplicon. Tijdens deze thesis wordt gewerkt met het Applied
Biosystems 7500 Fast Real-Time PCR systeem en de geassocieerde Applied Biosystems
HRM software v2.0. De analyse start met een PCR van de te onderzoeken regio in de
aanwezigheid van een fluorescente kleurstof (Meltdoctor HRM dye) dat een hoog fluorescent
signaal geeft bij de binding aan dsDNA en een laag fluorescent signaal in de niet-gebonden
toestand (als het DNA enkelstrengig is, na denaturatie) [57]. De standaard mix die gebruikt
wordt bij een HRM experiment staat vermeld in tabel 2.6. De mix wordt uiteindelijk
aangevuld met dH2O tot een totaalvolume van 20 µl per staal.
Tabel 2.6: Standaard HRM mix
Product (concentratie) Hoeveelheid (concentratie)
Buffer (10x) 2 µl (1x)
MgCl2 (25mM) 1.2 µl (1.5mM)
dNTP Mix (10mM) 0.4 µl (0.2mM)
Forward primer (5µM) 1.2 µl (0.3µM)
Reverse primer (5µM) 1.2 µl (0.3µM)
Meltdoctor HRM Dye (20x) 1 µl (1x)
AmpliTaq Gold 360 DNA Polymerase (5U/µl) 0.2 µl (0.05 U/µl, 1U)
DNA (25 ng/µl) 1 µl (1 ng/µl)
De smeltcurve ontstaat door het DNA traag te denaturen (uitsmelten). Tijdens deze
denaturatie wordt het dsDNA ssDNA waardoor de kleurstof van het DNA loskomt en een
vermindering in fluorescentie wordt waargenomen. Door het detecteren van de
fluorescentiegraad in functie van de temperatuur wordt een smeltpatroon gevormd (figuur 2.1)
[57]. Het standaard HRM programma wordt weergegeven in tabel 2.7.
8 2% agarosegel: 2 g agarose + 100 ml TBE (0.5x)
Materialen en Methoden
23
Tabel 2.7: Standaard HRM programma
Stadium Reactiestap Temperatuur
(°C) Tijd
Precyclische fase Enzym activatie 95 10 min
Cyclische fase (40 cycly) Denaturatie 95 15 sec
Annealing/Extentie 60 1 min
Smelt curve/ dissociatie Denaturatie 95 10 sec
Annealing 60 1 min
HRM 95 15 sec
Annealing 60 15 sec
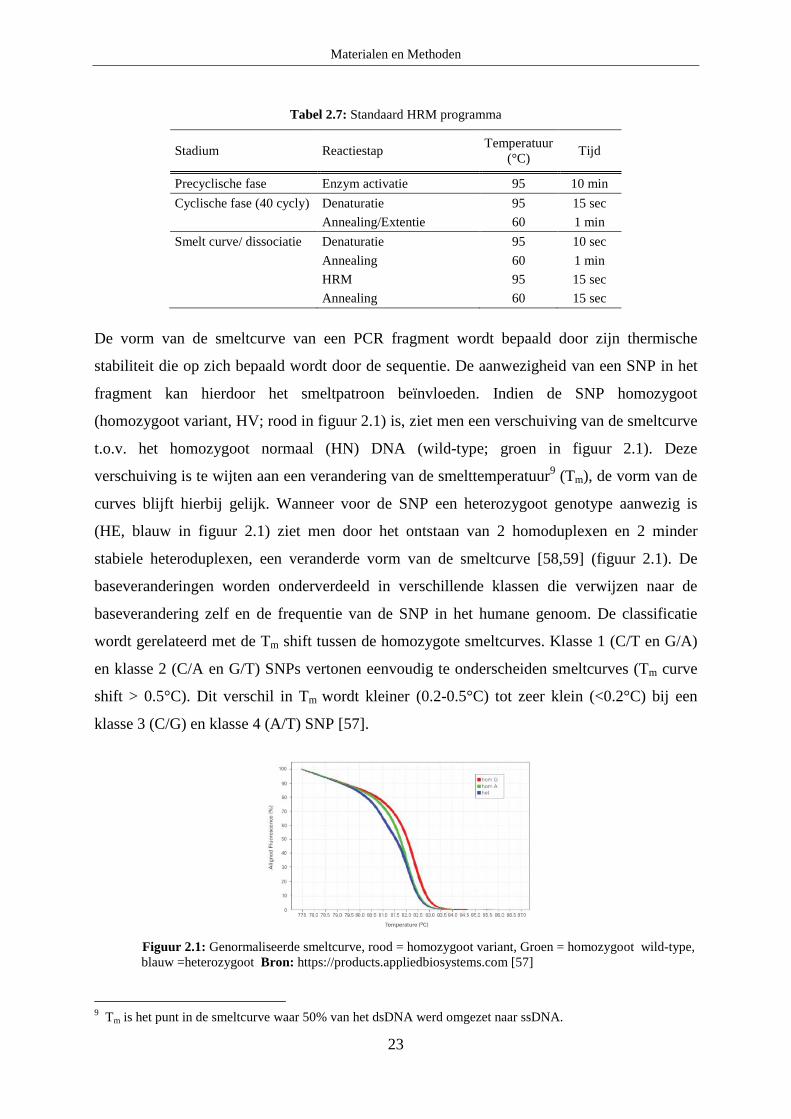
De vorm van de smeltcurve van een PCR fragment wordt bepaald door zijn thermische
stabiliteit die op zich bepaald wordt door de sequentie. De aanwezigheid van een SNP in het
fragment kan hierdoor het smeltpatroon beïnvloeden. Indien de SNP homozygoot
(homozygoot variant, HV; rood in figuur 2.1) is, ziet men een verschuiving van de smeltcurve
t.o.v. het homozygoot normaal (HN) DNA (wild-type; groen in figuur 2.1). Deze
verschuiving is te wijten aan een verandering van de smelttemperatuur9 (Tm), de vorm van de
curves blijft hierbij gelijk. Wanneer voor de SNP een heterozygoot genotype aanwezig is
(HE, blauw in figuur 2.1) ziet men door het ontstaan van 2 homoduplexen en 2 minder
stabiele heteroduplexen, een veranderde vorm van de smeltcurve [58,59] (figuur 2.1). De
baseveranderingen worden onderverdeeld in verschillende klassen die verwijzen naar de
baseverandering zelf en de frequentie van de SNP in het humane genoom. De classificatie
wordt gerelateerd met de Tm shift tussen de homozygote smeltcurves. Klasse 1 (C/T en G/A)
en klasse 2 (C/A en G/T) SNPs vertonen eenvoudig te onderscheiden smeltcurves (Tm curve
shift > 0.5°C). Dit verschil in Tm wordt kleiner (0.2-0.5°C) tot zeer klein (<0.2°C) bij een
klasse 3 (C/G) en klasse 4 (A/T) SNP [57].
Figuur 2.1: Genormaliseerde smeltcurve, rood = homozygoot variant, Groen = homozygoot wild-type, blauw =heterozygoot Bron: https://products.appliedbiosystems.com [57]
9 Tm is het punt in de smeltcurve waar 50% van het dsDNA werd omgezet naar ssDNA.

Materialen en Methoden
24
7.2.1 Primer bepaling
Bij de ontwikkeling van primers voor HRM moet men rekening houden dat extra
polymorfismen in het PCR fragment een invloed hebben op het smeltpatroon. Tijdens deze
thesis zal HRM gebruikt worden voor de genotypering van 1 SNP per fragment en wordt de
aanwezigheid van een andere polymorfisme in het amplicon vermeden. Hiernaast mogen ook
geen SNPs in de primersequentie voorkomen omdat dit hun binding zou kunnen beïnvloeden.
De aanwezigheid van SNPs kan gecontroleerd worden via de Ensembl genoom browser [60].
Verder moet er bij de keuze van het primerpaar ook rekening gehouden worden met de
primerlengte (±20 nt elk), de fragmentlengte (optimaal 80-100 nt), de smelttemperatuur van
de primers (58°C-60°C), het % GC (30-80%) en moeten herhalingen (> 3) worden vermeden.
Deze karakteristieken kunnen samen met de zelf-complementariteit van de primers bepaald
worden via de Oligonucleotide Properties Calculator (Oligo Calc) [61]. Voor het ontwerpen
van de primers wordt de SNP eerst, via NCBI, in de referentiesequentie van het gen gezocht
[36]. De uiteindelijke primerontwikkeling kan manueel of automatisch via Primer3Plus
gebeuren [62]. Na het ontwerpen van de primers dient hun specificiteit getest te worden, dit
kan d.m.v. een in silico PCR via de UCSC genoom browser [63]. Deze software laat toe te
controleren of de primers op de correcte plaats in het genoom binden en het correcte fragment
genereren. Bij de optimalisatie van een HRM experiment worden telkens meerdere
primerparen besteld waaruit dan het optimale paar wordt bepaald. De gebruikte HRM primers
staan vermeld in tabel 2.8.
Tabel 2.8: Gebruikte HRM primers
Nr. Naam Sequentie Primerlengte
(nt) Tm
(°C) PCR
fragment (nt)
Ku70 c.-1310 C>G
145 Ku70_HRM_F2 5’-GTCGTGGCCCAAGTCTCC -3’ 18 61.7
100 147 Ku70_HRM_R 5’-CCATCATGCTGGGTGAGG -3’ 18 61.1
XRCC3 c.722 C>T
124 XRCC3_HRM_F1 5’-GGCCAGGCATCTGCAGTC-3’ 18 63
87 122 XRCC3_HRM_R1 5’-CTCACCTGGTTGATGCACAG-3’ 20 60.3
7.2.2 Optimalisatie HRM protocols
De optimalisatie van de HRM protocols van Ku70 c.-1310 C>G en XRCC3 c.722 C>T
vormen één van de doelstelling van deze thesis. Tijdens deze optimalisatie worden

Materialen en Methoden
25
verschillende HRM mixen en HRM programma’s geprobeerd om tot een optimale smeltcurve
te komen waarin de verschillende genotypes konden onderscheiden worden.
8. Kwaliteitscontroles
Om de juistheid van de reacties na te gaan volgt steeds een kwaliteitscontrole na genotypering
van alle stalen. Hierbij worden per SNP ongeveer 15% van de stalen opnieuw gegenotypeerd.
Indien afwijkingen worden gedetecteerd, worden alle stalen van deze reactie opnieuw
geanalyseerd.
9. Statistische Analyse
Voor de statistische analyse wordt gewerkt met de SPSS 15.0 software. Voor elke
onderzochte weefselcomplicatie (xerostomie, fibrose, dermatitis, acute dysfagie, late dysfagie
en acute mucositis) worden de patiënten op basis van de CTC-scores opgedeeld in twee
groepen. Bij de verdere analyse wordt de niet- tot matig radiosensitieve groep (CTC0-2, voor
acute complicaties en CTC0-1 voor late complicaties) vergeleken met de sterk radiosensitieve
groep (CTC3+ voor acute complicaties en CTC2+ voor late complicaties).
Om na te gaan of patiëntgerelateerde parameters (leeftijd, rookgedrag10 en alcoholgebruik) en
therapiegerelateerde parameters (dosis ter hoogte van kritische weefsels) op zich een effect
hebben op de opdeling van de patiënten in de matig tot ernstig radiosensitieve groep wordt
een Mann-Whitney U test uitgevoerd. Het eventuele effect van het geslacht, chemotherapie en
chirurgie op het optreden van ernstige weefselcomplicaties wordt nagegaan d.m.v. de chi-
kwadraat test. Voor alle SNPs wordt voor elke complicatie ook het Hardy-Weinberg
equilibrium (HWE) gecontroleerd.
Voor het vinden van een relatie tussen de genotypefrequenties en het optreden van ernstige
radiosensitiviteit wordt gebruik gemaakt van logistieke regressie analyse. Om het effect van
therapie- en patiëntgerelateerde factoren uit te sluiten wordt in de analyses gecorrigeerd voor
die parameters die significante verschillen vertonen tussen de twee groepen. Bij de analyses
wordt telkens het HN genotype gebruikt als referentie. De genotypefrequenties worden
geëvalueerd d.m.v. de odds ratio (OR) en de p-waarde. Een p kleiner of gelijk aan 0.05 wordt
als statistisch significant beschouwd.
10 Het rookgedrag wordt uitgedrukt in pack-years (PY): ( aantal sigaretten per dag/20) x aantal jaren gerookt.

Resultaten
26
Resultaten
1. Reeds geoptimaliseerde protocols
De protocols voor de bepaling van de SNPs in het TGFβ1 gen werden reeds geoptimaliseerd
binnen de onderzoeksgroep waardoor de patiëntenstalen eenvoudig bepaald konden worden.
Voor de genotypering van de drie onderzochte polymorfismen TGFβ1 c.-509 C>T, c.-
800G>A en c.74G>C SNP werd de RFLP techniek toegepast. Na inwerking van het RE kan
via de lengte van de bekomen fragmentjes een duidelijk onderscheid gemaakt worden tussen
de mogelijke genotypes. Voorbeelden van digest gels voor elk van de SNPs worden
weergegeven in figuur 3.1. De volledige resultaten van de genotypering staan beschreven in
bijlage D.
Figuur 3.1: Digest gels voor de genotypering van TGFβ1 c.-800 G>A (I), TGFβ1 c.-509C>T (II) en TGFβ1 c.74G>C (III). Per gel van links naar rechts: ladder, ongeknipt product en de 3 mogelijke genotypes.
2. Optimalisatie HRM Protocols
De genotypering van de XRCC3 c.722C>T en Ku70 c.-1310C>G polymorfismen werd binnen
de onderzoeksgroep reeds geoptimaliseerd met behulp van de RFLP techniek. Het proces van
PCR en RFLP, inclusief het lopen van de controlegels, neemt respectievelijk 8 en 9uur in
beslag. Om in de toekomst efficiënter te werken zou het handig zijn dat de genotypering op
een veel snellere manier kan gebeuren. Hiervoor biedt de recent ontwikkelde HRM
technologie een oplossing. Dit proces neemt slechts een 2-tal uur in beslag voor een reactie
waarbij PCR en uitsmelting van de amplicons gecombineerd worden. Het is bovendien een
high-througput techniek die toelaat 96 stalen in eens te genotyperen. Aangezien de
amplificatieplot en ampliconsmeltcurve in grafieken weergegeven worden, is er geen controle
van de PCR via gelelektroforese meer nodig. In deze scriptie werden de protocols voor de
XRCC3 c.722 en Ku70 c.-1310 SNPs geoptimaliseerd voor genotypering via HRM.
Resultaten
27
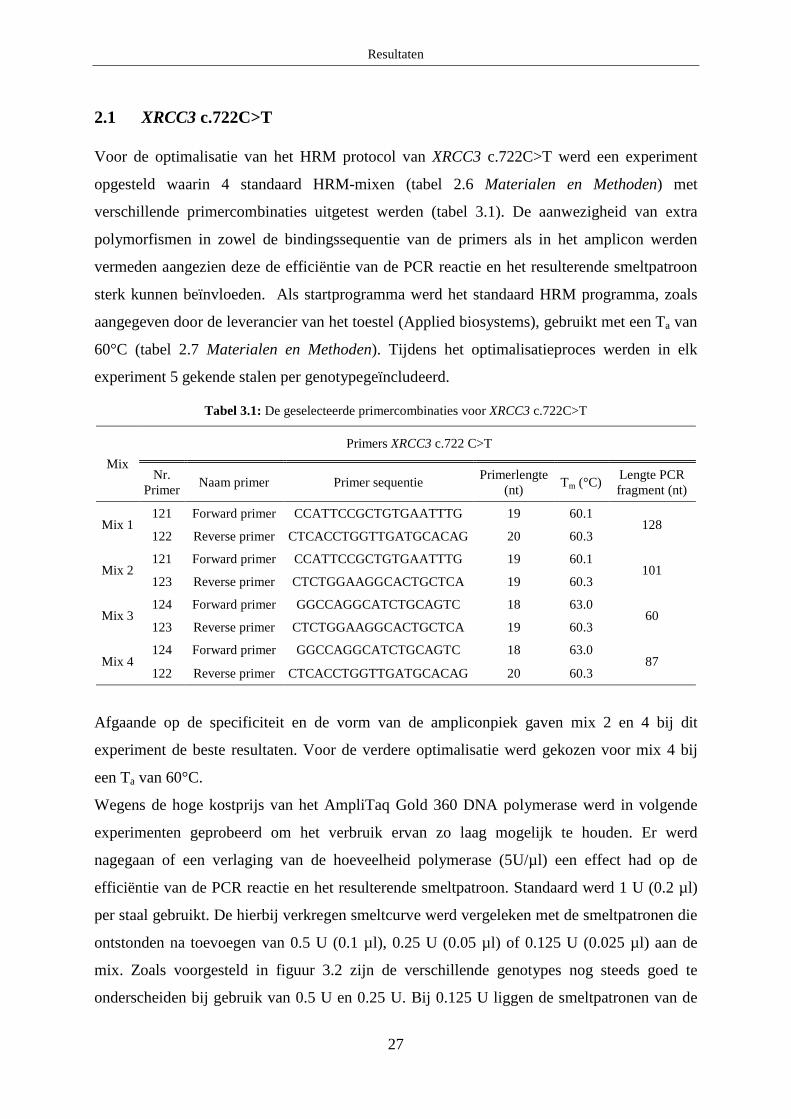
2.1 XRCC3 c.722C>T
Voor de optimalisatie van het HRM protocol van XRCC3 c.722C>T werd een experiment
opgesteld waarin 4 standaard HRM-mixen (tabel 2.6 Materialen en Methoden) met
verschillende primercombinaties uitgetest werden (tabel 3.1). De aanwezigheid van extra
polymorfismen in zowel de bindingssequentie van de primers als in het amplicon werden
vermeden aangezien deze de efficiëntie van de PCR reactie en het resulterende smeltpatroon
sterk kunnen beïnvloeden. Als startprogramma werd het standaard HRM programma, zoals
aangegeven door de leverancier van het toestel (Applied biosystems), gebruikt met een Ta van
60°C (tabel 2.7 Materialen en Methoden). Tijdens het optimalisatieproces werden in elk
experiment 5 gekende stalen per genotypegeïncludeerd.
Tabel 3.1: De geselecteerde primercombinaties voor XRCC3 c.722C>T
Mix
Primers XRCC3 c.722 C>T
Nr. Primer
Naam primer Primer sequentie Primerlengte
(nt) Tm (°C)
Lengte PCR fragment (nt)
Mix 1 121 Forward primer CCATTCCGCTGTGAATTTG 19 60.1
128 122 Reverse primer CTCACCTGGTTGATGCACAG 20 60.3
Mix 2 121 Forward primer CCATTCCGCTGTGAATTTG 19 60.1
101 123 Reverse primer CTCTGGAAGGCACTGCTCA 19 60.3
Mix 3 124 Forward primer GGCCAGGCATCTGCAGTC 18 63.0
60 123 Reverse primer CTCTGGAAGGCACTGCTCA 19 60.3
Mix 4 124 Forward primer GGCCAGGCATCTGCAGTC 18 63.0
87 122 Reverse primer CTCACCTGGTTGATGCACAG 20 60.3
Afgaande op de specificiteit en de vorm van de ampliconpiek gaven mix 2 en 4 bij dit
experiment de beste resultaten. Voor de verdere optimalisatie werd gekozen voor mix 4 bij
een Ta van 60°C.
Wegens de hoge kostprijs van het AmpliTaq Gold 360 DNA polymerase werd in volgende
experimenten geprobeerd om het verbruik ervan zo laag mogelijk te houden. Er werd
nagegaan of een verlaging van de hoeveelheid polymerase (5U/µl) een effect had op de
efficiëntie van de PCR reactie en het resulterende smeltpatroon. Standaard werd 1 U (0.2 µl)
per staal gebruikt. De hierbij verkregen smeltcurve werd vergeleken met de smeltpatronen die
ontstonden na toevoegen van 0.5 U (0.1 µl), 0.25 U (0.05 µl) of 0.125 U (0.025 µl) aan de
mix. Zoals voorgesteld in figuur 3.2 zijn de verschillende genotypes nog steeds goed te
onderscheiden bij gebruik van 0.5 U en 0.25 U. Bij 0.125 U liggen de smeltpatronen van de
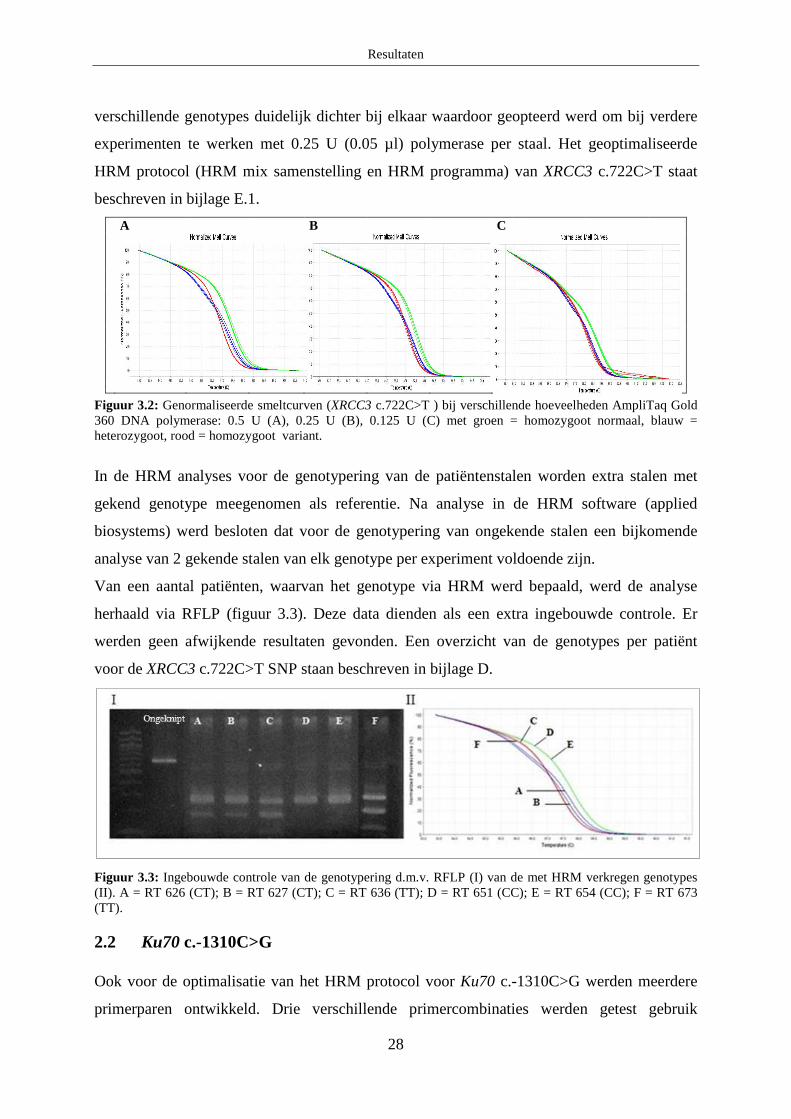
Resultaten
28
verschillende genotypes duidelijk dichter bij elkaar waardoor geopteerd werd om bij verdere
experimenten te werken met 0.25 U (0.05 µl) polymerase per staal. Het geoptimaliseerde
HRM protocol (HRM mix samenstelling en HRM programma) van XRCC3 c.722C>T staat
beschreven in bijlage E.1.
A B C
Figuur 3.2: Genormaliseerde smeltcurven (XRCC3 c.722C>T ) bij verschillende hoeveelheden AmpliTaq Gold 360 DNA polymerase: 0.5 U (A), 0.25 U (B), 0.125 U (C) met groen = homozygoot normaal, blauw = heterozygoot, rood = homozygoot variant.
In de HRM analyses voor de genotypering van de patiëntenstalen worden extra stalen met
gekend genotype meegenomen als referentie. Na analyse in de HRM software (applied
biosystems) werd besloten dat voor de genotypering van ongekende stalen een bijkomende
analyse van 2 gekende stalen van elk genotype per experiment voldoende zijn.
Van een aantal patiënten, waarvan het genotype via HRM werd bepaald, werd de analyse
herhaald via RFLP (figuur 3.3). Deze data dienden als een extra ingebouwde controle. Er
werden geen afwijkende resultaten gevonden. Een overzicht van de genotypes per patiënt
voor de XRCC3 c.722C>T SNP staan beschreven in bijlage D.
Figuur 3.3: Ingebouwde controle van de genotypering d.m.v. RFLP (I) van de met HRM verkregen genotypes (II). A = RT 626 (CT); B = RT 627 (CT); C = RT 636 (TT); D = RT 651 (CC); E = RT 654 (CC); F = RT 673 (TT).
2.2 Ku70 c.-1310C>G
Ook voor de optimalisatie van het HRM protocol voor Ku70 c.-1310C>G werden meerdere
primerparen ontwikkeld. Drie verschillende primercombinaties werden getest gebruik
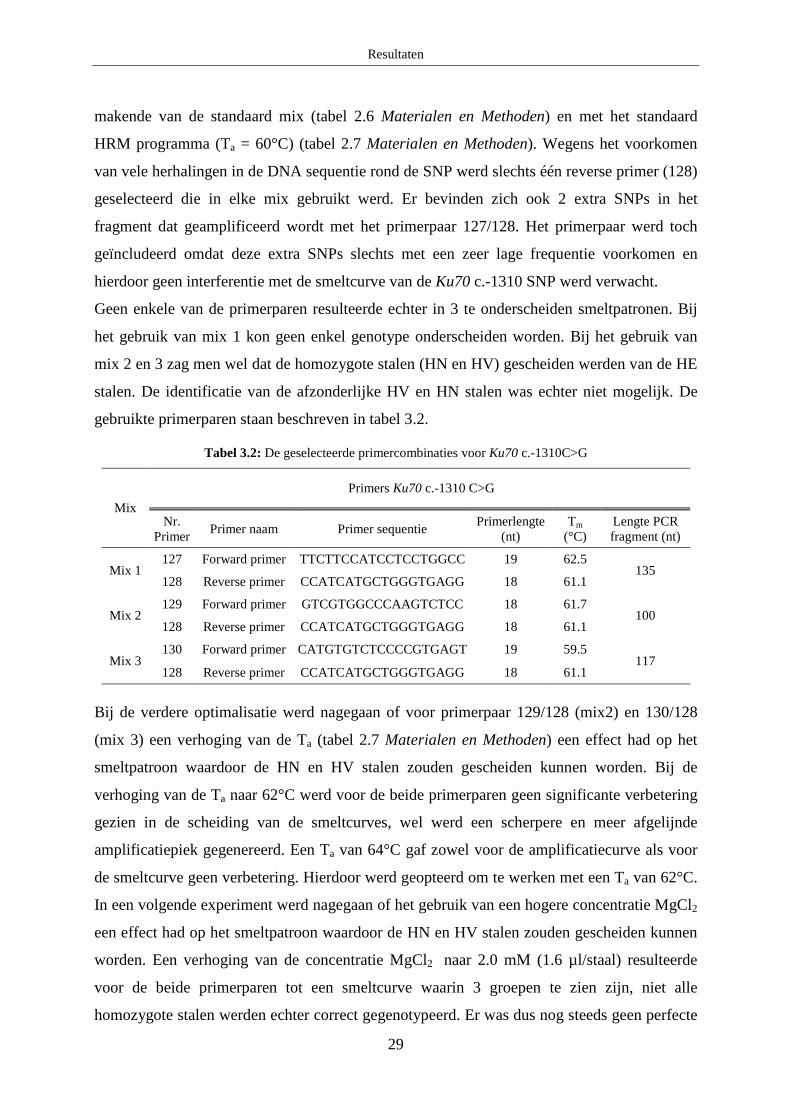
Resultaten
29
makende van de standaard mix (tabel 2.6 Materialen en Methoden) en met het standaard
HRM programma (Ta = 60°C) (tabel 2.7 Materialen en Methoden). Wegens het voorkomen
van vele herhalingen in de DNA sequentie rond de SNP werd slechts één reverse primer (128)
geselecteerd die in elke mix gebruikt werd. Er bevinden zich ook 2 extra SNPs in het
fragment dat geamplificeerd wordt met het primerpaar 127/128. Het primerpaar werd toch
geïncludeerd omdat deze extra SNPs slechts met een zeer lage frequentie voorkomen en
hierdoor geen interferentie met de smeltcurve van de Ku70 c.-1310 SNP werd verwacht.
Geen enkele van de primerparen resulteerde echter in 3 te onderscheiden smeltpatronen. Bij
het gebruik van mix 1 kon geen enkel genotype onderscheiden worden. Bij het gebruik van
mix 2 en 3 zag men wel dat de homozygote stalen (HN en HV) gescheiden werden van de HE
stalen. De identificatie van de afzonderlijke HV en HN stalen was echter niet mogelijk. De
gebruikte primerparen staan beschreven in tabel 3.2.
Tabel 3.2: De geselecteerde primercombinaties voor Ku70 c.-1310C>G
Mix
Primers Ku70 c.-1310 C>G
Nr. Primer
Primer naam Primer sequentie Primerlengte
(nt) Tm
(°C) Lengte PCR fragment (nt)
Mix 1 127 Forward primer TTCTTCCATCCTCCTGGCC 19 62.5
135 128 Reverse primer CCATCATGCTGGGTGAGG 18 61.1
Mix 2 129 Forward primer GTCGTGGCCCAAGTCTCC 18 61.7
100 128 Reverse primer CCATCATGCTGGGTGAGG 18 61.1
Mix 3 130 Forward primer CATGTGTCTCCCCGTGAGT 19 59.5
117 128 Reverse primer CCATCATGCTGGGTGAGG 18 61.1
Bij de verdere optimalisatie werd nagegaan of voor primerpaar 129/128 (mix2) en 130/128
(mix 3) een verhoging van de Ta (tabel 2.7 Materialen en Methoden) een effect had op het
smeltpatroon waardoor de HN en HV stalen zouden gescheiden kunnen worden. Bij de
verhoging van de Ta naar 62°C werd voor de beide primerparen geen significante verbetering
gezien in de scheiding van de smeltcurves, wel werd een scherpere en meer afgelijnde
amplificatiepiek gegenereerd. Een Ta van 64°C gaf zowel voor de amplificatiecurve als voor
de smeltcurve geen verbetering. Hierdoor werd geopteerd om te werken met een Ta van 62°C.
In een volgende experiment werd nagegaan of het gebruik van een hogere concentratie MgCl2
een effect had op het smeltpatroon waardoor de HN en HV stalen zouden gescheiden kunnen
worden. Een verhoging van de concentratie MgCl2 naar 2.0 mM (1.6 µl/staal) resulteerde
voor de beide primerparen tot een smeltcurve waarin 3 groepen te zien zijn, niet alle
homozygote stalen werden echter correct gegenotypeerd. Er was dus nog steeds geen perfecte
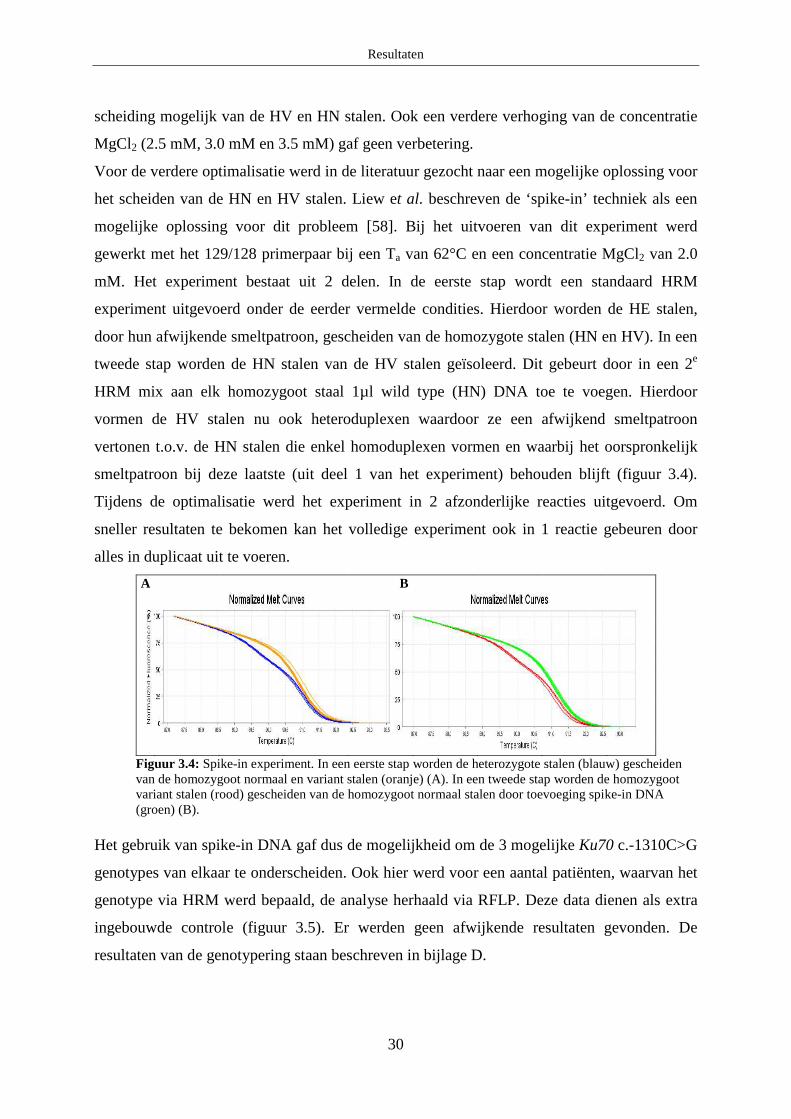
Resultaten
30
scheiding mogelijk van de HV en HN stalen. Ook een verdere verhoging van de concentratie
MgCl2 (2.5 mM, 3.0 mM en 3.5 mM) gaf geen verbetering.
Voor de verdere optimalisatie werd in de literatuur gezocht naar een mogelijke oplossing voor
het scheiden van de HN en HV stalen. Liew et al. beschreven de ‘spike-in’ techniek als een
mogelijke oplossing voor dit probleem [58]. Bij het uitvoeren van dit experiment werd
gewerkt met het 129/128 primerpaar bij een Ta van 62°C en een concentratie MgCl2 van 2.0
mM. Het experiment bestaat uit 2 delen. In de eerste stap wordt een standaard HRM
experiment uitgevoerd onder de eerder vermelde condities. Hierdoor worden de HE stalen,
door hun afwijkende smeltpatroon, gescheiden van de homozygote stalen (HN en HV). In een
tweede stap worden de HN stalen van de HV stalen geïsoleerd. Dit gebeurt door in een 2e
HRM mix aan elk homozygoot staal 1µl wild type (HN) DNA toe te voegen. Hierdoor
vormen de HV stalen nu ook heteroduplexen waardoor ze een afwijkend smeltpatroon
vertonen t.o.v. de HN stalen die enkel homoduplexen vormen en waarbij het oorspronkelijk
smeltpatroon bij deze laatste (uit deel 1 van het experiment) behouden blijft (figuur 3.4).
Tijdens de optimalisatie werd het experiment in 2 afzonderlijke reacties uitgevoerd. Om
sneller resultaten te bekomen kan het volledige experiment ook in 1 reactie gebeuren door
alles in duplicaat uit te voeren.
A
B
Figuur 3.4: Spike-in experiment. In een eerste stap worden de heterozygote stalen (blauw) gescheiden van de homozygoot normaal en variant stalen (oranje) (A). In een tweede stap worden de homozygoot variant stalen (rood) gescheiden van de homozygoot normaal stalen door toevoeging spike-in DNA (groen) (B).
Het gebruik van spike-in DNA gaf dus de mogelijkheid om de 3 mogelijke Ku70 c.-1310C>G
genotypes van elkaar te onderscheiden. Ook hier werd voor een aantal patiënten, waarvan het
genotype via HRM werd bepaald, de analyse herhaald via RFLP. Deze data dienen als extra
ingebouwde controle (figuur 3.5). Er werden geen afwijkende resultaten gevonden. De
resultaten van de genotypering staan beschreven in bijlage D.
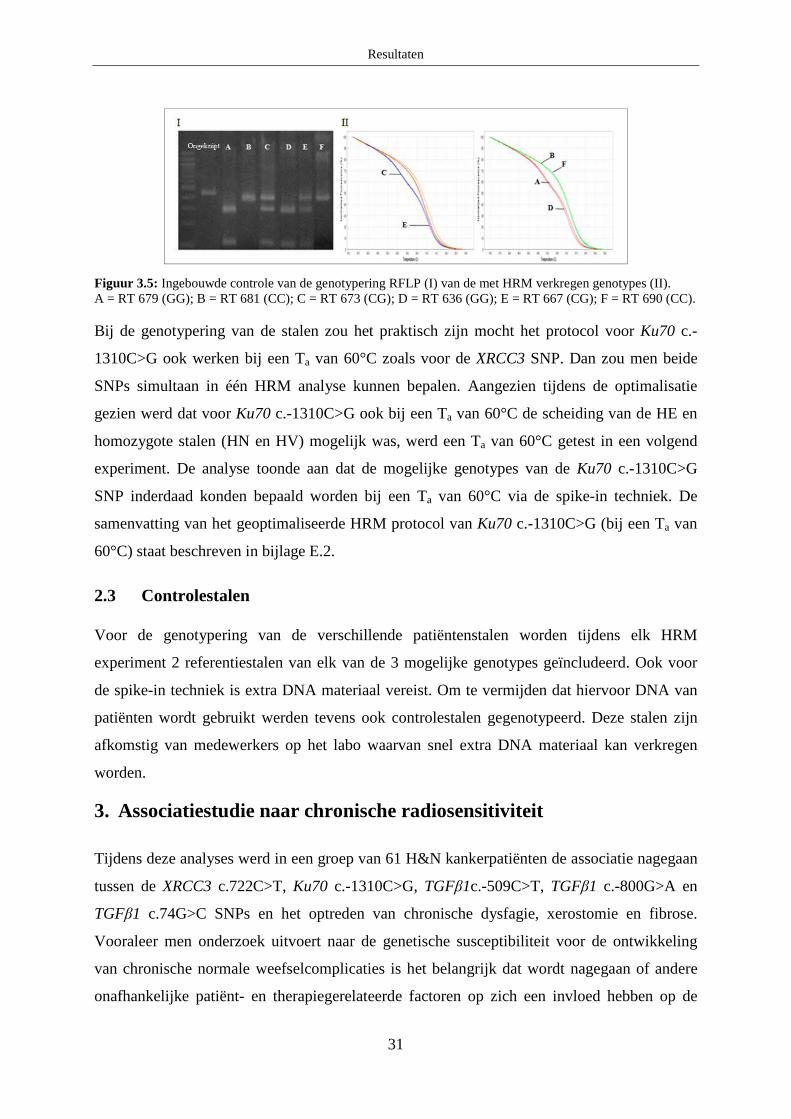
Resultaten
31
Figuur 3.5: Ingebouwde controle van de genotypering RFLP (I) van de met HRM verkregen genotypes (II). A = RT 679 (GG); B = RT 681 (CC); C = RT 673 (CG); D = RT 636 (GG); E = RT 667 (CG); F = RT 690 (CC). Bij de genotypering van de stalen zou het praktisch zijn mocht het protocol voor Ku70 c.-
1310C>G ook werken bij een Ta van 60°C zoals voor de XRCC3 SNP. Dan zou men beide
SNPs simultaan in één HRM analyse kunnen bepalen. Aangezien tijdens de optimalisatie
gezien werd dat voor Ku70 c.-1310C>G ook bij een Ta van 60°C de scheiding van de HE en
homozygote stalen (HN en HV) mogelijk was, werd een Ta van 60°C getest in een volgend
experiment. De analyse toonde aan dat de mogelijke genotypes van de Ku70 c.-1310C>G
SNP inderdaad konden bepaald worden bij een Ta van 60°C via de spike-in techniek. De
samenvatting van het geoptimaliseerde HRM protocol van Ku70 c.-1310C>G (bij een Ta van
60°C) staat beschreven in bijlage E.2.
2.3 Controlestalen
Voor de genotypering van de verschillende patiëntenstalen worden tijdens elk HRM
experiment 2 referentiestalen van elk van de 3 mogelijke genotypes geïncludeerd. Ook voor
de spike-in techniek is extra DNA materiaal vereist. Om te vermijden dat hiervoor DNA van
patiënten wordt gebruikt werden tevens ook controlestalen gegenotypeerd. Deze stalen zijn
afkomstig van medewerkers op het labo waarvan snel extra DNA materiaal kan verkregen
worden.
3. Associatiestudie naar chronische radiosensitiviteit
Tijdens deze analyses werd in een groep van 61 H&N kankerpatiënten de associatie nagegaan
tussen de XRCC3 c.722C>T, Ku70 c.-1310C>G, TGFβ1c.-509C>T, TGFβ1 c.-800G>A en
TGFβ1 c.74G>C SNPs en het optreden van chronische dysfagie, xerostomie en fibrose.
Vooraleer men onderzoek uitvoert naar de genetische susceptibiliteit voor de ontwikkeling
van chronische normale weefselcomplicaties is het belangrijk dat wordt nagegaan of andere
onafhankelijke patiënt- en therapiegerelateerde factoren op zich een invloed hebben op de

Resultaten
32
reactie na blootstelling aan IR. Voor de analyses werden de patiënten voor elke onderzochte
weefselcomplicatie opgedeeld in een niet- tot mild radiosensitieve groep (CTC0-1) en een
matig tot ernstig radiosensitieve groep (CTC2+). De niet- tot mild radiosensitieve groep werd
telkens als referentiegroep beschouwd.
3.1 Analyse genotype-onafhankelijke factoren
Als in deze analyses een p-waarde van kleiner dan 0.05 gevonden wordt, is het verschil tussen
de onderzochte groepen significant en wordt de onderzochte parameter als confounder
beschouwd in het onderzoek naar genetische risicofactoren voor de ontwikkeling van
chronische weefselcomplicaties.
3.1.1. Therapiegerelateerde parameters
De meeste patiënten die geïncludeerd werden in de studie, ondergingen naast de IMRT
behandeling ook chemotherapie en/of HK. De invloed van deze bijkomende behandelingen op
de ontwikkeling van matige- tot ernstige late weefselcomplicaties (CTC2+) werd onderzocht
voor elke complicatie beschouwd in deze studie. Ook de combinaties RT+chemo, RT+HK en
RT+chemo+HK werden onderzocht. Hiernaast werd ook het effect van de dosisparameters
onderzocht. Deze parameters werden berekend op basis van de verkregen DVH data voor de
kritische structuren met betrekking tot de verschillende weefselcomplicaties. De
dosisparameters onderzocht voor elk van de chronische eindpunten worden voorgesteld in
bijlage F.1. Geen enkele therapiegerelateerde parameter lijkt een invloed te hebben op de
ontwikkeling van late stralingstoxiciteiten.
3.1.2. Patiëntgerelateerde parameters
Met betrekking tot de patiënt specifieke parameters die mogelijk een rol spelen in het ontstaan
van ernstige normale weefselcomplicaties werden de niet- tot mild radiosensitieve en de matig
tot ernstig radiosensitieve groep vergeleken voor mogelijke verschillen in leeftijd, geslacht,
PY en drinks/week. Geen van deze parameters vertoonde significante verschillen tussen de
CTC0-1 en CTC2+ groep.
3.2 Analyse naar de associatie tussen SNPs en chronische radiosensitiviteit
Vooraleer deze analyse kon gebeuren, werd gecontroleerd of er voldaan werd aan het HWE.
Hierbij wordt nagegaan of de geobserveerde genotypefrequenties overeenkomen met wat
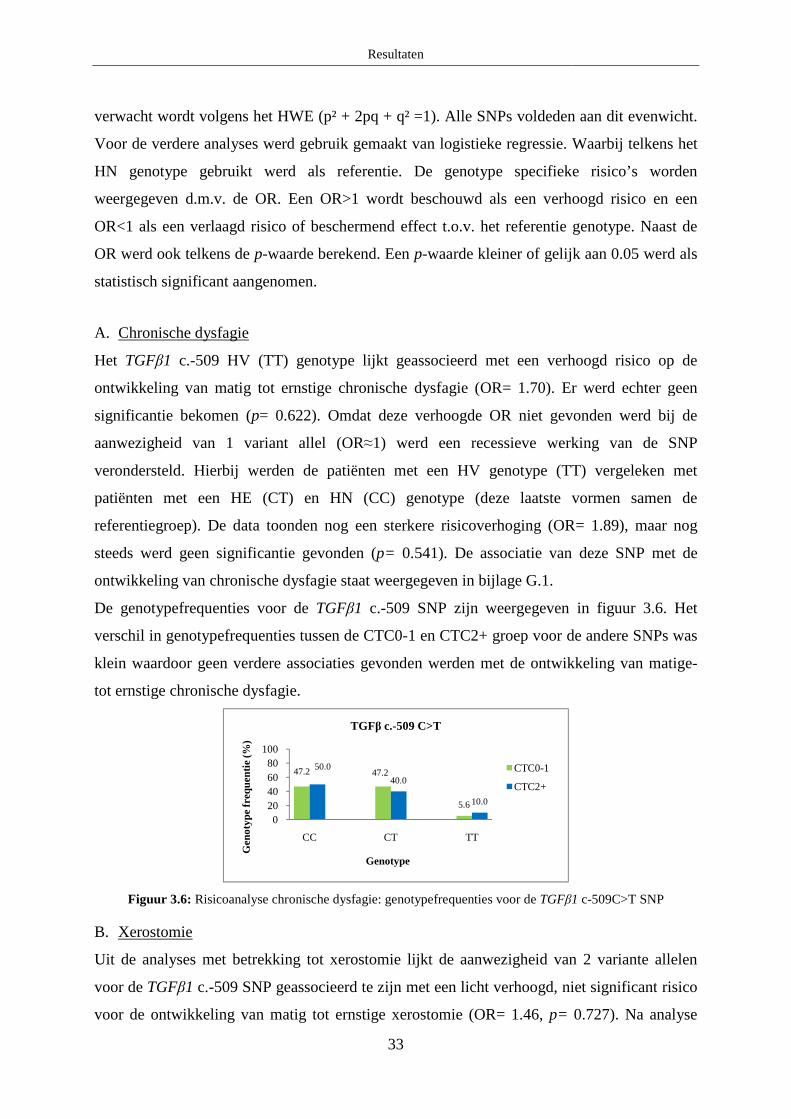
verwacht wordt volgens het HWE (p² + 2pq + q² =1).
Voor de verdere analyses werd gebruik gemaakt van logistieke regressie
HN genotype gebruikt werd
weergegeven d.m.v. de OR.
OR<1 als een verlaagd risico of
OR werd ook telkens de p-waarde berekend. Een
statistisch significant aangenomen.
A. Chronische dysfagie
Het TGFβ1 c.-509 HV (TT)
ontwikkeling van matig tot e
significantie bekomen (p= 0.622
aanwezigheid van 1 variant allel (OR
verondersteld. Hierbij werden de patiënten met een
patiënten met een HE (CT) en
referentiegroep). De data toonden nog een sterkere risicoverhoging (OR=
steeds werd geen significantie g
ontwikkeling van chronische dysfagie staat weergegeven in bijlage
De genotypefrequenties voor
verschil in genotypefrequenties tussen de CTC0
klein waardoor geen verdere associaties gevonden werden met de ontwikkeling van matige
tot ernstige chronische dysfagie.
Figuur 3.6: Risicoanalyse chronische dy
B. Xerostomie
Uit de analyses met betrekking tot xerostomie
voor de TGFβ1 c.-509 SNP geassocieerd te zijn met
voor de ontwikkeling van matig
020406080
100
Gen
otyp
e fr
eque
ntie
(%)
Resultaten
33
ens het HWE (p² + 2pq + q² =1). Alle SNPs voldeden aan dit evenwicht.
werd gebruik gemaakt van logistieke regressie. Waarbij
werd als referentie. De genotype specifieke risico’s worden
de OR. Een OR>1 wordt beschouwd als een verhoogd risico en een
OR<1 als een verlaagd risico of beschermend effect t.o.v. het referentie genotype. Naast de
waarde berekend. Een p-waarde kleiner of gelijk aan 0.05
tistisch significant aangenomen.
) genotype lijkt geassocieerd met een verhoogd risico op de
ernstige chronische dysfagie (OR= 1.70). Er werd echter geen
0.622). Omdat deze verhoogde OR niet gevonden werd bij de
aanwezigheid van 1 variant allel (OR≈1) werd een recessieve werking van de SNP
Hierbij werden de patiënten met een HV genotype (TT) vergeleken met
(CT) en HN (CC) genotype (deze laatste vormen
De data toonden nog een sterkere risicoverhoging (OR=
ie gevonden (p= 0.541). De associatie van deze SNP met de
ontwikkeling van chronische dysfagie staat weergegeven in bijlage G.1.
De genotypefrequenties voor de TGFβ1 c.-509 SNP zijn weergegeven in
verschil in genotypefrequenties tussen de CTC0-1 en CTC2+ groep voor de andere SNPs was
klein waardoor geen verdere associaties gevonden werden met de ontwikkeling van matige
tot ernstige chronische dysfagie.
chronische dysfagie: genotypefrequenties voor de TGFβ1
Uit de analyses met betrekking tot xerostomie lijkt de aanwezigheid van 2 variante allelen
geassocieerd te zijn met een licht verhoogd, niet significant
matig tot ernstige xerostomie (OR= 1.46, p=
47.2 47.2
5.6
50.0
40.0
10.0
CC CT TT
Genotype
TGFβ c.-509 C>T
CTC0-1
CTC2+
Alle SNPs voldeden aan dit evenwicht.
Waarbij telkens het
De genotype specifieke risico’s worden
beschouwd als een verhoogd risico en een
effect t.o.v. het referentie genotype. Naast de
kleiner of gelijk aan 0.05 werd als
geassocieerd met een verhoogd risico op de
Er werd echter geen
). Omdat deze verhoogde OR niet gevonden werd bij de
recessieve werking van de SNP
genotype (TT) vergeleken met
deze laatste vormen samen de
De data toonden nog een sterkere risicoverhoging (OR= 1.89), maar nog
. De associatie van deze SNP met de
weergegeven in figuur 3.6. Het
1 en CTC2+ groep voor de andere SNPs was
klein waardoor geen verdere associaties gevonden werden met de ontwikkeling van matige-
β1 c-509C>T SNP
de aanwezigheid van 2 variante allelen
, niet significant risico
= 0.727). Na analyse
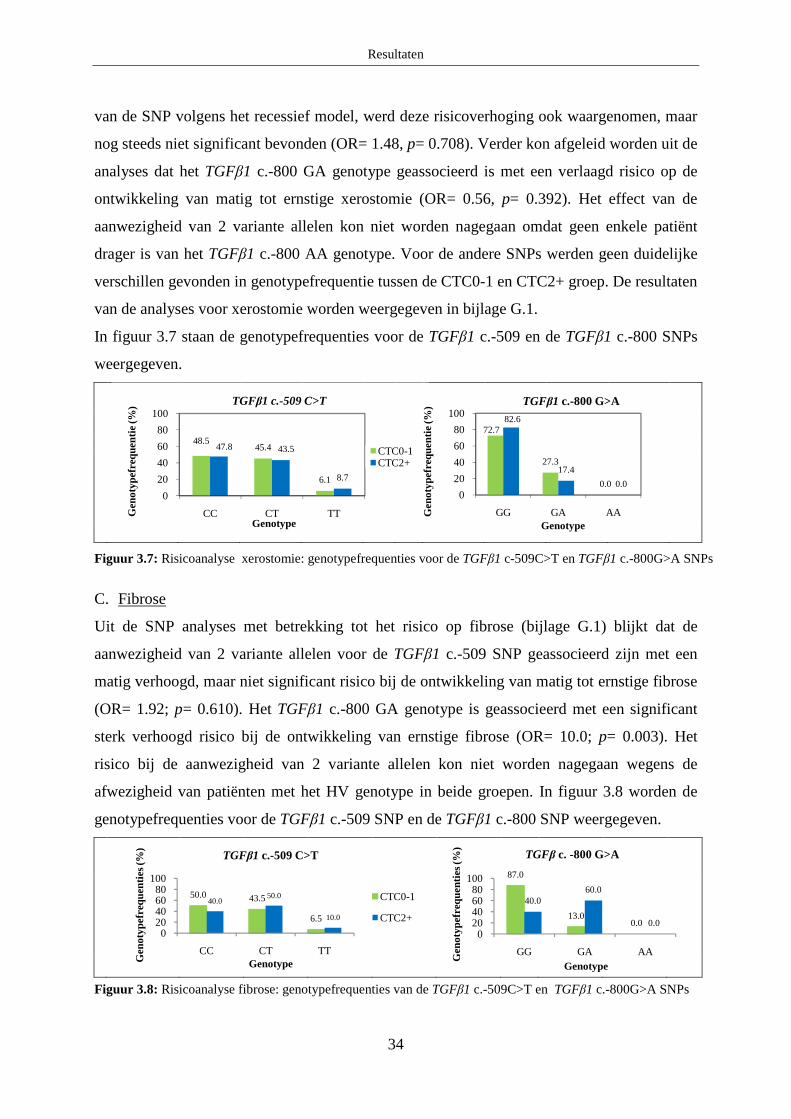
Resultaten
34
van de SNP volgens het recessief model, werd deze risicoverhoging ook waargenomen, maar
nog steeds niet significant bevonden (OR= 1.48, p= 0.708). Verder kon afgeleid worden uit de
analyses dat het TGFβ1 c.-800 GA genotype geassocieerd is met een verlaagd risico op de
ontwikkeling van matig tot ernstige xerostomie (OR= 0.56, p= 0.392). Het effect van de
aanwezigheid van 2 variante allelen kon niet worden nagegaan omdat geen enkele patiënt
drager is van het TGFβ1 c.-800 AA genotype. Voor de andere SNPs werden geen duidelijke
verschillen gevonden in genotypefrequentie tussen de CTC0-1 en CTC2+ groep. De resultaten
van de analyses voor xerostomie worden weergegeven in bijlage G.1.
In figuur 3.7 staan de genotypefrequenties voor de TGFβ1 c.-509 en de TGFβ1 c.-800 SNPs
weergegeven.
Figuur 3.7: Risicoanalyse xerostomie: genotypefrequenties voor de TGFβ1 c-509C>T en TGFβ1 c.-800G>A SNPs
C. Fibrose
Uit de SNP analyses met betrekking tot het risico op fibrose (bijlage G.1) blijkt dat de
aanwezigheid van 2 variante allelen voor de TGFβ1 c.-509 SNP geassocieerd zijn met een
matig verhoogd, maar niet significant risico bij de ontwikkeling van matig tot ernstige fibrose
(OR= 1.92; p= 0.610). Het TGFβ1 c.-800 GA genotype is geassocieerd met een significant
sterk verhoogd risico bij de ontwikkeling van ernstige fibrose (OR= 10.0; p= 0.003). Het
risico bij de aanwezigheid van 2 variante allelen kon niet worden nagegaan wegens de
afwezigheid van patiënten met het HV genotype in beide groepen. In figuur 3.8 worden de
genotypefrequenties voor de TGFβ1 c.-509 SNP en de TGFβ1 c.-800 SNP weergegeven.
Figuur 3.8: Risicoanalyse fibrose: genotypefrequenties van de TGFβ1 c.-509C>T en TGFβ1 c.-800G>A SNPs
50.0 43.5
6.5
40.050.0
10.0
020406080
100
CC CT TT
Gen
otyp
efre
quen
ties
(%)
Genotype
TGFβ1 c.-509 C>T
CTC0-1
CTC2+
87.0
13.00.0
40.060.0
0.00
20406080
100
GG GA AA
Gen
otyp
efre
quen
ties
(%)
Genotype
TGFβ c. -800 G>A
48.545.4
6.1
47.8 43.5
8.7
0
20
40
60
80
100
CC CT TTGen
otyp
efre
quen
tie (%
)
Genotype
TGFβ1 c.-509 C>T
CTC0-1CTC2+
72.7
27.3
0.0
82.6
17.4
0.00
20
40
60
80
100
GG GA AAGen
otyp
efre
quen
tie (%
)
Genotype
TGFβ1 c.-800 G>A

Resultaten
35
Voor geen enkele van de andere onderzocht SNPs werd een mogelijke associatie gevonden
met de ontwikkeling van matig tot ernstige fibrose.
4. Associatiestudie naar acute radiosensitiviteit
In dit deel van de thesis werd in een nieuw verzamelde groep H&N kankerpatiënten (n= 79)
de associatie nagegaan tussen de XRCC3 c.722C>T, Ku70 c.-1310C>G, TGFβ1 c.-509C>T,
TGFβ1 c.-800G>A en TGFβ1 c.74G>C SNPs en de ontwikkeling van acute dysfagie,
mucositis en dermatitis. Naast patiënten die de huidige standaard IMRT behandeling
ondergingen (standaardgroep, n= 33) omvat deze groep ook patiënten uit de DD studie (DD
groep, n= 46). Door het verschil in dosisprotocols tussen deze twee groepen zullen de
dosisvolume distributies van deze groepen ook afzonderlijk worden bekeken. Vooraleer het
onderzoek naar genetische susceptibiliteit kon gebeuren werd, net als voor de analyses naar
chronische weefselcomplicaties, eerst het eventuele effect van de verschillende genotype-
onafhankelijke factoren bestudeerd. Voor de verschillende analyses werden de patiënten voor
elke onderzochte weefselcomplicatie opgedeeld in een niet- tot matig radiosensitieve groep
(CTC0-2) en een sterk radiosensitieve groep (CTC3+). De niet- tot matig radiosensitieve
groep werd telkens als referentiegroep beschouwd.
4.1 Analyse genotype-onafhankelijke factoren
4.1.1 Therapiegerelateerde parameters
A. Acute dysfagie
Totale patiëntengroep
Uit de analyses naar het effect van bijkomende behandelingen (chemotherapie en HK) bleek
dat patiënten in de CTC3+ groep voor acute dysfagie significant meer behandeld werden met
concomitante chemotherapie (p= 0.048). Met betrekking tot de dosisparameters op de
kritische structuren werd voor ernstige acute dysfagie (SAD) enkel de Dmean op de SPC als
“borderline” significant bevonden (p= 0.052). Voor de andere acute eindpunten werden geen
significante verschillen in de beschouwde dosisparameters of bijkomende behandelingen
waargenomen tussen de CTC0-2 en CTC3+ groep (bijlage F.2).
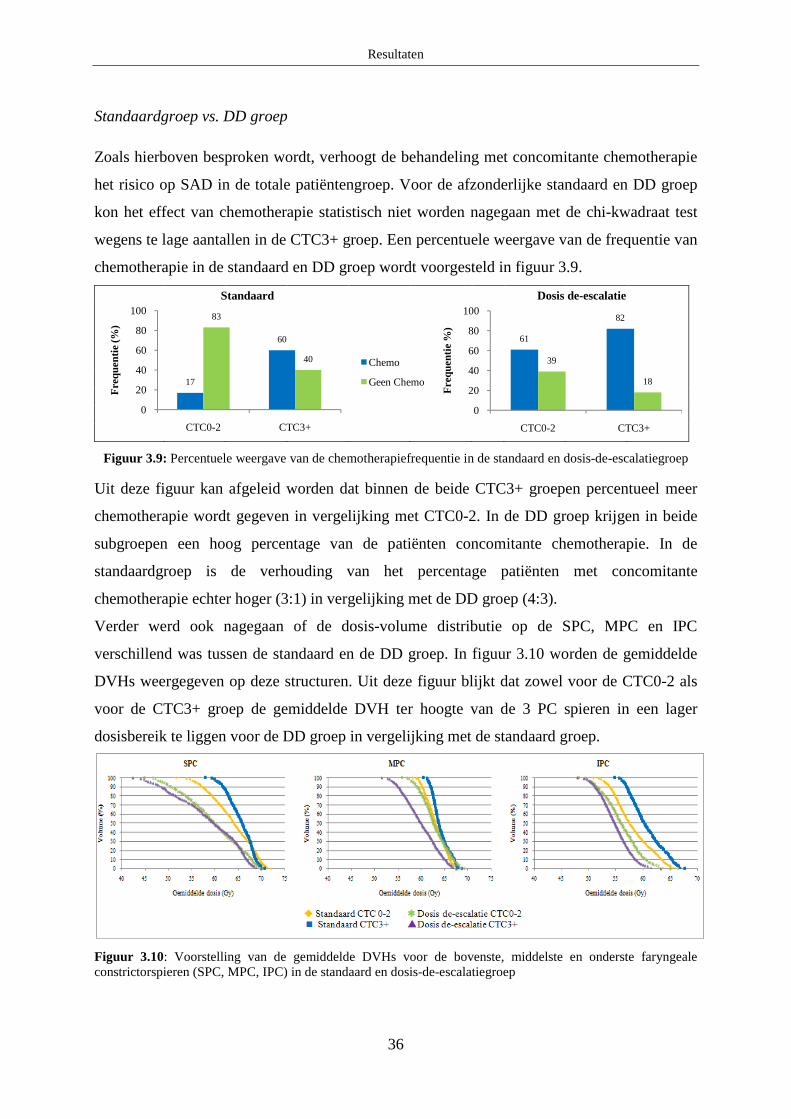
Resultaten
36
Standaardgroep vs. DD groep
Zoals hierboven besproken wordt, verhoogt de behandeling met concomitante chemotherapie
het risico op SAD in de totale patiëntengroep. Voor de afzonderlijke standaard en DD groep
kon het effect van chemotherapie statistisch niet worden nagegaan met de chi-kwadraat test
wegens te lage aantallen in de CTC3+ groep. Een percentuele weergave van de frequentie van
chemotherapie in de standaard en DD groep wordt voorgesteld in figuur 3.9.
Figuur 3.9: Percentuele weergave van de chemotherapiefrequentie in de standaard en dosis-de-escalatiegroep
Uit deze figuur kan afgeleid worden dat binnen de beide CTC3+ groepen percentueel meer
chemotherapie wordt gegeven in vergelijking met CTC0-2. In de DD groep krijgen in beide
subgroepen een hoog percentage van de patiënten concomitante chemotherapie. In de
standaardgroep is de verhouding van het percentage patiënten met concomitante
chemotherapie echter hoger (3:1) in vergelijking met de DD groep (4:3).
Verder werd ook nagegaan of de dosis-volume distributie op de SPC, MPC en IPC
verschillend was tussen de standaard en de DD groep. In figuur 3.10 worden de gemiddelde
DVHs weergegeven op deze structuren. Uit deze figuur blijkt dat zowel voor de CTC0-2 als
voor de CTC3+ groep de gemiddelde DVH ter hoogte van de 3 PC spieren in een lager
dosisbereik te liggen voor de DD groep in vergelijking met de standaard groep.
Figuur 3.10: Voorstelling van de gemiddelde DVHs voor de bovenste, middelste en onderste faryngeale constrictorspieren (SPC, MPC, IPC) in de standaard en dosis-de-escalatiegroep
61
82
39
18
0
20
40
60
80
100
CTC0-2 CTC3+
Fre
quen
tie %
)
Dosis de-escalatie
17
60
83
40
0
20
40
60
80
100
CTC0-2 CTC3+
Fre
quen
tie (%
)
Standaard
Chemo
Geen Chemo
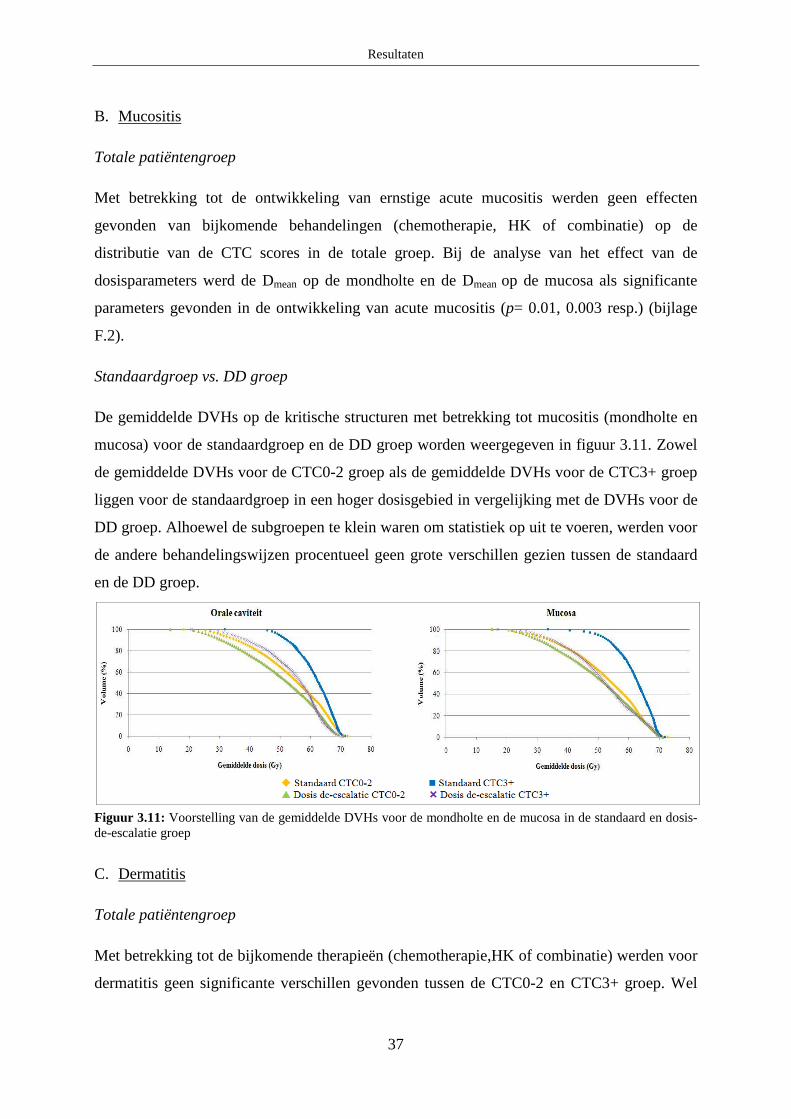
Resultaten
37
B. Mucositis
Totale patiëntengroep
Met betrekking tot de ontwikkeling van ernstige acute mucositis werden geen effecten
gevonden van bijkomende behandelingen (chemotherapie, HK of combinatie) op de
distributie van de CTC scores in de totale groep. Bij de analyse van het effect van de
dosisparameters werd de Dmean op de mondholte en de Dmean op de mucosa als significante
parameters gevonden in de ontwikkeling van acute mucositis (p= 0.01, 0.003 resp.) (bijlage
F.2).
Standaardgroep vs. DD groep
De gemiddelde DVHs op de kritische structuren met betrekking tot mucositis (mondholte en
mucosa) voor de standaardgroep en de DD groep worden weergegeven in figuur 3.11. Zowel
de gemiddelde DVHs voor de CTC0-2 groep als de gemiddelde DVHs voor de CTC3+ groep
liggen voor de standaardgroep in een hoger dosisgebied in vergelijking met de DVHs voor de
DD groep. Alhoewel de subgroepen te klein waren om statistiek op uit te voeren, werden voor
de andere behandelingswijzen procentueel geen grote verschillen gezien tussen de standaard
en de DD groep.
Figuur 3.11: Voorstelling van de gemiddelde DVHs voor de mondholte en de mucosa in de standaard en dosis- de-escalatie groep
C. Dermatitis
Totale patiëntengroep
Met betrekking tot de bijkomende therapieën (chemotherapie,HK of combinatie) werden voor
dermatitis geen significante verschillen gevonden tussen de CTC0-2 en CTC3+ groep. Wel
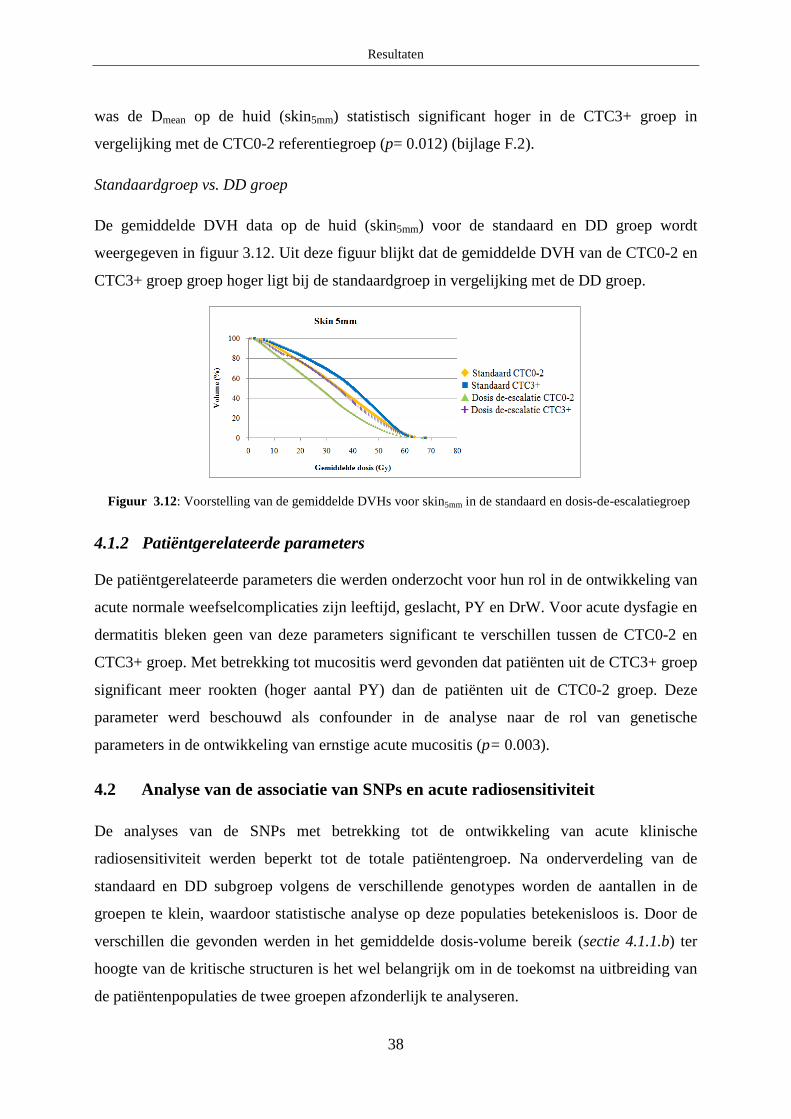
Resultaten
38
was de Dmean op de huid (skin5mm) statistisch significant hoger in de CTC3+ groep in
vergelijking met de CTC0-2 referentiegroep (p= 0.012) (bijlage F.2).
Standaardgroep vs. DD groep
De gemiddelde DVH data op de huid (skin5mm) voor de standaard en DD groep wordt
weergegeven in figuur 3.12. Uit deze figuur blijkt dat de gemiddelde DVH van de CTC0-2 en
CTC3+ groep groep hoger ligt bij de standaardgroep in vergelijking met de DD groep.
Figuur 3.12: Voorstelling van de gemiddelde DVHs voor skin5mm in de standaard en dosis-de-escalatiegroep
4.1.2 Patiëntgerelateerde parameters
De patiëntgerelateerde parameters die werden onderzocht voor hun rol in de ontwikkeling van
acute normale weefselcomplicaties zijn leeftijd, geslacht, PY en DrW. Voor acute dysfagie en
dermatitis bleken geen van deze parameters significant te verschillen tussen de CTC0-2 en
CTC3+ groep. Met betrekking tot mucositis werd gevonden dat patiënten uit de CTC3+ groep
significant meer rookten (hoger aantal PY) dan de patiënten uit de CTC0-2 groep. Deze
parameter werd beschouwd als confounder in de analyse naar de rol van genetische
parameters in de ontwikkeling van ernstige acute mucositis (p= 0.003).
4.2 Analyse van de associatie van SNPs en acute radiosensitiviteit
De analyses van de SNPs met betrekking tot de ontwikkeling van acute klinische
radiosensitiviteit werden beperkt tot de totale patiëntengroep. Na onderverdeling van de
standaard en DD subgroep volgens de verschillende genotypes worden de aantallen in de
groepen te klein, waardoor statistische analyse op deze populaties betekenisloos is. Door de
verschillen die gevonden werden in het gemiddelde dosis-volume bereik (sectie 4.1.1.b) ter
hoogte van de kritische structuren is het wel belangrijk om in de toekomst na uitbreiding van
de patiëntenpopulaties de twee groepen afzonderlijk te analyseren.
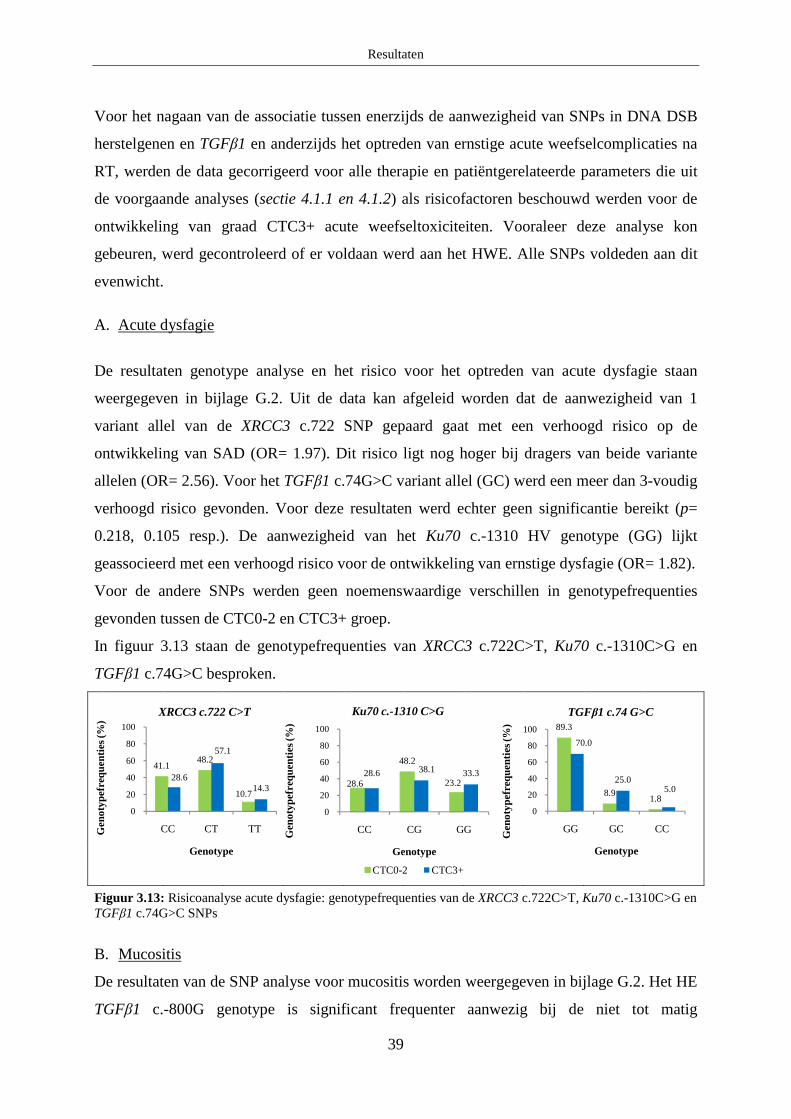
Resultaten
39
Voor het nagaan van de associatie tussen enerzijds de aanwezigheid van SNPs in DNA DSB
herstelgenen en TGFβ1 en anderzijds het optreden van ernstige acute weefselcomplicaties na
RT, werden de data gecorrigeerd voor alle therapie en patiëntgerelateerde parameters die uit
de voorgaande analyses (sectie 4.1.1 en 4.1.2) als risicofactoren beschouwd werden voor de
ontwikkeling van graad CTC3+ acute weefseltoxiciteiten. Vooraleer deze analyse kon
gebeuren, werd gecontroleerd of er voldaan werd aan het HWE. Alle SNPs voldeden aan dit
evenwicht.
A. Acute dysfagie
De resultaten genotype analyse en het risico voor het optreden van acute dysfagie staan
weergegeven in bijlage G.2. Uit de data kan afgeleid worden dat de aanwezigheid van 1
variant allel van de XRCC3 c.722 SNP gepaard gaat met een verhoogd risico op de
ontwikkeling van SAD (OR= 1.97). Dit risico ligt nog hoger bij dragers van beide variante
allelen (OR= 2.56). Voor het TGFβ1 c.74G>C variant allel (GC) werd een meer dan 3-voudig
verhoogd risico gevonden. Voor deze resultaten werd echter geen significantie bereikt (p=
0.218, 0.105 resp.). De aanwezigheid van het Ku70 c.-1310 HV genotype (GG) lijkt
geassocieerd met een verhoogd risico voor de ontwikkeling van ernstige dysfagie (OR= 1.82).
Voor de andere SNPs werden geen noemenswaardige verschillen in genotypefrequenties
gevonden tussen de CTC0-2 en CTC3+ groep.
In figuur 3.13 staan de genotypefrequenties van XRCC3 c.722C>T, Ku70 c.-1310C>G en
TGFβ1 c.74G>C besproken.
Figuur 3.13: Risicoanalyse acute dysfagie: genotypefrequenties van de XRCC3 c.722C>T, Ku70 c.-1310C>G en TGFβ1 c.74G>C SNPs
B. Mucositis
De resultaten van de SNP analyse voor mucositis worden weergegeven in bijlage G.2. Het HE
TGFβ1 c.-800G genotype is significant frequenter aanwezig bij de niet tot matig
41.148.2
10.7
28.6
57.1
14.3
0
20
40
60
80
100
CC CT TTGen
otyp
efre
quen
ties
(%)
Genotype
XRCC3 c.722 C>T
28.6
48.2
23.228.6 38.1 33.3
0
20
40
60
80
100
CC CG GGGen
otyp
efre
quen
ties
(%)
Genotype
Ku70 c.-1310 C>G
CTC0-2 CTC3+
89.3
8.91.8
70.0
25.05.0
0
20
40
60
80
100
GG GC CC
Gen
otyp
efre
quen
ties
(%)
Genotype
TGFβ1 c.74 G>C
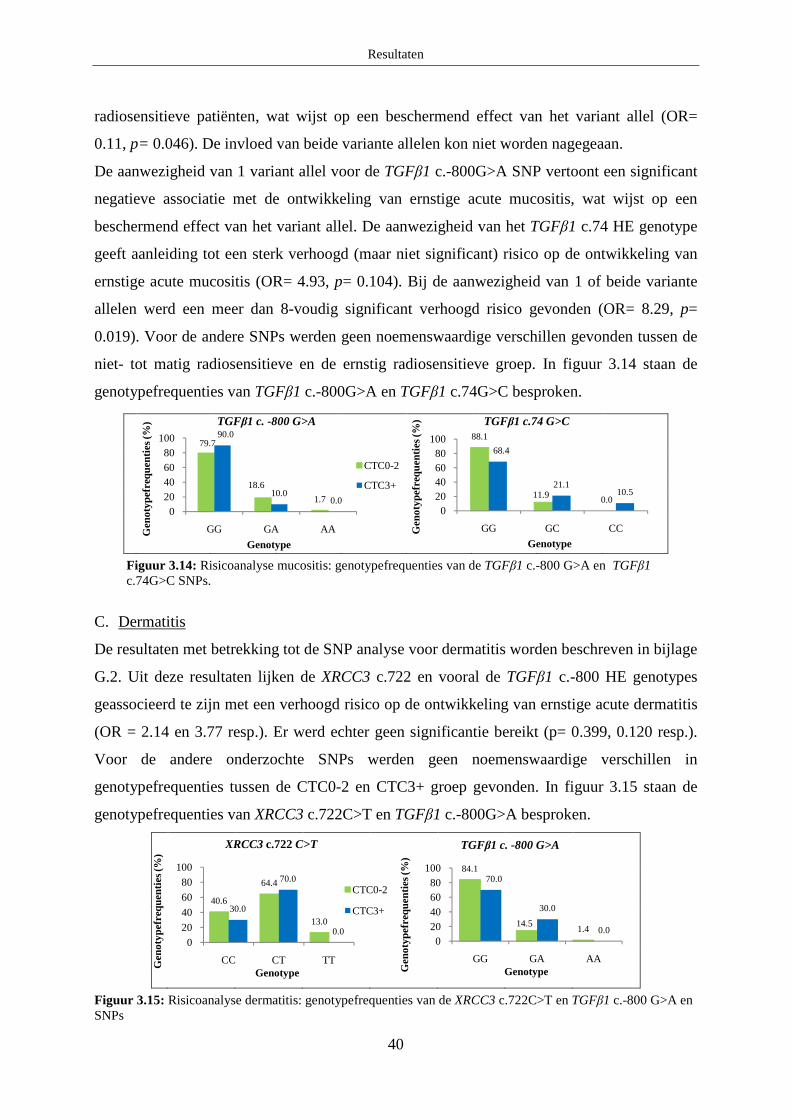
Resultaten
40
radiosensitieve patiënten, wat wijst op een beschermend effect van het variant allel (OR=
0.11, p= 0.046). De invloed van beide variante allelen kon niet worden nagegeaan.
De aanwezigheid van 1 variant allel voor de TGFβ1 c.-800G>A SNP vertoont een significant
negatieve associatie met de ontwikkeling van ernstige acute mucositis, wat wijst op een
beschermend effect van het variant allel. De aanwezigheid van het TGFβ1 c.74 HE genotype
geeft aanleiding tot een sterk verhoogd (maar niet significant) risico op de ontwikkeling van
ernstige acute mucositis (OR= 4.93, p= 0.104). Bij de aanwezigheid van 1 of beide variante
allelen werd een meer dan 8-voudig significant verhoogd risico gevonden (OR= 8.29, p=
0.019). Voor de andere SNPs werden geen noemenswaardige verschillen gevonden tussen de
niet- tot matig radiosensitieve en de ernstig radiosensitieve groep. In figuur 3.14 staan de
genotypefrequenties van TGFβ1 c.-800G>A en TGFβ1 c.74G>C besproken.
Figuur 3.14: Risicoanalyse mucositis: genotypefrequenties van de TGFβ1 c.-800 G>A en TGFβ1 c.74G>C SNPs.
C. Dermatitis
De resultaten met betrekking tot de SNP analyse voor dermatitis worden beschreven in bijlage
G.2. Uit deze resultaten lijken de XRCC3 c.722 en vooral de TGFβ1 c.-800 HE genotypes
geassocieerd te zijn met een verhoogd risico op de ontwikkeling van ernstige acute dermatitis
(OR = 2.14 en 3.77 resp.). Er werd echter geen significantie bereikt (p= 0.399, 0.120 resp.).
Voor de andere onderzochte SNPs werden geen noemenswaardige verschillen in
genotypefrequenties tussen de CTC0-2 en CTC3+ groep gevonden. In figuur 3.15 staan de
genotypefrequenties van XRCC3 c.722C>T en TGFβ1 c.-800G>A besproken.
Figuur 3.15: Risicoanalyse dermatitis: genotypefrequenties van de XRCC3 c.722C>T en TGFβ1 c.-800 G>A en SNPs
40.6
64.4
13.030.0
70.0
0.00
20
40
60
80
100
CC CT TTGen
otyp
efre
quen
ties
(%)
Genotype
XRCC3 c.722C>T
CTC0-2
CTC3+
84.1
14.51.4
70.0
30.0
0.00
20406080
100
GG GA AA
Gen
otyp
efre
quen
ties
(%)
Genotype
TGFβ1 c. -800 G>A
79.7
18.6
1.7
90.0
10.00.0
020406080
100
GG GA AAGen
otyp
efre
quen
ties
(%)
Genotype
TGFβ1 c. -800 G>A
CTC0-2
CTC3+
88.1
11.9 0.0
68.4
21.110.5
020406080
100
GG GC CCGen
otyp
efre
quen
ties
(%)
Genotype
TGFβ1 c.74 G>C

Discussie
41
Discussie
In deze thesis werd de associatie onderzocht tussen SNPs in DNA DSB herstelgenen (XRCC3
c.722C>T en Ku70 c.-1310C>G), TGFβ1 (c.-509C>T, c.-800G>A en c.74G>C) en de
ontwikkeling van acute (mucositis, dermatitis en dysfagie) en chronische (fibrose, xerostomie
en dysfagie) normale weefselcomplicaties. Een onderdeel van deze thesis was ook de
optimalisatie van de HRM protocols voor XRCC3 c.722C>T en Ku70 c.-1310C>G.
1. Optimalisatie HRM protocols
1.1 XRCC3 c.722 C>T
De optimalisatie van dit protocol verliep vlot. Het juiste primerpaar en reactiecondities werd
bij de eerste optimalisatiereactie gevonden, waardoor alle patiëntenstalen snel gegenotypeerd
konden worden. Een reden voor de snelle optimalisatie ligt in het feit dat de XRCC3
c.722C>T variant een klasse 1 SNP is, waarbij het verschil in smelttemperatuur tussen de HN
en HV stalen redelijk hoog ligt (>0.5°C) en dus gemakkelijk te onderscheiden is [57].
1.2 Ku70 c.-1310 C>G
De optimalisatie van het Ku70 c.-1310C>G (een klasse 3 SNP) protocol verliep minder vlot.
Dit polymorfisme behoort tot de klasse 3 SNPs, waarbij het verschil in Tm tussen de HN en
HV stalen kleiner is dan 0.2°C. Geen van de geselecteerde primerparen leek ideaal te zijn en
enkel de HE stalen konden geïdentificeerd worden. Ook na verhoging van de Ta en de MgCl2
concentratie konden de homozygote stalen niet onderscheiden worden. In de literatuur werd
de spike-in techniek voorgesteld als oplossing voor dit probleem. Toevoeging van een zelfde
hoeveelheid HN DNA maakte het uiteindelijk wel mogelijk alle stalen te genotyperen. Een
nadeel van deze techniek is echter dat het experiment dubbel moet worden uitgevoerd,
éénmaal zonder en éénmaal met gekend HN DNA waardoor deze methode meer materiaal
kost [57].
2. Associatiestudie naar chronische en acute radiosensitiviteit
De SNPs onderzocht in deze thesis zijn gelegen in de promotor of coderende regio’s van het
gen en kunnen dus een belangrijke invloed hebben op de hoeveelheid en de functionaliteit van
het geproduceerde eiwit.

Discussie
42
2.1 Chronische normale weefselcomplicaties
In het onderzoek naar chronische weefselcomplicaties speelden in deze thesis noch de
onderzochte dosisparameters en bijkomende behandelingen, noch de patiëntgerelateerde
parameters een rol in de ontwikkeling van late toxiciteiten in de gezonde weefsels.
2.1.1 XRCC3 c.722 C>T
Met betrekking tot de ontwikkeling van chronische weefselcomplicaties kon in deze thesis
geen associatie aangetoond worden met de XRCC3 c.722C>T SNP. In overeenstemming met
deze resultaten kon ook in de studie van Alsbeih et al. geen associatie aangetoond worden bij
het onderzoek naar de ontwikkeling van fibrose na RT bij patiënten met tumoren in de
nasofarynx [52]. In de studie van Andreassen et al. werd deze SNP wel geassocieerd met de
ontwikkeling van ernstige fibrose bij patiënten behandelt met RT na mastectomie [40]. Deze
associatie kon in een grotere validatie studie echter niet meer bevestigd worden[43]. Met
betrekking tot de ontwikkeling van ernstige late weefselcomplicaties na behandeling van
prostaatkankerpatiënten werd in de studie van Burri et al. een significante associatie
gevonden met deze SNP [51]. Het feit dat in deze thesis geen associatie gevonden werd kan
enerzijds te wijten zijn aan de mogelijks weefselspecifieke werking van het XRCC3 eiwit met
betrekking tot chronische toxiciteit. Anderzijds is XRCC3 een eiwit betrokken in het herstel
van DSBs, een proces dat werkzaam is in de periode tijdens en kort na de
stralingsblootstelling.
2.1.2 Ku70 c.-1310 C>G
In deze thesis kon geen associatie aangetoond worden tussen de ontwikkeling van chronische
weefselcomplicaties en de Ku70 c.-1310C>G SNP. Slechts in een enkele studie werd de rol
van deze SNP naar chronische toxiciteit onderzocht. Hierbij werd het variant allel
geassocieerd met een verhoogd risico op de ontwikkeling van chronische
weefselcomplicaties, meerbepaald bij de ontwikkeling van dysurie bij prostaat-
kankerpatiënten na RT [48]. De verschillen in protocol (indeling patiënten, type
weefselcomplicaties, gebruik van carbon ion RT) kunnen een verklaring zijn voor de
verschillende resultaten.

Discussie
43
2.1.3 TGFβ1 c.-509 C>T
In deze thesis werd het HV TGFβ1 c.-509 genotype (TT) geassocieerd met een
risicoverhoging voor de ontwikkeling van matig tot ernstige fibrose (OR= 1.92). Ook met
betrekking tot de ontwikkeling van CTC2+ graad chronische dysfagie en xerostomie werd het
TT genotype geassocieerd met een risicoverhoging (OR= 1.70; 1.46 resp.). De rol van deze
SNP in de ontwikkeling van xerostomie en chronische dysfagie bij H&N kankerpatiënten
werd echter nog niet onderzocht door andere onderzoeksgroepen, waardoor geen vergelijking
met literatuurdata mogelijk was voor deze weefselcomplicaties. Slechts een enkele studie met
betrekking tot fibrose bij H&N kankerpatiënten werd gevonden. Hier kon echter geen
associatie aangetoond worden [44].
Met betrekking tot andere kankertypes werden wel reeds verschillende studies uitgevoerd met
oog op het vinden van een associatie tussen dit polymorfisme en fibrose na de behandeling
met RT. In de studie van Quarmby et al. werd een significant positieve associatie gevonden
tussen het c.-509 TT genotype en het optreden van ernstige stralinggeïnduceerde fibrose bij
borstkankerpatiënten (OR= 3.4) [39]. In de studies van Andreassen et al. en Giotopoulos et al.
werden deze resultaten bevestigd. In deze laatste studie werd deze SNP zelfs geassocieerd met
een 14.7-voudige risicoverhoging (p< 0.001) [20,40]. Ook in het onderzoek van De Ruyck et
al. naar late klinische radiosensitiviteit bij gynaecologische kankerpatiënten werd dezelfde
trend gezien, maar kon geen significantie aangetoond worden (OR= 4.0). In overeenstemming
met deze thesis werd de recessieve werking van de SNP in deze studie ook gevonden [41].
Ondanks deze overtuigende publicaties werd deze associatie in een meer recent artikel van
Andreassen et al. (2006) niet meer gevonden [43]. Ook in de studie van Yuan et al. werd deze
SNP niet aangeduid als risicofactor bij de ontwikkeling van pneumotitis na behandeling van
longkankerpatiënten met RT [42]. Het is duidelijk dat omtrent deze SNP nog geen consensus
bestaat, hierbij is het ook belangrijk om de gevonden resultaten in een grotere groep patiënten
te valideren.
2.1.4 TGFβ1 c.-800 G>A
Uit de resultaten van de analyses voor de TGFβ1 c.-800G>A SNP met betrekking tot
chronische toxiciteiten bleek dat het c.-800 GA genotype significant geassocieerd is met een
sterk verhoogd risico met de ontwikkeling van matig tot ernstige fibrose (OR= 10.0; p=
0.003). Deze significantie is te wijten aan het zeer grote verschil in de frequentie van het

Discussie
44
variante allel tussen de CTC0-1 en CTC2+ groep (6.5% vs. 30.0%). Omdat de CTC2+ groep
slechts 10 patiënten telt, kan de hoge variante allelfrequentie te wijten zijn aan toeval. Daarom
is het belangrijk dat men deze sterke risicoverhoging nuanceert en dat er verder onderzoek
met een grotere groep patiënten gebeurt om een waardevol besluit te kunnen nemen over de
associatie van deze SNP en de ontwikkeling van ernstige fibrose.
Met betrekking tot de ontwikkeling van xerostomie vertoont het HE genotype (GA) een 2-
voudige protectieve rol. Dit gegeven werd echter niet significant bevonden. De frequenties
van het variante allel lagen voor zowel de CTC0-2 groep als de CTC3+ groep rond de
normale frequentie volgens NCBI (≈10%) [36].
De associatie van deze SNP met de ontwikkeling van chronische weefselcomplicaties bij
H&N kankerpatiënten na behandeling met RT werden tot nog toe niet onderzocht. Met
betrekking tot de ontwikkeling van late weefselcomplicaties na RT behandeling van
gynaecologische tumoren en borstkanker werd voor deze SNP in de studie van De Ruyck et
al. en Quarmby et al. geen associatie gevonden [39,41].
2.1.5 TGFβ1 c.74G>C
In deze thesis werd geen associatie gevonden tussen de TGFβ1 c.74G>C SNP en de
ontwikkeling van chronische weefselcomplicaties na RT bij H&N kankerpatiënten. De SNP
werd tevens onderzocht in de eerder vermelde studies van Quarmby et al., Andreassen et al.,
De Ruyck et al. en Yuan et al. [39-42]. Deze onderzoeken bevestigen de afwezigheid van een
associatie tussen deze SNP en de ontwikkeling van late weefselcomplicaties na RT
behandeling bij borst, gynaecologische- en long kankerpatiënten.
2.2 Acute normale weefselcomplicaties
Binnen de onderzoeksgroep werd reeds een associatie studie uitgevoerd naar de ontwikkeling
van acute radiosensitiviteit en de aanwezigheid van SNPs in DNA herstelgenen (waaronder
XRCC3 c.722C>T en Ku70 c.-1310C>G). Uit deze studie werden sterk significante
verschillen gevonden in de dosis distributie tussen de CTC0-2 en CTC3+ groep. Zowel de
Dmean als de Dmin_circumferentie op de 3 faryngeale constrictorspieren (SPC, MPC en IPC) lagen
significant hoger in de CTC3+ groep. Geen van deze dosisparameters werden echter in deze
thesis significant bevonden in de huidige populatie. Enkel de Dmean op de SPC was
‘borderline’ significant (p= 0.052).

Discussie
45
Wanneer de gemiddelde DVH data op deze structuren vergeleken werd tussen de patiënten uit
de studie van Werbrouck et al. en de patiënten uit deze thesis zag men dat het dosisbereik
voor zowel de CTC0-2 als de CTC3+ groepen lager lag bij de huidige populatie. Dit was ook
te verwachten aangezien de huidige patiëntengroep bestaat uit patiënten uit een dosis-de-
escalatie groep en patiënten behandeld met de huidige standaard (meer weefselsparende)
IMRT protocols. De gemiddelde DVHs van de CTC0-2 en CTC3+ groepen liggen ook minder
ver uiteen voor elk van deze acute weefselcomplicaties.
Uit de studie van Werbrouck et al. bleek chemotherapie geen rol te spelen bij de ontwikkeling
van SAD. Uit de resultaten van deze thesis bleek deze parameter echter wel een belangrijke
rol te spelen bij de ontwikkeling van SAD [19]. Omdat de ervaring in de kliniek aangetoond
heeft dat het geven van concomitante RT een meer efficiënte behandeling van H&N
kankerpatiënten geeft werd in de huidige behandelingsschema’s de toepassing van
concomitante chemotherapie opgedreven in vergelijking met de pilootstudie. Dit verschil
geeft een logische verklaring waarom chemotherapie nu wel een belangrijke parameter is bij
de ontwikkeling van SAD [9,19].
Met betrekking tot de ontwikkeling van ernstige acute mucositis bleek in de studie van
Werbrouck et al. enkel de Dmean ter hoogte van de mondholte significant hoger te liggen in de
CTC3+ groep. In deze thesis werd naast de Dmean ter hoogte van de mondholte ook de Dmean
op de mucosa significant verschillend gevonden tussen de CTC0-2 en CTC3+ groep (p=
0.001, 0.003 resp.) Verder bleek uit de huidige studie ook het rookgedrag een significante rol
te spelen bij de ontwikkeling van ernstige mucositis (p= 0.003).
Bij de ontwikkeling van ernstige dermatitis bleek uit deze thesis de Dmean op de huid (skin5mm)
een significante rol te spelen (p= 0.012), terwijl dat niet het geval was in de studie van
Werbrouck et al. [19].
2.2.1 XRCC3 c.722C>T
Slechts een enkele studie met betrekking tot de associatie van deze SNP en het optreden van
weefselcomplicaties bij H&N kankerpatiënten werd gepubliceerd. Dit is de pilootstudie van
Werbrouck et al. die binnen de onderzoeksgroep werd uitgevoerd. Uit deze studie bleek het
HE XRCC3 c.722 (CT) genotype significant geassocieerd te zijn met een verhoogd risico op
de ontwikkeling van SAD (OR= 4.47) [19]. Ook in de huidige studie werd deze SNP
geassocieerd met een risicoverhoging op SAD (OR= 2.56). Uit de resultaten van deze thesis
bleek dat het HE genotype ook positief geassocieerd is met de ontwikkeling van ernstige

Discussie
46
dermatitis (OR= 2.04). In de studie van Popanda et al. werd de associatie van deze SNP bij de
ontwikkeling van acute toxiciteit bevestigd bij RT-behandelde prostaatkankerpatiënten [50].
Ondanks deze resultaten werd in de literatuur aangetoond dat deze SNP geen invloed heeft op
de functie van het gen binnen de HR [64]. Een mogelijk verklaring voor de gevonden
associatie kan zijn dat het XRCC3 gen in linkage-equilibrium is met een andere (tot op
vandaag ongeïdentificeerde) SNP die verantwoordelijk is voor het ontstaan van de acute
weefseltoxiciteiten.
2.2.2 Ku70 c.-1310 C>G
In deze thesis werd een positieve associatie gevonden tussen de ontwikkeling van SAD en het
HV Ku70 c.-1310 genotype (OR= 1.82). Deze resultaten liggen in lijn met wat gevonden
werd in de studie van Werbrouck et al. waar de aanwezigheid van 1 of 2 variante allelen voor
de Ku70 c.-1310 SNP significant geassocieerd werden met een risicoverhoging bij de
ontwikkeling van SAD na RT bij H&N kankerpatiënten [19]. Geen andere studies die de
associatie tussen deze SNP en acute RT effecten onderzochten werden reeds gepubliceerd.
2.2.3 TGFβ1 c.-509 C>T
De onderzoeken met betrekking tot SNPs in TGFβ1 worden over het algemeen geplaatst in
het kader van late weefselcomplicaties. In deze thesis werd echter ook de invloed van deze
SNPs nagegaan bij de ontwikkeling van acute weefselcomplicaties. Er konden echter geen
associaties aangetoond worden. Ook in de literatuur werd deze SNP tot nog toe niet gelinkt
met de ontwikkeling van acute weefseltoxiciteiten.
2.2.4 TGFβ1 c.-800 G>A
Uit de resultaten van deze thesis bleek het TGFβ1 c.-800 HE genotype een verhoogd risico te
geven voor de ontwikkeling van ernstige acute dermatitis (OR= 3.77). Hierbij moet
opgemerkt worden dat het aantal patiënten in de CTC3+ groep zeer laag is (n= 10) en dat deze
risicoverhoging dus moet genuanceerd worden. Voor ernstige mucositis was het c.-800 GA
genotype significant geassocieerd met een sterk verlaagd risico voor deze complicatie (OR=
0.11; p= 0.046). Wegens de lage aantallen in de CTC3+ groepen, zeker in het onderzoek naar
acute dermatititis, is het noodzakelijk de studie uit te voeren bij een veel grotere groep

Discussie
47
patiënten. Deze SNP werd tot nog toe niet onderzocht naar zijn associatie met de
ontwikkeling van acute weefseltoxiciteit.
2.2.5 TGFβ1 c.74G>C
Uit deze thesis bleek het HE en HV genotype van de TGFβ1 c.74 G>C SNP geassocieerd te
zijn met een risicoverhoging voor de ontwikkeling van SAD (OR= 2.90). Verder bleek bij de
ontwikkeling van ernstige mucositis het TGFβ1 c.74 de aanwezigheid van 1 of beide variante
allelen significant geassocieerd met een sterke risicoverhoging (OR= 8.29; p= 0.019). Deze
sterke risicoverhoging moet ook hier genuanceerd worden door de relatief kleine CTC3+
groep waardoor de hoge variante allelfrequentie in deze groep aan het toeval te wijten kan
zijn.
Door het gebrek aan publicaties die de relatie van deze SNP en de ontwikkeling van acute
weefselcomplicaties onderzochten konden de resultaten van deze thesis met betrekking tot
acute toxiciteit niet worden vergeleken met de literatuur.
Besluit
Uit dit onderzoek lijkt zowel de XRCC3 c.722 SNP (bij dysfagie en dermatitis) als de Ku70
c.-1310 SNP (bij dysfagie) geassocieerd te zijn met de ontwikkeling van acute
weefselcomplicaties. Verder lijkt ook TGFβ1 (c.74, c.-800) een rol te spelen bij het optreden
van acute weefselcomplicaties na RT. Met betrekking tot de ontwikkeling van chronische
weefelcomplicaties bleek zowel de TGFβ1 c.-800 SNP (bij xerostomie en fibrose) als de
TGFβ1 c.-509 SNP ( bij xerostomie, fibrose en dysfagie) een mogelijke rol te spelen. De
associaties naar chronische toxiciteit moeten echter wel genuanceerd worden door het kleine
aantal patiënten in de CTC2+ groepen. Verder onderzoek bij een grotere patiëntenpopulaties
is nodig om de statistische power op te drijven.
Analyse van de dosisparameters toonde aan dat Dmean op de skin5mm, mucosa, mondholte en
SPC sterk verschillen tussen de subgroepen in de analyses naar de ontwikkeling van acute
toxiciteiten. Hoe hoger de dosis op deze structuren hoe sterker het risico op ernstige
dermatitis, mucositis en dysfagie respectievelijk. Ook het gebruik van concomitante
chemotherapie bleek significant geassocieerd met de ontwikkeling van SAD. In tegenstelling
hiermee lijken de therapiegerelateerde parameters echter geen rol te spelen in de ontwikkeling
van chronische toxiciteit in de H&N regio.

Referentielijst
48
Referentielijst
1. Argiris A, Karamouzis MV, Raben D, Ferris RL (2008). Head and neck cancer. Lancet 371(9625):1695-1709. 2. National Cancer Institute (http://www.cancer.gov/cancertopics/types/head-and-neck). 3. Pinar T, Akdur R, Tuncbilek A, Altundag K, Cengiz M (2007). The relationship between occupations and head
and neck cancers. Journal of the national medical association 99(1):64,68-71. 4. Vokes EE, Weichselbaum RR, Lippman SM, Hong WK (1993). Head and neck cancer. The new England journal
of medicine 328(3):184-194. 5. Stelow EB, Mills SE (2005). Squamous cell carcinoma variants of the upper aerodigestive tract. American
journal of clinical pathology 124(Suppl 1):96-109. 6. Werbrouck J, De Ruyck K, Duprez F, Van Eijkeren M, Rietzschel E, Bekaert S, Vral A, De Neve W, Thierens H
(2008). Single-nucleotide polymorphisms in DNA double-strand break repair genes: association with head and neck cancer and interaction with tobacco use and alcohol consumption. Mutation research 656(1-2):74-81.
7. Ang KK (2008). Multidisciplinary management of locally advanced SCCHN: optimizing treatment outcomes. The Oncologist 13(8):899-910.
8. Alderden RA, Hall MD, Hambley TW (2006). The discovery and development of cisplatin. Journal of chemical education 83(5):728-734.
9. Bernier J, Domenge C, Ozsahin M, Matuszewska K, Lefebvre JL, Greiner RH, Giralt J, Maingon P, Rolland F, Bolla M, Cognetti F, Bourhis J, Kirkpatrick A, van Glabbeke M (2004). Postoperative irradiation with or without concomitant chemotherapy for locally advanced head and neck cancer. The new England journal of medicine 350(19)):1945-1954.
10. Bonner JA, Harari PM, Giralt J, Azarnia N, Shin DM, Cohen RB, Jones CU, Sur R, Raben D, Jassem J, Ove R, Kies MS, Baselga J, Youssoufian H, Amellal N, Rowinsky EK, Ang KK (2006). Radiotherapy plus cetuximab for squamous-cell carcinoma of the head and neck. The new England journal of medicine 354(6):567-578.
11. Giro C, Berger B, Bölke E, Ciernik IF, Duprez F, Locati L, Maillard S, Ozsahin M, Pfeffer R, Robertson AG, Langedijk JA, Budach W (2009). High rate of severe radiation dermatitis during radiation therapy with concurrent cetuximab in head and neck cancer: Results of a survey in EORTC institutes. Radiotherapy and Oncology 90(2):166-171.
12. Hendry JH, Jeremic B, Zubizarreta EH (2006). Normal tissue complications after radiation therapy. Pan American journal of public health 20(2-3):151-160.
13. Hong Ts, Tomé WA, Harari PM (2005). Intensity-modulated radiation therapy in the management of head and neck cancer. Current opinion in oncology 17(3):231-235.
14. Bourguignon MH, Gisone PA, Perez MR, Michelin S, Dubner D, Giorgio MD, Carosella ED (2005). Genetic and epigenetic features in radiation sensitivity. Part 2: implications for clinical practice and radiation protection. European journal of nuclear medicine and molecular imaging 32(3):351-368.
15. Nguyen NP, Sallah S, Karlsson U, Antoine JE (2002). Combined chemotherapy and radiation therapy for head and neck malignancies: quality of life issues. Cancer 94(4):1131-1141.
16. Common terminology criteria for adverse events v3.0 (CTCAE) (http://ctep.cancer.gov/protocol Development/electronic_applications/docs/ctcaev3.pdf).
17. Barnett GC, West CM, Dunning AM, Elliott RM, Coles CE, Pharoah PD, Burnet NG (2009). Normal tissue reactions to radiotherapy: towards tailoring treatment dose by genotype. Nature reviews. Cancer 9(2):134-142.
18. Filippi AR, Franco P, Ricardi U (2006). Is clinical radiosensitivity a complex genetically controlled event? Tumori 92(2):87-91.
19. Werbrouck J, De Ruyck K, Duprez F, Veldeman L, Claes K, Van Eijkeren M, Boterberg T, Willems P, Vral A, De Neve W, Thierens H (2009). Acute normal tissue reactions in head-and-neck cancer patients treated with IMRT: influence of dose and association with genetic polymorphisms in DNA DSB repair genes. International journal of radiation oncology, biology, physics 73(4):1187-1195.
20. Giotopoulos G, Symonds RP, Foweraker K, Griffin M, Peat I, Osman A, Plumb M (2007). The late radiotherapy normal tissue injury phenotypes of telangiectasia fibrosis and atrophy in breast cancer patients have distinct genotype-dependent causes. British journal of cancer 96(6):1001-1007.
21. Stone HB, Coleman CN, Anscher MS, McBride WH (2003). Effects of radiation on normal tissue consequences and mechanisms. The lancet oncology 4(9):529-536.
22. Dörr W, Hendry JH (2001). Consequential late effects in normal tissue. Radiotherapy and oncology 61(3):223-231.
23. Murphy BA, Gilbert J, Cmelak A, Ridner SH (2007). Symptom control issues and supportive care for patients with head and neck cancer. Clinical advances in hematology & oncology: H&O 5(10):807-822.
24. Palazzi M, Tomatis S, Orlandi E, Guzzo M, Sangalli C, Potepan P, Fantini S, Bergamini C, Gavazzi C, Licitra L, Scaramellini G, Cantu G, Olmi P (2008). Effects of treatment intensification on acute local toxicity during

Referentielijst
49
radiotherapy for head and neck cancer: prospective observational study validating CTCAE, version 3.0, scoring system. International journal of radiation oncology, biology, physics 70(2):330-337.
25. Scully C, Epstein J, Sonis S (2003). Oral mucositis: a challenging complication of radiotherapy, chemotherapy, and radiochemotherapy: part 1, pathogenesis and prophylaxis of mucositis. Head & Neck 25(12):1057-1070.
26. Henson BS, Inglehart MR, Eisbruch A, Ship JA (2001). Preserved salivary output and xerostomia-related quality of life in head and neck cancer patients receiving parotid-sparing radiotherapy. Oral oncology 37(1):84-93.
27. Eisbruch A, Schwartz M, Rasch C, Vineberg K, Damen E, Van As CJ, Marsh R, Pameijer FA, Balm AJ (2004). Dysphagia and aspiration after chemoradiotherapy for head-and-neck cancer: which anatomic structures are affected and can they be spared with IMRT? International journal of radiation oncology, biology, physics 60(5):1425-1439.
28. Platteaux N, Dirix P, Dejaeger E, Nuyts S (2009). Dysphagia in head and neck cancer patients treated with chemoradiotherapy. Dysphagia in druk.
29. Murphy Ba, Gilberd J (2008). Dysphagia in head and neck cancer patients treated with radiation: assessment, sequelae and rehabilitation. Seminars in radiation oncology 19(1):35-42.
30. Martin M, Lefaix J, Delanian S (2000). TGF-beta1 and radiation fibrosis: a master switch and a specific therapeutic target? International journal of radiation oncology, biology, physics 47(2):277-290.
31. Sakata K, Matsumoto Y, Tauchi H, Satoh M, Oouchi A, Nagakura H, Koito K, Hosoi Y, Suzuki N, Komatsu K, Hareyama M (2001). Expression of genes involved in repair of DNA double-strand breaks in normal and tumor tissues. International journal of radiation oncology, biology, physics 49(1):161-167.
32. Weterings Eric, Chen DJ (2008). The endless tale of non-homologous end-joining. Cell research 18(1):114-124. 33. Jackson PS (2002). Sensing and repairing DNA double-strand breaks. Carcinogenesis 23(5):687-696. 34. Novakova-Jiresova A, Van Gameren MM, Coppes RP, Kampinga HH, Groen HJ (2004). Transforming growth
factor-β plasma dynamics and post-irradiation lung injury in lung cancer patients. Radiotherapy and oncology 71(2):183-189.
35. Bentzen SM (2006). Preventing or reducing late side effects of radiation therapy: radiobiology meets molecular pathology. Nature reviews. Cancer 6(9):702-713.
36. National center for biotechnology information (http://www.ncbi.nlm.nih.gov). 37. Savas S, Geraci J, Jurisica I, Liu G (2009). A comprehensive catalogue of functional genetic variations in the
EGFR pathway: protein-protein interaction analysis reveals novel genes and polymorphisms important for cancer research. International journal of cancer 125(6):1257-1265.
38. Grainger DJ, Heathcote K, Chiano M, Snieder H, Kemp PR, Metcalfe JC, Carter ND, Spector TD (1999). Genetic control of the circulating concentration of transforming growth factor typ β1. Human molecular genetics 8(1):93-97.
39. Quarmby S, Fakhoury H, Levine E, Barber J, Wylie J, Hajeer AH, West C, Stewart A, Magee B, Kumar S (2003). Association of transforming growth factor beta-1 single nucleotide polymorphisms with radiation-induced damage to normal tissues in breast cancer patients. International journal of radiation biology 79(2):137-143.
40. Andreassen CN, Alsner J, Overgaard M, Overgaard J (2003). Prediction of normal tissue radiosensitivity from polymorphisms in candidate genes. Radiotherapy and oncology 69(2):127-135.
41. De Ruyck K, Van Eijkeren M, Claes K, Bacher K, Vral A, De Neve W, Thierens H (2006). TGFβ1 polymorphisms and late clinical radiosensitivity in patients treated for gynecologic tumors. Radiation oncology, biology, physics 65(4):1240-1248.
42. Yuan X, Liao Z, Liu Z, Wang LE, Tucker SL, Mao L, Wang XS, Martel M, Komaki R, Cox JD, Milas L, Wei Q (2009). Single nucleotide polymorphism at rs1982073: T869C of the TGFβ1 gene is associated with the risk of radiation pneumonitis in patients with definitive radiotherapy. Journal of clinical oncology 27(20):3370-3378.
43. Andreassen CN, Alsner J, Overgaard M, Sørensen FB, Overgaard J (2006). Risk of radiation-induced subcutaneaous fibrosis in relation to single nucleotide polymorphisms in TGFB1, SOD2, XRCC1, XRCC3, APEX and ATM – a study based on DNA from formalin fixed paraffin embedded tissue samples. International journal of radiation biology 82(8):577-586.
44. Andreassen CN, Grou C, Alsner J, Overgaard J (2007). Are TGF-beta 1 polymorphisms potential predictors of fibrosis risk after radiotherapy? – a subset analysis from the DAHANCA 6 and 7 protocols. European journal of cancer supplements 5(4):119.
45. Awad MR, El-Gamel A, Hasleton P, Turner DM, Sinnott PJ, Hutchinson IV (1998). Genotypic variation in the transforming growth factor-[beta]1 gene: association with transforming growth factor-[beta]1 production, fibrotic lung disease and graft fibrosis after lung transplantation. Transplantation 66(8):1014-1020.
46. Omori S, Takiguchi Y, Suda A, Sugimoto T, Miyazawa H, Takiguchi Y, Tanabe N, Tatsumi K, Kimura H, Pardington PE, Chen F, Chen DJ, Kuriyama T (2002). Suppression of a DNA double-strand break repair gen, Ku70, increases radio-and chemosensitivity in a human lung carcinoma cell line. DNA repair 1(4):299-310.

Referentielijst
50
47. Gu Y, Jin S, Gao Y, Weaver DT, Alt FW (1997). Ku70-deficient embryonic stem cells have increased ionizing radiosensitivity , defective DNA end-binding activity, and inability to support V(D)J recombination. Proceedings of the national academy of sciences of the United States of America 94(15):8076-8081.
48. Suga T, Iwakawa M, Tsuji H, Ishikawa H, Oda E, Noda S, Otsuka Y, Ishikawa A, Ishikawa K, Shimazaki J, Mizoe JE, Tsujii H, Imai T (2008). Influence of multiple genetic polymorphisms on genitourinary morbidity after carbon ion radiotherapy for prostate cancer. International journal of radiation oncology, biology, physics 72(3):808-813.
49. Kurumizaka H, Enomoto R, Nakada M, Eda K, Yokoyama S, Shibata T(2003). Region and amino acid residues required for Rad51C binding in the human Xrcc3 protein. Nucleic acids research 31(14):4041-4050.
50. Popanda O, Marquardt JU, Chang-Claude J, Schmezer P (2009). Genetic variation in normal tissue toxicity induced by ionizing radiation. Mutation research 667(1-2):58-69.
51. Burri RJ, Stock RG, Cesaretti JA, Atencio DP, Peters S, Peters CA, Fan G, Stone NN, Ostrer H, Rosenstein BS (2008). Association of single nucleotide polymorphisms in SOD2, XRCC1 and XRCC3 with susceptibility for the development of adverse effects resulting from radiotherapy for prostate cancer. Radiation research 170(1):49-59.
52. Alsbeih G, Al-Harbi N, Al-Hadyan K, El-Sebaie M, Al-Rajhi N (2010). Association between normal tissue complications after radiotherapy and polymorphic variations in TGFB1 and XRCC1 genes. Radiation research 173(4):505-511.
53. Duprez F, Bonte K, De Neve W, Boterberg T, De Gersem W, Madani I (2010). Regional relapse after intensity-modulated radiotherapy for head-and-neck cancer. International journal of radiation oncology, biology, physics in druk..
54. Singleton P (2000). DNA methods in clinical microbiology. The Netherlands: Kluwer academic publishers. 55. Chakrabarti R, Schutt CE (2001). The enhancement of PCR amplification by low molecular-weight sulfones.
Gene 274(1-2):293-298. 56. RestrictionMapper version 3 (http://restrictionmapper.org). 57. A guide to high resolution melting (HRM) analysis (https://products.appliedbiosystems.com). 58. Liew M, Pryor R, Palais R, Meadows C, Erali M, Lyon E, Wittwer C (2004). Genotyping of single-nucleotide
polymorphisms by high-resolution melting of small amplicons. Clinical chemistry 50(7):1156-1164. 59. Montgomery J, Wittwer CT, Palais R, Zhou L (2007). Simultaneous mutation scanning and genotyping by high-
resolution DNA melting analysis. Nature protocols 2(1):59-66. 60. Ensembl genome browser (http://www.ensembl.org/index.html) 61. Oligonucleotide properties calculator (http://www.basic.northwestern.edu/biotools/oligocalc.html). 62. Primer3Plus (http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi). 63. UCSC genome bioinformatics (http://genome.ucsc.edu). 64. Araujo FD, Pierce AJ, Stark JM, Jasin M (2002). Variant XRCC3 implicated in cancer is functional in
homology-directed repair of double-strand breaks. Oncogene 21(26):4176-4180.
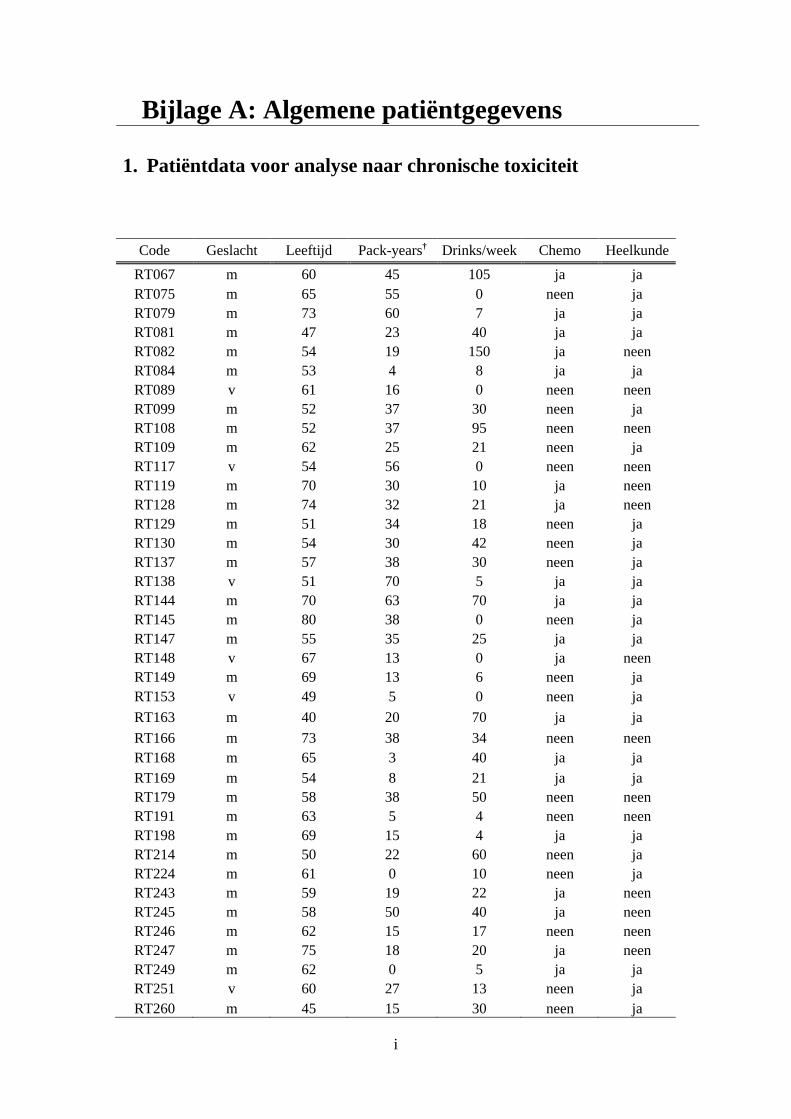
i
Bijlage A: Algemene patiëntgegevens
1. Patiëntdata voor analyse naar chronische toxiciteit
Code Geslacht Leeftijd Pack-years† Drinks/week Chemo Heelkunde
RT067 m 60 45 105 ja ja RT075 m 65 55 0 neen ja RT079 m 73 60 7 ja ja RT081 m 47 23 40 ja ja RT082 m 54 19 150 ja neen RT084 m 53 4 8 ja ja RT089 v 61 16 0 neen neen RT099 m 52 37 30 neen ja RT108 m 52 37 95 neen neen RT109 m 62 25 21 neen ja RT117 v 54 56 0 neen neen RT119 m 70 30 10 ja neen RT128 m 74 32 21 ja neen RT129 m 51 34 18 neen ja RT130 m 54 30 42 neen ja RT137 m 57 38 30 neen ja RT138 v 51 70 5 ja ja RT144 m 70 63 70 ja ja RT145 m 80 38 0 neen ja RT147 m 55 35 25 ja ja RT148 v 67 13 0 ja neen RT149 m 69 13 6 neen ja RT153 v 49 5 0 neen ja RT163 m 40 20 70 ja ja RT166 m 73 38 34 neen neen RT168 m 65 3 40 ja ja RT169 m 54 8 21 ja ja RT179 m 58 38 50 neen neen RT191 m 63 5 4 neen neen RT198 m 69 15 4 ja ja RT214 m 50 22 60 neen ja RT224 m 61 0 10 neen ja RT243 m 59 19 22 ja neen RT245 m 58 50 40 ja neen RT246 m 62 15 17 neen neen RT247 m 75 18 20 ja neen RT249 m 62 0 5 ja ja RT251 v 60 27 13 neen ja RT260 m 45 15 30 neen ja
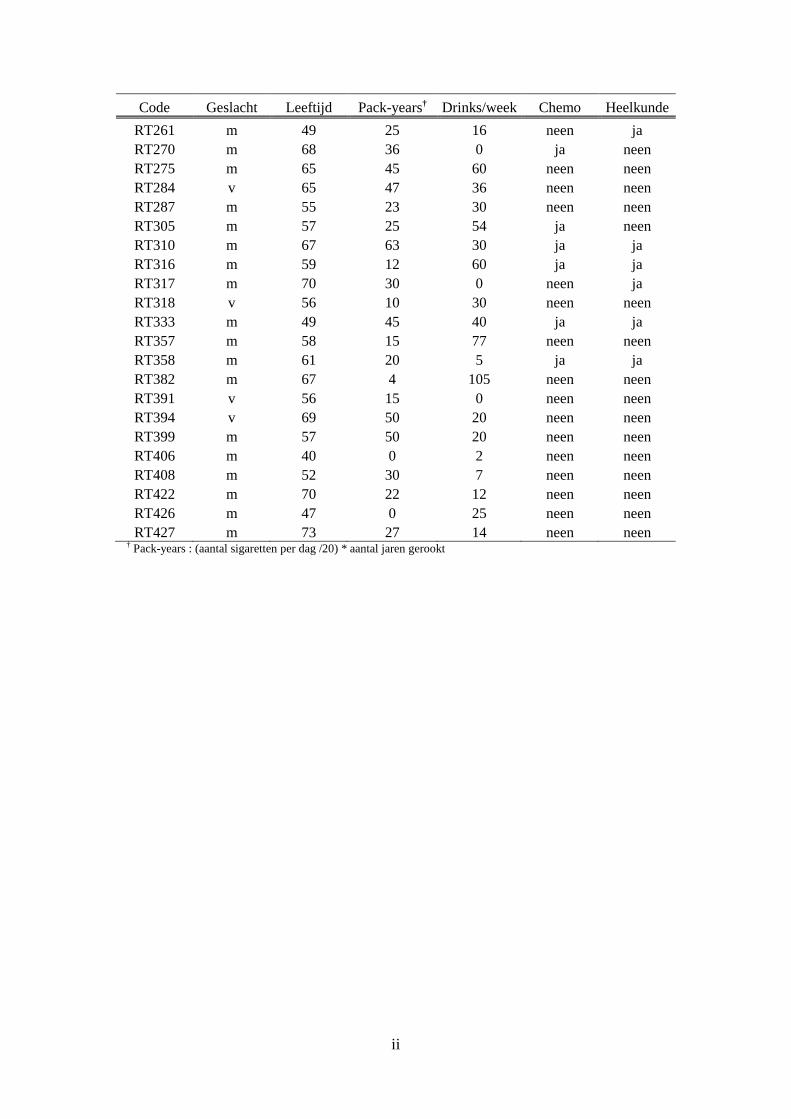
ii
Code Geslacht Leeftijd Pack-years† Drinks/week Chemo Heelkunde
RT261 m 49 25 16 neen ja RT270 m 68 36 0 ja neen RT275 m 65 45 60 neen neen RT284 v 65 47 36 neen neen RT287 m 55 23 30 neen neen RT305 m 57 25 54 ja neen RT310 m 67 63 30 ja ja RT316 m 59 12 60 ja ja RT317 m 70 30 0 neen ja RT318 v 56 10 30 neen neen RT333 m 49 45 40 ja ja RT357 m 58 15 77 neen neen RT358 m 61 20 5 ja ja RT382 m 67 4 105 neen neen RT391 v 56 15 0 neen neen RT394 v 69 50 20 neen neen RT399 m 57 50 20 neen neen RT406 m 40 0 2 neen neen RT408 m 52 30 7 neen neen RT422 m 70 22 12 neen neen RT426 m 47 0 25 neen neen RT427 m 73 27 14 neen neen
† Pack-years : (aantal sigaretten per dag /20) * aantal jaren gerookt
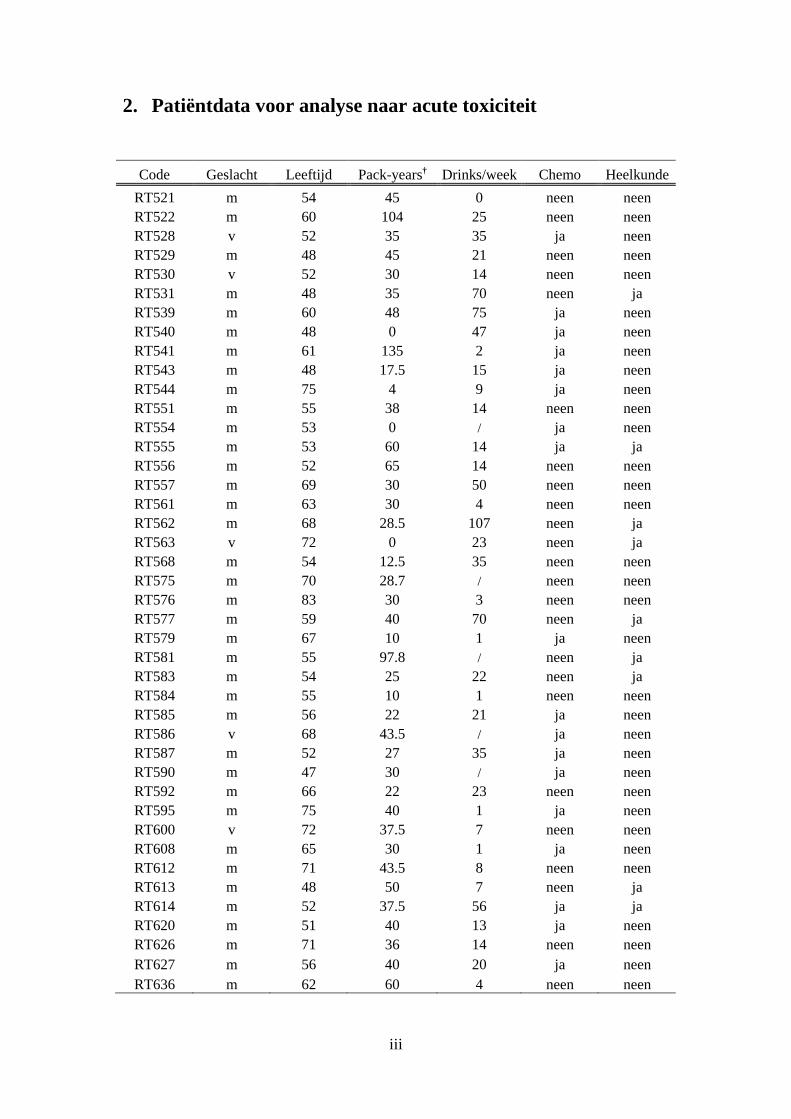
iii
2. Patiëntdata voor analyse naar acute toxiciteit
Code Geslacht Leeftijd Pack-years† Drinks/week Chemo Heelkunde
RT521 m 54 45 0 neen neen RT522 m 60 104 25 neen neen RT528 v 52 35 35 ja neen RT529 m 48 45 21 neen neen RT530 v 52 30 14 neen neen RT531 m 48 35 70 neen ja RT539 m 60 48 75 ja neen RT540 m 48 0 47 ja neen RT541 m 61 135 2 ja neen RT543 m 48 17.5 15 ja neen RT544 m 75 4 9 ja neen RT551 m 55 38 14 neen neen RT554 m 53 0 / ja neen RT555 m 53 60 14 ja ja RT556 m 52 65 14 neen neen RT557 m 69 30 50 neen neen RT561 m 63 30 4 neen neen RT562 m 68 28.5 107 neen ja RT563 v 72 0 23 neen ja RT568 m 54 12.5 35 neen neen RT575 m 70 28.7 / neen neen RT576 m 83 30 3 neen neen RT577 m 59 40 70 neen ja RT579 m 67 10 1 ja neen RT581 m 55 97.8 / neen ja RT583 m 54 25 22 neen ja RT584 m 55 10 1 neen neen RT585 m 56 22 21 ja neen RT586 v 68 43.5 / ja neen RT587 m 52 27 35 ja neen RT590 m 47 30 / ja neen RT592 m 66 22 23 neen neen RT595 m 75 40 1 ja neen RT600 v 72 37.5 7 neen neen RT608 m 65 30 1 ja neen RT612 m 71 43.5 8 neen neen RT613 m 48 50 7 neen ja RT614 m 52 37.5 56 ja ja RT620 m 51 40 13 ja neen RT626 m 71 36 14 neen neen RT627 m 56 40 20 ja neen RT636 m 62 60 4 neen neen
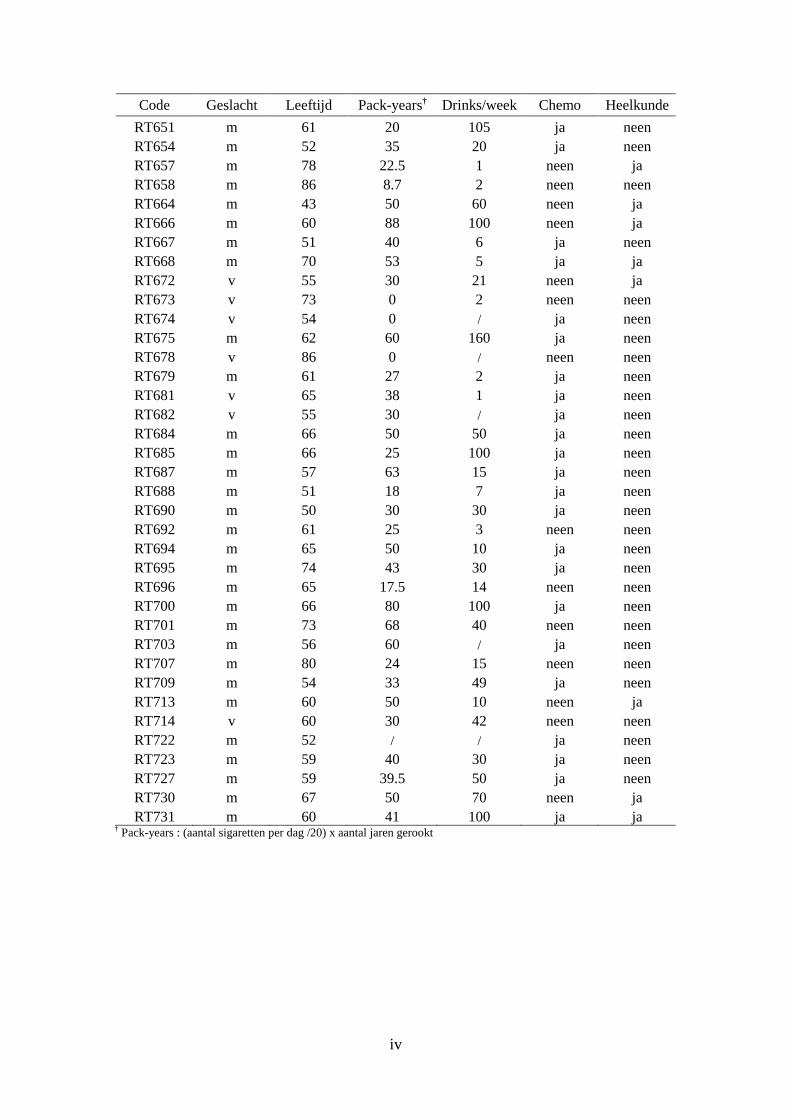
iv
Code Geslacht Leeftijd Pack-years† Drinks/week Chemo Heelkunde RT651 m 61 20 105 ja neen RT654 m 52 35 20 ja neen RT657 m 78 22.5 1 neen ja RT658 m 86 8.7 2 neen neen RT664 m 43 50 60 neen ja RT666 m 60 88 100 neen ja RT667 m 51 40 6 ja neen RT668 m 70 53 5 ja ja RT672 v 55 30 21 neen ja RT673 v 73 0 2 neen neen RT674 v 54 0 / ja neen RT675 m 62 60 160 ja neen RT678 v 86 0 / neen neen RT679 m 61 27 2 ja neen RT681 v 65 38 1 ja neen RT682 v 55 30 / ja neen RT684 m 66 50 50 ja neen RT685 m 66 25 100 ja neen RT687 m 57 63 15 ja neen RT688 m 51 18 7 ja neen RT690 m 50 30 30 ja neen RT692 m 61 25 3 neen neen RT694 m 65 50 10 ja neen RT695 m 74 43 30 ja neen RT696 m 65 17.5 14 neen neen RT700 m 66 80 100 ja neen RT701 m 73 68 40 neen neen RT703 m 56 60 / ja neen RT707 m 80 24 15 neen neen RT709 m 54 33 49 ja neen RT713 m 60 50 10 neen ja RT714 v 60 30 42 neen neen RT722 m 52 / / ja neen RT723 m 59 40 30 ja neen RT727 m 59 39.5 50 ja neen RT730 m 67 50 70 neen ja RT731 m 60 41 100 ja ja
† Pack-years : (aantal sigaretten per dag /20) x aantal jaren gerookt
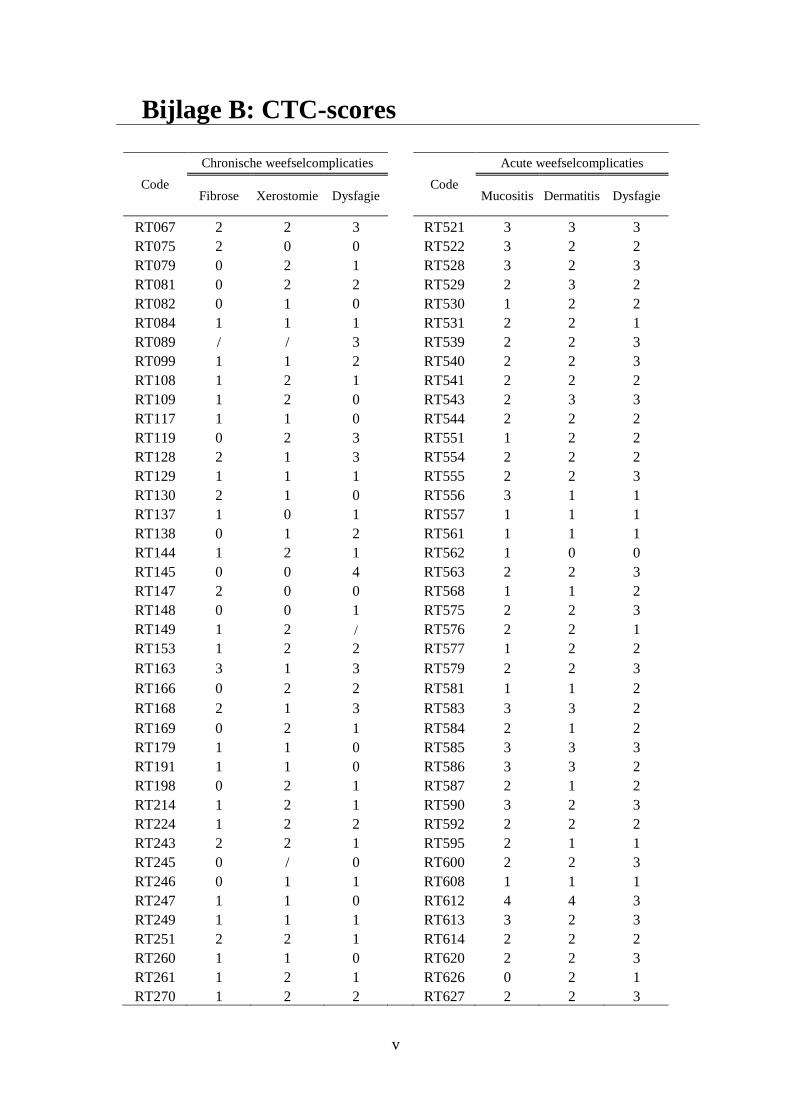
v
Bijlage B: CTC-scores
Code
Chronische weefselcomplicaties
Code
Acute weefselcomplicaties
Fibrose Xerostomie Dysfagie Mucositis Dermatitis Dysfagie
RT067 2 2 3 RT521 3 3 3 RT075 2 0 0 RT522 3 2 2 RT079 0 2 1 RT528 3 2 3 RT081 0 2 2 RT529 2 3 2 RT082 0 1 0 RT530 1 2 2 RT084 1 1 1 RT531 2 2 1 RT089 / / 3 RT539 2 2 3 RT099 1 1 2 RT540 2 2 3 RT108 1 2 1 RT541 2 2 2 RT109 1 2 0 RT543 2 3 3 RT117 1 1 0 RT544 2 2 2 RT119 0 2 3 RT551 1 2 2 RT128 2 1 3 RT554 2 2 2 RT129 1 1 1 RT555 2 2 3 RT130 2 1 0 RT556 3 1 1 RT137 1 0 1 RT557 1 1 1 RT138 0 1 2 RT561 1 1 1 RT144 1 2 1 RT562 1 0 0 RT145 0 0 4 RT563 2 2 3 RT147 2 0 0 RT568 1 1 2 RT148 0 0 1 RT575 2 2 3 RT149 1 2 / RT576 2 2 1 RT153 1 2 2 RT577 1 2 2 RT163 3 1 3 RT579 2 2 3 RT166 0 2 2 RT581 1 1 2 RT168 2 1 3 RT583 3 3 2 RT169 0 2 1 RT584 2 1 2 RT179 1 1 0 RT585 3 3 3 RT191 1 1 0 RT586 3 3 2 RT198 0 2 1 RT587 2 1 2 RT214 1 2 1 RT590 3 2 3 RT224 1 2 2 RT592 2 2 2 RT243 2 2 1 RT595 2 1 1 RT245 0 / 0 RT600 2 2 3 RT246 0 1 1 RT608 1 1 1 RT247 1 1 0 RT612 4 4 3 RT249 1 1 1 RT613 3 2 3 RT251 2 2 1 RT614 2 2 2 RT260 1 1 0 RT620 2 2 3 RT261 1 2 1 RT626 0 2 1 RT270 1 2 2 RT627 2 2 3
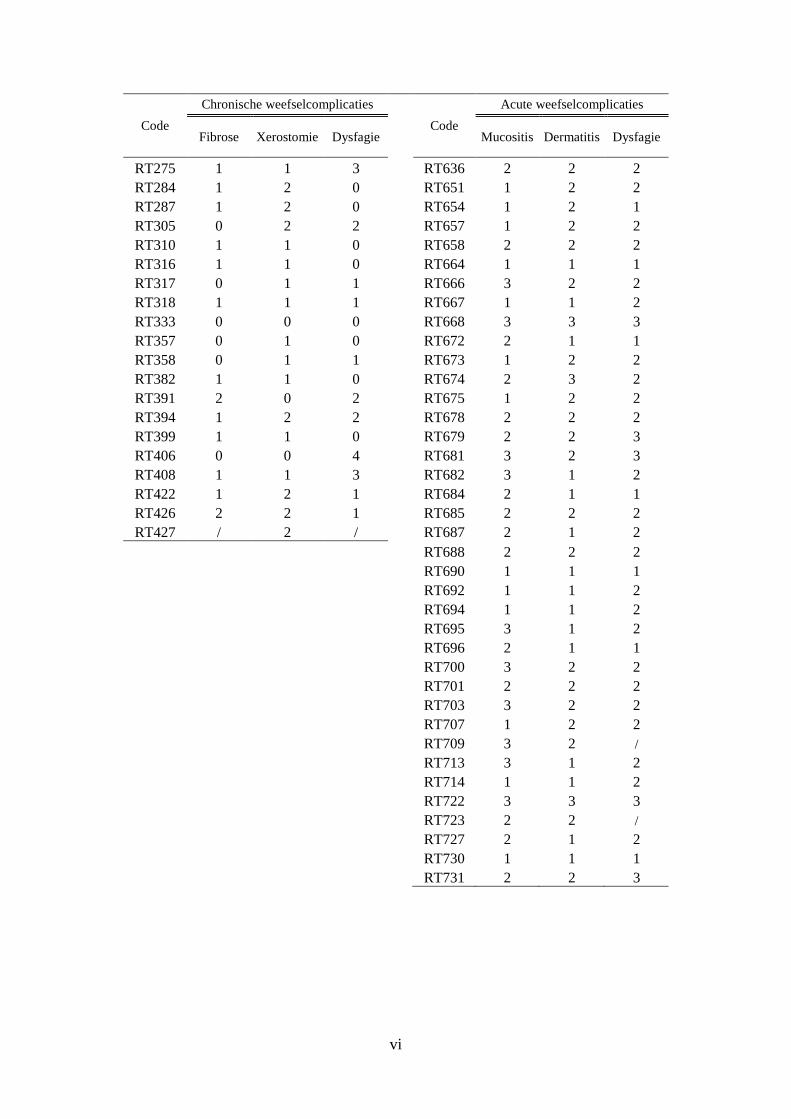
vi
Code
Chronische weefselcomplicaties
Code
Acute weefselcomplicaties
Fibrose Xerostomie Dysfagie Mucositis Dermatitis Dysfagie
RT275 1 1 3 RT636 2 2 2 RT284 1 2 0 RT651 1 2 2 RT287 1 2 0 RT654 1 2 1 RT305 0 2 2 RT657 1 2 2 RT310 1 1 0 RT658 2 2 2 RT316 1 1 0 RT664 1 1 1 RT317 0 1 1 RT666 3 2 2 RT318 1 1 1 RT667 1 1 2 RT333 0 0 0 RT668 3 3 3 RT357 0 1 0 RT672 2 1 1 RT358 0 1 1 RT673 1 2 2 RT382 1 1 0 RT674 2 3 2 RT391 2 0 2 RT675 1 2 2 RT394 1 2 2 RT678 2 2 2 RT399 1 1 0 RT679 2 2 3 RT406 0 0 4 RT681 3 2 3 RT408 1 1 3 RT682 3 1 2 RT422 1 2 1 RT684 2 1 1 RT426 2 2 1 RT685 2 2 2 RT427 / 2 / RT687 2 1 2
RT688 2 2 2 RT690 1 1 1 RT692 1 1 2 RT694 1 1 2 RT695 3 1 2 RT696 2 1 1 RT700 3 2 2 RT701 2 2 2 RT703 3 2 2 RT707 1 2 2 RT709 3 2 /
RT713 3 1 2 RT714 1 1 2 RT722 3 3 3 RT723 2 2 /
RT727 2 1 2 RT730 1 1 1 RT731 2 2 3
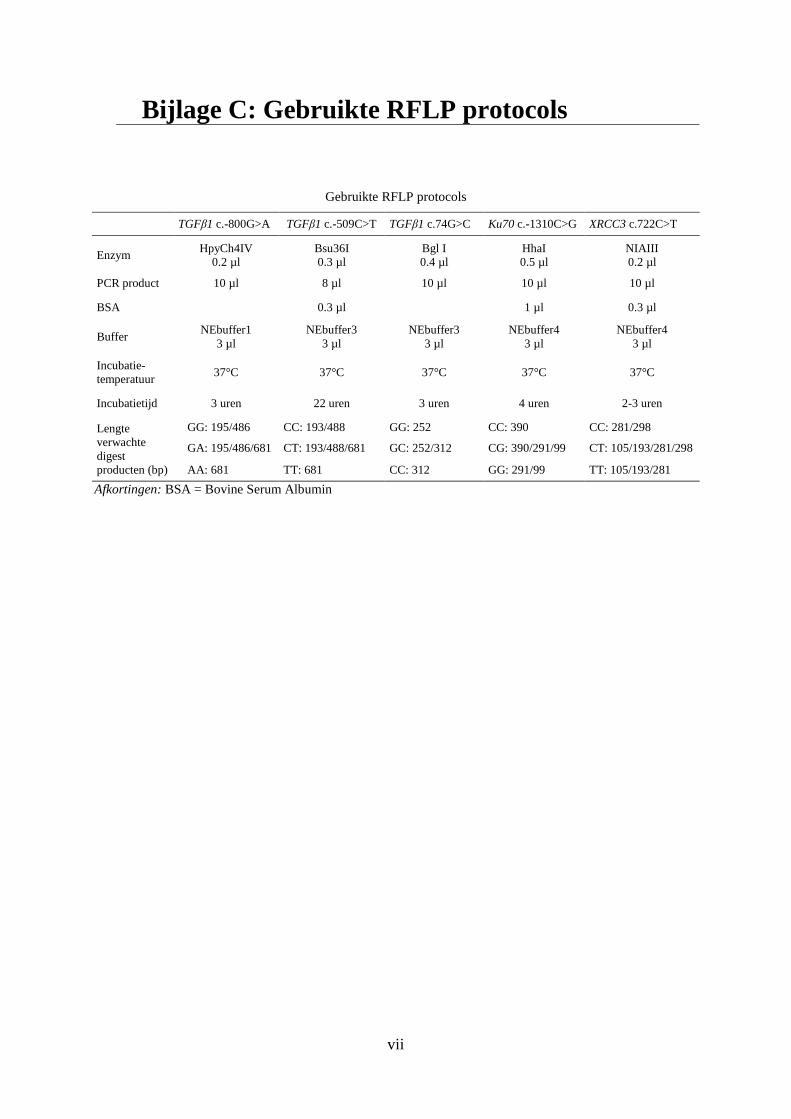
vii
Bijlage C: Gebruikte RFLP protocols
Gebruikte RFLP protocols
TGFβ1 c.-800G>A TGFβ1 c.-509C>T TGFβ1 c.74G>C Ku70 c.-1310C>G XRCC3 c.722C>T
Enzym
HpyCh4IV 0.2 µl
Bsu36I 0.3 µl
Bgl I 0.4 µl
HhaI 0.5 µl
NIAIII 0.2 µl
PCR product 10 µl 8 µl 10 µl 10 µl 10 µl
BSA
0.3 µl
1 µl 0.3 µl
Buffer
NEbuffer1 3 µl
NEbuffer3 3 µl
NEbuffer3 3 µl
NEbuffer4 3 µl
NEbuffer4 3 µl
Incubatie- temperatuur
37°C 37°C 37°C 37°C 37°C
Incubatietijd 3 uren 22 uren 3 uren 4 uren 2-3 uren
Lengte verwachte digest producten (bp)
GG: 195/486 CC: 193/488 GG: 252 CC: 390 CC: 281/298
GA: 195/486/681 CT: 193/488/681 GC: 252/312 CG: 390/291/99 CT: 105/193/281/298
AA: 681 TT: 681 CC: 312 GG: 291/99 TT: 105/193/281
Afkortingen: BSA = Bovine Serum Albumin
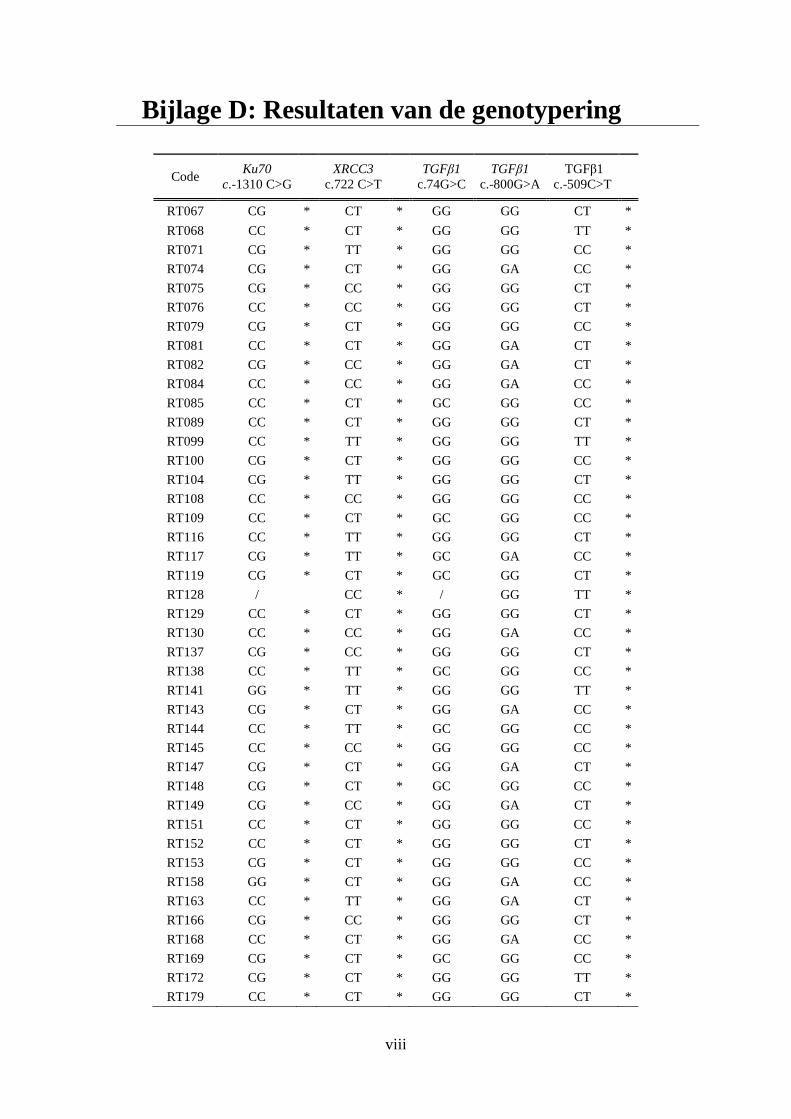
viii
Bijlage D: Resultaten van de genotypering
Code Ku70
c.-1310 C>G
XRCC3 c.722 C>T
TGFβ1
c.74G>C TGFβ1
c.-800G>A TGFβ1
c.-509C>T
RT067 CG * CT * GG GG CT *
RT068 CC * CT * GG GG TT *
RT071 CG * TT * GG GG CC *
RT074 CG * CT * GG GA CC *
RT075 CG * CC * GG GG CT *
RT076 CC * CC * GG GG CT *
RT079 CG * CT * GG GG CC *
RT081 CC * CT * GG GA CT *
RT082 CG * CC * GG GA CT *
RT084 CC * CC * GG GA CC *
RT085 CC * CT * GC GG CC *
RT089 CC * CT * GG GG CT *
RT099 CC * TT * GG GG TT *
RT100 CG * CT * GG GG CC *
RT104 CG * TT * GG GG CT *
RT108 CC * CC * GG GG CC *
RT109 CC * CT * GC GG CC *
RT116 CC * TT * GG GG CT *
RT117 CG * TT * GC GA CC *
RT119 CG * CT * GC GG CT *
RT128 /
CC * / GG TT *
RT129 CC * CT * GG GG CT *
RT130 CC * CC * GG GA CC *
RT137 CG * CC * GG GG CT *
RT138 CC * TT * GC GG CC *
RT141 GG * TT * GG GG TT *
RT143 CG * CT * GG GA CC *
RT144 CC * TT * GC GG CC *
RT145 CC * CC * GG GG CC *
RT147 CG * CT * GG GA CT *
RT148 CG * CT * GC GG CC *
RT149 CG * CC * GG GA CT *
RT151 CC * CT * GG GG CC *
RT152 CC * CT * GG GG CT *
RT153 CG * CT * GG GG CC *
RT158 GG * CT * GG GA CC *
RT163 CC * TT * GG GA CT *
RT166 CG * CC * GG GG CT *
RT168 CC * CT * GG GA CC *
RT169 CG * CT * GC GG CC *
RT172 CG * CT * GG GG TT *
RT179 CC * CT * GG GG CT *
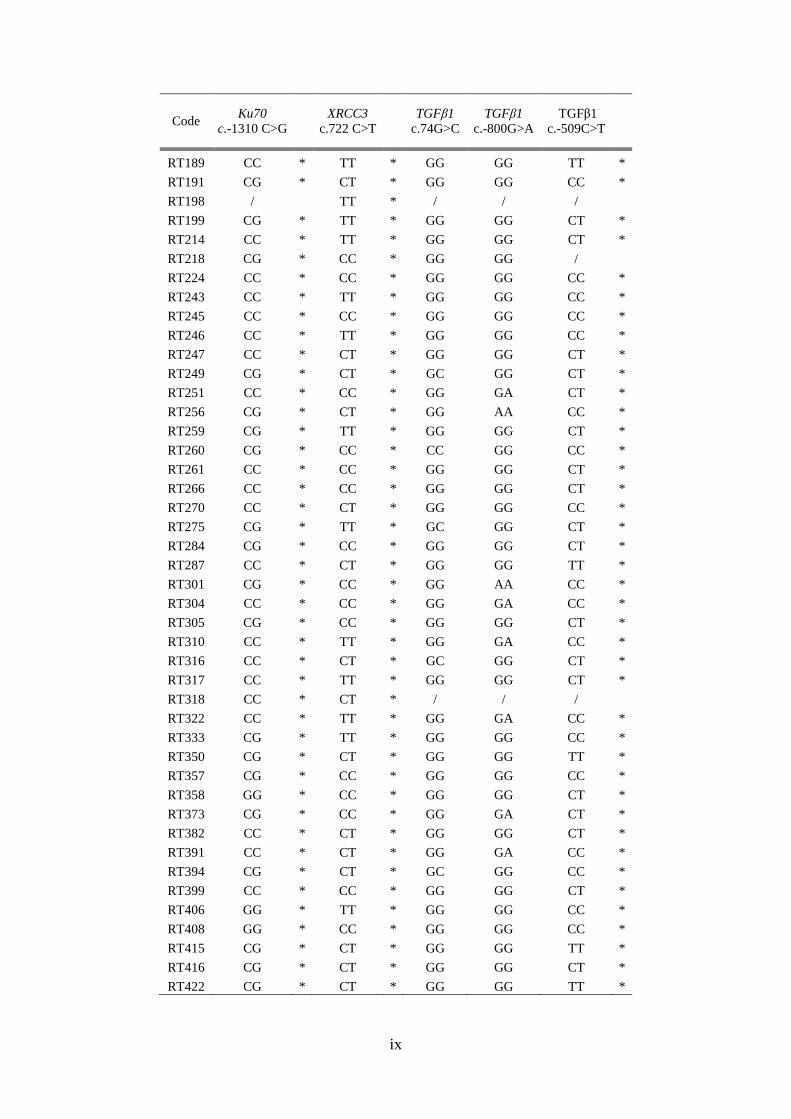
ix
Code Ku70
c.-1310 C>G
XRCC3 c.722 C>T
TGFβ1
c.74G>C TGFβ1
c.-800G>A TGFβ1
c.-509C>T
RT189 CC * TT * GG GG TT *
RT191 CG * CT * GG GG CC *
RT198 / TT * / / /
RT199 CG * TT * GG GG CT *
RT214 CC * TT * GG GG CT *
RT218 CG * CC * GG GG /
RT224 CC * CC * GG GG CC *
RT243 CC * TT * GG GG CC *
RT245 CC * CC * GG GG CC *
RT246 CC * TT * GG GG CC *
RT247 CC * CT * GG GG CT *
RT249 CG * CT * GC GG CT *
RT251 CC * CC * GG GA CT *
RT256 CG * CT * GG AA CC *
RT259 CG * TT * GG GG CT *
RT260 CG * CC * CC GG CC *
RT261 CC * CC * GG GG CT *
RT266 CC * CC * GG GG CT *
RT270 CC * CT * GG GG CC *
RT275 CG * TT * GC GG CT *
RT284 CG * CC * GG GG CT *
RT287 CC * CT * GG GG TT *
RT301 CG * CC * GG AA CC *
RT304 CC * CC * GG GA CC *
RT305 CG * CC * GG GG CT *
RT310 CC * TT * GG GA CC *
RT316 CC * CT * GC GG CT *
RT317 CC * TT * GG GG CT *
RT318 CC * CT * / / /
RT322 CC * TT * GG GA CC *
RT333 CG * TT * GG GG CC *
RT350 CG * CT * GG GG TT *
RT357 CG * CC * GG GG CC *
RT358 GG * CC * GG GG CT *
RT373 CG * CC * GG GA CT *
RT382 CC * CT * GG GG CT *
RT391 CC * CT * GG GA CC *
RT394 CG * CT * GC GG CC *
RT399 CC * CC * GG GG CT *
RT406 GG * TT * GG GG CC *
RT408 GG * CC * GG GG CC *
RT415 CG * CT * GG GG TT *
RT416 CG * CT * GG GG CT *
RT422 CG * CT * GG GG TT *
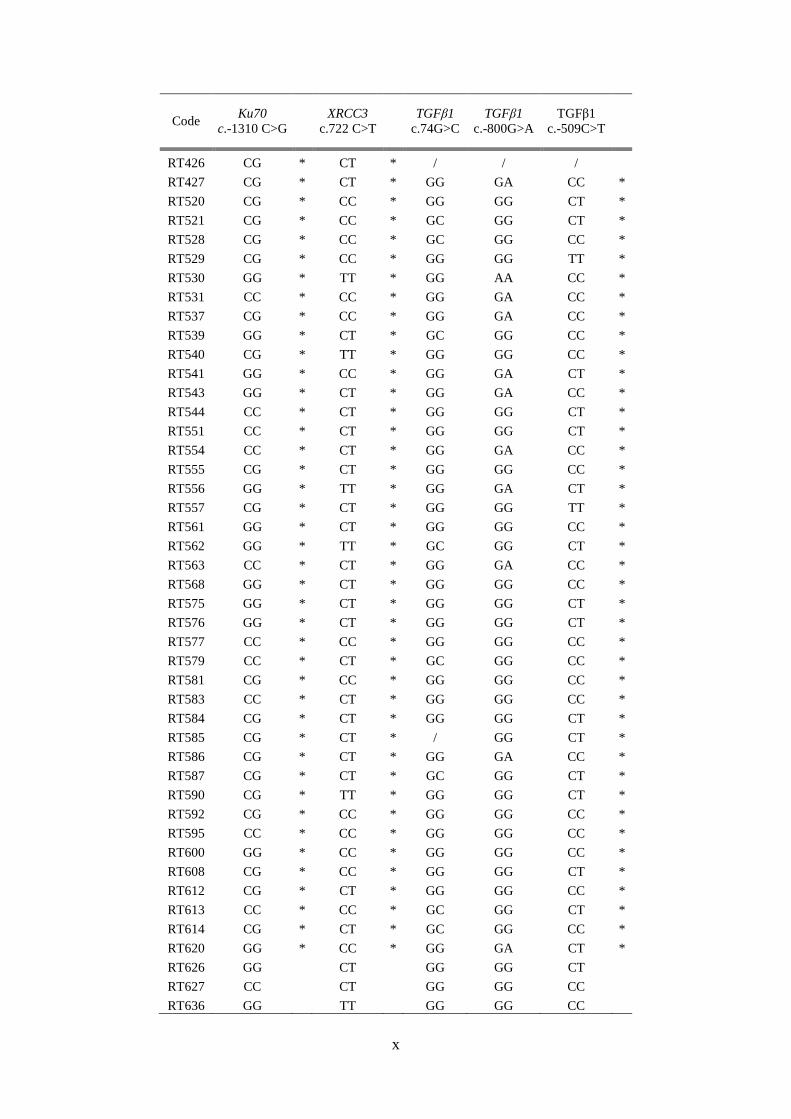
x
Code Ku70
c.-1310 C>G XRCC3
c.722 C>T TGFβ1
c.74G>C TGFβ1
c.-800G>A TGFβ1
c.-509C>T
RT426 CG * CT * / / /
RT427 CG * CT * GG GA CC *
RT520 CG * CC * GG GG CT *
RT521 CG * CC * GC GG CT *
RT528 CG * CC * GC GG CC *
RT529 CG * CC * GG GG TT *
RT530 GG * TT * GG AA CC *
RT531 CC * CC * GG GA CC *
RT537 CG * CC * GG GA CC *
RT539 GG * CT * GC GG CC *
RT540 CG * TT * GG GG CC *
RT541 GG * CC * GG GA CT *
RT543 GG * CT * GG GA CC *
RT544 CC * CT * GG GG CT *
RT551 CC * CT * GG GG CT *
RT554 CC * CT * GG GA CC *
RT555 CG * CT * GG GG CC *
RT556 GG * TT * GG GA CT *
RT557 CG * CT * GG GG TT *
RT561 GG * CT * GG GG CC *
RT562 GG * TT * GC GG CT *
RT563 CC * CT * GG GA CC *
RT568 GG * CT * GG GG CC *
RT575 GG * CT * GG GG CT *
RT576 GG * CT * GG GG CT *
RT577 CC * CC * GG GG CC *
RT579 CC * CT * GC GG CC *
RT581 CG * CC * GG GG CC *
RT583 CC * CT * GG GG CC *
RT584 CG * CT * GG GG CT *
RT585 CG * CT * / GG CT *
RT586 CG * CT * GG GA CC *
RT587 CG * CT * GC GG CT *
RT590 CG * TT * GG GG CT *
RT592 CG * CC * GG GG CC *
RT595 CC * CC * GG GG CC *
RT600 GG * CC * GG GG CC *
RT608 CG * CC * GG GG CT *
RT612 CG * CT * GG GG CC *
RT613 CC * CC * GC GG CT *
RT614 CG * CT * GC GG CC *
RT620 GG * CC * GG GA CT *
RT626 GG CT GG GG CT
RT627 CC CT GG GG CC
RT636 GG TT GG GG CC
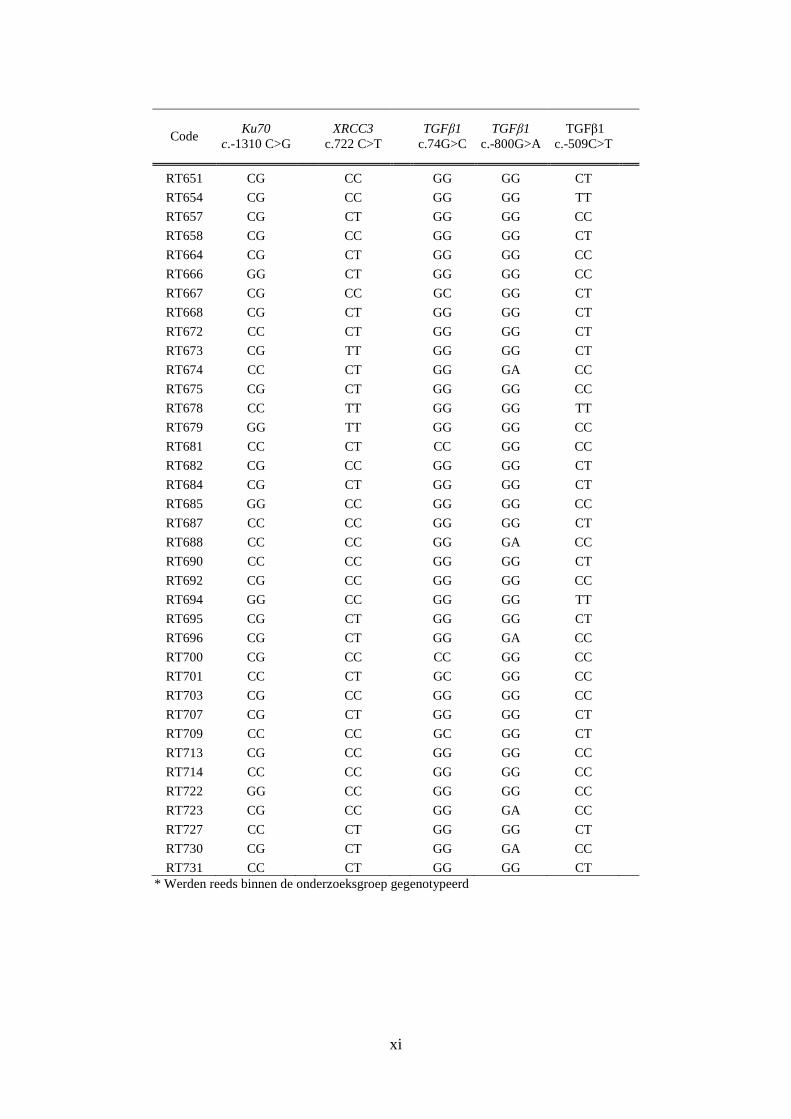
xi
Code Ku70
c.-1310 C>G
XRCC3 c.722 C>T
TGFβ1
c.74G>C TGFβ1
c.-800G>A TGFβ1
c.-509C>T
RT651 CG CC GG GG CT
RT654 CG CC GG GG TT
RT657 CG CT GG GG CC
RT658 CG CC GG GG CT
RT664 CG CT GG GG CC
RT666 GG CT GG GG CC
RT667 CG CC GC GG CT
RT668 CG CT GG GG CT
RT672 CC CT GG GG CT
RT673 CG TT GG GG CT
RT674 CC CT GG GA CC
RT675 CG CT GG GG CC
RT678 CC TT GG GG TT
RT679 GG TT GG GG CC
RT681 CC CT CC GG CC
RT682 CG CC GG GG CT
RT684 CG CT GG GG CT
RT685 GG CC GG GG CC
RT687 CC CC GG GG CT
RT688 CC CC GG GA CC
RT690 CC CC GG GG CT
RT692 CG CC GG GG CC
RT694 GG CC GG GG TT
RT695 CG CT GG GG CT
RT696 CG CT GG GA CC
RT700 CG CC CC GG CC
RT701 CC CT GC GG CC
RT703 CG CC GG GG CC
RT707 CG CT GG GG CT
RT709 CC CC GC GG CT
RT713 CG CC GG GG CC
RT714 CC CC GG GG CC
RT722 GG CC GG GG CC
RT723 CG CC GG GA CC
RT727 CC CT GG GG CT
RT730 CG CT GG GA CC
RT731 CC CT GG GG CT * Werden reeds binnen de onderzoeksgroep gegenotypeerd

xii
Bijlage E: Geoptimaliseerde HRM protocols
1. XRCC3 c.722C>T
HRM Mix (20µl)
Product Hoeveelheid
(µl)
Buffer (10x) 2
MgCl2 (25mM) 1.2
dNTP Mix (10mM) 0.4
124 F primer (5µM) 1.2
122 R primer (5µM) 1.2
Meltdoctor HRM Dye (20x) 1
AmpliTaq Gold 360 DNA Polymerase (5U/µl) 0.05
dH2O 11.95
DNA (25 ng/µl) 1
HRM programma
Stadium Reactiestap Temperatuur
(°C) Tijd
Precyclische fase Enzym activatie 95 10 min
Cyclische fase (40 cycles) Denaturatie 95 15 sec
Annealing/Extentie 60 1 min
Smeltcurve/ dissociatie Denaturatie 95 10 sec
Annealing 60 1 min
HRM 95 15 sec
Annealing 60 15 sec
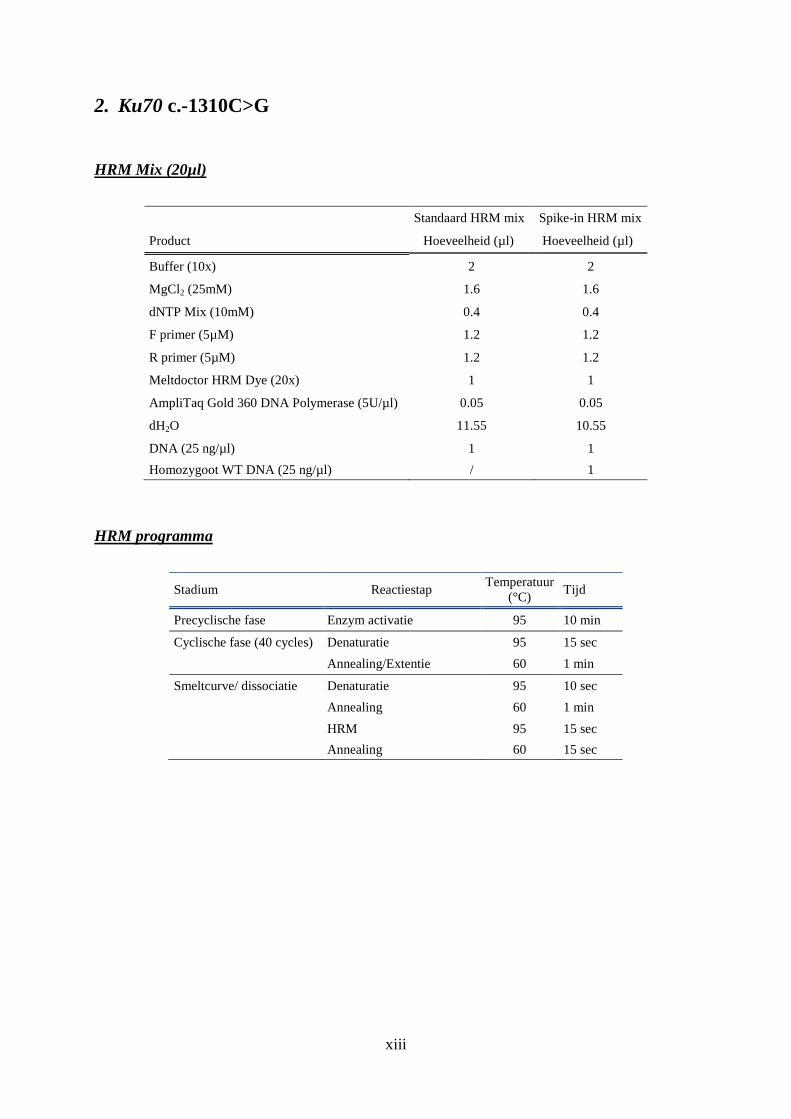
xiii
2. Ku70 c.-1310C>G
HRM Mix (20µl)
Standaard HRM mix Spike-in HRM mix
Product Hoeveelheid (µl) Hoeveelheid (µl)
Buffer (10x) 2 2
MgCl2 (25mM) 1.6 1.6
dNTP Mix (10mM) 0.4 0.4
F primer (5µM) 1.2 1.2
R primer (5µM) 1.2 1.2
Meltdoctor HRM Dye (20x) 1 1
AmpliTaq Gold 360 DNA Polymerase (5U/µl) 0.05 0.05
dH2O 11.55 10.55
DNA (25 ng/µl) 1 1
Homozygoot WT DNA (25 ng/µl) / 1
HRM programma
Stadium Reactiestap Temperatuur
(°C) Tijd
Precyclische fase Enzym activatie 95 10 min
Cyclische fase (40 cycles) Denaturatie 95 15 sec
Annealing/Extentie 60 1 min
Smeltcurve/ dissociatie Denaturatie 95 10 sec
Annealing 60 1 min
HRM 95 15 sec
Annealing 60 15 sec
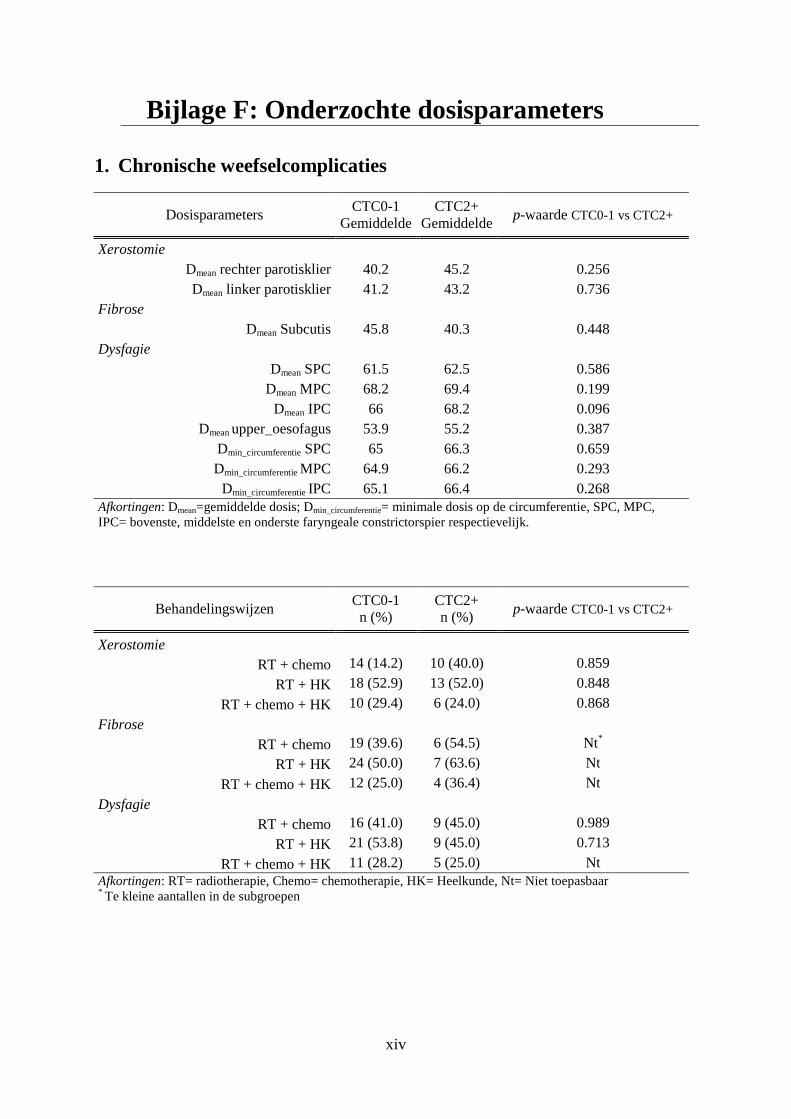
xiv
Bijlage F: Onderzochte dosisparameters
1. Chronische weefselcomplicaties
Dosisparameters CTC0-1 Gemiddelde
CTC2+ Gemiddelde p-waarde CTC0-1 vs CTC2+
Xerostomie
Dmean rechter parotisklier 40.2 45.2 0.256 Dmean linker parotisklier 41.2 43.2 0.736
Fibrose
Dmean Subcutis 45.8 40.3 0.448 Dysfagie
Dmean SPC 61.5 62.5 0.586
Dmean MPC 68.2 69.4 0.199 Dmean IPC 66 68.2 0.096
Dmean upper_oesofagus 53.9 55.2 0.387 Dmin_circumferentie SPC 65 66.3 0.659 Dmin_circumferentie MPC 64.9 66.2 0.293 Dmin_circumferentie IPC 65.1 66.4 0.268
Afkortingen: Dmean=gemiddelde dosis; Dmin_circumferentie= minimale dosis op de circumferentie, SPC, MPC, IPC= bovenste, middelste en onderste faryngeale constrictorspier respectievelijk.
Behandelingswijzen CTC0-1 n (%)
CTC2+ n (%) p-waarde CTC0-1 vs CTC2+
Xerostomie RT + chemo 14 (14.2) 10 (40.0) 0.859
RT + HK 18 (52.9) 13 (52.0) 0.848 RT + chemo + HK 10 (29.4) 6 (24.0) 0.868
Fibrose RT + chemo 19 (39.6) 6 (54.5) Nt*
RT + HK 24 (50.0) 7 (63.6) Nt RT + chemo + HK 12 (25.0) 4 (36.4) Nt
Dysfagie RT + chemo 16 (41.0) 9 (45.0) 0.989
RT + HK 21 (53.8) 9 (45.0) 0.713 RT + chemo + HK 11 (28.2) 5 (25.0) Nt
Afkortingen: RT= radiotherapie, Chemo= chemotherapie, HK= Heelkunde, Nt= Niet toepasbaar * Te kleine aantallen in de subgroepen
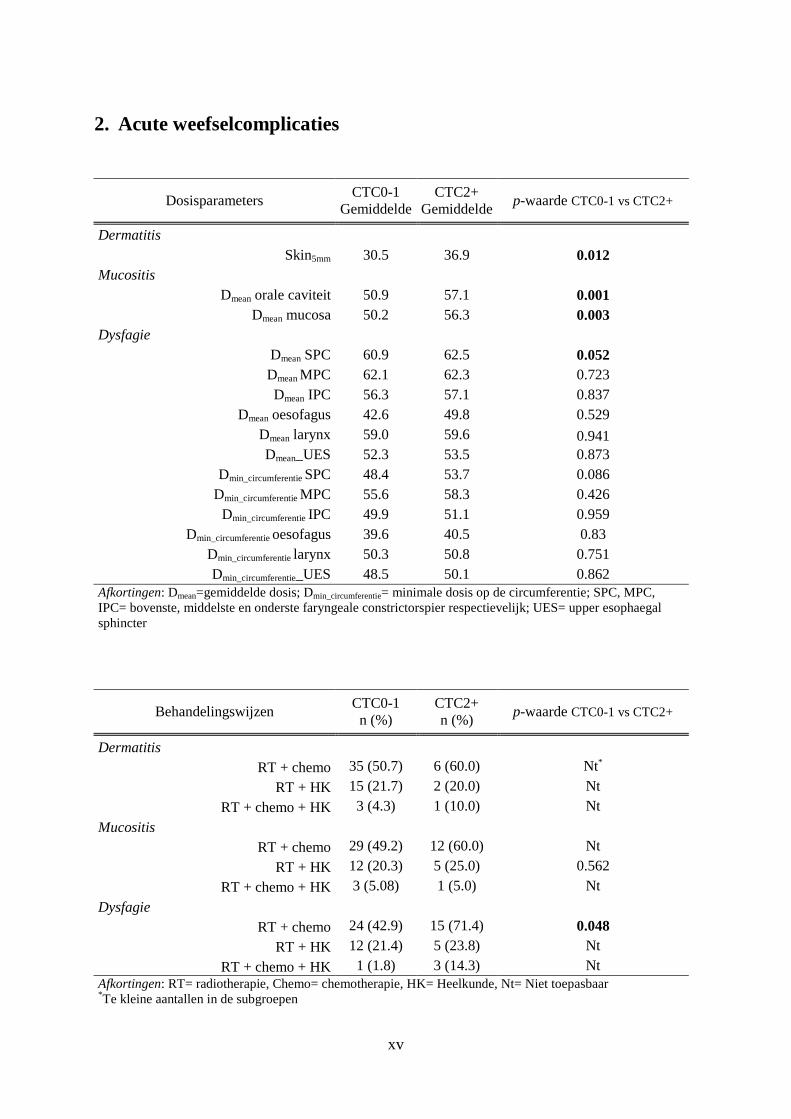
xv
2. Acute weefselcomplicaties
Dosisparameters CTC0-1 Gemiddelde
CTC2+ Gemiddelde p-waarde CTC0-1 vs CTC2+
Dermatitis
Skin5mm 30.5 36.9 0.012 Mucositis
Dmean orale caviteit 50.9 57.1 0.001
Dmean mucosa 50.2 56.3 0.003 Dysfagie
Dmean SPC 60.9 62.5 0.052 Dmean MPC 62.1 62.3 0.723 Dmean IPC 56.3 57.1 0.837
Dmean oesofagus 42.6 49.8 0.529 Dmean larynx 59.0 59.6 0.941 Dmean_UES 52.3 53.5 0.873
Dmin_circumferentie SPC 48.4 53.7 0.086 Dmin_circumferentie MPC 55.6 58.3 0.426 Dmin_circumferentie IPC 49.9 51.1 0.959
Dmin_circumferentie oesofagus 39.6 40.5 0.83 Dmin_circumferentie larynx 50.3 50.8 0.751 Dmin_circumferentie_UES 48.5 50.1 0.862
Afkortingen: Dmean=gemiddelde dosis; Dmin_circumferentie= minimale dosis op de circumferentie; SPC, MPC, IPC= bovenste, middelste en onderste faryngeale constrictorspier respectievelijk; UES= upper esophaegal sphincter
Behandelingswijzen CTC0-1 n (%)
CTC2+ n (%) p-waarde CTC0-1 vs CTC2+
Dermatitis RT + chemo 35 (50.7) 6 (60.0) Nt*
RT + HK 15 (21.7) 2 (20.0) Nt RT + chemo + HK 3 (4.3) 1 (10.0) Nt
Mucositis RT + chemo 29 (49.2) 12 (60.0) Nt
RT + HK 12 (20.3) 5 (25.0) 0.562 RT + chemo + HK 3 (5.08) 1 (5.0) Nt
Dysfagie RT + chemo 24 (42.9) 15 (71.4) 0.048
RT + HK 12 (21.4) 5 (23.8) Nt RT + chemo + HK 1 (1.8) 3 (14.3) Nt
Afkortingen: RT= radiotherapie, Chemo= chemotherapie, HK= Heelkunde, Nt= Niet toepasbaar *Te kleine aantallen in de subgroepen
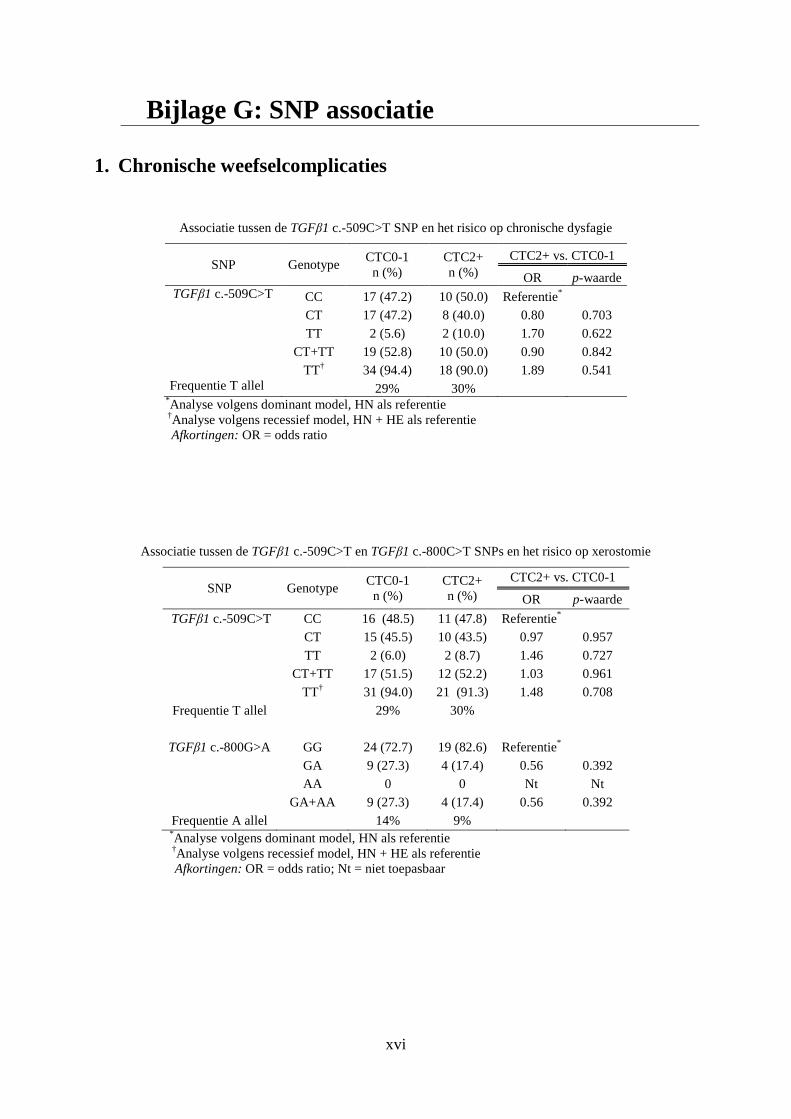
xvi
Bijlage G: SNP associatie
1. Chronische weefselcomplicaties
Associatie tussen de TGFβ1 c.-509C>T SNP en het risico op chronische dysfagie
SNP Genotype CTC0-1 n (%)
CTC2+ n (%)
CTC2+ vs. CTC0-1
OR p-waarde TGFβ1 c.-509C>T CC 17 (47.2) 10 (50.0) Referentie* CT 17 (47.2) 8 (40.0) 0.80 0.703 TT 2 (5.6) 2 (10.0) 1.70 0.622 CT+TT 19 (52.8) 10 (50.0) 0.90 0.842 TT† 34 (94.4) 18 (90.0) 1.89 0.541 Frequentie T allel 29% 30%
*Analyse volgens dominant model, HN als referentie †Analyse volgens recessief model, HN + HE als referentie
Afkortingen: OR = odds ratio
Associatie tussen de TGFβ1 c.-509C>T en TGFβ1 c.-800C>T SNPs en het risico op xerostomie
SNP Genotype CTC0-1 n (%)
CTC2+ n (%)
CTC2+ vs. CTC0-1
OR p-waarde
TGFβ1 c.-509C>T CC 16 (48.5) 11 (47.8) Referentie*
CT 15 (45.5) 10 (43.5) 0.97 0.957
TT 2 (6.0) 2 (8.7) 1.46 0.727
CT+TT 17 (51.5) 12 (52.2) 1.03 0.961
TT† 31 (94.0) 21 (91.3) 1.48 0.708
Frequentie T allel
29% 30%
TGFβ1 c.-800G>A GG 24 (72.7) 19 (82.6) Referentie*
GA 9 (27.3) 4 (17.4) 0.56 0.392
AA 0 0 Nt Nt
GA+AA 9 (27.3) 4 (17.4) 0.56 0.392 Frequentie A allel
14% 9%
*Analyse volgens dominant model, HN als referentie †Analyse volgens recessief model, HN + HE als referentie
Afkortingen: OR = odds ratio; Nt = niet toepasbaar
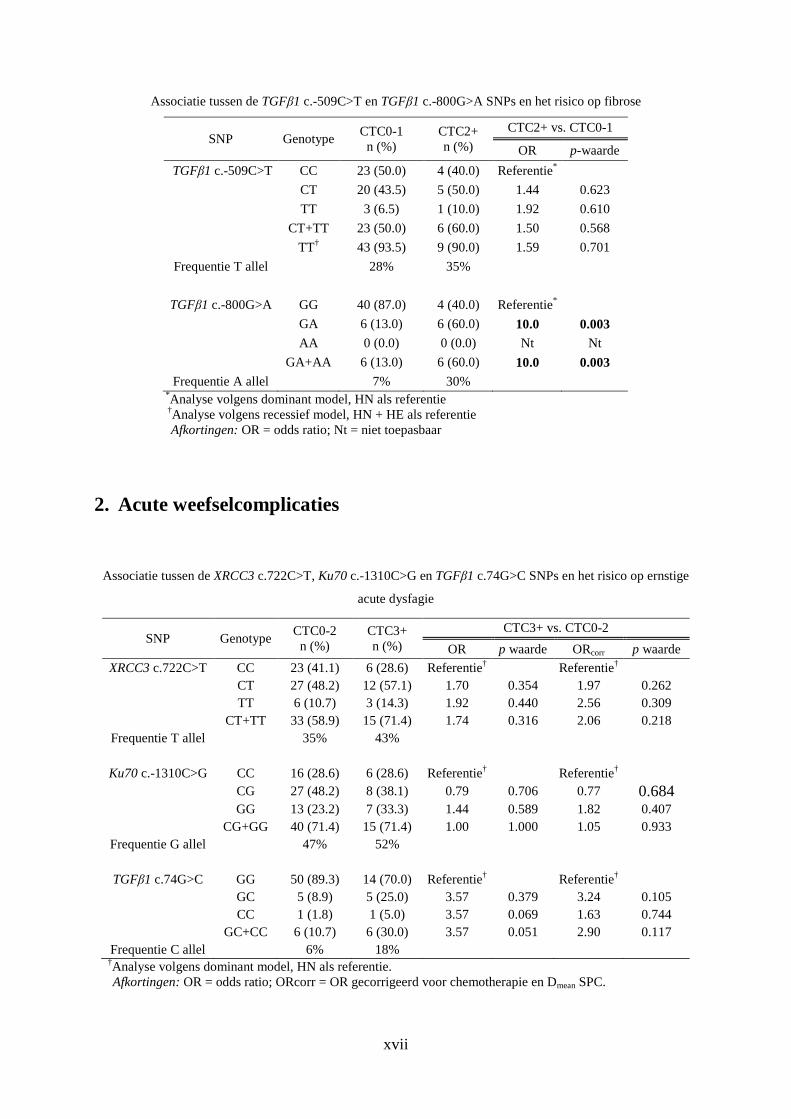
xvii
Associatie tussen de TGFβ1 c.-509C>T en TGFβ1 c.-800G>A SNPs en het risico op fibrose
SNP Genotype CTC0-1 n (%)
CTC2+ n (%)
CTC2+ vs. CTC0-1
OR p-waarde
TGFβ1 c.-509C>T CC 23 (50.0) 4 (40.0) Referentie*
CT 20 (43.5) 5 (50.0) 1.44 0.623
TT 3 (6.5) 1 (10.0) 1.92 0.610
CT+TT 23 (50.0) 6 (60.0) 1.50 0.568
TT† 43 (93.5) 9 (90.0) 1.59 0.701
Frequentie T allel
28% 35%
TGFβ1 c.-800G>A GG 40 (87.0) 4 (40.0) Referentie*
GA 6 (13.0) 6 (60.0) 10.0 0.003
AA 0 (0.0) 0 (0.0) Nt Nt
GA+AA 6 (13.0) 6 (60.0) 10.0 0.003 Frequentie A allel
7% 30%
*Analyse volgens dominant model, HN als referentie †Analyse volgens recessief model, HN + HE als referentie
Afkortingen: OR = odds ratio; Nt = niet toepasbaar
2. Acute weefselcomplicaties
Associatie tussen de XRCC3 c.722C>T, Ku70 c.-1310C>G en TGFβ1 c.74G>C SNPs en het risico op ernstige
acute dysfagie
SNP Genotype CTC0-2 n (%)
CTC3+ n (%)
CTC3+ vs. CTC0-2
OR p waarde ORcorr p waarde
XRCC3 c.722C>T CC 23 (41.1) 6 (28.6) Referentie†
Referentie†
CT 27 (48.2) 12 (57.1) 1.70 0.354 1.97 0.262
TT 6 (10.7) 3 (14.3) 1.92 0.440 2.56 0.309
CT+TT 33 (58.9) 15 (71.4) 1.74 0.316 2.06 0.218
Frequentie T allel
35% 43%
Ku70 c.-1310C>G CC 16 (28.6) 6 (28.6) Referentie†
Referentie†
CG 27 (48.2) 8 (38.1) 0.79 0.706 0.77 0.684
GG 13 (23.2) 7 (33.3) 1.44 0.589 1.82 0.407
CG+GG 40 (71.4) 15 (71.4) 1.00 1.000 1.05 0.933
Frequentie G allel
47% 52%
TGFβ1 c.74G>C GG 50 (89.3) 14 (70.0) Referentie†
Referentie†
GC 5 (8.9) 5 (25.0) 3.57 0.379 3.24 0.105
CC 1 (1.8) 1 (5.0) 3.57 0.069 1.63 0.744
GC+CC 6 (10.7) 6 (30.0) 3.57 0.051 2.90 0.117 Frequentie C allel
6% 18%
†Analyse volgens dominant model, HN als referentie.
Afkortingen: OR = odds ratio; ORcorr = OR gecorrigeerd voor chemotherapie en Dmean SPC.
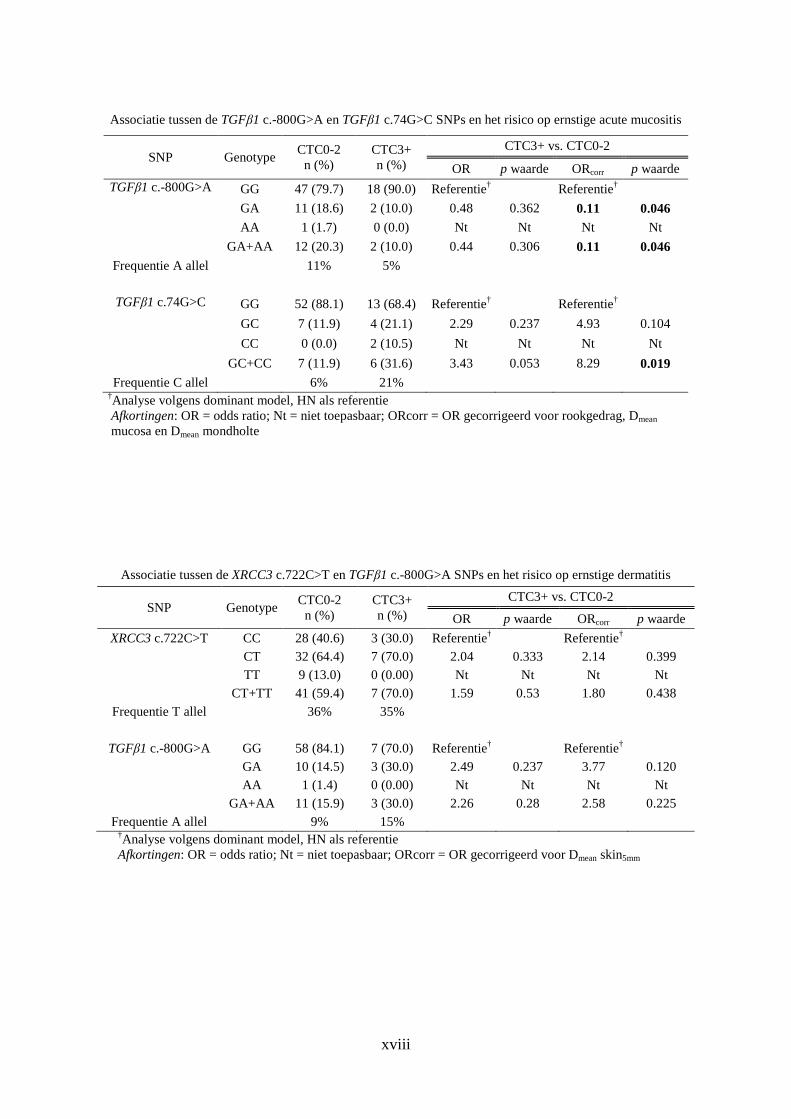
xviii
Associatie tussen de TGFβ1 c.-800G>A en TGFβ1 c.74G>C SNPs en het risico op ernstige acute mucositis
SNP Genotype CTC0-2 n (%)
CTC3+ n (%)
CTC3+ vs. CTC0-2
OR p waarde ORcorr p waarde TGFβ1 c.-800G>A
GG 47 (79.7) 18 (90.0) Referentie†
Referentie†
GA 11 (18.6) 2 (10.0) 0.48 0.362 0.11 0.046 AA 1 (1.7) 0 (0.0) Nt Nt Nt Nt
GA+AA 12 (20.3) 2 (10.0) 0.44 0.306 0.11 0.046 Frequentie A allel
11% 5%
TGFβ1 c.74G>C GG 52 (88.1) 13 (68.4) Referentie†
Referentie†
GC 7 (11.9) 4 (21.1) 2.29 0.237 4.93 0.104
CC 0 (0.0) 2 (10.5) Nt Nt Nt Nt
GC+CC 7 (11.9) 6 (31.6) 3.43 0.053 8.29 0.019 Frequentie C allel
6% 21%
†Analyse volgens dominant model, HN als referentie
Afkortingen: OR = odds ratio; Nt = niet toepasbaar; ORcorr = OR gecorrigeerd voor rookgedrag, Dmean mucosa en Dmean mondholte
Associatie tussen de XRCC3 c.722C>T en TGFβ1 c.-800G>A SNPs en het risico op ernstige dermatitis
SNP Genotype CTC0-2 n (%)
CTC3+ n (%)
CTC3+ vs. CTC0-2
OR p waarde ORcorr p waarde
XRCC3 c.722C>T CC 28 (40.6) 3 (30.0) Referentie†
Referentie†
CT 32 (64.4) 7 (70.0) 2.04 0.333 2.14 0.399
TT 9 (13.0) 0 (0.00) Nt Nt Nt Nt
CT+TT 41 (59.4) 7 (70.0) 1.59 0.53 1.80 0.438
Frequentie T allel
36% 35%
TGFβ1 c.-800G>A GG 58 (84.1) 7 (70.0) Referentie† Referentie†
GA 10 (14.5) 3 (30.0) 2.49 0.237 3.77 0.120
AA 1 (1.4) 0 (0.00) Nt Nt Nt Nt
GA+AA 11 (15.9) 3 (30.0) 2.26 0.28 2.58 0.225 Frequentie A allel
9% 15%
†Analyse volgens dominant model, HN als referentie
Afkortingen: OR = odds ratio; Nt = niet toepasbaar; ORcorr = OR gecorrigeerd voor Dmean skin5mm


















