
De waarde van medische beeldvorming bij de bepaling van de ...
Upload
pieter-charleCategory
view
113download
0
Faculteit Geneeskunde en Gezondheidswetenschappen
Opleiding Lichamelijke Opvoeding en Bewegingswetenschappen
Academiejaar 2012-2013
BEPALING VAN DE MORFOLOGIE VAN DE
BEENMUSCULATUUR BIJ TOPSPRINTERS EN -
DUURLOPERS
Masterproef voorgelegd tot het behalen van de graad van Master in de Lichamelijke
Opvoeding en Bewegingswetenschappen
Door: Pieter Charle en Griet Monkerhey
Promotor: Prof. Dr. W. Derave
Copromotor: Prof. Dr. D. De Clercq
Begeleider: Tine Bex

II

III
VOORWOORD
Na een bacheloropleiding van drie jaar aan de Arteveldehogeschool Gent strandden we
samen aan de universiteit. Vol goede moed, maar met een klein hartje betraden we in
september 2010 voor de eerste keer het HILO. De twee schakeljaren die ons stonden te
wachten, hadden de reputatie hard en moeilijk te zijn. Tijdens deze twee theoretische
jaren hebben we elkaar telkens trachten te helpen en vonden we steun bij elkaar wanneer
het wat moeilijker ging. Na twee jaar intense samenwerking leek samen een scriptie
schrijven dan ook de ultieme bekroning van onze masteropleiding.
Door onze drie jaar praktijkervaring op de hogeschool waren we het al snel eens dat onze
scriptie van wetenschappelijke aard zou zijn. Bij het doornemen van de
thesisonderwerpen was er één titel die er voor ons beiden uitsprong, namelijk bepaling
van de morfologie van de beenmusculatuur bij topsprinters en -duurlopers. Er waren
echter meerdere kandidaten voor hetzelfde onderwerp dus een goede motivatiebrief was
een must. We werden geselecteerd en dat betekende heel wat voor ons. Het
voortgangsrapport dat we schreven op het einde van het tweede schakeljaar was een
bevestiging dat we het juiste onderwerp hadden gekozen.
Naast de ontdekkingen die we deden in de literatuur deden de testen in het UZ en de
ontmoetingen met topatleten ons beseffen dat we deel uitmaakten van een toponderzoek.
We bedanken hier dan ook graag Prof. dr. Wim Derave die ons het vertrouwen gaf om
van dit onderzoek deel uit te maken. Door zijn innovatieve input en kennis waren we in
staat onze thesis naar een hoger niveau te tillen.
Uiteraard gaat er speciale dank uit naar onze thesisbegeleidster, Tine Bex. Haar
enthousiasme en professionaliteit werkten enorm aanstekelijk. Wanneer we een stuk
doorstuurden, kregen we vaak enkele uren later al een mailtje met de vraag wanneer we
op feedback konden komen. Altijd stond ze voor ons klaar, gaf ze ons opbouwende
kritiek, reikte vernieuwende ideeën aan en dit alles steeds met een glimlach. Tine, veel
succes verder met je doctoraat en een dikke merci!
Tenslotte nog een woord van dank voor de atleten, zonder wie dit onderzoek onmogelijk
was geweest. We waren verheugd dat zij in hun druk trainingsschema nog een gaatje
vonden om hun steentje bij te dragen aan de wetenschap.
Griet en Pieter

IV
SAMENVATTING
Doelstelling: Duurlopers en sprinters beschikken over een welbepaalde morfologie van de
beenmusculatuur. In dit onderzoek werd deze morfologie in beeld gebracht en vergeleken
tussen beide groepen atleten. De rode draad van deze scriptie bestaat uit een duurloop- en
sprintprestatieschema waarin de prestatiebepalende factoren voor de 60-100 m sprint en de 3-
10 km opgelijst worden. Het onderzoek ging de correctheid na van een deel van deze
prestatiebepalende factoren. Het ultieme doel was om met een kort onderzoek te kunnen
bepalen of een atleet geschikt is voor de duurloopprestatie of voor de sprintprestatie.
Onderzoeksmethode: 13 atleten van Belgisch subtop- en topniveau namen deel aan deze
observationele cross-sectionele studie. Zeven van hen waren duurlopers, de andere zes waren
sprinters. Het onderzoek was vierledig. Ten eerste werden een aantal antropometrische
eigenschappen onderzocht zoals lengte- en omtrekmaten van het onderste lidmaat en een
aantal vetmetingen. Vervolgens werden met behulp van een ultrasonografietoestel spierdikte,
pennatiehoek en fascicle lengte bepaald van meerdere spieren van het rechterbeen. Daarnaast
werden met behulp van MRI spiervolumes, spier- en botlengtes en massamiddelpunten in
kaart gebracht. Tenslotte werd aan de hand van H-MRS de carnosineconcentratie bepaald in
M. soleus en M. gastrocnemius.
Resultaten: Omtrekken ter hoogte van het bovenbeen, op 30 en 50% proximaal ten opzichte
van de trochanter major, waren groter bij sprinters dan bij duurlopers. Hierbij aansluitend was
de spierdikte groter bij sprinters dan bij duurlopers in M. rectus femoris en M. vastus lateralis.
Spiervolume in M. rectus femoris was eveneens groter bij sprinters. Fascicle lengte was
significant kleiner bij duurlopers in de M. vastus lateralis. Sprinters bezaten hogere waarden
voor carnosineconcentratie in M. soleus en M. gastrocnemius. Pennatiehoek, fysiologische
dwarsdoorsnede en spiermassamiddelpunt waren op geen enkele onderzochte plaats
verschillend tussen duurlopers en sprinters. De beste prestatie bij de sprinters werd in
verband gebracht met een groot spiervolume van de M. rectus femoris, een laag
massamiddelpunt van de M. soleus en van het volledige onderbeen.
Conclusie: Een aantal factoren uit het duurloop- en sprintprestatieschema verschilden
significant tussen duurlopers en sprinters. Slechts zeer weinig factoren correleerden echter
met de beste prestatie. In toekomstig onderzoek kan het nuttig zijn om meer proefpersonen te
recruteren, etnische subgroepen te maken en atleten van wereldtopniveau te gebruiken.

V
INHOUDSTAFEL
VOORWOORD............................................................................................................... III
SAMENVATTING ......................................................................................................... IV
INHOUDSTAFEL............................................................................................................. V
RODE DRAAD................................................................................................................ IX
I. LITERATUURSTUDIE ................................................................................................ 1
SITUERING ....................................................................................................................................................... 2
DEEL 1: DE DUURLOOPPRESTATIE (3-10 KM) ................................................................................................. 3
1. VO2max ................................................................................................................................................. 3
1.1 Zuurstoftransport ............................................................................................................................... 4
1.2 Zuurstofverbruik ............................................................................................................................... 4
1.2.1 Mitochondriën ............................................................................................................................... 4
1.2.2 Energiesystemen ........................................................................................................................... 6
2. Fractie VO2max .................................................................................................................................... 6
3. Loopeconomie ....................................................................................................................................... 7
3.1 Lichaamssamenstelling ..................................................................................................................... 8
3.1.1 Spiermassaverdeling ..................................................................................................................... 9
3.2 Lengtematen .................................................................................................................................... 10
3.2.1 Lichaamslengte ...................................................................................................................... 10
3.2.2 Beenlengte ............................................................................................................................. 11
3.2.3 Voetlengte .............................................................................................................................. 11
3.3 Looptechniek ................................................................................................................................... 13
3.3.1 Loophouding .......................................................................................................................... 13
3.3.2 Staplengte en stapfrequentie .................................................................................................. 13
3.4 Peeseigenschappen .......................................................................................................................... 13
3.5 Spiereigenschappen ......................................................................................................................... 13

VI
DEEL 2: DE SPRINTPRESTATIE (60-100 M) .................................................................................................... 15
1. Prestatiebepalende factoren, onafhankelijk van de sprintfasen .................................................... 15
1.1 Lichaamssamenstelling ................................................................................................................... 15
1.2 Techniek .......................................................................................................................................... 16
1.2.1 Beïnvloedende factoren van staplengte .................................................................................. 17
1.2.2 Beïnvloedende factoren van stapfrequentie ........................................................................... 18
2. Fasespecifieke factoren ...................................................................................................................... 21
2.1 Start ................................................................................................................................................. 21
2.2 Acceleratie ....................................................................................................................................... 22
2.2.1 Netto voorwaartse impuls ...................................................................................................... 22
2.3 Maximale snelheid en aanhouden van de snelheid .......................................................................... 28
DEEL 3: HYPOTHESEN EN ONDERZOEKSVRAGEN ......................................................................................... 29
II. METHODIEK ............................................................................................................. 31
1. Vergelijkende studie .......................................................................................................................... 32
1.1 Proefpersonen .................................................................................................................................. 32
1.2 Studiedesign .................................................................................................................................... 32
2. Meetinstrumenten .............................................................................................................................. 34
2.1 Huidplooicaliper .............................................................................................................................. 34
2.2 Ultrasonografie ................................................................................................................................ 35
2.3 NMR ................................................................................................................................................ 36
3. Analyse ................................................................................................................................................ 38
3.1 Data-analyse .................................................................................................................................... 38
3.1.1 Ultrasonografie ...................................................................................................................... 38
3.1.2 NMR ...................................................................................................................................... 39
3.2 Statistische analyse .......................................................................................................................... 41
III. RESULTATEN .......................................................................................................... 43
1. Inleiding .............................................................................................................................................. 44
2. Antropometrische metingen .............................................................................................................. 44
3. Ultrasonografie ................................................................................................................................... 48
4. NMR .................................................................................................................................................... 50
4.1 MRI ................................................................................................................................................. 50

VII
4.2 H-MRS ............................................................................................................................................ 59
IV. DISCUSSIE ................................................................................................................ 61
1. Proefpersonen en studiedesign .......................................................................................................... 62
2. Prestatiebepalende factoren bij duurlopers en sprinters ............................................................... 63
2.1 Lengtematen .................................................................................................................................... 64
2.2 Spiereigenschappen ......................................................................................................................... 65
2.3 Spiermassaverdeling ....................................................................................................................... 69
3. Conclusie ............................................................................................................................................. 74
V. BIBLIOGRAFIE ......................................................................................................... 79
VI. BIJLAGEN ................................................................................................................. 87
Bijlage 1: vragenlijst voor de atleet ............................................................................................................ 88

VIII

IX
RODE DRAAD
Aangezien de duurloop- en de sprintprestatie worden bepaald door talrijke factoren
trachten wij deze in een overzichtelijke structuur te ordenen. Het resultaat van deze
poging zijn twee schema’s die de duurloop- en sprintprestatie voor respectievelijk
drie tot tien kilometer en 60 tot 100 meter beschrijven (figuur 1 en 2). We proberen
hierbij volledigheid na te streven, maar beseffen dat hier en daar aanvullingen mogelijk
zijn.
Als eerste bespreken we de duurloopprestatie. Er zijn hierbij drie belangrijke
prestatiebepalende factoren, namelijk het fractioneel gebruik van de VO2max tijdens de
prestatie, de VO2max en de loopeconomie. Deze factoren kunnen echter niet los van
elkaar gezien worden. Er is steeds een onderlinge beïnvloeding. Het fractioneel gebruik
van de VO2max wordt voornamelijk bepaald door het aerobe en het anaerobe
energiesysteem. De lactaateliminatie speelt hierbij een cruciale rol. Vervolgens
onderscheiden we bij de VO2max drie fases, namelijk de maximale aanvoer, het
maximale transport en het maximale verbruik van zuurstof. In onze studie zullen de
laatste twee fases aan bod komen omdat vooral deze bepalend zijn voor de
duurloopprestatie. Onder deze fases bevinden zich nog een aantal andere factoren die
terug te vinden zijn in het schema. Loopeconomie, de laatste prestatiebepalende factor,
laat zich kenmerken door de lichaamssamenstelling, lengtematen, de looptechniek, de
peeseigenschappen en de spiereigenschappen van een atleet. Ook hier zijn de volgens ons
meest belangrijke onderliggende factoren terug te vinden in het schema. Ons onderzoek
zal zich voornamelijk focussen op de laatstgenoemde prestatiebepalende factor, namelijk
loopeconomie. Met behulp van antropometrische metingen, ultrasonografie, MRI en H-
MRS kunnen we de zaken die hieronder vallen nauwkeurig onderzoeken.
Vervolgens bespreken we de sprintprestatie. Deze prestatie laat zich kenmerken door
verschillende fases, wat het opstellen van een schema bemoeilijkt. We kozen ervoor om
de sprintprestatie op te delen in drie fases, namelijk de startfase, de acceleratiefase en de
fase waarin de maximale snelheid wordt bereikt en zo lang mogelijk wordt aangehouden.
Bij de startfase speelt voornamelijk de reactietijd een rol. Bij de acceleratie vervolgens is
de netto voorwaartse impuls de grootste prestatiebepalende factor. Deze wordt bepaald
door de contacttijd van de voet, de spierverkortingssnelheid en in mindere mate door de
peeseigenschappen. In de laatste fase spelen de energiesystemen een grote rol, waarbij

X
een opdeling kan gemaakt worden tussen aerobe en anaerobe energiesystemen. Daarnaast
zijn er factoren die op alle fases een invloed uitoefenen zoals de lichaamssamenstelling
en de techniek van een atleet. Ook hier zullen we de aangeduide factoren in het schema
met behulp van antropometrische metingen, ultrasonografie, MRI en H-MRS nauwkeurig
beschrijven.
Naast de opgesomde prestatiebepalende factoren zijn er zowel bij de duurloop- als bij de
sprintprestatie andere factoren die een potentiële invloed hebben op de prestatie. Hierbij
denken we bijvoorbeeld aan training, blessuregevoeligheid, omgeving, psychologische
factoren en nog vele andere. Deze zaken worden niet behandeld in onze scriptie omdat dit
ons te ver zou leiden.
We doen een poging om onze scriptie te structureren met behulp van de door ons
opgebouwde schema’s. Het ultieme doel is om met een kort onderzoek te kunnen bepalen
of een atleet geschikt is voor de duurloopprestatie of voor de sprintprestatie. Dit doen we
aan de hand van de door ons gekozen en onderzochte factoren.
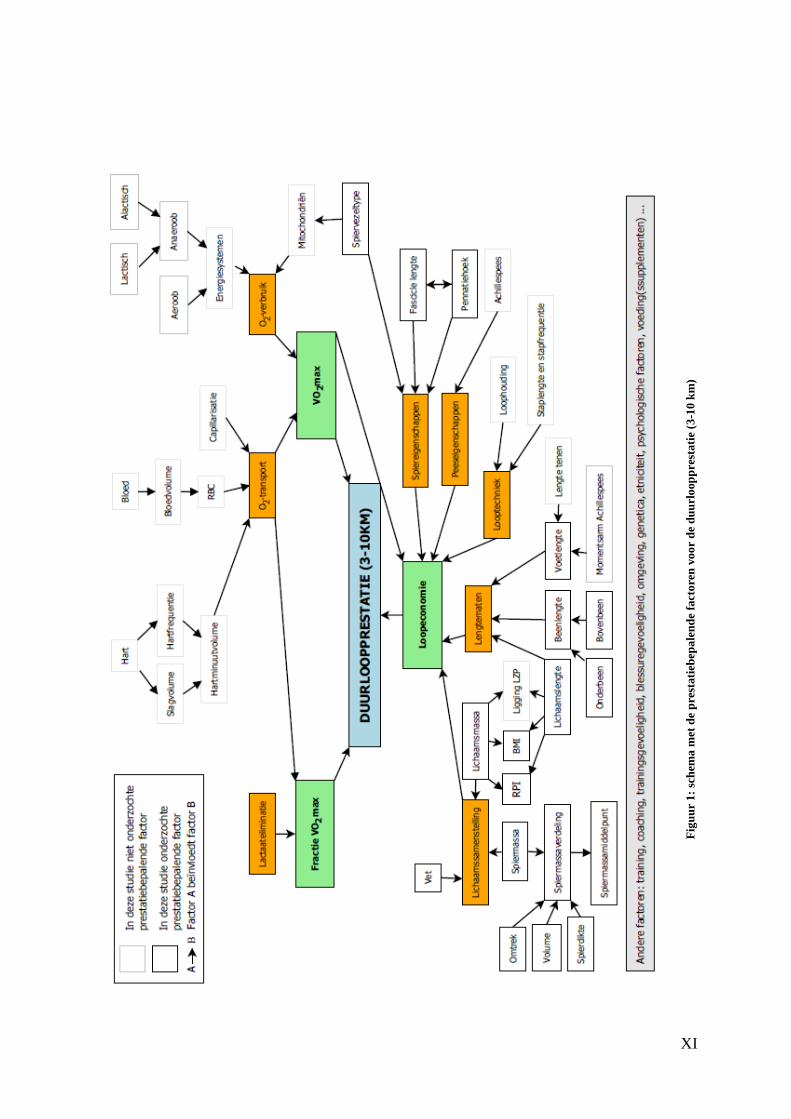
XI
Fig
uu
r 1
: sc
hem
a m
et d
e p
rest
ati
ebep
ale
nd
e fa
cto
ren
vo
or
de
du
url
oop
pre
sta
tie
(3-1
0 k
m)
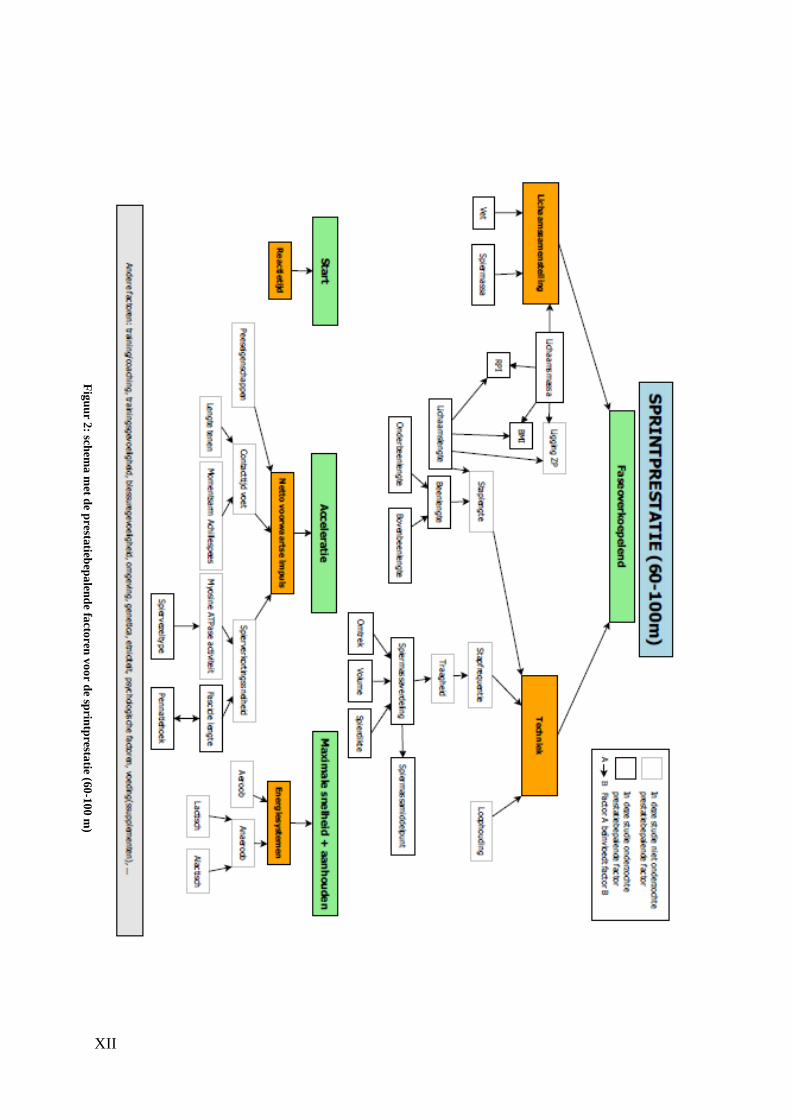
XII
Fig
uu
r 2: sch
ema
met d
e presta
tiebep
alen
de fa
ctoren
vo
or d
e sprin
tpresta
tie (60
-10
0 m
)

1
I. LITERATUURSTUDIE

Literatuurstudie
2
SITUERING
Wanneer een atleet in de Olympische finale van het jaar 1900 de
100 m sprint liep in een tijd van 11 seconden, dan werd dit als
een mirakel beschouwd (Quercetani & Pallicca, 2006). Vandaag
zou deze tijd niet eens volstaan voor een atleet om zich te
kwalificeren voor een deelname aan een Wereldkampioenschap
of de Olympische Spelen. Het huidige wereldrecord van Usain
Bolt van 9.58s suggereert een significante vooruitgang van de
prestatie over de tijd. Bij duurlopen zien we hetzelfde fenomeen.
Patrick Makau slaagde erin de marathon te lopen in 2u 03min
38s, terwijl het wereldrecord in 1954 nog 2u 17min 40s was.
Als er gekeken wordt naar het uiterlijk van bovenvermelde twee
atleten kan men duidelijke verschillen opmerken. Een
sprinterslichaam is over het algemeen krachtig en gespierd,
terwijl een typische duurloper eerder smal en pezig is. Komen
alle personen met bovenstaande uiterlijke kenmerken nu in
aanmerking om een goede sprinter of duurloper te worden of
spelen er nog andere dieperliggende mechanismen een rol? Veel
onderzoekers werden reeds gebeten door de vraag naar wat nu
precies de prestatiebepalende factoren zijn voor een goede
sprint- of duurloopprestatie.
In deze literatuurstudie worden de reeds onderzochte prestatiebepalende factoren op een
rijtje gezet. De literatuurstudie wordt opgedeeld in twee grote onderdelen. In het eerste
deel wordt de duurloopprestatie onder de loep genomen, in het tweede deel ligt de
focus op de sprintprestatie. Alle factoren uit de schema’s van de rode draad worden
aangehaald, maar er wordt enkel dieper ingegaan op diegene die in een latere fase ook
zullen onderzocht worden.
Opmerkelijk is dat in de literatuur vaak de focus ligt op het onderzoeken van de
sprintprestatie enerzijds of de duurprestatie anderzijds. Beide prestaties worden niet vaak
met elkaar vergeleken. In een latere fase van deze scriptie wordt getracht dergelijke
vergelijking te bewerkstelligen. Om beide groepen optimaal te kunnen vergelijken
worden dezelfde prestatiebepalende factoren onderzocht en in vraag gesteld.
Literatuurstudie
3
DEEL 1: DE DUURLOOPPRESTATIE (3-10 KM)
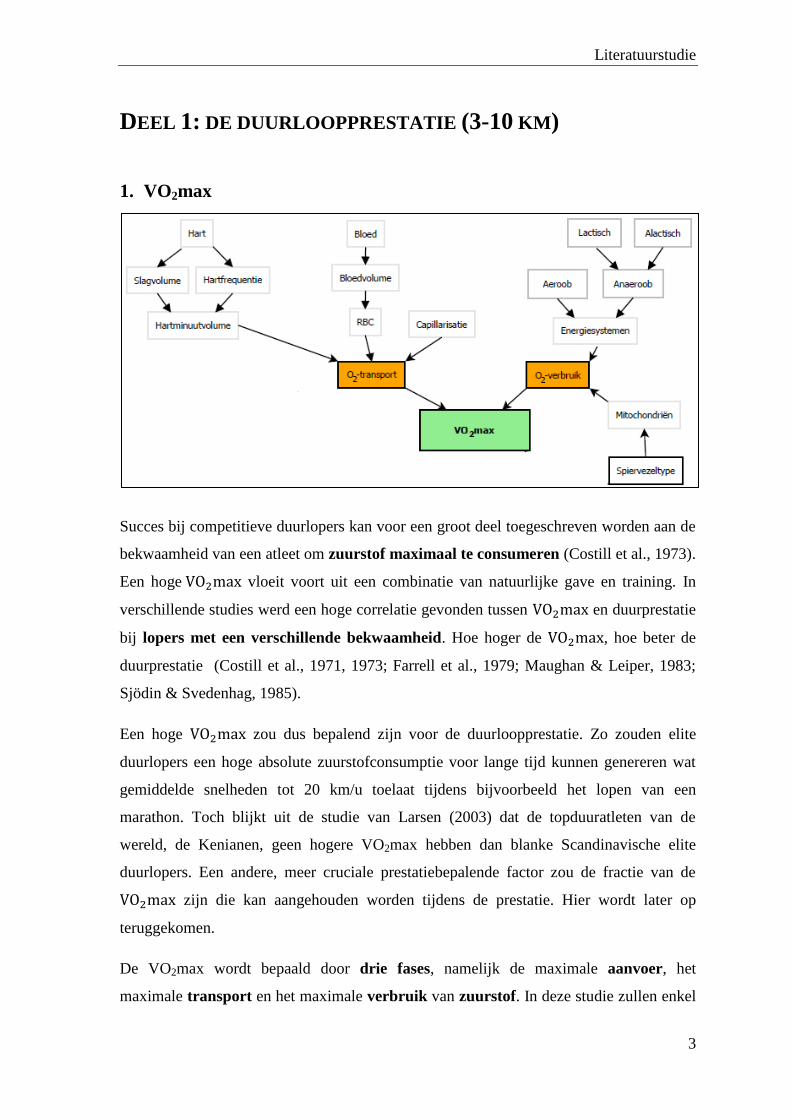
1. VO2max
Succes bij competitieve duurlopers kan voor een groot deel toegeschreven worden aan de
bekwaamheid van een atleet om zuurstof maximaal te consumeren (Costill et al., 1973).
Een hoge vloeit voort uit een combinatie van natuurlijke gave en training. In
verschillende studies werd een hoge correlatie gevonden tussen en duurprestatie
bij lopers met een verschillende bekwaamheid. Hoe hoger de , hoe beter de
duurprestatie (Costill et al., 1971, 1973; Farrell et al., 1979; Maughan & Leiper, 1983;
Sjödin & Svedenhag, 1985).
Een hoge zou dus bepalend zijn voor de duurloopprestatie. Zo zouden elite
duurlopers een hoge absolute zuurstofconsumptie voor lange tijd kunnen genereren wat
gemiddelde snelheden tot 20 km/u toelaat tijdens bijvoorbeeld het lopen van een
marathon. Toch blijkt uit de studie van Larsen (2003) dat de topduuratleten van de
wereld, de Kenianen, geen hogere VO2max hebben dan blanke Scandinavische elite
duurlopers. Een andere, meer cruciale prestatiebepalende factor zou de fractie van de
zijn die kan aangehouden worden tijdens de prestatie. Hier wordt later op
teruggekomen.
De VO2max wordt bepaald door drie fases, namelijk de maximale aanvoer, het
maximale transport en het maximale verbruik van zuurstof. In deze studie zullen enkel

Literatuurstudie
4
de laatste twee fases aan bod komen omdat vooral deze bepalend zijn voor de
duurloopprestatie.
1.1 Zuurstoftransport
Voor het hebben van een hoge VO2max is er een optimaal transport van zuurstof nodig.
Er zijn enkele factoren die dit transport op een gunstige manier kunnen beïnvloeden. Een
eerste is een groot hartminuutvolume (slagvolume x hartfrequentie). Één van de
trainingsadaptaties van uithoudingstraining is het groter en sterker worden van het hart.
Dit zorgt voor een vergroot slagvolume wat op zijn beurt een positief effect heeft op het
hartminuutvolume. Ten tweede zijn er aanpassingen door uithoudingstraining in het
bloed die het zuurstoftransport gunstig kunnen beïnvloeden. Zo zorgt een groot
bloedvolume voor een hogere absolute hoeveelheid rode bloedcellen. Een laatste
gunstige factor bij het zuurstoftransport is de capillarisatie. Volgens Larsen (2003)
correleert de capillaire densiteit positief met de loopsnelheid waarbij bloedlactaat begint
op te hopen. Er is een trend tot hogere capillarisatie bij Keniaanse elite lopers dan bij
Deense. Ongetrainde Keniaanse en Deense jongens met vergelijkbare VO2max hebben
gelijke capillarisatie.
1.2 Zuurstofverbruik
1.2.1 Mitochondriën
Naast zuurstoftransport zal ook het zuurstofverbruik een prestatiebepalende factor zijn
voor een hoge VO2max. Een eerste bepalende factor van het zuurstofverbruik is het
aantal mitochondriën. Hoe meer mitochondriën, hoe meer zuurstof kan verbruikt
worden. Het aantal mitochondriën wordt naast training mede bepaald door het type
spiervezels waarover een atleet beschikt.
Men onderscheidt bij zoogdieren, inclusief
de mens, twee hoofdcategorieën onder de
spiervezeltypes. Enerzijds bestaan er de
rode, hoog oxidatieve, trage spiervezels,
ook wel de type 1 of slow twitch (ST)
vezels genaamd. Deze vezels bevatten veel
mitochondriën. Anderzijds zijn er de
glycolytische, snelle spiervezels, kortweg
Figuur 3: ATPase-kleuring van een spierbiopsie met
aanduiding van volgende spiervezeltypes: ST (cirkel),
FTa (driehoek), FTx (sterretje) en ongeclassificeerd
(halve cirkel) (Saltin et al., 1977).
Literatuurstudie
5
de type 2 of fast twitch (FT) vezels. Deze vezels bevatten minder mitochondriën dan de
type 1 vezels. De type 2 vezels worden op hun beurt onderverdeeld in twee
subcategorieën, de type 2a vezels en de type 2x vezels. De type 2a vezels hebben een
oxidatief potentieel dat hoger is dan de type 2x vezels, terwijl de type 2x vezels dan weer
een glycolytisch potentieel hebben dat veel hoger ligt dan bij de type 2a vezels (Gollnick
& Matoba, 1984; Saltin et al., 1977).
Duurlopers hebben een hoger
percentage ST vezels in hun spieren
(Saltin et al., 1977; Wahren et al.,
1971; Nygaard & Goricke, 1976;
Gollnick et al., 1972; Costill et al.,
1976). Deze resultaten werden
bekomen door het gebruik van een
invasieve methode, namelijk de
spierbiopsie. Nadelen aan deze
methode zijn de grote variantie en
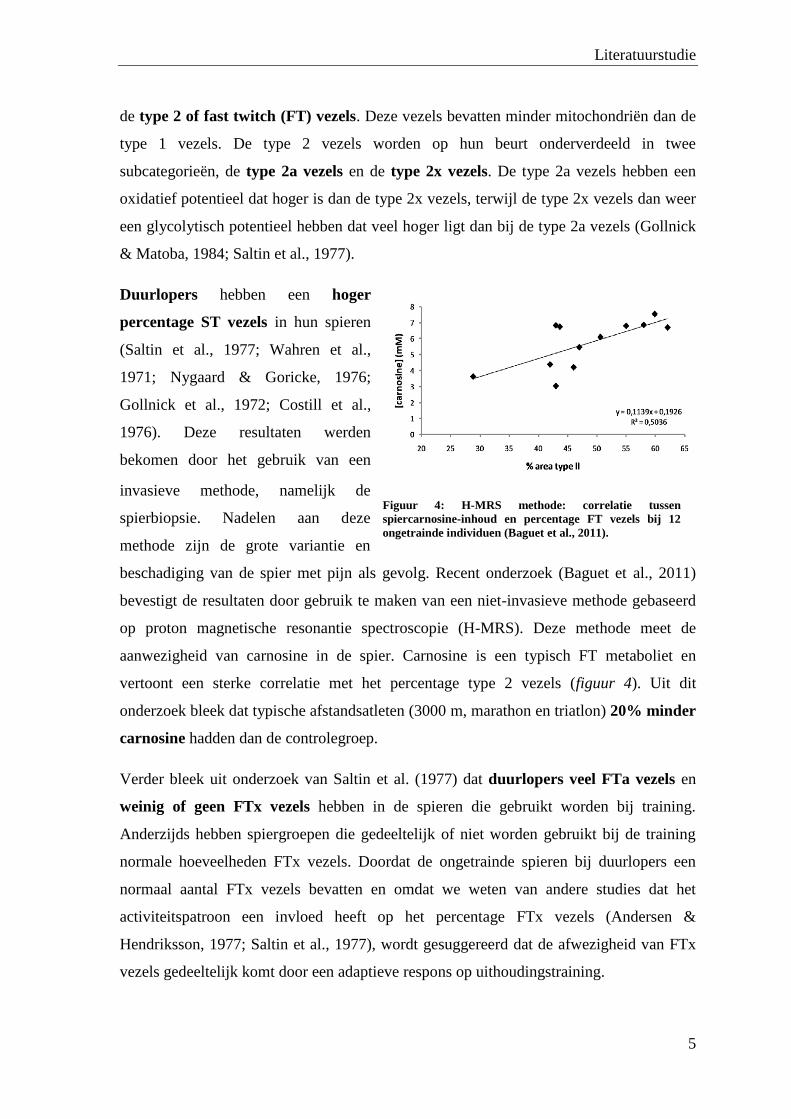
beschadiging van de spier met pijn als gevolg. Recent onderzoek (Baguet et al., 2011)
bevestigt de resultaten door gebruik te maken van een niet-invasieve methode gebaseerd
op proton magnetische resonantie spectroscopie (H-MRS). Deze methode meet de
aanwezigheid van carnosine in de spier. Carnosine is een typisch FT metaboliet en
vertoont een sterke correlatie met het percentage type 2 vezels (figuur 4). Uit dit
onderzoek bleek dat typische afstandsatleten (3000 m, marathon en triatlon) 20% minder
carnosine hadden dan de controlegroep.
Verder bleek uit onderzoek van Saltin et al. (1977) dat duurlopers veel FTa vezels en
weinig of geen FTx vezels hebben in de spieren die gebruikt worden bij training.
Anderzijds hebben spiergroepen die gedeeltelijk of niet worden gebruikt bij de training
normale hoeveelheden FTx vezels. Doordat de ongetrainde spieren bij duurlopers een
normaal aantal FTx vezels bevatten en omdat we weten van andere studies dat het
activiteitspatroon een invloed heeft op het percentage FTx vezels (Andersen &
Hendriksson, 1977; Saltin et al., 1977), wordt gesuggereerd dat de afwezigheid van FTx
vezels gedeeltelijk komt door een adaptieve respons op uithoudingstraining.
Figuur 4: H-MRS methode: correlatie tussen
spiercarnosine-inhoud en percentage FT vezels bij 12
ongetrainde individuen (Baguet et al., 2011).

Literatuurstudie
6
Ama et al. (1986) onderzochten het verschil tussen sedentaire Centraal- en West-
Afrikanen en blanken. Uit zijn studie bleek dat de Afrikanen significant meer type 2a
vezels hadden en dat de blanken over meer type 1 vezels beschikten. De studie van Abe
et al. (1999) bevestigt deze resultaten. Aangezien beide studies beschikten over
voldoende proefpersonen zijn deze resultaten aannemelijk. Ander onderzoek (Larsen,
2003) toonde aan dat Kenianen en blanken een gelijke proportie type 1 vezels bezitten.
Ook wanneer Nandi’s, de best presterende Kenianen, vergeleken werden met Denen werd
er geen significant verschil gevonden wat betreft type 1 vezels. Deze studie is niet
gepubliceerd dus de resultaten moeten met enige voorzichtigheid geïnterpreteerd worden.
1.2.2 Energiesystemen
Een tweede factor die het verbruik van zuurstof bepaalt, is het gebruik van
energiesystemen. Er wordt een onderscheid gemaakt tussen drie energiesystemen,
namelijk het anaeroob alactisch systeem (fosfageensysteem), het anaeroob lactisch
systeem (anaerobe glycolyse) en het aerobe systeem. Het laatst genoemde systeem
haalt energie uit de oxidatie van glycogeen (en bloedglucose) of vetten (en eiwitten)
waardoor de resynthese van ATP mogelijk is. Deze reacties vinden plaats in de
mitochondriën. Uit onderzoek blijkt dat het aerobe energiesysteem bij duurlopers sterk
ontwikkeld is. Zo zouden zij een 50% hogere oxidatieve capaciteit hebben dan sprinters
(Crowther et al., 2002). Verder onderzoek is echter vereist om de precieze aard van de
aerobe adaptatie te achterhalen.
Wanneer de enzymatische activiteit werd vergeleken tussen Kenianen en blanken, vond
men geen verschil in oxidatieve enzymes (Saltin et al., 1995). Wanneer in diezelfde studie
werd gekeken naar het beta-oxidatieve enzym HAD (3-hydroxyacyl-CoA-dehydrogenase
activity) dan vond men een hogere activiteit van dit enzym terug bij de Kenianen.
2. Fractie VO2max

Literatuurstudie
7
De fractie van de wijst op het percentage van de dat voor lange tijd kan
aangehouden worden tijdens wedstrijden zonder de accumulatie van lactaat in het
bloed. De lactaatconcentratie in het bloed is de resultante van de aanmaak van lactaat in
de spiercel en de afvoer van lactaat uit de bloedbaan. Ophoping van lactaat onstaat
wanneer de lactaateliminatie lager is dan de lactaataccumulatie. Het punt waarbij
accumulatie en eliminatie nog net in evenwicht zijn, wordt ook wel de anaerobe drempel
genoemd. Elite atleten hebben een hogere anaerobe drempel dan ongetrainden en kunnen
dus aan een hogere fractie van hun lopen tijdens wedstrijden (Costill et al.,
1973; Joyner & Coyle, 2008). Bij duurlopers met vergelijkbare bekwaamheid is het
fractioneel gebruik van de tijdens prestatie een betere predictor van succes
dan de .
3. Loopeconomie
Naast fractioneel gebruik van de tijdens
prestatie is ook de loopeconomie een betere
voorspeller van succes dan de bij
duurlopers van gelijk niveau (Conley &
Krahenbuhl, 1980; Saunders et al., 2004). De
loopeconomie drukt de energiebehoefte uit voor
een bepaalde submaximale loopsnelheid. In
figuur 5 heeft subject één een betere loopeconomie
dan subject twee want de steady-state
zuurstofconsumptie ( uitgedrukt in
Figuur 5: vergelijking in
) tussen twee 10km
lopers van internationaal niveau (Saunders et
al., unpublished data, 2003).

Literatuurstudie
8
) aan een gegeven loopsnelheid is lager bij subject één. Als we
lichaamsmassa in rekening brengen, gebruiken lopers met een betere loopeconomie
minder energie en dus minder zuurstof dan lopers met een slechtere loopeconomie aan
een gegeven snelheid. De zuurstofbehoefte aan een gegeven snelheid kan tot 30 à 40%
variëren tussen individuen (Joyner & Coyle, 2008; Saunders et al., 2004).
Er is een sterke associatie tussen loopeconomie en de prestatie bij duurlopers.
Onderzoek van Conley en Krahenbuhl (1980) toonde aan dat er een significante correlatie
bestaat tussen de aerobe vereiste aan een bepaalde snelheid en de looptijd voor 10 km.
Hoe hoger de aan een bepaalde snelheid, hoe langer de looptijd. 65,4% van de
variantie in loopprestatie voor de 10 km kon verklaard worden door loopeconomie.
Heel wat factoren kunnen op hun beurt de loopeconomie beïnvloeden, zoals te zien is op
het duurloopprestatieschema uit de rode draad. Er kunnen vijf beïnvloedende factoren
onderscheiden worden, namelijk lichaamssamenstelling, lengtematen, looptechniek,
peeseigenschappen en spiereigenschappen. Elk van deze wordt hieronder grondig
besproken.
3.1 Lichaamssamenstelling
Een eerste factor die de lichaamssamenstelling bepaalt, is het aanwezige lichaamsvet.
Vucetic et al. (2005) toonden aan dat duurlopers minder lichaamsvet hebben dan atleten
van de meeste andere sportdisciplines.
Daarnaast kan lichaamsmassa de lichaamssamenstelling beïnvloeden. De lichaamsmassa
van atleten die gespecialiseerd zijn in loopnummers tussen de 800 m en de 10 km was
omgekeerd gerelateerd aan de afstand: hoe langer de afstand, hoe lager de lichaamsmassa
(Weyand & Davis, 2005). De verdeling van de lichaamsmassa kan een invloed hebben op
de ligging van het lichaamszwaartepunt. Bovendien kunnen we met behulp van de
lichaamsmassa en de lichaamslengte (zie verder) de body mass index (BMI) berekenen.
Dit is een maat om de lichaamsstructuur te beschrijven. Bij atleten slaat het BMI
voornamelijk op de aanwezige spiermassa. Bij niet-atleten is het BMI een indicatie voor
het aanwezige lichaamsvet (Uth, 2005). Toch geeft BMI bij onderzoek naar atleten
weinig nuttige informatie, aangezien we de lichaamssamenstelling niet rechtstreeks uit de
body mass index kunnen afleiden. Vucetic et al. (2005) toonden aan dat onder de atleten
duurlopers het laagste BMI hebben.

Literatuurstudie
9
Als laatste kan de spiermassa gezien worden als een beïnvloedende factor van de
lichaamssamenstelling. Spiermassa is op zijn beurt gerelateerd aan de
spiermassaverdeling die hieronder uitgebreid besproken wordt.
3.1.1 Spiermassaverdeling
Om spiermassa en de verdeling ervan te meten worden verscheidene technieken gebruikt.
In deze literatuurstudie wordt enkel ingegaan op de spiermassaverdeling ter hoogte van
het onderste lidmaat. Uit de spiermassaverdeling kan vervolgens het
spiermassamiddelpunt berekend worden.
Een eerste methode om spiermassaverdeling te bepalen, is het meten van de omtrek van
het onderste lidmaat. Vucetic et al. (2005) stelden in een vergelijkende studie verschillen
vast in omtrek van de ledematen tussen sprinters (100 m en 200 m), uithoudingssprinters
(400 m), middellange afstandsatleten en lange afstandsatleten. Uit deze studie bleek
onder andere dat lange afstandsatleten een lagere dijomtrek hebben dan sprinters.
Een tweede manier om de spiermassaverdeling in kaart te brengen, is het gebruik van
volumemetingen. Middellange en lange afstandsatleten hebben gelijke waarden voor
lichaamsvolumemetingen die een stuk lager liggen dan deze van andere atleten (Vucetic
et al., 2005).
Bij bovenstaande twee methoden worden enkele kanttekeningen gemaakt. Zo kan de
omtrek van een lidmaat een vertekend beeld geven doordat het geen rekening houdt met
de verhouding spieren/vet, noch met de dikte van het bot. Wanneer we deze parameter
zouden combineren met metingen van de spierdikte, dan zouden hier meer zinvolle
conclusies uit getrokken kunnen worden. Een tweede opmerking heeft te maken met de
volumemetingen. Over deze methode is slechts weinig te vinden in de literatuur én
bovendien wordt ze weinig gebruikt. Een mogelijke verklaring hiervoor is de
moeilijkheid en tijdrovendheid van deze methode. Een betere, meer gebruikte methode is
het meten van de spierdikte. Deze methode wordt hieronder besproken.
De spierdikte stelt de diepte van een spier voor gemeten tussen vet en bot van een
gekozen spiergroep. Een veel gebruikte meetmethode hiervoor is ultrasonografie (B-mode
ultrasound). In verschillende onderzoeken werden spierdiktes bepaald ter hoogte van 13
anatomische sites (Abe et al., 1999; Abe et al., 2000; Abe et al., 2001; Kumagai et
al.,2000) verspreid over het gehele lichaam. In deze literatuurstudie wordt enkel ingegaan

Literatuurstudie
10
op de anatomische sites ter hoogte van het onderste lidmaat, zoals schematisch
voorgesteld in figuur 6.
Uit een studie van Abe et al. (2000) bleek dat de spierdikte significant groter was bij
duurlopers dan bij de controlegroep ter hoogte van de posterieure dij.
Naast spierdikte kan ook geïsoleerde spierdikte een indicator zijn van de
spiermassaverdeling. In tegenstelling tot spierdikte meet men bij geïsoleerde spierdikte
niet de dikte van een spiergroep op bepaalde anatomische sites, maar de dikte van één
bepaalde spier. Geïsoleerde spierdikte wordt eveneens bepaald aan de hand van B-mode
ultrasound.
3.2 Lengtematen
3.2.1 Lichaamslengte
Wat betreft de lichaamslengte van duurlopers zijn er geen eenduidige resultaten terug te
vinden. Onderzoek van Radford (1990) zegt dat duurlopers gemiddeld groter zijn dan
sprinters. Een studie van Weyand en Davis (2005) meent daarentegen dat de
Figuur 6: 13 anatomische sites waar spierdikte gemeten werd (gemeten vanaf proximaal).

Literatuurstudie
11
lichaamslengte daalt met het groter worden van de gelopen afstand, met uitzondering van
de 400 meter lopers die er als grootsten uitkwamen. Deze laatste studie lijkt de meest
plausibele te zijn als er gekeken wordt naar hedendaagse elite lange afstandsatleten.
3.2.2 Beenlengte
Voorlopig werden er in de literatuur geen significante verschillen teruggevonden tussen
duurlopers en andere atletenpopulaties wat betreft beenlengte, onderbeenlengte en
bovenbeenlengte.
3.2.3 Voetlengte
De teen- en hiellengte leveren een belangrijke bijdrage aan de voetlengte. Dit is meer dan
louter een antropometrisch gegeven. Het heeft een sterke invloed op het biomechanische
aspect bij lopen.
Bij duurlopers blijken kortere tenen een
voordeel op te leveren (Rolian et al., 2008).
Dit voordeel wordt uitgelegd aan de hand
van figuur 7. Bij twee identieke duurlopers
waarbij enkel de teenlengte verschilt, zal de
grootte van de grondreactiekracht (GRK)
gelijk zijn. Doordat persoon B een grotere
teenlengte heeft dan persoon A zal de
afstand (R’) tussen de distale falangen
(waar de GRK aangrijpt) en het rotatiepunt
(r) van het enkelgewricht langer zijn. Dit
impliceert een groter dorsiflexiemoment bij
persoon B. Om dit dorsiflexiemoment te
overwinnen zal persoon B een groter
plantair flexiemoment (F’ flexor) moeten
creëren dan persoon A. Dit leidt tot grotere
krachtimpulsen tijdens standfase en meer mechanisch werk per tijdseenheid, wat op zijn
beurt leidt tot een hogere metabole kost. Kortere tenen kunnen dus de locomotorische
prestatie verbeteren door het verlagen van de metabole kost voor de stabilisatie van de
metatarsofalangeale gewrichten en de krachtproductie van de digitale flexoren (Rolian et
al., 2008).
Figuur 7: schematisch model van de
musculoskeletale functie van de tenen tijdens de
propulsiefase (Rolian et al., 2008). GRF =
grondreactiekracht, R en R’ = afstand tussen
distale falangen en rotatiepunt enkelgewricht,
Fflexor = kracht die op afstand r t.o.v. het
rotatiepunt van het enkelgewricht werkt, fdl = pees
van flexor digitorum longus, MP = middelste
falangen, MT = metatarsalen, PP = proximale
falangen.
Literatuurstudie
12
Een tweede factor die de voetlengte bepaalt, is de
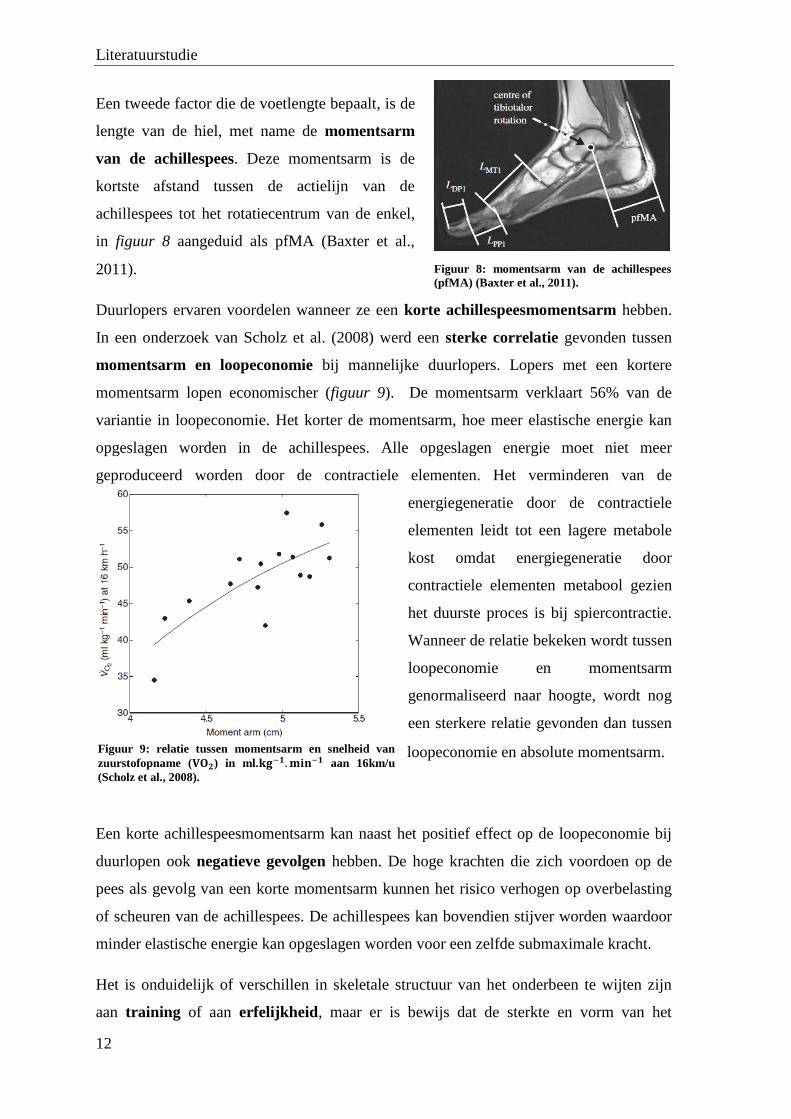
lengte van de hiel, met name de momentsarm
van de achillespees. Deze momentsarm is de
kortste afstand tussen de actielijn van de
achillespees tot het rotatiecentrum van de enkel,
in figuur 8 aangeduid als pfMA (Baxter et al.,
2011).
Duurlopers ervaren voordelen wanneer ze een korte achillespeesmomentsarm hebben.
In een onderzoek van Scholz et al. (2008) werd een sterke correlatie gevonden tussen
momentsarm en loopeconomie bij mannelijke duurlopers. Lopers met een kortere
momentsarm lopen economischer (figuur 9). De momentsarm verklaart 56% van de
variantie in loopeconomie. Het korter de momentsarm, hoe meer elastische energie kan
opgeslagen worden in de achillespees. Alle opgeslagen energie moet niet meer
geproduceerd worden door de contractiele elementen. Het verminderen van de
energiegeneratie door de contractiele
elementen leidt tot een lagere metabole
kost omdat energiegeneratie door
contractiele elementen metabool gezien
het duurste proces is bij spiercontractie.
Wanneer de relatie bekeken wordt tussen
loopeconomie en momentsarm
genormaliseerd naar hoogte, wordt nog
een sterkere relatie gevonden dan tussen
loopeconomie en absolute momentsarm.
Een korte achillespeesmomentsarm kan naast het positief effect op de loopeconomie bij
duurlopen ook negatieve gevolgen hebben. De hoge krachten die zich voordoen op de
pees als gevolg van een korte momentsarm kunnen het risico verhogen op overbelasting
of scheuren van de achillespees. De achillespees kan bovendien stijver worden waardoor
minder elastische energie kan opgeslagen worden voor een zelfde submaximale kracht.
Het is onduidelijk of verschillen in skeletale structuur van het onderbeen te wijten zijn
aan training of aan erfelijkheid, maar er is bewijs dat de sterkte en vorm van het
Figuur 8: momentsarm van de achillespees
(pfMA) (Baxter et al., 2011).
Figuur 9: relatie tussen momentsarm en snelheid van
zuurstofopname ( ) in ml. aan 16km/u
(Scholz et al., 2008).

Literatuurstudie
13
menselijk skelet kunnen veranderen door zekere vormen van training. Het is echter nog
niet aangetoond of menselijke teenlengte en lengte van achillespeesmomentsarm
beïnvloed worden door sporttraining.
3.3 Looptechniek
Om een goede loopeconomie te garanderen zal een looptechniek die zo weinig mogelijk
energie vraagt essentieel zijn. Onder looptechniek worden grosso modo twee zaken
onderscheiden, namelijk loophouding en staplengte en –frequentie.
3.3.1 Loophouding
Om economisch te lopen moet een goede houding gegarandeerd zijn. Het hoofd, de
ledematen en de romp moeten zich in de juiste positie bevinden. Omdat dit niet de
essentie is in dit onderzoek, wordt hier niet dieper op ingegaan.
3.3.2 Staplengte en stapfrequentie
Naast de loophouding van een atleet zullen staplengte en –frequentie ook een invloed
hebben op de loopeconomie. In de literatuur werden voorlopig geen significante
resultaten teruggevonden voor duurlopers in vergelijking met andere atletenpopulaties.
3.4 Peeseigenschappen
Lopen zorgt voor rekverkortingscyclussen in het spierpeescomplex van de onderste
ledematen. Onderzoek toonde aan dat peeseigenschappen belangrijk zijn en gerelateerd
zijn aan sportprestatie tijdens oefeningen betreffende de rekverkortingscyclus (Bojsen-
Moller et al, 2005; Kubo et al., 1999, 2000, 2007; Stafilidis en Arampatzis, 2007). In de
literatuur werden geen significante resultaten teruggevonden voor duurlopers in
tegenstelling tot andere atleten. Aangezien bij duurlopers de excentrische spieractiviteit
wordt aangesproken, lijkt het evident dat de peeseigenschappen een invloed zullen
hebben op de loopeconomie. Zo kan een dunne en lange achillespees bijvoorbeeld een
positieve invloed hebben op de rekverkortingscyclus.
3.5 Spiereigenschappen
Volgens Scott et al. (1993) wordt de pennatiehoek gedefinieerd als de hoek tussen de
fascicles en de actielijn van de spier (figuur 10). Maganaris et al. (1998) stellen dat de
grootte van de pennatiehoek een belangrijke determinant is voor de functionele
karakteristieken van de spier. Zo zorgt een grotere pennatiehoek voor een kortere

Literatuurstudie
14
vezellengte. Deze kortere vezellengte doet enerzijds afbreuk aan de verkortingssnelheid
en de bewegingsgrootte. Anderzijds zorgt ze ervoor dat meer contractiele elementen
parallel geplaatst worden, waardoor de maximale krachtgeneratie toeneemt. Om die reden
is de maximale kracht die geproduceerd wordt aan een welbepaalde vezellengte in de
richting van de vezels bij een
pennate spier groter dan de
maximale kracht die
geproduceerd wordt in de
richting van de vezels van een
parallel gevezelde spier bij
eenzelfde anatomische
dwarsdoorsnede en volume. Een
pennatiehoek boven 45 graden
zou volgens Maganaris et al.
(1998) niet voor een verhoogde
krachtproductie zorgen.
De pennatiehoek zou volgens Abe et al. (2000) groter zijn bij duurlopers in
vergelijking met sprinters. De grotere pennatiehoek bij duurlopers kan verklaard
worden door de kortere fascicle lengte en/of de stijging van de dwarsdoorsnede van de
spier als gevolg van een stijging van de dwarsdoorsnede van de trage spiervezels op
dezelfde volledige spieroppervlakte, geïnduceerd door afstandstraining. Deze
architecturale aanpassing zou voordelig zijn bij het duurlopen.
Figuur 10: 1= bovenste aponeurose, 2= onderste aponeurose, 3=
spiervezelrichting, α= pennatiehoek, d= afstand tussen 1 en 2.

Literatuurstudie
15
DEEL 2: DE SPRINTPRESTATIE (60-100 M)
Bij de sprintprestatie zijn voornamelijk de lichaamssamenstelling en de techniek van
groot belang, onafhankelijk van de fasen waarin de sprintprestatie kan onderverdeeld
worden. Bij het onderdeel techniek kan er bovendien een duidelijk onderscheid gemaakt
worden tussen staplengte, stapfrequentie en loophouding.
Daarnaast zijn er nog een aantal prestatiebepalende factoren die zich vooral manifesteren
in één bepaalde fase van de sprint. De startfase van de sprint wordt bijvoorbeeld
gedomineerd door de reactietijd. Bij de acceleratiefase speelt voornamelijk de netto
voorwaartse impuls een rol. Energiesystemen hebben dan weer een grote invloed op het
bereiken en aanhouden van de maximale snelheid.
1. Prestatiebepalende factoren, onafhankelijk van de sprintfasen
1.1 Lichaamssamenstelling
Net zoals bij duurlopers hebben ook sprinters minder lichaamsvet dan atleten van de
meeste andere sportdisciplines (Vucetic et al., 2005).
Daarnaast is eveneens bij sprinters de lichaamsmassa een bepalende factor van de
lichaamssamenstelling. Een te hoge of te lage lichaamsmassa bij sprinters zou een
limiterende factor kunnen zijn voor de prestatie. Een te hoge lichaamsmassa hindert de
sprinter doordat het een grotere kracht vereist bij het versnellen. Toch hebben goede

Literatuurstudie
16
sprinters veel spieren en daardoor zijn ze zwaarder dan minder goede sprinters. Vandaar
dat sprinters met een te lage lichaamsmassa waarschijnlijk minder spiermassa hebben en
daarom dus minder presteren. Sprinters hebben een grotere lichaamsmassa dan
duurlopers. Onderzoek van Weyand en Davis (2005) toonde aan dat korte afstandslopers
massiever waren dan lange afstandslopers. De meest massieve lopers waren diegenen van
de 100 m, 200 m en 400 m sprint. Specifiek voor sprinters toonde Uth (2005) aan dat er
een tendens is tot minder lichaamsmassavariabiliteit. Dit suggereert, net als bij
lichaamslengte (zie verder), dat er een gelimiteerde zone zou bestaan voor de
lichaamsmassa van sprinters. Zelfde onderzoekers toonden bovendien aan dat sprinters
het hoogste BMI hebben onder de atleten, in tegenstelling tot duurlopers die het laagste
BMI hebben. Aangezien beide groepen een gelijk percentage lichaamsvet hebben, wijst
een hoger BMI op een hogere vetvrije massa.
Bij sprinters is ook de reciprocal ponderal index (RPI) een significante factor voor
succes. De reciprocal ponderal index wordt als volgt berekend: RPI =
(Watts et al., 2012). Het is een maat voor lineariteit
met een hogere score voor slankere (grotere) atleten. Watts et al. (2012) toonden aan dat
de RPI kan beschouwd worden als een voorspeller van succes bij vrouwelijke
elitesprinters. Vanaf de jaren ’80 is de RPI ook bij mannelijke sprinters een voorspeller
van succes geworden. Meer lineaire (slankere en grotere) atleten presteren beter. Een
mogelijke verklaring hiervoor kan zijn dat meer lineaire atleten over het algemeen langere
benen hebben waardoor hun staplengte vergroot. Dit is een bepalende factor voor
snelheid (staplengte x stapfrequentie).
Naast bovenstaande factoren hebben ook spiermassa en de ligging van het
lichaamszwaartepunt een invloed op de lichaamssamenstelling. In een studie van Bejan
et al. (2010) verklaart men de snelheidsverschillen tussen blanken en zwarten met behulp
van het lichaamszwaartepunt. Zwarten zouden een hoger lichaamszwaartepunt hebben
waardoor hun voorwaartse valsnelheid hoger zou liggen en waardoor ze dus sneller
zouden lopen.
1.2 Techniek
Tijdens het trainen moet voldoende aandacht besteed worden aan de loophouding. Het
hoofd, de ledematen en de romp moeten zich immers in een correcte positie bevinden en
in de juiste richting bewegen op het gepaste moment.

Literatuurstudie
17
Naast loophouding, zijn ook een optimale staplengte en stapfrequentie van groot
belang. Deze twee factoren bepalen immers de loopsnelheid. Sprinters met langere benen
produceren de grootste staplengte en de laagste stapfrequentie (Moravec, 1988; Winter,
1990). Hoffman (1971) stelt dat wanneer de benen van een sprinter langer zijn dan de
optimale lengte, het moeilijker wordt om een voldoende hoog beentempo (stapfrequentie)
te produceren. Korte ledematen kunnen daarentegen sneller bewegen dan lange (grotere
stapfrequentie), maar ze kunnen een minder grote staplengte produceren. De beenlengte
moet dus zodanig zijn dat staplengte en stapfrequentie in evenwicht zijn, waarbij het
traagheidsmoment van het naar voor gebrachte been tijdens sprinten zo optimaal mogelijk
overwonnen wordt.
Grote lineaire sprinters zouden dus een nadeel hebben tijdens de eerste helft van de race
doordat ze bij de start minder kracht kunnen genereren. Tijdens de maximale
snelheidsfase zouden ze echter een voordeel hebben door hun grotere staplengte.
Explosieve sprinters met kortere beenlengte zouden hun voordeel halen uit de eerste
helft van de 100 m doordat ze voldoende kracht kunnen genereren om heel snel te kunnen
starten. Tijdens de tweede helft van de wedstrijd ondervinden zij een nadeel door hun
kleinere staplengte.
Usain Bolt heeft een lagere stapfrequentie (4.28Hz) dan zijn concurrenten, maar hij
compenseert dit met een erg grote staplengte die goed van pas komt na de
versnellingsfase van de sprint. Hij loopt de 100 m in minder stappen dan zijn
tegenstrevers (Beneke & Taylor, 2010).
1.2.1 Beïnvloedende factoren van staplengte
Zoals hierboven reeds vermeld zijn de lichaamslengte en de lengte van de benen bepalend
voor de staplengte.
Uth (2005) vond dat er een lagere lichaamslengtevariabiliteit is onder sprinters dan
onder de normale populatie. Er zou een optimale zone bestaan voor de lichaamslengte.
Deze zone ligt voor mannen tussen 1.68 en 1.91 m. Aangezien de lichaamslengte van de
meeste mannen zich in deze zone bevindt, is dit geen goede parameter om sprinters van
de normale populatie te onderscheiden. De meest succesvolle mannelijke sprinters van de
laatste tien jaar bevinden zich bovendien boven deze zone (Watts et al., 2012). Denk maar
aan Usain Bolt die met zijn 1.96 m het wereldrecord van de 100 m sprint op zijn naam
heeft staan (Beneke & Taylor, 2010). Volgens Charles en Bejan (2009) gaan grotere

Literatuurstudie
18
atleten sneller lopen én is de lichaamslengte gestegen doorheen de tijd. Dit heeft als
gevolg dat de snelheidsrecords doorheen de tijd zijn gestegen en nog zullen stijgen.
Wanneer de lichaamslengte zal stoppen met stijgen en welke impact dit zal hebben op de
snelheidsrecords kan niemand voorspellen. Wegens de tegenstrijdige data is de
lichaamslengte een interessant gegeven om in een latere fase te onderzoeken en te
vergelijken tussen sprinters en duurlopers.
Abe et al. (1999) vonden in hun studie waarin ze blanke en zwarte American footballers
vergeleken dat zwarten langere benen hebben ten opzichte van hun lichaamslengte dan
blanken.
Lee en Piazza (2009) toonden aan dat de lengte van het onderbeen 30 mm korter is bij
sprinters dan bij controles. Karamanidis et al. (2011) onderzochten het verschil tussen
snelle en trage sprinters, waarbij het onderbeen bij snelle sprinters 13 mm korter was dan
bij trage sprinters. In hetzelfde onderzoek werd een significante maar gematigde relatie
gevonden tussen onderbeenlengte en beste tijd voor de 100 m sprint. Hoe korter de
onderbeenlengte, hoe beter de tijd voor de 100 m sprint. Dit kan verklaard worden door
een kleiner traagheidsmoment bij een korter onderbeen, wat de loopsnelheid ten goede
komt.
1.2.2 Beïnvloedende factoren van stapfrequentie
De stapfrequentie van een atleet wijst op het aantal stappen dat hij kan zetten per minuut.
Cruciale factoren die de stapfrequentie mee helpen bepalen, zijn de stijfheid en de
traagheid van het been.
De traagheid van het been wordt bepaald door de beenmassa en de spiermassaverdeling.
Om een hoge frequentie te kunnen aanhouden (lage traagheid), mag de beenmassa niet te
groot zijn, maar is er wel voldoende spiermassa nodig op de juiste plaats. Net zoals bij
duurlopers wordt er ook hier een onderscheid gemaakt tussen omtrekmetingen,
volumemetingen en het meten van spierdikte en geïsoleerde spierdikte om de
spiermassaverdeling in kaart te brengen.
Vucetic et al. (2005) stelden in een vergelijkende studie vast dat de omtrek van het
onderbeen groter is bij sprinters (100 m en 200 m) dan bij uithoudingssprinters (400 m),
middellange afstandsatleten en lange afstandsatleten. De omtrek van de dij daarentegen
verschilt enkel tussen sprinters en lange afstandsatleten, waarbij de sprinters een grotere

Literatuurstudie
19
dijomtrek hebben. Deze resultaten zijn in overeenstemming met het feit dat sprinters een
andere spiervorm hebben dan duurlopers (zie verder): sprinters vertonen een dikker
bovenste gedeelte van quadriceps en hamstrings.
Aangezien sprinters meer spiermassa hebben ter hoogte van het bovenste gedeelte van
quadriceps en hamstrings, hebben ze daar ook de hoogste waarden voor volumemetingen
(Vucetic et al., 2005). Iets lagere waarden werden vastgesteld bij uithoudingssprinters.
Meerdere onderzoekers vonden reeds verschillen tussen sprinters onderling of tussen
sprinters en duurlopers of controles wat betreft spierdikte en geïsoleerde spierdikte.
Kumagai et al. (2000) onderzochten de spierdikte op eerdergenoemde 13 anatomische
sites (figuur 6 p.10) bij snelle sprinters (100 m record tussen 10 en 10.9 s) en trage
sprinters (100 m record tussen 11 en 11.70 s). Spierdikte was significant groter bij de
snelle sprinters in het bovenste gedeelte van de dij op volgende plaatsen: 30% anterieur
en 50% posterieur. Ook de spierdikte van het posterieure onderbeen was significant groter
bij snelle sprinters dan bij trage sprinters. Geïsoleerde spierdikte van de M. gastrocnemius
lateralis was significant groter bij de snelle sprinters dan bij de trage.
Uit een studie van Abe et al. (2000) bleek dat de spierdikte significant groter was bij
sprinters dan bij duurlopers voor alle sites ter hoogte van het onderste lidmaat
uitgezonderd 70% anterieure dij. Er was ook een significant verschil tussen sprinters en
de controlegroep, behalve bij 70% anterieure dij en 30% anterieure onderbeen. Volgend
uit bovenstaande resultaten is ook de geïsoleerde spierdikte van M. vastus lateralis, M.
gastrocnemius medialis en M. gastrocnemius lateralis significant groter bij sprinters in
vergelijking met duurlopers en controles, die gelijk waren.
Een vergelijkende studie van Kubo et al. (2011)
tussen 100 meter sprinters en een controlegroep
mat de spierdikte op 14 anatomische sites zoals
weergegeven op figuur 11. Ook hier werd gebruik
gemaakt van ultrasonografie. Bij de knie-
extensoren was er enkel een significant verschil
tussen sprinters en de controlegroep voor de
mediale zijde; spierdikte was groter bij sprinters
dan bij controles. De spierdikte van de plantaire Figuur 11: anatomische sites waar
spierdikte gemeten werd (Kubo et al., 2011).

Literatuurstudie
20
flexoren was op alle sites groter bij sprinters dan bij controles. De beste tijd op de 100 m
voor sprinters correleerde significant met de spierdikte van de mediale zijde van de knie-
extensoren. Er werden geen significante correlaties gevonden met andere gemeten
spierdiktes.
Er kan dus besloten worden dat er significante verschillen bestaan qua spierdikte tussen
sprinters, duurlopers en controles. Over het algemeen is de spierdikte groter bij sprinters
dan bij duurlopers en controles op bijna alle onderzochte anatomische sites. Geïsoleerde
spierdikte is eveneens significant groter bij sprinters dan bij duurlopers en controles, die
gelijke waarden vertoonden.
Zoals eerder beschreven, toonden Abe et al. (2000) aan dat de spierdikte significant groter
is bij sprinters dan bij duurlopers en controles ter hoogte van het proximale gedeelte van
de dij (30 en 50% anterieure dij). Er was echter geen significant verschil ter hoogte van
het distale gedeelte van de dij (70% anterieure dij). Gezien er geen verschil was in
dijlengte tussen de groepen, wijzen deze bevindingen op een verschil in spiervorm
tussen sprinters en de andere groepen. De quadriceps van sprinters hebben dus een andere
vorm dan deze van duurlopers en controles. Kumagai et al. (2000) vonden gelijkaardige
verschillen tussen snelle en trage sprinters. Spierdikte was significant groter bij de snelle
sprinters in het bovenste gedeelte van de dij op volgende plaatsen: 30% anterieur en 50%
posterieur. De spiervorm van quadriceps en hamstrings is dus verschillend tussen snelle
en trage sprinters. De hypothese bestaat dus dat een dikker proximaal gedeelte van
quadriceps en hamstrings geassocieerd is met een betere sprintprestatie. De grotere
spieren ter hoogte van het onderste lidmaat bij sprinters zijn nodig om krachtig te kunnen
afzetten van het startblok bij de start van een sprint. Ook in de versnellingsfase en bij het
onderhouden van de maximale snelheid komen grote krachtige spieren goed van pas.
Duurlopers hebben een heel groot trainingsvolume. Extra massa ter hoogte van de spieren
betekent voor hen geen voordeel. Dit zou een onnodige last vormen. Het is opmerkelijk
dat sprinters geen extra spiermassa vertonen ter hoogte van het distale deel van de dij.
Een verklaring hiervoor kan zijn dat distale massa zich verder van het massacentrum
bevindt dan proximale massa. Distale massa zou dus het traagheidsmoment vergroten en
het slingeren van de benen tijdens het lopen bemoeilijken. Distale massa verhoogt dus de
aerobe vraag meer dan massa die dicht bij het massacentrum gesitueerd is (Saltin et al.,
1995; Saunders et al., 2004).

Literatuurstudie
21
Zijn verschillen in spierdikte en spiervorm een gevolg van genetische variatie of zijn ze
te wijten zijn aan spiertraining? Een studie van Abe (2002) bij identieke tweelingen
toonde aan dat er geen verschil is in spierdikte tussen beiden bij M. gastrocnemius
medialis en lateralis. Dit kan betekenen dat omgevingsfactoren zoals training geen
invloed uitoefenen op de spiervorm. De studie werd uitgevoerd bij negen tweelingen. Bij
drie tweelingen was één persoon actief en de andere sedentair. De zes andere tweelingen
waren sedentair. Bijkomend wetenschappelijk onderzoek rond dit thema is echter nodig
want deze studieresultaten zijn gebaseerd op een zeer laag aantal proefpersonen. Om echt
sluitende resultaten te behalen omtrent de invloed van training op de spiervorm zou ook
het experiment moeten veranderd worden. Zo zou er een studie moeten worden opgezet
waarbij telkens één persoon van de tweeling actief is en de andere sedentair.
Abe et al. (1999) vonden ook verschillen op etnisch vlak omtrent de spierdikte. In een
vergelijkende studie tussen blanke en zwarte American footballers vond hij dat de
spierdikte significant groter was bij de zwarten op 30%-quadriceps en 50%-hamstrings
(bovenste positie van de dij), terwijl de andere plaatsen van de dij niet significant
verschilden. Verder was er voor de geïsoleerde spierdikte geen verschil voor de M. vastus
lateralis en de M. gastrocnemius medialis tussen beide groepen.
2. Fasespecifieke factoren
2.1 Start
Snel begonnen is half gewonnen. Een snelle reactietijd is essentieel voor een goede
sprintprestatie. Wanneer een persoon sneller dan 0,100 seconden reageert op het
startsignaal, dan is er sprake van een valse start. Toch zijn er voorbeelden van atleten die
sneller dan 0,100 seconden kunnen reageren.
Literatuurstudie
22
2.2 Acceleratie
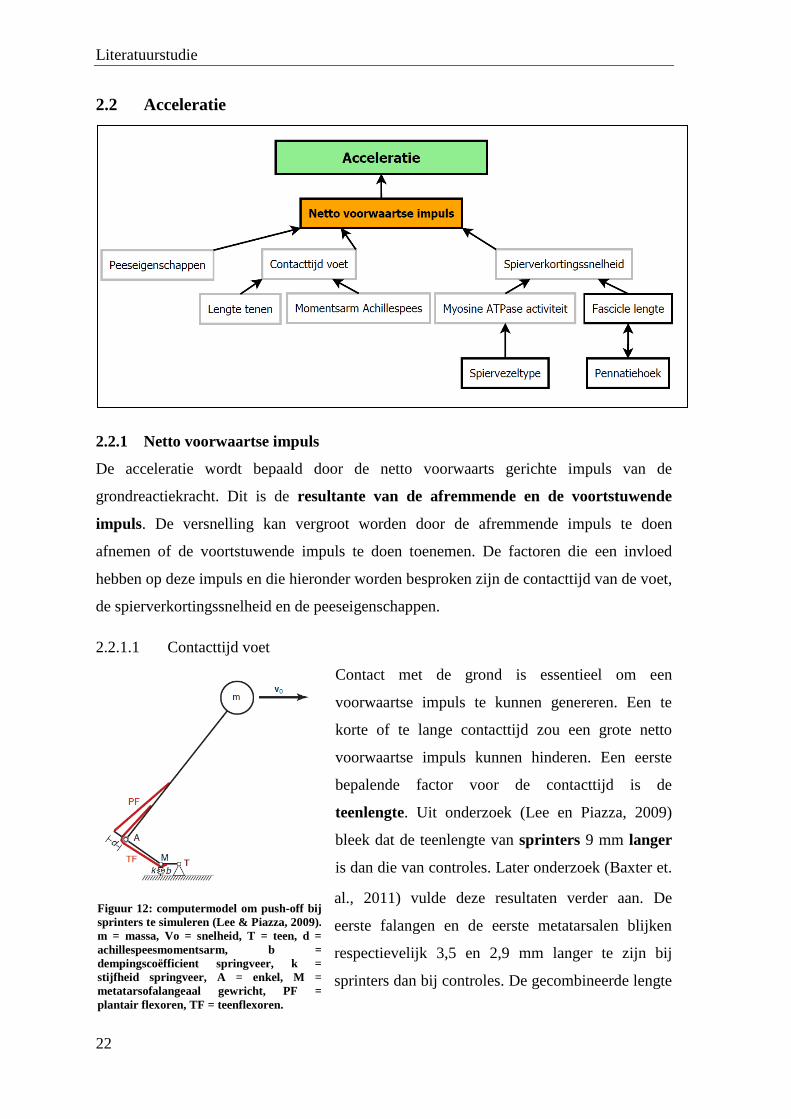
2.2.1 Netto voorwaartse impuls
De acceleratie wordt bepaald door de netto voorwaarts gerichte impuls van de
grondreactiekracht. Dit is de resultante van de afremmende en de voortstuwende
impuls. De versnelling kan vergroot worden door de afremmende impuls te doen
afnemen of de voortstuwende impuls te doen toenemen. De factoren die een invloed
hebben op deze impuls en die hieronder worden besproken zijn de contacttijd van de voet,
de spierverkortingssnelheid en de peeseigenschappen.
2.2.1.1 Contacttijd voet
Contact met de grond is essentieel om een
voorwaartse impuls te kunnen genereren. Een te
korte of te lange contacttijd zou een grote netto
voorwaartse impuls kunnen hinderen. Een eerste
bepalende factor voor de contacttijd is de
teenlengte. Uit onderzoek (Lee en Piazza, 2009)
bleek dat de teenlengte van sprinters 9 mm langer
is dan die van controles. Later onderzoek (Baxter et.
al., 2011) vulde deze resultaten verder aan. De
eerste falangen en de eerste metatarsalen blijken
respectievelijk 3,5 en 2,9 mm langer te zijn bij
sprinters dan bij controles. De gecombineerde lengte
Figuur 12: computermodel om push-off bij
sprinters te simuleren (Lee & Piazza, 2009).
m = massa, Vo = snelheid, T = teen, d =
achillespeesmomentsarm, b =
dempingscoëfficient springveer, k =
stijfheid springveer, A = enkel, M =
metatarsofalangeaal gewricht, PF =
plantair flexoren, TF = teenflexoren.
Literatuurstudie
23
van falangen en metatarsalen was eveneens langer bij sprinters.
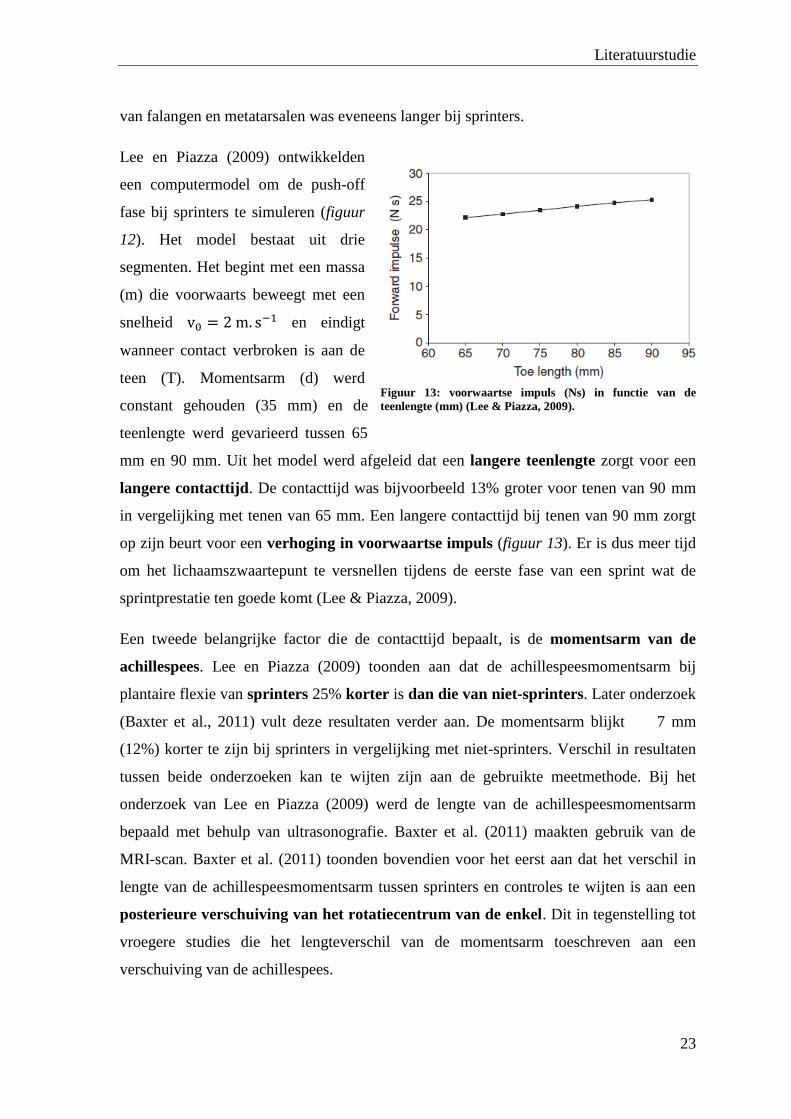
Lee en Piazza (2009) ontwikkelden
een computermodel om de push-off
fase bij sprinters te simuleren (figuur
12). Het model bestaat uit drie
segmenten. Het begint met een massa
(m) die voorwaarts beweegt met een
snelheid en eindigt
wanneer contact verbroken is aan de
teen (T). Momentsarm (d) werd
constant gehouden (35 mm) en de
teenlengte werd gevarieerd tussen 65
mm en 90 mm. Uit het model werd afgeleid dat een langere teenlengte zorgt voor een
langere contacttijd. De contacttijd was bijvoorbeeld 13% groter voor tenen van 90 mm
in vergelijking met tenen van 65 mm. Een langere contacttijd bij tenen van 90 mm zorgt
op zijn beurt voor een verhoging in voorwaartse impuls (figuur 13). Er is dus meer tijd
om het lichaamszwaartepunt te versnellen tijdens de eerste fase van een sprint wat de
sprintprestatie ten goede komt (Lee & Piazza, 2009).
Een tweede belangrijke factor die de contacttijd bepaalt, is de momentsarm van de
achillespees. Lee en Piazza (2009) toonden aan dat de achillespeesmomentsarm bij
plantaire flexie van sprinters 25% korter is dan die van niet-sprinters. Later onderzoek
(Baxter et al., 2011) vult deze resultaten verder aan. De momentsarm blijkt 7 mm
(12%) korter te zijn bij sprinters in vergelijking met niet-sprinters. Verschil in resultaten
tussen beide onderzoeken kan te wijten zijn aan de gebruikte meetmethode. Bij het
onderzoek van Lee en Piazza (2009) werd de lengte van de achillespeesmomentsarm
bepaald met behulp van ultrasonografie. Baxter et al. (2011) maakten gebruik van de
MRI-scan. Baxter et al. (2011) toonden bovendien voor het eerst aan dat het verschil in
lengte van de achillespeesmomentsarm tussen sprinters en controles te wijten is aan een
posterieure verschuiving van het rotatiecentrum van de enkel. Dit in tegenstelling tot
vroegere studies die het lengteverschil van de momentsarm toeschreven aan een
verschuiving van de achillespees.
Figuur 13: voorwaartse impuls (Ns) in functie van de
teenlengte (mm) (Lee & Piazza, 2009).
Literatuurstudie
24
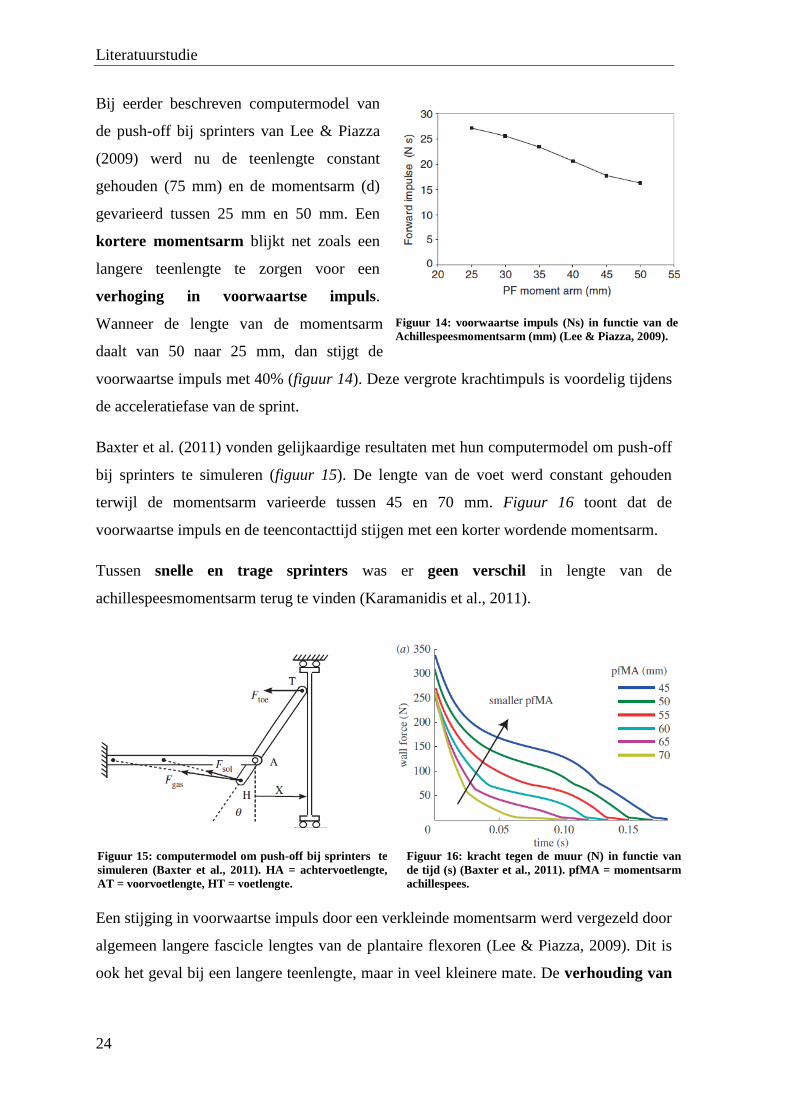
Bij eerder beschreven computermodel van
de push-off bij sprinters van Lee & Piazza
(2009) werd nu de teenlengte constant
gehouden (75 mm) en de momentsarm (d)
gevarieerd tussen 25 mm en 50 mm. Een
kortere momentsarm blijkt net zoals een
langere teenlengte te zorgen voor een
verhoging in voorwaartse impuls.
Wanneer de lengte van de momentsarm
daalt van 50 naar 25 mm, dan stijgt de
voorwaartse impuls met 40% (figuur 14). Deze vergrote krachtimpuls is voordelig tijdens
de acceleratiefase van de sprint.
Baxter et al. (2011) vonden gelijkaardige resultaten met hun computermodel om push-off
bij sprinters te simuleren (figuur 15). De lengte van de voet werd constant gehouden
terwijl de momentsarm varieerde tussen 45 en 70 mm. Figuur 16 toont dat de
voorwaartse impuls en de teencontacttijd stijgen met een korter wordende momentsarm.
Tussen snelle en trage sprinters was er geen verschil in lengte van de
achillespeesmomentsarm terug te vinden (Karamanidis et al., 2011).
Een stijging in voorwaartse impuls door een verkleinde momentsarm werd vergezeld door
algemeen langere fascicle lengtes van de plantaire flexoren (Lee & Piazza, 2009). Dit is
ook het geval bij een langere teenlengte, maar in veel kleinere mate. De verhouding van
Figuur 14: voorwaartse impuls (Ns) in functie van de
Achillespeesmomentsarm (mm) (Lee & Piazza, 2009).
Figuur 15: computermodel om push-off bij sprinters te
simuleren (Baxter et al., 2011). HA = achtervoetlengte,
AT = voorvoetlengte, HT = voetlengte.
Figuur 16: kracht tegen de muur (N) in functie van
de tijd (s) (Baxter et al., 2011). pfMA = momentsarm
achillespees.

Literatuurstudie
25
de fascicle lengte tot de achillespeesmomentsarm is 50% groter bij sprinters dan bij
controles.
2.2.1.2 Spierverkortingssnelheid
De netto voorwaartse impuls die kan geleverd worden in een bepaalde tijdspanne hangt af
van de spierverkortingssnelheid. Doordat de contacttijd bij sprinten zeer kort is, is een
hoge spierverkortingssnelheid essentieel. De spierverkortingssnelheid wordt bepaald
door biochemische (myosine ATPase activiteit die beïnvloed wordt door het
spiervezeltype en door training) en architecturale karakteristieken (vezellengte; het
aantal sarcomeren in serie) (Kumagai et al., 2000).
a) Biochemische karakteristieken
Een eerste belangrijk element voor het bereiken van een hoge spierverkortingssnelheid is
het beschikken over snelle spiervezels, ook wel fast twitch of type 2 vezels genaamd.
Onderzoek uit de jaren 70 toonde aan dat sprinters meer FT vezels in hun beenspieren
hebben. Spiervezeltype werd hier bepaald aan de hand van een invasieve methode (Saltin
et al., 1977; Wahren et al., 1971; Nygaard & Goricke, 1976; Gollnick et al., 1972; Costill
et al., 1976). Recent onderzoek (Baguet et al., 2011) bevestigde deze resultaten met
behulp van een niet-invasieve estimatiemethode, de proton magnetische resonantie
spectroscopie (H-MRS). De studie toonde aan dat de spiercarnosine-inhoud 30% hoger
is bij explosieve atleten zoals sprinters dan bij de controlegroep. Alle 100 tot 400 m
lopers hadden een hogere spiercarnosine-inhoud dan het populatiegemiddelde.
Als we sprinters vergelijken met duurlopers blijkt dat sprinters (100 – 400 m)
gemiddeld een 1,9 keer hogere carnosine-inhoud hebben dan marathonlopers en triatleten
(p < 0.001) (Baguet et al., 2011). Een punt dat dikwijls over het hoofd gezien wordt, is
dat de variabiliteit in vezelsamenstelling bij iedere subgroep van atleten verschillend is.
Atleten kunnen dus succesvol zijn in uithoudingsactiviteiten, ook al bevatten hun spieren
een relatief hoog percentage type 2 vezels. Het is belangrijk om hier te benadrukken dat
type 2 vezels hoog oxidatief kunnen zijn en dat met uithoudingstraining het oxidatieve
potentieel kan verhoogd worden tot een punt waar het dat van de type 1 vezels van
sedentairen benadert (Henriksson & Reitman, 1976).

Literatuurstudie
26
b) Architecturale karakteristieken
Naast het spiervezeltype wordt de spierverkortingssnelheid ook nog bepaald door
architecturale eigenschappen zoals de fascicle lengte en de pennatiehoek van een spier.
Ten eerste blijkt uit onderzoek dat de fascicle lengte van de beenspieren (M. vastus
lateralis, M. gastrocnemius lateralis, M. gastrocnemius medialis) groter is bij elite
sprinters dan bij elite duurlopers (Abe et al., 2000; Abe et al., 2001; Lee & Piazza, 2009).
In een studie (Kumagai et al., 2000) waar sprinters werden opgesplitst in trage (11.00 –
11.70 s) en snelle (10.00 – 10.90 s) sprinters, werden voor alle onderzochte spieren
langere fascicle lengtes gevonden voor de snelle sprinters. Deze correleerden significant
met de beste prestatie op de 100 m sprint.
Er kan dus geconcludeerd worden dat
een langere fascicle lengte geassocieerd
kan worden met een betere
sprintprestatie. Er zijn twee
mogelijkheden waarbij een langere
fascicle lengte leidt tot een betere
sprintprestatie. Ten eerste zorgen
langere fascicles voor een grotere
maximale verkortingssnelheid. Het
gevolg hiervan is een grotere kracht die
zou bijdragen aan de sprintprestatie. Een
tweede mogelijkheid is dat een grotere
fascicle lengte ervoor zorgt dat er een
grotere krachtoutput is aan een identieke verkortingssnelheid doordat de sarcomeren
aan een lagere snelheid kunnen werken. Dit laatste kan worden uitgelegd aan de hand
van figuur 17. Voor een gegeven peesuitrekking aan een verkortingssnelheid van 1 cm
per seconde, zou fascicle (B) zijn individuele sarcomeer moeten verkorten met 1.0 µm,
terwijl fascicle (A) de verkortingsafstand tussen de sarcomeren zou kunnen verdelen
waardoor iedere sarcomeer slechts 0.5 µm zou moeten verkorten. De fysiologische
implicatie hiervan is dat fascicle (A) zou kunnen verkorten aan een lagere snelheid (0.5
µm/sec) dan fascicle (B) (1 µm/sec) en zo meer kracht kan leveren op basis van de
kracht-snelheidscurve (Abe et al., 2001).
Figuur 17: het effect van een langere fascicle lengte op de
spierverkortingssnelheid en de kracht-snelheidscurve (Abe
et al., 2001).

Literatuurstudie
27
De redenen voor de verschillen in fascicle lengte tussen sprinters en duurlopers zijn
onduidelijk, maar er bestaan verschillende verklaringen (Abe et al., 2000). Als eerste is er
de genetische aanleg. Het is mogelijk dat individuen geboren worden met kortere of
langere fascicles en dat men op basis daarvan de “juiste” sport koos. Een andere
mogelijkheid is dat de verschillen in fascicle lengte te wijten zijn aan training. Bewijzen
voor deze laatste verklaring zijn enkel nog maar gevonden in dierenstudies. Er moet nog
verder onderzoek gedaan worden om de mogelijkheid en de rol van fascicleverlenging of
-verkorting als een adaptatie op looptraining volledig te begrijpen.
Ten tweede blijkt uit de studie van Abe et al. (2000) dat de pennatiehoek van sprinters
significant kleiner is dan die van duurlopers voor M. vastus lateralis en M.
gastrocnemius medialis. Dit blijkt niet het geval te zijn voor de M. gastrocnemius
lateralis. In de studie waar snelle sprinters vergeleken werden met trage sprinters
(Kumagai et al., 2000) vond men kleinere pennatiehoeken bij de snelle sprinters. Uit
deze studies kan geconcludeerd worden dat een kleine pennatiehoek voordeel oplevert bij
het sprinten.
2.2.1.3 Peeseigenschappen
Ook de peeseigenschappen hebben een invloed op de netto voorwaartse impuls. In een
studie van Kubo et al. (2011) werden 100 meter sprinters vergeleken met een
controlegroep wat betreft hun peeseigenschappen. Concreet werd de verlenging van de
peesstructuren bepaald in knie-extensoren bij isometrische knie-extensie en in plantaire
flexoren bij plantaire flexie. Ook werd de dikte van de patellapees (op 50%, gemeten
vanaf proximaal) en achillespees (30 mm proximaal van de calcaneus) gemeten.
Verlenging van de peesstructuren bij een krachtproductie hoger dan 400N was
significant groter bij sprinters dan bij controles voor de knie-extensoren. Bij de
plantaire flexoren was er geen verschil merkbaar. Maximale verlenging van de pezen
was significant groter bij sprinters voor knie-extensoren, maar niet voor plantaire
flexoren. De stijfheid van de peesstructuren was lager bij sprinters voor knie-
extensoren, maar niet voor plantaire flexoren. Er was geen significant verschil in
spierdikte tussen sprinters en controles voor zowel de patellapees als de achillespees.
In overeenstemming met vorige onderzoeken werd gevonden dat de peesstructuren van de
knie-extensoren soepeler zijn bij snelle sprinters dan bij trage sprinters en ongetrainden.
Meer soepele peesstructuren zouden helpen om hogere krachten in de standfase te

Literatuurstudie
28
ontwikkelen. Kubo et al. (2000) en Stafilidis en Arampatzis (2007) vonden eerder al een
significante relatie tussen peeseigenschappen in de knie-extensoren en de prestatie op
de 100 m sprint.
2.3 Maximale snelheid en aanhouden van de snelheid
Wanneer de sprinter zijn maximale snelheid heeft bereikt, tracht hij deze zo lang mogelijk
aan te houden. Om dit vol te houden maakt de atleet onder andere gebruik van het
fosfageensysteem. In de literatuur werd gevonden dat het fosfaatmetabolisme,
verantwoordelijk voor korte maximale inspanningen van 10 seconden of minder, beter
ontwikkeld zou zijn bij sprinters, mits toepassing van de juiste, specifieke trainingen
(Ross & Leveritt, 2001). Wanneer werd gekeken naar de snelheid van afbraak en
heropbouw van PCr, dan bleek dat sprinters PCr sneller afbreken dan afstandsatleten,
maar dat de resynthese bij duurlopers sneller verloopt dan bij sprinters (Crowther et al.,
2002).
De glycolytische ATP-productie, die zowel voorkomt bij de anaerobe als aerobe
glycolyse, is zowel voor sprinters als voor duurlopers van groot belang. Uit de studie van
Costill et al. (1976) blijkt dat de glycolytische enzymatische activiteit bij sprinters dubbel
zo groot is als bij duurlopers. De functionele betekenis van deze verhoging is echter nog
niet aangetoond. Dikwijls werden verhogingen van glycolytische enzymen vastgesteld
zonder een verbetering van de sprintprestatie (Jacobs et al., 1987).
Specifiek voor het aerobe energiesysteem blijkt dat duurlopers een 50% hogere
oxidatieve capaciteit hebben dan sprinters (Crowther et al., 2002). Toch zou sprinttraining
ook kunnen zorgen voor de stijging van bepaalde enzymen van het aerobe
energiesysteem, maar daarvoor zouden sprints van relatief langere afstand nodig zijn
(Ross & Leveritt, 2001). Verder onderzoek is echter vereist om de precieze aard van de
aerobe adaptatie te achterhalen.

Literatuurstudie
29
DEEL 3: HYPOTHESEN EN ONDERZOEKSVRAGEN
Er is al veel onderzoek verricht naar de prestatiebepalende factoren voor atleten en meer
specifiek voor sprinters en duurlopers. Het onderzoek van deze scriptie tracht de huidige
resultaten aan te vullen en mogelijke hiaten op te lossen. In deze studie worden sprinters
en duurlopers van verschillende niveaus (Belgische top en subtop) onderzocht. De
meetmethodes die worden gebruikt, zijn enkele antropometrische metingen waarbij
vetpercentages, lengte- en omtrekmaten worden bepaald en ultrasonografie waarbij de
focus ligt op het bepalen van fascicle lengte, pennatiehoek en spierdikte. Vervolgens
wordt ook gebruik gemaakt van de NMR-scanner voor de bepaling van spiervolumes,
spiermassamiddelpunten, botlengtes en spiervezeltype.
Het doel van ons onderzoek is het maken van een vergelijkende studie tussen duurlopers
en sprinters. Daarbij wordt getracht antagonistische karakteristieken aan te tonen. Er
wordt gekeken of de door ons onderzochte items uit de rode draad significant verschillen
tussen duurlopers en sprinters. Op antropometrisch vlak verwachten we enkele duidelijke
verschillen tussen duurlopers en sprinters. Hierbij stellen we de volgende hypothesen.
Een eerste hypothese stelt dat de omtrek van onderbeen en bovenbeen groter zal zijn bij
sprinters dan bij duurlopers. Ten tweede verwachten we verschillen in beenlengte en
lichaamslengte tussen beide groepen atleten.
Vervolgens stellen we enkele hypothesen met betrekking tot spierarchitecturale
verschillen. Fascicle lengte van M. gastrocnemius medialis en lateralis en M. vastus
lateralis bij sprinters is groter dan bij duurlopers terwijl de pennatiehoek van deze spieren
kleiner is bij sprinters dan bij duurlopers. Daarnaast is de spierdikte van M.
gastrocnemius medialis en lateralis groter bij sprinters dan bij duurlopers. Spierdikte van
M. rectus femoris, M. vastus medialis en lateralis is eveneens groter bij sprinters behalve
op 70% anterieure dij. Deze verschillen worden duidelijker naarmate het niveau van de
atleten hoger is. Vervolgens is het spiervolume van M. soleus, M. gastrocnemius medialis
en lateralis, M. rectus femoris en M. vastus medialis en lateralis groter bij sprinters dan
bij duurlopers. Fysiologische dwarsdoorsnede van M. gastrocnemius medialis en lateralis
en M. vastus lateralis is ook groter bij sprinters dan bij duurlopers. Als voorlaatste ligt het
spiermassamiddelpunt van M. soleus, M. gastrocnemius medialis en lateralis, M. rectus

Literatuurstudie
30
femoris en M. vastus medialis en lateralis hoger bij duurlopers dan bij sprinters. Tenslotte
hebben duurlopers meer type 1 vezels in vergelijking met sprinters.
Onze onderzoeksvraag is tweeledig. Ten eerste willen we onze schema’s voor de
duurloop- en sprintprestatie justifiëren. Hierbij willen we de correctheid nagaan van de
prestatiebepalende factoren en hun onderlinge relaties. Ten tweede willen we nagaan of
we met de door ons onderzochte factoren een atleet eerder kunnen classificeren als een
typische duurloper of eerder als een typische sprinter.

31
II. METHODIEK

Methodiek
32
1. Vergelijkende studie
1.1 Proefpersonen
Om een vergelijkende studie te kunnen uitvoeren, werden duurlopers en sprinters van
Belgisch subtop- en topniveau gerekruteerd. Uiteindelijk bestond de onderzoeksgroep uit
13 mannelijke atleten, waarvan 7 duurlopers en 6 sprinters.
Alle atleten moesten voorafgaand aan het onderzoek een vragenlijst invullen (bijlage 1).
De leeftijd, discipline en persoonlijke records werden onder andere opgevraagd. Hieruit
bleek dat de duurlopers een gemiddelde leeftijd hadden van 22,71 (± 3,77) jaar. Hun beste
tijd voor de 3000 m bevond zich tussen 8 min 05’ 06’’ en 8 min 53’ 20’’. De gemiddelde
leeftijd van de onderzochte sprinters was 21,83 (± 4,71) jaar. Ze liepen de 60 m tussen
6,79 s en 7,07 s en de 100 m tussen 10,71 s en 10,99 s. Bij het bevragen van de inname
van voedingssupplementen en het voedingspatroon bleek dat niemand de afgelopen drie
maanden beta-alanine of carnosine innam als voedingssupplement en geen enkele atleet
vegetariër was.
Bij het rekruteren van de proefpersonen werden ziekte, blessure of een dopingverleden als
exclusiecriteria gehanteerd.
1.2 Studiedesign
Ons onderzoek bestond uit een observationele cross-sectionele studie om verschillen na
te gaan tussen sprinters en duurlopers ter hoogte van het onderste lidmaat. Hierbij werd
iedere atleet eenmaal onderzocht in de periode van januari tot en met maart. Het
onderzoek nam ongeveer één uur en een kwartier in beslag en was drieledig. De studie
werd goedgekeurd door het Ethisch Comité van het Universitair Ziekenhuis te Gent. Voor
de start van de testen werd aan de proefpersonen het testverloop uitgelegd en werd hen
gevraagd een informed consent te ondertekenen.
Het eerste deel van het onderzoek bestond uit een aantal antropometrische metingen.
Het gewicht van de proefpersonen werd genoteerd, net als hun lichaamslengte,
voetlengtes en enkele andere lengtematen, omtrekken en vetmetingen zoals weergegeven
in tabel 1. De vetmetingen werden uitgevoerd met een huidplooicaliper. Voor al deze
metingen bevonden de proefpersonen zich in de anatomische houding.

Methodiek
33
Tabel 1: antropometrische metingen: lengtematen rechterbeen, omtrekken rechterbeen en vetmetingen.
LENGTEMATEN RECHTERBEEN
Onderbeenlengte Afstand van de malleolus lateralis tot de kop van de tibia
Bovenbeenlengte Afstand van de laterale condyl tot de trochanter major van de
femur
Beenlengte Afstand van de trochanter major tot op de grond
OMTREKKEN RECHTERBEEN
Onderbeen Zone 20% Zone 30% Maximale
onderbeenomtrek
Bovenbeen Zone 30% Zone 50% Zone 70%
VETMETINGEN
Biceps Triceps Subscapulair Supra-iliacaal Kuit
Een volgend deel van het onderzoek bestond uit
het bepalen van de spierdikte, pennatiehoek en
fascicle lengte op een aantal anatomische
plaatsen ter hoogte van de kuit en de quadriceps
met behulp van ultrasonografie. Voorafgaand
aan de echo werden in totaal 13 meetplaatsen
aangeduid op het rechterbeen terwijl de
proefpersoon zich in anatomische houding
bevond zoals weergegeven op figuur 18. Om de
exacte meetplaatsen op het onderbeen en
bovenbeen te bepalen, moest de proefpersoon
respectievelijk zijn kuit en quadriceps opspannen. Op deze manier werden de spierbuiken
van M. gastrocnemius medialis en lateralis, M. rectus femoris en M. vastus medialis en
lateralis zichtbaar en konden de meetplaatsen aangeduid worden. Er moest telkens gel
aangebracht worden op de transducent van het ultrasoundtoestel. Daarna werd de
transducent op de meetplaatsen geplaatst en lichtjes bewogen (zonder te duwen) tot er een
goed beeld verscheen op de computer. Dit beeld werd voor alle meetplaatsen opgeslagen
om dan in een latere fase spierdikte, pennatiehoek en fascicle lengte te bepalen.
Figuur 18: 13 meetplaatsen voor
ultrasonografie (gebaseerd op Kubo et al.,
2011).

Methodiek
34
Tenslotte moesten de
proefpersonen twee maal 20
minuten in de NMR-scanner
liggen (figuur 19). Vooraleer
ze de scanruimte mochten
betreden, moesten de atleten
zich ontdoen van voorwerpen
die metaal bevatten zoals een
horloge, broeksriem, gsm …
Vervolgens moest men in
ruglig op de scantafel gaan
liggen met hun hoofd weg van
de scanner. Eén of beide benen werden geïmmobiliseerd in een spoel. Het hoofd werd
ondersteund door een kussen en de proefpersonen kregen een koptelefoon en oordopjes
tegen de geluidshinder. De atleten moesten zo stil mogelijk liggen. Tijdens een eerste
scan werd de imagingtechniek (MRI) toegepast om spiervolumes in kaart te brengen van
M. soleus, M. gastrocnemius medialis en lateralis, M. rectus femoris en M. vastus
medialis en lateralis van beide benen. Ook botlengtes, spiervolumes en
spiermassamiddelpunten konden met deze techniek bepaald worden. Tijdens deze scan
werd een perifere beenspoel gebruikt die het volledige onderlichaam omvat. Tussen beide
scans moesten de proefpersonen even uit de scanner zodat de spoelen konden verwisseld
worden. Vervolgens werd met behulp van proton magnetic resonance spectroscopy (H-
MRS) het spiervezeltype bepaald aan de hand van de gemeten carnosineconcentratie in de
M. soleus en de M. gastrocnemius. Hierbij werd een kuitspoel gebruikt die enkel het
rechter onderbeen omvat.
2. Meetinstrumenten
2.1 Huidplooicaliper
De vetmetingen gebeurden met behulp van een
huidplooicaliper (Harpenden Skinfold Caliper) zoals
weergegeven op figuur 20. Er werd gemeten ter hoogte
van de biceps, de triceps, de kuit, supra-iliacaal en
Figuur 19: proefpersoon in NMR scanner met perifere beenspoel.
Figuur 20: huidplooicaliper

Methodiek
35
subscapulair. Hieruit werd vervolgens het totale vetpercentage berekend met behulp van
de tabel van Durnin en Womersley.
2.2 Ultrasonografie
In dit onderzoek werd een draagbaar
echosysteem gebruikt, namelijk de
Telemed Echo Blaster 128Z (figuur 21).
De werking van dit toestel berust op
ultrasoon geluid, kortweg ultrageluid.
Dit geluid heeft een zeer hoge frequentie
waardoor het voor de mens niet
waarneembaar is. Via een transducent
wordt het geluid in het lichaam
gebracht. Tussen de transducent en de
huid moet een gel worden aangebracht
omdat de geluidsgolven niet door een luchtspleet kunnen. De transducent zendt aan een
heel hoge snelheid geluidsgolven in het lichaam. Afhankelijk van de tijd die het signaal
nodig heeft om de transducent opnieuw te
bereiken en de sterkte waarmee ze deze
bereikt zal een beeld worden gevormd. De
door het lichaam gereflecteerde
geluidsgolven worden opgevangen en
omgezet in elektrische wisselspanning. Deze
elektrische echosignalen kunnen door een
computersysteem (scan converter) worden
omgezet in videobeelden (figuur 22). Het
voordeel van de echografie is dat er een goed beeld kan geschetst worden van organen die
uit zacht weefsel bestaan, zoals spieren. Deze bevatten immers geen lucht. Een nadeel is
dat ultrageluiden met een frequentie hoger dan 1 MHz bijna volledig door lucht worden
weerkaatst. Hierdoor is alles dat zich achter een luchtlaag bevindt onzichtbaar. Een ander
nadeel is dat ultrageluiden ook niet door bot kunnen dringen door de sterke reflectie tegen
de bovenzijde.
Figuur 22: afbeelding ultrasonografie.
Figuur 21: draagbaar echosysteem: Telemed Echo Blaster
128Z.

Methodiek
36
2.3 NMR
Aan de hand van de 3 Tesla whole body NMR scanner (Siemens Trio Erlangen) werden
in deze studie twee zaken bepaald. Enerzijds werd het onderlichaam aan de hand van
MRI (proton magnetic resonance imaging) in beeld gebracht om op deze manier zaken
als spiervolumes en botlengtes te berekenen. MRI gebruikt magnetische resonantie om
een beeld te vormen van het lichaam.
Waterstofatomen hebben slechts één proton in de kern. Deze protonen hebben zeer
specifieke magnetische eigenschappen. Zo kan een proton met een externe magneet mee-
of tegenwerken. Wanneer een magnetisch veld wordt ingeschakeld zal het proton zich
richten naar het magnetisch veld. Deze fase wordt ook wel de “spin-up” genoemd (figuur
23).
Onder invloed van fotonen die door de radiofrequentiespoel (RF-spoel) worden
uitgezonden kan de energietoestand veranderen. Op deze manier kan het proton naar een
“spin-down” toestand gebracht worden waarbij het proton tegen het magnetisch veld
wordt gericht (figuur 24).
.
Wanneer de stralingen van de RF-spoel worden uitgezet, zullen de protonen terugvallen
naar de begintoestand. Bij deze terugval komt de opgenomen energie vrij en wordt er een
foton uitgezonden (figuur 25).
Figuur 23: spin-up fase
Figuur 24: spin-down fase

Methodiek
37
Door deze fotonen te meten, kan bepaald worden op welke plaats hoeveel
waterstofkernen zitten. Al deze metingen kunnen via de computer verwerkt worden tot
een driedimensionaal beeld dat het waterstofgehalte van de weefsels aangeeft. Doordat
ieder weefsel een eigen specifieke waterstofdichtheid bezit, kunnen anatomische details
worden waargenomen.
Als tweede werd met behulp van H-MRS (proton
magnetic resonance spectroscopy) de
carnosineconcentratie bepaald. Hiervoor werd in
twee spieren (de M. soleus en de M. gastrocnemius)
een voxel geplaatst zoals weergegeven op figuur 26,
op een plaats met zo weinig mogelijk vetweefsel. De
voxelgrootte bij de duurlopers varieerde tussen 11,2
en 14,4 g in de M. soleus en tussen 12 en 14,4 g in de
M. gastrocnemius. Bij de sprinters was de
voxelgrootte in beide spieren telkens 14,4 g. De voxel werd gescand met en zonder
wateronderdrukking. Het ontvangen signaal
werd in een computer ingevoerd en de
beelden en waarden werden op het scherm
geprojecteerd. De wateronderdrukking is
noodzakelijk om een beeld te krijgen van
de C2-H en C4-H-imidazole pieken van
carnosine (figuur 27). Shimming van het
signaal was nodig om het magnetisch veld
homogeen te maken zodat de pieken zo
smal en duidelijk mogelijk waren. De ‘best
Figuur 25: terugval van het proton met
opgenomen energie die vrijkomt.
Figuur 26: plaatsing voxel
Figuur 27: C2-H en C4-H-imidazole pieken van
carnosine.

Methodiek
38
shimm’ varieerde voor de duurlopers tussen 23,2 Hz en 31,7 Hz en voor de sprinters
tussen 24 Hz en 27,6 Hz.
Er werd gebruik gemaakt van twee radiofrequentievolumespoelen. Deze spoelen
garanderen een homogeen RF veld binnen de spoel. In de spoel ligt het te onderzoeken
lichaamsdeel. In deze studie werd zoals reeds vermeld, gebruik gemaakt van de kuitspoel
en de perifere beenspoel.
3. Analyse
3.1 Data-analyse
3.1.1 Ultrasonografie
De opgeslagen beelden die verkregen werden bij de ultrasound metingen op de 13
meetplaatsen van het rechterbeen konden geanalyseerd worden met Echomaster 3.0.
Ten eerste kon de spierdikte
bepaald worden. Hiervoor
werden de bovenste en
onderste aponeurose
aangeduid met een lijn.
Daarna werden drie verticale
lijnen getrokken tussen beide
aponeurosen; één links op het
beeld (meer distaal), één in
het midden en één rechts
(meer proximaal). Spierdiktes
werden verkregen door het
meten van de lengte van deze
lijnen (figuur 28).
Vervolgens kon de pennatiehoek gemeten worden. Hiervoor werden opnieuw de
aponeurosen gebruikt. Daarnaast werd een lijn getrokken evenwijdig met de
spiervezelrichting (figuur 28). Via het commando ‘angle’ berekende het programma de
hoek tussen de bovenste aponeurose en de spiervezelrichting en tussen de onderste
aponeurose en de spiervezelrichting. Daarna werd het gemiddelde van deze twee hoeken
Figuur: 28: beeldverwerking ultrasonografie M. vastus lateralis, zone
30%. 1: bovenste aponeurose; 2: onderste aponeurose; 3, 4 en 5:
spierdiktes; 6: aanduiding spiervezelrichting.
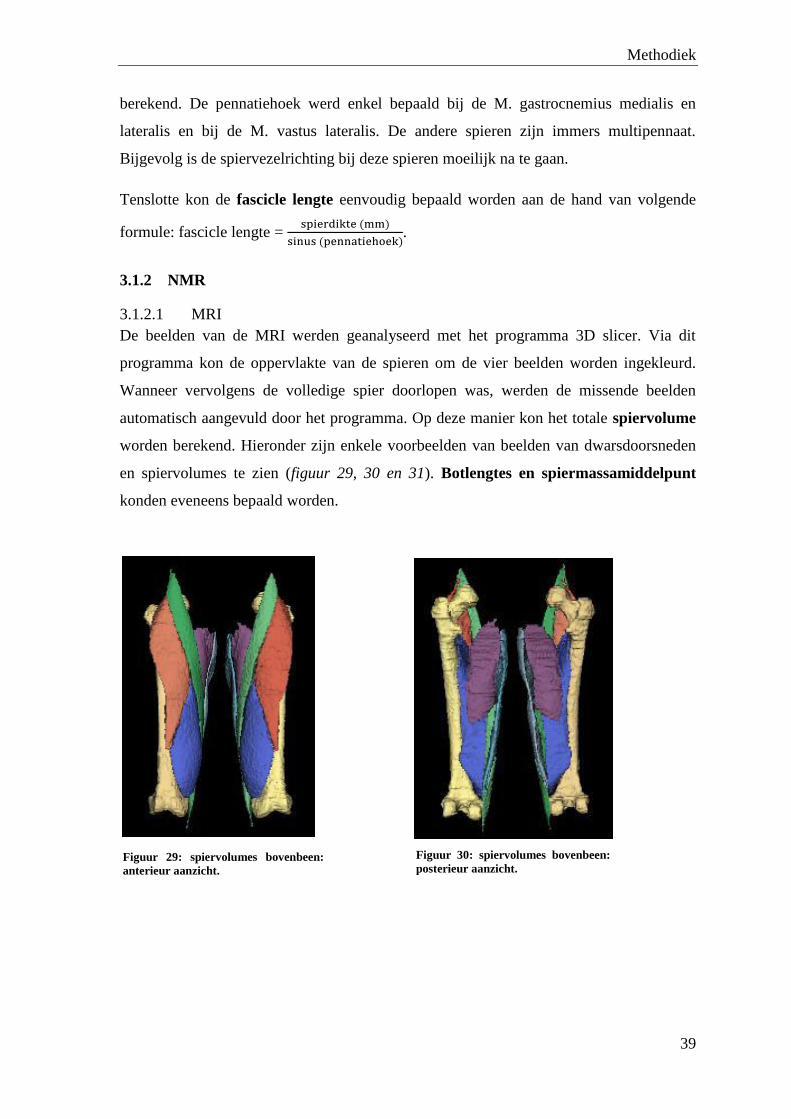
Methodiek
39
berekend. De pennatiehoek werd enkel bepaald bij de M. gastrocnemius medialis en
lateralis en bij de M. vastus lateralis. De andere spieren zijn immers multipennaat.
Bijgevolg is de spiervezelrichting bij deze spieren moeilijk na te gaan.
Tenslotte kon de fascicle lengte eenvoudig bepaald worden aan de hand van volgende
formule: fascicle lengte =
.
3.1.2 NMR
3.1.2.1 MRI
De beelden van de MRI werden geanalyseerd met het programma 3D slicer. Via dit
programma kon de oppervlakte van de spieren om de vier beelden worden ingekleurd.
Wanneer vervolgens de volledige spier doorlopen was, werden de missende beelden
automatisch aangevuld door het programma. Op deze manier kon het totale spiervolume
worden berekend. Hieronder zijn enkele voorbeelden van beelden van dwarsdoorsneden
en spiervolumes te zien (figuur 29, 30 en 31). Botlengtes en spiermassamiddelpunt
konden eveneens bepaald worden.
Figuur 30: spiervolumes bovenbeen:
posterieur aanzicht.
Figuur 29: spiervolumes bovenbeen:
anterieur aanzicht.
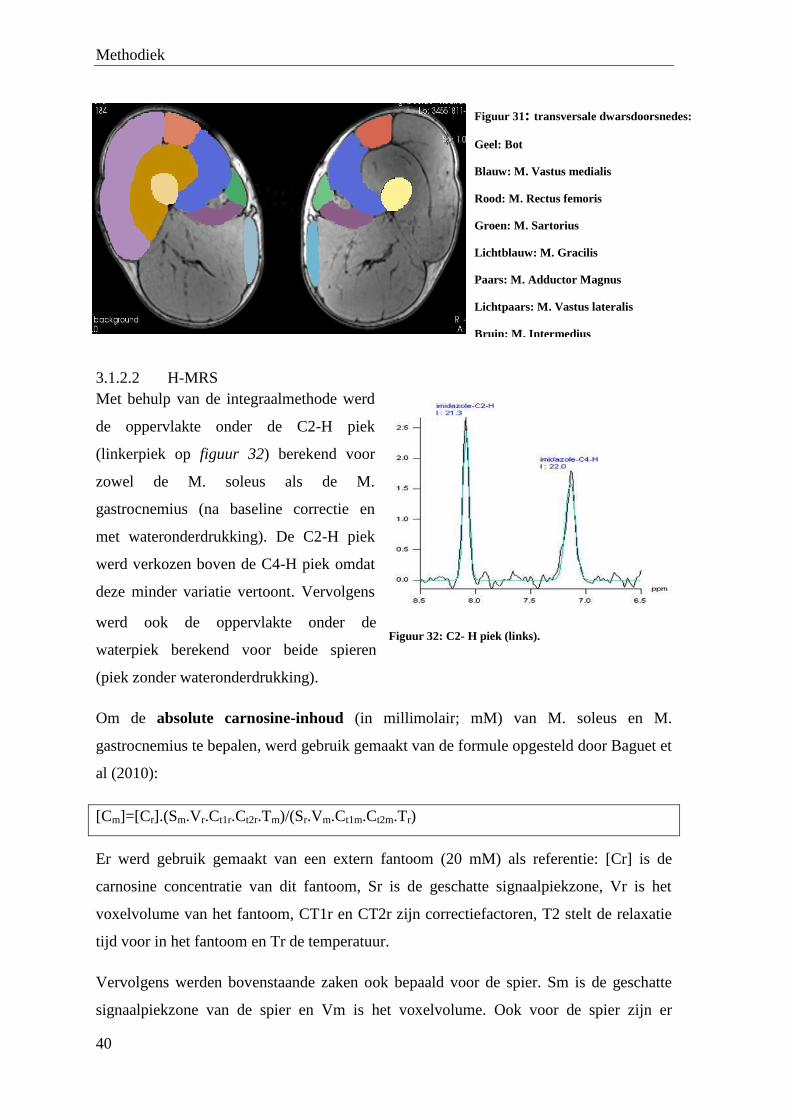
Methodiek
40
3.1.2.2 H-MRS
Met behulp van de integraalmethode werd
de oppervlakte onder de C2-H piek
(linkerpiek op figuur 32) berekend voor
zowel de M. soleus als de M.
gastrocnemius (na baseline correctie en
met wateronderdrukking). De C2-H piek
werd verkozen boven de C4-H piek omdat
deze minder variatie vertoont. Vervolgens
werd ook de oppervlakte onder de
waterpiek berekend voor beide spieren
(piek zonder wateronderdrukking).
Om de absolute carnosine-inhoud (in millimolair; mM) van M. soleus en M.
gastrocnemius te bepalen, werd gebruik gemaakt van de formule opgesteld door Baguet et
al (2010):
[Cm]=[Cr].(Sm.Vr.Ct1r.Ct2r.Tm)/(Sr.Vm.Ct1m.Ct2m.Tr)
Er werd gebruik gemaakt van een extern fantoom (20 mM) als referentie: [Cr] is de
carnosine concentratie van dit fantoom, Sr is de geschatte signaalpiekzone, Vr is het
voxelvolume van het fantoom, CT1r en CT2r zijn correctiefactoren, T2 stelt de relaxatie
tijd voor in het fantoom en Tr de temperatuur.
Vervolgens werden bovenstaande zaken ook bepaald voor de spier. Sm is de geschatte
signaalpiekzone van de spier en Vm is het voxelvolume. Ook voor de spier zijn er
Figuur 32: C2- H piek (links).
Figuur 31: transversale dwarsdoorsnedes:
Geel: Bot
Blauw: M. Vastus medialis
Rood: M. Rectus femoris
Groen: M. Sartorius
Lichtblauw: M. Gracilis
Paars: M. Adductor Magnus
Lichtpaars: M. Vastus lateralis
Bruin: M. Intermedius

Methodiek
41
correctiefactoren: CT1m en CT2m. T1 is de relaxatie tijd in de spier en Tm stelt de
temperatuur van de spier voor.
Wanneer al bovenstaande zaken ingevuld werden in de formule, werd de absolute
carnosineconentratie [Cm] gevonden.
3.2 Statistische analyse
De statistische analyse gebeurde aan de hand van het programma SPSS Statistics 21.
Voor de verwerking van de resultaten van zowel de antropometrische metingen, de
ultrasonografiebeelden en de gegevens verkregen met de NMR-scanner (MRI en H-MRS)
werd gebruik gemaakt van twee statistische technieken.
Ten eerste werd de Independent-Samples T-test gebruikt om gemiddelden te vergelijken
tussen twee groepen, namelijk duurlopers en sprinters. In dit geval is er dus één
kwantitatieve variabele (vb. lichaamsmassa) waarvoor het gemiddelde berekend wordt in
de ene populatie (duurlopers) en ook in de andere populatie (sprinters). Nadien worden
beide gemiddelden met elkaar vergeleken. Indien de p-waarde significant is (p < 0,05),
wil dit zeggen dat er een significant verschil bestaat tussen beide groepen. Het
gemiddelde van de ene groep is dus duidelijk hoger dan dat van de andere groep.
Als tweede statistische techniek werd de Pearson correlatie uitgevoerd om het verband
te berekenen tussen twee kwantitatieve variabelen (vb. verband tussen fascicle lengte en
looptijd). Een correlatiecoëfficiënt bevindt zich steeds tussen -1 en +1. Een grotere
absolute waarde wijst op een groter verband tussen de twee variabelen. Het verband kan
positief zijn (vb. hoe groter de fascicle lengte, hoe langer de looptijd) of negatief (vb. hoe
groter de fascicle lengte, hoe korter de looptijd). Dit positief of negatief zijn wordt
aangeduid door het teken van de correlatiecoëfficiënt. Het verband tussen de twee
variabelen is significant indien de p-waarde kleiner is dan 0,05.

42

43
III. RESULTATEN

Resultaten
44
1. Inleiding
Bij het beschrijven van de resultaten worden de structuur en de schema’s van de rode
draad niet gevolgd. De resultaten worden weergegeven per meetinstrument om het
overzicht te bewaren.
In deze studie werden duurlopers (n = 7) vergeleken met sprinters (n = 6). De gemiddelde
leeftijd bij de duurlopers bedroeg 22,71 (± 3,77) jaar. De sprinters hadden een
gemiddelde leeftijd van 21,83 (± 4,71) jaar. Er was geen significant verschil in leeftijd
tussen beide groepen.
De International Association of Athletics Federations ontwierp een puntensysteem (IAAF
scoring tables of athletics) waarmee prestaties van atleten van verschillende disciplines
met elkaar kunnen vergeleken worden. Iedere prestatie van een bepaald atletiekevent
krijgt een bepaalde score toegekend; een hogere score duidt op een betere prestatie. De
onderzochte duurlopers scoorden met hun tijden voor de 3000 m tussen de 767 en 1027
punten. Gemiddeld hadden ze een puntenwaarde van 894,43 (± 93,65). Sprinters scoorden
voor hun prestatie op de 60 m tussen de 876 en 970 punten met een gemiddelde van
920,83 punten (± 30,44). Er waren geen uitbijters onder de puntenwaarden van de
individuele atleten. Tussen de gemiddelde scores van duurlopers en sprinters was geen
significant verschil terug te vinden.
2. Antropometrische metingen
Uit tabel 2 blijkt dat de gemiddelde massa niet verschilde tussen beide groepen. BMI en
RPI verschilden echter wel. De duurlopers hadden een BMI van 20,43 (± 1,75) die
significant lager (p = 0,013) was dan die van de sprinters (23,06 ± 1,39). De RPI van de
duurlopers lag significant hoger (p = 0,003) dan de RPI van de sprinters (respectievelijk
44,71 ± 0,86 en 42,55 ± 1,19). Wanneer de vetpercentages werden vergeleken tussen
beide groepen werden geen significante verschillen vastgesteld. Dit was het geval voor
zowel het totale vetpercentage als voor de metingen op de verschillende plaatsen.
Resultaten
45
Tabel 2: massa, BMI, RPI en vetpercentage bij duurlopers en sprinters.
Duurlopers Sprinters
Massa (kg) 67,53 ± 10,15 71,75 ± 3,55
BMI 20,43 ± 1,75* 23,06 ± 1,39*
RPI 44,71 ± 0,86* 42,55 ± 1,19*
Totale vetpercentage (%) 8,24 ± 1,68 9,32 ± 1,20
* = p<0,05; duurlopers vs. sprinters
Bij de analyse van de lengtematen (tabel 3) werd geen verschil in lichaamslengte
vastgesteld tussen beide groepen. De totale beenlengte en onderbeenlengte verschilden
eveneens niet. Bij de bovenbeenlengte werd een trend tot significantie (p = 0,097)
vastgesteld waarbij de duurlopers een grotere bovenbeenlengte hadden dan de sprinters
(respectievelijk 44,80 cm ± 2,21 en 42,60 cm ± 2,15) (grafiek 1). Wanneer de been-,
onderbeen- en bovenbeenlengte relatief werden bekeken ten opzichte van de
lichaamslengte werden geen significante verschillen tussen beide groepen gevonden. De
lengte van de linker- en rechtervoet verschilde niet tussen beide groepen.
Zowel bij duurlopers als bij sprinters werden geen significante correlaties teruggevonden
tussen één van de lengtematen en de beste prestatie.
Tabel 3: lichaamslengte en andere lengtematen ter hoogte van onderste lidmaat bij duurlopers en sprinters.
Lengte Duurlopers Sprinters
Lichaamslengte (cm) 181,34 ± 6,24 176,52 ± 5,14
Beenlengte (cm) 90,91 ± 4,56 88,08 ±4,31
Onderbeenlengte (cm) 40,10 ± 1,64 39,28 ± 2,05
Bovenbeenlengte (cm) 44,80 ± 2,21~ 42,60 ± 2,15~
Beenlengte/lichaamslengte 0,50 ± 0,01 0,50 ± 0,02
Onderbeenlengte/lichaamslengte 0,22 ± 0,003 0,22 ± 0,01
Bovenbeenlengte/lichaamslengte 0,25 ± 0,01 0,24 ± 0,01
Lengte rechtervoet (cm) 26,63 ± 1,27 26,40 ± 0,60
Lengte linkervoet (cm) 26,57 ± 1,11 26,20 ± 0,75
~ = p<0,10: duurlopers vs. sprinters
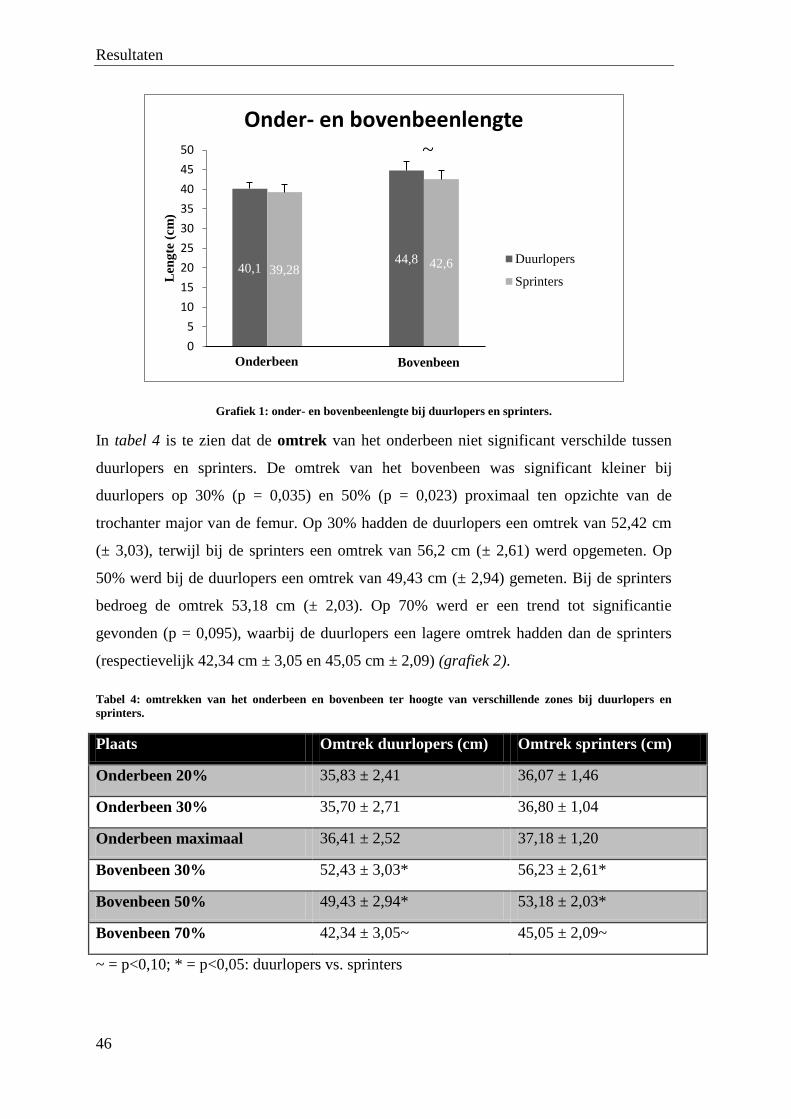
Resultaten
46
Grafiek 1: onder- en bovenbeenlengte bij duurlopers en sprinters.
In tabel 4 is te zien dat de omtrek van het onderbeen niet significant verschilde tussen
duurlopers en sprinters. De omtrek van het bovenbeen was significant kleiner bij
duurlopers op 30% (p = 0,035) en 50% (p = 0,023) proximaal ten opzichte van de
trochanter major van de femur. Op 30% hadden de duurlopers een omtrek van 52,42 cm
(± 3,03), terwijl bij de sprinters een omtrek van 56,2 cm (± 2,61) werd opgemeten. Op
50% werd bij de duurlopers een omtrek van 49,43 cm (± 2,94) gemeten. Bij de sprinters
bedroeg de omtrek 53,18 cm (± 2,03). Op 70% werd er een trend tot significantie
gevonden (p = 0,095), waarbij de duurlopers een lagere omtrek hadden dan de sprinters
(respectievelijk 42,34 cm ± 3,05 en 45,05 cm ± 2,09) (grafiek 2).
Tabel 4: omtrekken van het onderbeen en bovenbeen ter hoogte van verschillende zones bij duurlopers en
sprinters.
Plaats Omtrek duurlopers (cm) Omtrek sprinters (cm)
Onderbeen 20% 35,83 ± 2,41 36,07 ± 1,46
Onderbeen 30% 35,70 ± 2,71 36,80 ± 1,04
Onderbeen maximaal 36,41 ± 2,52 37,18 ± 1,20
Bovenbeen 30% 52,43 ± 3,03* 56,23 ± 2,61*
Bovenbeen 50% 49,43 ± 2,94* 53,18 ± 2,03*
Bovenbeen 70% 42,34 ± 3,05~ 45,05 ± 2,09~
~ = p<0,10; * = p<0,05: duurlopers vs. sprinters
40,1 44,8
39,28 42,6
0
5
10
15
20
25
30
35
40
45
50
Len
gte
(cm
)
Onder- en bovenbeenlengte
Duurlopers
Sprinters
Onderbeen Bovenbeen
~
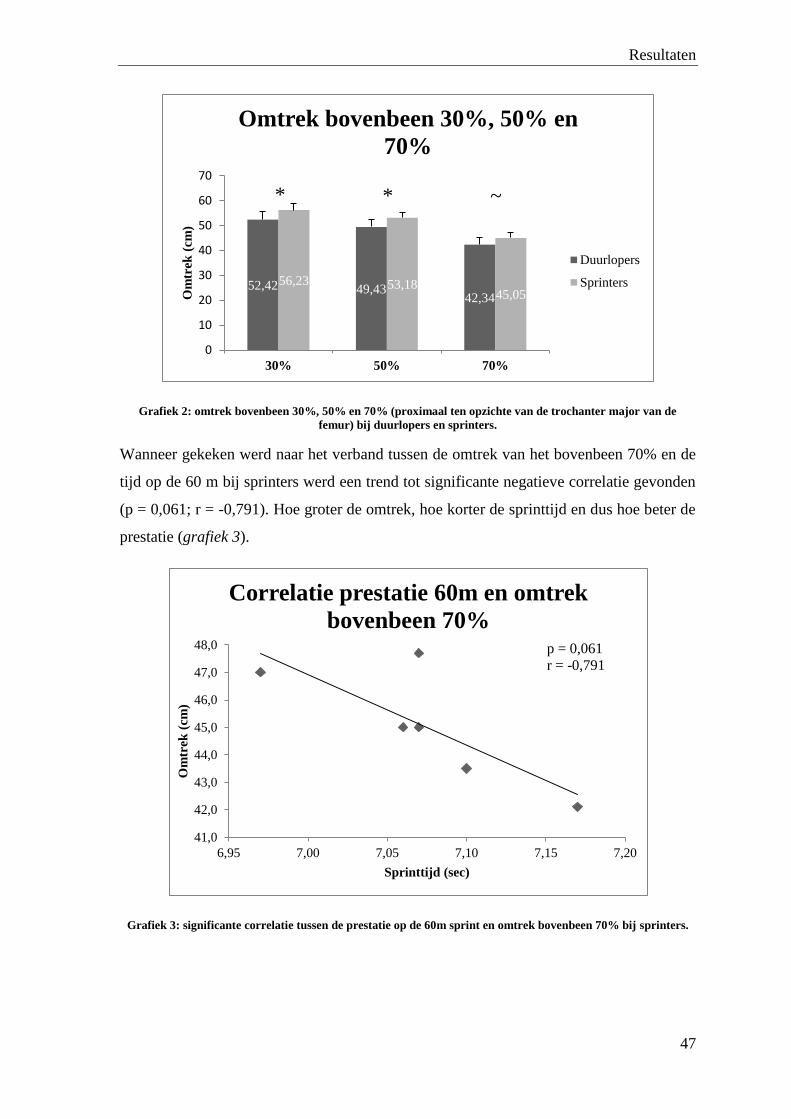
Resultaten
47
Grafiek 2: omtrek bovenbeen 30%, 50% en 70% (proximaal ten opzichte van de trochanter major van de
femur) bij duurlopers en sprinters.
Wanneer gekeken werd naar het verband tussen de omtrek van het bovenbeen 70% en de
tijd op de 60 m bij sprinters werd een trend tot significante negatieve correlatie gevonden
(p = 0,061; r = -0,791). Hoe groter de omtrek, hoe korter de sprinttijd en dus hoe beter de
prestatie (grafiek 3).
Grafiek 3: significante correlatie tussen de prestatie op de 60m sprint en omtrek bovenbeen 70% bij sprinters.
52,42 49,43 42,34
56,23 53,18 45,05
0
10
20
30
40
50
60
70
30% 50% 70%
Om
trek
(cm
)
Omtrek bovenbeen 30%, 50% en
70%
Duurlopers
Sprinters
* * ~
41,0
42,0
43,0
44,0
45,0
46,0
47,0
48,0
6,95 7,00 7,05 7,10 7,15 7,20
Om
trek
(cm
)
Sprinttijd (sec)
Correlatie prestatie 60m en omtrek
bovenbeen 70% p = 0,061
r = -0,791
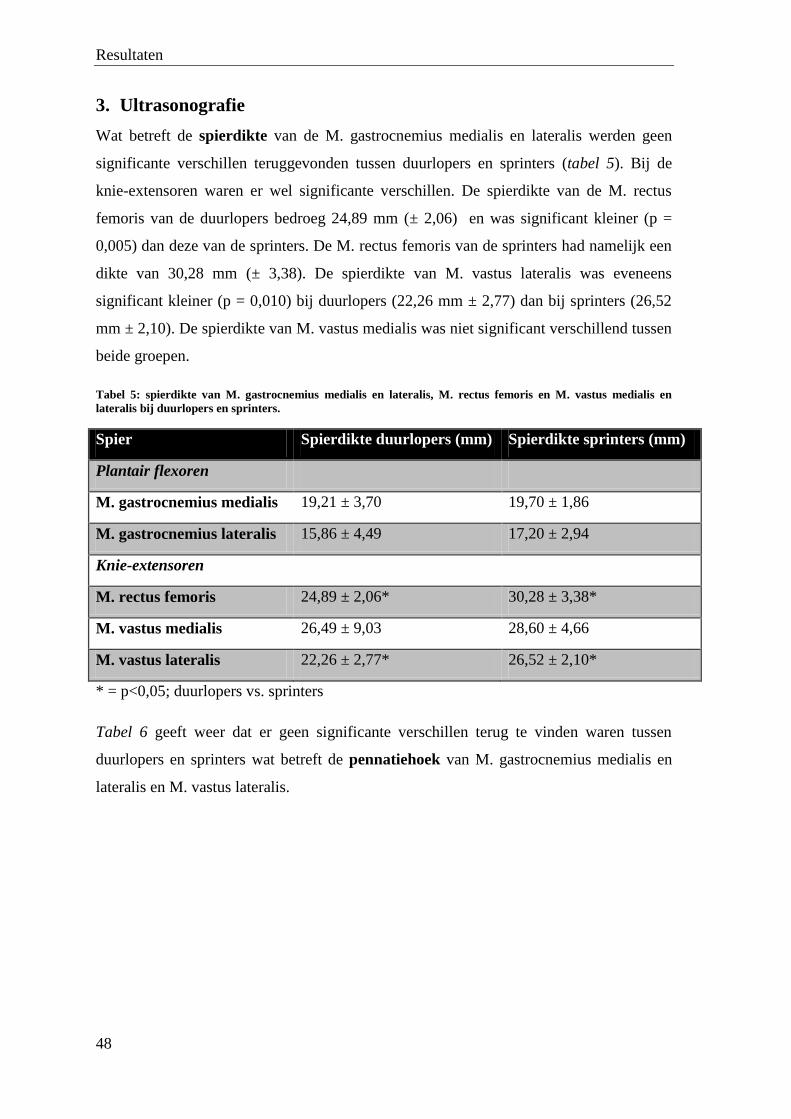
Resultaten
48
3. Ultrasonografie
Wat betreft de spierdikte van de M. gastrocnemius medialis en lateralis werden geen
significante verschillen teruggevonden tussen duurlopers en sprinters (tabel 5). Bij de
knie-extensoren waren er wel significante verschillen. De spierdikte van de M. rectus
femoris van de duurlopers bedroeg 24,89 mm (± 2,06) en was significant kleiner (p =
0,005) dan deze van de sprinters. De M. rectus femoris van de sprinters had namelijk een
dikte van 30,28 mm (± 3,38). De spierdikte van M. vastus lateralis was eveneens
significant kleiner (p = 0,010) bij duurlopers (22,26 mm ± 2,77) dan bij sprinters (26,52
mm ± 2,10). De spierdikte van M. vastus medialis was niet significant verschillend tussen
beide groepen.
Tabel 5: spierdikte van M. gastrocnemius medialis en lateralis, M. rectus femoris en M. vastus medialis en
lateralis bij duurlopers en sprinters.
Spier Spierdikte duurlopers (mm) Spierdikte sprinters (mm)
Plantair flexoren
M. gastrocnemius medialis 19,21 ± 3,70 19,70 ± 1,86
M. gastrocnemius lateralis 15,86 ± 4,49 17,20 ± 2,94
Knie-extensoren
M. rectus femoris 24,89 ± 2,06* 30,28 ± 3,38*
M. vastus medialis 26,49 ± 9,03 28,60 ± 4,66
M. vastus lateralis 22,26 ± 2,77* 26,52 ± 2,10*
* = p<0,05; duurlopers vs. sprinters
Tabel 6 geeft weer dat er geen significante verschillen terug te vinden waren tussen
duurlopers en sprinters wat betreft de pennatiehoek van M. gastrocnemius medialis en
lateralis en M. vastus lateralis.
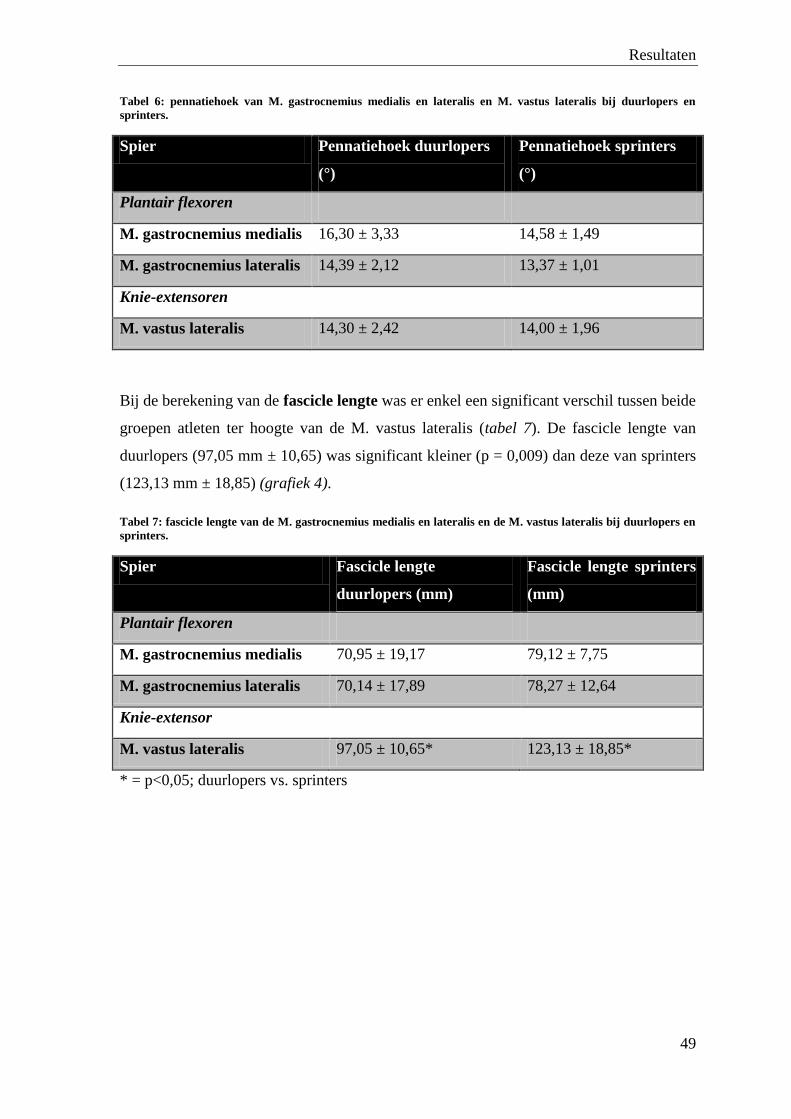
Resultaten
49
Tabel 6: pennatiehoek van M. gastrocnemius medialis en lateralis en M. vastus lateralis bij duurlopers en
sprinters.
Spier Pennatiehoek duurlopers
(°)
Pennatiehoek sprinters
(°)
Plantair flexoren
M. gastrocnemius medialis 16,30 ± 3,33 14,58 ± 1,49
M. gastrocnemius lateralis 14,39 ± 2,12 13,37 ± 1,01
Knie-extensoren
M. vastus lateralis 14,30 ± 2,42 14,00 ± 1,96
Bij de berekening van de fascicle lengte was er enkel een significant verschil tussen beide
groepen atleten ter hoogte van de M. vastus lateralis (tabel 7). De fascicle lengte van
duurlopers (97,05 mm ± 10,65) was significant kleiner (p = 0,009) dan deze van sprinters
(123,13 mm ± 18,85) (grafiek 4).
Tabel 7: fascicle lengte van de M. gastrocnemius medialis en lateralis en de M. vastus lateralis bij duurlopers en
sprinters.
Spier Fascicle lengte
duurlopers (mm)
Fascicle lengte sprinters
(mm)
Plantair flexoren
M. gastrocnemius medialis 70,95 ± 19,17 79,12 ± 7,75
M. gastrocnemius lateralis 70,14 ± 17,89 78,27 ± 12,64
Knie-extensor
M. vastus lateralis 97,05 ± 10,65* 123,13 ± 18,85*
* = p<0,05; duurlopers vs. sprinters
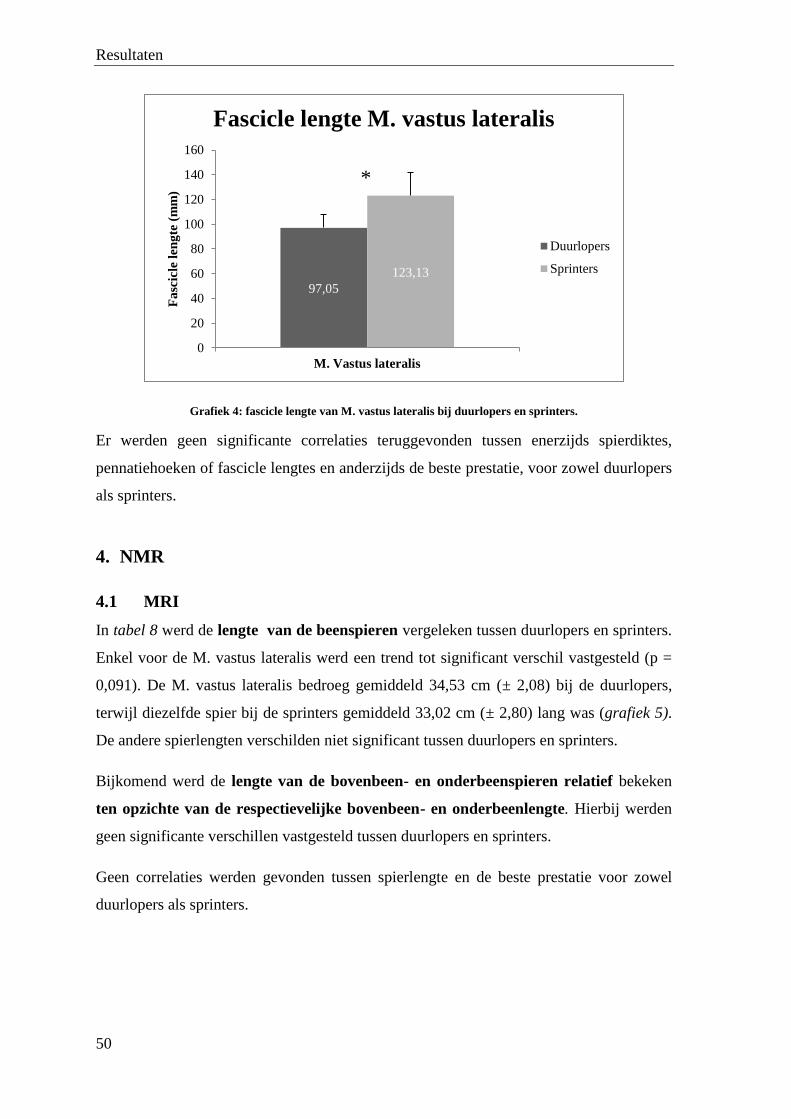
Resultaten
50
Grafiek 4: fascicle lengte van M. vastus lateralis bij duurlopers en sprinters.
Er werden geen significante correlaties teruggevonden tussen enerzijds spierdiktes,
pennatiehoeken of fascicle lengtes en anderzijds de beste prestatie, voor zowel duurlopers
als sprinters.
4. NMR
4.1 MRI
In tabel 8 werd de lengte van de beenspieren vergeleken tussen duurlopers en sprinters.
Enkel voor de M. vastus lateralis werd een trend tot significant verschil vastgesteld (p =
0,091). De M. vastus lateralis bedroeg gemiddeld 34,53 cm (± 2,08) bij de duurlopers,
terwijl diezelfde spier bij de sprinters gemiddeld 33,02 cm (± 2,80) lang was (grafiek 5).
De andere spierlengten verschilden niet significant tussen duurlopers en sprinters.
Bijkomend werd de lengte van de bovenbeen- en onderbeenspieren relatief bekeken
ten opzichte van de respectievelijke bovenbeen- en onderbeenlengte. Hierbij werden
geen significante verschillen vastgesteld tussen duurlopers en sprinters.
Geen correlaties werden gevonden tussen spierlengte en de beste prestatie voor zowel
duurlopers als sprinters.
97,05
123,13
0
20
40
60
80
100
120
140
160
M. Vastus lateralis
Fa
scic
le l
eng
te (
mm
)
Fascicle lengte M. vastus lateralis
Duurlopers
Sprinters
*
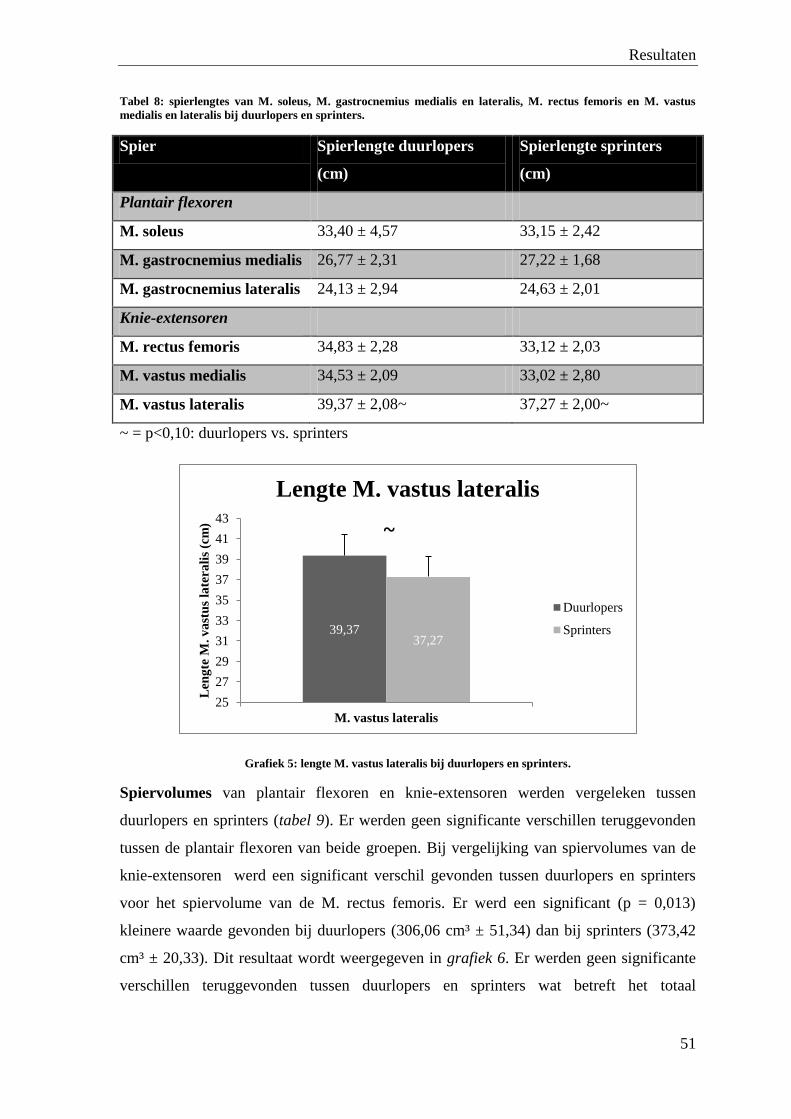
Resultaten
51
Tabel 8: spierlengtes van M. soleus, M. gastrocnemius medialis en lateralis, M. rectus femoris en M. vastus
medialis en lateralis bij duurlopers en sprinters.
Spier Spierlengte duurlopers
(cm)
Spierlengte sprinters
(cm)
Plantair flexoren
M. soleus 33,40 ± 4,57 33,15 ± 2,42
M. gastrocnemius medialis 26,77 ± 2,31 27,22 ± 1,68
M. gastrocnemius lateralis 24,13 ± 2,94 24,63 ± 2,01
Knie-extensoren
M. rectus femoris 34,83 ± 2,28 33,12 ± 2,03
M. vastus medialis 34,53 ± 2,09 33,02 ± 2,80
M. vastus lateralis 39,37 ± 2,08~ 37,27 ± 2,00~
~ = p<0,10: duurlopers vs. sprinters
Grafiek 5: lengte M. vastus lateralis bij duurlopers en sprinters.
Spiervolumes van plantair flexoren en knie-extensoren werden vergeleken tussen
duurlopers en sprinters (tabel 9). Er werden geen significante verschillen teruggevonden
tussen de plantair flexoren van beide groepen. Bij vergelijking van spiervolumes van de
knie-extensoren werd een significant verschil gevonden tussen duurlopers en sprinters
voor het spiervolume van de M. rectus femoris. Er werd een significant (p = 0,013)
kleinere waarde gevonden bij duurlopers (306,06 cm³ ± 51,34) dan bij sprinters (373,42
cm³ ± 20,33). Dit resultaat wordt weergegeven in grafiek 6. Er werden geen significante
verschillen teruggevonden tussen duurlopers en sprinters wat betreft het totaal
39,37 37,27
25
27
29
31
33
35
37
39
41
43
M. vastus lateralis
Len
gte
M. v
ast
us
late
ra
lis
(cm
)
Lengte M. vastus lateralis
Duurlopers
Sprinters
~
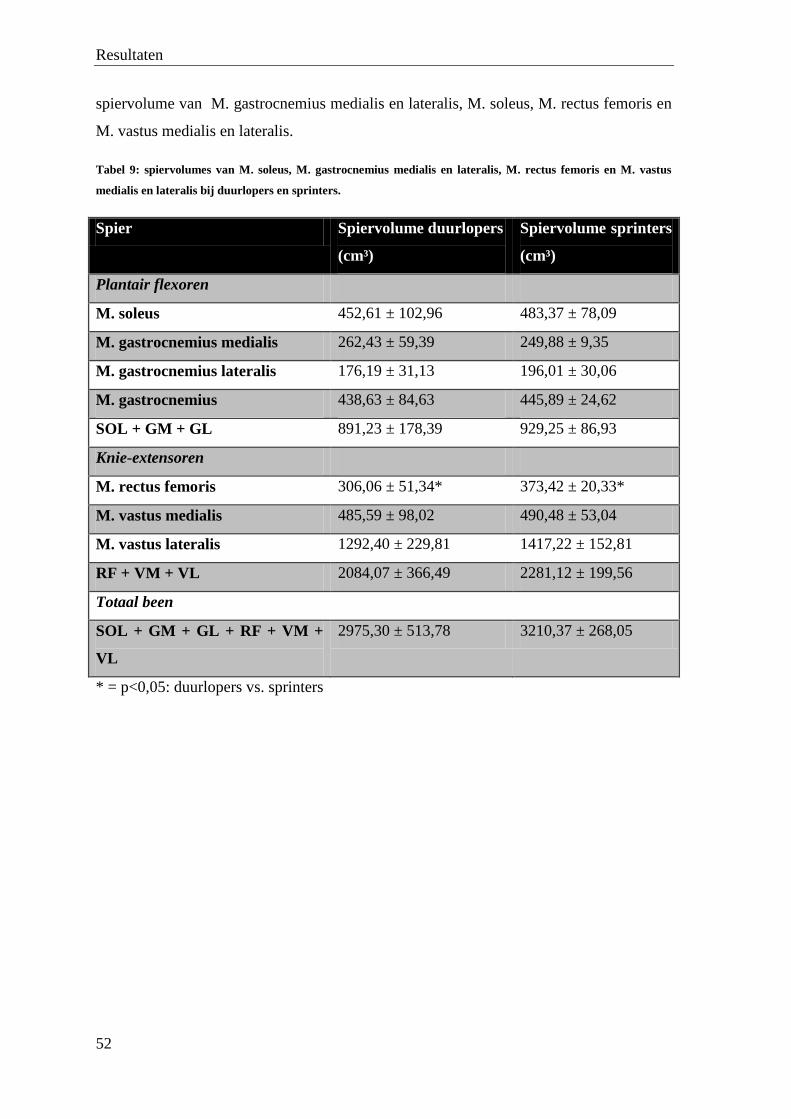
Resultaten
52
spiervolume van M. gastrocnemius medialis en lateralis, M. soleus, M. rectus femoris en
M. vastus medialis en lateralis.
Tabel 9: spiervolumes van M. soleus, M. gastrocnemius medialis en lateralis, M. rectus femoris en M. vastus
medialis en lateralis bij duurlopers en sprinters.
Spier Spiervolume duurlopers
(cm³)
Spiervolume sprinters
(cm³)
Plantair flexoren
M. soleus 452,61 ± 102,96 483,37 ± 78,09
M. gastrocnemius medialis 262,43 ± 59,39 249,88 ± 9,35
M. gastrocnemius lateralis 176,19 ± 31,13 196,01 ± 30,06
M. gastrocnemius 438,63 ± 84,63 445,89 ± 24,62
SOL + GM + GL 891,23 ± 178,39 929,25 ± 86,93
Knie-extensoren
M. rectus femoris 306,06 ± 51,34* 373,42 ± 20,33*
M. vastus medialis 485,59 ± 98,02 490,48 ± 53,04
M. vastus lateralis 1292,40 ± 229,81 1417,22 ± 152,81
RF + VM + VL 2084,07 ± 366,49 2281,12 ± 199,56
Totaal been
SOL + GM + GL + RF + VM +
VL
2975,30 ± 513,78 3210,37 ± 268,05
* = p<0,05: duurlopers vs. sprinters
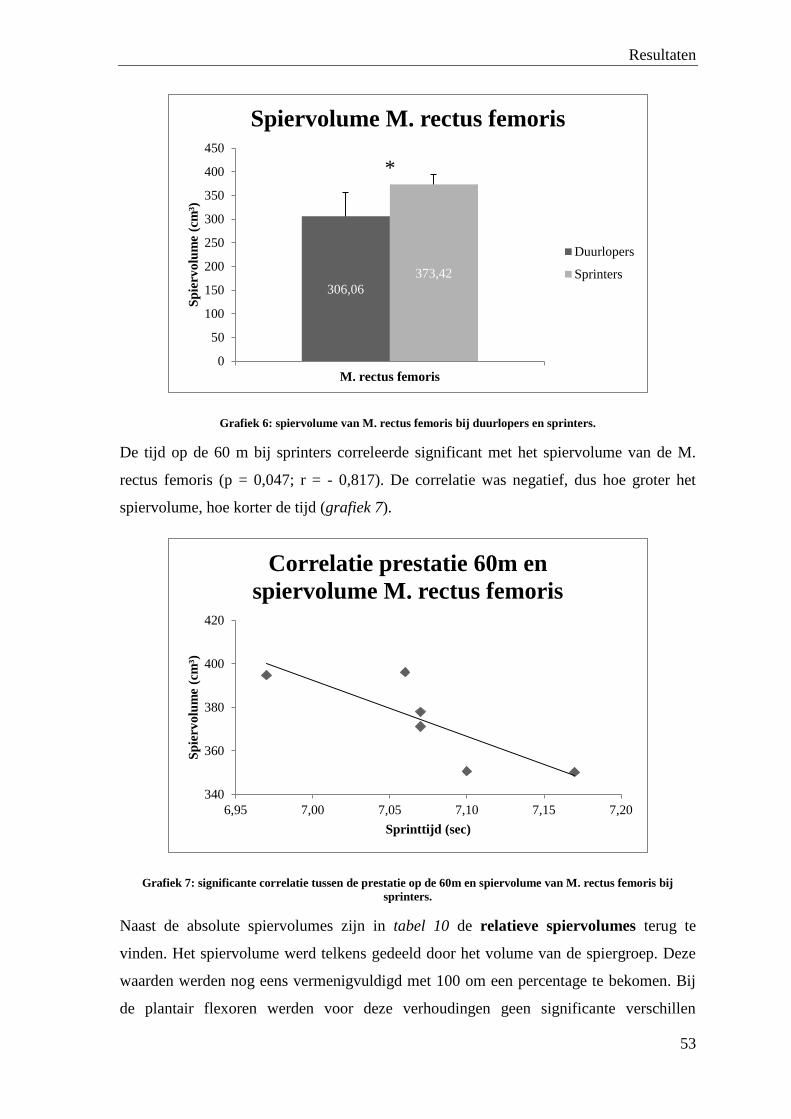
Resultaten
53
Grafiek 6: spiervolume van M. rectus femoris bij duurlopers en sprinters.
De tijd op de 60 m bij sprinters correleerde significant met het spiervolume van de M.
rectus femoris (p = 0,047; r = - 0,817). De correlatie was negatief, dus hoe groter het
spiervolume, hoe korter de tijd (grafiek 7).
Grafiek 7: significante correlatie tussen de prestatie op de 60m en spiervolume van M. rectus femoris bij
sprinters.
Naast de absolute spiervolumes zijn in tabel 10 de relatieve spiervolumes terug te
vinden. Het spiervolume werd telkens gedeeld door het volume van de spiergroep. Deze
waarden werden nog eens vermenigvuldigd met 100 om een percentage te bekomen. Bij
de plantair flexoren werden voor deze verhoudingen geen significante verschillen
306,06
373,42
0
50
100
150
200
250
300
350
400
450
M. rectus femoris
Sp
ierv
olu
me
(cm
³)
Spiervolume M. rectus femoris
Duurlopers
Sprinters
*
340
360
380
400
420
6,95 7,00 7,05 7,10 7,15 7,20
Sp
ierv
olu
me
(cm
³)
Sprinttijd (sec)
Correlatie prestatie 60m en
spiervolume M. rectus femoris
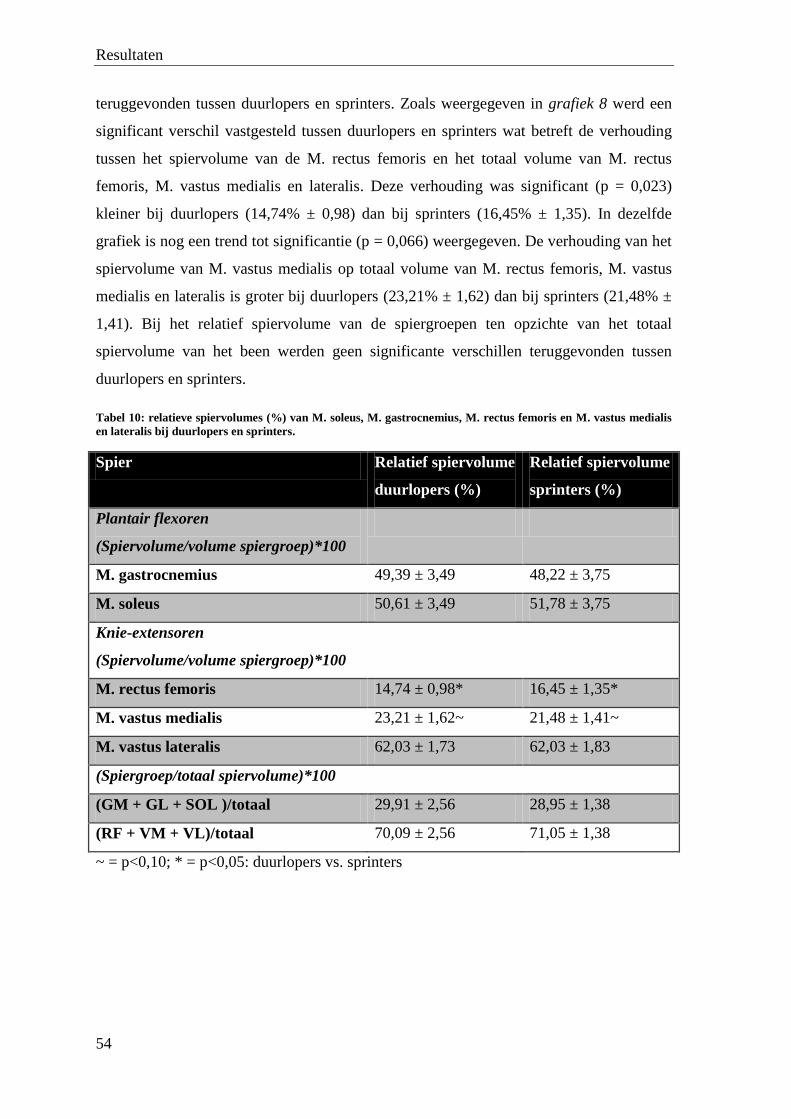
Resultaten
54
teruggevonden tussen duurlopers en sprinters. Zoals weergegeven in grafiek 8 werd een
significant verschil vastgesteld tussen duurlopers en sprinters wat betreft de verhouding
tussen het spiervolume van de M. rectus femoris en het totaal volume van M. rectus
femoris, M. vastus medialis en lateralis. Deze verhouding was significant (p = 0,023)
kleiner bij duurlopers (14,74% ± 0,98) dan bij sprinters (16,45% ± 1,35). In dezelfde
grafiek is nog een trend tot significantie (p = 0,066) weergegeven. De verhouding van het
spiervolume van M. vastus medialis op totaal volume van M. rectus femoris, M. vastus
medialis en lateralis is groter bij duurlopers (23,21% ± 1,62) dan bij sprinters (21,48% ±
1,41). Bij het relatief spiervolume van de spiergroepen ten opzichte van het totaal
spiervolume van het been werden geen significante verschillen teruggevonden tussen
duurlopers en sprinters.
Tabel 10: relatieve spiervolumes (%) van M. soleus, M. gastrocnemius, M. rectus femoris en M. vastus medialis
en lateralis bij duurlopers en sprinters.
Spier Relatief spiervolume
duurlopers (%)
Relatief spiervolume
sprinters (%)
Plantair flexoren
(Spiervolume/volume spiergroep)*100
M. gastrocnemius 49,39 ± 3,49 48,22 ± 3,75
M. soleus 50,61 ± 3,49 51,78 ± 3,75
Knie-extensoren
(Spiervolume/volume spiergroep)*100
M. rectus femoris 14,74 ± 0,98* 16,45 ± 1,35*
M. vastus medialis 23,21 ± 1,62~ 21,48 ± 1,41~
M. vastus lateralis 62,03 ± 1,73 62,03 ± 1,83
(Spiergroep/totaal spiervolume)*100
(GM + GL + SOL )/totaal 29,91 ± 2,56 28,95 ± 1,38
(RF + VM + VL)/totaal 70,09 ± 2,56 71,05 ± 1,38
~ = p<0,10; * = p<0,05: duurlopers vs. sprinters
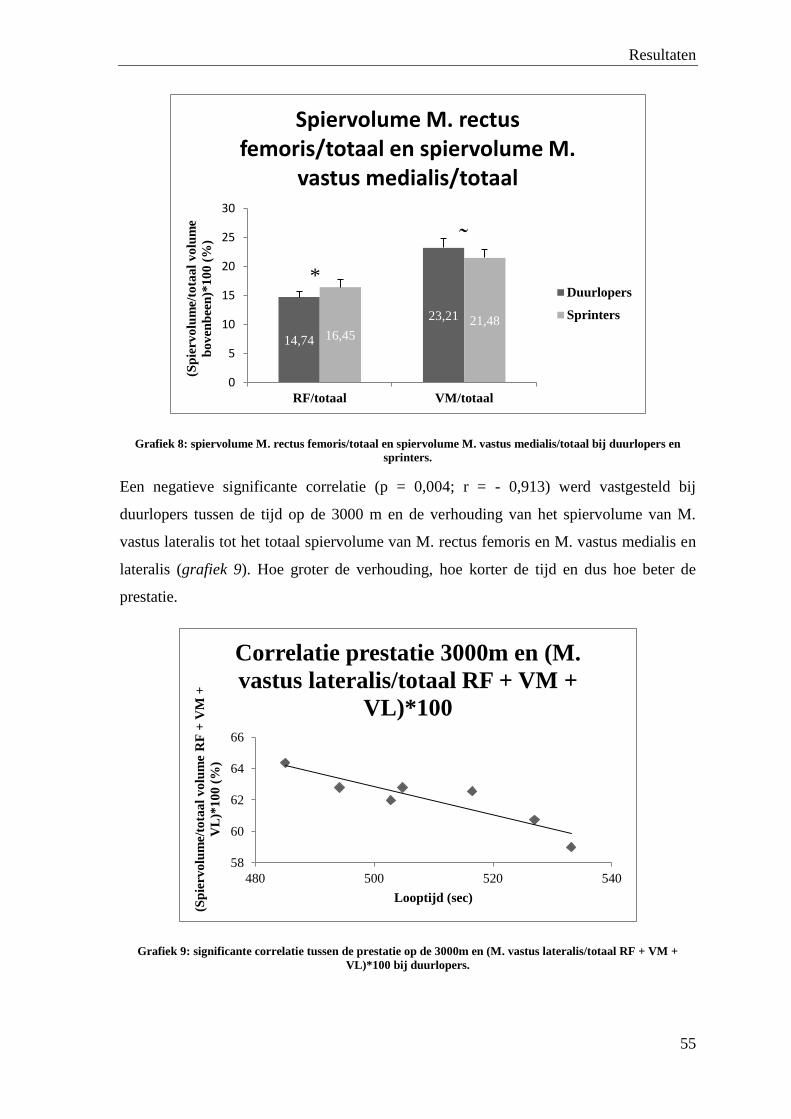
Resultaten
55
Grafiek 8: spiervolume M. rectus femoris/totaal en spiervolume M. vastus medialis/totaal bij duurlopers en
sprinters.
Een negatieve significante correlatie (p = 0,004; r = - 0,913) werd vastgesteld bij
duurlopers tussen de tijd op de 3000 m en de verhouding van het spiervolume van M.
vastus lateralis tot het totaal spiervolume van M. rectus femoris en M. vastus medialis en
lateralis (grafiek 9). Hoe groter de verhouding, hoe korter de tijd en dus hoe beter de
prestatie.
Grafiek 9: significante correlatie tussen de prestatie op de 3000m en (M. vastus lateralis/totaal RF + VM +
VL)*100 bij duurlopers.
14,74
23,21
16,45 21,48
0
5
10
15
20
25
30
RF/totaal VM/totaal
(Sp
ierv
olu
me/
tota
al
vo
lum
e
bo
ven
bee
n)*
10
0 (
%)
Spiervolume M. rectus femoris/totaal en spiervolume M.
vastus medialis/totaal
Duurlopers
Sprinters
*
˜
58
60
62
64
66
480 500 520 540
(Sp
ierv
olu
me/
tota
al
vo
lum
e R
F +
VM
+
VL
)*1
00
(%
)
Looptijd (sec)
Correlatie prestatie 3000m en (M.
vastus lateralis/totaal RF + VM +
VL)*100
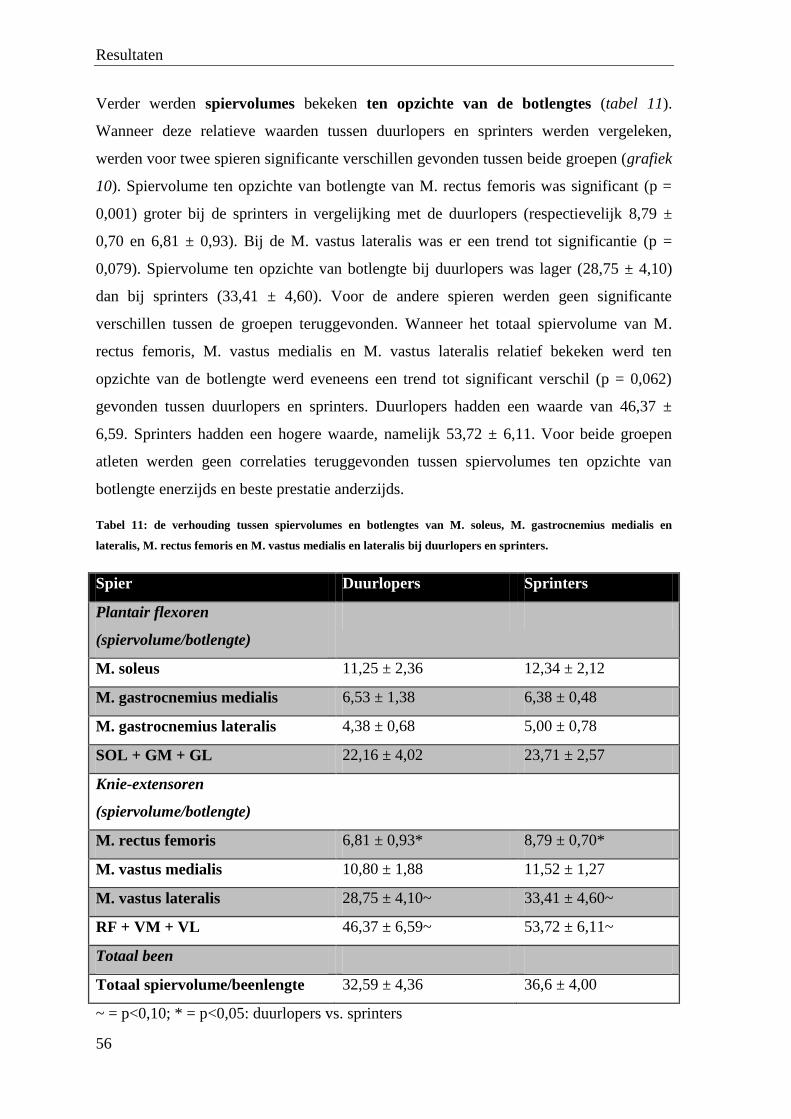
Resultaten
56
Verder werden spiervolumes bekeken ten opzichte van de botlengtes (tabel 11).
Wanneer deze relatieve waarden tussen duurlopers en sprinters werden vergeleken,
werden voor twee spieren significante verschillen gevonden tussen beide groepen (grafiek
10). Spiervolume ten opzichte van botlengte van M. rectus femoris was significant (p =
0,001) groter bij de sprinters in vergelijking met de duurlopers (respectievelijk 8,79 ±
0,70 en 6,81 ± 0,93). Bij de M. vastus lateralis was er een trend tot significantie (p =
0,079). Spiervolume ten opzichte van botlengte bij duurlopers was lager (28,75 ± 4,10)
dan bij sprinters (33,41 ± 4,60). Voor de andere spieren werden geen significante
verschillen tussen de groepen teruggevonden. Wanneer het totaal spiervolume van M.
rectus femoris, M. vastus medialis en M. vastus lateralis relatief bekeken werd ten
opzichte van de botlengte werd eveneens een trend tot significant verschil (p = 0,062)
gevonden tussen duurlopers en sprinters. Duurlopers hadden een waarde van 46,37 ±
6,59. Sprinters hadden een hogere waarde, namelijk 53,72 ± 6,11. Voor beide groepen
atleten werden geen correlaties teruggevonden tussen spiervolumes ten opzichte van
botlengte enerzijds en beste prestatie anderzijds.
Tabel 11: de verhouding tussen spiervolumes en botlengtes van M. soleus, M. gastrocnemius medialis en
lateralis, M. rectus femoris en M. vastus medialis en lateralis bij duurlopers en sprinters.
Spier Duurlopers Sprinters
Plantair flexoren
(spiervolume/botlengte)
M. soleus 11,25 ± 2,36 12,34 ± 2,12
M. gastrocnemius medialis 6,53 ± 1,38 6,38 ± 0,48
M. gastrocnemius lateralis 4,38 ± 0,68 5,00 ± 0,78
SOL + GM + GL 22,16 ± 4,02 23,71 ± 2,57
Knie-extensoren
(spiervolume/botlengte)
M. rectus femoris 6,81 ± 0,93* 8,79 ± 0,70*
M. vastus medialis 10,80 ± 1,88 11,52 ± 1,27
M. vastus lateralis 28,75 ± 4,10~ 33,41 ± 4,60~
RF + VM + VL 46,37 ± 6,59~ 53,72 ± 6,11~
Totaal been
Totaal spiervolume/beenlengte 32,59 ± 4,36 36,6 ± 4,00
~ = p<0,10; * = p<0,05: duurlopers vs. sprinters
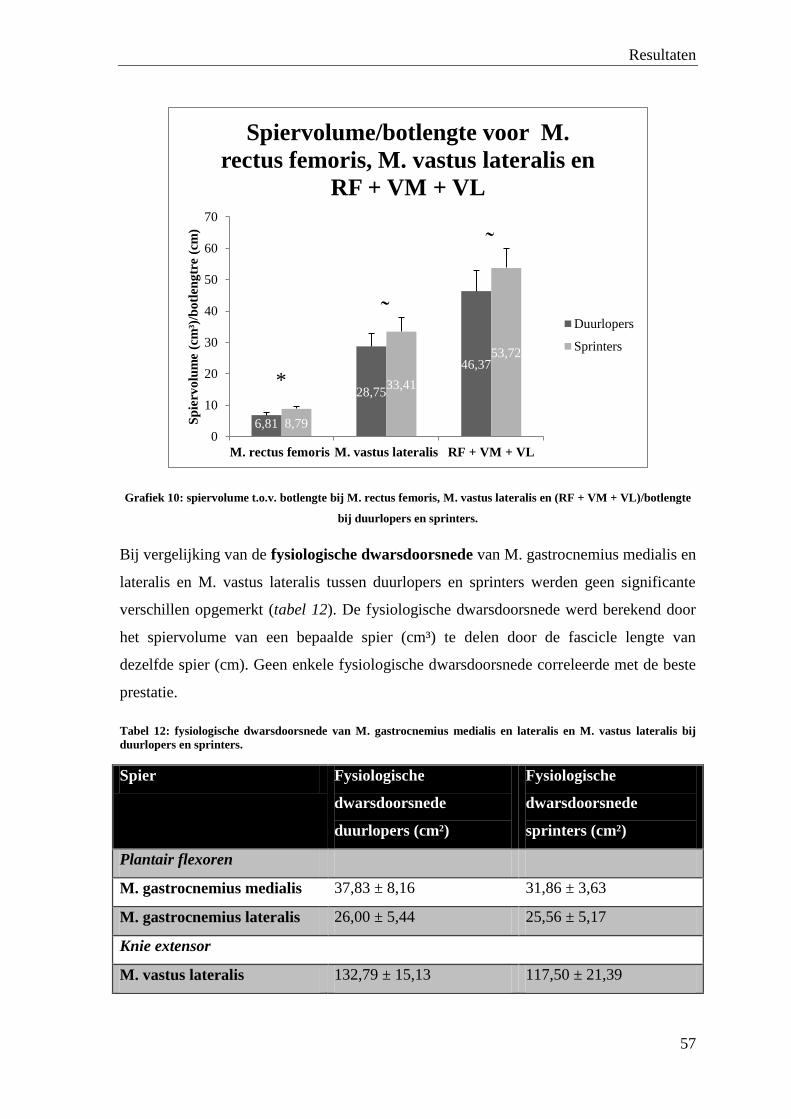
Resultaten
57
Grafiek 10: spiervolume t.o.v. botlengte bij M. rectus femoris, M. vastus lateralis en (RF + VM + VL)/botlengte
bij duurlopers en sprinters.
Bij vergelijking van de fysiologische dwarsdoorsnede van M. gastrocnemius medialis en
lateralis en M. vastus lateralis tussen duurlopers en sprinters werden geen significante
verschillen opgemerkt (tabel 12). De fysiologische dwarsdoorsnede werd berekend door
het spiervolume van een bepaalde spier (cm³) te delen door de fascicle lengte van
dezelfde spier (cm). Geen enkele fysiologische dwarsdoorsnede correleerde met de beste
prestatie.
Tabel 12: fysiologische dwarsdoorsnede van M. gastrocnemius medialis en lateralis en M. vastus lateralis bij
duurlopers en sprinters.
Spier Fysiologische
dwarsdoorsnede
duurlopers (cm²)
Fysiologische
dwarsdoorsnede
sprinters (cm²)
Plantair flexoren
M. gastrocnemius medialis 37,83 ± 8,16 31,86 ± 3,63
M. gastrocnemius lateralis 26,00 ± 5,44 25,56 ± 5,17
Knie extensor
M. vastus lateralis 132,79 ± 15,13 117,50 ± 21,39
6,81
28,75
46,37
8,79
33,41
53,72
0
10
20
30
40
50
60
70
M. rectus femoris M. vastus lateralis RF + VM + VL
Sp
ierv
olu
me
(cm
³)/b
otl
eng
tre (
cm)
Spiervolume/botlengte voor M.
rectus femoris, M. vastus lateralis en
RF + VM + VL
Duurlopers
Sprinters
*
˜
˜
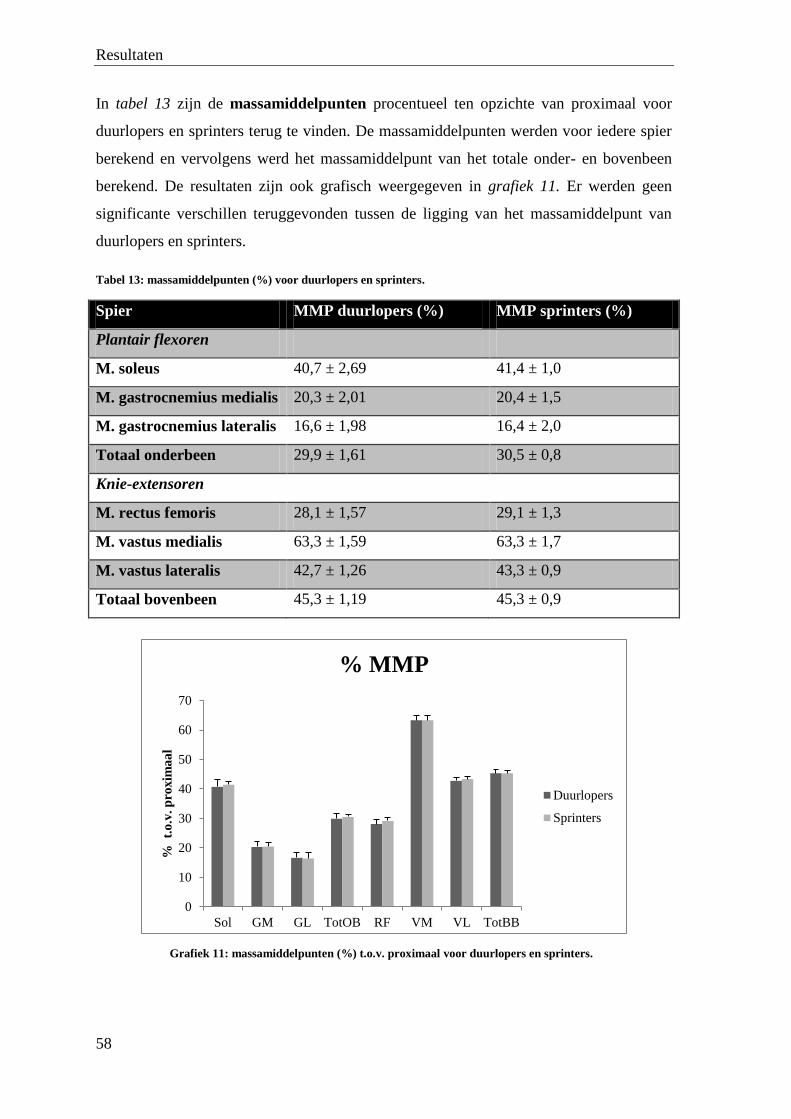
Resultaten
58
0
10
20
30
40
50
60
70
Sol GM GL TotOB RF VM VL TotBB
%
t.o
.v. p
roxim
aa
l
% MMP
Duurlopers
Sprinters
In tabel 13 zijn de massamiddelpunten procentueel ten opzichte van proximaal voor
duurlopers en sprinters terug te vinden. De massamiddelpunten werden voor iedere spier
berekend en vervolgens werd het massamiddelpunt van het totale onder- en bovenbeen
berekend. De resultaten zijn ook grafisch weergegeven in grafiek 11. Er werden geen
significante verschillen teruggevonden tussen de ligging van het massamiddelpunt van
duurlopers en sprinters.
Tabel 13: massamiddelpunten (%) voor duurlopers en sprinters.
Spier MMP duurlopers (%) MMP sprinters (%)
Plantair flexoren
M. soleus 40,7 ± 2,69 41,4 ± 1,0
M. gastrocnemius medialis 20,3 ± 2,01 20,4 ± 1,5
M. gastrocnemius lateralis 16,6 ± 1,98 16,4 ± 2,0
Totaal onderbeen 29,9 ± 1,61 30,5 ± 0,8
Knie-extensoren
M. rectus femoris 28,1 ± 1,57 29,1 ± 1,3
M. vastus medialis 63,3 ± 1,59 63,3 ± 1,7
M. vastus lateralis 42,7 ± 1,26 43,3 ± 0,9
Totaal bovenbeen 45,3 ± 1,19 45,3 ± 0,9
Grafiek 11: massamiddelpunten (%) t.o.v. proximaal voor duurlopers en sprinters.
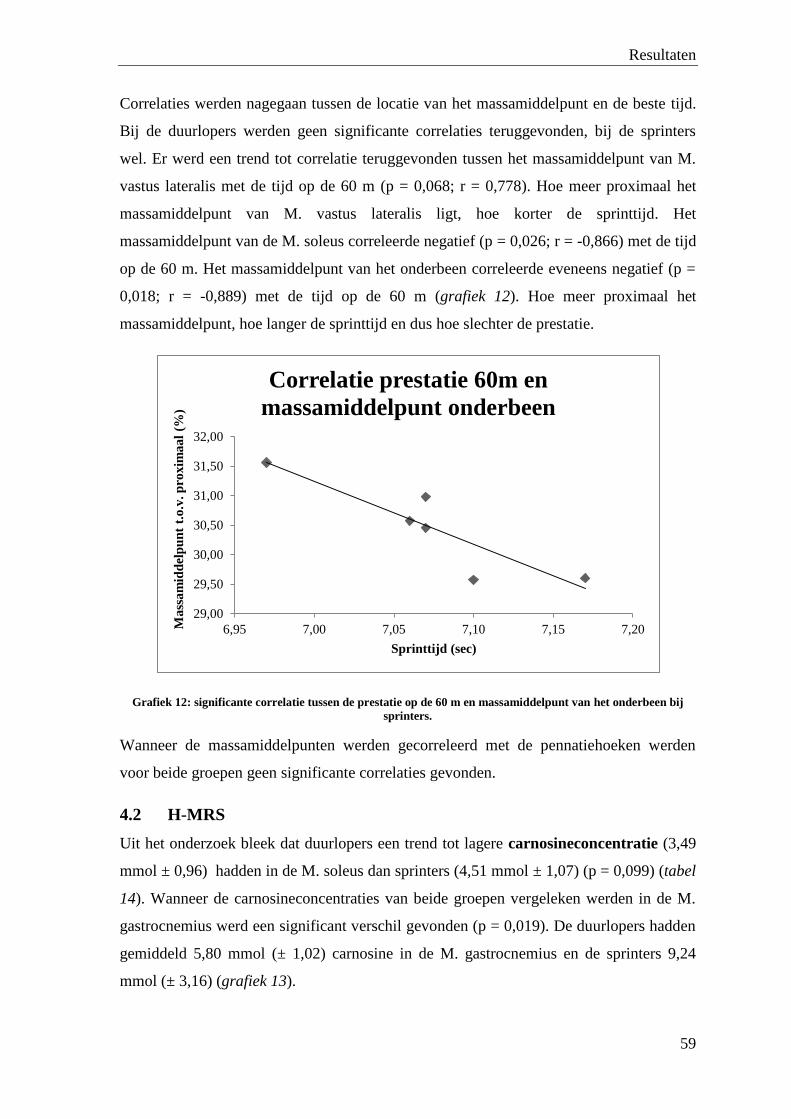
Resultaten
59
Correlaties werden nagegaan tussen de locatie van het massamiddelpunt en de beste tijd.
Bij de duurlopers werden geen significante correlaties teruggevonden, bij de sprinters
wel. Er werd een trend tot correlatie teruggevonden tussen het massamiddelpunt van M.
vastus lateralis met de tijd op de 60 m (p = 0,068; r = 0,778). Hoe meer proximaal het
massamiddelpunt van M. vastus lateralis ligt, hoe korter de sprinttijd. Het
massamiddelpunt van de M. soleus correleerde negatief (p = 0,026; r = -0,866) met de tijd
op de 60 m. Het massamiddelpunt van het onderbeen correleerde eveneens negatief (p =
0,018; r = -0,889) met de tijd op de 60 m (grafiek 12). Hoe meer proximaal het
massamiddelpunt, hoe langer de sprinttijd en dus hoe slechter de prestatie.
Grafiek 12: significante correlatie tussen de prestatie op de 60 m en massamiddelpunt van het onderbeen bij
sprinters.
Wanneer de massamiddelpunten werden gecorreleerd met de pennatiehoeken werden
voor beide groepen geen significante correlaties gevonden.
4.2 H-MRS
Uit het onderzoek bleek dat duurlopers een trend tot lagere carnosineconcentratie (3,49
mmol ± 0,96) hadden in de M. soleus dan sprinters (4,51 mmol ± 1,07) (p = 0,099) (tabel
14). Wanneer de carnosineconcentraties van beide groepen vergeleken werden in de M.
gastrocnemius werd een significant verschil gevonden (p = 0,019). De duurlopers hadden
gemiddeld 5,80 mmol (± 1,02) carnosine in de M. gastrocnemius en de sprinters 9,24
mmol (± 3,16) (grafiek 13).
29,00
29,50
30,00
30,50
31,00
31,50
32,00
6,95 7,00 7,05 7,10 7,15 7,20Ma
ssa
mid
del
pu
nt
t.o
.v. p
roxim
aa
l (%
)
Sprinttijd (sec)
Correlatie prestatie 60m en
massamiddelpunt onderbeen
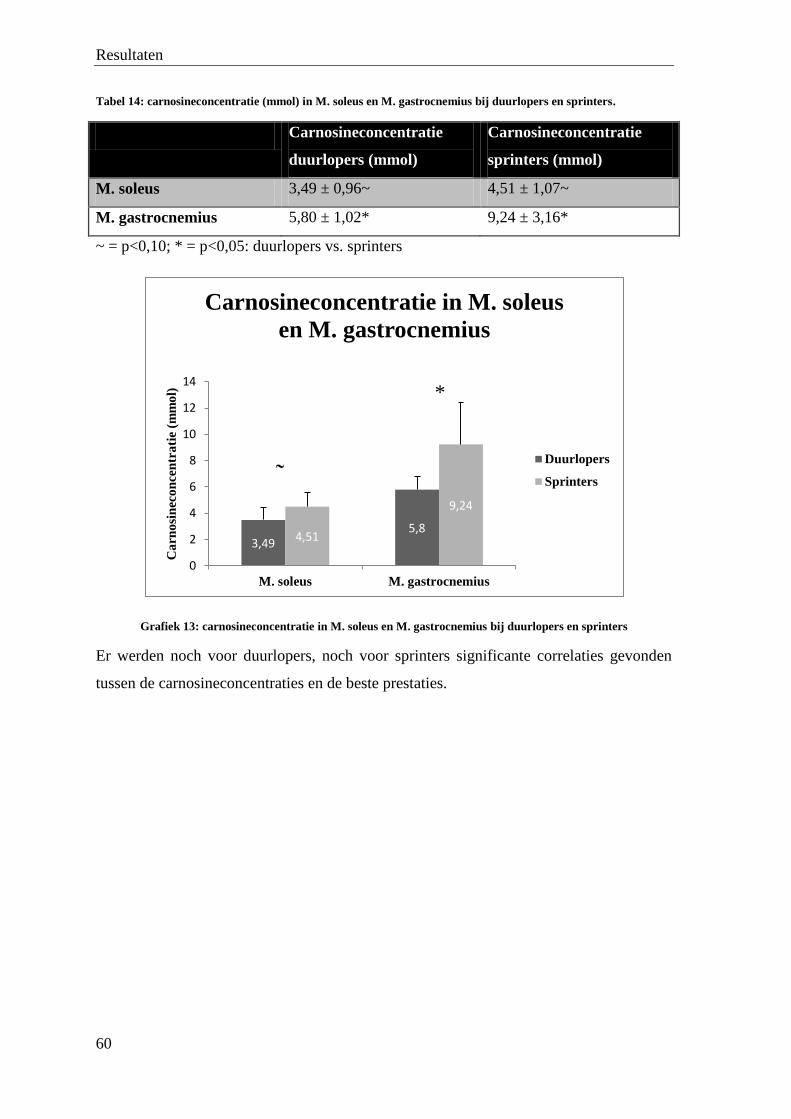
Resultaten
60
Tabel 14: carnosineconcentratie (mmol) in M. soleus en M. gastrocnemius bij duurlopers en sprinters.
Carnosineconcentratie
duurlopers (mmol)
Carnosineconcentratie
sprinters (mmol)
M. soleus 3,49 ± 0,96~ 4,51 ± 1,07~
M. gastrocnemius 5,80 ± 1,02* 9,24 ± 3,16*
~ = p<0,10; * = p<0,05: duurlopers vs. sprinters
Grafiek 13: carnosineconcentratie in M. soleus en M. gastrocnemius bij duurlopers en sprinters
Er werden noch voor duurlopers, noch voor sprinters significante correlaties gevonden
tussen de carnosineconcentraties en de beste prestaties.
3,49 5,8
4,51
9,24
0
2
4
6
8
10
12
14
M. soleus M. gastrocnemius
Ca
rno
sin
eco
nce
ntr
ati
e (m
mo
l)
Carnosineconcentratie in M. soleus
en M. gastrocnemius
Duurlopers
Sprinters˜
*

61
IV. DISCUSSIE

Discussie
62
1. Proefpersonen en studiedesign
In deze vergelijkende observationele cross-sectionele studie werden 13 proefpersonen
onderzocht: zeven duurlopers en zes sprinters. Aangezien er op basis van het
puntensysteem, opgesteld door de International Association of Athletics Federations,
geen uitbijters gevonden werden onder de atleten kan besloten worden dat geen enkele
atleet extreem beter of slechter was dan de anderen. Bovendien was er geen significant
verschil terug te vinden tussen de gemiddelde scores van de onderzochte duurlopers en
sprinters. Beide groepen atleten kunnen dus terecht met elkaar vergeleken worden.
Het aantal proefpersonen in deze studie was relatief laag waardoor significantie soms
moeilijk aan te tonen was. De reden van dit lage aantal is drieledig. Ten eerste zijn de
atleten allemaal van subtop- en topniveau. Wegens hun drukbezette leven zijn deze
atleten moeilijk te overtuigen om deel te nemen aan wetenschappelijke studies.
Bovendien was de NMR- scanner drukbezet. Vaak kon deze enkel ’s avonds en in het
weekend gebruikt worden. Als laatste vergt deze onderzoekstechniek een zeer intensieve
en tijdrovende dataverwerking. De proefpersonen moesten allen getest worden tussen
begin januari en eind maart waardoor meer dan 13 proefpersonen praktisch niet haalbaar
was. Daarentegen kunnen met deze studie heel wat interessante zaken aangetoond
worden.
In een later stadium kan de studie nog uitgebreid worden met meer proefpersonen.
Onder de atleten kan in de toekomst een onderscheid gemaakt worden tussen subtoppers
en toppers. Het zou bovendien interessant zijn om een controlegroep toe te voegen. Deze
controlepersonen kunnen sedentaire personen zijn. Er kan echter ook gebruik gemaakt
worden van tienkampers als controles. Volgens Van Damme et al. (2002) kunnen
tienkampers beschouwd worden als generalisten. Ze scoren in de tien onderdelen van de
tienkamp goed, maar ze blinken niet uit in één bepaald atletieknummer. Toch kan de
bedenking gemaakt worden dat tienkampers morfologisch dichter bij sprinters aanleunen
dan bij duurlopers. Een andere mogelijkheid is het gebruik van halve fond lopers (800 tot
1500 m) als controles. Hun loopafstand bevindt zich namelijk tussen de voorkeurafstand
van de onderzochte duurlopers (3000 m) en sprinters (60 – 100 m).
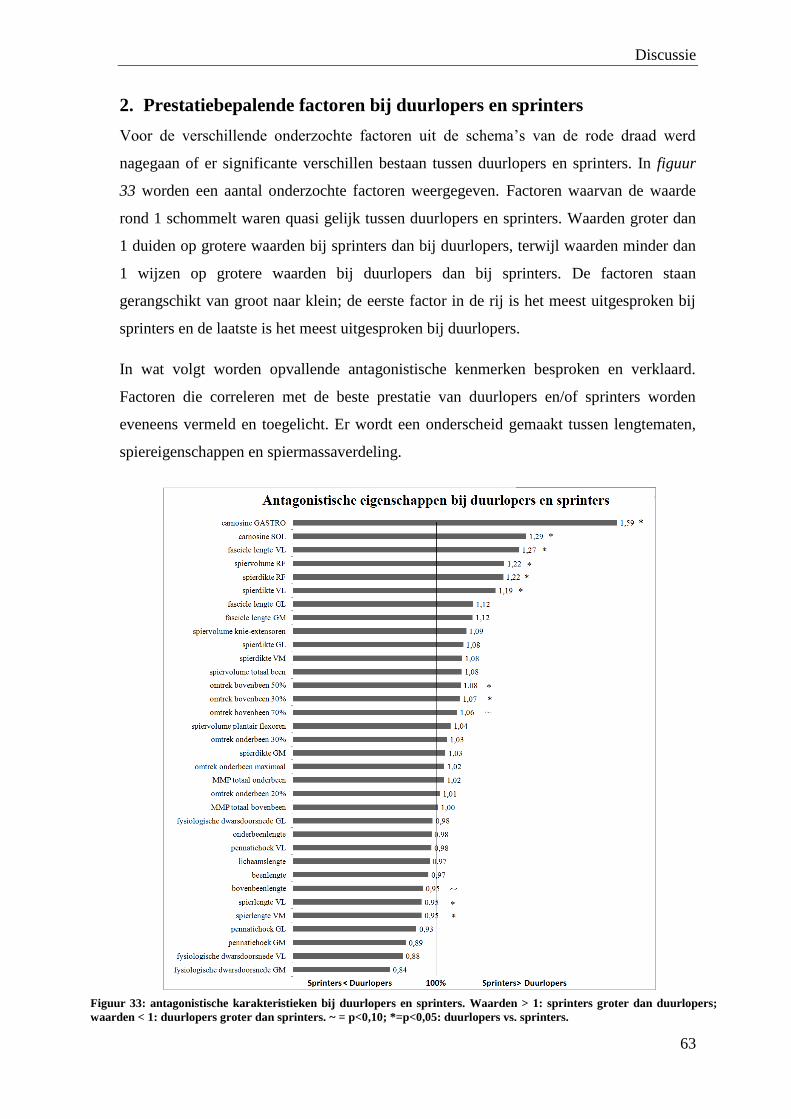
Discussie
63
2. Prestatiebepalende factoren bij duurlopers en sprinters
Voor de verschillende onderzochte factoren uit de schema’s van de rode draad werd
nagegaan of er significante verschillen bestaan tussen duurlopers en sprinters. In figuur
33 worden een aantal onderzochte factoren weergegeven. Factoren waarvan de waarde
rond 1 schommelt waren quasi gelijk tussen duurlopers en sprinters. Waarden groter dan
1 duiden op grotere waarden bij sprinters dan bij duurlopers, terwijl waarden minder dan
1 wijzen op grotere waarden bij duurlopers dan bij sprinters. De factoren staan
gerangschikt van groot naar klein; de eerste factor in de rij is het meest uitgesproken bij
sprinters en de laatste is het meest uitgesproken bij duurlopers.
In wat volgt worden opvallende antagonistische kenmerken besproken en verklaard.
Factoren die correleren met de beste prestatie van duurlopers en/of sprinters worden
eveneens vermeld en toegelicht. Er wordt een onderscheid gemaakt tussen lengtematen,
spiereigenschappen en spiermassaverdeling.
Figuur 33: antagonistische karakteristieken bij duurlopers en sprinters. Waarden > 1: sprinters groter dan duurlopers;
waarden < 1: duurlopers groter dan sprinters. ~ = p<0,10; *=p<0,05: duurlopers vs. sprinters.

Discussie
64
2.1 Lengtematen
Wat betreft de lichaamslengte van duurlopers en sprinters zijn er in de literatuur geen
eenduidige resultaten terug te vinden. Onderzoek van Radford (1990) toonde aan dat
duurlopers gemiddeld groter zijn dan sprinters, terwijl een studie van Weyand en Davis
(2005) net het omgekeerde beweerde. In dit onderzoek werd geen verschil teruggevonden
tussen de lichaamslengte van duurlopers en sprinters. De gemiddelde lichaamslengte van
de sprinters (176,52 cm ± 5,14) lag in de optimale zone beschreven door Uth (2005).
Deze zone is echter zo ruim (gaande van 168 tot 191 cm) dat dit gegeven niet
verwonderlijk is. Uit bovenstaande tegenstrijdige onderzoeken en niet-significante
resultaten van het onderzoek kan besloten worden dat de lichaamslengte op subtopniveau
vermoedelijk een weinig prestatiebepalende rol speelt. Als gekeken wordt naar de
lichaamslengte van enkele wereldtoppers kan een verschil verwacht worden tussen
topduurlopers en topsprinters. Topduurlopers zoals Daniel Komen en Kenenisa Bekele
meten respectievelijk 170 cm en 174 cm. Topsprinter Usain Bolt heeft een veel grotere
lengte van 196 cm, Asafa Powell meet 190 cm. In de toekomst kan het nuttig zijn om
verschillen te zoeken in lichaamslengte tussen topatleten èn tussen de verschillende
rassen op wereldtopniveau. Hoogstwaarschijnlijk komt er daar wel een significant
verschil naar boven.
Naast lichaamslengte werden ook op het vlak van onder- en bovenbeenlengte een aantal
resultaten gevonden in dit onderzoek die tegenstrijdig leken aan de literatuur. Lee en
Piazza (2009) vonden een gematigde correlatie tussen de onderbeenlengte en de beste tijd
voor de 100 m sprint: een kleinere onderbeenlengte bij sprinters zou zorgen voor een
betere prestatie. Deze correlatie kon echter met dit onderzoek niet bevestigd worden. Er
werd een trend tot significant verschil gevonden tussen de bovenbeenlengte van beide
groepen atleten met een langere bovenbeenlengte bij duurlopers (44,80 cm ± 2,21) dan bij
sprinters (42,60 cm ± 2,15). Aangezien dit slechts aangetoond werd op het 90%
betrouwbaarheidsniveau (p = 0,097), zou dit kunnen wijzen op een toevalligheid. Relatief
ten opzichte van de lichaamslengte is er bovendien geen verschil in bovenbeenlengte
tussen duurlopers en sprinters gevonden. Verder onderzoek hieromtrent is nodig.
Bij onderzoek van de spierlengtes werd een grotere spierlengte van de M. vastus lateralis
gevonden bij duurlopers dan bij sprinters (p = 0,091). Wegens de grotere
bovenbeenlengte bij duurlopers, lijkt een grotere spierlengte hieruit een logisch gevolg.

Discussie
65
Er kan besloten worden dat de lichaamslengte niet significant verschilt tussen duurlopers
en sprinters van subtopniveau en dus waarschijnlijk ook geen prestatiebepalende factor is.
In de toekomst kan lichaamslengte vergeleken worden tussen atleten van verschillende
rassen van topniveau. Bovendien kan het nuttig zijn om aanvullend onderzoek te doen
rond de ligging van het lichaamszwaartepunt en een eventueel verschil tussen rassen. Een
hoger lichaamszwaartepunt zou voordelig kunnen zijn bij het lopen door de hogere
voorwaartse valsnelheid dat dit gegeven met zich meebrengt (Bejan et al., 2010).
Hierdoor zou dus sneller gelopen kunnen worden. Onderzoek van de lengte van de tenen
en de momentsarm van de achillespees zijn eveneens te onderzoeken factoren voor een
toekomstig onderzoek.
2.2 Spiereigenschappen
Als hypothese werd gesteld dat duurlopers zouden beschikken over kortere fascicle
lengtes en grotere pennatiehoeken in de M. gastrocnemius lateralis en medialis en in de
M. vastus lateralis. Bij duurlopers zouden bovenstaande zaken een invloed hebben op de
loopeconomie wat op zijn beurt zorgt voor een betere prestatie. Bij sprinters moeten deze
factoren zorgen voor een zo groot mogelijke spierverkortingssnelheid om de netto
voorwaartse impuls zo groot mogelijk te maken tijdens de acceleratiefase. Een tweede
hypothese beweert dat duurlopers over minder type 2 vezels beschikken dan sprinters. In
wat volgt wordt nagegaan of bovenstaande hypotheses kloppen.
Enkel bij de M. vastus lateralis is de fascicle lengte significant kleiner bij duurlopers dan
bij sprinters. Wel werden ook voor de M. gastrocnemius medialis en de M. gastrocnemius
lateralis kleinere waarden gevonden voor de fascicle lengte bij duurlopers in vergelijking
met sprinters. Dit verschil kon echter niet significant worden aangetoond. Een mogelijke
verklaring voor de kortere fascicles bij duurlopers is de hogere metabole kost die een
langere fascicle lengte met zich meebrengt. Een langere fascicle lengte betekent immers
dat er meer sarcomeren in serie staan. Elk van deze sarcomeren moet van ATP voorzien
worden om te kunnen samentrekken tijdens spiercontractie. De energiebehoefte is dus
groter wat nadelig is voor de loopeconomie en bijgevolg ook voor de duurprestatie. Uit de
resultaten van dit onderzoek bleek bovendien dat de pennatiehoek van de M.
gastrocnemius medialis, de M. gastrocnemius lateralis en de M. vastus lateralis van de
duurlopers telkens groter was dan die van de sprinters. Helaas kon dit verschil niet
significant aangetoond worden.

Discussie
66
Tijdens het onderzoek werd het percentage type 2 vezels indirect bepaald aan de hand
van de gemeten carnosineconcentratie in de M. soleus en M. gastrocnemius. Er was een
trend tot lagere carnosineconcentratie bij duurlopers in de M. soleus en een significant
lagere carnosineconcentratie in de M. gastrocnemius. Lagere carnosineconcentraties
werden gevonden in de M. soleus. Dit is evident want de M. soleus is een trage spier met
een hoog oxidatief potentieel. Deze spier bevat meer mitochondriën en bijgevolg meer
type 1 vezels dan de M. gastrocnemius die als een snelle spier beschouwd wordt met een
hoog glycolytisch potentieel en meer type 2 vezels. Deze spier produceert kracht voor
kortere periodes dan de M. soleus en is bijgevolg sneller moe. Uit de literatuur (Baguet et
al., 2011) bleek dat sprinters gemiddeld 1,9 keer hogere carnosine-inhouden hadden dan
duuratleten in de M. gastrocnemius. In dit onderzoek bleek de carnosine-inhoud van
sprinters gemiddeld 1,59 keer groter dan die van de duurlopers. De lagere waarde in dit
onderzoek kan verklaard worden door het feit dat de duurlopers in de studie van Baguet et
al. (2001) marathonlopers en triatleten waren. Zij bezaten gemiddeld een lagere
carnosineconcentratie dan de duurlopers uit ons onderzoek. Aan de hand van de
regressievergelijking (y = 0,1139x + 0,1926) gevonden door Baguet et al. (2011) waarbij
y staat voor de carnosineconcentratie in mmol en x voor het percentage type 2 vezels kon
een schatting gemaakt worden van het percentage type 2 vezels. Zo hadden duurlopers
gemiddeld ongeveer 37% type 2 vezels in de M. gastrocnemius. Bij de sprinters was dit
ongeveer 60%. De percentages gevonden in de M. gastrocnemius komen overeen met de
literatuur (Baguet et a., 2011; Costill et al., 1976) (figuur 34).
Er kan geconcludeerd worden dat de hypotheses (duurlopers beschikken over kortere
fascicle lengtes, grotere pennatiehoeken en minder type 2 vezels dan sprinters) niet
volledig kunnen bevestigd worden. Duurlopers beschikken enkel over kortere fascicle
lengtes in de M. vastus lateralis. Pennatiehoeken lijken groter bij duurlopers dan bij
sprinters maar dit kon niet significant bevestigd worden. De hypothese dat duurlopers
minder type 2 vezels hebben, werd wel bevestigd aan de hand van de gemeten
carnosineconcentratie. Dit laatste kan gezien worden als een duidelijke antagonistische
factor waarbij duurlopers lage waarden hebben terwijl bij sprinters hoge waarden worden
vastgesteld.
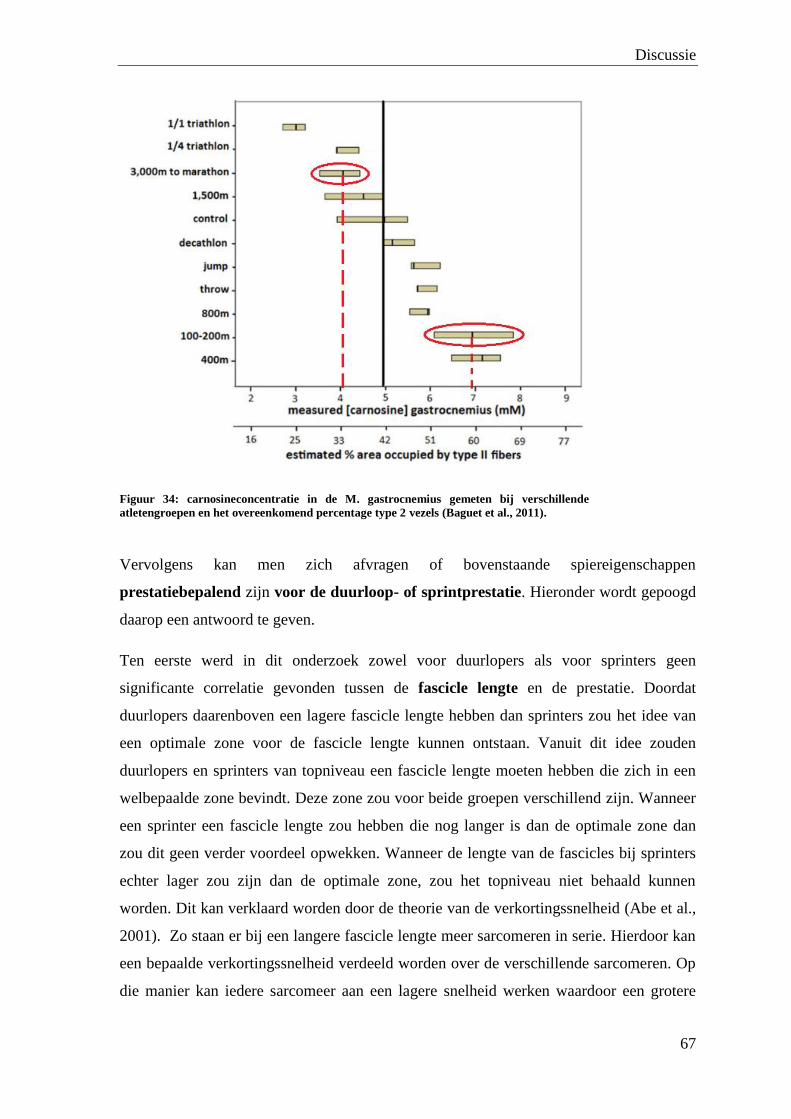
Discussie
67
Vervolgens kan men zich afvragen of bovenstaande spiereigenschappen
prestatiebepalend zijn voor de duurloop- of sprintprestatie. Hieronder wordt gepoogd
daarop een antwoord te geven.
Ten eerste werd in dit onderzoek zowel voor duurlopers als voor sprinters geen
significante correlatie gevonden tussen de fascicle lengte en de prestatie. Doordat
duurlopers daarenboven een lagere fascicle lengte hebben dan sprinters zou het idee van
een optimale zone voor de fascicle lengte kunnen ontstaan. Vanuit dit idee zouden
duurlopers en sprinters van topniveau een fascicle lengte moeten hebben die zich in een
welbepaalde zone bevindt. Deze zone zou voor beide groepen verschillend zijn. Wanneer
een sprinter een fascicle lengte zou hebben die nog langer is dan de optimale zone dan
zou dit geen verder voordeel opwekken. Wanneer de lengte van de fascicles bij sprinters
echter lager zou zijn dan de optimale zone, zou het topniveau niet behaald kunnen
worden. Dit kan verklaard worden door de theorie van de verkortingssnelheid (Abe et al.,
2001). Zo staan er bij een langere fascicle lengte meer sarcomeren in serie. Hierdoor kan
een bepaalde verkortingssnelheid verdeeld worden over de verschillende sarcomeren. Op
die manier kan iedere sarcomeer aan een lagere snelheid werken waardoor een grotere
Figuur 34: carnosineconcentratie in de M. gastrocnemius gemeten bij verschillende
atletengroepen en het overeenkomend percentage type 2 vezels (Baguet et al., 2011).

Discussie
68
kracht geproduceerd kan worden volgens de kracht-snelheidscurve. Bij duurlopers
daarentegen zou een fascicle lengte die groter is dan de optimale zone nadelig kunnen zijn
vanwege de grotere metabole kost. Een fascicle lengte die zich onder de optimale zone bij
duurlopers bevindt zou dan weer voor een te kleine krachtproductie zorgen.
In de literatuur (Abe et al., 2000) vond men dat de pennatiehoek van sprinters een
positieve correlatie vertoonde met de sprinttijd op de 100 m. Een kleinere pennatiehoek
zou dus zorgen voor een betere prestatie. In deze studie werden zowel voor de sprinters
als voor de duurlopers geen correlaties gevonden tussen pennatiehoek en prestatie. Het
ontbreken van correlaties in deze studie zou kunnen liggen aan het relatief lage aantal
proefpersonen (7 duurlopers en 6 sprinters) in vergelijking met de studie van Abe et al.
(2000) waar men gebruik maakte van 24 duurlopers en 23 sprinters. De pennatiehoek zou
een prestatiebepalende factor kunnen zijn. Doordat er in deze studie geen correlaties
konden aangetoond worden, terwijl dit in andere studies wel het geval was, is verder
onderzoek vereist.
Bij duurlopers bleek tussen de carnosineconcentratie en de prestatie geen significante
negatieve correlatie te bestaan zowel in de M. soleus als de M. gastrocnemius. Bijgevolg
bestaat er ook geen positieve correlatie tussen het percentage type 1 vezels en de
prestatie. Dit is tegenstrijdig aan de literatuur. Costill et al. (1976) vonden immers wel
een positieve correlatie tussen het percentage type 1 vezels en de duurloopprestatie: hoe
meer type 1 vezels, hoe beter de prestatie. Een verklaring voor de afwezigheid van een
correlatie in dit onderzoek kan zijn dat een duurloper met wat minder type 1 vezels toch
nog over veel type 2a vezels kan beschikken. Type 2a vezels hebben een veel hoger
oxidatief potentieel dan type 2x vezels. Met uithoudingstraining kan het oxidatief
potentieel nog verhoogd worden tot een punt waar het dat van de type 1 vezels van
sedentairen benadert (Henriksson & Reitman, 1976). Op deze manier kan een atleet met
minder type 1 vezels toch een goede duurloper zijn, met als voorwaarde dat hij over een
bepaald minimum aan type 1 vezels beschikt. Zo kan iemand met 60% type 1 vezels in de
M. gastrocnemius een even goede duurloper zijn als iemand met 80% type 1 vezels
vanwege de compensatie door de oxidatieve type 2a vezels. Iemand met 30% type 1
vezels kan echter nooit een goede duurloper zijn.
Bij sprinters bleek er eveneens geen correlatie te bestaan tussen de carnosineconcentratie
en de prestatie. De carnosineconcentratie is wel hoger dan bij duurlopers zoals eerder

Discussie
69
reeds vermeld. Het ontbreken van een correlatie kan wijzen op het bestaan van een
optimale zone voor de carnosineconcentratie. Eenmaal de carnosineconcentratie een
bepaalde waarde overstijgt, zal een verdere verhoging van de carnosine geen verdere
verbetering van de prestatie met zich meebrengen. Voorzichtigheid bij deze conclusie is
echter geboden wegens het lage aantal proefpersonen.
Samenvattend kan gesteld worden dat er geen significante correlaties werden gevonden
tussen de spiereigenschappen (fascicle lengte, pennatiehoek en spiervezeltype) en de
prestatie zowel bij duurlopers als bij sprinters. Toch moet het idee van een optimale zone
in rekening gebracht worden bij de fascicle lengte. Vervolgens werd reeds aangetoond in
de literatuur (Abe et al., 2000) dat de pennatiehoek van sprinters een significante
positieve correlatie vertoonde met de sprinttijd op de 100 m. Dit werd in dit onderzoek
niet teruggevonden. Bij de duurlopers werd eveneens geen correlatie teruggevonden.
Verder onderzoek zal hier eenduidigheid moeten brengen. Als laatste konden
carnosineconcentratie en prestatie niet gecorreleerd worden met elkaar. Toch bleek uit
een eerdere studie (Abe et al., 2000) dat de carnosineconcentratie bij duurlopers als een
prestatiebepalende factor beschouwd kan worden. De afwezigheid van een significante
correlatie tussen de carnosineconcentratie en de prestatie zou echter kunnen verklaard
worden door het hoge oxidatieve potentieel van de type 2a vezels. Een minimum aantal
type 1 vezels is hier uiteraard vereist. Bij de sprinters wordt het idee van een optimale
zone aangewend. De sprinters moeten een bepaalde carnosineconcentratie bezitten om bij
de top te behoren, maar een verdere verhoging zou geen verder voordeel opleveren.
2.3 Spiermassaverdeling
Voor de beschrijving van de spiermassaverdeling werd in dit onderzoek gebruik gemaakt
van omtrekken, spierdiktes en spiervolumemetingen. Als eerste werden voor het
onderbeen geen verschillen in omtrek gevonden tussen de duurlopers en de sprinters
(figuur 35). Dit stemt niet overeen met de resultaten van Vučetić et al. (2005). Uit dit
onderzoek bleek namelijk dat sprinters een grotere onderbeenomtrek hadden dan
controles én afstandslopers. De omtrekken van het bovenbeen op 30 % en 50%
proximaal ten opzichte van de trochanter major, waren in dit onderzoek echter wel groter
bij de sprinters dan bij de duurlopers (Figuur 35). Op 70% was er een trend tot grotere
omtrek bij sprinters dan bij duurlopers.
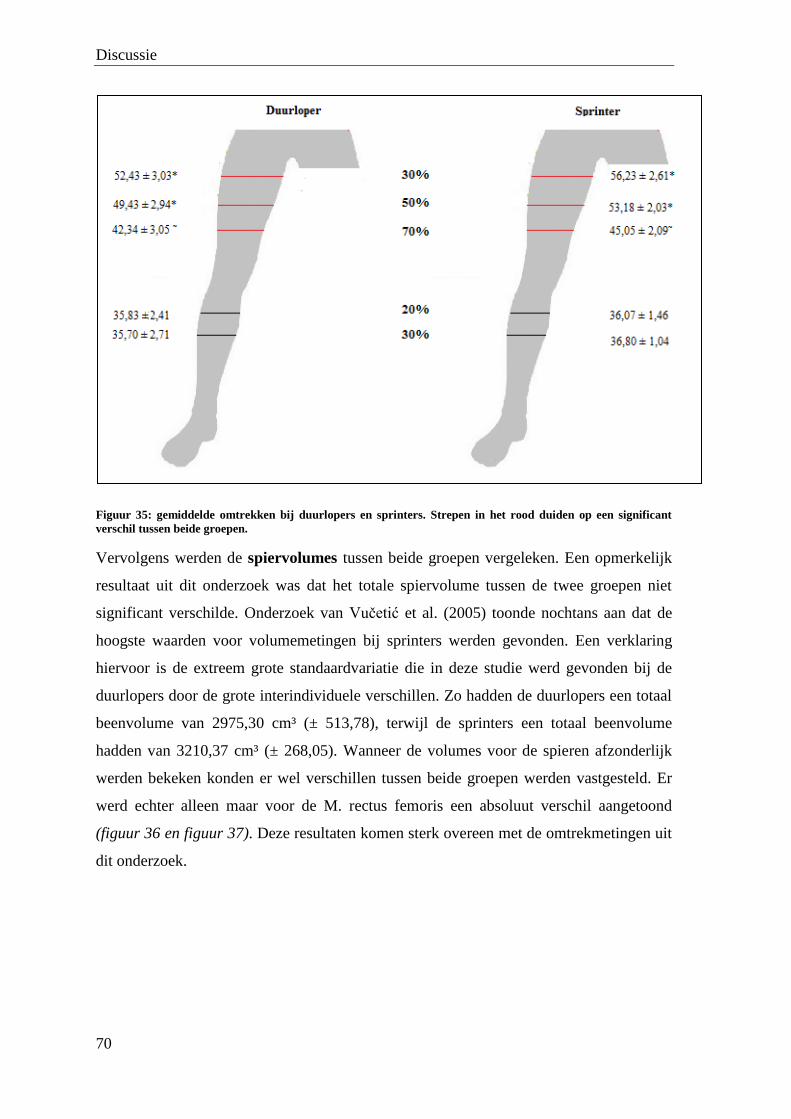
Discussie
70
Figuur 35: gemiddelde omtrekken bij duurlopers en sprinters. Strepen in het rood duiden op een significant
verschil tussen beide groepen.
Vervolgens werden de spiervolumes tussen beide groepen vergeleken. Een opmerkelijk
resultaat uit dit onderzoek was dat het totale spiervolume tussen de twee groepen niet
significant verschilde. Onderzoek van Vučetić et al. (2005) toonde nochtans aan dat de
hoogste waarden voor volumemetingen bij sprinters werden gevonden. Een verklaring
hiervoor is de extreem grote standaardvariatie die in deze studie werd gevonden bij de
duurlopers door de grote interindividuele verschillen. Zo hadden de duurlopers een totaal
beenvolume van 2975,30 cm³ (± 513,78), terwijl de sprinters een totaal beenvolume
hadden van 3210,37 cm³ (± 268,05). Wanneer de volumes voor de spieren afzonderlijk
werden bekeken konden er wel verschillen tussen beide groepen werden vastgesteld. Er
werd echter alleen maar voor de M. rectus femoris een absoluut verschil aangetoond
(figuur 36 en figuur 37). Deze resultaten komen sterk overeen met de omtrekmetingen uit
dit onderzoek.
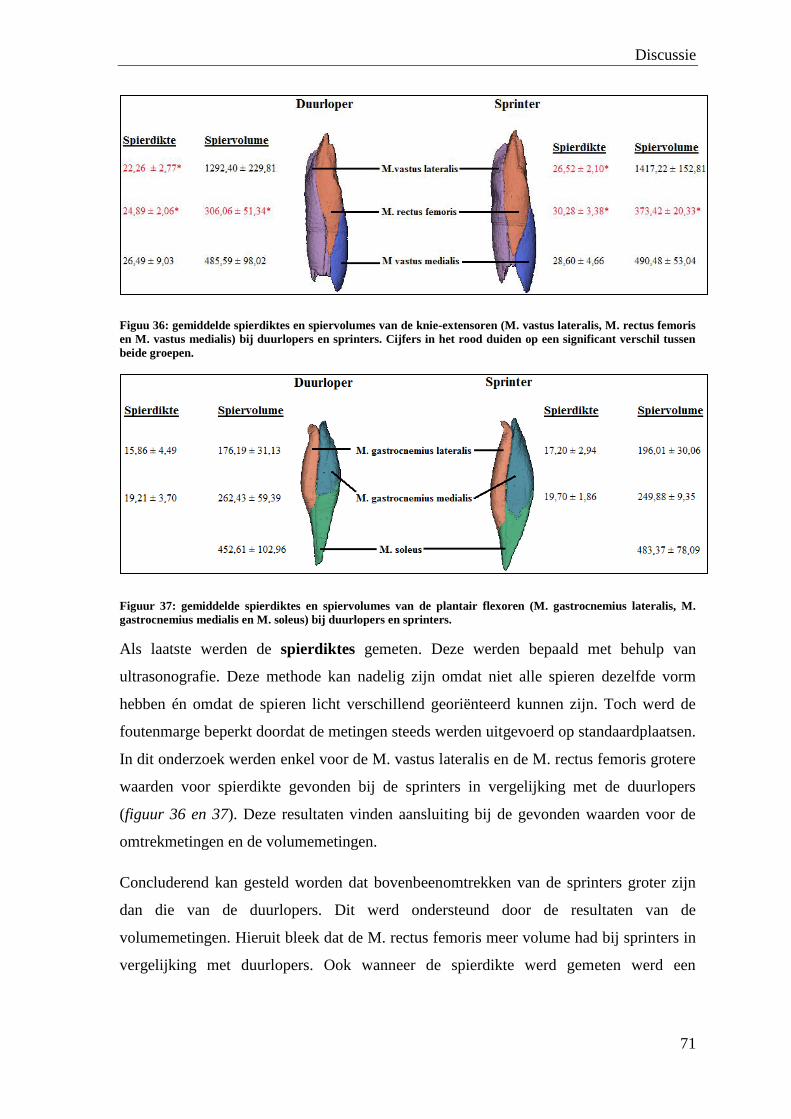
Discussie
71
Figuu 36: gemiddelde spierdiktes en spiervolumes van de knie-extensoren (M. vastus lateralis, M. rectus femoris
en M. vastus medialis) bij duurlopers en sprinters. Cijfers in het rood duiden op een significant verschil tussen
beide groepen.
Figuur 37: gemiddelde spierdiktes en spiervolumes van de plantair flexoren (M. gastrocnemius lateralis, M.
gastrocnemius medialis en M. soleus) bij duurlopers en sprinters.
Als laatste werden de spierdiktes gemeten. Deze werden bepaald met behulp van
ultrasonografie. Deze methode kan nadelig zijn omdat niet alle spieren dezelfde vorm
hebben én omdat de spieren licht verschillend georiënteerd kunnen zijn. Toch werd de
foutenmarge beperkt doordat de metingen steeds werden uitgevoerd op standaardplaatsen.
In dit onderzoek werden enkel voor de M. vastus lateralis en de M. rectus femoris grotere
waarden voor spierdikte gevonden bij de sprinters in vergelijking met de duurlopers
(figuur 36 en 37). Deze resultaten vinden aansluiting bij de gevonden waarden voor de
omtrekmetingen en de volumemetingen.
Concluderend kan gesteld worden dat bovenbeenomtrekken van de sprinters groter zijn
dan die van de duurlopers. Dit werd ondersteund door de resultaten van de
volumemetingen. Hieruit bleek dat de M. rectus femoris meer volume had bij sprinters in
vergelijking met duurlopers. Ook wanneer de spierdikte werd gemeten werd een

Discussie
72
gelijkaardig resultaat gevonden. Hierbij was de M. vastus lateralis en de M. rectus
femoris significant dikker bij sprinters in vergelijking met duurlopers.
In wat volgt worden enkele opmerkelijke resultaten besproken. Hiervoor wordt telkens
een mogelijke verklaring gezocht. Als eerste blijkt uit de volume- en spierdiktemetingen
dat de M. rectus femoris dikker is bij sprinters. Verder werd een negatieve correlatie
gevonden tussen het volume van de M. rectus femoris en de sprinttijd voor de 60 m. Hoe
groter het spiervolume, hoe beter de prestatie.
Uit deze gegevens kan besloten worden dat de M. rectus femoris een van dé
belangrijkste spieren is bij sprinters. Deze bi-articulaire spier zorgt voor een
kniestrekkend en een heupbuigend moment. Deze twee bewegingen zijn essentieel bij de
sprintprestatie. Zo zorgt de voorovergebogen positie bij de acceleratiefase ervoor dat het
kniestrekkend moment maximaal kan zijn. De voorovergebogen positie zorgt er namelijk
voor dat er in de heup weinig kracht moet aangewend worden, waardoor het overgrote
deel van de kracht naar het kniestrekkend moment kan gaan. Wanneer de sprinter
vervolgens in de maximale snelheidsfase meer rechtop komt, zorgt het heupbuigend
moment er dan weer voor dat het been snel en krachtig omhoog geheven kan worden.
Ten tweede werden bij de vergelijking van de fysiologische dwarsdoorsnedes van M.
gastrocnemius medialis en lateralis en M. vastus lateralis tussen duurlopers en sprinters
geen significante verschillen opgemerkt. De waarden lagen zelfs iets hoger bij duurlopers,
terwijl men eerder hogere waarden bij de sprinters zou verwachten wegens hun grotere
spierdikte. Fysiologische dwarsdoorsnede is een maat voor de spierkracht. Bij de
sprintprestatie moet in een heel korte tijd heel veel kracht aangewend worden, dus het
lijkt logisch dat sprinters baat hebben bij een grote fysiologische dwarsdoorsnede. Een
mogelijke verklaring voor het niet vinden van een significant verschil tussen duurlopers
en sprinters kan de volgende zijn. De fysiologische dwarsdoorsnede is gelijk aan het
oppervlak van de doorsnede loodrecht op de spiervezelrichting. Aangezien duurlopers
beschikken over grotere pennatiehoeken (al werd dit niet significant aangetoond) zou dit
kunnen leiden tot een gelijke fysiologische doorsnede bij duurlopers en sprinters ondanks
de grotere spierdikte bij sprinters (figuur 38).

Discussie
73
Figuur 38: schets van een spier met aanduiding van mogelijke spiervezelrichting en fysiologische
dwarsdoorsnede bij duurlopers en sprinters
Vervolgens vertonen duurlopers en sprinters volgens de literatuur een verschil in
spiervorm (Abe et al., 2000). Beide groepen hadden verschillende spierdiktes op 30% en
50% van het bovenbeen, terwijl er geen verschil werd teruggevonden op 70% van het
bovenbeen. In dit onderzoek werden echter zowel op 30%, 50% en 70% van het
bovenbeen verschillen gevonden tussen duurlopers en sprinters. Bovendien werd er geen
verschil teruggevonden in de positie van het massamiddelpunt bij sprinters en duurlopers.
Een verschillende spiervorm kan hier dus niet bevestigd worden. Toch was het verschil
op 70% maar aantoonbaar op het 10% betrouwbaarheidsniveau (p=0,095). Aangezien
deze trend zo zwak is, zou dit verschil een toevalligheid kunnen zijn en zou de spiervorm
van het bovenbeen dus wel kunnen verschillen tussen beide groepen. Hierdoor zouden de
resultaten uit de studie van Abe et al. (2000) wel bevestigd kunnen worden.
Een vierde aspect dat aandacht verdient om te bespreken, is de correlatie tussen de
ligging van het massamiddelpunt en de prestatie bij sprinters. Zo bleek een
significante correlatie te bestaan tussen het massamiddelpunt van het volledige onderbeen
en de prestatie. Een lager massamiddelpunt zou zorgen voor een betere tijd. Dit lijkt niet
logisch omdat een lager massamiddelpunt zou zorgen voor een groter traagheidsmoment.
Blijkbaar weegt het positieve effect van de extra distale massa en de daarbij horende
krachtlevering meer door dan het negatieve effect dat deze extra massa op het
traagheidsmoment uitoefent. De extra distale massa zou kunnen verklaard worden door de
langere fascicle lengtes die werden teruggevonden bij sprinters. Bij de duurlopers werd
verwacht dat een hoger gelegen massamiddelpunt in het onderbeen positief zou correleren

Discussie
74
met de beste prestatie. Dit werd gedacht omdat duurlopers vaak heel smalle benen hebben
waarbij de spiermassa zich vooral aan de proximale kant van het onderbeen situeert. Dit
gegeven valt vooral op bij de absolute top van de duurlopers zoals de Kenianen en andere
Oost-Afrikanen. Deze correlatie ontbrak echter in dit onderzoek. Het zou dus kunnen dat
een hoog gelegen massamiddelpunt van ondergeschikt belang is om te kunnen
meedraaien aan de Belgische top.
Een laatste opmerkelijk resultaat omvat de negatieve correlatie die in deze studie werd
gevonden tussen de omtrek van het bovenbeen 70 % en de sprinttijd voor de 60 m bij
sprinters. Volgens deze correlatie zorgt een grotere omtrek voor een kortere sprinttijd en
dus voor een betere prestatie.
Zoals hierboven reeds aangereikt, zorgt distaal gelegen massa voor een negatieve invloed
op het traagheidsmoment bij het naar voor slingeren van het been tijdens het lopen.
Blijkbaar weegt ook hier de grotere spiermassa met daarbij horende grotere
krachtlevering meer door dan het negatief effect op het traagheidsmoment. Verder
onderzoek omtrent deze prestatiebepalende factor bij sprinters is aangewezen.
3. Conclusie
Om tot een schematische conclusie te komen in dit onderzoek worden de schema’s uit de
rode draad opnieuw aangehaald. Hierbij zijn echter enkele aanvullingen gedaan (figuur 39
en 40). Zo zijn de antagonistische eigenschappen aangeduid, alsook de prestatiebepalende
factoren, die in dit onderzoek bevestigd werden. Er werd ervoor gekozen de factoren aan
te duiden waarbij ofwel significantie ofwel trend tot significantie werd gevonden. Het
opnemen van de trends was een bewuste keuze omdat de kans op significantie groot is bij
onderzoek met een groter aantal proefpersonen.
Ten eerste was het doel van dit onderzoek om de schema’s voor de duurloop- en
sprintprestatie te justifiëren. Hierbij was het de bedoeling de correctheid na te gaan van de
prestatiebepalende factoren en hun onderlinge relaties. Zoals uit de schema’s blijkt,
werden enkel correlaties met de beste prestatie teruggevonden bij de factor
spiermassaverdeling. Bij de sprinters kan geconcludeerd worden dat het volume van de
M. rectus femoris een prestatiebepalende factor is waarbij een groter volume zorgt voor
een betere prestatie. Wat de duurlopers betreft werden op één trend na geen opmerkelijke

Discussie
75
correlaties gevonden. In totaliteit werden dus weinig factoren gevonden die correleren
met de prestatie, zowel bij duurlopers als bij sprinters.
Vervolgens was een tweede opzet van dit onderzoek om na te gaan of de onderzochte
factoren een atleet konden classificeren als typische duurloper of eerder als typische
sprinter. Zowel voor spierdikte als spiervolume werden lagere waarden gevonden bij de
duurlopers. De fascicle lengte bij duurlopers ligt lager dan bij sprinters. Voor de
duurlopers zou een te grote fascicle lengte voor een te grote metabole kost zorgen. Bij de
sprinters daarentegen laat een langere fascicle lengte toe om te werken op een lagere
verkortingssnelheid waardoor de krachtproductie wordt bevorderd. Een laatste verschil
tussen duurlopers en sprinters is eveneens de meest uitgesproken en de meest beschreven,
namelijk de carnosineconcentratie. Zo blijken sprinters een hogere carnosineconcentratie
te bezitten dan duurlopers. De grootste verschillen blijken zich dus te situeren ter hoogte
van de spiermassaverdeling en de spiereigenschappen. Dit werd schematisch
weergegeven in figuur 39 en 40.
Tot slot rijst de vraag of de gevonden verschillen te wijten zijn aan training of eerder
een genetische oorsprong hebben. In dit onderzoek werden atleten onderzocht van
Belgisch subtop- en topniveau. Geen enkele van deze atleten heeft medaillekansen op een
Europees of Wereldkampioenschap. Om op dit Belgische niveau te kunnen meedraaien,
zou het kunnen dat vooral goede trainingsmethoden een grote rol spelen en genetica
eerder van ondergeschikt belang is. Als echter gekeken wordt naar de wereldtop, worden
de sprintnummers duidelijk gedomineerd door atleten van West-Afrikaanse oorsprong.
Duuratleten op wereldniveau zijn voornamelijk Oost-Afrikanen en heel dikwijls
Kenianen. Aangezien de trainingsmethoden in Afrika waarschijnlijk niet beter zijn dan
deze in Europa, kan gesuggereerd worden dat naast een goede trainingsmethode
morfologische erfelijkheid van de onderste ledematen op dit niveau een doorslaggevende
rol speelt.
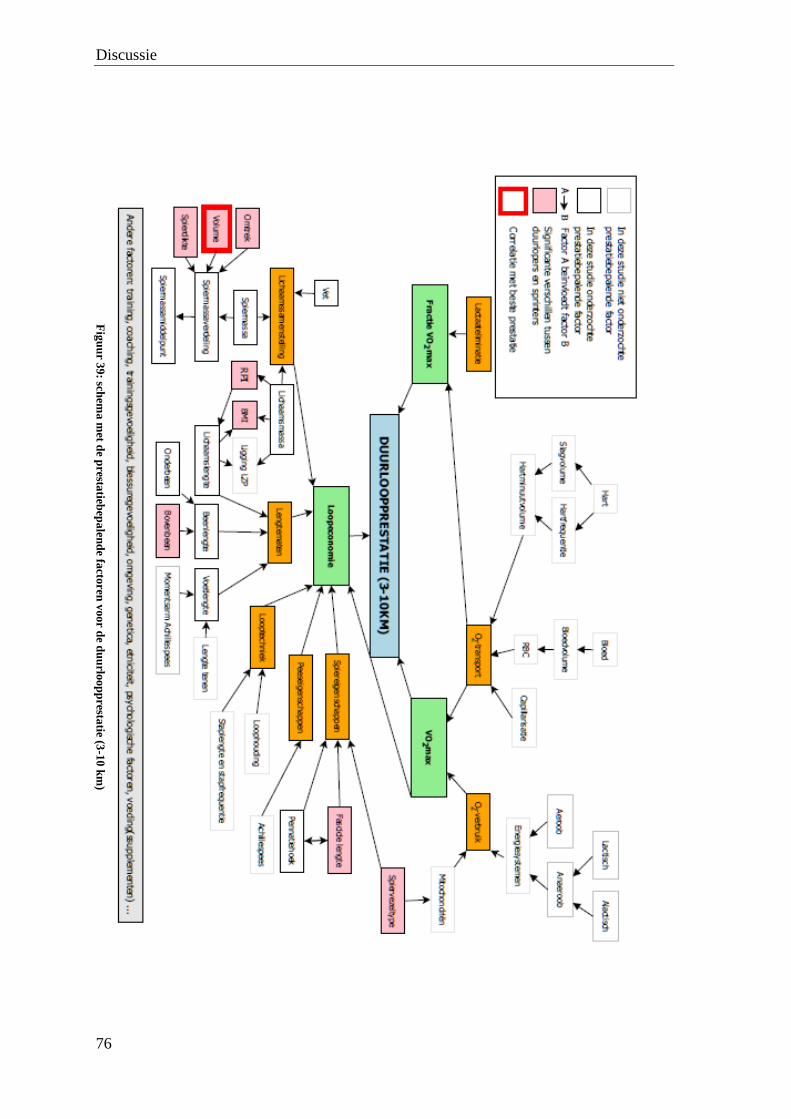
Discussie
76
Fig
uu
r 39
: schem
a m
et de p
restatieb
epa
lend
e facto
ren v
oo
r de d
uu
rloo
pp
restatie (3
-10
km
)
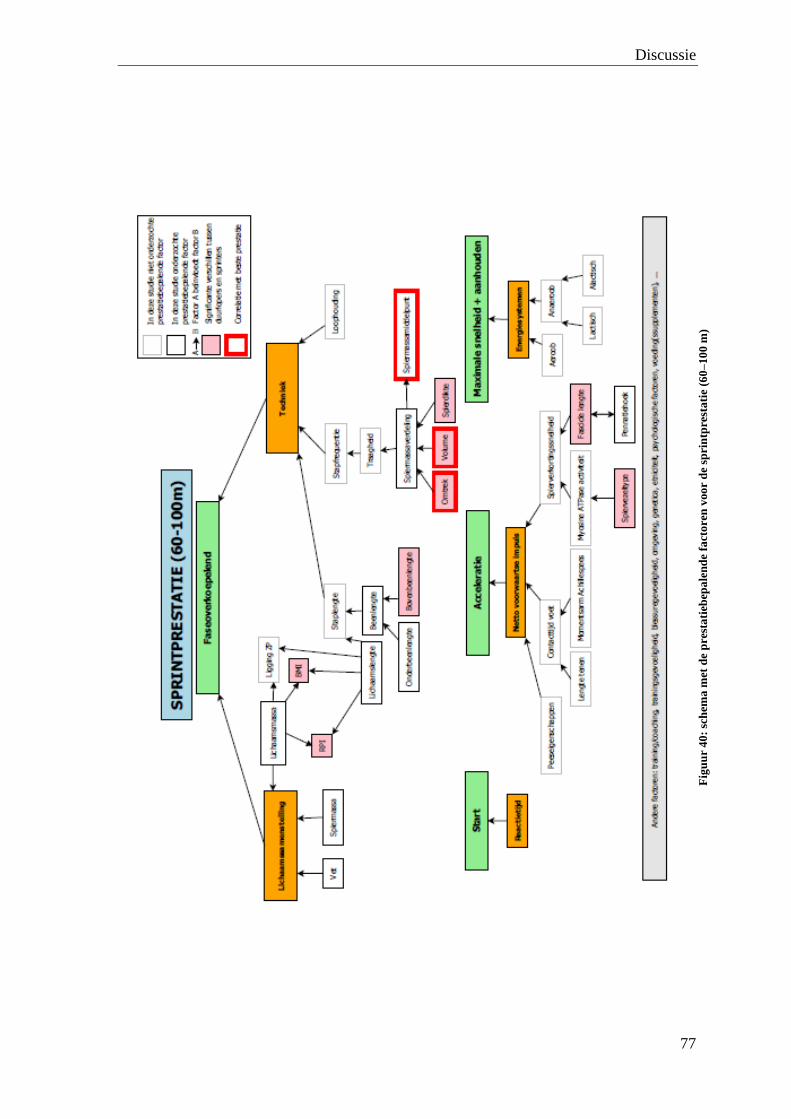
Discussie
77
Fig
uu
r 4
0:
sch
ema
met
de
pre
sta
tieb
epa
len
de
fact
ore
n v
oo
r d
e sp
rin
tpre
sta
tie
(60
–10
0 m
)

78

79
V. BIBLIOGRAFIE

Bibliografie
80
Abe, T. (2002). Fascicle length of gastrocnemius muscles in monozygous twins. Journal
of Physiological Anthropology, 21(6), 291-295
Abe, T., Brown, J.B. and Brechue, W.F. (1999). Architectural characteristics of muscle in
black and white college football players. Medicine & Science in Sports & Exercise,
31(10), 1448-1452
Abe, T., Fukashiro, S., Harada, Y. and Kawamoto, K. (2001). Relationship between sprint
performance and muscle fascicle length in female sprinters. Journal of Physiological
Anthropology, 20 (2), 141-147
Abe, T., Kumagai, K. and Brechue, W.F. (2000). Fascicle length of leg muscles is greater
in sprinters than distance runners. Medicine & Science in Sports & Exercise, 32(6), 1125-
1129
Adams, G.R., Foley, J.M. and Meyer, R.A. (1990). Muscle buffer capacity estimated
from pH changes during rest-to-work transitions. Journal of Applied Physiology, 69, 968-
972
Adams, G.R., Foley, J.M. and Meyer, R.A. (1990). Muscle buffer capacity estimated
from pH changes during rest-to-work transitions. Journal of Applied Physiology, 69, 968-
972
Ama, P.F., Simoneau, J.A., Boulay, M.R., Serresse, O., Theriault, J. and Bouchard, C.
(1986). Skeletal muscle characteristics in sedentary black and Caucasian males. Journal
of Applied Physiology, 61, 1758-1761
Baguet, A., Bourgois, J., Vanhee, L., Achten, E. and Derave, W. (2010) Important role of
muscle carnosine in rowing performance. Journal of Applied Physiology, 109, 1096–1101
Baguet, A., Everaert, I., Hespel, P., Petrovic, M., Achten, E. and Derave, W. (2011). A
new method for non-invasive estimation of human muscle fiber type composition. Plos
one, 6(7), 1-6
Baxter, J.R., Novack, T.A., Van Werkhoven, H., Pennell, D.R. and Piazza, S.J. (2012).
Ankle joint mechanics and foot proportions differ between human sprinters and non-
sprinters. Proceedings of the Royal Society, 279, 2018-2024

Bibliografie
81
Bejan, A., Jones, E.C. and Charles, J.D. (2010). The evolution of speed and athletics: why
the fastest runners are black and swimmers white. International Journal of Design &
Nature, 5(0), 1-13
Beneke, R. and Taylor, M.J.D. (2010). What gives Bolt the edge – A.V. Hill knew it
already! Journal of Biomechanics, 43, 2241-2243
Bernús, G., González de Suso, J.M., Alonso, J., Martin, P.A., Prat, J.A. and Arús, C.
(1993). 31P-MRS of quadriceps reveals quantitative differences between sprinters and
long-distance runners Medicine and Science in Sports and Exercise, 25(4), 479-484
Bogdanis, G.C., Nevill, M.E., Lakomy, H.K. and Boobis, L.H. (1998). Power output and
muscle metabolism during and following recovery from 10 and 20s of maximal sprint
exercise in humans. Acta Physiologica Scandinavica, 163, 261-272
Boobis, L.H., Williams, C. and Wooton, S.A. (1983). Influence of sprint training on
muscle metabolism during brief maximal exercise in man. Journal of Physiology, 342,
36-37
Buchthal, F. and Schmalbruch, H. (1969). Spectrum of contraction times of different fibre
bundles in the brachial biceps and triceps muscles of man. Nature, 89, 670-672
Charles, J.D. and Bejan, A. (2009). The evolution of speed, size and shape in modern
athletics. The Journal of Experimental Biology, 212, 2419-2425
Conley, D.L. and Krahenbuhl, G.S. (1980). Running economy and distance running
performance of highly trained athletes. Medicine and Science in Sports and Exercise,
12(5), 357-360
Costill, D.L., Daniels, J., Evans, W., Fink, W., Krahenbuhl, G. and Saltin, B. (1976).
Skeletal muscle enzymes and fiber composition in male and female track athletes.
Journal of Apllied Physiology, 40, 149-154
Costill, D.L., Fink, W.J. and Pollock M.L. (1976). Muscle fiber composition and enzyme
activities of elite distance runners. Medicine and Science in Sports, 8(2), 96-100
Costill, D.L., Thomason, H. and Roberts, E. (1973). Fractional utilization of the aerobic
capacity during distance running. Medicine and Science in Sports, 5(4), 248-252

Bibliografie
82
Crowther, G.J., Jubrias, S.A., Gronka, R.K. and Conley, K.E. (2002). A "functional
biopsy" of muscle properties in sprinters and distance runners. Medicine and Science in
Sports and Exercice, 34(11), 1719-1724
Dawson, B., Fitzsimons, M., Green, S., Goodman, C., Carey, M. and Cole, K. (1998).
Changes in performance, muscle metabolites, enzymes and fibre types after short sprint
training. European Journal of Applied Physiology and Occupational Physiology, 78, 163-
9
Eriksson, B.O., Gollnick, P.D. and Saltin, B. (1973). Muscle metabolism and enzyme
activities after training in boys 11-13 years old. Acta Physiologica Scandinavica, 87(4),
485–497
Gollnick, P.D and Matoba, H. (1984). The muscle fiber composition of skeletal muscle as
a predictor of athletic success. An overview. The American Journal of Sports Medicine,
12(3), 212-217
Gollnick, P.D., Oakley, C.R., Bayly, W.M., Allen, J.R. and Matoba, H. (1985).
Comparison of fiber types in skeletal muscles from ten animal species based on
sensitivity of the myofibrillar actomyosin ATPase to acid or copper. Histochemistry,
82(2), 175-183
Gollnick, P.D., Armstrong, R.B., Saltin B., Saubert C.W. IV, Sembrowich, W.L. and
Shepherd, R.E. Effect of training on enzyme activity and fibre composition of human
skeletal muscle. Journal of Applied Physiology, 34, 107-111
Gollnick, P.D., Armstrong, R.B., Saubert, C.W. IV, Piehl, K. and Saltin, B. (1972).
Enzyme activity and fiber composition in skeletal muscle of untrained and trained men.
Journal of Applied Physiology, 33(3), 312-319
Hautier, C.A., Wouassi, D., Arsac, L.M., Bitanga, E., Thiriet, P. and Lacour, J.R. (1994).
Relationships between postcompetition blood lactate concentration and average running
velocity over 100m and 200m races. European Journal of Applied Physiology and
Occupational Physiology, 68(6), 508-513
Henriksson, J. and Reitman, J.S. (1976). Quantitative measures of enzyme activities in
type I and type II muscle fibres of man after training. Acta Physiologica Scandinavica,
97(3), 392-397

Bibliografie
83
Hirvonen, J., Rehunen, S., Rusko, H. and Härkonen, M. (1987). Breakdown of high
energy phosphate compounds and lactate accumalation duringshort supramaximal
exercise. European Journal of Applied Physiology and Occupational Physiology, 56(3),
253-259
Jacobs, I., Esbjörnsson, M., Sylvén, C., Holm, I. and Jansson, E. (1987). Sprint training
effects on muscle myoglobin, enzymes, fiber types and blood lactate. Medicine and
Science in Sports and Exercise, 19(4), 368-374
Joyner, M.J. and Coyle, E.F. (2008). Endurance exercise performance: the physiology of
champions. Journal of Physiology, 586(1), 35-44
Karamanidis, K., Albracht, K., Braunstein, B., Catala, M.M., Goldmann, J.P. and
Brüggemann, G.P. (2011). Lower leg musculoskeletal geometry and sprint performance.
Gait & Posture, 34, 138-141
Kubo, K., Ikebukuro, T., Yata, H., Tomita, M. and Okada, M. (2011). Morphological and
mechanical properties of muscle and tendon in highly trained sprinters. Journal of
Applied Biomechanics, 27, 336-344
Kumagai, K., Abe, T., Brechue, W.F., Ryushi, T., Takano, S. and Mizuno, M. (2000).
Sprint performance is related to muscle fascicle length in male 100-m sprinters. Journal
of Applied Physiology, 88, 811-816
Kuno, S., Katsuta, S., Inouye, T., Anno, I., Matsumoto, K. and Akisada, M. (1988).
Relationship between MR relaxation time and muscle fiber composition. Radiology,
169(2), 567-568
Larsen, B. (2003). Kenyan dominance in distance running. Comparative Biochemistry
and Physiology, 136, 161-170
Lee, S.S.M. and Piazza, S.J. (2009). Built for speed: musculoskeletal structure and
sprinting ability. The Journal of Experimental Biology, 212, 3700-3707
Linnosier, M.T., Domois, D., Perier, C., Frey, J., Geyssant, A. and Denis, C. (1997).
Enzyme adaptations of human skeletal muscle during bicycle short-sprint training and
detraining. Acta Physiologica Scandinavica, 161(4), 439-445

Bibliografie
84
Locatelli, E. and Arsac, L. (1995). The mechanics and energetic of the 100m sprint. New
Stud Athletics, 10(1), 81-87
Maganaris, C.N., Baltzopoulos, V., Sargeant A.J. (1998). In vivo measurements of the
triceps surae complex architecture in man: implications for muscle function. Journal of
Physiology, 512(2), 603-614
McCully, K.K., Vandenborne, K., DeMeirleir, K., Posner, J.D. and Leigh, J.S Jr. (1992).
Muscle metabolism in track athletes, using 31P magnetic resonance spectroscopy.
Canadian Journal of Physiology and Pharmacology, 70(10), 1353-1359
Nygaard, E. and Goricke, T. (1976). Morphological studies of skeletal muscles in women.
August Krogh Institute
Parra, J., Cadefeau, J.A., Rodas, G., Amigo, N. and Cusso, R. (2000). The distribution of
rest periods affects performance and adaptations of energy metabolism induced by high-
intensity training in human muscle. Acta Physiologica Scandinavica, 169, 157-165
Pitsiladis, Y.P. and Scott, R. (2005). The makings of the perfect athlete. Medicine and
sport. 366, 16-17
Rolian, C., Lieberman, D.E., Hamill, J., Scott, J.W. and Werbel, W. (2008). Walking,
running and the evolution of short toes in humans. The Journal of Experimental Biology,
212, 713-721
Ross, A. and Leveritt, M. (2001). Long-term metabolic and skeletal muscle adaptations to
short-sprint training: implications for sprint training and tapering. Sports Medicine,
31(15), 1063-1082
Saltin, B., Henriksson, J., Nygaard, E, Andersen, P. and Jansson, E. (1977). Fiber types
and metabolic potentials of skeletal muscles in sedentary man and endurance runners.
Annals of the New York Academy of Sciences, 301, 3-29
Saunders, P.U., Pyne, B., Telford, R.D. and Hawley, J.A. (2004). Factors affecting
running economy in trained distance runners. Sports Med, 34(7), 465-485
Scholz, M.N., Bobbert, M.F., van Soest, A.J., Clark, J.R. and van Heerden, J. (2008).
Running biomechanics: shorter heels, better economy. The Journal of Experimental
Biology, 211, 3266-3271

Bibliografie
85
Scott, S.H., Engstrom, C.M. and Loeb, G.E. (1993). Morphometry of human thigh
muscles. Determination of fascicle architecture by magnetic resonance imaging. Journal
of Anatomy, 182, 249-257
Spiriev, B. and Spiriev, A. (2011). IAAF scoring tables of athletics.
Thorstensson, A., Larsson, L., Tesch, P. and Karlsson, J. (1977). Muscle strength and
fiber composition in athletes and sedentary men. Medicine and Science in Sports, 9(1),
26-30
Thorstensson, A., Sjodin, B. and Karlsson, J. (1975). Enzyme activities and muscle
strength after ‘sprint training’ in man. Acta Physiologica Scandinavica, 94(3), 313-318
Uth, N. (2005). Anthropometric comparison of world-class sprinters and normal
population. Journal of Sports Science and Medicine, 4, 608-616
Van Damme, R., Wilson, R.S, Vanhooydonck, B. and Aerts, P. (2002). Performance
constraints in decathletes. Nature, 415, 755-756
Vucetic, V., Babic, V., Sentija, D. and Nekic, B. (2005). Anthropometric and
morphological characteristics of runners. Science and profession – challenge for the
future, 612-615
Watts, A.S., Coleman, I. and Nevill, A. (2012). The changing shape characteristics
associated with success in world-class sprinters. Journal of Sports Sciences, 30(11), 1085-
1095
Weyand, P.G. and Davis, J.A. (2005). Running performance has a structural basis. The
Journal of Experimental Biology, 208, 2625-2631

86

87
VI. BIJLAGEN

Bijlagen
88
Bijlage 1: vragenlijst voor de atleet
Gegevens:
Naam:
Geboortedatum:
Gewicht:
Lengte:
Trainingsinfo:
Persoonlijke records (= jaartal):
Aantal jaar training:
Disciplineshift? + jaartal:
2011-12 actief atleet of gekwetst:
Aantal trainingen per week/dag:
Hoeveel daarvan snelheid: kracht: uithouding :
Welk soort training heb je gisteren en/of vandaag gedaan?

Bijlagen
89
Dieet:
Voedingssupplementen:
Welke voedingssupplementen nam je de laatste 3 maand?
Heb je ooit creatine genomen? Wanneer, hoeveel, hoelang?
Heb je ooit beta-alanine genomen? Wanneer, hoeveel, hoelang?
Heb je ooit carnosine genomen? Wanneer, hoeveel, hoelang?
Voedingspatroon:
Volg je een specifiek dieet? Zoja, welk?
Ben je vegetarisch of niet?
Eet je zowel rood vlees als kip en kalkoen?
Hoeveel keer per week gemiddeld eet je kip/ kalkoen?
Hoeveel vlees eet je ongeveer per dag (aantal gram?)


















