Web viewPunctated equilibrum. Ontbrekende fossielen. 29 oktober 2013 Jente Ottenburghs 2. Fossielen...
37
Punctated equilibrum Ontbrekende fossielen 29 oktober 2013 Jente Ottenburghs 2 Fossielen worden voortdurend aangehaald als bewijs voor de evolutietheorie. Maar in de ogen van Charles Darwin vormden ze tegelijkertijd een uitdaging voor zijn theorie. Want waarom zijn er zo weinig overgangsvormen die de trage en gelijkmatige werking van natuurlijke selectie illustreren, ontdekt? Darwin loste dat vraagstuk op door te stellen dat het fossielenbestand incompleet is. Maar klopt dat wel? Gebaseerd op klassieke Darwiniaanse processen (mutatie, natuurlijke selectie en genetische drift) formuleerden paleontologen een model om de oorsprong van taxa te verklaren: het fyletisch gradualisme. In dit model ontstaan nieuwe soorten door de trage transformatie van een volledige populatie in een andere door geleidelijke veranderingen
Transcript of Web viewPunctated equilibrum. Ontbrekende fossielen. 29 oktober 2013 Jente Ottenburghs 2. Fossielen...

Punctated equilibrum
Ontbrekende fossielen
29 oktober 2013 Jente Ottenburghs 2
Fossielen worden voortdurend aangehaald als bewijs voor de evolutietheorie. Maar in de ogen van Charles Darwin vormden ze tegelijkertijd een uitdaging voor zijn theorie. Want waarom zijn er zo weinig overgangsvormen die de trage en gelijkmatige werking van natuurlijke selectie illustreren, ontdekt?
Darwin loste dat vraagstuk op door te stellen dat het fossielenbestand incompleet is. Maar klopt dat wel?
Gebaseerd op klassieke Darwiniaanse processen (mutatie, natuurlijke selectie en genetische drift) formuleerden paleontologen een model om de oorsprong van taxa te verklaren: het fyletisch gradualisme.
In dit model ontstaan nieuwe soorten door de trage transformatie van een volledige populatie in een andere door geleidelijke veranderingen (anagenese). In combinatie met cladogenesis (het splitsen van evolutionaire lijnen) verklaart dit evolutionaire model de diversiteit van het leven op aarde.

Twee voorspellingenDeze theorie doet twee voorspellingen: ten eerste, het fossielenbestand voor de oorsprong van nieuwe soorten bestaat uit een lange reeks continue, langzaam veranderende tussenvormen die een afstammeling verbinden met zijn voorouder. En ten tweede, morfologische breuken in deze reeksen zijn het gevolg van imperfecties in het fossielenbestand.
De twee theorieën: fyletisch gradualisme en punctuated equilibria. Afbeelding: Miguel Chavez (via Wikimedia Commons).
Het Alternatief: Punctuated Equilibria
In 1972 stelden Stephen Jay Gould en Niles Eldredge een nieuw model voor:punctuated equilibria (onderbroken evenwichten). Deze theorie geeft aan dat de meeste soorten geen grote evolutionaire veranderingen ondergaan tijdens hun geologische geschiedenis. Maar zulke periodes van stasis worden onderbroken door onverwachte gebeurtenissen van snelle evolutionaire verandering (op een geologische tijdschaal) waarin nieuwe soorten ontstaan. De gaten die we in het fossielenbestand zien, zijn dus geen ontbrekende fossielen, maar het gevolg van periodes met snelle evolutionaire veranderingen.
Waarschijnlijk zijn beide theorieën complementair ,
er is bewijs gevonden voor zowel fyletisch gradualisme als voor punctuated equilibria. Ze sluiten elkaar ook niet volledig uit. Het is perfect mogelijk dat bepaalde taxa zich ontwikkeld hebben volgens het ene model en andere taxa volgens het andere model. Malmgren en collega’s combineerden beide theorieën zelfs (het zogenaamde punctuated gradualisme) om de evolutie van planktonische foraminiferen te verklaren.

Er zijn echter wetenschappers die het fyletisch gradualisme als enige juiste model zien. Zo blijven evolutiebioloog Richard Dawkins en filosoof Daniel Dennett kritisch ten opzichte vanpunctuated equilibria. Verder onderzoek en nieuwe fossielen zullen hopelijk uitsluitsel geven in dit debat.Jente Ottenburghs (1988) heeft sinds zijn Master Evolutie en Gedragsbiologie aan de Universiteit van Antwerpen een brede interesse voor evolutionaire biologie. Sinds mei 2012 werkt hij als PhD-student bij de Resource Ecology Group aan de Universiteit van Wageningen. Meer informatie over zijn onderzoek vindt u hier.
Bronmateriaal:Darwin, C. (1859). On the Origin of Species. London: John Murray.Eldredge, N. & S. J. Gould (1972). Punctuated equilibria: an alternative to phyletic gradualism. In T.J.M. Schopf, ed., Models in Paleobiology. San Francisco: Freeman Cooper. pp. 82-115.Dawkins, R. (1996). The Blind Watchmaker. New York: W. W. Norton & Co..Dennett, D. (1995). Darwin's Dangerous Idea. New York: Simon & SchusterMalmgren, B. A., Berggren, W. A., & Lohmann, G. P. (1983). Evidence for punctuated gradualism in the late Neogene Globorotalia tumida lineage of planktonic foraminifera .GeoScienceWorld, 9(3), 377-389.De foto bovenaan dit artikel is gemaakt door Mike Beauregard (via Wikimedia Commons).
Punctuated Equilibrium is een ondertheorie van de evolutietheorie die een verklaring geeft voor plotselinge sprongetjes in evolutionaire lijnen van fossiele soorten, zoals we die kunnen volgen in de geologische kolom.
Er bestaat niet echt een Nederlandse vertaling van dit begrip, maar de aanduiding 'onderbroken evenwichten', wordt wel eens gebruikt, en de term 'punctualisme' voor de wetenschappelijke stroming die het omvat.
Korte beschrijving
Het idee achter punctuated equilibrium is dat soorten gedurende lange tijd geen of nauwelijks evolutionaire verandering kennen (een evenwicht of equilibrium), en dan gedurende geologisch korte tijd (bijvoorbeeld enkele duizenden tot honderdduizenden jaren) relatief sterk wijzigen (een onderbreking of punctuation).
Volgens de theorie zouden overgangen tussen soorten dus zeer zeldzaam zijn. Het heeft niets te maken met fossiele overgangsvormen tussen grotere categorie 챘 n organismen, maar alleen de evolutie binnen bepaalde soorten of geslachten.
Op hogere niveaus zijn er ook volgens het punctualisme tussenvormen.
Er heeft paleontologisch onderzoek plaatsgevonden om te kijken of punctualisme dan wel gradualisme (dat zegt dat soorten voortdurend, doch zeer langzaam veranderen) correct is.

Van beide modellen zijn voorbeelden gevonden die ze bevestigen.
De algemeen aanvaarde conclusie is daarom dat beide vormen van evolutie voorkomen.Een interessant aspect van punctualisme is dat de natuurlijke selectie die in de klassieke evolutietheorie als het ware als de motor van de evolutie fungeert, hier eerder als een rem gezien wordt.
Zolang de omgeving niet verandert, zo redeneert het punctualisme, zorgt de selectie ervoor dat soorten niet (meer) veranderen.
Zodra er echter een grote ecologische verandering plaatsvindt (bijvoorbeeld een ramp veroorzaakt door de inslag van een grote meteoriet) is de rem los en kunnen soorten in vrij korte tijd sterk veranderen, omdat mutaties niet meer onmiddellijk uitgewied worden.
Daarna volgt een periode van stabilisatie waar de selectie-rem weer in werking treedt.
Externe links
Punctuated equilibria: an alternative to phyletic gradualism. Niles Eldredge en Stephen Jay Gould, 1972. In Models in Paleobiology, San Francisco: Freeman, Cooper and Company, pp. 82-115. (pdf)
http://nl.wikipedia.org/wiki/Punctuated_equilibrium
Evolutiemodellenhttp://www.natuurinformatie.nl/asp/page.asp?alias=nnm.dossiers&id=i001385&view=natuurdatabase.nl
Volgens Darwins theorie gaat de evolutie met hele kleine stapjes. Dit noemen we het gradualistische model. Het model heeft veel kritiek gekregen, omdat de oorsprong van ingewikkelde organen zoals het oog moeilijk voorstelbaar is in kleine stapjes. Er zijn dan ook andere modellen bedacht.
Micro- en macro-evolutie
Naast kleine verschillen binnen een soort zien we grotere verschillen tussen soorten, nog grotere tussen geslachten, enzovoort. Voor sommigen was dat reden om aan te nemen dat er verschillende processen een rol spelen. Er zouden veranderingen zijn die niet leiden tot nieuwe soorten, ook wel microevolutie genoemd, en veranderingen die wel leiden tot nieuwe soorten, nieuwe kenmerken en bouwplannen, macro-evolutie genoemd. Vooral voor het ontstaan van grote verschillen en ingewikkelde structuren, zoals ogen, waarvan men zich het nut niet kan voorstellen in een opbouwfase, als het half af is, werden 'sprongmutaties' bedacht. De geneticus Goldschmidt dacht dat er nu en dan een hopeful monster ontstond, een volkomen nieuw type dat op een of andere manier toch in leven bleef. Hij vertelde er niet bij waar dit arme monster dan een partner kon vinden. Bovendien zouden dit soort monsters dan ook nu nog rond moeten lopen.
Punctuated equilibrium
Het idee van sprongsgewijze in plaats van graduele veranderingen lijkt ondersteund te worden door de fossielen. Hier zien we vaak soorten lange tijd nauwelijks veranderen, om er dan plotseling anders

uit te gaan zien, waarna ze weer lange tijd gelijk blijven. Het ontbreken van zulke schakels (missing links) tussen de typen was al lang bekend en werd toegeschreven aan de onvolledigheid van de fossielen. De kans dat een soort fossiliseert, is immers erg klein en afhankelijk van allerlei verschillende factoren. In 1972 bedachten de Amerikaanse paleontologen Eldredge en Gould dat de fossiele overlevering misschien helemaal niet zo onvolledig was, maar dat de soorten in zo korte tijd sterk veranderden dat de kans op het vinden van fossielen van die zogenaamde tussenvormen erg klein was. Er waren er simpelweg te weinig van. Zij noemden dit verschijnsel punctuated equilibrium, onderbroken evenwicht.
Langzame en snelle evolutie
Als ondersteuning van het model van punctuated equilibrium is wel aangehaald, dat het overeenkomt met wat men waarneemt als een populatie van weinig individuen plotseling explosief groeit: in korte tijd treden veel veranderingen op. We spreken dan echter wel over een heel andere tijdschaal. Een populatie kan in tien, vijftien jaar sterk veranderen. Bij punctuated equilibrium moeten we volgens Gould toch denken aan 챕챕 n procent of minder van de totale leefduur van een soort. Als een soort gemiddeld tien miljoen jaar zou bestaan, zou het bij punctuated equilibrium om een veranderingsperiode van 100.000 jaar gaan. Het is dan ook de vraag of het zinnig is deze vergelijking te maken. Een ander probleem bij dit model is, dat het vaak moeilijk is vast te stellen of opeenvolgende fossiele typen tot dezelfde ontwikkelingslijn behoren of onafhankelijke lijnen vertegenwoordigen.
Grote en kleine verschillen
Als we praten over grote en kleine verschillen, gaat het om subjectieve waarneming. Stel, je hebt een plantensoort met twee rassen: één met rode en één met gele bloemen. Er is een verwante soort met blauwe bloemen. Is het verschil tussen rood en geel kleiner dan tussen rood en blauw? Wellicht is het zinvoller door te dringen tot de kern van de verschillen, namelijk het DNA. Een maat voor verschillen zou het aantal verschillen in organische basen kunnen zijn. Maar ook dit geeft geen uitsluitsel, want basen kunnen meerdere malen vervangen worden zonder dat het later is terug te zien aan het DNA. Het lijkt het beste het hele verhitte debat over snelheid van verandering en grootte van verschillen voorlopig te laten voor wat het is en de aandacht te richten op de vraag: hoe ontstaan in het DNA nieuwe eigenschappen?
Punctuated equilibriumhttp://www.natuurinformatie.nl/nnm.dossiers/natuurdatabase.nl/i000653.html
Sinds Darwin gingen onderzoekers ervan uit dat evolutie een geleidelijk proces was. Dat dit proces niet altijd even snel verliep, was te zien aan levende fossielen, die gedurende miljoenen jaren onveranderd bleken te zijn. In de praktijk heeft men in sommige gevallen aan de hand van fossielen geleidelijke evolutie kunnen aantonen. Toch kleefde er aan dit idee een probleem. In reeksen fossielen die een bepaalde ontwikkeling van een groep te zien gaven, ontbraken vaak tussenvormen of 'missing links'. Men dacht dat dit kwam door de onvolledigheid van fossiele vondsten.
In 1972 kwamen de Amerikaanse fossielendeskundigen (paleontologen) Eldredge en Gould met het idee, dat de fossielen toch wel een goed beeld gaven, maar dat de evolutie met sprongen gebeurde.
Lange perioden waarin groepen min of meer gelijk bleven, wisselden zich af met korte perioden waarin er wel veranderingen optraden. Er is dus een evenwicht (equilibrium) dat af en toe verstoord (punctuated - onderbroken) wordt. Dit verschijnsel heet punctuated equilibrium.

Omdat de veranderingen zo snel gebeuren, is de kans op fossilisatie van tussenvormen gering en zullen deze niet of nauwelijks gevonden worden. Overigens moet men zich wel realiseren dat er bij een 'snelle verandering' toch altijd nog miljoenen jaren verstrijken. Een goed voorbeeld is de enorme toename in diversiteit van het zeeleven in het begin van het Cambrium, ongeveer 540 miljoen jaar geleden (de 'Cambrische explosie').
In een relatief korte periode van 5 tot 10 miljoen jaar, ontstonden alle hoofdgroepen van ongewervelde dieren, inclusief de voorloper van de gewervelde dieren. Er waren zelfs een aantal hoofdgroepen die later weer uitgestorven zijn. In de daarop volgende 550 miljoen jaar is er op die schaal eigenlijk weinig meer toegevoegd aan de diversiteit van het leven. De theorie van het 'punctuated equilibrium' lijkt in een aantal gevallen goed op te gaan.
Hoe het proces van evolutie precies verloopt, zullen we nooit weten.
Stephen J. Gould was bij leven een man die graag knuppels in het hoenderhok gooide. (Hij overleed in 2002 op zestigjarige leeftijd). Hij wist niet iedereen van zijn denkbeelden te overtuigen maar hij bracht wel veel (vruchtbare) discussies op gang. Een van zijn opzienbarende idee 챘 n betrof de snelheid waarmee evolutie plaatsvond. Darwin ging er van uit dat soortvorming een heel geleidelijk proces was dat honderdduizenden of miljoenen jaren in beslag nam. Aan de hand van een prachtig onderzoek aan populaties van trilobieten toonde hij, samen met N. Eldredge, aan dat evolutie ook zeer snel kan verlopen. Hij kwam met de theorie van het 'verbroken evenwicht' (punctuated equilibrium), die stelde dat soorten door een verstoring van het evenwicht in korte tijd ontstaan en daarna lange tijd vrijwel onveranderd blijven.

Meer weten?Biografie van Stephen J. Gould
Skelton: Evolution (1993)
In het schema hierboven stelt A het model van Darwin voor. Dit heet wel 'gradualisme' en daarin gaat alles heel geleidelijk en langzaam.C is het model van Gould (punctuated equilibrium). Daarin verandert een soort lange tijd niet, totdat ten gevolge van een verstoring van de leefomgeving in korte tijd zulke veranderingen optreden dat er een nieuwe soort ontstaat. Dit snelle ontstaan van soorten verklaart ook het ontbreken van fossiele tussenvormen, een probleem waar Darwin al mee worstelde. Goulds model kan voorkomen, maar soorten blijven alleen onveranderd als de omstandigheden heel stabiel zijn en dat komt niet vaak voor. Andere plant- en diersoorten oefenen meestal ook een druk uit, waardoor een soort verandert. Daarom benadert model B de werkelijkheid het meest: geleidelijke veranderingen en soms het in (vrij) korte tijd het ontstaan van een nieuwe soort.
Meer weten?Punctuated equilibrium (in het Nederlands)Evolutiemodellen
http://www.fossieleplanten.nl/Evolutie/evolu21.html
Creationisten en Steeman
De waarheid over 'Punctuated Equilibria' (Daarom : evolutie !)Fedor A. Steeman drs. Biologie
Creationisten wapperen vaak met citaten uit artikelen van Gould en Eldredge waarin zij hun theorie van Punctuated Equilibria uiteenzetten. Dit soort citaten moeten de indruk wekken dat zelfs gerespecteerde (en fel anti-creationistische) evolutionair-biologen toegeven dat het fossielenbestand waardeloos is m.b.t. de evolutie-theorie.
Ook wordt Gould en Eldredge's theorie vaak gemisinterpreteerd en als gevolg daarvan gekarikaturiseerd als een ongeloofwaardige theorie die uitgaat van absurde evolutionaire sprongen. Ik zal hieronder uiteenzetten waar deze misciteringen en misinterpretaties in feite op neerkomen.
Als je wat Gould en Eldredge met hun theorie willen zeggen in één zin zou kunnen samenvatten zou dat als volgt zijn:
"Het is een gegeven dat geleidelijke overgangen tussen de verschillende fossiele soorten niet of nauwelijks worden gevonden, maar daar is eigenlijk een hele logische (darwinistische) verklaring voor."
Zoals zij door creationisten vaak worden geciteerd doet overkomen dat ze het volgende willen zeggen:
"Het is een gegeven dat geleidelijke overgangen tussen de verschillende fossiele soorten niet of nauwelijks worden gevonden."

M.a.w. het laatste en meest essentiële gedeelte van hun betoog wordt weggelaten, waardoor je een vervorming krijgt van wat ze eigenlijk willen zeggen ! Daardoor wordt de indruk gewekt dat zij wijzen op iets wat niet in de evolutietheorie past, terwijl ze juist een mechanisme aangeven dat dit ontbreken van geleidelijke overgangen verklaart!
Welnu, is dit nog wel een eerlijke vorm van argumenteren van de kant van de creationisten? Ik zou zeggen van niet!
De verklaring voor het uitblijven van geleidelijke overgangen past uitstekend in de evolutie-theorie en vereist helemaal geen absurde gebeurtenissen zoals een kip die een ei legt waar een pauw uit komt. De verklaring is dat evolutie zich in zo'n beperkt deel van het verspreidingsgebied en/of in zo'n (geologisch gezien) korte tijd afspeelt dat het alleen maar logisch is dat de geleidelijke overgangen niet worden gevonden. Dit heeft te maken met de zogenaamde peripatrische (of allopatrische) speciatie, een model van soortsvorming waarbij de evolutie in een kleine, geïsoleerde populatie plaatsvindt, buiten de moederpopulatie.
Evolutie verloopt sneller in een kleine populatie, omdat nieuwe genen zich sneller kunnen verspreiden over de hele populatie. Bij herstelling van het contact met de moederpopulatie verspreiden ofwel de nieuwe eigenschappen (mits gunstig) zich door de populatie in relatief korte tijd, ofwel verdringt de door reproductieve isolatie nieuw ontstane vorm de voorouderlijke vorm in relatief korte tijd. Aangezien de evoluerende populatie zo klein en afgelegen is, is er maar een minieme kans is dat vertegenwoordigers hieruit in het fossielenbestand terechtkomen.
Dit verklaart waarom een soort lange tijd onveranderd en stabiel blijft en dan plotseling een 'sprongetje' lijkt te maken. Dit is natuurlijk geen echt sprongetje van generatie-op-generatie in één lijn, maar een nieuwe vorm die de oude vervangt binnen een klein aantal generaties.
Een andere denkfout van creationisten is dat zij deze 'sprongetjes' verwarren met het ontbreken van overgangsvormen. De sprongetjes vinden plaats tussen fossiele sequenties die goed gedocumenteerd zijn. Juist omdat ze goed gedocumenteerd zijn, kunnen we we waarnemen dat er om de

zoveel tijd een sprongetje plaatsvind. Deze sprongetjes vinden dus plaats binnen een enkele soort of geslacht en hebben niets te maken met het al of niet ontbreken van overgangsvormen tussen grotere diergroepen.
Als Gould en Eldredge dus zeggen dat geleidelijke overgangen ontbreken, doelen zij op het verschijnsel dat bepaalde, goed gedocumenteerde fossiele soorten/geslachten sprongetjes maken in hun evolutie, en niet dat er geen overgangsvormen bestaan tussen grotere groepen. Die laatste zijn er juist wel!
Punctuated Equilibra / Terrencevrijdag 27 augustus 2010
Links: Niles Eldredge, rechts: Stephen Jay Gould.
Ik kom steeds vaker mensen tegen op het internet die een bepaalde mening hebben over punctuated equilibria zonder dat ze begrijpen wat dit model inhoud of hoe deze tot stand kwam.
{(Laat) ik beschrijven wat punctuated equilibria inhoud. Het meeste komt uit de artikelen die Gould en Eldredge zelf schreven (ze waren goed in het uiteen zetten van hun idee, dus dit is alleen maar logisch, toch?).}
Het paleosoorten probleem. Een terugkerend onderwerp in de biologie is wat een soort eigenlijk is. De biologische soort (door Mayr) wordt vaak op deze manier gedefinieerd: een groep organismen waarbij de individuen onderling kunnen voortplanten in natuurlijke omstandigheden.
WAT IS EEN SOORT?
Nu heeft dat in de natuur al zijn limitaties (denk maar bijvoorbeeld aan aseksuele organismen), maar paleontologen hebben het helemaal moeilijk met hun fossielen.
Paleontologen kunnen bijna alleen gebruik maken van morfologische kenmerken. En alleen de kenmerken die fossileren (zachte onderdelen fossileren niet vaak) kunnen daarvoor worden gebruikt.
Het samen brengen van de concepten 'biologische soort' en 'paleosoort' leidt tot een eindeloze discussie. Echter is het concept van de biologische soort wel op een andere manier nuttig voor de paleontologie...

Hoe soortvorming vandaag de dag gaat er zijn 2 categorieën hoe een nieuwe soort ontstaat. De eerste is dat een gehele soort evolueert in een nieuwe soort (anagenese), de tweede is dat een soort (populatie) zich splitst in 2 populaties en uiteindelijk 2 soorten wordt (cladogenese).
Clade by John Wilkins at Evolving Thoughts
ANAGENESE
Wellicht zal anagenese genoeg voorkomen, maar daarmee krijg je niet de diversiteit van het leven. Alleen doordat evolutionaire lijnen zich splitsen (cladogenese) ontstaan er meer soorten waar er eerst 1 was. Cladogenese kwam dus in het verleden heel vaak voor....heel vaak.
De bekendste vorm van cladogenese is allopatrische soortvorming. Hierbij wordt de populatie gescheiden door een fysieke barrière. De 2 'nieuwe' populaties hebben een apart leefgebied en kunnen niet met elkaar in contact komen (er zit bijvoorbeeld een nieuwe rivier tussen). Beide groepen evolueren onafhankelijk van elkaar en het 'resultaat' is 2 nieuwe soorten. De grote van de 2 nieuwe populaties in het begin kan natuurlijk verschillen, maar des te kleiner de populatie, des te sneller evolutie plaats vind. Een andere vorm is peripatrische soortvorming. Hierbij migreert een relatief klein gedeelte van de populatie naar een nieuwe omgeving en leeft daar gescheiden van de oude populatie. De nieuwe populatie wordt een nieuwe soort en als ze bij elkaar komen dan kunnen ze zich niet onderling voortplanten. Sympatrische soortvorming komt veel minder voor en saltationele soortvorming (polyploïdie) komt bijna helemaal niet voor in dieren (wel vaak in planten).
POLYPLOIDIE
http://www.kennislink.nl/publicaties/soortvorming-terwijl-u-wacht(polyploidie)
Planten kruisingen ( polyploidie ) Polyploïdie Polyploidy
Polyploidy on Kimball's Biology Pages The polyploidy portal a community-editable project with information, research, education, and
a bibliography about polyploidy.
Allopatrie
Allopatry and sympatry by John Wilkins at Evolving ThoughtsSPECIATIE
Wat paleontologie kan leren van neontologie. Hoe soortvorming te werk gaat kan alleen worden bestudeert uit de levende wereld. Met geen mogelijkheid kun je een model opstellen door naar de fossielen te kijken. Modellen die men gebruikt in de paleontologie worden opgesteld in de levende wereld. Neontologie is de studie van de levende wezens en zo beargumenteren Gould en Eldredge dat paleontologie in het geval van soortvorming moet leren van de neontologie. Net zoals paleontologie altijd al leert van de neontologie. een voorbeeld: als je een fossielen schedel vind met kaken en je alleen de schedel bestudeert, kun je niet ontdekken wat de functie is van de kaken. Door levende wezens te bestuderen weet je wat de functie van kaken is en kun je die kennis toepassen op de fossielen. Er is een goed idee hoe soortvorming gaat in de levende natuur en daarmee kunnen voorspellingen worden gemaakt wat we zouden moeten zien in de aardlagen. Deze kunnen dan worden getest.
Een voorspelling voor allopatrische of peripatrische soortvorming in de aardlagen is dat een nieuwe soort niet ontstaat in hetzelfde gebied als de voorouder, maar er een eind vandaan. Het is dus bijna ondenkbaar om een bepaalde soort te zien splitsen in 2 soorten als we naar een lokaal (klein) gedeelte kijken van de aardlagen.

Een andere voorspelling is dat de morfologische verandering die in een afgescheiden populatie kort na het scheiden ontstaat of (in veel mindere mate) in de originele populatie aanwezig was. Deze morfologische eigenschappen komen in de originele populatie in minimale frequentie voor (zijn heel zeldzaam), maar kunnen in isolatie snel stijgen naar fixatie. Snelle verandering ontstaat dus tijdens het aanpassen aan de nieuwe omstandigheden en een sterkere invloed van genetische drift. Wanneer de soort stabiel is zal deze niet meer sneller veranderen als de oude soort waar de nieuwe soort vanaf is gesplitst.
Het begrip biologische soort is handig om soortvorming te bestuderen. En modellen van soortvorming kunnen dan toegepast worden op de fossielen. Dat is deel 1 in een notendop. In het volgende deel komen de sappige voorbeelden.
Species by John Wilkins at Evolving Thoughts
Punctuated Equilibria (2) - in de aardlagen.
maandag 27 september 2010
3
Afbeelding 1: links laat het patroon zien wat verwacht wordt van phyletic gradualism (graduele overgang) terwijl rechts het patroon laat zien wat verwacht wordt van punctuated equilibria. Dit geldt zo als je kijkt
naar de grote populatie.
In deel 1 kwam aan bod waarom Gould en Eldredge een nieuw model hadden over soortvorming voor de paleontologie. Paleontologie leert gewoonlijk van de neontologie (studie van de levende natuur, ten opzichte van paleontologie) en bij soortvorming zou dat niet anders moeten zijn. Cladogenese (en voornamelijk allopatrische en peripatrische soortvorming) is wat er in de natuur het meeste voor komt en wat in het verleden ook heel vaak moest gebeuren anders zouden we niet de enorme biodiversiteit krijgen die we nu hebben. Punctuated equilibria is dan dat een klein deel van de oorspronkelijke populatie afgesloten raakt, relatief snel evolueert en een nieuwe soort wordt. In dit deel komen aan bod: - de patronen in de aardlagen volgens punctuated equilibria - voorbeelden De patronen in de aardlagen volgens punctuated equilibria.

Afbeelding 2: mogelijke scenario's. A = blijvende isolatie, B = menging, C = overname door de nieuwe soort.
Als de nieuwe soort eenmaal is ontstaan zijn er 3 mogelijke scenario's belangrijk in de paleontologie (zie afbeelding 2). Scenario A is dat de twee soorten gescheiden blijven. Scenario B is dat beide soorten elkaar weer tegenkomen en naast elkaar leven (in verschillende niches). Scenario C is dat beide soorten elkaar weer tegenkomen, maar de nieuwe soort de oude soort verdringt (met het uitsterven van de oude soort als gevolg). Er zijn meer scenario's denkbaar (zoals dat de oude soort de nieuwe soort verdringt), maar deze zullen bijna nooit sporen achterlaten in de aardlagen. Wat zouden we moeten zien in de aardlagen van de meeste plekken? In het geval van scenario C zien we het patroon dat rechts afgebeeld is in afbeelding 1. In de eerste aardlagen heb je de ene soort en in de aardlagen erboven verschijnt plotseling de nieuwe soort. Scenario B is nagenoeg hetzelfde behalve dat de oude soort ook in de bovenste lagen te vinden is (naast de nieuwe soort). Er zijn echter plekken waar de overgang te vinden is (deze zijn zeldzaam) en waar wel overgangsfossielen te vinden zijn. Bovenaan in afbeelding 2 is het rondje dat van donkerblauw naar lichtblauw 'evolueert' geschikt om de tussenvormen te vinden. Dit betekend dat paleontologen niet zomaar één plek moeten onderzoeken, de kans is heel klein dat er dan overgangsvormen worden gevonden.
Voorbeelden. Voorbeelden uitwerken bleek moeilijker dan gedacht. Ik wilde er eerst een stuk of drie doen, maar het eerste voorbeeld duurt al zo lang om samen te vatten dat ik het daar maar bij houd. Mede hierdoor duurde het even voordat ik deze blog wilde plaatsen. Zo'n beetje het stokpaardje van Gould en Eldredge is Phacops rana, een trilobiet uit het Devoon (in dit tijdperk kwamen de gewervelden voor het eerst aan land, een mijlpaal). Het Devoon (416-359 miljoen jaar geleden) is weer onderverdeelt in verschillende

sudivisies. Tegenwoordig wordt bijna alleen nog de Europese onderverdeling gebruikt, maar vroeger was er nog een Noord-Amerikaanse onderverdeling. Gould en Eldredge gebruikten in hun grafieken de Noord-Amerikaanse onderverdeling, maar noemden wel de Europese variant als vergelijking (zie afbeelding 3).
Afbeelding 3: de Europese tegenover de Amerikaanse
onderverdeling. Uit Eldredge, 1971.
http://palaeo-electronica.org/2007_3/books/equal.htm
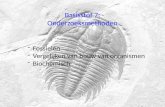
Figure 1
Species vs. Time: In this example, a snail species, A, is isolated by a geographic barrier to form subspecies X and Y. Eventually, this geographic separation leads to genetic isolation, and two distinct new species, B and C, form. © AMNH

http://www.amnh.org/learn/evolution/Resource1
http://www.science20.com/between_death_and_data/missing_link_fallacy-75791
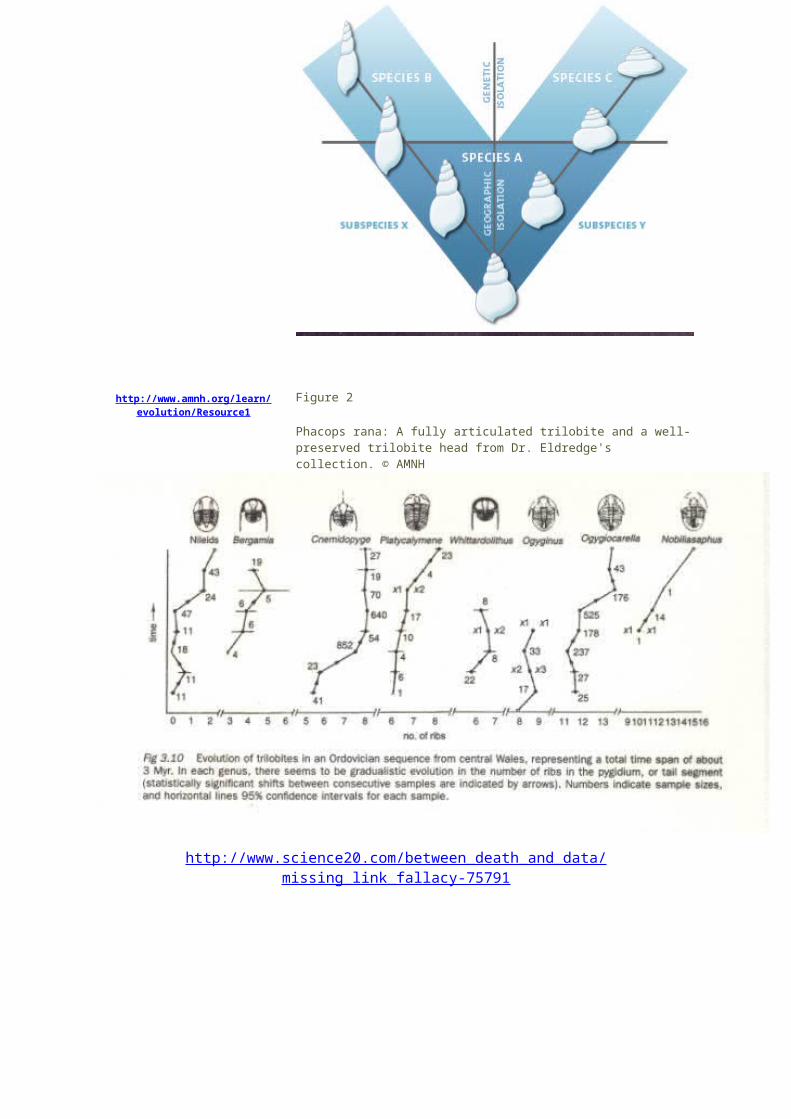
Figure 2
Phacops rana: A fully articulated trilobite and a well-preserved trilobite head from Dr. Eldredge's collection. © AMNH

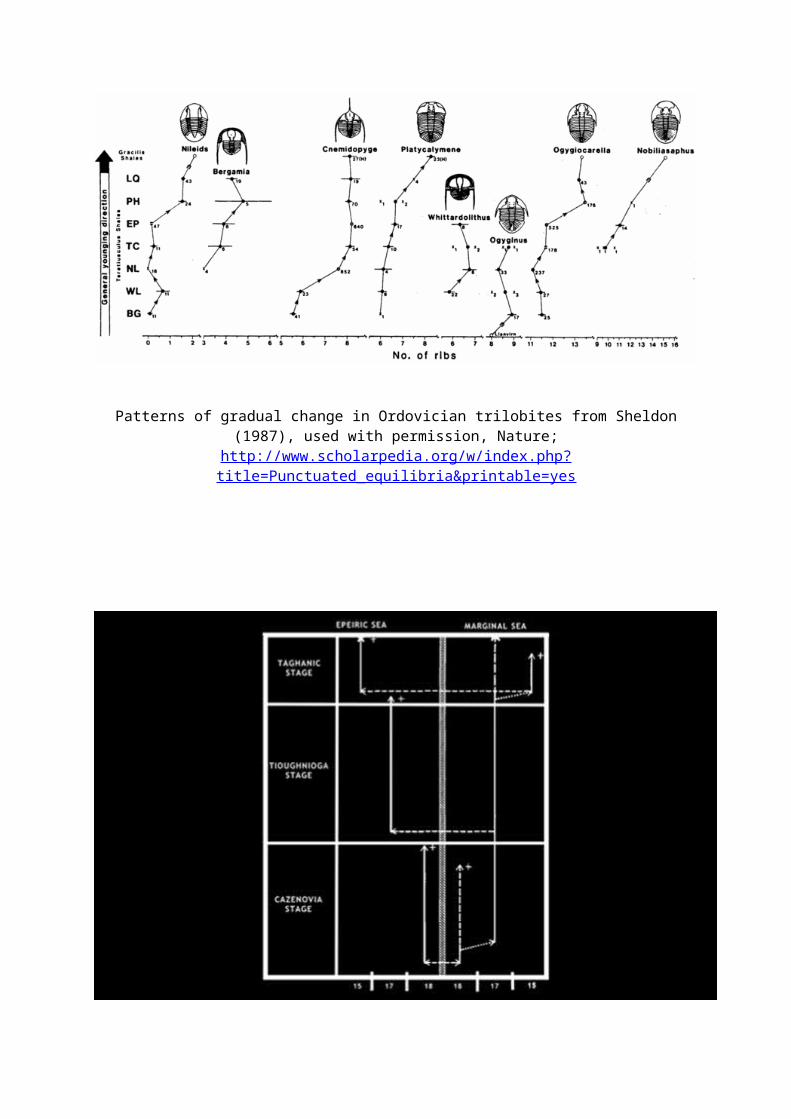
Patterns of gradual change in Ordovician trilobites from Sheldon (1987), used with permission, Nature;
http://www.scholarpedia.org/w/index.php?title=Punctuated_equilibria&printable=yes
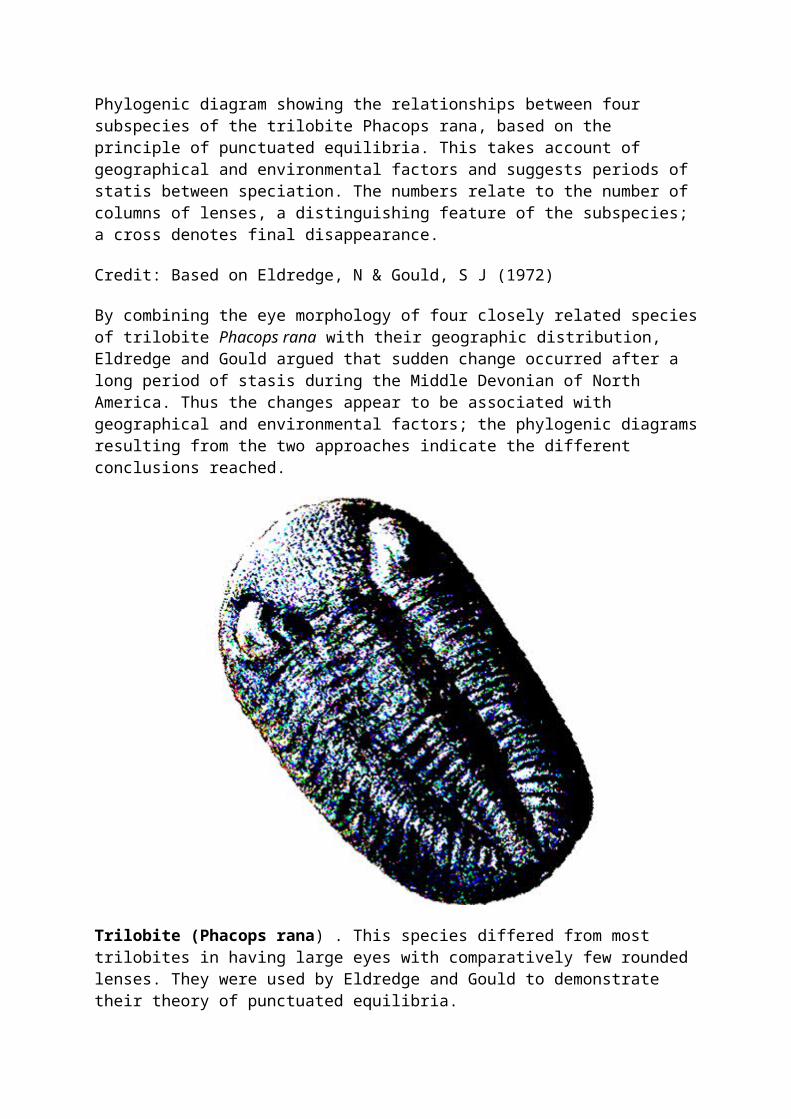
Phylogenic diagram showing the relationships between four subspecies of the trilobite Phacops rana, based on the principle of punctuated equilibria. This takes account of

geographical and environmental factors and suggests periods of statis between speciation. The numbers relate to the number of columns of lenses, a distinguishing feature of the subspecies; a cross denotes final disappearance.
Credit: Based on Eldredge, N & Gould, S J (1972)
By combining the eye morphology of four closely related species of trilobite Phacops rana with their geographic distribution, Eldredge and Gould argued that sudden change occurred after a long period of stasis during the Middle Devonian of North America. Thus the changes appear to be associated with geographical and environmental factors; the phylogenic diagrams resulting from the two approaches indicate the different conclusions reached.
Trilobite (Phacops rana) . This species differed from most trilobites in having large eyes with comparatively few rounded lenses. They were used by Eldredge and Gould to demonstrate their theory of punctuated equilibria.
Credit: courtesy of the Valdosta State University's Virtual Fossil Museum
Exquisitely-Preserved Eldredgeops Trilobite from New York
Phacops rana crassituberculata = Eldredgeops rana crassituberculata
Trilobite Order Phacopida, Family Phacopidae
Geological Time: Middle Devonian
Size: 41 mm long
Fossil Site: Hamilton Group, Moscow Formation, Erie County, New York
trilobite previously assigned to the genus Phacops, Phacops is particularly well known for its Schizochroal eyes. This type of trilobite eye typically has fewer lenses, but they are much larger, with each lens separated from its neighbors and having an individual cornea.
The details to this specimen are readily evident in the photos. The Moscow Formation is the youngest formation in the Hamilton Group, and is known for the largest known
examples of several trilobite taxa.
The genus name for this trilobite is variously called Phacops or Eldredgeops, with the latter named after Niles Eldredge, whose study of Phacops trilobites from Ohio led him to hypothesize that modifications to the arrangement of the trilobite’s eye lenses proceeded in rapid change followed by stasis over millions of years during the Devonian.
Eldredge's interpretation of the Phacops fossil record was that the aftermaths of the lens changes, but not the rapidly occurring evolutionary process, were fossilized. This and other data led Gould and Eldridge to publish the initial paper explaining punctuated equilibrium in 1971.
click trilobite pictures to enlarge
Afbeelding 4: Phacops rana, ontstaan van nieuwe vormen en migratie. Uit Eldredge, 1971 en Gould en Eldredge, 1972. Kleuren zijn overgetrokken voor verduidelijking.
LINK Gould Strikes Back At Creationistsclick here to read article
De afbeelding brengt de stamboom tevoorschijn op grond van veranderingen(stadia: Taghanic, Tioughnioga Cazenovia.) in Phacops rana doorgemaakt gedurende de tijd dat deze soort bestaat. Rechts hebben we allerlei lijntjes en pijltjes en een dikke zwarte lijn in het midden. Deze lijn scheidt de populaties die leefden in een epicontinentale zee (links) en een randzee (rechts)°*. Een epicontinentale zee is een ondiepe zee dat op een continent ligt, vaak doordat de zeespiegel snel is gestegen. Voorbeelden daarvan zijn de Perzische golf en de Hudsonbaai. Een randzee is een zee die voor een groot deel contact heeft met een oceaan en dan ook zeestromingen heeft. Een voorbeeld daarvan is de rode zee. De rode verticale lijnen geven aan hoelang een bepaalde vorm (ondersoort) van deze soort leefde (stase of stilstand). De groene horizontale lijnen geven aan dat er migratie plaats vond. De schuine gele lijnen geven aan dat een deel van de populatie zich afsplitste en de populatie een verandering doormaakt (snelle evolutie). De blauwe lijnen geven aan dat deze vorm van Phacops rana doorleefde terwijl er een nieuwe vorm vanaf gesplitst was (de vormen waren geografisch geïsoleerd van elkaar). De +jes geven aan dat deze bepaalde vorm in de hogere gesteenten niet meer voorkomt.
http://www.fossilmuseum.net/Evolution/TrilobiteArmsRace.htm
Schizochroal eyes of the Devonian
Schizochroal eyes of the
Devonian
The Schizochroa
l eyes of
Schizochroal eyes of the Devonian
Phacopid trilobite Reedops deckeri
Each lens has an
individual cornea that
isseparated
from adjacent lenses
Phacopid trilobite
Hollardops merocristat
aEach lens
has an individual
cornea that is
separated from
adjacent lenses
this Devonian Phacopid trilobite
Coltraenia oufatenensis are both high, with
many lenses, and they wrap around,
affording a panoramic
view
Phacopid trilobite Phacops
ranaEach lens
has an individual
cornea that is
separated from
adjacent lenses
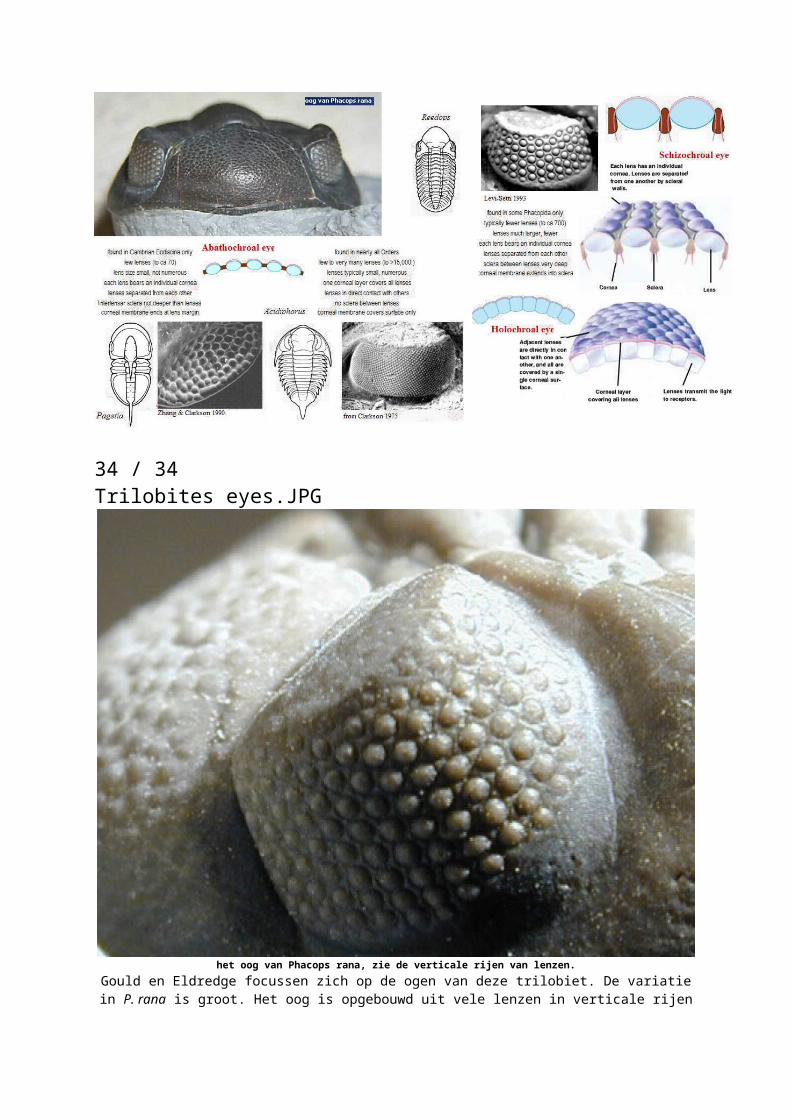
34 / 34Trilobites eyes.JPG

het oog van Phacops rana, zie de verticale rijen van lenzen. Gould en Eldredge focussen zich op de ogen van deze trilobiet. De variatie in P. rana is
groot. Het oog is opgebouwd uit vele lenzen in verticale rijen met meerdere lenzen per rij. In het begin heeft P. rana 18 verticale rijen. Bij de eerste gele pijl (afbeelding 5) ontstaat er een nieuwe ondersoort, deze heeft 17 verticale rijen. Bij de tweede gele pijl wordt het aantal rijen weer gereduceerd naar 15 (via 16). De getallen onderin de afbeelding geven
dit ook aan.
Onderin begint de grafiek met migratie; P. rana leeft dan in beide typen zeeën. Al snel evolueert er een tweede vorm uit de eerste vorm in de randzee. Beide vormen leven een tijdje door, maar voordat het Cazenovia eindigt sterft de voorouderlijke vorm uit. De vorm die in de epicontinentale zee leefde sterft uit aan het eind van het Cazenovia. In het Tioughnioga vind er een migratie plaats en P. rana is weer te vinden in beide typen zeeën. De gemigreerde groep gaat nog door tot aan het begin van het Taghanic. Er ontstaat in deze periode een nieuwe vorm uit de vorm die leefde in de randzee. Weer zijn er in de randzee wel twee vormen te vinden. Een deel van deze nieuwe vorm migreert naar de epicontinentale zeeën. Aan het eind van het Taghanic zijn alle vormen weer verdwenen. Als je alleen het gebied onderzoekt wat vroeger de epicontinentale zee was krijg je dat de eerste vorm opeens verdwijnt en kort erachter er een nieuwe vorm verschijnt. En dat gebeurd daarna nog een keer. Dit is echter maar een deel van het complete verhaal en wanneer je ook het gebied onderzoekt wat vroeger de randzee was, krijg je een veel beter beeld van wat er gebeurde met deze soort. De tussenvormen zijn daar ook te vinden. Nu gaat dit niet om nieuwe soorten, maar nieuwe ondersoorten. Gould en Eldredge zeggen echter dat het idee hetzelfde blijft. Een ander voorbeeld is te vinden in het geslacht Metrarabdotos (van de afdelingmosdiertjes of Bryozoa) over een tijdspanne van 24 miljoen jaar, zie afbeelding 6. Hier gaat het wel om nieuwe soorten.

Afbeelding 6: evolutie van het geslacht Metrarabdotos. De bedoeling van deze afbeelding is hetzelfde als afbeelding 4 over het patroon van de trilobiet Phacops rana. Uit McGraw-Hill's AccessScience.
Er zijn nog veel andere voorbeelden aan te wijzen waar dit patroon ook te vinden is, zoals bij Poecilozonites (een geslacht van landslakken), maar dan word me verhaal wel erg lang. Volgende deel is het punctuated equilibria vs. phyletic gradualism. En dan wil ik afsluiten met een quote uit de scholarpedia artikel:
"The recognition that species are individuals, and that macroevolution is basically about cladogenesis, indicates that species are evolutionarily significant entities, yet much of evolutionary theory has focused on within species patterns of population differentiation (Eldredge, 1985b). In short, punctuated equilibria entailed the need for a hierarchical expansion of evolutionary theory. Further, it suggested that the fossil record was one of the best venues to study evolution, partly because it was the only place where it is possible to directly observe what happens to groups of species over long periods of time." Eldredge, N. (1971). The allopatric model and phylogeny in Paleozoic invertebrates.Evolution 25: 156-167. Eldredge, N. & Gould, S. J. (1972). "Punctuated equilibria: an alternative to phyletic gradualism". In T.J.M. Schopf, ed., Models in Paleobiology. San Francisco: Freeman Cooper. pp. 82-115. Lieberman, B. S. & Eldredge, N. (2008). Punctuated equilibria. Scholarpedia, 3(1):3806 McGraw-Hill's Acces Science. Punctuated equilibria (evolutionary theory). *: Ik weet niet zeker of 'marginal sea' vertaald wordt naar randzee. Na veel zoeken dacht ik dat dat wel de beste vertaling was. De rest klopt gewoon (o.a. de voorbeelden).

Punctuated Equilibria (3) - phyletic gradualismvrijdag 1 oktober 2010 20:13 door Terrence
In deel 1 en 2 kwamen de theorie en de voorbeelden van punctuated equilibria aan bod. Eldredge en Gould bespreken ook phyletic gradualism, de zogenaamde grote tegenhanger van punctuated equilibria. Dit is zo'n beetje het laatste gedeelte, uit de twee vroege artikelen van Eldredge en Gould, dat belangrijk is voor deze 'serie'. Blijft er nog wat over van phyletic gradualism? Wat aan bod komt:
- hoe Eldredge en Gould phyletic gradualism zien
- voorbeelden die phyletic gradualism ondersteunen
- voorbeelden die niet strikt in 1 van de modellen passen
Hoe Eldredge en Gould phyletic gradualism zien.
Eldredge en Gould hebben de voorwaarden voor phyletic gradualism opgenoemd:
- nieuwe soorten ontstaan door de transformatie van oude soorten
- deze transformatie is langzaam en gelijkmatig
- deze transformatie gebeurt met een groot aantal individuen, vaak met de gehele populatie
- deze transformatie gebeurt over een groot gedeelte van de geografische spreiding van de voorouderlijke soorten
Hieruit volgen twee belangrijke conclusies volgens Eldredge en Gould:
- er zijn vele intermediaire vormen te vinden tussen de nieuwe en oude soort
- gaten in deze overgang komen door imperfecties van het fossielen verslag
De soortvorming in dit model gaat via anagenese (zie deel 1 onder het kopje: hoe soortvorming vandaag de dag gaat), de gehele populatie van een soort evolueert in een nieuwe soort. Zelfs
wanneer een populatie zich splitst in twee populaties (allopatrische soortvorming) zal de transformatie van elk van deze populaties gaan (zie afbeelding 1).
Afbeelding 1: hoe soortvorming eruit ziet volgens phyletic gradualism. Soorten B en C ontstaan langzaam uit soort A. Uiteindelijk bestaat soort A ook niet meer (de
gehele voorouderlijke populatie is veranderd in 2 nieuwe soorten).
Eldredge en Gould noemen nog dat Darwin zelf aanhanger was van dit model. Darwin schreef in Origin of Species al wel dat hij zelf niet dacht dat de transformatie van soorten gelijkwaardig verliep, dat het gebeurde met de gehele populatie en over de gehele geografische spreiding van de voorouderlijke populaties. Dit stuk hadden Eldredge en Gould er van mij dan ook uit kunnen laten.
Voor diegene die hier meer info over wil hebben, klik hier.
Voorbeelden die phyletic gradualism ondersteunen.
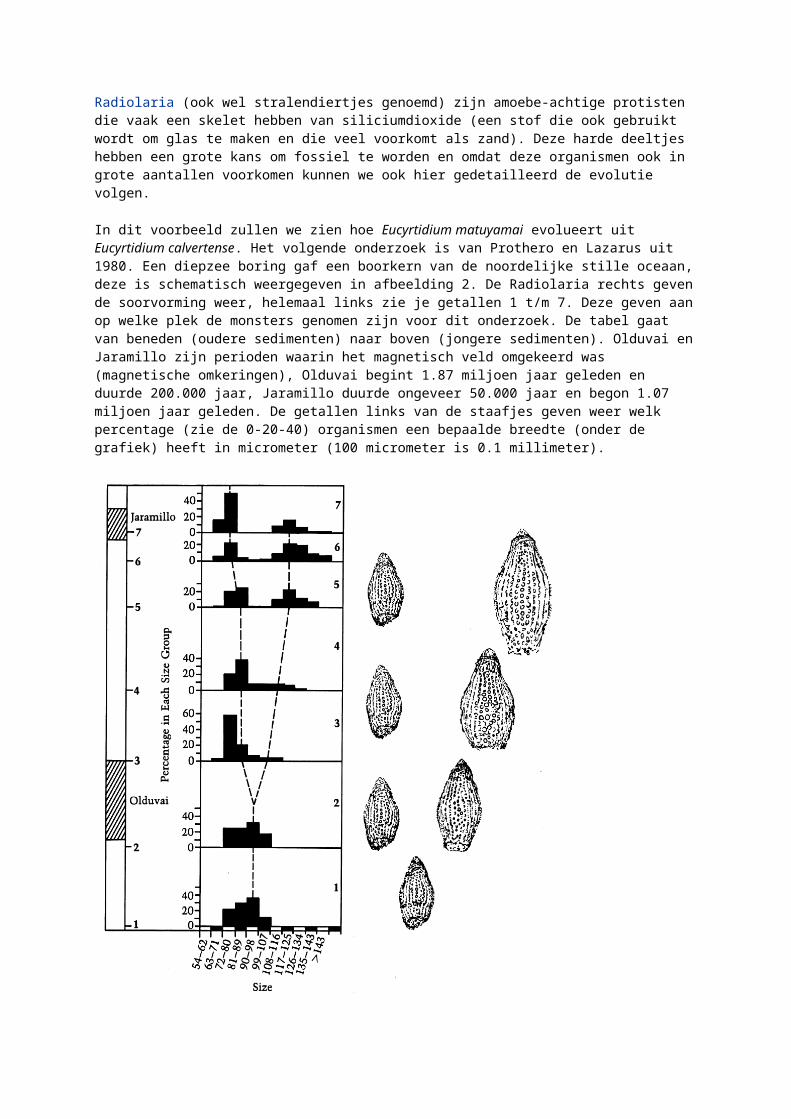
Radiolaria (ook wel stralendiertjes genoemd) zijn amoebe-achtige protisten die vaak een skelet hebben van siliciumdioxide (een stof die ook gebruikt wordt om glas te maken en die veel voorkomt als zand). Deze harde deeltjes hebben een grote kans om fossiel te worden en omdat deze organismen ook in grote aantallen voorkomen kunnen we ook hier gedetailleerd de evolutie volgen.
In dit voorbeeld zullen we zien hoe Eucyrtidium matuyamai evolueert uit Eucyrtidium calvertense. Het volgende onderzoek is van Prothero en Lazarus uit 1980. Een diepzee boring gaf een boorkern van de noordelijke stille oceaan, deze is schematisch weergegeven in afbeelding 2. De Radiolaria rechts geven de soorvorming weer, helemaal links zie je getallen 1 t/m 7. Deze geven aan op welke plek de monsters genomen zijn voor dit onderzoek. De tabel gaat van beneden (oudere sedimenten) naar boven (jongere sedimenten). Olduvai en Jaramillo zijn perioden waarin het magnetisch veld omgekeerd was (magnetische omkeringen), Olduvai begint 1.87 miljoen jaar geleden en duurde 200.000 jaar, Jaramillo duurde ongeveer 50.000 jaar en begon 1.07 miljoen jaar geleden. De getallen links van de staafjes geven weer welk percentage (zie de 0-20-40) organismen een bepaalde breedte (onder de grafiek) heeft in micrometer (100 micrometer is 0.1 millimeter).

Afbeelding 2: evolutie van ucyrtidium matuyamai uit Eucyrtidium calvertense.
In het eerste monster (van 2 miljoen jaar geleden) vinden we alleen E. calvertense met een breedte tussen 72 en 107 micrometer. Monster #2 (van ongeveer 1.9 miljoen jaar oud) bevat ook alleen E. calvertense met een breedte tussen de 72 en 107 micrometer.
Aan het eind van het Olduvai (1.67 miljoen jaar geleden) is monster #3 genomen. Hier zien we het begin van een nieuwe soort. De variatie ligt tussen 63 en 116 micrometer, maar boven de 90 micrometer zijn er weinig exemplaren. Monster #4 (ongeveer 1.55 miljoen jaar geleden) laat zien dat de variatie wat meer opschuift naar de grote kant, echter gaat het hier nog om één soort.
Bij monsters #5 en #6 (respectievelijk ongeveer 1.3 en 1.15 miljoen jaar geleden) zien we al een duidelijke scheiding, maar zijn er nog exemplaren te vinden van intermediaire grootte. Pas tijdens de Jaramillo periode (monster #7, ongeveer 1.05 miljoen jaar geleden) zien we dat alle intermediaire groottes verdwenen zijn en er duidelijk twee verschillende soorten zijn: E. matuyamai en E. calvertense. Deze soortvorming duurde dus ruim 600.000 jaar.
Nog een voorbeeld: Motoyama laat zien hoe Cycladophora davisiana tussen 2.8 en 2.4 miljoen jaar geleden uit C. sakaii evolueert via anagenese en hoe deze verder ontwikkeld (morfologisch) aan de hand van boorkern 192 van het Deep Sea Drilling Project. Deze soortvorming duurde 400.000 jaar.

Vooral in diepzee boringen (met eencellige organismen) vinden we het patroon van phyletic gradualism.
Een voorbeeld die niet strikt in één van de modellen past.
Orbulina universa en Globigerinoides trilobus zijn twee verschillende soorten Foraminifera (Gaatjesdragers worden ze ook genoemd) die vandaag nog steeds bestaan. Foraminifera zijn eencellige eukaryoten met een kalkskelet. Dit kalkskelet kent allerlei structuren en gaatjes waardoor de uitstulpingen van het cytoplasma naar buiten kunnen komen zodat het organisme zich kan voeden of verplaatsen. G. trilobus heeft drie grote lobben aan een vrij bolvormige kalkskelet. O. universa heeft echter een kalkskelet dat vanaf de buitenkant geheel bolvormig is. Als je O. universa openbreekt, vind je binnen dit bolvormige skelet een structuur die sterk lijkt op het uiterlijk van G. trilobus.
Afbeelding 3: evolutie van O. universa uit G. trilobus. Depth (linkerrij) is in meters. Age (rechterrij) is in miljoenen jaren.
Het onderzoek (Pearson et al., 1975) naar het ontstaan van O. universa uit G. trilobus was gedaan aan de hand van een kernboring van het Ocean Drilling Program (ODP) kern nummer 871A.
In deze kernboor (te zien in afbeelding 3) vinden we beide soorten (G. trilobus en O. universa) terug. G. trilobus is te vinden van begin tot einde terwijl O. universa pas te vinden is op 65 meter diepte. Tussen ongeveer 95 en 60 meter diepte vinden we ook andere soorten Foraminifera. We vinden Globigerinoides bisphericus tussen 95 en 62 meter diepte. Bij deze soort is de hoofdkamer in verhouding met de andere kamers (de lobben) een stuk groter dan bij G. trilobus.

Praeorbulina is een ander geslacht Foraminifera en soms worden ze ingedeeld in een enkele soort (met ondersoorten) en soms in 4 soorten. Deze (onder)soorten (P. sicana, P. curva, P. glomerosa en P. circularis) zijn te vinden tussen 80 en 60 meter diepte. P. sicana is de vorm die nog het meest lijkt op G. bisphericus. De meer geavanceerdere Praeorbulina lijken sterk op O. universa en hebben een bijna geheel bolvormig skelet. Deze komen na elkaar in de kernboring en volgen de voorouderlijke P. sicana op, maar sterven uit kort nadat O. universa (de geheel bolvormige Foraminifera) verschijnt.
Uiteindelijk blijven er dan twee soorten over en dat zijn Globigerinoides trilobus en Orbulina universa die tot op de dag van vandaag nog leven in de oceanen. De soortvorming van G. trilobus naar O. universa duurde bijna 2 miljoen jaar.
Afbeelding 4: evolutie van O. universe (#15) uit G. trilobus (#1 en #2).
In afbeelding 4 is te zien hoe al deze soorten Foraminifera eruit zien. Deze afbeelding laat de overgangsvormen zien tussen G. trilobus en O. universa. 1 en 2 zijn G. trilobus. 3 is G. bisphericus. 4,5 en 6 zijn P. sicana. 7 is P. curva. 8 is P. glomerosa. 9 en 10 zijn tussenvormen tussen P. glomerosa en P. circularis. 11 t/m 14 zijn P. circularis en alleen 15 (de enige gehele bolvormige van de 15) is O. universa.
De onderzoekers concluderen ook dat het hier gaat om sympatrische soortvorming. De lichte dieptescheiding tussen G. trilobus en O. universa die we nu aantreffen komt pas nadat O. universa is geëvolueerd. G. bisphericus (de eerste overgangsvorm) evolueert uit G. trilobus en laat dan een beetje stilstand zien. Daaruit evolueren opeenvolgend de Praeorbulina soorten (of ondersoorten) en daaruit O. universa. De soorten die nog overblijven veranderen nauwelijks en zien er tot op heden ongeveer hetzelfde uit. Het heeft kenmerken van punctuated equilibria. Echter is de soortvorming sympatrisch (dus niet volgens het standaard punctuated equilibria) en er is een geleidelijke lijn tussen G. trilobus en O. universa (voornamelijk wanneer de Praeorbulina soorten tevoorschijn komen). Deze overgang is dus niet volledig volgens punctuated equilibria of phyletic gradualism. En hier zijn ook meer voorbeelden van de vinden.
Beide modellen zijn bruikbaar in bepaalde gevallen en het een sluit het ander zeker niet uit. Ik kan zeker niet zeggen wat men vaker ziet in de aardlagen. Eldredge en Gould zeggen gelukkig ook dat beide modellen elkaar aanvullen.
In deze drie delen probeerde ik punctuated equilibria te beschrijven zoals Gould en Eldregde dat deden in hun eerste twee artikelen over dit onderwerp.

In het volgende (en laatste) deel behandel ik nog wat creationistische bezwaren tegen punctuated equilibria en dan moet volgens mij wel helemaal duidelijk zijn wat punctuated equilibria is en wat niet.
Referenties
Eldredge, N. & Gould, S. J. (1972). "Punctuated equilibria: an alternative to phyletic gradualism". In T.J.M. Schopf, ed., Models in Paleobiology. San Francisco: Freeman Cooper. pp. 82-115.
Motoyama, I. (1997). Origin and evolution of Cycladophora davisiana Ehrenberg (Radiolaria) in DSDP Site 192, Northwest Pacific. Marine Micropaleontology 30, pp. 45-63.
Pearson, P.N., Shackleton, N.J. & Hall, M.A. (1997). Stable isotopic evidence for the sympatric divergence of Globigerinoides trilobus and Orbulina universa (planktonic foraminifera). Journal of the Geological Society, London, Vol. 154, 1997 pp. 295-302
Prothero, D.R. & Lazarus, D.B. (1980). Planktonic microfossils and the recognition of ancestors. Systematic Zoology, 29: 119-129.
Punctated Equilibrum (Gould Eldridge )
ZIE OOK http://anticreato.multiply.com/notes/item/7
http://evodisku.multiply.com/journal/item/382/Punctuated_equilibrium
(Clik on image for zoom and link ) (Left) http://lhs2.lps.org/staff/sputnam/Biology/U6Evolution/U6Notes.htm (right) The diagram illustrates the now-famous pattern of punctuated equilibria as detected in the fossil record. http://sandwalk.blogspot.com/2009/03/stasis-is-data-says-don-mcleroy.html What it shows is that speciation by cladogenesis (splitting) is associated with morphological change. When a new species evolves from a parent species it does so quite rapidly. After the speciation event (horizontal lines on
the chart) the two species exist side-by-side in the same environment for millions of years without significant morphological change. Eventually they becomes extinct as shown by the termination of the vertical lines.
Species exist for 5-10 million years. During most of that time they do not change very much. This is what Eldredge and Gould called "stasis." The entire pattern is one of stasis interrupted by short periods of evolution at the time of speciation, or "punctuated equilibria."
The patterns in the fossil record raise interesting questions. One of the most important is whether it represents the normal pattern of evolution or whether it is confined to a minority of clades. What causes stasis? Why is change associated with cladogenesis? Why do species go extinct? All of these are widely discussed in the scientific literature.
None of the questions is a challenge to evolution. Punctuated equilibria is a pattern that might lead to an extension of evolutionary theory.

Home
Phylogenetic patterns within the Phacopidae based on Ramsköld and Werdelin (1991).
Tue, 08/04/2009 - 13:35 — Lieberman, Bruce

Taxa
Phacopidae
Photographer: Eldredge, Niles
Phyletic Gradualism and Punctuated Equilibrium

Phyletic Gradualism and Punctuated Equilibrium, these are the two theories to how new species develop over time, phyletic gradualism or gradualism is the idea that species develop slowly overtime into new species. Punctuated Equilibrium is the idea that for long periods of time there is equilibrium and followed by short periods of very fast evolution. This is described as a possible explanation for the Cambrian explosion.
http://sandwalk.blogspot.com/2011/03/punctuated-equilibria.html
http://www.youtube.com/embed/_YpAG3miURY
PEEC EEK misverstand
(naar prof Dr Gerdien de jong )http://tasmedes.web-log.nl/tasmedes/2009/03/everard-de-jong.html#comments# 5,6,7
'Plotselinge soortvorming' wordt door ID-ers en Creationisten vaak gesteld tegenover 'Darwin (die) zelf nog een geleidelijke groei van de ene soort uit de andere veronderstelde'.
Echter Een geleidelijk verandering en punctuated equilibrium gaan goed samen.
Die ‘plotselinge overgangen van de ene soort naar de andere soort’ is een verkeerde weergave van punctuated equilibria, waarbij geologisch snel gelijk gesteld wordt aan biologisch snel."
Het verschil in geologisch snel en biologisch snel heeft niets met de snelheid van afzetting van lagen

te maken, maar met de tijdschaal waarin mensen in de verschillende vakgebieden gewend zijn te denken.
Een snelle overgang bij punctuated equilibria is bv een overgang in een tijd van 10.000 - 20.000 jaar.
Voor een paleontoloog/geoloog mag dat snel zijn, voor een bioloog die aan soortvorming werkt is dat niet snel.
Voor een bioloog is snel: binnen 100 generaties.Voor een bioloog is 10000 jaar 10.000 - 100.000 generaties, en voor soortvorming is dat tijd zat.
http://scienceblogs.com/evolvingthoughts/2009/02/myth_4_darwin_was_a_gradualist.php