
HET BELANG VAN NIET-HUMANE PRIMATEN ALS...
30
UNIVERSITEIT GENT FACULTEIT DIERGENEESKUNDE Academiejaar 2011-2012 HET BELANG VAN NIET-HUMANE PRIMATEN ALS RESERVOIR VOOR MALARIA door Hannelore MILBOUW Promotor : Dr. Bruno Levecke Literatuurstudie in het kader van de Masterproef
Transcript of HET BELANG VAN NIET-HUMANE PRIMATEN ALS...

UNIVERSITEIT GENT
FACULTEIT DIERGENEESKUNDE
Academiejaar 2011-2012
HET BELANG VAN NIET-HUMANE PRIMATEN ALS RESERVOIR VOOR MALARIA
door
Hannelore MILBOUW
Promotor : Dr. Bruno Levecke Literatuurstudie in het kader
van de Masterproef


UNIVERSITEIT GENT
FACULTEIT DIERGENEESKUNDE
Academiejaar 2011-2012
HET BELANG VAN NIET-HUMANE PRIMATEN ALS RESERVOIR VOOR MALARIA
door
Hannelore MILBOUW
Promotor : Dr. Bruno Levecke Literatuurstudie in het kader
van de Masterproef

De auteur en de promotor(en) geven de toelating deze studie als geheel voor consultatie beschikbaar te stellen
voor persoonlijk gebruik. Elk ander gebruik valt onder de beperkingen van het auteursrecht, in het bijzonder met
betrekking tot de verplichting de bron uitdrukkelijk te vermelden bij het aanhalen van gegevens uit deze studie.Het
auteursrecht betreffende de gegevens vermeld in deze studie berust bij de promotor(en). Het auteursrecht
beperkt zich tot de wijze waarop de auteur de problematiek van het onderwerp heeft benaderd en
neergeschreven. De auteur respecteert daarbij het oorspronkelijke auteursrecht van de individueel geciteerde
studies en eventueel bijhorende documentatie, zoals tabellen en figuren. De auteur en de promotor(en) zijn niet
verantwoordelijk voor de behandelingen en eventuele doseringen die in deze studie geciteerd en beschreven zijn.

VOORWOORD
Het schrijven dan deze literatuurstudie was geen gemakkelijke opdracht. Ik zou dan ook graag enkele
personen willen bedanken die me geholpen hebben bij het verwezenlijken van dit werk.
Eerst en vooral wil ik mijn promotor, Dr. Bruno Levecke, danken voor zijn goede raad en tips. Ik zou
hem ook willen bedanken voor het herlezen en het kritisch evalueren van deze literatuurstudie.
Vervolgens zou ik mijn familie, en in het bijzonder mijn zus Soetkin Milbouw, willen bedanken. Zij
hebben niet enkel deze studie herhaaldelijk nagelezen, zij hebben mij ook langsheen de hele weg
gesteund. Hun onuitputtelijke steun helpt mij steeds vooruit. Bedankt.

VOORWOORD
INHOUDSOPGAVE
SAMENVATTING .................................................................................................................................... 1
INLEIDING ............................................................................................................................................... 2
LITERATUURSTUDIE ............................................................................................................................. 3
1. Plasmodium ........................................................................................................................................ 3
1.1. Taxonomie ................................................................................................................................... 3
1.2. De levenscyclus ........................................................................................................................... 5
1.2.1. De cyclus in de eindgastheer ............................................................................................... 5
1.2.2. De cyclus in de vector .......................................................................................................... 6
1.3. Morfologie .................................................................................................................................... 7
2. Klinische symptomen .......................................................................................................................... 8
2.1. Klinische symptomen bij de mens ............................................................................................... 8
2.2. Klinische symptomen bij niet-humane primaten ........................................................................ 10
3. Zoönose ............................................................................................................................................ 10
3.1. Transmissie van niet-humane malariaparasieten naar de mens .............................................. 11
3.2. Transmissie van humane malariaparasieten naar niet-humane primaten ................................ 13
3.3. De mug als link tussen mens en aap ........................................................................................ 14
4. Diagnose .......................................................................................................................................... 16
4.1. Invasieve methoden .................................................................................................................. 16
4.1.1. Dikke druppelpreparaten en uitstrijkpreparaten ................................................................. 16
4.1.2. Quantative buffy coat test ................................................................................................... 16
4.1.3. Immunochromatografische testen ...................................................................................... 17
4.1.4. Real-Time Polymerase Chain Reaction (PCR) test ........................................................... 17
4.1.5. Indirecte testen ................................................................................................................... 18
4.2. Niet-invasieve methoden ........................................................................................................... 18
BESPREKING ....................................................................................................................................... 19
REFERENTIES ..................................................................................................................................... 21

1
SAMENVATTING
Malaria is een infectieziekte die jaarlijks miljoenen mensen treft over heel de wereld. De parasiet,
verantwoordelijk voor de ziekte, werd in het begin van de twintigste eeuw geïdentificeerd. Sindsdien
werd de parasiet wereldwijd het onderwerp van menig wetenschappelijk onderzoek.
In het verleden kende men slechts vier malariaspecies die ziekte konden veroorzaken bij de mens,
met name Plasmodium falciparum, P. ovale, P. vivax en P. malariae. Recent heeft men echter, met
behulp van moderne moleculaire technieken, een vijfde malariaspecies kunnen aanduiden in humane
patiënten in Zuid-Oost Azië, namelijk P. knowlesi. Dit species heeft, in tegenstelling tot de vier andere
malariasoorten, niet de mens als natuurlijke eindgastheer, maar de oude wereldapen. P. knowlesi is
bijgevolg het eerste malariaspecies waarvan het zoönotische karakter bewezen is. De betrokken niet-
humane primaten ondervinden weinig hinder van de parasiet, waardoor zij kunnen fungeren als een
reservoir voor het malariaspecies. De aanwezigheid van een zodanig reservoir heeft belangrijke
repercussies op de hedendaagse eradicatieprogramma’s voor malaria. Het efficiënt uitroeien van
malaria zal men enkel kunnen bekomen door ook de niet-humane primaten mee in rekening te
brengen. Bovendien vermoedt men dat in de toekomst, naast P. knowlesi, meerdere niet-humane
species teruggevonden zullen worden bij humane patiënten. Momenteel worden verscheidene
malariaparasieten sterk verdacht van zoönotische eigenschappen, met name P. cynomogli, P. inui, P.
simium en P. brasilianum.
Hiernaast toonde onderzoek aan dat humane malariaspecies eveneens teruggevonden kunnen
worden bij niet-humane primaten. Hetgeen tevens van belang is bij het uitroeien van malaria.
Sleutelwoorden: Controle – Malaria – Niet-humane primaten – Plasmodium – Zoönose

2
INLEIDING
Malaria is één van de belangrijkste infectieuze ziekten uit onze tijd. Ook duizenden jaren geleden
speelde malaria reeds een prominente rol in het leven van de mens, waardoor het ook tot één van de
oudste infectieziekten kan gerekend worden. Vele antieke schrijvers beschrijven in hun teksten
ziekten die doen denken aan malaria. Zo beschreef Hippocrates, een geneesheer die leefde in de
vijfde eeuw voor Christus, zorgvuldig de typische symptomen van malaria, die bestaan uit
intermitterende koortsopstoten. Hij wordt dan ook beschouwd als de eerste malarioloog. Vele
geleerden na hem gaven blijk van dezelfde waarnemingen en stelden hypothesen op over de
mogelijke oorzaak. Een uiteindelijk antwoord, omtrent de oorzaak van de ziekte, werd pas in 1880
gevonden, toen de malariaparasiet voor het eerst in de geschiedenis werd geïdentificeerd. Deze
baanbrekende ontdekking werd verricht door Charles Louis Alphonse Laveran, een Franse arts. Hij
ontdekte de malariaparasiet in een vers bloedstaal van een patiënt in Algerije. Sindsdien is onze
kennis omtrent de parasiet aanzienlijk verbreed aan de hand van intensief onderzoek. Toch blijven
vele vragen tot op heden onbeantwoord.
Malaria heeft niet enkel in het verleden vele slachtoffers geëist, ook op dit moment vormt malaria een
gezondheidsprobleem op wereldschaal. Uit onderzoek van het World Health Organization (WHO)
bleek malaria in 2010 endemisch te zijn in 106 landen, wat wil zeggen dat ongeveer de helft van de
wereldbevolking elke dag het risico loopt geïnfecteerd te worden (WHO, 2011). Deze cijfers
benadrukken het belang van de ontwikkeling van gedegen controleprogramma’s.
In het verleden werden vier malariaspecies aangetoond als oorzaak van malaria-infecties bij de mens,
namelijk Plasmodium vivax, P. ovale, P. malariae en P. falciparum. Recent onderzoek toonde echter
aan dat ook een vijfde malariaspecies hiervoor verantwoordelijk kan worden gesteld. Desbetreffend
malariaspecies, P. knowlesi, is een malariaparasiet van niet-humane primaten en werd teruggevonden
bij talrijke humane patiënten in Zuid-Oost Azië. Dit bewijst dat malaria een zoönose is. De ontdekking
dat ook P. falciparum afkomstig is van de gorilla, pleit voor deze stelling.
Men weet echter nog niet in welke mate dit zoönotisch karakter relevant is voor de algemene
volksgezondheid. Vragen over hoeveel malariasoorten een interspecies transmissie kennen en in
welke mate dit onder natuurlijke omstandigheden voorkomt, dienen hiervoor eerst verder onderzocht
en beantwoord te worden. Ook het voorkomen van niet-humane primaten als reservoir voor zowel
humane als niet-humane malariapasieten is een belangrijk onderzoekspunt.
Recent ontwikkelde moleculaire technieken en niet-invasieve methoden, helpen de wetenschap voort
bij dit zware onderzoek. Zij maken het namelijk mogelijk om op grote schaal Plasmodium spp. in
wildlevende niet-humane primaten op te sporen. Op deze manier hoopt de wetenschap stap voor
stap zijn inzicht te verruimen omtrent de verschillende facetten van de malariaproblematiek, met in het
bijzonder het belang van diens zoönotische karakter.

3
LITERATUURSTUDIE
1. PLASMODIUM
Malaria is een infectieziekte, dewelke veroorzaakt wordt door een heterotrofe eukaryote intracellulaire
parasiet van het geslacht Plasmodium. Dit geslacht maakt deel uit van de familie Plasmodiidae, de
orde Haemosporida, de klasse Coccidea, de stam Apicomplexa en behoort uiteindelijk tot het rijk van
de ééncellige parasieten, de Protozoa (Taylor et al., 2007).
1.1. TAXONOMIE
Wereldwijd komen circa 180 verschillende species Plasmodium voor (Taylor M. et al., 2007). Elke
soort heeft één of meerdere specifieke vertebrate eindgastheren. De belangrijkste soorten
Plasmodium van de niet-humane primaten kunnen opgedeeld worden naargelang hun natuurlijke
gastheer, met name de nieuwe wereldapen, de oude wereldapen en de mensapen (tabel 2).
In tegenstelling tot de niet-humane primaten, waarbij men een grote verscheidenheid aan
malariaparasieten kan terugvinden, is de mens slechts de natuurlijke gastheer van vier
malariaparasieten. Dit zijn met name Plasmodium falciparum, P. vivax, P. ovale en P. malariae. P.
falciparum en P. vivax zijn sterk verspreid in de (sub)tropische gebieden. P. malariae en P. ovale
daarentegen kennen een minder wijde distributie (tabel 1; figuur 1).
Tabel 1: Globale distributie (in percentage) van humane malaria in 2002 (naar Carter et al., 2002)
Species West-, Centraal-Afrika
Oost-, Zuid-Afrika
Azië Centraal Azië, Midden-oosten
Zuid-Oost Azië, Oceanië
Oceanië Centraal-Amerika en de Carrabieën
Zuid-Amerika
P. falciparum 88,2 78,8 4,2 19,8 51,4 43 12,9 29,2
P. vivax 1,2 9,8 95,6 80,2 48,6 56,1 87,1 70,6
P. malariae 2,2 3,0 0 0,9 0 0,2
P. ovale 8,4 8,4 0,2 0 0 0
Figuur 1: Globale risicogebieden van malaria bij de mens in 2010 opgesteld door het WHO. In de donker blauw gekleurde gebieden komen malaria-infecties veelvuldig voor. In de licht blauw gekleurde gebieden is het risico op infectie slechts gering. (http://gamapserver.who.int/mapLibrary/Files/Maps/Global_Malaria_2010.png)
4
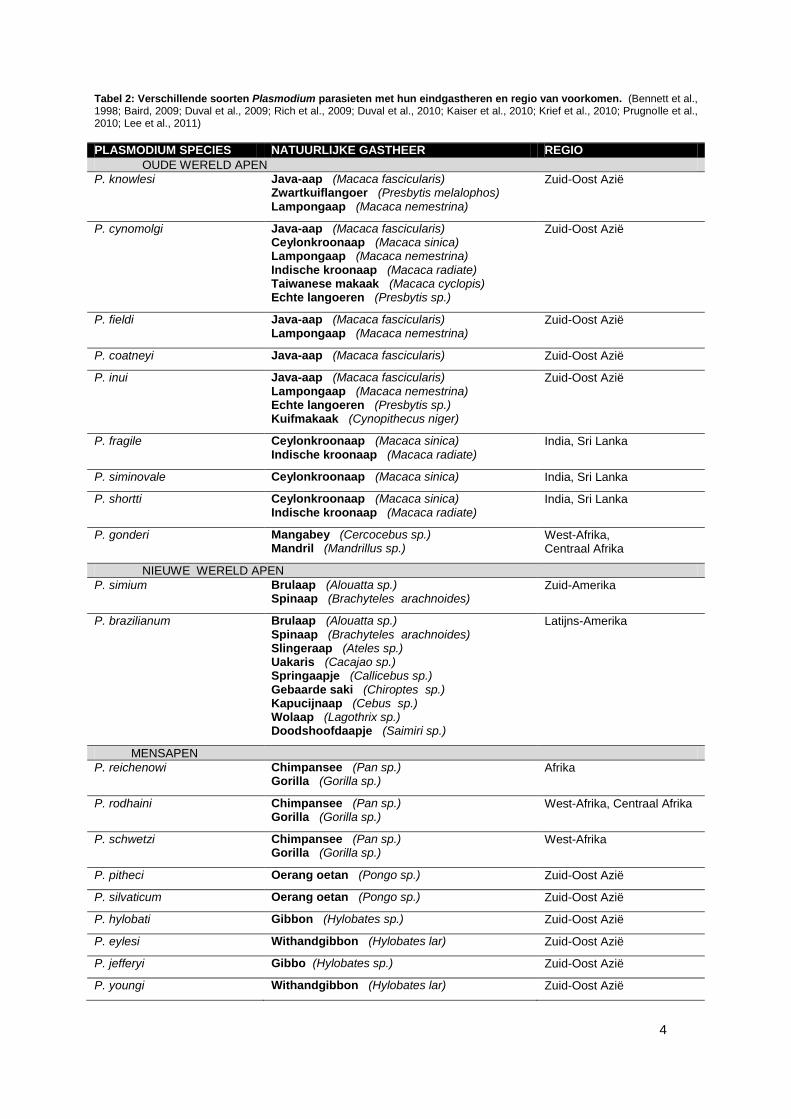
Tabel 2: Verschillende soorten Plasmodium parasieten met hun eindgastheren en regio van voorkomen. (Bennett et al., 1998; Baird, 2009; Duval et al., 2009; Rich et al., 2009; Duval et al., 2010; Kaiser et al., 2010; Krief et al., 2010; Prugnolle et al., 2010; Lee et al., 2011)
PLASMODIUM SPECIES NATUURLIJKE GASTHEER REGIO
OUDE WERELD APEN
P. knowlesi Java-aap (Macaca fascicularis) Zwartkuiflangoer (Presbytis melalophos) Lampongaap (Macaca nemestrina)
Zuid-Oost Azië
P. cynomolgi Java-aap (Macaca fascicularis) Ceylonkroonaap (Macaca sinica) Lampongaap (Macaca nemestrina) Indische kroonaap (Macaca radiate) Taiwanese makaak (Macaca cyclopis) Echte langoeren (Presbytis sp.)
Zuid-Oost Azië
P. fieldi Java-aap (Macaca fascicularis) Lampongaap (Macaca nemestrina)
Zuid-Oost Azië
P. coatneyi Java-aap (Macaca fascicularis)
Zuid-Oost Azië
P. inui Java-aap (Macaca fascicularis) Lampongaap (Macaca nemestrina) Echte langoeren (Presbytis sp.) Kuifmakaak (Cynopithecus niger)
Zuid-Oost Azië
P. fragile Ceylonkroonaap (Macaca sinica) Indische kroonaap (Macaca radiate)
India, Sri Lanka
P. siminovale Ceylonkroonaap (Macaca sinica)
India, Sri Lanka
P. shortti Ceylonkroonaap (Macaca sinica) Indische kroonaap (Macaca radiate)
India, Sri Lanka
P. gonderi Mangabey (Cercocebus sp.) Mandril (Mandrillus sp.)
West-Afrika, Centraal Afrika
NIEUWE WERELD APEN
P. simium Brulaap (Alouatta sp.) Spinaap (Brachyteles arachnoides)
Zuid-Amerika
P. brazilianum Brulaap (Alouatta sp.) Spinaap (Brachyteles arachnoides) Slingeraap (Ateles sp.) Uakaris (Cacajao sp.) Springaapje (Callicebus sp.) Gebaarde saki (Chiroptes sp.) Kapucijnaap (Cebus sp.) Wolaap (Lagothrix sp.) Doodshoofdaapje (Saimiri sp.)
Latijns-Amerika
MENSAPEN
P. reichenowi Chimpansee (Pan sp.) Gorilla (Gorilla sp.)
Afrika
P. rodhaini Chimpansee (Pan sp.) Gorilla (Gorilla sp.)
West-Afrika, Centraal Afrika
P. schwetzi Chimpansee (Pan sp.) Gorilla (Gorilla sp.)
West-Afrika
P. pitheci Oerang oetan (Pongo sp.)
Zuid-Oost Azië
P. silvaticum Oerang oetan (Pongo sp.)
Zuid-Oost Azië
P. hylobati Gibbon (Hylobates sp.)
Zuid-Oost Azië
P. eylesi Withandgibbon (Hylobates lar)
Zuid-Oost Azië
P. jefferyi Gibbo (Hylobates sp.)
Zuid-Oost Azië
P. youngi Withandgibbon (Hylobates lar)
Zuid-Oost Azië

5
1.2. DE LEVENSCYCLUS
De levenscyclus van Plasmodium is een indirecte cyclus. De
parasiet wordt namelijk via invertebrate vectoren overgebracht naar
gewervelde eindgastheren. De vectoren zijn steekmuggen behorend
tot de familie Culicidae en de subfamilies Culicinae en Anophelinae
(figuur 2). Circa zeventig muggensoorten van het genus Anopheles
staan in voor de overdracht van malaria bij de mens (Service en
Townson, 2002). De gewervelde eindgastheren maken deel uit van
de familie van de zoogdieren, vogels of reptielen.
De levenscyclus van de malariaparasiet kan men onderverdelen in een cyclus die plaatsgrijpt in de
vector en een cyclus in de gewervelde eindgastheer (figuur 3) (Nelson en Williams, 2006).
1.2.1. De cyclus in de eindgastheer
Tijdens een bloedmaal injecteert de geïnfecteerde vector Plasmodium in de gewervelde eindgastheer
onder de vorm van sporozoïeten (Nelson en Williams, 2006). Deze sporozoïeten bevinden zich
namelijk in het speeksel van de vector, dewelke via de steeksnuit van het insect in contact komt met
het bloed van de eindgastheer. De vectoren zijn namelijk solenofagen. Vervolgens migreren de
sporozoïeten langsheen de bloedbaan naar de lever en diens cellen. In de hepatocyten ondergaan de
sporozoïeten schizogonie. Het betreft een aseksuele vermenigvuldiging van de intracellulaire
parasiet. De kern van de schizont zal herhaaldelijk delen en het cytoplasma zal tijdens deze fase
enorme proporties aannemen met een verdrukking van de kern van de levercel tot gevolg. Na een
gemiddelde van tien dagen zal de geïnfecteerde hepatocyt en de intracellulaire schizont openbarsten
met de afgifte van een grote hoeveelheid merozoïeten tot gevolg (Nelson en Williams, 2006). De
merozoïeten komen op die manier in de bloedbaan terecht, waar zij vervolgens de rode bloedcellen
van de gastheer invaderen. Dit betekent het einde van het exo-erythrocyte stadium van de cyclus.
Bij bepaalde Plasmodium species, zoals P. vivax en P. ovale, is het mogelijk dat de geïnfecteerde
levercellen niet openbarsten na de gemiddelde periode van tien dagen. Men spreekt in dit geval van
hypnozoïeten of ‘slapende sporozoïeten’ (Krotoski et al., 1982). Deze inactieve sporozoïeten zullen
pas na een verlengde periode gereactiveerd worden, met de vorming van merozoïeten in de
bloedbaan. Dit fenomeen verklaart het feit dat sommige patiënten pas klinische symptomen vertonen
na verscheidene maanden na contact met de malariaparasiet.
Nadat de merozoïeten de erythrocyten van de gastheer binnen zijn gedrongen, veranderen zij in
trofozoïeten. Vervolgens deelt de kern van de trofozoïet zich aseksueel, met de vorming van een
meerkernige schizont (Nelson en Williams, 2006). Op dit moment spreekt men van erythrocyte
schizogonie. Daarna deelt de schizont zich op in verschillende éénkernige merozoïeten die bij het
openscheuren van de rode bloedcel vrijkomen in de bloedbaan. Deze cyclus verloopt synchroon in
het lichaam, hetgeen betekent dat alle merozoïeten gelijktijdig vrijgesteld worden uit de geïnfecteerde
rode bloedcellen en nieuwe erythrocyten invaderen. In een relatief korte periode kunnen zo grote
Figuur 2: De Anopheles mug als vector voor de malariaparasiet.

6
aantallen rode bloedcellen aangetast worden door de Plasmodium parasiet, hetgeen uiteindelijk zal
resulteren in klinische symptomen.
Niet alle merozoïeten in de erythrocyten zullen echter schizogonie ondergaan. Een gedeelte zal
namelijk gametocyten produceren en op die manier deel uitmaken van de seksuele vermenigvuldiging
van de parasiet (Nelson en Williams, 2006). Zowel mannelijke als vrouwelijke gametocyten zullen
aanwezig zijn in de rode bloedcellen van de eindgastheer. Wanneer een mug van de familie Culicidae
vervolgens bloed zuigt van deze gastheer, zullen de gametocyten mee opgenomen worden.
1.2.2. De cyclus in de vector
Na opname van de mannelijke en vrouwelijke gametocyten in het gastro-intestinaal systeem van de
vector, worden beiden omgezet in gameten. Vervolgens vindt er een bevruchting plaats, met de
vorming van een zygote. De zygote ontwikkelt zich verder tot een mobiele oökinete die migreert
doorheen de maagwand van de vector en terecht≠ komt op de buitenwand van het darmstelsel. De
oöcyt zal herhaaldelijk mitose ondergaan en uiteindelijk openbarsten met de vrijstelling van meerdere
sporozoïeten tot gevolg. Men spreekt van sporogonie. De sporozoïeten zullen vervolgens migreren
doorheen het lichaam van de vector naar de speekselklieren, zodat zij bij een volgend bloedmaal
geïnjecteerd kunnen worden in een nieuwe eindgastheer. Dit proces neemt ongeveer tien tot achttien
dagen in beslag, afhankelijk van de soort Plasmodium parasiet (Nelson en Williams, 2006).
Figuur 3: Levenscyclus Plasmodium (http://www.cdc.gov/malaria/about/biology/)
7
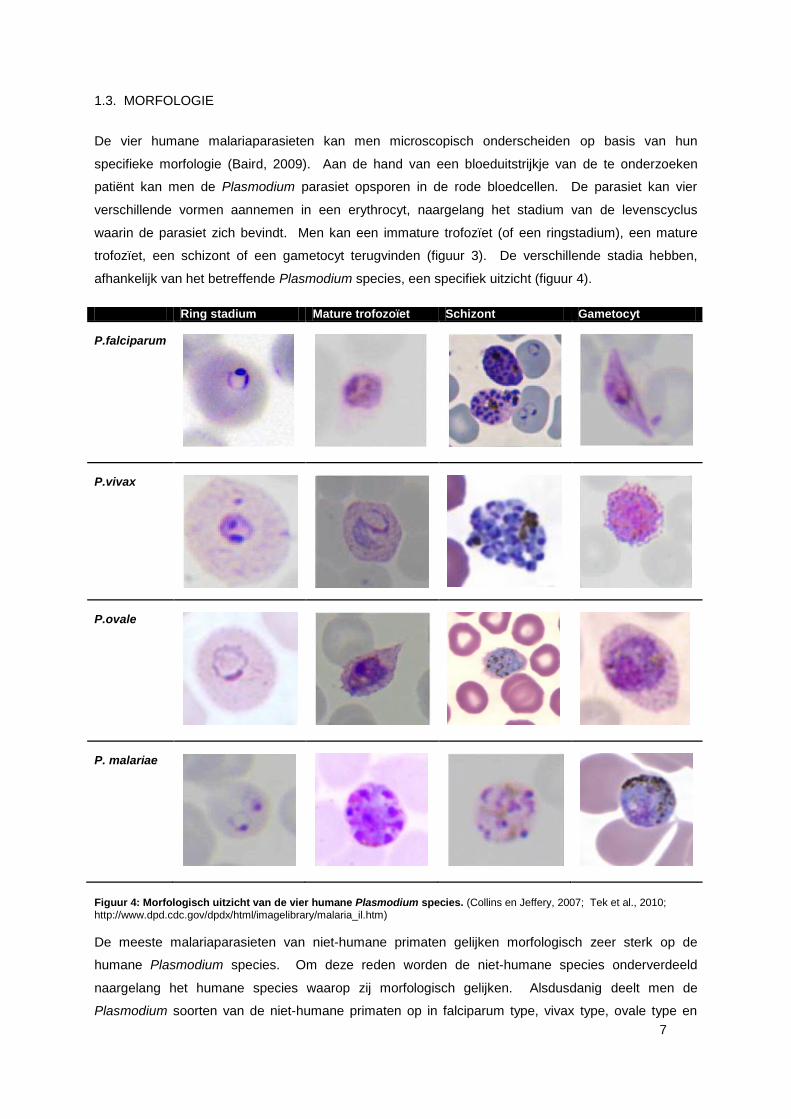
1.3. MORFOLOGIE
De vier humane malariaparasieten kan men microscopisch onderscheiden op basis van hun
specifieke morfologie (Baird, 2009). Aan de hand van een bloeduitstrijkje van de te onderzoeken
patiënt kan men de Plasmodium parasiet opsporen in de rode bloedcellen. De parasiet kan vier
verschillende vormen aannemen in een erythrocyt, naargelang het stadium van de levenscyclus
waarin de parasiet zich bevindt. Men kan een immature trofozïet (of een ringstadium), een mature
trofozïet, een schizont of een gametocyt terugvinden (figuur 3). De verschillende stadia hebben,
afhankelijk van het betreffende Plasmodium species, een specifiek uitzicht (figuur 4).
Ring stadium Mature trofozoïet Schizont Gametocyt
P.falciparum
P.vivax
P.ovale
P. malariae
Figuur 4: Morfologisch uitzicht van de vier humane Plasmodium species. (Collins en Jeffery, 2007; Tek et al., 2010; http://www.dpd.cdc.gov/dpdx/html/imagelibrary/malaria_il.htm)
De meeste malariaparasieten van niet-humane primaten gelijken morfologisch zeer sterk op de
humane Plasmodium species. Om deze reden worden de niet-humane species onderverdeeld
naargelang het humane species waarop zij morfologisch gelijken. Alsdusdanig deelt men de
Plasmodium soorten van de niet-humane primaten op in falciparum type, vivax type, ovale type en

8
malariae type parasieten (tabel 3) (Baird, 2009). Deze opdeling staat enkel voor de morfologische
gelijkenissen tussen de species en niet voor de klinische veruiterlijking van de specifieke infecties.
Tabel 3: Verdeling van niet-humane malariasoorten naargelang hun morfologische eigenschappen. (naar Baird, 2009)
Falciparum type Vivax type Ovale type Malariae type P. coatneyi
P. fragile
P. reichenowi
P. cynomolgi
P. eylesi
P. gonderi
P. hylobati
P. jefferyi
P. pitheci
P. silvaticum
P. schwetzi
P. simium
P. youngi
P. fieldi
P. simiovale
P. inui
P. rodhaini
P. brasilianum
P. knowlesi, een malariaparasiet van de oude wereldapen, past echter niet in één van de
bovenstaande types. De immature trofozoïeten van P. knowlesi gelijken namelijk sterk op P.
falciparum. Later in zijn cyclus gaat de parasiet, meer bepaald de oudere trofozoïeten en schizonten,
daarentegen meer gelijken op P. malariae (Baird, 2009).
Het gevolg van de morfologische gelijkenissen tussen de malariasoorten bij niet-humane primaten en
de mens is dat men bij het stellen van een diagnose niet enkel mag berusten op de resultaten van een
microscopisch onderzoek van een bloeduitstrijkje. De ervaren onderzoeker zal aan de hand van
microscopie een goed onderscheid kunnen maken tussen de verschillende humane species, maar zal
een mogelijke zoönose niet kunnen onderkennen. Dit kan vervolgens leiden tot een incorrecte
diagnose en prognose en mogelijks een insufficiënt therapeutisch plan (Baird, 2009; Galinski en
Barnwell, 2009).
2. KLINISCHE SYMPTOMEN
2.1. KLINISCHE SYMPTOMEN BIJ DE MENS
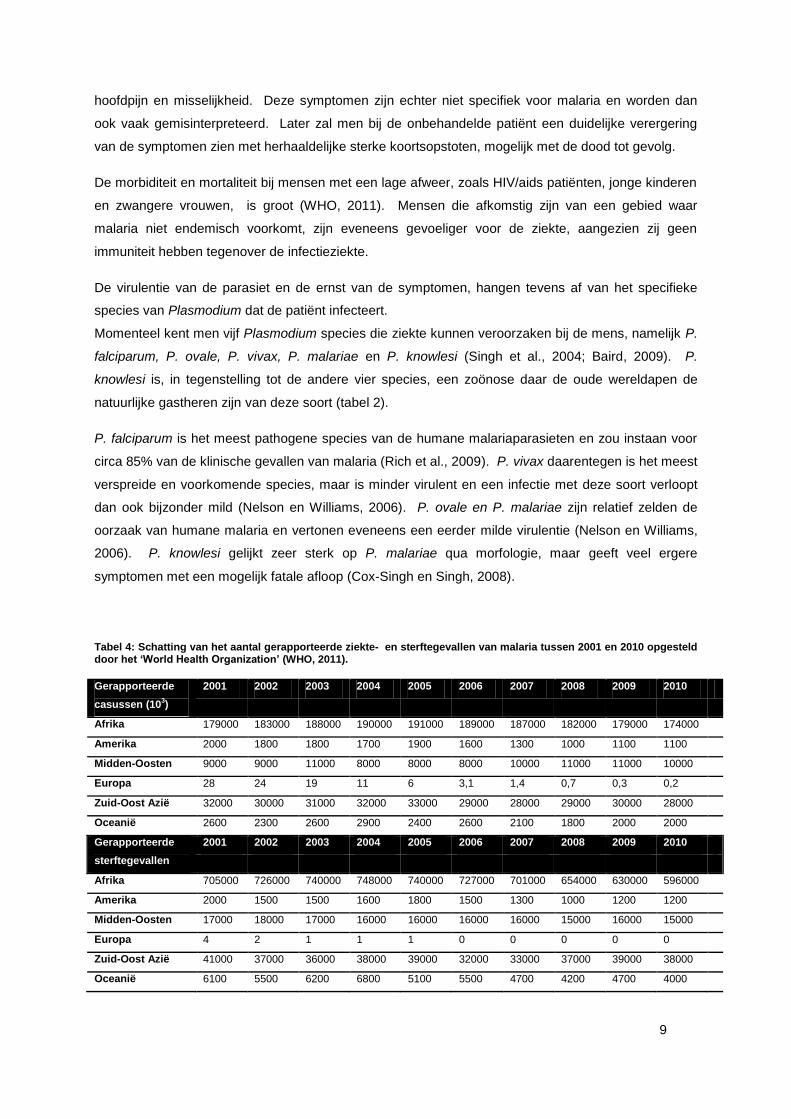
Malaria is een ziekte die jaarlijks miljoenen mensen treft over heel de wereld (tabel 4). Volgens het
‘World Malaria Report 2011’, opgesteld door het WHO (World Health Organization), waren er circa
216 miljoen gevallen van malaria in het jaar 2010, waarbij ongeveer 655 000 mensen stierven. Het
WHO bemerkt ook dat voornamelijk Afrika getroffen werd door malaria, daar in Afrika 81% van de
gerapporteerde patiënten woonden en er 91% van de fataal verlopende infecties plaatsvond (WHO,
2011).
Gemiddeld één tot twee weken na infectie met een malaria gerelateerde parasiet, ziet men bij niet-
immune patiënten vage symptomen. De patiënten hebben griepachtige klachten met koorts,
9
hoofdpijn en misselijkheid. Deze symptomen zijn echter niet specifiek voor malaria en worden dan
ook vaak gemisinterpreteerd. Later zal men bij de onbehandelde patiënt een duidelijke verergering
van de symptomen zien met herhaaldelijke sterke koortsopstoten, mogelijk met de dood tot gevolg.
De morbiditeit en mortaliteit bij mensen met een lage afweer, zoals HIV/aids patiënten, jonge kinderen
en zwangere vrouwen, is groot (WHO, 2011). Mensen die afkomstig zijn van een gebied waar
malaria niet endemisch voorkomt, zijn eveneens gevoeliger voor de ziekte, aangezien zij geen
immuniteit hebben tegenover de infectieziekte.
De virulentie van de parasiet en de ernst van de symptomen, hangen tevens af van het specifieke
species van Plasmodium dat de patiënt infecteert.
Momenteel kent men vijf Plasmodium species die ziekte kunnen veroorzaken bij de mens, namelijk P.
falciparum, P. ovale, P. vivax, P. malariae en P. knowlesi (Singh et al., 2004; Baird, 2009). P.
knowlesi is, in tegenstelling tot de andere vier species, een zoönose daar de oude wereldapen de
natuurlijke gastheren zijn van deze soort (tabel 2).
P. falciparum is het meest pathogene species van de humane malariaparasieten en zou instaan voor
circa 85% van de klinische gevallen van malaria (Rich et al., 2009). P. vivax daarentegen is het meest
verspreide en voorkomende species, maar is minder virulent en een infectie met deze soort verloopt
dan ook bijzonder mild (Nelson en Williams, 2006). P. ovale en P. malariae zijn relatief zelden de
oorzaak van humane malaria en vertonen eveneens een eerder milde virulentie (Nelson en Williams,
2006). P. knowlesi gelijkt zeer sterk op P. malariae qua morfologie, maar geeft veel ergere
symptomen met een mogelijk fatale afloop (Cox-Singh en Singh, 2008).
Tabel 4: Schatting van het aantal gerapporteerde ziekte- en sterftegevallen van malaria tussen 2001 en 2010 opgesteld door het ‘World Health Organization’ (WHO, 2011).
Gerapporteerde
casussen (103)
2001 2002 2003 2004 2005 2006 2007 2008 2009 2010
Afrika 179000 183000 188000 190000 191000 189000 187000 182000 179000 174000
Amerika 2000 1800 1800 1700 1900 1600 1300 1000 1100 1100
Midden-Oosten 9000 9000 11000 8000 8000 8000 10000 11000 11000 10000
Europa 28 24 19 11 6 3,1 1,4 0,7 0,3 0,2
Zuid-Oost Azië 32000 30000 31000 32000 33000 29000 28000 29000 30000 28000
Oceanië 2600 2300 2600 2900 2400 2600 2100 1800 2000 2000
Gerapporteerde
sterftegevallen
2001 2002 2003 2004 2005 2006 2007 2008 2009 2010
Afrika 705000 726000 740000 748000 740000 727000 701000 654000 630000 596000
Amerika 2000 1500 1500 1600 1800 1500 1300 1000 1200 1200
Midden-Oosten 17000 18000 17000 16000 16000 16000 16000 15000 16000 15000
Europa 4 2 1 1 1 0 0 0 0 0
Zuid-Oost Azië 41000 37000 36000 38000 39000 32000 33000 37000 39000 38000
Oceanië 6100 5500 6200 6800 5100 5500 4700 4200 4700 4000

10
2.2. DE KLINISCHE SYMPTOMEN BIJ NIET-HUMANE PRIMATEN
Malaria-infecties in niet-humane primaten zijn, in tegenstelling tot bij de mens, zelden fataal verlopend.
Zo ziet men in Zuid-Oost Azië binnen de stammen van de java-aap (Macaca fascicularis) en de
lampongaap (Macaca nemestrina), dewelke de natuurlijke gastheer zijn voor de meeste Plasmodium
spp. in deze regio (tabel 2), dat malaria-infecties eerder mild en chronisch verlopen (Galinski en
Barnwell, 2009). Dit geldt ook voor P. knowlesi infecties bij deze apen. Wel heeft dit species bij zijn
niet-natuurlijke gastheren, zoals de mens en de rhesus aap (Macaca mulatta), een hoge
mortaliteitsgraad (Galinski en Barnwell, 2009).
Eveneens vermoedt men dat malaria-infecties bij de mensapen in Afrika mild verlopen (Kaiser et al.,
2010). Onderzoek toonde bovendien aan dat de malariaparasiet prominent aanwezig is in de
chimpansee (Pan troglodytes) en in de westelijke laaglandgorilla (Gorilla gorilla gorilla) (tabel 5) (Liu et
al., 2010). Deze hoge cijfers zijn enkel mogelijk indien de parasiet de geïnfecteerde mensapen
slechts weinig hindert (Liu et al., 2010). Opmerkelijk is dat geen enkel species van Plasmodium
teruggevonden werd in de oostelijke laaglandgorilla (Gorilla beringei graueri) en in de bonobo (Pan
paniscus).
Tabel 5: Prevalentie van Plasmodium spp. infecties in wilde apen in Afrika (naar Liu et al., 2010).
Niet-humane primaat Prevalentie Plasmodium spp.
Chimpansee van Nigeria-Kameroen
(Pan troglodytes ellioti) 32% (23-46%)
Centraal-Afrikaanse chimpansee (Pan troglodytes troglodytes)
48% (44-5%)
Oostelijke chimpansee (Pan troglodytes schweinfurthii)
34% (30-40%)
Westelijke laaglandgorilla (Gorilla gorilla gorilla)
37% (32-41%)
Oostelijke laaglandgorilla (Gorilla beringei graueri)
0% (0%-4%)
Bonobo (Pan paniscus)
0% (0-6%)
Een belangrijke opmerking bij bovenstaande prevalenties is dat zij bekomen zijn aan de hand van een
niet-invasieve methode, meer specifiek een faecesonderzoek (Liu et al., 2010). De cijfers kunnen
daarom enkel beschouwd worden als schattingen van de werkelijkheid, aangezien men nog niet weet
in welke mate de parasiet exact wordt uitgescheiden in de mest (Rayner et al., 2011).
3. ZOONOSE
Oorspronkelijk werd malaria niet aanzien als zijnde een zoönose. Men erkende malaria bij de mens
en bij niet-humane primaten, maar men dacht dat deze niet overdraagbaar was tussen beide soorten.
Hedentendage weet men echter dat de grenzen tussen de verschillende gastheren van de
malariaparasiet niet zo strikt zijn als men aanvankelijk vermoedde (Baird, 2009; Rayner et al., 2011).
Toch is de kennis omtrent dit gegeven nog steeds zeer miniem. Vele vragen blijven onopgelost. Met
moderne technologieën en methoden wordt er intensief gezocht naar zoönotische Plasmodium
species. Het opsporen en identificeren van deze malariasoorten is niet enkel interessant vanuit een

11
puur wetenschappelijk oogpunt, maar ook vanuit het oogpunt van de volksgezondheid. De
hedendaagse ecologische veranderingen, zoals de enorme ontbossingen in vele tropische gebieden,
en de stijgende bevolkingspopulatie, zorgen er namelijk voor dat het contact tussen de wildlevende
niet-humane primaten en de mens steeds intensiever wordt (Tan et al., 2007; Rayner et al., 2011).
Het risico op transmissie van ziekten tussen beiden wordt hierdoor steeds groter.
3.1. TRANSMISSIE VAN NIET-HUMANE MALARIAPARASIETEN NAAR DE MENS
In het verleden dacht men dat slechts vier malariaspecies ziekte konden veroorzaken bij de mens,
namelijk P. falciparum, P. vivax, P. ovale en P. malariae. In het begin van vorige eeuw rees echter de
vraag of ook andere niet-humane species de mens konden infecteren. Dit vermoeden ontstond nadat
verscheidene humane patiënten, op experimentele basis, succesvol geïnfecteerd werden met niet-
humane species van de malariaparasiet (Baird, 2009). In 1932 ontdekte men namelijk voor het eerst
dat dit mogelijk was voor P. knowlesi (Knowles en Das Gupta, 1932). Hierbij werd een proefpersoon
geïnoculeerd met geïnfecteerd bloed van een niet-humane primaat. Gelijkaardige proeven met bloed
geïnfecteerd met P. rhodaini, P. inui en P. schwetzi kenden eveneens een positieve uitkomst (Das
Gupta, 1938; Rhodain, 1940; Rhodain en Dellaert, 1955).
Deze bevindingen leidden in 1960 tot een grootschalig onderzoek naar het zoönotisch karakter van P.
knowlesi in Zuid-Oost Azië (Warren et al., 1970). Hierbij werd bloed van 1117 humane patiënten
geïnoculeerd in Rhesus apen (Macaca mulatta). De Rhesus aap is geen natuurlijke gastheer van P.
knowlesi, waardoor infecties met dit species zeer virulent zijn (Vythilingam et al., 2006; Galinski en
Barnwell, 2009). Het experiment gaf echter uitsluitend negatieve resultaten, waardoor het belang van
P. knowlesi als zoönose onder natuurlijke omstandigheden als minimaal werd ingeschat en
gedeeltelijk vergeten werd (Cox-Singh en Singh, 2008; Galinski en Barnwell, 2009).
In 1960 toonde men, tijdens een experimentele studie, aan dat P. cynomolgi mensen eveneens kon
infecteren via de mug (Eyles et al., 1960). Gelijkaardige infecties lukten vervolgens ook met P.
brasilianum en P. inui (Contacos et al., 1963; Coatney et al., 1966).
In 1965 werd nadien een geval gerapporteerd van een natuurlijk geïnfecteerde humane patiënt met P.
knowlesi, maar het werd aangenomen dat dit slechts om een alleenstaande, eerder zeldzame
gebeurtenis ging (Chin et al., 1965; Tan et al., 2008; Galinski en Barnwell, 2009). In 1966 werd in
Brazilië een tweede natuurlijke infectie met P. simium aangetroffen in een humane patiënt (Deane et
al., 1966).
Bovenstaande bevindingen duidden malaria aan als een zoönose, maar men kreeg pas een begrip
van de uitgebreidheid van malaria als zoönose in 2004. Singh et al. toonde in dat jaar aan, met
behulp van een PCR-test, dat P. knowlesi veelvuldig teruggevonden kon worden in humane patiënten
in Borneo, bij wie de parasiet klinische symptomen en mogelijks sterfte veroorzaakte (Singh et al.,
2004; Galinski en Barnwell, 2009). De sequenties van P. knowlesi die men terugvond bij de humane
patiënten, bleken bovendien polymorf te zijn, hetgeen een recent bekomen zoönotisch karakter uitsluit
(Lee et al., 2011). Dit leidde tot een cascade van onderzoeken in Zuid-Oost Azië, waarbij P. knowlesi

12
tevens aangetoond kon worden bij patiënten in Maleisië, Thailand, Myanmar, Singapore, de Filipijnen,
Vietnam en Indonesië (Lee et al., 2011). Sindsdien wordt P. knowlesi aanzien als het vijfde
Plasmodium species dat malaria bij de mens kan induceren. Men besluit dus dat P. Knowlesi een
belangrijke zoönose in Zuid-Oost Azië is, met wilde makaken als reservoir (Galinski en Barnwell,
2009; Kaiser et al., 2010).
Deze bevindingen deden wetenschappers vermoeden dat P. knowlesi niet de enige niet-humane
malariaparasiet is die veelvuldig onder natuurlijke omstandigheden overgebracht kan worden naar
mensen, maar tot op heden is nog geen ander niet-humaan malariaspecies aangeduid.
Een analyse, verricht door Baird (2009), duidt naast P. knowlesi tevens P. cynomolgi en P. inui aan
als eventuele zoönotische species in Zuid-Oost Azië. Beide species zijn, volgens de auteur, sterk
verdacht, daar uit meerdere studies blijkt dat zij beiden goed gedijen in humane patiënten na
experimentele of accidentele natuurlijke infecties (Baird, 2009). Bovendien worden de twee species
overgedragen door dezelfde vectoren als P. knowlesi, met name de Anopheles species van de
leucosphyrus groep (tabel 6) (Tan et al., 2008; Galinski en Barnwell, 2009). Bewijzen om beide
species als ware zoönotische soorten aan te duiden, zijn echter nog niet gevonden. Verder
onderzoek zal hierover uitsluitsel moeten geven.
P. fieldi, P. semiovale en P. fragile worden in de studie van Baird (2009) aangeduid als een matig
risico voor de mens. Deze drie species worden eveneens door dezelfde vectoren overgedragen als P.
knowlesi. Men weet echter nog te weinig over de karakteristieken en biologische eigenschappen van
deze parasieten om een mogelijke zoönose aan te kunnen duiden of uit te sluiten (Baird, 2009).
P. coatneyi is waarschijnlijk geen zoönotisch species. Nochtans heeft dit species dezelfde gastheren
als de bovenstaande malariasoorten, maar studies waarbij men trachtte patiënten te infecteren met dit
species kenden allen een negatief resultaat (Coatney et al., 1971).
Men vermoedt dat ook de bevolking van Zuid-Amerika een risico loopt op infectie met niet-humane
malariaspecies. P. simium en P. brasilianum worden immers beide sterk verdacht van zoönotische
eigenschappen. Deze twee species, met de nieuwe wereldapen als natuurlijke gastheren, zijn
waarschijnlijk anthropozoönosen, die oorspronkelijk ontstaan zijn uit infecties van apen met P. vivax
en P. malaria (Baird, 2009).
Naast de biologische eigenschappen van de malariaparasieten speelt ook de grootte van de populatie
van de natuurlijke gastheren een rol. Wanneer de natuurlijke gastheer slechts in geringe mate
voorkomt zal de kans op het optreden van een zoönose uiterst klein zijn. Zulke zoönosen zijn
bijgevolg niet relevant voor de algemene volksgezondheid (Baird, 2009).
Het aantonen van interspecies transmissies is niet vanzelfsprekend. Dit is te wijten aan verschillende
oorzaken (Rayner et al., 2011). Ten eerste geven beide infecties vaak gelijkaardige symptomen,
waardoor patiënten, geïnfecteerd met humane malariasoorten, klinisch niet gedifferentieerd kunnen
worden van patiënten die geïnfecteerd zijn met niet-humane soorten. Deze laatsten zullen bovendien

13
bij behandeling betrekkelijk goed reageren op de medicatie aangezien de niet-humane malariasoorten
niet resistent zijn aan de gebruikte farmaca. Ten tweede wordt vaak een verkeerde diagnose gesteld
aan de hand van microscopisch onderzoek. Moderne diagnostische methoden, zoals polymerase
chain reactie (PCR) technieken, zijn nodig om het juiste species aan te duiden. Deze technieken
kunnen echter niet op routinematige basis uitgevoerd worden in klinieken omwille van hun hoge
kostprijs en de nood aan gespecialiseerde technici (Rayner et al., 2011).
3.2. TRANSMISSIE VAN HUMANE MALARIAPARASIETEN NAAR NIET-HUMANE PRIMATEN
Rond 1970 ontdekte men malariaparasieten in het bloed van wilde primaten dewelke sterk leken op
humane soorten (Coatney, 1971). Het vermoeden groeide dat overdracht van humane
malariaparasieten naar niet-humane primaten werkelijk voorkwam in bepaalde regio’s. Aangezien
men op dat moment slechts beschikte over de klassieke diagnostische methoden kon men dit
vermoeden niet bewijzen.
De moderne moleculaire technieken kunnen daarentegen malariasequenties in geïnfecteerde
erythrocyten zonder twijfel identificeren. Op deze manier heeft men humane malariaparasieten
kunnen aanduiden in wilde apen.
Uit een studie, verricht door Kaiser et al. (2010), bleek dit mogelijk voor P. vivax, P. malariae en P.
ovale. De stalen, onderzocht in deze studie, waren afkomstig van zowel gedomesticeerde als van in
het wild levende mensapen. In beide groepen vond men de humane malariaparasieten terug. Het feit
dat ook de wilde primaten, die geen contact hadden met mensen, geïnfecteerd waren, heeft de vraag
doen ontstaan of de wilde niet-humane primaten een natuurlijk reservoir vormen voor humane
malariaparasieten (Kaiser et al., 2010).
Ook P. falciparum kan teruggevonden worden in niet-humane primaten (Liu et al., 2010). Jarenlang
kon men geen exacte oorsprong aanduiden van dit species en trachtte men verschillende hypothesen
over zijn ontstaan te bewijzen.
In 1994 toonde men aan dat P. reichenowi, die voorkomt bij de chimpansee, het sterkst gerelateerd is
aan P. falciparum (Escalante en Ayla, 1994). Drie mogelijke hypothesen kan deze graad van
verwantschap verklaren (Rich et al., 2009). Ten eerste was het mogelijk dat de twee parasieten
afkomstig waren van dezelfde voorouder vijf tot zeven miljoen jaar geleden. Ten tweede kon P.
falciparum afkomstig zijn van P. reichenowi doordat een mens geïnfecteerd werd door de
chimpansee. Daartegenover kon P. reichenowi ook een menselijke afkomst hebben. De mening van
Escalante en Ayla (1994) stemden overeen met de eerste hypothese.
Rich et al. (2009) weerlegden deze theorie echter, aangezien zij, aan de hand van nieuwe isolaten
van P. reichenowi, stelden dat P. falciparum afkomstig was van P. reichenowi en men dus van een
dierlijke oorsprong kon spreken. Zij geloofden in de tweede hypothese, daar zij via de genetische
analyse van beide soorten opmerkten dat de genetische diversiteit van P. falciparum volledig binnen
de veel bredere genetische diversiteit van P. reichenowi viel (Rich et al., 2009).
Recent werd deze theorie weerom weerlegd door Liu et al. (2010). Zij voerden een grootschalige
moleculaire epidemiologische studie uit, waarbij circa 3000 faecesstalen onderzocht werden in

14
Centraal-Afrika. De gecollecteerde faecesstalen waren afkomstig van wilde niet-humane primaten,
met name de chimpansee (Pan Troglodytes), de westerse gorilla (Gorilla Gorilla), de oosterse gorilla
(Gorilla Beringei) en de bonobo (Pan Paniscus) (tabel 5). Uit de bekomen resultaten bleek dat al de
humane sequenties van P. falciparum, die men tot op heden bij de mens heeft verzameld, vallen
binnen de groep van de gorilla parasieten (figuur 5). Men kan aldusdanig besluiten dat de humane
malariaparasiet P. falciparum afkomstig is van een malariaparasiet bij de gorilla en dat de gorilla een
reservoir vormt voor de dodelijkste malariaparasiet die de mensheid teistert (Liu et al., 2010).
Bovendien wordt er vermoed dat aan de basis hiervan slechts een éénmalige overdracht van de
malariaparasiet tussen de gorilla en de mens ligt, daar de sequenties van de humane parasiet allen
sterk op elkaar gelijken (Liu et al., 2010).
Ook in Zuid-Amerika werden nieuwe wereldapen aangetroffen die besmet waren met humane
malariaparasieten, meer specifiek P. malariae (Fandeur et al., 2000).
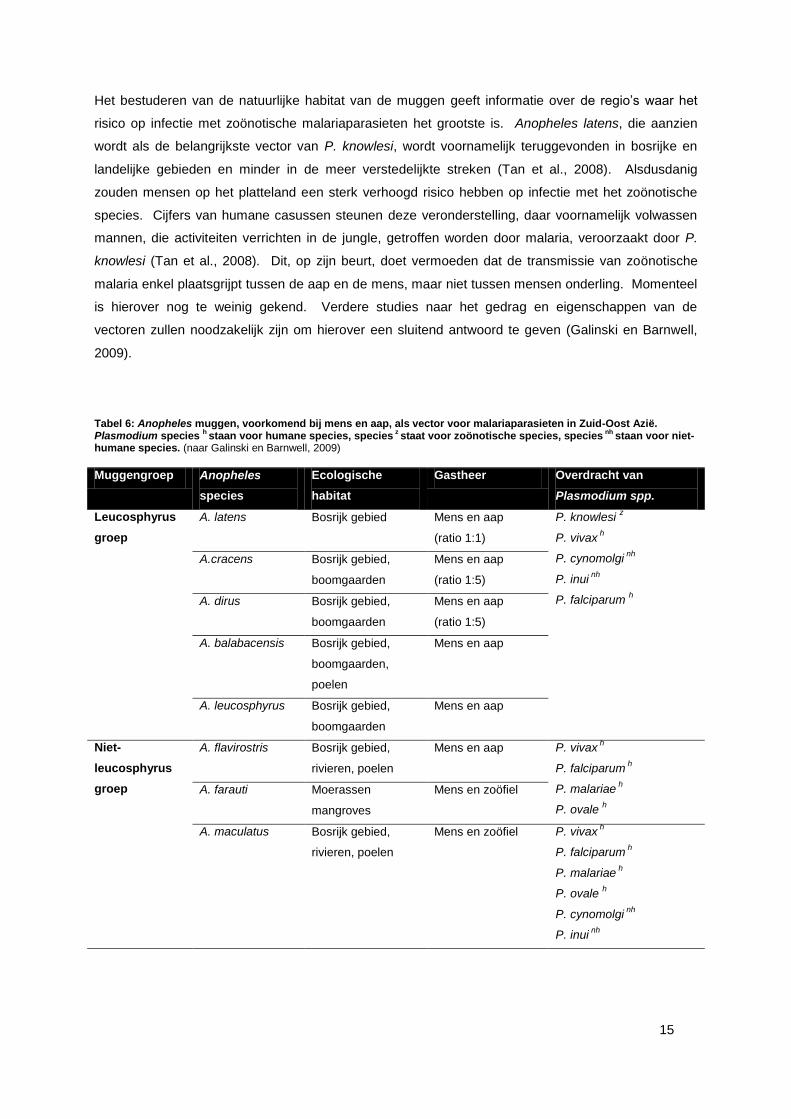
3.3. DE MUG ALS LINK TUSSEN MENS EN AAP
Rond 1970 startte men onderzoek naar het gedrag van muggen behorende tot de Anopheles
leucosphyrus groep. Men vroeg zich af in welke mate deze muggen mee aan de basis liggen van het
zoönotisch optreden van malaria door bloed te zuigen bij zowel de mens als de aap (Coatney, 1971).
In Zuid-Oost Azië vond men verscheidene Anopheles species terug, die inderdaad beide zoogdieren
parasiteren (tabel 6) (Galinski en Barnwel, 2009). Aangezien de muggen eveneens de drager zijn van
P. knowlesi, kunnen zij zorgen voor het optreden van zoönosen. Bij de eradicatie van malaria als
zoönose, zouden de desbetreffende muggen een mogelijk aangrijpingspunt kunnen vormen.
Figuur 5: De figuur toont de dierlijke oorsprong van Plasmodium falciparum. Het geeft ook de relatie weer tussen de strengen geïsoleerd bij de chimpansee (blauw, C1-C3) en de gorilla (rood, G1-G3). (Uit Holmes E.C.,
2010)
15
Het bestuderen van de natuurlijke habitat van de muggen geeft informatie over de regio’s waar het
risico op infectie met zoönotische malariaparasieten het grootste is. Anopheles latens, die aanzien
wordt als de belangrijkste vector van P. knowlesi, wordt voornamelijk teruggevonden in bosrijke en
landelijke gebieden en minder in de meer verstedelijkte streken (Tan et al., 2008). Alsdusdanig
zouden mensen op het platteland een sterk verhoogd risico hebben op infectie met het zoönotische
species. Cijfers van humane casussen steunen deze veronderstelling, daar voornamelijk volwassen
mannen, die activiteiten verrichten in de jungle, getroffen worden door malaria, veroorzaakt door P.
knowlesi (Tan et al., 2008). Dit, op zijn beurt, doet vermoeden dat de transmissie van zoönotische
malaria enkel plaatsgrijpt tussen de aap en de mens, maar niet tussen mensen onderling. Momenteel
is hierover nog te weinig gekend. Verdere studies naar het gedrag en eigenschappen van de
vectoren zullen noodzakelijk zijn om hierover een sluitend antwoord te geven (Galinski en Barnwell,
2009).
Tabel 6: Anopheles muggen, voorkomend bij mens en aap, als vector voor malariaparasieten in Zuid-Oost Azië. Plasmodium species
h staan voor humane species, species
z staat voor zoönotische species, species
nh staan voor niet-
humane species. (naar Galinski en Barnwell, 2009)
Muggengroep Anopheles
species
Ecologische
habitat
Gastheer Overdracht van
Plasmodium spp.
Leucosphyrus
groep
A. latens Bosrijk gebied Mens en aap
(ratio 1:1)
P. knowlesi z
P. vivax h
P. cynomolgi nh
P. inui nh
P. falciparum h
A.cracens Bosrijk gebied,
boomgaarden
Mens en aap
(ratio 1:5)
A. dirus Bosrijk gebied,
boomgaarden
Mens en aap
(ratio 1:5)
A. balabacensis Bosrijk gebied,
boomgaarden,
poelen
Mens en aap
A. leucosphyrus Bosrijk gebied,
boomgaarden
Mens en aap
Niet-
leucosphyrus
groep
A. flavirostris Bosrijk gebied,
rivieren, poelen
Mens en aap P. vivax h
P. falciparum h
P. malariae h
P. ovale h
A. farauti Moerassen
mangroves
Mens en zoöfiel
A. maculatus Bosrijk gebied,
rivieren, poelen
Mens en zoöfiel P. vivax h
P. falciparum h
P. malariae h
P. ovale h
P. cynomolgi nh
P. inui nh

16
4. DIAGNOSE
Malaria kan opgespoord worden aan de hand van twee methoden, een invasieve en een non-
invasieve methode.
4.1. INVASIEVE METHODEN
In het verleden beschikte men enkel over invasieve methoden om een malariaparasiet te
diagnosticeren bij een patiënt. Men maakt hierbij voornamelijk gebruik van bloedstalen. Het
bloedstaal kan op verschillende manieren geanalyseerd worden. Hieronder worden kort de
verschillende methoden besproken met hun voor- en nadelen.
4.1.1. Dikke druppelpreparaten en uitstrijkpreparaten
Deze methode is de meest eenvoudige methode om malariaparasieten op te sporen in bloed.
Jarenlang was dit de enige manier om Plasmodium in een patiënt te diagnosticeren. Men gaat, met
behulp van een lichtmicroscoop, op zoek naar verschillende ontwikkelingsstadia van de parasiet in het
perifere bloed, namelijk de trofozoïeten, schizonten en gametocyten (De jonge et al., 1999).
Het dikke druppelpreparaat wordt specifiek gebruikt om de parasiet op te sporen. Het bloeduitstrijkje
daarentegen wordt gebruikt om het species van de parasiet aan te duiden aan de hand van zijn
specifieke morfologie (figuur 4).
Deze methode heeft als voordeel niet duur te zijn, maar vereist daarentegen wel een ervaren
onderzoeker. Bovendien kan men zowel vals negatieve als vals positieve resultaten bekomen.
Zoönotische species worden eveneens niet onderscheiden van de humane malariaparasieten (Baird,
2009).
Evenwel wordt deze methode nog steeds aanzien als de gouden standaard bij de diagnose van
malaria (Schindler et al., 2001; Moody, 2002; Nelson en Williams, 2006). Toch heeft men nood aan
modernere technieken die malaria op een efficiëntere en snellere wijze kunnen opsporen, zodat men
snel een gepaste therapie kan starten (Murray et al., 2003).
4.1.2. Quantative buffy coat test
De ‘quantative buffy coat test’ (QBC test) is een meer recent ontwikkelde labotest dewelke
bloedparasieten kan opsporen. Bij deze test maakt men gebruik van een tube die gecoat is met de
kleurstof Acridine oranje en kalium oxalaat. Acridine oranje heeft de eigenschap doorheen
membranen te kunnen migrereren om vervolgens te binden aan genetisch materiaal (DNA of RNA).
Daarenboven absorbeert de kleurstof UV-licht waarbij het vervolgens zichtbaar licht terug uitzendt.
De tube met het bloedstaal zal men centrifugeren waardoor, volgens het principe van centrifugatie, de
verschillende cellen aanwezig in het staal zich zullen schikken naargelang hun densiteit.
Geparasiteerde rode bloedcellen hebben een lagere densiteit dan gezonde rode bloedcellen waardoor
zij zich zullen bevinden op de grens tussen de gezonde niet-geïnvadeerde erythrocyten en de

17
leukocyten. Door deze regio met behulp van een fluorescentie microscoop te onderzoeken, kan men
geïnfecteerde rode bloedcellen opsporen en op die manier een diagnose stellen.
Uit onderzoek blijkt dat de sensitiviteit van de QBC test hoger is dan de sensitiviteit van het
conventionele onderzoek van een dikke druppelpreparaat (Damodar, 1996). De test is eveneens zeer
gemakkelijk uitvoerbaar en kan op een zeer korte tijd worden verricht (Damodar, 1996; Schindler et
al., 2001).
Een nadeel van de test is de hoge kostprijs van de benodigde materialen, maar dit weegt volgens
Damodar (1996) niet op tegen de prominent aanwezige voordelen van de test.
4.1.3. Immunochromatografische testen
Deze eenvoudig uitvoerbare testen kenden hun opkomst rond 1990 (Shiff et al., 1993; Murray et al.,
2003). Ze kunnen op een uiterst snelle wijze Plasmodium antigenen aanduiden in een bloedstaal. De
test is op de markt aanwezig als strips of dipsticks waarop men het te onderzoeken bloed moet
aanbrengen. Vervolgens kan men hierop meteen het resultaat aflezen. Momenteel maken deze
testen gebruik van vier verschillende antigenen, namelijk het histidine-rijke proteïne 2 (HRP-2), lactaat
dehydrogenase, aldolase en een vierde antigen van P. vivax dewelke niet verder geïdentificeerd is
(Murray et al., 2003). Het lactaat dehydrogenase en aldolase zijn enzymen dewelke in specifieke
stadia van de levenscyclus van de parasiet gevormd worden.
De voordelen van deze testen zijn dat men bij het stellen van een diagnose geen nood heeft aan een
speciaal uitgerust labo. Men kan deze test dus ook gebruiken in het veld. Tevens is deze test zeer
geschikt om Plasmodium op te sporen in gebieden waar de parasiet niet frequent voorkomt,
aangezien geen ervaren onderzoekers nodig zijn om de diagnosemethode uit te voeren (Murray et al.,
2003).
Het nadeel daarentegen is dat de gevoeligheid van de testen behoorlijk laag is. Sommige testen,
zoals de ParaSight®-F test dat het antigen HRP-2 opspoort, zullen enkel P. falciparum kunnen
aanduiden aangezien het antigen enkel bij dit species voorkomt (Shiff et al., 1993; Moody, 2002;
Murray et al., 2003). Andere testen daarentegen hebben als nadeel dat zij P. falciparum niet kunnen
onderscheiden van andere minder pathogene Plasmodium species (Murray et al., 2003).
4.1.4. Real-Time Polymerase Chain Reaction (PCR) test
Deze test spoort de aanwezigheid van genetische materiaal van de parasiet op. De test heeft een
hoge gevoeligheid en sensitiviteit (Mangold et al., 2005). Patiënten met een lage parasitaemie zullen
met behulp van deze methode ook opgespoord kunnen worden (Mangold et al., 2005).
De nadelen echter zijn de hoge kostprijs van de test en de nood aan gespecialiseerde technici
(Rayner et al., 2011).

18
4.1.5. Indirecte testen
Deze testen sporen antistoffen op in het bloed gericht tegen antigenen van Plasmodium. De
antistoffen kunnen aangetoond worden met behulp van indirecte immunofluorescentie of enzym-linked
immunosorbant assay (ELISA).
Deze testen kunnen geen recente infecties aantonen aangezien op dat moment nog geen antistoffen
aangemaakt zijn door het lichaam.
4.2. NIET-INVASIEVE METHODEN
Niet-invasieve methoden werden recent gebruikt bij het onderzoek naar het apen immunodeficiëntie
virus (SIV) (Neel et al., 2009). In dit onderzoek werden faecesstalen van wilde niet-humane primaten
getest op de aanwezigheid van specifieke SIVgor antilichamen en nucleïnezuren (Neel et al., 2009).
Uitgaande van de methoden die men hanteerde bij dit onderzoek, ontdekte men dat ook Plasmodium
opgespoord kan worden in faeces van malariapatiënten wanneer men gebruik maakt van een
conventionele en single-template PCR amplificatie methode (Liu et al., 2010). Het voordeel van zulke
methoden is dat men geen direct contact dient te hebben met de patiënt, hetgeen ideaal is bij het
onderzoeken van wilde dieren. Liu et al. (2010) maakten gebruik van deze methode voor het
uitvoeren van hun grootschalige epidemiologische studie naar malaria in wilde niet-humane primaten.
Alsdusdanig hebben zij de oorsprong van P. falciparum kunnen achterhalen.

19
BESPREKING
Ondanks de steeds groter wordende kennis omtrent malaria, tast de wetenschap op sommige vlakken
nog steeds in het duister. Het vermoeden dat malaria zoönotisch is, is bevestigd. De mate waarin
deze zoönose een gevaar kan vormen voor de gezondheid van de mens is echter nog niet volledig in
kaart gebracht, noch heeft men voor vele malariaspecies voldoende bewijsmateriaal gevonden om
deze aan te duiden als ware zoönosen.
In Zuid-Oost Azië werd voor het eerst duidelijk dat malaria als zoönose een belangrijke rol speelt.
Onderzoekers vonden namelijk P. knowlesi veelvuldig terug in humane patiënten. P. knowlesi
infecties bij mensen kunnen uiterst ernstig zijn, in tegenstelling tot de infecties die het malariaspecies
veroorzaakt bij zijn natuurlijke gastheer, de makaken. Men neemt daarom ook aan dat makaken een
reservoir kunnen vormen voor deze welbepaalde malariaparasiet.
Er zijn nog geen harde bewijzen gevonden om andere malariaspecies van de oude wereld apen aan
te duiden als zijnde zoönotisch.
In Afrika werd de dodelijkste malariaparsiet van de mens, P. falciparum, teruggevonden in
verscheidene wilde mensapen. Dit species zou bovendien ontstaan zijn door een zoönotische infectie
van één mens met een malariaspecies van de gorilla vele jaren geleden.
Naast P. falciparum vond men ook de drie andere humane malariaspecies (P. malariae, P. vivax, P.
ovale) terug in wildlevende chimpansees. Het feit dat humane malariaspecies teruggevonden werden
in mensapen, die waarschijnlijk nooit in contact geweest zijn met de mens, suggereert dat zij een
natuurlijk reservoir vormen voor de humane Plasmodium species. Bovendien vertoonden de
geïnfecteerde mensapen geen ziektesymptomen.
Het is mogelijk dat de humane malariaparasieten afstammen van malariaspecies bij de Afrikaanse
mensapen. Onderzoek zal hierover in de toekomst meer duidelijkheid moeten brengen.
De nieuwe wereldapen in het Amazone regenwoud worden eveneens sterk verdacht van het dragen
van zowel niet-humane malariasoorten die een gevaar kunnen vormen voor de mens, als voor
humane malariaspecies.
Men kan alsdusdanig besluiten dat in elk continent waar malaria heerst, niet-humane primaten met
alle waarschijnlijkheid een reservoir vormen voor malariaspecies die ziekte kunnen veroorzaken bij de
mens. Om die reden dient men in te zien dat maatregelen, die getroffen worden ter uitroeiing van
malaria, zich niet enkel mogen richten op de mens. De reservoirs kunnen namelijk zorgen voor het
continu opnieuw insluipen van malaria bij de mens.
Om het inzicht in deze gehele problematiek te vergroten, is het aangewezen verscheidene
maatregelen te treffen (Rayner et al., 2011).
Eerst en vooral dient men wilde niet-humane primaten te blijven screenen op aanwezigheid van
malaria. Zodanig dat men een beter beeld krijgt van de prevalentie van de Plasmodium spp. in de
wildlevende zoogdieren. Daarenboven kan men op die manier informatie over de sequenties van de
malariaparasieten vergaren, opdat de oorsprong en eventuele vroegere interspecies transmissies

20
kunnen worden aangeduid. Zulke screenings maken het eveneens mogelijk nieuwe
malariapararsieten te ontdekken (Rayner et al., 2011).
Naast het screenen van niet-humane primaten dienen ook mensen die leven in risicogebieden getest
te worden op de aanwezigheid van zoönotische infecties. Moderne diagnostische technieken zullen
hierbij nodig zijn om zoönotische species te kunnen onderscheiden van humane species. Bovendien
zal men zo ook bij gemengde infecties, die veelvuldig voorkomen, de afzonderlijke malariaspecies
kunnen identificeren (Rayner et al., 2011).
Tenslotte zal een meer uitgebreide kennis over de vectoren van malaria de wetenschap helpen in zijn
queeste tegen malaria.

21
REFERENTIES
Baird J.K. (2009). Malaria zoonoses. Travel Medicine and Infectious Disease 7, 269-277
Bennett B.T., Abee C.R., Henrickson R. (1998). Nonhuman primates in biomedical research.
Academic Press. p 124-126
Carter R., Mendis K.N. (2002). Evolutionary and historical aspects of the burden of malaria. Clinical
microbiology reviews 15(4), 564-594
Chin W., Contacos P.G., Coatney G.R., Kimball H.R (1965). A naturally acquired quotidian-type
malaria in man transferable to monkeys. Science. 149, 865
Coatney G.R., Chin W., Contacos P.G., King H.K. (1966). Plasmodium inui a quartan-type malaria
parasite of Old world monkeys transmissible to man. Journal of parasitology 52, 660-663
Coatney G.R. (1971). The simian malarias: zoonoses, anthroponoses or both? The American journal
of tropical medicine and hygiene 20, 795-803
Coatney G.R., Collins W.E., Contacos P.G. (1971). The primate malarias. US government printing
office. Washington
Collins W.E., Jeffery G.M. (2007). Plasmodium malariae: parasite and disease. Clinical microbiology
reviews 20(4), 579-592
Contacos P.G., Lunn J.S., Coatney G.R., Kilpatrick J.W., Jones F.E. (1963). Quartan type malaria
parasites of New World monkeys transmissible to man. Science 142, 676
Cormier L.A. (2010). The historical ecology of human and wild primate malarias in the new world.
Diversity 2, 256-280
Cox-Singh J., Davis T.M.E., Lee K.-S., Shamsul S.S.G., Matusop A., Ratnam S., Rahman H.A.,
Conway D.J., Singh B. (2008). Plasmodium knowlesi malaria in humans is widely distributed and
potentially life threatening. Clinical Infectious Diseases 46,165–171
Cox-Singh J., Singh B. (2008). Knowlesi malaria: newly emergent and of public health importance?
Trends in Parasitology. 24(9), 406-410
Damodar S.U. (1996). Evaluation of acridine - orange staining of centrifuged parasites in
malarialinfection. Indian journal of medical sciences 50(7), 228-30
Das Gupta B.M. (1938). Transmission of Plasmodium inui to man. Proceedings of the National
Institute of Sciences of India 4, 241-244
Deane L.M., Deane M.P., Neto J.F. (1966). Studies on transmission of simian malaria and report of a
natural infection of man with Plasmodium simium in Brazil. Bulletin of the World Health Organization
35, 805-808

22
De jonge N., Polderman A.M., Verhave J.P. (1999). Diagnose van malaria. Nederlands tijdschrift
klinische chemie 24, 34-37
Duval L., Nerrienet E., Rousset D., Sadeuh Mba S.A., Houze S., Fourment M., Le Bras J., Robert V.,
Ariey F. (2009). Chimpanzee malaria parasites related to Plasmodium ovale in Africa. PLoS
pathogens 4, e5520
Duval L., Fourment M., Nerrienet E., Rousset D., Sadeuh S.A., Goodman S.M., Andriaholinirina N.V.,
Randreianarivelojosia M., Paul R.E., Robert V., Ayala F.J., Ariey F. (2010). African apes as reservoirs
of Plasmodium falciparum and the origin and diversification of the Laverania subgenus. Proceedings
of the national Academy of Sciences (PNAS) 107, 10561-10566
Escelanta A.A., Ayala F.J. (1994). Phylogeny of the malarial genus Plasmodium, derived from rRNA
gene sequences. Proceedings of the National Academy of Sciences of the United States of America
91, 11373-11377
Escelanta A.A., Barrio E., Ayala F.J. (1995). Evolutionary origin of human and primate malarias:
Evidence from the circumsporozoite protein gene. Molecular Biology and Evolution 12 616-626
Eyles D.E., Coatney G.R., Getz M.E. (1960). Vivax-type malaria parasite of macaques transmissible to
man. Science 132, 1812-1813
Fandeur T., Volney B., Peneau C., de Thoisy B (2000). Monkeys of the rainforest in French Guiana
are natural reservoirs for P. brasilianum/P. malariae malaria. Parasitology 120, 11-21
Galinski M.R., Barnwell J.W. (2009). Monkey malaria kills four humans. Trends in Parasitology 25,
200-204
Holmes E.C. (2010). Malaria: The gorilla connection. Nature 467, 404-405
Kaiser M., Löwa A., Ulrich M., Ellerbrok H., Goffe A.S., Blasse A., Zommers Z., Couacy-Hymann E.,
Babweteera F., Zuberbühler K., Metzger S., Geidel S., Boesch C., Gillespie T.R., Leendertz F.H.
(2010). Wild chimpanzees infected with 5 Plasmodium species. Emerging infectious diseases CDC
12, 1956-1959
Kappe S.H.I., Vaughan A.M., Boddey J.A., Cowman A.F. (2010). That was then but this is now:
malaria research in the time of an eradication agenda. Science 328(862), 862-866
Knowles R., Das Gupta B.M. (1932). A study of monkey-malaria, and its experimental transmission to
man. Indian Medical Gazette 67, 301-320. Bron: Coatney G.R., Collins W.E., Contacos P.G. (1971).
The primate malarias. US government printing office. Washington.
Krief S., Escalante A.A., Pacheco M.A., Mugisha L., André C., Halbwax M., Fischer A., Krief J-M.,
Kasenene J.M., Crandfield M., Cornejo O.E., Chavatte J-M., Lin C., Letourneur F., Grüner A.C.,
McCutchan T.F., Rénia L., Snounou G. (2010). On the diversity of malaria parasites in African apes
and the origin of Plasmodium falciparum from bonobos. PLoS pathogens 6, e1000765

23
Krotoski W.A., Collins W.E., Bray R.S., Garnham P.C.C., Cogswell F.B., Gwadz R.W., Killick-Kendrick
R., Wolf R., Sinden R., Koontz L.C., Stanfill P.S. (1982). Demonstration of hypnozoites in sporozoite-
transmitted Plasmodium vivax infection. The American journal of tropical medicine and hygiene 31(6),
1291-1293
Lee K.S., Divis P.C.S., Zakaria S.K., Matusop A., Julin R.A., Conway D.J., Cox-Singh J., Singh B.
(2011). Plasmodium Knowlesi: Reservoir hosts and tracking the emergence in humans and
macaques. PLoS pathogens 7, e1002015
Liu W., Li Y., Learn G.H., Rudicell R.S., Robertson J.D., Keele B.F., Ndjango J.N., Sanz C.M., Morgan
D.B., Locatelli S., Gonder M.K., Kranzusch P.J., Walsh P.D., Delaporte E., Mpoudi-Ngole E., Georgiev
A.V., Muller M.N., Shaw G.M., Peeters M., Sharp M.P., Rayner J.C., Hahn B.H. (2010). Origin of the
human malaria parasite Plasmodium falciparum in gorillas. Nature 467, 420-425
Mangold K.A., Manson R.U., Koay E.S.C., Stephens L., Regner M., Thomson R.B., Peterson L.R.,
Kaul K.L. (2005). Real-Time PCR for detection and identification of Plasmodium spp. Journal of
clinical microbiology 43(5), 2435-2440
Moody A. (2002). Rapid diagnostic test for Malaria Parasites. Clinical microbiology reviews. 15(1), 66-
78
Murray C.K., Bell D., Gasser R.A., Wongsrichanalai C. (2003). Rapid diagnostic testing for malaria.
Tropical Medicine and International Health 8(10), 876–883
Neel C., Etienne L., Li Y., Takehisa J., Rudicell R.S., Bass I..N., Moudindo J., Mebenga A., Esteban
A., Van Heuverswyn F., Liegeois F., Kranzusch P.J., Walsh P.D., Sanz C.M., Morgan D.B., Ndjango
J.-B. N., Plantier J.-C., Locatelli S., Gonder M.K., Leendertz F.H., Boesch C., Todd A., Delaporte E.,
Mpoudi-Ngole E. (2009). Molecular epidemiology of simian immunodeficiency virus infection in wild-
living gorillas. Journal of Virology 84(3) 1464–1476
Nelson K.E., Williams C.M. (2006). Infectious Disease Epidemiology, Theory and practice, 2nd
ed.
Jones and Bartlett publishers. p1113-1114
Prugnolle F., Durand P., Neel C., Ollomo B., Ayala F.J., Arnathau C., Etienne L., Mpoudi-Ngole E.,
Nkoghe D., Leroy E., Delaporte E., Peeters M., Renaud F. (2010). African great apes are natural
hosts of multiple related malaria species, including Plasmodium falciparum. PNAS 107, 1458-1463
Rayner J.C., Liu W., Peeters M., Sharp P.M., Hahn B.H. (2011). A plethora of Plasmodium species in
wild apes: a source of human infection? Trends in Parasitology 27(5), 222-229
Rhodain J. (1940). Les plasmodiums des anthropoids de l’Afrique Centrale et leur relations avec les
plasmodiums humains. Réceptivité de l’homme au Plasmodium malariae (Plasmodium rhodaini
Brumpt) du chimpanzee. C. R. Soc. Biol. 133, 276-277
Rhodain J., Dellaert R. (1955). L’infection à Plasmodium schwetzi chez l’homme. Annales de la
Societé Belge de Médecine Tropicale. 35, 757-776

24
Rich S.M., Leendertz F.H., Xu G., LeBreton M., Djoko C.F., Aminake M.N., Takang E.E., Diffo J.L.D.,
Pike B.L., Rosenthal B.M., Formenty P., Boesch C., Ayala F.J., Wolfe N.D. (2009). The origin of
malignant malaria. PNAS 106, 14902-14907
Ruch T.C. (1959). Disease of laboratory primates. W.B. Saunders Company, Philadelphia, London
p329
Schindler H.C., Montenegro L., Mentenegro R., Carvalho A.B., Abath F.G.C., Jaureguiberry G. (2001).
Development and optimization of polymerase chain reaction-based malaria diagnostic methods and
their comparison with quantitative buffy coat assay. The American journal of tropical medicine and
hygiene 65(4), 355-361
Service M.W., Townson H. (2002): The anopheles vector. In Essential Malariology, fourth edition,
Oxford University press: New York
Shiff C.J., Premji Z., Minjas J.N. (1993). The rapid manual ParaSight®-F test. A new diagnostic tool for
Plasmodium falciparum infection. Transactions of the Royal Society of Tropical Medicine and Hygiene.
87 646-648
Singh B., Lee K.S., Matusop A., Radhakrishnan A., Shamsul S.S.G., Cox-Singh J. (2004). A large
focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet 363, 1014-1024
Tan C.H., Vythilingam I., Matusop A., Chan S.T., Singh B. (2008). Bionomics of Anopheles lateens in
Kapit, Sarawak, Malaysian Borneo in relation to the transmission of zoonotic simian malaria parasite
Plasmodium knowlesi. Malaria Journal 7, 52
Taylor M., Coop R., Wall R. (2007). Veterinary Parasitology. Third edition. Blackwell Publishing Ltd.
Oxford
Tek F.B., Dempster A.G., Kale I. (2010). Parasite detection and identification for automated thin blood
film malaria diagnosis. Computer Vision and Image understanding 114(1), 21-32
Vythilingam I., Tan C.H., Asmad M., Chan S.T., Lee K.S., Singh B. (2006). Natural transmission of
Plasmodium knowlesi to humans by Anopheles lateens in Sarawak, Malaysia. Transactions of the
Royal Society of Tropical Medicine and Hygiene 100(11), 1087-1088
Warren M., Cheong W.H., Fredericks H.K., Coatney G.R. (1970). Cycles of jungle malaria in west
Malaysia. American Journal of Tropical Medicine and Hygiene 19, 383-393
WHO (2011). World malaria report 2011. World Health Organization, Geneva














![Ontwerp van stents met pyFormex - Ghent Universitylib.ugent.be/fulltxt/RUG01/001/312/417/RUG01... · [1] P. Mortier, Eindige elementen simulatie van ballon-stent interactie, 2006](https://static.fdocuments.nl/doc/165x107/604547f01b3cb8607238015e/ontwerp-van-stents-met-pyformex-ghent-1-p-mortier-eindige-elementen-simulatie.jpg)



