GENETISCHE RELATIE TUSSEN SCHISIS EN … · palatum wordt gevormd uit dezelfde processi die zorgen...
47
Annelies Kellens, 3 e jaar tandarts. Genetische relatie tussen schisis en tandagenesie? GENETISCHE RELATIE TUSSEN SCHISIS EN TANDAGENESIE? Annelies Kellens Verhandeling aangeboden tot het behalen van het diploma van Tandarts Promotor: Prof. C. Carels - 2008 - KATHOLIEKE UNIVERSITEIT LEUVEN FACULTEIT GENEESKUNDE SCHOOL VOOR TANDHEELKUNDE, MONDZIEKTEN EN KAAKCHIRURGIE
Transcript of GENETISCHE RELATIE TUSSEN SCHISIS EN … · palatum wordt gevormd uit dezelfde processi die zorgen...

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
GENETISCHE RELATIE TUSSEN SCHISIS EN TANDAGENESIE?
Annelies Kellens
Verhandeling aangeboden tot
het behalen van het diploma
van Tandarts
Promotor: Prof. C. Carels
- 2008 -
KATHOLIEKE UNIVERSITEIT
LEUVEN
FACULTEIT GENEESKUNDE SCHOOL VOOR TANDHEELKUNDE, MONDZIEKTEN EN KAAKCHIRURGIE

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
ANNELIES KELLENS
Genetische relatie tussen schisis en tandagenesie?
Promotor: Prof. C. Carels
- 2008 -
FACULTEIT GENEESKUNDE SCHOOL VOOR TANDHEELKUNDE, MONDZIEKTEN EN KAAKCHIRURGIE
KATHOLIEKE UNIVERSITEIT
LEUVEN

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
I
Dankwoord
Dank aan Prof C. Carels voor de steun en begeleiding bij het schrijven van mijn capita.
Dank aan mama en papa voor jullie steun en geloof in alles wat ik doe, dank u voor alle
kansen die jullie mij gegeven hebben. Dank u mama voor alle goede zorgen
tijdens de blok- en examenperiodes. Dank u voor de steun bij het tot stand komen
van mijn capita.
Dank aan broers en zus voor alle fijne momenten en voor de steun van de afgelopen
jaren.
Dank aan Kristof voor al zijn steun, vriendschap en liefde. Bedankt om er altijd voor
mij te zijn.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
II
Inhoudsopgave Algemene Inleiding…………………………………………………………………...…1
Hoofdstuk 1 : Schisis ....................................................................................................... 2
1.1 Definitie ............................................................................................................ 2
1.2 Prevalentie en incidentie ................................................................................... 3
1.3 Normale ontwikkeling van de kaken................................................................. 4
1.4 Etiologie ............................................................................................................ 4
1.4.1 Erfelijkheid ................................................................................................ 5
1.4.2 Exogene factoren ....................................................................................... 9
1.5 Herhalingsrisico .............................................................................................. 11
Hoofdstuk 2 : Agenesie ................................................................................................ 13
2.1 Defintie ............................................................................................................ 13
2.2 Prevalentie ....................................................................................................... 13
2.3 Normale tandontwikkeling .............................................................................. 14
2.4 Etiologie van agenesie..................................................................................... 16
2.4.1 Erfelijkheid .............................................................................................. 16
2.4.2 Exogene factoren ..................................................................................... 19
Hoofdstuk 3 : Onderzoek naar de erfelijkheid van schisis en agenesie ....................... 20
3.1 Tweelingenonderzoek ..................................................................................... 20
3.1.1 Waarom tweelingenonderzoek? .............................................................. 20
3.1.2 Zygositeit van de tweeling ...................................................................... 20
3.1.3 Soorten tweelingenonderzoek ................................................................. 22
3.2 Resultaten van tweelingenonderzoek in verband met schisis ......................... 25
3.3 Resultaten van tweelingenonderzoek in verband met agenesie ...................... 26
Hoofdstuk 4 : Zijn schisis en agenesie genetisch gerelateerd? .................................... 29
4.1 Studies naar een genetische link tussen niet-syndromale schisis en agenesie 29
4.1.1 Studies met muizen ................................................................................. 29
4.1.2 Familie-onderzoek .......................... Fout! Bladwijzer niet gedefinieerd.
4.2 Chromosomale afwijkingen en single gene disorders ..................................... 33
4.2.1 Van der Woude syndroom ...................................................................... 33

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
1
Algemene inleiding
Schisis en tandagenesie zijn frequent voorkomende congenitale afwijkingen. Het is dan
ook logisch dat veel onderzoek gebeurt naar de etiologie van beide afwijkingen.
Voornamelijk genetische maar ook omgevingsfactoren worden onderzocht.
Tweelingenonderzoek is één methode om de relatieve bijdrage van genetische en
omgevingsfactoren te ontrafelen, vooral bij multifactorieel bepaalde afwijkingen. Ook zijn
deze onderzoeken van grote waarde voor de analyse van de discordantie tussen een
tweelingenpaar voor een bepaalde afwijking.
Doel van dit werk is om de huidige stand van zaken over de kennis omtrent de etiologie
van schisis en agenesie te beschrijven. Tevens zoek ik in de wetenschappelijke literatuur
naar evidentie over een eventuele genetische link tussen beide kenmerken.
Ik start met een overzicht van de etiologie van schisis. Vervolgens ga ik dieper in op de
pathogenese van tandagenesie en behandel ik tweelingenonderzoek om te eindigen met
een bespreking van studies die een genetische link tussen beide afwijkingen suggereren.
Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
2
Hoofdstuk 1 : Schisis
1.1 Definitie
Schisis is een Grieks woord en betekent spleet/splijting (Stegenga. 2000). Een spleet wijst
op een stoornis in de ontwikkeling van het embryo (Devriendt et al. 2008).
In medische zin gebruikt men deze term voor een groep aangeboren afwijkingen waarbij
de lip, kaak en/of gehemelte gespleten kunnen zijn (Stegenga. 2000).
Orofaciale spleten worden ingedeeld in syndromaal of niet-syndromaal (geïsoleerd),
afhankelijk van de aanwezigheid van nog andere congenitale defecten of
ontwikkelingsstoornissen (Lidral et al. 2004).
De spleten kunnen uni- of bilateraal voorkomen en kennen meerdere verschijningsvormen
(Stegenga. 2000):
Lipspleet (cheiloschisis)
Lip– en kaakspleet (cheilognathoschisis)
Lip–, kaak– en gehemeltespleet (cheilognatopalatoschisis)
Geïsoleerde gehemeltespleet (palatoschisis)
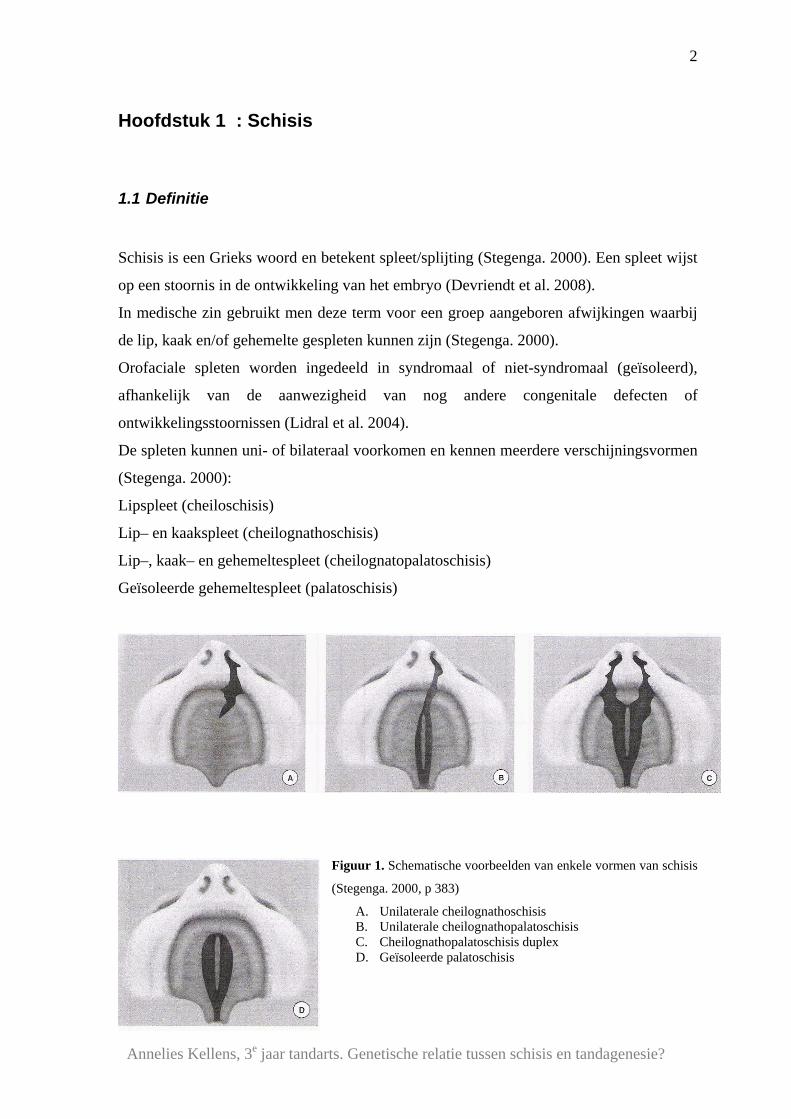
Figuur 1. Schematische voorbeelden van enkele vormen van schisis
(Stegenga. 2000, p 383)
A. Unilaterale cheilognathoschisis B. Unilaterale cheilognathopalatoschisis C. Cheilognathopalatoschisis duplex D. Geïsoleerde palatoschisis

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
3
1.2 Prevalentie en incidentie
Lip, kaak en/of gehemeltespleten behoren tot de meest frequente geboortedefecten. De
incidentie van lip, lipkaak en lipkaakgehemelte spleet bij pasgeborenen is ongeveer 1 op
1000 per jaar en doet zich 2 keer zoveel voor bij jongens dan bij meisjes (Stegenga. 2000).
Geïsoleerde gehemeltespleet is minder voorkomend, volgens Lidral et al. (2004) ongeveer
1 op 5000 volgens Stegenga. (2000) ongeveer 1 op 2000 geboortes. Het komt meer voor
bij het vrouwelijk geslacht (Lidral et al. 2004, Stegenga. 2000).
In 50% van de gevallen gaat het om een gecombineerde vorm van schisis (lipkaak of
lipkaakgehemelte spleet (CL/P)). Een geïsoleerde gehemeltespleet (CP) of lipspleet (CL)
doet zich voor in 25% van de gevallen (Stegenga. 2000).
De prevalentie van orofaciale schisis is afhankelijk van de etnische achtergrond,
geografische origine (hoogste incidentie bij de Aziatische bevolking) en socio–
economische status (Lidral et al. 2004, Van den Boogaard et al. 2000, Warrington et al.
2006).
Syndromale schisis zien we voor lip, lipkaak en lipkaakgehemelte spleet in 1% en voor
gehemeltespleet in 8% van de gevallen. In de andere 99% en 92% doet schisis zich voor
als een niet-syndromale afwijking (Stegenga. 2000). Bij deze individuen zien we bepaalde
dentale karateristieken, waaronder hypodontie, frequenter dan bij gewone individuen
(Eerens et al. 2001).
Uitzonderlijke gevallen zoals mediane lipspleet komen maar zelden voor (Van Esch et al.
2008).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
4
1.3 Normale ontwikkeling van de kaken
Het basisprincipe van de normale processen die betrokken zijn bij de vorming van
gehemelte en gelaat, is het met elkaar versmelten van uitgroeiende mesenchymale
processi (Stegenga. 2000).
In het gelaat ontstaan deze zwellingen in de loop van de 5de en 6de week van de
embryonale ontwikkeling. De processus nasalis lateralis, processus nasalis medialis,
processus maxillaris en processus mandibularis fusioneren zowel mediaan als
paramediaan waaruit de structuren van neus, filtrum, bovenlip, bovenkaak en onderkaak
worden gevormd (Van Esch et al. 2008, Stegenga. 2000).
Rond de 12de week ontstaat het gehemelte op gelijke wijze. Het primaire deel van het
palatum wordt gevormd uit dezelfde processi die zorgen voor de vorming van neus en
filtrum. Het secundaire palatum ontstaat door de fusie van de processi palatini (Van Esch
et al. 2008, Stegenga. 2000).
Het verhemelte, de bovenste processus alveolaris en de bovenlip worden dus deels uit
dezelfde structuren gevormd. Dit verklaart waarom een lipspleet samen kan voorkomen
met een kaakspleet en/of met een verhemeltespleet (Devriendt et al. 2008).
1.4 Etiologie
De sluiting van lip, kaak en gehemelte is één van de meest complexe morfogenetische
processen. Een spleet ter hoogte van deze structuren is het resultaat van een onvolledige of
afwezige fusie van processi. Deze onvolledige fusie vindt meestal niet plaats op de
middenlijn maar lateraal ervan, zowel links en rechts waar deze twee structuren met elkaar
versmelten (Devriendt et al. 2008).
Schisis ontstaat reeds vroeg in de zwangerschap aangezien het gelaat en gehemelte zich in
de vroeg–embryonale periode ontwikkelen (tussen de drieëneenhalf en twaalf weken)
(Stegenga. 2000).
Naar oorzaak van niet–syndromale orofaciale schisis onderscheiden we twee aparte
aandoeningen. Enerzijds bestaat er een verhemeltespleet (CP), anderzijds is er een
aandoening waarbij er steeds een lipspleet is, al of niet gecomplexeerd met een kaak-

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
5
en/of verhemeltespleet (CL/P) (Devriendt et al. 2008). Het foramen incisivum vormt in
feite het scheidingspunt tussen de lipkaakspleten en de gehemeltespleten. Spleten die
zowel voor als achter dit foramen liggen noemt men de lipkaakgehemeltespleten
(Devriendt et al. 2008, Stegenga. 2000)
Er wordt dus een onderscheid gemaakt tussen syndromale en niet-syndromale schisis.
Men neemt aan dat beide een verschil hebben in de etiologie (Lidral et al. 2004).
Syndromale schisis heeft een Mendeliaanse wijze van overerving en is dus makkelijk te
linken aan ziektegenen (Lidral et al. 2004). De niet-syndromale schisis daarentegen wordt
algemeen verondersteld familiaal te zijn maar er is geen eenvoudig patroon van
overerving gekend. Bij CL/P is de oorsprong meer complex en verloopt de genetische
overdracht niet volgens de klassieke regels van Mendel. Niet–syndromale schisis wordt
gekarakteriseerd door een genetische heterogeniciteit, ongetwijfeld veroorzaakt door
verschillende factoren, waaronder zowel genetische- als omgevingsfactoren (Karsten et al.
2005, Mitchell et al. 2002). Meerdere epidemiologische studies hebben de invloed van
beide op het risico van geboortedefecten, waaronder schisis, aangetoond (Christensen et
al. 1992, Devriendt et al. 2008, Modesto et al. 2006, Van den Boogaard et al. 2000). De
genetische invloed op schisis is reeds geruime tijd gekend. Hoewel duidelijk is dat deze
factoren een belangrijke oorzakelijke rol hebben, is de bepaling van de invloed van de
genetica versus die van de externe factoren in de etiologie van schisis moeilijk (Nordstrom
et al. 1996).
1.4.1 Erfelijkheid
Eén van de belangrijkste oorzaken van schisis is een afwijking in het genetisch materiaal.
Bij de overerving kan sprake zijn van multifactoriële, monofactoriële en chromosomale
storingen/afwijkingen (Stegenga. 2000).
Multifactoriële erfelijkheid
De sluiting van lip,kaak en gehemelte is een ingewikkeld proces en mogelijks zijn hierbij
honderden genen betrokken. De optelsom van de verschillende bijdrage van deze genen
bepaalt het ontwikkelingsproces van de bovengenoemde structuren. Men spreekt van een
additief multifactorieel bepaald ontwikkelingsproces (Stegenga. 2000).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
6
Hoewel er geen specifieke oorzakelijke gen mutaties geïdentificeerd zijn bij niet–
syndromale schisis, zijn er wel een aantal kandidaat genen (Karsten et al. 2005). Recent
onderzoek suggereert dat 3 tot 14 interagerende genen een goed model voorzien voor de
genetische effecten in schisis (Vieira et al. 2005, Van den Boogaard et al. 2000, Modesto
et al. 2006). In veel van deze genen kunnen varianten voorkomen. Deze resulteren niet
zonder meer in pathologische gevolgen omdat de overige normale genen dit kunnen
opvangen. Het compensatiemechanisme is echter niet gelimiteerd. Wanneer de bijdrage
van de gezonde genen in de groep afneemt tot beneden een bepaalde drempel, wordt het
genetische defect klinisch zichtbaar. Dit wil dus zeggen dat elke variant maar in beperkte
mate bijdraagt tot het ontstaan van schisis en dat deze kleine defecten vaak moeilijk op te
sporen zijn. Het voorkomen van een CL/P en CP is afhankelijk van de additieve effecten
van verschillende genen, elk met een minor effect samen met één of meerdere
omgevingsfactoren (Nordstrom et al. 1996). Onderzoek naar al deze varianten is
noodzakelijk maar dan nog is de aanwezigheid van een genetische variant op zich
onvoldoende om de oorzaak te bepalen (Devriendt et al. 2008).
Monogenetische overerving
Er zijn ook een aantal individueel functionerende genen betrokken bij de morfogenese van
de bovenlip, kaak en gehemelte. Omdat deze genen ook op andere plaatsen in het
organisme van specifieke betekenis zijn, zal een mutatie van deze genen zich klinisch
manifesteren als een erfelijke aandoening met een karakteristiek patroon en afwijkingen.
Men noemt dit een pleiotroop gendefect. Dit verklaart het optreden van schisis als
onderdeel van een groot aantal syndromen en aangeboren afwijkingen. De
overdrachtspatronen van dit soort mutaties volgen de klassieke Mendeliaanse modellen
van autosomale of geslachtsgekoppelde dominante of recessiviteive overerving. Bij
familiaal aanwezige afwijkingen zijn de herhalingsrisico’s in de orde van grootte van 25-
50% en, anders dan bij de multifactoriële erfelijkheid, bovendien constant (Stegenga.
2000).
Vb: IRF 6 (Van der Woude syndroom) (Lidral et al. 2004).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
7
Chromosomale afwijkingen
Uit het voorgaande blijkt dat één genmutatie mogelijk ingrijpende pathologische gevolgen
kan hebben. Ook kwantitatieve afwijkingen van één chromosoom, zoals vb trisomie,
zonder verandering van de op dit chromosoom aanwezige genen, kunnen uitgebreide
aangeboren afwijkingen veroorzaken. Hoe complexer een ontwikkelingsproces is, hoe
groter de kans dat genen, die dit proces mee sturen, op vele van de 23 chromosomenparen
aanwezig zijn. Het is dan ook niet verwonderlijk dat schisis bij een groot aantal
chromosomale syndromen wordt aangetroffen. Palatoschisis komt in samenhang met deze
syndromen vaker voor dan lip, kaak en lipkaakgehemelte schisis (Stegenga. 2000).
Syndromale schisis is dus het gevolg van monogenetische overerving of chromosomale
afwijkingen. Vele auteurs zijn echter geïnteresseerd in de etiologie van niet-syndromale
schisis. Dat verschillende genen en omgevingsfactoren aan de basis liggen van deze
afwijking is reeds geweten. Verschillende studies toonden reeds een aantal genen aan die
geassocieerd kunnen worden met het voorkomen van niet-syndromale orofaciale schisis.
Van den Boogaard et al. (2000) screende verschillende kandidaat genen voor orofaciale
schisis, onder andere TGFA, BCL3, DLX2, MSX1 en TGFB3. Associatie werd gevonden
tussen CL/P met MSX1 en TGFB3, ook tussen CP en MSX1. Vieira et al. (2005) vond bij
het screenen van 20 mogelijk oorzakelijke genen resultaten die dit bevestigde. Verder
werden SPRY2 en TBX10 ook geassocieerd met schisis en werd er daarenboven een link
aangetoond tussen FGFR1 met zowel de syndromale als de geïsoleerde vorm van schisis.
Ook in de studie van Etheredge et al. (2005) kwamen mutaties in MSX1, TGF en RARA
in de cases met CL/P aan bod.
Warrington et al. (2006) vond mutaties in de genen TGFA, MSX1, TGFB3, RARA,
CLPTM1, BCL3 die geassocieerd kunnen worden met schisis.
MSX1 is een non-clustered homeobox gen en afwijkingen van dit gen zijn mogelijk één
van de belanrijkste oorzaken voor niet-syndromale CL/P bij mensen (Modesto et al.
2006).
Doorheen de jaren zijn er zowel studies op mensen als dieren die een associatie
aantoonden tussen schisis en MSX1 (Modesto et al. 2006, Lidral et al. 2004). Zowel
vormen van familiale schisis (Vastardis et al. 1996) als geïsoleerde schisis (Jezewski et al.
2003) werden gelinkt met dit gen.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
8
Verder tonen data van genetische studies een bijdrage van het MSX1 gen aan bij CL/P in
combinatie met agenesie (Slatyton et al. 2003) en bij geïsoleerde agenesie (Viera et al.
2004).
Vieira et al. (2005) vond bij het screenen van kandidaat genen bij cases verschillende
mutaties van MSX1 die niet gevonden werden bij de meer dan 1000 controles.
Afwijkingen van de genen SPRY2 en TBX10 zijn ook reeds geassocieerd met schisis.
Viera et al. (2005) vond mutaties van beide genen bij een aantal van de cases maar ook bij
een aantal controles waar geen orofaciale schisis aanwezig was. Dit kan er op wijzen dat
deze mutatie een toegevoegde functie heeft in de etiologie van CL/P. Warrington et al.
(2006) daarentegen vond dezelfde mutaties ook terug bij een aantal cases en bij geen
enkele van de controles.
De associatie van het TGFA gen met CL/P is zeer variabel. Vieira et al. (2005) steunde
deze associatie, zij het als een kleiner risico. Etheredge et al. (2005) vond verschillende
varianten van dit gen in cases met CL/P. Vergeleken met eerdere studies (Chenevix-
Trench et al. 1992, Ardinger et al. 1989) zijn er bepaalde varianten die vaker voorkomen
in associatie met schisis dan andere.
Van IRF6 is bewezen is dat het een sterk oorzakelijke kandidaat is voor een vorm van
syndromale schisis, namelijk het Van der Woude syndroom (VWS). Het leidt ertoe om te
kijken op genetisch niveau of IRF6 ook direct kan betrokken zijn bij niet-syndromale
schisis of dat het kan dienen als “modifier”, een gen dat niet uit zichzelf de oorzaak is van
de spleet maar wel de graad van ernstigheid bepaalt. Aangezien er zoveel klinische
overlap is tussen beide, verwachten we dat gelijkaardige genen en zelfs dezelfde genen
een rol kunnen spelen bij zowel de syndromale als de niet syndromale orofaciale spleten
(Kondo et al. 2002).
Het gen behoort tot een familie van genen die betrokken zijn bij de immuunrespons aan
virale infecties. Murray et al. (2002) is op zoek naar virale infecties tijdens de vroege
zwangerschap die een rol zouden kunnen spelen in de oorzaak van CL/P en CP. Maar daar
is tot op heden nog geen enkel sterk bewijs voor.
Vieira et al. (2005) had eerder al aangehaald dat IRF6 een mogelijke veroorzaker was
voor het VWS, maar volgens een andere studie van Vieria et al. (2005) kon het ook een
modificerende functie uitoefenen bij geïsoleerde schisis.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
9
Verder zouden volgens verschillende studies één of meerdere genen op 19q13 een
oorzakelijke rol kunnen spelen (Modesto et al. 2006).
PVR is een menselijk poliovirus receptor waarvan gezegd wordt dat het een rol zou
kunnen spelen in de etiologie van niet–syndromale CL/P (Warrington et al. 2006).
PVRL1 is een poliovirus receptor related 1 en wordt vaak gelinkt aan het ontstaan van
ectodermale dysplasie en syndromale schisis. Data suggereren dat wanneer men
heterozygoot zou zijn voor een mutatie, men dan een verhoogd risico is voor niet-
syndromische schisis (Warrington et al. 2006).
PVRL2 is een poliovirus receptor related 2 die oorspronkelijk gevonden werd bij
knaagdieren, homoloog aan het menselijk PVR. Het is een kandidaat gen die mogelijk een
rol speelt in de etiologie van CL/P waarvan de functie niet gekend is (Warrington et al.
2006).
Andere genen op 19q13 die mogelijk verband houden met de pathogenese van orofaciale
schisis zijn CLPTM, BCL3 en APOC2 (Warrington et al. 2006).
Er zijn heel wat verschillen in de resultaten van de onderzoeken, meestal is dit een
reflectie van het verschil in de studiepopulatie. Ook zijn er verschillende methodes voor
een genetische benadering tot identificatie van de ziektegenen (Lidral et al. 2004, Mitchell
et al. 2002).
1.4.2 Exogene factoren
Tot de exogene factoren die een invloed kunnen hebben op de etiologie van schisis
behoren onder andere de periconceptale gezondheid van beide ouders, inname van
medicatie, voedingsgewoonte van de moeder en alles wat doorgaat als risicogedrag
(roken, alcoholmisbruik…) van zowel de vader als moeder tijdens de periconceptieperiode
(Karpels et al. 2006).
Omgevingsfactoren op zich zijn niet schadelijk, enkel in combinatie met genetische
factoren kunnen zij tot een afwijking leiden. Het zijn stoffen waaraan we allemaal bloot
gesteld worden tijdens de prenatale periode. Enkel bij genetisch gevoelige embryo’s

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
10
zullen deze “schadelijk” stoffen mogelijks bijdragen tot een aangeboren afwijking
(Devriendt et al. 2008).
Toch zijn er een aantal gevallen waar zuivere milieu-effecten een rol spelen bij het
ontstaan van schisis, dit is echter uitzonderlijk (Devriendt et al. 2008). Meerdere door
exogene effecten veroorzaakte syndromen en andere aangeboren afwijkingen, waarvan
schisis een onderdeel vormt, zijn beschreven. Vb foetale methotrexaat, foetaal
alcoholsyndromen. Reeds 25 jaar staat de invloed van foliumzuur (in het bijzonder de
foliumzuurdeficiëntie) op het ontstaan van schisis sterk in de belangstelling (Stegenga.
2000).
De studie van Etheredge et al. (2005) wou de invloeden van gen–omgevingsinteractie op
schisis aantonen. Dit is niet zo simpel omdat er voor zo een case–control studie een groot
aantal patiënten noodzakelijk is en het vaak moeilijk is om individuele factoren te
achterhalen die van invloed kunnen zijn op de afwijking. Hoewel Etheredge et al. (2005)
geen duidelijk bewijs kon leveren dat een bepaald genotype als oorzaak moest aangehaald
worden of dat het risico voor CL/P beïnvloedt werd door de gen-omgeving interacties,
stelde hij toch de volgende zaken vast: het risico op CL/P steeg bij nakomelingen van
vrouwen die rookten in vergelijking met vrouwen die niet rookten (Christensen et al.
1999, Wyszynski et al. 1997) en het daalde bij de nakomelingen van vrouwen die alcohol
gebruikten in vergelijking met de nakomelingen van vrouwen die geen alcohol gebruikten
(Mitchell et al. 2003). Dit is tegenstrijdig met een studie van Munger et al. (1996) waar
een hoger risico was voor geïsoleerde orofaciale spleten bij vrouwen die alcohol
gebruikten tijdens de zwangerschap. Ook zo de nakomelingen van vrouwen die dagelijks
multivitaminen namen in vergelijking met vrouwen die dit niet dagelijks namen (Mitchell
et al. 2003). Veel vrouwen veranderen wel hun “gedrag“ zodra ze weten dat ze zwanger
zijn. Daarom moet men een goed onderscheid maken tussen de eerste maanden van de
zwangerschap en de daarop volgende periode (Ehteredge et al. 2005, Christensen et al.
1999).
De volgende tabel toont ons de waardes die Etheredge et al. (2005) vond bij het screenen
van cases en controls voor verschillende kandidaat genen van schisis in combinatie met
roken, alcohol en gebruik van vitaminen.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
11
Tabel 1. Genotype verdeling van cases en controles bij blootstelling aan bepaalde omgevingsfactoren
(Etheredge et al. 2005)
1.5 Herhalingsrisico
De grote genetische heterogeniciteit van schisis heeft duidelijke implicaties voor het
herhalingsrisico. Dit risico is ook groter wanneer schisis onderdeel is van een aangeboren
afwijking (Stegenga. 2000).
Schisis wordt meestal niet veroorzaakt door één erfelijke factor, afkomstig van één of
beide ouders maar ontstaat zoals reeds aangehaald door een samenloop van
omstandigheden (Devriendt et al. 2008). Bij multifactoriële erfelijkheid met drempel is het
berekenen van herhalingsrisico’s zeer moeilijk. Hier wordt het risico gekoppeld aan het
aantal verwanten met schisis (Stegenga. 2000). Dit verklaart dan ook dat de kans op een
volgend kindje met schisis eigenlijk relatief klein is. De kans dat dit kindje opnieuw
dezelfde combinatie van meerdere genen overerft en opnieuw bepaalde
omgevingsinvloeden ondergaat is in de grootorde van 3 à 5%. In het verleden zijn er
verschillende studies gebeurd waarbij men de kansen onderzocht op een volgend kind met
schisis. Men kwam tot de volgende vaststellingen: (Devriendt et al. 2008)
1) Het herhalingsrisico op schisis is groter naarmate een familielid meer verwant is.
Familieleden hebben namelijk een hogere graad van gemeenschappelijke genen dan niet
verwante personen (Devriendt et al. 2008).
Wanneer de ouders normaal zijn en één kind met schisis hebben, is de kans op herhaling
slechts enkele procenten. Dergelijke berekeningen vereisen de nodige deskundigheid en
worden door een geneticus uitgevoerd (Stegenga. 2000).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
12
2) Het herhalingsrisico is afhankelijk van het aantal aangedane personen in de familie
(meerdere personen wijst op een grotere vatbaarheid in de familie) (Devriendt et al.
2008). Als twee eerstegraads verwanten van een kind schisis hebben dan verdubbelt het
herhalingsrisico (Stegenga. 2000).
3) Het herhalingsrisico is groter naarmate het defect meer uitgesproken is (Stegenga.
2000)
4) Het herhalingsrisico is afhankelijk van het geslacht van de persoon met de aandoening.
CP komt vaker voor bij meisjes en CL/P zien we meer bij jongens (Stegenga. 2000).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
13
Hoofdstuk 2 : Agenesie
2.1 Defintie
Agenesie of hypodontie is de congenitale afwezigheid van één of meerdere
gebitselementen. Zijn er zes tanden of meer niet aangelegd (met uitzondering van de
wijsheidstanden) dan spreekt men van oligodontie (Adeboye et al. 2006).
Wanneer het gaat om de totale afwezigheid van tanden noemen we dit anodontie. Dit
komt maar zelden voor en is vaak geassocieerd met hypohydrotische ectodermale
dysplasie of een andere X-gebonden recessieve aandoening (Adeboye et al. 2006).
Agenesie kan voorkomen als een geïsoleerde familiale of sporadische vorm of in
associatie met andere afwijkingen (Adeboye et al. 2006). Ongeveer 10% van de gevallen
tonen nog andere tandabnormaliteiten zoals hypoplastisch glazuur, peg-shaped laterale
snijtanden van de bovenkaak, microdontie etc. (Vieira et al. 2004).
2.2 Prevalentie
Hypodontie komt vaker voor in het definitieve gebit dan in het melkgebit. Bij de
melktanden ontbreken het vaakst de snijtanden van de onderkaak. Bij de definitieve
elementen vooral de 3de molaren, 2de premolaren, en laterale snijtanden (afwezigheid van
de 1ste en 2de molaren is zeldzaam) (Adeboye et al. 2006, Stegenga. 2000). Townsend et
al. (2005) haalde aan dat deze tanden het vaakst ontbreken omdat de ontwikkeling over
een langere periode gebeurt, wat een grotere kans kan geven tot agenesie.
De verschillen in prevalentie hangen nauw samen met de diagnostische criteria, de wijze
waarop de steekproef bepaald wordt, de raciale variatie en het populatietype. Uit eerdere
studies is ook gebleken dat dergelijke afwijkingen meer voorkomen bij meisjes dan bij
jongens ( 3 op 2) (Adeboye et al. 2006, Stegenga. 2000).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
14
Bij de prevalentie van hypodontie zien we dat deze groter is binnen familiale kring dan bij
de gewone populatie, wat wijst op een erfelijk verband van hypodontie (de proportie van
verwanten hangt af van de graad van de hypodontie) (Adeboye et al. 2006).
2.3 Normale tandontwikkeling
De eerste tekenen van de tandontwikkeling doen zich voor tussen de zesde en zevende
week van de ontwikkeling na de bevruchting (Gartner. 2001), nadat de neurale lijstcellen
gemigreerd zijn naar de plaats waar het dentofaciale complex zich gaat ontwikkelen
(Carels. 2005-2006). De aanzet tot de tandformatie vormen de lokale
celvermenigvuldigingen ter hoogte van het orale epitheel. Door interacties met het
onderliggende mesenchyme wordt een signaalcentrum gevormd die tot de verschillende
weefselcomponenten leiden en uiteindelijk tot de formatie van zowel de melk– als
definitieve elementen (Carels. 2005-2006). Het resultaat van deze mitotische activiteit is
de vorming van een hoefijzervormige band van epitheel cellen, de dentale lamina
genoemd (Garnter. 2001). Deze lokale veranderingen worden geïnduceerd door cellen van
de neurale lijst die naar het orale mesenchym zijn gemigreerd en die daar de plaats
bepalen waar de gebitselementen zich al dan niet zullen ontwikkelen.
Het onderzoek naar de erfelijke factoren bij muizen heeft aangetoond dat de
tandontwikkeling gereguleerd wordt door interacties tussen epitheliale en mesenchymale
cellen en dat deze afhankelijk zijn van een aantal genen. Deze genen coderen voor
transcriptiefactoren en signaalmoleculen (Adeboye et al. 2006) wat leidt tot de initiatie,
morfogenese en differentiatie van de ontwikkeling van de tanden (Towsend et al. 2005).
Vorming van de kroon
De knopfase
Kort na het verschijnen van de dentale lamina, stijgt de mitotische activiteit in het
onderste deel van de epitheliale band van elke tandboog. Deze activiteit is
verantwoordelijk voor de vorming van 10 epitheliale structuren, de knoppen. Zij leiden tot
de vorming van de melktanden van zowel de boven- als onderkaak.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
15
Verdere ontwikkeling is gelijkaardig voor elke tand, maar deze zijn wel asynchroon
(Gartner. 2001).
De kapfase
Zodra de cellen van de knop gaan prolifereren, veranderen deze niet alleen van grootte
maar ook van vorm. We krijgen zo een drielagige configuratie, de kap.
Er volgt vervolgens een ingewikkeld moleculair biologisch proces waarbij multipele
interacties optreden tussen het epitheel en het mesenchym; de eerste noodzakelijke
cellulaire signalen zijn afkomstig van het epitheel, met name de vrijzetting van Sonic
Hedgehoc (SHH), Bone Morphogenetic Protein 4 (BMP-4), BPM-2, BMP-7 en
Fibroblastic Growth Factor 8 (FGF-8) naar de dentale mesenchymcellen (Carels. 2005-
2006). Om het proces van de tandontwikkeling verder normaal te laten verlopen is een
respons noodzakelijk vanuit het dentale mesenchym, waarbij –onder andere– MSX1 en
PAX9 een rol spelen. De mesenchymcellen vuren dus op hun beurt terug signalen af naar
de epitheelcellen, die dan wederom een ander expressiepatroon gaan vertonen.
Tijdens de kapfase van de tandontwikkeling ontstaat een stevige streng van epitheliale
cellen uit de dentale lamina. Deze cellen gaan prolifereren en groeien in de diepte (Allan.
1997).
De klokfase
Deze laatste fase van de kroonvorming wordt ook wel de fase van de morfo- en
histodifferentiatie genoemd.
Verschillende cellen differentiëren tot ameloblasten of odontoblasten die zorgen voor de
productie van glazuur en dentine (Gartner. 2001).
Wanneer het glazuur en het coronale dentine gevormd is, gaan de tandkiemen naar een
volgende fase van tandvorming, nl de wortelvorming (Gartner. 2001).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
16
2.4 Etiologie van agenesie
Meer dan honderden genen spelen een rol bij de tandontwikkeling en elk van deze zou een
oorzakelijke rol kunnen spelen bij het ontstaan van tandagenesie (Viera. 2003, Nieminen
et al. 1998). Hoewel tandagenesie occasioneel veroorzaakt wordt door omgevingsfactoren
(vb trauma), chemo– of radiotherapie, is de meerderheid echter te wijten aan genetische
factoren (Näsman et al. 1997).
Brook et al. (1984) kwam tot de conclusie dat de prevalentie van hypodontie hoger is in
familiale kring dan in de algemene populatie, wat de hypothese over het bestaan van
erfelijke factoren bij agenesie ondersteunt.
Townsend et al. (2005) heeft aan de hand van stambomen aangetoond dat familiale
tandagenesie autosomaal dominant, recessief of X–gebonden kan zijn.
2.4.1 Erfelijkheid
Onderzoeken zijn gedaan naar het verband tussen hypodontie en de verschillende
kandidaat genen die betrokken zijn bij de odontogenese (Townsend et al. 2005). Hoewel
nog niet alle moleculen gekend zijn, werden de laatste jaren vooral mutaties van de genen
PAX9, MSX1 en AXIN2 geassocieerd met tandagenesie (Viera. 2003). Het zijn
transcriptiefactoren die actief zijn in de vroeg epitheliaal–mesenchymale interacties
(Carels. 2005-2006).
PAX 9
Dit gen is actief tijdens de embryogenese. Het speelt onder andere een belangrijke rol bij
de epitheliale en mesenchymale celcascades tijdens de odontogenese (Swinnen et al. 2008,
Peters et al. 1999). Mutaties in PAX9 kunnen als gevolg hebben dat er een fout optreedt in
de tandontwikkeling (Mensah et al. 2003).
Tot nu zijn er reeds vijftien heterozygote mutaties van PAX9 (Nieminen et al. 2001,
Frazier-Bowers et al. 2002, Das et al. 2003, Mensah et al. 2003, Stockton et al. 2000)
geïdentificeerd die geassocieerd kunnen worden met een familiale niet–syndromale vorm
van tandagenesie (Swinnen et al. 2008).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
17
Mostowska et al. (2003) toonde aan dat mutaties in de coderende regio’s van PAX9 vaak
geassocieerd worden met een uniek fenotype van familiale tandagenesie, het ontbreken
van vnl de molaren. Dit werd o.a. bevestigd door Rice. (2004) en Mensah et al. (2003).
Ook de studie van Frazier-Bowers et al. (2002) toonde bij een familie, met een erfelijke
vorm van hypodontie, een associatie tussen een mutatie van PAX9 en agenesie van de
molaren.
Hoewel door Vieira et al. (2004) de link tussen PAX9 en agenesie aangetoond was, was
het onmogelijk om met volledige zekerheid de associatie aan te tonen met de afwezige
molaren.
Een heterozygote deletie van PAX9 wordt ook al eens aangetroffen bij ernstige
hypodontie, meestal betreft het dan afwezigheid van de melkmolaren en definitieve
molaren (Adeboye et al. 2006, Vieira et al. 2004). Dit werd ook aangetoond bij muizen
door Peters et al (1998/1999). Een heterozygote deletie had hier hypodontie tot gevolg
zonder dat er zich andere afwijkingen vertoonden. Een homozygote deletie van PAX9
daarentegen leidde tot het ontbreken van alle tanden en tekorten in de craniofaciale
ontwikkeling waaronder schisis van het palatum en ademhalingsproblemen (Peters et al.
1998/1999).
MSX1
Dit gen is net zoals PAX9 een transcriptiefactor en komt tot expressie in verschillende
embryologische processen, waaronder de odontogenese (Viera. 2003). Mutaties van
MSX1 kunnen bijgevolg leiden tot fouten in de tandontwikkeling (Viera. 2003).
Tot nu zijn er reeds 7 heterozygote mutaties van dit gen gevonden die geassocieerd
kunnen worden met tandagenesie (Vastardis et al. 1996, De Muynck et al. 2004). Satokata
en Maas (1994) stelden bij een studie op muizen een mutatie vast die leidde tot schisis,
tandagenesie en deficiëntie in de ontwikkeling van het kaakbot. Ook Van den Boogaard et
al. (2000) vond een mutatie die een link vormt tussen schisis en agenesie.
Vastardis et al. (1996/2000) vond dan weer een mutatie die verantwoordelijk was voor een
autosomaal dominante vorm van agenesie van de 2de premolaren en 3de molaren in een
familie. Lidral en Reising (2002) kwam tot dezelfde conclusie. Ook de resultaten van de

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
18
studie van Frazier-Bowers et al. (2001) toonde een associatie tussen een mutatie van
MSX1 met agenesie van de premolaren.
Wat opvalt bij deze laatste studies is dat mutaties van het MSX1 gen vaak geassocieerd
worden met agenesie van de 2de premolaren en 3de molaren (Viera. 2003).
Vieira et al. (2004) kwam tot dezelfde vaststelling. Maar omdat de populatie aantallen van
deze studie niet groot genoeg waren kon hij deze conclusie niet met zekerheid bevestigen.
Muizen met een deletie van MSX1 vertoonden craniofaciale defecten, defecten van het
kaakbot en verstoringen van de tandontwikkeling (Peters et al. 1999, Satokata en Maas
1994).
Zowel Adeboye et al. (2006) als Mensah et al. (2003) hebben cases gevonden waar
hypodontie aanwezig was in afwezigheid van mutaties in zowel het PAX9 gen als in
MSX1. Dit wijst er op dat ook andere genen verantwoordelijk moeten zijn voor de
vorming van de tanden.
AXIN2
Zo werd de laatste jaren AXIN2 geassocieerd met agenesie (Vieira. 2003). Dit gen
behoort tot de regulatoren van het Wnt-signalisering pad, het is een grote familie van
signaalmoleculen die een rol hebben tijdens de embryologische ontwikkeling en expressie
van de tanden (Cobourne. 2007). Om de exacte functie te kennen van AXIN2 in de
odontogenese, is er in de toekomst gedetailleerd onderzoek van elke fase van de
tandontwikkeling noodzakelijk (Mostowska et al. 2006).
Verschillende mutaties van dit gen werden door stamboomonderzoek reeds geassocieerd
met hypodontie (Stockton et al. 2000, Mostowska et al. 2006, Cobourne. 2007). In
tegenstelling tot PAX9 en MSX1 heeft men mutaties van AXIN2 tot nu toe nog niet
kunnen linken met de afwezigheid van een bepaald type tand.
Verder zijn er reeds ook verschillende polymorfismen geïdentificeerd die een verhoogd
risico vormen op tandagenesie (Cobourne. 2007).
Ook werden mutaties van FGF8 en BMP4 gezien als mogelijke veroorzakers (Adeboye et
al. 2006, Mensah et al. 2003). Vieira et al. (2004) zag bij zijn studie een relatie tussen
afwijkingen van het TGFA gen en agenesie. Thomas et al. (1997) vond een mutatie van
DLX1 en DLX2 die geassocieerd kon worden met agenesie van de molaren in de

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
19
bovenkaak. Ferguson et al. (1998) vond bij muizen een mutatie in TGF waarvan de
molaren van de bovenkaak ontbraken.
Ook al zijn deze genen geïdentificeerd met een mogelijke rol in de etiologie van
hypodontie, toch is het noodzakelijk dat we de relatie aantonen tussen de genetische
afwijking en zijn fenotype. Epigenetica, wat zorgt voor een verandering van de
genexpressie zonder dat er veranderingen zijn in de nucleotide sequentie, is een
belangrijke term waar nog maar weinig van geweten is. Vergelijking van monozygote
tweelingen die dezelfde genen delen en een verschillende fenotype expressie vertonen
kunnen een verklaring zijn van de epigenetische invloeden op het fenotype (Townsend et
al. 2005).
2.4.2 Exogene factoren
Omgevingsfactoren kunnen occasioneel een rol spelen in de etiologie van tandagenesie.
Mogelijke factoren zijn chemo- of radiotherapie, infectieziekten op jonge leeftijd,
endocriene stoornissen, trauma etc. Toch zijn deze van minder belang en is de
meerderheid van agenesie te wijten aan genetische factoren (Näsman et al. 1997).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
20
Hoofdstuk 3 : Onderzoek naar de erfelijkheid van schisis en agenesie
3.1 Tweelingenonderzoek
3.1.1 Waarom tweelingenonderzoek?
Tweelingen vormen voor wetenschappers een goede studiepopulatie (De Fruyt et al.
2000). Zij worden frequent gebruikt bij medisch onderzoek en zijn vooral van belang bij
het bepalen van de relatieve bijdrage van genetisch versus niet genetische componenten in
de etiologie van bepaalde afwijkingen (Mansilla et al. 2005, Eufinger et al. 1993,
Christensen et al. 1993, Van Esch et al. 2008).
Een dergelijke studie met tweelingen brengt geen duidelijk wijze van overerving aan het
licht, maar wanneer informatie over het herhalingsrisico gecombineerd wordt met goede
informatie over de tweeling concordantie kan dat leiden tot een vaststelling van het aantal
en grootte van het effect van bepaalde genen (Christensen et al. 1993).
3.1.2 Zygositeit van de tweeling
Dizygote tweelingen ( DZ)
Twee-eiige of dizygote tweelingen ontstaan uit de bevruchting van twee verschillende
eicellen met twee verschillende zaadcellen. Deze kinderen delen in termen van erfelijk
materiaal evenveel als hun biolgische broers en zussen en zijn dus erfelijk verschillend
(De Fruyt et al. 2000).
Monozygote Tweelingen (MZ)
Eeneiige of monozygote tweelingen ontstaan door de bevruchting van één eicel met één
zaadcel, waarna splitsing optreedt. Hierbij zijn de twee helften genetisch identiek.
Monozygote tweelingen zijn dus per definitie van hetzelfde geslacht, terwijl dizygote
tweelingen van hetzelfde of van verschillend geslacht kunnen zijn (De Fruyt et al. 2000).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
21
Bepalen van de zygositeit
Een exacte bepaling van de zygositeit van de tweelingen is de basis voor een goed
onderzoek. Aangezien er verschillende methodes bestaan om de zygositeit van tweelingen
te achterhalen kan dit leiden tot verschillende waarnemingen tussen de onderzoeken
(Eufinger et al. 1993). Methodes om de zygositeit van een tweelingenpaar te achterhalen
zijn o.a.: algemeen voorkomen, genetische merkers zoals DNA, vingerafdrukken,
bloedgroep, serumproteïnen en onderzoek van placenta bij de geboorte (Eufinger et al.
1993, De Fruyt et al. 2000). Correcte bepaling van de zygositeit kan niet altijd op grond
van uiterlijke overeenkomsten of verschillen. Meestal is het zo dat eeneiige tweelingen op
elkaar gelijken en dat twee-eiige tweelingen uiterlijk verschillend zijn, zoals broers en
zussen, maar dit is niet altijd het geval (De Fruyt et al. 2000).
Concordantie
Van tweelingen wordt gezegd dat ze concordant zijn als ze beide de genetische
karaktertrek hebben en discordant als één van beide de karaktertrek heeft en de andere
niet. In vele studies over genetische karaktertrekken wordt een grotere concordantie tussen
MZ tweelingen dan DZ tweelingen getoond. Sun-wei et al. (2001) gaf aan wat door vele
anderen bevestigd wordt, dat een grotere concordantie, dwz een groter fenotype
gelijkheid, bij MZ dan DZ tweelingen vermoedelijk te wijten is aan de grotere genetische
gelijkheid. Waar vroeger dus aangenomen werd dat elk fenotype verschil tussen MZ
tweelingen een invloed van de omgeving was, heeft recente onderzoekswerk deze
veronderstelling in vraag gesteld. Van de frequentie en oorsprong van elke genetische
discordantie tussen MZ tweelingen blijven echter nog veel vragen onbeantwoord
(Eufinger et al. 1993, Kato et al. 2005, Singh et al. 2002).
MZ tweelingen kunnen discordant zijn voor geïsoleerde afwijkingen, chromosomale
afwijkingen of single gene disorders. Een discordantie voor een geïsoleerde afwijking kan
het gevolg zijn van postzygotische splitsing mutaties, verschillende
omgevingsblootstelling in de baarmoeder, epigentische variatie in de imprinting, X-
chromosoom inactivatie etc. (Eufinger et al. 1993).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
22
3.1.3 Soorten tweelingenonderzoek
Sinds MZ tweelingen een belangrijke waarde voorzien in de studie naar de rol van
genetische factoren, is de methode van tweelingenstudie continu geëvolueerd. Gaande van
klassieke tweelingenstudies tot recent tweelingenonderzoek (Kato et al. 2005).
Klassiek tweelingenonderzoek
Dit is gebaseerd op het onderscheid tussen de genetische- en de omgevingsfactoren door
de vergelijking van de concordantie tussen MZ en DZ tweelingen. Deze tweelingen zijn
samen opgegroeid en hebben dezelfde omgevingsfactoren gedeeld. Het uitgangspunt bij
deze studies is de veronderstelling dat MZ tweelingen genetisch identiek zijn terwijl DZ
tweelingen gemiddeld slechts 50% van hun genen delen (Christensen et al. 1993, De Fruyt
et al. 2000, Townsend et al. 2005, Kato et al. 2005).
Gelijkaardigheid bij MZ en DZ tweelingen kunnen we vaststellen door het bepalen van de
concordantie (De Fruyt et al. 2000). Fenotypes die een grotere graad van gelijkheid
hebben bij MZ dan DZ tweelingen worden verondersteld een genetisch etiologisch
gelijkaardige component te hebben (Kato et al. 2005).
Recent tweelingenonderzoek
Dit onderzoek wil de genetische en/of epigenetische verschillen tussen discordante MZ
identificeren (Kato et al. 2005).
Epigenetica is het verschil in genexpressie zonder veranderingen in de DNA sequentie
(Townsend et al. 2005). Er is een studie met bewijs dat gekloonde dieren fenotypisch
kunnen verschillen ondanks een zelfde genoom. Deze verschillen in fenotype zouden het
resultaat zijn van epigenetische verschillen. Gelijkaardig kunnen we veronderstellen dat
discordantie tussen MZ tweelingen kan veroorzaakt zijn door epigenetische verschillen
(Kato et al. 2005, Petronis et al. 2003). Omgevingsfactoren kunnen de discordantie tussen
MZ tweelingen beïnvloeden, aangezien deze een invloed kunnen hebben op de
epigenetica (Kato et al. 2005).
Epigenetische regulatie van een genactiviteit is onderworpen aan het
ontwikkelingsstadium, is weefselspecifiek, leeftijdsafhankelijk en afhankelijk van
omgevingsinvloeden (Petronis et al. 2003).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
23
Voorlopige classificatie van de mogelijke genetische- en epigenetische factoren die
kunnen leiden tot verschillen tussen MZ:
Genetisch
1) Puntmutatie Een puntmutatie kan leiden tot een verschil tussen MZ. Dit soort van genetisch verschil
kan tijdens of na de splitsing ontstaan (Kato et al. 2005).
2) Chromosomale abnormaliteit Een verschillende afwijking van chromosomen tussen MZ tweelingen kan leiden tot
discordantie tussen beide (Kato et al. 2005).
3) Mitochondriaal DNA (mtDNA) heteroplasmy
Hierbij bestaat het gemuteerde mtDNA meestal samen met een wild type mtDNA (Kato et
al. 2005). Klinische fenotypes kunnen wijzigen met de verhouding en de weefselverdeling
van de mutatie (Singh et al. 2002).
4) Triplet repeats
De lengte van de triplet repeat kan verschillen tussen MZ als gevolg van een expansie
(van generatie op generatie). Deze expansie kan veroorzaakt worden door het uitzetten
tijdens de meiose of door het uitbreiden van de repeats tijdens de mitose (Kato et al.
2005).
Epigenetisch
1) X chromosoon inactivatie
Discordantie tussen MZ tweelingen voor X-gebonden genen kan het gevolg zijn van 2
fenomen. Ten eerste is er de willekeurige inactivatie en ten tweede kan methylatie leiden
tot een toevoegende factor van discordantie tussen beide.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
24
Aangezien mannen maar één X chromosoom en de vrouwen twee X chromosomen
hebben, wordt er bij de vrouwen één geïnactiveerd om de ongebalanceerde expressie
tussen beide te vermijden. Bij de vrouwen wordt de vaderlijke X geïnactiveerd tijdens de
embryogenese, nadien wordt deze gereactiveerd en wordt er achteraf willekeurig één van
beide X-en geïnactiveerd. Aangezien deze inactivatie willekeurig verloopt kan dit een
heterozygositeit veroorzaken in de weefsels. Ten gevolge van deze chromosoom
inactivatie kunnen X-gebonden ziektes soms discordant zijn bij vrouwelijke MZ
tweelingen. Vb mentale retardatie, rood-groen kleurenblindheid etc. (Mansilla et al. 2005,
Kato et al. 2005).
2) Verlies van imprinting
Genomische imprinting is een fenomeen waarbij één van de twee allelen, van moederlijke
of vaderlijke origine, geïnactiveerd wordt door DNA methylatie. Het gemethyleerd en
geïnactiveerd allel wordt vermeld als “imprinted”. Veel van de imprinted genen zijn
gerelateerd aan celgroei en verlies van imprinting is ook een oorzaak van kanker. Een
ziekte waarbij verlies van imprinting vaak als oorzaak gezien wordt is het Beckwith–
Wiedemann syndroom, dit kan ook bij vrouwelijke MZ tweelingen en dan meestal in
discordantie (Mansilla et al. 2005, Kato et al. 2005).
Om te achterhalen of het gaat om een genetisch of epigenetisch verschil tussen
discordante tweelingen, is het belangrijk om tweelingen te vinden die kunnen dienen voor
de studie. Zij moeten aan een aantal voorwaardes voldoen :
Ten eerste, als één van beide de ziekte een paar jaar eerder heeft dan de ander dan zijn ze
niet volledig discordant voor deze ziekte.
Ten tweede, cases waar er sprake is van omgevingsinvloeden zoals infecties, perinatale
complicaties etc. zijn ook niet bruikbaar voor een genetisch/epigenetisch onderzoek (Kato
et al. 2005).
De ideale bron voor epigenetische analyse is hersenweefsel, maar dit is praktisch niet
haalbaar in de studie van discordante tweelingen (Kato et al. 2005, Petronis et al. 2003).
Perifere bloedcellen (Kato et al. 2005) of de met teelt voortgebrachte dieren (Petronis et
al. 2003) worden als alternatief gebruikt.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
25
3.2 Resultaten van tweelingenonderzoek in verband met schisis
Christensen et al. (1993) had als doel de relatieve bijdrage van de genen en omgeving bij
het ontstaan van CL/P bij tweelingen en het effect van een enkele locus op het risico van
CL/P te achterhalen. De zygositeitsbepaling van de tweelingen gebeurde aan de hand van
bloed, serum en enzymes, waarbij 60% concordant was voor schisis bij MZ tweelingen en
10% bij de DZ. In deze studie was er geen verschil tussen CL/P bij tweelingen en gewone
individuen wat betreft frequentie, geslachtsverdeling en type van spleet. Deze vaststelling
wijst erop dat er geen verschil is in de pathogenese tussen beide. Het significante verschil
in de concordantie tussen de MZ en DZ indiceert een grote genetische invloed maar het
feit dat er MZ paren discordant waren voor CL/P, ondanks de concordante
zygositeitsbepaling, wijst er op dat nog andere factoren belangrijk zijn in de oorzaak van
CL/P waaronder epigenetisch en omgevingsinvloeden. Toch vermoedt men dat de
omgevingsinvloeden van minder belang zijn dan de genetische factoren.
Het onderzoek van Nordström et al. (1996) is vergelijkbaar met de vorige studie
aangezien het hier ook gaat om data van tweelingen van een specifieke populatie
gedurende een bepaalde periode (40 jaar). Hier heeft men de zygositeit bepaald aan de
hand van het geslacht, de fysische overeenkomsten en door bloedgroep, serumproteïnen,
erythrocyten of isoenzyme systemen. De zygositeit MZ/DZ komt overeen met de studie
van Christensen et al. (1993). Nordström et al. (1996) heeft ook de CP/CL(P) ratio
vergeleken met andere studies. Zij kwamen tot de vaststelling dat er verschillen zijn
tussen de populaties, wat wijst op een geografische invloed op het soort van cleft.
Het doel van Mansilla et al. (2005) was genen te identificeren die een rol zouden kunnen
spelen in de etiologie van orofaciale spleten. Zygositeitsbepaling gebeurde hier door de
analyse van 15 markers. De DNA sequentie van elk tweelingenpaar werd bekomen en
men deed een analyse van de genen die gekend waren als mogelijke oorzakers voor
schisis.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
26
Deze tabel geeft het aantal MZ en DZ
tweelingen van deze studie weer. Bij
een deel van de MZ tweelingen zien we
dat er discordantie aanwezig was voor
schisis.
Tabel 2. Tweelingen gebruikt voor sequentie in deze studie (Mansilla et al. 2005)
Aangezien DNA sequentie geen significante verschillen toonde bij discordante MZ
tweelingen zijn er, zoals eerder aangehaald, andere factoren die een rol spelen bij de
etiologie van schisis. Voorbeelden hiervan zijn postzygotische splitsing mutaties,
omgevingsinvloeden,X-chromosoominactivatie….
3.3 Resultaten van tweelingenonderzoek in verband met agenesie
Kindelan et al. (1998) stelde zich tot doel aan te tonen dat genetische codering niet de
enige oorzaak is voor agenesie. Door middel van DNA en vinderafdrukken stelde men
vast dat de tweeling van deze case monozygoot was, toch toonden zij een verschil in
agenesie. Persoon 1 miste buiten de overeenkomende elementen ook element 25 en had
een grotere overjet dan persoon 2 in afwezigheid van een uitlokkende factor.
Verondersteld werd dat epigenetische of omgevingsinvloeden hier een rol zouden spelen.
Adeboye et al. (2006) besprak een case van ernstige hypodontie bij een drieling. Deze
toonde een grote graad van concordantie in het patroon en aantal tanden van agenesie,
hetgeen wijst op een sterke genetische link. Verder was er ook een positieve geschiedenis
van agenesie aan de kant van de vader. Een hiaat aan deze case was het tekort aan
informatie voor wat betreft de zygositeit van de drieling.
Wat opviel was dat er bij één van de drie jongens vijf tanden meer ontbraken, daar kunnen
we als mogelijk oorzaak een genmutatie of een verschil in genexpressie uit
veronderstellen.
Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
27
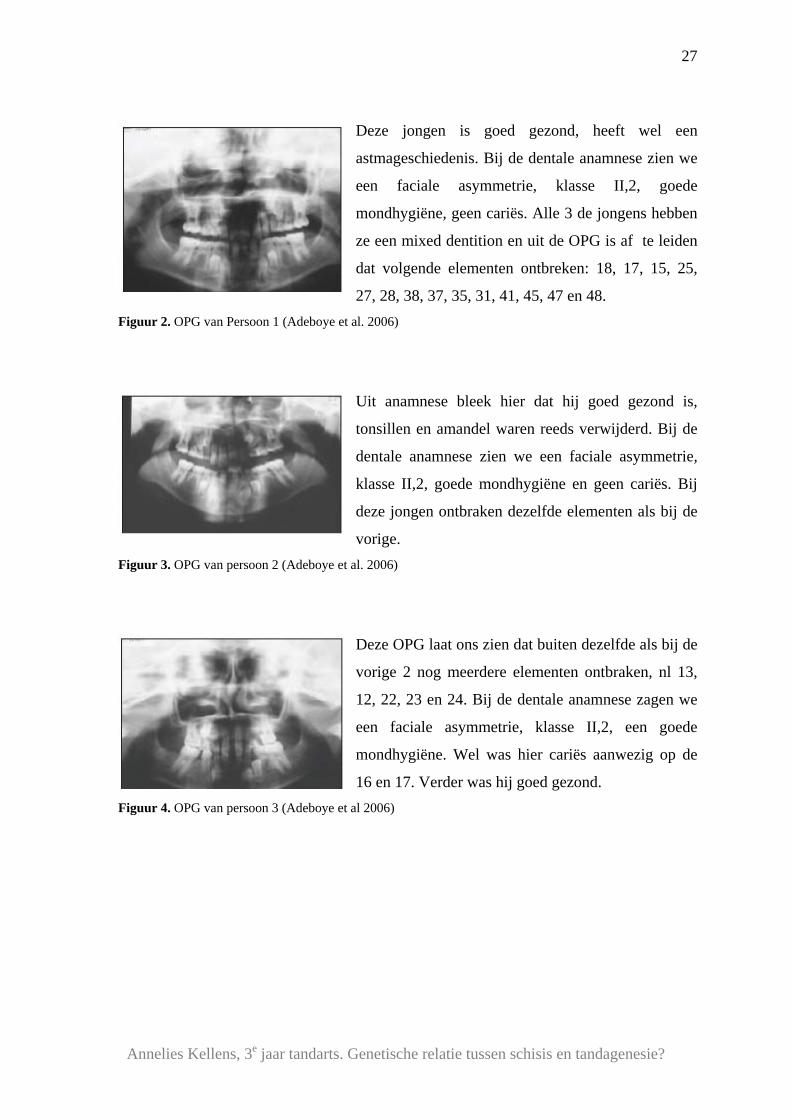
Deze jongen is goed gezond, heeft wel een
astmageschiedenis. Bij de dentale anamnese zien we
een faciale asymmetrie, klasse II,2, goede
mondhygiëne, geen cariës. Alle 3 de jongens hebben
ze een mixed dentition en uit de OPG is af te leiden
dat volgende elementen ontbreken: 18, 17, 15, 25,
27, 28, 38, 37, 35, 31, 41, 45, 47 en 48. Figuur 2. OPG van Persoon 1 (Adeboye et al. 2006)
Uit anamnese bleek hier dat hij goed gezond is,
tonsillen en amandel waren reeds verwijderd. Bij de
dentale anamnese zien we een faciale asymmetrie,
klasse II,2, goede mondhygiëne en geen cariës. Bij
deze jongen ontbraken dezelfde elementen als bij de
vorige. Figuur 3. OPG van persoon 2 (Adeboye et al. 2006)
Deze OPG laat ons zien dat buiten dezelfde als bij de
vorige 2 nog meerdere elementen ontbraken, nl 13,
12, 22, 23 en 24. Bij de dentale anamnese zagen we
een faciale asymmetrie, klasse II,2, een goede
mondhygiëne. Wel was hier cariës aanwezig op de
16 en 17. Verder was hij goed gezond. Figuur 4. OPG van persoon 3 (Adeboye et al 2006)

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
28
Townsend et al. (2005) liet de term epigenetica vallen bij de etiologie van agenesie. In
deze studie werd de zygositeit van de tweelingen bevestigd door verschillende
onderzoeken.
Deze tabel laat zien dat 24 van de
278 tweelingparen gescreend voor
dit onderzoek discordant waren voor
agenesie, wat neer komt op 8.6%.
Deze hoge prevalentie van
discordantie was in strijd met
sommige andere studies, waaronder
Markovic. (1982) waar een grote
concordantie voor agenesie
gevonden werd bij de meeste
tweelingen. We zien hier dat vooral
de laterale snijtanden boven en
premolaren ontbreken.
Tabel 3. Tweelingen met agenesie van de laterale snijtand bovenkaak en premolaren (Townsend et al. 2005)
De vraag blijft dan wat als oorzaak kan aangehaald worden voor deze verschillen?
Townsend et al. (2005) heeft uit dit onderzoek besloten dat er een genetische bijdrage ligt
in de oorzaak van de variatie van de menselijke tandmorfologie. MZ tweelingparen
kunnen kleine verschillen tonen in de tandmorfologie omwille van epigenetische
invloeden.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
29
Hoofdstuk 4 : Zijn schisis en agenesie genetisch gerelateerd?
In dit laatste hoofdstuk ga ik op zoek naar een genetische link tussen beide orofaciale
defecten. Het bewijs voor een gelijke genetische etiologie, voor fenotypes die gelijktijdig
zowel schisis als agenesie hebben, komt van een aantal verschillende bronnen. Enerzijds
zijn er de muizen studies naar geïsoleerde vormen van schisis in combinatie met
tandagenesie (Satokata en Maas 1994, Peters et al. 1998), anderzijds is er
familieonderzoek dat een link aantoont tussen schisis en agenesie (Eerens et al. 2001).
Ook zijn er aantal single–gene-disorders en chromosomale afwijkingen waarbij bij de
fenotypes zowel schisis als agenesie aanwezig is (Ranta. 1983).
4.1 Studies naar een genetische link tussen niet-syndromale schisis en
agenesie
4.1.1 Studies met muizen
Het MSX1 gen is reeds geassocieerd met het voorkomen van orofaciale spleten en
erfelijke tandagenesie (Vastardis et al. 1996, Van den Boogaard et al. 2000). Het heeft een
belangrijke rol in de epitheel-mesenchymale reacties van de ontwikkeling van het
craniofaciale bot en de odontogense (Satokata en Maas 1994, Modesto et al. 2006).
Muizen met een homozygote deletie van dit gen toonden vaak verschillende craniofaciale
defecten zoals spleten van het palatum, deficiëntie van het alveolaire bot van boven- en
onderkaak en tandagenesie (Satokata en Maas 1994, Modesto et al. 2006). Deze
observatie stelt dus vast dat bepaalde mutaties van MSX1 een oorzaak zouden zijn voor
zowel schisis als selectieve tandagenesie. In studies van Maestri et al. (1997) en Lidral et
al. (1998) werd deze associatie bevestigd .
Peters et al. (1998) stelde vast dat muizen met een PAX9 deficiëntie verstoringen
vertoonde van de craniofaciale ontwikkeling. Kist et al. (2007) bevestigde dit. Hoewel de
muizen met een homozygote deletie van het PAX9 gen kort sterven na de geboorte heeft
men toch kunnen aantonen dat er in afwezigheid van dit gen verschillende craniofaciale

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
30
defecten aanwezig zijn, waaronder spleten van het palatum afwezigheid van de tanden
(Peters et al. 1998, Kist et al. 2007)
4.1.2 Familieonderzoek
De volgende figuur toont ons een Nederlandse familie waarvan twaalf familieleden
verschillende combinaties van CL, CP en tandagenesie vertoonden.
Figuur 5. Stamboom en symptomen van een Nederlandse familie met schisis en tandagenesie (Van den
Boogaard et al. 2000)
Zoals af te leiden uit bovenstaande figuur zien we dat vier familieleden schisis hebben
(CL of CLP) al dan niet in combinatie met agenesie. Bij elf van de twaalf ontbraken een
aantal definitieve elementen. Bij de meeste van hen ontbraken de 2e premolaren, zowel in
de boven– als onderkaak, ook was er vaak afwezigheid van de wijsheidstand. In het
algemeen waren de tandagenesiën bilateraal symmetrisch. Een deel van de familie werd
gescreend op MSX1 en bij de twaalf affected familieleden vond men dat zij heterozygoot
waren voor een mutatie van MSX1, de welke men niet vond bij de unaffected gescreende
familieleden. De verschillen in schisis bij deze vier familieleden, hoewel heterozygoot
voor de mutatie van MSX1 wijst op de variabiliteit van het tot expressie komen van
schisis. Verschillen in genetische achtergrond of omgeving kunnen verantwoordelijk zijn
voor de schisis verschillen.
We concluderen hieruit dat het mutant type van MSX1 in deze familie de oorzaak is van
de schisis en de tandagenesie, resultaten die eerder ook werden aangetoond bij studies met
muizen. Toch erop wijzen dat er andere mutaties van MSX1 zijn die tot nu toe alleen maar
gelinkt werden een ofwel tandagenesie ofwel schisis (Van den Boogaard et al. 2000).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
31
Uit een steekproef van Eerens et al. (2001) bleek dat sommige genetische factoren voor
schisis gerelateerd zijn aan agenesie. Door de dentale ontwikkeling te vergelijken bij een
groep met schisis, een groep familieleden en een controlegroep kwam men tot de
volgende conclusie: de groep met schisis en de verwantengroep hadden een hogere
frequentie van agenesie dan de controlegroep. De schisisgroep toonde significant meer
hypodontie dan de verwantengroep. Hieruit kan men concluderen dat agenesie van de
definitieve tanden buiten de spleetregio vaker voorkomt bij individuen met schisis in
vergelijking met de populatie zonder schisis. De tanden die het vaakst ontbreken zijn de
2de premolaren in onder- en bovenkaak en de laterale snijtanden in de bovenkaak in de
niet spleet regio (Eerens et al. 2001, Ranta. 1986). Eenzelfde etiologische factor of
factoren zouden dus aan de basis van schisis en agenesie kunnen liggen (Eerens et al.
2001). Er wordt verondersteld dat buiten de genetische basis van agenesie, ook een aantal
prenatale factoren die betrokken zijn bij schisis, kunnen resulteren in een stijgende
incidentie van agenesie bij kinderen met CL/P of CP.
Daarenboven was er een significante correlatie tussen de grootte van de spleet en
agenesie. Bij spleten die een derde of meer van het harde palatum overspande, was
agenesie in 45% van de gevallen aanwezig, terwijl bij spleten die minder dan een derde
van het harde palatum innamen maar agenesie aanwezig was in 18% van de gevallen (Van
Esch et al. 2008, Viera. 2003).
Slayton et al. (2003) wou met de volgende studie aantonen dat de genen die reeds
geïdentificeerd waren als kandidaat genen voor schisis in eerdere studies, ook
geassocieerd konden worden met hypodontie buiten de cleft regio. Slayton et al (2003)
baseerde zich op de populatie gebaseerde case control studie van de Iowa Craniofacial
Anomalies Clinic. In deze studie was er een schisisgroep met agenesie buiten de
spleetregio, een groep met schisis zonder agenesie buiten deze regio en een controlegroep.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
32
Uit deze tabel kunnen we afleiden
dat de frequentie van hypodontie
buiten de spleet regio het grootst is
bij CL/P.
Tabel 4. Frequentie van type spleet en voorkomen van hypodontie buiten deze regio (Slayton et al. 2003)
De frequentie van hypodontie buiten de cleft regio in deze studie was 30%, wat
vergelijkbaar is met resultaten uit eerdere onderzoeken o. a. Eerens et al. 2001.
Met de achterliggende gedachte dat de frequentie van hypodontie verschilt naar gelang het
type schisis, hebben Stayton et al. (2003) de allel frequenties van verschillende kandidaat
genen berekend voor elke type van spleet. Hoewel er geen significante associaties
gevonden waren voor schisis met agenesie en met de controlegroep, was er toch een
duidelijke associatie van de MSX1 1.3 merker en TGFB3 CA merker voor schisis met
agenesie. Ondanks het feit dat eerdere studies deze veronderstelling steunen is verder
onderzoek in de toekomst naar mutaties van de oorzakelijke genen noodzakelijk (Slayton
et al. 2003).
Modesto et al. (2006) gelooft dat het voorkomen van CL/P en tandagenesie bij sommige
individuen veroorzaakt wordt door een zelfde genetische mutatie. Zoals vele andere reeds
aanhaalde dat mutaties van MSX1 een oorzakelijk factor kunnen zijn voor schisis en
agenesie, ging ook Modesto et al. (2006) van deze veronderstelling uit.
Bij dit onderzoek observeerde met 52 niet verwante personen. 33 van hen hadden
geïsoleerde schisis en 19 hadden een vorm van schisis samen met agenesie. Verder was er
een controlegroep van een op populatie gebaseerde case-control studie van de universiteit
van Iowa Craniofacial Anomalies. Bij de meeste individuen ontbrak één tand, met een
maximum van acht tanden. Er werden geen nieuwe mutaties of varianten van MSX1
gevonden. Toch werden reeds gekende varianten van MSX1 gezien bij de onderzochte
personen. Eén bepaalde variant zag men vaker bij schisis en een andere meer bij schisis en
agenesie. Het blijkt dat deze varianten markers zijn die genetisch bijdragen aan deze
fenotypes, waaruit dus blijkt dat er een associatie is tussen MSX1 met zowel schisis als
schisis en agenesie.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
33
Gen-gen interacties kunnen een mechanisme zijn voor het ontwikkelen van CL/P met
agenesie. Er is genetische bewijs dat MSX1 interageert met PAX9 bij geïsoleerde
tandagenesie (Viera et al. 2004) en dat PAX9 de genexpressie van MSX1 reguleert bij
muizen (Peeters et al. 1998). Verder onderzoek naar de MSX1 varianten van deze studies
met PAX9 zijn nodig in de toekomst om de hypothese te testen dat deze 2 genen en rol
spelen bij schisis met agenesie (Modesto et al. 2006).
4.2 Chromosomale afwijkingen en single gene disorders
Buiten het voorkomen van geïsoleerde schisis met tandagenesie zijn er een aantal
syndromen waarbij zowel schisis als agenesie aanwezig zijn. Hier ligt één oorzakelijke
factor aan de basis ofwel een chromosomale afwijking ofwel een single-gene-disorder.
Een bekend syndroom waar zowel schisis als agenesie aanwezig is en het niet altijd
makkelijk is om een onderscheid te maken met de geïsoleerde vorm van beide is het Van
der Woude syndroom. Verder zijn er een vormen van het syndroom van Down, Wolf-
Hirschorn syndroom en ectodermale dysplasie beschreven waarbij orofaciale spleten en
fouten in de tandontwikkeling aanwezig zijn.
4.2.1 Van der Woude syndroom
Het Van der Woude syndroom (VWS) is een autosomaal dominant syndroom dat
evenveel voorkomt bij mannen als bij vrouwen. Het is een voorbeeld van een single–gen–
disorder dat CL/P of CP veroorzaakt en sterk lijkt op de niet–syndromale vorm van
schisis.
Dit syndroom wordt gekenmerkt door verschillende anomaliën, waaronder de orofaciale
afwijkingen het belangrijkste zijn. Het meest voorkomend is een combinatie van schisis
en agenesie. De orofaciale spleten kunnen uni- of bilateraal zijn en kunnen in graad van
ernstigheid verschillen. Agenesie doet zich voor in 10 tot 81% van de gevallen en vooral
van de 2de premolaren in onder- en bovenkaak.
Andere intra– en extra- orale symptomen zoals ankyloglossus, congenitale hartdefecten,
ledemaatafwijkinge etc. doen zich maar zelden voor.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
34
De oorzakelijke mutatie is geïdentificeerd in de interferon regulatory factor 6 ( IRF 6)
maar het exacte mechanisme is nog niet gekend. Ongeveer 1 à 2% van de mensen met een
schisis hebben het VWS (Conners. 2007).

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
35
Algemeen Besluit
Uit deze literatuurstudie blijkt dat men voor de meeste gevallen van niet-syndromale
schisis en –tandagenesie een multifactoriële etiologie veronderstelt. Multifactoriële
overerving betekent dat meerdere genetische factoren alsook omgevingsfactoren
meespelen bij het tot stand komen van de afwijking.
Wat betreft de genetische invloeden in de etiologie van niet-syndromale schisis heeft men
polymorfismen van een aantal genen gevonden. Deze kunnen geassocieerd worden met
een groter risico op het voorkomen van schisis. Voor de syndromale vorm zijn enkele
genen aangetoond die in geval van mutatie schisis veroorzaken.
Wat betreft niet-syndromale hypodontie zijn er drie genen aangetoond, namelijk MSX1,
PAX9 en AXIN2 die een oorzakelijke rol hebben in de etiologie. Toch zijn er een aantal
gevallen waarin agenesie niet veroorzaakt wordt door een mutatie van één van deze drie.
Aangezien een grote hoeveelheid genen een rol spelen in de tandformatie, kunnen zij
allemaal een invloed hebben op het ontstaan deze afwijking. De meest voorkomende
syndromale vorm van tandagenesie is deze die geassocieerd wordt met ectodermale
dysplasie.
Door tweelingenonderzoek is het mogelijk de oorzaken van fenotypische variatie te
achterhalen. Zowel voor schisis als voor agenesie vond men dat een groot aantal van de
MZ tweelingen, die verondersteld worden genetisch identiek te zijn, discordant waren
voor deze kenmerken. Hierbij moet evenwel het tijdstip van de splitsing in acht genomen
worden: hoe later de splitsing, hoe geringer de kans op genetische verschillen. Of de
discordantie te wijten is aan de omgevingsinvloeden of aan epigenetische factoren moet
nog verder onderzocht worden. De meeste auteurs blijken ervan uit te gaan dat de invloed
van de omgeving van minder belang zou zijn.
Er zijn een aantal aanwijzingen dat er mogelijks een genetische link bestaat tussen de
factoren die schisis veroorzaken en deze die tot tandagenesie leiden. Deze evidentie komt
vooral uit dierenonderzoek. Hieruit blijkt namelijk dat als men het MSX1 gen
uitgeschakeld (knock-out Mouse) er zich dan niet alleen agenesieën, maar ook schisis
voordoet bij deze knock-out muizen. Anderzijds werd er door familieonderzoek, waarbij
kinderen aanwezig waren die zowel schisis als tandagenesie hebben, aangetoond dat

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
36
verwanten zonder schisis meer agenesie vertoonden dan niet verwanten personen. Dit zijn
twee argumenten om aan een genetische link tussen de oorzakelijke factoren van beide
afwijkingen te denken.
Hoewel we nu kunnen veronderstellen dat genetica een grote rol speelt bij zowel niet-
syndromale schisis als –tandagenesie, is verder onderzoek op niveau van het genoom
noodzakelijk om een genetische link tussen beide met zekerheid aan te tonen.

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
37
Bronnen
Adeboye S.O. , Cole B.O.I. , Hobson R.S. , Wright M.J. Severe hypodontia in a set of triplets. Britisch Dental Journal Volume 201 No. 2 Jul 22 2006 Allan Evens, James Lowe. Histologie van de mens. Bohn staffleu van Loghum. Londen. 1997:182-186
Ardinger HH, Bluetow KH, Bell GI, Bardach J, Vandemark DR, Murray JC. Association of genetic variation of the transforming growth factor – alpha gene with cleft lip and palate. Am J Hum Genet 1989;45:348-353
Brook AH. A unifying aetiological explanation for anomalies of human tooth number and size. Arch Oral Biol 1984;29:373-8
Carels Carine. Orhtodontie en dentofaciale orthopedie. KUL 2005-2006
Chenevix-Trenck G, Jones K, Green AC. Cleft lip with or without cleft palate: associations with transforming growth factor alpha and retinoic acid receptor loci. Am J Human Genet 1992;51:1377-1385
Christensen K, Holm NV, Olsen J, Kock K, Fogh-Anderson P. Selection bias in genetic-epidemiological studies of cleft lip and palata. Am J Hum Genet 1992;51:654-659
Christensen K, Fogh-Andersen P. Isolated cleft palate in Danish multiple births, 1970-1990. Cleft Palate Craniofac J. 1993 Sep;30(5):469-74
Christensen K. , Fogh – Anderson P. Cleft lip (+/- cleft palate) in Danisch twins , 1970 -1990 Am J Med Genet 1993 NOV1;47(6):910-6
Christensen K, Olsen J, Norgaard-Pedersen B. Oral clefts, transforming growth factor alpha gene variants, and maternal smoking: a population-based case-control studye in Denmark, 1991-1994. Am J Epidemiol 1999;149:248-255 Cobourne MT. Familial human hypodontia, is it all in the genes? Br Dent J. 2007 AYG25;203(4):208-8
Conners GP. Van der Woude syndroom. 23 augustus 2007 www.emedicine.com
Das P, Hai M, Elcock C, Leal SM, Brown DT, Brook AH et al Novel missense mutations and 288 bp exonic insertion in PAX9 in families with autosomal dominant hypodontia. Am J Genet 2003;118A:35-42
De Fruyt F et al. De ontwikkeling van meerlingen. Zoekindex Caleidsoscoop. 2000;12:5,4
De Muynck S, Schollen E, Matthijs G, Verdonck A, Devriendt K, Carels C. A novel MSX1 mutation in hypodontia. Am J Med Genet 2004;128A:18-20
Devriendt et al . Erfelijkheid van lipspleet en verhemeltespleet. www.uzleuven.be/schisisteam 2008
Eerens K, Vlietinck R, Heidbücel K, Van Olmen A, Derom V, Willems G, Carels C. Hypodontia and tooth formation in groups of children with cleft, siblings without cleft and nonrelated controls. Cleft Palate Craniofacial Journal July 2001, Vol38 No 4

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
38
Etheredge AJ , Christensen K , Del Junco D , Murray JC , Mitchell LE . Evaluation of two methods for assessing gene – environment interactions using data from de Danish case – control study of facial clefts. Birth Defects Res A Clin Mol Teratol. 2005 Aug;73(8):541-6
Eufinger H, Rand S, Scholz W, Machtens E. Clefts of the lip and palate in twins: use of DNA fingerprinting for zygosity determination. Cleft Palate Craniofac J. 1993 Nov;30(6):564-8.
Ferguson CA, Trucker AS, Christensen L, Lau AL, MatzukMM, Sharpe PT. Avtivin is an essential early mesenchymal signal in tooth development that is required for patterning of the murine dentition. Genes Dev 1998;12:2636-2649
Frazier-Bowers SA, Guo DC, Gavender A, Xue L, Evans B, King T et al. A novel mutation in human PAX9 causes molar oligodontia. J Dent Res 2002;81:129-33
Frazier-Bowers SA, Scott MR, Cavender A, Mensah J, D’Souza RN. Mutational analysis of families affected with molar oligodontia. Connective Tussue Research. 43:296-300,2002
Gartner LP, Hiatt JL. Color textbook of Histology. Second edition. Saunders,USA. 2001:365-373
Jezewski PA, Vieira AR, Nishimura C, et al. Complete sequencing shows a role for MSX1 in non-syndromic cleft lip and palta. J Med Genet 2003;40:399-407
Karsten A. , Larson M. , Larson O. . Length of the cleft in relation to the incidence of hypodontia of inheritance of cleft lip and palate in children with isolated cleft palate. Scan J Plast Reconstr Hand Surg,2005; 39: 283-286
Kato T. , Iwamoto K. , Kakiuchi C. , Kuratomi G. , Okazaki Y. Genetic or epigenetic difference causing discordance between monozygotic twins as aclue to molecular basis of mental disorders. Mol Psychiatry. 2005Jul;10(7):622-630
Kindelan JD, Rysiecki G, Childs WP. Hypodontia: genotype or environment? A case report of monozygotic twins. Br J Orthod. 1998 Aug;25(3):175-8. Kist R., Greally E., Peters H. Derivation of a mouse model for conditional inactivation of PAX9. Genesis 2007 Jul;45(7):460-4
Kondo S, Schutte BC, Richardson RJ, Bjork BC, Knight AS, Watanabe Y. Mutations in IRF6 cause Van der Woude and popliteal ptrygium syndromes. Nat Genet 2002;32:271-277
Lidral AC, Romitti PA, Basart AM, Doetschman T, Leysens NJ, Daack-hirsch S. Association of MSX1 and TGFB3 with nonsyndromic clefting in humans. Am J Hum Genet 1998;63:557-568
Lidral AC, Reising BC. The role of MSX1 in human tooth agenesis. J Dent Res 2002;81:274-278
LidraL AC, Murray J.C. Genetic approaches to identify disease genes for birth defects with cleft lip / palate as a model. Birth defects research ( part A) 70:893-901(2004)
Maestri NE, Beaty TH, Hetmanski JB. Application of transmission disequilibrium tests to nonsyndromic oral clefts: including candidate genes and environmental exposures in the models. Am J Human Genet 1997;39:399-403

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
39
Mansilla MA , Kimani J , Mitchell LE , Christensen K , Boomsa DI , Daack – Hirsch S , Nepomucena B , Wyszynski DF , Felix TM , Martin NG , Murray JC; Discordant MZ twins with cleft lip and palate : a model for identifying genes in complex traits. Twin Res Hum Genet 2005Feb;8(1):39-46
Markovic M. Hypodontia in twins. Swed Dent J Suppl 1982;15:153-162
Mensah J. K. , Ogawa T. , Kapadia H. , Cavender A.C. , D’Souza R.N. Functional analysis of a mutation in PAX9 associated with familial tooth agenesis in humans. JBC papers in press, November 7, 2003
Mitchell LE, Beaty TH, Lidral AC, Munger RG, Murray JC, Saal HM, Wyszynski DF; International Consortium for Oral Clefts Genetics. Guidelines for the design and analysis of studies on nonsyndromic cleft lip and cleft palate in humans: summary report from a Workshop of the International Consortium for Oral Clefts Genetics Cleft Palate Craniofac J. 2002 Jan;39(1):93-100
Mitchell LE , Murray JC , O’Brien S , Christensen K . Retinoic acid receptor alpha gene variants , multivitamin use , and liver intake as risk factors for oral clefts : a population – based case – control study in Denmark. Am J Epidemiol 2003;158:69-76
Modesto A. , Moreno L.M. , Krahn K. , King S. , LIdral A.C. MSX1 and orofacial clefting with and without tooth agenesis. J Dent Res 85(6):542-546,2006
Mostowska A, Kobielak A, Biedzak B, Trzeciak WH. Novel mutation in the paired bos sequence of PAX9 gene in a sporadic form of oligodontia. Eur J Oral Sci 2003;111:272-6
Mostowska A, Kobielak A, Biedzak B, Trzeciak WH. Molecular basis of non-syndromic tooth agenesis : mutations of MSX1 and PAX9 reflect their role in patterning human dentition. Eur J Sci 2003;111:365-70
Mostowska A, Biedzak B, Jagodzinski PP. Axis inhibition protein2 ( AXIN2) polymorphisms may be a risk factor for selective tooth agenesis. J Hum Genet 2006;51:262-6
Murray J C , Schutte B . Interview in the Inside scoop october 2002
Näsman M, Forsberg CM, Dahllöf G. Long-term dental development in children after treatment for malignant disease. Eur J Orthod 1997Apr;19(2):151-9
Nieminen P, Arte S, Tanner D, Paulin L, Alaluusua S, Thesleff I et al. Identification of a nonsense mutation in the PAX9 gene in molar oligodontia. Eur J Hum Genet 2001;9:743-6
Nieminen P, Pekkanen M, Aberg T, Thesleff I. A graphical www- database on gene expression in tooth. Eur J Oral Sci 1998;106:7-11
Nordström RE, Laatikainen T, Juvonen TO, Ranta RE. Cleft-twin sets in Finland 1948-1987. Cleft Palate Craniofac J. 1996 Jul;33(4):340-7.
Peters H, Neubüser A, Kratochwil K, Balling R. Pax9 deficient mica lack pharyngeal pouch derivatives and teeth and exhibit craniofacial and limb abnormalities. Genes Dev 1998; 12:2735-2747
Peters H, Bailling R. Teeth. Where and how to make them. Trends Genet 1999;15:59-65
Petronis A, Irving I. Qottesman, Kan P, Kennedy J L ,Basile V S , Paterson A D , Popendikyte V . Monozygotic Twins Exhibit Numerous Epigenetic Differences: Clues to Twin Discordance? Schizophrenia Bulletin, Vol. 29, No. 1:169 - 178, 2003

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
40
Ranta R. Hypodontia and delayed development of the second premolars in cleft palate children. Cleft Palate J. 1983;20:163-165
Ranta R. A review of tooth formation in children with cleft lip/palate. Am J Orthod Denfofac Orhtop 1986;90:11-18
Rice R, Spencer-Dene B, Conner EC. Clin Inves 2004;113:1692-1700
Sato kata I, Maas R. MSX1 deficient mice exhibit cleft palate and abnormalities of craniofacial and tooth development. Nat genet 1994;6:348-356
Singh SM , Murphy B , O’Reilly R. Epigenetic contributors to the discordance of monozygotic twins. Clin Genet 2002 : 62 : 97 – 103
Slayton RL, Willimas L, Murray JC, Wheeler JJ, Lidral AC, Nishimura AJ. Genetic association studies of cleft lip and/or palate with hypodontia outside the cleft ragion. Cleft palate – Craniofacial Journal, May 2003, Vol 40 No.3
Stegenga B. , Vissink A. , De Bont L.G.M. Mondziekten en Kaakchirurgie 2000: 383-389
Stockton DW, Das P, Goldenberg M, D’Souza RN, Patel PI. Mutation of PAX9 is associated with oligodontia. Nat Genet 2000;24:18-20
Sun – Wei Guo . Does higher concordance in monozygotic twins than in dizygotic twins suggest a genetic component? Hum Hered 2001;51:121-132; 2001
Swinnen S, Bailleul-Forestier I, Arte S., Nieminen P, Devriendt K, Carels C . Investigating the etiology of multiple tooth agenesis in three sisters with severe oligodontia. Orthod Craniofac Res 2008;11:24-31
Thomas BL, Tucker AS, Qiu M, Ferguson CA, Hardcastle Z, Rubenstein JL. Role of dlx1 and dlx2 genes in patterning of the murine dentition. Development 1997;124:4811-4818
Townsend GC. , Richards L. , Hughes T. , Pinkerton S. , Schwerdt W. Epigenetic influences may explain dental differences in monozygotic twin pairs. Australian Dental Journal 2005:50:(2):95-100
Van den Boogaard M.H. , Dorland M. , Beemer F.A. , Van Amstel H.K.P. MSX1 mutation is associated with orofacial clefting and tooth agenesis in humans. Nature genetics 24,342-343(2000)
Van Esch H, Verdonck A, Carels C. Erfelijkheid van niet syndromale orofaciale schisis. Ned Tijdschr Tandheelkunde 2008;115:101-104
Vastardis H. The genetics of human tooth agenesis: new discoveries for understanding dental anomalies. Am J Ortho Dentofacial Orthop 2000;117:650-6
VastardisH, Karimbux N, Guthua SW, Seidmann JG, Seidmann CE. A Human MSX1 homeodomain missense mutation causes evidence tooth agenesis. Nat Genet 1996;13:417-21
Vieira AR. Oral clefts and syndromic formsof tooth agenesis as modals for genetics of isolated tooth agenesis. J Dent Res 82(3):162-165,2003

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
41
Vieira AR, Avila JR, Daack-Hirsch S, Dragan E, Félix TM, Rahimov F, Harrington J, Schultz RR, Watanabe Y, Johnson M, Fang J, O'Brien SE, Orioli IM, Castilla EE, Fitzpatrick DR, Jiang R, Marazita ML, Murray JC. Medical sequencing of candidate genes for nonsyndromic cleft lip and palate . PLoS Genet. 2005 Dec;1(6):e64. Epub 2005 Dec 2
Vieira A.R. , Meira R. , Modesto A. , Murray J.C. MSX1 , PAX9 and TGFA contribute to tooth agenesis in humans. J dent Res 83(9):723-727 , 2004
Warrington A , Vieira A R , Christensen K , Oriolo I M , Castilla E E , Romitti P A , Murray J C . Genetic evidence for the role of loci at 19q13 in cleft lip and palate. J Med Genet 2006;43:26
Wyszynski MD, Beaty TH, Maestri. Genetics of nonsyndromic oral clefts revisisted. Cleft Palate-Craniofacial Journal, September 1996, Vol.33 No.5

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
42

Annelies Kellens, 3e jaar tandarts. Genetische relatie tussen schisis en tandagenesie?
43