
Studie van het ontstaansmechanisme van autor eactiviteit...
57
Studie van h autoreactivitei in lymfotoxin en C Scriptie voorgedrag FACULTEIT GE GEZONDHEIDSW Academiejaar 2008-2009 het ontstaansmechanism it tegenover nucleaire a ne deficiënte muizensta validatie van de LIA Inge VANASSCHE Promotor: Prof. Dr. D. Elewaut Co-promotor: Dr. Jens Van Praet gen in de 2 de Master in het kader van de ople ARTS ENEESKUNDE EN WETENSCHAPPEN me van antigenen ammen eiding tot
Transcript of Studie van het ontstaansmechanisme van autor eactiviteit...

Studie van het ontstaansmechanisme van
autoreactiviteit tegenover nucleaire
in lymfotoxine deficiënte muizenstammen
en validatie van de LIA
Co
Scriptie voorgedragen
FACULTEIT GENEESKUND GEZONDHEIDSWETENSCHA
Academiejaar 2008-2009
Studie van het ontstaansmechanisme van
eactiviteit tegenover nucleaire antigenen
in lymfotoxine deficiënte muizenstammen
en validatie van de LIA
Inge VANASSCHE
Promotor: Prof. Dr. D. Elewaut
Co-promotor: Dr. Jens Van Praet
Scriptie voorgedragen in de 2de Master in het kader van de opleiding tot
ARTS
FACULTEIT GENEESKUNDE EN GEZONDHEIDSWETENSCHAPPEN
Studie van het ontstaansmechanisme van
antigenen
in lymfotoxine deficiënte muizenstammen
in het kader van de opleiding tot

Voorwoord
Toen ik 3 jaar geleden mijn thesis schreef tijdens mijn opleiding biomedische wetenschappen, had ik nooit kunnen denken dat ik hier vandaag weer zou staan, met een thesis voor de opleiding arts. Pas enkele weken na mijn afstuderen begon het te kriebelen, want hoe graag ik als SLE-patiënte ook onderzoek had willen doen naar auto-immuunziekten, het bleef me dwarszitten dat ik niet echt tussen de patiënten zou kunnen staan. En zo ben ik er, onder mijn motto ‘if you can dream it, you can do it’, gewoon voor gegaan. Al was het niet de gemakkelijkste weg, ik heb er nog geen enkel ogenblik spijt van gehad. Integendeel, vandaag kan ik alleen maar trots vaststellen dat ik die droom steeds meer in vervulling zie gaan. Natuurlijk zou dit nooit gelukt zijn zonder de hulp van velen, die ik hierbij graag van harte wil bedanken. Ten eerste zou ik mijn promotor, Prof. Dr. Dirk Elewaut, hartelijk willen bedanken voor de kans die hij me gaf een tweede thesis over SLE te maken, het ‘vervolg’ op mijn vorige thesis. En natuurlijk ook bedankt voor het antwoord op al mijn vragen. Daarnaast bedank ik ook prof. Dr. Filip Dekeyser voor alle hulp. Mijn begeleider, Jens Van Praet, zou ik ongelooflijk willen bedanken voor alle steun de voorbije 2 jaar, voor alle kennis die hij me heeft bijgebracht, voor het antwoord op mijn vele vragen, voor de hulp bij mijn geklungel met de computer en de statistiek, en natuurlijk voor het met engelengeduld verbeteren van mijn schrijfsels, ondanks z’n heel drukke agenda. Ook een grote dankjewel aan Nele en Annette, die me altijd met een glimlach en veel geduld hielpen bij het labowerk als ik het weer eens bijna aan het verknoeien was. Ook zou ik de andere fijne mensen van het labo willen bedanken, die altijd voor een heel gezellige sfeer zorgden in het labo, en waarbij ik ook altijd terecht kon met vragen. En dan sta ik hier tenslotte weer met een dankwoordje voor mijn ouders, zelfs nog groter dan de vorige keer, want ik besef maar al te goed dat niet iedereen het geluk heeft om nog een compleet nieuwe studierichting te mogen beginnen na 4 jaar studeren. Ongelooflijk bedankt voor de kans die jullie me hiertoe gaven, de nooit aflatende steun en schouderklopjes al die jaren, en al hetgeen jullie doen om mijn werk ook maar een beetje te verlichten. Voor mij verdienen jullie een heel grote pluim! Ook aan Marjan, Peter, vrienden, vriendinnen en familie een heel dikke merci!
Inge Vanassche, april 2009

Inhoudstafel
VOORWOORD
INHOUDSTAFEL
AFKORTINGEN
1 ABSTRACT ...................................................................................................................... 1
2 INLEIDING ...................................................................................................................... 3
2.1 HUMANE SLE EN SYSTEEMSCLEROSE ........................................................................................ 3
2.2 MUIZENMODELLEN VOOR SLE EN SYSTEEMSCLEROSE ............................................................. 52.2.1 Muizenmodellen voor SLE .............................................................................................................. 5
2.2.2 Muizenmodellen voor systeemsclerose ............................................................................................ 7
2.3 DETECTIE VAN AUTO-ANTILICHAMEN ........................................................................................ 92.3.1 Inleiding .......................................................................................................................................... 9
2.3.2 Indirecte immuunfluorescentie (IIF) ............................................................................................... 9
2.3.3 ELISA ............................................................................................................................................ 10
2.3.4 Western blotting ............................................................................................................................ 11
2.3.5 LIA ................................................................................................................................................. 11
2.4 ROL VAN DE LYMFOTOXINE- EN TNFR-SIGNAALWEG IN SLE ................................................. 142.4.1 Inleiding ........................................................................................................................................ 14
2.4.2 De liganden van de LTβReceptor-signaalweg .............................................................................. 15
2.4.3 Signaaltransductie via de TNFR1- en LTβReceptor-signaalweg .................................................. 15
2.4.4 Functie van de LTβReceptor-signaalweg ...................................................................................... 16
2.5 DOELSTELLING EN BENADERING .............................................................................................. 182.5.1 Validatie LIA ................................................................................................................................. 18
2.5.2 Rol van de LTβReceptor-signaalweg in het ontstaan van anti-ENA ............................................. 18
2.5.3 Weefselspecifieke antilichamen ..................................................................................................... 19
3 METHODOLOGIE ....................................................................................................... 20
3.1 MUIZEN .................................................................................................................................... 20
3.2 SERUMSTALEN .......................................................................................................................... 21
3.3 ELISA ...................................................................................................................................... 21
3.4 LIA (LINE IMMUNOASSAY) ...................................................................................................... 21
3.5 INDIRECTE IMMUUNFLUORESCENTIE OP LKS-COUPES ............................................................ 22
3.6 STATISTISCHE ANALYSE ........................................................................................................... 23
4 RESULTATEN ............................................................................................................... 24

4.1 VALIDATIE VAN DE LIA ........................................................................................................... 244.1.1 C57BL/6 muizen ............................................................................................................................ 24
4.1.2 MRL/lpr muizen ............................................................................................................................. 24
4.1.3 BWF1 muizen ................................................................................................................................ 27
4.1.4 Pristane-induced lupus muizen ..................................................................................................... 29
4.2 ROL VAN DE LTΒRECEPTOR-SIGNAALWEG IN HET ONTSTAAN VAN ANTI-ENA ...................... 314.2.1 LTβReceptor-/- muizen .................................................................................................................... 31
4.2.2 LTβReceptor-Fc fusieproteïne behandelde muizen ....................................................................... 32
4.2.3 Thymustransplanten ...................................................................................................................... 33
4.3 WEEFSELSPECIFIEKE ANTILICHAMEN ...................................................................................... 34
5 DISCUSSIE ..................................................................................................................... 36
5.1 VALIDATIE VAN DE LIA ........................................................................................................... 365.1.1 MRL/lpr muizen ............................................................................................................................. 36
5.1.2 BWF1 muizen ................................................................................................................................ 37
5.1.3 Pristane-induced lupus muizen ..................................................................................................... 37
5.1.4 Kinetiek van de anti-ENA antilichamen ........................................................................................ 38
5.1.5 Besluit en toekomstperspectieven .................................................................................................. 38
5.2 LTΒRECEPTOR-SIGNAALWEG ................................................................................................... 395.2.1 Rol centrale tolerantie via thymus transplantatie experimenten ................................................... 39
5.2.2 Rol structurele defecten en levensfase ........................................................................................... 40
5.2.3 Toekomstperspectieven .................................................................................................................. 41
5.3 WEEFSELSPECIFIEKE ANTILICHAMEN ...................................................................................... 42
6 REFERENTIELIJST ..................................................................................................... 43
BIJLAGEN

Afkortingen
ANA anti-nucleaire antilichamen AMA anti-mitochondriale antilichamen APCA anti-pariëtaalcel antilichamen ASMA ‘anti-smooth muscle antibodies’ BCIP/NBT 5-bromo-4-chloro-3-inodyl fosfaat/nitro blue tetrazolium DC dendritische cel dsDNA dubbelstrenging DNA ELISA enzyme-linked immunosorbent assay ENA extractable nuclear antigen FITC fluoresceïne-isothiocynaat Hep-2 en -2000 cellijn human epithelial-2 en -2000 cellijn His-fusieproteïne histidine fusieproteïne HRP horseradish-peroxidase HVEM herpesvirus entry mediator IIF indirecte immuunfluorescentie Ig immunoglobuline IVC individual ventilated cages KO knock-out LIA line immunoassay LKS-coupes ‘liver-kidney-stomach’ coupes LTα en β lymfotoxine α en β LTβReceptor lymfotoxine β receptor NK- cellen natural killer cellen iNKT-cellen invariant natural killer T-cellen PBS phosphate buffered saline rpm rounds per minute s.e.m. standard error of the mean SLE systemische lupus erythematosus SSc systeemsclerose TNFα tumor necrosis factor α TNFR1 en 2 tumor necrosis factor receptor 1 en 2 Tsk-1 en -2 muizen tight skin 1 en 2 muizen

Abstract ___________________________________________________________________________
1
1 Abstract
Systemische lupus erythematosus (SLE) is een auto-immuunziekte gekarakteriseerd door
een brede waaier aan klinische manifestaties zoals huidrash, artritis, glomerulonefritis en
vasculitis. Biochemisch zijn verhoogd complement verbruik, pancytopenie, proteïnurie en de
aanwezigheid van antinucleaire antilichamen (ANA en anti-ENA) de belangrijkste kenmerken
van deze ziekte. Deze antilichamen zijn voornamelijk gericht tegen dsDNA, histonen, Sm,
RNP, SS-A/B en histonen. Ook systeemsclerose (SSc) is een auto-immuunaandoening
gekenmerkt door vaataantasting en excessieve fibrose van de huid en inwendige organen. Ook
hier komen bij meer dan 90% van de patiënten autoantilichamen voor. Het gaar hier
voornamelijk om antisclero70, anticentromeer en anti-RNA polymerase III antistoffen.
Onderzoek naar het voorkomen van gelijkaardige auto-immuunfenomenen bij muizen is een
grote hulp om de etiopathogenese van deze ziekten te achterhalen, gezien de meeste
immunologische abnormaliteiten die fundamenteel zijn voor humane auto-immuunziekten
ook voorkomen in de muis. Zo hebben verschillende muizenmodellen voor SLE en SSc ons
de laatste jaren belangrijke inzichten verschaft in het ontstaansmechanisme van deze ziekten.
Een eerste doel van deze thesis was de validatie van de ANA INNO-LIA Update (kortweg
LIA) voor het gebruik bij muizenstalen in plaats van humane stalen. Het grote voordeel van
deze test ten opzichte van andere testen is dat hiermee een hele reeks anti-ENA antilichamen
tegelijk kunnen gedetecteerd worden. Eerder werd al aangetoond dat het hier om een
veelbelovende methode kon gaan, en dit werd hier verder uitgewerkt. Dit gebeurde door een
aantal gekende muizenmodellen voor SLE als positieve controle te testen met de LIA,
namelijk de MRL/lpr, BWF1 en pristane-induced lupus muizen, en de C57BL/6 muizen als
negatieve controle. De bekomen resultaten werden vergeleken met hetgeen in de literatuur
beschreven staat omtrent prevalenties van autoantilichamen in deze muizenmodellen, en dit
bleek in grote lijnen overeen te komen. Bijgevolg zal deze test in de toekomst nog verder
gevalideerd worden, door deze stalen opnieuw te testen met western blotting, en nog enkele
andere muizenmodellen (Tsk-1, NOD muizen) te onderzoeken via LIA en western blotting.
Een tweede doel van dit werk was de rol van de LTβReceptor-signaalweg in het ontstaan
van anti-ENA antilichamen na te gaan. Hierbij werd eerst de rol van de centrale tolerantie
nagegaan, door thymustransplanten met een LTβReceptor-/- thymus en hun controles te
onderzoeken via de LIA. Bij deze dieren konden geen anti-ENA antilichamen aangetoond

Abstract ___________________________________________________________________________
2
worden, hoewel in de literatuur bij de thymustransplanten een gestoorde thymusarchitectuur
beschreven wordt, waaruit we dus besluiten dat een afwijking in de centrale tolerantie op zich
niet voldoende is voor het ontstaan van deze anti-ENA antilichamen.
Vervolgens werd de rol van structurele defecten in het ontstaan van deze anti-ENA
antilichamen nagegaan, en in welke levensfase de LTβReceptor-signaalweg belangrijk is om
het ontstaan van deze antilichamen te vermijden. Dit gebeurde door onderzoek van de
LTβReceptor-/- muizen en muizen waarbij pre- en/of postnataal op verschillende tijdstippen
een solubel LTβReceptor-Fc fusieproteïne werd toegediend, en door deze met elkaar te
vergelijken. Bij de LTβReceptor-/- muizen ontbreekt deze signaalweg dus zowel pre- en
postnataal, en bij deze dieren werden afwezige lymfeknopen en Peyerse platen beschreven,
evenals een grondig verstoorde miltarchitectuur. Met de LIA werden bij deze dieren anti-ENA
aangetoond. Bij de muizen die voornamelijk postnataal een solubel LTβReceptor-Fc
fusieproteïne kregen toegediend, ontbreekt deze signaalweg dus hoofdzakelijk postnataal en
hierbij zijn de lymfeknopen wel aanwezig, de Peyerse platen ontbreken echter en de
miltarchitectuur is ook verstoord. Ook bij deze dieren werden met de LIA anti-ENA
aangetoond. Bij de muizen die enkel prenataal behandeld werden met het LTβReceptor-Fc
fusieproteïne ontbreken de Peyerse platen en bijna alle lymfeknopen, maar de milt
architectuur is volledig normaal. Bij deze dieren konden via LIA geen anti-ENA aangetoond
worden. Hieruit kunnen we dus besluiten dat de LTβReceptor-signaalweg voornamelijk
postnataal van belang is om auto-immuniteit te voorkomen, en dat afwezige lymfeknopen dus
geen anti-ENA antilichamen doen ontstaan. Gezien bij beide groepen met een gestoorde
miltarchitectuur echter wel anti-ENA werden aangetoond, kunnen we besluiten dat een
gestoorde miltarchitectuur mogelijks betrokken is bij het ontstaan van deze antilichamen. De
rol van deze pathway zal in de toekomst nog verder onderzocht worden aan de hand van
LIGHT-/- muizen, celspecifieke LTβ-/- muizen en Toll-like receptor 7-/- muizen.
Tenslotte werden de sera van deze LTβReceptor-/- muizen onderzocht via IIF op LKS
coupes. Hierbij werd nagegaan of bij deze dieren naast anti-ENA antilichamen, die gericht
zijn tegen algemene bestanddelen van alle cellen, nog andere antilichamen aanwezig zijn, die
gericht zijn tegen een bepaald celtype. Hierbij krijgt men een meer volledig beeld over de
auto-immuniteit aanwezig in deze dieren. Zo konden bij bijna alle dieren op 6 maand ASMA
(anti-smooth muscle antibodies) aangetoond worden, die in de humane situatie gecorreleerd
zijn met auto-immuunhepatitis type 1. Ook konden bij een deel van deze dieren een
transaminasestijging aangetoond worden, wat wijst op een mogelijk pathofysiologische rol
van deze ASMA.

Inleiding ___________________________________________________________________________
3
2 Inleiding
2.1
Systemische lupus erythematosus (SLE) is een multi-systeem auto-immuunziekte
gekarakteriseerd door een brede waaier aan klinische manifestaties zoals huidrash, artritis,
glomerulonefritis, vasculitis en neurologische symptomen. De ziekte komt wereldwijd voor in
een frequentie tussen 1 en 10 per 100.000. Vooral adolescente en jong volwassen vrouwen
worden getroffen. De vrouw:man ratio bedraagt 9:1. Biochemisch zijn verhoogd complement
verbruik, pancytopenie, proteïnurie en de aanwezigheid van hoge titers auto-antilichamen
tegen nucleaire componenten, de antinucleaire antilichamen (ANA), de belangrijkste
kenmerken van deze ziekte (D’Cruz et al., 2007). Vooral antilichamen tegen dsDNA
(dubbelstrengig DNA) zijn kenmerkend voor SLE, en fluctuaties in de dsDNA antilichaam
titers worden gebruikt als merkers voor de ziekteactiviteit en renale opstoten (Linnik et al.,
2005). Daarnaast kunnen bij deze patiënten vaak antilichamen teruggevonden worden tegen
histonen, Sm, SS-A/B, RNP en histonen (Fritzler and salazar, 1991). De ANA kunnen met
verscheidene antigenen immuuncomplexen vormen, die neerslaan in de nieren, bloedvaten en
andere organen. Deze immuuncomplexen activeren complement en promoten cytokine-
responsen, waardoor lokale inflammatie en weefselschade ontstaat (Cortés-Hernández et al.,
2004). De behandeling van SLE varieert van antimalariamiddelen tot corticosteroïden en
immuunsuppressie. De oorzaak van de ziekte is nog onduidelijk, gezien het om een complexe
multifactoriële aandoening gaat. Verschillende hypothesen werden reeds vooropgesteld,
waaronder de rol van een gestoorde apoptose in de etiologie van SLE (Vanassche, 2006).
Momenteel wordt hieromtrent veel onderzoek uitgevoerd, met als doel een meer gerichte
behandeling van SLE.
Humane SLE en systeemsclerose
Ook systeemsclerose (SSc) is een chronische auto-immuunaandoening van nog onbekende
oorsprong, gekenmerkt door vaataantasting en excessieve fibrose van de huid en inwendige
organen. Deze kan zowel de longen, het hart, als het maag-darmstelsel aantasten. Klinisch kan
de vaataantasting zich naargelang het aangetaste vaatbed uiten in een Raynaud fenomeen,
ulcera ter hoogte van de vingers, pulmonale arteriële hypertensie of een niercrisis (Abraham
and Varga, 2005). Ook hier gaat het om een auto-immuunaandoening, gezien de
aanwezigheid van specifieke antinucleaire antilichamen bij meer dan 90% van de patiënten.
Het gaat in hoofdzaak om anti-sclero70 (of anti-topoisomerase I), anti-centromeer en anti-

Inleiding ___________________________________________________________________________
4
RNA polymerase III antistoffen (Fritzler and Salazar, 1991). Hoewel deze antinucleaire
antilichamen klinisch zeer nuttig zijn als biomerker voor de diagnose en prognose, is er geen
duidelijke evidentie voor hun rol in de pathogenese. De prevalentie van de ziekte bedraagt
ongeveer 1 op 5000. Zoals bij vele andere auto-immuunziekten, is ook SSc meest prevalent in
vrouwen, en de ziekte ontstaat gemiddeld rond de leeftijd van 40-45 jaar (Eckes et al., 2007).
Onderzoek naar het voorkomen van gelijkaardige auto-immuunfenomenen bij muizen zou het
etiopathogenetisch onderzoek naar deze auto-immuunziekten verder vooruit kunnen helpen,
en een grote hulp zijn in de ontwikkeling en het testen van verscheidene immunologische
concepten. Immers, de meeste immunologische abnormaliteiten die fundamenteel zijn voor
humane auto-immuunziekten, komen ook voor in de muis (Theofilopoulos and Dixon, 1985).
Door de pathologie en etiologie van SLE en SSc bij muizen beter te begrijpen, hoopt men ook
meer inzicht te krijgen in de humane vormen van deze aandoeningen, en in auto-immuniteit
en immuunregulatie in het algemeen.
Inleiding ___________________________________________________________________________
5
2.2
2.2.1 Muizenmodellen voor SLE
Muizenmodellen voor SLE en systeemsclerose
De laatste 30 jaar hebben verschillende muizenmodellen ons belangrijke inzichten bijgebracht
in de pathogenese van SLE. Men kan ze onderverdelen in 4 groepen volgens hun
ontstaanswijze:
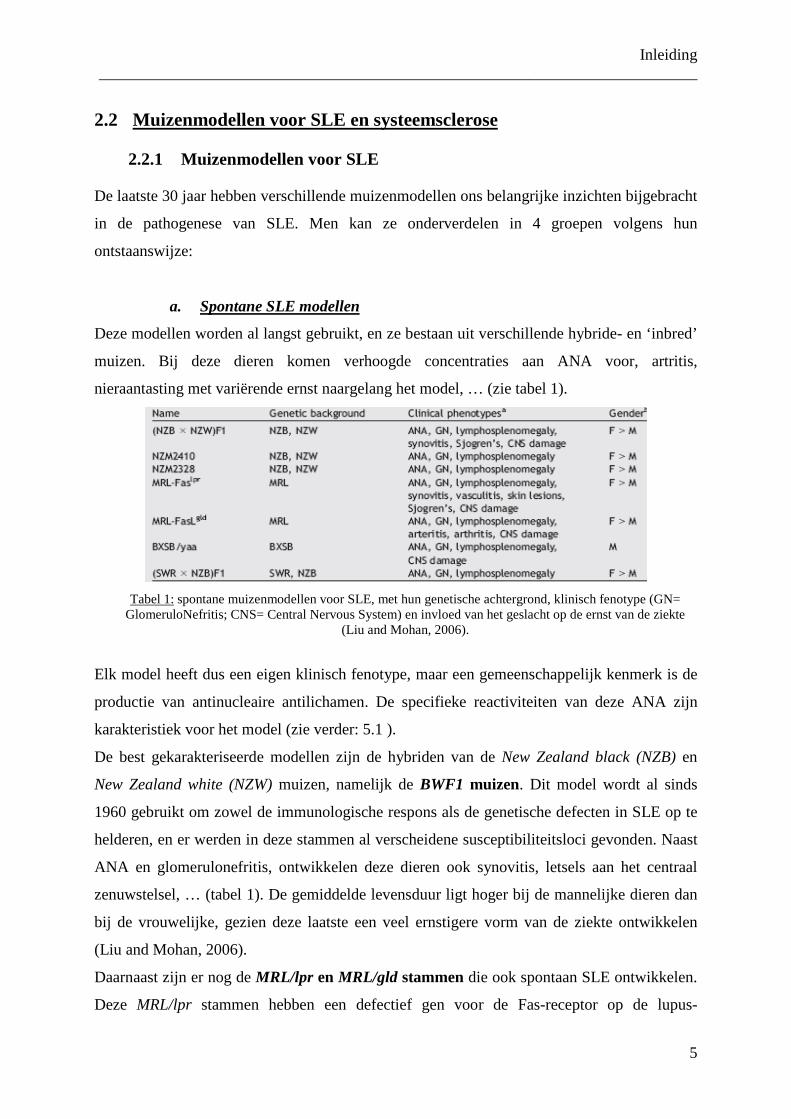
a. Deze modellen worden al langst gebruikt, en ze bestaan uit verschillende hybride- en ‘inbred’
muizen. Bij deze dieren komen verhoogde concentraties aan ANA voor, artritis,
nieraantasting met variërende ernst naargelang het model, … (zie tabel 1).
Spontane SLE modellen
Tabel 1:
Daarnaast zijn er nog de MRL/lpr en MRL/gld stammen die ook spontaan SLE ontwikkelen.
Deze MRL/lpr stammen hebben een defectief gen voor de Fas-receptor op de lupus-
spontane muizenmodellen voor SLE, met hun genetische achtergrond, klinisch fenotype (GN= GlomeruloNefritis; CNS= Central Nervous System) en invloed van het geslacht op de ernst van de ziekte
(Liu and Mohan, 2006).
Elk model heeft dus een eigen klinisch fenotype, maar een gemeenschappelijk kenmerk is de
productie van antinucleaire antilichamen. De specifieke reactiviteiten van deze ANA zijn
karakteristiek voor het model (zie verder: 5.1 ).
De best gekarakteriseerde modellen zijn de hybriden van de New Zealand black (NZB) en
New Zealand white (NZW) muizen, namelijk de BWF1 muizen. Dit model wordt al sinds
1960 gebruikt om zowel de immunologische respons als de genetische defecten in SLE op te
helderen, en er werden in deze stammen al verscheidene susceptibiliteitsloci gevonden. Naast
ANA en glomerulonefritis, ontwikkelen deze dieren ook synovitis, letsels aan het centraal
zenuwstelsel, … (tabel 1). De gemiddelde levensduur ligt hoger bij de mannelijke dieren dan
bij de vrouwelijke, gezien deze laatste een veel ernstigere vorm van de ziekte ontwikkelen
(Liu and Mohan, 2006).

Inleiding ___________________________________________________________________________
6
voorbestemde MRL-achtergrond, en de MRL/gld stammen een defectief gen voor het Fas-
ligand op de MRL-achtergrond. Naast deze 2 loci zijn er nog verscheidene additionele loci
recentelijk geïdentificeerd in hun achtergrond genoom (Liu and Mohan, 2006). De MRL/lpr
dieren worden gekenmerkt door enorme lymfosplenomegalie, glomerulonefritis, de
aanwezigheid van ANA, aantasting van longen en vaten,… Ook hier overleven de mannelijke
dieren enkele weken langer dan de vrouwelijke (Murphy and Roths, 1978).
Volledigheidshalve dient te worden vermeld dat er ook nog andere spontane muizenmodellen
bestaan (tabel 1), maar hierop wordt niet dieper ingegaan aangezien van deze muizenmodellen
minder gebruik wordt gemaakt. In dit werk werd gebruik gemaakt van de BWF1 en MRL/lpr
muizenstam.
b. Een andere manier om de functie of betrokkenheid van genen in een ziekte te achterhalen is
via de knock-out- (KO-) technologie, waarbij men in een dier een gen uitschakelt en
vervolgens kijkt welk fenotype zich ontwikkelt. Op die manier heeft men ontdekt dat de
deficiëntie in veel verschillende eiwitten kan leiden tot een pathologie gelijkaardig aan SLE,
bijv. in eiwitten betrokken bij apoptose, in eiwitten die de B- en T-cel activatie en proliferatie
controleren,… Ook dit is een krachtige en goede strategie, maar toch zijn er beperkingen bij
dit soort modellen. Zo kan met deze strategie bijvoorbeeld moeilijk achterhaald worden wat
de fenotypische consequenties van kleine polymorfismen in kandidaatgenen zijn (Liu and
Mohan, 2006). Ook in dit werk werd gebruik gemaakt van zo’n KO-muizen, namelijk de
LTβReceptor-/- muizen.
KO-muizen
c. Via linkage analyse zijn er al heel wat loci geïdentificeerd die geassocieerd zijn met bepaalde
polygene ziekten, maar via welk mechanisme de genen op deze loci dan uiteindelijk bijdragen
tot de ziekte, is dikwijls nog niet bekend. Congene diermodellen zijn een goeie strategie om
hierin wel inzichten te verwerven. Hierbij gaat men de verschillende gevonden loci die
bijdragen tot een ziekte, één voor één inbrengen in een collectie van unieke substammen,
waarbij elke substam dus een bepaalde locus draagt. Op die manier werden al verscheidene
susceptibiliteitsgenen voor SLE ingebracht in het genoom van ’lupus-resistente stammen’,
zoals de C57BL/6 en de BALB/c stam. Deze nieuw gegenereerde muisstammen dragen bij tot
een unieke en krachtige strategie om de lupus pathogenese te achterhalen (Liu and Mohan,
2006). Van deze muizenmodellen werd in dit werk geen gebruik gemaakt.
Congene SLE muizenmodellen
Inleiding ___________________________________________________________________________
7
d. Tenslotte kan intraperitoneale injectie van het adjuvans pristane ook in muizenmodellen die
genetisch niet voorbeschikt zijn voor auto-immuniteit een SLE-muizenmodel induceren,
bijvoorbeeld in de lupus-resistente stammen C57BL/6 en BALB/c (Satoh et al., 1995; Savarese
et al., 2008). Hiervoor is dus geen voor lupus voorbeschikte achtergrond nodig, in
tegenstelling tot bijvoorbeeld de eerder beschreven MRL/lpr muizen, die de lupus prone MRL
achtergrond vereisen. Deze pristane-injectie leidt immers tot een chronische peritonitis met de
vorming van ectopische lymfoïde weefsels, wat ook in verschillende auto-immuunziekten
wordt beschreven. Muizen geïnjecteerd met pristane vormen sequentieel antilichamen tegen
ss-DNA, ds-DNA, Sm en RNP antigenen, gelijkaardig aan de humane lupus. Na 6 maand
ontstaan bij deze dieren ook immuun-complex-gemedieerde glomerulonefritis, gevolgd door
artritis. Deze modellen lijken dus op de humane drug-induced lupus, met het grote verschil
dat bij deze dieren de ontstane ANA een veel breder spectrum hebben (Savarese et al, 2008).
Ook in dit werk werden deze modellen gebruikt.
Drug induced SLE muizenmodellen
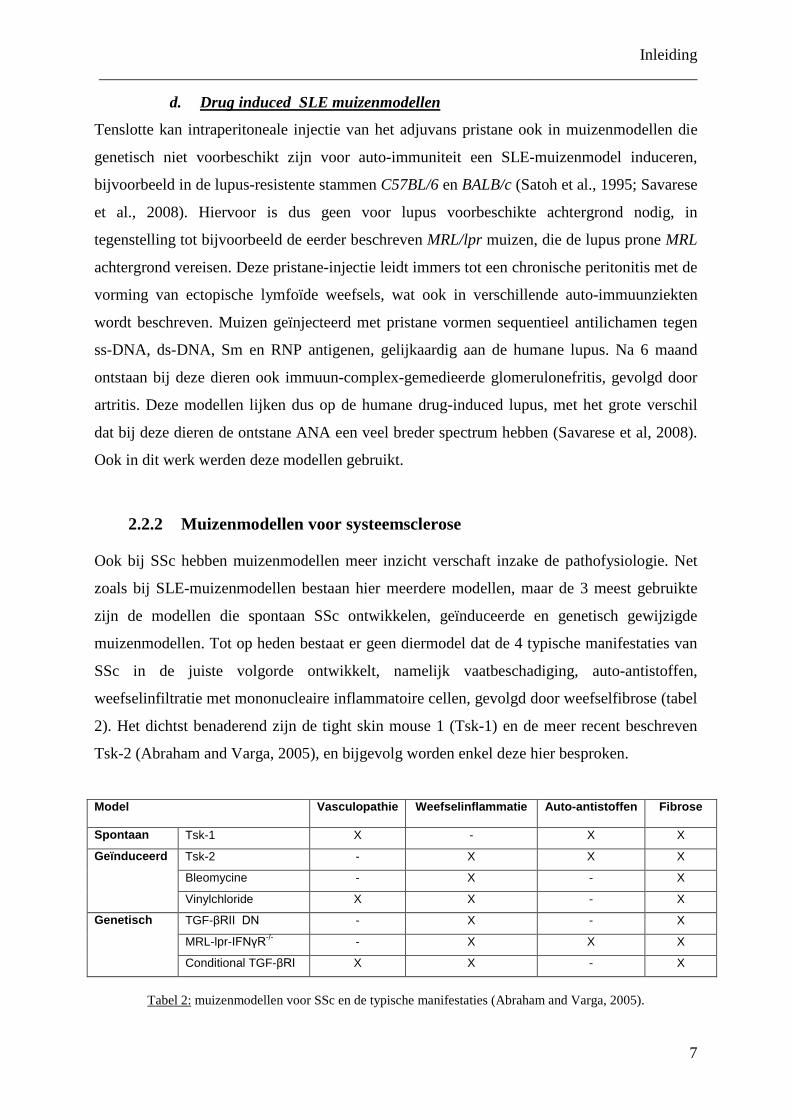
2.2.2 Muizenmodellen voor systeemsclerose
Ook bij SSc hebben muizenmodellen meer inzicht verschaft inzake de pathofysiologie. Net
zoals bij SLE-muizenmodellen bestaan hier meerdere modellen, maar de 3 meest gebruikte
zijn de modellen die spontaan SSc ontwikkelen, geïnduceerde en genetisch gewijzigde
muizenmodellen. Tot op heden bestaat er geen diermodel dat de 4 typische manifestaties van
SSc in de juiste volgorde ontwikkelt, namelijk vaatbeschadiging, auto-antistoffen,
weefselinfiltratie met mononucleaire inflammatoire cellen, gevolgd door weefselfibrose (tabel
2). Het dichtst benaderend zijn de tight skin mouse 1 (Tsk-1) en de meer recent beschreven
Tsk-2 (Abraham and Varga, 2005), en bijgevolg worden enkel deze hier besproken.
Model
Vasculopathie Weefselinflammatie Auto-antistoffen Fibrose
Spontaan Tsk-1 X - X X
Geïnduceerd Tsk-2 - X X X
Bleomycine - X - X
Vinylchloride X X - X
Genetisch TGF-βRII DN - X - X
MRL-lpr-IFNγR-/- - X X X
Conditional TGF-βRI X X - X
Tabel 2: muizenmodellen voor SSc en de typische manifestaties (Abraham and Varga, 2005).

Inleiding ___________________________________________________________________________
8
Bij het Tsk-1 model is er spontaan een duplicatie opgetreden van het fibrilline-1 gen. Deze
mutatie is lethaal indien zij homozygoot voorkomt, en deze dieren sterven dan ook in utero na
8 à 10 dagen zwangerschap. De heterozygote dieren daarentegen overleven, en vertonen
cutane en viscerale afwijkingen, naast biochemische en moleculaire abnormaliteiten
vergelijkbaar met de humane SSc (Jimenez and Christner, 2002). In tegenstelling tot de
humane SSc, komt bij deze dieren echter geen inflammatoir infiltraat voor in de huid en is
enkel de hypodermis verdikt, terwijl bij de humane vorm de dermis aangetast is. Ook wordt
bij dit model longemfyseem gevonden in plaats van de humane longfibrose (Hasegawa et al.,
2006).
Ook bij het Tsk-2 model gaat het om longemfyseem in plaats van fibrose, en deze dieren
vertonen ook geen vaataantasting. Het inflammatoir infiltraat komt hier echter wel in de
dermis voor, en het wordt gevolgd door fibrose. Daarnaast vertonen deze dieren ook
glomerulonefritis (Gentiletti et al., 2006). Dit model werden bekomen door toediening van het
mutagene ethylnitrosourea.

Inleiding ___________________________________________________________________________
9
2.3
2.3.1 Inleiding
Detectie van auto-antilichamen
Auto-immuunziekten worden gekarakteriseerd door de vorming van antilichamen tegen auto-
antigenen. Deze auto-antigenen kunnen op verschillende manieren geclassificeerd worden,
zoals celtype of weefsel waarin het voorkomt, functie en cellulaire lokalisatie. Bij SLE en SSc
komen volgens deze laatste indeling onder meer antilichamen tegen nucleaire bestanddelen
voor. De indirecte immuunfluorescentie (IIF) op HEp-2 of HEp-2000 cellen is momenteel de
screeningstest die gebruikt wordt om deze ANA aan te tonen. Een positieve fluorescente
kleuring toont de aanwezigheid van ANA aan, maar verder geeft het weinig informatie
omtrent de precieze identificatie van deze ANA, met uitzondering van anticentromeer
antistoffen. Hiervoor zijn bijkomende tests nodig, zoals ELISA en line immunoassay (LIA),
die dan in typische gevallen anti-dsDNA, anti-Sm, anti-histon, anti-RNP en anti-SS-A/B
antilichamen aantonen in geval van SLE, en hoofdzakelijk antisclero70 (of anti-
topoisomerase I), anticentromeer en anti-RNA polymerase III antistoffen in geval van SSc
(Fritzler and Salazar, 1991). IIF wordt ook gebruikt om autoantilichamen tegen bepaalde
celtypes te detecteren, zoals bijvoorbeeld anti-pariëtaalcel antilichamen. Hieronder wordt een
overzicht gegeven van de gebruikte detectietechnieken en hun toepasbaarheid.
2.3.2 Indirecte immuunfluorescentie (IIF)
Bij deze techniek laat men natieve antigenen in situ reageren met de eventueel aanwezige
antilichamen in het serum, waarbij na binding van FITC-gelabeld secundair antilichaam met
deze AG-AL complexen visualisatie mogelijk wordt met een fluorescentie-microscoop.
Om ANA te detecteren werden oorspronkelijk heel wat celtypes gebruikt, zoals nier- of
leverweefsel afkomstig van ratten of muizen. Om de test meer te standaardiseren zijn deze nu
vervangen door in cultuur gebrachte cellijnen, zoals de HEp-2 (human epithelial-2) cellijn,
afkomstig van larynx epithelioma kanker, en de HEp-2000 cellijn die bestaat uit dezelfde
HEp-2 cellen, getransfecteerd met SSA/Ro60 cDNA (Pollock and Toh, 1999). Met deze
cellijnen kunnen verschillende antilichamen tegen heel wat celkerncomponenten gedetecteerd
worden, zoals de antilichamen tegen DNA, histonen, RNP, Sm, SSA/Ro en centromeren,
maar met lage positief predictieve waarde. Daarbovenop kunnen ook enkele cytoplasmatische
antilichamen geïdentificeerd worden. Deze screeningstest wordt ook gebruikt om de
aanwezigheid van ANA aan te tonen in muizenserum. Omwille van de hoge

Inleiding ___________________________________________________________________________
10
conserveringsgraad over de species heen van deze belangrijke nucleaire componenten, binden
de aanwezige muis-antinucleaire antilichamen met een gelijkaardige affiniteit aan de
reactiviteiten aanwezig in HEp-2/HEp-2000. Dit werd al in heel wat studies toegepast
(Gentiletti et al., 2006; Bygrave et al., 2004). Echter, er werd aangetoond dat deze test vrij
aspecifiek is (Vanassche, 2006) net zoals in de humane setting, en bijgevolg werd er in dit
werk geen gebruik van gemaakt.
Om met IIF autoantilichamen die gericht zijn tegen welbepaalde celtypes te detecteren, maakt
men gebruik van coupes bestaande uit lever-, nier- en maagweefsel afkomstig van ratten,
namelijk de LKS-coupes. Door deze 3 verschillende weefsels tegelijk te gebruiken, kunnen de
verschillende types auto-antilichamen makkelijker gedifferentieerd worden van elkaar, door
hun aanwezigheid te vergelijken op de 3 weefsels. De 3 belangrijkste antilichamen die
hiermee kunnen aangetoond worden zijn naast de ANA, de anti-mitochondriale antilichamen
(AMA), de “anti-smooth muscle antibodies”(ASMA) en de anti-pariëtaalcel antilichamen
(APCA) (Bradwell et al., 1997). Ook in dit werk werd deze techniek toegepast.
2.3.3 ELISA
De ‘indirecte ELISA’ is een volgende methode waarbij men antilichamen kan detecteren.
Hierbij werkt men met platen waarop het antigen gecoat is, men voegt het serum met de
antilichamen toe, en men maakt de antigen-antilichaam-complexen zichtbaar door een
gelabeld antilichaam toe te voegen. De ELISA-techniek kan toegepast worden voor de
detectie van antilichamen in humane sera, maar ook in het serum van allerlei andere species,
waaronder muizen, waarbij men een passend gelabeld antilichaam gebruikt, met reactiviteit
tegen het geteste species. De ELISA detecteert zowel lage als hoge affiniteitsantilichamen,
zodat het sensitiever is dan de andere methoden. Het is echter wel heel belangrijk met puur
antigen te werken om een hoge specificiteit te verkrijgen, en deze zijn in het geval van ANA
moeilijk te verkrijgen aangezien de meeste auto-antigenen deel uitmaken van cellulaire
nucleoproteïnen of multicellulaire complexen zoals splicing complexen en nucleosomen, die
moeilijk op te zuiveren zijn. Een ander nadeel van ELISA is dat men slechts de aanwezigheid
van 1 antigen tegelijk kan testen, wat dus neerkomt op een heel arbeidsintensieve procedure
(Goldsby et al., 2003).

Inleiding ___________________________________________________________________________
11
2.3.4 Western blotting
Bij western blotting maakt men gebruik van gel elektroforese om eiwitten, bijvoorbeeld
antilichamen, te scheiden van andere eiwitten op basis van de lengte of 3D-structuur van het
polypeptide. Vervolgens worden de eiwitten overgebracht naar een membraan, waar ze na
binding met gelabelde antilichamen kunnen gedetecteerd worden (Kurien and Scofield, 2006).
In dit werk werd hier geen gebruik van gemaakt, en bijgevolg wordt hier ook niet dieper op
ingegaan.
2.3.5 LIA
Bij de ANA INNO-LIA Update tenslotte, of kortweg LIA (Line ImmunoAssay), kunnen een
hele reeks anti-ENA antilichamen tegelijk worden gedetecteerd, en deze methode zou een
gelijke of zelfs hogere sensitiviteit en specificiteit hebben dan de eerder vermelde
conventionele technieken (Meheus et al., 1999).
a. De INNO-LIATM ANA Update is gebaseerd op het principe van een line immunoassay, en is
gelijkaardig aan de ELISA, met als groot verschil dat de LIA niet kwantitatief is. Het te testen
staal wordt geïncubeerd op een teststrip, waarop de antigenen en een positieve controle lijn
(bestaande uit IgG) in dunne parallelle lijntjes gecoat zijn (figuur 1). Hierop kunnen de
aanwezige antigenen in het serum binden, en dit antigen-antilichaam complex wordt zichtbaar
gemaakt door een antilichaam gelabeld met alkaline fosfatase toe te voegen, gevolgd door het
chromogeen BCIP/NBT dat door dit alkaline fosfatase omgezet wordt in een bruine kleur. De
intensiteit van de gevormde kleur is proportioneel aan de hoeveelheid specifiek antilichaam in
het staal. De commerciële kit gebaseerd op deze technologie, wordt geproduceerd door
Innogenetics (Zwijnaarde, België).
Principe van LIA
b. De antigenen gecoat op de strip zijn ofwel recombinante of natuurlijke proteïnen, ofwel
peptiden. De recombinante antigenen werden getransfecteerd als His-fusieproteïne in E. coli.
Vervolgens werden deze His-fusieproteïnen gezuiverd via nikkel-geimmobiliseerde metaal
affiniteitschromatografie (Ni-IMAC). Deze antigenen kwamen dus als ‘full-length’ proteïnen
tot expressie, en werden als dusdanig op de strip gecoat.
Antigenen

Inleiding ___________________________________________________________________________
12
anti-Sm (Sm
anti-RNP (ribonucleoproteïn) →
ith: patiëntnaam) →
anti-SS-A/B (Soluble Substance A /B) anti-Ro (Robert: patiëntnaam) → →anti-La (La
anti-Cenp-B (
ne: patiëntnaam)
cenanti-Scl 70 (
tromeer proteïne B) Scleroderma 70
anti-
) → →anti-TopoI(TopoisomeraseI) anti-Jo 1 (patiëntnaam) → →anti-histidyl tRNA synthetase
Ribosomal
Phosphoproteïn →
c. Zoals hoger vermeld, is het belangrijkste voordeel van INNO-LIATM ANA Update ten
opzichte van andere detectietechnieken dat een heel gamma aan specifieke reactiviteiten in
één en dezelfde test kunnen gedetecteerd worden. De INNO-LIATM ANA Update werd echter
gevalideerd en ontworpen voor gebruik van humane sera. Ook in de muis-context zou een
dergelijke techniek een belangrijke doorbraak kunnen betekenen naar screening van nucleaire
antilichamen in verschillende muizenmodellen. Gezien de hoge conserveringsgraad van de
verschillende antigenen, kan men kruisreactiviteit verwachten van muis-antilichamen. In dit
werk wordt uitgezocht of deze INNO-LIATM ANA Update mits de nodige aanpassingen ook
kan gebruikt worden om de aanwezigheid van deze specifieke anti-ENA antilichamen te
testen in muizenstalen.
LIA: toepasbaar op muizenstalen?
Ten eerste zou het substraat (menselijke antigenen) geen probleem mogen vormen, wegens
de hoge conserveringsgraad van de nucleaire antigenen over verschillende species heen
(zie ook IIF: 2.3.2). De muizen-antilichamen aanwezig in de verdunde stalen die hierop
toegevoegd worden, zouden moeten binden op deze humane antigenen.
Vervolgens kan men het anti-humaan IgG- specifiek antilichaam gelabeld met alkaline
fosfatase, vervangen door anti-muis antilichaam gelabeld met alkaline fosfatase, dit als
polyreactief of IgG-specifiek antilichaam. Als dan vervolgens eenzelfde hoeveelheid
chromogeen BCIP/NBT toegevoegd wordt, reageert dit met het alkaline fosfatase en de
kleurreactie treedt op. Deze wordt vervolgens gestopt met eenzelfde hoeveelheid en
Figuur 1: Line Immunoassay gecoat met natuurlijke, synthetische en recombinante antigenen; met de oorsprong van de naam (www.innogenetics.be).

Inleiding ___________________________________________________________________________
13
concentratie zwavelzuur. Om zoveel mogelijk reactiviteiten te detecteren, werd hier
gebruik gemaakt van een polyreactief anti-muis antilichaam.
Het positieve controle lijntje bovenaan de strip bestaat uit humane IgG antilichamen. In
geval van muizenstalen gebruikt men een anti-muis antilichaam, maar door kruis-
reactiviteit zal dit toch kunnen reageren met deze gecoate humane IgG’s, en ook de
aanwezige ANA in de muizenstalen zullen hiermee dus kunnen reageren, waardoor het
controlelijntje positief zal zijn.
In de humane situatie worden de resultaten van de LIA vergeleken met een cut-off strook.
Deze cut-off werd bepaald om met een zo hoog mogelijke sensitiviteit en specificiteit de aan-
of afwezigheid van ziekte te voorspellen. Elke concentratie hoger dan de cut-off (of dus met
een donkerdere kleur op de strip dan deze van de cut-off strip) wordt als positief beschouwd,
elke concentratie lager dan de cut-off of dus minder gekleurd, als negatief. Het is dus logisch
dat deze ‘humane’ cut-off niet kan toegepast worden bij de muizenstalen, en bijgevolg werd
een nieuwe cut-off gedefinieerd via uitverdunnen van een sterk positief staal, waarbij de
hoogste verdunning geen vals positieve testen gaf bij de negatieve controlestalen (zie 4.1.1).

Inleiding ___________________________________________________________________________
14
2.4
2.4.1 Inleiding
Rol van de lymfotoxine- en TNFR-signaalweg in SLE
Cytokines zijn signaalmoleculen die betrokken zijn in verscheidene fysiologische processen,
zoals hematopoëse, celdifferentiatie en de initiatie, het behoud en de modulatie van
defensiemechanismen van de gastheer. Deze cytokines worden meestal gesecreteerd door één
bepaald celtype om te binden aan een bepaalde receptor op een targetcel, die zorgt voor
signaaltransductie, en zo onder andere celactivatie, proliferatie, maturatie, differentiatie,
adhesie, migratie en de synthese van acute fase proteïnes op gang brengt. Daarnaast zorgen ze
ook voor de productie van andere cytokines en hun antagonisten en receptoren, zodat een heel
‘cytokine signaalnetwerk’ ontstaat. Individuele cytokines kunnen meerdere functies hebben,
die uniek zijn of ook voorkomen bij andere cytokines (Shields et al., 1995). Al deze cytokines
worden ondergebracht in verschillende families op basis van hun structuur. Eén van deze
families is de TNF-familie, die heel wat leden bevat, zoals TNFα, lymfotoxine α, lymfotoxine
β, LIGHT, CD40 ligand en Fas ligand (Janeway et al., 2005) (figuur 2).
Figuur 2: Liganden en receptoren van het TNF/ lymfotoxine systeem (Gommerman and Browning, 2003).

Inleiding ___________________________________________________________________________
15
2.4.2 De liganden van de LTβReceptor-signaalweg
LTα wordt geproduceerd en gesecreteerd door macrofagen, NK-cellen en T-cellen en
associeert tot een homotrimeer (LTα3). Dit homotrimeer lijkt structureel op TNFα, en ze
kunnen beiden binden op de TNF-receptor I en II. Daarnaast kan LTα na associatie met LTβ
ook binden op de LTβReceptor (figuur 2). Dit LTβ is een transmembranair proteïne dat
voorkomt op de celmembraan van geactiveerde B- en T-lymfocyten, NK-cellen en op een
bepaalde groep van rustende B-cellen (Browning et al., 1993). 2 eenheden LTβ associëren
met 1 LTα-eenheid, en zo vormt het een membraangebonden heterotrimeer (LTα1β2). Dit
LTα1β2 bindt enkel op de LTβReceptor (figuur 2), die voornamelijk voorkomt op myeloïde en
niet-hematopoëtische celtypes. Op die manier functioneert LTβ als een communicatie link
tussen lymfocyten en stromale cellen (Gommerman and Browning, 2003).
Daarnaast kan ook LIGHT op de LTβReceptor binden. Dit transmembranair proteïne komt
tot expressie op geactiveerde T-cellen en immature DC’s, en kan naast de LTβReceptor ook
nog binden op de herpesvirus entry mediator (HVEM), die tot expressie komt op T-cellen
(Granger and Ware, 2001).
De functie van deze liganden wordt besproken aan de hand van de functie van de LTβR-
signaalweg (zie 2.4.4).
2.4.3 Signaaltransductie via de TNFR1- en LTβReceptor-signaalweg
Na binding van het ligand op de TNFR1 of LTβReceptor, treedt receptor aggregatie op. Dit is
de eerste stap van een hele signaalcascade, waarbij telkens nieuwe eiwitten en domeinen
geactiveerd worden (figuur 3).
Figuur 3: signaaltransductiepathways via TNFR1- en LTβR-pathway (Ware, 2005).

Inleiding ___________________________________________________________________________
16
Voor de TNFR-signaalweg kan via caspase 8 activatie enerzijds apoptose gestart worden,
maar via TRAF2 en IκB-degradatie kan ook de canonische NF-κB pathway geactiveerd
worden. Deze controleert veel inflammatoire genen en p100 synthese. Ook na binding op
LTβReceptor kan deze laatste pathway geactiveerd worden, maar daarnaast ook de NF-κB2
pathway via processing van p100. Deze leidt tot transcriptie van genen die belangrijk zijn in
de ontwikkeling (figuur 3) (Ware, 2005).
2.4.4 Functie van de LTβReceptor-signaalweg
Gezien LTα1β2 en LIGHT binden op de LTβReceptor, en zo de signaalweg starten, kan men
via KO-muizen met een deficiëntie voor LTα , LTβ, LTβReceptor of LIGHT de functies van
deze signaalweg afleiden. In dit verband werden al heel wat studies verricht.
Een eerste studie toonde een rol aan van de LTβReceptor-signaalweg in het ontstaan van
auto-antilichamen via stoornissen in de centrale tolerantie (Martins et al., 2008). Dit is een
proces dat optreedt tijdens de ontwikkeling, waarbij autoreactieve T-cellen in de thymus
geëlimineerd worden wanneer ze een antigeen, dat door een HLA molecule wordt
gepresenteerd, herkennen. De medullaire epitheelcellen in de thymus brengen immers perifere
weefselantigenen ectopisch tot expressie, en zo wordt hier de tolerantie tegen deze self-
antigenen geïnduceerd. In deze studie onderzocht men LTβReceptor-deficiënte muizen, en
deze vertoonden een aberrante distributie van medullaire epitheelcellen in de thymus,
waardoor hun 3-dimensionele structuur verstoord is, met als gevolg een gestoorde interactie
tussen deze epitheelcellen en de T-cellen. Bijgevolg zouden dus minder autoreactieve T-
cellen verwijderd worden via negatieve selectie, en op die manier zou auto-immuniteit kunnen
onstaan.
Ook Boehm et al. (2003) toonden de rol van de pathway aan in het ontstaan van auto-
immuniteit, waarbij er antilichamen tegen speekselklier, pancreas en maag werden
aangetoond in LTβReceptor-/- muizen en niet in hun controles.
Ook werden eerder al ANA en anti-ENA (extractable nuclear antigen) antilichamen
aangetoond in muizen deficiënt voor LTα en LTβ (Vanassche, 2006). Bij de LTα-/- muizen
ging het enerzijds om antilichamen tegen dsDNA, en anderzijds tegen ENA zoals anti-Sm-B
en anti-RNP-A. Hierbij dient echter wel te worden vermeld dat bij deze dieren niet enkel de
LTβReceptor-pathway gedeeltelijk geblokkeerd wordt door afwezigheid van binding LTα1β2
van op de LTβReceptor, maar ook de TNFReceptor-signaalweg door afwezigheid van binding

Inleiding ___________________________________________________________________________
17
van LTα3 op de 2 TNFR’s. Bij de LTβ-/- muizen ging het enkel om anti-ENA antilichamen,
namelijk anti-RNP, anti-Cenp-B en anti-TopoI/Scl70. In dit werk concludeerde men bijgevolg
dat blokkade van de TNF-pathway voornamelijk verantwoordelijk zou zijn voor het ontstaan
van anti-DNA antilichamen, terwijl blokkade van de LTβReceptor-pathway daarentegen
voornamelijk anti-ENA antilichamen zou doen ontstaan.
Verder is deze pathway belangrijk in de ontwikkeling van de lymfeknopen. In verschillende
studies bij muizen deficiënt voor LTα, LTβ of LTβReceptor, zag men dat de lymfeknopen bij
deze dieren volledig ontbraken, met uitzondering van de LTβ-/- dieren, waar toch nog de
meeste mesenterische lymfeknopen aanwezig waren (Ware, 2005).
Daarnaast zou de LTβR-signaalweg ook een rol spelen in het ontstaan en behoud van
secundaire lymfoïde structuren. Hun unieke architectuur is cruciaal in de ontmoeting en
herkenning van antigenen. Zo zorgt deze pathway voor de ontwikkeling van Peyerse platen en
voor de juiste ontwikkeling en de structuur van de milt en zijn B-cel follikels (Fu and
Chaplin, 1999; Futterer et al., 1998).
Vervolgens speelt ze ook een rol in de ontwikkeling van NK-cellen (Iizuka et al., 1999) en
export van iNKT cellen vanuit de thymus (Franki, 2006).

Inleiding ___________________________________________________________________________
18
2.5
2.5.1 Validatie LIA
Doelstelling en benadering
Een eerste doel van deze thesis was de validatie van een multiparameter test (ANA INNO-
LIA Update) voor anti-ENA antilichamen in een aantal muizenmodellen voor lupus en
systeemsclerose. Eerder werd al aangetoond dat het hier om een veelbelovende methode kon
gaan, door overeenkomst aan te tonen tussen de resultaten bekomen met IIF op Hep-2000 en
deze bekomen met LIA in LTα-/-, LTβ-/- en BWF1-muizen (Vanassche, 2006). In dit werk
werd dit verder uitgewerkt.
De gouden standaard om een diagnostische test te valideren is een aantal zieke en gezonde
muizen testen om zo de sensitiviteit en specificiteit te berekenen van de nieuwe test. Daarom
werden een aantal gekende diermodellen gebruikt als positieve controle, namelijk MRL/lpr,
BWF1 en pristane-induced lupus muizen. Voor dit laatste model werden C57BL/6 muizen
éénmalig intraperitoneaal geïnjecteerd met pristane op de leeftijd van 8 weken. Als
controlegroepen voor de BWF1 en MRL/lpr werden de C57BL/6 muizen gebruikt, en voor de
pristane-induced lupus muizen de C57BL/6 behandeld met PBS. Vervolgens werden de
bekomen resultaten vergeleken met hetgeen in de literatuur beschreven staat omtrent
prevalenties van auto-antilichamen in deze muizenmodellen.
2.5.2 Rol van de LTβReceptor-signaalweg in het ontstaan van anti-ENA
Naast het gebruik van bestaande muizenmodellen die spontaan anti-ENA antilichamen
ontwikkelen, werd hier ook gebruik gemaakt van muizenmodellen waarbij de LTβReceptor-
signaalweg selectief en op verschillende ogenblikken uitgeschakeld werd. Op die manier werd
enerzijds nagaan in welke levensfase deze pathway van belang is om auto-immuniteit te
vermijden, en anderzijds wat de rol is van de aanwezige structurele defecten in het ontstaan
van deze anti-ENA antilichamen. Daarnaast werd ook nagegaan of de gestoorde centrale
tolerantie zoals beschreven bij de LTβReceptor-/- muizen een rol speelt in het ontstaan van
deze antilichamen.
Om de rol van structurele afwijkingen na te gaan en de fase waarin de signaalweg van belang
is, werden de LTβReceptor-/- muizen gebruikt, waarbij de pathway al vanaf de embryonale
fase ontbreekt, en deze werden vergeleken met muizen waarbij pre- en postnataal op
verschillende tijdstippen een solubel LTβreceptor-Fc fusieproteïne werd toegediend. Dit

Inleiding ___________________________________________________________________________
19
eiwit blokkeert de signaalweg door binding op het membranaire LTα1β2 en op LIGHT,
waardoor er geen signaaloverdracht via LTβReceptor plaatsvindt. Het werkingsmechanisme is
te vergelijken met dat van de humane TNF remmer etanercept, die binding van solubel TNF
en LTα3 op de TNF receptoren verhindert. Bij de behandelde dieren werd op regelmatige
tijdstippen een bloedafname verricht, en het serum werd vervolgens onderzocht op de
aanwezigheid van anti-ENA antilichamen.
De rol van de centrale tolerantie werd nagegaan via thymus transplantatie experimenten,
de gouden standaard om een stoornis van de centrale immuuntolerantie aan te tonen (Chin et
al., 2006). Hierbij werd aan muizen die een thymectomie hadden ondergaan, een totale
lichaamsbestaling aan hoge dosis toegediend zodat alle beenmergcellen afstierven.
Vervolgens kregen deze nieuw beenmerg afkomstig van congene muizen toegediend, gevolgd
door een thymustransplantie met een thymus afkomstig van ofwel een LTβReceptor-/- ofwel
een C57BL/6 muis, nadat hieruit alle cellen vernietigd werden. Op die manier ontstonden dus
bij beide groepen nieuwe T-cellen uit vergelijkbaar congeen beenmerg en deze circuleerden in
dieren van dezelfde stam, met het enige verschil dat bij de ene groep deze T-cellen uitrijpten
in een normale thymus van een C57BL/6, en bij de andere groep in een thymus afkomstig van
een LTβReceptor-/-. Indien de LTβReceptor-signaalweg dus een rol zou spelen in de centrale
tolerantie voor ENA, verwacht men in de muizen met de LTβReceptor-/- thymus
autoantilichamen, en niet bij de dieren met de C57BL/6 thymus. Dit werd in dit werk
nagegaan via LIA.
2.5.3 Weefselspecifieke antilichamen
Aan de hand van de indirecte immuunfluorescentie (IIF) op LKS coupes werden de sera van
de LTβReceptor-/- muizen onderzocht. Het ging hier om de lever, maag en nier, die
geïncubeerd worden met FITC-gelabeld anti-muis polyreactief antilichaam (zie 2.3.2.). De
sera afkomstig van C57BL/6 muizen werden hier gebruikt als negatieve controle. Het doel van
het experiment was na te gaan of bij deze dieren naast anti-ENA antilichamen, die gericht zijn
tegen algemene bestanddelen van alle cellen, nog andere antilichamen aanwezig zijn, die
gericht zijn tegen een bepaald celtype. Hierbij krijgt men een meer volledig beeld over de
auto-immuniteit aanwezig in deze dieren.

Methodologie ___________________________________________________________________________
20
3 Methodologie
3.1
De spontane SLE muizenmodellen BWF1 en MRL/lpr werden hier gebruikt ter validatie
van de LIA.
Muizen
Voor het pristane-induced lupus model werden C57BL/6 muizen van 8 weken oud
éénmalig intraperitoneaal geïnjecteerd met 500 µl pristane (2,6,10,14-
tetramethylpentadecaan, Sigma) (Satoh et al.,1995). Als controle werden C57BL/6 muizen
van 8 weken oud éénmalig geïnjecteerd met 500 µl PBS (Phosphate Buffered Saline).
Daarnaast werd ook een KO-muizenmodel voor SLE gebruikt, namelijk de LTβReceptor-/-
muizen. Als negatieve controle werden hierbij de C57BL/6 muizen gebruikt.
In dit werk werd ook gebruik gemaakt van muizen waarbij pre- en postnataal op
verschillende tijdstippen een solubel LTβReceptor-Fc fusieproteïne werd toegediend, in
dit werk gebeurde dit volgens 2 verschillende injectieschema’s. Bij een eerste schema
gebeurde deze toediening (100 µg) op dag 11 en 15 van de zwangerschap (schema D11,15),
bij een tweede op dag 18 (100 µg) van de zwangerschap en vervolgens na de geboorte
wekelijks (25 µg) tot de leeftijd van 6 weken (schema D18+verlengd 6w). Deze schema’s
werden bepaald op basis van wat in de literatuur beschreven staat over de ontwikkeling van
specifieke structuren op elk van deze tijdstippen in en na de zwangerschap. Als controle
werden hier C57BL/6 muizen gebruikt, die op dezelfde leeftijd dezelfde hoeveelheid
Gammagard® toegediend kregen, een mengsel van humane immunoglobulines,
Tenslotte werden hier ook thymustransplanten getest. Hiervoor werd bij 6 pasgeboren
C57BL/6 en 6 pasgeboren LTβReceptor-/- muizen de thymus verwijderd. Vervolgens werden
deze thymi in een medium gebracht bestaande uit 9 volumes cDMEM (Invitrogen) en 1
volume verdund 2-deoxyguanosine aan een concentratie van 0,0361 g 2-deoxyguanosine
(Sigma-Aldrich) in 10 ml PBS, waardoor alle aanwezige T-cellen gedood werden. Na 1
week werden deze 12 thymi getransplanteerd onder het nierkapsel in 12 C57BL/6
mannelijke muizen, die een thymectomie hadden ondergaan op 6 weken (Harlan Europe)
gevolgd door een totale lichaamsbestraling (10 Gy) en beenmergtransplantatie met
beenmerg afkomstig uit 30 congene muizen. Een bestraling aan deze dosis zorgde ervoor
dat alle beenmergcellen gedood werden. Op die manier werden dus 12 thymustransplanten
verkregen, waarvan 6 de thymus hadden afkomstig van de LTβReceptor-/- en 6 deze van de
C57BL/6 muizen, en waarbij alle muizen beenmerg hadden afkomstig van congene muizen.

Methodologie ___________________________________________________________________________
21
Als controle voor de bepaling van IgG levels werden 10 mannelijke C57BL/6 muizen van
12 weken oud gekocht (Harlan Europe), die in IVC (individual ventilated cages) waren
opgegroeid. De IgG levels van muizen zijn immers sterk afhankelijk van hun omgeving.
De muizen werden gehuisvest in het Centraal Animalarium van de faculteit Geneeskunde en
Gezondheidswetenschappen onder pathogeen-vrije condities. Ze werden behandeld zoals het
ethisch comité het voorschrijft.
3.2
De serumstalen van de verschillende muizen werden verkregen door na vaste tijdsintervallen
een bloedafname uit te voeren bij alle hierboven beschreven dieren. Deze bloedafname werd
uitgevoerd via de staartsnede. Afhankelijk van de leeftijd van de muizen werd 100µl tot
maximaal 200µl bloed afgenomen. Dit bloed werd gecentrifugeerd met een microcentrifuge
op 8000 rpm bij 4°C gedurende 5 min. Vervolgens werd het serum afgenomen, gealiquoteerd
en ingevroren.
Serumstalen
3.3
IgG titers werden bepaald in de sera van de thymustransplanten met ELISA volgens de
instructies van de producent (Bethyl Laboratories).
ELISA
3.4 LIA (Line ImmunoAssay)
Principe
Zie 2.3.5.a: ‘Principe van LIA’
Aan iedere cup werd 2 ml sample diluent en 10 μl staal toegevoegd, en het geheel werd
gemengd gedurende enkele minuten. Vervolgens werden de teststrips in de cupjes gebracht en
ondergedompeld. Na een incubatietijd van 1 uur bij kamertemperatuur op een shaker, werden
de strips 3 keer gewassen met 2 ml wasbuffer (1/5 verdunning van wasoplossing uit de kit met
gedestilleerd water) voor 3 min. Vervolgens werd aan elke cup 2 ml conjugaatbuffer
toegevoegd. Deze bestaat uit het anti-muis polyreactief antilichaam gelabeld met alkaline
fosfatase (Chemicon International, Temecula, CA) dat 5000x verdund werd met
Uitvoering

Methodologie ___________________________________________________________________________
22
conjugaatdiluent uit de kit. (Deze verdunning werd bepaald aan de hand van een
verdunningsreeks met verdunningen 1/5000, 1/25.000 en 1/50.000; 1/5000 was voldoende
sensitief en zorgde ook niet voor een te hoge achtergrondkleuring). Na een incubatietijd van
30 min bij kamertemperatuur op een shaker, werd elke strip weer 2 keer gewassen met
wasbuffer voor 3 min, en vervolgens een 3e keer met substraatbuffer uit de kit voor 3 min.
Hierna werd aan elke cup 2 ml substraatoplossing toegevoegd (1/100 verdunning van
substraatconcentraat uit de kit met substraatbuffer uit de kit) en dit werd gedurende 30 min
geïncubeerd. Uiteindelijk werd de vloeistof geaspireerd, en aan elke cup werd 2 ml
stopoplossing uit de kit toegevoegd om de kleurontwikkeling te stoppen. Na een incubatietijd
van 20 min op de shaker, werd elke strip nog 1 maal gewassen met 2 ml gedestilleerd water
voor 3 min, alle vloeistof werd vervolgens geaspireerd en de strips werden aan de lucht
gedroogd en afgedekt met papier gedurende enkele uren. Tenslotte werden de strips uit de
cupjes gehaald, en de resultaten werden afgelezen en vergeleken met de nieuw gedefinieerde
cut-off (zie 4.1.1).
3.5 Indirecte immuunfluorescentie op LKS-coupes
Principe
Voor de detectie van weefselspecifieke antilichamen, worden lever-, nier- en maagweefsel
afkomstig van ratten gecoat op een glazen plaatje, gebruikt als antigen-substraat. Als het staal
positief is, gaan specifieke antilichamen aanwezig in het verdunde staal reageren met de
antigenen aanwezig in het substraat. Vervolgens worden de gebonden antilichamen gekleurd
met FITC-gelabeld anti-muis antilichamen en gevisualiseerd met een fluorescentie-
microscoop.
Voor het bepalen van deze weefselspecifieke antilichamen werd gebruik gemaakt van slides
voorgecoat met lever-, nier- en maagweefsel afkomstig van ratten. Deze kits werden
verkregen bij de firma The binding site Ltd, Birningham. Alle stalen werden 1/20 verdund, en
per well werd 50 µl verdund staal toegevoegd. Na 30 min incubatietijd op kamertemperatuur
werden de slides 10 min gespoeld met PBS om de niet-specifieke bindingen te verwijderen,
en vervolgens 30 min geïncubeerd met FITC-gelabeld geit anti-muis IgG specifiek
antilichaam (BD PharmingenTM), in een verdunning van 1/100. Vervolgens werden de slides 5
min gespoeld in PBS, en daarna 3 min gespoeld in PBS met Evans Blue als tegenkleuring. Na
Uitvoering

Methodologie ___________________________________________________________________________
23
het spoelen van de slides in PBS om de overtollige counterstain weg te wassen, werden de
plaatjes gemonteerd. De gevormde complexen werden gevisualiseerd met een fluorescentie-
microscoop. De precieze lokalisatie van de fluorescentie ter hoogte van de specifieke
weefsels, maken het de gevonden antilichamen te identificeren.
3.6
Voor elke onderzochte continue variabele werd nagegaan of de verdeling al
dan niet normaal was. De symmetrie werd beoordeeld door een box-and-whiskers
plot en de standaard descriptieve variabelen van elke distributie (gemiddelde, 5% trimmed
gemiddelde, mediaan, kurtosis en scheefheid). Een Kolmogorov-Smirnov test werd
uitgevoerd en de Q-Q plot beoordeeld. Bij een normaalverdeling werd gebruik gemaakt van
de Student T-test, bij niet normale verdeling van de Mann Whitney U test. p-waarden kleiner
dan 0.05 werden als statistisch significant beschouwd.
Wanneer het ging om categorische variabelen, werd de Chi kwadraat test gebruikt. Ook hier
werden p-waarden kleiner dan 0.05 als statistisch significant beschouwd. Om de s.e.m.
(standard error of the mean) te bepalen, werd gebruik gemaakt van volgende formule:
Statistische analyse
s.e.m.=
waarbij p hier de proportie positieve stalen voorstelt, en n het aantal onderzochte stalen.

Resultaten ___________________________________________________________________________
24
4 Resultaten
4.1
Om de multiparameter test ANA INNO-LIA Update te valideren voor gebruik bij muizensera,
werden een aantal zieke muizen getest: de SLE muizenmodellen MRL/lpr, BWF1 en pristane-
induced lupus. Deze werden vergeleken met sera van gezonde dieren, de C57BL/6 muizen. Er
werden foto’s genomen van de LIA’s, die terug te vinden zijn in de bijlage (foto’s 1 - 4).
Validatie van de LIA
4.1.1 C57BL/6 muizen
Zoals eerder vermeld (zie 2.3.5.c) kan de humane cut-off strook niet gebruikt worden bij de
interpretatie van muizenstalen. Kwantificatie van de intensiteit van de bekomen
kleurverandering gebeurde bijgevolg via een standaard strook met 6 verschillende
kleurintensiteiten, bekomen via uittitreren van een positief serumstaal met toenemende
verdunning van het conjugaat (1/12800, 1/6400, 1/3200, 1/1600, 1/800 en 1/200). Vervolgens
werden de C57BL/6 muizenstammen getest via de LIA. Er werden 15 muizen getest op een
leeftijd van 24 weken, 8 vrouwelijke en 7 mannelijke dieren, en allen bleken een resultaat te
geven met lagere intensiteit dan deze op de standaardstrook overeenkomend met de grootste
verdunning (1/12800). Gezien deze C57BL/6 muizenstammen in de literatuur als ‘lupus-
resistente stammen’ beschreven worden (Liu and Mohan, 2006), werden deze resultaten als
negatief beschouwd. Bijgevolg werd een testresultaat als positief beschouwd zodra op de
strookjes een kleurverandering te onderscheiden was met minstens dezelfde intensiteit als de
laagste intensiteit van de standaardstrook (verdunning 1/12800). (zie ook foto 1 in bijlage).
4.1.2 MRL/lpr muizen
Een eerste muizenmodel dat bestudeerd werd zijn de MRL/lpr muizen. Hiervoor werden sera
afkomstig van muizen op verschillende leeftijden onderzocht. Het aantal stalen per leeftijd
wordt in onderstaande tabel weergegeven (tabel 3).
Leeftijd 6 weken 8 weken 12 weken 16 weken 20 weken 24 weken Aantal stalen 11 5 19 20 2 12
Tabel 3: Aantal onderzochte stalen per leeftijd bij de MRL/lpr muizen.
Resultaten ___________________________________________________________________________
25
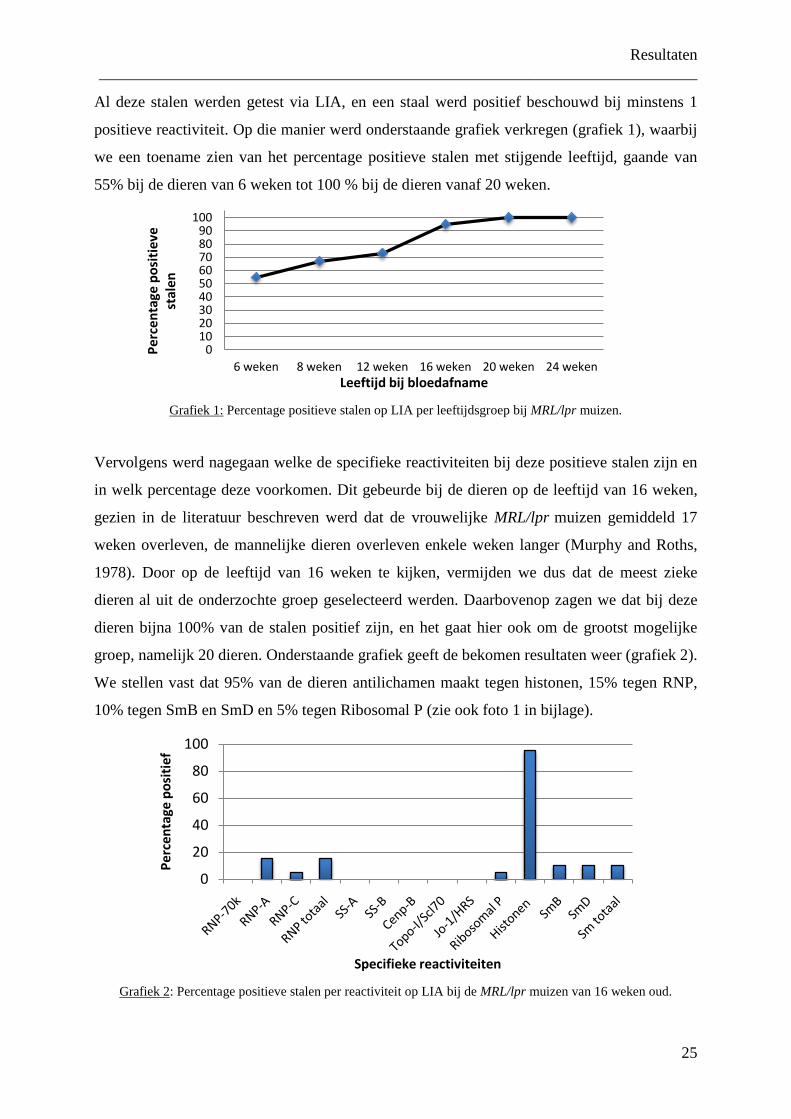
Al deze stalen werden getest via LIA, en een staal werd positief beschouwd bij minstens 1
positieve reactiviteit. Op die manier werd onderstaande grafiek verkregen (grafiek 1), waarbij
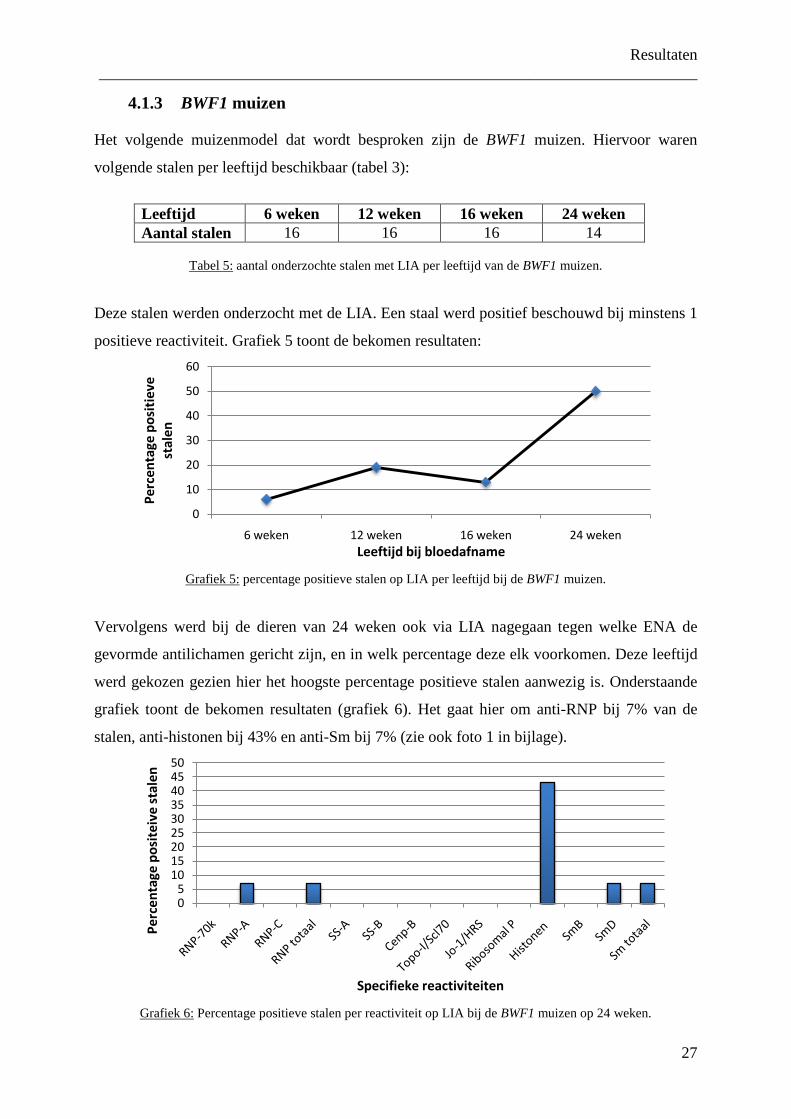
we een toename zien van het percentage positieve stalen met stijgende leeftijd, gaande van
55% bij de dieren van 6 weken tot 100 % bij de dieren vanaf 20 weken.
Grafiek 1:
Percentage positieve stalen op LIA per leeftijdsgroep bij MRL/lpr muizen.
Vervolgens werd nagegaan welke de specifieke reactiviteiten bij deze positieve stalen zijn en
in welk percentage deze voorkomen. Dit gebeurde bij de dieren op de leeftijd van 16 weken,
gezien in de literatuur beschreven werd dat de vrouwelijke MRL/lpr muizen gemiddeld 17
weken overleven, de mannelijke dieren overleven enkele weken langer (Murphy and Roths,
1978). Door op de leeftijd van 16 weken te kijken, vermijden we dus dat de meest zieke
dieren al uit de onderzochte groep geselecteerd werden. Daarbovenop zagen we dat bij deze
dieren bijna 100% van de stalen positief zijn, en het gaat hier ook om de grootst mogelijke
groep, namelijk 20 dieren. Onderstaande grafiek geeft de bekomen resultaten weer (grafiek 2).
We stellen vast dat 95% van de dieren antilichamen maakt tegen histonen, 15% tegen RNP,
10% tegen SmB en SmD en 5% tegen Ribosomal P (zie ook foto 1 in bijlage).
Grafiek 2
: Percentage positieve stalen per reactiviteit op LIA bij de MRL/lpr muizen van 16 weken oud.
0102030405060708090
100
6 weken 8 weken 12 weken 16 weken 20 weken 24 weken
Perc
enta
ge p
osit
ieve
st
alen
Leeftijd bij bloedafname
0
20
40
60
80
100
Perc
enta
ge p
osit
ief
Specifieke reactiviteiten
Resultaten ___________________________________________________________________________
26
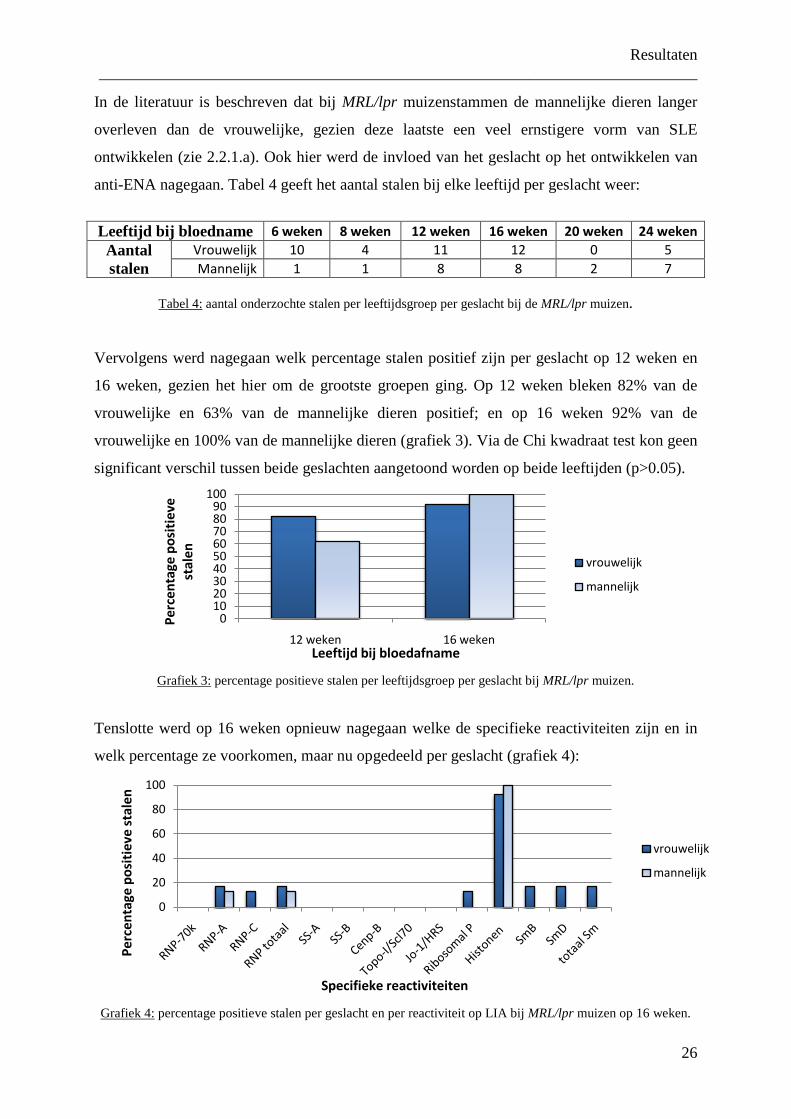
In de literatuur is beschreven dat bij MRL/lpr muizenstammen de mannelijke dieren langer
overleven dan de vrouwelijke, gezien deze laatste een veel ernstigere vorm van SLE
ontwikkelen (zie 2.2.1.a). Ook hier werd de invloed van het geslacht op het ontwikkelen van
anti-ENA nagegaan. Tabel 4 geeft het aantal stalen bij elke leeftijd per geslacht weer:
Leeftijd bij bloedname 6 weken 8 weken 12 weken 16 weken 20 weken 24 weken
Aantal stalen
Vrouwelijk 10 4 11 12 0 5 Mannelijk 1 1 8 8 2 7
Tabel 4:
aantal onderzochte stalen per leeftijdsgroep per geslacht bij de MRL/lpr muizen.
Vervolgens werd nagegaan welk percentage stalen positief zijn per geslacht op 12 weken en
16 weken, gezien het hier om de grootste groepen ging. Op 12 weken bleken 82% van de
vrouwelijke en 63% van de mannelijke dieren positief; en op 16 weken 92% van de
vrouwelijke en 100% van de mannelijke dieren (grafiek 3). Via de Chi kwadraat test kon geen
significant verschil tussen beide geslachten aangetoond worden op beide leeftijden (p>0.05).
Grafiek 3:
percentage positieve stalen per leeftijdsgroep per geslacht bij MRL/lpr muizen.
Tenslotte werd op 16 weken opnieuw nagegaan welke de specifieke reactiviteiten zijn en in
welk percentage ze voorkomen, maar nu opgedeeld per geslacht (grafiek 4):
Grafiek 4: percentage positieve stalen per geslacht en per reactiviteit op LIA bij MRL/lpr muizen op 16 weken.
0102030405060708090
100
12 weken 16 weken
Perc
enta
ge p
osit
ieve
st
alen
Leeftijd bij bloedafname
vrouwelijk
mannelijk
0
20
40
60
80
100
Perc
enta
ge p
osit
ieve
sta
len
Specifieke reactiviteiten
vrouwelijk
mannelijk
Resultaten ___________________________________________________________________________
27
4.1.3 BWF1 muizen
Het volgende muizenmodel dat wordt besproken zijn de BWF1 muizen. Hiervoor waren
volgende stalen per leeftijd beschikbaar (tabel 3):
Leeftijd 6 weken 12 weken 16 weken 24 weken Aantal stalen 16 16 16 14
Tabel 5:
aantal onderzochte stalen met LIA per leeftijd van de BWF1 muizen.
Deze stalen werden onderzocht met de LIA. Een staal werd positief beschouwd bij minstens 1
positieve reactiviteit. Grafiek 5 toont de bekomen resultaten:
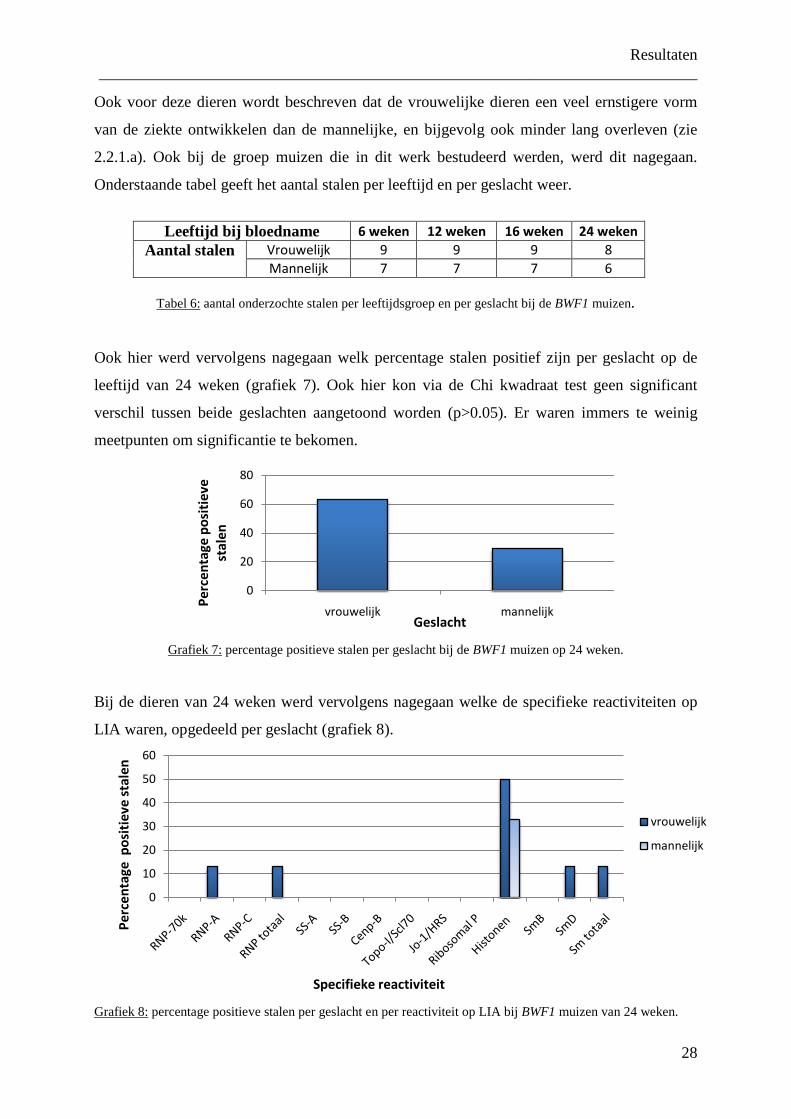
Grafiek 5:
percentage positieve stalen op LIA per leeftijd bij de BWF1 muizen.
Vervolgens werd bij de dieren van 24 weken ook via LIA nagegaan tegen welke ENA de
gevormde antilichamen gericht zijn, en in welk percentage deze elk voorkomen. Deze leeftijd
werd gekozen gezien hier het hoogste percentage positieve stalen aanwezig is. Onderstaande
grafiek toont de bekomen resultaten (grafiek 6). Het gaat hier om anti-RNP bij 7% van de
stalen, anti-histonen bij 43% en anti-Sm bij 7% (zie ook foto 1 in bijlage).
Grafiek 6: Percentage positieve stalen per reactiviteit op LIA bij de BWF1 muizen op 24 weken.
0
10
20
30
40
50
60
6 weken 12 weken 16 weken 24 weken
Perc
enta
ge p
osit
ieve
st
alen
Leeftijd bij bloedafname
05
101520253035404550
Perc
enta
ge p
osit
eive
sta
len
Specifieke reactiviteiten
Resultaten ___________________________________________________________________________
28
Ook voor deze dieren wordt beschreven dat de vrouwelijke dieren een veel ernstigere vorm
van de ziekte ontwikkelen dan de mannelijke, en bijgevolg ook minder lang overleven (zie
2.2.1.a). Ook bij de groep muizen die in dit werk bestudeerd werden, werd dit nagegaan.
Onderstaande tabel geeft het aantal stalen per leeftijd en per geslacht weer.
Leeftijd bij bloedname 6 weken 12 weken 16 weken 24 weken
Aantal stalen Vrouwelijk 9 9 9 8 Mannelijk 7 7 7 6
Tabel 6:
aantal onderzochte stalen per leeftijdsgroep en per geslacht bij de BWF1 muizen.
Ook hier werd vervolgens nagegaan welk percentage stalen positief zijn per geslacht op de
leeftijd van 24 weken (grafiek 7). Ook hier kon via de Chi kwadraat test geen significant
verschil tussen beide geslachten aangetoond worden (p>0.05). Er waren immers te weinig
meetpunten om significantie te bekomen.
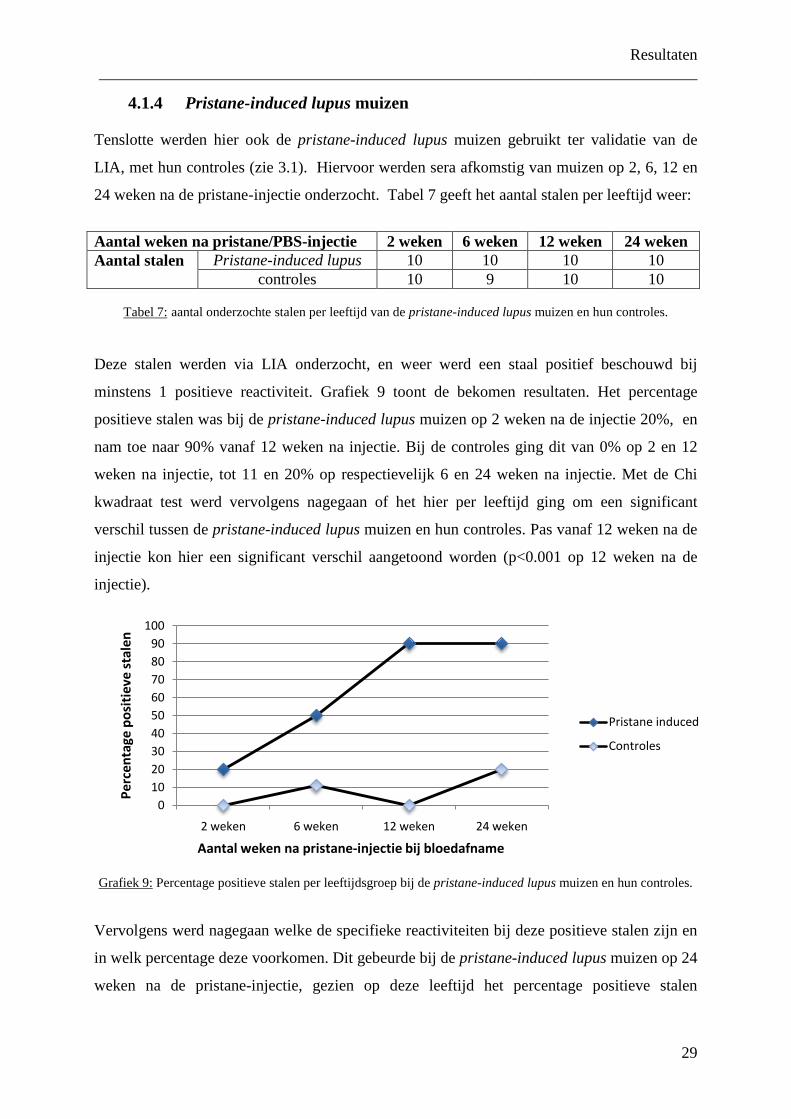
Grafiek 7:
percentage positieve stalen per geslacht bij de BWF1 muizen op 24 weken.
Bij de dieren van 24 weken werd vervolgens nagegaan welke de specifieke reactiviteiten op
LIA waren, opgedeeld per geslacht (grafiek 8).
Grafiek 8: percentage positieve stalen per geslacht en per reactiviteit op LIA bij BWF1 muizen van 24 weken.
0
20
40
60
80
vrouwelijk mannelijk
Perc
enta
ge p
osit
ieve
st
alen
Geslacht
0
10
20
30
40
50
60
Perc
enta
ge p
osit
ieve
sta
len
Specifieke reactiviteit
vrouwelijk
mannelijk
Resultaten ___________________________________________________________________________
29
4.1.4 Pristane-induced lupus muizen
Tenslotte werden hier ook de pristane-induced lupus muizen gebruikt ter validatie van de
LIA, met hun controles (zie 3.1). Hiervoor werden sera afkomstig van muizen op 2, 6, 12 en
24 weken na de pristane-injectie onderzocht. Tabel 7 geeft het aantal stalen per leeftijd weer:
Aantal weken na pristane/PBS-injectie 2 weken 6 weken 12 weken 24 weken Aantal stalen Pristane-induced lupus 10 10 10 10
controles 10 9 10 10
Tabel 7:
aantal onderzochte stalen per leeftijd van de pristane-induced lupus muizen en hun controles.
Deze stalen werden via LIA onderzocht, en weer werd een staal positief beschouwd bij
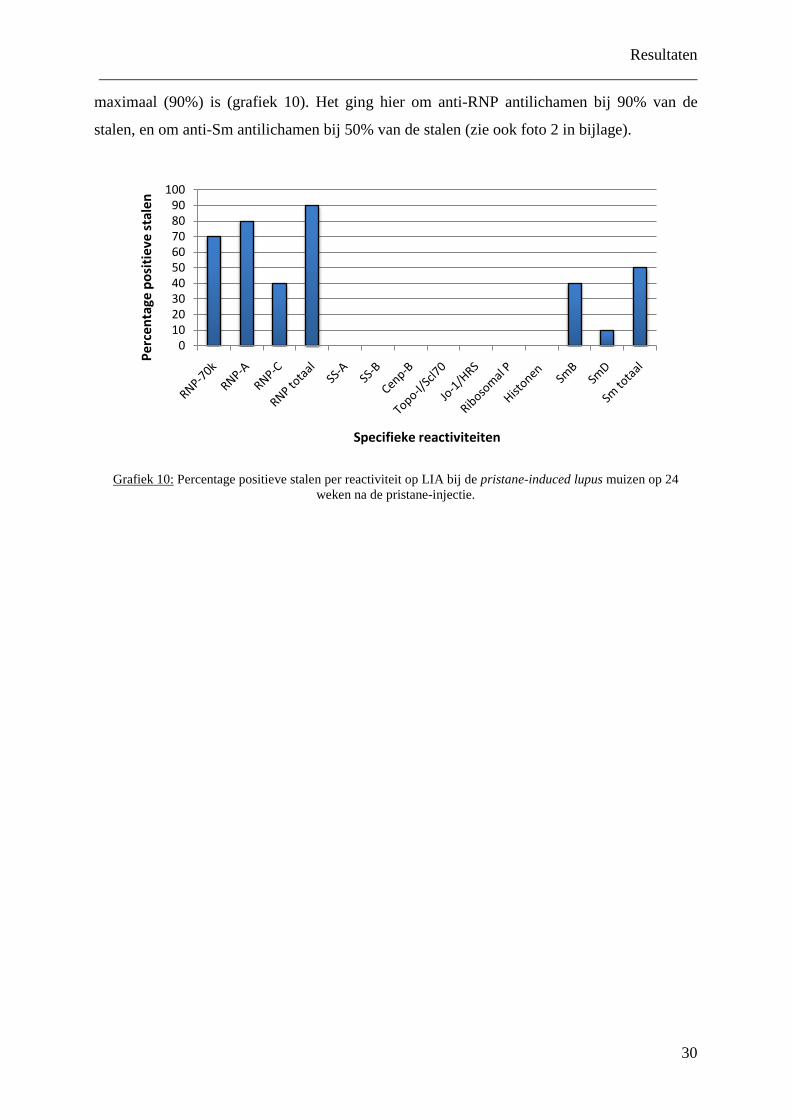
minstens 1 positieve reactiviteit. Grafiek 9 toont de bekomen resultaten. Het percentage
positieve stalen was bij de pristane-induced lupus muizen op 2 weken na de injectie 20%, en
nam toe naar 90% vanaf 12 weken na injectie. Bij de controles ging dit van 0% op 2 en 12
weken na injectie, tot 11 en 20% op respectievelijk 6 en 24 weken na injectie. Met de Chi
kwadraat test werd vervolgens nagegaan of het hier per leeftijd ging om een significant
verschil tussen de pristane-induced lupus muizen en hun controles. Pas vanaf 12 weken na de
injectie kon hier een significant verschil aangetoond worden (p<0.001 op 12 weken na de
injectie).
Grafiek 9:
Vervolgens werd nagegaan welke de specifieke reactiviteiten bij deze positieve stalen zijn en
in welk percentage deze voorkomen. Dit gebeurde bij de pristane-induced lupus muizen op 24
weken na de pristane-injectie, gezien op deze leeftijd het percentage positieve stalen
Percentage positieve stalen per leeftijdsgroep bij de pristane-induced lupus muizen en hun controles.
0102030405060708090
100
2 weken 6 weken 12 weken 24 weken
Perc
enta
ge p
osit
ieve
sta
len
Aantal weken na pristane-injectie bij bloedafname
Pristane induced
Controles
Resultaten ___________________________________________________________________________
30
maximaal (90%) is (grafiek 10). Het ging hier om anti-RNP antilichamen bij 90% van de
stalen, en om anti-Sm antilichamen bij 50% van de stalen (zie ook foto 2 in bijlage).
Grafiek 10: Percentage positieve stalen per reactiviteit op LIA bij de pristane-induced lupus muizen op 24
weken na de pristane-injectie.
0102030405060708090
100
Perc
enta
ge p
osit
ieve
sta
len
Specifieke reactiviteiten
Resultaten ___________________________________________________________________________
31
4.2
Zoals eerder beschreven (zie 2.5.2.), werden 3 muizengroepen onderzocht om de rol van deze
LTβReceptor-signaalweg in het ontstaan van auto-antilichamen na te gaan:
Rol van de LTβReceptor-signaalweg in het ontstaan van anti-ENA
4.2.1 LTβReceptor-/- muizen
Van deze stam werden 17 stalen op een leeftijd van 3 maand getest via de LIA, en 15 stalen
op een leeftijd van 6 maand. Deze werden vervolgens vergeleken met hun controles, de
C57BL/6 muizen, waarvan op 3 maand ook 17 stalen getest werden en op 6 maand ook 15
stalen. Onderstaande tabel toont de bekomen resultaten, met de overeenkomstige ‘standard
error of the mean’ (s.e.m.). Via de Chi kwadraat test werd nagegaan of de percentages
positieve stalen bij beide leeftijdsgroepen van LTβReceptor-/- muizen significant verschillen
van deze van hun controles, en dit bleek in beide groepen zo (p=0.033 op 3 maand, en
p=0.032 op 6 maand).
Stam Frequentie positieve
stalen op 3 maand (%) s.e.m. Frequentie positieve
stalen op 6 maand (%) s.e.m.
LTβReceptor-/- 23.5 10.3 26.7 11.4 C57BL/6 0 0 0 0
Tabel 8:
Percentage positieve stalen op LIA en s.e.m. bij de LTβReceptor-/- en C57BL/6 per leeftijdsgroep.
Vervolgens werd per groep en per leeftijd het percentage positieve stalen per reactiviteit
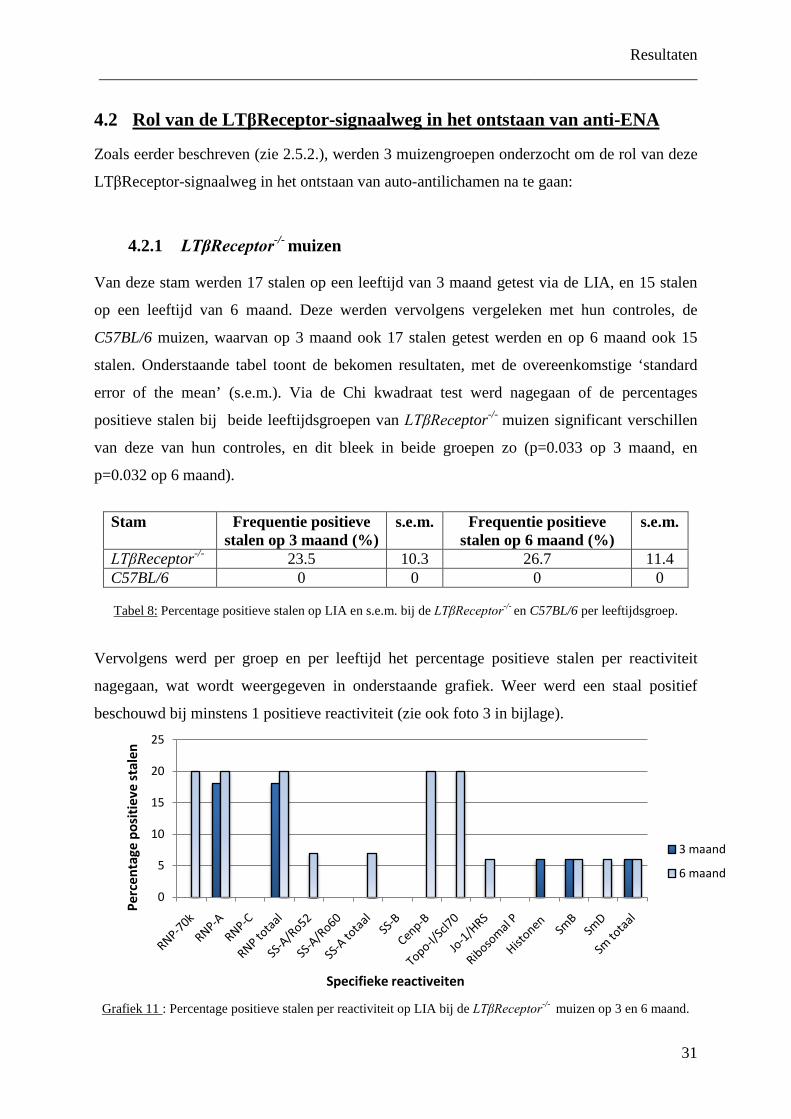
nagegaan, wat wordt weergegeven in onderstaande grafiek. Weer werd een staal positief
beschouwd bij minstens 1 positieve reactiviteit (zie ook foto 3 in bijlage).
Grafiek 11 : Percentage positieve stalen per reactiviteit op LIA bij de LTβReceptor-/- muizen op 3 en 6 maand.
0
5
10
15
20
25
Perc
enta
ge p
osit
ieve
sta
len
Specifieke reactiveiten
3 maand
6 maand
Resultaten ___________________________________________________________________________
32
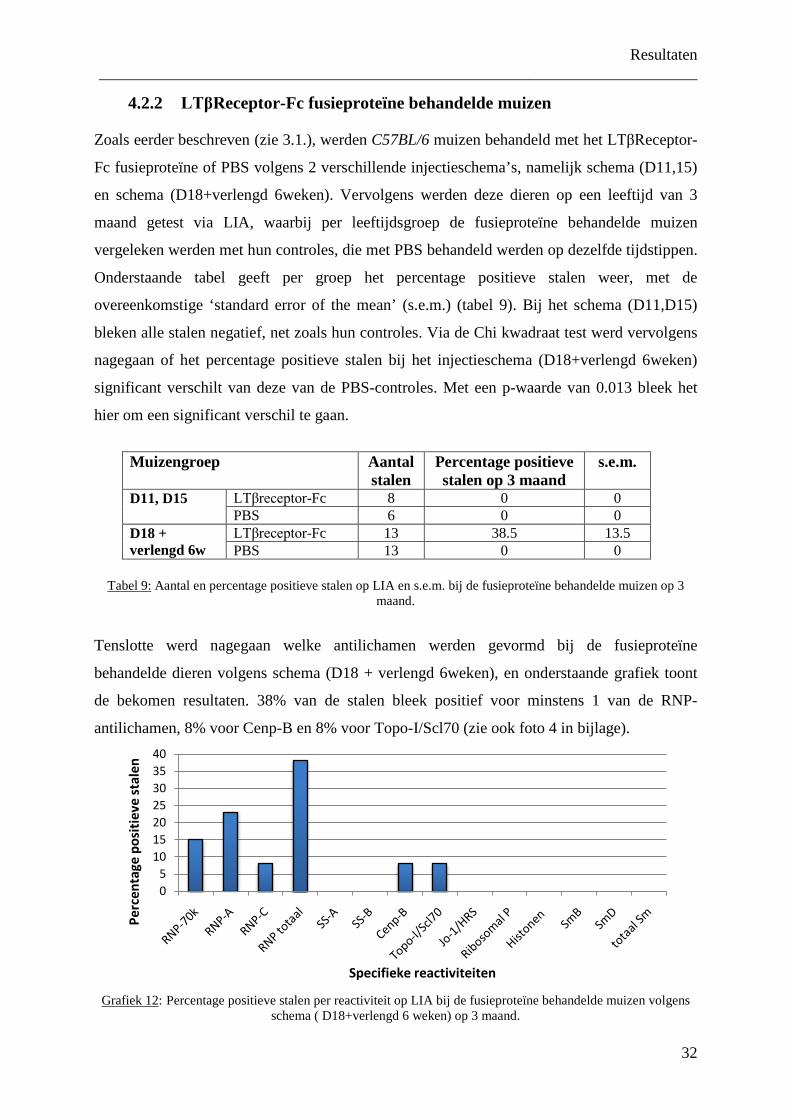
4.2.2 LTβReceptor-Fc fusieproteïne behandelde muizen
Zoals eerder beschreven (zie 3.1.), werden C57BL/6 muizen behandeld met het LTβReceptor-
Fc fusieproteïne of PBS volgens 2 verschillende injectieschema’s, namelijk schema (D11,15)
en schema (D18+verlengd 6weken). Vervolgens werden deze dieren op een leeftijd van 3
maand getest via LIA, waarbij per leeftijdsgroep de fusieproteïne behandelde muizen
vergeleken werden met hun controles, die met PBS behandeld werden op dezelfde tijdstippen.
Onderstaande tabel geeft per groep het percentage positieve stalen weer, met de
overeenkomstige ‘standard error of the mean’ (s.e.m.) (tabel 9). Bij het schema (D11,D15)
bleken alle stalen negatief, net zoals hun controles. Via de Chi kwadraat test werd vervolgens
nagegaan of het percentage positieve stalen bij het injectieschema (D18+verlengd 6weken)
significant verschilt van deze van de PBS-controles. Met een p-waarde van 0.013 bleek het
hier om een significant verschil te gaan.
Muizengroep Aantal
stalen Percentage positieve stalen op 3 maand
s.e.m.
D11, D15 LTβreceptor-Fc 8 0 0 PBS 6 0 0
D18 + verlengd 6w
LTβreceptor-Fc 13 38.5 13.5 PBS 13 0 0
Tabel 9:
Aantal en percentage positieve stalen op LIA en s.e.m. bij de fusieproteïne behandelde muizen op 3 maand.
Tenslotte werd nagegaan welke antilichamen werden gevormd bij de fusieproteïne
behandelde dieren volgens schema (D18 + verlengd 6weken), en onderstaande grafiek toont
de bekomen resultaten. 38% van de stalen bleek positief voor minstens 1 van de RNP-
antilichamen, 8% voor Cenp-B en 8% voor Topo-I/Scl70 (zie ook foto 4 in bijlage).
Grafiek 12: Percentage positieve stalen per reactiviteit op LIA bij de fusieproteïne behandelde muizen volgens schema ( D18+verlengd 6 weken) op 3 maand.
05
10152025303540
Perc
enta
ge p
osit
ieve
sta
len
Specifieke reactiviteiten
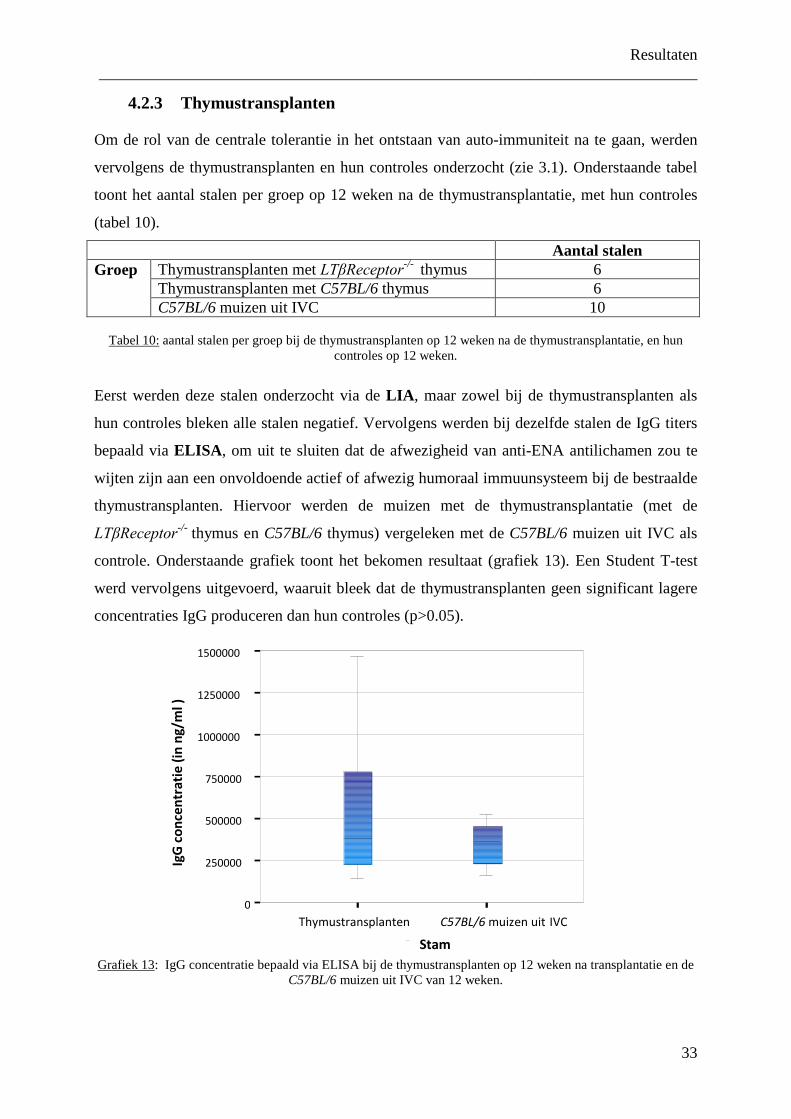
Resultaten ___________________________________________________________________________
33
4.2.3 Thymustransplanten
Om de rol van de centrale tolerantie in het ontstaan van auto-immuniteit na te gaan, werden
vervolgens de thymustransplanten en hun controles onderzocht (zie 3.1). Onderstaande tabel
toont het aantal stalen per groep op 12 weken na de thymustransplantatie, met hun controles
(tabel 10).
Aantal stalen Groep
Thymustransplanten met LTβReceptor-/- thymus 6 Thymustransplanten met C57BL/6 thymus 6 C57BL/6 muizen uit IVC 10
Tabel 10:
aantal stalen per groep bij de thymustransplanten op 12 weken na de thymustransplantatie, en hun controles op 12 weken.
Eerst werden deze stalen onderzocht via de LIA, maar zowel bij de thymustransplanten als
hun controles bleken alle stalen negatief. Vervolgens werden bij dezelfde stalen de IgG titers
bepaald via ELISA, om uit te sluiten dat de afwezigheid van anti-ENA antilichamen zou te
wijten zijn aan een onvoldoende actief of afwezig humoraal immuunsysteem bij de bestraalde
thymustransplanten. Hiervoor werden de muizen met de thymustransplantatie (met de
LTβReceptor-/- thymus en C57BL/6 thymus) vergeleken met de C57BL/6 muizen uit IVC als
controle. Onderstaande grafiek toont het bekomen resultaat (grafiek 13). Een Student T-test
werd vervolgens uitgevoerd, waaruit bleek dat de thymustransplanten geen significant lagere
concentraties IgG produceren dan hun controles (p>0.05).
Grafiek 13: IgG concentratie bepaald via ELISA bij de thymustransplanten op 12 weken na transplantatie en de C57BL/6 muizen uit IVC van 12 weken.
1,00 2,00 Stam
0
250000
500000
750000
1000000
1250000
1500000
Thymustransplanten C57BL/6 muizen uit IVC
Stam
IgG
con
cent
rati
e (in
ng/
ml )
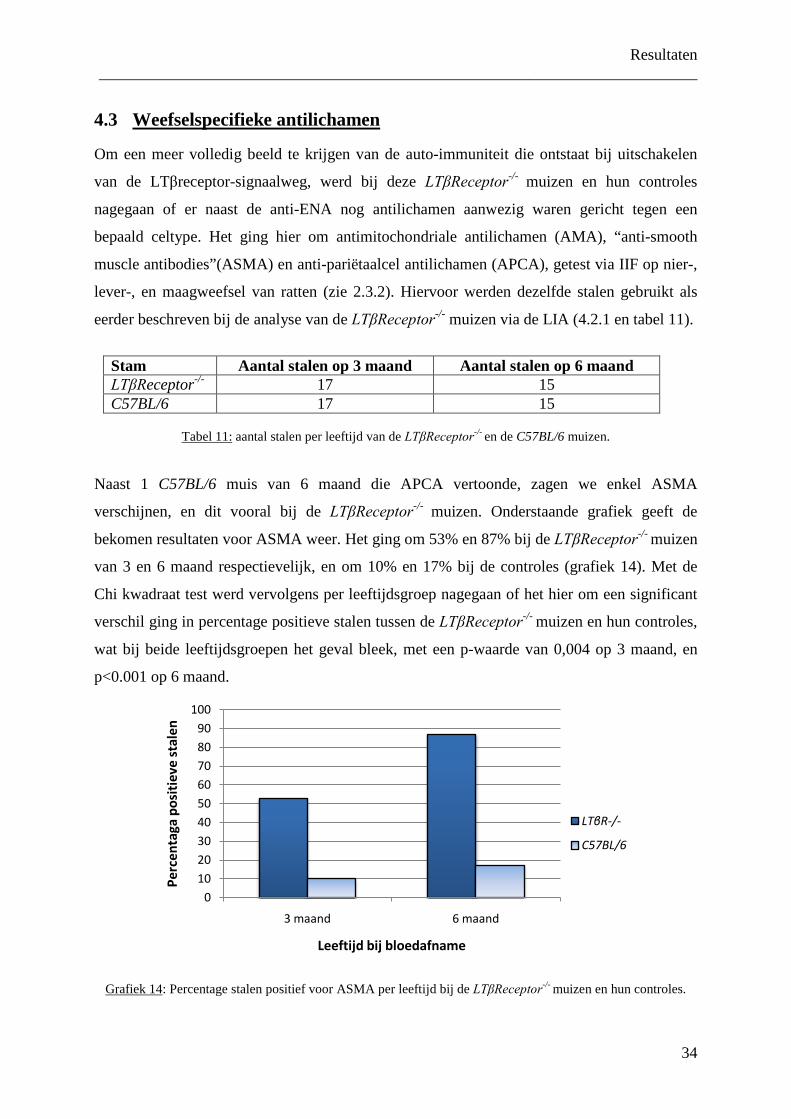
Resultaten ___________________________________________________________________________
34
4.3
Om een meer volledig beeld te krijgen van de auto-immuniteit die ontstaat bij uitschakelen
van de LTβreceptor-signaalweg, werd bij deze LTβReceptor-/- muizen en hun controles
nagegaan of er naast de anti-ENA nog antilichamen aanwezig waren gericht tegen een
bepaald celtype. Het ging hier om antimitochondriale antilichamen (AMA), “anti-smooth
muscle antibodies”(ASMA) en anti-pariëtaalcel antilichamen (APCA), getest via IIF op nier-,
lever-, en maagweefsel van ratten (zie 2.3.2). Hiervoor werden dezelfde stalen gebruikt als
eerder beschreven bij de analyse van de LTβReceptor-/- muizen via de LIA (4.2.1 en tabel 11).
Weefselspecifieke antilichamen
Stam Aantal stalen op 3 maand Aantal stalen op 6 maand LTβReceptor-/- 17 15 C57BL/6 17 15
Tabel 11:
aantal stalen per leeftijd van de LTβReceptor-/- en de C57BL/6 muizen.
Naast 1 C57BL/6 muis van 6 maand die APCA vertoonde, zagen we enkel ASMA
verschijnen, en dit vooral bij de LTβReceptor-/- muizen. Onderstaande grafiek geeft de
bekomen resultaten voor ASMA weer. Het ging om 53% en 87% bij de LTβReceptor-/- muizen
van 3 en 6 maand respectievelijk, en om 10% en 17% bij de controles (grafiek 14). Met de
Chi kwadraat test werd vervolgens per leeftijdsgroep nagegaan of het hier om een significant
verschil ging in percentage positieve stalen tussen de LTβReceptor-/- muizen en hun controles,
wat bij beide leeftijdsgroepen het geval bleek, met een p-waarde van 0,004 op 3 maand, en
p<0.001 op 6 maand.
Grafiek 14
: Percentage stalen positief voor ASMA per leeftijd bij de LTβReceptor-/- muizen en hun controles.
0102030405060708090
100
3 maand 6 maand
Perc
enta
ga p
osit
ieve
sta
len
Leeftijd bij bloedafname
LTβR-/-
C57BL/6
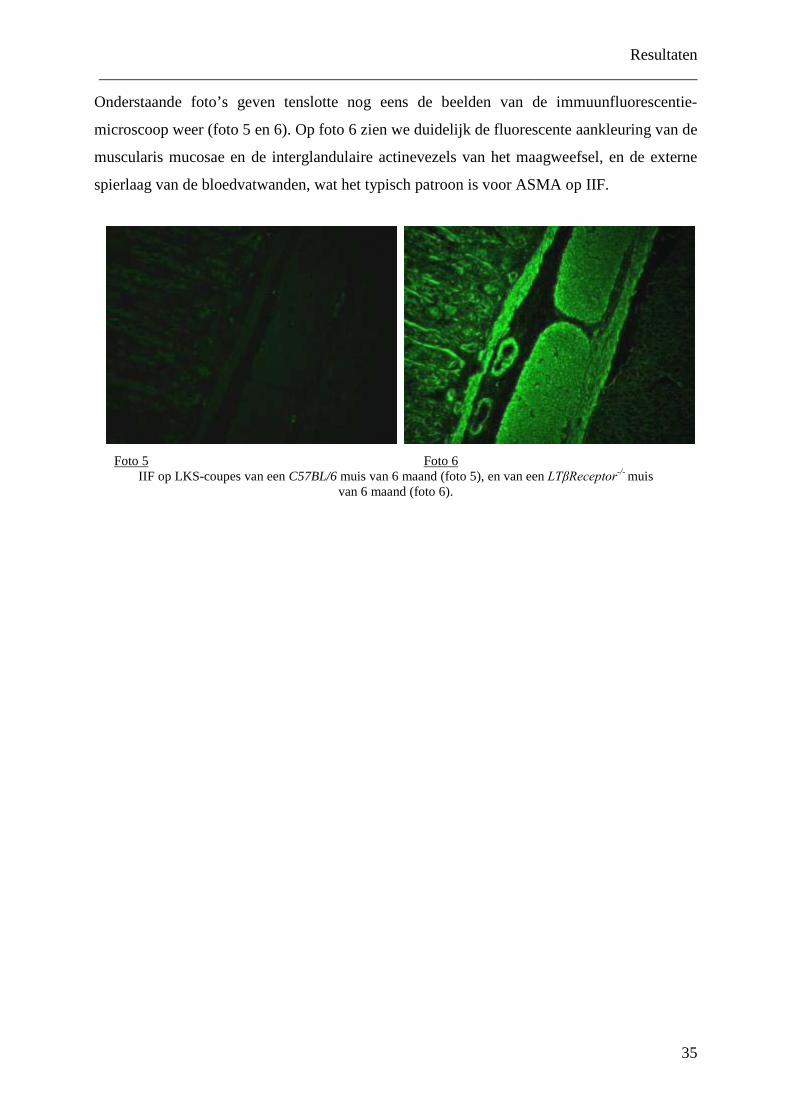
Resultaten ___________________________________________________________________________
35
Onderstaande foto’s geven tenslotte nog eens de beelden van de immuunfluorescentie-
microscoop weer (foto 5 en 6). Op foto 6 zien we duidelijk de fluorescente aankleuring van de
muscularis mucosae en de interglandulaire actinevezels van het maagweefsel, en de externe
spierlaag van de bloedvatwanden, wat het typisch patroon is voor ASMA op IIF.
Foto 5
Foto 6 IIF op LKS-coupes van een C57BL/6 muis van 6 maand (foto 5), en van een LTβReceptor-/- muis
van 6 maand (foto 6).

Discussie ___________________________________________________________________________
36
5 Discussie
5.1
Een eerste doel van dit werk was de validatie van de ANA INNO-LIA Update (kortweg LIA)
voor anti-ENA in een aantal muizenmodellen voor SLE en SSc. Vanassche (2006) toonde
eerder al aan dat het hier om een veelbelovende techniek kon gaan, maar dit diende nog
verder uitgewerkt te worden. Hiervoor werden in dit werk een aantal muizenmodellen voor
SLE (MRL/lpr, BWF1 en pristane-induced lupus) en gezonde muizen (C57BL/6) getest via
LIA. Vervolgens werden de bekomen resultaten vergeleken met hetgeen in de literatuur
beschreven staat omtrent prevalenties van autoantilichamen in deze muizenmodellen.
Validatie van de LIA
5.1.1 MRL/lpr muizen
In de literatuur werden bij deze dieren antilichamen tegen Ribosomal P, RNP, Sm en
histonen beschreven, wat overeenkomt met de resultaten van onze groep muizen onderzocht
met de LIA (zie 4.1.2). Daarnaast werd 1 artikel gevonden dat anti-SS-A antilichamen
beschreef, zowel tegen SS-A/Ro52 als tegen SS-A/Ro60 via western blotting en ELISA
(Wahren et al., 1994). Dit kan mogelijks verklaard worden door een lagere sensitiviteit van de
LIA voor anti-SS-A antilichamen, maar dit dient in de toekomst verder onderzocht te worden.
Vervolgens werden de frequenties van de antilichamen, die in dit werk beschreven werden
voor de 20 muizen van 16 weken, vergeleken met de frequenties die in de literatuur
beschreven werden voor vergelijkbare leeftijden. Met de LIA werd in 5% van de stalen anti-
ribosomal P aangetoond, terwijl in de studie van Bonfa et al. (1988) 11% van de stalen via
ELISA en immunoblotting positief bleken, maar het ging hier om iets oudere leeftijden (5 à 7
maand). Laderach et al. (2002) toonden met radioligand studies bij 50% van de 12 MRL/lpr
muizen anti-RNP antilichamen aan, terwijl dit via de LIA 15% was. Ook hier waren de
muizen enkele weken ouder (17-20 weken) dan deze onderzocht via LIA, en het ging hier
slechts om een kleine groep (12 muizen), wat het verschil kan verklaren. Via LIA werden
vervolgens 10% van de onderzochte stalen positief bevonden voor anti-Sm, waarvan 17% bij
de vrouwelijke dieren. Eisenberg et al. (1978) vond anti-Sm antilichamen bij 10% van de
vrouwelijke dieren van 16 weken met immunodiffusie. Ook voor anti-SmB waren de
resulaten gelijkaardig: via LIA waren 17% vrouwelijke dieren positief, terwijl Laderach et al.
(2003) 30% anti-SmB aantoonden bij de vrouwelijke dieren van 17-20 weken via radioligand

Discussie ___________________________________________________________________________
37
studies. Tenslotte werden via LIA in 95% van de geteste stalen anti-histon antilichamen
aangetoond. In de literatuur werden geen percentages gevonden omtrent het voorkomen van
antilichamen tegen alle histonproteïnen, maar wel tegen aparte eiwitten hiervan. Bijgevolg
kunnen we deze data helaas niet volledig vergelijken met onze data. Costa en Monier (1986)
onderzochten via immunoblotting welk percentage positief was voor de histon-eiwitten H1-
H4, en vond hierbij percentages van 47% tot 71%, afhankelijk van welk eiwit onderzocht
werd. Dit gebeurde bij muizen van 3 tot 5 maand.
5.1.2 BWF1 muizen
Bij deze dieren werden in de literatuur enkel anti-Sm, anti-RNP en anti-histonen
antilichamen beschreven, wat weer overeenkomt met hetgeen gevonden werd met de LIA. In
dit werk werden met de LIA de specifieke reactiviteiten bij de 14 stalen van BWF1 muizen
van 24 weken onderzocht (zie 4.1.3).
Met de LIA werden hier bij 7% van de dieren anti-Sm antilichamen aangetoond, terwijl
Eisenberg et al. (1978) met immunodiffusie bij jongere muizen van 1-17 weken bij 3% anti-
Sm kon aantonen. Laderach et al. (2003) beschreven vervolgens met radioligand studies bij
10% van hun 40-55 weken oude dieren anti-RNP antilichamen, terwijl deze met de LIA bij
7% van onze dieren kon aangetoond worden. Tenslotte werden in dit werk bij 43% van de
muizen anti-histon antilichamen aangetoond. Gioud et al. (1983) toonde bij dieren van
dezelfde leeftijd bij 55% van de dieren deze antilichamen aan.
5.1.3 Pristane-induced lupus muizen
Tenslotte werd in de literatuur nagegaan welke antilichamen de pristane-induced lupus
muizen produceren, en het bleek enkel te gaan om anti-Sm en anti-RNP antilichamen, wat
overeenkomt met de resultaten bekomen via LIA. Bij de 10 met pristane behandelde C57BL/6
muizen op 24 weken na de pristane-injectie bleek 90% van de stalen positief voor anti-RNP
en 50% voor anti-Sm (zie 4.1.4.). Savarese et al. (2008) toonde bij 50% van de met pristane
behandelde C57BL/6 muizen antilichamen aan tegen Sm/RNP via ELISA, ook op 24 weken
na de pristane-injectie. Satoh et al.(1995) werkte met BALB/c muizen in plaats van C57BL/6,
een andere lupusresistente stam, en toonde met ELISA bij 64% van de dieren anti-Sm/RNP
antilichamen aan, ook op 6 maand na de pristane-injectie. Hier werden geen gegevens
gevonden omtrent de frequentie van het apart voorkomen van beide antilichamen.

Discussie ___________________________________________________________________________
38
5.1.4 Kinetiek van de anti-ENA antilichamen
Tenslotte werd van elk van deze muizenmodellen de kinetiek van de anti-ENA antilichamen
nagegaan, door het percentage positieve stalen in de tijd te volgen. Zowel bij de MRL/lpr,
BWF1 als de pristane-induced lupus muizen nam de prevalentie aan positieve stalen toe met
de leeftijd. Ook dit komt overeen met wat in de literatuur wordt beschreven voor deze
stammen. Gioud et al. (1983) toonden dit aan bij de BWF1 muizen, Eisenberg and Dixon
(1978) voor de MRL/lpr muizen, en tenslotte Satoh et al. (2000) voor de pristane-induced
muizen. Ook bij andere stammen werd dit fenomeen in de literatuur beschreven (Laderach et
al., 2003; Eisenberg and Dixon, 1978).
5.1.5 Besluit en toekomstperspectieven
We kunnen hier dus besluiten dat de resultaten bekomen met de LIA in grote lijnen
overeenkomen met wat beschreven staat in de literatuur voor deze muizenmodellen. De
afwijkingen die voorkomen kunnen in enkele gevallen verklaard worden door een verschil in
leeftijd, gezien oudere dieren in hogere percentages antilichamen produceren dan jongere.
Ook omvatten de onderzochte groepen muizen zowel in dit werk als deze beschreven in de
literatuur vaak slechts maximum 20 muizen, wat bijgevolg zorgt voor grote variaties.
De LIA ziet er dus een veelbelovende techniek uit voor gebruik bij muizenstalen, maar dit
dient in de toekomst nog verder uitgewerkt te worden. Hiervoor zullen ten eerste de stalen die
onderzocht werden met de LIA nog bijkomend onderzocht worden met western blotting,
waarna de bekomen resultaten zullen vergeleken worden met deze van de LIA. Daarnaast
dienen ook nog andere muizenmodellen onderzocht te worden, waarvan beschreven is dat ze
antilichamen produceren tegen SS-A, SS-B, Cenp-B, Topo-1/Scl70 en Jo-1/HRS. Op die
manier kan nagegaan worden bij alle antilichamen die in theorie aangetoond kunnen worden
met de LIA of ze ook effectief positief zijn op LIA. Van de Tsk-1 muizen is beschreven dat ze
anti-Topo-1/Scl70 antilichamen produceren (Jimenez and Christner, 2002), en bij de NOD-
muizen worden anti-SS-A (Skarstein et al., 1995) en anti-Sm antilichamen (Baxter et al.,
1994) teruggevonden. Bijgevolg zullen deze 2 stammen in de toekomst ook onderzocht
worden met LIA. Het zal waarschijnlijk niet mogelijk zijn alle antilichamen die gedetecteerd
kunnen worden met LIA op die manier te testen, gezien voor sommige antilichamen (anti-SS-
B, Cenp-B en Jo-1/HRS) geen muizenmodellen beschikbaar zijn.

Discussie ___________________________________________________________________________
39
5.2
In dit werk werd de rol van de LTβReceptor-signaalweg nagegaan in het ontstaan van auto-
immuniteit. Omtrent het ontstaan van auto-immuniteit in het algemeen zijn immers
verschillende mechanismen beschreven. Enerzijds kan een gestoorde centrale tolerantie
hiertoe leiden, en ook kunnen structurele afwijkingen in lymfoïde organen auto-immuniteit
induceren, naast nog vele andere mechanismen. In dit werk werd nagegaan of blokkade van
de LTβReceptor-signaalweg leidt tot een gestoorde centrale tolerantie of tot structurele
afwijkingen, en of deze op hun beurt zorgen voor autoantilichamen.
LTβReceptor-signaalweg
5.2.1 Rol centrale tolerantie via thymus transplantatie experimenten
Met de thymus transplantatie experimenten werd de rol van de centrale tolerantie
nagegaan. Met de LIA konden noch bij de muizen met de LTβReceptor-/- thymus, noch bij
deze met de C57BL/6 thymus anti-ENA antilichamen aangetoond worden. Dit ondanks de
studie van Martins et al. (2008), die bij LTβR-deficiënte muizen een aberrante distributie van
medullaire epitheelcellen in de thymus aantoonden, met als gevolg een gestoorde interactie
tussen deze epitheelcellen en de T-cellen tijdens de negatieve selectie. Daarnaast werden met
IgG ELISA bij de thymustransplanten geen lagere IgG levels aangetoond dan bij hun
controles, wat bewijst dat deze afwezigheid van ENA in elk geval niet te wijten is aan een
verminderde of afwezige humorale immuniteit bij deze bestraalde transplanten. Hieruit
kunnen we dus besluiten dat een afwijking in de centrale tolerantie op zich niet voldoende is
voor het ontstaan van anti-ENA antilichamen.
Nochtans toonden Chin et al. (2006) in een gelijkaardig experiment antilichamen aan tegen
type II collageen en insuline in dezelfde soort muizen met dezelfde getransplanteerde thymi.
Een mogelijke verklaring hiervoor is dat het type antigen hier een rol zou spelen: zo zouden
perifere weefselantigenen zoals collageen en insuline enkel tot expressie gebracht worden in
de medullaire epitheelcellen van de thymus, waarbij de negatieve selectie dan auto-immuniteit
verhindert. Bij de nucleaire antigenen daarentegen zijn naast deze negatieve selectie
waarschijnlijk nog andere mechanismen aanwezig die auto-immuniteit onderdrukken.
Bijgevolg zou voor de perifere antigenen een abnormale thymus zorgen voor afwezigheid van
negatieve selectie, en zo voor het ontstaan van auto-antilichamen, terwijl dit bij de nucleaire
antigenen niet het geval is gezien deze tolerantie tegen eigen antigenen ook nog elders kan
geïnduceerd worden.

Discussie ___________________________________________________________________________
40
5.2.2 Rol structurele defecten en levensfase
Vervolgens werd aan de hand van LTβReceptor-/- muizen en muizen behandeld met het
solubel LTβReceptor-Fc fusieproteïne nagegaan wat de rol van structurele defecten is in het
ontstaan van auto-immuniteit, en in welke levensfase deze pathway van belang is om auto-
immuniteit te vermijden.
Eerst worden de de LTβReceptor-/- muizen besproken, waarbij zowel pre- als postnataal deze
LTβReceptor-signaalweg geblokkeerd is. Zoals eerder beschreven, ontbreken bij deze dieren
alle lymfeknopen en Peyerse platen, en is de miltarchitectuur ook grondig verstoord. In dit
werk konden bij 24% van deze dieren op 3 maand met LIA autoantilichamen aangetoond
worden, gericht tegen RNP, histonen en Sm; en op 6 maand zagen we antilichamen tegen
RNP, Cenp-B en Topo-1/Scl70, maar ook tegen Sm en Jo-1/HRS in lage percentages.
Om verder na te gaan welke van deze structurele defecten bijdragen tot het ontstaan van anti-
ENA antilichamen, en om na te gaan in welke levensfase de LTβReceptor-signaalweg vereist
is om auto-immuniteit te vermijden, werden vervolgens de muizen behandeld met het solubel
LTβReceptor-Fc fusieproteïne onderzocht.
Bij het eerste injectieschema waarbij het fusieproteïne op dag 11 en dag 15 prenataal werd
toegediend, kon men via LIA geen autoantilichamen aantonen. Rennert et al. (1999) voerden
dezelfde testen uit bij BALB/c muizen en kon bij muizen behandeld met het fusieproteïne op
dag 11 prenataal aantonen dat alle lymfeknopen afwezig waren, behalve de mesenterische.
Ook ontbraken de Peyerse platen bij deze dieren. Bij behandeling op dag 15 prenataal waren
naast de mesenterische, ook nog de brachiale en axillaire aanwezig. Ook hier waren de
Peyerse platen afwezig. Bij beide was de miltarchitectuur normaal. Dus kunnen we aannemen
dat bij de muizen in dit werk die op dag 11 en dag 15 prenataal behandeld werden met het
fusieproteïne geen Peyerse platen hebben en geen lymfeknopen, behalve de mesenterische. De
miltarchitectuur is bij deze muizen normaal. Gezien bij deze dieren via LIA geen
autoantilichamen konden aangetoond worden, ondanks de afwezigheid van lymfeknopen en
Peyerse platen, kunnen we dus besluiten dat deze aanwezige structurele defecten op zich niet
leiden tot het ontstaan van auto-immuniteit.
Bij het tweede injectieschema werd het fusieproteïne op dag 18 prenataal toegediend, en
postnataal wekelijks tot 6 weken na de geboorte. Rennert et al. (1999) beschreef bij de

Discussie ___________________________________________________________________________
41
BALB/c muizen die hetzelfde injectieschema toegediend kregen, dat alle lymfeknopen
aanwezig waren, de Peyerse platen afwezig en dat er duidelijke stoornissen waren in de
miltarchitectuur. In dit werk konden met de LIA autoantilichamen aangetoond worden bij
39% van deze dieren op 3 maanden, die gericht waren tegen RNP, Cenp-B en Topo-1/Scl70.
We vinden hier dus een iets hoger percentage aan antilichamen op 3 maand dan bij de
LTβReceptor-/- muizen van dezelfde leeftijd (39% ten opzichte van 24%). Mogelijks is dit te
verklaren doordat bij de dieren die enkel postnataal fusieproteïne kregen de lymfeknopen wél
aanwezig waren, en deze kunnen op die manier zorgen voor een efficiëntere immuunrespons
door ontmoeting en herkenning van antigenen te faciliteren, wat kan leiden tot meer
antilichaamproductie ten opzichte van de LTβReceptor-/- muizen waar alle lymfeknopen
ontbreken. Maar gezien het hier niet gaat om een significant verschil, kan dit ook aan toeval
en de geringe grootte van de onderzoeksgroepen te wijten zijn.
Hieruit kunnen we dus besluiten dat de blokkade van de LTβReceptor-signaalweg
voornamelijk postnataal van belang is in het ontstaan van anti-ENA antilichamen. Daarnaast
wordt hier nogmaals bevestigd dat het ontbreken van lymfeknopen dus niet noodzakelijk is
voor het ontstaan van auto-immuniteit. Ook kunnen we besluiten dat een verstoorde
miltarchitectuur mogelijks betrokken is bij het ontstaan van anti-ENA antilichamen gezien we
enkel deze antilichamen zien ontstaan in dieren waarbij de miltarchitectuur verstoord is, dus
zowel bij de LTβReceptor-/- muizen als bij de muizen behandeld met het fusieproteïne op dag
18 prenataal en postnataal wekelijks tot 6 weken.
5.2.3 Toekomstperspectieven
Om de rol van de LTβReceptor-signaalweg in het ontstaan van auto-immuniteit en het
ontstaansmechanisme verder te ontrafelen, dienen in de toekomst nog andere experimenten
uitgevoerd te worden. Zo zal enerzijds nagegaan worden op welk celtype de afwezigheid van
LTα1β2 vereist is voor het ontstaan van autoantilichamen, via de bepaling van de serologische
afwijkingen in LTβ celspecifieke knock-out muizen. Ook zullen LIGHT-/- muizen bestudeerd
worden, ter uitsluiting van betrokkenheid van andere liganden van de LTβReceptor-
signaalweg in het ontstaan van anti-ENA antilichamen. Anderzijds zullen ook muizen
onderzocht worden waarbij het gen voor de Toll-like Receptor 7 ontbreekt, gezien Savarese et
al. (2008) aantoonden dat de productie van anti-snRNP antilichamen (die RNA bevatten)
afhankelijk is van deze receptor.

Discussie ___________________________________________________________________________
42
5.3
In dit werk werden via IIF op LKS-coupes significant hogere percentages ASMA (anti-
smooth muscle antibodies) aangetoond bij de LTβReceptor-/- muizen ten opzichte van hun
controles, en dit zowel op 3 als 6 maanden. In de literatuur wordt beschreven dat deze ASMA
reageren met F-actine bij humane auto-immuunhepatitis type 1, en het aantonen van deze F-
actine-ASMA binding via IIF op LKS-coupes is bijgevolg een heel sensitieve en specifieke
test voor auto-immuun hepatitis type 1 in de humane situatie (Johnson et al., 1965). Daarnaast
reageren deze ASMA ook met een hele range andere cytoskeletale antigenen, zoals actine en
vimentine, desmine,… Bijgevolg kunnen ASMA ook aangetoond worden bij allerlei andere
aandoeningen dan auto-immuunhepatitis type 1, zoals maligniteiten, hepatitis B, mycoplasma
infectie, thyreoiditis, systemische auto-immuunziekten,… en zelfs bij gezonde individuen
(Silvestrini and Benson, 2001). ASMA tegen antigenen anders dan F-actine kleuren aan
volgens een ander fluorescentiepatroon dan dat van F-actine, waardoor een onderscheid
tussen beide mogelijk is. In dit werk kwam het fluorescentiepatroon overeen met dat van
ASMA tegen F-actine, waardoor bij deze dieren mogelijks een auto-immuunhepatitis type 1
aanwezig is.
Weefselspecifieke antilichamen
Bij humane auto-immuunhepatitis type 1 treft men hier ook vaak ANA aan, wat ook het geval
is bij deze dieren. Daarnaast vindt men in de humane situatie ook verhoogde leverenzymen
(Gish and Mason, 2001), en bijgevolg werd dit ook nagegaan bij deze muizen. De resultaten
worden besproken in de thesis van Sigurd Delanghe. Bij 20% van de dieren van 6 maand
werden significant verhoogde concentraties ALT/AST aangetoond terwijl dit bij geen enkele
C57BL/6 controle het geval was, wat wijst op een mogelijke pathofysiologische rol van de
ASMA antilichamen.

Referentielijst ___________________________________________________________________________
43
6 Referentielijst
ABRAHAM D.J., VARGA J.: Scleroderma: from cell and molecular mechanisms to disease models. Trends Immunol.,2005, 26 (11), 587-595. ARDBUCKLE M.R., JAMES J.A., KOLHASE K.F., RUBERTONE M.V., DENNIS G.J., HARLEY J.B.: Development of anti-dsDNA autoantibodies prior to clinical diagnosis of systemic lupus erythematosus. Scand. J. Immunol., 2001,54, 211-219. BAXTER A.G., HORSFALL A.C., HEALEY D., OZEGBE P., DAY S., WILLIAMS D.G., COOKE A.: Mycobacteria precipitate an SLE-like syndrome in diabetes-prone NOD mice. Immunology, 1994, 83, 227-231. BOEHM T., SCHEU S., PFEFFER K., BLEUL C.C. : Thymic medullary epithelial cell differentiation, thymocyte emigration, and the control of autoimmunity require lympho-epithelial cross talk via LTβR. J. Exp. Med., 2003, 198: 757-769. BONFA E., MARSHAK-ROTHSTEIN A., WEISSBACH H., BROT N., ELKON K.; Frequency and epitope recognition of anti-ribosome P antibodies from humans with systemic lupus erythematosus and MRL/lpr mice are similar. J. Immunol., 1988, 140 (10): 3434-3437. BRADWELL A.R., STOKES R.P., JOHNSON G.D. : Atlas of autoantibody patterns on tissues. KNP Group LTD, Redditch, 1997. BROWNING J.L., NGAMEK A., LAWTON P., DEMARINIS J., TIZARD R., CHOW E.P.C., HESSION C., O’BRINE-GRECO B., FOLEY S.F., WARE C.F.: Lymphotoxin-beta, a novel member of the TNF-family that forms a heterotrimeric complex with lymphotoxin on the cell-surface. Cell, 1993, 72 (6): 847-856. BYGRAVE A.E., ROSE K.L., CORTES-HERMANDEZ J., WARREN J., RIGBY R.J., COOK H.T., WALPORT M.J., VYSE T.J., BOTTO M.: Spontaneous autoimmunity in 129 and C57BL/6 mice- Implications for autoimmunity described in gene-targeted mice. PLoS Biol., 2004, 2 (8): 1081-1090. CHIN R.K., ZHU M., CHRISTIANSEN P.A., LIU W., WARE C., PELTONEN L., ZHANG X., GUO L., HAN S., ZHENG B., FU Y.X. : Lymphotoxin pathway-directed, autoimmune regulator-independent central tolerance to arthritogenic collagen. J Immunol., 2006, 177(1), 290-297.

Referentielijst ___________________________________________________________________________
44
COSTA O., MONIER J.C.: Antihistone antibodies detected by micro-ELISA and immunoblotting in mice with lupus-like syndrome (MRL/l, MRL/n, PN and NZB strains). Clin. Immunol. Immunopathol., 1986, 40(2), 276-82.
CORTES-HERNANDEZ J., ORDI-ROS J., LABRADOR M., BUJAN S., BALADA E., SEGARRA A., VILARDELL-TARRES M.: Antihistone and anti–double-stranded deoxyribonucleic acid antibodies are associated with renal disease in systemic lupus erythematosus. Am. J. Med., 2004, 116, 165-173.
D’CRUZ D.P, KHAMASHTA M.A., HUGHES G.R.V.: Systemic lupus erythematosus. Lancet, 2007, 369, 587-596. ECKES B., HUNZELMANN N., MOINZADEH P., KRIEG T.: Scleroderma - news to tell. Arch. Dermatol. Res., 2007, 299, 139-144. EISENBERG R.A., TAN E.M., DIXON F.J.: Presence of anti-Sm reactivity in autoimmune mouse strains.J. Exp. Med., 1987, 147: 582-587. FU Y., CHAPLIN D.: Development and maturation of secondary lymphoid tissues. Annu. Rev. Immunol., 1999, 17: 399-433. FUTTERER A., MINK K., LUZ A., KOSCO-VILBOIS M.H., PFEFFER K. : The lymphotoxin beta receptor controls organogenesis and affinity maturation in peripheral lymphoid tissues. Immunity, 1998, 9 (1): 59-70. FRANKI A.S., VAN BENEDEN K., DEWINT P., HAMMOND K.L.J., LAMBRECHT S., LECLERCQ G., KRONENBERG M., DEFORCE D., ELEWAUT D.: A unique lymphotoxin αβ dependent pathway regulates thymic emigration of Vα14i NKT cells. Proc. Natl. Acad. Sci. USA, 2006, 103 (24): 9160-9165. FRITZLER J.M., SALAZAR M.: Diversity and origin of rheumatologic autoantibodies. Clin Microbiol. Rev., 1991, 4, 256-269. JIMENEZ S.A., CHRISTNER P.J.: Murine animal models of systemic sclerosis. Curr. Opin. Rheumatol., 2002, 14: 671-680. GENTILETTI J., AYITEY S., CHRISTNER P.J.: Pathologic alterations in the internal organs of the Tsk2/+ mouse. ACR 2006. GIOUD M., KOTZIN B.L., RUBIN R.L., JOSLIN F.G., TAN E.M.: In vivo and in vitro production of anti-histone antibodies in NZB/NZW mice. J. Immunol., 1983, 131: 269-274.

Referentielijst ___________________________________________________________________________
45
GISH R.G., MASON A.: Autoimmune liver disease: Current standards, future directions. Clin. Liver Dis., 2001, 5, 287-314. GRANGER S.W., WARE C.F.: Turning on LIGHT. J. Clin. Invest., 2001, 108, 1741-1742. GOLDSBY R.A., KINDT T.J., OSBORNE B.A., KUBY J.: Enzyme-Linked Immuno-sorbent Assay. In: Immunology, 5th ed., W. H. Freeman, New York, 2003, 148-150. GOMMERMAN J.L., BROWNING J.L.: Lymphotoxin/light, lymphoid microenvironments and auto-immmune disease. Nat. Rev. Immunol., 2003, 3 (8): 642-688. HASEGAWA M., HAMAGUCHI Y., YANABA K., BOUAZIZ J.D., UCHIDA J., FUJIMOTO M., MATSUSHITA T., MATSUSHITA Y., HORIKAWA M., KOMURA K., TAKEHARA K., SATO S., TEDDER T.F.: B-lymphocyte depletion reduces skin fibrosis and autoimmunity in the tight-skin mouse model for systemic sclerosis. Am. J. Path., 2006, 169 (3): 954-966. IIZUKA K., CHAPLIN D.D., WANG Y., WO Q., PEGG L.E., YOKOYAMA W.M., et al. : Requirement for membrane lymphotoxin in natural killer cell development. Proc. Natl. Acad. Sci. USA, 1999, 96 (11):6336-6340. JANEWAY C.A., TRAVERS P., WALPORT M., SHLOMCHIK M.J.: Immunobiology: the immuunsystem in health and disease, 6th ed. Garland Science Publishing, New York, 2005. JIMENEZ S.A., CHRISTNER P.J.: Murine animal models of systemic sclerosis. Curr. Opin. Rheumatol., 2002, 14: 671-680. JOHNSON G.D., HOLBOROW E.J., GLYNN L.E.: Antibody to smooth muscle in patients with liver disease. Lancet, 1965, 2: 878-879. KURIEN B.T., SCOFIELD R.H.: Western blotting. Methods, 2006, 38(4), 878-879. LADERACH D., KOUTOUZOV S., BACH J.F., YAMAMOTO A.M.: Concomitant early appearance of anti-ribonucleoprotein and anti-nucleosome antibodies in lupus prone mice. J. autoimmun., 2003, 20: 161-170. LINNIK M.D., HU J.Z., HEILBRUNN K.R., STRAND V., HURLEY F.L., JOH T.: Relationship between anti-double-stranded DNA antibodies and exacerbation of renal disease in patients with systemic lupus erythematosus. Arthritis Rheum., 2005, 52 (4), 1129-1137. LIU K., MOHAN C.: What do mouse models teach us about human SLE? Clin. Immunol., 2006, 119, 123-130.

Referentielijst ___________________________________________________________________________
46
MARTINS V.C., BOEHM T., BLEUL C.C.: LTβR signaling does not regulate aire-dependent transcripts in medullary thymic epithelial cells. J. Immunol., 2008, 181: 400-407. MEHEUS L., VAN VENROOIJ W.J., WIIK A., CHARLES P.J., TZIOUFAS A.G., MEYER O., STEINER G., GIANOLA D., BOMBARDIERI S., UNION A., DE KEYSER S., VEYS E., DE KEYSER F.: Multicenter validation of recombinant, natural and sysnthetic antigens used in a single multiparamter assay for the detection of specific anti-nuclear autoantibodies in connective tissue disorders. Clin. Exp. Rheumatol., 1999, 17, 205-214. MURPHY E.D., ROTHS J.B. : Autoimmunity and lymphoproliferation: Induction by mutant gene lpr, and acceleration by a male-associated factor in strain BXSB mice. (1978). In: Genetic control of autoimmune disease, Eds. N.R. Rose, P.E. Bigazzi, L.N. Warner LN., Elsevier North-Holland, New York, 1978, 207-221. POLLOCK W., TOH B.: Routine immunofluorescence detection of Ro/SS-A autoantibody using Hep-2 cells transfected with human 60kDa Ro/SS-A. J. Clin. Pathol., 1999, 52, 684-687. SATOH M., KUMAR A., KANWAR Y.S., REEVES W.H.: Antinuclear antibody production and immune-complex glomerulonephritis in BALB/c mice treated with pristane. Proc. Natl. Acad. Sci. USA, 1995, 92(24), 10934–10938. SATOH M., RICHARDS H.B., SHAHEEN V.M., YOSHIDA H., SHAW M., NAIM J.O., WOOLEY P.H., REEVES W.H.: Widespread susceptibility among inbred mouse strains to the production of lupus autoantibodies by pristane. Clin. Exp. Immunol., 2000, 121, 399-405. SAVARESE E., STEINBERG C., PAWAR R.D., REINDL W., SHIZUO A., ANDERS H.J., KRUG A.: Requirement of Toll-like Receptor 7 for pristine-induced production of autoantibodies and development of murine lupus nephritis. Arthritis Rheum., 2008, 58: 1107-1115. SHIELDS D.C., HARMON D.L., NUNEZ F., WHITEHEADL A.S.: The evolution of haematopoetic cytokine/receptor complexes. Cytokine, 1995, 7 (7), 679-688. SILVESTRINI R.A., BENSON E.M.: Whither smooth muscle antibodies in the third millennium? J. Clin. Pathol., 2001, 54, 677-678. SKARSTEIN K., WAHREN M., ZAURA E., HATTORI M., JONSSON R.: Characterization of T cell receptor repertoire and anti-Ro/SSA autoantibodies in relation to sialadenitis of NOD mice. Autoimmunity, 1995, 22, 9-16.

Referentielijst ___________________________________________________________________________
47
THEOFILOPOULOS A.N., DIXON F.J.: Murine models of systemic lupus erythematosus. Adv. Imuunol., 1985, 37, 269-390. VANASSCHE I.: Ontstaansmechanismen van SLE en voorkomen en detectie van antinucleaire antilichamen in verschillende muizenstammen. Scriptie Biomed. Wet. Ugent, 2006. WAHREN M., SKARSTEIN K., BLANGE I., PETTERSSON I., JONSSON R.: MRL/lpr mice produce anti-Ro 52000 MW antibodies: detection, analysis of specificity and site of production. Immunology, 1994, 83 (1): 9-15. WARE C.F.: Network Communications: Lymphotoxins, LIGHT and TNF. Annu. Rev. Immunol., 2005, 23: 787-819.

Bijlagen Dit addendum bevat:
- Foto 1:
Immunoreactieve patronen via LIA van enkele C57BL/6 en enkele BWF1 muizen op
een leeftijd van 24 weken; en van alle MRL/lpr muizen op een leeftijd van 16
weken
- Foto 2:
Immunoreactieve patronen via LIA van de pristane-induced lupus muizen en hun
controles op 24 weken na de pristane/PBS-injectie.
- Foto 3:
Immunoreactieve patronen via LIA van de LTβR-/- muizen op een leeftijd van 3
maand en 6 maand.
- Foto 4:
Immunoreactieve patronen via LIA van enkele fusieproteïne behandelde muizen
(prenataal op dag 18 en verlengd tot 6 weken postnataal) en enkele van hun
controles op een leeftijd van 24 weken.

C57BL/6 BWF1 MRL/lpr (24 weken) (24 weken) (16 weken)
_________________ __________________ _________________________________________________
Foto 1: Immunoreactieve patronen via LIA van enkele C57BL/6 en enkele BWF1 muizen op een leeftijd van 24 weken; en van alle MRL/lpr muizen op een leeftijd van 16 weken.

Pristane-induced lupus controles voor de muizen pristane-induced lupus muizen
(24 weken na de pristane-injectie) (24 weken na de PBS injectie) _____________________________________________ _______________________________________________
Foto 2: Immunoreactieve patronen via LIA van de pristane-induced lupus muizen en hun controles op 24 weken na de pristane/PBS-injectie.
Opmerking: Bij alle onderzochte groepen muizen werden telkens ook negatieve controles getest via LIA. Indien een
bepaalde reactiviteit bij deze negatieve controles positief bleek, werd deze reactiviteit bij de onderzochte groep op basis hiervan niet als positief beschouwd.
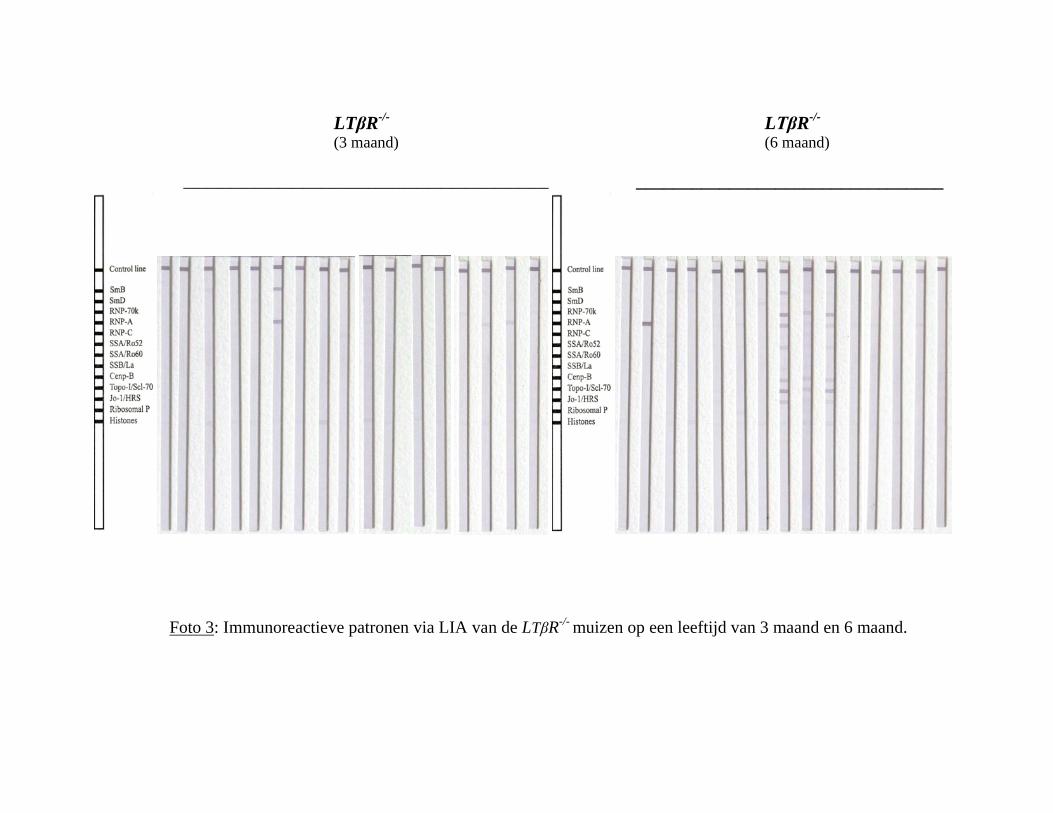
LTβR-/- LTβR-/-
(3 maand) (6 maand)
_____________________________________________________________ _________________________________
Foto 3: Immunoreactieve patronen via LIA van de LTβR-/- muizen op een leeftijd van 3 maand en 6 maand.

Fusieproteïne behandelde muizen controles volgens schema D18,verlengd 6w
(24 weken) (24 weken) ________________________________ ____________________________________
Foto 4: Immunoreactieve patronen via LIA van enkele fusieproteïne behandelde muizen (prenataal op dag 18 en
verlengd tot 6 weken postnataal) en enkele van hun controles op een leeftijd van 24 weken.




![[KT] Hitman Reborn 391 Fr (fairynopiece.shonenblog.com)](https://static.fdocuments.nl/doc/165x107/568c48a61a28ab49169100bf/kt-hitman-reborn-391-fr-fairynopieceshonenblogcom.jpg)













