RÉVISION SYSTÉMATIQUE DES LEPTODIRINAE ...3 Révision des Leptodirinae souterrains du bassin de...
19
RÉVISION SYSTÉMATIQUE DES LEPTODIRINAE SOUTERRAINS DES MONTS ASPUSENI. VI. LE SOUS-GENRE PHOLEUON (S. STR.) DU BASSIN DE SOMEŞUL CALD (MONTS DU BIHOR) GHEORGHE RACOVIŢĂ Résumé. Des mesures biométriques entreprises sur 746 mâles et 760 femelles de Pholeuon (s. str.) angusticolle collectés de neuf grottes ont fourni les données brutes pour un traitement statistique réalisé par les mêmes méthodes multifactorielles que celles employées dans nos précédentes études régionales. Il est ainsi démontré qu’aux sous-espèces connues jusqu’à présent dans le bassin de Someşul Cald s’ajoutent trois autres, nouvelles pour la science: P. a. alunensis, P. a. longicornis et P. a. gujai. 1. INTRODUCTION Le Someşul Cald coule dans sa partie supérieure à travers une pittoresque région calcaire, à la limite nord des Monts du Bihor. Dans le bassin hydro- graphique de cette rivière 87 cavités souterraines ont été inventoriées jusqu’en 1981 (GORAN, 1982), pour qu’à présent leur nombre dépasse largement la centaine. Les Coléoptères Leptodirinae y sont cependant représentés par une seule espèce troglobie, Pholeuon (s. str.) angusticolle Hampe, 1856, endémique pour ce périmètre et comportant quatre sous-espèces (JEANNEL, 1924, DECU, 1964): − P. angusticolle (s. str.), de la grotte dite Peştera Zmeilor de la Onceasa; − P. a. mihoki Csiki, 1911, de la Peştera de la Alun et Şura Ponorului; − P. a. bihariense Csiki, 1912, de l’Izbucul de la Rădeasa; − P. a. arpadi Csiki, 1912, de la Peştera din Valea Firii. Différenciées au début par les procédés propres à la systématique classique, trois de ces sous-espèces (angusticolle, mihoki et bihariense) ont fait plus tard l’objet d’une étude de taxonomie quantitative par laquelle MOLDOVAN (1989) a validé leur existence. Quant à P. a. arpadi, cet auteur l’a considéré comme disparu de son unique station connue à l’époque, car il n’y a pas pu être repris ni même à l’aide de pièges appâtées. Et il n’est point exclu que pareille conclusion soit la raison pour laquelle ce taxon ne figure plus dans le catalogue de PERRAU (2000). En revanche, il est cité à nouveau dans un ample répertoire sur la faune de la Roumanie, publié par MOLDOVAN et al. (2007). Trav. Inst. Spéol. «Émile Racovitza», t. XLVIII, p. 11–29, Bucarest, 2009
Transcript of RÉVISION SYSTÉMATIQUE DES LEPTODIRINAE ...3 Révision des Leptodirinae souterrains du bassin de...

RÉVISION SYSTÉMATIQUE DES LEPTODIRINAE SOUTERRAINS DES MONTS ASPUSENI.
VI. LE SOUS-GENRE PHOLEUON (S. STR.) DU BASSIN DE SOMEŞUL CALD (MONTS DU BIHOR)
GHEORGHE RACOVIŢĂ
Résumé. Des mesures biométriques entreprises sur 746 mâles et 760 femelles de Pholeuon (s. str.) angusticolle collectés de neuf grottes ont fourni les données brutes pour un traitement statistique réalisé par les mêmes méthodes multifactorielles que celles employées dans nos précédentes études régionales. Il est ainsi démontré qu’aux sous-espèces connues jusqu’à présent dans le bassin de Someşul Cald s’ajoutent trois autres, nouvelles pour la science: P. a. alunensis, P. a. longicornis et P. a. gujai.
1. INTRODUCTION
Le Someşul Cald coule dans sa partie supérieure à travers une pittoresque région calcaire, à la limite nord des Monts du Bihor. Dans le bassin hydro-graphique de cette rivière 87 cavités souterraines ont été inventoriées jusqu’en 1981 (GORAN, 1982), pour qu’à présent leur nombre dépasse largement la centaine. Les Coléoptères Leptodirinae y sont cependant représentés par une seule espèce troglobie, Pholeuon (s. str.) angusticolle Hampe, 1856, endémique pour ce périmètre et comportant quatre sous-espèces (JEANNEL, 1924, DECU, 1964):
− P. angusticolle (s. str.), de la grotte dite Peştera Zmeilor de la Onceasa; − P. a. mihoki Csiki, 1911, de la Peştera de la Alun et Şura Ponorului; − P. a. bihariense Csiki, 1912, de l’Izbucul de la Rădeasa; − P. a. arpadi Csiki, 1912, de la Peştera din Valea Firii.
Différenciées au début par les procédés propres à la systématique classique, trois de ces sous-espèces (angusticolle, mihoki et bihariense) ont fait plus tard l’objet d’une étude de taxonomie quantitative par laquelle MOLDOVAN (1989) a validé leur existence. Quant à P. a. arpadi, cet auteur l’a considéré comme disparu de son unique station connue à l’époque, car il n’y a pas pu être repris ni même à l’aide de pièges appâtées. Et il n’est point exclu que pareille conclusion soit la raison pour laquelle ce taxon ne figure plus dans le catalogue de PERRAU (2000). En revanche, il est cité à nouveau dans un ample répertoire sur la faune de la Roumanie, publié par MOLDOVAN et al. (2007).
Trav. Inst. Spéol. «Émile Racovitza», t. XLVIII, p. 11–29, Bucarest, 2009

Gheorghe Racoviţă 2
12
2. MATÉRIEL
Le matériel faunistique disponible pour les mesures biométriques consiste en neuf échantillons, tous obtenus par suite de captures faites à l’aide des appâts. Tel que nous l’avons précédemment précisé dans nos travaux similaires, il s’agit d’échantillons ayant des effectifs suffisamment grands pour assurer un degré convenable de signification statistique, en cas idéal de 100 individus de chaque sexe. Les grottes de provenance sont énumérées ci-dessus, avec la spécification des abréviations utilisées en ce qui suit et des codes décimaux sous lesquels elles sont inscrites dans le Cadastre du karst de la Roumanie (GORAN, 1982).
− Izbucul din Alunul Mare (AM) – 3411/5 – 100 ♂ et 100 ♀ – Legit: O. Guja; − Peştera Onceasa (= Peştera Zmeilor de la Onceasa) (ZO) – 3414/1 – 100 ♂
et 100 ♀ – Legit: O. Guja; − Peştera lui Teo (Te) – 3414/44 – 100 ♂ et 100 ♀ – Legit: O. Guja; − Peştera Diaclazei (Dc) – 3414/6 – 100 ♂ et 100 ♀ – Legit: O. Guja; − Poarta Alunului (PA) – 3413/8 – 100 ♂ et 100 ♀ – Legit: O. Guja; − Peştera de la Alun (Al) – 3413/1 – 100 ♂ et 100 ♀ – Legit: O. Guja; − Peştera din Piatra Ponorului (PP) – 3414/5 – 100 ♂ et 100 ♀ – Legit: Liliana
Potra; − Peştera din Dealul Humpleu (DH) (= Peştera Mare din Valea Firii) –
3416/18 – 26 ♂ et 41 ♀ – Legit: Oana Moldovan et B. Onac; − Peştera Grama (PG) (= Corobaia Mare) – 3416/17 – 20 ♂ et 19 ♀ – Legit:
O. Guja.
Des P. angusticolle ont été trouvés également dans quatre autres grottes, mais le nombre d’exemplaires recueillis a été chaque fois trop faible (généralement inférieur à 10) pour que ces échantillons puissent être pris en considérations. A l’exception près, il s’agit de grottes d’où provenait le matériel ayant servi à l’étude faite par MOLDOVAN (1989), à savoir Izbucul de la Rădeasa (3410/3), Peştera cu Ceaţă (3410/64), Peştera Colţului (3410/57) et Peştera cu Oase (3413/3).
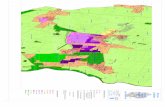
La position géographique de toutes ces 13 cavités, auxquelles s’ajoutent la Şura Ponorului (= Ponorul Văii Ponorului) (3414/2) et la Peştera cu Apă din Valea Firii (3416/5) citées par Jeannel (1924), est marquée sur la Fig. 1.
3. BIOMÉTRIE
Afin de conserver la méthodologie appliquée tout le long de notre révision des Leptodirinae, dans l’étude biométrique ont été pris en considération les mêmes caractères quantitatifs utilisés dans le cas des populations de Pholeuon (s. str.) localisées dans le bassin de l’Arieş et la zone de Padiş (RACOVIŢĂ, 2004–2005, 2006–2007). Nous rappelons qu’il s’agit de neuf caractères dimensionnels – longueur du

3 Révision des Leptodirinae souterrains du bassin de Someşul Cald
13
Fig.
1. –
Em
plac
emen
t dan
s le
bass
in su
périe
ur d
e So
meş
ul C
ald
des c
avité
s peu
plée
s par
Pho
leuo
n (s
. str.
) ang
ustic
olle
. Cel
les q
ui o
nt fo
urni
les
écha
ntill
ons u
tilis
és d
ans l
es a
naly
ses m
ultif
acto
rielle
s son
t rep
rése
ntée
s par
des
sym
bole
s noi
rs. L
a lig
ne é
pais
se p
onct
uée
mar
que
la li
mite
du
bass
in h
ydro
grap
hiqu
e.
1 =
Izbu
cul d
e la
Răd
easa
; 2 =
Peş
tera
cu
Cea
ţă; 3
= Iz
bucu
l din
Alu
nul M
are;
4 =
P. C
olţu
lui;
5 =
P. Z
mei
lor d
e la
Onc
easa
; 6 =
P. l
ui T
eo; 7
= P
. D
iacl
azei
; 8 =
Poa
rta A
lunu
lui;
9 =
P. d
e la
Alu
n; 1
0 =
P. c
u O
ase;
11 =
Şura
Pon
orul
ui; 1
2 =
P. d
in P
iatra
Pon
orul
ui; 1
3 =
P. d
in D
ealu
l Hum
pleu
; 14
= P
. cu
Apă
din
Val
ea F
irii;
15 =
P. G
ram
a.

Gheorghe Racoviţă 4
14
corps (Lc), longueur du pronotum (Lp), largeur du bord antérieur du pronotum (lap), largeur maximum du pronotum (lMp), largeur minimum du pronotum (lmp), largeur de la base du pronotum (lbp), longueur des élytres (Le), largeur des élytres (le) et longueur des antennes (La) –, à partir des quels ont été calculés les rapports définissant huit caractères morphométriques – Lp/lMp, lap/lMp, lmp/lMp, lbp/lmp, lap/lbp, Le/le, La/Lc et lMp/le. Une remarque faite au cours de mesures biométriques et ayant trait à la morphologie du pronotum nous a toutefois amené à en ajouter en tant que caractère morphométrique le rapport lbp/lMp. Les moyennes arithmétiques et les écarts standards de ces 18 paramètres biométriques sont inscrits dans les Tableaux 1 et 2.
Tableau 1 Moyennes arithmétiques (µ, en mm) et écarts standard (σ) des caractères dimensionnels
Échant. Lc Lp lap lMp lmp lbp Le le La Mâles
AM µ 4,518 1,113 0,776 1,175 1,013 1,032 3,105 1,893 3,082 n = 100 σ 0,1012 0,0301 0,0206 0,0325 0,0293 0,0292 0,0768 0,0421 0,0827
ZO µ 4,327 1,078 0,740 1,113 0,967 0,994 2,949 1,813 2,881 n = 100 σ 0,0929 0,0295 0,0204 0,0370 0,0316 0,0335 0,0699 0,0441 0,0735
Dc µ 4,403 1,093 0,744 1,141 0,991 1,009 3,010 1,799 3,041 n = 100 σ 0,0977 0,0304 0,0218 0,0356 0,0340 0,0337 0,0723 0,0480 0,0782
PA µ 4,345 1,077 0,733 1,126 0,985 1,004 2,968 1,773 3,020 n = 100 σ 0,1074 0,0348 0,0208 0,0345 0,0360 0,0372 0,0793 0,0515 0,0907
Te µ 4,371 1,086 0,734 1,130 0,987 1,003 2,985 1,777 3,029 n = 100 σ 0,0848 0,0265 0,0167 0,0288 0,0252 0,0271 0,0666 0,0386 0,0723
Al µ 4,411 1,091 0,741 1,144 0,994 1,011 3,020 1,810 3,085 n = 100 σ 0,0925 0,0304 0,0191 0,0308 0,0325 0,0330 0,0704 0,0425 0,0720
PP µ 4,304 1,065 0,734 1,101 0,955 0,961 2,939 1,716 3,027 n = 100 σ 0,0952 0,0294 0,0218 0,0350 0,0335 0,0339 0,0733 0,0421 0,0971
DH µ 4,282 1,057 0,713 1,096 0,951 0,966 2,925 1,722 2,968 n = 25 σ 0,0883 0,0247 0,0183 0,0277 0,0255 0,0256 0,0716 0,0434 0,0615
PG µ 4,274 1,074 0,727 1,129 1,006 1,021 2,900 1,744 2,939 n = 20 σ 0,0968 0,0249 0,0226 0,0290 0,0292 0,0274 0,0757 0,0386 0,0763
Femelles AM µ 4,763 1,147 0,822 1,248 1,089 1,109 3,315 2,045 2,854
n = 100 σ 0,1085 0,0323 0,0238 0,0385 0,0398 0,0403 0,0817 0,0527 0,0777 ZO µ 4,599 1,120 0,789 1,192 1,048 1,077 3,179 1,979 2,708
n = 100 σ 0,0917 0,0291 0,0204 0,0325 0,0306 0,0305 0,0674 0,0469 0,0658 Dc µ 4,635 1,115 0,783 1,198 1,058 1,070 3,220 1,942 2,749
n = 100 σ 0,0943 0,0285 0,0180 0,0311 0,0303 0,0321 0,0716 0,0487 0,0677 PA µ 4,576 1,108 0,775 1,185 1,059 1,072 3,168 1,923 2,748
n = 100 σ 0,1011 0,0277 0,0197 0,0316 0,0346 0,0320 0,0794 0,0500 0,0738 Te µ 4,590 1,110 0,784 1,187 1,053 1,070 3,180 1,921 2,730
n = 100 σ 0,0873 0,0274 0,0189 0,0294 0,0299 0,0279 0,0658 0,0445 0,0701 Al µ 4,636 1,114 0,785 1,196 1,059 1,077 3,222 1,956 2,827
n = 100 σ 0,1091 0,0324 0,0225 0,0339 0,0355 0,0348 0,0811 0,0542 0,0742 PP µ 4,517 1,090 0,773 1,159 1,027 1,020 3,128 1,857 2,742
n = 100 σ 0,0948 0,0295 0,0229 0,0346 0,0334 0,0346 0,0739 0,0492 0,0694 DH µ 4,498 1,078 0,758 1,157 1,022 1,039 3,120 1,857 2,687
n = 39 σ 0,1174 0,0330 0,0233 0,0399 0,0378 0,0369 0,0893 0,0545 0,0727 PG µ 4,523 1,095 0,761 1,185 1,064 1,078 3,128 1,882 2,650
n = 18 σ 0,0983 0,0379 0,0151 0,0397 0,0448 0,0418 0,0669 0,0418 0,0822

5 Révision des Leptodirinae souterrains du bassin de Someşul Cald
15
Tableau 2
Moyennes arithmétiques (µ) et écarts standard (σ) des caractères morphométriques
Echant. Lp/ lMp
lap/ lMp
lmp/ lMp
lbp/ lMp
lbp/ lmp
lap/ lbp
Le/ le
La/ Lc
lMp/ le
Mâles AM µ 0,948 0,661 0,862 0,879 1,019 0,752 1,641 0,682 0,621
n = 100 σ 0,0218 0,0148 0,0161 0,0177 0,0162 0,0198 0,0300 0,0114 0,0120 ZO µ 0,969 0,665 0,869 0,894 1,028 0,745 1,627 0,666 0,614
n = 100 σ 0,0235 0,0170 0,0165 0,0203 0,0190 0,0188 0,0290 0,0136 0,0163 Dc µ 0,958 0,652 0,869 0,884 1,018 0,738 1,674 0,691 0,635
n = 100 σ 0,0194 0,0160 0,0164 0,0179 0,0151 0,0192 0,0351 0,0126 0,0159 PA µ 0,957 0,651 0,875 0,891 1,019 0,731 1,675 0,695 0,636
n = 100 σ 0,0224 0,0145 0,0184 0,0219 0,0181 0,0207 0,0331 0,0143 0,0146 Te µ 0,961 0,650 0,873 0,888 1,017 0,732 1,681 0,693 0,636
n = 100 σ 0,0230 0,0131 0,0134 0,0203 0,0194 0,0178 0,0317 0,0121 0,0150 Al µ 0,954 0,648 0,869 0,884 1,017 0,733 1,669 0,700 0,632
n = 100 σ 0,0192 0,0138 0,0157 0,0185 0,0163 0,0193 0,0344 0,0136 0,0133 PP µ 0,967 0,667 0,867 0,872 1,007 0,765 1,713 0,703 0,642
n = 100 σ 0,0242 0,0162 0,0138 0,0175 0,0223 0,0214 0,0322 0,0153 0,0153 DH µ 0,965 0,651 0,868 0,882 1,017 0,739 1,699 0,693 0,636
n = 26 σ 0,0188 0,0149 0,0130 0,0208 0,0181 0,0185 0,0330 0,0102 0,0132 PG µ 0,952 0,644 0,891 0,905 1,015 0,712 1,663 0,688 0,647
n = 20 σ 0,0177 0,0130 0,0140 0,0142 0,0168 0,0133 0,0247 0,0142 0,0125 Femelles
AM µ 0,920 0,659 0,873 0,888 1,018 0,742 1,622 0,599 0,610 n = 100 σ 0,0205 0,0113 0,0179 0,0235 0,0169 0,0227 0,0248 0,0103 0,0139
ZO µ 0,940 0,662 0,880 0,904 1,028 0,733 1,607 0,589 0,602 n = 100 σ 0,0179 0,0126 0,0131 0,0215 0,0239 0,0179 0,0243 0,0105 0,0119
Dc µ 0,931 0,654 0,884 0,894 1,011 0,732 1,659 0,593 0,617 n = 100 σ 0,0163 0,0116 0,0136 0,0182 0,0165 0,0157 0,0299 0,0123 0,0113
PA µ 0,936 0,654 0,894 0,905 1,012 0,723 1,648 0,601 0,616 n = 100 σ 0,0175 0,0127 0,0206 0,0190 0,0183 0,0169 0,0298 0,0116 0,0120
Te µ 0,935 0,660 0,887 0,901 1,016 0,733 1,656 0,595 0,618 n = 100 σ 0,0197 0,0132 0,0160 0,0180 0,0140 0,0166 0,0302 0,0121 0,0145
Al µ 0,931 0,656 0,885 0,901 1,017 0,729 1,648 0,610 0,612 n = 100 σ 0,0172 0,0135 0,0142 0,0192 0,0196 0,0171 0,0291 0,0117 0,0117
PP µ 0,941 0,667 0,886 0,880 0,993 0,758 1,685 0,607 0,624 n = 100 σ 0,0176 0,0162 0,0165 0,0220 0,0205 0,0206 0,0297 0,0108 0,0153
DH µ 0,932 0,655 0,883 0,899 1,017 0,730 1,681 0,597 0,623 n = 39 σ 0,0185 0,0131 0,0166 0,0256 0,0235 0,0201 0,0301 0,0085 0,0175
PG µ 0,925 0,643 0,898 0,911 1,014 0,706 1,662 0,586 0,629 n = 18 σ 0,0209 0,0196 0,0183 0,0257 0,0201 0,0275 0,0211 0,0154 0,0134
Les valeurs moyennes du coefficient de variation sont de 2,71% pour les caractères dimensionnels et de 2,05% pour ceux morphométriques. Ces pourcentages sont inférieurs à ceux établis pour les échantillons de P. knirschi

Gheorghe Racoviţă 6
16
(3,15%, respectivement 2,34%), mais sans que la différence puisse avoir une certaine signification. Mentionnons aussi pour mémoire que ces valeurs sont pratiquement égales pour les deux sexes. En somme, ce qu’on doit retenir est que la variabilité individuelle n’est pas moins importante chez P. angusticolle. Le fait est mis clairement en évidence par le pronotum des individus faisant partie d’une même population et présentant les valeurs extrêmes (minima et maxima) des caractères morphométriques relatifs à ce segment du corps (Fig. 2).
Fig. 2. – Contour du pronotum tracé à partir des valeurs minima et maxima de quatre caractères
morphométriques (échantillon de mâles de la Poarta Alunului).
Calculées comme d’habitude pour tous les échantillons de mâles composés de 100 individus, les courbes normales qui reproduisent la distribution de fréquences des caractères quantitatifs (Fig. 3 et 4) s’avère être comparables à celles obtenues pour P. knirschi de la zone de Padiş (RACOVIŢĂ, 2006–2007). La ressemblance est déterminée surtout par deux éléments. Le premier est le groupement très sérés des courbes correspondant au rapport Lp/lMp, en concordance avec le fait que, tel que nous allons le voir, ce paramètre est moins utile dans les différenciations infra-spécifiques. Le second concerne la manière similaire dont sont réparties les courbes illustrant la longueur absolue et celle relative de l’antenne (La, respectivement La/Lc), plus ou moins espacées dans les deux cas. Apparemment banal, ce dernier aspect doit être néanmoins signalé du fait que, chez les populations de P. knirschi cantonnées dans le bassin de l’Arieş, les courbes en question étaient bien plus dispersées dans le cas du rapport La/Lc (RACOVIŢĂ, 2004–2005). En conséquence, nous avions considéré la longueur relative des antennes comme un très important caractère de diagnose, idée qui s’avère maintenant n’être point généralisable. Notons encore que la position que ces

7 Révision des Leptodirinae souterrains du bassin de Someşul Cald
17
Fig. 3. – Distributions normales de fréquences pour trois caractères dimensionnels
(abréviations dans le texte).
courbes occupent dans le système de coordonnées ne comporte pas de différences significatives entre les échantillons de mâles et ceux de femelles, quoi qu’il en soit le caractère auquel on se rapporte. En revanche, la longueur relative des antennes reste le caractère quantitatifs le plus représentatif au point de vue dimorphisme sexuel,

Gheorghe Racoviţă 8
18
Fig. 4. – Distributions normales de fréquences pour trois caractères morphométriques
(abréviations dans le texte).
tel que nous l’avons souligné dès le travail qui a marqué le début de la révision systématique du genre Pholeuon (RACOVIŢĂ, 1995). En effet, le paramètre statistique t (le test Student), calculé à partir de la différence entre les moyennes arithmétiques des valeurs biométriques individuelles pour les échantillons de mâles et de femelles, prend la valeur de loin la plus grande pour le rapport La/Lc (52,85

9 Révision des Leptodirinae souterrains du bassin de Someşul Cald
19
en moyenne), alors qu’il est inférieur à 26 pour n’importe quel autre caractère, y compris la longueur du corps.
4. TAXONOMIE NUMÉRIQUE
Compte tenu du fait que le codage des caractères quantitatifs a été réalisé de la même manière que celle appliqué dans toutes les autres analyses régionales, nous n’insistons plus sur cette séquence du protocole d’étude.
Dans la matrice taxons × taxons (Tableau 3), la population pour laquelle la moyenne réciproque des coefficients de similitude a la valeur la plus grande est celle de la Peştera lui Teo (Te). Par conséquent, c’est cette population qui constitue le «taxon central» et qui, de ce fait, occupe une position médiane dans le dendrogramme construit à partir de cette matrice (Fig. 5).
Tableau 3
Matrice taxons × taxons pour l’ensemble des échantillons (males + femelles)
AM ZO Dc PA Te Al PP DH PG Mr % AM – 70,83 75,00 71,53 71,53 79,17 63,89 70,14 77,78 72,48 ZO 102 – 79,86 84,03 84,03 79,17 74,31 80,56 81,25 79,25 Dc 108 115 – 88,89 95,14 95,14 76,39 86,11 85,42 85,24 PA 103 121 128 – 96,53 89,58 77,78 86,11 90,28 85,59 Te 103 121 137 139 – 90,97 79,17 86,11 86,11 86,20 Al 114 114 137 129 131 – 77,08 84,03 86,11 85,16 PP 92 107 110 112 114 111 – 91,67 81,94 77,78 DH 101 116 124 124 124 121 132 – 87,50 84,03 PG 112 117 123 130 124 124 118 126 – 84,55
L’examen de ce diagramme révèle en premier lieu l’isolement évident de la population cantonnée dans la résurgence de la vallée d’Alunul Mare (AM), qui est connectée aux autres échantillons au niveau de similitude le plus bas (72,5%). Dans une situation relativement semblable se trouve ensuite la population de la Peştera Zmeilor de la Onceasa (ZO), liée à un niveau un peu plus élevé, de 80,6%. Il s’ensuit que ces deux populations peuvent être attribuées chacune à des sous-espèces à part. Au pôle opposé se placent les populations de la Poarta Alunului (PA), Peştera lui Teo, Peştera Diaclazei (Dc) et Peştera de la Alun (Al), qui forment une tétrade assez homogène, avec une moyenne triangulaire1 de 92,71%. Il est donc possible que ces populations appartiennent à une troisième sous-espèce.
1 Terme proposé par SNEATH (1957) pour designer la moyenne des coefficients de similitude calculés en comparant successivement chaque échantillon avec tous les autres compris dans un certain secteur (le plus souvent triangulaire) de la matrice taxons × taxons.

Gheorghe Racoviţă 10
20
Fig. 5. – Dendrogramme global obtenu par analyse numérique (échantillons de mâles et de femelles
réunis). En ordonnée, niveau de similitude.
Quant au reste des populations, leur statut est moins clair. On voit d’abord que la population de la Peştera Grama (PG) est liée à la tétrade qu’on vient de définir à un niveau de similitude de 87,0%, un peu trop grand pour qu’on puisse affirmer sur cette unique base qu’il s’agit d’une sous-espèce différente. Mais, par ailleurs, on ne doit pas négliger le fait que cette cavité est située dans un autre sous-bassin hydrographique (Fig. 1), à une distance à laquelle l’isolement géographique devient tout à fait possible.
Dans une situation semblable se trouvent les derniers deux populations, celles de la Peştera din Piatra Ponorului (PP) et la Peştera din Dealul Humpleu (DH). Du fait qu’elles sont liées entre elles à un niveau de similitude parfaitement comparable avec celui établi pour l’échantillon Al, on serait en droit de les considérer comme formant ensemble une sous-espèce distincte. Mais cette fois aussi une telle option est contraire au critère biogéographique.
5. ANALYSE DES GROUPEMENTS PAR CLASSIFICATION HIÉRARCHIQUE
Le dendrogramme fournit par le logiciel STATISTICA à partir des mêmes données primaires que celles utilisées dans l’analyse numérique (Fig. 6) est très

11 Révision des Leptodirinae souterrains du bassin de Someşul Cald
21
semblable au précédent. Il ne comporte en effet que deux différences de structure. La première est que l’échantillon ZO s’associe maintenant à la tétrade PA–Al, position qui le rend moins isolé mais qui correspond en plus grande mesure à la réalité géographique. La seconde concerne l’échantillon PG, lié non pas au groupe PA–Al, mais au couple PP–DH, et cette liaison est elle aussi plus adéquate au même point de vue. Pour le reste, l’ordre différent dans lequel se succèdent les échantillons dans les deux diagrammes n’est que la conséquence normale et nécessaire du fait que, à l’encontre de l’analyse hiérarchique, le dendrogramme construit par analyse numérique représente la variante isomérique optimale, c’est-à-dire celle dans laquelle cette succession est établie en tenant compte de la similitude entre les échantillons voisins, mai appartenant à des groupes différents (les rapports de voisinage) (Şerban et al., 1979).
Fig. 6. – Dendrogramme global obtenu par classification hiérarchique des groupements
(en ordonnée, distance euclidienne).
6. ANALYSE EN COMPOSANTES PRINCIPALES
La projection dans le plan factoriel (1–2) reflète d’une manière acceptable la configuration du nuage statistique inscrit dans l’espace multidimensionnel, les deux premiers facteurs exprimant 76,5% de la variance totale. L’examen des corrélations

Gheorghe Racoviţă 12
22
multiples montre toutefois que la position de l’échantillon ZO n’est correctement reproduite que si on se rapporte au troisième facteur aussi, auquel revient 16,6% de la variance.
6.1. ANALYSE DANS L’ESPACE DES VARIABLES
Énumérées par ordre décroissant de leur saturation, les variables qui ont le poids le plus important dans la détermination des trois premiers axes factoriels sont:
– le (0,9720), Lp (0,9707), lMp (0,9278) et Lc (0,9194) pour le premier axe, – lap/lbp (0,9295) et lbp/lMp (0,9257) pour le deuxième axe et – La/Lc (0,5108) et lMp/le (0,4562) pour le troisième axe.
Tel qu’on le sait, ces variables sont aussi celles qui se placent le plus près de la circonférence du cercle des corrélations (Fig. 7). Par ailleurs, la distribution dans ce cercle de l’ensemble des variables suggère que les axes qui définissent le plan de projection peuvent être considérés comme des «facteurs formes», malgré la tendance manifeste des caractères dimensionnels de se grouper vers l’extrémité positive du F1.
Fig. 7. – Projection dans l’espace des variables selon le plan factoriel (1–2). Pour les caractères dimensionnels (cercles), abréviations dans le texte; pour les caractères morphométriques (carrés), les abréviations correspondant aux divers rapports sont:
lMP = Lp/lMp, laP = lap/lMp, lmP = lmp/lMp, lbP = lbp/lmp,
abP = lap/lbp, lE = Le/le, LA = La/Lc, lPE = lMp/le et
lbM = lbp/lMp.
6.2. ANALYSE DANS L’ESPCAE DES UNITÉS STATISTIQUES
La visualisation selon le plan factoriel (1–2) de la position des échantillons dans le nuage statistique (Fig. 8) confirme la plupart des faits relevés par les deux autres méthodes d’analyse multifactorielle, en ajoutant en même temps des éléments nouveaux et bien utiles. En effet, ce qu’on remarque en premier lieu dans cette représentation graphique est la position parfois encore plus isolée sur laquelle se

13 Révision des Leptodirinae souterrains du bassin de Someşul Cald
23
Fig. 8. – Projection dans le plan des unités statistiques selon le plan factoriel (1–2).
Abréviations dans le texte.
Fig. 9. – Projection dans le plan des unités statistiques selon le plan factoriel (1–3).

Gheorghe Racoviţă 14
24
placent les populations d’Izbucul din Alunul Mare (AM), Peştera Grama (PG), Peştera din Piatra Ponorului (PP) et Peştera din Dealul Humpleu (DH). L’idée que chacune de ces populations peut être attribuée à une sous-espèce distincte est ainsi entièrement confirmée. En opposition évidente avec ces quatre échantillons se trouve le groupement compact qui, outre la tétrade déjà mentionnée, renferme aussi la population de la grotte d’Onceasa (ZO). Nous venons cependant de noter que la position de celle-ci n’est correctement définie que si on se rapporte au troisième axe. Or, la visualisation selon le plan factoriel (1–3) montre bien que cette population s’écarte des autres au mois dans la même mesure que celle d’Izbucul din Alunul Mare (Fig. 9). Il devient ainsi certain qu’il s’agit une fois de plus d’une sous-espèce à part.
Ajoutons pour mémoire que l’essai d’appliquer l’analyse factorielle discriminante a échoué dans absolument toutes les variantes testées, le plus vraisemblablement à cause du trop grand nombre de groupes définis par une seule unité statistique.
7. DIFFÉRENCIATION TAXONOMIQUE DES POPULATIONS
Compte tenu des résultats concordants auxquels ont abouti les méthodes de traitement statistique des données biométriques, on peut affirmer sans risque d’erreur que les populations qui font l’objet de cette étude appartiennent aux sous-espèces suivantes, dont trois sont nouvelles pour la science:
− Pholeuon (s. str.) angusticolle alunensis n. ssp. – Izbucul din Alunul Mare; − Pholeuon (s. str.) angusticolle angusticolle Hampe, 1856 – Peştera Zmeilor de
la Onceasa; − Pholeuon (s. str.) angusticolle mihoki Csiki, 1911 – Peştera de la Alun
(station-type), Peştera Diaclază, Peştera lui Teo, Poarta Alunului et Peştera cu Oase;
− Pholeuon (s. str.) angusticolle longicornis n. ssp. – Peştera din Piatra Ponorului;
− Pholeuon (s. str.) angusticolle arpadi Csiki, 1912 – Peştera din Valea Firii (station-type) et Peştera din Dealul Humpleu;
− Pholeuon (s. str.) angusticolle gujai n. ssp – Peştera Grama.
Avant de préciser les caractères discriminants de ces sous-espèces il s’impose de faire plusieurs précisions.
D’abord, qu’à la liste ci-dessus on doit certainement ajouter Pholeuon (s. str.) angusticolle bihariense Csiki, 1912, d’une part parce que nous avons déjà mentionné que MOLDOVAN (1989) a confirmé sa validité par des méthodes de même quantitatives, et d’autre part, parce que la position géographique de sa

15 Révision des Leptodirinae souterrains du bassin de Someşul Cald
25
station-type, l’Izbucul de la Rădeasa (Fig. 1), justifie un tel point de vue. Pour des raisons analogues, la population de la Peştera cu Oase peut être inclue dans la sous-espèce mihoki. Quant à la population de la Peştera Colţului, considère comme appartenant à la même sous-espèce, nous sommes d’avis que son statut est plutôt incertain, en premier lieu à cause du fait que la cavité en question est plus éloignée et proche de celle d’Alunul Mare.
Ensuite, qu’il est raisonnable de considérer que la grotte de Humpleu et peuplée par la même sous-espèce que la grotte de Valea Firii, car tel qu’Orăşeanu (1996) l’a démontré par marquage des drainages souterrains, les deux cavités font partie du même système karstique. Malheureusement, les caractères de diagnose qu’on peut déduire de la clé dichotomique établie par JEANNEL (1923) ne nous fournissent aucun autre argument.
Enfin, que considérer la Peştera din Piatra Ponorului comme habitée par une sous-espèce autre que celle cantonnée dans les grottes avoisinantes n’est point une opinion hasardée du moment que la grotte d’Onceasa se trouve exactement dans la même situation. De plus, un problème similaire s’est posé dans le bassin de l’Arieş aussi, à l’égard des cavités situées sur le plateau karstique de Scărişoara (RACOVIŢĂ, 2004–2005).
Pareillement à ce que nous avons constaté dans le cas de Pholeuon (s. str.) knirschi, le critère de différenciation infra-spécifique utilisé dans la révision du sous-genre Parapholeuon (RACOVIŢĂ, 1996) ne fournit que peu de différences significatives au niveau de sous-espèce et en même temps valables pour les deux sexes et pour toutes les populations d’une sous-espèce donnée. Rappelons que ce critère imposait que 75% des individus d’un échantillon A diffèrent de 85% des individus d’un échantillon B. Plus ou moins prévisible puisqu’il ne s’agit que d’une autre espèce de Pholeuon (s. str.), l’inconvénient a été dépassé en prenant en considération y compris les différences significatives à un seuil de confiance de 0,1%, quand 75% des individus d’un échantillon A diffèrent d’environ 42% des individus d’un échantillon B (lorsque les courbes de distribution sont symétriques, ces deux fractions deviennent égales, d’environ 60%). C’est d’ailleurs le motif évident pour lequel, comme dans le cas présent aussi, le degré de similitude entre des populations attribuées à des sous-espèces différentes a été parfois plus élevé que d’ordinaire.
Il va sans dire que l’application d’une semblable solution pourrait donner lieu à des objections. Mais il est tout aussi vrai que la seule alternative envisageable est de faire complètement abstraction du critère biogéographique et de supprimer absolument toutes les sous-espèces de Pholeuon angusticolle, ce qui est encore plus douteux. Nous allons donc procéder dans cette occurrence aussi à la comparaison successive d’une sous-espèce avec toutes les autres, les principaux paramètres statistiques des caractères quantitatifs discriminants étant inscrits dans le Tableau 4

Gheorghe Racoviţă 16
26
Tableau 4
Moyennes arithmétiques et écarts standard de la moyenne (p = 1%) des caractères discriminants
Caractères alunensis angusticolle mihoki longicornis arpadi gujai Mâles
Lc (mm) 4,518 ± 0,026 4,327 ± 0,024 4,382 ± 0,025 4,304 ± 0,025 4,282 ± 0,050 4,274 ± 0,062 Lp/lMp 0,948 ± 0,006 0,969 ± 0,006 – 0,967 ± 0,006 – – lMp/le 0,621 ± 0,003 0,614 ± 0,004 0,635 ± 0,004 0,642 ± 0,004 0,636 ± 0,008 0,647 ± 0,008 lap/lMp 0,661 ± 0,004 0,665 ± 0,006 0,650 ± 0,004 0,667 ± 0,004 0,651 ± 0,008 0,644 ± 0,008 lmp/lMp – 0,869 ± 0,004 – – 0,868 ± 0,007 0,891 ± 0,009 lbp/lMp 0,879 ± 0,005 0,894 ± 0,005 0,887 ± 0,005 0,872 ± 0,005 – 0,905 ± 0,009 lbp/lmp 1,019 ± 0,004 1,028 ± 0,005 1,018 ± 0,004 1,007 ± 0,006 – – lap/lbp – 0,745 ± 0,005 0,733 ± 0,005 0,765 ± 0,006 0,739 ± 0,010 0,712 ± 0,009 Le/le 1,641 ± 0,008 1,627 ± 0,008 1,675 ± 0,009 1,713 ± 0,008 1,699 ± 0,019 1,663 ± 0,016 La/Lc 0,682 ± 0,003 0,666 ± 0,004 – 0,703 ± 0,004 0,693 ± 0,006 0,688 ± 0,009
Femelles Lc (mm) 4,763 ± 0,028 4,599 ± 0,024 4,609 ± 0,026 4,517 ± 0,025 4,498 ± 0,051 4,523 ± 0,068 Lp/lMp 0,920 ± 0,005 0,940 ± 0,005 – 0,941 ± 0,005 – – lMp/le 0,610 ± 0,004 0,602 ± 0,003 0,616 ± 0,003 0,624 ± 0,004 0,623 ± 0,008 0,629 ± 0,009 lap/lMp 0,659 ± 0,003 0,662 ± 0,003 0,656 ± 0,003 0,667 ± 0,005 0,655 ± 0,006 0,643 ± 0,013 lmp/lMp – 0,880 ± 0,004 – 0,883 ± 0,007 0,898 ± 0,013 lbp/lMp 0,888 ± 0,006 0,904 ± 0,006 0,900 ± 0,005 0,880 ± 0,006 – 0,911 ± 0,018 lbp/lmp 1,018 ± 0,004 1,028 ± 0,006 1,014 ± 0,004 0,993 ± 0,005 – – lap/lbp – 0,733 ± 0,005 0,729 ± 0,004 0,758 ± 0,005 0,730 ± 0,009 0,706 ± 0,019 Le/le 1,622 ± 0,006 1,607 ± 0,006 1,652 ± 0,008 1,685 ± 0,008 1,681 ± 0,013 1,662 ± 0,015 La/Lc 0,599 ± 0,003 0,589 ± 0,003 – 0,607 ± 0,003 0,597 ± 0,004 0,586 ± 0,011
Comparaison de P. a. alunensis avec:
− P. a. angusticolle: taille plus grande; pronotum moins allongé (Lp/lMp inférieur) et relativement plus large que les élytres (lMp/le supérieur), avec la base plus étroite (lbp/lMp inférieur) et les côtés moins divergents en arrière dans leur tiers postérieur (lbp/lmp inférieur); antennes relativement plus longues (La/Lc supérieur).
− P. a. mihoki: taille plus grande; élytres plus larges (Le/le inférieur). − P. a. longicornis: taille plus grande; pronotum moins allongé (Lp/lMp inférieur)
et relativement plus étroit que les élytres; élytres plus larges; antennes relativement plus courtes.
− P. a. arpadi: taille plus grande; pronotum relativement plus étroit que les élytres; élytres plus larges.
− P. a. gujai: taille plus grande; pronotum relativement plus étroit que les élytres, avec le bord antérieur plus large (lap/lMp supérieur) et la base plus étroite.

17 Révision des Leptodirinae souterrains du bassin de Someşul Cald
27
Comparaison de P. a. angusticolle avec:
− P. a. mihoki: pronotum relativement plus étroites que les élytres, avec le bord antérieur plus large; élytres plus larges; antennes relativement plus courtes.
− P. a. longicornis: pronotum relativement plus étroit que les élytres, moins rétréci à la base, avec les côtés plus divergents dans leur tiers postérieur et la base relativement plus large que le bord antérieur (lap/lbp inférieur); élytre plus larges; antennes relativement plus courtes.
− P. a. arpadi: pronotum relativement plus étroit que les élytres; élytres plus large; antennes relativement plus courtes.
− P. a. gujai: pronotum relativement plus étroit que les élytres, avec le bord antérieur plus large, les côtés plus profondément sinués et la base relativement plus étroite que le bord antérieur; élytres plus larges.
Comparaison de P. a. mihoki avec:
− P. a. longicornis: taille plus grande; pronotum moins rétréci à la base, avec le bord antérieur plus étroit, les côtés plus divergentes dans leur tiers postérieur et la base relativement plus large que le bord antérieur; élytres plus larges; antennes relativement plus courtes (à l’exception de la population de la Poarta Alunului).
− P. a. arpadi: taille plus grande; élytres plus larges. − P. a. gujai: pronotum plus rétréci à la base (à l’exception des femelles de la
Peştera lui Teo et de celles de la Poarta Alunului), avec le bord antérieur relativement plus large que la base.
Comparaison de P. a. longicornis avec: − P. a arpadi: pronotum plus rétréci à la base, avec le bord antérieur plus large. − P. a. gujai: pronotum moins rétréci à la base, avec le bord antérieur plus
large et la base relativement plus étroite que celui-ci; élytres plus étroits; antennes relativement plus longues.
Comparaison de P. a. arpadi avec:
− P. a. gujai: pronotum moins rétréci à la base, avec le bord antérieur relativement plus large par que celle-ci.
Outre ces caractères purement quantitatifs, il convient de noter en tant qu’élément discriminant que les côtés du pronotum sont parfois moins arrondis dans leur partie antérieure, par suite du fait que le rebord est ici plus ou moins effacé. Remarquée premièrement par Jeannel (1923) chez P. a. angusticolle, cette particularité morphologique se rencontre également chez P. a. alunensis.
Quant au pronotum «type», figuré à partir des valeurs moyennes que les caractères dimensionnels prennent pour chacune des six sous-espèces (Fig. 10), il est visiblement moins variable que celui des individus ayant les valeurs extrêmes

Gheorghe Racoviţă 18
28
des caractères morphométriques mais provenant de l’une et la même population (Fig. 2). Une fois de plus, il s’agit d’une constatation déjà faite chez P. knirschi de la zone de Padiş et qui a toutes les chances de pouvoir être généralisée au moins pour toutes les espèces de Pholeuon.
Fig. 10. – Pronotum «type» de six sous-espèces de P. angusticolle.
Cette variabilité si forte et que nous avons évoquée en maintes reprises se manifeste en égale mesure par des traits morphologiques qui ne peuvent plus être exprimés à l’aide des paramètres biométriques. Dans le cas du pronotum, il s’agit du fait qu’à l’intérieur de l’un et même échantillon, les côtés sont soit divergentes, soit parallèles, soit convexes vers leur extrémité postérieure. Bien plus, des asymétries assez nombreuses ont été observées à ce même point de vue, surtout chez les femelles d’Izbucul din Alunul Mare et de la Peştera lui Teo. A leur tour, les élytres peuvent être aussi bien elliptiques qu’ovales, avec la ponctuation homogène sur toute leur surface ou plus grossière dans la partie apicale. Il est donc hors de toute incertitude que l’aspect de la ponctuation a été employé à tort par JEANNEL (1923, 1924) pour différencier P. knirschi de P proserpinae.
Signalons en terminant qu’une femelle parasitée sur la face dorsale de son pronotum par un champignon (?) a été trouvée dans la population de Poarta Alunului.

19 Révision des Leptodirinae souterrains du bassin de Someşul Cald
29
REMERCIEMENTS. Ce travail n’aurait certainement pas pu être réalisé sans l’extrême obligeance de M. Ovidiu Guja, qui c’est volontairement chargé de prélever des échantillons de coléoptères cavernicoles des grottes qu’il a explorées dans le bassin de Someşul Cald. C’est pourquoi nous lui avons dédiée la sous-espèce de la grotte de Grama, en le priant de croire à notre vive et sincère reconnaissance.
B I B L I O G R A P H I E
DECU, V., Le catalogue des Coléoptères cavernicoles de Roumanie (Coleoptera). Acta Zool. Cracov., IX (7), 1964, 441–467.
GORAN, C., Catalogul sistematic al peşterilor din România. Edit. Cons. Naţ. Ed. Fiz. şi Sport, Bucureşti, 1982.
JEANNEL, R., Étude préliminaire des Coléoptères aveugles du Bihor. Bull. Soc. Sc. Cluj, I, 1923, 411–472.
JEANNEL, R., Monographie des Bathysciinae. Arch. Zool. exp. gén., 63 (1), 1924, 1–436. MOLDOVAN, O., Révision systématique de l’espèce Pholeuon (s. str.) angusticolle (Coleoptrea,
Catopidae, Bathysciinae). Trav. Inst. Spéol. «E. Racovitza», XXVIII, 1989, 29–41. MOLDOVAN, O., CÎMPEAN, M., BORDA, D., IEPURE, S., ILIE, V. (eds.), Lista faunistică a
României (Checklist of Romania Fauna). Edit. Casa Cărţii de Ştiinţă, Cluj-Napoca, 2007, 412 pp. ORĂŞANU, I., Contributions to the hydrogeology of the karst areas of the Bihor–Vlădeasa
Mountains (Romania). Theor. Appl. Karstol., 9, 1996, 185–214. PERREAU, M., Catalogue des Coléoptères Leiodidae Cholevinae et Plathypsyllinae. Mém. Soc. entomol.
Fr., 4, 2000, 460 pp. RACOVIŢĂ, G., Révision systématique des Bathysciinae souterrains des Monts Apuseni. I. Variabilité
individuelle et valeur taxonomique des caractères morphologiques dans la série phylétique de Drimeotus (Coleoptera, Bathysciinae). Trav. Inst. Spéol. «E. Racovitza», XXXIV, 1995, 103–129.
RACOVIŢĂ, G., Révision systématique des Leptodirinae souterrains des Monts Apuseni. II. Le sous-genre Parapholeuon Ganglb. du bassin de Crişul Repede (Monts Pădurea Craiului). Trav. Inst. Spéol. «E. Racovitza», XXXV, 1996, 69–105.
RACOVIŢĂ, G., Révision systématique des Leptodirinae souterrains des Monts Apuseni. IV. Le sous-genre Pholeuon (s. str.) du bassin de l’Arieş (Monts du Bihor). Trav. Inst. Spéol. «E. Racovitza», XLIII–XLIV, 2004–2005, 165–191.
RACOVIŢĂ, G., Révision systématique des Leptodirinae souterrains des Monts Apuseni. V. Le sous-genre Pholeuon (s. str.) de la zone de Padiş (Monts du Bihor). Trav. Inst. Spéol. «E. Racovitza», XLV–XLVI, 2006–2007, 13–30.
SNEATH, P.H.A., The application of computers to taxonomy. J. Gen. Microbiol., 17, 1957, 201–226. ŞERBAN, M., NEAGU, LIVIA, ALB, MARIA, RACOVIŢĂ, G., Application des méthodes numériques à
la systématique des Harpacticoïdes. 1. Analyse des formules d’armature des pattes natatoires en tant que critère taxonomique. Trav. Inst. Spéol. «E. Racovitza», XVIII, 1979, 33–52.
Institut de Spéologie «E. Racovitza» Département de Cluj-Napoca
Str. Clinicilor nr. 5, 400006 Cluj-Napoca, Roumanie