
V10-EF hele proefschrift - WUR
212
Transcript of V10-EF hele proefschrift - WUR



Eggsposed
Impact of maternally transferred POPs
on fish early life development
Edwin M. Foekema

Thesis committee
Promotor Prof. Dr. A.J. Murk Personal chair at the Sub-department of Environmental Technology Wageningen University Other members Prof. Dr. A.J. Hendriks, Radboud University Nijmegen Prof. Dr. P.J. van den Brink, Wageningen University Prof. Dr. N.M. van Straalen, VU University of Amsterdam Dr. M.G.D. Smit, Shell Global Solutions International BV, Rijswijk This research was conducted under the auspices of the Graduate School SENSE (Socio-Economic and Natural Sciences of the Environment).

Eggsposed
Impact of maternally transferred POPs
on fish early life development
Edwin M. Foekema
Thesis Submitted in fulfilment of the requirements for the degree of doctor
at Wageningen University by the authority of the Rector Magnificus
Prof. Dr. M.J. Kropff, in the presence of the
Thesis Committee appointed by the Academic Board to be defended in public
on Friday 4 October 2013 at 13:30 p.m. in the Aula

Edwin M. Foekema Eggsposed. Impact of maternally transferred POPs on fish early life development, 210 Pages PhD Thesis, Wageningen University, Wageningen, NL (2013) With references, with summaries in Dutch and English ISBN 978-94-6173-665-9

Voor Gerda, Lars en Ivar “Study to be quiet” Izaak Walton; The Complete Angler, 1655 “Je gaat het pas zien als je het doorhebt” Johan Cruijff


Eggsposed
7
Table of contents
Introduction ........................................................................... 9 CHAPTER 1
1.1 POPs in marine environment ................................................... 10 1.2 Fish early life development and effects of POPs .......................... 13 1.3 The European eel, a special case ............................................. 17 1.4 The common sole as test species ............................................. 17 1.5 Thesis outline ....................................................................... 20 1.6 References ........................................................................... 22
Prolonged ELS test with the marine flatfish sole (Solea solea) shows CHAPTER 2delayed toxic effects of previous exposure to PCB 126 ................ 29
2.1 Introduction ......................................................................... 31 2.2 Materials and methods ........................................................... 32 2.3 Results ................................................................................ 35 2.4 Discussion ............................................................................ 41 2.5 Conclusions .......................................................................... 44 2.6 References ........................................................................... 44
Critical body burden of maternally transferred methyl-triclosan for CHAPTER 3the flatfish Solea solea ........................................................... 49
3.1 Introduction ......................................................................... 51 3.2 Materials and methods ........................................................... 52 3.3 Results and discussion ........................................................... 53 3.4 References ........................................................................... 55
Toxic concentrations in fish early life stages peak at critical momentCHAPTER 4 .......................................................................................... 57
4.1 Introduction ......................................................................... 59 4.2 Material and methods ............................................................ 61 4.3 Results ................................................................................ 66 4.4 Discussion ............................................................................ 72 4.5 References ........................................................................... 77 Annex to Chapter 4 ........................................................................ 81
The risks of maternally transferred organic substances for fish early CHAPTER 5life development ................................................................... 83
5.1 Introduction ......................................................................... 85 5.2 Materials and methods ........................................................... 87

Eggsposed
8
5.3 Results ................................................................................ 91 5.4 Discussion ............................................................................ 97 5.5 References ......................................................................... 104 Annex to Chapter 5 ...................................................................... 109
Is the reproduction success of European Eel affected by maternally CHAPTER 6transferred dioxin-like compounds? ....................................... 117
6.1 Introduction ....................................................................... 119 6.2 Data collection and calculations ............................................. 121 6.3 Risk assessment ................................................................. 128 6.4 References ......................................................................... 131
The potential impact of reduced larval survival on fish population CHAPTER 7development ...................................................................... 135
7.1 Introduction ....................................................................... 137 7.2 Model description ................................................................ 139 7.3 Results .............................................................................. 141 7.4 Discussion .......................................................................... 145 7.5 Conclusions ........................................................................ 146 7.6 References ......................................................................... 146
General discussion ............................................................... 149 CHAPTER 88.1 Introduction ....................................................................... 150 8.2 Observed effects in the prolonged ELS test ............................. 151 8.3 Simulation of maternal transfer in the p-ELS fish test ............... 155 8.4 Characteristics that determine sensitivity ................................ 160 8.5 Identification of vulnerable species ........................................ 163 8.6 Conclusions ........................................................................ 169 8.7 Future perspectives ............................................................. 170 8.8 References ......................................................................... 172
Summary ........................................................................................ 181
Nederlandse samenvatting (speciaal voor niet ingewijden) ........................ 187
Dankwoord ........................................................................................ 193
Curriculum Vitae .................................................................................. 199
Publications ........................................................................................ 201
SENSE Certificate ................................................................................. 209

9
CHAPTER 1
Introduction
“The old man knew he was going far out and he left the smell of the land behind and rowed into the clean early morning smell of the ocean.”
Ernest Hemmingway: The Old Man and the Sea

Chapter 1 Eggsposed
10
1.1 POPs in marine environment
Although the application and emission of major persistent organic pollutants (POPs) has been strongly reduced during the last decades, their mass production in the past in combination with their persistence make that these pollutants still are globally present in the marine environment (Moore et al., 2002).
As a result of their persistency and low water solubility (high lipophilicity), highest concentrations in the aquatic environment are found in the organic matrix of sediments and in biota, while dissolved concentrations in water are low. The major exposure route for lipophilic POPs is through food uptake (Moore et al., 2002). As a result of biomagnification the highest levels of POPs have been identified in the tissue of predators, like eel, swordfish, sharks and marine mammals (Boon et al., 2002; de Boer et al., 2010; Kannan et al., 2002; Law et al., 2006). POP concentrations are often expressed on whole tissue bases, mostly to assess the risk for human exposure by (fish) consumption (Domingo and Bocio, 2007; Dorea, 2008; Gomara et al., 2005; van Leeuwen and de Boer, 2008). Lipid-rich fish generally have higher POP tissue concentrations. However, from a toxicological point of view the lipid normalised concentrations are more relevant to assess the risk for a toxic effect on the organism (Murk et al., 1998). The lipophilicity of a compound is expressed based on the equilibrium distribution of the substance between water and octanol, usually as the logarithm of the octanol-water partitioning coefficient (log Kow). The log Kow of lipophilic POPs roughly ranges between 4 and 7 (Hansen et al., 1999; Li et al., 2008). A log Kow of 7 indicates that in an equilibrium situation the concentration of this compound in the octanol phase will be 107 times higher than the concentration in the water phase.
Well known lipophilic POPs that are ubiquitous in the marine environment are polychlorinated biphenyls (PCBs) and brominated flame retardants such as polybrominated diphenyl ethers (PBDEs) and hexabromocyclododecane (HBCD). High volume production of PCBs started around 1930 and ceased around the 1990s. PCBs were applied especially in hydraulic fluids, electric transformers and as flame retardants (Breivik et al., 2004). They now are present in biota all over the world (Domingo and Bocio, 2007). In total 209 PCB congeners exist depending on the chlorine number and positions in the biphenyl molecule. The more chlorinated non- and mono-ortho substituted PCB congeners form the most persistent molecules (Beyer and Biziuk, 2009), and show toxic features comparable to polychlorinated dibenzo dioxins (PCDDs) and polychlorinated dibenzofurans (PCDFs), with tetrachlorinated dibenzodioxin (TCDD) as most toxic representative. For that reason non- and mono-substituted PCBs are often referred to as dioxin-like PCBs. The potency of the individual congeners to cause

Eggsposed Chapter 1
11
dioxin-like toxic effects depends on the molecular structure and is expressed as TCDD TEF (Toxic Equivalency Factor). The non-ortho PCB 126 with a TEF of 0.1 is the most potent dioxin-like PCB (Van den Berg et al., 1998; Van den Berg et al., 2006). Together with PCDD/Fs the dioxin-like PCBs add to the total dioxin-like toxic pressure that is be expressed as TEQ (TCDD Equivalent) (Ahlborg et al., 1994; Van den Berg et al., 1998; Van den Berg et al., 2006). Since differences in the sensitivity of taxonomic groups for dioxins have been found, TEF-systems have been proposed for mammals, fish, and birds (Van den Berg et al., 1998). In 2005, the TEF system for mammals was revised which resulted in lower TEF values for some PCBs (Van den Berg et al., 2006). The choice of the TEF-system to use thus has consequences for the resulting TEQ. As the far majority of the research on the presence of dioxin-like toxicity in fish focuses on the risk for human consumption, toxic potencies normally are expressed using the WHO-TEF system for mammals. These TEQ values however, overestimate the contribution of mono-ortho PCBs to the overall dioxin-like toxicity for fish.
The highest PCB concentrations in fish were reported around the 1980s in samples from the Great lakes, e.g. rainbow trout containing 4.8 µg/g wet weight (ww) (42 ug/g lipid weight (lw)) and white sucker 3.4 ug/g ww (55 µg/g lw) of total PCBs (no data on congeners given) respectively (Niimi, 1983), and lake trout ranging from 3.5 to 13.9 µg/g ww (Mac et al., 1993). Analyses of historical monitoring data from the year 1977 to 2006 revealed that PCB concentrations in eel in the Netherlands declined relatively slow during this 30 year period with a factor 5 to 10 (de Boer et al., 2010). The maximum PCB concentration, 48 µg/g lw (∑7 congeners), was measured in a sample from 1979. This value is comparable to values reported in rainbow trout and white sucker in the Great Lakes in 1983 (Niimi, 1983). The data-set of (de Boer et al., 2010) suggest that even higher PCB concentrations in eel tissue must have been present before 1977. In the year 2006 the PCB concentrations in eel from the same water basins ranged between 5 and 8 µg/g lw (de Boer et al., 2010). Other recent studies reported total PCB levels in Swordfish: 1.0 µg/g lw (∑57 congeners, (van Leeuwen et al., 2007), tuna: 1.4 µg/g lw (∑23 congeners, (Gomara et al., 2005)), salmon: 3.2 µg/g lw (∑19 congeners, (Svendsen et al., 2007)) Goby: 4 µg/g lw (∑25 congeners, (Voorspoels et al., 2004), Greenland shark 5.8 µg/g lw (Molde et al., 2013), and in brown trout after a migratory starving period: 12.6 µg/g lw (∑19 congeners, (Svendsen et al., 2007).
The highest published levels of dioxin-like pollutants in fish include 596 pg TEQ/g lw in pike from the Baltic sea (Isosaari et al., 2006), 3764 pg TEQ /g lw in flounder from the Greenland fjord (Knutzen et al., 2003). In both cases TEF systems for mammals were applied to calculate the reported TEQ values. In

Chapter 1 Eggsposed
12
Greenland sharks, the TEQ concentrations based on the TEF system for fish were reported to range from 460 to 579 pg TEQ/g (Storelli et al., 2011).
Polybrominated diphenyl ethers (PBDEs) are flame retardants with a chemical structure comparable to PCBs, but with chlorine replaced by bromine. As with PCBs 209 PBDE congeners can exist. Market introduction of PBDEs took place about 50 years ago, and although application of PBDEs now is restricted and shifted towards the higher brominated ones, these substances are still globally present (Borghesi et al., 2009; Hites, 2004). The European Food Safety Authority recommended monitoring of the (sum)concentrations of BDE 28, 47, 99, 100, 153, 154, 183 and 209 (EFSA, 2006). In fish samples the PBDE profile is often dominated by the poorly degradable BDE 47 (Voorspoels et al., 2003) (Borghesi et al., 2009; van Leeuwen and de Boer, 2008). In the Dutch eel monitoring programme PBDE 47 concentrations in eel tissue is measured since 1984. The maximum concentrations around 1.5 µg/g lw were recorded during the first years. Concentrations decreased 4 to 10 times a maximum concentration of 0.2 µg/g lw in 2006 (de Boer et al., 2010). Some of the highest PBDE levels that have been reported in fish muscle samples are 0.26 µg/g lw in salmon (Svendsen et al., 2007), 0.21 µg/g lw in bream (Isosaari et al., 2006) and 0.64 and 0.74 µg/g lw in feral carp from two Spanisch rivers (Labandeira et al., 2007).
Hexabromocyclododecanes (HBCD) also are brominated flame retardants, which mainly are applied in expanded and extruded polystyrene. HBCDs have been produced and used for about three decades. They were first detected in fish at the end of the 20th century and are now ubiquitous pollutants in the environment (Covaci et al., 2006). A UN-expert group recently (Oct. 2012) recommended that HBCD be eliminated from the global market to protect human health and the environment (http://chm.pops.int). HBCDs can be found in alpha, beta and gamma diastereomers of which alpha-HBCD is the dominant (>75%) congener present in fish tissue (Covaci et al., 2006; van Leeuwen and de Boer, 2008). Extremely high concentrations of 4 to 8 µg/g lw HBCD are reported in the tissue of pike sampled near point sources (textile industries) in Sweden (Sellstrom et al., 1998). In the Western Scheldt estuary concentrations up to 1.1 µg/g lw were detected in muscle of sole (Janak et al., 2005).
A wide variety of effects on fish and other taxa have been related to these elevated POP tissue concentrations, often indicating early development as the most sensitive life stage (Hutchinson et al., 1998; McKim, 1977).

Eggsposed Chapter 1
13
1.2 Fish early life development and effects of POPs
1.2.1 Fish reproduction and early development
The majority of the teleost fish species are ovo-parous with an iteroparous life strategy (Wooton, 1992), indication that an individual fish can produce multiple batches of eggs during a lifetime. This, in contrast to semelparous fishes that reproduce only once in their lifetime and die after spawning. Well known examples of the latter are eels and pacific salmon that undertake a long migration to the spawning grounds that are located in a completely different environment than where the fish reach maturity. Eels mature in freshwater and spawn at sea, for the pacific salmon species this is vice versa.
Although there are many exceptions, in general fish spend little energy on nursery of eggs and larvae. Especially in the marine environment most fish spawn in the pelagic zone where the eggs develop being suspended in the water column. In the freshwater, more species produce demersal eggs or eggs that attach to plants or other substrates (Wooton, 1992). Some teleost fish species are ovo-viviparous, indicating that eggs are fertilised and develop inside the mother’s body.
The reproduction success of a fish depends, as for other species, on the number of surviving offspring it can produce during its life-time. Bigger eggs have the advantages of producing bigger larvae that can take a wider range of prey, are less vulnerable to predation and better in withstanding periods of food shortage than smaller larvae. The majority of fish without parental care, however, produce small eggs in sometimes extremely high numbers (Wooton, 1992). A large number of offspring reduces the impact of predation and increases the chance of larvae encountering food and favourable conditions (Rothschild, 2000). Smaller clutch size and larger eggs are typically found in demersal spawners like salmonids, and species that show some form of parental care.
An egg is provided with all resources necessary for the development of the embryo into a larva until it is capable of external feeding. The majority of these resources is stored by the mother in the yolk during vitellogenesis. A review of this process is given by Mommsen and Walsh (1988). The protein vitellogenin is released from the liver in the blood stream and efficiently absorbed by the developing oocytes, where it is deposited in the yolk. Due to strong ion-binding properties of vitellogenin, it can act as an important vehicle for minerals to the oocyte. Vitellogenin itself consist in general of proteins and polar and neutral lipids (Johnson, 2009). Polar lipids (phospholipids) have a structural function in animal cells and are essential in cell membranes and neural tissue. Neutral lipids

Chapter 1 Eggsposed
14
are important as sources of energy and probably serve as storage place for essential fatty acids (Wiegand, 1996).
For species that do not feed during vitellogenesis like salmon and eel, the source of the lipids deposited in the eggs are the adipose reserves stored by the female fish. For other species dietary lipids can form an (additional) source.
After fertilisation, the egg chorion hardens as a protective shield allowing only limited exchange of water and molecules (Kamler, 2008). In many marine species the larvae hatch at an early stage of development. The mouth and jaws are not formed, the eye is not pigmented and a primordial finfold runs around the trunk in the median position. Most freshwater species and some marine species hatch at a further developed state, but newly hatched fish larvae are never ready to start external feeding. During the first days after hatching (the yolk-sac stage) the larvae still depend on the yolk resources. The mouth and intestine develop and enable the larvae to switch to exogenous feeding when the yolk is depleted. The duration of the yolk-sac stage differs per species. The Japenese Medaka fish can reach the free feeding state already one day after hatching (Villalobos et al., 2000), while for other species this will take several days. It has been suggested that the duration of the yolk-sac stage is related with egg size and temperature (Blaxter, 1988).
During the free-feeding stage organs and tissues develop further, and the larvae gain length and weight. At a species-dependent age, the free-feeding larvae metamorphose into a juvenile with the same appearance as the adult fish (Blaxter, 1988). This metamorphoses takes place in all teleost fish, but is most obvious in flatfish as larvae of this group undergo a drastic craniofacial remodelling (Figure 1.1), during which the symmetrical larvae with a pelagic life style change into a benthic asymmetrical juvenile (Schreiber, 2006).
1.2.2 Effects of POPs on Fish early life stages
POPs like dioxins, PCBs, PBDEs and HBCDs have the potential to cause a variety of toxic effects. From earlier research with fish larvae it is known that especially early life stages are very sensitive to the toxic action of dioxins, and dioxin-like substances such as PCDD/Fs and, and non- and mono-ortho PCBs of which the molecular structure can have a planar confirmation (Johnson et al., 1998; Walker and Peterson, 1994; Yamauchi et al., 2006). These substances are able to bind to and activate the aryl hydrocarbon (Ah) receptor, which binding has been associated with a variety of toxic responses including teratogenicity and changes in the thyroid function (Brouwer et al., 1999; Brown et al., 2004). A typical effect of dioxin-like toxicity in developing fish larvae is blue-sac disease that is characterised by pericardial and yolk-sac oedema and results in early

Eggsposed Chapter 1
15
mortality (Brinkworth et al., 2003; Hill et al., 2004; Stouthart et al., 1998; Zabel et al., 1995).
Some PCBs and brominated flame retardants have the potential to disrupt hormone systems. This includes the thyroid hormone system (Brouwer et al., 1999) that plays an essential role in metabolisms and differentiation during early life development, including metamorphosis. Exposure to these substances can therefore disturb the development of early life stages for a wide variety of taxa, as has been shown for echinoderms (Anselmo et al., 2012b), fish (Palace et al., 2010; Yu et al., 2011), and amphibians (Gutleb et al., 2007a; Gutleb et al., 2007b; Schriks et al., 2006).
Especially the hydroxylated (OH-) metabolites of PCBs (OH-PCBs) and PBDEs (OH-PBDEs) show a high potency for disruption of the thyroid hormone system as they have a structural similarity with thyroid hormones (Freitas et al., 2011; Zheng et al., 2012). The potency for hydroxylation of PCBs and PBDEs differs between species (Murk et al., 1994; Roberts et al., 2011). Juveniles of the common sole have been shown to metabolise PBDEs into OH-PBDEs (Munschy et al., 2010). It is however unclear during which stage of the development this ability develops. Embryos of Zebrafish did not form OH-PBDE 47 after exposure to PBDE 47 (Zheng et al., 2012).
Also for toxicants with less specific effects, the strongest individual responses to exposure are expected on early life stages (Hutchinson et al., 1998; McKim, 1977). This is believed to be related to the development of organs and tissues that can be disrupted easily, in combination with the lacking ability to compensate the impact of physiological processes affected by toxicant exposure (Crane et al., 2006). In addition, the small size and related large surface-volume ratio facilitates a faster increase of internal concentrations in fish larvae than in larger fish when exposed through the water phase.
1.2.3 Fish early life stage tests
As fish early life stages (ELS) are recognised as sensitive for toxic disruption, several test guidelines for fish ELS testing are available (OECD, 1992, 1998, 2006; USEPA, 1996). Despite this, guidance on the utilisation of marine species is scarce, especially with respect to species relevant for the West European region.
For multiple (practical, economical, and ethical) reasons the majority of the fish ELS tests that are applied are terminated just before or soon after the larvae become free-feeding, thus without covering the metamorphoses into a juvenile fish. In addition, none of the recommended test species undergoes an obvious

Chapter 1 Eggsposed
16
metamorphosis that is typical for certain taxonomical fish groups such as flatfish (Yamano, 2005). Tests with amphibians showed that developmental effects of thyroid hormone disrupting chemicals only become visual in prolonged ELS tests where metamorphosis is included (Gutleb et al., 2007a). For this reason a prolonged ELS test with a native West European marine species that undergoes obvious metamorphoses was developed and applied for the research presented in this thesis.
1.2.4 Exposure of fish early life stages to POPs in the natural environment
When the toxicity for fish larvae of a poor water soluble pollutant is being tested, most of the time a solvent/carrier is applied to maintain the water concentration of the test substance at the appropriate level. In the natural environment water concentrations of such lipophilic POPs will be very low. As the eggs and larvae of the far majority of marine fish species develop suspended in the water column, external exposure to POPs is very limited until the larvae become free feeding and might feed on contaminated food items.
Due to bioaccumulation of POPs during life, tissue concentrations can build up in the mother fish, and accumulated substances will be transferred with the yolk to the eggs (Hammerschmidt and Sandheinrich, 2005; Johnson et al., 1998; Johnston et al., 2001; Miller, 1993; Monteverdi and Di Giulio, 2000; Nyholm et al., 2008; Russell et al., 1999; Serrano et al., 2008; Wang et al., 2011; Yu et al., 2011). For POPs this results in concentrations in the eggs that on a lipid bases are comparable with the mother’s tissue (Russell et al., 1999). Via maternal transfer the developing embryo is thus exposed to relatively high levels of POPs, including those substances that may be specifically harmful during early life development. The importance of maternal transfer as exposure route has already been studied and recognised for dioxins and dioxin-like PCBs, especially in research with trout from the Great Lakes that contained extremely high PCB levels in the late 20th century (Mac et al., 1993; Niimi, 1983; Walker and Peterson, 1994). From this research it was concluded that the high concentration of PBCs contributed to the poor reproduction success of local trout population (Wilson and Tillitt, 1996).
The exposure rate of embryos and yolk-sac larvae to lipophilic POPs depends thus strongly on the concentration of these substances in the mother fish tissue, or actually in her lipids. Through bioaccumulation tissue concentrations can build op with time, and during periods of starvation the poor water soluble substances that can hardly be excreted will concentrate in the remaining tissues. This is for instance the case for fish that do not feed during spawning migration like salmon

Eggsposed Chapter 1
17
and eel, and has consequences for the amounts of POPs that are deposited in the eggs (Debruyn et al., 2004; Kelly et al., 2007).
1.3 The European eel, a special case
Around 1980 PCB concentrations were detected in the tissue of eel (Anguilla Anguilla) from the Netherlands that were among the highest ever recorded in fish (de Boer et al., 2010). In the same period the numbers of juveniles (glass eel) of the European eel that reach the European coasts started to decline rapidly and nowadays these are only 10% of what it used to be before the 1980’s (Dekker, 2003). A clear explanation for this extreme strong decline is not available at this moment; multiple causes are suggested, including effects of accumulated toxic pollutants.
The European eel combines different features that make it potentially vulnerable to the effects of POPs. After entering from sea as a glass eel, the eels remain in the fresh water for many years where they live in close contact with the sediment and its pore water. In the sediments the POPs accumulate to sometimes very high levels. The eel diet consisting of macro invertebrates and fish (Tesch, 2003), and their very high lipid content facilitates bioaccumulation of POPs. Eels do not reproduce during their stay in the freshwater, thus no POPs are excreted through spawning of lipid-rich eggs, which in some species results in reduced tissue concentrations (Madenjian, 2011). After spending many years in the freshwater environment the eels start a long marine migration to the spawning grounds that are believed to be located in the Sargasso Sea, at a distance of 5000 to 6000 km (Tesch, 2003). As they do not feed during migration weight loss will occur, probably leading to concentration of the POPs in the tissue lipids of the fish, conform observations in migrating pacific salmon (Debruyn et al., 2004; Kelly et al., 2011). In this case the POP concentrations in the tissue of the eel that has reached the spawning ground will exceed the concentration at the start of the migration, and these concentrations will be transferred to the eggs (Russell et al., 1999). Following this line of thinking the eel embryos will thus be exposed to the highest levels of lipophilic POPs that were ever present in the parent fish.
1.4 The common sole as test species
The common sole (Solea solea) is chosen as appropriate candidate for the development of a new fish test to study early life development including metamorphoses. It is a native European species of both ecological and economic importance, with landings ranging between 10 - 15 000 tons per year for the

Chapter 1 Eggsposed
18
North Sea (ICES FishMap Sole, http://www.ices.dk/ (accessed November 2012). Because sole is a valuable consumption fish, it is aquacultured and fertilised sole eggs were available at different moments throughout the year from the IMARES department of aquaculture.
All flatfishes (Order Pleuronectiformes) hatch as symmetrical larvae but undergo an obvious metamorphosis during early life development, that is characterised by the relocation of (in general) the left eye to the right side of the head (Figure 1.1). This metamorphosis is regulated by the thyroid hormone (Klaren et al., 2008). During the free-feeding stage the first thyroid follicles develop and the thyroid hormone tissue concentrations start to rise. The moment that the maximum thyroid hormone concentrations are reached coincides with the moment of metamorphosis (Figure 1.1). This is comparable to the metamorphosis of amphibians and sea urchins that has been shown to be easily disrupted by toxic compounds (Anselmo et al., 2012a; Gutleb et al., 2007a; Gutleb et al., 2007b). The early life development of flatfish is well studied by various researchers (Klaren et al., 2008; Martinez and Bolker, 2003; Schreiber, 2006), and the description of the different stages, including the metamorphosis is more or less standardised (Martinez and Bolker, 2003). The moment of metamorphoses marks the change from a pelagic to a benthic life style. At approximately 14°C, the eggs hatch 5-6 days after fertilisation (dpf), the larvae became free feeding around 10 dpf, and the first larvae show signs of the onset of metamorphoses around 25 dpf. Metamorphosis typically will be completed for normally developing individuals within 40 dpf. This rate of development is strongly temperature dependent. Sole reared at 18°C can complete the full development into a juvenile within 31 dpf (Palazzi et al., 2006). A full ELS developmental test with sole takes at least a 30-40 day observation period, which is still relatively short when compared with an amphibian test that requires more than 100 days to cover the full development (Gutleb et al., 2007a).
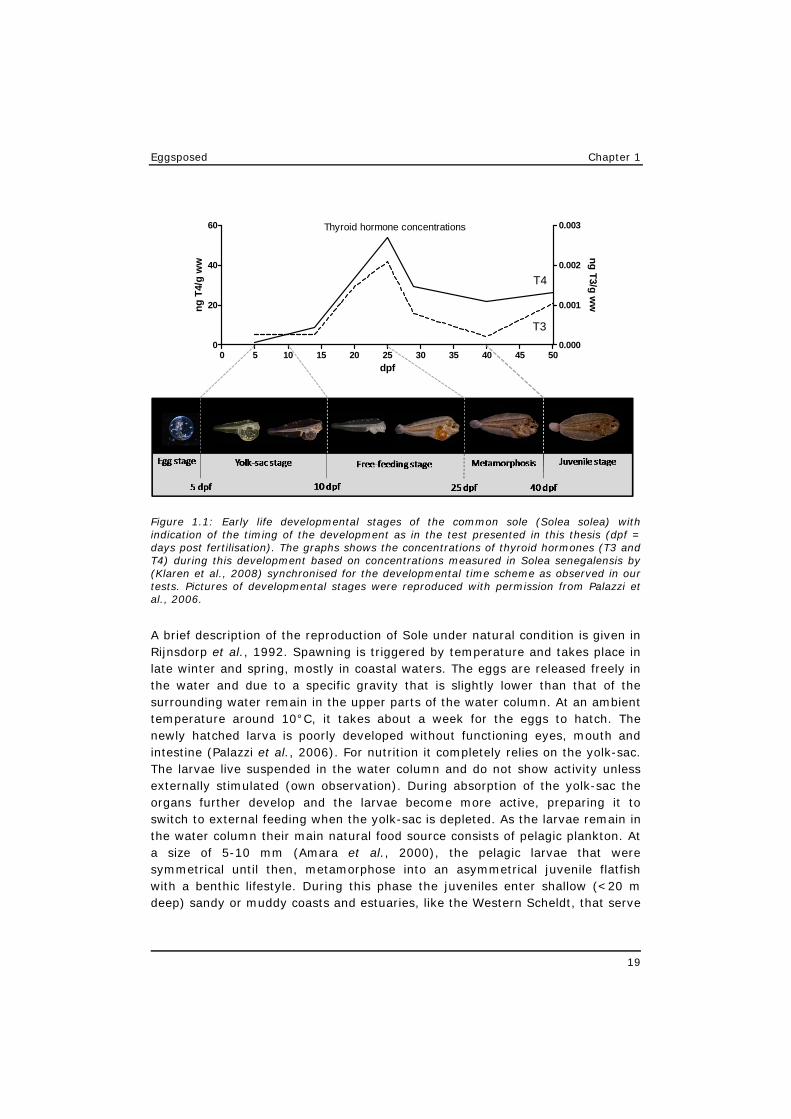
Eggsposed Chapter 1
19
Figure 1.1: Early life developmental stages of the common sole (Solea solea) with indication of the timing of the development as in the test presented in this thesis (dpf = days post fertilisation). The graphs shows the concentrations of thyroid hormones (T3 and T4) during this development based on concentrations measured in Solea senegalensis by (Klaren et al., 2008) synchronised for the developmental time scheme as observed in our tests. Pictures of developmental stages were reproduced with permission from Palazzi et al., 2006.
A brief description of the reproduction of Sole under natural condition is given in Rijnsdorp et al., 1992. Spawning is triggered by temperature and takes place in late winter and spring, mostly in coastal waters. The eggs are released freely in the water and due to a specific gravity that is slightly lower than that of the surrounding water remain in the upper parts of the water column. At an ambient temperature around 10°C, it takes about a week for the eggs to hatch. The newly hatched larva is poorly developed without functioning eyes, mouth and intestine (Palazzi et al., 2006). For nutrition it completely relies on the yolk-sac. The larvae live suspended in the water column and do not show activity unless externally stimulated (own observation). During absorption of the yolk-sac the organs further develop and the larvae become more active, preparing it to switch to external feeding when the yolk-sac is depleted. As the larvae remain in the water column their main natural food source consists of pelagic plankton. At a size of 5-10 mm (Amara et al., 2000), the pelagic larvae that were symmetrical until then, metamorphose into an asymmetrical juvenile flatfish with a benthic lifestyle. During this phase the juveniles enter shallow (<20 m deep) sandy or muddy coasts and estuaries, like the Western Scheldt, that serve
dpf
ng
T4/
g w
wn
g T
3/g w
w
0 5 10 15 20 25 30 35 40 45 500
20
40
60
0.000
0.001
0.002
0.003Thyroid hormone concentrations
T4
T3

Chapter 1 Eggsposed
20
as nursery areas. Here, the fish remain for the next one or two years, after which they take part in spawning for the first time and spend the rest of their life in deeper waters. The relative fecundity of the common sole in the North Sea ranges between 400 and 900 eggs per gram body weight, and the estimated reproductive efficiency, being the number of 1 year old fish (recruits) per egg produced, is estimated at approximately 2.5·10-6. There are strong indications that the number of recruits per stock is dictated by the surface of the available nursery areas (Rijnsdorp et al., 1992).
1.5 Thesis outline
The research presented in this thesis aims at the determination of the impact of maternally transferred POPs on development and survival of fish early life stages, in order to assess if this exposure route can significantly impact the development of a fish population at current environmental concentrations.
In line with this aim the following 4 research questions will be addressed in this thesis:
1. What are critical egg concentrations for PCBs, PBDEs, HBCDs and MTCS for larval development, and how do these relate to field (fish) concentrations?
2. Do the above mentioned (groups of) substances represent the major risk in case of maternal transfer, or are there indications for substantial impact of other (unknown) substances?
3. What is the influence of ecophysiological characteristics on the impact of maternally accumulated compounds transferred to the eggs on the developing larvae?
4. To what extent can the larval mortality from maternally transferred toxicants affect population development, possibly in combination with high fishing pressure?
Chapter 1, the introduction, gives some background information on the topic of the present thesis, defines the aims and research questions, and presents the outline of the thesis.
Chapter 2 presents the p-ELS test that was developed to determine the impact of lipophilic POPs on the development of fertilised sole eggs into a fully metamorphosed juvenile fish. The performance of the test is illustrated by showing the effect of the dioxin-like PCB 126.

Eggsposed Chapter 1
21
In Chapter 3 the response to methyltriclosan was tested in the p-ELS test. Methyltriclosan is a degradation product of triclosan that has the potential to accumulate in fish tissue and disrupt the thyroid hormone system.
Chapter 4 reveals that POPs that are present in the eggs, concentrate during yolk-sac depletion to levels that can be up to 5 times higher than concentrations in the adult fish, and as such presents a possible explanation for the delayed effects that were observed in the p-ELS tests. A mathematical model (called ELS-OMEGA) based on the exciting OMEGA model (Hendriks et al,. 2001) is adapted and validated with experimental results and gives insight in the development of body residues of maternally transferred POPs in fish larvae under different development scenarios.
In Chapter 5 it is shown that the effects of mixtures of POPs extracted from sole from the Western Scheldt estuary and the North Sea can be predicted based on the effect levels of individual POPs in the mixture. Dioxin-like toxicity and polar narcosis were identified as responsible for most of the observed effects, and no indications were found for synergistic effects or for a substantial contribution of unknown lipophilic substances.
Chapter 6 estimates the potential risk of maternally transferred POPs for the European eel (Anguilla Anguilla) taking into account the specific biology of the species and European field POP concentrations. Assuming a relatively high sensitivity of the eel larvae, the results indicate that it is likely that eel from specific contaminated locations experience poor larval survival.
Chapter 7 explores the relevance of early life stage mortality as could be caused by maternally transferred substances, for population development in combination with no, low or high fishing pressure. A simple population model was developed for this purpose, based on the life history of the common sole. The model was used to identify the biological features that determine the vulnerability of species for enhanced larval mortality.
Chapter 8 contains the general discussion of the presented research. It discusses the overall results and strengths and weaknesses of the research performed. Based on the insights gained species that are potentially vulnerable to the effects of maternally transferred POPs are identified and discussed. Finally, the research questions are addressed and perspectives for future research are given.

Chapter 1 Eggsposed
22
1.6 References
Ahlborg, U.G., Becking, G.C., Birnbaum, L.S., Brouwer, A., Derks, H., Feeley, M., Golor, G., Hanberg, A., Larsen, J.C., Liem, A.K.D., Safe, S.H., Schlatter, C., Waern, F., Younes, M., Yrjanheikki, E., 1994. Toxic equivalency factors for dioxin-like PCBs - Report on a WHO-ECEH and IPCS consultation, December 1993. Chemosphere 28, 1049-1067.
Amara, R., Lagardere, F., Desaunay, Y., Marchand, J., 2000. Metamorphosis and estuarine colonisation in the common sole, Solea solea (L.): Implications for recruitment regulation. Oceanol. Acta 23, 469-484.
Anselmo, H.M.R., Koerting, L., Devito, S., van den Berg, J.H.J., Dubbeldam, M., Kwadijk, C., Murk, A.J., 2012a. Early life developmental effects of marine persistent organic pollutants on the sea urchin Psammechinus miliaris (vol 74, pg 2182, 2011). Ecotoxicology and Environmental Safety 80, 401-401.
Anselmo, H.M.R., Van Den Berg, J.H.J., Rietjens, I.M.C.M., Murk, A.T.J., 2012b. Inhibition of cellular efflux pumps involved in multi xenobiotic resistance (MXR) in echinoid larvae as a possible mode of action for increased ecotoxicological risk of mixtures. Ecotoxicology 21, 2276-2287.
Beyer, A., Biziuk, M., 2009. Environmental Fate and Global Distribution of Polychlorinated Biphenyls, Reviews of Environmental Contamination and Toxicology, Vol 201, pp. 137-158.
Blaxter, J.H.S., 1988. Pattern and variety in development, in: Randall, H. (Ed.), Fish physiology Volume XI Part A, Eggs and larvae.
Boon, J.P., Lewis, W.E., Tjoen-A-Choy, M.R., Allchin, C.R., Law, R.J., de Boer, J., ten Hallers-Tjabbes, C.C., Zegers, B.N., 2002. Levels of polybrominated diphenyl ether (PBDE) flame retardants in animals representing different trophic levels of the North Sea food web. Environmental Science & Technology 36, 4025-4032.
Borghesi, N., Corsolini, S., Leonards, P., Brandsma, S., de Boer, J., Focardi, S., 2009. Polybrominated diphenyl ether contamination levels in fish from the Antarctic and the Mediterranean Sea. Chemosphere 77, 693-698.
Breivik, K., Alcock, R., Li, Y.F., Bailey, R.E., Fiedler, H., Pacyna, J.M., 2004. Primary sources of selected POPs: regional and global scale emission inventories. Environmental Pollution 128, 3-16.
Brinkworth, L.C., Hodson, P.V., Tabash, S., Lee, P., 2003. CYP1A induction and blue sac disease in early developmental stages of rainbow trout (Oncorhynchus mykiss) exposed to retene. Journal of Toxicology and Environmental Health-Part A 66, 627-646.
Brouwer, A., Longnecker, M.P., Birnbaum, L.S., Cogliano, J., Kostyniak, P., Moore, J., Schantz, S., Winneke, G., 1999. Characterization of potential endocrine-related health effects at low-dose levels of exposure to PCBs. Environ. Health Perspect. 107, 639-649.
Covaci, A., Gerecke, A.C., Law, R.J., Voorspoels, S., Kohler, M., Heeb, N.V., Leslie, H., Allchin, C.R., de Boer, J., 2006. Hexabromocyclododecanes (HBCDs) in the

Eggsposed Chapter 1
23
environment and humans: A review. Environmental Science & Technology 40, 3679-3688.
Crane, H.M., Pickford, D.B., Hutchinson, T.H., Brown, J.A., 2006. The effects of methimazole on development of the fathead minnow, pimephales promelas, from embryo to adult. Toxicological Sciences 93, 278-285.
de Boer, J., Dao, Q.T., van Leeuwen, S.P.J., Kotterman, M.J.J., Schobben, J.H.M., 2010. Thirty year monitoring of PCBs, organochlorine pesticides and tetrabromodiphenylether in eel from The Netherlands. Environmental Pollution 158, 1228-1236.
Debruyn, A.M.H., Ikonomou, M.G., Gobas, F., 2004. Magnification and toxicity of PCBs, PCDDs, and PCDFs in upriver-migrating Pacific salmon. Environmental Science & Technology 38, 6217-6224.
Dekker, W., 2003. Did lack of spawners cause the collapse of the European eel, Anguilla anguilla? Fisheries Management and Ecology 10, 365-376.
Domingo, J.L., Bocio, A., 2007. Levels of PCDD/PCDFs and PCBs in edible marine species and human intake: A literature review. Environment International 33, 397-405.
Dorea, J.G., 2008. Persistent, bioaccumulative and toxic substances in fish: Human health considerations. Science of the Total Environment 400, 93-114.
Freitas, J., Cano, P., Craig-Veit, C., Goodson, M.L., Furlow, J.D., Murk, A.J., 2011. Detection of thyroid hormone receptor disruptors by a novel stable in vitro reporter gene assay. Toxicology in Vitro 25, 257-266.
Gomara, B., Bordajandi, L.R., Fernandez, M.A., Herrero, L., Abad, E., Abalos, M., Rivera, J., Gonzalez, M.J., 2005. Levels and trends of polychlorinated dibenzo-p-dioxins/furans (PCDD/Fs) and dioxin-like polychlorinated biphenyls (PCBs) in Spanish commercial fish and shellfish products, 1995-2003. J. Agric. Food Chem. 53, 8406-8413.
Gutleb, A.C., Mossink, L., Schriks, M., van den Berg, H.J.H., Murk, A.J., 2007a. Delayed effects of environmentally relevant concentrations of 3,3 ',4,4 '-tetrachlorobiphenyl (PCB-77) and non-polar sediment extracts detected in the prolonged-FETAX. Science of the Total Environment 381, 307-315.
Gutleb, A.C., Schriks, M., Mossink, L., van den Berg, J.H.J., Murk, A.J., 2007b. A synchronized amphibian metamorphosis assay as an improved tool to detect thyroid hormone disturbance by endocrine disruptors and apolar sediment extracts. Chemosphere 70, 93-100.
Hammerschmidt, C.R., Sandheinrich, M.B., 2005. Maternal diet during oogenesis is the major source of methylmercury in fish embryos. Environmental Science and Technology 39, 3580-3584.
Hansen, B.G., Paya-Perez, A.B., Rahman, M., Larsen, B.R., 1999. Qsars for K-ow and K-oc of PCB congeners: A critical examination of data, assumptions and statistical approaches. Chemosphere 39, 2209-2228.
Hill, A.J., Bello, S.M., Prasch, A.L., Peterson, R.E., Heideman, W., 2004. Water permeability and TCDD-induced edema in zebrafish early-life stages. Toxicological Sciences 78, 78-87.

Chapter 1 Eggsposed
24
Hites, R.A., 2004. Polybrominated diphenyl ethers in the environment and in people: A meta-analysis of concentrations. Environmental Science & Technology 38, 945-956.
Hutchinson, T.H., Solbe, J., Kloepper-Sams, P.J., 1998. Analysis of the ECETOC aquatic toxicity (EAT) database - III - Comparative toxicity of chemical substances to different life stages of aquatic organisms. Chemosphere 36, 129-142.
Isosaari, P., Hallikainen, A., Kiviranta, H., Vuorinen, P.J., Parmanne, R., Koistinen, J., Vartiainen, T., 2006. Polychlorinated dibenzo-p-dioxins, dibenzofurans, biphenyls, naphthalenes and polybrominated diphenyl ethers in the edible fish caught from the Baltic Sea and lakes in Finland. Environmental Pollution 141, 213-225.
Janak, K., Covaci, A., Voorspoels, S., Becher, G., 2005. Hexabromocyclododecane in marine species from the Western Scheldt Estuary: Diastereoisomer- and enantiomer-specific accumulation. Environmental Science & Technology 39, 1987-1994.
Johnson, R.D., Tietge, J.E., Jensen, K.M., Fernandez, J.D., Linnum, A.L., Lothenbach, D.B., Holcombe, G.W., Cook, P.M., Christ, S.A., Lattier, D.L., Gordon, D.A., 1998. Toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin to early life stage brook trout (Salvelinus fontinalis) following parental dietary exposure. Environmental Toxicology and Chemistry 17, 2408-2421.
Johnston, T.A., Bodaly, R.A., Latif, M.A., Fudge, R.J.P., Strange, N.E., 2001. Intra- and interpopulation variability in maternal transfer of mercury to eggs of walleye (Stizostedion vitreum). Aquatic Toxicology 52, 73-85.
Kamler, E., 2008. Resource allocation in yolk-feeding fish. Reviews in Fish Biology and Fisheries 18, 143-200.
Kannan, K., Corsolini, S., Imagawa, T., Focardi, S., Giesy, J.P., 2002. Polychlorinated -naphthalenes, -biphenyls, -dibenzo-p-dioxins, -dibenzofurans and p,p '-DDE in bluefin tuna, swordfish, cormorants and barn swallows from Italy. Ambio 31, 207-211.
Kelly, B.C., Gray, S.L., Ikonomou, M.G., MacDonald, J.S., Bandiera, S.M., Hrycay, E.G., 2007. Lipid reserve dynamics and magnification of persistent organic pollutants in spawning sockeye salmon (Oncorhynchus nerka) from the Fraser River, British Columbia. Environmental Science & Technology 41, 3083-3089.
Kelly, B.C., Ikonomou, M.G., MacPherson, N., Sampson, T., Patterson, D.A., Dubetz, C., 2011. Tissue residue concentrtions of organohalogens and trace elements in adult pacific salmon returning to the fraser river, Britisch Colombia, Canada . Environmental Toxicology and Chemistry 30, 367-376.
Klaren, P.H.M., Wunderink, Y.S., Yufera, M., Mancera, J.M., Flik, G., 2008. The thyroid gland and thyroid hormones in Senegalese sole (Solea senegalensis) during early development and metamorphosis. General and Comparative Endocrinology 155, 686-694.
Knutzen, J., Bjerkeng, B., Naes, K., Schlabach, M., 2003. Polychlorinated dibenzofurans/dibenzo-p-dioxins (PCDF/PCDDs) and other dioxin-like substances in marine organisms from the Grenland fjords, S. Norway, 1975-2001: present contamination levels, trends and species specific accumulation of PCDF/PCDD congeners. Chemosphere 52, 745-760.

Eggsposed Chapter 1
25
Labandeira, A., Eljarrat, E., Barcelo, D., 2007. Congener distribution of polybrominated diphenyl ethers in feral carp (Cyprinus carpio) from the Llobregat River, Spain. Environmental Pollution 146, 188-195.
Law, R.J., Allchin, C.R., de Boer, J., Covaci, A., Herzke, D., Lepom, P., Morris, S., Tronczynski, J., de Wit, C.A., 2006. Levels and trends of brominated flame retardants in the European environment. Chemosphere 64, 187-208.
Li, L.N., Xie, S.D., Cai, H., Bai, X.T., Xue, Z., 2008. Quantitative structure-property relationships for octanol-water partition coefficients of polybrominated diphenyl ethers. Chemosphere 72, 1602-1606.
Mac, M.J., Schwartz, T.R., Edsall, C.C., Frank, A.M., 1993. Polychlorinated-bipenyls in Great-Lakes laketrout and theur eggs-Relations to survival and cengener composition 1979-1988. Journal of Great Lakes Research 19, 752-765.
Madenjian, C.P., 2011. Sex effect on polychlorinated biphenyl concentrations in fish: a synthesis. Fish and Fisheries 12, 451-460.
Martinez, G.M., Bolker, J.A., 2003. Embryonic and larval staging of summer flounder (Paralichthys dentatus). Journal of Morphology 255, 162-176.
McKim, J.M., 1977. Evaluation of tests with eraly life stages of fish for predicting long term toxicity. Journal of the Fisheries Research Board of Canada 34, 1148-1154.
Miller, M.A., 1993. Maternal transfer or organochlosine compounds in salmonines to their eggs. Canadian Journal of Fisheries and Aquatic Sciences 50, 1405-1413.
Molde, K., Ciesielski, T.M., Fisk, A.T., Lydersen, C., Kovacs, K.M., Sormo, E.G., Jenssen, B.M., 2013. Associations between vitamins A and E and legacy POP levels in highly contaminated Greenland sharks (Somniosus microcephalus). Science of the Total Environment 442, 445-454.
Mommsen, T.P., Walsh, P.J., 1988. Vitellogenesis and oocyte assembly in: Randall, H. (Ed.), Fish physiology, Volume XI The physiology of developing fish Part A, Eggs and larvae. .
Monteverdi, G.H., Di Giulio, R.T., 2000. Oocytic accumulation and tissue distribution of 2,3,7,8-tetrachlorodibenzo-p-dioxin and benzo a pyrene in gravid Fundulus heteroclitus. Environmental Toxicology and Chemistry 19, 2512-2518.
Moore, M.R., Vetter, W., Gaus, C., Shaw, G.R., Muller, J.F., 2002. Trace organic compounds in the marine environment. Marine Pollution Bulletin 45, 62-68.
Munschy, C., Heas-Moisan, K., Tixier, C., Pacepavicius, G., Alaee, M., 2010. Dietary exposure of juvenile common sole (Solea solea L.) to polybrominated diphenyl ethers (PBDEs): Part 2. Formation, bioaccumulation and elimination of hydroxylated metabolites. Environmental Pollution 158, 3527-3533.
Murk, A., Morse, D., Boon, J., Brouwer, A., 1994. In-vitro metabolism of 3,3',4,4'-tertrachlorobiphenyl in relation to ethoxyresorufin-o-deethylase activity in liver microsomes of some wildlife species and rat. European Journal of Pharmacology-Environmental Toxicology and Pharmacology Section 270, 253-261.
Murk, A.J., Leonards, P.E.G., van Hattum, B., Luit, R., van der Weiden, M.E.J., Smit, M., 1998. Application of biomarkers for exposure and effect of polyhalogenated aromatic hydrocarbons in naturally exposed European otters (Lutra lutra). Environmental Toxicology and Pharmacology 6, 91-102.

Chapter 1 Eggsposed
26
Niimi, A.J., 1983. Biological and Toxicological Effects of Environmental Contaminants in Fish and Their Eggs. Canadian Journal of Fisheries and Aquatic Sciences 40, 306-312.
Nyholm, J.R., Norman, A., Norrgren, L., Haglund, P., Andersson, P.L., 2008. Maternal transfer of brominated flame retardants in zebrafish (Danio rerio). Chemosphere 73, 203-208.
OECD, 1992. Guidline for testing of chemicals 210: Fish, Early-life Stage Toxicity Test Organisation for Economic Co-operation and Development (OECD), Paris OECD, 1998. Guidline for testing of chemicals 212: Fish, Short-term toxicity test on
embryo and sac-fry stages Organisation for Economic Co-operation and Development (OECD), Paris OECD, 2006. Draft proposal for a new guideline: Fish Embryo Toxicity (FET) Test Organisation for Economic Co-operation and Development (OECD), Paris Palace, V., Park, B., Pleskach, K., Gemmill, B., Tomy, G., 2010. Altered thyroxine
metabolism in rainbow trout (Oncorhynchus mykiss) exposed to hexabromocyclododecane (HBCD). Chemosphere 80, 165-169.
Palazzi, R., Richard, J., Bozzato, G., Zanella, L., 2006. Larval and juvenile rearing of common sole (Solea solea L.) in the Northern Adriatic (Italy). Aquaculture 255, 495-506.
Rijnsdorp, A.D., Vanbeek, F.A., Flatman, S., Millner, R.M., Riley, J.D., Giret, M., Declerck, R., 1992. Recriutment of sole stocks, Solea solea (L), in the Northeast Atlantic Neth. J. Sea Res. 29, VOL 29, p 173-
Roberts, S.C., Noyes, P.D., Gallagher, E.P., Stapleton, H.M., 2011. Species-Specific Differences and Structure-Activity Relationships in the Debromination of PBDE Congeners in Three Fish Species. Environmental Science & Technology 45, 1999-2005.
Rothschild, B.J., 2000. "Fish stocks and recruitment": the past thirty years. Ices Journal of Marine Science 57, 191-201.
Russell, R.W., Gobas, F., Haffner, G.D., 1999. Maternal transfer and in ovo exposure of organochlorines in oviparous organisms: A model and field verification. Environmental Science & Technology 33, 416-420.
Schreiber, A.M., 2006. Asymmetric craniofacial remodeling and liberalized behavior in larval flatfish. Journal of Experimental Biology 209, 610-621.
Schriks, M., Zvinavashe, E., Furlow, J.D., Murk, A.J., 2006. Disruption of thyroid hormone-mediated Xenopus laevis tadpole tail tip regression by hexabromocyclododecane (HBCD) and 2,2 ',3,3 ',4,4 ',5,5 ',6-nona brominated diphenyl ether (BDE206). Chemosphere 65, 1904-1908.
Sellstrom, U., Kierkegaard, A., de Wit, C., Jansson, B., 1998. Polybrominated diphenyl ethers and hexabromocyclododecane in sediment and fish from a Swedish river. Environmental Toxicology and Chemistry 17, 1065-1072.
Serrano, R., Blanes, M.A., Lopez, F.J., 2008. Maternal transfer of organochlorine compounds to oocytes in wild and farmed gilthead sea bream (Sparus aurata). Chemosphere 70, 561-566.

Eggsposed Chapter 1
27
Storelli, M.M., Barone, G., Storelli, A., Marcotrigiano, G.O., 2011. Levels and congener profiles of PCBs and PCDD/Fs in blue shark (Prionace glauca) liver from the South-Eastern Mediterranean Sea (Italy). Chemosphere 82, 37-42.
Stouthart, X., Huijbregts, M.A.J., Balm, P.H.M., Lock, R.A.C., Bonga, S.E.W., 1998. Endocrine stress response and abnormal development in carp (Cyprinus carpio) larvae after exposure of the embryos to PCB 126. Fish Physiol. Biochem. 18, 321-329.
Svendsen, T.C., Vorkamp, K., Ronsholdt, B., Frier, J.O., 2007. Organochlorines and polybrominated diphenyl ethers in four geographically separated populations of Atlantic salmon (Salmo salar). Journal of Environmental Monitoring 9, 1213-1219.
Tesch, F.W., 2003. The eel. Blackwell Science. USEPA, 1996. Ecological Effects Test Guidelines: OPPTS 850.1400 Fish Early-Life Stage
Toxicity Test. United States Environmental Protection Agency (USEPA). Van den Berg, M., Birnbaum, L., Bosveld, A.T.C., Brunstrom, B., Cook, P., Feeley, M.,
Giesy, J.P., Hanberg, A., Hasegawa, R., Kennedy, S.W., Kubiak, T., Larsen, J.C., van Leeuwen, F.X.R., Liem, A.K.D., Nolt, C., Peterson, R.E., Poellinger, L., Safe, S., Schrenk, D., Tillitt, D., Tysklind, M., Younes, M., Waern, F., Zacharewski, T., 1998. Toxic equivalency factors (TEFs) for PCBs, PCDDs, PCDFs for humans and wildlife. Environ. Health Perspect. 106, 775-792.
Van den Berg, M., Birnbaum, L.S., Denison, M., De Vito, M., Farland, W., Feeley, M., Fiedler, H., Hakansson, H., Hanberg, A., Haws, L., Rose, M., Safe, S., Schrenk, D., Tohyama, C., Tritscher, A., Tuomisto, J., Tysklind, M., Walker, N., Peterson, R.E., 2006. The 2005 World Health Organization reevaluation of human and mammalian toxic equivalency factors for dioxins and dioxin-like compounds. Toxicological Sciences 93, 223-241.
van Leeuwen, S.P.J., de Boer, J., 2008. Brominated flame retardants in fish and shellfish - levels and contribution of fish consumption to dietary exposure of Dutch citizens to HBCD. Mol. Nutr. Food Res. 52, 194-203.
van Leeuwen, S.P.J., Leonards, P.E.G., Traag, W.A., Hoogenboom, L.A.P., de Boer, J., 2007. Polychlorinated dibenzo-p-dioxins, dibenzofurans and biphenyls in fish from the Netherlands: concentrations, profiles and comparison with DR CALUX (R) bioassay results. Analytical and Bioanalytical Chemistry 389, 321-333.
Villalobos, S.A., Hamm, J.T., Teh, S.J., Hinton, D.E., 2000. Thiobencarb-induced embryotoxicity in medaka (Oryzias latipes): stage-specific toxicity and the protective role of chorion. Aquatic Toxicology 48, 309-326.
Voorspoels, S., Covaci, A., Maervoet, J., De Meester, I., Schepens, P., 2004. Levels and profiles of PCBs and OCPs in marine benthic species from the Belgian North Sea and the Western Scheldt Estuary. Marine Pollution Bulletin 49, 393-404.
Voorspoels, S., Covaci, A., Schepens, P., 2003. Polybrominated diphenyl ethers in marine species from the Belgian North Sea and the Western Scheidt Estuary: Levels, profiles, and distribution. Environmental Science & Technology 37, 4348-4357.
Walker, M.K., Peterson, R.E., 1994. Toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin to brook trout (Salvelinus fontinalis) during early life development. Environmental Toxicology and Chemistry 13, 817-820.

Chapter 1 Eggsposed
28
Wang, M.Y., Chen, J.F., Lin, K.F., Chen, Y.H., Hu, W., Tanguay, R.L., Huang, C.J., Dong, Q.X., 2011. Chronic zebrafish PFOS exposure alters seks ration and metarnal related effects in F1-offspring. Environmental Toxicology and Chemistry 30, 2073-2080.
Wiegand, M.D., 1996. Composition, accumulation and utilization of yolk lipids in teleost fish. Reviews in Fish Biology and Fisheries 6, 259-286.
Wilson, P.J., Tillitt, D.E., 1996. Rainbow trout embryotoxicity of a complex contaminant mixture extracted from Lake Michigan lake trout. Mar. Environ. Res. 42, 129-134.
Wooton, R.J., 1992. Ecology of teleost Fishes. Chapman 7 Hall, London. Yamano, K., 2005. The role of thyroid hormone in fish development with reference to
aquaculture. Jarq-Japan Agricultural Research Quarterly 39, 161-168. Yamauchi, M., Kim, E.Y., Iwata, H., Shima, Y., Tanabe, S., 2006. Toxic effects of 2,3,7,8-
tetrachlorodibenzo-p-dioxin (TCDD) in developing red seabream (Pagrus major) embryo: An association of morphological deformities with AHR1, AHR2 and CYP1A expressions. Aquatic Toxicology 80, 166-179.
Yu, L.Q., Lam, J.C.W., Guo, Y.Y., Wu, R.S.S., Lam, P.K.S., Zhou, B.S., 2011. Parental Transfer of Polybrominated Diphenyl Ethers (PBDEs) and Thyroid Endocrine Disruption in Zebrafish. Environmental Science & Technology 45, 10652-10659.
Zabel, E.W., Cook, P.M., Peterson, R.E., 1995. Potency of 3,3',4,4',5-pentachlorobiphenyl (PCB-126), alone and in combination with 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD), to produce lake trout early life stage mortality. Environmental Toxicology and Chemistry 14, 2175-2179.
Zheng, X.M., Zhu, Y.T., Liu, C.S., Liu, H.L., Giesy, J.P., Hecker, M., Lam, M.H.W., Yu, H.X., 2012. Accumulation and Biotransformation of BDE-47 by Zebrafish Larvae and Teratogenicity and Expression of Genes along the Hypothalamus-Pituitary-Thyroid Axis. Environmental Science & Technology 46, 12943-12951.

29
CHAPTER 2
Prolonged ELS test with the marine flatfish sole (Solea solea) shows delayed toxic
effects of previous exposure to PCB 126
E.M. Foekemaa, C.M. Deerenbergb, and A.J. Murka,c
Foekema, E.M., C.M. Deerenberg, A.J. Murk. (2008): Prolonged ELS test with the marine flatfish sole (Solea solea) shows delayed toxic effects of previous exposure to PCB 126. Aquatic Toxicology, 90(3): p. 197-203.
a IMARES, Department Experimental Ecology b IMARES, Department Fish c Wageningen University, Division of Toxicology

Chapter 2 Eggsposed
30
Abstract
The effect of the dioxin-like PCB 126 (3,3',4,4',5-pentachlorobiphenyl) on the early development of the marine flatfish sole (Solea solea) was tested in a newly developed early life stage (ELS) test that includes the metamorphosis of the symmetric larvae into an asymmetrical flatfish. Early life stages of sole were exposed to a concentration series of PCB 126 in seawater until 4, 8, 10 and 15 days post fertilisation (dpf). Subsequently the development of the larvae was registered under further unexposed conditions. The LC50s at the start of the free-feeding stage (12 dpf) ranged between 39 and 83 ng PCB 126/L depending on exposure duration. After the fish had completed the metamorphosis, the LC50 values ranged between 1.7 and 3.7 ng PCB 126/L for the groups exposed for 4, 8 and 10 dpf respectively. Thus exposure for only 4 days, covering only the egg stage, was sufficient to cause adverse effects during a critical developmental phase two weeks later. The internal dosages of these larvae, determined by means of an in-vitro gene reporter assay as dioxin-equivalent values (TEQ), revealed an internal lethal concentration ILC50 of 1 ng TEQ/g lipid, which is within the same order of magnitude as TEQ levels found in fish from highly polluted areas. This study indicates that ELS fish tests that are terminated shortly after the fish becomes free-feeding underestimate the toxic potential of compounds with low acute toxicity such as PCBs. Our prolonged ELS with this native marine flatfish suggests that reproductive success of fish populations at contaminated sites can be affected by persistent compounds that are accumulated by the female fish and passed on to the eggs.

Eggsposed Chapter 2
31
2.1 Introduction
The development of an embryo into a juvenile fish is considered as the period of the fish‘s life that is the most sensitive to disturbances (Hutchington et al., 1998). During this early life stage critical development of organs and tissues takes place, a process which can easily be disrupted by unfavourable environmental conditions, including exposure to toxic compounds. The young stages lack the ability to compensate the impact of physiological processes affected by toxicant exposure and are therefore more sensitive to adverse effects than fully grown fish (Crane et al., 2006). Moreover, with their small body volume and relatively large surface it is expected that the internal concentration of toxic compounds will rapidly increase during exposure to contaminated water.
For this reason toxicity tests with early life stages of fish, the so-called ELS (Early Life Stage)-fish tests, are often applied for assessing the toxic potential of substances and environmental samples. Besides their sensitivity, these tests have the practical advantages over tests with older fish that they require less test volume and space. This enables the use of higher numbers of test organisms and replicates and so improves the statistical power of the test results (Wedekind et al., 2007).
The available test guidelines for fish ELS testing have their emphasis on fresh water species from temperate or tropical regions (OECD, 1992; EPA, 1996; OECD, 1998; OECD, 2006). Guidance on the utilisation of marine species is scarce, especially with respect to species relevant for the West European region. In addition, none of the test species recommended in these guidelines undergoes an obvious metamorphosis, that is typical for certain taxonomical fish groups such as flatfish and eels (Yamano, 2005) and that could be very sensitive to toxic compounds as has been shown for amphibians (Gutleb, 2006; Gutleb et al., 2007b). Therefore we investigated the applicability of the marine flatfish sole (Solea solea) as a test species in full development tests. Sole is a native European species of both ecological and economical importance. It is bred under controlled conditions for aquaculture purposes and undergoes a full metamorphosis during the development into a juvenile (see for detailed descriptions for instance: Martinez & Bolker, 2003; Schreiber, 2006).
The development from egg to larva is known to be sensitive to dioxin-like toxicity in zebrafish (Murk et al., 1996a) and amphibians (Gutleb, 1999). We used PCB 126 for studying dioxin-like toxicity on early life stages of sole. PCB 126 is a good model compound, as it is hardly metabolised and known to bind to the Ah-receptor with high potency. This binding has been associated with a

Chapter 2 Eggsposed
32
variety of toxic responses including teratogenicity, possibly induced via disruption of vitamin A homeostasis and changes in the thyroid function (Brouwer et al., 1995, Brown et al., 2004). The dioxin-like toxic potency of dioxins and PCBs, is expressed as TEF, being the Toxic Equivalency Factor; relative to that of the most potent congener 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) (Safe, 1992). Since the TEF is related to a biological response, it is not only compound-specific, but also taxon-specific. Based on available literature Van den Berg and co-authors established TEFs for mammals, birds and fish (Van den Berg et al.,1998), that were adopted by the World Health Organisation (WHO). The TEF-concept assumes additivity of the dioxin-like toxicity of compounds in a mixture, and thus enables the expression of the total dioxin-like toxic potency of the mixture of contaminants present in an environmental sample as Toxicity Equivalents (TEQ) of the toxicity of TCDD.
The dioxin-like toxic potency of compounds present in fish is generally assessed in the light of human risk assessment, and therefore calculated based on the WHO-TEF values for mammals. In vitro reporter gene assays based on Ah-receptor activation have been developed and chemically validated for the direct quantification of the dioxin-like potency of mixtures of contaminants in sediments and biota (Murk et al., 1996a; 1998; Stronkhorst et al., 2002; Hoogenboom et al., 2006; Leeuwen et al., 2007). In these in vitro reporter gene assays the relative potency of compounds to activate the Ah-receptor is related to that of TCDD, not the toxicity compared to that of TCDD. Although the in vitro TEFs are not based on toxicological experiments, in practise they compare well with the mammalian in vivo TEFs for dioxin-like compounds. The mammalian TEF for PCB 126 is well established and consequently set at 0.1 in various assessments including the in vitro-reporter gene systems (Safe, 1992; Ahlborg et al., 1992; Van den Berg et al., 1998; Stronkhorst et al., 2002).
This paper describes the effects of PCB 126 on the early development of the marine fish Solea solea in a newly developed bioassay that covers the whole development from fertilised egg into a fully metamorphosed flatfish.
2.2 Materials and methods
2.2.1 Animals
The fertilised eggs used in this study all originated from one group of parent sole (Solea solea). Spawning and fertilisation were temperature-induced and took place overnight at the facilities of the commercial sole farm Solea BV in IJmuiden, the Netherlands. The next morning, the eggs were transported to the laboratory in Den Helder in plastic bags containing seawater. During the ca. 1

Eggsposed Chapter 2
33
hour transport period, the water temperature was maintained around 12ºC, the temperature in the spawning tank. The eggs and larvae were kept in natural seawater (salinity 32 ‰) collected from the Eastern Scheldt, a relatively pristine bay of the North Sea, that is often used as a reference site in marine ecotoxicological research in the Netherlands (e.g. Vethaak et al., 2005; Kuiper et al., 2007).
2.2.2 Stock solutions and exposure media
Stock solutions of PCB 126 (3,3’,4,4’5-pentachlorobiphenyl, purity 99.1%, supplier Promochem) in DMSO (purity 99.9% A.C.S. spectrophotometric grade, supplier SIGMA-Aldrich) were used to spike the seawater that was used as exposure medium. The final DMSO concentration always was 0.1 % vv. The stock solutions were prepared in sufficient volumes for all exposures at the start of the first test and were stored in dark at room temperature.
2.2.3 Test procedure
Immediately after arrival at the laboratory (<12h after fertilisation), the sole eggs were randomly divided into groups of ca. 200 eggs that were placed in glass beakers containing 800 ml seawater spiked with the test concentrations. The glass beakers were placed in a temperature controlled cabinet at 12-13°C, with a 16 hour photoperiod with dimmed light (ca. 100 LUX) per day. After 48 hours the floating fertilised eggs were collected and the non-fertilised eggs at the bottom discarded. Out of each exposure group twenty fertilised eggs were put together in a 100 ml glass beaker containing 80 ml of a fresh solution of the exposure concentration. Each exposure condition was duplicated. The test beakers were further incubated under the above described conditions.
Every other day, about 50 ml (ca. 60%) of the water in each test beaker was pipetted off and replaced with fresh seawater at the appropriate concentration.
From the moment the first larvae had desorbed their yolk-sac and became free-feeding, all test beakers were provided with an ad libitum amount of Artemia nauplii (Artemia salina) on a daily basis. On an almost daily basis the dead individuals were removed and faeces and surplus food was removed as much as possible. Each fish that had completed the metamorphosis was removed, narcotised in a MS222-sollution (100 mg MS222 = Ethyl 3-aminonebzoate methanesulfonic acid salt (purity 98%, ACROS Organics) and 200 mg NaHCO3 (purity 99.0%, Fluka Chemika) in 1 l seawater) measured and checked for malformations using a stereo microscope. While still being narcotised, the fish was then killed and preserved in a 4%-formaldehyde solution.

Chapter 2 Eggsposed
34
Two exposure studies were performed between February and May 2007, with some differences. In the first test, the eggs and larvae were exposed until 15 days post fertilisation (15-dpf), the moment that all fish had become free-feeding. The exposure concentrations ranged from 3 to 1000 ng PCB 126/l, chosen based on test results from ELS tests with Zebra fish (Murk et al., 1996a) and tadpoles (Gutleb et al., 1999). On day 15, the surviving larvae were carefully transferred with a polyethylene pipette to another glass beaker with clean seawater and further reared unexposed until the end of the metamorphosis. During the first 19 days of this test the water temperature was maintained on 12 ± 1ºC, and on 13ºC thereafter. The fish were considered fully metamorphosed when both eyes were located at one side of the head.
Based on the results of the first experiment, the second test series was performed with lower test concentrations ranging from 0.1 to 100 ng PCB 126/l. Moreover, the exposure duration was reduced to 4, 8 or 10 days post fertilisation (dpf). The 4-dpf period covered only the egg stage, the 8-dpf exposure period covered most of the yolk-sac stage, whereas exposure of the 10-dpf group ended when the first fish became free-feeding. The water temperature was maintained at 13 ± 1ºC during the whole test period. The criterion for ‘fully metamorphosed’ now was not based on eye migration only, but also on the development of the dorsal fin that passes the left eye after this has reached the right side of the head.
2.2.4 Internal concentrations
To assess the actual uptake of PCB 126 into the larvae, an additional 200 eggs were exposed to a selection of test concentrations in the same way as the 4-dpf group. After two days of exposure, 150 fertilised eggs were collected from each exposure concentration and placed in 600 ml of new test medium at the appropriate test concentration, thus maintaining the ratio of 4 ml exposure media per egg. As soon as possible after hatching (<12 hr) at least 140 larvae per test concentration were collected with a polyethylene pipette and placed in clean seawater for maximal 15 minutes to rinse off exposure water before they were pipetted into a glass tube with a minimum of seawater and stored frozen.
The total wet weight per sample was about 100 mg, hardly sufficient for accurate chemical analysis of the PCB 126 concentration and expression on a lipid basis. Therefore, a sensitive in vitro reporter gene assay for dioxin-like toxic potency in H4IIE rat hepatoma cells (Murk et al., 1998) was applied for the direct quantification of the in vitro TCDD-equivalent (TEQ) levels on a lipid basis in these very small samples. This DRE-H4IIE.Luc assay (sometimes also referred to as DR CALUX) has been chemically validated before and produces a response

Eggsposed Chapter 2
35
that is linear with the concentration of compounds with dioxin-like toxicity (Murk et al., 1998; Stronkhorst et al., 2002; Besselink et al., 2004; Hoogenboom et al., 2006; Van Leeuwen et al., 2007).
2.2.5 Calculations and statistical analysis
All statistical calculations were performed using GraphPad Prism (version 4.03, January 2005). To determine LC50-values, a sigmoid dose response curve with variable slope was fitted through the experimental data.
The significance of differences between the LC50 values of the different treatments at various moments of observation was calculated with a two-way ANOVA with Bonferroni post test. For this analysis data from both experiments were used for observations on days (dpf) 12, 15, and 20, while from the second experiments also data from dpf 10 and 56 were available. Due to this difference in the available data from both experiments, two ANOVA analyses were performed: the first included all days but excluded the data from the first experiment; the second included all experiments but excluded observations on days 10 and 56.
The relation between exposure concentrations and internal TEQ-level in the fish larvae was fitted using the same GraphPad Prism software.
2.3 Results
2.3.1 First experiment
In the first test, the sole eggs started to hatch 5 days after fertilisation. Five days later (dpf 10), the first larvae became free-feeding and on dpf 14 all larvae had reached that stage. On day 33, the first fish completed metamorphosis. The test was terminated on dpf 47 when all surviving fish had completed metamorphosis.
The development of the eggs/fish in the test beakers with clean seawater (controls) was not significantly different from that in the solvent controls (0.1% DMSO). At the end of this experiment 75% of the eggs from both controls had developed in normally metamorphosed juvenile flatfish (Figure 2.1). The mortality in the controls occurred almost exclusively before day 25.
Until day 9 no exposure related effects were observed and mortality was below 20% in all test beakers. However, within the next 5 days a high mortality rate was seen in the exposure concentrations of 32 ng PCB 126/l and above. At the moment all larvae were free-feeding, on day 15, less than 20% of the larvae
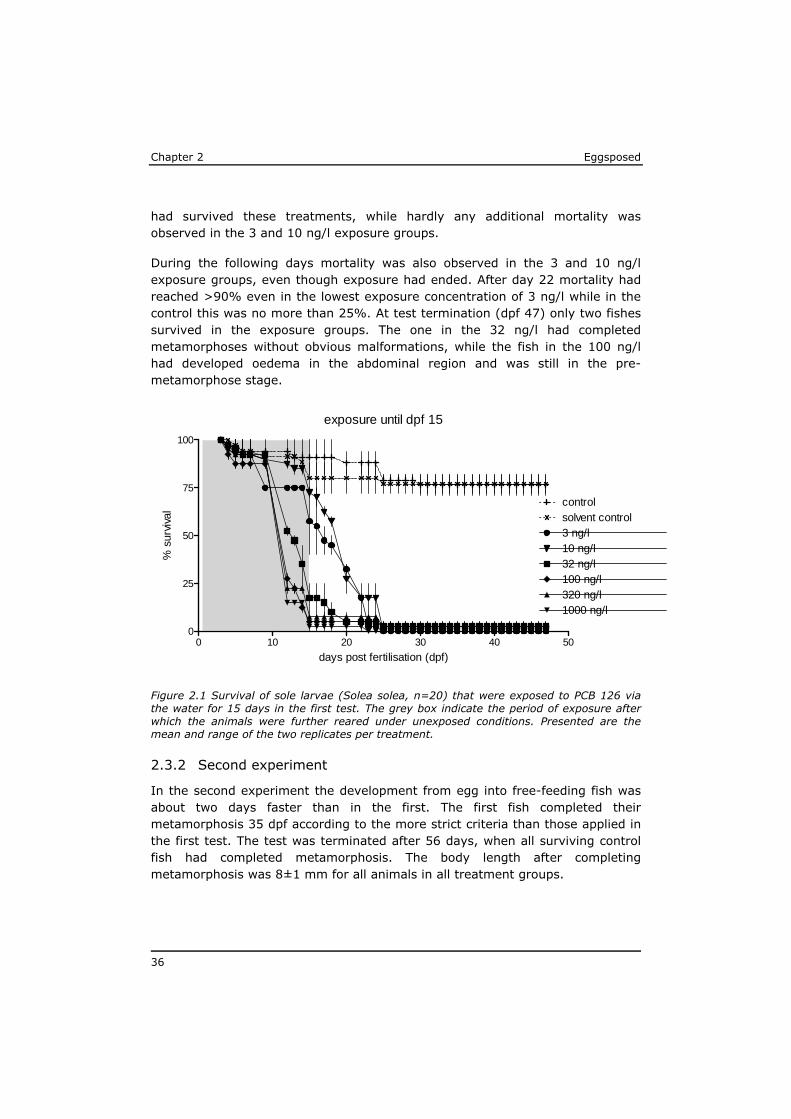
Chapter 2 Eggsposed
36
had survived these treatments, while hardly any additional mortality was observed in the 3 and 10 ng/l exposure groups.
During the following days mortality was also observed in the 3 and 10 ng/l exposure groups, even though exposure had ended. After day 22 mortality had reached >90% even in the lowest exposure concentration of 3 ng/l while in the control this was no more than 25%. At test termination (dpf 47) only two fishes survived in the exposure groups. The one in the 32 ng/l had completed metamorphoses without obvious malformations, while the fish in the 100 ng/l had developed oedema in the abdominal region and was still in the pre-metamorphose stage.
Figure 2.1 Survival of sole larvae (Solea solea, n=20) that were exposed to PCB 126 via the water for 15 days in the first test. The grey box indicate the period of exposure after which the animals were further reared under unexposed conditions. Presented are the mean and range of the two replicates per treatment.
2.3.2 Second experiment
In the second experiment the development from egg into free-feeding fish was about two days faster than in the first. The first fish completed their metamorphosis 35 dpf according to the more strict criteria than those applied in the first test. The test was terminated after 56 days, when all surviving control fish had completed metamorphosis. The body length after completing metamorphosis was 8±1 mm for all animals in all treatment groups.
exposure until dpf 15
0 10 20 30 40 500
25
50
75
100
controlsolvent control3 ng/l10 ng/l32 ng/l100 ng/l320 ng/l1000 ng/l
days post fertilisation (dpf)
% s
urvi
val
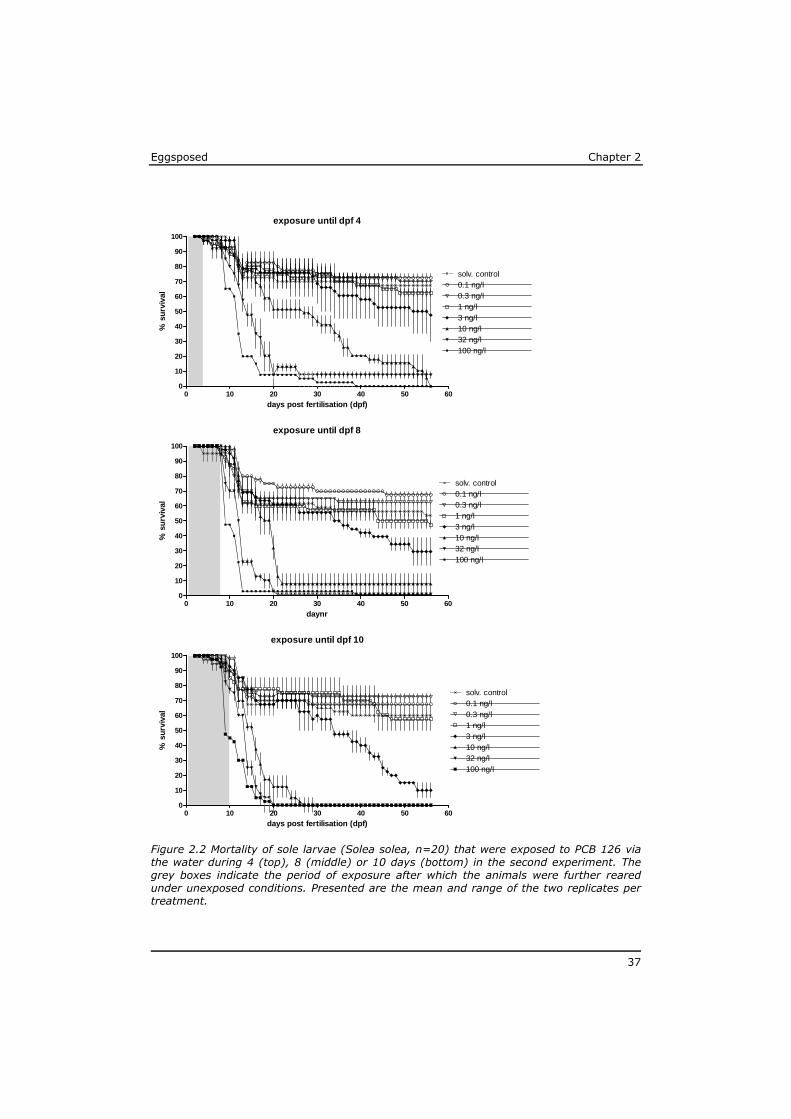
Eggsposed Chapter 2
37
Figure 2.2 Mortality of sole larvae (Solea solea, n=20) that were exposed to PCB 126 via the water during 4 (top), 8 (middle) or 10 days (bottom) in the second experiment. The grey boxes indicate the period of exposure after which the animals were further reared under unexposed conditions. Presented are the mean and range of the two replicates per treatment.
exposure until dpf 4
0 10 20 30 40 50 600
10
20
30
40
50
60
70
80
90
100
solv. control0.1 ng/l0.3 ng/l1 ng/l3 ng/l10 ng/l32 ng/l100 ng/l
days post fertilisation (dpf)
% s
urv
ival
exposure until dpf 8
0 10 20 30 40 50 600
10
20
30
40
50
60
70
80
90
100
solv. control0.1 ng/l0.3 ng/l1 ng/l3 ng/l10 ng/l32 ng/l100 ng/l
daynr
% s
urv
ival
exposure until dpf 10
0 10 20 30 40 50 600
10
20
30
40
50
60
70
80
90
100
solv. control0.1 ng/l0.3 ng/l1 ng/l3 ng/l10 ng/l32 ng/l100 ng/l
days post fertilisation (dpf)
% s
urv
ival

Chapter 2 Eggsposed
38
The survival of the control animals was ca. 60% at completion of metamorphosis. Again, the mortality in the controls occurred mainly during the first 25 days of the test, followed by only incidental mortality later (Figure 2.2).
The timing of main mortality was the same as in the first experiment. The highest test concentrations (32 and 100 ng PCB 126/l) induced almost complete mortality of the larvae at the end of the yolk-sac/beginning of the free-feeding stage. This occurred for all exposure durations, even in the groups that were only exposed during the first 4 days. In the 3 and 10 ng PCB 126/l exposure groups mortality occurred at a later moment depending on exposure duration, but this was in all cases days after exposure had ended. When around day 35, the first fish had started the metamorphosis, other surviving fish in the 1, 3 and 10 ng PCB 126/l exposure groups that were still in the pre-metamorphosis stage, developed oedema in the abdominal region. On day 41, oedema was observed in more than 50% of the surviving pre-metamorphic fish in the 8-dpf and 10-dpf treatments at 3 ng/l and in ca. 20% of the survivors in the 4dpf group. In the 4-dpf 10 ng/l treatment this was even 100%. Remarkably, no fish with oedema were seen at the same test concentration in the 8-dpf group. At the same concentration in the 10-dpf treatments complete mortality had occurred at that moment. All fish with oedema died without any indication that the metamorphosis started. The surviving fish completed the metamorphosis without exposure-related malformations or delay.
2.3.3 Effect concentrations
The data set allowed the calculation of LC50 values between dpf 12 and 20 for all 4 exposure durations (Figure 2.3). Due to the increasing mortality in time it was not possible to calculate LC50s after day 20 for the 15-dpf group, and no data were available from day 10.
Statistical analysis revealed that treatment only affected the LC50s in a significant way at early observation. Only on day 10 the LC50 of the 4-pdf group was significantly (p<0.05) higher than other treatments. During all later observations the LC50 values of all treatment groups were statistically comparable.
The moment of observation had an extremely significant effect (p<0.0001). On day 12, the LC50 values for the various treatments ranged between 39 and 82 ng PCB 126/l. Although exposure stopped after a period of 4 to 15 days all LC50 values rapidly decreased with increasing observation time. At the end of the test all LC50 values ranged between 1.7 and 3.7 ng PCB 126/l and did not differ significantly. They were, however, significantly more than 20 times lower than the LC50 values on day 12.
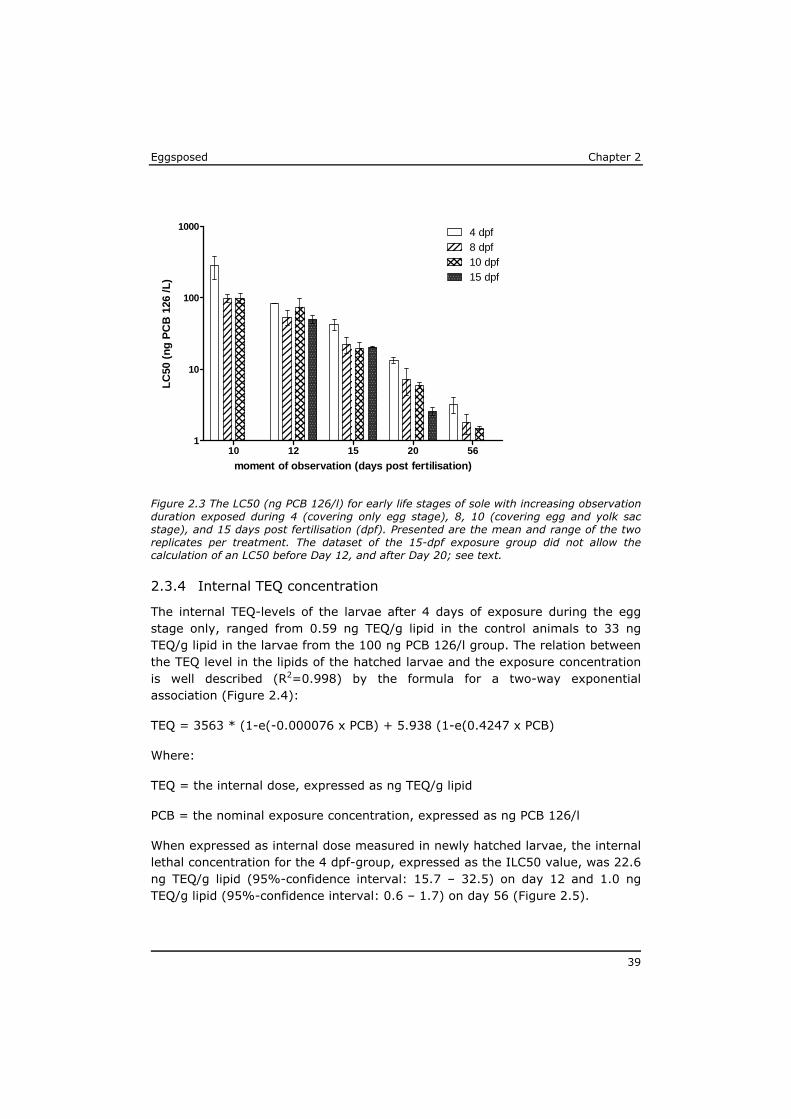
Eggsposed Chapter 2
39
Figure 2.3 The LC50 (ng PCB 126/l) for early life stages of sole with increasing observation duration exposed during 4 (covering only egg stage), 8, 10 (covering egg and yolk sac stage), and 15 days post fertilisation (dpf). Presented are the mean and range of the two replicates per treatment. The dataset of the 15-dpf exposure group did not allow the calculation of an LC50 before Day 12, and after Day 20; see text.
2.3.4 Internal TEQ concentration
The internal TEQ-levels of the larvae after 4 days of exposure during the egg stage only, ranged from 0.59 ng TEQ/g lipid in the control animals to 33 ng TEQ/g lipid in the larvae from the 100 ng PCB 126/l group. The relation between the TEQ level in the lipids of the hatched larvae and the exposure concentration is well described (R2=0.998) by the formula for a two-way exponential association (Figure 2.4):
TEQ = 3563 * (1-e(-0.000076 x PCB) + 5.938 (1-e(0.4247 x PCB)
Where:
TEQ = the internal dose, expressed as ng TEQ/g lipid
PCB = the nominal exposure concentration, expressed as ng PCB 126/l
When expressed as internal dose measured in newly hatched larvae, the internal lethal concentration for the 4 dpf-group, expressed as the ILC50 value, was 22.6 ng TEQ/g lipid (95%-confidence interval: 15.7 – 32.5) on day 12 and 1.0 ng TEQ/g lipid (95%-confidence interval: 0.6 – 1.7) on day 56 (Figure 2.5).
10 12 15 20 561
10
100
10004 dpf8 dpf10 dpf15 dpf
moment of observation (days post fertilisation)
LC
50 (
ng
PC
B 1
26 /
L)
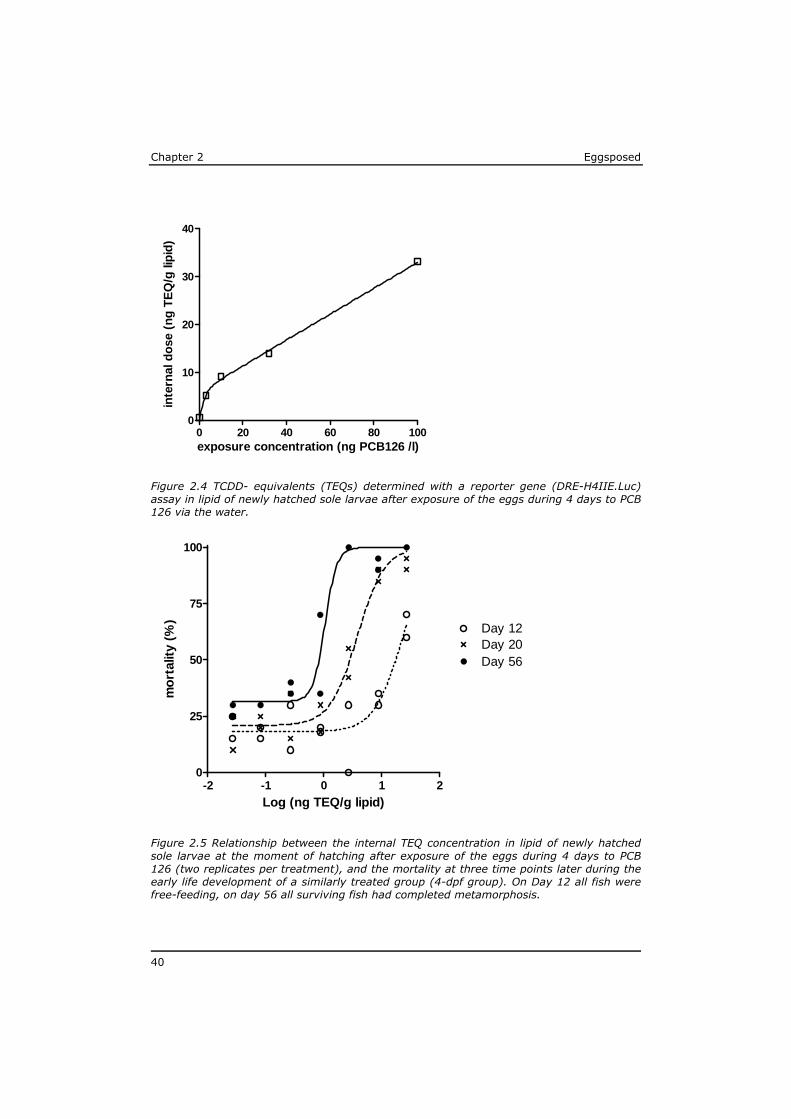
Chapter 2 Eggsposed
40
Figure 2.4 TCDD- equivalents (TEQs) determined with a reporter gene (DRE-H4IIE.Luc) assay in lipid of newly hatched sole larvae after exposure of the eggs during 4 days to PCB 126 via the water.
Figure 2.5 Relationship between the internal TEQ concentration in lipid of newly hatched sole larvae at the moment of hatching after exposure of the eggs during 4 days to PCB 126 (two replicates per treatment), and the mortality at three time points later during the early life development of a similarly treated group (4-dpf group). On Day 12 all fish were free-feeding, on day 56 all surviving fish had completed metamorphosis.
0 20 40 60 80 1000
10
20
30
40
exposure concentration (ng PCB126 /l)
inte
rnal
do
se (
ng
TE
Q/g
lip
id)
-2 -1 0 1 20
25
50
75
100
Day 12
Day 56Day 20
Log (ng TEQ/g lipid)
mo
rtal
ity
(%)

Eggsposed Chapter 2
41
2.4 Discussion
2.4.1 Experimental set-up
The moment of hatching and becoming free-feeding differed 1-2 days between experiments. This might be related to the 1º C difference in temperature, as the development of early life stages of sole (and other fish) is strongly temperature dependent (Imsland et al., 2003). The time required by the surviving fish to complete metamorphosis seemed longer in the second experiment, but this was only due to the application of the stricter criterion for ‘complete metamorphosis’.
As the test compound was applied in the water column, the only uptake route for the exposure periods 4 dpf and 8 dpf which ended before the fish became free-feeding, was via the water. With longer exposure conditions some indirect exposure will have occurred via PCB adsorbed to the food, the artemia nauplii. This period with an additional route of exposure lasted 1 and 5 days for the 10-dpf and 15-dpf group, respectively. However, when the LC50 values for the various exposure durations are compared (Figure 2.3) there are no indications that indirect exposure via the food affected the outcome of the tests. The differences between LC50 values are related to the exposure duration and in absolute sense strongly determined by the moment of observation.
2.4.2 Effect type and effect concentrations
At the moment that all fish were free-feeding, which is the moment that a standard ELS fish test (EPA, 1996; OECD, 1992) could have been terminated, the LC50 values ranged between 39 and 82 ng PCB 126/l. However, with a prolonged observation period the LC50-values in the sole test were revealed to be 22 times lower (1.7 to 3.7 ng PCB 126/l). Since exposure to the test substance had already stopped after 4 to 15 days, this is not due to higher body burdens. Internal concentrations will even have decreased in time due to growth dilution. The first 4 dpf appear to be the main critical period for exposure to PCB 126. The delayed effects become apparent during the first week that the fish are free-feeding and resulted in an increased mortality rate.
A substantial number of the exposed fish that survived this first critical period developed oedema in the abdominal region about 20 days later, and died before metamorphosis. Oedema is an indication of a disturbed water balance (Stouthart et al., 1998; Hill et al., 2004) and is often found in ELS fish tests with dioxin-like toxicants. It is then referred to as blue-sac disease that is characterised by pericardial and yolk-sac oedema (Stouthart et al., 1998; Zabel et al., 1995; Hill et al., 2004; Brinkworth et al., 2003). The indications of blue-sac disease develop at early larval stages or directly after the absorption of the yolk-sac. In

Chapter 2 Eggsposed
42
our tests the oedema was observed in a much later stage of the development which might, amongst other reasons, be related to the low test concentrations that we applied in combination with a long observation period.
Delayed effects of PCB exposure (including oedema) have also been described for Xenopus laevis tadpoles where the exposure of embryos to PCB 126, PCB 77 or PCB-mixtures during 4 dpf did not affect larval development during exposure, but resulted in increased mortality and delayed metamorphosis several weeks later (Gutleb et al., 1999; 2007a). Based on these results a prolonged FETAX (Frog Embryo Teratogenic Assay - Xenopus) was proposed for compounds without acute effects (Gutleb et al., 2007a).
The clearly delayed adverse effect of early exposure, as we found in our tests, is also not covered in the standardised ELS fish tests described in OECD 210 and 212, nor in the new embryo fish test that is currently being developed (OECD, 2006). Therefore, these tests may seriously underestimate the toxic potency of single compounds or environmental extracts without acute teratogenicity.
2.4.3 Field relevance
The 4-dpf group that was only exposed during the egg stage already showed considerable effects with effect concentrations in the same order of a magnitude as the groups that were exposed during a substantial part of the early development. This makes the reproduction success of the fish susceptible for lipophilic persistent compounds that are accumulated by the mother fish and transported to the eggs during gametogenesis (Solbakken et al., 1984; Vodicnic & Peterson, 1985).
When expressed as internal TEQ concentration in the newly hatched larvae the ILC50 after 56 days is 1.0 ng TEQ/g lipid for the 4-dpf group. This level can be compared with TEQ levels on lipid basis in adult fish to get an indication of the field relevance of these effect concentrations. The TEQ levels in field-caught fish range from 4 pg TEQ/g lipid in sardinella (Leonards et al., 2000) up to 3764 pg TEQ/g lipid in flounder from the heavily polluted Grenland fjords in Norway (Knutzen et al, 2003) on the basis of by no means an exhaustive literature review. Fourteen (20%) out of the 70 samples that we identified in our short literature search contained more than 250 pg TEQ /g lipid (Table 2.1). This is 25% of the ILC50 we determined in our prolonged sole ELS test after exposure during 4 dpf. The TEQ levels detected in the two flounder samples from the Grenland fjords even exceeded our ILC50.
It is important to realise that exposure of the sole eggs in our experiment only started at ca. 12 hours instead of immediately after fertilisation as is the case
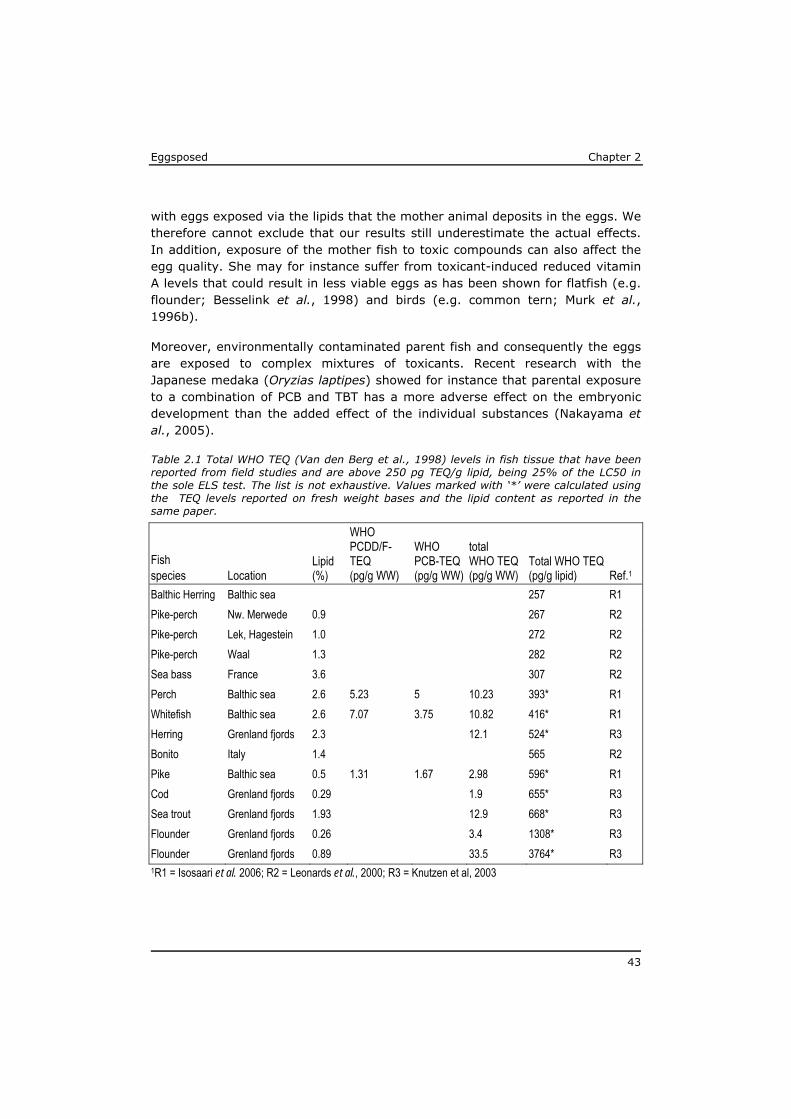
Eggsposed Chapter 2
43
with eggs exposed via the lipids that the mother animal deposits in the eggs. We therefore cannot exclude that our results still underestimate the actual effects. In addition, exposure of the mother fish to toxic compounds can also affect the egg quality. She may for instance suffer from toxicant-induced reduced vitamin A levels that could result in less viable eggs as has been shown for flatfish (e.g. flounder; Besselink et al., 1998) and birds (e.g. common tern; Murk et al., 1996b).
Moreover, environmentally contaminated parent fish and consequently the eggs are exposed to complex mixtures of toxicants. Recent research with the Japanese medaka (Oryzias laptipes) showed for instance that parental exposure to a combination of PCB and TBT has a more adverse effect on the embryonic development than the added effect of the individual substances (Nakayama et al., 2005).
Table 2.1 Total WHO TEQ (Van den Berg et al., 1998) levels in fish tissue that have been reported from field studies and are above 250 pg TEQ/g lipid, being 25% of the LC50 in the sole ELS test. The list is not exhaustive. Values marked with ‘*’ were calculated using the TEQ levels reported on fresh weight bases and the lipid content as reported in the same paper.
Fish species Location
Lipid (%)
WHO PCDD/F-TEQ (pg/g WW)
WHO PCB-TEQ (pg/g WW)
total WHO TEQ (pg/g WW)
Total WHO TEQ (pg/g lipid) Ref.1
Balthic Herring Balthic sea 257 R1
Pike-perch Nw. Merwede 0.9 267 R2
Pike-perch Lek, Hagestein 1.0 272 R2
Pike-perch Waal 1.3 282 R2
Sea bass France 3.6 307 R2
Perch Balthic sea 2.6 5.23 5 10.23 393* R1
Whitefish Balthic sea 2.6 7.07 3.75 10.82 416* R1
Herring Grenland fjords 2.3 12.1 524* R3
Bonito Italy 1.4 565 R2
Pike Balthic sea 0.5 1.31 1.67 2.98 596* R1
Cod Grenland fjords 0.29 1.9 655* R3
Sea trout Grenland fjords 1.93 12.9 668* R3
Flounder Grenland fjords 0.26 3.4 1308* R3
Flounder Grenland fjords 0.89 33.5 3764* R3 1R1 = Isosaari et al. 2006; R2 = Leonards et al., 2000; R3 = Knutzen et al, 2003

Chapter 2 Eggsposed
44
2.5 Conclusions
This study shows that the flatfish sole is a suitable marine test organism to perform early life stage tests including the phase of full metamorphosis. The sole is a test species that is relevant for the North Sea area and sole eggs are becoming more easily available since aquaculture activities are increasing world-wide.
We demonstrated that, at least for sole, the moment of observation has a significant impact on the effect concentration, even without further exposure. Exposure of only the eggs to non-acutely toxic compounds such as the dioxin-like PCB 126 resulted in delayed effects that are not covered in the standardised ELS fish tests. Therefore, these tests may seriously underestimate the toxicity of single compounds or environmental mixtures without acute toxicity.
Our study results suggest that the reproductive success of fish populations at contaminated sites can be expected to be negatively affected by (mixtures of) persistent compounds including dioxin-like compounds that are accumulated by the female fish and passed on to the eggs.
2.6 References
Ahlborg, U.G., Brouwer, A., Fingerhut, M.A., Jacobsen, J.L., Jacobsen, S.W., Kennedy, S.W., Kettrup, A.A.F., Koeman, J.H., Poiger, H., Rappe, C., Safe, S.H., Seegal, R.F., Tuomisto, J., and van den Berg, M. 1992. Impact of polychlorinated dibenzo-p-dioxins, dibenzofurans, dioxine and biphenyls on human and environmental health, with special emphasis on application of the toxic equivalency factor concept. Eur. J. Pharm. Environ. Toxicol. 228, 179–199.
Besselink H.T., Flipsen, E.M.T.E., Eggens, M.L., Vethaak, D., Koeman, J.H., Brouwer, A., 1998. Alterations in plasma and hepatic retinoid levels in flounder (Platichthys flesus) after chronic exposure to contaminated harbour sludge in a mesocosm study. Aquat. Toxicol. 42, 271–285.
Besselink H.T., Schipper, C., Klamer, H., Leonards, P., Verhaar, H., Felzel, E., Murk, A.J., Thain, J., Hosoe, K., Schoeters, G., Legler, J., Brouwer, B., 2004. Intra- and interlaboratory calibration of the DR Calux bioassay for the analysis of dioxins and dioxin-like chemicals in sediments. Environ. Toxicol. Chem., 23, 2781–2789.
Brinkworth, L.C., Hodson, P.V., Tabash, S., Lee P., 2003. CYP1a Induction and blue-sac disease in early developmental stages of rainbow trout (Oncorhynchus mykiss) exposed to retene. J. Toxicol. Environ. Health. 66, 526-646.
Brouwer, A., Ahlborg, U.G., van den Berg, M., Birnbaum, L.S., Boersma, R.E., Bosveld, B., Denison, M.S., Gray, L.E., Hagmar, L., Holene, E., Huisman, M., Jacobson, S.W., Jacobson, J.L., Koopman-Esseboom, C., Koppe, J.G., Kulig, B.M., Morse, D.C., Muckle, G., Peterson, R.E., Sauer, P.J.J., Seegal, R.F., Smits-van Prooije, A.E.,

Eggsposed Chapter 2
45
Touwen, B.C.L., Weisglas-Kuperus, N., Winneke G., 1995. Functional aspects of developmental toxicology of polyhalogenated aroma-tic hydrocarbons in experimental animals and human infants. Eur. J. Pharmacol., 293, 1-40.
Brown S.B., Adams, B.A., Cyr, D.G., , Eales J.G., 2004. Contaminant effects on the teleost fish thyroid. Environ. Toxicol. Chem. 23, 1680-1701.
Crane H.M., Pickford, D.B., Hutchinson, T.H., Brown, J.A., 2006. The effect of methimazole on development of the fathead minnow Pimephales promelas, from embryo to adult. Toxicol. Sci. 93, 278-285.
EPA, 1996. Ecological Effects Test Guidelines OPPTS 850.1400 Fish Early-Life Stage Toxicity Test. Environmental Protection Agency. Washington, USA.
Gutleb, A.C., Appelman J., Bronkhorst M.C. 1999. Delayed effects of pre- and early-life time exposure to polychlorinated biphenyls on tadpoles of two amphibian species (Xenopus laevis and Rana temporaria). Environ. Toxicol. Pharmacol. 8, 1-14.
Gutleb, A.C., 2006. Detecting the effects of environmentally relevant concentrations of thyroid hormone disrupting compounds on amphibian development. Thesis Wageningen University, the Netherlands.
Gutleb, A.C., Mossinka, L., Schriks, M., van den Berg, H.J.H., Murk, A.J. (2007a): Delayed effects of environmentally relevant concentrations of 3,3′,4,4′-tetrachlorobiphenyl (PCB-77) and non-polar sediment extracts detected in the prolonged-FETAX. Sci. Total Environ. 381, 307–315.
Gutleb, A.C., Schriks, M., Mossink, L., van den Berg, H.J.H., Murk, A.J., 2007b. A synchronized amphibian metamorphosis assay as an improved tool to detect thyroid hormone disturbance by endocrine disruptors and apolar sediment extracts. Chemosphere 70 (1), 93-100.
Hill A.J., Bello, S.M., Prasch, A.L., Peterson, R.E., Heidemann, W., 2004. Water permeability and TCDD-induced edema in zebrafish early life stages. Toxicol. Sci. 78, 78-87.
Hoogenboom R., Bovee, T., Traag, W., Hoogerbrugge, R., Baumann, B., Portier, L., van de Weg, G., de Vries, J., 2006. The use of DRE-CALUX bioassay and indicator polychlorinated biphenyles for screening of elevated levels of dioxins and dioxin-like polychlorinated biphenyls in eel. Mol. Nutr. Food. Res. 50, 945-957.
Hutchington T.H., Solbe, J., Kloepper-Sams, P., 1998. Analysis of the ecetoc aquatic toxicity (EAT) database. III-Comparative toxicity of chemical substances to different life stages of aquatic organisms. Chemosphere Vol. 36 No. 1, 129-142.
Imsland, A.K., Foss, A., Conceicao, L.E.C., Dinis, M.T., Delbare, D., Schram, E., Kamstra, A., Rema, P. , White, P., 2003. A review of the culture potential of Solea solea and S. senegalensis. Reviews in fish biology and fisheries 13, 379-407.
Isosaari, P., Hallikainen, A., Kiviranta, H., Vuorinen, P.J., Parmanne, R., Koistinen, J., Vartiainen, T., 2006. Polychlorinated dibenzo-p-dioxins, dibenzofurans, biphenyls, naphthalenes and polybrominated diphenyl ethers in the edible fish caught from the Baltic Sea and lakes in Finland. Environmental Pollution 141, 213-225.
Knutzen, J., Bjerkeng, B., Naes, K., Slabach, M., 2003. Polychlorinated dibenzofurans/dibenzo-p-dioxins (PCDF/PCDDs) and other dioxin-like substances in marine organisms from the Greenland fjords, S. Norway, 1975-2001: present

Chapter 2 Eggsposed
46
contamination levels and specific accumulation of PCDF/PCDD congeners. Chemosphere 52, 745-760.
Kuiper, R.V., Cantón, R.F., Leonards, P.E.G., Jenssen, B.M., Dubbeldam, M., Wester, P.W., van den Berg, M., Vethaak, A.D., 2007. Long-term exposure of European flounder (Platichthys flesus) to the flame-retardants tetrabromobisphenol A (TBBPA) and hexabromocyclododecane (HBCD). Ecotoxicology and Environmental Safety 67 (3), 349-360.
Leonards, P.E.G., Lohman, M., de Wit, M.M., Booy, G., Brandsma, S.H., de Boer, J., 2000. Actual situation on chlorinated dioxins, furans and PCBs in fisheries products: Quick- en Full-Scan. RIVO-Report C034/00 (in Dutch), IJmuiden, the Netherlands.
Martinez, G.M., Bolker, J.A., 2003. Embryonic and larval staging of summer flounder (Paralichtys dentatus). Journal of Morphology 255, 162-176.
Murk, A.J., Legler, J., Denison, M.S., Giesy, J.P. van de Guchte, C., Brouwer A., 1996a. Chemical-activated luciferase expression (CALUX): a novel in-vitro bioassay for Ah receptor active compounds in sediments and pore water. Fundam. Appl. Toxicol. 33, 1, 149-160.
Murk, A.J., Boudewijn, T.J., Meininger, P.L., Bosveld, A.T.C., Rossaert, G., Ysebaert, T., Meire, P., Dirksen, S. 1996b. Effects of Polyhalogenated Aromatic Hydrocarbons and Related Contaminants on Common Tern Reproduction: Integration of Biological, Biochemical and Chemical Data. Arch. Environ. Contam. Toxicol. 31, 128-140.
Murk, A.J., Leonards, P.E.G., van Hattum, B., Luit, R., van der Weiden, M.E.J., Smit, M., 1998. Application of biomarkers for exposure and effect of polyhalogenated aromatic hydrocarbons in naturally exposed European otters (Lutra lutra). Environmental toxicology and pharmacology 6, 91-102.
Nakayama, K., Oshima, Y., Nagafuchi, K., Hano, T., Shimasaki, Y., Honjo, T., 2005. Early-life stage toxicity in offspring from exposed parent medaka, Oryzias laptipes, to mixtures of tributyltin and polychlorinated biphenyls. Environmental Toxicology and Chemistry 24, no. 3, 591-596.
OECD, 1992. OECD Guideline for testing of chemicals 210: Fish, Early Life Stage Toxicity Test. Adopted 17.07.92. Organization for Economic Co-operation and Development, Paris.
OECD, 1998. OECD Guideline for testing of chemicals 212: Fish, short term Toxicology test on Embryo and sac-fry Stages. Adopted 21st September 1998. Organization for Economic Co-operation and Development, Paris.
OECD, 2006. Fish Embryo Toxicity (FET) test. Draft proposal for a new guideline May 30, 2006 (1st Version). Organization for Economic Co-operation and Development, Paris.
Safe S,. (1992): Development, validation and limitations of toxic equivalency factors. Chemosphere 25, issues 1-2, 61-64.
Solbakken, J.E., Ingebrigtsen, K., Palmork, K.H., 1984. Comparative study on the fate of the polychlorinated biphenyl 2,4,5,2,4,5-hexachlorobiphenyl and the polycyclic aromatic hydrocarbon phenanthrene in flounder (Platichthys flesus), determined by liquid scintillation counting and autoradiography. Marine Biology 83, 239-246.

Eggsposed Chapter 2
47
Schreiber, A.M., 2006. Asymmetric craniofacial remodelling and lateralized behaviour in larval flatfish. The Journal of Experimental Biology 209, 610-621.
Stronkhorst, J., Leonards, P., Murk, A.J., 2002. Using the dioxin receptor-CALUX in vitro bioassay to screen marine harbour sediments for compounds with a dioxin-like mode of action. Environmental Toxicology and Chemistry, Vol. 21, No. 12, 2552-2561.
Stouthart, X.J.H.X., Huijbregts, M.A.J., Balm, P.H.M., Lock, R.A.C. , Bonga, W., 1998.Endocrine stress response and abnormal development in carp (Cyprinus carpio) larvae after exposure of the embryos to PCB 126. Fish Physiology and Biochemistry 18: 321-329.
Van den Berg, M., Birnbaum, L., Bosveld, A.T.C., Brunström, B., Cook, P., Feeley, M., Giesy, J.P, Hanberg, A., Hasegawa, R., Kennedy, S.W., Kubiak, T., Larsen, J.C., van Leeuwen, F.X.R., Liem, A.K.D, Nolt, C., Peterson, R.E., Poellinger, L., Safe, S., Schrenk, D., Tillitt, D., Tysklind, M., Younes, M., Waern, F., Zacharewski T. 1998. Toxic Equivalency Factors (TEFs) for PCBs, PCDDs, PCDFs for humans and wildlife. Environ. Health Persp. 106 No. 12, 775-792.
Van Leeuwen, S.P.J., Leonards, P.E.G., Traag, W.A., Hoogenboom, L.A.P., de Boer, V.C.J., 2007. Polychlorinated dibenzo-p-dioxins, dibenzofurans and biphenyls in fish from the Netherlands: concentrations, profiles and comparison with DR CALUX bioassay results. Analytical and Bioanalytical Chemistry 389 (1), 321 - 333.
Vethaak, A.D. , Lahr, J., Schrap, S.M., Belfroid, A.C., Rijs, G.B.J., Gerritsen, A., De Boer, J., Bulder, A.S., Grinwis, G.C.M., Kuiper, R.V., Legler, J., Murk, T.A.J., Peijnenburg, W., Verhaar, H.J.M., De Voogt, P., 2005. An integrated assessment of estrogenic contamination and biological effects in the aquatic environment of The Netherlands. Chemosphere, Vol. 59, Issue 4, 511-524.
Vodicnic, M.J., Peterson, R.E., 1985. The enhancing effect of spawning on elimination of a persistent polychlorinated biphenyl from female yellow perch. Fundamental and Applied Toxicology 5, 770-776.
Wedekind, C., von Siebenthal, B., Gingold, R., 2007. The weaker points of fish acute toxicity tests and how tests on embryos can solve some issues. Environmental Pollution 148, 385-389.
Yamano, K., 2005. The role of thyroid hormone in fish development with reference to aquaculture. Japan Agricultural Research Quarterly, 39 (3), 106-168.
Zabel, E.W., Cook, P.M., Peterson, R.E., 1995. Potency of 3,3,4,4,5-pentachlorobiphenyl (PCB 126), alone and in combination with 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD), to produce lake trout early life-stage mortality Environmental Toxicology and Chemistry. Vol. 14, No 12, 2175-2179.


49
CHAPTER 3
Critical body burden of maternally transferred methyl-triclosan for the flatfish
Solea solea
E.M. Foekemaa, M. de Graafb, C. Kwadijkc, and A.J. Murkd
Submitted in extended form
a IMARES, Department Experimental Ecology b University of Groningen, Department of Marine Benthic Ecology and Evolution c IMARES, Department Fish d Wageningen University, Division of Toxicology

Chapter 3 Eggsposed
50
Abstract
In order to mimic exposure through maternal transfer, fertilized eggs of the marine flatfish Solea solea were exposed to a concentration series of methyltriclosan (MTCS) in water until hatching (6 days post fertilization, dpf), and were then allowed to develop further under unexposed conditions until completed metamorphoses, 40 dpf. Mortality occurred in the higher treatment levels until 20 dpf, and all surviving larvae completed metamorphoses without problems. Internal effect concentrations (IEC), reached in larvae at the end of the exposure (6 dpf), were 5.8 mg/g lipid weight (lw) and 2.1 mg/g lw for IEC50 and IEC10 respectively. These internal effect concentrations are at least 200 times higher than concentrations that due to maternally transfer can be expected in the eggs of highly exposed fish in a field situation. Our results thus do not indicate a high risk from maternally transferred MTCS for fish at the current field concentrations. However, the impact of mixtures including MTCS, the parent compound triclosan, and other persistent lipophilic substances still needs further assessment.

Eggsposed Chapter 3
51
3.1 Introduction
Methyltriclosan (MTCS) is formed by microbiological methylation of triclosan (TCS) that is commonly used as bactericide in a wide variety of human care products. Effluents from sewage treatment plants form a major source of TCS and MTCS to the environment (Coogan et al., 2007; Coogan and La Point, 2008; Dann and Hontela, 2011; Fernandes et al., 2011), and traces have been detected in the marine environment (Andresen et al., 2007; Fernandes et al., 2011; Xie et al., 2008). MTCS is suggested to be more persistent than the parent compound (Leiker et al., 2009), and this persistence combined with a log Kow of 5.0 to 5.5 (Valo and Salkinojasalonen, 1986) gives concern about uptake in the food chain. Indeed MTCS has been detected in tissue of fresh water fish (Adolfsson-Erici et al., 2002;Balmer et al., 2004; Leiker et al., 2009), and therefore it is suggested that further research is needed, especially with regards to the bioaccumulation potential and toxicity (Dann and Hontela, 2011), and with respect to the potential of MTCS to interfere with the thyroid hormone system (Hinther et al., 2011).
A typical exposure route for bioaccumulation substances is maternal transfer. Substances like MTCS, that have accumulated in the mother’s tissues are transferred to the eggs (Russell et al., 1999) causing an early exposure of the developing embryo. Since the thyroid hormone system plays an important role during early development in many species including fish (e.g. Klaren et al., 2008; Yu et al., 2011), maternally transferred MTCS poses a potential risk for developing fish larvae. The aim of the study was to determine critical internal (no-)effect levels for MTCS in fish eggs on the survival and development. By a short exposure of fertilized eggs, maternal transfer of the contaminants to the egg was mimicked. Earlier research with PCBs and TCDD has indicated that such exposure results in comparable effects on the developing larvae as actual maternal exposure (Walker and Peterson, 1994).
As far as we know critical body burdens for MTCS have not been determined so far. The common sole (Solea solea) was chosen as test species, as during early development this species undergoes the obvious metamorphosis typical for all flatfishes. As this metamorphosis is thyroid hormone mediated (Klaren et al., 2008) it is potentially sensitive for disruption by MTCS (Hinther et al., 2011).

Chapter 3 Eggsposed
52
3.2 Materials and methods
3.2.1 Test procedure
The test was performed according to Foekema et al., 2008 where details of the test procedure are reported. In summary, fertilized eggs from Solea solea were exposed to a concentration series of MTCS (Methyltriclosan, purity 99.7%, Sigma-Aldrich) in 0.1% DMSO (Dimethylsulfoxide, purity 99.9%, A.C.S. spectrophotometric grade, supplier SIGMA-Aldrich) until full hatching 6 days post fertilization (dpf). Nominal exposure concentrations were 4, 12, 37, 111, 333 and 1000 µg MTCS/L.
Per test concentration two groups of 15 larvae were allowed to further develop until the end of the metamorphosis at 40 dpf in glass beakers containing clean seawater without DMSO or MTCS. From 9 dpf onwards, when the larvae are about to start external feeding, they were fed ad libitum with newly hatched nauplli of Artemia salina on a daily basis. Every other day, approximately 75% of the water volume in each glass beaker was replaced. On an almost daily bases mortality and development was recorded, and dead larvae, faeces and surplus food items were removed. Fish that completed metamorphosis were taken out of the test, narcotized and subsequently killed in a solution consisting of 500 mg of ethyl 3-aminonebzoate methanesulfonic acid salt (purity 98%, ACROS Organics) and 200 mg NaHCO3 (purity 99.0%, Fluka Chemika) in 1 liter seawater). The moment of completed metamorphosis, body size and observable malformations were recorded.
The experiments were approved by the Animal Care Committee of the Animal Sciences Group of Wageningen UR.
3.2.2 Chemical analysis
Those larvae that were not transferred to clean water on 6 dpf were sacrificed for chemical analysis of the lipid-normalized MTCS concentration. For that they were taken from the exposure beakers with a polyethylene pipette and transferred to clean seawater to rinse remaining exposure water, and within 15 minutes collected by filtration on a Whatman GF/C filter with known wet (seawater) weight. The filter with the larvae was weighed again and stored at -20°C until preparation for analysis. For that, the filters containing the larvae were subsequently extracted 3 times by ultrasonification with 4 ml of a mixture of 50% pentane – 50% dichloromethane (both substances picograde; LGC standards, Germany). The extract was transferred to a test tube and evaporated to dryness under a gentle stream of nitrogen. The residue was reconstituted in 1 ml injection standard (PCB112, purity >99%, LGC standards) and transferred to

Eggsposed Chapter 3
53
a vial for GC/MS analysis using an Agilent 6890 gas chromatograph (GC) coupled to an Agilent 5973 MSD quadropole mass spectrometer (MS) detector (Amstelveen, The Netherlands). Analysis was performed in electron impact (EI) mode monitoring m/z 302 and 304 for MTCS and m/z 326 and 324 for PCB112 as quantifier and qualifier ion.
3.2.3 Data analysis
Internal effect concentrations (IEC) for survival and normal development were determined by fitting the formula for a 'sigmoid dose response curve with variable slope' as incorporated in GraphPad Prism (version 4.03) to the dataset. Both the IEC50 with 95%-confidence intervals as well as the IEC10 for survival and development were determined from the curve.
3.3 Results and discussion
In the solvent controls more than 90% of the larvae developed into normal juvenile fish. Hatching success at the end of the exposure period (6 dpf) was not affected by any of the treatments. The newly hatched larvae in treatments 333 µg MTCS/l and 1000 µg MTCS/l however, were in poor condition and complete mortality occurred in these groups within the next two days (Figure 3.1a). In this stage no increased mortality was seen in 111 µg MTCS/l, the next lower exposure group. Between 15 and 20 dpf, when yolk had been fully absorbed, mortality increased substantially. After 20 dpf no further mortality occurred and all surviving fish developed normally, with onset of metamorphoses around 25 dpf, comparable to the controls. At the end of the test no fish were malformed and therefore effect concentrations for survival and normal development were comparable.
The ILC50 and ILC10 expressed as lipid normalized body burden on 6 dpf (Figure 3.1b) were 5.8 mg/g lw (95% confidence intervals 4.3 to 7.9 mg/g lw, r2=0.969) and 2.1 mg/g lw, respectively.
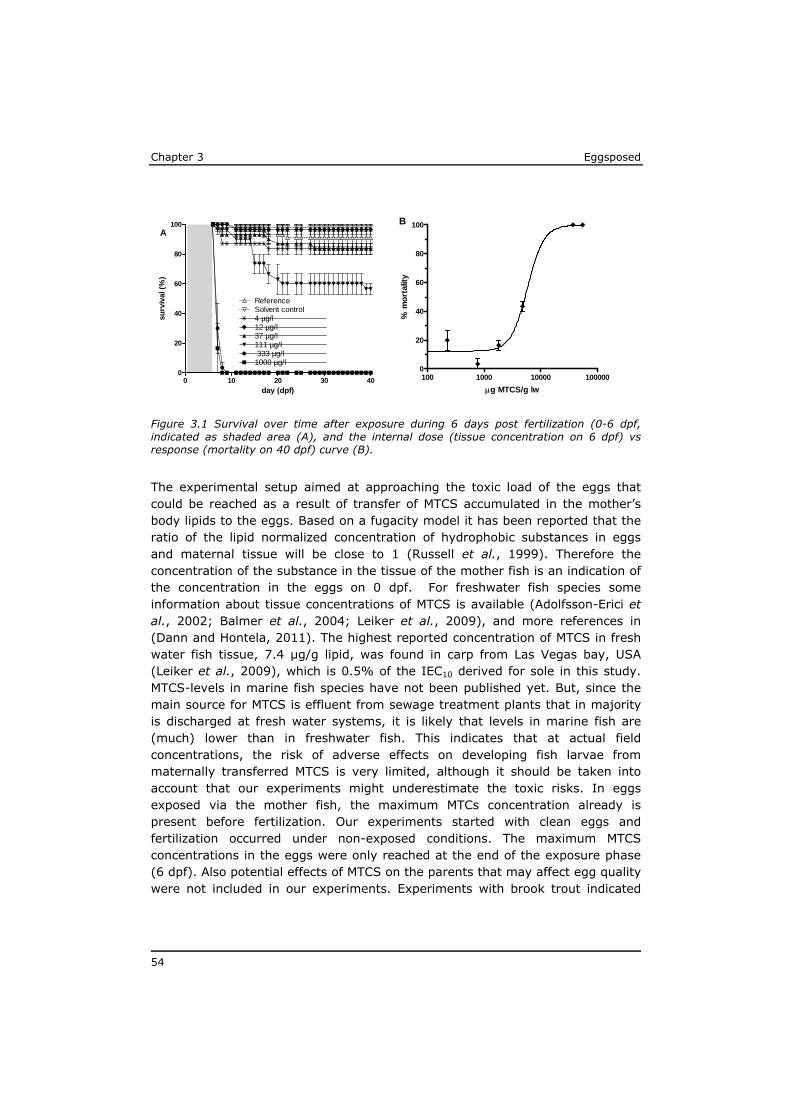
Chapter 3 Eggsposed
54
Figure 3.1 Survival over time after exposure during 6 days post fertilization (0-6 dpf, indicated as shaded area (A), and the internal dose (tissue concentration on 6 dpf) vs response (mortality on 40 dpf) curve (B).
The experimental setup aimed at approaching the toxic load of the eggs that could be reached as a result of transfer of MTCS accumulated in the mother’s body lipids to the eggs. Based on a fugacity model it has been reported that the ratio of the lipid normalized concentration of hydrophobic substances in eggs and maternal tissue will be close to 1 (Russell et al., 1999). Therefore the concentration of the substance in the tissue of the mother fish is an indication of the concentration in the eggs on 0 dpf. For freshwater fish species some information about tissue concentrations of MTCS is available (Adolfsson-Erici et al., 2002; Balmer et al., 2004; Leiker et al., 2009), and more references in (Dann and Hontela, 2011). The highest reported concentration of MTCS in fresh water fish tissue, 7.4 μg/g lipid, was found in carp from Las Vegas bay, USA (Leiker et al., 2009), which is 0.5% of the IEC10 derived for sole in this study. MTCS-levels in marine fish species have not been published yet. But, since the main source for MTCS is effluent from sewage treatment plants that in majority is discharged at fresh water systems, it is likely that levels in marine fish are (much) lower than in freshwater fish. This indicates that at actual field concentrations, the risk of adverse effects on developing fish larvae from maternally transferred MTCS is very limited, although it should be taken into account that our experiments might underestimate the toxic risks. In eggs exposed via the mother fish, the maximum MTCs concentration already is present before fertilization. Our experiments started with clean eggs and fertilization occurred under non-exposed conditions. The maximum MTCS concentrations in the eggs were only reached at the end of the exposure phase (6 dpf). Also potential effects of MTCS on the parents that may affect egg quality were not included in our experiments. Experiments with brook trout indicated
0 10 20 30 400
20
40
60
80
100
ReferenceSolvent control4 µg/l12 µg/l37 µg/l111 µg/l 333 µg/l1000 µg/l
day (dpf)
surv
ival
(%
)
A
100 1000 10000 1000000
20
40
60
80
100
g MTCS/g lw
% m
ort
alit
y
B

Eggsposed Chapter 3
55
that the effects and internal effect concentrations of PCBs and TCDD were comparable for eggs that were exposed via the water phase, by injection or by maternal exposure (Walker and Peterson, 1994). However, this does not have to be true for MTCS, that can interfere with the thyroid hormone system (Hinther et al., 2011) of the parent fish. For other substances (polybrominated diphenyl ethers, PBDEs) with the potential to disrupt this hormone system, it has been shown that parental exposure can influence the levels of thyroid hormones in the eggs, resulting in decreased hatching and increased malformations (Yu et al., 2011). Finally, the effect levels we derived were based on single compounds, while in field situations fish are always exposed to cocktails of pollutants. This can give rise to mixture toxicity, e.g. when some compounds inhibit the cellular efflux pumps, an important defense mechanism against toxic compounds, as has been shown for the parent compound TCS (Anselmo et al., 2012).
We conclude that, although our results do not indicate a direct risk of MTCS for fish larvae through maternal exposure at the current field concentrations, the issues mentioned above should be taken into account and studied further. Special attention should go to the impact of mixtures of TCS and MTCS and other substances, especially those that are common in effluents from sewage treatment plants like metals and (residues of) pharmaceuticals.
3.4 References
Adolfsson-Erici, M., Pettersson, M., Parkkonen, J., Sturve, J., 2002. Triclosan, a commonly used bactericide found in human milk and in the aquatic environment in Sweden. Chemosphere 46, 1485-1489.
Andresen, J.A., Muir, D., Ueno, D., Darling, C., Theobald, N., Bester, K., 2007. Emerging pollutants in the north sea in comparison to Lake Ontario, Canada, data. Environmental Toxicology and Chemistry 26, 1081-1089.
Anselmo, H.M.R., Van Den Berg, J.H.J., Rietjens, I.M.C.M., Murk, A.T.J., 2012. Inhibition of cellular efflux pumps involved in multi xenobiotic resistance (MXR) in echinoid larvae as a possible mode of action for increased ecotoxicological risk of mixtures. Ecotoxicology 21, 2276-2287.
Balmer, M.E., Poiger, T., Droz, C., Romanin, K., Bergqvist, P.A., Muller, M.D., Buser, H.R., 2004. Occurrence of methyl triclosan, a transformation product of the bactericide triclosan, in fish from various lakes in Switzerland. Environ. Sci. Technol. 38, 390-395.
Coogan, M.A., Edziyie, R.E., La Point, T.W., Venables, B.J., 2007. Algal bioaccumulation of triclocarban, triclosan, and methyl-triclosan in a North Texas wastewater, treatment plant receiving stream. Chemosphere 67, 1911-1918.
Coogan, M.A., La Point, T.W., 2008. Snail bioaccumulation of triclocarban, triclosan, and methyltriclosan in a North Texas, USA, stream affected by wastewater treatment plant runoff. Environmental Toxicology and Chemistry 27, 1788-1793.

Chapter 3 Eggsposed
56
Dann, A.B., Hontela, A., 2011. Triclosan: environmental exposure, toxicity and mechanisms of action. Journal of Applied Toxicology 31, 285-311.
Fernandes, M., Shareef, A., Kookana, R., Gaylard, S., Hoare, S., Kildea, T., 2011. The distribution of triclosan and methyl-triclosan in marine sediments of Barker Inlet, South Australia. Journal of Environmental Monitoring 13, 801-806.
Foekema, E.M., Deerenberg, C.M., Murk, A.J., 2008. Prolonged ELS test with the marine flatfish sole (Solea solea) shows delayed toxic effects of previous exposure to PCB 126. Aquatic Toxicology 90, 197-203.
Hinther, A., Bromba, C.M., Wulff, J.E., Helbing, C.C., 2011. Effects of Triclocarban, Triclosan, and Methyl Triclosan on Thyroid Hormone Action and Stress in Frog and Mammalian Culture Systems. Environ. Sci. Technol. 45, 5395-5402.
Klaren, P.H.M., Wunderink, Y.S., Yufera, M., Mancera, J.M., Flik, G., 2008. The thyroid gland and thyroid hormones in Senegalese sole (Solea senegalensis) during early development and metamorphosis. General and Comparative Endocrinology 155, 686-694.
Leiker, T.J., Abney, S.R., Goodbred, S.L., Rosen, M.R., 2009. Identification of methyl triclosan and halogenated analogues in male common carp (Cyprinus carpio) from Las Vegas Bay and semipermeable membrane devices from Las Vegas Wash, Nevada. Sci. Total Environ. 407, 2102-2114.
Russell, R.W., Gobas, F., Haffner, G.D., 1999. Maternal transfer and in ovo exposure of organochlorines in oviparous organisms: A model and field verification. Environ. Sci. Technol. 33, 416-420.
Valo, R., Salkinojasalonen, M., 1986. Microbial transformtation of polychlorinated phenoxy phenols. Journal of General and Applied Microbiology 32, 505-517.
Walker, M.K., Peterson, R.E., 1994. Toxicity of 2,3,7,8-tetrachlorodibenzo-P-dioxin to brook trout (Salvelinus fontinalis) during early development. Environmental Toxicology and Chemistry 13, 817-820.
Xie, Z.Y., Ebinghaus, R., Floser, G., Caba, A., Ruck, W., 2008. Occurrence and distribution of triclosan in the German Bight (North Sea). Environmental Pollution 156, 1190-1195.
Yu, L.Q., Lam, J.C.W., Guo, Y.Y., Wu, R.S.S., Lam, P.K.S., Zhou, B.S., 2011. Parental Transfer of Polybrominated Diphenyl Ethers (PBDEs) and Thyroid Endocrine Disruption in Zebrafish. Environ. Sci. Technol. 45, 10652-10659.

57
CHAPTER 4
Toxic concentrations in fish early life stages peak at critical moment
E.M. Foekemaa, A. Fischerb, M. Lopez Parronb, C. Kwadijkc, P. de Vriesd, and A.J. Murkb
Foekema EM, Fischer A, Parron ML, Kwadijk C, de Vries P, Murk AJ (2012): Toxic concentrations in fish early life stages peak at a critical moment. Environmental Toxicology and Chemistry, 31: 1381-1390.
a IMARES, Department Experimental Ecology b Wageningen University, Division of Toxicology c IMARES, Department Fish d IMARES, Department Maritime

Eggsposed Chapter 4
58
Abstract
During the development of an embryo into a juvenile, the physiology and behavior of a fish strongly change, affecting exposure to and uptake of environmental pollutants. Based on experimental data with sole (Solea solea) an existing bioaccumulation model was adapted and validated to calculate the development of concentrations of persistent organic pollutants in the tissue of the developing fish. Simulation revealed that toxic tissue concentrations of pollutants with log Kow >5 peak at the moment the larvae become free-feeding, when the lipid reserves are depleted. This may explain the delayed effects observed in fish early life stage experiments with exposed eggs. In the field, eggs can be exposed through maternal transfer to adult pollutant tissue concentrations, which will increase in the larva to peak tissue concentrations exceeding those of the adult fish. The results demonstrate the risk of underestimating the effects of lipophilic persistent organic pollutants with log Kow >5 in short term early life stage fish tests, and underline the importance of maternal transfer as an exposure route in the field situation.

Chapter 4 Eggsposed
59
4.1 Introduction
Early life stages of fish, as well as other organisms, are in general considered to be more sensitive to toxicants than fully-developed individuals from the same species (Hutchinson et al., 1998). This is related to a combination of critical developments during early life stages, the limited possibilities to compensate the impact of physiological processes affected (Crane et al., 2006), and large changes in the composition of the developing tissue that affect internal bioavailability. Early life stages of oviparous species have a unique exposure route as pollutants accumulated by the mother fish can be deposited in the egg together with lipids and proteins (Kelly et al., 2011; Russell et al., 1999). Maternal transfer is especially relevant for lipophilic persistent organic pollutants (POPs), such as polychlorinated biphenyls (PCBs) and dioxins that accumulate in lipids of the mother fish.
In general, bioaccumulation/exposure models focus on the fate of pollutants in fully developed fish in a stable environment (Bodiguel et al., 2009; Eichinger et al., 2010; Veltman et al., 2005). Sometimes the impact of seasonal fluctuations and life cycles on levels in the adult fish are included as well (Ng and Gray, 2009; Sijm et al., 1992; Veltman et al., 2005), but the early life stages never are addressed as such. This, however, is an interesting phase due to the drastic biological and physiological changes that take place in the animals, sometimes in the presence of maternally transferred pollutants. More insight in the development of pollutant tissue concentration will help the interpretation of the results from standard fish early life stages (ELS) toxicity tests (EPA, 1996; OECD, 1992, 1998, 2006) in which exposure mostly takes place through the water phase only.
In the present study we measured and modeled the water and tissue concentrations of POPs during the development of an embryo into a juvenile fish. The exposure experiment was conducted with eggs of aquacultured common sole (Solea solea), an oviparous marine flatfish. The fate of a selection of PCBs with a range of lipophilicities (log Kow values of ranging from 5.9 to 6.8) from the technical mixture Arochlor 1254 was measured in the exposure water and larvae. Based on knowledge of the main developmental stages of fish larvae (relevant information is given below), the existing OMEGA (Optimal Modeling for EcotoxicoloGical Applications) bioaccumulation model (Hendriks et al., 2001) was adapted to calculate the development of POP-concentrations in the tissue of the developing fish. After validation with measured concentrations, different exposure scenarios were simulated using the developed ELS- OMEGA model to calculate the development of the tissue concentrations of POPs in sole larvae in time.

Eggsposed Chapter 4
60
4.1.1 Fish developmental stages
In early life development of fish several stages can be distinguished (Blaxter, 1988; Jobling, 2002) of which three are most relevant for the present study: the egg stage, the yolk-sac stage and the free-feeding stage. The biological characteristics and possible exposure routes of those stages briefly described below, apply for the majority of oviparous fish species. The most obvious difference between species is the duration of the different stages (Blaxter, 1988), which also is temperature related. Below, the timing of the stages for our model fish Solea solea under experimental conditions is indicated.
At spawning, a fish egg contains all resources necessary for the development of the embryo into a larvae that is capable of external feeding. The majority of these resources is stored in the yolk by the mother fish during vitellogenesis (as reviewed by (Walsh, 1988) and during this process lipophilic pollutants are transferred to the eggs as well (De Bruyn et al., 2004; Serrano et al., 2008; Daley et al., 2009; Kelly et al., 2011). For our model calculations we assume a lipid normalized maternal-to-egg transfer ratio of 1 for all persistent organic substances, conform the fugacity model (Russel et al., 1999). Immediately after spawning, the eggs are fertilized and the chorion becomes a protective shield against physical damage. The uptake and excretion of pollutants in a fertilized fish egg is low compared to uptake in newly hatched larvae (Villalobos et al., 2000; Gonzalez-Doncel et al., 2003). This probably is related to the lack of actively circulated fluids through or near the chorion and the relatively small surface area-volume ratio of a fish egg in comparison to that of a fish larva (Petersen and Kristensen, 1988). In our calculations we assume that for the egg and the larva exposure and excretion are through the water column. This is obvious for the majority of the eggs from marine fish species, including Solea solea, that are more or less buoyant. But even for demersal eggs it is plausible that transfer of pollutants from sediments occurs via the (pore) water phase, as is also the case with organisms that have a more intense contact with the sediment (Lu et al., 2011).
Five days post fertilization (5 dpf), the sole eggs hatch. The so-called yolk-sac larvae are not capable of external feeding. As they completely rely on the yolk for nutrition, exposure and excretion still is limited to the water phase. The exchange with water is more intense than before hatching, for reasons mentioned above and will further intensify with developing gills. During the yolk-sac stage, the mouth and intestine develops, which enables the larvae to switch from endogenous to exogenous feeding before the yolk is depleted. For the sole larvae in our experiment this critical moment takes place around 10 dpf. From then on, the larva is referred to as a free-feeding larva and food is an additional
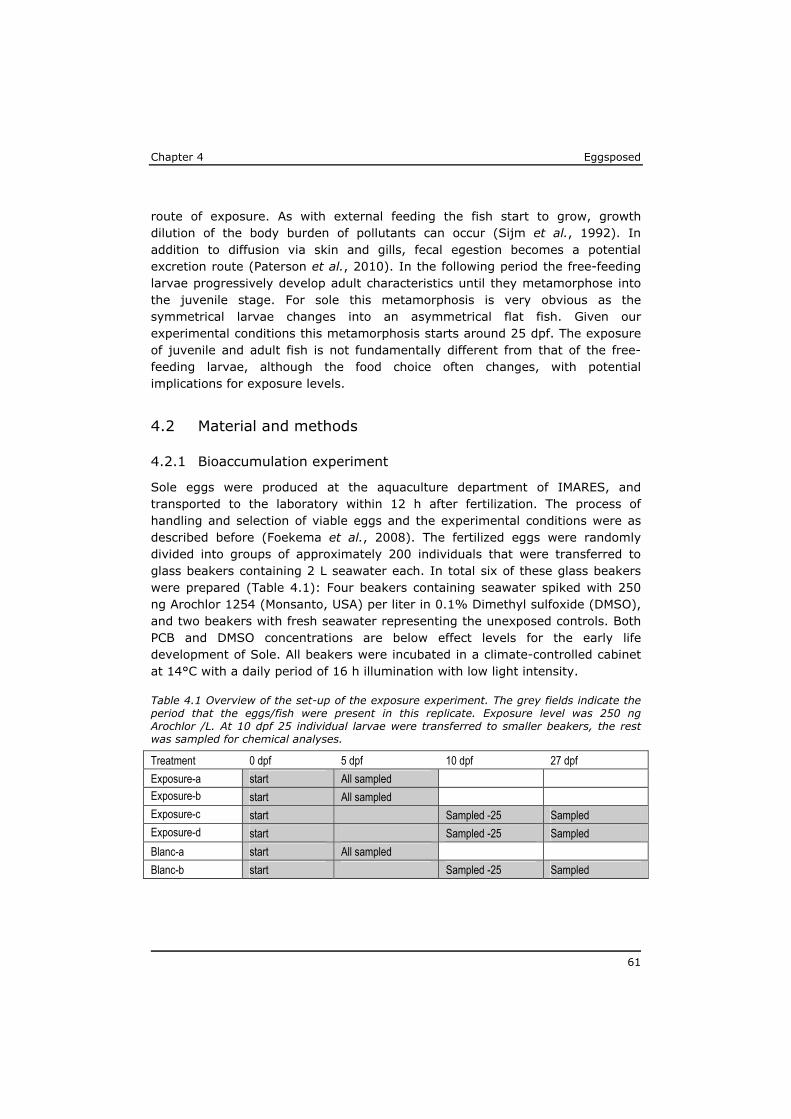
Chapter 4 Eggsposed
61
route of exposure. As with external feeding the fish start to grow, growth dilution of the body burden of pollutants can occur (Sijm et al., 1992). In addition to diffusion via skin and gills, fecal egestion becomes a potential excretion route (Paterson et al., 2010). In the following period the free-feeding larvae progressively develop adult characteristics until they metamorphose into the juvenile stage. For sole this metamorphosis is very obvious as the symmetrical larvae changes into an asymmetrical flat fish. Given our experimental conditions this metamorphosis starts around 25 dpf. The exposure of juvenile and adult fish is not fundamentally different from that of the free-feeding larvae, although the food choice often changes, with potential implications for exposure levels.
4.2 Material and methods
4.2.1 Bioaccumulation experiment
Sole eggs were produced at the aquaculture department of IMARES, and transported to the laboratory within 12 h after fertilization. The process of handling and selection of viable eggs and the experimental conditions were as described before (Foekema et al., 2008). The fertilized eggs were randomly divided into groups of approximately 200 individuals that were transferred to glass beakers containing 2 L seawater each. In total six of these glass beakers were prepared (Table 4.1): Four beakers containing seawater spiked with 250 ng Arochlor 1254 (Monsanto, USA) per liter in 0.1% Dimethyl sulfoxide (DMSO), and two beakers with fresh seawater representing the unexposed controls. Both PCB and DMSO concentrations are below effect levels for the early life development of Sole. All beakers were incubated in a climate-controlled cabinet at 14°C with a daily period of 16 h illumination with low light intensity.
Table 4.1 Overview of the set-up of the exposure experiment. The grey fields indicate the period that the eggs/fish were present in this replicate. Exposure level was 250 ng Arochlor /L. At 10 dpf 25 individual larvae were transferred to smaller beakers, the rest was sampled for chemical analyses.
Treatment 0 dpf 5 dpf 10 dpf 27 dpf
Exposure-a start All sampled Exposure-b start All sampled Exposure-c start Sampled -25 Sampled Exposure-d start Sampled -25 Sampled
Blanc-a start All sampled
Blanc-b start Sampled -25 Sampled

Eggsposed Chapter 4
62
After hatching, 5 d post fertilization (5 dpf), the larvae from two beakers of the exposure group and one of the control group were sampled for chemical analyses following the procedure described below. At 10 dpf, at the end of the yolk-sac stage, 25 larvae were randomly collected from each of the remaining three beakers (one control and two exposure groups) and transferred to smaller (200 ml) glass beakers containing clean seawater for the ‘control’ group and seawater spiked at 250 ng Arochlor1254 /L for the exposed group. The remaining fish larvae from the 2 L beakers were sampled for chemical analysis. The 3 x 25 selected larvae were fed ad libitum daily with newly hatched Artemia salina nauplii. Before being fed to the exposure group, the Artemia were placed in a 250 ng Arochlor 1254 /L solution for approximately 2 h. At 27 dpf, when the first fish reached the climax of metamorphoses, all fish were collected for chemical analyses.
Between 0 and 10 dpf, while the egg/larvae were kept in the 2 L-beakers, every second day approximately 75% of the water volume was replaced with new spiked or fresh seawater. After 10 dpf, the 25 fish in the smaller beakers were transferred every 48 h with a polyethylene pipet to a new beaker that contained seawater spiked at the appropriate level. In this way, contamination of the beakers with uneaten food, microbial fouling, and feces was minimized.
The experiment was approved by the Animal Care Committee of the Animal sciences group of Wageningen UR.
4.2.2 Sampling for chemical analyses
Using a semi-static test set up, exponential dissipation of the PCBs from the water column with time was expected (Koponen et al., 1998; Breitholtz et al., 2006). In order to get an impression of the range of the PCB concentrations in the test beakers, water samples were collected from the freshly made exposure solution at 0 dpf to determine the initial exposure concentration. At 5, 10 and 23 dpf water samples were collected from each individual test beaker. Sampling on these days was performed just before water replacement took place and these samples were thus indicative for the minimum exposure concentration.
For the determination of the background PCB concentration, unexposed eggs were sampled and stored at -20°C. The early life stages were sampled for chemical analyses immediately after hatching (5 dpf), at the end of the yolk-sac stage (10 dpf), and at the end of the experiment (27 dpf). The larvae were collected with a polyethylene pipette and while counting, transferred for maximum 15 min to unexposed water containing a narcotizing solution (100 mg Ethyl 3-aminonebzoate methanesulfonic acid salt -MS222, purity 98%, ACROS Organics-, and 200 mg NaHCO3 -purity 99.0%, Fluka Chemika- per 1 L

Chapter 4 Eggsposed
63
seawater). This also rinsed off the exposure water that initially came with the larvae. The narcotized larvae were collected by filtration on a pre-(wet)weighed filter (Whatman GF/C, glass microfiber filter) removing as much water as possible. After determination of the total wet weight, the filters with the larvae were stored at -20°C until chemical analyses.
Two independent samples of the spiked Artemia nauplii used as food source were collected by filtration, weighted, stored, and analyzed as the fish samples.
4.2.3 PCB analysis
In total, 29 PCB congeners were analysed following an accredited in-house method. In the present study we focus on the following six PCB congeners: PCB 52, 101, 110, 118, 138 and 153. The basis behind this selection are the seven indicator PCBs (PCB 28, 52, 101, 118, 138, 153, 180) applied by the International Council for the Exploration of the Sea (ICES), minus PCB 28 and 180, which levels in the exposure water were below the analytical limit of detection. PCB 110 was included to create a series with the range log Kow 5.9 to 6.8. Kow values for the selected PCBs were taken from Hansen et al., 1999 (Table 4.2).
PCB congeners as standards for chemical analysis (>99% purity) were purchased from Sigma-Aldrich, The Netherlands and CN Schmidt, The Netherlands. Pentane (picograde), dichloromethane (picograde) and isooctane (picograde) were purchased from LGC standards, Germany. Before analysis, the samples were spiked with 50 ng PCB 143 in 200 µl isooctane as internal standards and were then subsequently extracted three times by ultrasonification with 4 ml of a mixture of pentane - dichloromethane (50:50). The extract was transferred to a test tube, after which it was evaporated to dryness under a gentle stream of nitrogen. The residue was weighted to determine the lipid content of the sample and was then reconstituted in 1 ml injection standard and transferred to a vial for gas chromatography-mass spectrometry (GC/MS) analysis using a Agilent 6890 gas chromatograph (GC) coupled to an Agilent 5973 MSD quadrupole mass spectrometer (MS) detector (Amstelveen, The Netherlands) as described before. PCBs were determined using a 60 m x0.25 mm i.d. HT8 column with a film thickness of 0.25 µm. A subsample of 1 µl was injected using split/splitless mode with an injection temperature of 290 °C. Transfer line, source and quadrupole temperatures were 290, 230 and 150 °C, respectively. The oven program was as follows: 90°C, hold for 3 min, then 30 °C/min to 170, 1.5 °C/min to 270, 30 °C/min to 300 °C hold for 3 min. Ionization was performed using electron impact ionization (EI) mode. PCB 52 was quantified on m/z 292 using 290 as qualifier ion, PCBs 101, 110, and 118
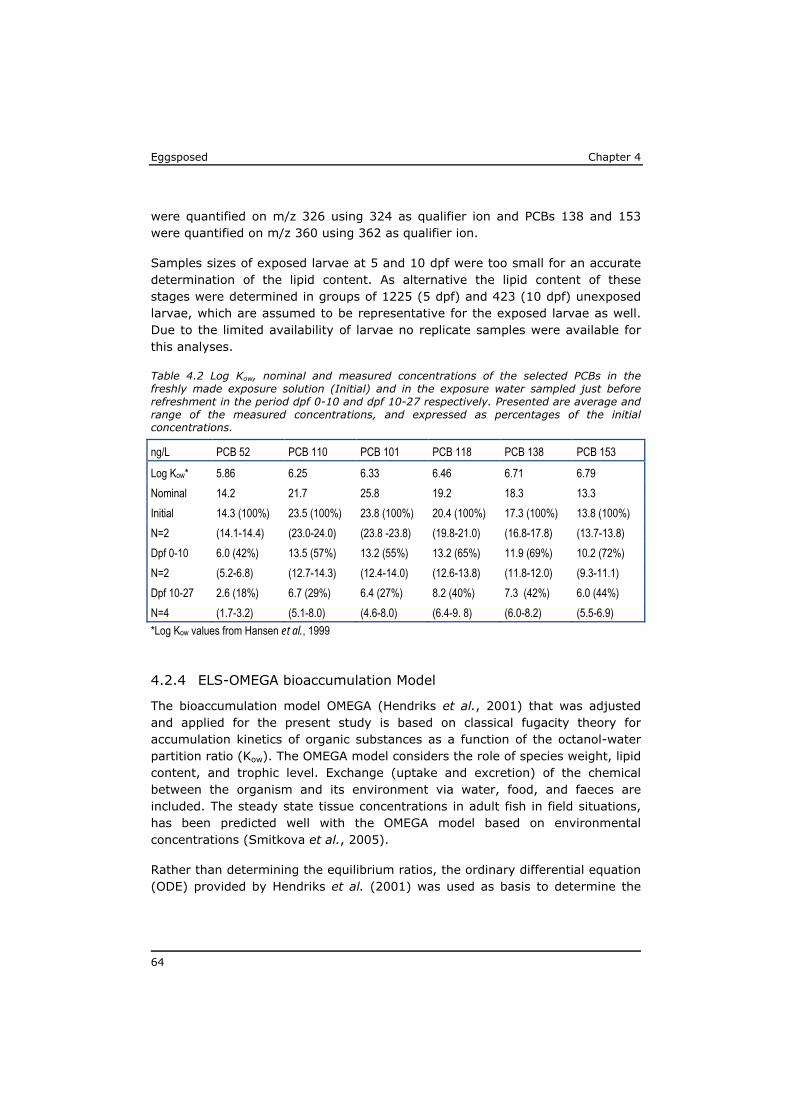
Eggsposed Chapter 4
64
were quantified on m/z 326 using 324 as qualifier ion and PCBs 138 and 153 were quantified on m/z 360 using 362 as qualifier ion.
Samples sizes of exposed larvae at 5 and 10 dpf were too small for an accurate determination of the lipid content. As alternative the lipid content of these stages were determined in groups of 1225 (5 dpf) and 423 (10 dpf) unexposed larvae, which are assumed to be representative for the exposed larvae as well. Due to the limited availability of larvae no replicate samples were available for this analyses.
Table 4.2 Log Kow, nominal and measured concentrations of the selected PCBs in the freshly made exposure solution (Initial) and in the exposure water sampled just before refreshment in the period dpf 0-10 and dpf 10-27 respectively. Presented are average and range of the measured concentrations, and expressed as percentages of the initial concentrations.
ng/L PCB 52 PCB 110 PCB 101 PCB 118 PCB 138 PCB 153
Log Kow* 5.86 6.25 6.33 6.46 6.71 6.79
Nominal 14.2 21.7 25.8 19.2 18.3 13.3
Initial 14.3 (100%) 23.5 (100%) 23.8 (100%) 20.4 (100%) 17.3 (100%) 13.8 (100%)
N=2 (14.1-14.4) (23.0-24.0) (23.8 -23.8) (19.8-21.0) (16.8-17.8) (13.7-13.8)
Dpf 0-10 6.0 (42%) 13.5 (57%) 13.2 (55%) 13.2 (65%) 11.9 (69%) 10.2 (72%)
N=2 (5.2-6.8) (12.7-14.3) (12.4-14.0) (12.6-13.8) (11.8-12.0) (9.3-11.1)
Dpf 10-27 2.6 (18%) 6.7 (29%) 6.4 (27%) 8.2 (40%) 7.3 (42%) 6.0 (44%)
N=4 (1.7-3.2) (5.1-8.0) (4.6-8.0) (6.4-9. 8) (6.0-8.2) (5.5-6.9)
*Log Kow values from Hansen et al., 1999
4.2.4 ELS-OMEGA bioaccumulation Model
The bioaccumulation model OMEGA (Hendriks et al., 2001) that was adjusted and applied for the present study is based on classical fugacity theory for accumulation kinetics of organic substances as a function of the octanol-water partition ratio (Kow). The OMEGA model considers the role of species weight, lipid content, and trophic level. Exchange (uptake and excretion) of the chemical between the organism and its environment via water, food, and faeces are included. The steady state tissue concentrations in adult fish in field situations, has been predicted well with the OMEGA model based on environmental concentrations (Smitkova et al., 2005).
Rather than determining the equilibrium ratios, the ordinary differential equation (ODE) provided by Hendriks et al. (2001) was used as basis to determine the

Chapter 4 Eggsposed
65
internal PCB concentrations in the egg/larvae at any given time. To this end, the ‘deSolve’ package (Soetaert et al., 2010) in R (The R Foundation for Statistical Computing) was used to solve the ODE numerically, using Euler’s method.
The OMEGA model optimized for fish early life stages will further be referred to as the ELS-OMEGA model.
For those model parameters that are assumed not to be life stage or species specific (respiration coefficient, rate exponent, body temperature correction factor, water layer diffusion resistance and lipid layer permeation resistance), the default and typical values that are presented for animals in (Hendriks et al., 2001) were applied (Table 4.4). For the water absorption-excretion coefficient (γ0) the default value was applied after hatching at 5 dpf. The value of the egg stage γ0 was calculated on the basis of the relative uptake of PCB 118 during the egg stage and the yolk-sac stage respectively, both covering a time frame of 5 d. PCB 118 was used for this calibration since its Kow was the closest to the average Kow of the PCBs tested. As γ0 is typical for a species/life stage, the thus calculated value is assumed valid for all PCBs.
Parameters describing the development of body weight and lipid content were derived from experimental data.
Mortality was not included since the ELS model describes the development of an individual at non-lethal exposure levels. Biotransformation of the persistent PCBs that were used in the present study will be close to zero.
Parameters related to feeding, like food ingestion coefficient (γ1), fraction of ingested food assimilated (P1), lipid fraction of food (Pch2, i-1) and pollutant concentration in food (Ci-1) are not applicable as long as fish are not free-feeding, and were thus set at zero until 10 dpf. After 10 dpf, γ1 and P1 were set at the default values (Hendriks et al., 2001). Lipid fraction of food (Pch2, i-1) and PCB concentration (Ci-1) were based on the results of the chemical analyses of the food source Artemia nauplii.
For the validation of the ELS-OMEGA model and the input parameters including γ0 based on PCB 118, the average exposure concentration of each PCB was predicted and compared to the measured range of concentrations in the exposure water. Since exposure conditions differed between dpf 0 to 10 and dpf 10 to 27, due to the application of smaller exposure beakers and the presence of food and faeces after dpf 10, the exposure concentrations for these two periods were calculated individually.
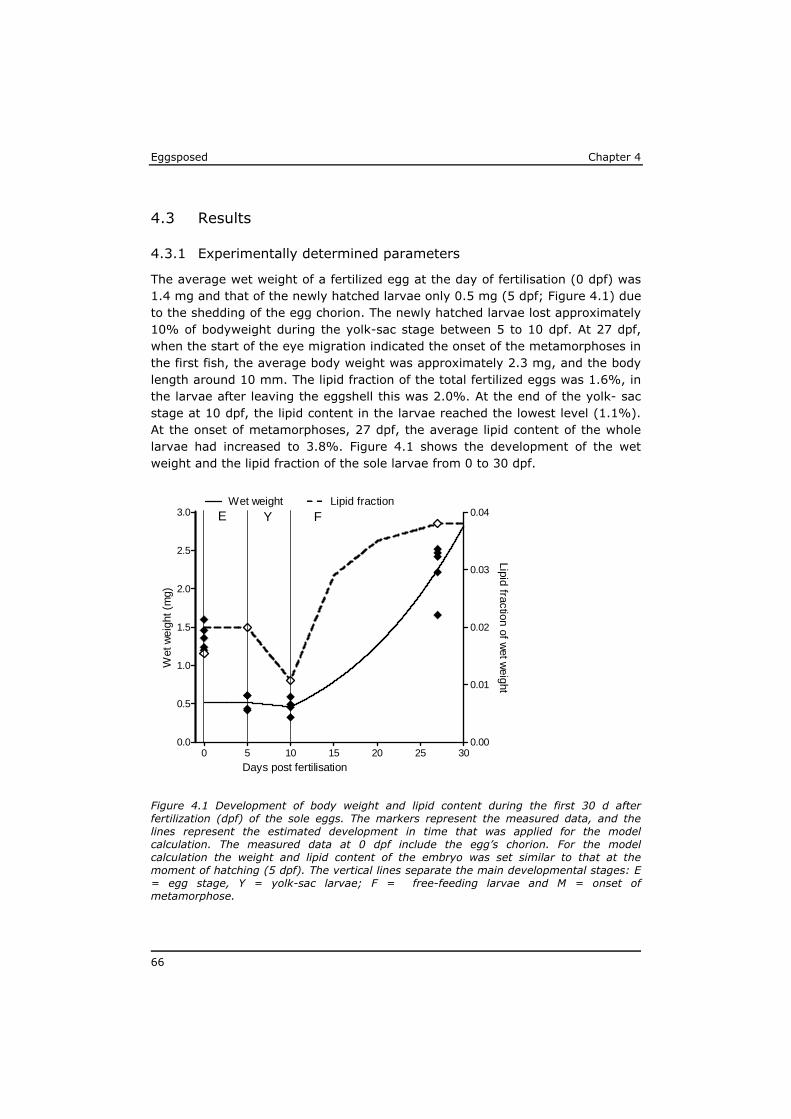
Eggsposed Chapter 4
66
4.3 Results
4.3.1 Experimentally determined parameters
The average wet weight of a fertilized egg at the day of fertilisation (0 dpf) was 1.4 mg and that of the newly hatched larvae only 0.5 mg (5 dpf; Figure 4.1) due to the shedding of the egg chorion. The newly hatched larvae lost approximately 10% of bodyweight during the yolk-sac stage between 5 to 10 dpf. At 27 dpf, when the start of the eye migration indicated the onset of the metamorphoses in the first fish, the average body weight was approximately 2.3 mg, and the body length around 10 mm. The lipid fraction of the total fertilized eggs was 1.6%, in the larvae after leaving the eggshell this was 2.0%. At the end of the yolk- sac stage at 10 dpf, the lipid content in the larvae reached the lowest level (1.1%). At the onset of metamorphoses, 27 dpf, the average lipid content of the whole larvae had increased to 3.8%. Figure 4.1 shows the development of the wet weight and the lipid fraction of the sole larvae from 0 to 30 dpf.
Figure 4.1 Development of body weight and lipid content during the first 30 d after fertilization (dpf) of the sole eggs. The markers represent the measured data, and the lines represent the estimated development in time that was applied for the model calculation. The measured data at 0 dpf include the egg’s chorion. For the model calculation the weight and lipid content of the embryo was set similar to that at the moment of hatching (5 dpf). The vertical lines separate the main developmental stages: E = egg stage, Y = yolk-sac larvae; F = free-feeding larvae and M = onset of metamorphose.
0 5 10 15 20 25 300.0
0.5
1.0
1.5
2.0
2.5
3.0
0.00
0.01
0.02
0.03
0.04
Days post fertilisation
Wet weight Lipid fractionE Y F
Wet
wei
ght (
mg)
Lipid fraction of wet w
eight
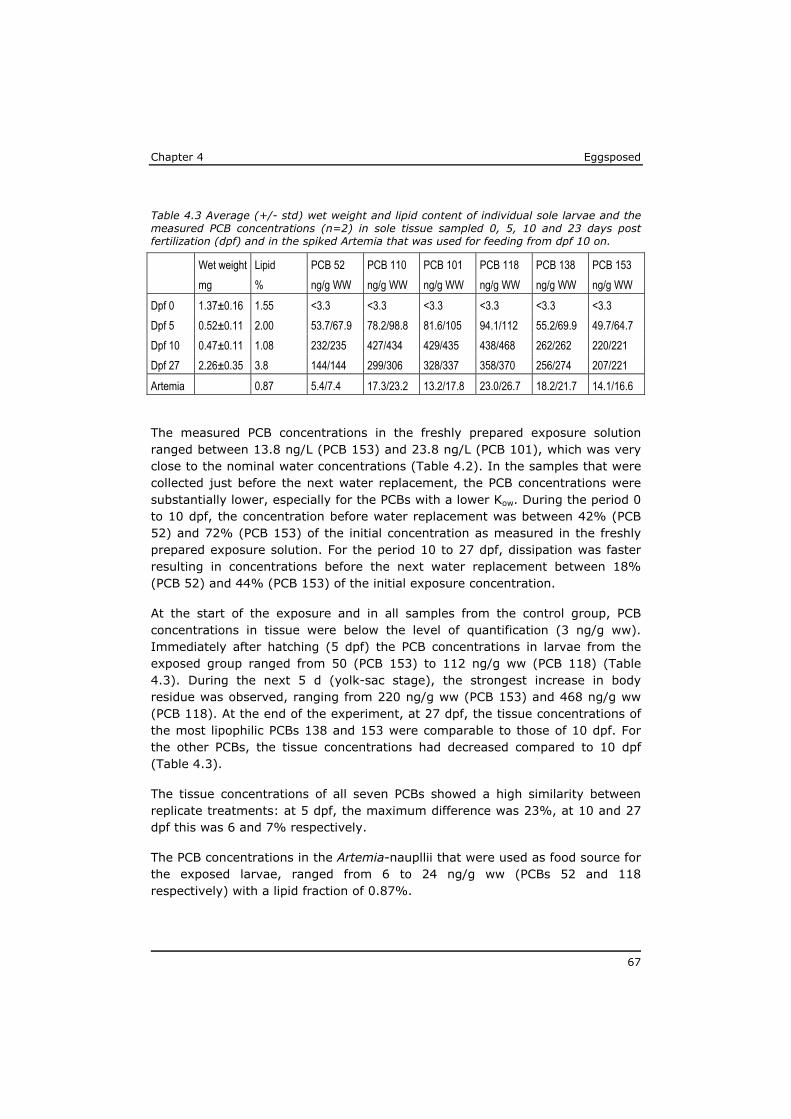
Chapter 4 Eggsposed
67
Table 4.3 Average (+/- std) wet weight and lipid content of individual sole larvae and the measured PCB concentrations (n=2) in sole tissue sampled 0, 5, 10 and 23 days post fertilization (dpf) and in the spiked Artemia that was used for feeding from dpf 10 on.
Wet weight Lipid PCB 52 PCB 110 PCB 101 PCB 118 PCB 138 PCB 153
mg % ng/g WW ng/g WW ng/g WW ng/g WW ng/g WW ng/g WW
Dpf 0 1.37±0.16 1.55 <3.3 <3.3 <3.3 <3.3 <3.3 <3.3
Dpf 5 0.52±0.11 2.00 53.7/67.9 78.2/98.8 81.6/105 94.1/112 55.2/69.9 49.7/64.7
Dpf 10 0.47±0.11 1.08 232/235 427/434 429/435 438/468 262/262 220/221
Dpf 27 2.26±0.35 3.8 144/144 299/306 328/337 358/370 256/274 207/221
Artemia 0.87 5.4/7.4 17.3/23.2 13.2/17.8 23.0/26.7 18.2/21.7 14.1/16.6
The measured PCB concentrations in the freshly prepared exposure solution ranged between 13.8 ng/L (PCB 153) and 23.8 ng/L (PCB 101), which was very close to the nominal water concentrations (Table 4.2). In the samples that were collected just before the next water replacement, the PCB concentrations were substantially lower, especially for the PCBs with a lower Kow. During the period 0 to 10 dpf, the concentration before water replacement was between 42% (PCB 52) and 72% (PCB 153) of the initial concentration as measured in the freshly prepared exposure solution. For the period 10 to 27 dpf, dissipation was faster resulting in concentrations before the next water replacement between 18% (PCB 52) and 44% (PCB 153) of the initial exposure concentration.
At the start of the exposure and in all samples from the control group, PCB concentrations in tissue were below the level of quantification (3 ng/g ww). Immediately after hatching (5 dpf) the PCB concentrations in larvae from the exposed group ranged from 50 (PCB 153) to 112 ng/g ww (PCB 118) (Table 4.3). During the next 5 d (yolk-sac stage), the strongest increase in body residue was observed, ranging from 220 ng/g ww (PCB 153) and 468 ng/g ww (PCB 118). At the end of the experiment, at 27 dpf, the tissue concentrations of the most lipophilic PCBs 138 and 153 were comparable to those of 10 dpf. For the other PCBs, the tissue concentrations had decreased compared to 10 dpf (Table 4.3).
The tissue concentrations of all seven PCBs showed a high similarity between replicate treatments: at 5 dpf, the maximum difference was 23%, at 10 and 27 dpf this was 6 and 7% respectively.
The PCB concentrations in the Artemia-naupllii that were used as food source for the exposed larvae, ranged from 6 to 24 ng/g ww (PCBs 52 and 118 respectively) with a lipid fraction of 0.87%.

Eggsposed Chapter 4
68
4.3.2 Input parameters for the model
As the model aims at estimating the PCB levels in the developing embryo, the weight and lipid fraction during the egg stage were set equal to the values of the freshly hatched larvae at 5 dpf. Thus, it is assumed that the observed differences in weight and lipid content between the egg at 0 dpf and the freshly hatched embryo at 5 dpf were due to the loss of the chorion alone and not the result of changes of the embryo. Bioamplification of the PCBs during the egg stage (Daley et al., 2009) was therefore not covered in our calculations. The weight loss during the yolk-sac stage (5 to10 dpf) is described by a negative biomass production coefficient (γ2) of -0.006 kgκ/d. The rapidly increasing body mass after 10 dpf corresponds to γ2 = 0.002985 kgκ/d.
The change in lipid fraction between 5 and 10 dpf when the yolk-sac is absorbed, is described as a constant decline of -0.18%/d. Between 10 and 27 dpf, the lipid content in the developing larvae builds up from 1.1% to 3.8%. This development is described by an exponential association with maximum value of 3.8%, assuming that the lipid content does not change after 27 dpf. For practical mathematical reasons, this development was described in the ELS-OMEGA model in four periods in which the lipid fraction increased with a constant factor: 10 to 15 dpf: +0.36%/d; 15 to 20 dpf: +0.12%/d; 20 to 27 dpf: +0.04%/d; and >27 dpf: 0%/d.
For the calculation of the egg stage γ0, the uptake of PCB 118 during egg-stage and yolk-sac stage, both covering a 5 day period, were compared. During egg stage the uptake was 27% and 31% of that during the yolk-sac stage, for the duplicated treatments respectively. With γ0 after hatching set at the default value of 200 kgκ/d (Hendriks et al., 2001), this corresponds to an egg stage γ0 of 34 and 40 kgκ/d respectively (see Figure S4.1 in Annex to Chapter 4). The difference in the model outcomes calculated with egg stage γ0 of 34 or 40 kgκ/d was less than 5%. Therefore, it was decided to use the average value of 37 kgκ/d as egg stage γ0 in the ELS-OMEGA model.
All input values (and their origin) that were used in the ELS-OMEGA model are presented in Table 4.4.
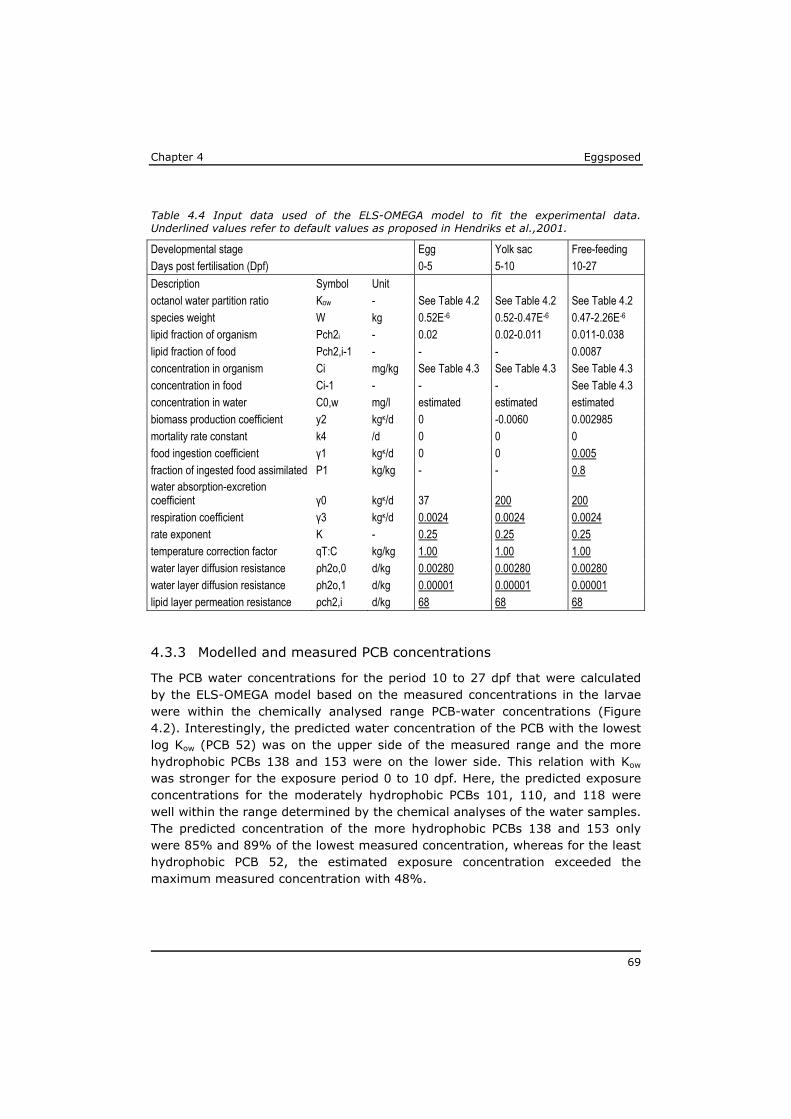
Chapter 4 Eggsposed
69
Table 4.4 Input data used of the ELS-OMEGA model to fit the experimental data. Underlined values refer to default values as proposed in Hendriks et al.,2001.
Developmental stage Egg Yolk sac Free-feeding Days post fertilisation (Dpf) 0-5 5-10 10-27
Description Symbol Unit octanol water partition ratio Kow - See Table 4.2 See Table 4.2 See Table 4.2 species weight W kg 0.52E-6 0.52-0.47E-6 0.47-2.26E-6 lipid fraction of organism Pch2i - 0.02 0.02-0.011 0.011-0.038 lipid fraction of food Pch2,i-1 - - - 0.0087 concentration in organism Ci mg/kg See Table 4.3 See Table 4.3 See Table 4.3 concentration in food Ci-1 - - - See Table 4.3 concentration in water C0,w mg/l estimated estimated estimated biomass production coefficient y2 kgκ/d 0 -0.0060 0.002985 mortality rate constant k4 /d 0 0 0 food ingestion coefficient γ1 kgκ/d 0 0 0.005 fraction of ingested food assimilated P1 kg/kg - - 0.8 water absorption-excretion coefficient γ0 kgκ/d 37 200 200 respiration coefficient γ3 kgκ/d 0.0024 0.0024 0.0024 rate exponent K - 0.25 0.25 0.25 temperature correction factor qT:C kg/kg 1.00 1.00 1.00 water layer diffusion resistance ρh2o,0 d/kg 0.00280 0.00280 0.00280 water layer diffusion resistance ρh2o,1 d/kg 0.00001 0.00001 0.00001 lipid layer permeation resistance ρch2,i d/kg 68 68 68
4.3.3 Modelled and measured PCB concentrations
The PCB water concentrations for the period 10 to 27 dpf that were calculated by the ELS-OMEGA model based on the measured concentrations in the larvae were within the chemically analysed range PCB-water concentrations (Figure 4.2). Interestingly, the predicted water concentration of the PCB with the lowest log Kow (PCB 52) was on the upper side of the measured range and the more hydrophobic PCBs 138 and 153 were on the lower side. This relation with Kow was stronger for the exposure period 0 to 10 dpf. Here, the predicted exposure concentrations for the moderately hydrophobic PCBs 101, 110, and 118 were well within the range determined by the chemical analyses of the water samples. The predicted concentration of the more hydrophobic PCBs 138 and 153 only were 85% and 89% of the lowest measured concentration, whereas for the least hydrophobic PCB 52, the estimated exposure concentration exceeded the maximum measured concentration with 48%.
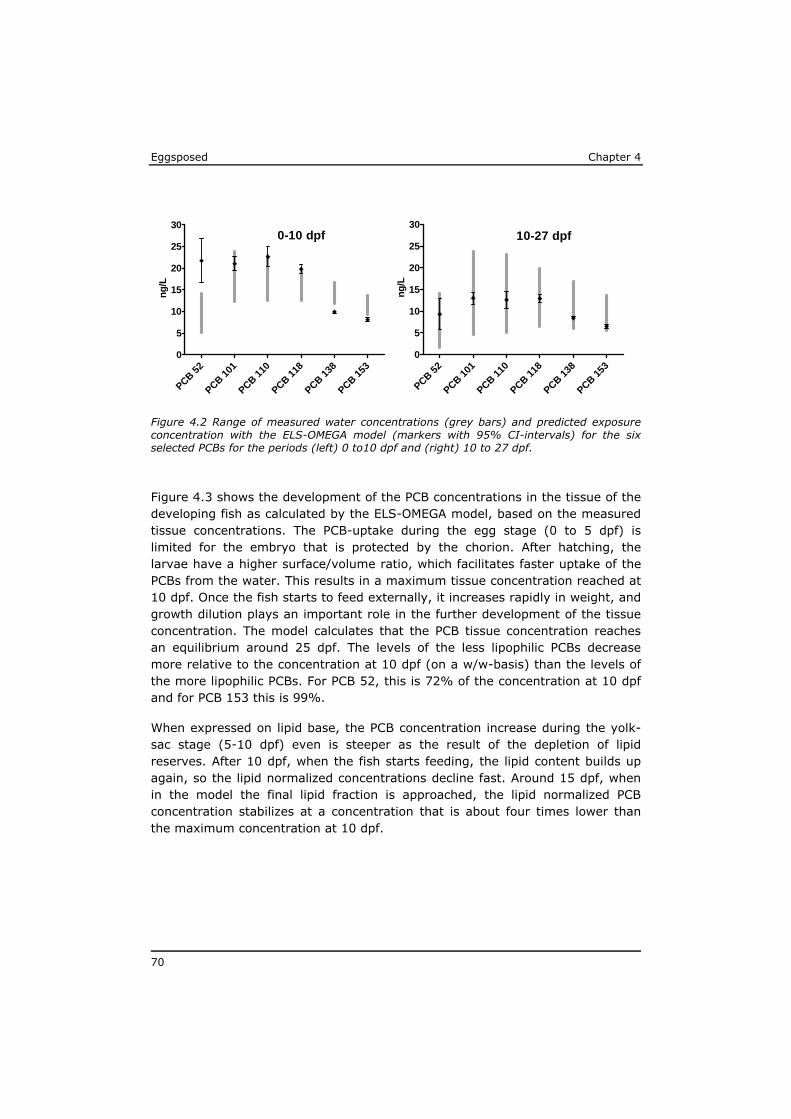
Eggsposed Chapter 4
70
Figure 4.2 Range of measured water concentrations (grey bars) and predicted exposure concentration with the ELS-OMEGA model (markers with 95% CI-intervals) for the six selected PCBs for the periods (left) 0 to10 dpf and (right) 10 to 27 dpf.
Figure 4.3 shows the development of the PCB concentrations in the tissue of the developing fish as calculated by the ELS-OMEGA model, based on the measured tissue concentrations. The PCB-uptake during the egg stage (0 to 5 dpf) is limited for the embryo that is protected by the chorion. After hatching, the larvae have a higher surface/volume ratio, which facilitates faster uptake of the PCBs from the water. This results in a maximum tissue concentration reached at 10 dpf. Once the fish starts to feed externally, it increases rapidly in weight, and growth dilution plays an important role in the further development of the tissue concentration. The model calculates that the PCB tissue concentration reaches an equilibrium around 25 dpf. The levels of the less lipophilic PCBs decrease more relative to the concentration at 10 dpf (on a w/w-basis) than the levels of the more lipophilic PCBs. For PCB 52, this is 72% of the concentration at 10 dpf and for PCB 153 this is 99%.
When expressed on lipid base, the PCB concentration increase during the yolk-sac stage (5-10 dpf) even is steeper as the result of the depletion of lipid reserves. After 10 dpf, when the fish starts feeding, the lipid content builds up again, so the lipid normalized concentrations decline fast. Around 15 dpf, when in the model the final lipid fraction is approached, the lipid normalized PCB concentration stabilizes at a concentration that is about four times lower than the maximum concentration at 10 dpf.
PCB 52
PCB 101
PCB 110
PCB 118
PCB 138
PCB 153
0
5
10
15
20
25
300-10 dpf
ng
/L
PCB 52
PCB 101
PCB 110
PCB 118
PCB 138
PCB 153
0
5
10
15
20
25
3010-27 dpf
ng
/L
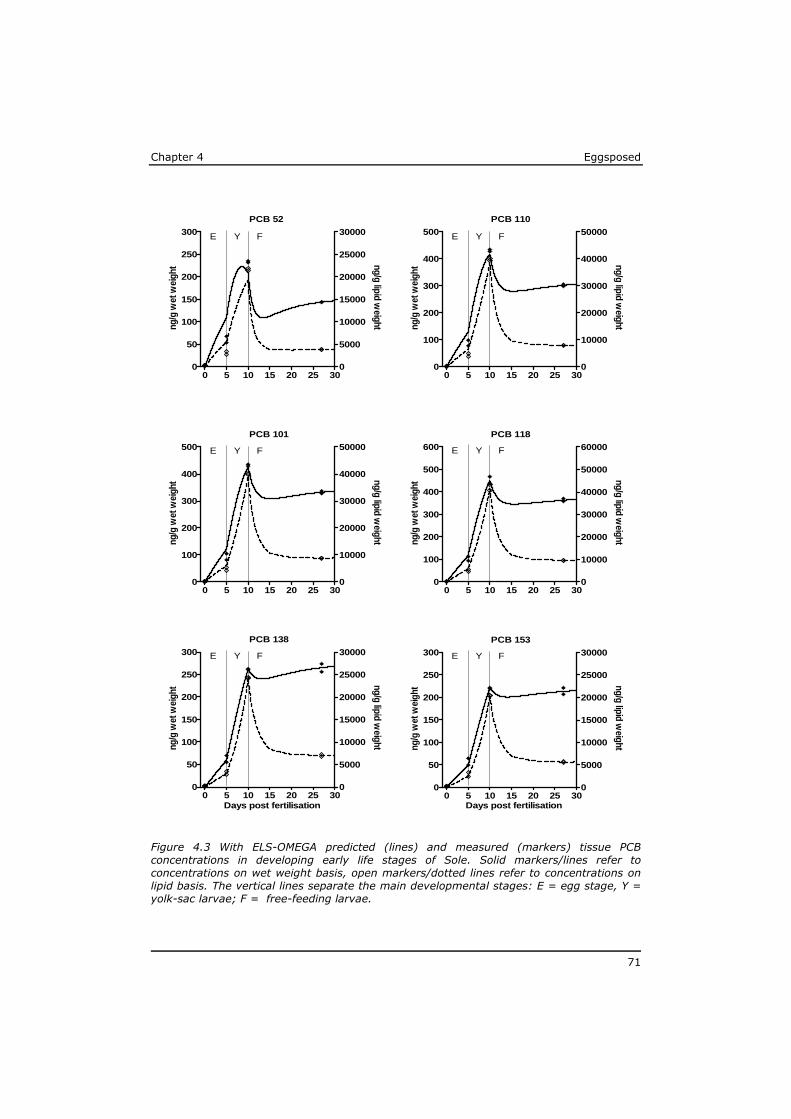
Chapter 4 Eggsposed
71
Figure 4.3 With ELS-OMEGA predicted (lines) and measured (markers) tissue PCB concentrations in developing early life stages of Sole. Solid markers/lines refer to concentrations on wet weight basis, open markers/dotted lines refer to concentrations on lipid basis. The vertical lines separate the main developmental stages: E = egg stage, Y = yolk-sac larvae; F = free-feeding larvae.
0 5 10 15 20 25 300
50
100
150
200
250
300
0
5000
10000
15000
20000
25000
30000
PCB 52
E Y F
ng/g
wet
wei
ght ng/g lipid w
eight
0 5 10 15 20 25 300
100
200
300
400
500
0
10000
20000
30000
40000
50000
PCB 110
E Y F
ng/g
wet
wei
ght ng/g lipid w
eight
0 5 10 15 20 25 300
100
200
300
400
500
0
10000
20000
30000
40000
50000
PCB 101
E Y F
ng/g
wet
wei
ght ng/g lipid w
eight
0 5 10 15 20 25 300
100
200
300
400
500
600
0
10000
20000
30000
40000
50000
60000
PCB 118
E Y F
ng/g
wet
wei
ght ng/g lipid w
eight
0 5 10 15 20 25 300
50
100
150
200
250
300
0
5000
10000
15000
20000
25000
30000
Days post fertilisation
PCB 138
E Y F
ng/g
wet
wei
ght ng/g lipid w
eight
0 5 10 15 20 25 300
50
100
150
200
250
300
0
5000
10000
15000
20000
25000
30000
Days post fertilisation
PCB 153
E Y F
ng/g
wet
wei
ght ng/g lipid w
eight

Eggsposed Chapter 4
72
4.4 Discussion
The OMEGA model for bioaccumulation of POPs in adult fish, was refined to provide an ELS-OMEGA model for calculation of the POP tissue concentrations during the development of fish eggs, via yolk-sac larvae into a free-feeding juvenile. Application of this model and validation with PCB-exposed sole eggs and larvae revealed that the internal levels rise to reach a peak at the end of the yolk-sac stage. Below the developed ELS-OMEGA model is discussed and subsequently a number of different realistic exposure scenarios are modelled and discussed.
4.4.1 Model validation
Validation of the ELS-OMEGA model was performed by comparing the external exposure concentrations that were calculated based on measured tissue concentrations with measured external exposure concentrations. This sequence is opposite to the more standard approach in bioaccumulation modelling where water- and/or food concentrations are used to try to predict the tissue concentrations. We choose this approach being aware of the fact that it is impossible to maintain a stable exposure concentration of PCBs in semi-static tests (Breitholtz et al., 2006; Rufli et al., 1998) due to exponential dissipation of the PCBs from the water column with time (Koponen et al., 1998; Bteitholtz and Wollenberger, 2003).
The measured tissue concentrations were very similar between the independent replicates (Table 4.3) and can thus be considered to give a more reliable representation than the measured water concentrations that indicated strong dissipation between refreshments. Most of the predicted water concentrations fell within, or were close to the measured ranges of the water concentrations (Figure 4.6). An exception was the less lipophilic PCB 52 water concentration during the egg and yolk- sac stage (dpf 0 – 10) that was predicted twice as high as what was measured. This could indicate that the uptake of this less lipophilic PCB during these early stages is more efficient than assumed by the ELS-OMEGA model especially during the yolk-sac stage, since the tissue concentration at the moment of hatching seems overestimated.
4.4.2 Model parameter values and sensitivity
In the ELS-OMEGA model, the parameter values applied for respiration coefficient, rate exponent, water layer diffusion resistance, and lipid layer permeation resistance were for all developmental stages kept constant at the default values proposed by Hendriks (Hendriks et al., 2001). Although one could

Chapter 4 Eggsposed
73
argue if some of these values might change during the early life development, the impact of these parameters on the model outcome was limited (no more than 5% difference) as long as they were maintained within the ranges given by Hendriks et al. (2001).
Based on the experimental data for PCB 118, the water exchange rate during the egg stage, expressed as the water absorption-excretion coefficient γ0 in the model, was calculated to be 17-20% (34-40 kgκ/d) of the default value that was applied after hatching (200 kgκ/d) (Hendriks et al., 2001) . This relative difference between egg and larval stage is in accordance with observations with zebra fish early life stages, where uptake rate constants for PAHs with log Kow 4.5 to 6.1 during the egg stage were 16 to 20% of those after hatching (Lu et al., 2011). The impact, however, of the egg stage γ0 on the outcome of the ELS-OMEGA model for sole is limited, as it only affects a short, 5 d period.
The model outcome is mostly depending on (changes in) the lipid content of the organisms. The lipid content we found for the freshly hatched larvae (2% of wet weight) is in accordance with the 1.9% (20.3% of dry weight) reported for the closely related species Solea senegalesis (Mourente et al.,1996). In the same study the total lipid fraction during the yolk- sac stage was reduced with 33%, while in our study this was 45%. Our juvenile sole consisted of 3.8% lipid at 27 dpf. As this value is on the high side of the 1% to 3.5% that is presented in literature for adult sole muscle tissue (Baeyens et al., 2003 and Bordajandi et al., 2006 respectively), we assume in the model that our 3.8 % is representative for a fully developed sole. This is somewhat higher than the lipid content of three total fish samples composed of juvenile sole from the Western Scheldt estuary (2.2, 2.9 and 3.3%, average 2.8%; unpublished data IMARES, Netherlands, 2005, 2006), which could be related to the life in captivity without food limitation.
The exposure scenarios presented below, were also performed with alternative lipid dynamics (egg 1.9%; end yolk-sac 1.3%; juvenile 2.8%; Figure 4.4) based on the data presented above. This does not fundamentally change the outcome, but shows how the relative height of the exposure peak and the relative depletion of the lipid are related. Results are presented as Figures S4.2 and S4.3 in Annex to Chapter 4.
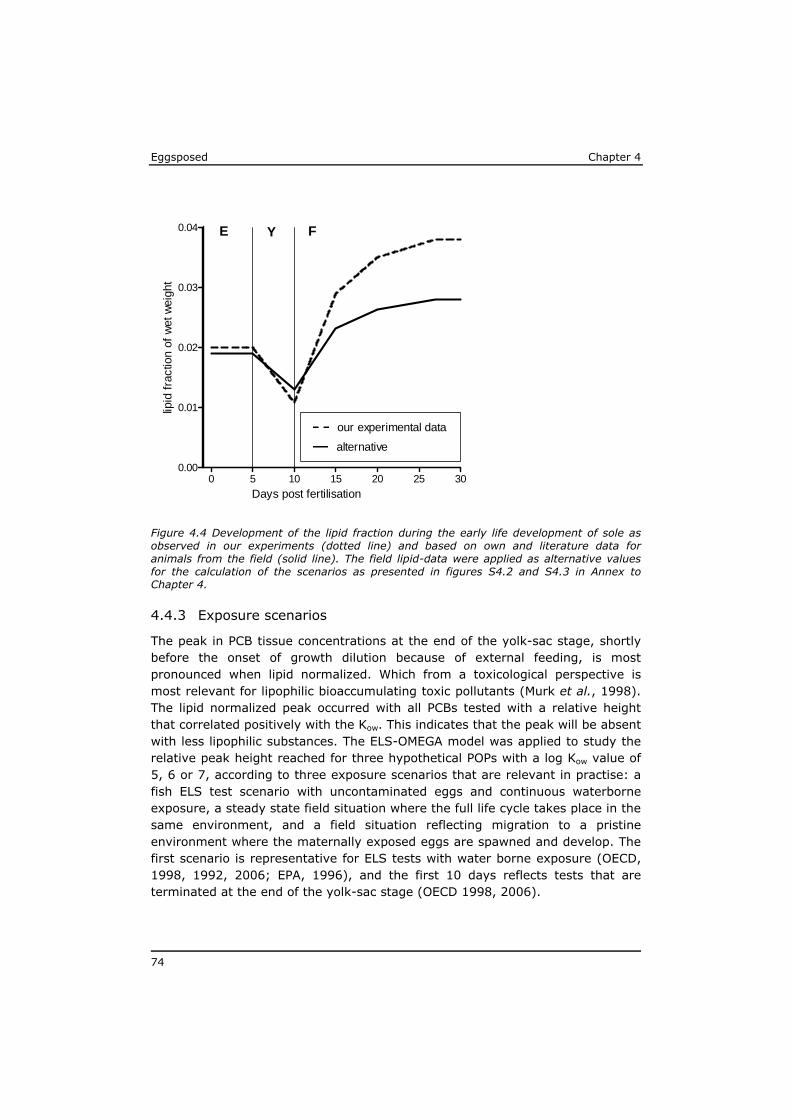
Eggsposed Chapter 4
74
Figure 4.4 Development of the lipid fraction during the early life development of sole as observed in our experiments (dotted line) and based on own and literature data for animals from the field (solid line). The field lipid-data were applied as alternative values for the calculation of the scenarios as presented in figures S4.2 and S4.3 in Annex to Chapter 4.
4.4.3 Exposure scenarios
The peak in PCB tissue concentrations at the end of the yolk-sac stage, shortly before the onset of growth dilution because of external feeding, is most pronounced when lipid normalized. Which from a toxicological perspective is most relevant for lipophilic bioaccumulating toxic pollutants (Murk et al., 1998). The lipid normalized peak occurred with all PCBs tested with a relative height that correlated positively with the Kow. This indicates that the peak will be absent with less lipophilic substances. The ELS-OMEGA model was applied to study the relative peak height reached for three hypothetical POPs with a log Kow value of 5, 6 or 7, according to three exposure scenarios that are relevant in practise: a fish ELS test scenario with uncontaminated eggs and continuous waterborne exposure, a steady state field situation where the full life cycle takes place in the same environment, and a field situation reflecting migration to a pristine environment where the maternally exposed eggs are spawned and develop. The first scenario is representative for ELS tests with water borne exposure (OECD, 1998, 1992, 2006; EPA, 1996), and the first 10 days reflects tests that are terminated at the end of the yolk-sac stage (OECD 1998, 2006).
0 5 10 15 20 25 300.00
0.01
0.02
0.03
0.04
Days post fertilisation
our experimental data
E Y F
lipid
fra
ctio
n of
wet
wei
ght
alternative
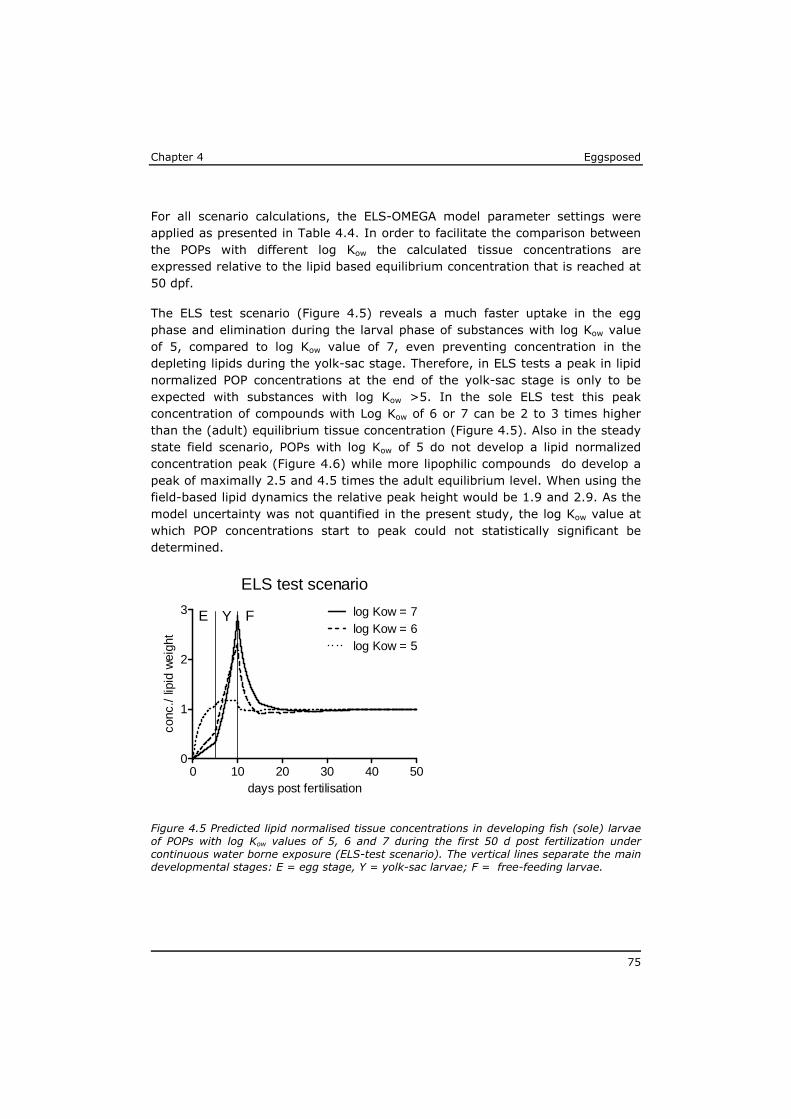
Chapter 4 Eggsposed
75
For all scenario calculations, the ELS-OMEGA model parameter settings were applied as presented in Table 4.4. In order to facilitate the comparison between the POPs with different log Kow the calculated tissue concentrations are expressed relative to the lipid based equilibrium concentration that is reached at 50 dpf.
The ELS test scenario (Figure 4.5) reveals a much faster uptake in the egg phase and elimination during the larval phase of substances with log Kow value of 5, compared to log Kow value of 7, even preventing concentration in the depleting lipids during the yolk-sac stage. Therefore, in ELS tests a peak in lipid normalized POP concentrations at the end of the yolk-sac stage is only to be expected with substances with log Kow >5. In the sole ELS test this peak concentration of compounds with Log Kow of 6 or 7 can be 2 to 3 times higher than the (adult) equilibrium tissue concentration (Figure 4.5). Also in the steady state field scenario, POPs with log Kow of 5 do not develop a lipid normalized concentration peak (Figure 4.6) while more lipophilic compounds do develop a peak of maximally 2.5 and 4.5 times the adult equilibrium level. When using the field-based lipid dynamics the relative peak height would be 1.9 and 2.9. As the model uncertainty was not quantified in the present study, the log Kow value at which POP concentrations start to peak could not statistically significant be determined.
Figure 4.5 Predicted lipid normalised tissue concentrations in developing fish (sole) larvae of POPs with log Kow values of 5, 6 and 7 during the first 50 d post fertilization under continuous water borne exposure (ELS-test scenario). The vertical lines separate the main developmental stages: E = egg stage, Y = yolk-sac larvae; F = free-feeding larvae.
ELS test scenario
0 10 20 30 40 500
1
2
3
days post fertilisation
conc
./ li
pid
wei
ght
E Y F log Kow = 7
log Kow = 5log Kow = 6
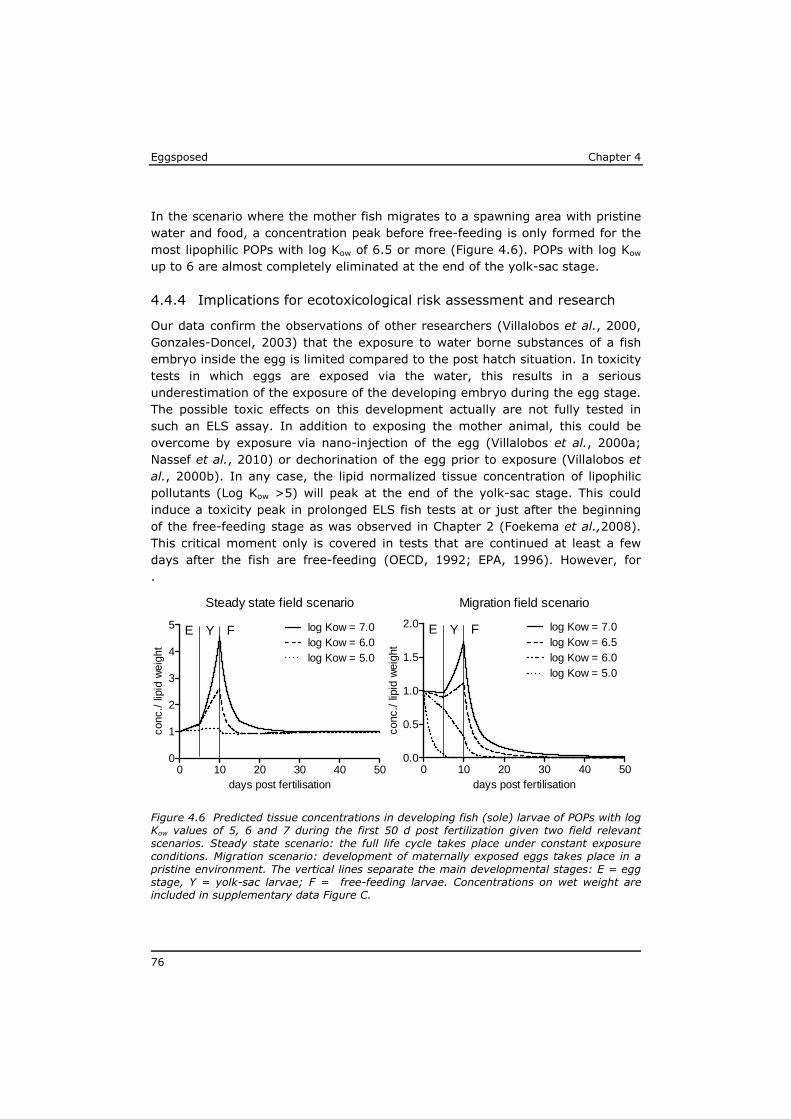
Eggsposed Chapter 4
76
In the scenario where the mother fish migrates to a spawning area with pristine water and food, a concentration peak before free-feeding is only formed for the most lipophilic POPs with log Kow of 6.5 or more (Figure 4.6). POPs with log Kow up to 6 are almost completely eliminated at the end of the yolk-sac stage.
4.4.4 Implications for ecotoxicological risk assessment and research
Our data confirm the observations of other researchers (Villalobos et al., 2000, Gonzales-Doncel, 2003) that the exposure to water borne substances of a fish embryo inside the egg is limited compared to the post hatch situation. In toxicity tests in which eggs are exposed via the water, this results in a serious underestimation of the exposure of the developing embryo during the egg stage. The possible toxic effects on this development actually are not fully tested in such an ELS assay. In addition to exposing the mother animal, this could be overcome by exposure via nano-injection of the egg (Villalobos et al., 2000a; Nassef et al., 2010) or dechorination of the egg prior to exposure (Villalobos et al., 2000b). In any case, the lipid normalized tissue concentration of lipophilic pollutants (Log Kow >5) will peak at the end of the yolk-sac stage. This could induce a toxicity peak in prolonged ELS fish tests at or just after the beginning of the free-feeding stage as was observed in Chapter 2 (Foekema et al.,2008). This critical moment only is covered in tests that are continued at least a few days after the fish are free-feeding (OECD, 1992; EPA, 1996). However, for .
Figure 4.6 Predicted tissue concentrations in developing fish (sole) larvae of POPs with log Kow values of 5, 6 and 7 during the first 50 d post fertilization given two field relevant scenarios. Steady state scenario: the full life cycle takes place under constant exposure conditions. Migration scenario: development of maternally exposed eggs takes place in a pristine environment. The vertical lines separate the main developmental stages: E = egg stage, Y = yolk-sac larvae; F = free-feeding larvae. Concentrations on wet weight are included in supplementary data Figure C.
Steady state field scenario
0 10 20 30 40 500
1
2
3
4
5
days post fertilisation
conc
./ li
pid
wei
ght
E Y F log Kow = 7.0log Kow = 6.0log Kow = 5.0
Migration field scenario
0 10 20 30 40 500.0
0.5
1.0
1.5
2.0
days post fertilisation
conc
./ li
pid
wei
ght
E Y F log Kow = 7.0
log Kow = 6.0log Kow = 5.0
log Kow = 6.5

Chapter 4 Eggsposed
77
practical and ethical/legal reasons, the majority of ELS fish tests are terminated (just) before the end of the yolk-sac stage (OECD 1998, 2006). Our data indicate that this is valid for testing substances with log Kow <5. But when more hydrophobic substances or (mixtures of) unknown substances as present in effluents or sediments (Hollert et al., 2003; Kosmehl et al., 2006), are being tested, the effects of lipophilic substances will be seriously underestimated.
Our scenarios reveal that maternally transferred high lipophilic POPs (log Kow >6.5) are hardly excreted, and reach peak concentrations at the end of the yolk-sac stage, the critical moment when the larvae has to switch to external feeding. Current levels of dioxin-like and other teratogenic pollutants may therefore cause serious adverse effects on developing fish at realistic maternal concentrations (Foekema et al., 2008). The amount of maternally transferred POPs to the eggs can be estimated based on the lipid normalized maternal concentrations (Russel et al., 1999). By applying the ELS-OMEGA model the peak exposure concentration for the larvae can be calculated for species specific lipid dynamics and local (exposure) circumstances. Our data indicate that the lipid normalized peak concentration of substances with log Kow >5 at the end of the yolk-sac stage may exceed 2 to 4 times the concentration in the spawning parent fish. The latter concentration could also be greatly enhanced in species with reproductive migration without feeding like salmon and eel. The greatly increased (lipid normalized) POP concentrations in the maternal tissue and eggs (Debruyn et al., 2004; Van Ginneken et al.,2009; Kelly et al., 2011) in combination with the further concentration in the developing larvae as shown in the present study, could result in extreme body burdens during critical stages of larval development with serious consequences for larval survival, even when eggs and larvae can develop in pristine areas.
4.5 References
Bodiguel, X., O. Maury, C. Mellon-Duval, F. Roupsard, A.M. Le Guellec, V. Loizeau. 2009. A dynamic and mechanistic model of PCB bioaccumulation in the European hake (Merluccius merluccius). Journal of Sea Research, 62(2-3): p. 124-134.
Baeyens, W., Leermakers, M., Papina, T., Saprykin, A., Brion, N., Noyen, J., De Gieter, M., Elskens, M., Goeyens, L. 2003. Bioconcentration and biomagnification of mercury and methylmercury in North Sea and Scheldt estuary fish. Archives of Environmental Contamination and Toxicology, 45, (4), 498-508.
Berg, V., J.L. Lyche, C. Karlsson, B. Stavik, R. Nourizadeh-Lillabadi, N. Hardnes, J. U. Skaare, P. Alestrøm, E. Lie, E. Ropstad. 2011. Accumulation and Effects of Natural Mixtures of Persistent Organic Pollutants (POP) in Zebrafish after Two Generations of Exposure. Journal of Toxicology and Environmental Health-Part a-Current Issues, 74(7-9): p. 407-423.

Eggsposed Chapter 4
78
Blaxter, J.H.S. 1988. Pattern and Variety in Development, in: W.S Hoar & D.J Randall (eds.) Fish Physiology , Academic press, inc. California USA. p. 1-58.
Bordajandi, L. R., Martin, I., Abad, E., Rivera, J., Gonzalez, M. J. 2006. Organochlorine compounds (PCBs, PCDDs and PCDFs) in seafish and seafood from the Spanish Atlantic southwest coast. Chemosphere, 64, (9), 1450-1457.
Breitholtz, M., L. Wollenberger. 2003. Effects of three PBDEs on development, reproduction and population growth rate of the harpacticoid copepod Nitocra spinipes. Aquatic Toxicology, 64(1): p. 85-96.
Breitholtz, M., C. Rudén, S. O. Hansson, B. E. Bengtsson. 2006. Ten challenges for improved ecotoxicological testing in environmental risk assessment. Ecotoxicology and Environmental Safety, 63(2): p. 324-335.
Crane H.M. D.B.Pickford, T.H.Hutchinson, J.A. Brown. 2006. The effect of methimazole on development of the fathead minnow Pimephales promelas, from embryo to adult. Toxicol. Sci. 93, 278-285.
Daley, J.M., T.A. Leadley, and K.G. Drouillard. 2009. Evidence for bioamplification of nine polychlorinated biphenyl (PCB) congeners in yellow perch (Perca flavascens) eggs during incubation. Chemosphere, 75(11): p. 1500-1505.
Debruyn, A.M.H., M.G. Ikonomou, and F. Gobas. 2004. Magnification and toxicity of PCBs, PCDDs, and PCDFs in upriver-migrating Pacific salmon. Environmental Science & Technology, 38(23): p. 6217-6224.
Eichinger, M., V. Loizeau, F. Roupsard, A.M. Le Guellec, C. Bacher. 2010. Modelling growth and bioaccumulation of Polychlorinated biphenyls in common sole (Solea solea). Journal of Sea Research, 64(3): p. 373-385.
EPA. 1996. Ecological Effects Test Guidelines OPPTS 850.1400 Fish Early-Life Stage Toxicity Test. Environmental Protection Agency. Washington, USA.
Foekema, E.M., C.M. Deerenberg, A.J. Murk. 2008. Prolonged ELS test with the marine flatfish sole (Solea solea) shows delayed toxic effects of previous exposure to PCB 126. Aquatic Toxicology, 90(3): p. 197-203.
González-Doncel, M., M. Larrea, S. Sánchez-Fortún, D.E. Hinton. 2003. Influence of water hardening of the chorion on cadmium accumulation in medaka (Oryzias latipes) eggs. Chemosphere, 52(1): p. 75-83.
Hansen, B. G., Paya-Perez, A. B., Rahman, M., Larsen, B. R. 1999. Qsars for K-ow and K-oc of PCB congeners: A critical examination of data, assumptions and statistical approaches. Chemosphere, 39, (13), 2209-2228..
Hendriks, A.J., A. van der Linde, G. Cornelissen, D.T.H.M. Sijm. 2001. The power of size. 1. Rate constants and equilibrium ratios for accumulation of organic substances related to octanol-water partition ratio and species weight. Environmental Toxicology and Chemistry, 20(7): p. 1399-1420.
Hollert, H., S. Keiter, n. Konig, M. Rudolf, M. Ulrich, T. Braunbeck. 2003. A new sediment contact assay to assess particle-bound pollutants using zebrafish (Danio rerio) embryos. Journal of Soils and Sediments, 3(3): p. 197-207.
Hutchinson, T.H., J. Solbe, and P.J. Kloepper-Sams. 1988. Analysis of the ECETOC aquatic toxicity (EAT) database - III - Comparative toxicity of chemical substances to different life stages of aquatic organisms. Chemosphere, 36(1): p. 129-142.

Chapter 4 Eggsposed
79
Jobling, M. 2002. Environmental factors and rates of development and growth, in: Handbook of fish biology and fisheries, J.B.H.J.D. Reynolds (ed.), Blackwell Publishing.
Kelly, B.C., M.G. Ikonomou, N. MacPherson, T. Sampson, D.A. Patterson, and C. Dubetz. 2011. Tissue residue concentrations of organohalogens and trace elements in adult Pacific salmon returning to the Fraser River, British Columbia, Canada. Environmental Toxicology and Chemistry 30(2): 367-376.
Koponen, K., J.V.K. Kukkonen, P. Lindstrom-Seppa. 1998. Chemical accumulation and biotransformation enzyme activities of rainbow trout embryos in waterborne exposure to PCB-77. Marine Environmental Research, 46(1-5): p. 475-478.
Kosmehl, T., Reifferscheid, G., Manz, W., Braunbeck, T., Hollert, H. 2006. A novel contact assay to assess genotoxic burden of sediments using the comet assay with zebrafish larvae. Environ. Toxicol. Chem. 25: 2097-2106.
Lu, X., A. Skwarski, B. Drake, D. D. Reible. 2011. Predicting bioavailability of PAHs and PCBs with porewater concentrations measured by solid-phase microextraction fibers. Environmental Toxicology and Chemistry, 30(5): p. 1109-1116.
Mourente, G., R. Vazques. 1996. Changes in the content of total lipids, lipid classes and their fatty acids of developing eggs and unfed larvae of the Senegal Sole, Solea senegalensis Kaup. Fish Physiol. Biochem. 15:221-235.
Murk, A.J., Leonards, P.E.G., van Hattum, B., Luit, R., van der Weiden, M.E.J., Smit, M. 1998. Application of biomarkers for exposure and effect of polyhalogenated aromatic hydrocarbons in naturally exposed European otters (Lutra lutra). Environmental toxicology and pharmacology 6, 91-102.
Nassef, M., S.G. Kim, M. Seki, I. J. Kang, T. Hano, Y. Shimasaki, Y. Oshima. 2010. In-ovo nanoinjection of triclosan, diclofenac and carbamazepine affects embryonic development of medaka fish (Oryzias latipes). Chemosphere, 79(9): p. 966-973.
Ng, C.A. and K.A. Gray. 2009. Tracking bioaccumulation in aquatic organisms: A dynamic model integrating life history characteristics and environmental change. Ecological Modelling, 220(9-10): p. 1266-1273.
OECD. 1992. OECD Guideline for testing of chemicals 210: Fish, Early-Life Stage Toxicology test. Adopted 17 July 1992. Organization for Economic Co-operation and Development, Paris.
OECD. 1998. OECD Guideline for testing of chemicals 212: Fish, short term Toxicology test on Embryo and sac-fry Stages. Adopted 21st September 1998. Organization for Economic Co-operation and Development, Paris.
OECD. 2006. Fish Embryo Toxicity (FET) test. Draft proposal for a new guideline May 30, 2006 (1st Version). Organization for Economic Co-operation and Development, Paris.
Paterson, G., J. Lieu, G.D. Haffner, K.G. Drouillard. 2010. Contribution of fecal egestion to the whole body elimination of polychlorinated biphenyls by Japanese koi (Cyprinus carpio). Environ. Sci. Technol. 44:5769-5774.
Petersen, G.I., P. Kristensen. 1998. Bioaccumulation of lipophilic substances in fish early life stages. Environmental Toxicology and Chemistry, 17(7): p. 1385-1395.
Rufli, H., P. R. Fisk, A. E. Girling, J. M. King, R. Länge, X. Lejeune, N. Stelter, C. Stevens, P. Suteau, J. Tapp, J. Thus, D. J. Versteeg, H. J. Niessen. 1998. Aquatic toxicity

Eggsposed Chapter 4
80
testing of sparingly soluble, volatile, and unstable substances and interpretation and use of data. Ecotoxicology and Environmental Safety, 39(2): p. 72-77.
Russell, R.W., F. Gobas, G.D. Haffner. 1999. Maternal transfer and in ovo exposure of organochlorines in oviparous organisms: A model and field verification. Environmental Science & Technology, 33(3): p. 416-420.
Serrano, R., M.A. Blanes, and F.J. Lopez. 2008. Maternal transfer of organochlorine compounds to oocytes in wild and farmed gilthead sea bream (Sparus aurata). Chemosphere, 70(4): p. 561-566.
Sijm, D., W. Seinen, A. Opperhulzen. 1992. Life-cycle biomagnification study in fish. Environmental Science & Technology, 1992. 26(11): p. 2162-2174.
Smitkova, H., M.A.J. Huijbregts, and A.J. Hendriks. 2005. Comparison of three fish bioaccumulation models for ecological and human risk assessment and validation with field data. Sar and Qsar in Environmental Research, 16(5): p. 483-493.
Soetaert, K., T. Petzoldt, R.W. Setzer. 2010. Solving Differential Equations in R: Package deSolve. Journal of Statistical Software, 33(9): p. 1-25.
Van Ginneken, V., A. Palstra, P. Leonards, M. Nieveen, H. Van Den Berg, G. Flik, T. Spanings, P. Niemantsverdriet, G. Van Den Thillart, A.J. Murk. 2009. PCBs and the energy cost of migration in the European eel (Anguilla anguilla L.). Aquatic Toxicology,. 92(4): p. 213-220.
Veltman, K., A.J. Hendriks, M.A.J. Huijbregts, M.J. van den Heuvel-Greve, A.D. Vethaak. 2005. Accumulation of organochlorines and brominated flame retardants in estuarine and marine food chains: Field measurements and model calculations. Marine Pollution Bulletin, 50(10): p. 1085-1102.
Villalobos, S. A., Hamm, J. T., Teh, S. J., Hinton, D. E. 2000a. Thiobencarb-induced embryotoxicity in medaka (Oryzias latipes): Stage- specific toxicity and the protective role of chorion. Aquatic Toxicology, 48, (2-3), 309-326.
Villalobos, S.A., Papoulias, D.M., Meadows, J., Blankenship, A.L., Pastva, S.D., Kannan, K., Hinton, D.E., Tillitt, D.E., Giesy, J.P. 2000b. Toxic responses of medaka, d-rR strain, to polychlorinated naphthalene mixtures after embryonic exposure by in ovo nanoinjection: A partial life- cycle assessment. Environmental Toxicology and Chemistry, 19(2): p. 432-440.
Walsh, T.P.M.P.J. 1988. Vitlogenesis and oocyte assembly, in: Fish physiology, W.S.H.D.J. Randall (ed.), Academic press, inc. California USA.
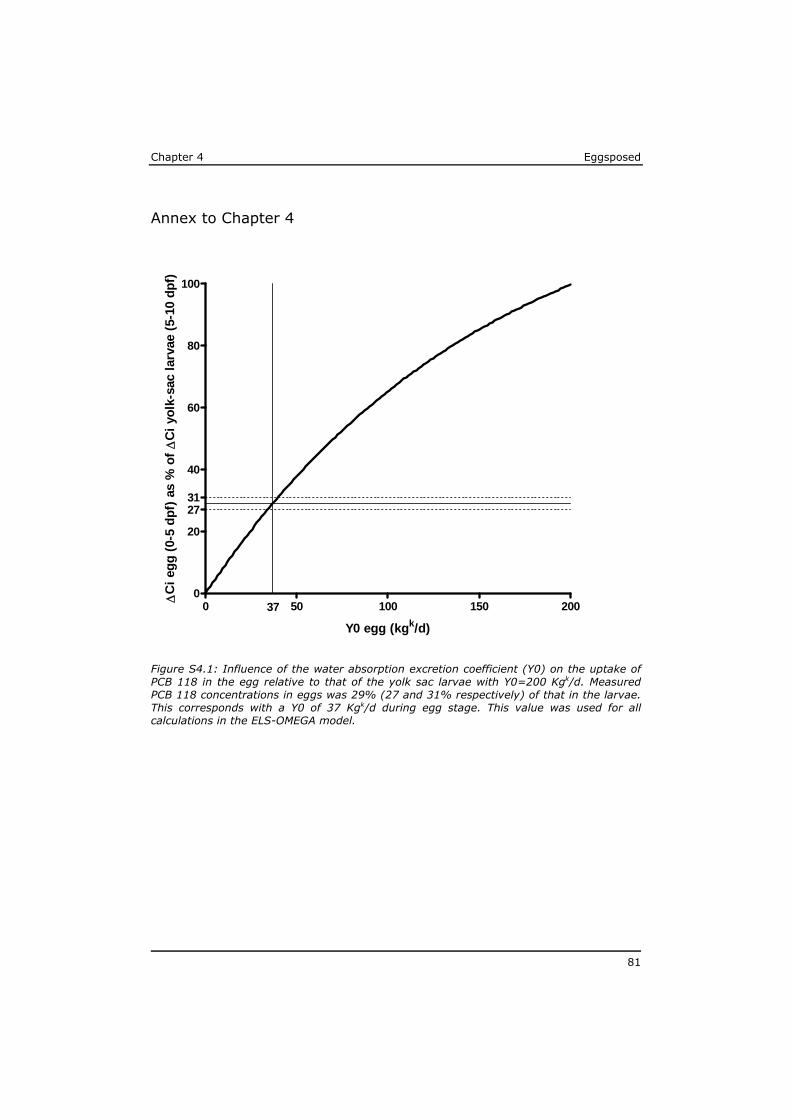
Chapter 4 Eggsposed
81
Annex to Chapter 4
0 50 100 150 2000
20
40
60
80
100
Y0 egg (kgk/d)
C
i eg
g (
0-5
dp
f) a
s %
of
Ci
yolk
-sac
lar
vae
(5-1
0 d
pf)
2731
37
Figure S4.1: Influence of the water absorption excretion coefficient (Y0) on the uptake of PCB 118 in the egg relative to that of the yolk sac larvae with Y0=200 Kgk/d. Measured PCB 118 concentrations in eggs was 29% (27 and 31% respectively) of that in the larvae. This corresponds with a Y0 of 37 Kgk/d during egg stage. This value was used for all calculations in the ELS-OMEGA model.
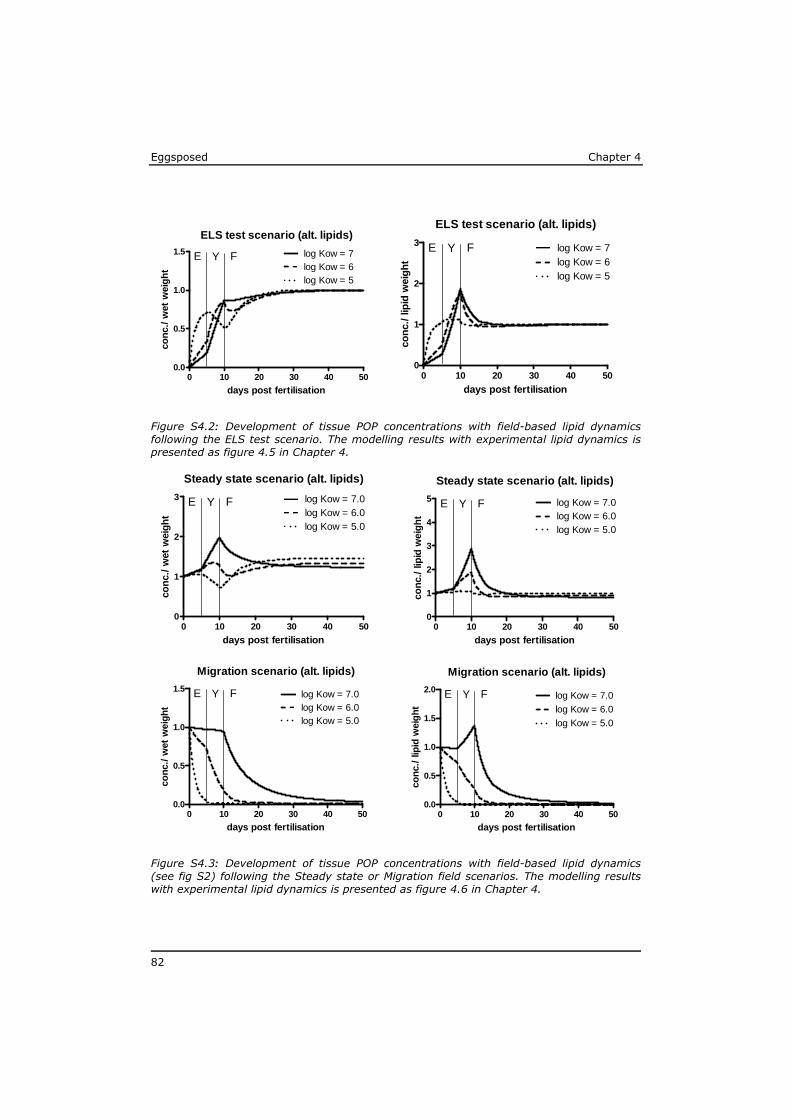
Eggsposed Chapter 4
82
ELS test scenario (alt. lipids)
0 10 20 30 40 500.0
0.5
1.0
1.5
days post fertilisation
con
c./
wet
wei
gh
t
E Y F log Kow = 7log Kow = 6log Kow = 5
ELS test scenario (alt. lipids)
0 10 20 30 40 500
1
2
3
days post fertilisation
con
c./
lipid
wei
gh
t
E Y F log Kow = 7
log Kow = 5log Kow = 6
Figure S4.2: Development of tissue POP concentrations with field-based lipid dynamics following the ELS test scenario. The modelling results with experimental lipid dynamics is presented as figure 4.5 in Chapter 4.
Steady state scenario (alt. lipids)
0 10 20 30 40 500
1
2
3
log Kow = 6.0log Kow = 7.0
days post fertilisation
con
c./
wet
wei
gh
t
log Kow = 5.0
E Y F
Steady state scenario (alt. lipids)
0 10 20 30 40 500
1
2
3
4
5
days post fertilisation
con
c./
lipid
wei
gh
t
E Y F log Kow = 7.0log Kow = 6.0log Kow = 5.0
Migration scenario (alt. lipids)
0 10 20 30 40 500.0
0.5
1.0
1.5
log Kow = 6.0
days post fertilisation
con
c./
wet
wei
gh
t
log Kow = 5.0
E Y F log Kow = 7.0
Migration scenario (alt. lipids)
0 10 20 30 40 500.0
0.5
1.0
1.5
2.0
days post fertilisation
con
c./
lipid
wei
gh
t
E Y F log Kow = 7.0
log Kow = 6.0
log Kow = 5.0
Figure S4.3: Development of tissue POP concentrations with field-based lipid dynamics (see fig S2) following the Steady state or Migration field scenarios. The modelling results with experimental lipid dynamics is presented as figure 4.6 in Chapter 4.

83
CHAPTER 5
The risks of maternally transferred organic substances for fish early life development
E.M. Foekemaa, M. Lopez Parronb, M.T. Mergiab, E.R.M. Carolusb, J.H.J. vd Bergb, C. Kwadijkc, Q. Daoc , and A.J. Murkb
Submitted
a IMARES, Department Experimental Ecology b Wageningen University, Division of Toxicology c IMARES, Department Fish

Chapter 5 Eggsposed
84
Abstract
The present study investigates the likelihood that early life development of fish from contaminated areas is affected by maternally transferred persistent organic substances (POPs). The common sole (Solea solea) was used as model species and maternal transfer was mimicked by exposing fertilised eggs via the water until hatching. The newly hatched larvae were allowed to develop further under unexposed conditions until the end of the metamorphosis. Effects on the larvae were determined for the dioxin-like PCB 126, the technical PCB-mixture Arochlor 1254, PBDEs and HBCDs, for an artificial mixture of PCBs and PBDEs, and for ‘field mixtures’ extracted from sole from the North Sea and the contaminated Western Scheldt estuary. Effect levels were expressed as maximum tissue concentrations in the newly hatched larvae at the end of the exposure period. Exposure to PCBs, PBDEs and the artificial and field mixtures caused mortality that started to occur shortly after the larvae became free-feeding (10 days post fertilization) and continued to increase until the onset of metamorphoses, 15 days later. The effects induced by the field mixtures correlated well with the ∑PCB concentrations in the tissue of the exposed larvae. No indications were found for synergistic effects or for substantial contribution of other (unknown) substances in the field mixtures. HBCD did not induce toxic effects. POP levels in sole from the Western Scheldt estuary are about 20 times lower than the larval tissue concentration that produced 50% early life stage mortality. Levels in North Sea sole are an order of a magnitude lower. At more heavily contaminated sites negative effect of PCBs, especially of those with dioxin-like toxicity can be expected.

Eggsposed Chapter 5
85
5.1 Introduction
Although bulk production and use has been stopped for persistent organic pollutants (POPs), compounds like polychlorinatedbiphenyls (PCBs), brominated flame retardants (polybrominated diphenyl ethers PBDEs, hexabromocyclododecane HBCDs), and chlorinated pesticides (dichloro-diphenyl-trichloroethane DDT, Lindane, Dieldrin, etc) still are present in the environment (de Boer et al., 2010; Maes et al., 2008; Voorspoels et al., 2004a). As a result of the low water solubility/high lipophilicity in combination with high persistence these substances concentrate in sediments and accumulate in biota, including fish. During oogenesis, POPs that were accumulated in female lipid tissue are transferred to the eggs (Miller, 1993; Monteverdi and Di Giulio, 2000; Nyholm et al., 2008; Serrano et al., 2008), and can cause negative effects on the developing off-spring after fertilisation (Tietge et al., 1998; Olsson et al., 1999; Nakayama et al., 2005; Ishibashi et al., 2006).
Due to a homogeneous distribution of the POPs over the lipids in the female tissue, the lipid normalised concentration in the eggs will be comparable to the mothers tissue (Russell et al., 1999). Thus, the POP concentration in the tissue of the mother fish represents the minimum toxic pressure for the developping offspring. For lipophilic substances (log Kow>5), the lipid normalised concentration will further increase in the developping larvae when the lipids stored in the yolk are depleted. This results in an exposure peak to these substances at the end of the yolk-sac stage that is the highest to be expected during the life time of the fish (Chapter 4; Foekema et al., 2012). Compared to fully developed fish, larvae are relatively sensitive to toxicants (Hutchinson et al., 1998) as a concequence of the critical development of organs and tissues during this life phase.
In an earlier study we determined that in a prolonged Early Life Stage (pELS) test with the common sole (Solea solea) the effect concentrations of dioxin-like compounds in the larvae were in the same order of magnitude as levels found in tissue of fish from highly contaminated areas. This implies that in these areas maternal transfer of such compounds can negatively affect the larval development (Foekema et al., 2008). In addition to dioxin-like compounds also other POPs are found in fish from polluted field locations, and the present paper assesses the effect of well-known field-relevant lipophilic POPs (PCBs, PBDEs, and HBCDs) that will be maternally transferred, on early life development of Solea solea. These POPs have, among others, been identified as potential endocrine disrupters (Zimmer et al., 2011), including the thyroid hormone system (Brouwer et al., 1999; Noyes et al., 2011; Palace et al., 2010; Schriks et

Chapter 5 Eggsposed
86
al., 2006; Yu et al., 2011) which plays an essential role in early development and (flat)fish metamorphosis (Klaren et al., 2008).
The common sole (Solea solea) was chosen as model species for this study as it is available from aquaculture which ensures almost year round availability of fertilized eggs. Also, sole is a common marine species along the North Sea coast, that uses the contaminated Western Scheldt estuary (Baeyens et al., 2003; De Vijver et al., 2003; Roosens et al., 2008; Voorspoels et al., 2004a; Voorspoels et al., 2004b; Voorspoels et al., 2003) as nursery area and feeding ground until first reproduction (Rijnsdorp et al., 1992). Further advantage of using sole is that, as a flatfish, it undergoes an obvious thyroid hormone mediated metamorphosis (Klaren et al., 2008) and thus is potentially sensitive to thyroid hormone disruption as has been described for other species (Noyes et al., 2011; Palace et al., 2010; Schriks et al., 2006; Yu et al., 2011; Gutleb et al., 1999). Last but not least, an optimized protocol is available for testing the impact of substances on the full development from a fertilized egg into a metamorphosed flat fish (Foekema et al., 2008).
In this study first the effect concentrations are determined for the selected POPs PCBs, PBDEs and HBCD, and secondly, a test with an artificial mixture of PCBs and PBDEs is performed in order to determine possible mixture effects. Finally, sole eggs are exposed to two ‘field mixtures’ of organic lipophilic pollutants extracted from tissue of sole from the North Sea and the Western Scheldt estuary. Invertebrates and fish from the Western Scheldt estuary are known to be contaminated with PCBs and brominated flame retardants (Baeyens et al., 2003; De Vijver et al., 2003; Roosens et al., 2008; Voorspoels et al., 2004a; Voorspoels et al., 2004b; Voorspoels et al., 2003; Janak et al., 2005), but also with other substances as perfluorooctane sulfonic acid (PFOS; De Vijver et al., 2003) and organo-chlorine pesticides (OCPs; Voorspoels et al., 2004a).
The comparison of the effects of the field mixtures with those induced by the artificial mixture, composed of only PCBs and PBDEs, indicates to what extent other (unknown) substances present in the cocktail of POPs extracted from the fish contribute to an effect on the developing larvae.
In summary, the present study aims to assess the likelihood that maternally transferred POPs negatively affect early life development of fish from contaminated areas with special focus on PCBs, HBCD and PBDEs, and (other) POPs present in sole from the Western Scheldt estuary.

Eggsposed Chapter 5
87
5.2 Materials and methods
5.2.1 Test organisms and seawater
Fertilized Sole eggs were produced at IMARES (location IJmuiden, the Netherlands) where a group of mature male and female soles is kept together in large spawning tanks. Spawning and fertilization was temperature-induced and took place overnight at water temperatures around 12 ºC. The following morning, the fertilized eggs were transported in plastic bags with oxygen saturated seawater to the IMARES laboratory in Den Helder, the Netherlands, were the tests were performed. Together with the eggs, artificial seawater (Instant Ocean Aquarium Salt in demineralized water, salinity ~35‰) from the spawning tank was transported to the laboratory. Here, the water was stored in a container were it was continuously aerated and circulated through a particle filter and UV-disinfection system. This water was used for the start of the tests. During the test the artificial seawater was gradually replaced with natural seawater (salinity ~32 ‰) collected from the Eastern Scheldt, a relatively pristine bay of the North Sea, often used as a reference site in marine ecotoxicological research in the Netherlands (e.g. Kuiper et al., 2007; Foekema et al., 2008; Foekema et al., 2012).
For the tests presented in this paper three batches of eggs were used, one for the tests with the PCB mixture Arochlor 1254, PCB 126 and HBCD, and two others for testing the PBDE and the artificial and field mixtures.
5.2.2 Sole ELS test
The sole prolonged early life stage (pELS) test was performed according to Foekema et al., 2008, with some slight modifications to improve test performance or to simplify the method.
Immediately after delivery at the laboratory, fertilized eggs were transferred to 5 L glass beakers and placed in a temperature controlled (15°C) room. The beakers were left untouched for about one hour after which non-fertilized/dead eggs had sunk to the bottom of the beaker. Exposure started approximately 12 hours after fertilization by transferring 500 fertilized eggs in approximately 50-100 ml seawater to the glass ‘exposure beakers’ containing 2 L seawater with the desired exposure concentration.
The exposure beakers were covered with individual glass lids and kept in a temperature controlled cabinet at 14.5 ±0.5°C, with a 16 hour photoperiod (ca. 100 LUX). During the following 5 days, about 60-75% of the water volume was replaced every other day with freshly spiked seawater. Dead eggs were removed

Chapter 5 Eggsposed
88
daily. The first larvae hatched around 5 days post fertilization (dpf). At 6 dpf, from each exposure group, two times 15 larvae were randomly selected and transferred to two smaller glass beakers containing 80 ml of clean seawater. This was done by means of a polyethylene pipette, with as little exposure water as possible. The larvae were then allowed to develop further under similar, yet unexposed conditions. Every other day, approximately 75% of the water volume in each glass beaker was replaced with fresh seawater, and almost daily, feces, surplus food items, and where appropriate dead larvae were removed with a polyethylene pipette. From 9 dpf onwards, when pigmentation of the eyes indicates that the larvae are about to start external feeding, they were fed ad libitum with newly hatched naupllii of Artemia salina on a daily basis.
The development of the fish was recorded following the stages described for the development from fertilized egg to a fully metamorphosed summer flounder (Paralichthys dentatus) (Martinez and Bolker, 2003). At the given test conditions sole metamorphosis (stage G) starts around 25 dpf, and is completed (stage I) around 40 dpf. Fish that completed metamorphoses were taken out of the test, narcotized and subsequently killed in a strong MS222-solution (500 mg of ethyl 3-aminonebzoate methanesulfonic acid salt, purity 98%, ACROS Organics, and 200 mg NaHCO3, purity 99.0%, Fluka Chemika, in 1 liter seawater). Moment of completed metamorphosis, body length, and morphological deviations, if any, were recorded.
All experiments were approved by the Animal Experimentation Board of Wageningen UR.
5.2.3 Chemicals, stock solutions and exposure concentrations
The technical PCB mixture Arochlor 1254 (Monsanto, USA) was used to prepare the stock solutions for the treatment that will be further referred to as ARO. The contribution of the different PCB congeners in this mixture was chemically analyzed (Table S4.2 in Annex to Chapter 5). The PBDE mixture was composed of standards of five different congeners: BDE 28, 47, 99, 100 and 153 (Accustandard, USA, purity 99.3-100%). The rationale behind the chosen composition (Table S5.2 in Annex to Chapter 5) was that exposure of the eggs will result in internal mixture of the individual congeners with a composition that is comparable to what is found in fish tissue from the Western Scheldt estuary. The HBCD and PCB 126 that were used in this study were obtained from Albermarle and Promochem respectively. Some background information on the substances selected for this study is given in the supplementary data, including the molecular weights and the log Kow values that were used for calculations (Table S5.1 in Annex to Chapter 5).

Eggsposed Chapter 5
89
For each exposure concentration a 1000x concentrated stock solution was prepared in dimethyl sulfoxide (DMSO, purity 99.9% A.C.S. spectrophotometric grade, supplier SIGMA-Aldrich), so that addition of 1 ml of this stock solution in 1 liter seawater produced the appropriate exposure concentration in 0.1% DMSO. In every test series triplicate solvent controls and seawater controls were included.
The stock solution for the test with the artificial mixture was prepared by combining Arochlor-1254, PCB 126 and the PBDE mixture, in such proportions that, based on accumulation kinetics as a function of the octanol-water partition ratio (Kow), will result in a composition that is comparable to what is found in tissue of fish from the Western Scheldt estuary.
Tests with PCBs, PBDEs and HBCD were performed twice in duplicate. The exposure concentrations (Table 5.1) were determined based on a range finding tests (data not shown). The field mixtures only were tested once, due to the limited amount of fish extract available.
PBDEs, the artificial mixture (Mix), and the field mixtures were tested in five concentrations, while for arochlor (ARO), PCB 126 (P126) and HBCD only three concentrations could be tested due to the limited number of available eggs.
Table 5.1 Nominal exposure concentrations of PCB 126, ∑PCBs (Arochlor 1254), PBDEs and HBCD that were used in the tests with the individual substances.
Code Unit (µg/L) C1 C2 C3 C4 C5
P126 PCB 126 0.003 0.01 0.03
ARO ∑ 25 PCBs 2.1 7 21
PBDE ∑ 5 PBDEs 0.4 1.1 3.3 10 30
HBCD ∑ HBCD 25 80 250
5.2.4 Extraction of field mixtures from field samples
Sole from the Western Scheldt estuary was collected by beam trawl fishing at two locations near the places Terneuzen (WS1) and Hansweert (WS2), in the autumn of the year 2006. From each location, about 60 individual Sole with a length between 10 and 20 cm, (1 to 2 years of age) were collected to compose a sample of about 1 kg wet weight (ww). A sample of sole from the North Sea (NS) with a length between 25 and 40 cm was obtained from commercial fishermen. Filets were used to compose a sample of about 3 kg ww. Spawn that was present in female fish from the North Sea sample was collected separately (NS-spawn).

Chapter 5 Eggsposed
90
The samples were freeze-dried, ground, mixed with dried Na2SO4 and in portions extracted in a Soxhlet extractor with n-pentane and dichloromethane (1:1). The extract was concentrated in a rotary evaporator with iso-octane as a keeper. Lipid was removed on an Al2O3 column eluted with n-pentane. For a more detailed method description see De Boer & Hagel, 1994.
A sub sample of all extracts was used for chemical analyses of the concentrations of PCBs, HBCDs, and PBDEs. The residues were evaporated to almost dryness and dissolved in DMSO to be used for exposure in the p-ELS test. Because of the limited volume, the extract of the NS spawn was only used for chemical analyses and was not used in the p-ELS test.
5.2.5 Chemical analysis of larvae
All larvae that were not included for observation of the development in the P-ELS test were sampled for chemical analysis at the end of the exposure period (6 dpf). They were transferred with a polyethylene pipette from the exposure beakers to clean seawater to rinse remaining exposure water, and within 15 minutes collected by filtration on a Whatman GF/C filter with known wet (seawater) weight. The filter with the larvae was weighed again and stored at -20°C. To prepare for analysis the filters with the larvae were subsequently extracted three times with 4 ml of a mixture of pentane : dichloromethane (1:1, both picograde LGC standards, Germany) assisted with ultra-sonification. The extracts were combined in a test tube and evaporated to dryness under a gentle stream of nitrogen. The residue was reconstituted in 1 ml injection standard (PCB112, purity >99%, LGC standards; 13C-α, β,γ HBCD (purity >99%, Wellington, >99%), and BDE58 (Wellington, >99%) and transferred to a vial for GC/MS analysis of 25 PCBs, PCB 126, total HBCD and 5 PBDEs using an Agilent 6890 gas chromatograph (GC) coupled to an Agilent 5973 MSD quadropole mass spectrometer (MS) detector (Amstelveen, The Netherlands).
5.2.6 Expression of results, calculations and statistical analysis
Since all test substances were very lipophilic (log Kow >5; (Hansen et al., 1999; Li et al., 2008)(see Table S5.2 in Annex to Chapter 5), the analyzed tissue concentrations were expressed on lipid weight (lw) (Murk et al., 1998). Survival and development were plotted against this internal dose to estimate the internal EC50 and LC50 (IEC50 and ILC50), using GraphPad Prism (version 4.03, January 2005).
Based on the IEC50 values for the individual chemical groups the toxic contributions were quantified as Toxic Units (TU = tissue concentration / IEC50) for substances found in the larvae exposed to the artificial and field mixtures.

Eggsposed Chapter 5
91
The total toxicity of the compounds found in the larvae exposed to the mixtures was then expressed as the ∑TU. Since the TU are calculated as fraction of the IEC50 value 50% effect can be expected at ∑TU =1. This approach follows the concept of concentration addition and can be regarded as a conservative method to calculate the toxicity of mixtures (Backhaus and Faust, 2012).
5.3 Results
5.3.1 General test performance
Hatching success was normal for cultured sole, about 50% in all test series, and not related to the exposures. After hatching the average survival in the seawater controls in all test series, was 87% (73-100%, n=9) at the end of the test (40 dpf). In the DMSO controls this was slightly higher (average 91%, range 80-100%, n=9) without being significantly different. Around 25 dpf the first fish reached the onset of metamorphoses, and all surviving fish in the controls had completed the metamorphoses without problems at 40 dpf.
Table 5.2 Summary of the test results with PCB 126, Arochlor, PBDEs and HBCD. Presented are the range of tissue concentrations in the larvae from the different exposure groups at the end of the exposure (6 dpf), and characteristics of the internal-dose vs response curves on 40 dpf (IEC50 with 95% confidence intervals and R square).
Mortality Disrupted development
PCB 126
Larvae 6 dpf (range µg/g lw) 4.3-52 4.3-52
IEC50 (µg/g lw) 1.3 (0.20 to 8.36) <1.3
R2 0.76 No fit
∑ 25 PCBs
Larvae 6 dpf (range µg/g lw) 320-502 320-502
IEC50 (µg/g lw) 374 (330 to 424) 341 (328 to 356)
R2 0.88 0.98
∑ 5 PBDEs
Larvae 6 dpf (range µg/g lw) 81 - 1950 81 - 1950
IEC50 (µg/g lw) 1777 (1717 to 1839) 1636 (1570 to 1704)
R2 0.99 0.98
HBCD
Larvae 6 dpf (range µg/g lw) 2280-12400 2280-12400
IEC50 (µg/g lw) >12400 >12400
R2 No fit No fit

Chapter 5 Eggsposed
92
5.3.2 Effects of selected POPs
The concentrations of the selected substances in tissue of the larvae at the end of the exposure period (6 dpf), clearly reflected the exposure concentration (Figure 5.1). The ARO (∑PCB), and PBDE concentrations seemed to approach a plateau at higher exposure levels. Concentrations were below the limit of detection in larvae from non-exposed controls and reached relatively high concentrations at the highest exposure levels: 52, 502, 1950 and 12400 µg/g lw for PCB 126, ∑PCBs (Arochlor), ∑PBDEs and HBCD respectively (Table 5.2).
The larvae that had been exposed to PCB 126, Arochlor, and PBDEs, showed the first indications of increased mortality around 10 dpf, the end of the yolk-sac / start of the free- feeding stage (Figure 5.2, left column). The timing of the increased mortality was comparable for all these substances, clearly dose related, and continued at least until the fish reached the onset of the metamorphoses (25-30 dpf). Only in individual cases mortality occurred after metamorphoses had started. Complete mortality was observed in the highest exposure level of PCB 126 (0.03 µg/L; P126-C3). In the lower exposure levels (0.01 and 0.003 µg PCB 126/L) overall survival was 10 and 25% respectively, and less than half of these survivors completed metamorphoses. At all exposure levels of PCB 126, some larvae developed oedema before dying.
All larvae in the highest exposure level of Arochlor (21 µg/L; ARO-C3) died during the observation period. At the lower exposure levels survival was higher, and in ARO-C1 (2.1 µg/L) 70% of the larvae survived the 40 day test period. The majority (86%) of these larvae successfully completed metamorphoses. In the intermediate ARO-C2 27% of the larvae survived, but only 7% had completed metamorphoses at 40 dpf.
Survival and development of the larvae at exposure levels up to 3.3 µg PBDE/L (PBDE-C1 to –C3) was comparable with the controls. In PBDE-C4 (10 µg/L) survival on dpf 40 was 67%, and in PBDE-C5 (30 µg/L) this was 36%. Only in PBDE-C5 some of the surviving larvae were arrested in an early developmental stage, and in total 27% of the hatched larvae developed in normally metamorphosed juveniles within 40 dpf. HBCD did not induce any mortality or adverse effects.
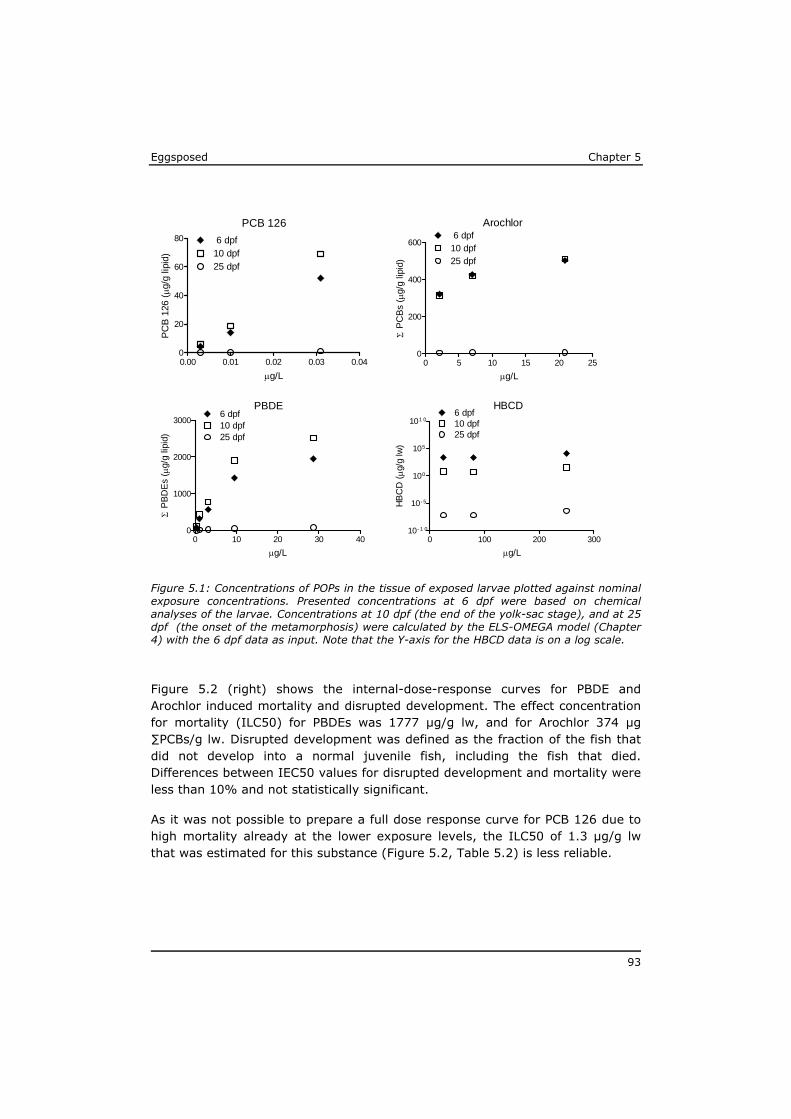
Eggsposed Chapter 5
93
Figure 5.1: Concentrations of POPs in the tissue of exposed larvae plotted against nominal exposure concentrations. Presented concentrations at 6 dpf were based on chemical analyses of the larvae. Concentrations at 10 dpf (the end of the yolk-sac stage), and at 25 dpf (the onset of the metamorphosis) were calculated by the ELS-OMEGA model (Chapter 4) with the 6 dpf data as input. Note that the Y-axis for the HBCD data is on a log scale.
Figure 5.2 (right) shows the internal-dose-response curves for PBDE and Arochlor induced mortality and disrupted development. The effect concentration for mortality (ILC50) for PBDEs was 1777 µg/g lw, and for Arochlor 374 µg ∑PCBs/g lw. Disrupted development was defined as the fraction of the fish that did not develop into a normal juvenile fish, including the fish that died. Differences between IEC50 values for disrupted development and mortality were less than 10% and not statistically significant.
As it was not possible to prepare a full dose response curve for PCB 126 due to high mortality already at the lower exposure levels, the ILC50 of 1.3 µg/g lw that was estimated for this substance (Figure 5.2, Table 5.2) is less reliable.
0.00 0.01 0.02 0.03 0.040
20
40
60
80
g/L
PC
B 1
26 (g
/g li
pid) 10 dpf
6 dpf
PCB 126
25 dpf
0 5 10 15 20 250
200
400
600
g/L
P
CB
s (
g/g
lipid
)
10 dpf 6 dpf
Arochlor
25 dpf
0 10 20 30 400
1000
2000
3000
g/L
P
BD
Es
( g/
g lip
id)
10 dpf6 dpf
PBDE
25 dpf
0 100 200 30010- 1 0
10- 5
100
105
101 0
g/L
HB
CD
(g
/g lw
)
10 dpf6 dpf
HBCD
25 dpf
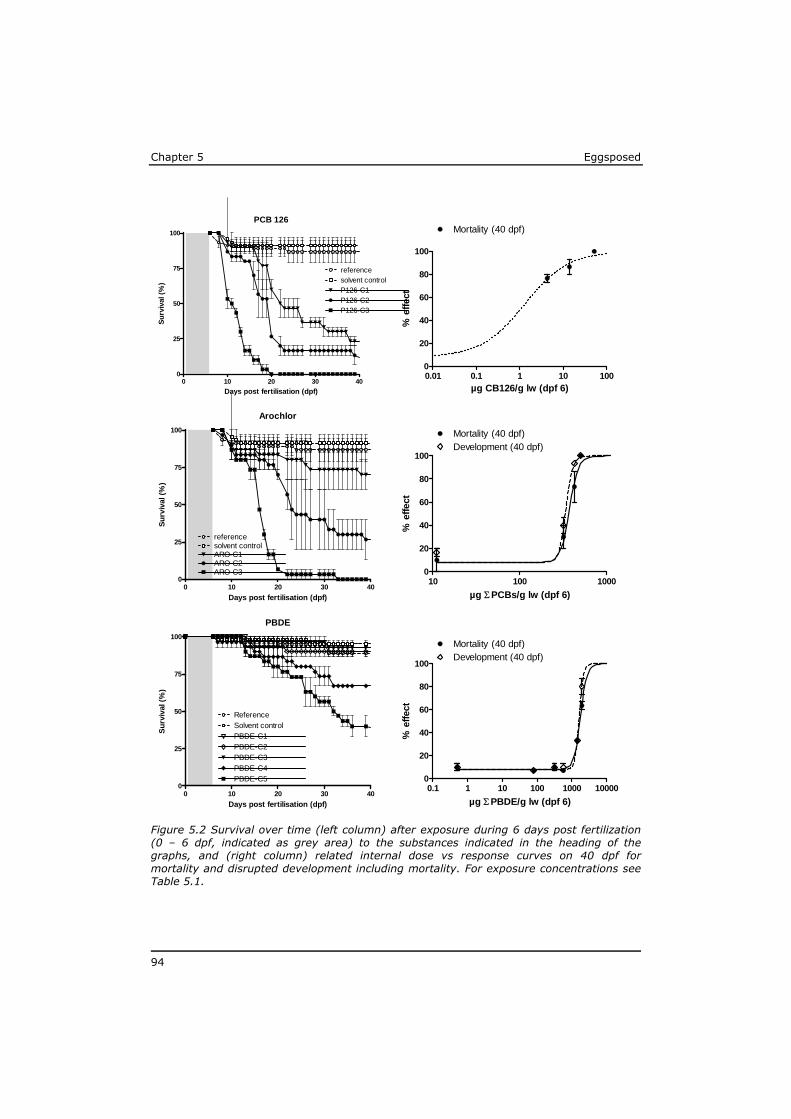
Chapter 5 Eggsposed
94
Figure 5.2 Survival over time (left column) after exposure during 6 days post fertilization (0 – 6 dpf, indicated as grey area) to the substances indicated in the heading of the graphs, and (right column) related internal dose vs response curves on 40 dpf for mortality and disrupted development including mortality. For exposure concentrations see Table 5.1.
PCB 126
0 10 20 30 400
25
50
75
100
referencesolvent controlP126-C1P126-C2P126-C3
Days post fertilisation (dpf)
Su
rviv
al (
%)
µg CB126/g lw (dpf 6)
% e
ffec
t
0.01 0.1 1 10 1000
20
40
60
80
100
Mortality (40 dpf)
Arochlor
0 10 20 30 400
25
50
75
100
referencesolvent controlARO-C1ARO-C2ARO-C3
Days post fertilisation (dpf)
Su
rviv
al (
%)
µg PCBs/g lw (dpf 6)
% e
ffec
t
10 100 10000
20
40
60
80
100
Mortality (40 dpf)Development (40 dpf)
PBDE
0 10 20 30 400
25
50
75
100
ReferenceSolvent controlPBDE-C1PBDE-C2PBDE-C3
Days post fertilisation (dpf)
Su
rviv
al (
%)
PBDE-C4PBDE-C5
µg PBDE/g lw (dpf 6)
% e
ffec
t
0.1 1 10 100 1000 100000
20
40
60
80
100
Mortality (40 dpf)Development (40 dpf)
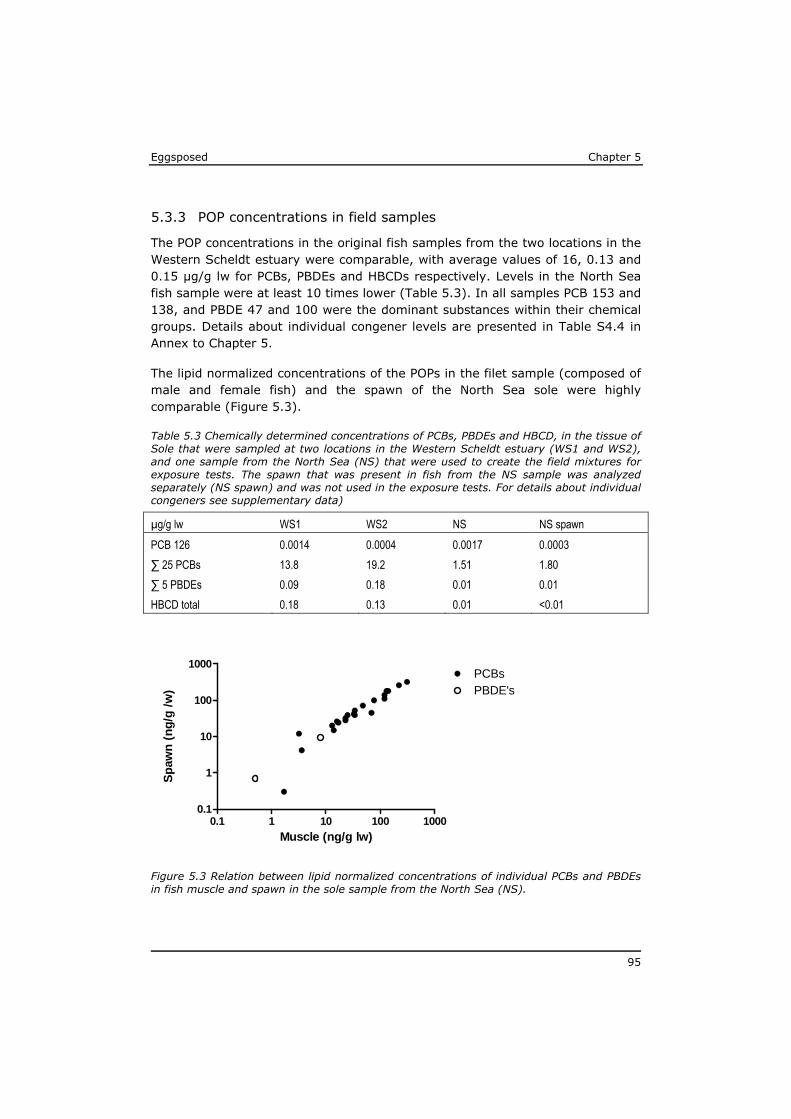
Eggsposed Chapter 5
95
5.3.3 POP concentrations in field samples
The POP concentrations in the original fish samples from the two locations in the Western Scheldt estuary were comparable, with average values of 16, 0.13 and 0.15 µg/g lw for PCBs, PBDEs and HBCDs respectively. Levels in the North Sea fish sample were at least 10 times lower (Table 5.3). In all samples PCB 153 and 138, and PBDE 47 and 100 were the dominant substances within their chemical groups. Details about individual congener levels are presented in Table S4.4 in Annex to Chapter 5.
The lipid normalized concentrations of the POPs in the filet sample (composed of male and female fish) and the spawn of the North Sea sole were highly comparable (Figure 5.3).
Table 5.3 Chemically determined concentrations of PCBs, PBDEs and HBCD, in the tissue of Sole that were sampled at two locations in the Western Scheldt estuary (WS1 and WS2), and one sample from the North Sea (NS) that were used to create the field mixtures for exposure tests. The spawn that was present in fish from the NS sample was analyzed separately (NS spawn) and was not used in the exposure tests. For details about individual congeners see supplementary data)
µg/g lw WS1 WS2 NS NS spawn
PCB 126 0.0014 0.0004 0.0017 0.0003
∑ 25 PCBs 13.8 19.2 1.51 1.80
∑ 5 PBDEs 0.09 0.18 0.01 0.01
HBCD total 0.18 0.13 0.01 <0.01
Figure 5.3 Relation between lipid normalized concentrations of individual PCBs and PBDEs in fish muscle and spawn in the sole sample from the North Sea (NS).
Muscle (ng/g lw)
Sp
awn
(n
g/g
/w
)
0.1 1 10 100 10000.1
1
10
100
1000PCBsPBDE's
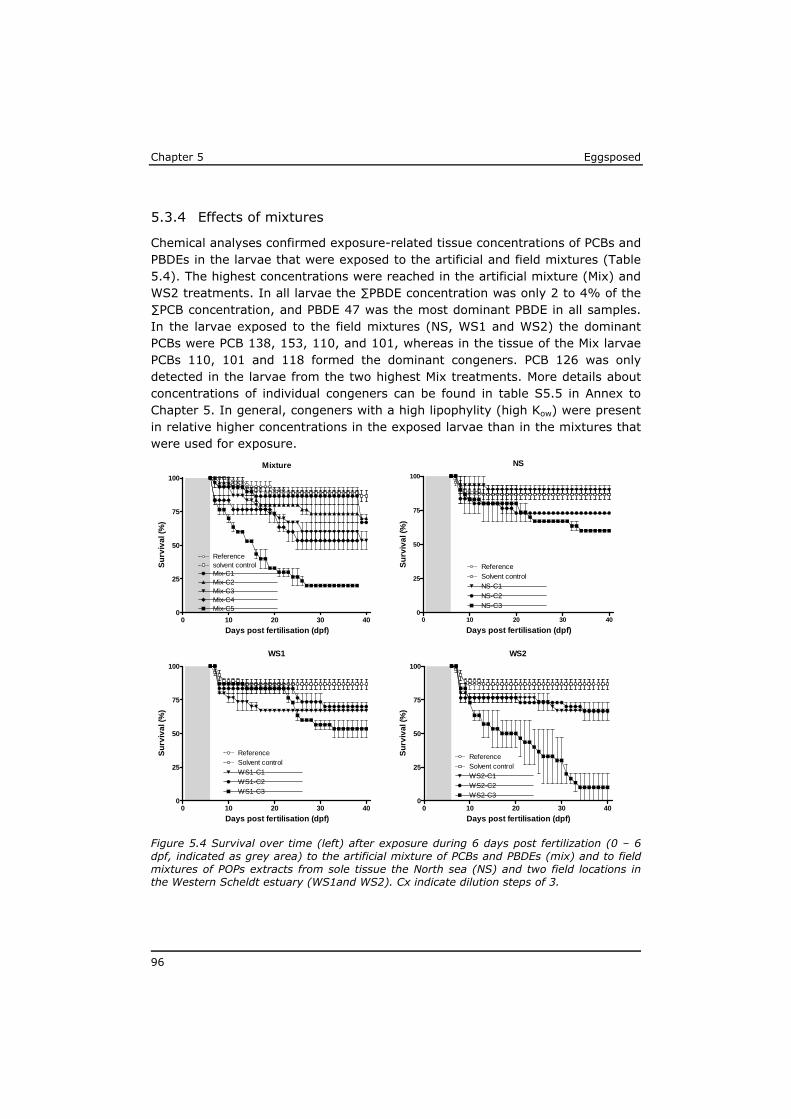
Chapter 5 Eggsposed
96
5.3.4 Effects of mixtures
Chemical analyses confirmed exposure-related tissue concentrations of PCBs and PBDEs in the larvae that were exposed to the artificial and field mixtures (Table 5.4). The highest concentrations were reached in the artificial mixture (Mix) and WS2 treatments. In all larvae the ∑PBDE concentration was only 2 to 4% of the ∑PCB concentration, and PBDE 47 was the most dominant PBDE in all samples. In the larvae exposed to the field mixtures (NS, WS1 and WS2) the dominant PCBs were PCB 138, 153, 110, and 101, whereas in the tissue of the Mix larvae PCBs 110, 101 and 118 formed the dominant congeners. PCB 126 was only detected in the larvae from the two highest Mix treatments. More details about concentrations of individual congeners can be found in table S5.5 in Annex to Chapter 5. In general, congeners with a high lipophylity (high Kow) were present in relative higher concentrations in the exposed larvae than in the mixtures that were used for exposure.
Figure 5.4 Survival over time (left) after exposure during 6 days post fertilization (0 – 6 dpf, indicated as grey area) to the artificial mixture of PCBs and PBDEs (mix) and to field mixtures of POPs extracts from sole tissue the North sea (NS) and two field locations in the Western Scheldt estuary (WS1and WS2). Cx indicate dilution steps of 3.
Mixture
0 10 20 30 400
25
50
75
100
Referencesolvent controlMix-C1Mix-C2Mix-C3
Days post fertilisation (dpf)
Su
rviv
al (
%)
Mix-C4Mix-C5
NS
0 10 20 30 400
25
50
75
100
ReferenceSolvent controlNS-C1NS-C2NS-C3
Days post fertilisation (dpf)
Su
rviv
al (
%)
WS1
0 10 20 30 400
25
50
75
100
ReferenceSolvent controlWS1-C1WS1-C2WS1-C3
Days post fertilisation (dpf)
Su
rviv
al (
%)
WS2
0 10 20 30 400
25
50
75
100
ReferenceSolvent controlWS2-C1WS2-C2WS2-C3
Days post fertilisation (dpf)
Su
rviv
al (
%)

Eggsposed Chapter 5
97
For all exposures to mixtures, survival of the developing larvae was clearly dose-related (Figure 5.4). The highest mortality was observed in the highest exposure levels of Mix and WS2. The development of the mortality over time was comparable in all tests; in most cases it started at the end of the yolk sac stage, thus a few days after the exposure was terminated, and continued until the onset of the metamorphoses starting around 25 dpf. Some of the larvae developed oedema before dying.
5.4 Discussion
Egg stage exposure to PCB 126, Arochlor and PBDE followed by unexposed further development resulted in increased mortality of the developing sole after the end of the yolk-sac stage. Although some exposed larvae were arrested in an early developmental stage and/or developed oedema, effect concentrations for mortality and development (including mortality) did not significantly differ. Exposure to high levels of HBCD did not lead to effects. Exposure to dilutions of an artificial mixture of PCBs and PBDEs, and to a field mixture of POPs extracted from sole filet from the Western Scheldt estuary and to a lesser extent the North Sea, induced comparable effects as exposure to PCBs and PBDEs.
5.4.1 General test performance
With the current test protocol about 90% of the larvae transferred to the smaller beakers on 6 dpf developed normally in clean seawater (reference) as well as in the solvent control (0.1% DMSO). This is a significant improvement compared to the first version of this assay where this was only about 70% (Chapter 2; Foekema et al., 2008). The main improvement was achieved by the use of the artificial seawater from the tank where spawning and fertilization took place during the first days of the test, followed by gradual replacement with natural seawater. In the earlier version of the protocol the fertilized eggs were immediately transferred to the natural seawater. Apparently the embryos are sensitive to abrupt changes in water characteristics. Disinfection of the seawater with UV before use further added to a good survival in the control groups.
Hatching success was not affected by any of the treatments in this study. Under the given test conditions sole larvae deplete the (lipid) resources in the yolk around dpf 10, which results in a peak in internal exposure for substances with log Kow>5 (Chapter 4; Foekema et al., 2012). Indeed, mortality rates increased beyond this moment, as was earlier reported in a similar experiment with the dioxin-like PCB 126 (Chapter 2; Foekema et al., 2008). The far majority of the surviving larvae completed metamorphoses without observable malformations. For this reason the differences between lethal and non-lethal effect

Chapter 5 Eggsposed
98
concentrations were very small. For calculation of the toxic units the more robust mortality data were preferred over the development data.
5.4.2 Exposure conditions and effect concentrations
The aim of the experimental approach was to simulate maternal transfer of bio accumulated substances, by a short term exposure of the eggs. For that reason the exposure concentrations used in the tests are much higher than water concentrations in even the highest contaminated field situation, and exposure was terminated after hatching. For PBDEs and Arochlor the relation between nominal water concentration and tissue concentration in the exposed larvae was not linear as would be expected assuming equilibrium between external and internal concentrations. The most likely explanation for this is that the short exposure time was not sufficient to reach equilibrium in the higher exposure levels, which is supported by reports that uptake rate during the egg stage is limited (Villalobos et al., 2000; Gonzalez-Doncel et al., 2003).
The reliability of the estimated ILC50 for PCB 126 is low since the lowest tested concentrations already caused 76% mortality. The ILC50 for PCB 126 of 1.3 µg/g lw (95% confidence intervals 0.2-8.4; Table 4.2) that was estimated in this study, is high compared to earlier work in a more or less comparable experiment with PCB 126 where a ILC50 of 1 ng TCDD-TEQ/g lw was determined using an in-vitro reporter gene assay (Foekema et al., 2008). Applying the WHO TCDD-TEF value of 0.1 for PCB 126 (Van den Berg et al., 2006), this TCDD TEQ would correspond to a PCB 126 concentration of 10 ng/g lw, which is below the lower 95%-confidence limit that was calculated in the present study. This suggests that the IEC50 value of PCB 126 from the present study is likely to be an underestimation of the real toxicity of PCB 126.
The timing of the observed effects, with mortality starting around the end of the yolk-sac stage (10 dpf) was comparable for all substances and mixtures tested, and comparable to earlier observations (Foekema et al., 2008). Most of the larvae that did not survive died before the onset of the metamorphosis that started around 25 dpf.
At high concentrations, accumulation of organic pollutants in biological membranes can affect cell integrity (vanWezel et al., 1996; Qin et al., 2010; Escher and Hermens, 2002; Escher and Schwarzenbach, 2002). This type of effect is referred to as narcosis or baseline toxicity and can be regarded as the minimum toxicity of a substance. Lethal body burdens of non-polar chemicals without specific toxicity are comparable and additive on molecular basis (vanWezel et al., 1996; Kipka and Di Toro, 2009). When the tissue concentration in the larvae at 6 dpf from our experiments is plotted on a
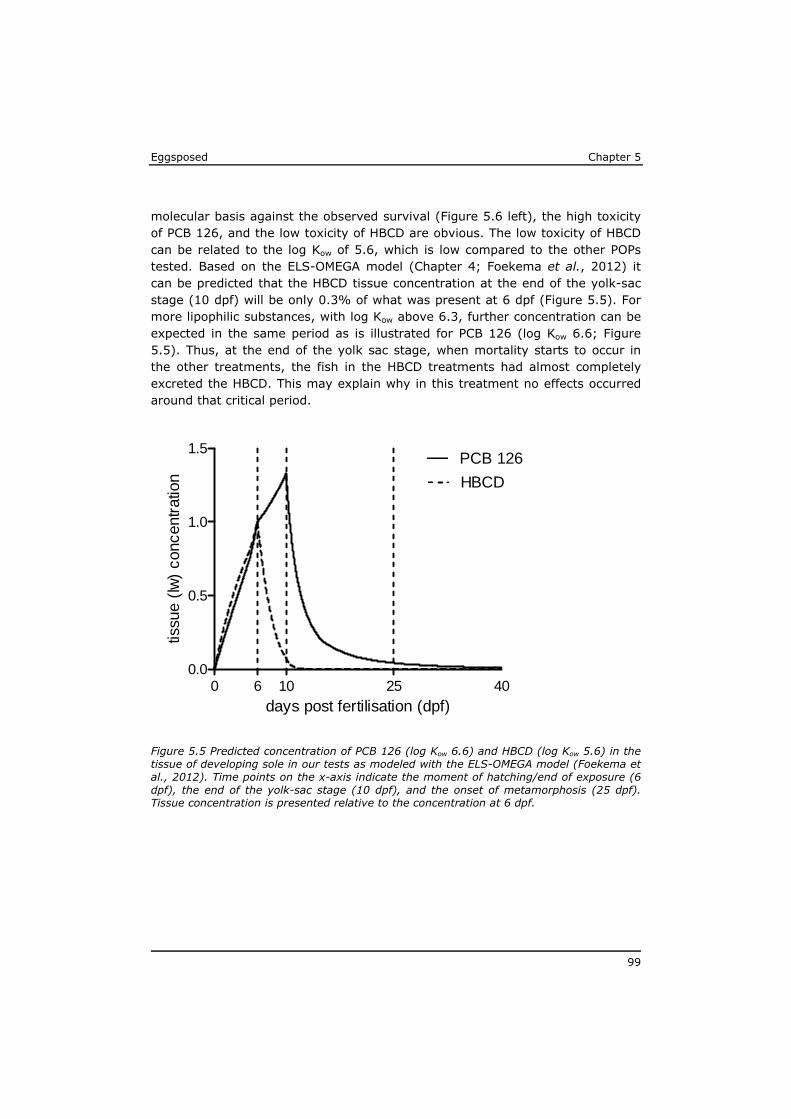
Eggsposed Chapter 5
99
molecular basis against the observed survival (Figure 5.6 left), the high toxicity of PCB 126, and the low toxicity of HBCD are obvious. The low toxicity of HBCD can be related to the log Kow of 5.6, which is low compared to the other POPs tested. Based on the ELS-OMEGA model (Chapter 4; Foekema et al., 2012) it can be predicted that the HBCD tissue concentration at the end of the yolk-sac stage (10 dpf) will be only 0.3% of what was present at 6 dpf (Figure 5.5). For more lipophilic substances, with log Kow above 6.3, further concentration can be expected in the same period as is illustrated for PCB 126 (log Kow 6.6; Figure 5.5). Thus, at the end of the yolk sac stage, when mortality starts to occur in the other treatments, the fish in the HBCD treatments had almost completely excreted the HBCD. This may explain why in this treatment no effects occurred around that critical period.
Figure 5.5 Predicted concentration of PCB 126 (log Kow 6.6) and HBCD (log Kow 5.6) in the tissue of developing sole in our tests as modeled with the ELS-OMEGA model (Foekema et al., 2012). Time points on the x-axis indicate the moment of hatching/end of exposure (6 dpf), the end of the yolk-sac stage (10 dpf), and the onset of metamorphosis (25 dpf). Tissue concentration is presented relative to the concentration at 6 dpf.
0.0
0.5
1.0
1.5
HBCD
days post fertilisation (dpf)
tissu
e (lw
) co
ncen
trat
ion
PCB 126
0 6 10 25 40
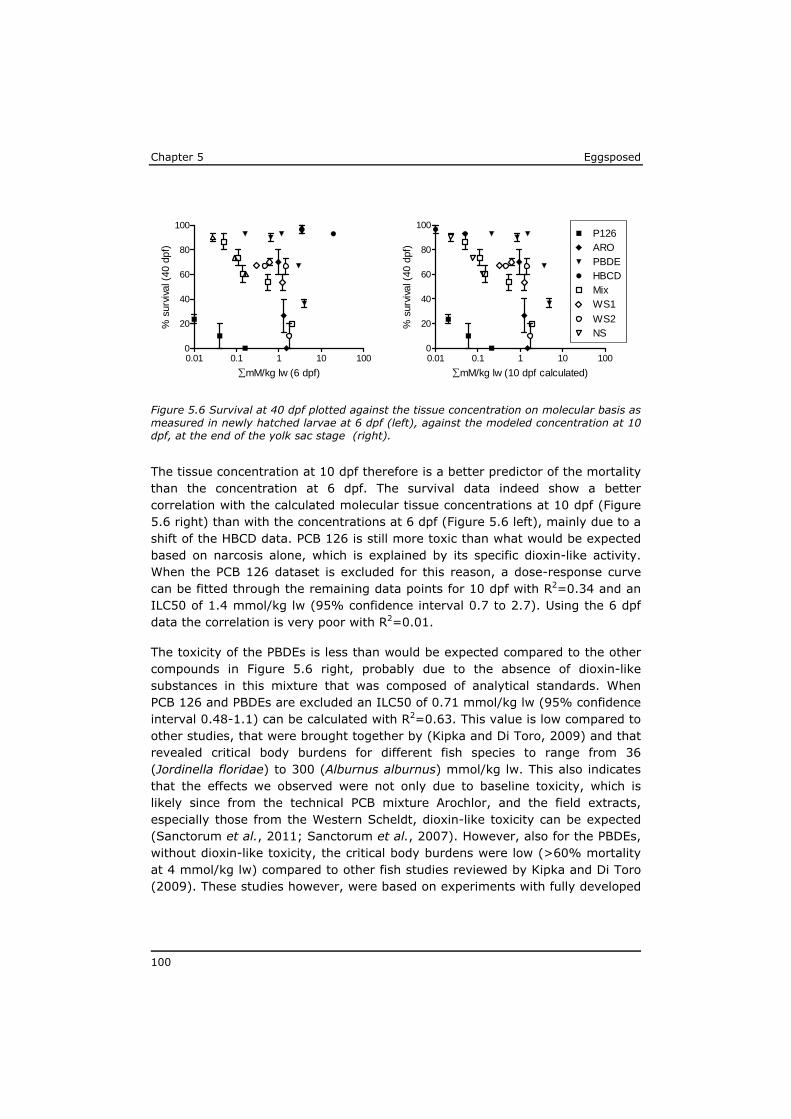
Chapter 5 Eggsposed
100
Figure 5.6 Survival at 40 dpf plotted against the tissue concentration on molecular basis as measured in newly hatched larvae at 6 dpf (left), against the modeled concentration at 10 dpf, at the end of the yolk sac stage (right).
The tissue concentration at 10 dpf therefore is a better predictor of the mortality than the concentration at 6 dpf. The survival data indeed show a better correlation with the calculated molecular tissue concentrations at 10 dpf (Figure 5.6 right) than with the concentrations at 6 dpf (Figure 5.6 left), mainly due to a shift of the HBCD data. PCB 126 is still more toxic than what would be expected based on narcosis alone, which is explained by its specific dioxin-like activity. When the PCB 126 dataset is excluded for this reason, a dose-response curve can be fitted through the remaining data points for 10 dpf with R2=0.34 and an ILC50 of 1.4 mmol/kg lw (95% confidence interval 0.7 to 2.7). Using the 6 dpf data the correlation is very poor with R2=0.01.
The toxicity of the PBDEs is less than would be expected compared to the other compounds in Figure 5.6 right, probably due to the absence of dioxin-like substances in this mixture that was composed of analytical standards. When PCB 126 and PBDEs are excluded an ILC50 of 0.71 mmol/kg lw (95% confidence interval 0.48-1.1) can be calculated with R2=0.63. This value is low compared to other studies, that were brought together by (Kipka and Di Toro, 2009) and that revealed critical body burdens for different fish species to range from 36 (Jordinella floridae) to 300 (Alburnus alburnus) mmol/kg lw. This also indicates that the effects we observed were not only due to baseline toxicity, which is likely since from the technical PCB mixture Arochlor, and the field extracts, especially those from the Western Scheldt, dioxin-like toxicity can be expected (Sanctorum et al., 2011; Sanctorum et al., 2007). However, also for the PBDEs, without dioxin-like toxicity, the critical body burdens were low (>60% mortality at 4 mmol/kg lw) compared to other fish studies reviewed by Kipka and Di Toro (2009). These studies however, were based on experiments with fully developed
mM/kg lw (6 dpf)
% s
urvi
val (
40 d
pf)
0.01 0.1 1 10 1000
20
40
60
80
100
mM/kg lw (10 dpf calculated)
% s
urvi
val (
40 d
pf)
0.01 0.1 1 10 1000
20
40
60
80
100
PBDEHBCD
ARO
MixWS1
WS2
P126
NS

Eggsposed Chapter 5
101
fish, thus without covering the critical period at the end of the yolk-sac stage as in our tests.
As baseline toxicity is the result of accumulation of substances in the biological membranes, the observed effects are actually related to the concentration of these substances in the relevant lipids in these membranes (Escher and Hermens, 2002; Escher and Schwarzenbach, 2002). Body burdens are often (also in our study) expressed on total lipids, but to relate the sensitivity of organisms the amount of membrane (polar) lipids compared to neutral lipids should be known. Neutral lipids mainly serve as energy storage pool which is being used after hatching in response of increased energy demand (Kamler, 2008). Upon lipid mobilization neutral lipids were depleted 3 times faster than polar lipids in larvae of the Senegal Sole (Solea senegalensis) (Mourente and Vazquez, 1996). Hence, at the end of the yolk-sac stage the contribution of the neutral lipids to the total lipid content reaches a minimum, and as a consequence POPs that are hardly excreted due to their high lipophilicity (Foekema et al., 2012) are concentrated in the polar lipids, including bio-membranes. This can thus be regarded as a worst case situation for the development of baseline toxicity, which might explain the low critical body burdens, expressed on total lipid basis, in our study compared to studies with fully developed fish.
Indications for effects of HBCD and PBDEs on the thyroid hormone dependent metamorphosis were not observed. Although the potency of PBDEs and HBCDs to disrupt thyroid hormone mediated metamorphosis has been shown in tests with echinoderms (Anselmo et al., 2012) and amphibians (Schriks et al., 2006). The ELS-OMEGA calculations predict that at 25 dpf, the onset of the metamorphoses, lipid normalized tissue concentrations of the POPs will be only fractions of those at 6 dpf (Figure 5.5) (ranging between 3.7% for ∑PBDEs and less than 0.01% for HBCD, Figure 5.1). This suggests that in our test, tissue concentrations were too low to affect the thyroid hormone system at the moment of metamorphoses. Apparently the relatively high concentrations between 6 and 10 dpf did not result in irreversible disruption of the thyroid hormone system, or other delayed adverse effects. In the tests reported by (Anselmo et al., 2012) and (Schriks et al., 2006) exposure continued during the metamorphoses.
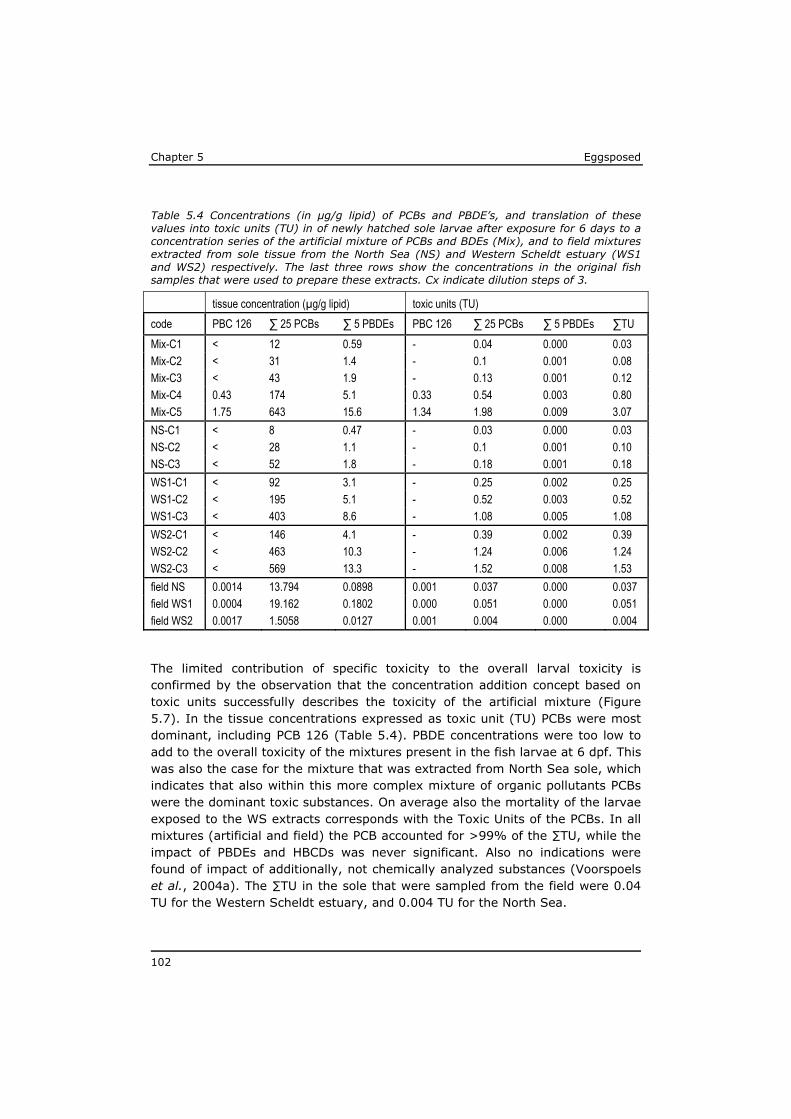
Chapter 5 Eggsposed
102
Table 5.4 Concentrations (in µg/g lipid) of PCBs and PBDE’s, and translation of these values into toxic units (TU) in of newly hatched sole larvae after exposure for 6 days to a concentration series of the artificial mixture of PCBs and BDEs (Mix), and to field mixtures extracted from sole tissue from the North Sea (NS) and Western Scheldt estuary (WS1 and WS2) respectively. The last three rows show the concentrations in the original fish samples that were used to prepare these extracts. Cx indicate dilution steps of 3.
tissue concentration (µg/g lipid) toxic units (TU)
code PBC 126 ∑ 25 PCBs ∑ 5 PBDEs PBC 126 ∑ 25 PCBs ∑ 5 PBDEs ∑TU
Mix-C1 < 12 0.59 - 0.04 0.000 0.03 Mix-C2 < 31 1.4 - 0.1 0.001 0.08 Mix-C3 < 43 1.9 - 0.13 0.001 0.12 Mix-C4 0.43 174 5.1 0.33 0.54 0.003 0.80 Mix-C5 1.75 643 15.6 1.34 1.98 0.009 3.07
NS-C1 < 8 0.47 - 0.03 0.000 0.03 NS-C2 < 28 1.1 - 0.1 0.001 0.10 NS-C3 < 52 1.8 - 0.18 0.001 0.18
WS1-C1 < 92 3.1 - 0.25 0.002 0.25 WS1-C2 < 195 5.1 - 0.52 0.003 0.52 WS1-C3 < 403 8.6 - 1.08 0.005 1.08
WS2-C1 < 146 4.1 - 0.39 0.002 0.39 WS2-C2 < 463 10.3 - 1.24 0.006 1.24 WS2-C3 < 569 13.3 - 1.52 0.008 1.53
field NS 0.0014 13.794 0.0898 0.001 0.037 0.000 0.037 field WS1 0.0004 19.162 0.1802 0.000 0.051 0.000 0.051 field WS2 0.0017 1.5058 0.0127 0.001 0.004 0.000 0.004
The limited contribution of specific toxicity to the overall larval toxicity is confirmed by the observation that the concentration addition concept based on toxic units successfully describes the toxicity of the artificial mixture (Figure 5.7). In the tissue concentrations expressed as toxic unit (TU) PCBs were most dominant, including PCB 126 (Table 5.4). PBDE concentrations were too low to add to the overall toxicity of the mixtures present in the fish larvae at 6 dpf. This was also the case for the mixture that was extracted from North Sea sole, which indicates that also within this more complex mixture of organic pollutants PCBs were the dominant toxic substances. On average also the mortality of the larvae exposed to the WS extracts corresponds with the Toxic Units of the PCBs. In all mixtures (artificial and field) the PCB accounted for >99% of the ∑TU, while the impact of PBDEs and HBCDs was never significant. Also no indications were found of impact of additionally, not chemically analyzed substances (Voorspoels et al., 2004a). The ∑TU in the sole that were sampled from the field were 0.04 TU for the Western Scheldt estuary, and 0.004 TU for the North Sea.

Eggsposed Chapter 5
103
5.4.3 Implication for wild Sole populations
Our comparison between the concentration of PCBs and PBDEs in filet and spawn of the North Sea sole confirms observations by other researchers (e.g. reviewed by Russell et al., 1999) that POPs present in the tissue of the mother fish are in general transferred to the eggs with a lipid normalized 1:1 ratio. Therefore the lipid normalized tissue concentration in the mother fish is a good descriptor of the POP concentrations in the eggs.
The Western Scheldt is considered to be one of the most contaminated estuaries of the North Sea with main pollution sources in the heavily industrialized area upstream along the river Scheldt (De Vijver et al., 2003; Voorspoels et al., 2004a; Sanctorum et al., 2011; Sanctorum et al., 2007). When the concentrations of PCBs, PBDEs and HBCD measured in the sole from the Western Scheldt estuary are translated into ∑TU levels for sole early life development, these do not exceed 0.05 (Table 5.4). In order to cause 50% mortality of the developing off-spring, the ∑TU needs to be 20 times higher (Figure 5.7). In the sole sampled from the North Sea the ∑TU was one order of a magnitude lower (Table 5.4). Higher PCB levels in sole tissue have been reported from the Mediterranean with a maximum concentration of 85 µg/g lw (∑31 PCBs) (Dierking et al., 2009). This translates to ∑TU=0.23, and is comparable to the concentration in larvae from treatment WS1, where 33% mortality was observed.
For several reasons it can be argued that our estimated effect concentrations underestimate the actual risk. Our experiments started with clean healthy eggs from parental fish that were not affected by toxic compounds, and exposure to the test substances commenced about 12 hours after spawning and fertilization. As a consequence the first 12 hours of the development of the embryo occurred under non-exposed conditions, and once the exposure started the internal concentration can be expected to have built up gradually. In a natural situation where maternal transfer is involved, potentially harmful substances will be present in the egg from the beginning. In our experiments only lipophilic POPs were ‘transferred’ to the eggs. In nature also metabolites of these POPs (Montano et al., 2013) will be deposited in the eggs, together with other substances that are known to be able to interfere with early life development, as for instance methylmercury (Latif et al., 2010) or tributyltinoxide (Nakayama et al., 2005).
Thus, although our data indicate that in a relatively contaminated estuary like the Western Scheldt the direct impact of maternally transferred POPs on early life development is limited, impact of the POPs on the whole reproduction process cannot be ruled out.

Chapter 5 Eggsposed
104
Figure 5.7 Larval mortality at the end of the tests with the artificial mixture (Mix) and the field mixtures (NS, WS1 and WS2) plotted against the tissue concentrations of PCBs, PBDEs and HBCDs on dpf 6 expressed as total toxic units. The crossing of the dotted lines indicates the point where theoretically 1 Toxic unit should result in 50% larval mortality. ‘NS’and ‘WS’ on the x-axis indicate the TU concentration that was present in the sole sampled from the North Sea and Western Scheldt estuary respectively.
5.5 References
Backhaus, T., Faust, M., 2012. Predictive Environmental Risk Assessment of Chemical Mixtures: A Conceptual Framework. Environ. Sci. Technol. 46, 2564-2573.
Baeyens, W., Leermakers, M., Papina, T., Saprykin, A., Brion, N., Noyen, J., De Gieter, M., Elskens, M., Goeyens, L., 2003. Bioconcentration and biomagnification of mercury and methylmercury in North Sea and Scheldt estuary fish. Archives of Environmental Contamination and Toxicology 45, 498-508.
Brouwer, A., Longnecker, M.P., Birnbaum, L.S., Cogliano, J., Kostyniak, P., Moore, J., Schantz, S., Winneke, G., 1999. Characterization of potential endocrine-related health effects at low-dose levels of exposure to PCBs. Environmental Health Perspectives 107, 639-649.
de Boer, J., Dao, Q.T., van Leeuwen, S.P.J., Kotterman, M.J.J., Schobben, J.H.M., 2010. Thirty year monitoring of PCBs, organochlorine pesticides and tetrabromodiphenylether in eel from The Netherlands. Environmental Pollution 158, 1228-1236.
De Vijver, K.I.V., Hoff, P.T., Van Dongen, W., Esmans, E.L., Blust, R., De Coen, W.M., 2003. Exposure patterns of perfluorooctane sulfonate in aquatic invertebrates from the Western Scheldt estuary and the southern North Sea. Environmental Toxicology and Chemistry 22, 2037-2041.
Dierking, J., Wafo, E., Schembri, T., Lagadec, V., Nicolas, C., Letourneur, Y., Harmelin-Vivien, M., 2009. Spatial patterns in PCBs, pesticides, mercury and cadmium in the common sole in the NW Mediterranean Sea, and a novel use of contaminants as biomarkers. Marine Pollution Bulletin 58, 1605-1614.
Toxic Units (dpf 6)
% m
orta
lity
0.001 0.01 0.1 1 10
0
25
50
75
100
WS1WS2
MixNS
WSNS

Eggsposed Chapter 5
105
Escher, B.I., Hermens, J.L.M., 2002. Modes of action in ecotoxicology: Their role in body burdens, species sensitivity, QSARs, and mixture effects. Environ. Sci. Technol. 36, 4201-4217.
Escher, B.I., Schwarzenbach, R.P., 2002. Mechanistic studies on baseline toxicity and uncoupling of organic compounds as a basis for modeling effective membrane concentrations in aquatic organisms. Aquatic Sciences 64, 20-35.
Foekema, E.M., Deerenberg, C.M., Murk, A.J., 2008. Prolonged ELS test with the marine flatfish sole (Solea solea) shows delayed toxic effects of previous exposure to PCB 126. Aquatic Toxicology 90, 197-203.
Foekema, E.M., Fischer, A., Parron, M.L., Kwadijk, C., de Vries, P., Murk, A.J., 2012. Toxic concentrations in fish early life stages peak at a critical moment. Environmental Toxicology and Chemistry 31, 1381-1390.
Gonzalez-Doncel, M., Larrea, M., Sanchez-Fortun, S., Hinton, D.E., 2003. Influence of water hardening of the chorion on cadmium accumulation in medaka (Oryzias latipes) eggs. Chemosphere 52, 75-83.
Gutleb, A.C., Appelman, J., Bronkhorst, M.C., van den Berg, J.H.J., Spenkelink, A., Brouwer, A., Murk, A.J., 1999. Delayed effects of pre- and early-life time exposure to polychlorinated biphenyls on tadpoles of two amphibian species (Xenopus laevis and Rana temporaria). Environmental Toxicology and Pharmacology 8, 1-14.
Hansen, B.G., Paya-Perez, A.B., Rahman, M., Larsen, B.R., 1999. Qsars for K-ow and K-oc of PCB congeners: A critical examination of data, assumptions and statistical approaches. Chemosphere 39, 2209-2228.
Ishibashi, H., Hirano, M., Matsumura, N., Watanabe, N., Takao, Y., Arizono, K., 2006. Reproductive effects and bioconcentration of 4-nonylphenol in medaka fish (Oryzias latipes). Chemosphere 65, 1019-1026.
Janak, K., Covaci, A., Voorspoels, S., Becher, G., 2005. Hexabromocyclododecane in marine species from the Western Scheldt Estuary: Diastereoisomer- and enantiomer-specific accumulation. Environ. Sci. Technol. 39, 1987-1994.
Kamler, E., 2008. Resource allocation in yolk-feeding fish. Reviews in Fish Biology and Fisheries 18, 143-200.
Kipka, U., Di Toro, D.M., 2009. Technical basis for polar and non-polar narcotic chemicals and polycyclic aromatic hydrocarbon criteria; III a polyparameter model for target lipid partitioning. Environmental Toxicology and Chemistry 28, 1429-1438.
Klaren, P.H.M., Wunderink, Y.S., Yufera, M., Mancera, J.M., Flik, G., 2008. The thyroid gland and thyroid hormones in Senegalese sole (Solea senegalensis) during early development and metamorphosis. General and Comparative Endocrinology 155, 686-694.
Kuiper, R.V., Canton, R.F., Leonards, P.E.G., Jenssen, B.M., Dubbeldam, M., Wester, P.W., van den Berg, M., Vos, J.G., Vethaak, A.D., 2007. Long-term exposure of European flounder (Platichthys flesus) to the flame-retardants tetrabromobisphenol A (TBBPA) and hexabromocyclododecane (HBCD). Ecotoxicology and Environmental Safety 67, 349-360.

Chapter 5 Eggsposed
106
Latif, M.A., Bodaly, R.A., Johnston, T.A., Fudge, R.J.P., 2001. Effects of environmental and maternally derived methylmercury on the embryonic and larval stages of walleye (Stizostedion vitreum). Environmental Pollution 111, 139-148.
Li, L.N., Xie, S.D., Cai, H., Bai, X.T., Xue, Z., 2008. Quantitative structure-property relationships for octanol-water partition coefficients of polybrominated diphenyl ethers. Chemosphere 72, 1602-1606.
Maes, J., Belpaire, C., Goemans, G., 2008. Spatial variations and temporal trends between 1994 and 2005 in polychlorinated biphenyls, organochlorine pesticides and heavy metals in European eel (Anguilla anguilla L.) in Flanders, Belgium. Environmental Pollution 153, 223-237.
Martinez, G.M., Bolker, J.A., 2003. Embryonic and larval staging of summer flounder (Paralichthys dentatus). Journal of Morphology 255, 162-176.
Miller, M.A., 1993. Maternal transfer of organochlorine compounds in salmonines to their eggs. Canadian Journal of Fisheries and Aquatic Sciences 50, 1405-1413.
Montano, M., Gutleb, A.C., murk, A.J., 2013 in press. Persistent Toxic Burdens of Halogenated Phenolic Compounds in Humans and Wildlife. Environ. Sci. Technol.
Monteverdi, G.H., Di Giulio, R.T., 2000. Oocytic accumulation and tissue distribution of 2,3,7,8-tetrachlorodibenzo-p-dioxin and benzo a pyrene in gravid Fundulus heteroclitus. Environmental Toxicology and Chemistry 19, 2512-2518.
Mourente, G., Vazquez, R., 1996. Changes in the content of total lipid, lipid classes and their fatty acids of developing eggs and unfed larvae of the Senegal sole, Solen senegalensis Kaup. Fish Physiology and Biochemistry 15, 221-235.
Murk, A.J., Leonards, P.E.G., van Hattum, B., Luit, R., van der Weiden, M.E.J., Smit, M., 1998. Application of biomarkers for exposure and effect of polyhalogenated aromatic hydrocarbons in naturally exposed European otters (Lutra lutra). Environmental Toxicology and Pharmacology 6, 91-102.
Nakayama, K., Oshima, Y., Nagafuchi, K., Hano, T., Shimasaki, Y., Nakayamaj, K., 2005. Early-life-stage toxicity in offspring from exposed parent medaka, Oryzias latipes, to mixtures of tributyltin and polychlorinated biphenyls. Environmental Toxicology and Chemistry 24, 591-596.
Noyes, P.D., Hinton, D.E., Stapleton, H.M., 2011. Accumulation and Debromination of Decabromodiphenyl Ether (BDE-209) in Juvenile Fathead Minnows (Pimephales promelas) Induces Thyroid Disruption and Liver Alterations. Toxicological Sciences 122, 265-274.
Nyholm, J.R., Norman, A., Norrgren, L., Haglund, P., Andersson, P.L., 2008. Maternal transfer of brominated flame retardants in zebrafish (Danio rerio). Chemosphere 73, 203-208.
Olsson, P.E., Westerlund, L., Teh, S.J., Billsson, K., Berg, A.H., Tysklind, M., Nilsson, J., Eriksson, L.O., Hinton, D.E., 1999. Effects of maternal exposure to estrogen and PCB on different life stages of zebrafish (Danio rerio). Ambio 28, 100-106.
Palace, V., Park, B., Pleskach, K., Gemmill, B., Tomy, G., 2010. Altered thyroxine metabolism in rainbow trout (Oncorhynchus mykiss) exposed to hexabromocyclododecane (HBCD). Chemosphere 80, 165-169.

Eggsposed Chapter 5
107
Qin, W.C., Su, L.M., Zhang, X.J., Qin, H.W., Wen, Y., Guo, Z., Sun, F.T., Sheng, L.X., Zhao, Y.H., Abraham, M.H., 2010. Toxicity of organic pollutants to seven aquatic organisms: effect of polarity and ionization. SAR QSAR Environ. Res. 21, 389-401.
Rijnsdorp, A.D., Vanbeek, F.A., Flatman, S., Millner, R.M., Riley, J.D., Giret, M., Declerck, R., 1992. RECRUITMENT OF SOLE STOCKS, SOLEA-SOLEA (L), IN THE NORTHEAST ATLANTIC (NETHERLANDS JOURNAL OF SEA RESEARCH, VOL 29, PG 173, 1992). Netherlands Journal of Sea Research 29, U402-U402.
Roosens, L., Dirtu, A.C., Goemans, G., Belpaire, C., Gheorghe, A., Neels, H., Blust, R., Covaci, A., 2008. Brominated flame retardants and polychlorinated biphenyls in fish from the river Scheldt, Belgium. Environment International 34, 976-983.
Russell, R.W., Gobas, F., Haffner, G.D., 1999. Maternal transfer and in ovo exposure of organochlorines in oviparous organisms: A model and field verification. Environ. Sci. Technol. 33, 416-420.
Sanctorum, H., Elskens, M., Leermakers, M., Gao, Y., Charriau, A., Billon, G., Goscinny, S., De Cooman, W., Baeyens, W., 2011. Sources of PCDD/Fs, non-ortho PCBs and PAHs in sediments of high and low impacted transboundary rivers (Belgium-France). Chemosphere 85, 203-209.
Sanctorum, H., Windal, I., Hanot, V., Goeyens, L., Baeyens, W., 2007. Dioxin and dioxin-like activity in sediments of the belgian coastal area (Southern north sea). Archives of Environmental Contamination and Toxicology 52, 317-325.
Schriks, M., Zvinavashe, E., Furlow, J.D., Murk, A.J., 2006. Disruption of thyroid hormone-mediated Xenopus laevis tadpole tail tip regression by hexabromocyclododecane (HBCD) and 2,2 ',3,3 ',4,4 ',5,5 ',6-nona brominated diphenyl ether (BDE206). Chemosphere 65, 1904-1908.
Serrano, R., Blanes, M.A., Lopez, F.J., 2008. Maternal transfer of organochlorine compounds to oocytes in wild and farmed gilthead sea bream (Sparus aurata). Chemosphere 70, 561-566.
Tietge, J.E., Johnson, R.D., Jensen, K.M., Cook, P.M., Elonen, G.E., Fernandez, J.D., Holcombe, G.W., Lothenbach, D.B., Nichols, J.W., 1998. Reproductive toxicity and disposition of 2,3,7,8-tetrachlorodibenzo-p-dioxin in adult brook trout (Salvelinus fontinalis) following a dietary exposure. Environmental Toxicology and Chemistry 17, 2395-2407.
Van den Berg, M., Birnbaum, L.S., Denison, M., De Vito, M., Farland, W., Feeley, M., Fiedler, H., Hakansson, H., Hanberg, A., Haws, L., Rose, M., Safe, S., Schrenk, D., Tohyama, C., Tritscher, A., Tuomisto, J., Tysklind, M., Walker, N., Peterson, R.E., 2006. The 2005 World Health Organization reevaluation of human and mammalian toxic equivalency factors for dioxins and dioxin-like compounds. Toxicological Sciences 93, 223-241.
vanWezel, A.P., deVries, D.A.M., Sijm, D., Opperhuizen, A., 1996. Use of the lethal body burden in the evaluation of mixture toxicity. Ecotoxicology and Environmental Safety 35, 236-241.
Villalobos, S.A., Hamm, J.T., Teh, S.J., Hinton, D.E., 2000. Thiobencarb-induced embryotoxicity in medaka (Oryzias latipes): stage-specific toxicity and the protective role of chorion. Aquatic Toxicology 48, 309-326.

Chapter 5 Eggsposed
108
Voorspoels, S., Covaci, A., Maervoet, J., De Meester, I., Schepens, P., 2004a. Levels and profiles of PCBs and OCPs in marine benthic species from the Belgian North Sea and the Western Scheldt Estuary. Marine Pollution Bulletin 49, 393-404.
Voorspoels, S., Covaci, A., Maervoet, J., Schepens, P., 2004b. PBDEs in marine and freshwater sediments from Belgium: levels, profiles and relations with biota. Journal of Environmental Monitoring 6, 914-918.
Voorspoels, S., Covaci, A., Schepens, P., 2003. Polybrominated diphenyl ethers in marine species from the Belgian North Sea and the Western Scheidt Estuary: Levels, profiles, and distribution. Environ. Sci. Technol. 37, 4348-4357.
Yu, L.Q., Lam, J.C.W., Guo, Y.Y., Wu, R.S.S., Lam, P.K.S., Zhou, B.S., 2011. Parental Transfer of Polybrominated Diphenyl Ethers (PBDEs) and Thyroid Endocrine Disruption in Zebrafish. Environ. Sci. Technol. 45, 10652-10659.
Zimmer, K.E., Montano, M., Olsaker, I., Dahl, E., Berg, V., Karlsson, C., Murk, A.J., Skaare, J.U., Ropstad, E., Verhaegen, S., 2011. In vitro steroidogenic effects of mixtures of persistent organic pollutants (POPs) extracted from burbot (Lota lota) caught in two Norwegian lakes. Sci. Total Environ. 409, 2040-2048.
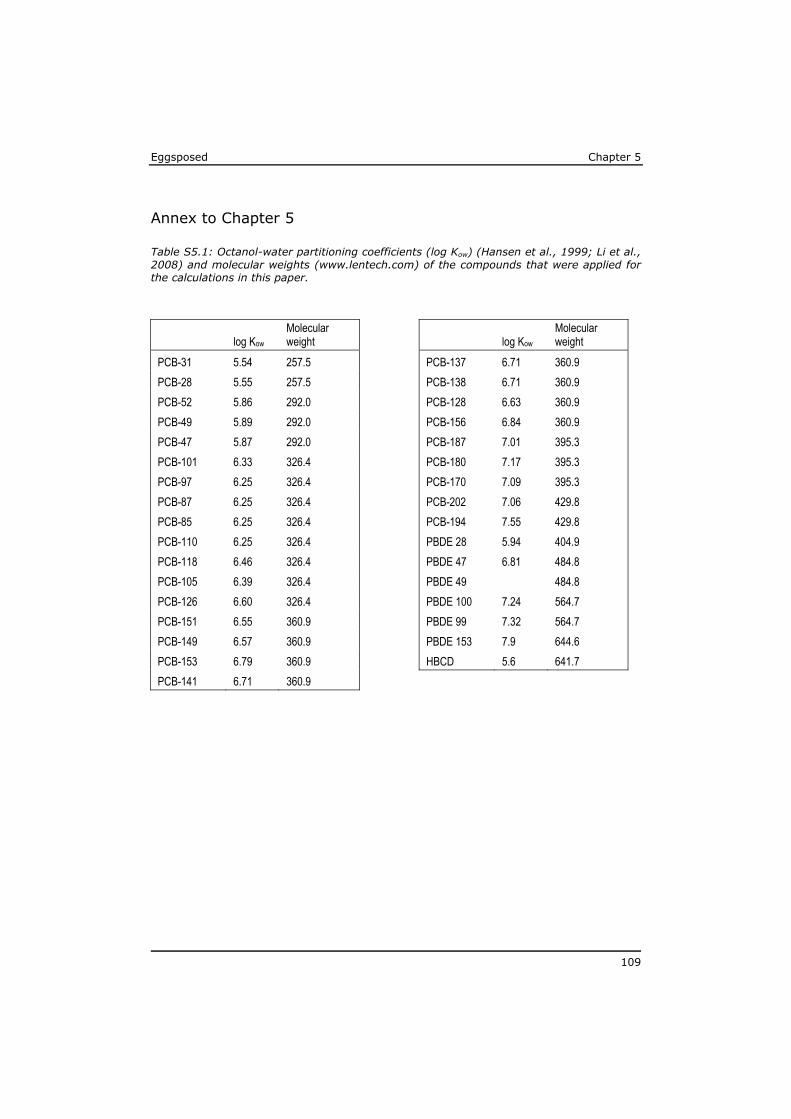
Eggsposed Chapter 5
109
Annex to Chapter 5
Table S5.1: Octanol-water partitioning coefficients (log Kow) (Hansen et al., 1999; Li et al., 2008) and molecular weights (www.lentech.com) of the compounds that were applied for the calculations in this paper.
log Kow Molecular weight
PCB-31 5.54 257.5
PCB-28 5.55 257.5
PCB-52 5.86 292.0
PCB-49 5.89 292.0
PCB-47 5.87 292.0
PCB-101 6.33 326.4
PCB-97 6.25 326.4
PCB-87 6.25 326.4
PCB-85 6.25 326.4
PCB-110 6.25 326.4
PCB-118 6.46 326.4
PCB-105 6.39 326.4
PCB-126 6.60 326.4
PCB-151 6.55 360.9
PCB-149 6.57 360.9
PCB-153 6.79 360.9
PCB-141 6.71 360.9
log Kow Molecular weight
PCB-137 6.71 360.9
PCB-138 6.71 360.9
PCB-128 6.63 360.9
PCB-156 6.84 360.9
PCB-187 7.01 395.3
PCB-180 7.17 395.3
PCB-170 7.09 395.3
PCB-202 7.06 429.8
PCB-194 7.55 429.8
PBDE 28 5.94 404.9
PBDE 47 6.81 484.8
PBDE 49 484.8
PBDE 100 7.24 564.7
PBDE 99 7.32 564.7
PBDE 153 7.9 644.6
HBCD 5.6 641.7
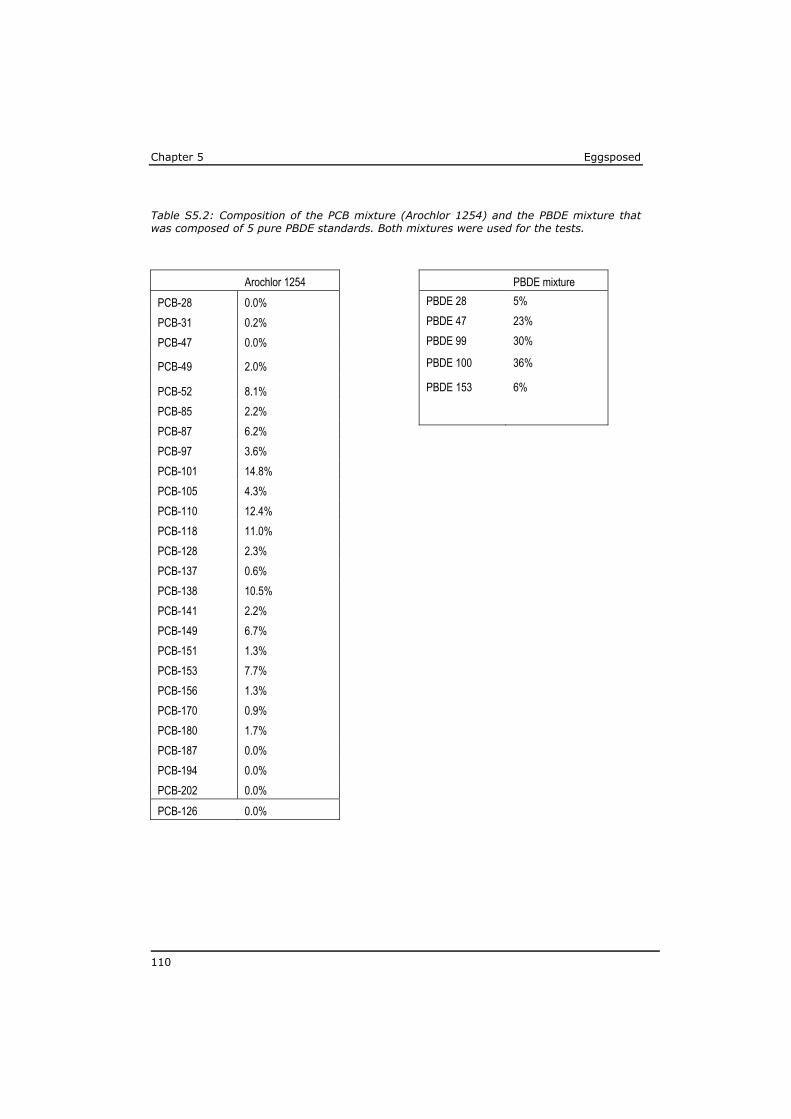
Chapter 5 Eggsposed
110
Table S5.2: Composition of the PCB mixture (Arochlor 1254) and the PBDE mixture that was composed of 5 pure PBDE standards. Both mixtures were used for the tests.
Arochlor 1254
PCB-28 0.0%
PCB-31 0.2%
PCB-47 0.0%
PCB-49 2.0%
PCB-52 8.1%
PCB-85 2.2%
PCB-87 6.2%
PCB-97 3.6%
PCB-101 14.8%
PCB-105 4.3%
PCB-110 12.4%
PCB-118 11.0%
PCB-128 2.3%
PCB-137 0.6%
PCB-138 10.5%
PCB-141 2.2%
PCB-149 6.7%
PCB-151 1.3%
PCB-153 7.7%
PCB-156 1.3%
PCB-170 0.9%
PCB-180 1.7%
PCB-187 0.0%
PCB-194 0.0%
PCB-202 0.0%
PCB-126 0.0%
PBDE mixture
PBDE 28 5%
PBDE 47 23%
PBDE 99 30%
PBDE 100 36%
PBDE 153 6%
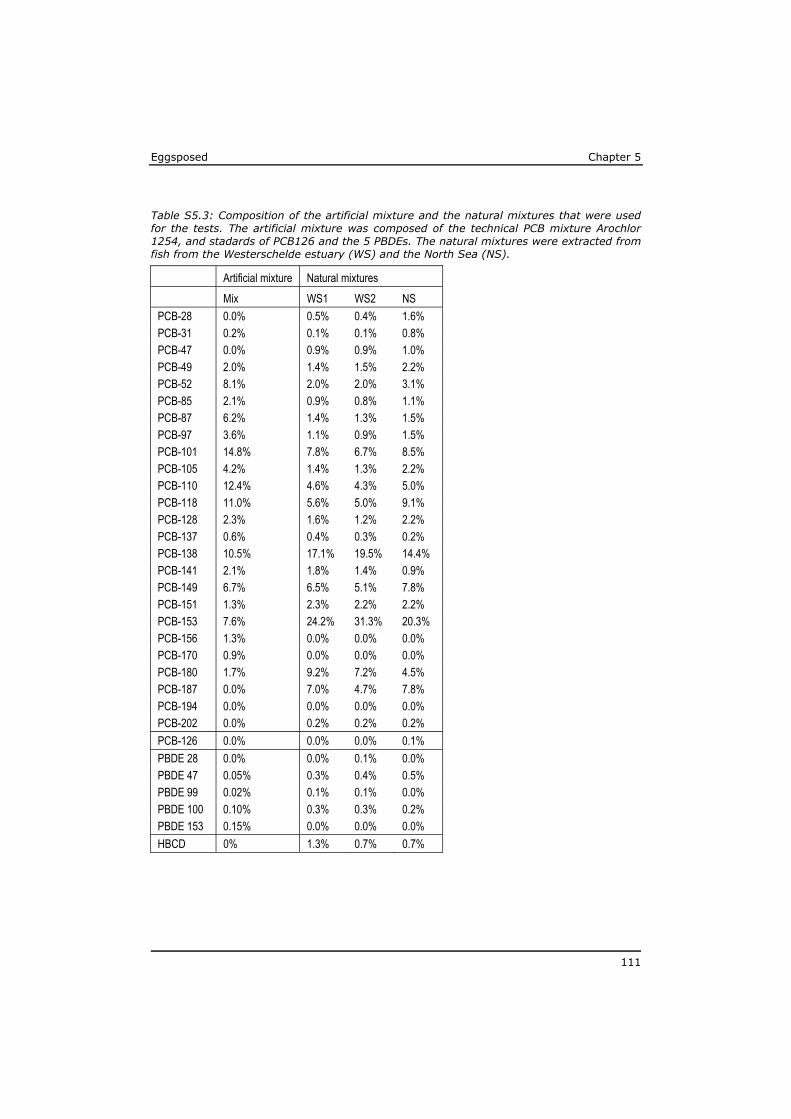
Eggsposed Chapter 5
111
Table S5.3: Composition of the artificial mixture and the natural mixtures that were used for the tests. The artificial mixture was composed of the technical PCB mixture Arochlor 1254, and stadards of PCB126 and the 5 PBDEs. The natural mixtures were extracted from fish from the Westerschelde estuary (WS) and the North Sea (NS).
Artificial mixture Natural mixtures
Mix WS1 WS2 NS
PCB-28 0.0% 0.5% 0.4% 1.6% PCB-31 0.2% 0.1% 0.1% 0.8% PCB-47 0.0% 0.9% 0.9% 1.0% PCB-49 2.0% 1.4% 1.5% 2.2% PCB-52 8.1% 2.0% 2.0% 3.1% PCB-85 2.1% 0.9% 0.8% 1.1% PCB-87 6.2% 1.4% 1.3% 1.5% PCB-97 3.6% 1.1% 0.9% 1.5% PCB-101 14.8% 7.8% 6.7% 8.5% PCB-105 4.2% 1.4% 1.3% 2.2% PCB-110 12.4% 4.6% 4.3% 5.0% PCB-118 11.0% 5.6% 5.0% 9.1% PCB-128 2.3% 1.6% 1.2% 2.2% PCB-137 0.6% 0.4% 0.3% 0.2% PCB-138 10.5% 17.1% 19.5% 14.4% PCB-141 2.1% 1.8% 1.4% 0.9% PCB-149 6.7% 6.5% 5.1% 7.8% PCB-151 1.3% 2.3% 2.2% 2.2% PCB-153 7.6% 24.2% 31.3% 20.3% PCB-156 1.3% 0.0% 0.0% 0.0% PCB-170 0.9% 0.0% 0.0% 0.0% PCB-180 1.7% 9.2% 7.2% 4.5% PCB-187 0.0% 7.0% 4.7% 7.8% PCB-194 0.0% 0.0% 0.0% 0.0% PCB-202 0.0% 0.2% 0.2% 0.2%
PCB-126 0.0% 0.0% 0.0% 0.1%
PBDE 28 0.0% 0.0% 0.1% 0.0% PBDE 47 0.05% 0.3% 0.4% 0.5% PBDE 99 0.02% 0.1% 0.1% 0.0% PBDE 100 0.10% 0.3% 0.3% 0.2% PBDE 153 0.15% 0.0% 0.0% 0.0%
HBCD 0% 1.3% 0.7% 0.7%
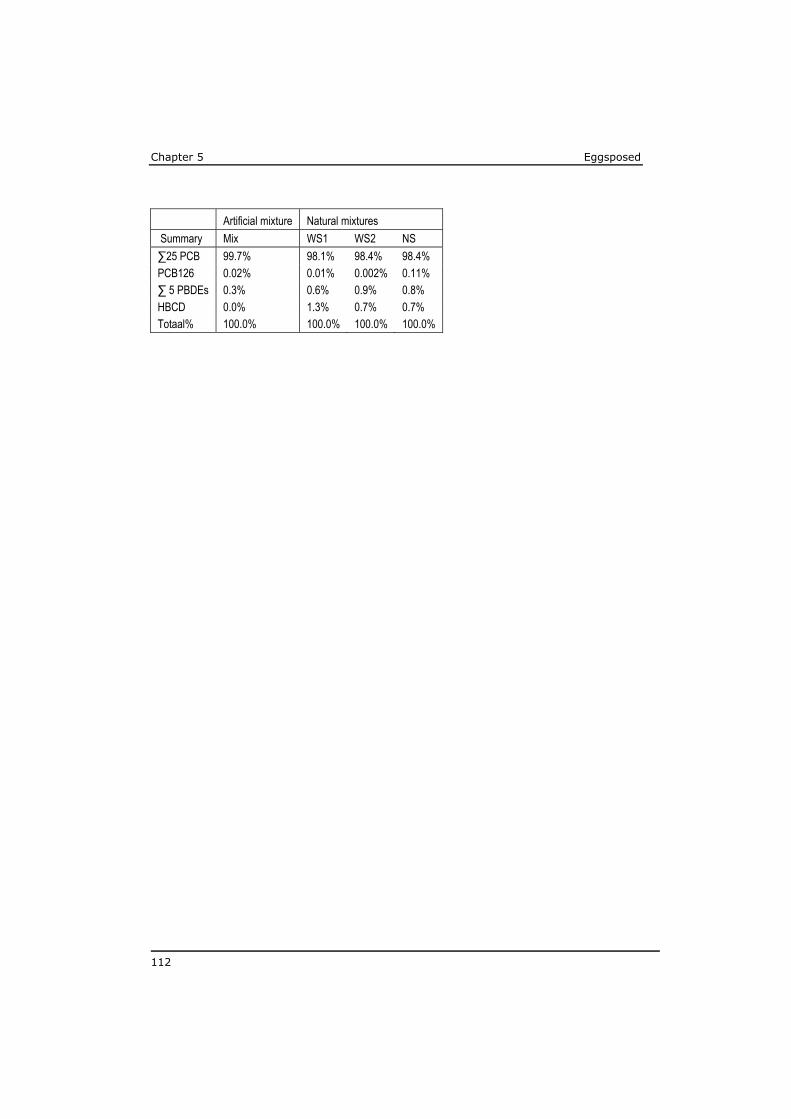
Chapter 5 Eggsposed
112
Artificial mixture Natural mixtures
Summary Mix WS1 WS2 NS
∑25 PCB 99.7% 98.1% 98.4% 98.4% PCB126 0.02% 0.01% 0.002% 0.11% ∑ 5 PBDEs 0.3% 0.6% 0.9% 0.8% HBCD 0.0% 1.3% 0.7% 0.7% Totaal% 100.0% 100.0% 100.0% 100.0%
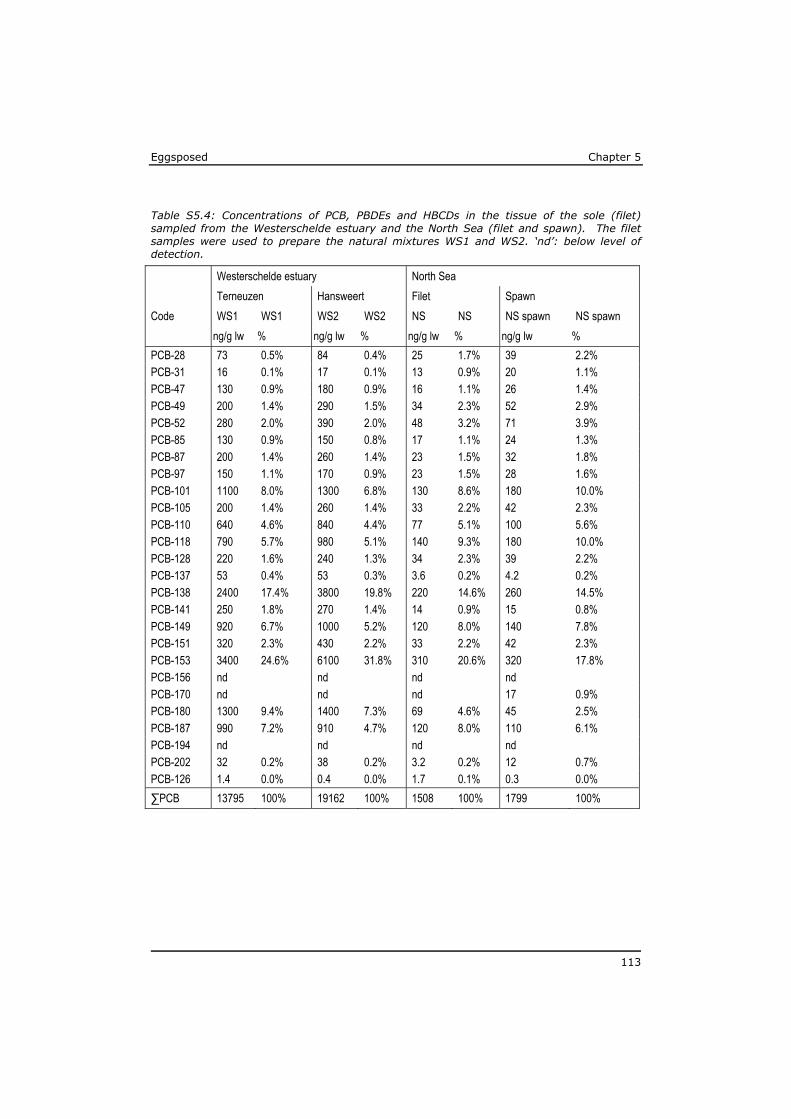
Eggsposed Chapter 5
113
Table S5.4: Concentrations of PCB, PBDEs and HBCDs in the tissue of the sole (filet) sampled from the Westerschelde estuary and the North Sea (filet and spawn). The filet samples were used to prepare the natural mixtures WS1 and WS2. ‘nd’: below level of detection.
Westerschelde estuary North Sea
Terneuzen Hansweert Filet Spawn
Code WS1 WS1 WS2 WS2 NS NS NS spawn NS spawn
ng/g lw % ng/g lw % ng/g lw % ng/g lw %
PCB-28 73 0.5% 84 0.4% 25 1.7% 39 2.2% PCB-31 16 0.1% 17 0.1% 13 0.9% 20 1.1% PCB-47 130 0.9% 180 0.9% 16 1.1% 26 1.4% PCB-49 200 1.4% 290 1.5% 34 2.3% 52 2.9% PCB-52 280 2.0% 390 2.0% 48 3.2% 71 3.9% PCB-85 130 0.9% 150 0.8% 17 1.1% 24 1.3% PCB-87 200 1.4% 260 1.4% 23 1.5% 32 1.8% PCB-97 150 1.1% 170 0.9% 23 1.5% 28 1.6% PCB-101 1100 8.0% 1300 6.8% 130 8.6% 180 10.0% PCB-105 200 1.4% 260 1.4% 33 2.2% 42 2.3% PCB-110 640 4.6% 840 4.4% 77 5.1% 100 5.6% PCB-118 790 5.7% 980 5.1% 140 9.3% 180 10.0% PCB-128 220 1.6% 240 1.3% 34 2.3% 39 2.2% PCB-137 53 0.4% 53 0.3% 3.6 0.2% 4.2 0.2% PCB-138 2400 17.4% 3800 19.8% 220 14.6% 260 14.5% PCB-141 250 1.8% 270 1.4% 14 0.9% 15 0.8% PCB-149 920 6.7% 1000 5.2% 120 8.0% 140 7.8% PCB-151 320 2.3% 430 2.2% 33 2.2% 42 2.3% PCB-153 3400 24.6% 6100 31.8% 310 20.6% 320 17.8% PCB-156 nd
nd
nd
nd
PCB-170 nd
nd
nd
17 0.9%
PCB-180 1300 9.4% 1400 7.3% 69 4.6% 45 2.5% PCB-187 990 7.2% 910 4.7% 120 8.0% 110 6.1% PCB-194 nd
nd
nd
nd
PCB-202 32 0.2% 38 0.2% 3.2 0.2% 12 0.7% PCB-126 1.4 0.0% 0.4 0.0% 1.7 0.1% 0.3 0.0%
∑PCB 13795 100% 19162 100% 1508 100% 1799 100%
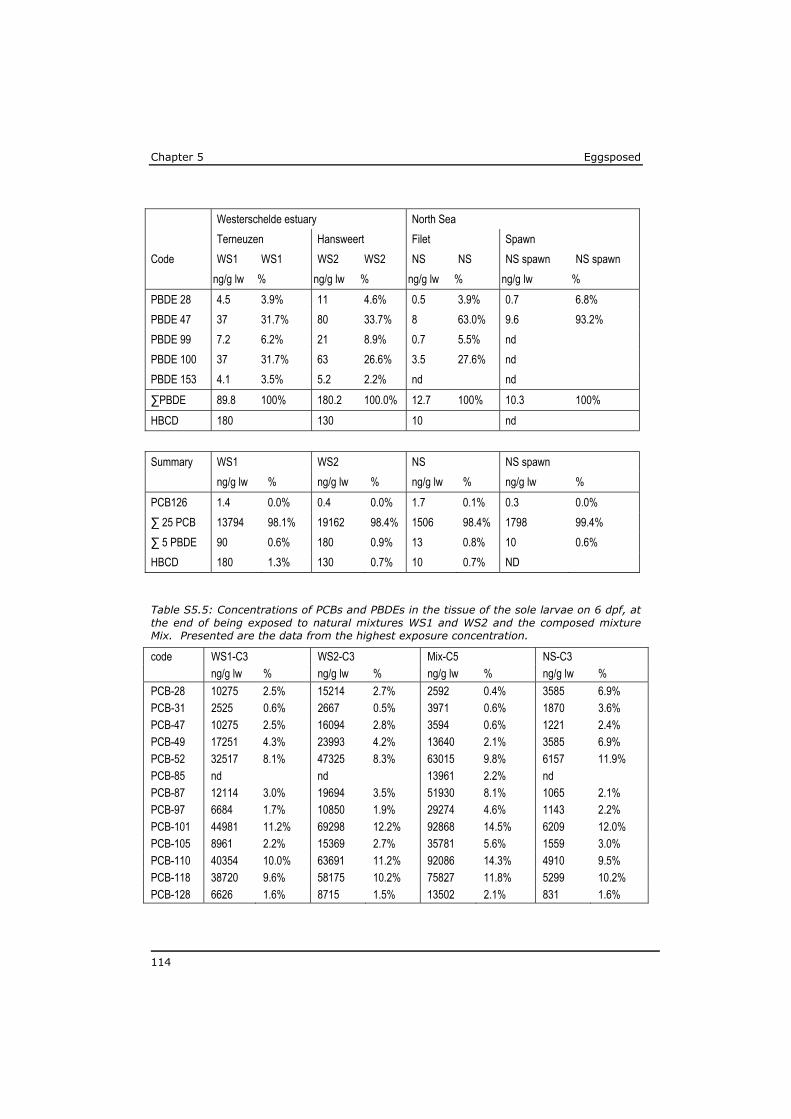
Chapter 5 Eggsposed
114
Westerschelde estuary North Sea
Terneuzen Hansweert Filet Spawn
Code WS1 WS1 WS2 WS2 NS NS NS spawn NS spawn
ng/g lw % ng/g lw % ng/g lw % ng/g lw %
PBDE 28 4.5 3.9% 11 4.6% 0.5 3.9% 0.7 6.8%
PBDE 47 37 31.7% 80 33.7% 8 63.0% 9.6 93.2%
PBDE 99 7.2 6.2% 21 8.9% 0.7 5.5% nd
PBDE 100 37 31.7% 63 26.6% 3.5 27.6% nd
PBDE 153 4.1 3.5% 5.2 2.2% nd
nd
∑PBDE 89.8 100% 180.2 100.0% 12.7 100% 10.3 100%
HBCD 180
130
10
nd
Summary WS1 WS2 NS NS spawn
ng/g lw % ng/g lw % ng/g lw % ng/g lw %
PCB126 1.4 0.0% 0.4 0.0% 1.7 0.1% 0.3 0.0%
∑ 25 PCB 13794 98.1% 19162 98.4% 1506 98.4% 1798 99.4%
∑ 5 PBDE 90 0.6% 180 0.9% 13 0.8% 10 0.6%
HBCD 180 1.3% 130 0.7% 10 0.7% ND
Table S5.5: Concentrations of PCBs and PBDEs in the tissue of the sole larvae on 6 dpf, at the end of being exposed to natural mixtures WS1 and WS2 and the composed mixture Mix. Presented are the data from the highest exposure concentration.
code WS1-C3 WS2-C3 Mix-C5 NS-C3 ng/g lw % ng/g lw % ng/g lw % ng/g lw %
PCB-28 10275 2.5% 15214 2.7% 2592 0.4% 3585 6.9% PCB-31 2525 0.6% 2667 0.5% 3971 0.6% 1870 3.6% PCB-47 10275 2.5% 16094 2.8% 3594 0.6% 1221 2.4% PCB-49 17251 4.3% 23993 4.2% 13640 2.1% 3585 6.9% PCB-52 32517 8.1% 47325 8.3% 63015 9.8% 6157 11.9% PCB-85 nd nd 13961 2.2% nd PCB-87 12114 3.0% 19694 3.5% 51930 8.1% 1065 2.1% PCB-97 6684 1.7% 10850 1.9% 29274 4.6% 1143 2.2% PCB-101 44981 11.2% 69298 12.2% 92868 14.5% 6209 12.0% PCB-105 8961 2.2% 15369 2.7% 35781 5.6% 1559 3.0% PCB-110 40354 10.0% 63691 11.2% 92086 14.3% 4910 9.5% PCB-118 38720 9.6% 58175 10.2% 75827 11.8% 5299 10.2% PCB-128 6626 1.6% 8715 1.5% 13502 2.1% 831 1.6%
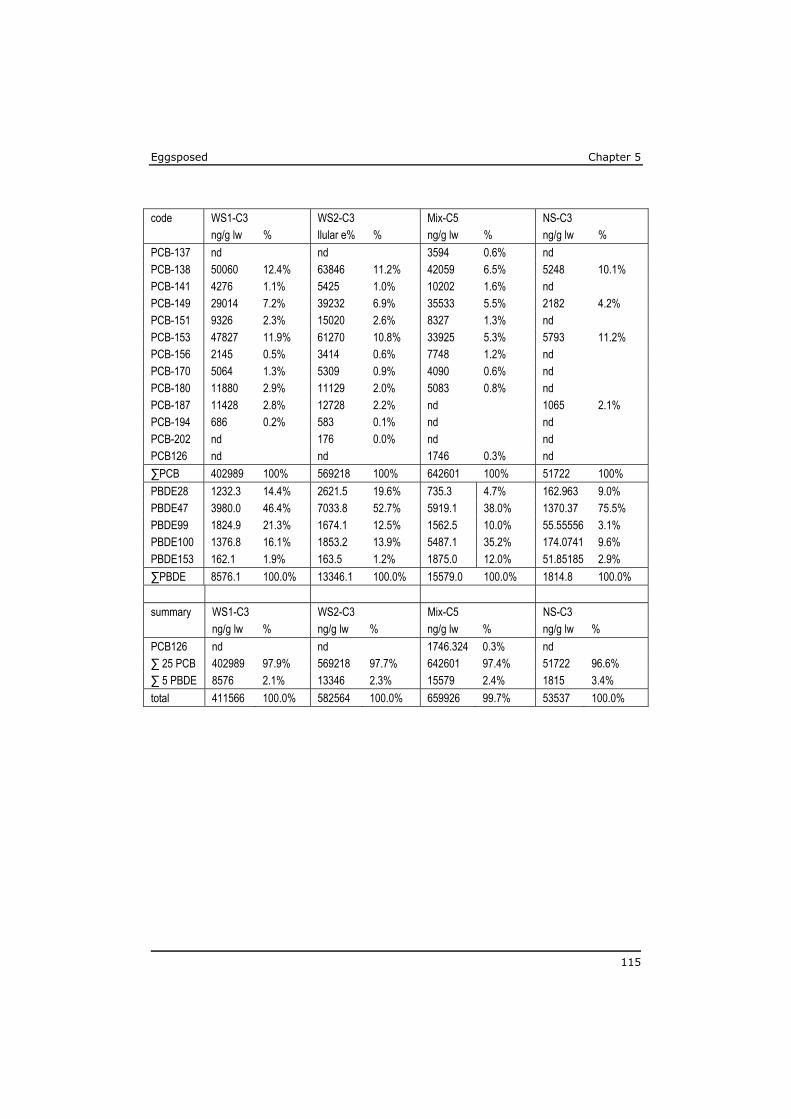
Eggsposed Chapter 5
115
code WS1-C3 WS2-C3 Mix-C5 NS-C3 ng/g lw % llular e% % ng/g lw % ng/g lw %
PCB-137 nd nd 3594 0.6% nd PCB-138 50060 12.4% 63846 11.2% 42059 6.5% 5248 10.1% PCB-141 4276 1.1% 5425 1.0% 10202 1.6% nd PCB-149 29014 7.2% 39232 6.9% 35533 5.5% 2182 4.2% PCB-151 9326 2.3% 15020 2.6% 8327 1.3% nd PCB-153 47827 11.9% 61270 10.8% 33925 5.3% 5793 11.2% PCB-156 2145 0.5% 3414 0.6% 7748 1.2% nd PCB-170 5064 1.3% 5309 0.9% 4090 0.6% nd PCB-180 11880 2.9% 11129 2.0% 5083 0.8% nd PCB-187 11428 2.8% 12728 2.2% nd 1065 2.1% PCB-194 686 0.2% 583 0.1% nd nd PCB-202 nd 176 0.0% nd nd PCB126 nd nd 1746 0.3% nd
∑PCB 402989 100% 569218 100% 642601 100% 51722 100%
PBDE28 1232.3 14.4% 2621.5 19.6% 735.3 4.7% 162.963 9.0% PBDE47 3980.0 46.4% 7033.8 52.7% 5919.1 38.0% 1370.37 75.5% PBDE99 1824.9 21.3% 1674.1 12.5% 1562.5 10.0% 55.55556 3.1% PBDE100 1376.8 16.1% 1853.2 13.9% 5487.1 35.2% 174.0741 9.6% PBDE153 162.1 1.9% 163.5 1.2% 1875.0 12.0% 51.85185 2.9%
∑PBDE 8576.1 100.0% 13346.1 100.0% 15579.0 100.0% 1814.8 100.0%
summary WS1-C3 WS2-C3 Mix-C5 NS-C3 ng/g lw % ng/g lw % ng/g lw % ng/g lw %
PCB126 nd nd 1746.324 0.3% nd ∑ 25 PCB 402989 97.9% 569218 97.7% 642601 97.4% 51722 96.6% ∑ 5 PBDE 8576 2.1% 13346 2.3% 15579 2.4% 1815 3.4%
total 411566 100.0% 582564 100.0% 659926 99.7% 53537 100.0%


117
CHAPTER 6
Is the reproduction success of European Eel affected by maternally transferred
dioxin-like compounds?
E.M. Foekemaa, M. Kottermanb, P. de Vriesc, A.J. Murkd
a IMARES, Department Experimental Ecology b IMARES, Department Fish c IMARES, Department Maritime d Wageningen University, Division of Toxicology

Chapter 6 Eggsposed
118
Abstract
Reported levels of dioxin-like compounds accumulated in European eel (Anguilla anguilla) were used to perform a risk assessment for eel larval survival, taking into account a modelled amplification of tissue concentrations with a factor of 1.33 or 2 during spawning migration. The calculated levels of dioxin-like compounds finally deposited in the eggs were compared to the internal effect concentrations for survival of early life stages of European eel as estimated from a sensitivity distribution based on literature data. As no toxicity data are available for eel larvae, it was assumed that they belong to the 5% or 1% most sensitive teleost fish species.
A risk assessment was performed for direct effect (lethality) of maternally transferred dioxin-like compounds for the developing embryos and larvae. Given concentrations of dioxin-like pollutants, and following the worst case scenarios, it can be expected that larvae of eel from highly contaminated locations in The Netherlands and Belgium will experience more than 50% mortality due to maternally transferred dioxin-like toxicants. Potential effects of other compounds or effects on the migration, condition and fertility of the parental animals were not taken into account. It is important to further study the overall impact of toxicants on the reproduction success of the European eel, as this may have been underestimated until now.

Eggsposed Chapter 6
119
6.1 Introduction
The numbers of juveniles (glass eel) of the European eel (Anguilla Anguilla) that reach the European coasts to populate the fresh water bodies, has dropped to a level below 10% of what it used to be before the 1980’s (Dekker, 2003). This decline gives rise to worries that the European eel may be on the edge of extinction. Over-exploitation of glass and mature eel, global changes in currents and environmental conditions, parasite infections, loss of habitat, and the presence of migration barriers such as dams and hydropower cyclones are mentioned as possible causes, and also the presence of anthropogenic toxicants in the eel’s environment is suggested to contribute (Palstra et al., 2006; Geeraerts et al., 2011; Brusle, 1991; Robinet and Feunteun, 2002). Due to the specific life history and eco-physiological challenges, eel could be higher exposed and more sensitive to the adverse effects of environmental pollutants than other fish species. After entering from sea as a glass eel, the eels remain in the fresh water for many years where they live in and near the, sometimes highly contaminated, sediment. With increasing body length, their diet shifts from macro invertebrates to fish, turning them into relative top predators (Tesch, 2003). The high lipid content of eel (10-30%) facilitates bioaccumulation of especially lipophilic substances, and chemical analysis confirm the presence of mixtures of PCBs, dioxins, brominated flame retardants, PAHs, organo-chlorine pesticides, TBT and perfluorinated compounds in eels (Kwadijk et al., 2010; Macgregor et al., 2010; Maes et al., 2008; Mariottini et al., 2006; McHugh et al., 2010; Roosens et al., 2010; Stachel et al., 2007; Tapie et al., 2011; de Boer et al., 2010; Geeraerts et al., 2011). This fed the idea that the observed collapse of the eel populations might, at least partly, be related to toxicological effects (Palstra et al., 2006; Geeraerts et al., 2011; Brusle, 1991; Robinet and Feunteun, 2002), for instance by reducing the chance for successful migration to the spawning grounds. After their stay in the freshwater environment as ‘yellow eels’, eels undergo a metamorphosis (silvering), during which the body is prepared for the long migration in marine water to the spawning grounds, most likely located in the Sargasso Sea, at 5000-6000 km distance (Tesch, 2003). As the fish, that now are referred to as ‘silver eels’, do not feed after silvering, they use their bodily resources as fuel during migration, for the sexual maturation, and to produce the eggs including the lipid-rich yolk. This makes efficient lipid storage before silvering essential for successful migration and reproduction. Eel monitoring programs in the Netherlands and Belgium from the last 15 years show a decline in the lipid content which has been suggested to be an effect of PCB exposure (Belpaire et al., 2009). Indeed, several environmental contaminants including PCBs, dioxins and some pesticides, have been shown to

Chapter 6 Eggsposed
120
interfere with fat accumulation and energy consumption (Robinet and Feunteun, 2002; van Ginneken et al., 2009).
For pacific salmon it has been shown that POPs accumulated in the tissue of the fish concentrate during spawning migration (Debruyn et al., 2004; Kelly et al., 2011). For eel this only has been shown experimentally during a simulated migration of 4 weeks (Ginneken et al., 2009). The increased POP concentrations could result in adverse effects on survival and sexual maturation of the eel, as such compounds have been shown to interfere with steroidogenesis in vitro (Montano et al., 2011; Zimmer et al., 2011), and with reproduction in fish (Daouk et al., 2011). In addition, female eel will deposit their lipids in the eggs which could then affect the developing larvae (Johnson et al., 1998; Latif et al., 2001; Nakayama et al., 2005; Yu et al., 2011). Lipophilic substances tend to evenly distribute over the lipids in the mother fish, including the gonads and hence the lipid normalised POP concentrations of the maternal tissues are close to those in the eggs (Russell et al., 1999).
During development of the early life stages the yolk-POP concentrations increase and peak when yolk lipids are depleted, shortly before the larvae become free feeding (Foekema et al., 2012). This implies that during larval development, tissue POP concentrations become higher than the concentrations in the adult mother fish, even if the eggs/larvae can develop under relative pristine conditions such as in the Sargasso Sea for the European eels. Embryonic and larval life stages are more sensitive to toxicants than fully developed individuals of the same species (Hutchinson et al., 1998) due to critical processes taking place such as development of tissues and the endocrine system.
Experiments with early life stages of the marine flatfish Solea solea that were exposed as eggs to mixtures of lipophilic POPs extracted from adult sole from a heavily contaminated estuary, indicated that mainly dioxin-like PCBs were responsible for the observed mortality (Chapter 5). Based on these findings, and the information that fish early life stages in general are very sensitive to dioxin-like toxicity (King-Heiden et al., 2012; Elonen et al., 1998; Foekema et al., 2008 –Chapter 2-), we here focus on the risk that dioxin-like compounds may pose to the early life stages of eel. The substances known to be mainly responsible for dioxin-like toxicity, PCDDs, PCDFs and some PCBs (Van den Berg et al., 1998; Van den Berg et al., 2006) still are ubiquitous in the environment. Their high lipophilicity (log Kow >6) facilitates bio accumulation, maternal transfer, and the development of a relative high internal peak concentration in the larval tissue at the end of the yolk sac stage (Foekema et al., 2012-Chapter 4-).

Eggsposed Chapter 6
121
In the present paper we investigate to what extent the larval survival of European eel could be negatively affected by maternally transferred dioxin-like compounds. For this purpose, we collected and calculated the following information:
1. Lipid-normalised concentrations of dioxin-like compounds in European eel tissue.
2. Development of concentrations of POPs with different log Kow in eel tissue during migration. For this, information about weight loss and changes in tissue composition during simulated migration in a laboratory set-up (Palstra and van den Thillart, 2010; van Ginneken et al., 2007; van Ginneken and Maes, 2005) was used to calculate the concentration of dioxin-like compounds in the lipids during migration using a mathematical model for the distribution of POPs in aquatic organisms (Hendriks et al., 2001).
3. Estimated sensitivity of eel early life stages. As successful larval development of the European eel in captivity still is not possible, no exposure studies can be performed to study the sensitivity of eel larvae to dioxin-like compounds. Therefore, the sensitivity of eel larvae for dioxin-like compounds was estimated based on the sensitivity distribution of larvae of other teleost fish species as published in Steevens et al., 2005.
Based on this information a risk assessment was performed for larval survival as consequence of maternal concentrations of dioxin-like compounds.
6.2 Data collection and calculations
6.2.1 Concentrations in European eel
A non-exhaustive literature search was performed to collect data about concentrations of dioxin-like pollutants (PCDDs, PCDFs, and PCBs) in European eel during the last decade. For the Netherlands also unpublished information was included from eel chemical monitoring programs performed by our own institute IMARES (previously RIVO). Most monitoring results of eel contaminant levels are reported on wet weight basis to be able to assess risks for human consumption, and often no indication of the lipid level is given. For our application only lipid-based tissue concentrations are useable, and therefore our datasets were limited to these data that were reported on lipid-base, or where reported lipid levels enable lipid-based normalisation. Furthermore, only datasets were included that were based on chemical analyses of the 12 dioxin-like PCBs (4 non-ortho and 8

Chapter 6 Eggsposed
122
mono-ortho) and 17 PCDDs/PCDFs as recommended by the WHO for the calculation of dioxin-like toxic potency.
The levels of individual congeners can be expressed as TCDD equivalency (TEQ) by multiplication of the concentration with Toxic Equivalency Factors (TEF). Based on differences in the sensitivity of taxonomic groups TEF-systems have been proposed for mammals, fish, and birds (Van den Berg et al., 1998). In 2005, the TEF system for mammals was revised (Van den Berg et al., 2006). The choice of the TEF-system to use has consequences for the resulting TEQ. As the far majority of the research on the presence of dioxin-like toxicity in fish focuses on the risk for human consumption, results are normally expressed using the WHO-TEF system for mammals as proposed in 1998 ((Van den Berg et al., 1998) and/or revised in 2005 (Van den Berg et al., 2006). The TEQ values for mammals however, overestimate the contribution of mon-ortho PCBs to the overall toxicity for fish. Therefore, in our study the toxicity of the measured compounds was expressed as TEQ by using the WHO Toxic Equivalence Factors (TEF) for fish (Van den Berg et al., 1998).
Information about dioxin-like toxicants in eel tissue for seven European countries is presented in Table 6.1. The most extended datasets were found for Belgium and the Netherlands both covering about 40 sites including heavily contaminated and relatively clean locations. The three included Norwegian locations are all highly contaminated (Grenland fjord), and the dataset for Germany originates from 10 locations in one river (Elbe). The dataset for Ireland, Poland and Portugal covered 7, 3 and 1 location(s) respectively, without indications that these sites were selected for their background contamination levels.
Table 6.1 Concentrations (median and range) of dioxin-like toxicants (in pg/g lw) in eel tissue from different locations in European countries expressed as Dioxin Toxic Equivalents (TEQ) calculated from tissue concentrations applying WHO1998 Toxic Equivalence factors (TEFs) for fish and mammals respectively. Presented are the averages and range of the reported values per location per country, sorted from low to high mammalian TEF-based TEQ values. The calculations are based on 12 WHO PCBs (4 non ortho and 8 mono-ortho) and 17 WHO PCDD/Fs
Country Period n locations TEQ’98fish TEQ’98mammals Reference
Portugal 2009 1 no data 4.7 Nunes et al 2011 Poland 2000-2008 3 no data 10.0 (2.5-31.4) Szlinder-Richert 2010 Ireland 2005-2007 7 3.3 (1.2-24.1) 15.6 (1.7-23.6) Mchugh et al 2010 Norway 2000-2001 3 no data 60.7 (53.4-170) Knutzen et al 2003 Belgium 2000-2007 38 11.3 (2.3-1565) 103 (12.3-5093) Geeraerts 2011 Germany 2002-2005 10 no data 122 (15.2-289) Stachel et al 2007 Netherlands 2004- 2011 42 20.4 (1.5-285) 120 (12.6-352) IMARES unpubl data

Eggsposed Chapter 6
123
The dioxin-like toxicity of all datasets was reported as TEQ applying the mammalian TEF system from 1998. Although not directly relevant for our calculations these data are included in Table 5.1 for rating the relative rank of pollution. It shows that the lowest TEQ levels reported are from the locations in Portugal, Poland, and Ireland, and that eel with the highest levels originate from Belgium, Germany, and the Netherlands. Detailed information on the underlying chemical dataset that allowed calculation of TEQs for fish was only available for Ireland, Belgium and the Netherlands. On average the TEF system for mammals resulted in 5 to 9 times higher TEQ levels than the TEF system for fish, but the ranking of the countries did not differ (Table 6.1).
6.2.2 Development of tissue POP levels during migration
The bioaccumulation model OMEGA (Hendriks et al., 2001) was applied in the present study to estimate the developments of POP concentrations in tissue of migrating silver eel. This model is based on classical fugacity theory for accumulation kinetics of organic substances as a function of the octanol-water partition ratio (Kow), and considers the role of species weight, lipid content and trophic level. Exchange (uptake and excretion) of the POP between the organism and its environment via water, food and faeces are included. The OMEGA model has been shown to well-predict steady state tissue concentrations in adult fish in field situations based on environmental concentrations (Veltman et al., 2005; Smitkova et al., 2005). Rather than determining the equilibrium ratios, the ordinary differential equation (ODE) provided by Hendriks and colleagues (Hendriks et al., 2001) was used as basis to determine the POP concentrations in the eel’s tissue in time. To this end, the ‘deSolve’ package (Soetaert et al., 2010) in R (The R Foundation for Statistical Computing, Vienna) was used to solve the ODE numerically, using Euler’s method, as proven successful before to calculate the development of PCB levels in sole larvae (Foekema et al., 2012-Chapter 4-).
For the model parameters that are assumed not to be species specific (respiration coefficient, rate exponent, body temperature correction factor, water layer diffusion resistance and lipid layer permeation resistance) the default and typical values presented in Hendriks et al., 2001 were applied (Table 5.2). Food intake (y1) was set at zero, as migrating eels do not feed. Mortality rate (k4) was set at zero since the model describes the development in an individual eel until spawning. As the major part of the migration takes place in a more or less pristine oceanic environment, the water concentration of POPs (C0,W) also was set at zero. The POP concentration in the tissue of the eel at the start of the migration was set at 1, so that the model outcome can be interpreted as the fraction of this concentration.

Chapter 6 Eggsposed
124
The remaining parameters for a standard silver eel were set at a body weight of 1 kg (W) and a lipid content of 25% (Pch2i = 0.25). We assumed that migration over 5500 km will take normally 180 days (Palstra and van den Thillart, 2010), but at optimal swimming speed this distance could be covered in 100 days (van Ginneken et al., 2005). During simulated migration experiments in swim tunnels with a continuous flow of fresh water covering the same distance in 180 days, an average loss of body weight of 19.7% was determined (van Ginneken et al., 2005). Later experiments from the same research group showed that the costs of transport are 20% higher when similar experiments are performed in seawater (Palstra et al., 2008). As almost the entire migration of the eel takes place in seawater, we assumed an average total weight loss of 25%. For a worst case situation we assume 50% weight loss during migration. The composition of the tissues does not change during the simulated migration, indicating that fat, protein and carbohydrate were metabolized in the same proportion (van Ginneken et al., 2005). Therefore, in our calculations, the lipid fraction was assumed not to change during migration.
The OMEGA model was used to predict the development of the POP concentration in the eel’s tissue for four migration scenarios. Two scenarios assume 25% weight loss during 180 (scenario 1) and 100 days (scenario 2) of migration, and two scenarios assuming maximum (50%) weight loss during 180 (scenario 3) and 100 days (scenario 4) of migration (Table 6.2).
The developments of the tissue concentrations are calculated for POPs with a log Kow of 4, 5, 6, and 7. As can be seen in Figure 6.1, model simulations indicate that tissue concentrations of POPs with a log Kow of 5 or less decrease during the migration. The removal mechanism involved is passive excretion through the water phase which is more effective for more water soluble substances. For POPs with log Kow 5 the model predicts a loss of approximately 50% of the initial concentration by the time the eel reaches the spawning grounds. Substances with log Kow of 4 and below can be expected to be completely washed out by then. Concentrations of POPs with log Kow values of 6 and higher are predicted to further magnify due to weight loss. Assuming there is no excretion or metabolism of a compound during migration, the tissue concentration at the end of the migration period can increase to 133% (Figure 6.1) and 200% of the initial tissue concentration for 25% and 50% weight loss respectively.
Changing the migration duration from 180 days into 100 days assuming optimal swimming speed (van Ginneken et al., 2005), does not affect the magnification of substances with log Kow 6 and above, as long as the total weight loss at the end of migration is not changed (Figure 6.2). Only the excretion of less lipophilic compounds (log Kow <6) is less with a shorter migration duration.
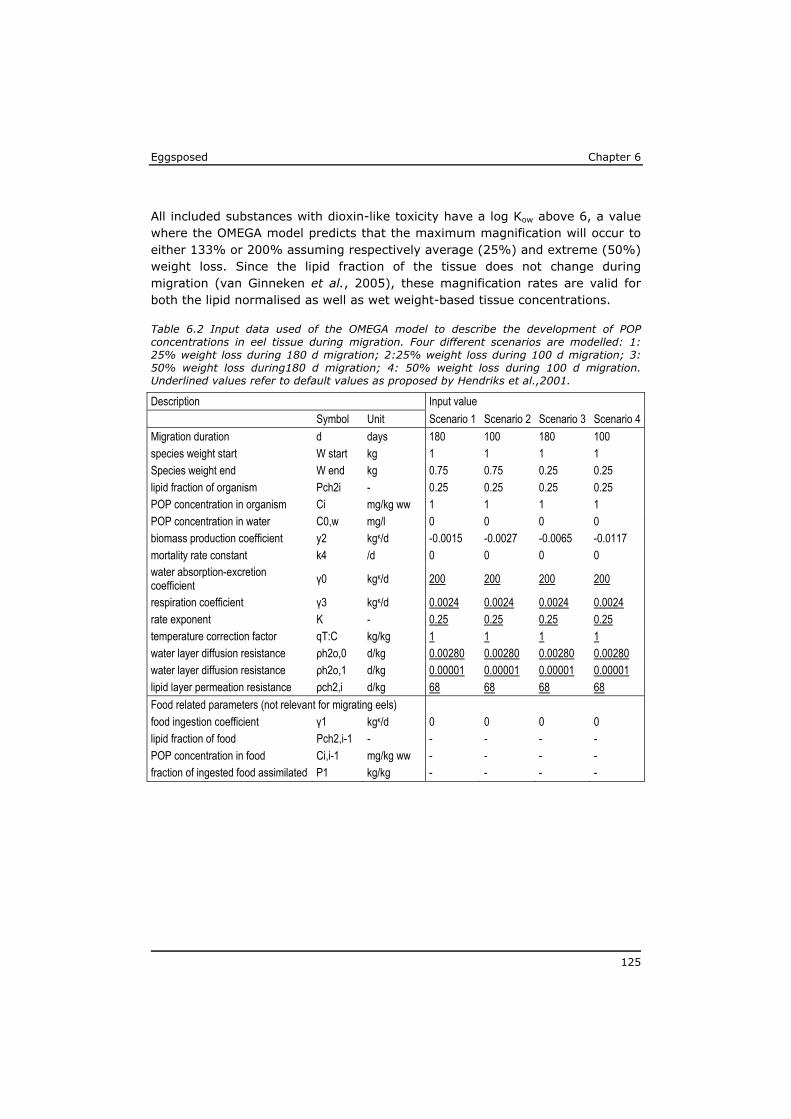
Eggsposed Chapter 6
125
All included substances with dioxin-like toxicity have a log Kow above 6, a value where the OMEGA model predicts that the maximum magnification will occur to either 133% or 200% assuming respectively average (25%) and extreme (50%) weight loss. Since the lipid fraction of the tissue does not change during migration (van Ginneken et al., 2005), these magnification rates are valid for both the lipid normalised as well as wet weight-based tissue concentrations.
Table 6.2 Input data used of the OMEGA model to describe the development of POP concentrations in eel tissue during migration. Four different scenarios are modelled: 1: 25% weight loss during 180 d migration; 2:25% weight loss during 100 d migration; 3: 50% weight loss during180 d migration; 4: 50% weight loss during 100 d migration. Underlined values refer to default values as proposed by Hendriks et al.,2001.
Description Input value
Symbol Unit Scenario 1 Scenario 2 Scenario 3 Scenario 4
Migration duration d days 180 100 180 100 species weight start W start kg 1 1 1 1 Species weight end W end kg 0.75 0.75 0.25 0.25 lipid fraction of organism Pch2i - 0.25 0.25 0.25 0.25 POP concentration in organism Ci mg/kg ww 1 1 1 1 POP concentration in water C0,w mg/l 0 0 0 0 biomass production coefficient y2 kgκ/d -0.0015 -0.0027 -0.0065 -0.0117 mortality rate constant k4 /d 0 0 0 0 water absorption-excretion coefficient
γ0 kgκ/d 200 200 200 200
respiration coefficient γ3 kgκ/d 0.0024 0.0024 0.0024 0.0024 rate exponent K - 0.25 0.25 0.25 0.25 temperature correction factor qT:C kg/kg 1 1 1 1 water layer diffusion resistance ρh2o,0 d/kg 0.00280 0.00280 0.00280 0.00280 water layer diffusion resistance ρh2o,1 d/kg 0.00001 0.00001 0.00001 0.00001 lipid layer permeation resistance ρch2,i d/kg 68 68 68 68
Food related parameters (not relevant for migrating eels) food ingestion coefficient γ1 kgκ/d 0 0 0 0 lipid fraction of food Pch2,i-1 - - - - - POP concentration in food Ci,i-1 mg/kg ww - - - - fraction of ingested food assimilated P1 kg/kg - - - -
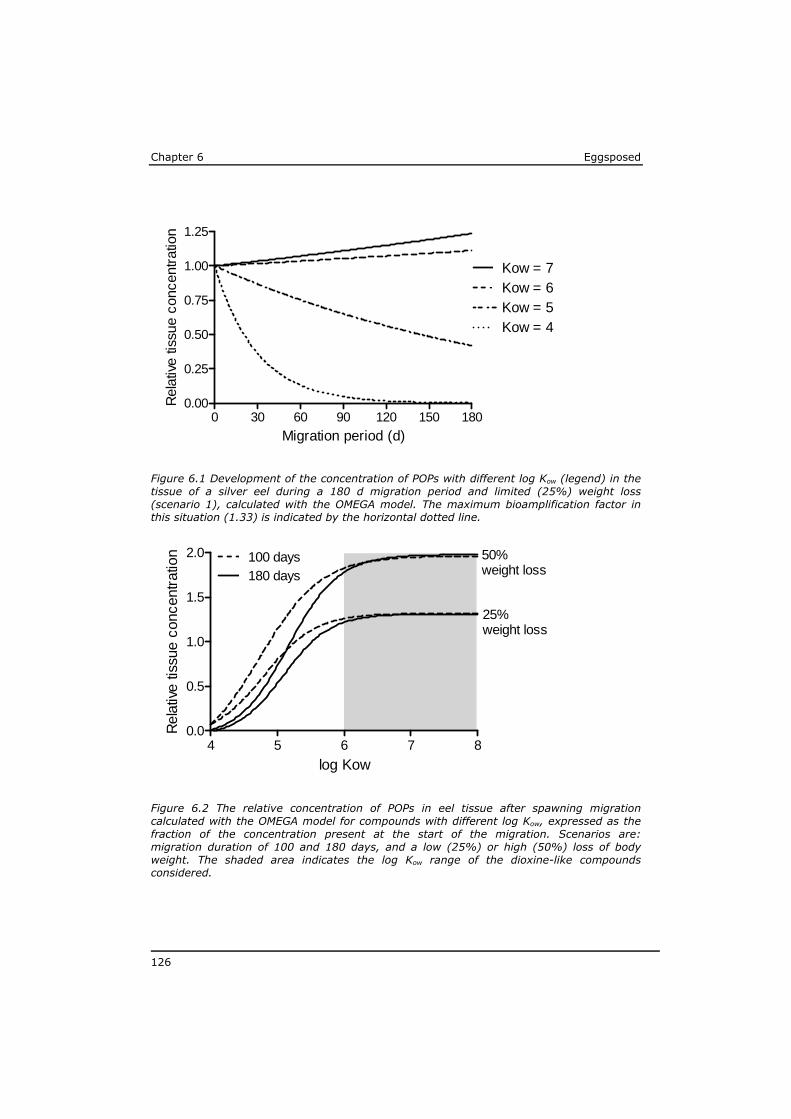
Chapter 6 Eggsposed
126
Figure 6.1 Development of the concentration of POPs with different log Kow (legend) in the tissue of a silver eel during a 180 d migration period and limited (25%) weight loss (scenario 1), calculated with the OMEGA model. The maximum bioamplification factor in this situation (1.33) is indicated by the horizontal dotted line.
Figure 6.2 The relative concentration of POPs in eel tissue after spawning migration calculated with the OMEGA model for compounds with different log Kow, expressed as the fraction of the concentration present at the start of the migration. Scenarios are: migration duration of 100 and 180 days, and a low (25%) or high (50%) loss of body weight. The shaded area indicates the log Kow range of the dioxine-like compounds considered.
Migration period (d)
Rel
ativ
e tis
sue
conc
entr
atio
n
0 30 60 90 120 150 1800.00
0.25
0.50
0.75
1.00
1.25
Kow = 4Kow = 5Kow = 6Kow = 7
log Kow
Rel
ativ
e tis
sue
conc
entr
atio
n
4 5 6 7 80.0
0.5
1.0
1.5
2.0
180 days100 days
25%weight loss
50%weight loss

Eggsposed Chapter 6
127
6.2.3 Estimating the sensitivity of eel early life stages
The sensitivity of eel larvae is estimated based on the distribution of the sensitivities of other teleost fish species as determined in 12 laboratory tests with TCDD, and 1 test with TCDD-like toxicants following egg exposure via water, injection or parental exposure and observation of the developing larvae at least until the end of the yolk sac stage (Steevens et al., 2005). From this distribution Steevens and colleagues derived egg concentrations below which 90 % (Hazard Concentration 10; HC10), 95%, or 99% (HC1) of the teleost fish species larvae do not exceed the median 50 % internal lethal concentration (ILC50) or the no/lowest observed internal (INOEC/ILOEC) concentration respectively (Table 6.3). The difference between ILC50 and INOEC/INOEC were not statistically significantly under the exposure conditions applied. For our worst-case approach we applied the 99% values (HC1), thus assuming that eel larvae belong to the 1% most sensitive teleost fish species.
The dataset assembled by Steevens et al., 2005 shows a positive correlation between the sensitivity of the species and the lipid content (1.7-8%) of the eggs. This is likely to be due to magnification of the lipophilic toxicant concentrations during depletion of the yolk lipids. The greater the lipid content, the greater the magnification can become. For example, in lean eggs (2% lipid) from Solea solea the increase in POP concentration at the end of the yolk-sac stage was a factor 2 to 3 (Foekema et al., 2012), while for the more lipid rich (8% lipids) eggs of Oncorhynchus tshawytscha this was around a factor 5 (Daley et al., 2012). As the sensitivity of larvae for dioxin-like compounds will mainly depend on the internal concentration reached, larvae of fish species with higher lipid content will be more vulnerable given the same POP-concentration in the egg yolk. Eel produce eggs with a relatively high lipid content (4.8-5.3% for A. japonica derived from Furuita et al., 2006; 5-7.5% for A. Anguilla, (personal comment M. Kotterman IMARES) This justifies the assumption that eel larvae belong to the more sensitive larvae among fish species.
Table 6.3 Dioxin-equivalent (TEQ) effect concentrations in fish eggs below which 99% (HC1), 95% (HC5) or 90% (HC10) of the teleost fish species are expected to experience 50% larval mortality or less (<ILC50), or no adverse effects on larval survival (<INOEC/ILOEC), with 95% confidence limits. Data from Steevens et al., 2005.
Species effect level pg TEQ/g lw in egg 99% <ILC50 (HC1) 58 (0.3 - 382) 95% <ILC50 (HC5) 386 (11.7 - 1430) 90% <ILC50 (HC10) 909 (58.3 - 2640)
99% <INOEC/ILOEC (HC1) 57 (15 – 201) 95% <INOEC/ILOEC (HC5) 321 (88 – 1050) 90% <INOEC/ILOEC (HC10) 699 (199 – 2220)

Chapter 6 Eggsposed
128
Observations of survival time of European eel embryos in relation to dioxin-like toxicity also suggested a high sensitivity with reduced survival already at 4 pg/g ww in the gonads of the parent fish (Palstra et al., 2006). However, these values were derived under sub-optimal conditions and all embryos, including the non-exposed died before hatching.
6.3 Risk assessment
To indicate theoretical tissue concentration in the female eel at the start of the migration that will result in 50% larval mortality, the critical TEQ-level in eggs were divided by 1.33 or 2.0 being the maximum magnification factor that may be expected during migration to the spawning grounds with 25% and 50% weight loss respectively. In the worst case scenario assuming that European eels are among the 1% most sensitive fish species, and the female eel loses half her body weight during migration, 50% larval mortality due to maternally transferred dioxin-like toxicants can be expected for tissue concentrations in silvering eel of 28.5 pg TEQ/g lw. At 29% of the sampled sites in the Netherlands and 21% of the Belgium sites these levels are exceeded (Figure 6.3A), sometimes even 10-50 fold. All samples from Ireland are below this critical level.
Assuming a lower weight loss of 25%, the critical tissue concentration increases to 43 pg TEQ/g lw. Obviously, all Irish locations are below this critical level. But at 11% of the locations in Belgium and 14% in the Netherlands also these levels are exceeded and according to this scenario more than 50% larval mortality may be expected for eggs from eel coming from these locations. At the most polluted locations in the Netherlands and Belgium this critical tissue concentration is exceeded 6 to 35 times. Even if we assume that eel larvae are amongst the 5% of the most sensitive fish species these critical levels for 50% larval mortality are exceeded at specific locations in the Netherlands and Belgium. Given the fact that historical POP levels in eel tissue from the Netherlands were 3 to 20 times higher in the year 1977 compared to 2006 (de Boer et al., 2010) these calculations indicate a serious risk for the larval survival of dioxin-like compounds in eel that manage to reach the spawning ground in the Sargasso sea.
Figure 6.3B shows the TEQs for the same eel samples from Ireland, Belgium and the Netherlands plus a number of other countries, now calculated with the mammalian TEF system (Van den Berg et al., 1998). Compared to the TEQsfish the TEQsmammalian are higher since in the mammalian TEF system the mono-ortho PCBs are assigned higher TEF values and these compounds are relatively

Eggsposed Chapter 6
129
abundant in aquatic species. Based on the TEQ(mammalian), the majority of eel sampled in Germany, Norway, Belgium and The Netherlands would exceed the critical tissue values. This reveals that it is very important to use the relevant TEF system, and also suggests that the TEQs in eel pose a serious risk to mammalian consumers.
In this study we only focus on the potential effects of dioxin-like substances, based on the indications that fish early life stages are especially sensitive to this group of compounds. However, many other POPs also have been shown to be accumulated in eel, including other compounds such as DDT/DDE and PBDEs (de Boer et al., 2010) with a log Kow of 6 or more. This means that these compounds too will be further concentrated during migration. For known accumulated POPs with lower log Kow values, such as HCH and HBCD, the body burdens are predicted to decrease during migration (Figure 6.2), and during larval development until free feeding as has been shown before (Foekema et al., 2012). The fate of other compounds is harder to predict. For example, perfluorinated compounds such as perfluorooctanesulfonate (PFOS) have been shown in high concentrations in eel tissue (Kwadijk et al., 2010). With both hydrophilic and hydrophobic physical properties PFOS behaves like fatty acids and therefore are not accumulated in lipids but in blood and cell membranes. These compounds are extremely persistent and have been shown to be maternally transferred also to fish eggs and to cause adverse effects in developing larvae (Wang et al., 2011). An important group of compounds that also have not been considered here are the hydroxylated POP-metabolites. Their effects on developing embryos can only be tested by exposing the mother fish to the parent compounds. A recent meta-analysis has shown that these metabolites still do occur at effect levels in several species, including humans that are exposed to POPs via the marine food chain (Montano et al., 2013).
Our risk assessment only was performed for direct effect (lethality) of maternally transferred dioxin-like compounds on the developing embryo and larvae. The potential impact of the presence of other toxicants including their metabolites was not included.
Potential effects of accumulated compounds and their metabolites on the migration, condition and fertility of the parental animals were not taken into account. It is important to further study the overall impact of toxicants on the reproduction success of the European eel, as this may have been underestimated until now.
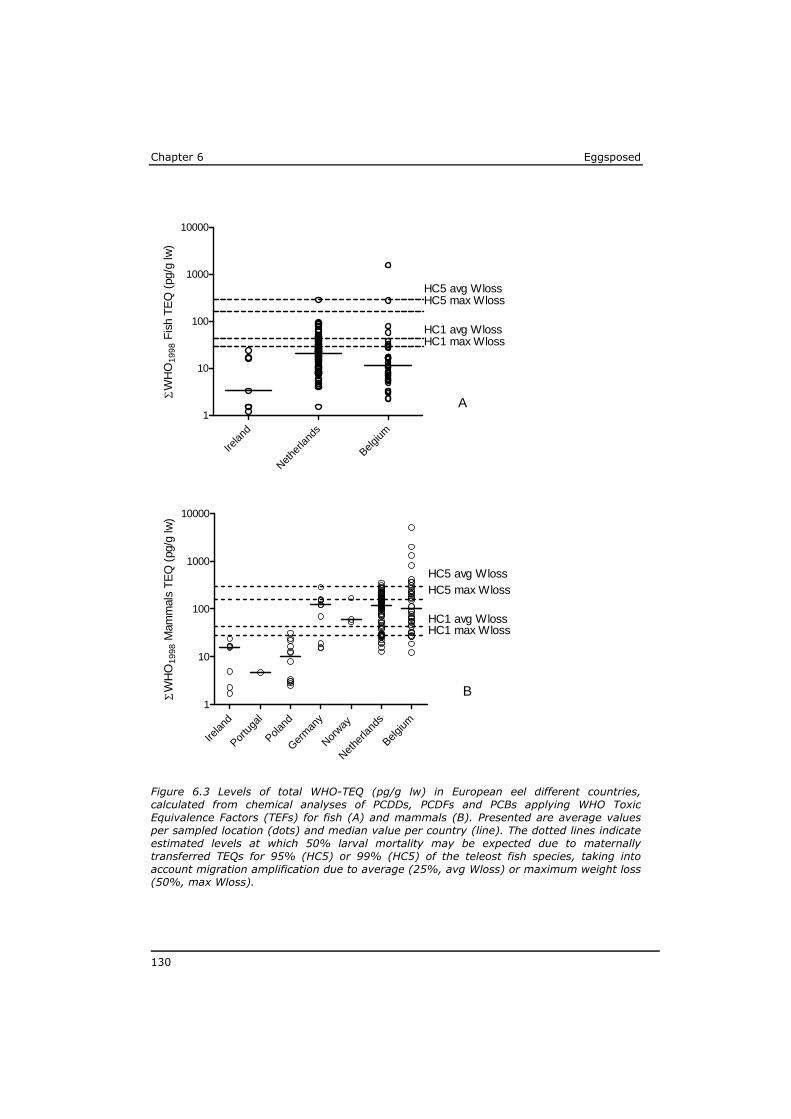
Chapter 6 Eggsposed
130
Figure 6.3 Levels of total WHO-TEQ (pg/g lw) in European eel different countries, calculated from chemical analyses of PCDDs, PCDFs and PCBs applying WHO Toxic Equivalence Factors (TEFs) for fish (A) and mammals (B). Presented are average values per sampled location (dots) and median value per country (line). The dotted lines indicate estimated levels at which 50% larval mortality may be expected due to maternally transferred TEQs for 95% (HC5) or 99% (HC5) of the teleost fish species, taking into account migration amplification due to average (25%, avg Wloss) or maximum weight loss (50%, max Wloss).
W
HO
19
98 F
ish
TE
Q (
pg/g
lw)
Irelan
d
Nethe
rland
s
Belgium
1
10
100
1000
10000
HC1 avg Wloss
HC5 avg Wloss
HC1 max Wloss
HC5 max Wloss
A
W
HO
19
98 M
amm
als
TE
Q (
pg/g
lw)
Irelan
d
Portu
gal
Poland
Germ
any
Norway
Nethe
rland
s
Belgium
1
10
100
1000
10000
HC1 avg Wloss
HC5 avg Wloss
HC1 max Wloss
HC5 max Wloss
B

Eggsposed Chapter 6
131
6.4 References
Belpaire CGJ, Goemans G, Geeraerts C, Quataert P, Parmentier K, Hagel P, et al. 2009. Decreasing eel stocks: survival of the fattest? Ecology of Freshwater Fish; 18
Brusle J. The eel (Anguilla sp.) and organic chemical pollutants. 1991. Science of the Total Environment; 102: 1-19.
Daley JM, Leadley TA, Pitcher TE, Drouillard KG. 2012. Bioamplification and the Selective Depletion of Persistent Organic Pollutants in Chinook Salmon Larvae. Environmental Science & Technology; 46: 2420-2426.
Daouk T, Larcher T, Roupsard F, Lyphout L, Rigaud C, Ledevin M, et al. 2011. Long-term food-exposure of zebrafish to PCB mixtures mimicking some environmental situations induces ovary pathology and impairs reproduction ability. Aquatic Toxicology; 105
de Boer J, Dao QT, van Leeuwen SPJ, Kotterman MJJ, Schobben JHM. 2010. Thirty year monitoring of PCBs, organochlorine pesticides and tetrabromodiphenylether in eel from The Netherlands. Environmental Pollution; 158: 1228-1236.
Debruyn AMH, Ikonomou MG, Gobas F. 2004. Magnification and toxicity of PCBs, PCDDs, and PCDFs in upriver-migrating Pacific salmon. Environmental Science & Technology; 38: 6217-6224.
Dekker W. 2003. Did lack of spawners cause the collapse of the European eel, Anguilla anguilla? Fisheries Management and Ecology; 10: 365-376.
Elonen GE, Spehar RL, Holcombe GW, Johnson RD, Fernandez JD, Erickson RJ, et al. 1998. Comparative toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin to seven freshwater fish species during early life-stage development. Environmental Toxicology and Chemistry; 17: 472-483.
Foekema EM, Fischer A, Parron ML, Kwadijk C, de Vries P, Murk AJ. 2012. Toxic concentrations in fish early life stages peak at a critical moment. Environmental Toxicology and Chemistry; 31: 1381-1390.
Furuita H, Unuma T, Nomura K, Tanaka H, Okuzawa K, Sugita T, et al. 2006. Lipid and fatty acid composition of eggs producing larvae with high survival rate in the Japanese eel. Journal of Fish Biology; 69: 1178-1189.
Geeraerts C, Focant JF, Eppe G, De Pauw E, Belpaire C. 2011. Reproduction of European eel jeopardised by high levels of dioxins and dioxin-like PCBs? Science of the Total Environment; 409: 4039-4047.
Hendriks AJ, van der Linde A, Cornelissen G, Sijm D. 2001. The power of size. 1. Rate constants and equilibrium ratios for accumulation of organic substances related to octanol-water partition ratio and species weight. Environmental Toxicology and Chemistry; 20: 1399-1420.
Hutchinson TH, Solbe J, Kloepper-Sams PJ. 1998. Analysis of the ECETOC aquatic toxicity (EAT) database - III - Comparative toxicity of chemical substances to different life stages of aquatic organisms. Chemosphere; 36: 129-142.
Kelly BC, Ikonomou MG, MacPherson N, Sampson T, Patterson DA, Dubetz C. 2011. Tissue residue concentrations of organohalogens and trace elements in adult pacific salmon returning to the Fraser river, Britisch Colombia, Canada. Environmental Toxicology and Chemistry; 30: 367-376.

Chapter 6 Eggsposed
132
King-Heiden TC, Mehta V, Xiong KM, Lanham KA, Antkiewicz DS, Ganser A, et al. 2012. Reproductive and developmental toxicity of dioxin in fish. Molecular and Cellular Endocrinology; 354: 121-138.
Kwadijk C, Korytar P, Koelmans AA. 2010. Distribution of Perfluorinated Compounds in Aquatic Systems in The Netherlands. Environmental Science & Technology; 44: 3746-3751.
Macgregor K, Oliver IW, Harris L, Ridgway IM. Persistent organic pollutants (PCB, DDT, HCH, HCB & BDE) in eels (Anguilla anguilla) in Scotland: Current levels and temporal trends. Environmental Pollution 2010; 158: 2402-2411.
Maes J, Belpaire C, Goemans G. 2010. Spatial variations and temporal trends between 1994 and 2005 in polychlorinated biphenyls, organochlorine pesticides and heavy metals in European eel (Anguilla anguilla L.) in Flanders, Belgium. Environmental Pollution 2008; 153: 223-237.
Mariottini M, Corsi I, Focardi S. 2006. PCB levels in European eel (Anguilla anguilla) from two coastal lagoons of the Mediterranean. Environmental Monitoring and Assessment; 117: 519-528.
McHugh B, Poole R, Corcoran J, Anninou P, Boyle B, Joyce E, et al. 2010. The occurrence of persistent chlorinated and brominated organic contaminants in the European eel (Anguilla anguilla) in Irish waters. Chemosphere; 79: 305-313.
Montano M, Zimmer KE, Dahl E, Berge V, Olsaker I, Skaare JU, et al. 2011. Effects of mixtures of persistent organic pollutants (POPs) derived from cod liver oil on H295R steroidogenesis. Food and Chemical Toxicology; 49: 2328-2335.
Palstra A, van Ginneken V, van den Thillart G. 2008. Cost of transport and optimal swimming speed in farmed and wild European silver eels (Anguilla anguilla). Comparative Biochemistry and Physiology a-Molecular & Integrative Physiology; 151: 37-44.
Palstra AP, van den Thillart G. 2010. Swimming physiology of European silver eels (Anguilla anguilla L.): energetic costs and effects on sexual maturation and reproduction. Fish Physiology and Biochemistry 2010; 36: 297-322.
Palstra AP, van Ginneken VJT, Murk AJ, van den Thillart G. 2006. Are dioxin-like contaminants responsible for the eel (Anguilla anguilla) drama? Naturwissenschaften; 93: 145-148.
Robinet TT, Feunteun EE. 2002. Sublethal effects of exposure to chemical compounds: A cause for the decline in Atlantic eels? Ecotoxicology; 11: 265-277.
Roosens L, Geeraerts C, Belpaire C, Van Pelt I, Neels H, Covaci A. 2010. Spatial variations in the levels and isomeric patterns of PBDEs and HBCDs in the European eel in Flanders. Environment International; 36: 415-423.
Russell RW, Gobas F, Haffner GD. 1999. Maternal transfer and in ovo exposure of organochlorines in oviparous organisms: A model and field verification. Environmental Science & Technology; 33: 416-420.
Smitkova H, Huijbregts MAJ, Hendriks AJ. 2005. Comparison of three fish bioaccumulation models for ecological and human risk assessment and validation with field data. Sar and Qsar in Environmental Research; 16: 483-493.

Eggsposed Chapter 6
133
Soetaert K, Petzoldt T, Setzer RW. 2010. Solving Differential Equations in R: Package deSolve. Journal of Statistical Software; 33: 1-25.
Stachel B, Christoph EH, Goetz R, Herrmann T, Krueger F, Kuehn T, et al. 2007. Dioxins and dioxin-like PCBs in different fish from the river Elbe and its tributaries, Germany. Journal of Hazardous Materials; 148: 199-209.
Steevens JA, Reiss MR, Pawlisz AV. 2005. A methodology for deriving tissue residue benchmarks for aquatic biota: a case study for fish exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin and equivalents. Integrated environmental assessment and management; 1: 142-151.
Tapie N, Le Menach K, Pasquaud S, Elie P, Devier MH, Budzinski H. 2011. PBDE and PCB contamination of eels from the Gironde estuary: From glass eels to silver eels. Chemosphere; 83: 175-185.
Van den Berg M, Birnbaum L, Bosveld ATC, Brunstrom B, Cook P, Feeley M, et al. 1998. Toxic equivalency factors (TEFs) for PCBs, PCDDs, PCDFs for humans and wildlife. Environmental Health Perspectives; 106: 775-792.
Van den Berg M, Birnbaum LS, Denison M, De Vito M, Farland W, Feeley M, et al. 2006. The 2005 World Health Organization reevaluation of human and mammalian toxic equivalency factors for dioxins and dioxin-like compounds. Toxicological Sciences; 93: 223-241.
van Ginneken V, Antonissen E, Muller UK, Booms R, Eding E, Verreth J, et al. 2005. Eel migration to the Sargasso: remarkably high swimming efficiency and low energy costs. Journal of Experimental Biology; 208: 1329-1335.
van Ginneken V, Dufour S, Sbaihi M, Balm P, Noorlander K, de Bakker M, et al. 2007. Does a 5500-km swim trial stimulate early sexual maturation in the European eel (Anguilla anguilla L.)? Comparative Biochemistry and Physiology a-Molecular & Integrative Physiology; 147: 1095-1103.
van Ginneken V, Palstra A, Leonards P, Nieveen M, van den Berg H, Flik G, et al. 2009. PCBs and the energy cost of migration in the European eel (Anguilla anguilla L.). Aquatic Toxicology; 92: 213-220.
van Ginneken VJT, Maes GE. 2005. The european eel (Anguilla anguilla, Linnaeus), its lifecycle, evolution and reproduction: a literature review. Reviews in Fish Biology and Fisheries; 15: 367-398.
Veltman K, Hendriks J, Huijbregts M, Leonards P, van den Heuvel-Greve M, Vethaak D. 2005. Accumulation of organochlorines and brominated flame retardants in estuarine and marine food chains: Field measurements and model calculations. Marine Pollution Bulletin; 50: 1085-1102.
Wang MY, Chen JF, Lin KF, Chen YH, Hu W, Tanguay RL, et al. 2011. Chronic zebrafish PFOS exposure alters sex ratio and maternal related effects in F1 offspring. Environmental Toxicology and Chemistry; 30: 2073-2080.
Zimmer KE, Montano M, Olsaker I, Dahl E, Berg V, Karlsson C, et al. 2011. In vitro steroidogenic effects of mixtures of persistent organic pollutants (POPs) extracted from burbot (Lota lota) caught in two Norwegian lakes. Science of the Total Environment; 409: 2040-2048.


135
CHAPTER 7
The potential impact of reduced larval survival on fish population development
E.M. Foekemaa, J.J. Poosb, K. van der Wolfshaarb, and A.J. Murkc
a IMARES, Department of Experimental Ecology b IMARES, Department of Fishery c Wageningen University, Division of Toxicology

Chapter 7 Eggsposed
136
Abstract
The potential impact of (toxicant induced) early life stage mortality on the population development of sole (Solea solea) was explored by application of an age structured matrix model. Sole was chosen for this exercise since the life history is well known and it is applied as model species in ecotoxicological research. The model is used to explore the population response to a combination of larval mortality and fishing-related mortality of mature fish. The results indicate that the impact of larval mortality that occurs before metamorphosis is very low, even in combination with high fishing pressure. This is the result of the combination of the high fecundity and the fact that the larval mortality occurs before the moment when the number of recruits is limited by the carrying capacity of the nursery areas. Mortality that occurs after the nursery areas are populated will have a more pronounced impact on population development. The results further imply that population development of pelagic fish species that do not concentrate in nursery areas and species with low fecundity is more vulnerable for disturbance through mortality of early life stages.

Eggsposed Chapter 7
137
7.1 Introduction
When present in high enough concentrations in the tissue of adult fish, maternally transferred organic substances will have a negative impact on the survival and development of the larvae into juveniles and eventually adult fish. Our experiments, performed with the common Sole (Solea solea) as model species, show an increased mortality rate before the onset of the metamorphosis (Chapter 2 and 5). Due to bio amplification of lipophilic pollutants such as dioxins and PCBs, fish larvae will experience the highest exposure and therefore the strongest acute toxicity at the end of the yolk-sac stage.
Translation from impact on specific life stages to ecological relevant endpoints is not straightforward, as the most sensitive endpoint is not necessarily the most important for population development (Forbes et al., 2010). Forbes and co-workers argue that this makes evolutionary sense as it dampens variation in population growth rates. Hence, to be able to determine the relevance of a toxic effect on early life stages at population level, it is important to know the population dynamics of the species.
Survival rates of fish early life stages in nature are generally low, at least in the case of oviparous species without parental care like sole and most other commercially exploited species. This low survival rate is compensated by the production of great numbers of eggs. Large changes in fecundity can be compensated by changes in mortality or survival rate that are often density dependent (Rothschild, 2000).
An analysis of datasets of 104 exploited marine species revealed that the population resilience at low adult abundance varies strongly between species. Many species exhibited strong compensatory dynamics with increasing reproduction at low adult densities (Keith and Hutchings, 2012). One of the mechanisms behind the compensation is described as the concentration hypothesis: Species that make a shift from a pelagic to a benthic habitat during the early development, experience an immediate concentration of their habitat from three into two dimensions, which could lead to density dependent mortality. The stronger this effect, the better the population can compensate for changes in population size (Beverton, 1995). The maximum number of recruits that can be sustained in a certain habitat is limited, and therefore a limited number of adults can produce sufficient amounts of eggs. Data analyses confirmed this hypothesis, revealing low compensation rates for fully pelagic species like mackerel and sardine, and intermediate compensation for gadoid species like cod and saithe of which many juveniles life close to the seabed. The highest compensation rates were found for flatfishes like sole and plaice of which
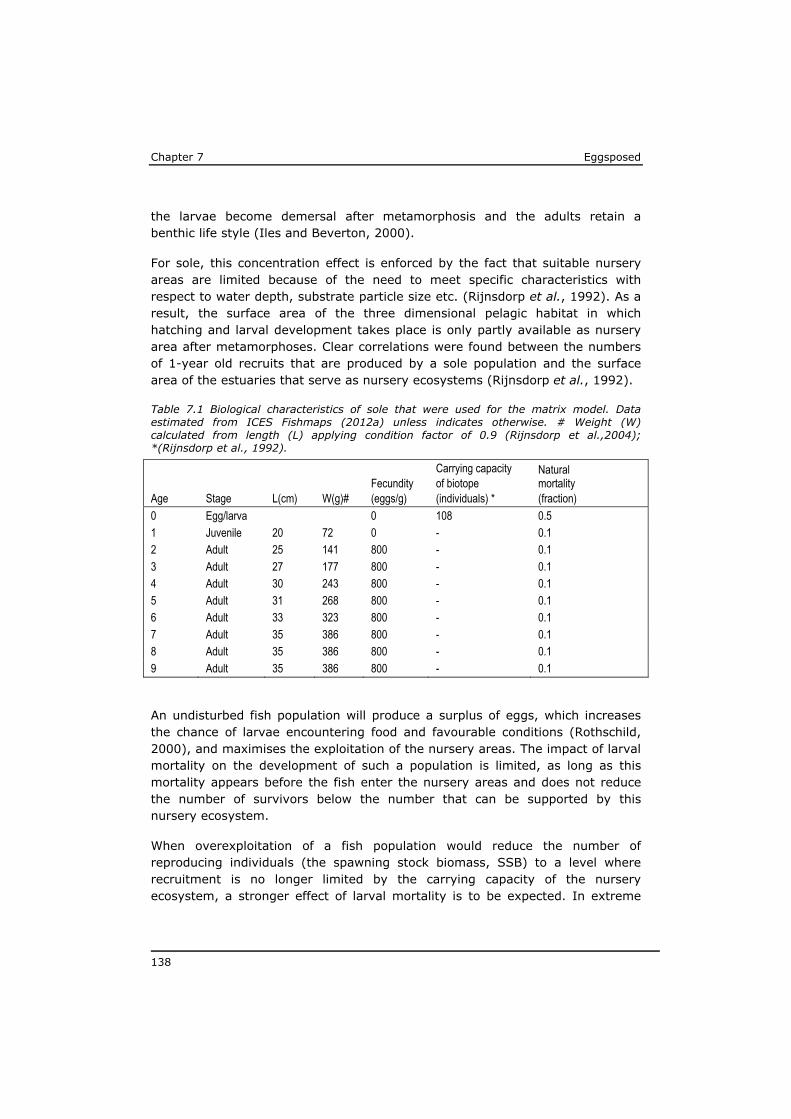
Chapter 7 Eggsposed
138
the larvae become demersal after metamorphosis and the adults retain a benthic life style (Iles and Beverton, 2000).
For sole, this concentration effect is enforced by the fact that suitable nursery areas are limited because of the need to meet specific characteristics with respect to water depth, substrate particle size etc. (Rijnsdorp et al., 1992). As a result, the surface area of the three dimensional pelagic habitat in which hatching and larval development takes place is only partly available as nursery area after metamorphoses. Clear correlations were found between the numbers of 1-year old recruits that are produced by a sole population and the surface area of the estuaries that serve as nursery ecosystems (Rijnsdorp et al., 1992).
Table 7.1 Biological characteristics of sole that were used for the matrix model. Data estimated from ICES Fishmaps (2012a) unless indicates otherwise. # Weight (W) calculated from length (L) applying condition factor of 0.9 (Rijnsdorp et al.,2004); *(Rijnsdorp et al., 1992).
Age Stage L(cm) W(g)# Fecundity (eggs/g)
Carrying capacity of biotope (individuals) *
Natural mortality (fraction)
0 Egg/larva 0 108 0.5 1 Juvenile 20 72 0 - 0.1 2 Adult 25 141 800 - 0.1 3 Adult 27 177 800 - 0.1 4 Adult 30 243 800 - 0.1 5 Adult 31 268 800 - 0.1 6 Adult 33 323 800 - 0.1 7 Adult 35 386 800 - 0.1 8 Adult 35 386 800 - 0.1 9 Adult 35 386 800 - 0.1
An undisturbed fish population will produce a surplus of eggs, which increases the chance of larvae encountering food and favourable conditions (Rothschild, 2000), and maximises the exploitation of the nursery areas. The impact of larval mortality on the development of such a population is limited, as long as this mortality appears before the fish enter the nursery areas and does not reduce the number of survivors below the number that can be supported by this nursery ecosystem.
When overexploitation of a fish population would reduce the number of reproducing individuals (the spawning stock biomass, SSB) to a level where recruitment is no longer limited by the carrying capacity of the nursery ecosystem, a stronger effect of larval mortality is to be expected. In extreme

Eggsposed Chapter 7
139
situations this could be simply because the remaining SSB is not enough to produce sufficient numbers of eggs. In addition, the reproduction of fish populations could decline stronger than predicted only based on the population density (Kramer et al., 2009; Keith and Hutchings, 2012) when the density of mates is too low to ensure participation of all mature animals in reproduction (Rowe and Hutchings, 2003), a so-called ‘Allee effect’ (Kramer et al., 2009).
The ecotoxicological experiments presented in chapters 2 and 5 of this thesis, show that the impact on sole larvae of maternally transferred lipophilic toxicants as studied, appears early in the development. For sole this is before the onset of the metamorphosis. Thus, before the fish colonise the nursery areas where density dependent processes become effective (Rijnsdorp et al., 1992). Potential effects that might develop in a later stage were not considered. In order to explore the potential impact of this toxicant induced early life stage mortality on the population development of sole, a matrix population model was developed based on the life history of sole in the North Sea. The model was kept as simple as possible and does not take into account different factors known to be important in the recruitment of fish, such as ambient water temperature (Fassler et al., 2011; Rijnsdorp et al., 1992), food availability (Nash and Dickey-Collas, 2005), currents and predation, or Allee effects (Rowe and Hutchings, 2003; Kramer et al., 2009). The results must therefore be regarded as purely indicative for the relevancy of (toxicity induced) early life mortality for sole population development.
7.2 Model description
The model describes the numerical development of a sole population per age class, with a maximum age of 9 years (Figure 7.1). Although sole can live up to 40 years individuals older than 10 years nowadays are scarce as a result of high fishing pressure (ICES FishMap, 2012a). At age 2 (‘Age-2’ in Figure 7.1 all individuals mature. Adults annually produce a number of eggs (‘Age-0’) based on the age specific body weight and a general fecundity of 800 eggs per gram female fish (Table 7.1) (ICES 2012a; Rijnsdorp et al., 1992). A 1:1 sex ratio is assumed; hence for the calculation of the population fecundity adult biomass is multiplied by 0.5 to model egg production.
Of all eggs entering the ‘Age-0’ box, 50% are not viable, for instance because they are not fertilised or of poor quality. The 50% hatching success of the spawned eggs is normal in sole aquaculture where maturation, spawning and fertilisation are achieved under more or less natural conditions (pers. comm. A. Kamstra IMARES). If toxicity related mortality (‘Tmort’) occurs, a fraction of the

Chapter 7 Eggsposed
140
hatched larvae will not further develop. The remaining larvae have the potential to develop into juveniles that are assumed not to suffer from additional toxicity. However, the number that can be transferred to the ‘Age-1’ box is limited by the carrying capacity of the nursery ecosystem, and the surplus is assumed to die. The carrying capacity of the nursery ecosystem hence sets the maximum number of individuals in the population of age 1 and older. For North Sea sole the geometric mean of the recruitment level was determined at approximately 94 million individuals (Rijnsdorp et al., 1992), for our model this was rounded off at 100 million individuals, and set as ceiling for the number of ‘Age-0’ individuals that can develop into ‘Age-1’ juveniles. From Age-1 onwards, natural yearly mortality was set at 10%, conform the average value that is applied by ICES for sole stock management advices (ICES, 2011). After Age-1 the survival can be further reduced by fishing related mortality (‘Fmort’). The fishing mortality of sole in the North Sea that was used as default value in the model was 30%, being the estimated average fishing mortality in 2011 (ICES, 2012).
Figure 7.1 Overview of the matrix model that was used for the calculations. The grey arrows indicate the external conditions that affect the survival.
Age‐0(egg/larv.)
Age‐1(juvenile)
Age‐2(adult)
Age‐9(adult)
Age‐n(adult)
A
B
C
B
B
C
C
= Age‐0 * 0.5 * (1‐Tmort)If A> carrying capacity, then A= carrying capacity
= Age‐n * (1‐(Nmort + Fmort))
= Age‐n * 0.5 * Weight‐n * Fecundity
Age‐n = number of individualsof age‐nWeight‐n = weight of individualsof age‐nNmort = fraction naturalmortalityTmort = fraction mortality caused by toxicityFmort = fraction mortality caused by fishing
A
B
C

Eggsposed Chapter 7
141
Different scenarios were run to explore the response of the modelled sole population to changes in (fishing related) mortality of adults, and (toxicity related) mortality of larvae. In addition the impact of changes in the fecundity and the carrying capacity on these model responses was explored. The model describes the development of a sole population during a 50 year period, expressed as development of the biomass of adult fish (∑Age-2 to 9; spawning stock biomass, SSB), and the number of Age-1 recruits.
7.3 Results
In the default situation with a fishing mortality of 30%, no toxicity related mortality, and a carrying capacity of 100 million recruits, the population reached equilibrium at 31 717 tonnes SSB (Figure 7.2). This is close to the 35 000 to 46 700 tonnes that is estimated as the SSB of sole in the North Sea in 2011 and 2012 respectively (ICES, 2012), indicating that despite the simplicity of the model, the order of magnitude of the outcome corroborates with other estimates. Applying a natural mortality of 5 and 0%, instead of the default 10% leads to equilibrium SSBs of 41 987 and 55 472 tonnes respectively.
When, starting from the equilibrium default situation the fishing mortality is changed, new equilibrium SSB levels develop (Figure 7.2). These equilibria logically show a negative correlation with fishing mortality as a consequence of the mortality being a fixed fraction of the population. Increasing fishing mortality above 90% in combination with 10% natural mortality, leads to immediate extinction of the modelled population. Also changes in the carrying capacity for maximum numbers Age-1 recruits result in new equilibrium values of SSB. These are positively correlated with the carrying capacity of the nursery ecosystem as this carrying capacity sets the maximum number of survivors to Age 1.
The impact of larval mortality was explored for populations that were in equilibrium in the absence of fishing mortality, and with 30% and 80% fishing mortality respectively. In all of these situations the population development was very insensitive to the effects of larval mortality (Figure 7.3). Without fishing pressure only 99.999% mortality resulted in a population decline. In combination with fishing mortality of 30% or more, population development is affected when larval mortality is more than 99.995%, thus almost complete and then leads to extinction of the population. Even with 80% fishing mortality only a larval mortality of more than 99.9% contributes to population decline.
It can be recognised that the impact of toxicity increases with increasing fishing mortality. However, these differences are of no significance.
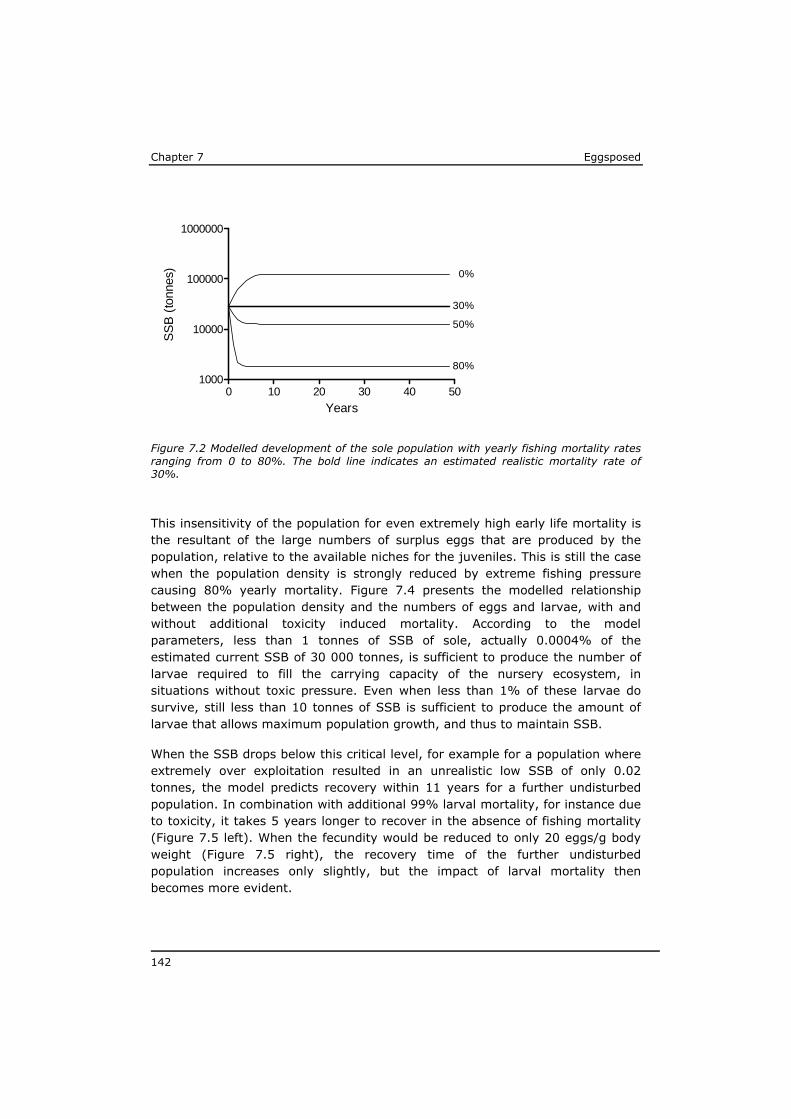
Chapter 7 Eggsposed
142
Figure 7.2 Modelled development of the sole population with yearly fishing mortality rates ranging from 0 to 80%. The bold line indicates an estimated realistic mortality rate of 30%.
This insensitivity of the population for even extremely high early life mortality is the resultant of the large numbers of surplus eggs that are produced by the population, relative to the available niches for the juveniles. This is still the case when the population density is strongly reduced by extreme fishing pressure causing 80% yearly mortality. Figure 7.4 presents the modelled relationship between the population density and the numbers of eggs and larvae, with and without additional toxicity induced mortality. According to the model parameters, less than 1 tonnes of SSB of sole, actually 0.0004% of the estimated current SSB of 30 000 tonnes, is sufficient to produce the number of larvae required to fill the carrying capacity of the nursery ecosystem, in situations without toxic pressure. Even when less than 1% of these larvae do survive, still less than 10 tonnes of SSB is sufficient to produce the amount of larvae that allows maximum population growth, and thus to maintain SSB.
When the SSB drops below this critical level, for example for a population where extremely over exploitation resulted in an unrealistic low SSB of only 0.02 tonnes, the model predicts recovery within 11 years for a further undisturbed population. In combination with additional 99% larval mortality, for instance due to toxicity, it takes 5 years longer to recover in the absence of fishing mortality (Figure 7.5 left). When the fecundity would be reduced to only 20 eggs/g body weight (Figure 7.5 right), the recovery time of the further undisturbed population increases only slightly, but the impact of larval mortality then becomes more evident.
Years
SS
B (
tonn
es)
0 10 20 30 40 501000
10000
100000
1000000
0%
30%
50%
80%
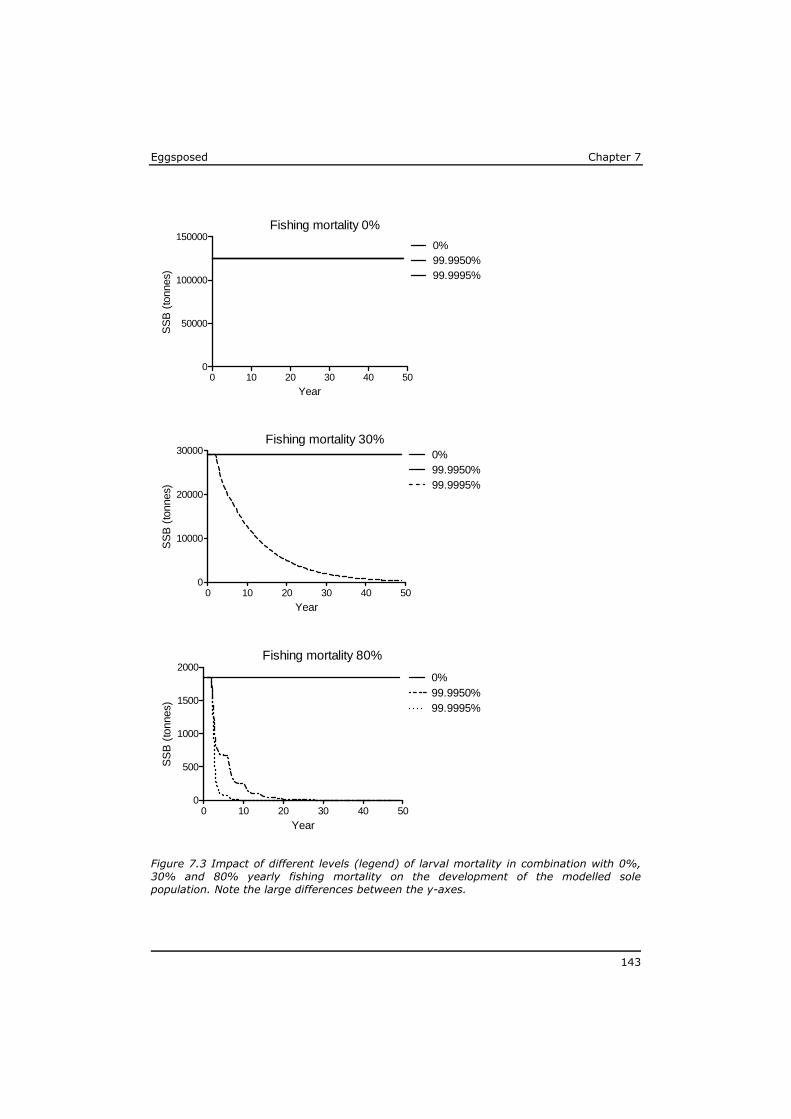
Eggsposed Chapter 7
143
Figure 7.3 Impact of different levels (legend) of larval mortality in combination with 0%, 30% and 80% yearly fishing mortality on the development of the modelled sole population. Note the large differences between the y-axes.
Year
SS
B (
tonn
es)
0 10 20 30 40 500
50000
100000
1500000%
Fishing mortality 0%
99.9995%99.9950%
Year
SS
B (
tonn
es)
0 10 20 30 40 500
10000
20000
30000 0%Fishing mortality 30%
99.9995%99.9950%
Year
SS
B (
tonn
es)
0 10 20 30 40 500
500
1000
1500
20000%99.9950%
Fishing mortality 80%
99.9995%
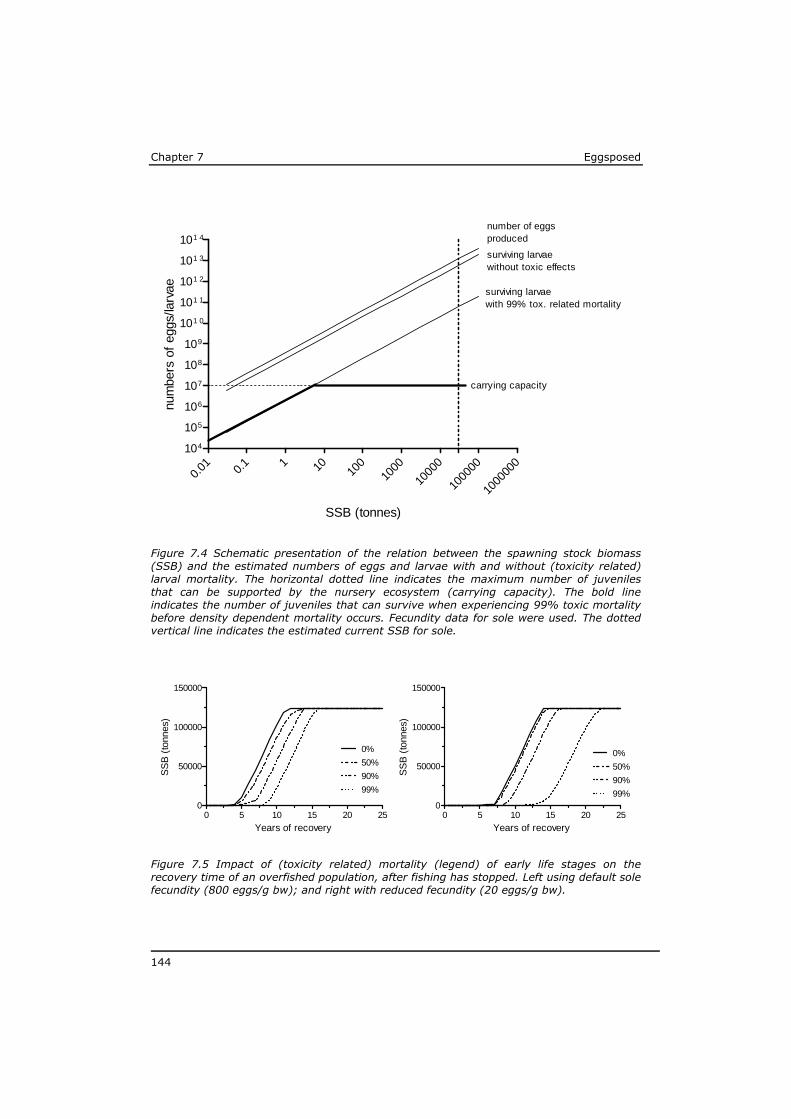
Chapter 7 Eggsposed
144
Figure 7.4 Schematic presentation of the relation between the spawning stock biomass (SSB) and the estimated numbers of eggs and larvae with and without (toxicity related) larval mortality. The horizontal dotted line indicates the maximum number of juveniles that can be supported by the nursery ecosystem (carrying capacity). The bold line indicates the number of juveniles that can survive when experiencing 99% toxic mortality before density dependent mortality occurs. Fecundity data for sole were used. The dotted vertical line indicates the estimated current SSB for sole.
Figure 7.5 Impact of (toxicity related) mortality (legend) of early life stages on the recovery time of an overfished population, after fishing has stopped. Left using default sole fecundity (800 eggs/g bw); and right with reduced fecundity (20 eggs/g bw).
SSB (tonnes)
num
bers
of
eggs
/larv
ae
0.01 0.
1 1 10 100
1000
1000
0
1000
00
1000
000
104
105
106
107
108
109
101 0
101 1
101 2
101 3
101 4
surviving larvaewith 99% tox. related mortality
number of eggsproduced
surviving larvaewithout toxic effects
carrying capacity
Years of recovery
SS
B (
tonn
es)
0 5 10 15 20 250
50000
100000
150000
0%
90%
50%
99%
Years of recovery
SS
B (
tonn
es)
0 5 10 15 20 250
50000
100000
150000
0%
90%
50%
99%

Eggsposed Chapter 7
145
7.4 Discussion
The model outcomes suggest that sole populations are able to compensate high levels of larval mortality that occurs before the nursery areas are colonised. This is related to the high degree of concentration of individuals that takes place when the pelagic sole larvae change to a demersal lifestyle while entering the nursery grounds, of which the carrying capacity is limited by the surface area. Compared to other fish species the concentration effect is relatively large for flatfish (Iles and Beverton, 2000). Species of which the whole life cycle takes place in the pelagic, like mackerel, sardine, and anchovies can be assumed to be more sensitive to early life stage mortality. Whereas the degree of concentration for gadoid species like cod and saithe forms an intermediate between flatfish and pelagic species (Iles and Beverton, 2000). For herring, also a pelagic fish, but with demersal eggs, the situation might be different. At high population density, the number of produced eggs can be limited by availability of suitable substrates needed by this species for egg deposition. Once the carrying capacity of these substrates is reached a further increase of the population will not result in more compensation for early life stage mortality.
The maternally transferred lipophilic toxicants that we studied caused mortality at an early stage of development. The impact of this type of larval mortality on our modelled sole population only became apparent at very low population levels. It is questionable if such a population would be able to sustain, even without impact of toxicants. Our model overestimates the recovery potential of the population since it assumes that all surviving larvae will be able to reach the nursery grounds, which in reality is not the case (Bolle et al., 2009).
Besides this, the number of viable eggs produced by a female is not affected by so called Allee effects in our model, although that can be expected at low population densities (Rowe and Hutchings, 2003; Kramer et al., 2009; Keith and Hutchings, 2012). The combination of these simplifications in the model makes that only one single adult fish would be able to produce sufficient eggs to start a sustainable population.
In addition factors like water temperature (Fassler et al., 2011; Rijnsdorp et al., 1992), food availability (Nash and Dickey-Collas, 2005), predation (Nash and Geffen, 2012), that have large impact on the recruitment process are not included in our model. Therefore, the outcome should only be considered as purely explorative for the potential role of (toxicity related) larval mortality on fish population development. Despite this, the general conclusion that sole is protected against a high level of toxic effects on early life stages by the combination of high fecundity and the concentration of the older larvae in the

Chapter 7 Eggsposed
146
nursery areas seems valid. These findings are in line with the outcome of a literature review that compensatory interactions and hence buffering of toxicant effects on the dynamics of density limited populations are likely (Forbes et al., 2001). Pelagic species can be expected to be more sensitive as here larval mortality is not compensated by concentration in nursery areas. This would especially be the case for species with relatively low fecundity.
7.5 Conclusions
The model results indicate that a high fecundity in combination with density dependent regulation of the maximum numbers of recruits, protects sole and other (flatfish) species with comparable reproductive biology, against the impact of (toxicant induced) high early life stage mortality. At least as long as this mortality occurs before density dependent processes take place, which may be expected with the type of toxic effects that are associated with maternally transferred substances.
Population development of pelagic species and of species with low fecundity can be considered more sensitive to the impact of early life stage mortality.
7.6 References
Beverton, R.J.H., 1995. Spatial limitation of population size; The concentration hypothesis. Neth. J. Sea Res. 34, 1-6.
Bolle, L.J., Dickey-Collas, M., van Beek, J.K.L., Erftemeijer, P.L.A., Witte, J.I.J., van der Veer, H.W., Rijinsdorp, A.D., 2009. Variability in transport of fish eggs and larvae. III. Effects of hydrodynamics and larval behaviour on recruitment in plaice. Marine Ecology Progress Series 390, 195-211.
Fassler, S.M.M., Payne, M.R., Brunel, T., Dickey-Collas, M., 2011. Does larval mortality influence population dynamics? An analysis of North Sea herring (Clupea harengus) time series. Fisheries Oceanography 20, 530-543.
Forbes, V.E., Olsen, M., Palmqvist, A., Calow, P., 2010. Environmentally sensitive life-cycle traits have low elasticity: implications for theory and practice. Ecological Applications 20, 1449-1455.
Forbes, V.E., Sibly, R.M., Calow, P., 2001. Toxicant impacts on density-limited populations: A critical review of theory, practice, and results. Ecological Applications 11, 1249-1257.
ICES (2011) WGNSSK REPORT 2011 Sec 10 Sole in Subarea IV ICES (2012) ICES Advice 2012, Book 6 http://www.ices.dk/committe/acom/comwork
/report/2012/2012/sol-nsea.pdf ICES (2012a) ICES FishMap Sole, http://www.ices.dk/marineworld/fishmap/ices/pdf/
sole.pdf (accessed November 2012)

Eggsposed Chapter 7
147
Iles, T.C., Beverton, R.J.H., 2000. The concentration hypothesis: The statistical evidence. Ices Journal of Marine Science 57, 216-227.
Keith, D.M., Hutchings, J.A., 2012. Population dynamics of marine fishes at low abundance (vol 69, pg 1150, 2012). Canadian Journal of Fisheries and Aquatic Sciences 69, 1722-1722.
Kramer, A.M., Dennis, B., Liebhold, A.M., Drake, J.M., 2009. The evidence for Allee effects. Population Ecology 51, 341-354.
Nash, R.D.M., Dickey-Collas, M., 2005. The influence of life history dynamics and environment on the determination of year class strength in North Sea herring (Clupea harengus L.). Fisheries Oceanography 14, 279-291.
Nash, R.D.M., Geffen, A.J., 2012. Mortality through the early life-history of fish: What can we learn from European plaice (Pleuronectes platessa L.)? J. Mar. Syst. 93, 58-68.
Rijnsdorp, A.D., Vanbeek, F.A., Flatman, S., Millner, R.M., Riley, J.D., Giret, M., Declerck, R., 1992. Recruitment of sole stocks, Solea solea (L) in the North East Atlantic. Neth. J. Sea Res. 29, p 173.
Rothschild, B.J., 2000. "Fish stocks and recruitment": the past thirty years. Ices Journal of Marine Science 57, 191-201.
Rowe, S., Hutchings, J.A., 2003. Mating systems and the conservation of commercially exploited marine fish. Trends in Ecology & Evolution 18, 567-572.


149
CHAPTER 8
General discussion
“You did not kill the fish only to keep alive and to sell for food, he thought. You killed him for pride and because you are a fisherman. You loved him when he was alive and you loved him after. If you love him, it is not a sin to kill him.
Or is it more?”
Ernest Hemmingway: The Old Man and the Sea

Chapter 8 Eggsposed
150
8.1 Introduction
The overall aim of the research presented in this thesis was to investigate whether and how maternal transfer of lipophilic POPs, that are ubiquitous in the marine environment, can affect the development and survival of fish larvae. And if so, to assess if this could result in reduced population development taking into account fishing as major pressure for fish populations. The focus was on lipophilic POPs that can cause dioxin-like toxicity or that are known to disrupt the thyroid hormone system. Both effect types are relevant for early life development. In addition, mixtures of lipophilic POPs were tested as they are accumulated in fish tissue.
In the study, fish early life stage tests were performed with exposure during the egg stage only, followed by observation until metamorphosis into juveniles. Results clearly show delayed mortality days after exposure had stopped, when the fish already are free-feeding. Crucial in this aspect is the peak in exposure that develops for highly lipophilic pollutants at the end of the yolk-sac stage. The height of this peak is the resultant of a combination of the lipophilicity of the pollutant and the utilisation of the yolk lipids by the developing larvae. Dioxin-like toxicity and baseline toxicity are the most likely toxicological mechanisms causing the observed effects. The test set up, with the common sole (Solea solea) as model species, did not provide indications for disruption of the thyroid hormone mediated metamorphosis. The effects of mixtures of lipophilic POPs extracted from the tissue of fish collected at contaminated field locations could be explained based on the individual pollutants present. No indications were found for synergistic mixture effects or for a substantial role of other lipophilic POPs in the mixtures than the PCBs and the brominated flame retardants that were subject of this study. Based on this research it is estimated that the levels of lipophilic POPs currently present in sole from the relatively contaminated Western Scheldt estuary pose little risk (∑TU=0.05; Chapter 5) for causing larval mortality related to maternal transfer of these pollutants. For other species including European eel, seriously reduced larval survival can be expected when the mother fish originates from highly contaminated locations. A simplified population model was applied to explore to what extent reduced larval survival may result in effects on population development of the common sole, in combination with and without (high) fishing pressure. Due to the high fecundity and the density dependent survival of juvenile stages that occurs after metamorphosis, the development of a sole population is insensitive to even very high larval mortality. Species that combine a low fecundity with density independent mortality of early life stages will be most sensitive to the type of mortality that can be expected as a result of maternal transferred lipophilic POPs.

Eggsposed Chapter 8
151
This chapter discusses the results presented in this thesis. First the results obtained in the current p-ELS test are discussed, also in comparison with the results that would (not) be observed in standard fish ELS tests with a shorter observation period. Next, the comparability of the chosen experimental set-up with the effects of maternally transferred POPs is discussed followed by the potential consequences of ELS mortality for population development of fish species. The characteristics that determine the vulnerability of a fish population are indicated plus some potentially sensitive fish species based on the most relevant characteristics identified in this thesis. Finally, in a concluding paragraph the research questions are addressed and future perspectives are discussed.
8.2 Observed effects in the prolonged ELS test
8.2.1 Timing of effects revealed with the p-ELS
With exception of HBCD, that induced no effects at any of the test concentrations, all compounds tested caused larval mortality that started a few days after yolk-sac absorption and continued in most cases until the onset of metamorphosis. The dose-related delayed mortality resulted from egg exposure only and occurred during further development of the larvae under unexposed conditions. Actually, mortality continued when the larvae already were free feeding and tissue concentrations of the toxicants already were declining due to growth dilution. The timing of the effects are highly reproducible (Chapters 2 and 5). At the lower test concentrations, this mortality occurred without initial indications for negative effects. Figure 8.1 shows the development in tissue concentrations of POPs depending on their log Kow in developing sole larvae exposed during the egg stage (6 days) in the p-ELS tests. The concentrations are as calculated with the ELS-OMEGA model (Chapter 4) and expressed relative to the concentration at 6 dpf. Pollutants with a log Kow of 6 and above are hardly excreted during the utilisation of the yolk lipids resulting in an exposure peak at the end of the yolk-sac stage. As a result the threshold level for toxicity can then be exceeded which can explain the timing of the observed mortality a few hours to days later (depending on the exposure concentration). Interestingly, similar timing of effects occurred after exposure to MTCS for which the ELS-OMEGA model (Figure 8.1) predicts no peak in toxicity at the end of the yolk-sac stage as this substance has a log Kow <5.5 (Valo and Salkinojasalonen, 1986). This suggests that MTCS may have some interaction with tissue proteins thus reducing the passive diffusion based on lipophilicity. For TCS it has been shown that it blocks the cellular efflux pumps (Anselmo et al., 2012b), thereby
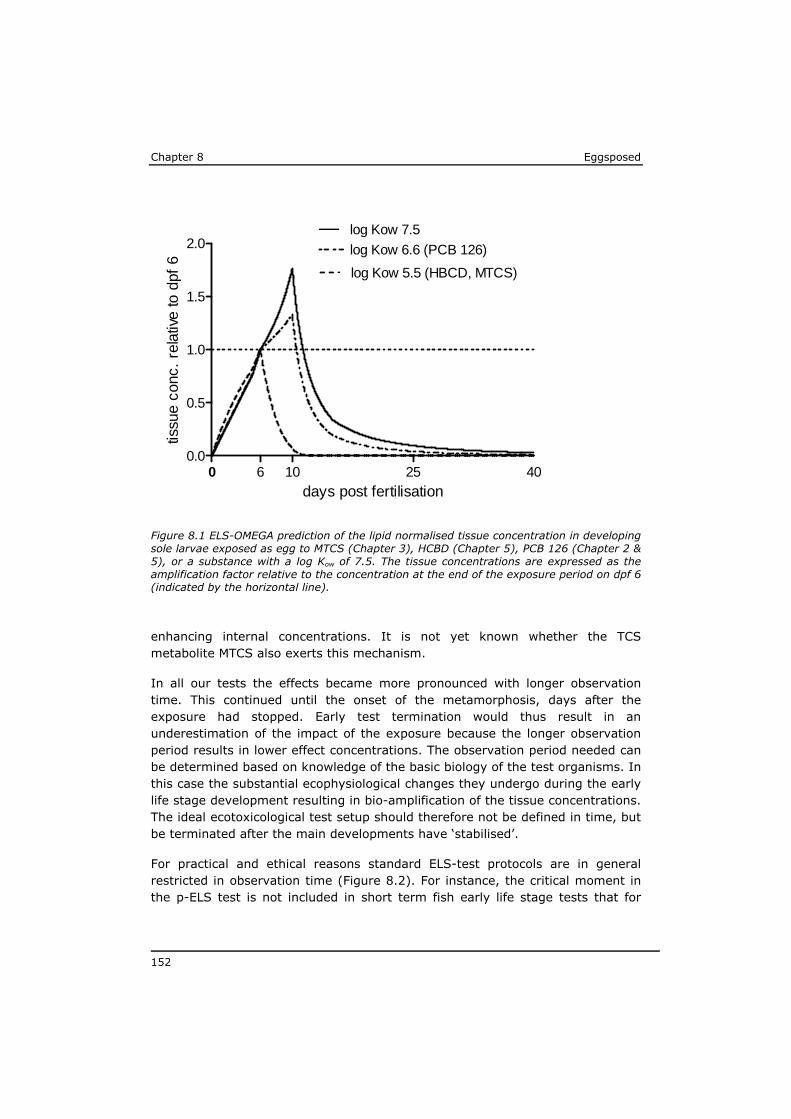
Chapter 8 Eggsposed
152
Figure 8.1 ELS-OMEGA prediction of the lipid normalised tissue concentration in developing sole larvae exposed as egg to MTCS (Chapter 3), HCBD (Chapter 5), PCB 126 (Chapter 2 & 5), or a substance with a log Kow of 7.5. The tissue concentrations are expressed as the amplification factor relative to the concentration at the end of the exposure period on dpf 6 (indicated by the horizontal line).
enhancing internal concentrations. It is not yet known whether the TCS metabolite MTCS also exerts this mechanism.
In all our tests the effects became more pronounced with longer observation time. This continued until the onset of the metamorphosis, days after the exposure had stopped. Early test termination would thus result in an underestimation of the impact of the exposure because the longer observation period results in lower effect concentrations. The observation period needed can be determined based on knowledge of the basic biology of the test organisms. In this case the substantial ecophysiological changes they undergo during the early life stage development resulting in bio-amplification of the tissue concentrations. The ideal ecotoxicological test setup should therefore not be defined in time, but be terminated after the main developments have ‘stabilised’.
For practical and ethical reasons standard ELS-test protocols are in general restricted in observation time (Figure 8.2). For instance, the critical moment in the p-ELS test is not included in short term fish early life stage tests that for
0.0
0.5
1.0
1.5
2.0
log Kow 5.5 (HBCD, MTCS)
days post fertilisation
tissu
e co
nc.
rela
tive
to d
pf 6
log Kow 6.6 (PCB 126)
log Kow 7.5
0 6 10 25 40

Eggsposed Chapter 8
153
practical reasons are terminated before the larvae become free feeding (OECD, 1998), or even before hatching (OECD, 2006). As this risk is recognised it is advised to perform what is called a full early-life-stage test (OECD guideline 210) when substances with log Kow >4 are being tested (OECD, 1998). However, OECD guideline 210, and the comparable EPA guideline OPPTS 850.1400 (USEPA, 1996) prescribe that this test continuous at least until the control fish are free feeding. Although in both guidelines recommendations are made for longer observation, this still leaves possibilities that delayed observable effects will be missed, and the toxicity of a substance is underestimated.
Insight in the mechanisms that determine the fate of the test compound in the tissue of the exposed organism through modelling (Chapter 4) helps to determine the minimal test duration based on the known characteristics of the substances to be tested. When mixtures with unknown composition are tested, like industrial effluents or extracts of tissues or contaminated sediments (Hallare et al., 2005; Rocha et al., 2011; Zielke et al., 2011) the observation period should best last until metamorphosis, to prevent underestimation of the risk posed by the most lipophilic substances (Log Kow >6).
8.2.2 (No) effects on thyroid hormone dependent metamorphosis
When starting this research it was expected that exposure to compounds like HBCD, PBDEs, and MTCS that are known for their potential to affect the thyroid hormone system (Anselmo et al., 2012a; Hinther et al., 2011; Legler, 2008; Palace et al., 2010; Yu et al., 2011), would result in disturbance of the sole’s metamorphosis. It is thyroid hormone mediated (Klaren et al., 2008), and has been shown to respond on manipulated thyroid hormone levels (Okada et al., 2005; Tagawa and Aritaki, 2005). However, none of the tests showed dose related effects on the onset or duration of the metamorphosis as has been shown for frogs or sea urchins (Gutleb et al., 1999; Anselmo et al., 2012a). Measurements of PCBs as model compounds and calculations with the ELS-OMEGA model revealed that at the onset of metamorphosis residue tissue concentrations only are a fraction of what they were earlier (Figure 8.1). Apparently, tissue concentrations were too low at the moment of metamorphoses to disturb the process.
It is likely that pollutants like PBDEs and MTCS that affect the thyroid mediated metamorphosis in other species under continuous exposure (Anselmo et al., 2012a), are also able to disturb the metamorphoses of flatfish when tissue concentrations are above a critical level when the organism becomes competent to start the metamorphosis. This tissue concentration must then be supported by external exposure. Maternal transfer of these pollutants on its own can never

Chapter 8 Eggsposed
154
result in such tissue concentrations during the metamorphosis without causing mortality in an earlier stage of development, as was shown by the results presented in Chapter 5, and indicated by the calculations with the ELS-OMEGA model. In field situation, external additional exposure after hatching could occur when larvae develop in an area with contaminated food.
8.2.3 Dioxin-like toxicity
The sole larvae appeared relatively sensitive for dioxin-like toxicity. Compared to the dataset compiled by (Steevens et al., 2005) consisting of 13 records covering 10 fish species with internal LC50 (ILC50) values ranging from 0.5 to 150 ng TEQ/g lw in eggs, sole belongs to the more sensitive species (ILC50 1 ng TEQ/g lw; Chapter 2).
A comparison of ELS test results did not reveal significant differences between TCCD ILC50 values for trout eggs that were exposed via the water phase, by nano-injection, or eggs that were produced by mother fish that were exposed during more than three months before spawning (Walker and Peterson, 1994; Johnson et al., 1998). This indicates that the TCDD exposure of the mother fish did not add to the effect of the TCDD that was deposited in the eggs. For various reasons this is not remarkable. First TCDD is hardly metabolised, which excludes parental transfer of metabolites as a factor. Due to its high teratogenicity, the direct effects on the early life stages overshadow the more subtle effects that could be caused by an affected parental condition. And finally, the effects of TCDD occur at a moment far after the tissue concentration via egg exposure is build up. This implies that the high comparability between the results of test with water exposed eggs and eggs from exposed parent fish that were found for TCDD does not necessarily apply for other pollutants.
It cannot be excluded that TCDD has an impact through epigenetic effects (King Heiden et al., 2009). However, the observation periods in the tests mentioned above (Figure 8.2), as well as in the p-ELS test were too short to be able to reveal these.
8.2.4 Baseline toxicity
Although it is difficult to separate the contribution of dioxin-like toxicity and baseline toxicity in the mortality caused by the PCBs and the mixtures, the data suggests that the sole p-ELS test is also relatively sensitive for base-line toxicity compared to other fish tests not restricted to early life stages (4 mmol/kg lw (Chapter 5) vs 36-300 mmol/kg lw (Kipka and Di Toro, 2009). This could indicate a special sensitivity to this type of toxicity for fish early life stages. This might be explained by the fast depletion of neutral lipids during the yolk-sac

Eggsposed Chapter 8
155
stage as energy source (Kamler, 2008), compared to the polar lipids that have a more structural function, for instance in cell membranes . Upon lipid mobilization neutral lipids were depleted three times faster than polar lipids in larvae of the Senegal Sole (Solea senegalensis ; Mourente and Vazquez, 1996). This will result in bioamplification of the lipophilic pollutants in the cell membrane lipids that are the target lipids for baseline toxicity (Escher and Hermens, 2002; Escher and Schwarzenbach, 2002). Although it does not show when tissue concentrations are normalised on total lipid content, the relative stronger concentration of lipophilic pollutants in the cell membranes could further enhance the impact of the toxicity peak at the end of the yolk-sac stage. Indeed, it has been recognised that models that in general are able to predict critical body burdens for a variety of aquatic species within acceptable ranges (Kipka and Di Toro, 2009), tend to underestimate the sensitivity of fish early life stages (Barron et al., 2004). The presence of specific toxic effects (Barron et al., 2004), or biotransformation of the mother compounds (Mathew et al., 2008) are mentioned as explanation for the deviation of the model prediction and experimental results. In both these studies that were performed with crude oils and early life stages of pacific herring (Carls et al., 1999) and pink salmon (Heintz et al., 1999), tissue concentrations were expressed on wet weight basis, and changes in the lipid content of the developing larvae were not determined. Also in the model calculations (Kipka and Di Toro, 2009) tissue lipid concentrations are assumed to be stable during larval development. Hence, although the presented explanations can very well be valid for a complex mixture as oil, it is likely that the changing lipid content and lipid composition that occurs during yolk absorption forms an additional factor explaining the relatively high sensitivity of fish early life stages to organic lipophilic compounds.
8.3 Simulation of maternal transfer in the p-ELS fish test
The p-ELS test procedure developed for this research intends to mimic, to a certain extent, maternal transfer of lipophilic POPs in order to study their effects on the development of an embryo into a juvenile fish. The metamorphosis into a juvenile fish was included in the test design as this process is thyroid hormone regulated and therefore potentially sensitive to thyroid hormone disrupting compounds (Okada et al., 2005; Tagawa and Aritaki, 2005).
The p-ELS revealed toxic effects that would have been missed with more standard ELS tests observing until free feeding. However, notwithstanding its advantages, also the p-ELS test cannot fully cover all effects of maternally transferred POPs on developing larvae.

Chapter 8 Eggsposed
156
The most reliable way to study the impact of maternally transferred pollutants on the developing offspring is by exposure of the parent fish; long term when effects on egg quality are to be included. This however, was not feasible given the resources available for this research. As alternative the possibilities of in-ovo injection (González-Doncel et al., 2003; Gonzalez-Doncel et al., 2003) were explored. But this extremely labour intensive procedure resulted in too high embryo mortality, probably related to a combination of the small egg size (max. 1 mm) and the very resistant chorion. Therefore it was decided to expose the eggs via the water until the moment of hatching, realising that this implied that the exposure situation in the test differs from real maternal exposure at least with respect to:
The time needed to reach the desired internal POP concentrations in the egg.
The absence of parental POP metabolites. The absence of impact of parental condition on egg quality. Non detectable epigenetic effects
These topics and their implications for the results of the p-ELS test are briefly discussed below.
8.3.1 Time needed to reach the desired internal POP concentrations in the egg
Figure 8.2 schematically shows the development of the lipid normalised POP concentration in an adult fish, in the produced eggs, and in the subsequent life stages. It is assumed that the POP concentration is transferred 1:1 from the mother’s tissue to the eggs (Russell et al., 1999). As can be seen the concentration peaks at the end of the yolk-sac stage, then declines due to growth dilution when the larva starts external feeding (Chapter 4). Finally the POP concentration gradually increases due to bioaccumulation/-magnification as the fish grows older.
Eggs that are produced by exposed female fish contain the POPs already before spawning and fertilisation. The p-ELS test starts with clean eggs, in which POP concentrations only start to build up after fertilisation. Exposure of early embryonic stages is thus low in the water exposed eggs compared to eggs produced by exposed parents. Hence, effects on fertilisation or very early embryonic development are not included in the p-ELS test.

Eggsposed Chapter 8
157
Figure 8.2 Schematic presentation of the tissue concentrations of POPs in eggs and subsequent life stages of fish in the field and in the p-ELS tests as applied in this thesis. The solid line indicates the development of POP levels, the dotted line of the possible concentrations of POP OH-metabolites. Below the observation periods covered in the p-ELS test and some standard Fish ELS tests are indicated.
During waterborne exposure the embryo is protected to a certain extent by the presence of the chorion, which limits exchange with the environment and so reduces the exposure of the embryo (González-Doncel et al., 2003; Gonzalez-Doncel et al., 2003). This could be overcome by enzymatic dechorionation of the eggs (Mizell and Romig, 1997; Usenko et al., 2011). Comparative studies show effects at lower water concentrations for dechorionated eggs (Villalobos et al., 2000), or for eggs that were exposed before fertilisation and consequent chorion hardening (González-Doncel et al., 2003; Gonzalez-Doncel et al., 2003). In our test set-up, effect concentrations were based on chemical analyses of the larvae immediately after hatching. The impact of the protective chorion thus did not affect the effect concentrations that were expressed as tissue concentration of the embryo.
Waterborne exposure resulted in a clear dose related larval accumulation of the lipophilic pollutants that were subject of the experiments presented in this thesis, MTCS, PCBs, PBDEs, and HBCDs. However, we have clear indications that

Chapter 8 Eggsposed
158
waterborne exposure of eggs with intact chorion does not work with surface active compounds. Despite extremely high exposure levels of perfluorooctanesulfonate (PFOS) during the egg stage, no effects on the developing sole larvae were recorded (unpublished data), while in a study with parental exposure to the same compound clear effects on the developing offspring of zebrafish were observed (Wang et al., 2011). It is to be expected that the surface active PFOS concentrates on/in the surface of the egg shell during exposure without penetrating the chorion. For this type of pollutants actual maternal transfer is necessary to deliver it inside the egg.
8.3.2 Absence of parental POP metabolites
In the p-ELS test eggs were exposed only to the parent compound. With this, it is ignored that some POPs can be transformed by fish into often more toxic metabolites that will also be deposited in the eggs bound to the deposited proteins. This is the case for halogenated phenolic compounds (HPCs) including the metabolites of PCBs (OH-PCBs) and PBDEs (OH-PBDEs). These metabolites are present in plasma of many organisms, including fish, at concentrations that exceed no effect levels in in-vitro assays (Montano et al.,2013). HPCs show a structural resemblance to thyroid hormones, and are in this way actively transported to the eggs. There, OH-PBDEs can induce developmental arrest in a concentration-dependent manner as was shown after exposure of dechorinated eggs of zebra fish (Usenko et al., 2012). Maternal transfer will probably be an important exposure route for these metabolites.
Juveniles of the common sole have been shown to form hydroxylated metabolites after PBDE exposure (Munschy et al., 2010), it is not yet known in what phase of development this ability occurs. Exposure of zebrafish embryos to PBDE47 did not result in formation of OH-PBDE47 (Zheng et al., 2012). Although species specific differences cannot be excluded (Roberts et al., 2011), these results might indicate that the ability for hydroxylation in fish develops somewhere between hatching and the juvenile stage. As biotransformation is a mechanism to protect an organism against the accumulation of poorly water soluble (natural) substances in their diet, it seems reasonable to assume that this ability will develop when the larvae starts external feeding. In the natural situation hydroxylated POPs are already present in the egg before fertilisation via maternal transfer (Figure 8.2, dotted line). In the p-ELS test, the metabolites only can be present after the larva itself has developed the capability for biotransformation of the mother compound, thus during the free-feeding stage. As a consequence the p-ELS test underestimates the potential impact of these metabolites compared to the natural situation.

Eggsposed Chapter 8
159
8.3.3 Impact of parental condition on egg quality
Thyroid hormones (T3 and T4) are deposited in eggs by the mother fish (Tagawa et al., 1990; Yamano, 2005). Altered thyroid hormone levels in parent fish can be reflected in the eggs, as shown after parental exposure of zebra fish with PBDEs (Yu et al., 2011). Since thyroid hormones are deposited in considerable amounts in fish eggs a critical role in early development of the embryo can be expected (Tagawa et al., 1990). Administration of thyroid hormones to eggs or newly hatched larvae sometimes results in better survival and development (Yamano, 2005). The effects of hydroxylated POP metabolites are mostly related to the blocking of the TH transport proteins and blocking or activating TH receptors (Montano et al., 2013). Lowering of TH levels will only become a problem when the reduction is extreme, as otherwise the strong feedback mechanisms in the TH homeostasis is able to compensate. For example in medaka eggs that contained only 10% of the normal total TH levels due to chemical repression of the thyroid hormone production by the mother fish, no effects on hatching, survival and development was observed (Tagawa and Hirano, 1991). The level of the active thyroid hormone (T3) probably still was high enough, but at that time this could not be analysed separately.
Finally, impact of exposure of parents on ovary histology (Daouk et al., 2011; Mochida et al., 2010), sperm quality (King Heiden et al., 2009; Dietrich et al., 2010) or behaviour that could affect reproduction success are not included in our p-ELS test set-up.
8.3.4 Not detectable epigenetic effects
Toxic compounds can induce effects that only become visible later in life, e.g. when certain functions become relevant such as reproduction (Vandegehuchte and Janssen, 2011). Early life exposure even can lead to effects on the next generation as has been shown with zebra fish that were exposed during the first 7 weeks of their life to TCDD (King Heiden et al., 2009). The second generation that could only have been exposed as gametes, showed increased mortality, reduced egg production, and reduced fertility. This suggests an transgenerational epigenetic effect (Youngson and Whitelaw, 2008) of the parental exposure. Epigentic effects are the result of changes in gene function without changing the DNA sequence, with DNA methylation as the most studied phenomenon. Evidence for epigenetic effects on among others fish, has been reported for pollutants as Benzo(a)pyrene, mercury, tributyltin, triphenyltin, zinc, cadmium, oestradiol and HBCD (reviewed in Vandegehuchte and Janssen, 2011. In some cases these alterations have been shown to be inheritable to the next generation (Vandegehuchte et al., 2010). In these situations the overall

Chapter 8 Eggsposed
160
impact of such a pollutant would be underestimated when based only on observation of development until metamorphosis of larvae exposed as eggs.
Some situations are described where chronic exposure has resulted in resistance of a population for a specific stressor. This was shown for fish populations from field locations with high concentrations of Ah-receptor agonists such as dioxin-like PCB and PAHs. Apparently the negative effects from these AhR active compounds were so strong that there was a fitness/evolutionary advantage for losing the activity of the physiologically relevant AhR. The developed resistance to the effects of these pollutants was genetic and transmitted to at least the F2 generation (Wirgin and Waldman, 2004; Wirgin et al., 2011).
8.4 Characteristics that determine sensitivity
Specific characteristics can make a species more vulnerable to the effect of maternally transferred POPs than others that share the same environment. This is the resultant of differences in toxicokinetic, toxicodynamic and population dynamic characteristics. The following paragraphs discuss these characteristics, illustrated with some examples.
8.4.1 Toxicokinetic and toxicodynamic characteristics
Due to biomagnification maximum POP concentrations are to be expected in the tissue of top predators and long living species (Moore et al., 2002), this implies that female top predatory fish deposit the highest POP loads in their eggs. As lipid normalised concentrations of POPs in the tissue of the mother fish will in general be comparable with her eggs (Chapter 5; (Russell et al., 1999), it is the lipid normalised POP concentration that determines the exposure of the eggs. This means that lipid-rich fish species that have high POP-levels per g of filet (as is relevant for human consumption fish) could transfer lower POP loads to their eggs than lean fish (such as e.g. cod) with lower POP levels per g of filet but high levels per g of lipid.
Some fish species like eels and salmon undertake a spawning migration, during which they cease feeding. As the fish loses weight during migration, POPs accumulated in the tissue will concentrate. Bio-amplification factors of 2 to 8 in gonads of pacific salmon (Debruyn et al., 2004), and of 2 to 5 for sockeye salmon (Ewald et al., 1998) have been established. Compared to these values, the amplification factor calculated for European eel of 1.3 to 2 (Chapter 6) is relatively low. This is related to the rather unique fact that the tissue composition of the eel does not change during migration. Lipids, protein and carbohydrates are utilised as energy source in the same proportion as they are

Eggsposed Chapter 8
161
present in the tissue (van Ginneken et al., 2005). Similar use of bodily tissues during migration has been reported for some reef fish. But in most fish species proteins are conserved, resulting in an clear decrease of the lipid content of the tissue (Stobutzki, 1997). In all cases migration can be expected to result in higher POP concentrations in the eggs.
Once the POPs have been deposited in the egg, they can affect the developing larvae. The sensitivity of the larvae, and thus the impact of the transferred POPs can vary between species. This is due to differences in biochemistry and (eco)physiology between species. It is this interspecies variation that forms the bases of the species sensitivity distribution (SSD) theory that is applied to estimate the critical concentration of a toxicant for causing effects on the community level (Posthuma et al., 2010 ). Normally these SSDs are based on external exposure concentrations (eg. Selck et al., 2002; Smit et al., 2009). This implies that interspecies variation in these datasets is not only determined by variation in the toxicological response of the organism once the toxicant reaches the target organs, but also by variation in internal exposure that can depend on the biological/physiological characteristics of the species. By expressing the effects on internal tissue concentrations during the exposure peak, the latter source of variation is excluded. In the SSD as produced by Steevens et al.(2005) the concentration in the yolk was used as measure of exposure, as was the case in this thesis when determining the critical tissue concentration of TCDD for fish early life stages (Chapter 6). Still, the variation in sensitivity between species (expressed as ILC50) ranges from 0.5 to 150 ng TCDD/g lw. Although already expressed on lipid bases, the effect concentrations shows a strong correlation (p<0.0001) with the egg’s lipid content (Figure 8.3). The lower effect concentrations of larvae from eggs with higher lipid content is likely to be related with stronger bio-amplification of the TCDD at the end of the yolk-sac stage (Chapter 4; Daley et al., 2012) when the majority of the neutral lipids is used as energy source. The larvae that hatch from eggs with high lipid content can thus be considered more sensitive, because they are higher exposed, to the impact of maternally transferred lipophilic POPs. It is interesting to note that zebra fish, a standard species in fish ELS tests, shows the lowest sensitivity to TCDD in this dataset (Elonen et al., 1998; Steevens et al., 2005), which is in accordance with the lowest lipid content (1.7%) for zebra fish of all the eggs included.
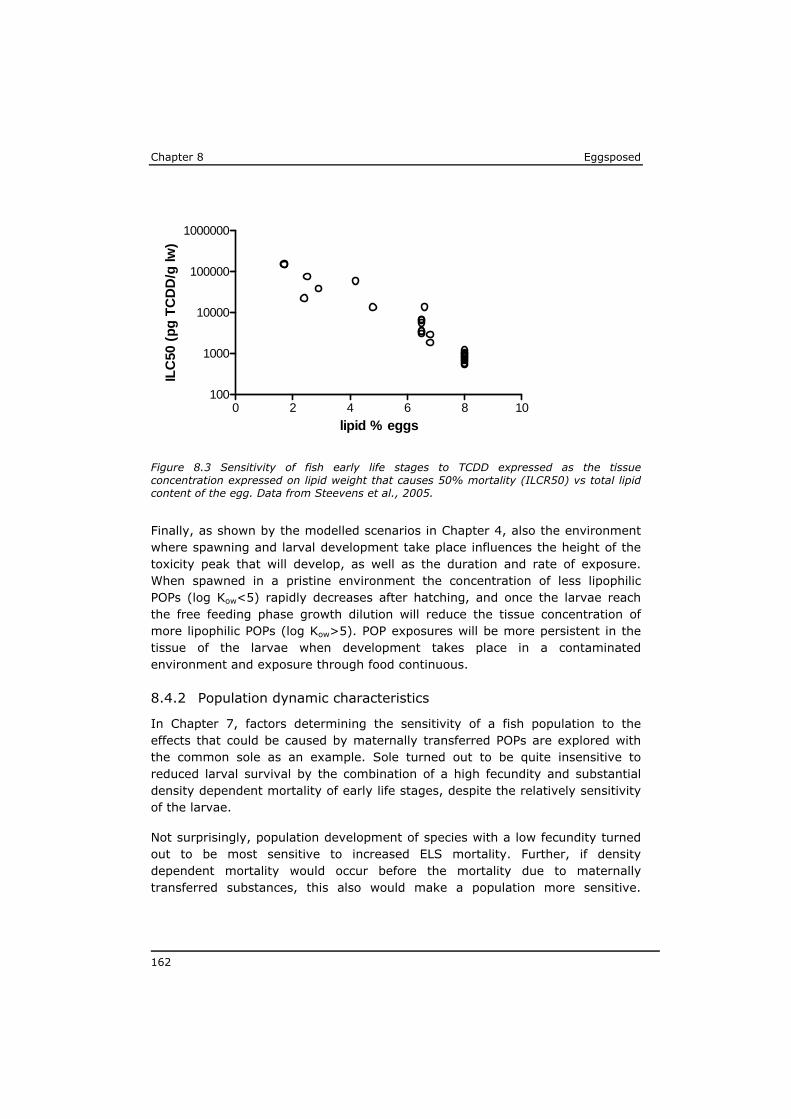
Chapter 8 Eggsposed
162
Figure 8.3 Sensitivity of fish early life stages to TCDD expressed as the tissue concentration expressed on lipid weight that causes 50% mortality (ILCR50) vs total lipid content of the egg. Data from Steevens et al., 2005.
Finally, as shown by the modelled scenarios in Chapter 4, also the environment where spawning and larval development take place influences the height of the toxicity peak that will develop, as well as the duration and rate of exposure. When spawned in a pristine environment the concentration of less lipophilic POPs (log Kow<5) rapidly decreases after hatching, and once the larvae reach the free feeding phase growth dilution will reduce the tissue concentration of more lipophilic POPs (log Kow>5). POP exposures will be more persistent in the tissue of the larvae when development takes place in a contaminated environment and exposure through food continuous.
8.4.2 Population dynamic characteristics
In Chapter 7, factors determining the sensitivity of a fish population to the effects that could be caused by maternally transferred POPs are explored with the common sole as an example. Sole turned out to be quite insensitive to reduced larval survival by the combination of a high fecundity and substantial density dependent mortality of early life stages, despite the relatively sensitivity of the larvae.
Not surprisingly, population development of species with a low fecundity turned out to be most sensitive to increased ELS mortality. Further, if density dependent mortality would occur before the mortality due to maternally transferred substances, this also would make a population more sensitive.
lipid % eggs
ILC
50 (
pg
TC
DD
/g lw
)
0 2 4 6 8 10100
1000
10000
100000
1000000

Eggsposed Chapter 8
163
Maternally transferred lipophilic POPs however as we tested, resulted in larval mortality in an early stage of the development, and thus before most density dependent mechanisms become effective, that are often related to increased predation pressure or competition for food and/or space.
8.5 Identification of vulnerable species
With the insight in the critical characteristics presented above it is possible to predict the vulnerability of species for the effects of maternally transferred POPs.
The most vulnerable species combine:
1. A position high in the food chain or a narrow food choice of relatively highly exposed food items facilitating biomagnification of POPs.
2. A period of starvation prior to spawning, for instance during spawning migration, resulting in bio-amplification of accumulated POPs.
3. Eggs with high lipid content, resulting in strong bio-amplification of POPs at the end of the yolk-sac stage.
4. A low fecundity. 5. Density dependent mortality or concentration of early life stages after
increased ELS mortality due to maternally transferred substances.
As an example, some of these characteristics are listed in Table 7.1 for selected marine fish species. This is not an exhaustive overview but an impression based on readily available data on fecundity of fish species collected by Mertz and Myers (1996). Only a limited number of orders are included, sometimes with only a single species per order. The dataset, for instance does not include large predatory fish like tuna and swordfish, known for high bioaccumulation of POPs (Kannan et al., 2002; Storelli et al., 2008).
Within this dataset the lowest fecundity is found in the order of Squaliformes (sharks). In combination with a diet that mainly consists of fish and higher organisms this makes this order theoretically vulnerable for effects of maternally transferred POPs. Salmonids also have a low fecundity. As facultative predators, their place in the food chain is in general lower than sharks, but many salmonid species undertake a spawning migration without feeding. The only other order in this dataset that undertakes such a migration is the Anguilliformes. The estimation of the vulnerability on the individual level is described for the European eel in Chapter 6. In the following paragraphs an attempt is made to determine the sensitivity for effects on a population level of this species, as well as for salmonids and sharks.
Chapter 8 Eggsposed
164
Table 8.1 Some characteristics that determine the vulnerability of fish species for the effects of maternally transferred POPs. Presented are the range (min-max) in fecundity based on the available data for that order, the species with the lowest fecundity per order, the main diet of that species and whether this species undertakes spawning migration without feeding. Fecundity data from Mertz and Myers (1996) and expressed per individual fish year. Data for Anguiliformes were taken from MacNamara and McCarthy (2012), as fecundity per fish and divided by 10 in order to calculate fecundity per year assuming a 10 year lifespan for eels. The fecundity data for Lophiiformes and Squaliformes consisted only of a single species. Diet information from sources as indicated.
Order
range fecundity x103/ind/yr Species with lowest fecundity Diet Migration
Lophiiformes 1320 Lophius piscatorius (Monkfish) Fish (Laurenson and Priede, 2005) No
Pleuronectiformes 80-2500 Pleuronectes platessa (Plaice) Benthos polychaetes (Schuckel et al., 2012) No
Gadiformes 79-9500 Merluccius productus (Pacific hake) Fish (Helser and Alade, 2012) No
Anguilliformes 70-3480 Anguilla anguilla (European eel) Opportunistic macrofauna – fish Yes
Clupeiformes 25-335 Engraulis sp (Anchovy) Zooplankton (Bacha and Amara, 2012) No
Perciformes 15-1000 Ammodytes marinus (Sandeel) Zooplankton (Eliasen et al., 2011) No
Salmoniformes 1.5-40 Oncorhynchus gorbuscha (Pink salmon)
Opportunistic zooplankton- fish (Ruggerone and Nielsen, 2004) Yes
Squaliformes 0.003 Squalus acanthias (Spiny dogfish) Fish (Dunn et al., 2013) No
8.5.1 European eel
The European eel (Anguilla Anguilla) is often mentioned as a species sensitive to the effect of organic contaminants (Belpaire et al., 2009; Clevestam et al., 2011; Geeraerts and Belpaire, 2010; Geeraerts et al., 2011; Palstra et al., 2006; Robinet and Feunteun, 2002; van Ginneken et al., 2009). This is primarily based on the relatively high contents of POPs that can be detected in the lipid rich tissue of the eel in comparison with other species from the same area, and the typical life history making this species potentially vulnerable to accumulating substances. For this reason an estimation of the chance that larvae of the European eel will be negatively affected by maternally transferred POPs was performed in Chapter 6. We assumed a high sensitivity of the larvae to dioxin like compounds based on a high lipid content of the eggs, and calculated a reasonable amplification of the accumulated POPs during spawning migration.

Eggsposed Chapter 8
165
The results indicate that 50% larval mortality is likely for eel from highly contaminated locations, due to maternally transferred dioxin-like pollutants (Chapter 6).
Since only limited knowledge is available about the reproductive biology of the European eel it is unclear to what extend the eel population is able to compensate for increased larval mortality. Eels have a very high fecundity ranging from 1600-3600 eggs/g body weight (MacNamara and McCarthy, 2012). However, it must be taken into account that eel most likely is a semelparous species, indicating that it only spawns once in a lifetime (Palstra and van den Thillart, 2010). Assuming an average life span of 10 year, the fecundity ranges between 160 and 360 eggs/g body weight per year. This is lower than for instance sole, that as fish with an iteroparous life strategy produces very year about 800 eggs/g body weight (Rijnsdorp et al., 1992).
Based on modelling studies it is estimated that it will take the larvae 1 to 3 years to reach the European coast (Righton et al., 2012). High mortality, due to natural causes can be expected to occur during this migration. Whether also density related mortality occurs is unclear. It seems reasonable to assume that some concentration of the glass eels occurs when they leave the ‘infinite’ ocean to enter the fresh water environment. This concentration thus takes place long after the moment that larval mortality due to maternal transferred POPs would occur. The relative high fecundity in combination with concentration of early life stages may result in not too high vulnerability of the population development for reduced larval survival. But whether in addition to high glass eel fishing mortality (Dekker, 2004) also density dependent natural mortality occurs is unknown.
Chapter 6 indicated that a strong negative impact of maternally transferred dioxin-like POPs can be expected for eels from highly contaminated locations. Effects of other pollutants like polyfluorinated compounds (PFCs), and halogenated phenolic compounds (HPCs) were not included in this study as they have completely different toxicokinetics than the lipophilic POPs. PFCs however, have been detected in high concentrations in eel tissue (Kwadijk et al., 2010), and can cause effects by maternal transfer (Wang et al., 2011). HPCs including OH-PCBs and OH-PBDEs have been identified in the blood of fish at levels that exceed effect concentrations in in-vitro assays, and can also be maternally transferred (Montano et al., 2013 in press)). In addition, early life exposure to dioxins could result in transgenerational epigenetic effects (King Heiden et al., 2009). Finally, toxic substances can have adverse effects on juvenile and adult fish, especially eel migration and reproduction could be negatively affected by for reduced fitness of the parent fish (Belpaire et al., 2009; Clevestam et al.,

Chapter 8 Eggsposed
166
2011; Geeraerts and Belpaire, 2010; Geeraerts et al., 2011; Robinet and Feunteun, 2002; van Ginneken et al., 2009).
The European eel that inhabits the European and North African waters form one population that all share the same spawning grounds (Als et al., 2011). As indicated in Chapter 6, a part of this population probably experiences reduced larval mortality due to maternally transferred dioxin-like POPs, but the exact proportion of the impact of these and other pollutants on the population remains unclear. It is therefore uncertain whether pollutants contribute to the decline of the eel population that has been observed since the 1980 (Dekker, 2003).
8.5.2 Salmonids
The impact of POPs on the condition and reproduction of salmonids has been studied for a long time. In the past the majority of these studies focussed on the North American Great Lakes and indications were found for reduced reproduction as a consequence of bioaccumulated dioxin-like POPs (Giesy et al., 2002).
A substantial part of the salmonid species is anadromous, thus migrate from salt to freshwater to spawn. The Atlantic salmon (Salmo salar) returns to sea after spawning, but all pacific salmon species (Oncorynchus sp.) are semelparous and die at the spawning grounds. As in this semelparous life strategy it is not essential to save energy for the migration back to sea, energy and thus lipid resources are strongly depleted when spawning grounds are reached (Wootton, 1992). This results in relatively high bio-amplification factors in the fish. For sockeye salmon (Oncorhynchus nerka) reported bio-amplification factors ange from 2 to 12 (Ewald et al., 1998; Debruyn et al., 2004; Kelly et al., 2007) which correlates positively with the migration distance (Debruyn et al., 2004; Kelly et al., 2007). Although it is expected that in an equilibrium situation the lipid normalised concentrations of POPs in tissue and eggs are comparable (Russell et al., 1999), concentrations were 50 to 20% lower in eggs than in tissue of salmon that had reached the spawning grounds. Possibly the redistribution of especially the more lipophilic substances during the migration is not fast enough to reach an equilibrium between bodily tissues including the eggs (Kelly et al., 2007). Hence, a 1 to 1 translation from tissue to egg after spawning migration, most likely overestimates the concentrations in the eggs. Nonetheless chemical analyses revealed that the concentration of dioxin-like pollutants in the gonads of the sockeye salmon still concentrate more than 3 times during migration (Debruyn et al., 2004; Kelly et al., 2007).
Salmonids produce relatively large eggs with high lipid contents, ranging from 6.5 to 8.6% for brook trout, lake trout, rainbow trout and Chinook salmon

Eggsposed Chapter 8
167
(Johnson et al., 1998; Walker and Peterson, 1994; Ballestrazzi et al., 2003; Daley et al., 2012). These large lipid rich eggs supply the developing embryos during the long egg and yolk-sac stages that can last up to several months (Milner et al., 2003). Especially after hatching, yolk lipids are applied as energy source for the developing larvae. Eggs with an initial lipid content of 8.6% produced larvae with a lipid content of 1.2% at the end of their yolk-sac stage 168 days post fertilisation. During this period POPs with log Kow >5.8 showed a 5 fold bio-amplification (Daley et al., 2012). Since dioxin-like POPs in fish all have a log Kow above 5.8, the lipid normalised concentrations of dioxin-like compounds at the end of the yolk-sac stage can be more than 15 times higher than in the maternal concentration at the start of the spawning migration. Measured concentrations in roe from salmon returning to the Frazer river in some cases exceeded the threshold level above which salmonid larval mortality can be expected (Kelly et al., 2011).
Salmonids have a relatively low fecundity (Table 7.1) which makes them potentially sensitive for increased larval mortality. The populations are regulated by density-dependent mortality that takes place typically during the early stages of free-feeding life after fry emerge from spawning gravels (Elliott, 1989) and display aggressive, territorial behaviour (Milner et al., 2003). Larval mortality due to maternally transferred POPs would occur during the same period, thus reducing the pressure of competition for space. It is to be expected that in a healthy population a surplus of larvae is produced so larval mortality due to toxic compounds will mostly relieve the competition. However, when salmon stocks are minimised due to overexploitation or migration barriers the impact of reduced larval mortality may become relevant.
8.5.3 Greenland shark, the worst case scenario?
The highest levels of dioxin-like pollutants in tissues of shark species were reported for the Greenland shark (Somniosus microcephalus) and ranged from 460 to 579 pg TEQ/g lw based on the TEF system for fish (Storelli et al., 2011). These tissue concentrations in adult fish fall within the range where at least 50% larval mortality may be expected for 5 to 10% of all teleost fish species (Chapter 6). No other fish species from the Atlantic ocean (17-88 pg TEQ/g lw), other top predator fish like swordfish (9 pg/g lw) or dolphin species from the Mediterranean (42-270 pg TEQ/g lw) have such high tissue concentrations of dioxin-like pollutants (Storelli et al., 2011).
Also other POP concentrations are extremely high in Greenland shark tissue. PCB concentrations in liver and muscle have been reported to range from 990 to 10 000 ng/g lw (Strid et al., 2007), while mean concentrations of DDT and

Chapter 8 Eggsposed
168
chlordane pesticides in livers of 7159 and 1815 ng/g lw respectively have been reported (Fisk et al., 2002). Sum PBDE concentrations range between 7-200 ng/g lipid, with means of 35 and 41 ng/g lw for liver and muscle respectively (Strid et al., 2010). The POP concentrations in Greenland sharks are amongst the highest reported for an Arctic fish and are in the range of top Arctic marine predators (MacNeil et al., 2012). Lipid normalised concentrations of PCBs, DDT and other chlordane pesticides in plasma of Greenland sharks were even higher than the values reported for plasma of polar bears and white whales from the same region (Svalbard, Norway; Molde et al., 2013). It reflects the diet of the Greenland shark that for substantial part consists of marine mammals (Yano et al., 2007; Fisk et al., 2002; Leclerc et al., 2012).
Relatively little is known about the biology of Greenland sharks (Leclerc et al., 2012). The species is found from the North Atlantic Ocean to the Arctic Ocean, but given their tolerance for extremely cold water (up to -2ºC) and great water depths (>2000 m) the distribution range is potentially unlimited throughout the deep sea (MacNeil et al., 2012). Greenland sharks are considered to have a low growth rate, and females mature only at a length above 450 cm (Yano et al., 2007). As many shark species, Greenland sharks are ovoviviparous; the eggs are fertilised and hatch inside the female's body. After absorption of the yolk-sac, the young are believed to feed on non-developed eggs and each other. In one bearing up to 10 young are born (Yano et al., 2007) as fully developed juveniles with a length probably ranging from 35 to 100 cm (MacNeil et al., 2012).
As the developing embryo/larva rely on their yolk for resources, the development of a toxic peak at the end of the yolk-sac stage is likely. Unlike oviparous fish, the shark eggs/larvae do not develop in relatively clean seawater. Hence, passive excretion that would prevent the development of a toxic peak for POPs with log Kow <6, is much lower for the ovo-vivi-parous sharks than for ovo-parous fish. Not only will this result in a higher bio-amplification factor at the end of the yolk-sac stage, but in addition it is likely that brominated flame retardants like HBCD and PBDEs with the potential to disrupt the thyroid hormone system maintain relatively high levels during the full early life development, whereas in e.g. sole larvae the concentrations of these substances rapidly decline. In addition, after yolk absorption the young sharks do not feed on less polluted zooplankton as most other fish, but first use undeveloped eggs and each other as food source. Given the tissue concentrations of the Greenland shark this must be the most contaminated food source that can be found.

Eggsposed Chapter 8
169
The additional impact of these adverse ecophysiological parameters on the development and survival of the young is not known, but it adds to the estimated 50% larval mortality from the dioxin-like toxicants only, as explained above. Here we assume that the shark embryos have a comparable sensitivity for dioxin-like compounds as other fish species. However, being a member of another infraclass of fish, the Chondrichthyes, for which the effects of dioxin-like compounds on the early life development has not been studied yet, it is not known how the sensitivity relates to that of the well- studied fish species. There are indications that the vitamin A and E systems of Greenland sharks are affected by high levels of POPs (Molde et al., 2013), which could result in reduced quality of the eggs, and thus add additional pressure on the developing larvae.
As Greenland sharks still are relatively abundant (Leclerc et al., 2012) it is apparently able to compensate for this toxic pressure. Favourable is that the species experiences relatively little fishing pressure, as the flesh is toxic for mammals for reasons that are not completely clear, but are most likely related to specific natural metabolites (MacNeil et al., 2012). Only for the production of oil from the sharks liver some extensive fisheries took place during the first half of the 20th century. Nowadays fishing mortality is limited to accidental by-catches. There is, however, concern about the impact of global warming for this cold water species. Even if the warming of their habitat does not directly affect the physiology of the species, it could lead to increasing fishing pressure with the retraction of the ice cover resulting in northward migration of commercial fish stocks. With its slow growing life-history, S. microcephalus may not be able to cope with increasing losses through by-catches (MacNeil et al., 2012) in combination with reduced survival of early life stages due to the high exposure to POPs.
8.6 Conclusions
To conclude the answers to the research questions posed in Chapter 1 are briefly summarised below.
1. What are critical egg concentrations for PCBs, PBDEs, HBCDs and MTCS for larval development, and how do these relate to field (fish) concentrations?
Critical egg concentrations were determined for all substances except for HBCD that did not cause observable effects in our test. With exception of dioxin like substances at highly contaminated locations, the reported tissue concentrations in fish are lower than the critical effect concentrations for larval survival.

Chapter 8 Eggsposed
170
2. Do the above mentioned (groups of) substances represent the major risk in case of maternal transfer, or are there indications for substantial impact of other (unknown) substances?
The highest risk is revealed for dioxin-like substances and very lipophillic substances that can induce narcosis. No indications were found that other lipophillic POPs extracted from Dutch sole substantially contribute to the reduced larval survival due to maternally transferred POPs.
3. What is the influence of ecophysiological characteristics on the impact of maternally accumulated compounds transferred to the eggs on the developing larvae?
Both spawning migration and high lipid content of eggs contribute to bio-amplification of lipophilic compounds to levels in the larvae that are higher than what they will ever reach during adult life. Through depletion of the yolk lipids maternally transferred lipophilic POPs with a log Kow around 6 and above concentrate in the remaining lipids, resulting to higher available concentrations and higher concentrations in cell membranes. As a result larval mortality starts at the end of the yolk sac stage and commences until the onset of metamorphosis. Due to growth dilution and passive excretion concentrations of maternally transferred substances have then reached such low tissue concentrations that the metamorphosis is not affected.
4. To what extent can the larval mortality from maternally transferred toxicants affect population development, possibly in combination with high fishing pressure?
The combination of high fecundity and density dependent survival of early life stages make most fish species capable to compensate for large variations in early life mortality. Since the effects of the maternally transferred substances studied occur in a stage when these density dependent mechanisms are not yet acting, the impact is limited, even in combination with extreme fishing pressure. Species with low fecundity and less density dependent regulation of population growth can be considered potentially vulnerable. It cannot be excluded that effects of maternally transferred substances that act at a later moment in life will have more impact on the population development.
8.7 Future perspectives
This thesis revealed the effects of maternally transferred dioxin-like compounds on the survival of fish larvae between hatching and metamorphosis. For some

Eggsposed Chapter 8
171
fish species, a significant effect on larval survival is to be expected. To be able to assess the full risk of POPs accumulated in the tissues of these fish, the effects on other important life cycle parameters need to be assessed. In particular effects on adult survival during spawning migration, and on fertility and egg quality are important to study. Also toxic effects of POPs that were not included in this study because their maternal transfer cannot be mimicked via water exposure deserve further attention. For example PFCs and OH-POPs should either be injected in the developing egg or exposure should take place via the mother animal. Exposure via the mother animal also offers the advantage of including effects on egg quality and on the very early embryonic development in the egg.
Further, it is important to investigate if the larvae that develop without obvious adverse effects in normal juveniles after early life exposure to POPs, do not experience negative effects at a later moment for instance through trans generational epigenetic effects (Vandegehuchte and Janssen, 2011; Wirgin and Waldman, 2004; Youngson and Whitelaw, 2008). To predict such delayed effects biomarker techniques could be applied that already can indicate sub-lethal changes in an early stage. Focus of such studies could be effects that reduce reproductive performance, hormone disruption, behavioural changes (Mennigen et al., 2011; Soffker and Tyler, 2012) or age of maturation (Hutchings et al., 2012).
The sensitivity of sharks, especially the Greenland shark, deserves more attention. Little is known about the sensitivity of early life stages of shark species, and more specifically on ovoviviparous species. Further, the insights gained in the characteristics what makes species vulnerable to maternally transferred substances can be applied to other orders and species that were not included in the exploration presented above. New potentially vulnerable species can be identified and for targeted species, population modelling can be applied to identify the most critical life phase for population development. Given the high potency of most fish populations to compensate for high early life mortality through high fecundity and density dependent survival mechanisms of the early life stages, population development probably will be more affected by toxic substances that impair the condition or reproductive efficiency of mature fish. For such species, it is recommended to focus future research on adult fish, including impact of age of maturation (Hutchings et al., 2012), hormone disruption, behavioural changes (Mennigen et al., 2011; Soffker and Tyler, 2012), and (transgenerational) epigenetic effects (Vandegehuchte and Janssen, 2011; Wirgin and Waldman, 2004; Youngson and Whitelaw, 2008).

Chapter 8 Eggsposed
172
Besides toxicants, additional stressors should also be included in the assessment of the sensitivity of species, as an organism will always experience multiple stressors. Relevant additional stressors for fish can for instance be fishing pressure, global warming, ocean acidification, loss of specific habitat like spawning ground or nursery areas, or the ability to reach these, and competition with invasive species may make species more vulnerable to toxicological effects that would be far less relevant in otherwise less impacted conditions.
8.8 References
Als, T.D., Hansen, M.M., Maes, G.E., Castonguay, M., Riemann, L., Aarestrup, K.I.M., Munk, P., Sparholt, H., Hanel, R., Bernatchez, L., 2011. All roads lead to home: panmixia of European eel in the Sargasso Sea. Molecular Ecology 20, 1333-1346.
Anselmo, H.M.R., Koerting, L., Devito, S., van den Berg, J.H.J., Dubbeldam, M., Kwadijk, C., Murk, A.J., 2012a. Early life developmental effects of marine persistent organic pollutants on the sea urchin Psammechinus miliaris (vol 74, pg 2182, 2011). Ecotoxicology and Environmental Safety 80, 401-401.
Anselmo, H.M.R., van den Berg, J.H.J., Rietjens, I., Murk, A.J., 2012b. Inhibition of cellular efflux pumps involved in multi xenobiotic resistance (MXR) in echinoid larvae as a possible mode of action for increased ecotoxicological risk of mixtures. Ecotoxicology 21, 2276-2287.
Bacha, M., Amara, R., 2012. Inter-cohort differences in growth, condition and feeding of juvenile anchovy (Engraulis encrasicolus) in the Gulf of Bejaia (Algerian coast, SW Mediterranean): Implications for recruitment success. Fisheries Research 129, 73-81.
Ballestrazzi, R., Rainis, S., Tulli, F., Bracelli, A., 2003. The effect of dietary coconut oil on reproductive traits and egg fatty acid composition in rainbow trout (Oncorhynchus mykiss). Aquaculture International 11, 289-299.
Barron, M.G., Carls, M.G., Heintz, R., Rice, S.D., 2004. Evaluation of fish early life-stage toxicity models of chronic embryonic exposures to complex polycyclic aromatic hydrocarbon mixtures. Toxicological Sciences 78, 60-67.
Belpaire, C.G.J., Goemans, G., Geeraerts, C., Quataert, P., Parmentier, K., Hagel, P., De Boer, J., 2009. Decreasing eel stocks: survival of the fattest? Ecology of Freshwater Fish 18, 197-214.
Carls, M.G., Rice, S.D., Hose, J.E., 1999. Sensitivity of fish embryos to weathered crude oil: Part I. Low-level exposure during incubation causes malformations, genetic damage, and mortality in larval Pacific herring (Clupea pallasi). Environmental Toxicology and Chemistry 18, 481-493.
Clevestam, P.D., Ogonowski, M., Sjoberg, N.B., Wickstrom, H., 2011. Too short to spawn? Implications of small body size and swimming distance on successful migration and maturation of the European eel Anguilla anguilla. Journal of Fish Biology 78, 1073-1089.

Eggsposed Chapter 8
173
Daley, J.M., Leadley, T.A., Pitcher, T.E., Drouillard, K.G., 2012. Bioamplification and the Selective Depletion of Persistent Organic Pollutants in Chinook Salmon Larvae. Environmental Science & Technology 46, 2420-2426.
Daouk, T., Larcher, T., Roupsard, F., Lyphout, L., Rigaud, C., Ledevin, M., Loizeau, V., Cousin, X., 2011. Long-term food-exposure of zebrafish to PCB mixtures mimicking some environmental situations induces ovary pathology and impairs reproduction ability. Aquatic Toxicology 105, 270-278.
Debruyn, A.M.H., Ikonomou, M.G., Gobas, F., 2004. Magnification and toxicity of PCBs, PCDDs, and PCDFs in upriver-migrating Pacific salmon. Environmental Science & Technology 38, 6217-6224.
Dekker, W., 2003. Did lack of spawners cause the collapse of the European eel, Anguilla anguilla? Fisheries Management and Ecology 10, 365-376.
Dekker, W., 2004. Slipping through our hands. Population dynamics of the European eel. University of Amsterdam, Amsterdam.
Dietrich, G.J., Dietrich, M., Kowalski, R.K., Dobosz, S., Karol, H., Demianowicz, W., Glogowski, J., 2010. Exposure of rainbow trout milt to mercury and cadmium alters sperm motility parameters and reproductive success. Aquatic Toxicology 97, 277-284.
Dunn, M.R., Stevens, D.W., Forman, J.S., Connell, A., 2013. Trophic Interactions and Distribution of Some Squaliforme Sharks, Including New Diet Descriptions for Deania calcea and Squalus acanthias. PLoS One 8.
Eliasen, K., Reinert, J., Gaard, E., Hansen, B., Jacobsen, J.A., Gronkjaer, P., Christensen, J.T., 2011. Sandeel as a link between primary production and higher trophic levels on the Faroe shelf. Marine Ecology Progress Series 438, 185-194.
Elliott, J.M., 1989. Mechanisms responsible for population regulation in young migratory trout, Salmo trutta. I. the critical time for survival. Journal of Animal Ecology 58, 987-1001.
Elonen, G.E., Spehar, R.L., Holcombe, G.W., Johnson, R.D., Fernandez, J.D., Erickson, R.J., Tietge, J.E., Cook, P.M., 1998. Comparative toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin to seven freshwater fish species during early life-stage development. Environmental Toxicology and Chemistry 17, 472-483.
Escher, B.I., Hermens, J.L.M., 2002. Modes of action in ecotoxicology: Their role in body burdens, species sensitivity, QSARs, and mixture effects. Environmental Science & Technology 36, 4201-4217.
Escher, B.I., Schwarzenbach, R.P., 2002. Mechanistic studies on baseline toxicity and uncoupling of organic compounds as a basis for modeling effective membrane concentrations in aquatic organisms. Aquatic Sciences 64, 20-35.
Ewald, G., Larsson, P., Linge, H., Okla, L., Szarzi, N., 1998. Biotransport of organic pollutants to an inland Alaska lake by migrating sockeye salmon (Oncorhynchus nerka). Arctic 51, 40-47.
Fisk, A.T., Tittlemier, S.A., Pranschke, J.L., Norstrom, R.J., 2002. Using anthropogenic contaminants and stable isotopes to assess the feeding ecology of greenland sharks. Ecology 83, 2162-2172.

Chapter 8 Eggsposed
174
Geeraerts, C., Belpaire, C., 2010. The effects of contaminants in European eel: a review. Ecotoxicology 19, 239-266.
Geeraerts, C., Focant, J.F., Eppe, G., De Pauw, E., Belpaire, C., 2011. Reproduction of European eel jeopardised by high levels of dioxins and dioxin-like PCBs? Science of the Total Environment 409, 4039-4047.
Giesy, J.P., Jones, P.D., Kannan, K., Newsted, J.L., Tillitt, D.E., Williams, L.L., 2002. Effects of chronic dietary exposure to environmentally relevant concentrations of 2,3,7,8-tetrachlorodibenzo-p-dioxin on survival, growth, reproduction and biochemical responses of female rainbow trout (Oncorhynchus mykiss). Aquatic Toxicology 59, 35-53.
González-Doncel, M., de la Peña, E., Barrueco, C., Hinton, D.E., 2003. Stage sensitivity of medaka (Oryzias latipes) eggs and embryos to permethrin. Aquatic Toxicology 62, 255-268.
Gonzalez-Doncel, M., Larrea, M., Sanchez-Fortun, S., Hinton, D.E., 2003. Influence of water hardening of the chorion on cadmium accumulation in medaka (Oryzias latipes) eggs. Chemosphere 52, 75-83.
Gutleb, A.C., Appelman, J., Bronkhorst, M.C., van den Berg, J.H.J., Spenkelink, A., Brouwer, A., Murk, A.J., 1999. Delayed effects of pre- and early-life time exposure to polychlorinated biphenyls on tadpoles of two amphibian species (Xenopus laevis and Rana temporaria). Environmental Toxicology and Pharmacology 8, 1-14.
Hallare, A.V., Kosmehl, T., Schulze, T., Hollert, H., Kohler, H.R., Triebskorn, R., 2005. Assessing contamination levels of Laguna Lake sediments (Philippines) using a contact assay with zebrafish (Danio rerio) embryos. Science of the Total Environment 347, 254-271.
Heintz, R.A., Short, J.W., Rice, S.D., 1999. Sensitivity of fish embryos to weathered crude oil: Part II. Increased mortality of pink salmon (Oncorhynchus gorbuscha) embryos incubating downstream from weathered Exxon Valdez crude oil. Environmental Toxicology and Chemistry 18, 494-503.
Helser, T.E., Alade, L., 2012. A retrospective of the hake stocks off the Atlantic and Pacific coasts of the United States: Uncertainties and challenges facing assessment and management in a complex environment. Fisheries Research 114, 2-18.
Hinther, A., Bromba, C.M., Wulff, J.E., Helbing, C.C., 2011. Effects of Triclocarban, Triclosan, and Methyl Triclosan on Thyroid Hormone Action and Stress in Frog and Mammalian Culture Systems. Environmental Science & Technology 45, 5395-5402.
Hutchings, J.A., Myers, R.A., Garcia, V.B., Lucifora, L.O., Kuparinen, A., 2012. Life-history correlates of extinction risk and recovery potential. Ecological Applications 22, 1061-1067.
Johnson, R.D., Tietge, J.E., Jensen, K.M., Fernandez, J.D., Linnum, A.L., Lothenbach, D.B., Holcombe, G.W., Cook, P.M., Christ, S.A., Lattier, D.L., Gordon, D.A., 1998. Toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin to early life stage brook trout (Salvelinus fontinalis) following parental dietary exposure. Environmental Toxicology and Chemistry 17, 2408-2421.
Kamler, E., 2008. Resource allocation in yolk-feeding fish. Reviews in Fish Biology and Fisheries 18, 143-200.

Eggsposed Chapter 8
175
Kannan, K., Corsolini, S., Imagawa, T., Focardi, S., Giesy, J.P., 2002. Polychlorinated -naphthalenes, -biphenyls, -dibenzo-p-dioxins, -dibenzofurans and p,p '-DDE in bluefin tuna, swordfish, cormorants and barn swallows from Italy. Ambio 31, 207-211.
Kelly, B.C., Gray, S.L., Ikonomou, M.G., MacDonald, J.S., Bandiera, S.M., Hrycay, E.G., 2007. Lipid reserve dynamics and magnification of persistent organic pollutants in spawning sockeye salmon (Oncorhynchus nerka) from the Fraser River, British Columbia. Environmental Science & Technology 41, 3083-3089.
Kelly, B.C., Ikonomou, M.G., MacPherson, N., Sampson, T., Patterson, D.A., Dubetz, C., 2011. Tissue residue concentrations of organohalogens and trace elements in adult pacific salmon returning to the Fraser River, Britisch Colombia, Canada. Environmental Toxicology and Chemistry 30, 367-376.
King Heiden, T.C., Spitsbergen, J., Heideman, W., Peterson, R.E., 2009. Persistent Adverse Effects on Health and Reproduction Caused by Exposure of Zebrafish to 2,3,7,8-Tetrachlorodibenzo-p-dioxin During Early Development and Gonad Differentiation. Toxicological Sciences 109, 75-87.
Kipka, U., Di Toro, D.M., 2009. Technical basis for polar and non-polar narcotic chemicals ans polycyclic aromaic hydrocarbon criteria. III. A polyparameter model for target lipid partitioning. Environmental Toxicology and Chemistry 28, 1429-1438.
Klaren, P.H.M., Wunderink, Y.S., Yufera, M., Mancera, J.M., Flik, G., 2008. The thyroid gland and thyroid hormones in Senegalese sole (Solea senegalensis) during early development and metamorphosis. General and Comparative Endocrinology 155, 686-694.
Kwadijk, C., Korytar, P., Koelmans, A.A., 2010. Distribution of Perfluorinated Compounds in Aquatic Systems in The Netherlands. Environmental Science & Technology 44, 3746-3751.
Laurenson, C.H., Priede, I.G., 2005. The diet and trophic ecology of anglerfish Lophius piscatorius at the Shetland Islands, UK. J. Mar. Biol. Assoc. U.K. 85, 419-424.
Leclerc, L.M.E., Lydersen, C., Haug, T., Bachmann, L., Fisk, A.T., Kovacs, K.M., 2012. A missing piece in the Arctic food web puzzle? Stomach contents of Greenland sharks sampled in Svalbard, Norway. Polar Biology 35, 1197-1208.
Legler, J., 2008. New insights into the endocrine disrupting effects of brominated flame retardants. Chemosphere 73, 216-222.
MacNamara, R., McCarthy, T.K., 2012. Size-related variation in fecundity of European eel (Anguilla anguilla). Ices Journal of Marine Science 69, 1333-1337.
MacNeil, M.A., McMeans, B.C., Hussey, N.E., Vecsei, P., Svavarsson, J., Kovacs, K.M., Lydersen, C., Treble, M.A., Skomal, G.B., Ramsey, M., Fisk, A.T., 2012. Biology of the Greenland shark Somniosus microcephalus. Journal of Fish Biology 80, 991-1018.
Mathew, R., McGrath, J.A., Di Toro, D.M., 2008. Modeling polycyclic aromatic hydrocarbon bioaccumulation and metabolism in time-variable early life-stage exposures. Environmental Toxicology and Chemistry 27, 1515-1525.
Mennigen, J.A., Stroud, P., Zamora, J.M., Moon, T.W., Trudeau, V.L., 2011. Pharmaceuticals as neuroendocrine disrupters: Lessons learned from fish on PROZAC. Journal of Toxicology and Environmental Health-Part B-Critical Reviews 14, 387-412.

Chapter 8 Eggsposed
176
Mertz, G., Myers, R.A., 1996. Influence of fecundity on recruitment variability of marine fish. Canadian Journal of Fisheries and Aquatic Sciences 53, 1618-1625.
Milner, N.J., Elliott, J.M., Armstrong, J.D., Gardiner, R., Welton, J.S., Ladle, M., 2003. The natural control of salmon and trout populations in streams. Fisheries Research 62, 111-125.
Mizell, M., Romig, E.S., 1997. The aquatic vertebrate embryo as a sentinel for toxins: Zebrafish embryo dechorionation and perivitelline space microinjection. International Journal of Developmental Biology 41, 411-423.
Mochida, K., Ito, K., Kono, K., Onduka, T., Kakuno, A., Fujii, K., 2010. Effect of tributyltin oxide exposure on the F-0 and F-1 generations of a marine teleost, the mummichog Fundulus heteroclitus. Fisheries Science 76, 333-341.
Molde, K., Ciesielski, T.M., Fisk, A.T., Lydersen, C., Kovacs, K.M., Sormo, E.G., Jenssen, B.M., 2013. Associations between vitamins A and E and legacy POP levels in highly contaminated Greenland sharks (Somniosus microcephalus). Science of the Total Environment 442, 445-454.
Montano, M., Gutleb, A.C., murk, A.J., 2013 in press. Persistent Toxic Burdens of Halogenated Phenolic Compounds in Humans and Wildlife. Environ. Sci. Technol.
Moore, M.R., Vetter, W., Gaus, C., Shaw, G.R., Muller, J.F., 2002. Trace organic compounds in the marine environment. Marine Pollution Bulletin 45, 62-68.
Mourente, G., Vazquez, R., 1996. Changes in the content of total lipid, lipid classes and their fatty acids of developing eggs and unfed larvae of the Senegal sole, Solen senegalensis Kaup. Fish Physiol. Biochem. 15, 221-235.
Munschy, C., Heas-Moisan, K., Tixier, C., Pacepavicius, G., Alaee, M., 2010. Dietary exposure of juvenile common sole (Solea solea L.) to polybrominated diphenyl ethers (PBDEs): Part 2. Formation, bioaccumulation and elimination of hydroxylated metabolites. Environmental Pollution 158, 3527-3533.
OECD, 1998. Guidline for testing of chemicals 212: Fish, Short-term toxicity test on embryo and sac-fry stages
Organisation for Economic Co-operation and Development (OECD), Paris OECD, 2006. Draft proposal for a new guideline: Fish Embryo Toxicity (FET) Test Organisation for Economic Co-operation and Development (OECD), Paris Okada, N., Morita, T., Tanaka, M., Tagawa, M., 2005. Thyroid hormone deficiency in
abnormal larvae of the Japanese flounder Paralichthys olivaceus. Fisheries Science 71, 107-114.
Palace, V., Park, B., Pleskach, K., Gemmill, B., Tomy, G., 2010. Altered thyroxine metabolism in rainbow trout (Oncorhynchus mykiss) exposed to hexabromocyclododecane (HBCD). Chemosphere 80, 165-169.
Palstra, A.P., van den Thillart, G., 2010. Swimming physiology of European silver eels (Anguilla anguilla L.): energetic costs and effects on sexual maturation and reproduction. Fish Physiol. Biochem. 36, 297-322.
Palstra, A.P., van Ginneken, V.J.T., Murk, A.J., van den Thillart, G., 2006. Are dioxin-like contaminants responsible for the eel (Anguilla anguilla) drama? Naturwissenschaften 93, 145-148.

Eggsposed Chapter 8
177
Posthuma, L., Suter, G.W., Traas, T.P., 2010 Species Sensitivity Distributions in Ecotoxicology. Taylor & Francis.
Righton, D., Aarestrup, K., Jellyman, D., Sebert, P., van den Thillart, G., Tsukamoto, K., 2012. The Anguilla spp. migration problem: 40 million years of evolution and two millennia of speculation. Journal of Fish Biology 81, 365-386.
Rijnsdorp, A.D., Vanbeek, F.A., Flatman, S., Millner, R.M., Riley, J.D., Giret, M., Declerck, R., 1992. Recruitment of sole stocks, Solea solea (L), in the North East Atlantic. Neth. J. Sea Res. 29, p. 173.
Roberts, S.C., Noyes, P.D., Gallagher, E.P., Stapleton, H.M., 2011. Species-Specific Differences and Structure-Activity Relationships in the Debromination of PBDE Congeners in Three Fish Species. Environmental Science & Technology 45, 1999-2005.
Robinet, T.T., Feunteun, E.E., 2002. Sublethal effects of exposure to chemical compounds: A cause for the decline in Atlantic eels? Ecotoxicology 11, 265-277.
Rocha, P.S., Bernecker, C., Strecker, R., Mariani, C.F., Pompeo, M.L.M., Storch, V., Hollert, H., Braunbeck, T., 2011. Sediment-contact fish embryo toxicity assay with Danio rerio to assess particle-bound pollutants in the Tiete River Basin (Sao Paulo, Brazil). Ecotoxicology and Environmental Safety 74, 1951-1959.
Ruggerone, G.T., Nielsen, J.L., 2004. Evidence for competitive dominance of Pink salmon (Oncorhynchus gorbuscha) over other Salmonids in the North Pacific Ocean. Reviews in Fish Biology and Fisheries 14, 371-390.
Russell, R.W., Gobas, F., Haffner, G.D., 1999. Maternal transfer and in ovo exposure of organochlorines in oviparous organisms: A model and field verification. Environmental Science & Technology 33, 416-420.
Schuckel, S., Sell, A.F., Kroncke, I., Reiss, H., 2012. Diet overlap among flatfish species in the southern North Sea. Journal of Fish Biology 80, 2571-2594.
Selck, H., Riemann, B., Christoffersen, K., Forbes, V.E., Gustavson, K., Hansen, B.W., Jacobsen, J.A., Kusk, O.K., Petersen, S., 2002. Comparing sensitivity of ecotoxicological effect endpoints between laboratory and field. Ecotoxicology and Environmental Safety 52, 97-112.
Smit, M.G.D., Bechmann, R.K., Hendriks, A.J., Skadsheim, A., Larsen, B.K., Baussant, T., Bamber, S., Sanni, S., 2009. Relating biomarkers to whole organism effects using species sensitivity distributions: A pilot study for marine species exposed to oil. Environmental Toxicology and Chemistry 28, 1104-1109.
Soffker, M., Tyler, C.R., 2012. Endocrine disrupting chemicals and sexual behaviors in fish - a critical review on effects and possible consequences. Critical Reviews in Toxicology 42, 653-668.
Steevens, J.A., Reiss, M.R., Pawlisz, A.V., 2005. A methodology for deriving tissue residue benchmarks for aquatic biota: a case study for fish exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin and equivalents. Integr Environ Assess Manag 1, 142-151.
Stobutzki, I.C., 1997. Energetic cost of sustained swimming in the late pelagic stages of reef fishes. Marine Ecology Progress Series 152, 249-259.

Chapter 8 Eggsposed
178
Storelli, M.M., Barone, G., Storelli, A., Marcotrigiano, G.O., 2011. Levels and congener profiles of PCBs and PCDD/Fs in blue shark (Prionace glauca) liver from the South-Eastern Mediterranean Sea (Italy). Chemosphere 82, 37-42.
Storelli, M.M., Casalino, E., Barone, G., Marcotrigiano, G.O., 2008. Persistent organic pollutants (PCBs and DDTs) in small size specimens of bluefin tuna (Thunnus thynnus) from the Mediterranean Sea (Ionian Sea). Environment International 34, 509-513.
Strid, A., Athanassiadis, I., Athanasiadou, M., Svavarsson, J., Papke, O., Bergman, A., 2010. Neutral and phenolic brominated organic compounds of natural and anthropogenic origin in Northeast Atlantic Greenland shark (Somniosus microcephalus). Environmental Toxicology and Chemistry 29, 2653-2659.
Strid, A., Jorundsdottir, H., Papke, O., Svavarsson, J., Bergman, A., 2007. Dioxins and PCBs in Greenland shark (Somniosus microcephalus) from the North-East Atlantic. Marine Pollution Bulletin 54, 1514-1522.
Tagawa, M., Aritaki, M., 2005. Production of symmetrical flatfish by controlling the timing of thyroid hormone treatment in spotted halibut Verasper variegatus. General and Comparative Endocrinology 141, 184-189.
Tagawa, M., Hirano, T., 1991. Effects of thyroid hormone deficiency in eggs on early developmnet of the Medaka, Orysias latipes. Journal of Experimental Zoology 257, 360-366.
Tagawa, M., Tanaka, M., Matsumoto, S., Hirano, T., 1990. Thyroid hormones in eggs of various freswater, marine and diadromous teleosts and their changs during egg development. Fish Physiol. Biochem. 8, 515-520.
Usenko, C.Y., Hopkins, D.C., Trumble, S.J., Bruce, E.D., 2012. Hydroxylated PBDEs induce developmental arrest in zebrafish. Toxicology and Applied Pharmacology 262, 43-51.
Usenko, C.Y., Robinson, E.M., Usenko, S., Brooks, B.W., Bruce, E.D., 2011. PBDE DEVELOPMENTAL EFFECTS ON EMBRYONIC ZEBRAFISH. Environmental Toxicology and Chemistry 30, 1865-1872.
USEPA, 1996. Ecological Effects Test Guidelines: OPPTS 850.1400 Fish Early-Life Stage Toxicity Test. United States Environmental Protection Agency (USEPA).
Valo, R., Salkinojasalonen, M., 1986. Microbial transformation of polychlorinated phenoxy phenols. Journal of General and Applied Microbiology 32, 505-517.
van Ginneken, V., Antonissen, E., Muller, U.K., Booms, R., Eding, E., Verreth, J., van den Thillart, G., 2005. Eel migration to the Sargasso: remarkably high swimming efficiency and low energy costs. Journal of Experimental Biology 208, 1329-1335.
van Ginneken, V., Palstra, A., Leonards, P., Nieveen, M., van den Berg, H., Flik, G., Spanings, T., Niemantsverdriet, P., van den Thillart, G., Murk, A., 2009. PCBs and the energy cost of migration in the European eel (Anguilla anguilla L.). Aquatic Toxicology 92, 213-220.
Vandegehuchte, M.B., De Coninck, D., Vandenbrouck, T., De Coen, W.M., Janssen, C.R., 2010. Gene transcription profiles, global DNA methylation and potential transgenerational epigenetic effects related to Zn exposure history in Daphnia magna. Environmental Pollution 158, 3323-3329.

Eggsposed Chapter 8
179
Vandegehuchte, M.B., Janssen, C.R., 2011. Epigenetics and its implications for ecotoxicology. Ecotoxicology 20, 607-624.
Villalobos, S.A., Hamm, J.T., Teh, S.J., Hinton, D.E., 2000. Thiobencarb-induced embryotoxicity in medaka (Oryzias latipes): stage-specific toxicity and the protective role of chorion. Aquatic Toxicology 48, 309-326.
Walker, M.K., Peterson, R.E., 1994. Toxicily of 2,3,7,8-tetrachlorodibenzo-p-dioxin to brook trout (Salvelinus fontinalis) during early development. Environmental Toxicology and Chemistry 13, 817-820.
Wang, M.Y., Chen, J.F., Lin, K.F., Chen, Y.H., Hu, W., Tanguay, R.L., Huang, C.J., Dong, Q.X., 2011. Chonic zebrafish PFOS exposure alters sex ration and maternal related effects in F1 offspring. Environmental Toxicology and Chemistry 30, 2073-2080.
Wirgin, I., Roy, N.K., Loftus, M., Chambers, R.C., Franks, D.G., Hahn, M.E., 2011. Mechanistic Basis of Resistance to PCBs in Atlantic Tomcod from the Hudson River. Science 331, 1322-1325.
Wirgin, I., Waldman, J.R., 2004. Resistance to contaminants in North American fish populations. Mutation Research-Fundamental and Molecular Mechanisms of Mutagenesis 552, 73-100.
Yamano, K., 2005. The role of thyroid hormone in fish development with reference to aquaculture. Jarq-Japan Agricultural Research Quarterly 39, 161-168.
Yano, K., Stevens, J.D., Compagno, L.J.V., 2007. Distribution, reproduction and feeding of the Greenland shark Somniosus (Somniosus) microcephalus, with notes on two other sleeper sharks, Somniosus (Somniosus) pacificus and Somniosus (Somniosus) antarcticus. Journal of Fish Biology 70, 374-390.
Youngson, N.A., Whitelaw, E., 2008. Transgenerational epigenetic effects, Annual Review of Genomics and Human Genetics, pp. 233-257.
Yu, L.Q., Lam, J.C.W., Guo, Y.Y., Wu, R.S.S., Lam, P.K.S., Zhou, B.S., 2011. Parental Transfer of Polybrominated Diphenyl Ethers (PBDEs) and Thyroid Endocrine Disruption in Zebrafish. Environmental Science & Technology 45, 10652-10659.
Zheng, X.M., Zhu, Y.T., Liu, C.S., Liu, H.L., Giesy, J.P., Hecker, M., Lam, M.H.W., Yu, H.X., 2012. Accumulation and Biotransformation of BDE-47 by Zebrafish Larvae and Teratogenicity and Expression of Genes along the Hypothalamus-Pituitary-Thyroid Axis. Environmental Science & Technology 46, 12943-12951.
Zielke, H., Seiler, T.B., Niebergall, S., Leist, E., Brinkmann, M., Spira, D., Streck, G., Brack, W., Feiler, U., Braunbeck, T., Hollert, H., 2011. The impact of extraction methodologies on the toxicity of sediments in the zebrafish (Danio rerio) embryo test. Journal of Soils and Sediments 11, 352-363.


181
Summary
“He stopped for a moment and looked back and saw in the reflection from the street light the great tail of the fish standing up well behind the skiff’s tern.”
Ernest Hemmingway: The Old man and the Sea

Summary Eggsposed
182
Persistent organic pollutants (POP), with well-known representatives as polychlorinated biphenyls (PCBs), dioxins, and brominated flame retardants as polybrominated diphenyl ethers (PBDEs) and hexabromocyclododecanes (HBCD), are still globally present in the marine environment, This despite the substantial reduction of application and emission that was achieved during the last decades. Apart from their persistency these compounds share low water solubility and a high lipophilicity which make that the highest concentrations in the aquatic environment are found in the organic matrix of sediments and in biota. Dissolved water concentrations are low. Hence, intake of contaminated food items forms the major source for POPs exposure of aquatic organisms, and through biomagnification the highest concentrations can be found in the tissue of top predators. POPs have the potency to cause a variety of toxic effects, among which endocrine disruption and teratogenic effects that especially apply to early life stages. As the early life stages of most fish species develop suspended in the water column, exposure to POPs may be considered relatively low, at least until the larvae start feeding after yolk absorption. However, POPs accumulated in the tissue of the mother are transferred to the eggs. The research presented in this thesis aims at the determination of the impact of such maternally transferred POPs on development and survival of fish early life stages, in order to assess if this exposure route can significantly impact the development of a fish population at current environmental concentrations, especially in combination with high fishing pressure.
For this purpose a bioassay was developed with the common sole (Solea solea). The advantages for this research of this new bioassay above standard fish early life stage (ELS) tests are that sole is a native West European species that as all flatfishes undergoes an obvious metamorphosis. The test set-up includes this metamorphosis that is thyroid hormone mediated and therefore expected to be easily disrupted by POPs, based on research with amphibians. The prolonged Early Life Stage test (p-ELS) with sole is presented in chapter 2. Early life stages were exposed to a concentration series of the dioxin-like PCB 126 (3,3',4,4',5-pentachlorobiphenyl) in seawater until 4, 8,10 and 15 days post fertilisation (dpf). Subsequently the development of the larvae was registered under further unexposed conditions. The LC50s at the start of the free-feeding stage (12 dpf) ranged between 39 and 83 ng PCB 126/l depending on exposure duration. After the fish had completed the metamorphosis, the LC50 values ranged between 1.7 and 3.7 ng PCB 126/l for the groups exposed for 4, 8 and 10 dpf respectively. Thus exposure for only 4 days, covering only the egg stage, was sufficient to cause adverse effects during a critical developmental phase two weeks later. This study indicates that ELS fish tests that are terminated shortly after the fish becomes free-feeding underestimate the toxic potential of compounds with low acute toxicity such as PCBs. The internal dosages of these larvae at the end of

Eggsposed Summary
183
the exposure, determined by means of an in-vitro gene reporter assays as dioxin-equivalent values (TEQ), revealed an internal lethal concentration, ILC50 of 1 ng TEQ/g lipid, which is within the same order of magnitude as TEQ levels found in fish from highly polluted areas. This suggests that larval survival of fish populations at contaminated sites can be affected by persistent compounds that are accumulated by the female fish and passed on to the eggs.
Based on these first results the p-ELS test procedure was improved to reach a better control performance. The exposure period was terminated when all larvae had hatched (6 dpf), this in order to mimic exposure through maternal transfer as good as possible without exposing parent fish or manipulation the eggs. In a second test (Chapter 3) the eggs were exposed to a concentration series of methyltriclosan (MTCS), a metabolite of triclosan (TCS) that is commonly used as bactericide in a wide variety of human care products. MTCS and TCS are discharged with waste water, bioaccumulate in fish tissue, and are known to have the potency to disrupt the thyroid hormone system. Mortality occurred in the higher treatment levels until 20 dpf. Indications for thyroid hormone disruption were not observed; all surviving larvae completed metamorphoses without problems. Internal effect concentrations, reached in larvae at the end of the exposure (6 dpf), were 5.8 mg/g lipid weight (lw) and 2.1 mg/g lw for ILC50 and ILC10 respectively. These internal effect concentrations are at least 200 times higher than concentrations that due to maternally transfer can be expected in the eggs of highly exposed fish in a field situation. Our results thus do not indicate a high risk from maternally transferred MTCS for fish at the current field concentrations.
In order to get more insight in the fate of the POPs in the larvae, in Chapter 4 the existing bioaccumulation model OMEGA was adjusted for sole early life stages and validated with experimental data with PCBs. This study revealed, that tissue concentrations of compounds with log Kow >6, peak in the tissues of developing sole at the end of the yolk-sac stage, when lipid reserves are depleted. As a result, just before the larvae become free feeding, the peak tissue concentrations of the pollutants in the larvae exceed that of the adult fish. This also explains at least partly, the delayed effects that were observed in Chapter 2 (and 5).
Chapter 5 assesses the likelihood that early life development of fish from contaminated areas is affected by maternally transferred POPs. Following the p-ELS test protocol, effects on sole larvae were determined for the dioxin-like PCB 126, the technical PCB-mixture Arochlor 1254, PBDEs and HBCDs, for an artificial mixture of PCBs and PBDEs, and for ‘field mixtures’ extracted from sole collected from the North Sea and in the contaminated Western Scheldt estuary.

Summary Eggsposed
184
As was earlier observed with PCB126 and MTCS, exposure to PCBs, PBDEs and the artificial and field mixtures caused mortality that started to occur shortly after the larvae became free-feeding and continued to increase until the onset of metamorphoses. The effects induced by the field mixtures correlated well with the ∑PCB concentrations in the tissue of the exposed larvae. No indications were found for synergistic effects or for substantial contribution of other (unknown) substances in the field mixtures. HBCD did not induce toxic effects. POP levels in sole from Western Scheldt estuary are about 20 times lower than the ILC50, the larval tissue concentration that produced 50% early life stage mortality. Levels in North Sea sole are an order of a magnitude lower.
Chapter 6 describes a risk assessment for toxicant induced larval survival for European eel (Anguilla anguilla). Eels are considered sensitive for the effect of POPs that can accumulate to high levels in their lipid rich tissue. During spawning migration without feeding high lipophilic dioxin-like POPs in the eel’s tissue were estimated to increase 1.33 or 2 fold, due to weight loss. As no toxicity data are available for eel larvae, the critical egg concentrations for larval survival was estimated from a sensitivity distribution based on literature data of other species. It was assumed that eel larvae belong to the 5% or 1% most sensitive teleost fish species. Given concentrations of dioxin-like pollutants as reported for European eel, and following the worst case scenarios with respect to sensitivity of the larvae and bio amplification during migration, it can be expected that larvae of eel from highly contaminated locations in The Netherlands and Belgium will experience more than 50% mortality due to maternally transferred dioxin-like toxicants.
Chapter 7 explores the potential impact of (toxicant induced) early life stage mortality on the population development of sole by application of a simple age structured matrix model. The model is used to explore the population response to a combination of (toxicant induced) larval mortality and fishing-related mortality of mature fish. The results indicate that the impact of larval mortality that occurs before metamorphosis is very low, even in population subject to high fishing pressure. This is the result of the combination of a high fecundity and the fact that the larval mortality occurs before the moment when the number of recruits is limited by the carrying capacity of the nursery areas. When colonising the nursery areas the, until than pelagic sole larvae metamorphose into flatfishes with a benthic life style. The individuals hence concentrate from the three dimensional pelagic environment to a two dimensional benthic environment, which caused density dependent mortality. This concentration of early life stages is typical for flatfish. Mortality that occurs after the nursery areas are populated will have a more pronounced impact on population development. The results further imply that population development of pelagic

Eggsposed Summary
185
fish species that do not concentrate in nursery areas, and species with low fecundity is more vulnerable for disturbance through mortality of early life stages.
Chapter 8 synthesises and discusses the outcome of the research. It is stressed that short term fish tests, often covering only the embryonic development, will underestimate the real risk of lipophilic substances. Toxicity of these substances will peak after yolk sac absorption when these tests have already been ended. When the characteristics of the test substance are known this risk is predictable with for instance the ELS-OMEGA model. However, especially when mixtures of unknown composition (effluents, sediment extracts) are being tested one must realise that the contribution of lipophilic substances may be underestimated in test that are terminated before, or too soon after the fish larvae are free feeding.
The absence of effects on metamorphosis in our P-ELS test is explained by the prediction of the ELS-OMEGA model that the POPs concentrations in the larvae, had reached too low concentrations at the moment of metamorphoses to disrupt the thyroid hormone system. This was due to passive excretion (for substances with log Kow <6) and growth dilution.
It must be realised that the experimental set-up that was followed to mimic the effects of maternally transferred POPs does not include potential effects of maternally transferred metabolites of these POPs that can be formed by the parent fish and that are often more toxic than the mother compounds. Also effects of the mother’s condition on the quality of the eggs and epigenetic effects were not included. This implies that the results of the tests as performed in some cases might underestimate the actual effects of these substances.
The species most vulnerable to the effects of maternally transferred POPs share a high exposure, low fecundity and the absence of density dependent mortality of early life stages. According to these criteria sharks and especially the Greenland shark (Somniosus microcephalus) that is highly exposed to POPs can be considered as highly vulnerable. It is therefore recommended to investigate the actual sensitivity of this species, in order to get more insight in the potential vulnerability of the populations.


187
Nederlandse samenvatting (speciaal voor niet-ingewijden)

Samenvatting Eggsposed
188
De hoeveelheden slecht afbreekbare organische verontreinigende stoffen (‘persistent organic pollutants ‘ kortweg POPs) die in het milieu terecht komen zijn de afgelopen decennia sterk teruggebracht. Vanwege de grote hoeveelheden die in het verleden zijn geproduceerd en hun slechte afbreekbaarheid zijn deze stoffen echter nog steeds in het milieu aanwezig. Bekende voorbeelden hiervan zijn PCBs en dioxines. Omdat zij beter in vet oplossen dan in water hopen deze stoffen zich op in organismen en via stapeling in de gehele voedselketen. POPs die zijn opgehoopt in het lichaam van een vrouwelijke vis worden aan haar eieren doorgegeven via het vet in de dooier. De eieren zijn dus al verontreinigd als zij het moederlichaam verlaten.
Het onderzoek beschreven in dit proefschrift is gericht op de effecten van deze aan eieren doorgegeven POPs op de ontwikkeling van jonge vis. Onderzocht is welke type effecten kunnen optreden, en of deze effecten ook te verwachten zijn bij de concentraties zoals die in vissen in Nederland en elders aanwezig zijn.
Uit de omvangrijke groep POPs zijn voor dit onderzoek stoffen gekozen die veel worden aangetroffen in het mariene milieu. Hieruit zijn stoffen geselecteerd die dioxine-achtige giftigheid vertonen of het schildklierhormoonsysteem kunnen verstoren. Dioxines en dioxine-achtige stoffen veroorzaken ernstige effecten bij ontwikkelende vislarven, zoals oedeem en misvormingen. Het schildklierhormoon speelt een rol bij verschillende onderdelen van de ontwikkeling van vislarven, maar is vooral belangrijk bij de metamorfose van de larve naar een juveniele vis. Dit is ook het geval bij de metamorfose van een kikkervisje in een kikker, waarbij is aangetoond dat verstoring van het schildklierhormoonsysteem de metamorfose kan verstoren, met misvormingen tot gevolg.
Om deze effecten te onderzoeken is een test ontwikkeld met de platvis tong. Dit is hiervoor een geschikte testsoort, omdat tong net als alle platvissen een duidelijke metamorfose ondergaat. Hierbij verandert de tot dan toe symmetrische larve in een asymmetrische platvis, perfect aangepast voor een leven op de zeebodem. Bovendien is tong een belangrijke commerciële vissoort voor de Noordzee, en zijn bevruchte eieren beschikbaar uit aquacultuur. Voor het onderzoek zijn geen moederdieren aan POPs blootgesteld. De eieren werden zo snel mogelijk na de bevruchting aan de teststof blootgesteld. Na 5 tot 6 dagen kwamen de eieren uit, en in de meeste testen werd op dat moment de blootstelling aan de teststof beëindigd. De pas uitgekomen larven bezitten geen ogen, mond of ingewanden en zijn voor hun voedingstoffen volledig aangewezen op de dooier. Deze periode wordt aangeduid als het ‘dooierzak stadium’ en duurt ca. 5 dagen. In dit stadium wordt het grootste deel van de dooier gebruikt en ontwikkelt de larve onder andere organen, ogen, een mond en ingewanden,

Eggsposed Samenvatting
189
waardoor het zelfstandig kan gaan eten. De larve, nu in het ‘free-feeding stadium’, begint duidelijk te groeien en heeft ca. 25 dagen na de bevruchting een lengte van iets minder dan een centimeter bereikt. Op dat moment beginnen de eerste vissen aan de metamorfose waarbij het linkeroog naar de rechterzijde verschuift om de vis optimaal aan te passen aan een leven op de zeebodem. Onder normale omstandigheden volbrengen de tonglarven de metamorfose binnen 40 dagen na de bevruchting.
Het eerste experiment (Hoofdstuk 2) werd uitgevoerd met PCB-126, een stof waarvan dioxine-achtige giftigheid verwacht mag worden. De eieren werden via het water blootgesteld gedurende 4, 8, 10 en 15 dagen. Opvallend was dat in alle gevallen sterfte van de larven pas zichtbaar werd nadat de larven de dooierzak hadden verbruikt en overgingen op externe voeding, ca. 10 dagen na de bevruchting. In sommige gevallen was dit dus lang nadat de blootstelling was beëindigd. In alle gevallen bleef de sterfte toenemen totdat het moment van metamorfose bereikt was. Vrijwel alle overlevende larven doorliepen de metamorfose zonder problemen.
De volgende testen (Hoofdstuk 3) zijn uitgevoerd met het potentieel schildklierhormoonverstorende “methyltriclosan”. Hoewel van deze stof dus een heel ander effect verwacht werd dan van PCB 126, waren de resultaten vergelijkbaar. Wederom vond de sterfte vooral plaats nadat de dooierzak was geabsorbeerd en de overlevende larven voltooiden de metamorfose zonder problemen.
Om beter te begrijpen hoe deze effecten tot stand komen wordt in Hoofdstuk 4 een rekenkundig model gepresenteerd dat het verloop van de concentratie van POPs in een zich ontwikkelende vislarve beschrijft. Hiervoor is het al bestaande OMEGA model aangepast op basis van experimenten waarin het gehalte van PCBs in tonglarven werd gevolgd. De resultaten laten zien dat de slecht in water oplosbare POPs die in de dooier aanwezig zijn de vislarven nauwelijks verlaten. Naarmate de dooier meer geabsorbeerd wordt neemt dus de concentratie van deze POPs toe, totdat het maximum bereikt wordt aan het eind van het dooierzakstadium. Dit verklaart waarom de effecten pas na dat moment zichtbaar worden. Het model laat ook zien dat de POP concentraties voornamelijk door verdunning dalen zodra het visje begint te eten en toeneemt in gewicht. Wanneer de larven klaar zijn voor de metamorfose zijn de gehalten zo laag geworden dat er geen effecten meer zullen optreden.
In Hoofdstuk 5 wordt het effect van andere POPs onderzocht die ook veel in het mariene milieu aanwezig zijn. Het betreft PCBs en gebromeerde brandvertragers. Naast het effect van de individuele stoffen is ook het effect van

Samenvatting Eggsposed
190
mengsels van stoffen onderzocht. Hiervoor werd niet alleen een mengsel in het laboratorium gemaakt, maar werden ook extracten gemaakt van tong afkomstig uit de verontreinigde Westerschelde en de Noordzee. Zeker van de extracten van de vis uit de Westerschelde mag verwacht worden dat zij meer stoffen bevatten dan alleen PCBs en gebromeerde vlamvertragers.
De meeste stoffen en mengsels lieten weer hetzelfde type effect zien: sterfte na het dooierzak stadium en geen effect op de metamorfose. De waargenomen effecten van de mengsels konden worden voorspeld aan de hand van de concentraties aan PCBs. De sterfte van de vislarven na het dooierzak stadium is waarschijnlijk het gevolg van de ophoping van deze stoffen in (het vet van) de celmembranen, waardoor deze hun functie verliezen. Dit type effect wordt als basis toxiciteit beschouwd en kan worden veroorzaakt door alle POPs die makkelijk in vet oplossen. Stoffen met dioxine-achtige werking zijn vele malen giftiger. De resultaten uit hoofdstuk 5 laten verder zien dat de POP gehalten van tong uit de Westerschelde veel lager zijn dan de gehalten waarbij doorgifte aan de eieren tot effecten op de larven zal leiden. In tong uit de Noordzee zijn deze gehalten nog 10 maal lager. Elders in Europa zijn wel locaties bekend waar verwacht kan worden dat de voortplanting van de vissen daar verstoord is. Het betreft dan wel zwaar verontreinigde gebieden.
Hoofdstuk 6 richt zich op de Europese aal, de paling. In vergelijking tot vóór 1980 zijn de aantallen jonge alen (glasaal) die jaarlijks vanuit zee het zoete water intrekken met ruim 90% afgenomen. In Hoofdstuk 6 wordt onderzocht of het mogelijk is dat deze afname (deels) veroorzaakt wordt door in de eieren aanwezige POPs. De aal is potentieel gevoelig omdat deze vis zich slechts eenmaal in zijn leven voortplant en daarvoor lange tijd heeft gehad om potentieel giftige stoffen op te hopen in het vetrijke weefsel. Omdat de aal niet eet tijdens de ruim 5000 km lange, maanden durende migratie naar de Sargasso zee zal gewichtsverlies optreden. Op basis van elders uitgevoerde zwemproeven met alen wordt geschat dat door dit gewichtsverlies de concentratie van slecht wateroplosbare POPs in het aalweefsel met 33 tot 100% toeneemt voordat de POPs aan de eieren worden doorgegeven. Omdat aal (nog) niet gekweekt kan worden, kan niet worden vastgesteld hoe gevoelig aallarven voor POPs zijn. Deze gevoeligheid is daarom geschat op basis van gegevens van andere vissoorten. Wanneer wordt aangenomen dat een aal(larve) tot de meest gevoelige vissoorten behoort, dan zijn de gehalten van dioxine-achtige POPs zoals aanwezig in alen uit verontreinigde locaties in Nederland en België hoog genoeg om sterfte onder de larven te veroorzaken.
In de vorige hoofdstukken is vastgesteld dat larvale sterfte kan voorkomen bij vissen van ernstig vervuilde locaties. Hoofdstuk 7 onderzoekt of dergelijke

Eggsposed Samenvatting
191
sterfte van invloed kan zijn op de ontwikkeling van een vispopulatie, en welke soortspecifieke eigenschappen daarbij een rol spelen. Het effect zou groter kunnen zijn als een populatie ook nog zwaar (over)bevist wordt. Het rekenmodel dat hiervoor werd opgesteld op basis van de biologie van de tong, laat zien dat juist platvispopulaties goed bestand zijn tegen larvale sterfte. Dit hangt samen met de grote hoeveelheden eieren die geproduceerd worden (enkele honderden per gram lichaamsgewicht) en het gegeven dat de jonge visjes zich na de metamorfose op de bodem van de opgroeigebieden verzamelen. Doordat de ruimte in de opgroeigebieden beperkt is, vindt hier hoe dan ook grote sterfte plaats onder de pas gemetamorfoseerde visjes. Door gifstoffen veroorzaakte sterfte onder de larven heeft daardoor weinig invloed. Dit blijft zelfs het geval bij extreme visserijdruk. Uit deze analyse blijkt dat populaties van soorten die weinig eieren produceren en waarbij geen dichtheidsafhankelijk sterfte onder de jongen optreedt het meest kwetsbaar zijn voor de effecten van larvale sterfte en visserijdruk.
In Hoofdstuk 8 worden de bevindingen uit de voorgaande hoofdstukken gecombineerd om tot algemene conclusies te komen en aanbevelingen voor verder onderzoek te formuleren. Omdat duidelijk is dat de grootste effecten van POPs op vislarven tot uiting komen aan het eind van het dooierzakstadium, wordt in laboratoriumtesten die eerder worden beëindigd de giftigheid onderschat. Dit is vooral een risico als de giftigheid moet worden bepaald van (mengsels met) onbekende stoffen, zoals mogelijk aanwezig in afvalwater of sedimenten. Ook wordt besproken waarom de resultaten in dit proefschrift waarschijnlijk een onderschatting zijn van de effecten die kunnen optreden wanneer de moedervis vóór het leggen van de eieren aan de POPs zou zijn blootgesteld. In dit geval zijn de gifstoffen immers al in het ei aanwezig voordat de bevruchting plaatsvindt. Bovendien worden ook door de moedervis aangemaakte afbraakproducten van gifstoffen (metabolieten) aan de eieren doorgegeven. Daarnaast kan de kwaliteit van de eieren minder zijn wanneer de moedervis zelf last ondervindt van de POPs.
Uit een vergelijking van verschillende vissoorten lijkt de Groenlandse haai bijzonder kwetsbaar voor de effecten van door de moedervis aan de jongen doorgegeven POPs. Vanwege hun plaats aan de top van de voedselketen bevat het weefsel van deze haaien hoge POP concentraties, die aan de eieren worden doorgegeven. Bovendien verblijven de jongen gedurende hun hele ontwikkeling tot juveniel in de moedervis. Na het verteren van de dooierzak voeden zij zich met de niet ontwikkelde eieren en mogelijk met andere jongen. De blootstelling blijft daardoor hoog, zeker in vergelijking met vissen waarvan de ontwikkeling in relatief schoon water plaatsvindt, met relatief schoon voedsel. Bovendien is het aantal jongen per worp beperkt, waardoor het onwaarschijnlijk is dat

Samenvatting Eggsposed
192
vroegtijdige sterfte van een deel van de jongen door minder dichtheidsafhankelijke sterfte wordt gecompenseerd. Nader onderzoek naar de effecten van POPs op de voortplanting van de Groenlandse haai wordt daarom aanbevolen.

193
Dankwoord
’Keep the blanket around you’, the boy said. ‘You’ll not fish without eating while I’m alive’.
‘Then live a long time and take care of yourself’, the old man said.
Ernest Hemmingway: The Old Man and the Sea

Dankwoord Eggsposed
194
In 2006 besloot ik om dit promotietraject te starten. Het is nu duidelijk dat ik de tijd, aandacht en energie die nodig waren om deze klus naast mijn normale werk te voltooien ernstig heb onderschat. De knelpunten die daardoor ontstonden konden alleen worden opgelost door (vaak ongevraagd) een beroep te doen op het begrip en de loyaliteit van de mensen in mijn directe omgeving.
Gerda, mijn lief, jij leverde zonder twijfel het grootste offer. Dat ik vaak ’s avonds en in de weekenden op het lab was, was misschien nog het minst erg. Maar zeker de laatste 2 jaar waren mijn gedachten ook als ik wel thuis was, vaak niet bij de zaken die we eigenlijk samen zouden moeten doen. Zonder jou was dit proefschrift er misschien ook wel gekomen. Maar dan zeker ten koste van meer dan een proefschrift ooit waard kan zijn. Deze promotie is daarmee onze gezamenlijke prestatie. Wat zal ik trots zijn als ik tijdens de ceremonie jou, geflankeerd door onze prachtige zonen Lars en Ivar op de eerste rij zie zitten. Ik draag dit proefschrift aan jullie op, maar eigenlijk verdien jij het om als co-auteur vermeld te worden. En dat staat dan nog los van je bijdrage aan de vormgeving van de omslag van het proefschrift.
De weg die uiteindelijk naar dit proefschrift heeft geleid begint in 1987 toen ik door het toenmalig afdelingshoofd Willem Christiaan (Pim) de Kock als veldmedewerker ecologisch onderzoek werd aangenomen bij TNO in Den Helder. Zijn eigenzinnige kijk op de wereld heb ik altijd erg gewaardeerd. Ik had me er daarom op verheugd om dit proefschrift straks aan Pim te kunnen aanbieden. Helaas overleed hij onverwacht in de week dat ik het akkoord van de leescommissie ontving.
Na mij aanstelling werd Martin Scholten al snel hoofd van de onderzoeksgroep. Martin, door het vertrouwen en de ruimte die je me gaf in het werk en de persoonlijke manier waarop je mij coachte, heb je een enorme bijdrage geleverd aan mijn ontwikkeling. Dit betrof niet alleen praktische zaken als projectleiding en rapportage, maar zeker ook het besef dat je met goed teamwerk en een gezamenlijk doel het meeste kunt bereiken. In 2001 bepleitte jij met succes bij de directie van TNO dat ik, hoewel ‘slechts’ opgeleid op HBO niveau, op academisch niveau functioneerde en als zodanig beloond diende te worden. Ik was destijds al erg vereerd met deze erkenning, maar zou pas jaren later beseffen hoe bijzonder dat werkelijk was.
Mede gestimuleerd door Martin overwoog ik rond diezelfde periode al om een promotietraject te starten. Omdat toen echter ook Lars en later Ivar werden geboren, is dit plan niet doorgezet en verdween deze ambitie naar de achtergrond van mijn volle leven. Het werd echter weer aangewakkerd door de

Eggsposed Dankwoord
195
inspirerende gesprekken met mijn nieuwe IMARES collega Charlotte Deerenberg tijdens een lange autorit naar Zeeland in september 2006.
Niet lang daarna bezocht ik voor de eerste keer de Wageningen Universiteit om mijn plannen te bespreken met Tinka Murk, mijn latere promotor. Tinka, je wees me er toen direct al op dat mijn ideeën in elk geval nooit binnen één promotietraject verwezenlijkt zouden kunnen worden en adviseerde mij om de ambitie sterk in te perken. Zonder dat advies was mijn proefschrift nooit gereed gekomen. Ook wil ik je danken voor de effectieve training in het schrijven van wetenschappelijke publicaties, en voor je vertrouwen en bemoedigende woorden waarmee je me door de dipjes heen trok als de dubbele baan even teveel werd. Vanaf het begin inspireren onze gesprekken tot nieuwe ideeën en ik weet zeker dat we in de toekomst nog veel mooi werk samen zullen verzetten.
Vanaf 2007 was ik dus echt begonnen aan mijn promotieonderzoek. Omdat hiervoor slechts zeer beperkte ruimte was (binnen het EU-project ‘Modelkey’), was ik voor de uitvoering van de langdurige en arbeidsintensieve experimenten sterk afhankelijk van de inzet van vrijwilligers en studenten. Dear Henrique, Elisa, Astrid, Maria, Merlijn, Mekuria, and Bastiaan, your contributions were crucial for the research presented in this thesis. Apart from the practical work that you did, I really enjoyed your presence and sharing thoughts about science, art, politics, life, and the effect of swearing on the quality of rice. Thanks for that. I hope that you will be able to develop the personal career that fulfils your dreams.
Ewout Blom en Jan van der Heul wil ik danken voor het leveren van de tongeieren, en voor de adviezen die zij samen met Andries Kamstra gaven over de verzorging van de eieren en de larven. Mede hierdoor kon de p-ELS bioassay behoorlijk snel worden ontwikkeld.
Hans van den Berg toonde zich een vakman door in monsters van slechts enkele milligrammen tonglarven toch betrouwbare TEQ gehalten te kunnen bepalen. Christiaan Kwadijk, en de analisten van het chemisch lab van IMARES in IJmuiden leverden vergelijkbare prestaties door dezelfde monsters chemisch te analyseren. Allemaal hartelijk bedankt, en excuses voor al die keren dat ik toch nog een keer een extra analyse wilde of de meetgegevens, eigenwijs als ik ben, nog een keer extra ging doorrekenen. Michiel Kotterman dank ik voor de adviezen over het gedrag van PCBs, en de inspirerende discussies over al dan niet proefschrift gerelateerde zaken.
Naast deze mensen die vrij direct betrokken waren bij mijn onderzoek, zijn er nog veel meer collega’s waar ik veel aan heb gehad. Al mijn collega’s op locatie

Dankwoord Eggsposed
196
Ambachtsweg dank ik voor hun interesse in mijn werk en de bereidwilligheid om altijd bij te springen als dat nodig was, maar bovenal voor het begrip voor het feit dat ik me het laatste jaar wat afzonderde om zaken af te kunnen ronden. Ik wil hierbij Klaas Kaag en Andrea Sneekes met name noemen die het meeste last hebben gehad van mijn verminderde betrokkenheid omdat onze werkvelden sterk overlappen. Zij pakten zonder morren op wat ik liet liggen of had laten vallen.
Uit de vele goede collega’s heb ik Gerrit Hoornsman en Diana Slijkerman gevraagd mij als paranimfen te vergezellen tijdens de promotieceremonie. Siem en Diaan, ik ben vereerd dat jullie dit met zoveel enthousiasme hebben aanvaard.
Gedurende mijn promotieperiode heb ik onder verschillende (ad-interim) afdelingshoofden gewerkt. In chronologische volgorde: John, Henk, Floris en Mark. Hoewel de middelen beperkt waren, hebben Julie mij toch enige ruimte gelaten om mijn promotieonderzoek te volbrengen. Bedankt voor dit vertrouwen.
De opmaak van dit proefschrift is mij uit handen genomen door een collega die niet met naam genoemd wil worden, maar die ik daarvoor toch heel hartelijk wil bedanken. Liesbeth, je weet wel wie ik bedoel.
IMARES en de Wageningen Universiteit dank ik voor de bijdragen aan de kosten voor het produceren van dit proefschrift en de overige zaken die samenhangen met de afronding van een promotie.
De leden van de promotiecommissie wil ik danken voor de tijd die zij hebben besteed aan het kritisch lezen van dit proefschrift, vermoedelijk gedeeltelijk in hun vakantietijd.
Mijn familie en vrienden waren op de achtergrond belangrijk door met interesse te informeren naar de voortgang en de bevindingen van het onderzoek. Ik ben er bovendien van overtuigd dat elk onderzoek erbij gebaat is wanneer de relevantie van de resultaten met niet-wetenschappers wordt bediscussieerd. En dat kun je natuurlijk nergens beter doen dan met vrienden of familie onder het genot van een drankje, met vismaten aan het water, of met je volleybalteam in een kantine.
Helemaal aan de wieg van dit alles staan natuurlijk mijn ouders. Pap en mam, jullie hebben jullie kinderen altijd gestimuleerd en geholpen om zichzelf te ontwikkelen, met de vrijheid om daarin zelf de richting te bepalen. De belangstelling voor het onderwaterleven was bij mij al jong aanwezig en ik kreeg

Eggsposed Dankwoord
197
alle ruimte om dit met aquaria, netten en hengels uit te bouwen. De kliederzooi die dat met mijn experimenteerdrang regelmatig opleverde, leidde wel eens tot gemopper maar nooit tot een verbod. Deze ‘praktijkervaring’, in combinatie met de bij de opvoeding meegegeven instelling dat je (bijna) alles kunt als je maar hard genoeg je best doet (“Kan Niet ligt op het kerkhof, en Wil Niet ligt ernaast.”), is een goede opmaat gebleken voor mijn professionele loopbaan. Hierdoor ben ik nu al meer dan 25 jaar werkzaam in een vakgebied waarin werken vaak niet als werk voelt. Mij promotie is hiervan de bekroning die ook op jullie afstraalt.
Als je zoals ik bijna 50 jaar oud bent, is het onmogelijk om in een dankwoord, dat nu eigenlijk al te lang geworden is, volledig te zijn. Daarom wil ik tenslotte iedereen bedanken die niet met name genoemd is, maar wel in mijn gedachten is nu ik dit schrijf. Bedankt voor de bijdrage die jullie op welke wijze dan ook hebben geleverd aan mijn persoonlijke ontwikkeling en mijn plezier in het leven.


199
Curriculum Vitae

Curriculum vitae Eggsposed
200
Edwin Matheus Foekema werd geboren op 7 November 1963 in Wormerveer, tegenwoordig opgenomen in de gemeente Zaanstad. Na afronding van zijn HAVO opleiding aan het St. Michael College te Zaandam, begon hij in 1981 aan de HBO-opleiding tot Ecologisch analist aan het Ir. Van den Broek Instituut te Amsterdam. In 1984 ronde hij deze opleiding met succes af en werkte vervolgens tot 1986 als analist bij de vakgroep Aquatische Ecologie van de Universiteit van Amsterdam. Gedreven door het geringe banenaanbod binnen zijn vakgebied tijdens de toen heersende economische crisis, volgde hij daarna gedurende 6 maanden een omscholingscursus in de ICT. Direct na het behalen van dit certificaat kon hij aan het werk als programmeur bij het NIPO in Amsterdam. Daar werd al snel duidelijk dat het verwerken van enquête-gegevens hem weinig voldoening gaf. Groot was dan ook de opluchting toen hij binnen 3 maanden deze werkplek kon verlaten om te starten als ecologisch veldmedewerker bij de afdeling Maatschappelijke Technologie van TNO, met als standplaats de Ambachtsweg in Den Helder. Na 4 jaar ervaring te hebben opgedaan als projectmedewerker in verschillende takken van het milieuonderzoek, werd hij in 1991 aangesteld als projectleider van experimenteel ecotoxicologisch onderzoek. Sindsdien heeft hij leiding gegeven aan een veelvoud van projecten op het gebied van ecotoxicologie, veelal met een experimenteel karakter en vaak uitgevoerd in experimentele ecosystemen (mesocosms). In 1996 behaalde hij in deeltijd het diploma HLO-milieuchemie aan de Hogeschool IJsselland te Deventer. Hij zet zijn werk als onderzoeker en projectleider voort bij IMARES Wageningen UR, het instituut dat in 2006 werd opgericht door een fusie van het RIVO, onderdelen van Alterra en de TNO afdeling aan de Ambachtsweg in Den Helder.

201
Publications

Publications Eggsposed
202
Scientific publications
Vries, P. de; Tamis, J.E.; Foekema, E.M.; Klok, T.C.; Murk, A.J. (2013) Towards quantitative ecological risk assessment of elevated carbon dioxide levels in the marine environment (online first) Marine Pollution Bulletin.
Besseling, E.; Wegner, A.; Foekema, E.M.; Heuvel_Greve, van den, M.J.; Koelmans, A.A. (2013) Effects of microplastic on fitness and PCB bioaccumulation by the lugworm Arenicola marina Environmental Science and Technology 47 (1). - p. 593 - 600.
Foekema, E.M.; Gruijter, C. de; Mergia, M.T.; Franeker, J.A. van; Murk, A.J.; Koelmans, A.A. (2013) Plastic in North Sea Fish Environmental Science and Technology 47 (15). - p. 8818 - 8824.
Koelmans, A.A.; Besseling, E.; Wegner, A.; Foekema, E.M. (2013) Plastic as a Carrier of POPs to Aquatic Organisms: A Model Analysis Environmental Science and Technology 47 (14). - p. 7812 - 7820.
Foekema, E.M.; Fischer, A.; Lopez Parron, M.; Kwadijk, C.; Vries, P. de; Murk, A.J. (2012) Toxic concentrations in fish early life stages peak at a critical moment. Environmental Toxicology and Chemistry 31 (6). - p. 1381 - 1390.
Wegner, A.; Besseling, E.; Foekema, E.M.; Kamermans, P.; Koelmans, A.A. (2012) Effects of nanopolystyrene on the feeding behavior of the blue mussel (Mytilus edulis L.) Environmental Toxicology and Chemistry 31 (11). - p. 2490 - 2497.
Hein, M.; Rotter, S.; Schmitt-Jansen, M.; Ohe, P.C. von der; Brack, W.; Deckere, E. de; Zwart, D. de; Foekema, E.M. (2010) MODELKEY Key findings and recommendations for reaching the EU Water Framework Directive's quality objectives Umweltwissenschaften und Schadstoff-Forschung 22 (3). - p. 217 - 228.
Brack, W.; Apitz, S.E.; Borchardt, D.; Brils, J.; Cardos, A.C.; Foekema, E.M.; Verdonschot, P.F.M. (2009) Toward a Holistic and Risk-Based Management of European River Basins Integrated Environmental Assessment and Management 5 (1). - p. 5 - 10.
Foekema, E.M.; Deerenberg, C.M.; Murk, A.J. (2008) Prolonged ELS test with the marine flatfish sole (Solea solea) shows delayed toxic effects of previous exposure to PCB 126Aquatic Toxicology 90 (3). - p. 197 - 203.
Foekema E.M (2005): Improving environmental realism in aquatic pond mesocosms. SETAC-Globe 5(5):29-30
Hamwijk, C.; Schouten, A.; Foekema, E.M.; Ravenberg, J.C.; Collombon, M.T.; Schmidt, K.; Kugler, M. (2005) Monitoring of the booster biocide dichlofluanid in water and marine sediment of Greek marinas. Chemosphere 60 (9). - p. 1316 - 1324
Foekema E.M., N.H.B.M. Kaag, D.M. van Hussel, R.G. Jak, M.C.Th. Scholten & C. van der Guchte (1997): Mesocosm observations on the ecological reponse of an aquatic community to sediment contamination. Water Sci. and Technol.,37(6-7):249-256.

Eggsposed Publications
203
Kaag N.H.B.M., E.M. Foekema, M.C.Th. Scholten & N.M. van Straalen (1997): Comparison of contaminant accumulation in three species of marine invertebrates with different feeding habits. Environ. Toxicol. Chem. 16: 837-842.
Looise B.A.S., D.A. Holwerda & E.M. Foekema (1996): Induction of gluthathione S-transferase in the freshwater bivalve Sphaerium corneum as a biomarker for short-term toxicity tests? comp. Biochem. Physiol. 113C: 103-107.
Kaag N.H.B.M., E.M. Foekema & C.T. Bowmer (1994): An new approach for testing contaminated marine sediments: fertilization success of lugworms following parental exposure. J. Aquat Ecosyst. Health 3: 177-184.
Bowmer C.T., H.A. Jenner, E. Foekema & M. van der Meer (1994): The detection of chronic biological effects in the marine intertidal bivalve Cerastoderma edule, in model ecosystem studies with pulverised fuel ash: Reproduction and histopathology. Environ. Pollut. 85: 191-204.
Book (chapters)
Scholten M.C.Th., E.M. Foekema, H.P. van Dokkum, N.H.M.B. Kaag & R.G. Jak (2005): Eutrophication management and ecotoxicology. Environmental science series, Spinger-Verlag Berlin Heidelberg 2005
Kramer K.J.M. & E.M. Foekema (2000): The musselmonitor as biological early warning system: The first decade. In: Biomonitors and biomarkers as indicators of environmental change: A Handbook, Vol. II F.M. Buttervorth, M.E. Gonsebatt-Bonaparte & A. Gunatilaka (eds.). Plenum, New York.
Foekema E.M., H.P.M. Schobben, J. Marquenie & M.C.Th. Scholten (1996): A comparison between the toxicity of produced oil and gas condensate using a new sediment bioassay for deposited oil. In: M. Reed & S. Johnsen (eds.), Produced Water 2. Environmental issues and mitigation technologies. Environmental Science Research Vol. 52, Plenum Press, New York. pp. 61-68.
Popular publications
Foekema, E.M.; Winter, H.V.; Kleissen, F.; Slijkerman, D.M.E. (2012) Rwzi-effluent gebruiken voor vismigratie H2O: 45 (1). - p. 34 - 36.
Foekema E.M. (2012): Schaamte. De Nederlandse Vliegvisser, no. 100. Foekema, E.M. (2009) Over fosfaat en de vermeende teloorgang van de voorn. De
Nederlandse Vliegvisser no. 90. - p. 62 - 63. Foekema, E., O. van Hulst & B. Reeze (2008): Massale sterfte onder Aziatische
korfmosselen in de Maas. H2O 41 (3): 45-49 Foekema E.M. (2008): Poor man’s flats. De Nederlandse Vliegvisser, no. 84. P-16-17

Publications Eggsposed
204
Slijkerman D., E. Foekema, R. van der Oost & R. Kampf (2006): Hormoonverstoring bij vis punt van aandacht in het zuiveringsmoeras. H2O 39 (25/26): 34-37
Foekema E.M., A. Sneekes & J. Graansma (2006): Hoog fosfaatgehalte in rwzi-effluenten voorspelt remming van de algengroei. H2O 39 (12): 35-38
Foekema E.M. & C. Karman (2006): Het aandeel van bouwmetaal aan de totale zinkbelasting van het oppervlaktewater in Amsterdam. H2O 39 (1):38-40
Dokkum H. van, E.M. Foekema & J. Graansma (2005): De ecologische impact van het effluent van een rwzi. H2O 38 (22): 45-48
Foekema E., M. Oosterhuis, R. Peters & J. Nonnekens (2005): TIE als opsporingsinstrument bij het zuiveren van afvalwater. H2O
Scholten M. & E. Foekema (1994) Het kuifeendenonderzoek. In: T. Boudwijn, S. Dirksen & M. Ohm (Eds)., zichtbare effecten van onzichtbare stoffen. Overzicht onderzoek aan watervogels in het benedenrivierengebied 1982-1992. Bureau Waardenburg BV/RWS Dir. Zuid-Holland. pp. 9-15.
Reports on mesocosm research (first author only)
Foekema E.M., D.M.E. Slijkerman, P. de Vries, J.E. Tamis, G. Hoornsman, E.M. van der Vlies & W.E. Lewis (2007): The first step towards Dynamic Aquatic Mesocosms: The impact of timing of pesticide application. IMARES report C093/07
Foekema E.M., W.E. Lewis, G. Hoornsman, H. van het Groenewoud & E.M. van der Vlies (2007): Determination of the fate and biological effects of Dodine 400SC in outdoor ponds. TNO IMARES report C111/07
Foekema E.M., D.M.E. Slijkerman, G. Hoornsman, H. van het Groenewoud & E.M. van der Vlies (2005): The determination of the biological effects of a single pulse of technical acetochlor in outdoor ponds. TNO report B&O-DH - R2005/253
Foekema E.M., H.P. van Dokkum, A.C. Sneekes, G. Hoornsman, P. Koelma & A. Kreike (2005):Mesocosm studie naar het effect van rwzi-effluent op het ontvangend oppervlaktewater.Resultaten 2004 en discussie.TNO rapport TNO B&O-DH - R2005/179
Foekema E.M., M.T. Collombon & G. Hoornsman (2004): Determination of the effect of one day exposure to technical acetochlor on Elodea canadensis from an outdoor pond. TNO report R2004/056,
Foekema E.M., D.M.E. Slijkerman, A.C. Sneekes, G. Hoornsman, P. Koelma, A. Kreike & H.P. van Dokkum (2004): Effecten van het effluent van rwzi Oostoever op het ontvangend oppervlaktewater getest in mesocosms.TNO-rapport R2004/058
Foekema E.M., M.T. Collombon & G. Hoornsman (2004): Determination of the effect of a single application of technical acetochlor in a static test on Elodea canadensis from a commercial supplier. TNO report R2004/055
Foekema E.M., M.T. Collombon & G. Hoornsman (2004): Determination of the effect of a single application of technical acetochlor in a static test on Elodea canadensis from an outdoor pond. TNO report R2004/054

Eggsposed Publications
205
Foekema E.M., M.T. Collombon, H. van het Groenewoud, A.C. Sneekes & G. Hoornsman (2003): Determination of the biological effects and fate of technical acetochlor in outdoor ponds according to HARAP (1999) and CLASIC (2002) guidelines. TNO report R2003/373.
Foekema E.M, P.C. Goedhart & G. Hoornsman (2003): De beoordeling van effluent van rwz Ameland met de planktonecoassay en chemische analyses. TNO rapport R2003/036
Foekema E.M., A.C. Sneekes, H. van het Groenewoud, M. Collombon, E.M. van der Vlies, I. van der Velde (2002): Determination of the biological effect and fate of MON 39801 (43% w/w/ Alachlor) in outdoor ponds according to HARAP (1999) and CLASSIC (2000) guidance documents. R2002/280
Foekema E.M., H. van het Groenewoud, A.C. Sneekes, E.M. van der Vlies, M. van der Meer & G. Hoornsman (2002): The fate and biological effects of Rugby 200 CS/cadusafos in small outdoor mesocosms. R2001/562
Foekema E.M., H. van het Groenewoud, G. Hoornsman, A.C. Sneekes & M. van der Meer (2001): The fate and biological effects of Marshal 25 CS/carbosulfan in small outdoor mesocosms. R2001/072
Foekema E.M., R.G. Jak & M.G.D. Smit (2000): Biologische beoordeling van de kwaliteit van uitslagwater uit Flevoland met behulp van de planktonecoassay. R2000/402
Foekema E.M. & G. Hoornsman (1999): Klein-kaliber projectielen in water-sediment systemen.TNO rapport R99/208.
Foekema E.M., R.G. Jak & M.C.Th. Scholten (1999): Determination of the effect of copper on a freshwater plankton community in the plankton ecoassay. TNO report R 99/020
Foekema E.M., R.G. Jak & M.C.Th. Scholten (1999): Determination of the effect of fluoranthene on a freshwater plankton community in the plankton ecoassay. TNO report R 99/019
Foekema E.M., R.G. Jak & M.C.Th. Scholten (1999): Toetsing van enkele veldexperimentele technieken in de mesocosms behandeld met mevinfos. R98/158
Foekema E.M. & R.G. Jak (1999): The fate and biological effects of Flufenacet WG 60 in aquatic indoor microcosms. R99/423
Foekema E.M., N.H.B.M. Kaag & R.G. Jak (1998): De invloed van benzo(a)pyreen op koude tolerantie van de halfgeknotte strandschelp (Spisula subtruncata). TNO-MEP rapport R98/077.
Foekema E.M. & C.T. Bowmer (1998): A 7 day indoor, multispecies plankton assay with the substances Trimangol 80 WP (Maneb) and Penncozeb 80 WP (mancozeb) following TNO Standard Operating Procedures DTOX/MEP/101. V98.172
Foekema E.M. & C.T. Bowmer (1996): The evaluation of contaminated freshwater sediments using experimental mesocosmsystems: De toetsorganismen; biologie, accumulatie en effecten TNO rapport R 96/445

Publications Eggsposed
206
Foekema E.M., R.G. Jak & M.C.Th. Scholten (1996): Toetsing van enkele veldexperimenten in microcosms behandeld met dimethoaat. R96/070
Foekema E.M. & G. Hoornsman (1993): Mesocosmonderzoek ter beoordeling van slib uit het slufterdamdepot. Biologische beschikbaarheid en biologische effecten van contaminanten in slib afkomstig uit het Hollands Diep. R93/027
Foekema E.M., D.P.C. van der Veen & L. van der Vlies (1992): The evaluation of contaminanted freshwater sediment using experimental mesocosm systems No. 13. R92/394
Reports on bioassays and toxicity identification (first author only)
Foekema E.M. & A.C. Sneekes (2010): Bepaling van de toxiciteit van vier diergeneesmiddelen met bioassays. IMARES C059/10
Foekema E.M. & A.C. Sneekes (2007): Assessment of the toxicity of solid coating PV1 in marine environment, using biotests with algae, a rotifer and bacteria. IMARES C059/07
Foekema E.M. & A.C. Sneekes (2006): Nader onderzoek naar de effecten van rwzi-effluent op algen. TNO rapport 2006-DH-R0041
Foekema E.M., G. Hoornsman & G. Mitchell (2005): The determination of the effect concentration of Rugby 200CS (active ingredient: cadusafos) for Daphnia longispina under various food conditions. TNO rapport R&I-A R 2005/032
Foekema E.M. & A.C. Sneekes (2005): Toxiciteitsidentificatie onderzoek met het influent van rwzi Coevorden van zomer 2005. TNO rapport B&O-DH - R2005/258
Foekema E.M. & A.C. Sneekes (2005): Verkenning van de mogelijkheden voor toxiciteitsidentificatieonderzoek (TIE) binnen de CTT. TNO rapport B&O-DH - R2005/252
Foekema E.M., A.C. Sneekes, R. de Vos & R.J.B. Peters (2004): Onderzoek naar de oorzaak van periodieke toxiciteit van efluent van rwzi Enschede-West in januari 2004. TNO-rapport R2004/060
Foekema E.M., R. de Vos & V.G. Blankendaal (2003): Ontwikkeling van TIE technieken voor munitie gerelateerde stoffen. Verslag van de werkzaamheden 2001-2002. TNO-rapport R2004/474
Foekema E.M. & N.H.B.M. Kaag (2001): Biologische beoordeling van baggerspecie uit het Zeehavenkanaal te Delfzijl. R2001/543
Foekema E.M. & J.E. Brils (2000): Toxiciteitsidentificatieonderzoek aan afvalwater van Eurometaal NV te Zaandam. Fase 2: nadere analyse van het gefiltreerde effluent. R2000/486
Foekema E.M., J.E. Brils & R. Bernhard (2000): Toxiciteitsidentificatieonderzoek aan afvalwater van Eurometaal NV te Zaandam. Fase 1: effect van pH-manipulatie, EDTA en filtratie. TNO rapport R2000/403

Eggsposed Publications
207
Reports on water technology related research (first author only)
Foekema, E.M.; Sneekes, A.C.; Koelemij, E.I.; Tjon Atsoi, M.M.G.; Hoornsman, G.; Bakker, A.G. (2012) WIPE: De invloed van moerassystemen op de milieukwaliteit van rwzi effluent en aanbevelingen tot optimalisering Den Helder: IMARES, (Rapport C005/12) - p. [324].
Foekema E.M. & R. Kampf (2005): Overzicht van werkzaamheden in het kader van het Kwekelbaarsjesproject in de periode 1998-2002. TNO rapport B&O-DH - R2005/257
Foekema E.M., A. Weber, D.M.E. Slijkerman, A.C. Sneekes, G. Hoornsman, P. Koelma, A. kreike & H.P. van Dokkum (2004): De invloed van een mosselfilter op de effecten van het effluent van rwzi Oostoever op het ontvangend oppervlaktewater getest in mesocosms. TNO-rapport R2004/059
Foekema E.M., V.G. Blankendaal, P.C. Goedhart & G. Hoornsman (2003): Ecotoxicologische aspecten bij de nabehandeling van RWZI-effluenten met behulp van biomassa kweek. Stichting Toegepast Onderzoek Waterbeheer, Utrecht. Rapport 2003-12.
Foekema E.M., V.G.Blankendaal, P.C. Goedhart & G. Hoornsman (2003): Ecotoxicologische aspecten bij nabehandeling van RWZI-effluenten met behulp van biomassa kweek. TNO-rapport R2003/032
Foekema E.M. & R. Kampf (2002): Gebruik van effluentvan de rwzi de Cocksdorp voor natuurdoeleinden: Onderzoek naar de kweek van watervlooien in het kwekelbaarsjes systeem. DH02/007
Foekema E.M., J.W. Assink & M.C.Th. Scholten (1999): Verkennende studie naar de mogelijkheden van duurzaam waterbeheer van de nieuwe walrussen accommodatie. R99/053
Foekema E.M., G. Hoornsman & M.G.D. Smit (1999): Mosselen en walrusfeces: laboratoriumonderzoek naar filterefficientie. R99/386
Foekema E.M. &. K.J.M. Kramer (1994): Toepassingsmogelijkheden voor de Mosselmonitor bij het WRK waterwinstation ir. Cornelis Biemond. R94/088
Reports on other research topics (first author only)
Foekema, E.M.; Winter, H.V.; Rippen, A.D.; Slijkerman, D.M.E.; Keeken, O.A. van; Kleissen, F. (2011) Pluim voor vismigratie: deelonderzoeken Den Helder: IMARES, (Rapport C078/11).
Foekema, E.M.; Rippen, A.D.; Slijkerman, D.M.E. (2011) Inventarisatie van lozingspluimen als potentiele migratie-barriere Den Helder: IMARES, (Rapport C078/11) - p. 28.

Publications Eggsposed
208
Foekema, E.M.; Winter, H.V.; Kleissen, F.; Keeken, O.A. van; Rippen, A.D.; Slijkerman, D.M.E. (2011) Vismigratie en lozingspluimen: Samenvattend rapport Den Helder: IMARES, (Rapport / IMARES C077/11) - p. 25.
Foekema E.M., D.M.E. Slijkerman & N.H.B.M. Kaag (2007): Mogelijke oorzaken Corbicula sterfte in de Maas: een verkennend onderzoek naar beschikbare informatie in 2006. IMARES rapport C069/07
Foekema E.M., A.C. Sneekes & J.T. van der Wal (2005): De verspreiding van munitiegerelateerde stoffen in verschillende milieucompartimenten. Modelberekeningen met PEARL en SimpleBox. TNO rapport B&O-DH - R2005/185
Foekema E.M. & C.C. Karman (2005): Zink in stedelijk oppervlaktewater TNO rapport B&O-DH - R2005/176
Foekema E.M. & M.G.D. Smit (2004): Mogelijkheden voor het opnemen van veerkracht in het Corophium model. TNO-rapport R2004/479
Foekema E.M., R.G. Jak, J.E. Tamis & G.Hoornsman (2000): De potentiele milieubezwaarlijkheid voor het aquatisch milieu van verfdeeltjes afgespoten van drijvende objecten R2000/487
Foekema E.M. & H.P. van Dokkum (2000): Evaluatie van de blauwalgenbloei in de geestmerambachtplas in 1999. R2000/177
Foekema E.M., E.M.van der Vlies & M.G.D. Smit (2000): Inzet van de Mosselmonitor bij de proefboring consessie Middelie. TNO rapport R2000/201.
Foekema E.M. & H. van het Groenewoud (1998): Environmental baseline survey F2 Hanzefield. R98/499
Foekema E.M., A. Bos, P. Verstappen & C.C. Karman (1998): Field validation of the CHARM-model. TNO report R98/317
Foekema E.M., S.L. Huwer, C.C. Karman & M.C.Th. Scholten (1997): Pilot study on the dose-effect responses of petroleum hydrocarbons in sediments (SLURP*olie). Report phase III: Evaluation of results. R97/419
Foekema E.M., M.C.Th. Scholten, R.G. Jak, P. Hernandez, D. Prats, B. Clement, G. Blake, G. Hoornsman, D.P.C. v.d. Veen, M. v.d. Meer, J.C. Asensi, M.J. Navarro & P. Ant (1996): Phosphorus-algal response curves: a critical analysis of the Vollenweider model. Technical report of the 1995 research programma. R96/030
Foekema E.M. & D.P.C. van der Veen (1996): Plaatsgebonden blootstelling van paling in het Ketelmeer.DH96/23
Foekema E.M., H.P.M. Schobben, E.A. Vik & A. Bos (1996): Experimental validation of the CHARM-model. CHARM Technical Note 37. DH96/07
Foekema E.M., G. Hoornsman, D.P.C. van der Veen & E.M. van der Vlies (1995): Biologische beschikbaarheid van zink in produktiewater. R95/108a
Foekema E.M., M.C.Th. Scholten, G. Hoornsman, M. v.d. Meer & B. Born (1993): Milieuaspecten maricultuurcentrum Neeltje Jans. R93/029
Foekema E. & M.C.Th. Scholten (1988): De effecten van een verhoogd gehalte aan microverontreinigingen in het voedsel, op de conditie en voortplanting van Kuifeenden. TNO-rapport R88/145.




















