
Ruimtelijke distributie van bodemfungi: evidentie voor de...
94
Faculteit Wetenschappen Vakgroep Biologie Onderzoeksgroep Terrestrische Ecologie Academiejaar 2012-2013 Ruimtelijke distributie van bodemfungi: evidentie voor de Janzen-Connell hypothese? Charlotte Watteyn Scriptie voorgelegd tot het behalen van de graad Master in de Biologie Promoter: Dr. Valérie Lehouck Co-promoter: Dr. Eduardo de la Peña Begeleiders: Dr. Valérie Lehouck Dr. Eduardo de la Peña
Transcript of Ruimtelijke distributie van bodemfungi: evidentie voor de...

Faculteit Wetenschappen
Vakgroep Biologie
Onderzoeksgroep Terrestrische Ecologie
Academiejaar 2012-2013
Ruimtelijke distributie van bodemfungi: evidentie voor de
Janzen-Connell hypothese?
Charlotte Watteyn
Scriptie voorgelegd tot het behalen
van de graad Master in de Biologie
Promoter: Dr. Valérie Lehouck
Co-promoter: Dr. Eduardo de la Peña
Begeleiders: Dr. Valérie Lehouck
Dr. Eduardo de la Peña

Dankwoord
Dankwoord
Om deze thesis tot een goed einde te brengen heb ik een lange, en dit mag ook letterlijk genomen worden,
reis achter de rug. Ik zou dan ook graag een woordje van dank richten aan de mensen die mij langsheen
dit leerrijke pad geholpen en gesteund hebben.
Allereerst wil ik Valérie en Eduardo bedanken voor de uitstekende begeleiding. Ik kon bij hen steeds
terecht met vragen of problemen. Valérie, bedankt voor het nalezen en verbeteren van mijn teksten en het
helpen bij de dataverwerking, die het geduld van een mens toch vaak op de proef stelt. Eduardo, een dikke
merci om mij te helpen met het veldwerk in Kenia en al de rompslomp van vergunningen te regelen om de
wortel- en bodemstalen zo legaal mogelijk op het vliegtuig te krijgen. Ook in België stond je steeds paraat
me te helpen waar nodig.
Peter, thank you for helping me with the fieldwork! Without you as a guide I would definitely be lost in
the Taita Hills. You are a quit but friendly and hard-working person who learned me a lot about the huge
amount of Taita tree and bird species. Rebecca and Mwadime, after I finally found the Taita Research
Station in Wundanyi, you welcomed me with open arms. I really enjoyed my stay with you guys. Thomas
en Stien, bedankt voor de fantastische tijd in de Taita Hills. Na het veldwerk terug krachten opdoen met
een overheerlijke chapatti, de avondlijke bush baby ‘jacht’ en de leuke uitstapjes op zondag zijn slechts
enkele van de vele toffe momenten in het vriendelijke en vredevolle stadje Wundanyi.
Jane en Pieter, jullie hebben me enorm geholpen met de moleculaire analyses. Zonder jullie kennis en het
mogen gebruiken van de chique toestellen van het ILVO, zou ik ‘mijn schimmeltjes’ nooit zo efficiënt
geïdentificeerd hebben. Ook met vragen kon ik steeds bij jullie terecht.
Mama, papa en zussen, jullie mogen uiteraard niet ontbreken in het lijstje. Jullie waren steeds bereid mij
te helpen waar nodig. Het nemen van foto’s, het nalezen van teksten en het regelmatig aanhoren van mijn
‘gezaag’ was jullie nooit te veel gevraagd. Het is niet echt jullie vakgebied, maar toch waren jullie steeds
geïnteresseerd in alles wat ik deed. Ook bij mijn beste maatjes kon ik steeds terecht voor een babbel en de
nodige ontspanningsmomenten.
Bedankt allemaal!
Charlotte

Lijst met afkortingen en definities
ANOSIM = Analysis Of Similarity
BLAST = Basic Local Alignment Sequence Tool
CFU = Colony Forming Units
= Aantal getelde kolonies
Cha = Bosfragment Chawia
CNDD = Conspecific Negative Density / Distance Dependence
= Conspecifieke negatieve densiteits- / afstandsafhankelijkheid
DBH = Diameter at Breast Height
= Diameter, gemeten op borsthoogte (± 130 cm)
Fur = Bosfragment Fururu
Identity (%) = Overeenkomstpercentage tussen eigen DNA-sequentie en een in de databank
aanwezige DNA-sequentie, gebaseerd op de nucleotide-nucleotide alineëring
ITS = Internal Transcribed Spacer
= DNA-sequentie, algemeen aanwezig in rDNA van fungi
J-C = Janzen-Connell
Mac = Bosfragment Macha
MMMeans = Michaelis-Menten Means
= Biodiversiteitsindex
Mwa = Bosfragment Mwachora
Ndi = Bosfragment Ndiwenyi
Nga = Bosfragment Ngangao
NMDS = Non-metric Multi-Dimensional Scaling
NTP = Nucleoside tri-fosfaat
PCA = Principal Component Analysis
PCR = Polymerase Chain Reaction
= Polymerase kettingreactie

Lijst met afkortingen en definities
Query cover (%) = Percentage van de eigen DNA-sequentie die gebruikt wordt bij het zoeken naar
naar overeenkomst met een DNA-sequentie in de databank.
R = Nearest neighbour index (Nearest neighbor analyse in ArcGIS)
Of
= Teststatistiek die de mate van verschil weergeeft in gemeenschaps-
samenstelling. De waarde varieert tussen -1 en 1. Een waarde groter dan 1 wijst op
variatie in gemeenschapssamenstelling (NMDS analyse in Primer).
rDNA = Ribsosomaal DNA
Relatieve densiteit = De ratio van de densiteit van adulte X. monospora bomen ten opzichte
van de densiteit aan alle adulte bomen (X. monospora en andere soorten)
Relatieve dominantie = De ratio van de som van de stamoppervlaktes (berekend op basis van de
gemeten DBH) van alle adulte X. monospora bomen ten opzichte van de
som van de stamoppervlaktes van alle aanwezige bomen
SD = Standaard Deviatie

Inhoudsopgave
Inhoudsopgave
1. Inleiding .......................................................................................................... 1
1.1.De onschatbare waarde van biodiversiteit ............................................... 1
1.1.1. Biotische interacties en tropische plantendiversiteit ..................... 2
1.2. Plant-bodem terugkoppelingsmechanismen ........................................... 4
1.2.1. De Janzen-Connell hypothese ....................................................... 6
1.2.1.1. Plant-pathogeen interacties .............................................. 8
1.3. Habitatfragmentatie ................................................................................ 10
1.4. Aanleiding tot deze masterproef ............................................................. 12
2. Doelstellingen ................................................................................................. 14
3. Materiaal en methoden ................................................................................... 15
3.1.Studiegebied en studiesoort ..................................................................... 15
3.1.1. Studiegebied ................................................................................. 15
3.1.2. Studiesoort .................................................................................... 16
3.2.Studiedesign ............................................................................................ 18
3.2.1. Staalname ...................................................................................... 18
3.2.2. Karakterisering bodemfungi ......................................................... 21
3.3.Statistische dataverwerking ..................................................................... 23
3.3.1. Ruimtelijke distributie van X. monospora .................................... 23
3.3.2. Ruimtelijke distributie van bodemfungi gemeenschap ................. 24
3.3.2.1. Soortensamenstelling en diversiteit ................................. 24
3.3.2.2. Gemeenschapsanalyse ...................................................... 24
4. Resultaten ....................................................................................................... 26
4.1.Ruimtelijke distributie X. monospora ...................................................... 26
4.2.Ruimtelijke distributie bodemfungi gemeenschap .................................. 28
4.2.1. Soortensamenstelling en diversiteit .............................................. 28
4.2.2. Gemeenschapsanalyse .................................................................. 31
4.2.2.1.Variatie tussen fragmenten en plots .................................. 33
4.2.2.2.Variatie in functie van de afstand tot moederboom .......... 36
4.3.Verband X. monospora en bodemfungi distributie .................................. 38
5. Discussie ......................................................................................................... 39
5.1. Ruimtelijke distributie bodemfungi gemeenschap en X. monospora .... 39
5.1.1. Functionele diversiteit ............................................................. 39
5.1.2. Variatie tussen fragmenten ...................................................... 40
5.1.3. Variatie tussen plots ................................................................. 42
5.1.4. Variatie in functie van afstand tot moederboom...................... 43
5.2. Studiedesign: moeilijkheden en eventuele verbeteringen ...................... 46
5.2.1. Staalname ................................................................................. 46
5.2.2. Karakterisering bodemfungi .................................................... 47

Inhoudsopgave
5.3. Suggesties voor verder onderzoek .......................................................... 48
6. Conclusie ........................................................................................................ 50
7. Samenvatting .................................................................................................. 51
8. Summary ......................................................................................................... 54
9. Referenties ...................................................................................................... 56
10. Bijlagen ........................................................................................................... 62

Inleiding
- 1 -
1. Inleiding
1.1. De onschatbare waarde van biodiversiteit
Biologische diversiteit, kortweg biodiversiteit, is meer dan alleen de diversiteit aan soorten.
Het wijst enerzijds op de variabiliteit aan levende organismen, afkomstig uit zeer
uiteenlopende habitats, maar anderzijds ook op de ecologische systemen waarvan zij deel
uitmaken. Dit omvat zowel diversiteit binnen de soort, tussen de soorten als van ecosystemen.
Biodiversiteit wijst dus op de verscheidenheid aan leven op onze planeet, en dit op drie
verschillende niveaus: genetisch, soort- en ecosysteemniveau (Loreau, 2010). Het onderwerp
‘biodiversiteit’ komt de laatste jaren steeds vaker aan bod in de media. Jammer genoeg is de
reden hiervoor de wereldwijde achteruitgang als gevolg van een alsmaar toenemende impact
van menselijke activiteiten.
Binnen een ecosysteem interageren biotische en abiotische componenten met elkaar door
middel van een brede range aan biologische, fysische en chemische processen (Loreau, 2010).
De (a)biotische samenstelling, structuur en functionaliteit van een ecosysteem wordt bijgevolg
bepaald door een aantal gerelateerde omgevingsfactoren, zoals nutriëntenbeschikbaarheid,
temperatuur, lichtintensiteit, begrazingsintensiteit en populatiedensiteit. Veranderingen in
deze factoren ten gevolge van verstoringen (ziektes, invasies van exotische soorten,
klimaatsverandering) destabiliseren de huidige dynamiek van ecosystemen, en aangezien de
aanwezigheid van gezonde populaties van diverse soorten een belangrijke voorwaarde is voor
het optimaal functioneren van ecosystemen, fungeert biodiversiteit als een buffer tegen deze
verstoringen. Het idee dat een toename in biodiversiteit leidt tot een toename in stabiliteit van
ecosystemen vormt dan ook de basis van de biodiversiteit-stabiliteit hypothese. Een
ecosysteem wordt gekenmerkt door interacties tussen soorten onderling, waardoor de
achteruitgang en eventuele extinctie van een bepaalde soort kan leiden tot de achteruitgang of
secundaire extinctie (co-extinctie) van andere soorten enerzijds (Koh et al., 2004) en de
vooruitgang van competitoren of prooien anderzijds (Morris, 2010). De voornaamste
drijfveren achter het verlies van biodiversiteit binnen een ecosysteem, en zodoende ook
ecosysteemfuncties en -diensten, zijn veranderingen in landgebruik, habitatverlies en -
fragmentatie, introductie van exotische soorten, vervuiling, overexploitatie (o.a. jacht en
visvangst) en meer recent ook klimaatsverandering (Morris, 2010). Hoe meer soorten
verdwijnen, hoe groter de kans op het wijzigen of verloren gaan van interacties en hoe groter
de negatieve impact op de structuur en functionaliteit van een ecosysteem. Dit in acht

Inleiding
- 2 -
genomen, kan verondersteld worden dat een soortenrijk systeem stabieler en robuuster is en
dus beter bestand is tegen externe invloeden (Hooper et al., 2005; Hector & Bagchi, 2007;
Loreau et al., 2010; Mori et al., 2013).
1.1.1. Biotische interacties en tropische plantendiversiteit
De afname in biologische diversiteit met toenemende breedtegraad, is een van de meest
opmerkelijke waargenomen patronen op onze planeet (Mittelbach et al., 2007; Schemske et
al., 2009). Een meta-analyse van Hillebrand (2004) toonde twee zaken aan. Ten eerste, de
soortenrijkdom van de meeste taxonomische groepen stijgt naar de evenaar toe en ten tweede,
deze trend verschilt niet tussen de Noordelijke of Zuidelijke hemisfeer, mariene of
terrestrische ecosystemen, taxa van verschillende trofische niveaus en ectotherme of
endotherme taxa. Ondanks het feit dat men al tientallen jaren op zoek is naar een universeel
aanvaardbare verklaring voor deze latitudinale biodiversiteitsgradiënt, bestaat er tot op heden
nog geen eensgezindheid over de achterliggende mechanismen ervan. Een antwoord bieden
op de vraag hoe het mogelijk is dat honderden plantensoorten samen kunnen voorkomen op
slechts één hectare tropisch bos is bijgevolg niet eenvoudig. Verscheidene meta-analyses (o.a.
Hill & Hill, 2001; Willig et al., 2003; Leigh et al., 2004) hebben geprobeerd de enorme
hoeveelheid aan hypothesen te bundelen. Eén van de vele opgestelde classificaties van
mogelijke mechanismen achter patronen van co-existentie wordt weergegeven in figuur 1.1
(Barot, 2004). Deze mechanismen zijn veelal compatibel en moeten idealiter in combinatie
met elkaar worden onderzocht (Wright, 2002). De focus in dit type van onderzoek wordt vaak
gelegd op zaailingen omdat de overgang van zaad naar reproductieve adult het meest cruciale
stadium is gedurende de levenscyclus van de plant (Bell et al., 2006; Freckleton et al., 2006).
Deze overgang wordt beïnvloed door een verscheidenheid aan (a)biotische factoren. Het is
dan ook niet verwonderlijk dat de wetenschappelijke literatuur is opgebouwd uit een groot
aantal, elkaar soms tegensprekende, verklaringen voor het bestaan van de soortenrijkdom en
ruimtelijke patronen van tropische bossen (Swamy, 2008).
Processen die de negatieve intraspecifieke interacties versterken ten opzichte van de negatieve
interspecifieke interacties zijn essentieel voor de co-existentie van soorten. Conspecifieke
negatieve densiteit- of afstandsafhankelijke (conspecific negative density or distance
dependence of CNDD) mortaliteit bijvoorbeeld wijst op een negatieve correlatie tussen de
densiteit aan of de afstand tussen conspecifieke (van dezelfde soort) planten en hun
overlevingssucces en wordt beschouwd als één van de belangrijkste mechanismen achter de
latitudinale biodiversiteitsgradiënt (Johnson et al., 2012). Densiteits- en afstandsafhankelijke

Inleiding
- 3 -
mortaliteit wordt vaak verward met elkaar. Afstandsafhankelijke mortaliteit wijst op een
toenemende overlevingskans verder weg van conspecifieke individuen, en dit is veelal naar
plaatsen met een lage conspecifieke densiteit, vandaar de verwarring (Terborgh et al., 2012).
Intraspecifieke competitie, dit is competitie binnen de soort, en de gevoeligheid voor
gastheerspecifieke vijanden zijn twee mogelijke mechanismen achter CNDD mortaliteit (Lin
et al., 2012; Terborgh, 2012). Intraspecifieke competitie bijvoorbeeld is verantwoordelijk
voor een toename in zaailingmortaliteit bij hoge conspecifieke densiteit in de nabijheid van de
moederboom aangezien deze zaailingen maken gebruik van dezelfde limiterende
hulpbronnen.
Figuur 1.1: Een van de vele classificaties van de mogelijke mechanismen achter de co-existentie van planten (Barot, 2004).
Het ontrafelen van de diverse interacties tussen levende organismen is noodzakelijk indien
men de werking van een ecosysteem beter wil begrijpen. Een review van Schemske et al.
(2009) schenkt aandacht aan de veronderstelling dat de sterkte van biotische interacties
toeneemt in de richting van de evenaar, en linkt deze hypothese met de hogere biodiversiteit
in de tropen. Sterke biotische interacties bevorderen co-evolutie, een fenomeen waarbij
interagerende soorten continu op elkaar inspelen via fenotypische en genotypische
aanpassingen. Het resultaat is een snellere adaptatie en speciatie. Tropische regio’s worden
gekenmerkt door de afwezigheid van seizoenen, waardoor pathogenen gedurende het hele jaar

Inleiding
- 4 -
kunnen accumuleren en er sprake kan zijn van een sterkere co-evolutie tussen pathogeen en
gastheer, en dus een snellere adaptatie en speciatie in vergelijking met gematigde streken. In
gematigde streken worden abiotische factoren beschouwd als de primaire mechanismen achter
adaptatie en speciatie. Deze factoren op zich evolueren niet, maar variëren in tijd en ruimte.
1.2. Plant-Bodem terugkoppelingsmechanismen
Zoals hierboven reeds kort aangehaald, worden ecosystemen gedefinieerd als functionele en
dynamische eenheden die het resultaat zijn van interacties tussen abiotische en biotische
componenten (Loreau, 2010). Bij het onderzoeken van correlaties tussen de diversiteit en het
functioneren van een ecosysteem, wordt de focus veelal gelegd op interacties binnen één
trofisch niveau, bijvoorbeeld tussen planten onderling. Het feit dat deze uitmaken van een
complex netwerk van multitrofe interacties en dat planten fungeren als mediatoren tussen
boven- en ondergrondse systemen, wordt vaak over het hoofd gezien (Eisenhauer, 2012).
Terrestrische ecosystemen zijn opgebouwd uit boven- en ondergrondse subsystemen die
interageren met elkaar, en dit op verschillende temporele en ruimtelijke schalen (van der
Putten et al, 2009). Studies die de complexiteit aan boven- en ondergrondse feedback en
interacties met de abiotische omgeving onder de loep nemen, leiden tot een beter inzicht
betreffende het belang van biodiversiteit en de impact van menselijke activiteiten op de
leefomgeving (Wardle et al, 2004, van der Putten et al, 2009). Het ondergrondse subsysteem
omvat de bodem met al zijn chemische, fysische en biologische eigenschappen. Deze laatsten
spelen een belangrijke rol bij de groei, productiviteit en reproductief succes van
plantindividuen, alsook de samenstelling en productiviteit van de gehele plantengemeenschap
(van der Putten et al., 2013). De bodem is een complexe en heterogene omgeving in termen
van poriegrootte, water- en luchtregime en substraatbeschikbaarheid, waardoor zowel op
ruimtelijke als temporele schaal een grote hoeveelheid aan niches wordt gecreëerd (Giller,
1996). De bodem kan dus beschouwd worden als een habitatmozaïek met een enorme
verscheidenheid aan soorten. Planten kunnen deze bodemkarakteristieken op verschillende
manieren beïnvloeden, bv. door meer of minder input van organisch materiaal, en hierdoor
wijzigingen teweegbrengen in de diversiteit en activiteit van bodemorganismen (Bardgett et
al.,1998). Deze veranderingen kunnen er op hun beurt toe leiden dat de bodem meer of juist
minder in staat is om deze zelfde plantindividuen te onderhouden, alsook andere individuen
van dezelfde of een andere soort. Samengevat, indien de door planten veroorzaakte
veranderingen in bodemeigenschappen een invloed hebben op de aanwezigheid van de

Inleiding
- 5 -
planten zelf, spreekt men over plant-bodem terugkoppelingsmechanismen (Wardle, 2004).
Deze laatste kunnen direct of indirect zijn, alsook positief, negatief of neutraal (figuur 2 en 3)
(van der Putten et al., 2013). Hoe sterk en met welke specificiteit deze bodemeigenschappen
worden gewijzigd, zal bepalen in welke mate plant-bodem terugkoppelingsmechanismen een
impact hebben op de dynamiek van de gehele plantengemeenschap, alsook de
ecosysteemfuncties. Door het feit dat planten fungeren als mediatoren tussen boven- en
ondergrondse subsystemen, kunnen eventueel ook veranderingen plaatsvinden in de
interacties tussen planten en bovengrondse organismen (van der Putten et al., 2013).
Wortelherbivorie bijvoorbeeld zal een fysiologische respons van de plant uitlokken, meer in
het bijzonder een verhoogde metabolietenconcentratie (suikers, aminozuren) in de stengel,
waardoor nu ook bovengrondse herbivoren worden aangetrokken tot de plant. De
aanwezigheid van een ondergrondse herbivoor faciliteert op deze manier een bovengrondse
herbivoor, zonder dat deze twee rechtstreeks met elkaar interageren.
Figuur 1.2: Plant-bodem terugkoppelingsmechanismen. Links: directe en indirecte mechanismen. Directe, ook wel
intraspecifieke of conspecifieke, mechanismen wijzen op het effect van een plantenindividu op zichzelf of op andere
individuen van dezelfde soort. Indirecte, ook wel interspecifieke of heterospecifieke, mechanismen vinden plaats tussen
individuen van verschillende soort. Rechts: positieve (blauwe pijl) en negatieve (rode pijl) plant-bodem
terugkoppelingsmechanismen. Positieve mechanismen verhogen de kans dat conspecifieke individuen een habitat kunnen
domineren en eventueel monopoliseren. Negatieve mechanismen maken de bodem minder optimaal voor conspecifieke
individuen, waardoor de kans op dominantie sterk wordt verlaagd (van der Putten et al., 2013).
De invloed van bodembiota op de bovengrondse levende gemeenschappen is heel divers
(Wardle et al., 2004; van der Heijden et al., 2008; van der Putten et al., 2009). De in de
rhizosfeer aanwezige herbivoren voeden zich met de wortels van de plant en oefenen op deze
manier een direct negatief effect uit op de plant. Ook bodempathogenen vormen een
zogenaamde antagonistische associatie met hun gastheerplant. Daarnaast zijn er tal van
directe positieve relaties tussen planten en bodemorganismen. Denk maar aan de
mutualistische interacties tussen planten en mycorrhiza of stikstoffixerende bacteriën (Wardle
et al, 2004). Daarenboven zijn bodemorganismen in staat de chemische eigenschappen van de
bodem, en hieruit volgend de nutriëntenbeschikbaarheid en absorptiecapaciteit van

Inleiding
- 6 -
plantenwortels, te wijzigen via decompositieprocessen en oefenen ze een indirect negatief of
positief effect uit op de plant (Wardle et al, 2004). Het is duidelijk dat de structuur en
abundantie van de ondergrondse gemeenschap, waaronder micro-organismen als bacteriën,
virussen, protisten, fungi en nematoden, een grote invloed uitoefent op de ecologie van
bovengronds levende planten en dieren (van der Putten et al., 2009). Het karakteriseren van
deze ondergrondse gemeenschappen is dan ook belangrijk om de werking van het volledige
ecosysteem beter te begrijpen.
Figuur 1.3: Terugkoppelingsmechanismen tussen boven- en ondergrondse systemen (Wardle et al., 2004).
1.2.1. De Janzen-Connell hypothese
Een 40-tal jaar geleden gaven Janzen (1970) en Connell (1971) onafhankelijk van elkaar een
mogelijke verklaring voor de waargenomen paradox dat er in tropische bossen minder
zaailingen aanwezig zijn in de buurt van een reproductieve boom, ondanks de abundante
zaadval in de omgeving van de moederboom. De veronderstelling dat deze waarneming te
wijten is aan de accumulatie van gastheerspecifieke vijanden nabij de moederboom vormt de
basis van de zogenaamde Janzen-Connell (J-C) hypothese. Een moederboom trekt naast
pollinatoren en zaaddispersers, ook herbivoren en zaadpredatoren aan en fungeert als een
reservoir voor gastheerspecifieke pestsoorten en pathogenen, die doorgegeven worden aan
dichtbij gewortelde zaailingen (Gilbert, 2002; Terborgh et al., 2008). Het resultaat van deze
positieve (pollinatie, zaaddispersie) en negatieve (herbivorie, zaadpredatie en pathologie)

Inleiding
- 7 -
effecten bepaalt wanneer, waar en met welk succes een boom zal reproduceren (Terborgh et
al., 2008). De J-C hypothese wordt al sinds de jaren ’80 proefondervindelijk ondersteund
(Gilbert, 2002 en alle hierin vermelde referenties met positief resultaat betreffende de J-C
hypothese) en vormt nog steeds een belangrijk concept bij het begrijpen van de manier
waarop natuurlijke vijanden (zaadpredatoren, bodempathogenen) de gemeenschapsstructuur
en diversiteit van planten beïnvloeden. Het idee achter de J-C hypothese baseert zich op het
principe van competitieve exclusie en zet de biotische interacties tussen planten en hun
natuurlijke vijanden centraal. Competitie resulteert in competitieve exclusie indien superieur
competitieve soorten zodanig toenemen in abundantie waardoor andere, minder competitieve
soorten worden benadeeld. Deze interspecifieke competitie, zijnde de competitie tussen
soorten, kan leiden tot een afname in diversiteit (Gilbert, 2002). Doch dit kan vermeden
worden door middel van mechanismen, die competitieve exclusie verstoren, uitstellen of
voorkomen (Palmer, 1994). De J-C hypothese is een voorbeeld van zo een mechanisme. In
tegenstelling tot dieren, beschikken planten, eens gevestigd, niet over de mogelijkheid om
zich voort te bewegen doorheen het landschap. Planten moeten zodoende op zoek gaan naar
alternatieven om te ontsnappen aan ongunstige omgevingscondities en de overleving van hun
nakomelingen te garanderen. Zaaddormantie en zaaddispersie, respectievelijk de temporele en
ruimtelijke ontsnapping aan natuurlijke vijanden, intraspecifieke competitie, enz., worden
hierbij beschouwd als belangrijke mechanismen van een plant om zich aan te passen aan
variabele omstandigheden (Augspurger, 1983). Ondanks de evolutie van complexe
zaaddispersie mechanismen (Howe et al., 1982), waarbij gebruik wordt gemaakt van
abiotische (wind, water) of biotische (frugivore zaadverbreiders) vectoren, worden de meeste
zaden afgezet in de rechtstreekse omgeving van de moederplant. De hoge zaaddensiteit die
hierdoor ontstaat, verhoogt de interspecifieke competitie en dus stressgevoeligheid.
Zaadpredatoren en bodempathogenen accumuleren maar al te graag op deze plaatsen. Naast
deze densiteitsafhankelijke mortaliteit spreekt men ook over afstandsafhankelijke mortaliteit,
waarbij de kans op zaadkieming, zaailingrekrutering en -overleving stijgt met toenemende
afstand ten opzichte van de moederplant. De ruimtelijke schaal waarop zaadpredatoren en
bodempathogenen actief zijn, is namelijk afhankelijk van de afstand waarover deze zich
kunnen verplaatsen en aangezien de meesten gekenmerkt worden door minimale
verspreidingsmogelijkheden, is hun aanwezigheid vaak beperkt tot plaatsen nabij de
moederboom, waar hoge zaad- en zaailingdensiteit heerst. Zaden die verplaatst worden naar
een ‘veilige’ plek, weg van de moederboom, hebben dus een hogere kans op overleven.
Hierdoor zal er selectie optreden voor langere zaaddispersie afstanden (Nathan & Casagrandi,

Inleiding
- 8 -
2004). De reeds vermelde paradox geeft aanleiding tot de zogenaamde recruitment curve
(figuur 4) en toont aan dat het J-C patroon enkel zal plaatsvinden indien de gemiddelde
afstand waarover pathogenen actief zijn lager is dan de gemiddelde afstand waarover de zaden
gedisperseerd worden (Schupp, 1992; Nathan & Casagrandi, 2004). De optimale,
soortspecifieke afstand voor zaadkieming is dus afhankelijk van de afstand waarover de zaden
gedisperseerd kunnen worden en de activiteitsradius van pathogenen of zaadpredatoren (Hill
& Hill, 2001).
Figuur 1.4: Grafische voorstelling van het Janzen-Connell model (Matthesius, 2006).
1.2.2.1. Plant-pathogeen interacties
Reeds lange tijd staan pathogenen berucht als verwoesters van landbouwgewassen, maar ook
hun regulerende functie in de dynamiek van natuurlijke plantengemeenschappen mag niet
onderschat worden (van der Heijden et al, 2008). De manieren waarop pathogenen hun
gastheren infecteren zijn enorm divers. Plant-pathogeen interacties en de gevolgen voor de
plant in termen van fitness zijn dan ook zeer complex en uiteenlopend (Mordecai, 2011).
Planten zijn in staat zich te beschermen tegen pathogene infecties via tal van resistentie- of
defensiemechanismen, zoals de productie van toxische metabolieten (chemische defensie) of
de vorming van haren of stekels (mechanische defensie). Pathogenen kunnen deze
mechanismen ontwijken. Sommige fungi soorten bijvoorbeeld manipuleren de hormoonbalans
van hun gastheer zodanig dat er een ideale omgeving wordt gecreëerd voor de fungus zelf.
Deze zogenaamde co-evolutionaire arms race tussen plant en pathogeen is uitermate

Inleiding
- 9 -
gevarieerd, waarbij beiden reageren op veranderingen bij hun tegenstander (Maor & Shirasu,
2005; Ingle et al., 2006). Afhankelijk van de gastheerspecificiteit worden pathogenen
doorgegeven tussen plantenindividuen van eenzelfde (specialisten) of verschillende
(generalisten) soort, waarbij zowel bodem, atmosfeer, water als levende organismen kunnen
optreden als transmissievectoren. Gastheerspecifieke pathogenen vallen één of enkele soorten
aan binnen de plantengemeenschap. Dit kan resulteren in een gereduceerde dominantie van
competitief superieure planten of de eliminatie van zeldzame en competitief inferieure planten
(Klironomos, 2002; Frecleton et al., 2006). Gastheerspecifieke pathogenen die zaailingen van
slechts één of enkele boomsoorten infecteren, accumuleren in de directe omgeving van de
moederboom aangezien de meeste zaden afgezet worden op deze plaatsen. Dit brengt een
verhoogde zaailingmortaliteit van die specifieke boomsoort met zich mee. Hierdoor zien
zaailingen van andere boomsoorten hun kans om te groeien op deze vrijgekomen plaatsen,
resulterend in een ensemble van zaailingen van verschillende boomsoorten. Hoe sterker de
suppressie van zaadkieming en zaailingrekrutering rondom de moederboom, hoe groter het
aantal soorten dat kan co-existeren in een boomgemeenschap (Hubbell, 1980). De
gastheerspecificiteit van pathogenen kan dus aanzien worden als een vereiste voor het
verkrijgen van de hoge biodiversiteit in tropische regio’s (Klironomos, 2002; Freckleton et al.,
2006). De verscheidenheid aan transmissiemechanismen, alsook de verschillende invloeden
van pathogenen op planten, maakt het moeilijk voorspellingen te maken omtrent het effect
van pathogenen op natuurlijke plantengemeenschappen (Hatcher et al, 2006). De pathogenen
zelf zijn normaal niet gevoelig aan top-down controle, maar worden gereguleerd door
omgevingscondities en gastheerbeschikbaarheid. Kleine veranderingen in deze factoren
kunnen leiden tot sterke veranderingen in de dynamiek van pathogenen (Thompson, 2010).
Informatie betreffende de verscheidenheid aan pathogene fungi, verantwoordelijk voor
densiteitsafhankelijke mortaliteit bij zaailingen is schaars (Swamy, 2008). Welgekende genera
zijn Phytium, Phytophthora, Fusarium en Rhizoctonia, verantwoordelijk voor de welgekende
damping-off disease bij zaailingen van zowel landbouwgewassen als in het wild voorkomende
plantensoorten (Augspurger, 1983; Bell et al., 2006; Freckleton et al., 2006, Machon et al.,
2009). De symptomen van deze ziekte zijn het plots verwelken van de bladeren, het naar
beneden buigen en zachter worden van de stengel en bladval, resulterend in een vroegtijdige
zaailingmortaliteit (Augspurger, 1983).
Dit alles in acht genomen, beletten gastheerspecifieke pathogenen dat één of enkele, meestal
algemene soorten gaan domineren (Mangan et al., 2010). Hierdoor wordt competitieve
exclusie verstoord, vertraagd of zelfs verhinderd (Freckleton et al., 2006), resulterend in een

Inleiding
- 10 -
co-existentie van honderden soorten per hectare tropisch bos (Bell et al., 2006; Lewis 2010).
De J-C hypothese werd voornamelijk getoetst in Neotropische regio’s, zoals Panama en Costa
Rica, maar ook in Mexico, het Amazonewoud, Azië (Maleisië, Frans-Guinea) en Noord-
Australië, terwijl Afrikaanse bossen vaak buiten beschouwing werden gehouden (Matthesius,
2006). De Janzen-Connell patronen spelen echter niet alleen een belangrijke rol bij het
behoud van diversiteit in tropische regio’s, maar zijn ook van toepassing in gematigde streken
(Packer & Clay, 2000), zij het minder prominent (Hyatt et al., 2003).
1.3. Habitatfragmentatie
De impact van allerhande stressfactoren op de biodiversiteit en functionaliteit van
ecosystemen is een prangende onderzoeksvraag binnen het domein van de conservation
biology (Mantyka-Pringle et al., 2012). Menselijke populatiegroei en de hiermee gepaard
gaande landbouwontwikkelingen, urbanisatie en industrialisatie hebben wereldwijd geleid tot
grootschalige ontbossingen, met habitatverlies en -fragmentatie als gevolg.
Habitatfragmentatie wordt enerzijds gedefinieerd als een proces, waarbij een continu habitat
gereduceerd wordt tot kleinere patches, geïsoleerd van elkaar door een matrix van habitats die
verschillen van het originele en bijgevolg (meestal) niet bruikbaar zijn voor de oorspronkelijk
aanwezige soorten (figuur 1.5) (Fahrig, 2003). Anderzijds spreekt men ook over
habitatfragmentatie als een patroon (Fahrig, 2003). Er zijn vier effecten die de basis vormen
van deze meer kwantitatieve meting van habitatfragmentatie: een reductie in
habitathoeveelheid en patchgrootte, een toename in patchaantal en patchisolatie. Het relatief
belang van elk effect afzonderlijk is echter moeilijk te achterhalen omwille van hun
onderlinge correlaties (Fahrig, 2003). Afhankelijk van de betrokken soorten en de context
(Lehouck et al., 2009a), kan habitatfragmentatie leiden tot veranderingen in de
gemeenschapsstructuur en -functie, inclusief verlies van soorten en verstoringen binnen de
voedselketen (Kruess & Tscharntke, 1994). Verandering in landgebruik, en de hiermee
gepaard gaande habitatfragmentatie, wordt beschouwd als een van de grootste bedreigingen
voor het overleven van wilde populaties fauna en flora in tropische regio’s. Studies
hieromtrent komen dan ook steeds meer centraal te staan in ecologisch onderzoek van
tropische bossen (Hooper et al, 2005, Morris, 2010).

Inleiding
- 11 -
Figuur 1.5: Het proces van habitatfragmentatie.
Habitatfragmentatie is verantwoordelijk voor een verkleining en meer ruimtelijke isolatie van
populaties (Fahrig, 2003) en de effecten ervan zijn vaak gebaseerd op het metapopulatie
paradigma. Hoe kleiner en meer geïsoleerd de patches, hoe hoger de snelheid van extinctie en
hoe lager de snelheid van rekolonisatie (McCallum & Dobson, 2002). De transfer van genen
tussen populaties van eenzelfde soort wordt namelijk sterk gereduceerd, wat resulteert in
minder genetische diversiteit binnen de populatie dan tussen de populaties onderling. Een
overmaat aan homozygote individuen verhoogt bijgevolg de kans op inteelt en de expressie
van recessieve allelen, en zodoende recessieve ziektes. De populaties verzwakken en worden
kleiner, met eventueel een populatie-extinctie als gevolg. Vooral kleine populaties zijn
gevoelig voor inteelt en genetische drift (het toevallig verlies van genen) waardoor deze
makkelijker in de zogenaamde extinctiespiraal kunnen terechtkomen (Young et al., 1996;
Bender et al., 1998; Fahrig, 2003). Een grote wijziging in landschapsstructuur zoals
habitatfragmentatie, kan leiden tot een afname in genetische en soortensamenstelling van
plantengemeenschappen en is hierdoor verantwoordelijk voor de verstoring van
mutualistische relaties tussen tropische boomsoorten en hun zaadverbreiders (Cordeiro &
Howe, 2003). Habitatfragmentatie kan dus, afhankelijk van de beschouwde
dispersiemechanismen, directe of indirecte effecten hebben op de zaadverbreiding en
vestiging van plantenpopulaties. Bij planten die zelf instaan voor hun verbreiding of
afhankelijk zijn van abiotische factoren (wind, water), kan habitatfragmentatie
verantwoordelijk zijn voor een sterke verstoring van de abiotische dispersiemechanismen of
ervoor zorgen dat de zaden terecht komen in minder gunstige omgevingscondities, en
hierdoor minder goed kunnen kiemen. Habitatfragmentatie kan er ook toe leiden dat
zaadverbreiders (bv.: frugivore vogels) afnemen in abundantie of zelfs volledig van het toneel

Inleiding
- 12 -
verdwijnen. Hierdoor kunnen verschuivingen optreden in soortensamenstelling ten voordele
van taxa met beter aangepaste dispersiemechanismen. Naast de bezoekratio, kunnen ook de
mobiliteit of het habitatgebruik van de disperseerders sterk gewijzigd worden ten gevolge van
habitatfragmentatie. Dit alles kan resulteren in een sterke reductie van het aantal verbreide
zaden.
Aangezien ondergrondse micro-organismen (bodembacteriën en -fungi, nematoden) zowel
een directe als indirecte invloed hebben op bovengrondse gemeenschappen, kunnen
veranderingen in de soortensamenstelling van bodemorganismen leiden tot wijzigingen van
de plantengemeenschap (Cordeiro & Howe, 2003; Reynolds et al., 2003; van der Putten et al.,
2013). Mangan et al. (2004) hebben reeds aangetoond dat habitatfragmentatie kan leiden tot
een shift in de samenstelling van arbusculaire mycorrhiza fungi (AMF). Nu stelt zich de vraag
welk effect habitatfragmentatie heeft op de abundantie, soortensamenstelling,
gastheerspecificiteit en virulentie van pathogene bodemfungi en hierdoor ook op de distributie
en diversiteit van bovengrondse gemeenschappen. De meeste studies veronderstellen een
hogere pathogenenabundantie in gefragmenteerde habitats ten gevolge van randeffecten
(Vögeli et al., 2011). Tylianakis et al. (2008) bijvoorbeeld toonden aan dat gefragmenteerde
habitats gekenmerkt worden door een hogere parasieten/gastheer ratio, een toegenomen graad
van parasitisme en meer gespecialiseerde parasieten. Vögeli et al. (2011) vonden echter dat de
aanwezigheid van pathogenen niet wordt beïnvloed door de ruimtelijke structuur van de
metapopulatie of gastheerdensiteit. Verder onderzoek moet meer duidelijk scheppen omtrent
de complexiteit en contextspecificiteit van de respons van soorten en ecosystemen op diverse
stressfactoren zoals habitatfragmentatie.
1.4. Aanleiding tot deze masterproef
Deze masterproef bouwt verder op het doctoraatsonderzoek van Valérie Lehouck (2009),
waarbij onder meer geconcludeerd werd dat kleine en sterk verstoorde bosfragmenten worden
gekenmerkt door de afwezigheid van enkele belangrijke zaadverbreiders, voornamelijk sterk
bosgebonden vogelsoorten met een beperkte mobiliteit. Het gereduceerde aantal
zaadverbreiders leidde tot een afname in zaadkieming, zaailingrekrutering en -overleving van
X. monospora, een algemeen voorkomende boomsoort in de tropische bossen van Afrika.
Enerzijds is dit het gevolg van lagere kiemkansen indien vruchten niet door het
spijsverteringskanaal van frugivore vogels passeren (Lehouck et al., 2011). Anderzijds blijven
niet verbreide zaden onder de moederboom liggen, waar gereduceerde zaadkieming,

Inleiding
- 13 -
zaailingrekrutering en -overleving mogelijks een gevolg is van de accumulatie van
gastheerspecifieke bodempathogenen dicht bij de moederboom (Lehouck et al., 2009c). De
ruimtelijke distributie van plantpathogenen in de bodem, meer specifiek deze van de
potentieel pathogene fungi (o.a. Pythium, Phytophthora, Fusarium en Rhizoctonia) zal in deze
studie meer in detail onderzocht worden en in relatie gebracht worden met de ruimtelijke
distributie van zaailingen en adulten van Xymalos monospora. Habitatfragmentatie vormt een
sterke bedreiging voor de integriteit van tropische bosecosystemen, onder meer door hun
eventuele invloed op de interactie tussen pathogenen en hun gastheren. Dit kan op zijn beurt
gevolgen hebben voor de dynamiek van een gemeenschap of volledig ecosysteem. Er is nog
maar weinig gekend over de bodembiota, en meer specifiek de bodemfungi, van de tropische
bossen van Afrika. Deze masterproef probeert aan de hand van een algemene boomsoort van
Afrotropische bossen een duidelijker beeld te krijgen van de ruimtelijke distributie en
soortensamenstelling van deze intrigerende organismen en hun eventuele rol bij J-C effecten.

Doelstellingen
14
2. Doelstellingen
Het hoofddoel van deze masterproef is het verwerven van meer kennis op het gebied van de
ruimtelijke distributiepatronen en soortensamenstelling van potentieel pathogene bodemfungi
in de sterk verstoorde bosfragmenten van de Taita Hills, ZO Kenia.
Hieronder volgt een overzicht van de in acht genomen doelstellingen.
1. Nagaan van de ruimtelijke distributie van adulte X. monospora bomen en hun
zaailingen in verschillende bosfragmenten.
2. Nagaan van de ruimtelijke distributie van de microbiële bodemgemeenschap. Meer in
het bijzonder zullen de potentieel pathogene fungigenera Pythium, Phytophthora,
Fusarium en Rhizoctonia vergeleken worden tussen de bosfragmenten en langsheen
een afstandsgradiënt ten opzichte van een X. monospora moederboom.
3. Een verband leggen tussen: de ruimtelijke patronen van zaailingen en adulten van de
boomsoort X. monospora met de eventueel waargenomen ruimtelijke variatie in
microbiële bodemgemeenschap, m.a.w. een verband leggen tussen de eerste en tweede
doelstelling
4. De potentieel pathogene fungi, die een rol kunnen spelen bij Janzen-Connell effecten,
identificeren.

Materiaal & Methoden
15
3. Materiaal & Methoden
3.1. Studiegebied en studiesoort
3.1.1. Studiegebied
Het veldwerk van deze masterproef werd uitgevoerd in de verstoorde bosfragmenten van de
Taita Hills, gelegen in zuidoost Kenia (03°25’S, 38°20’E, alt 1200-2208 m; neerslag 600-
2300 mm jaar-1
) (Pellikka et al., 2009). De Taita Hills bevinden zich in de intertropische
convergentiezone (ICZ) en hebben hierdoor een kort (november-december) en een lang
(maart-mei) regenseizoen (Pellikka et al., 2009). Desondanks zijn mist en neerslag elementen
die het hele jaar door aanwezig zijn en voor stabiele, vochtige condities zorgen op de
bergtoppen -en kammen (Beentje, 1988). De Taita Hills behoren tot het Taita-Taveta district
en maken deel uit van de Eastern Arc Mountains, een bergketen die erkend wordt als een van
de 25 biodiversiteit hotspots wereldwijd (Pellikka et al., 2009). Deze laatsten staan gekend
omwille van het grote aantal soorten, al dan niet endemisch, die ze herbergen. Hotspots
representeren bijgevolg een groot deel van de wereldwijde biodiversiteit waardoor hun
behoud van onschatbare waarde is (Myers et al., 2000). De Taita Hills waren oorspronkelijk
volledig begroeid met nevelwoud. Vanaf ongeveer 200 jaar geleden begon ontbossing en de
hiermee gepaard gaande habitatfragmentatie en –degradatie, waardoor dit Keniaans deel van
de Eastern Arc Mountains tot een van de meest bedreigde biodiversiteit hotspots in de wereld
behoort (Lovett, 1996). De bouw van een spoorweg en het introduceren van exotische soorten
(o.a. Cipres, Eucalyptus, Pinus) hebben geleid tot habitatverlies op grote schaal (Pellikka et
al., 2009). Momenteel blijft minder dan 500 ha inheems woud over, dit is minder dan 2 % van
het oorspronkelijk vegetatiedek. Het huidige landschap is een mozaïek van drie grotere
bosfragmenten (Mbololo, Ngangao en
Chawia, met elk minstens 80 ha inheems
bos) en een aantal kleinere bosfragmenten
(waaronder Fururu, Ndiwenyi, Macha en
Mwachora, met elk minder dan 10 ha
inheems bos), omgeven door menselijke
nederzettingen en landbouwgebied (Pellikka
et al., 2009). De resterende fragmenten van
het oorspronkelijk Afromontaan bos
Figuur 3.1: Bosfragment Macha. © Valerie Lehouck

Materiaal & Methoden
16
beperken zich tot de bergtoppen en steile hellingen omdat deze minder bereikbaar zijn voor de
mens en bijgevolg moeilijker om te zetten zijn in landbouwgebied (Aerts et al., 2011). De
voornaamste gewassen die er gecultiveerd worden zijn maïs, bonen, erwten, cassave,
aardappelen, tomaten, bananen en mango. De bossen zelf worden vandaag de dag gebruikt
voor het sprokkelen van hout, vervaardigen van houtskool en als graasgrond. Deze net
genoemde factoren zijn verantwoordelijk voor habitatverlies op lokale schaal. Ondanks het
enorme verlies aan habitat worden de Taita Hills nog steeds gekenmerkt door de
aanwezigheid van endemische soorten (Beentje, 1988; Pellikka et al., 2009), maar de snelle
populatiegroei en veranderingen in landgebruik vormen een aanhoudende bedreiging voor de
kenmerkende biodiversiteit en talrijke ecosysteemdiensten die de Taita Hills te bieden
hebben.
3.1.2. Studiesoort
Xymalos monospora (Harv.) Baill. (Monimiaceae) is een algemene tropische boomsoort,
groeiend in submontane, vochtige bossen van Afrika. Naar aanleiding van de typische
limoengeur die vrijkomt bij het kneuzen van de bladeren, wordt hij ook wel ‘limoenhout’
(lemonwood) genoemd. Deze tweehuizige boom is aanwezig in alle bosfragmenten van de
Taita Hills (Lehouck et al., pers. com.) en kan tot 25 meter hoog worden (Lehouck et al.,
2009a,b,c). De kleine bloemetjes van X. monospora hebben een groenachtige kleur en zijn
geschikt volgens een pluim of tros (Lehouck et al., 2009a,b,c). De éénzadige, geelrode
vruchten zijn vlezig en worden verbreid door vogels. Passage doorheen het darmkanaal van
de vogel zorgt voor het verwijderen van het vruchtvlees en verhoogt op deze manier het
kiemingssucces van de zaden (Lehouck et al., 2009c). De vruchten en zaden hebben een
gemiddelde grootte van respectievelijk 11,8 mm (± 0,4 mm) x 9,4 mm (± 0,3 mm) en 9,6 mm
(± 0,6 mm) x 7,3 mm (± 0,4 mm) (Lehouck et al., 2009b). De periode van vruchtzetting vindt
plaats van april tot en met november, met een piek gedurende de maanden juni tot en met
augustus. Vanuit de praktijk is echter gewaar geworden dat het tijdstip waarop de
vruchtzetting plaatsvindt sterk kan variëren doorheen de jaren heen (Lehouck et al., 2009b).
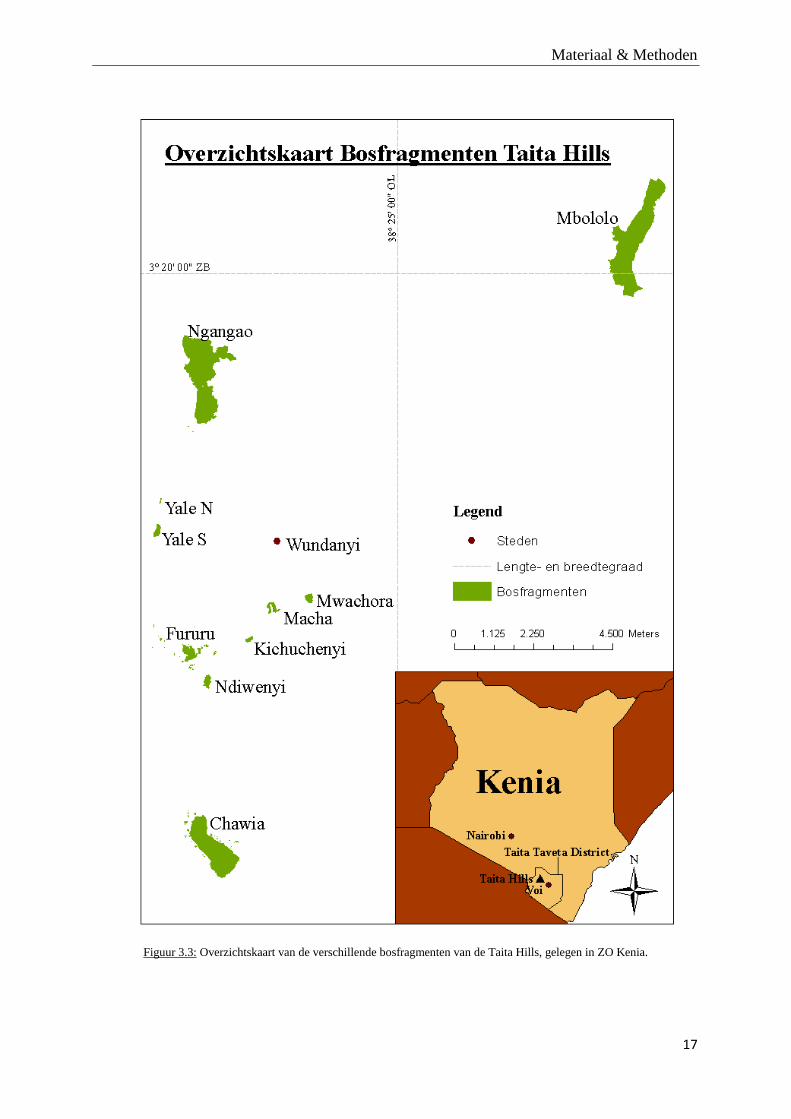
Figuur 3.2: Bladeren, vruchten en een zaailing van X. monospora. © Eduardo de la Peña en Valerie Lehouck
Materiaal & Methoden
17
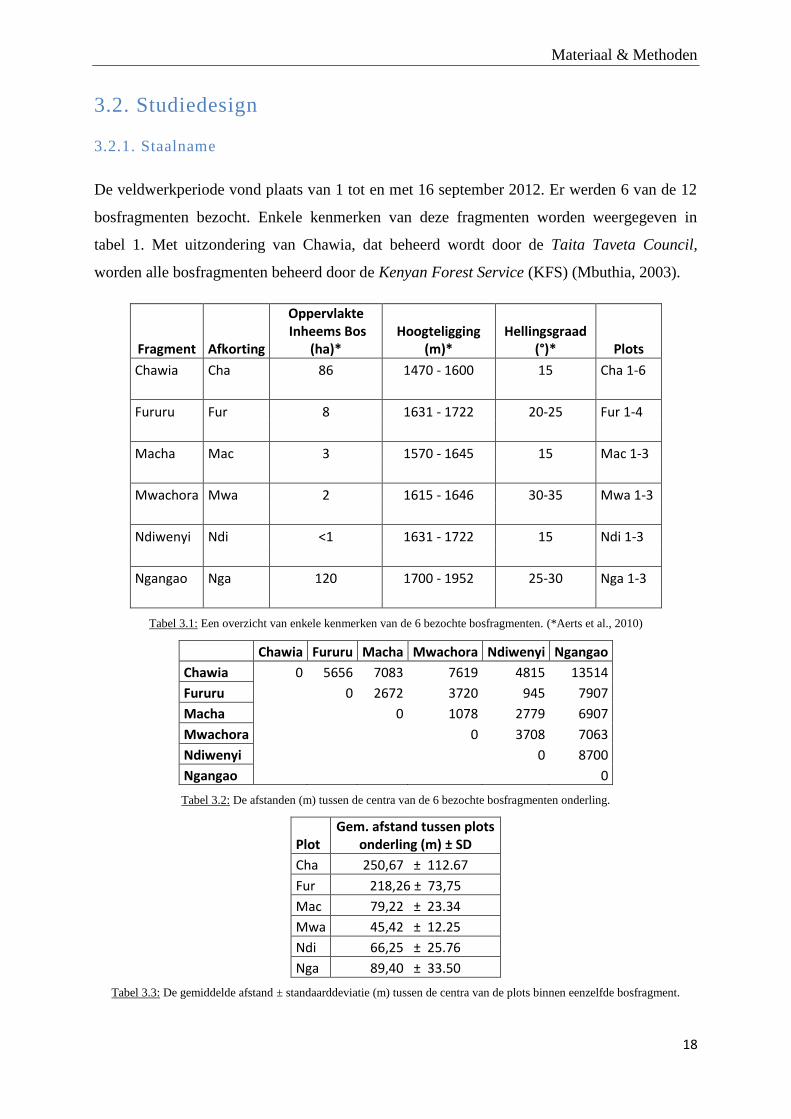
Figuur 3.3: Overzichtskaart van de verschillende bosfragmenten van de Taita Hills, gelegen in ZO Kenia.
Materiaal & Methoden
18
3.2. Studiedesign
3.2.1. Staalname
De veldwerkperiode vond plaats van 1 tot en met 16 september 2012. Er werden 6 van de 12
bosfragmenten bezocht. Enkele kenmerken van deze fragmenten worden weergegeven in
tabel 1. Met uitzondering van Chawia, dat beheerd wordt door de Taita Taveta Council,
worden alle bosfragmenten beheerd door de Kenyan Forest Service (KFS) (Mbuthia, 2003).
Fragment Afkorting
Oppervlakte Inheems Bos
(ha)* Hoogteligging
(m)* Hellingsgraad
(°)* Plots
Chawia Cha 86 1470 - 1600 15 Cha 1-6
Fururu Fur 8 1631 - 1722 20-25 Fur 1-4
Macha Mac 3 1570 - 1645 15 Mac 1-3
Mwachora Mwa 2 1615 - 1646 30-35 Mwa 1-3
Ndiwenyi Ndi <1 1631 - 1722 15 Ndi 1-3
Ngangao Nga 120 1700 - 1952 25-30 Nga 1-3
Tabel 3.1: Een overzicht van enkele kenmerken van de 6 bezochte bosfragmenten. (*Aerts et al., 2010)
Chawia Fururu Macha Mwachora Ndiwenyi Ngangao
Chawia 0 5656 7083 7619 4815 13514
Fururu
0 2672 3720 945 7907
Macha
0 1078 2779 6907
Mwachora
0 3708 7063
Ndiwenyi
0 8700
Ngangao 0
Tabel 3.2: De afstanden (m) tussen de centra van de 6 bezochte bosfragmenten onderling.
Plot Gem. afstand tussen plots
onderling (m) ± SD
Cha 250,67 ± 112.67
Fur 218,26 ± 73,75
Mac 79,22 ± 23.34
Mwa 45,42 ± 12.25
Ndi 66,25 ± 25.76
Nga 89,40 ± 33.50
Tabel 3.3: De gemiddelde afstand ± standaarddeviatie (m) tussen de centra van de plots binnen eenzelfde bosfragment.

Materiaal & Methoden
19
In alle bosfragmenten werden drie plots van 15 x 15 m uitgezet, met uitzondering van Fururu
(4 plots) en Chawia (6 plots), en binnen elke plot werden drie subplots van 3 x 3 m uitgezet.
Een schematische voorstelling van deze studiedesign kan teruggevonden worden in figuur 3.5.
Met behulp van een Global Positioning System toestel (GPS GARMIN 60CSX, gemiddelde
nauwkeurigheid 7.6 m) werden per plot 25 (indien mogelijk) Xymalos monospora zaailingen
en alle aanwezige adulte Xymalos monospora gekarteerd (zie verder).
In elke plot werden volgende parameters bepaald:
- De hellingsgraad en oriëntatie, gebruik makend van respectievelijk een clinometer en
kompas. Dit werd telkens bepaald onderaan de helling van de plot.
- Het aantal adulte bomen, zowel X. monospora als andere boomsoorten.
- De diameter van alle aanwezige adulte bomen, zowel van adulte monospora als andere
boomsoorten. Dit werd gemeten op borsthoogte (± 130 cm) en men spreekt dan ook
over de Diameter at Breast Height (DBH).
- De lengte van de X. monospora zaailingen, gemeten vanaf de hypocotyl tot en met de
groeitop.
- De inkomende lichtintensiteit. Hiervoor werden per plot drie foto’s (Nikon D3000)
genomen van de kruinlaag, en dit op eenzelfde ooghoogte, op drie random locaties
binnen de plot. Met behulp van het programma ImageJ (versie 1.32) werd de
verhouding witte ten opzichte van zwarte pixels berekend als proxy voor de graad van
kroonbedekking (%). Per plot het gemiddelde van de drie waarden berekend.
In de plots Fur1 en Ndi1 werd noch de diameter van andere adulten buiten X. monospora noch
de zaailinglengte opgemeten. Fur1 was onze eerste en dus ‘experimentele’ plot waarin nog
werd uitgezocht welke parameters er allemaal bepaald gingen worden. In Ndi1 werden geen
X. monospora zaailingen teruggevonden en werd vergeten de diameter van de adulte bomen
verschillend van X. monospora op te meten. Van de 25 zaailingen werd telkens een deel van
de wortel bewaard op 100% ethanol. Het resterende wortelmateriaal werd gedurende de nacht
gedroogd in een keukenoven op 50 °C en vervolgens convoluut bewaard in papieren zakjes
om rotting tegen te gaan. Bij een random subsample van 8 zaailingen per plot werden
bodemstalen van 10 ml genomen, en dit telkens in de bovenste 10 cm van de rhizosfeer. In
Ndi1 werden geen bodemstalen genomen omwille van de afwezigheid van zaailingen. De
bodemstalen van Cha6 liggen nog in Kenia. In elke subplot werd het aantal zaailingen en
saplingen van elke boomsoort bepaald. Een onderscheid tussen zaailing en sapling werd

Materiaal & Methoden
20
gemaakt op basis van lengte en stevigheid van de stengel. Een plantje met een lengte groter
dan 0.5 m en een min of meer stevig ontwikkelde stengel werd beschouwd als een sapling.
Aangezien op plotniveau niet alle aanwezige zaailingen geteld zijn, werd het gemiddelde van
het aantal X. monospora zaailingen, geteld in de drie verschillende subplots, gebruikt om de
densiteit aan zaailingen te bepalen per m2. Figuur 3.4 en 3.5 geven een schematische
voorstelling van de staalname design.
Figuur 3.4: Een overzicht van de gemeten variabelen op plot- en subplotniveau.
Figuur 3.5: Schematische voorstelling van de studiedesign. De figuur stelt 1 bosfragment voor. De grote vierkanten (15 x 15
m) stellen de plots voor, de kleinere vierkanten (3 x 3 m) de subplots.
15m
m
15m
15
m
3m
m
15m
3
m
m
15m
Hellingsgraad (°)
Oriëntatie
Graad van kroonbedekking (%)
Diameter adulte bomen (cm)
Lengte X. monospora zaailingen (cm)
Aantal zaailingen van elke boomsoort
Aantal saplingen van elke boomsoort
Plotniveau (15x15m)
Subplotniveau (3x3m)

Materiaal & Methoden
21
De verkregen GPS-coördinaten werden verwerkt met behulp van het geografisch
computerprogramma ArcGIS (Arcmap, versie 3.1). De ruimtelijke positie van de gekarteerde
X. monospora zaailingen en adulte bomen werden weergegeven per plot, binnen elk
bosfragment. Vervolgens werd van elke zaailing de Euclidische afstand tot de dichtstbijzijnde
adulte X. monospora bepaald met behulp van de Nearest neighbour analyse (Spatial Statistics
Tools). Aan de hand van dezelfde analyse werd het ruimtelijk distributiepatroon van X.
monospora adulten per plot nagegaan. Voor elke adulte X. monospora boom werd namelijk de
afstand bepaald tot zijn dichtstbijzijnde adulte X. monospora buur. Hiervan werd het
gemiddelde genomen en vergeleken met de verwachte afstand bij toevallige spreiding. De
verkregen Nearest neighbour index (R) is kleiner, gelijk of groter dan 1 en wijst
respectievelijk op een geclusterd, random of verspreid distributiepatroon. Tenslotte werd ook
de afstand tussen de zes bezochte fragmenten onderling en de verschillende plots binnen
eenzelfde bosfragment bepaald aan de hand van de nearest neighbour methode, waarbij
telkens gemeten werd vanuit het centrum van het fragment of de plot. De finale kaartjes
werden afgewerkt met behulp van ArcGIS (Arcview, versie 3.1).
3.2.2. Karakterisering bodemfungi
Uitplaten van de bodemstalen
De microbiële bodemgemeenschap langsheen een afstandsgradiënt van de moederboom werd
bepaald aan de hand van de klassieke cultuurafhankelijke methode. Alle bodemstalen werden
gedurende de periode vóór het uitplaten bewaard bij 8°C. Na het uitplaten werden deze
geïncubeerd bij kamertemperatuur gedurende twee tot drie dagen om vervolgens het aantal
gevormde kolonies te bepalen. Van de verzamelde bodemstalen werd telkens 0,5 g opgelost in
5 ml gedestilleerd en geautoclaveerd water. Vervolgens werd van deze 10% oplossing een
halve ml overgebracht in 4.5 ml water, resulterend in een 1:10 verdunning. In een volgende
stap werd 500 μl van deze verdunning uitgeplaat op een selectief medium door middel van de
strijkplaat methode. Rekening houdend met de bestaande literatuur omtrent potentieel
pathogene fungi van tropische boomsoorten (Augspurger, 2007), werden gekozen om te
werken met drie verschillende selectieve media: een selectief agarmedium voor de genera
Pythium en Phytophthora (Jeggers & Martin, 1986), een selectief agarmedium voor het genus
Fusarium (Komada, 1976) en een selectief agarmedium voor het genus Rhizoctonia (Ho &
hora, 1971). Per petri-plaatje en morfotype werd het aantal kolonies geteld (colony forming
units of CFU) en vermenigvuldig met 10 om het aantal kolonies van in 0.5 g bodem te

Materiaal & Methoden
22
kennen. Vervolgens werd van elk morfotype een kolonie overgeënt op potato dextrose agar
(PDA) medium, dit om een zuivere isolaat te verkrijgen. Er werden foto’s (Nikon D90)
genomen van de habitus van de verschillende morfotypes (bijlage 3).
Moleculaire Analyse van de isolaten
De moleculaire analyse van de isolaten van de 20 verschillende morfotypes vond plaats in het
Instituut voor Landbouw en Visserijonderzoek te Merelbeke (ILVO, onderzoeksgroep Plant).
Allereerst werd het DNA geëxtraheerd. Een kleine hoeveelheid afgeschraapt mycelium,
samen met 40 μl 0.25 M NaOH werd overgebracht in een 0.5 ml tube, die gedurende 30 s
werd ondergedompeld in kokend water. Vervolgens werden volgende reagentia toegevoegd:
40 µl 0.25 M HCl, 20 µl 0.5 M Tris-HCl (pH8), 20 µl 0.25 % Nonidet. De tube werd opnieuw
ondergebracht in kokend water, deze keer gedurende 2 minuten. Om te controleren of er wel
degelijk DNA aanwezig was in de tube, werd er 1 μl genanodropped met behulp van een
nanodrop spectrophotometer (ND-1000). Intussen werd het DNA bewaard op ijs, zodat het
intact bleef. Voor de uiteindelijke polymerase chain reaction (PCR) werd een mastermix
aangemaakt waarbij per tube van een 12-tube strip 5 μl magnesiumchloride buffer, 1 μl dNTP,
1 μl primer ITS1, 1 μl primer ITS2, 36.6 μl gedestilleerd water, 0.4 μl Taq-polymerase en 5 μl
van het geëxtraheerd DNA werd overgebracht. PCR is een veelvuldig gebruikte techniek om
kleine hoeveelheden van een bepaalde DNA-sequentie, in ons geval de internal transcribed
spacer (ITS) regio van ongeveer 600 basenparen (bp) en tevens de meest algemeen
gesequeneerde fungi DNA-sequentie (Schoch et al., 2012), te amplificeren via kunstmatige
DNA-replicatie. De reactie bestaat uit drie verschillende stappen. De eerste stap bestaat uit
een verhoging van de temperatuur tot ongeveer 90-95 °C, waardoor de waterstofbruggen
tussen de DNA-strengen worden verbroken. In de tweede stap worden de primers, die
complementair zijn aan het eerste stukje van het te kopiëren DNA, toegevoegd. De
temperatuur wordt verlaagd, zodat de vorming van nieuwe waterstofbruggen opnieuw
mogelijk wordt. In de derde stap maakt het enzym DNA-polymerase een nieuwe DNA-streng,
startend vanaf de primers en complementair aan de gesplitste strengen. Het resultaat is een
verdubbeling van de gewenste nucleotidesequentie. Deze drie verschillende stappen werden
30 keer herhaald, zodat de ITS-regio in voldoende hoge hoeveelheid aanwezig was om te
sequeneren. Het DNA-bandenpatroon werd verkregen via een Qiaxcel apparaat (Qiagen).
Deze capillaire gel-elektroforese is enorm gebruiksvriendelijk en dient ter vervanging van de
traditionele, arbeidsintensieve en tijdrovende gel-elektroforese. De, op basis van lengte, snelle
scheiding van DNA fragmenten gebeurt via een elektrische stroming, aangebracht op een met

Materiaal & Methoden
23
gel gevulde capillair. Een detector registreert de nucleïnezuren gedurende hun migratie naar
het positief geladen einde van de capillair. De resultaten worden vervolgens omgezet naar een
zogenaamd elektropherogram met behulp van specifieke software (BioCalculator).
Identificatie van de isolaten
De amplicons werden, samen met de twee primers, opgestuurd naar een gespecialiseerd
sequeneringslaboratorium in Amsterdam (Macrogen). Ter identificatie van de isolaten,
werden de bekomen DNA sequenties vergeleken met reeds aanwezige DNA sequenties in de
online beschikbare databank Basic Local Alignment Search Tool, kortweg BLAST
(http://blast.ncbi.nlm.nih.gov/). Het genus, of eventueel de soort, waarvan de ITS DNA-
sequentie, algemeen aanwezig in het ribosomaal DNA (rDNA) van fungi, het meeste
overeenkomst vertoonde met die van het geïsoleerd morfotype, werd geselecteerd. Hiervoor
werd gekeken naar zowel de mate van query cover (%) als identity (%).
3.3. Statistische dataverwerking
3.3.1. Ruimtelijke distributie van X. monospora
Gebruik makend van het statistisch programma R (versie 2.15) brachten univariate analyses
eventuele verschillen in gemeten parameters tussen de fragmenten aan het licht. De gebruikte
parameters waren: de densiteit van adulte bomen (zowel X. monospora als andere
boomsoorten), de relatieve densiteit, relatieve dominantie en diameter van X. monospora
adulten, de densiteit en lengte van X. monospora zaailingen en de graad van kroonbedekking.
De densiteit van adulte bomen (uitgedrukt per hectare) werd bepaald op basis van het aantal
adulten op plotniveau, dat vervolgens werd omgezet naar aantal adulten per hectare. De
relatieve densiteit van X. monospora adulten werd berekend als de verhouding van de
densiteit van adulte X. monospora bomen ten opzichte van de densiteit van alle adulte bomen.
De relatieve dominantie wijst op de ratio van de som van de stamoppervlaktes (berekend op
basis van de gemeten DBH) van alle adulte X. monospora bomen ten opzichte van de som van
de stamoppervlaktes van alle aanwezige adulte bomen. De densiteit aan X. monospora
zaailingen (uitgedrukt per m2)werd berekend op basis van het gemiddeld aantal zaailingen
over de drie subplots, dat vervolgens werd omgezet naar aantal zaailingen per m2. Een visuele
verkenning van de dataset werd uitgevoerd via staafdiagrammen (gemiddelde ±
standaarddeviatie) en boxplots (mediaan, 25-75 percentiel). Een one-way ANOVA

Materiaal & Methoden
24
(parametrisch) of Kruskal-Wallis test (niet-parametrisch) gaven aan of we al dan niet te
maken hadden met significante verschillen tussen de fragmenten onderling.
3.2.2. Ruimtelijke distributie van de bodemfungi gemeenschap
3.2.2.1. Soortensamenstelling en diversiteit
De γ-diversiteit, in ons geval de diversiteit aan fungi morfotypes binnen elk fragment, werd
geschat op basis van drie verschillende indices: het Michaelis-Menten gemiddelde
(MMMeans), Shannon Exponentieel gemiddelde en Simpson gemiddelde. Het Michaelis-
Menten gemiddelde is de index met de hoogste gevoeligheid ten opzichte van eerder
zeldzame fungi morfotypes (Colwell, 2009). Zowel het Shannon Exponentieel als het
Simpson gemiddelde geven meer gewicht aan veel voorkomende fungi morfotypes en zijn
hierdoor een maat voor het effectief aantal soorten binnen elk fragment (Jost, 2006). De
indices werden berekend via het programma EstimateS (versie 9). De Jaccard similarity index
werd gebruikt als maat voor de β-diversiteit en geeft de gelijkenis in fungi morfotype
soortenrijkdom weer tussen de fragmenten onderling. Deze index werd berekend op basis van
onderstaande formule.
Met
a = het aantal geïdentificeerde fungi morfotypes in fragment A
b = het aantal geïdentificeerde fungi morfotypes in fragment B
c = het aantal gemeenschappelijke fungi morfotypes in beide fragmenten.
3.2.2.2. Gemeenschapsanalyse
De fungi morfotypesamenstelling tussen de 6 fragmenten, de 22 plots en de verschillende
afstandsklassen (afstand zaailing ten opzichte van de dichtstbijzijnde moederboom) werd
vergeleken met behulp van multivariate analyses. Er werd gebruik gemaakt van het statistisch
programma Primer (versie 6) waarbij de abundantie van de geïsoleerde morfospecies
(kolommen) werd weergegeven per bodemstaal (rijen). Aangezien we te maken hebben met
een typisch ecologische dataset, d.w.z. met veel nul-waarden, werd deze getransformeerd
(square root transformation). Vervolgens werd een non-metric multi dimensional scaling
(NMDS) en analysis of similarity (ANOSIM) analyse uitgevoerd, gebaseerd op de Bray-
Curtis afstandsmatrix. Een NMDS-plot geeft de relatie weer tussen de objecten
(bodemstalen), en dit in een multidimensionale ruimte. Een ANOSIM analyse is een

Materiaal & Methoden
25
algemeen gebruikte, niet-parametrische methode van multivariate analyse die, in ons geval, de
variatie in fungi morfotype abundantie en samenstelling vergelijkt tussen de fragmenten
(Anderson, 2001). De NMDS werd meerdere keren uitgevoerd (met 100 iteraties) en dit om
de NMDS-plot met de laagst mogelijke stress te verkrijgen. De mate van stress geeft namelijk
weer hoe betrouwbaar de ordinatie-plot is (Sturrock & Rocha, 2001). Een stress-waarde hoger
dan 0.3 toont aan de dat de verkregen ordinatie met een zekere omzichtigheid moet
geïnterpreteerd worden omdat de kans groot is dat deze niet de werkelijkheid representeert. In
R (versie 2.15) werden Principal Component analyses (PCA) uitgevoerd om na te gaan in
welke mate de opgemeten variabelen (de densiteit van adulte X. monospora bomen en andere
boomsoorten, de relatieve densiteit aan X. monospora adulten, de densiteit van X. monospora
zaailingen, de graad van kroonbedekking) de eventueel waargenomen variatie tussen de
fragmenten, plots en afstandsklassen kunnen verklaren. De hoeveelheid en significantie van
verklaarde variatie wordt weergegeven door middel van r2 en bijhorende p-waarde.

Resultaten
26
4. Resultaten
4.1. De ruimtelijke distributie van X. monospora
De adulte X. monospora bomen vertonen een verspreid distributiepatroon. De bijhorende tabel
met R- en p-waarden op plotniveau en de kaartjes van de ruimtelijke distributie van adulte X.
monospora bomen en hun zaailingen kunnen teruggevonden worden in bijlage 1 en 2. Er zijn
slechts 25 zaailingen per plot gekarteerd omwille van tijdsgebrek. Hierdoor is het niet zinvol
de ruimtelijke distributiepatronen van de X. monospora zaailingen op plotniveau na te gaan.
Tabel 4.1 geeft een overzicht van de gemeten variabelen op fragmentniveau. Indien de data
normaal verdeeld zijn, wordt het gemiddelde ± standaarddeviatie weergegeven, indien niet
wordt de mediaan en het 25-75 percentiel getoond. Er werden significante verschillen
gevonden tussen de fragmenten in graad van kroonbedekking (%), DBH van X. monospora
adulten (cm) en lengte van X. monospora zaailingen (cm). De densiteit van alle adulte bomen,
X. monospora adulten en zaailingen, de relatieve densiteit en de relatieve dominantie van X.
monospora adulten verschillen niet significant tussen de fragmenten.
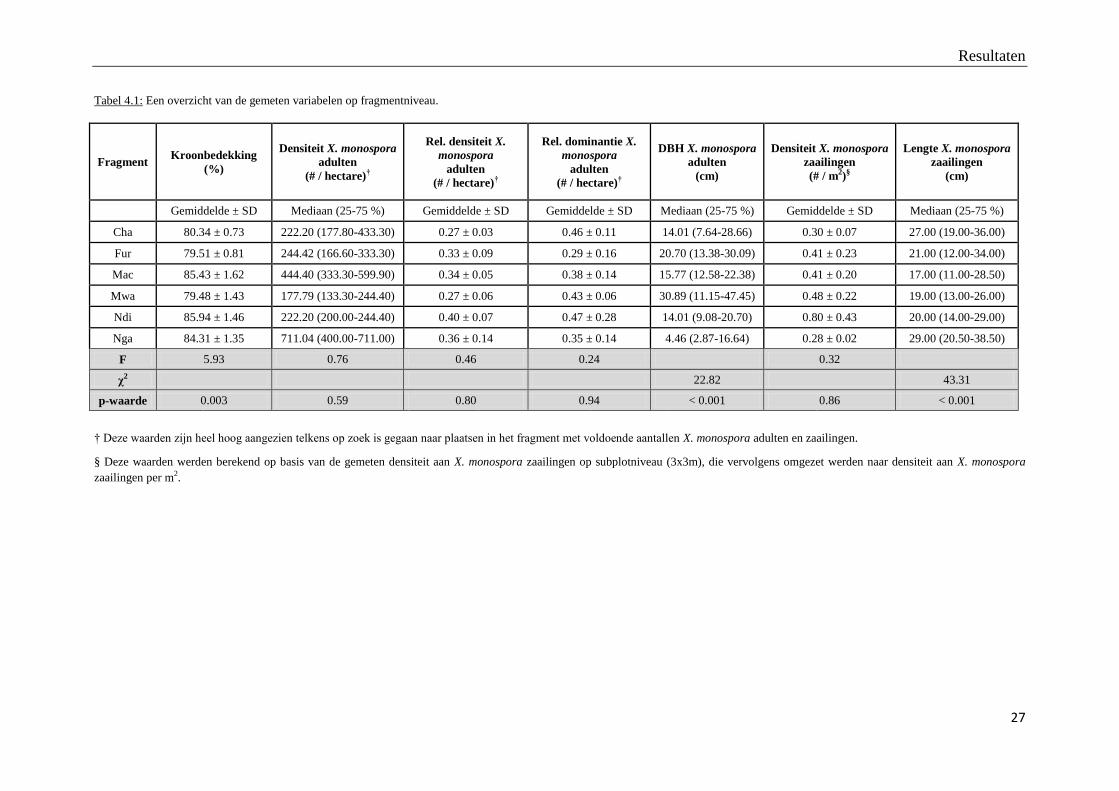
De afstand tussen X. monospora zaailingen en hun dichtstbijzijnde moederbomen verschilt
niet significant tussen de fragmenten (figuur 4.1).
Figuur 4.1: Vergelijking van de afstand tussen zaailing en dichtstbijzijnde moederboom tussen de fragmenten.
0
2
4
6
8
10
12
14
Cha Fur Mac Mwa Ndi Nga
Ge
mid
de
lde
afs
tan
d
zaai
ling-
mo
ed
erb
oo
m (
m)
Fragment
F = 0.68, Df = 5, p = 0.64
Resultaten
27
Tabel 4.1: Een overzicht van de gemeten variabelen op fragmentniveau.
Fragment Kroonbedekking
(%)
Densiteit X. monospora
adulten
(# / hectare)†
Rel. densiteit X.
monospora
adulten
(# / hectare)†
Rel. dominantie X.
monospora
adulten
(# / hectare)†
DBH X. monospora
adulten
(cm)
Densiteit X. monospora
zaailingen
(# / m2)§
Lengte X. monospora
zaailingen
(cm)
Gemiddelde ± SD Mediaan (25-75 %) Gemiddelde ± SD Gemiddelde ± SD Mediaan (25-75 %) Gemiddelde ± SD Mediaan (25-75 %)
Cha 80.34 ± 0.73 222.20 (177.80-433.30) 0.27 ± 0.03 0.46 ± 0.11 14.01 (7.64-28.66) 0.30 ± 0.07 27.00 (19.00-36.00)
Fur 79.51 ± 0.81 244.42 (166.60-333.30) 0.33 ± 0.09 0.29 ± 0.16 20.70 (13.38-30.09) 0.41 ± 0.23 21.00 (12.00-34.00)
Mac 85.43 ± 1.62 444.40 (333.30-599.90) 0.34 ± 0.05 0.38 ± 0.14 15.77 (12.58-22.38) 0.41 ± 0.20 17.00 (11.00-28.50)
Mwa 79.48 ± 1.43 177.79 (133.30-244.40) 0.27 ± 0.06 0.43 ± 0.06 30.89 (11.15-47.45) 0.48 ± 0.22 19.00 (13.00-26.00)
Ndi 85.94 ± 1.46 222.20 (200.00-244.40) 0.40 ± 0.07 0.47 ± 0.28 14.01 (9.08-20.70) 0.80 ± 0.43 20.00 (14.00-29.00)
Nga 84.31 ± 1.35 711.04 (400.00-711.00) 0.36 ± 0.14 0.35 ± 0.14 4.46 (2.87-16.64) 0.28 ± 0.02 29.00 (20.50-38.50)
F 5.93 0.76 0.46 0.24 0.32
χ2 22.82 43.31
p-waarde 0.003 0.59 0.80 0.94 < 0.001 0.86 < 0.001
† Deze waarden zijn heel hoog aangezien telkens op zoek is gegaan naar plaatsen in het fragment met voldoende aantallen X. monospora adulten en zaailingen.
§ Deze waarden werden berekend op basis van de gemeten densiteit aan X. monospora zaailingen op subplotniveau (3x3m), die vervolgens omgezet werden naar densiteit aan X. monospora
zaailingen per m2.

Resultaten
28
4.2. De ruimtelijke distributie van de bodemfungi gemeenschap
4.2.1. Soortensamenstelling en diversiteit
In totaal zijn er 20 verschillende fungi morfotypes geïsoleerd, gesequeneerd en
geïdentificeerd, waarvan één morfotype, geïdentificeerd als Galactomyces spp. (morfotype 10
in tabel 4.3), is verwijderd uit de dataset. Dit morfotype, slechts aanwezig in één staal (vierde
plot in bosfragment Chawia), is geen bodemfungus en de vondst is waarschijnlijk het gevolg
van contaminatie. De resultaten van de BLAST analyse worden weergegeven in tabel 4.2.
Foto’s van de habitus van de finaal 19 fungi morfotypes zijn terug te vinden in bijlage 3.
Morfotype Best genbank match Genbank accession Query score (%) Identity (%)
Morfotype 1 Mortierella spp.1 JF439489.1 85 98
Morfotype2 Mortierella spp.2 AY157495.1 93 89
Morfotype 3 Mortierella spp.3 HQ607844.1 97 99
Morfotype 4 Mortierella spp.4 GQ302682.1 95 98
Morfotype 5 Mortierella spp.5 HQ608097.1 99 98
Morfotype 6 Mucor fragilis FN650655.1 97 98
Morfotype 7 Trichoderma harzianum HM176572.1 98 99
Morfotype 8 Penicillium vancouverense JN617675.1 95 95
Morfotype 9 Trichoderma spirale JF439515.1 96 99
Morfotype 10 Galactomyces spp. JN974290.1 94 93
Morfotype 11 Ilyonectria radicicola FJ481036.1 95 99
Morfotype 12 Umbelopsis isabellina JN943789.1 91 97
Morfotype 13 Trichosporon porosum HF558656.1 95 99
Morfotype 14 Fusarium oxysporum 1 JX915255.1 98 99
Morfotype 15 Cylindrocarpon spp. AB725901.1 89 96
Morfotype 16 Fusarium oxysporum 2 GU934524.1 93 99
Morfotype 17 Fusarium spp. HQ130669.1 97 98
Morfotype 18 Fusarium lateritium FJ037744.1 98 99
Morfotype 19 Fusarium oxysporum 3 HM346538.1 97 99
Morfotype 20 Fusarium culmorum GU566271.1 98 99
Tabel 4.2: Resultaten van de BLAST analyse.
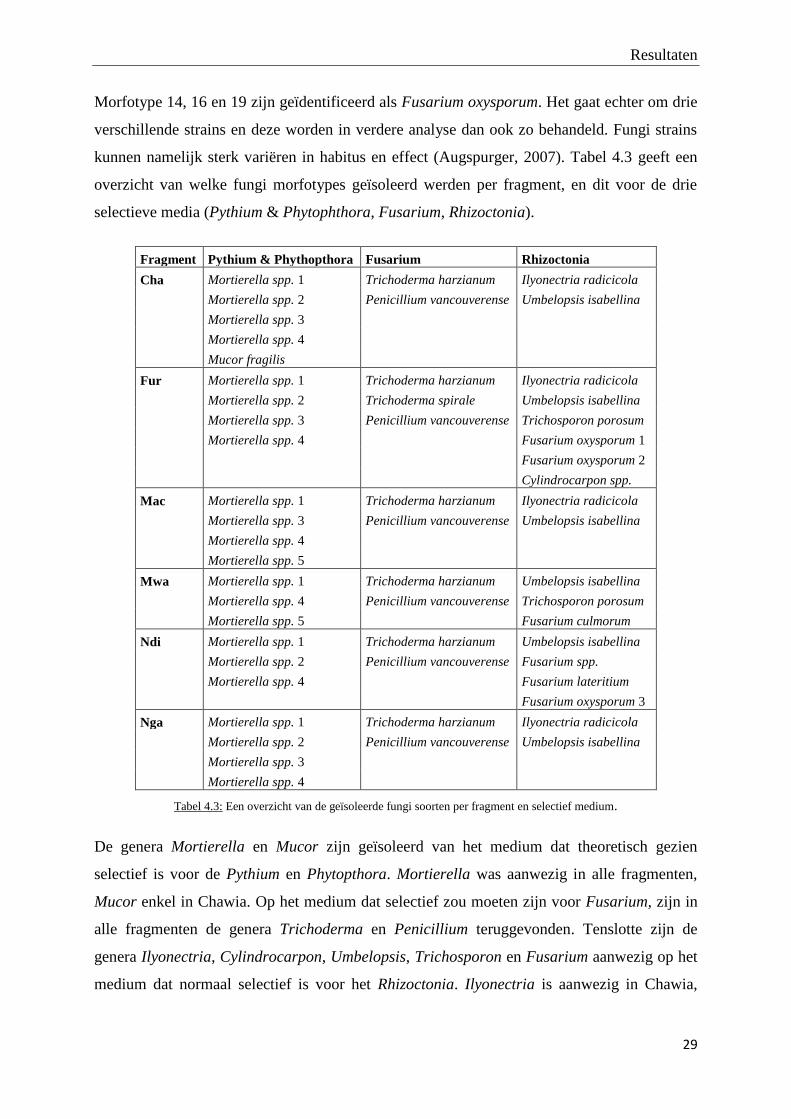
Resultaten
29
Morfotype 14, 16 en 19 zijn geïdentificeerd als Fusarium oxysporum. Het gaat echter om drie
verschillende strains en deze worden in verdere analyse dan ook zo behandeld. Fungi strains
kunnen namelijk sterk variëren in habitus en effect (Augspurger, 2007). Tabel 4.3 geeft een
overzicht van welke fungi morfotypes geïsoleerd werden per fragment, en dit voor de drie
selectieve media (Pythium & Phytophthora, Fusarium, Rhizoctonia).
Fragment Pythium & Phythopthora Fusarium Rhizoctonia
Cha Mortierella spp. 1 Trichoderma harzianum Ilyonectria radicicola
Mortierella spp. 2 Penicillium vancouverense Umbelopsis isabellina
Mortierella spp. 3
Mortierella spp. 4
Mucor fragilis
Fur Mortierella spp. 1 Trichoderma harzianum Ilyonectria radicicola
Mortierella spp. 2 Trichoderma spirale Umbelopsis isabellina
Mortierella spp. 3 Penicillium vancouverense Trichosporon porosum
Mortierella spp. 4 Fusarium oxysporum 1
Fusarium oxysporum 2
Cylindrocarpon spp.
Mac Mortierella spp. 1 Trichoderma harzianum Ilyonectria radicicola
Mortierella spp. 3 Penicillium vancouverense Umbelopsis isabellina
Mortierella spp. 4
Mortierella spp. 5
Mwa Mortierella spp. 1 Trichoderma harzianum Umbelopsis isabellina
Mortierella spp. 4 Penicillium vancouverense Trichosporon porosum
Mortierella spp. 5
Fusarium culmorum
Ndi Mortierella spp. 1 Trichoderma harzianum Umbelopsis isabellina
Mortierella spp. 2 Penicillium vancouverense Fusarium spp.
Mortierella spp. 4 Fusarium lateritium
Fusarium oxysporum 3
Nga Mortierella spp. 1 Trichoderma harzianum Ilyonectria radicicola
Mortierella spp. 2 Penicillium vancouverense Umbelopsis isabellina
Mortierella spp. 3
Mortierella spp. 4
Tabel 4.3: Een overzicht van de geïsoleerde fungi soorten per fragment en selectief medium.
De genera Mortierella en Mucor zijn geïsoleerd van het medium dat theoretisch gezien
selectief is voor de Pythium en Phytopthora. Mortierella was aanwezig in alle fragmenten,
Mucor enkel in Chawia. Op het medium dat selectief zou moeten zijn voor Fusarium, zijn in
alle fragmenten de genera Trichoderma en Penicillium teruggevonden. Tenslotte zijn de
genera Ilyonectria, Cylindrocarpon, Umbelopsis, Trichosporon en Fusarium aanwezig op het
medium dat normaal selectief is voor het Rhizoctonia. Ilyonectria is aanwezig in Chawia,

Resultaten
30
Fururu, Macha en Ngangao, Umbelopsis in alle fragmenten. Fusarium is enkel aanwezig in
Fururu, Mwachora en Ndiwenyi. Trichosporon was enkel aanwezig in Fururu en Mwachora.
Op fragmentniveau beschrijven het Michaelis-Menten gemidelde, Shannon Exponentieel
gemiddelde en het Simpson gemiddelde de fungi morfotype rijkdom binnen elk fragment
(figuur 4.2). Fururu wordt gekenmerkt door het hoogst aantal fungi morfotypes (MMMeans =
13.98). Chawia, Macha en Mwachora herbergen het laagst aantal morfotypes (MMMeans =
8.31, 8.46 en 8.49 resp.). Het aantal soorten in de fragmenten Ndiwenyi en Ngangao ligt
ergens tussenin (MMMeans = 10 en 9.74 resp.). Het verschil tussen de fragmenten wordt
kleiner indien gekeken wordt naar het effectieve aantal soorten binnen elk fragment (Shannon
Exponentieel gemiddelde en Simpson gemiddelde).
Figuur 4.2: Michaelis-Menten gemiddelde (MMMeans), Shannon (exp) en Simpson gem. vergeleken tussen de fragmenten
Tabel 4.4 geeft de Jaccard indices weer tussen elk
paar van de zes bezochte bosfragmenten. Deze index
geeft de mate van gelijkenis in morfotype
samenstelling tussen twee fragmenten onderling.
Chawia deelt bijvoorbeeld meer soorten met
Ngangao en Macha in plaats van met Fururu,
Ndiwenyi of Mwachora. Verder vertoont Fururu
sterke gelijkenissen met Ngangao, terwijl Macha een
hoge mate van overeenkomst vertoont met
Mwachora.
Tabel 4.4: Jaccard similarity indices tussen de zes bezochte fragmenten
0.00
2.00
4.00
6.00
8.00
10.00
12.00
14.00
16.00
Cha Fur Mac Mwa Ndi Nga
MMMeans
Shannon Exp. Mean
Simpson Mean
Fragment1 Fragment2 Jaccard index
Cha Fur 0.57
Cha Mac 0.70
Cha Mwa 0.42
Cha Ndi 0.50
Cha Nga 0.89
Fur Mac 0.50
Fur Mwa 0.40
Fur Ndi 0.38
Fur Nga 0.62
Mac Mwa 0.60
Mac Ndi 0.42
Mac Nga 0.50
Mwa Ndi 0.42
Mwa Nga 0.45
Ndi Nga 0.55
Resultaten
31
4.2.2. Gemeenschapsanalyse
Voor verdere analyse van de fungi bodemgemeenschap worden de 19 geïsoleerde morfotypes
ingedeeld in drie functionele groepen, en dit op basis van reeds gekende mogelijke effecten op
de fitness van de plant (tabel 4.5):
- Groep 1: Fungi met een potentieel pathogeen effect op de plantengemeenschap
- Groep 2: Fungi met een potentieel antagonistisch effect op pathogene fungi
- Groep 3: Saprotrofe fungi zonder gekende pathogene of antagonistische effecten
Groep 1 Groep 2 Groep 3
Cylindrocarpon spp. Penicillium spp. Mortierella spp.
Fusarium spp. Trichoderma spp. Mucor spp.
Ilyonectria spp. Umbelopsis spp. Trichosporon spp.
Tabel 4.5: Indeling van de geïsoleerde morfotypes op basis van hun potentieel effect op de plantengemeenschap.
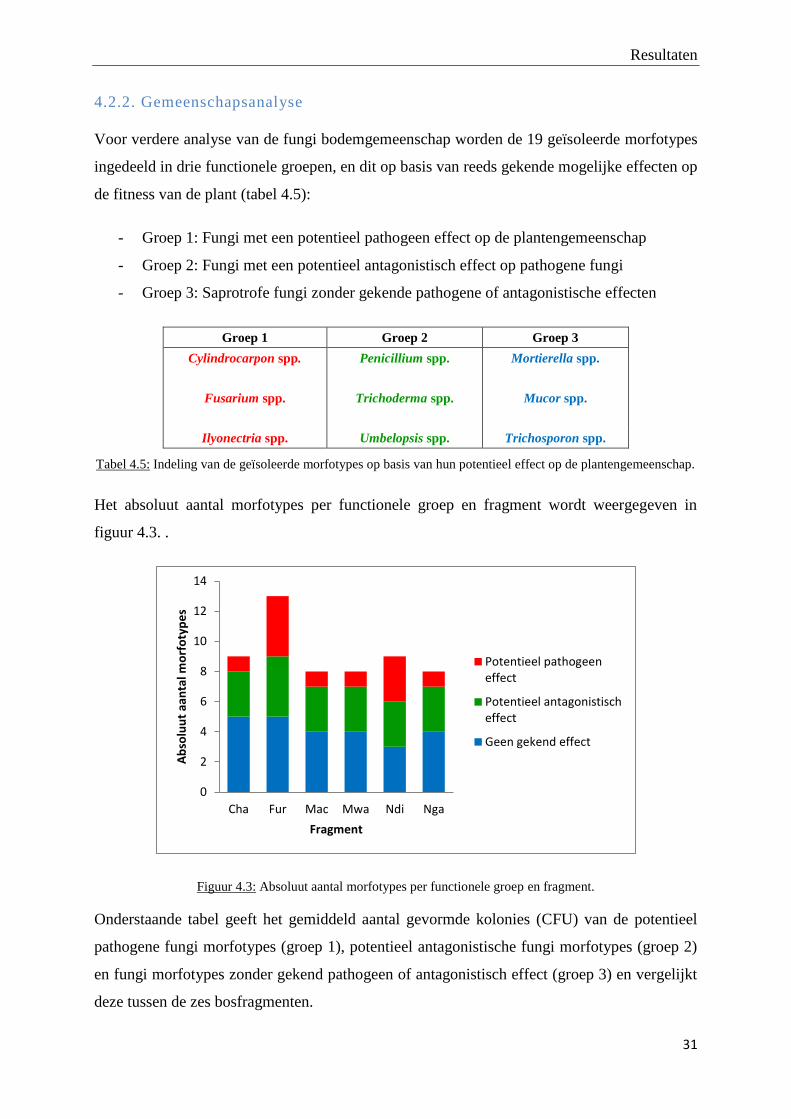
Het absoluut aantal morfotypes per functionele groep en fragment wordt weergegeven in
figuur 4.3. .
Figuur 4.3: Absoluut aantal morfotypes per functionele groep en fragment.
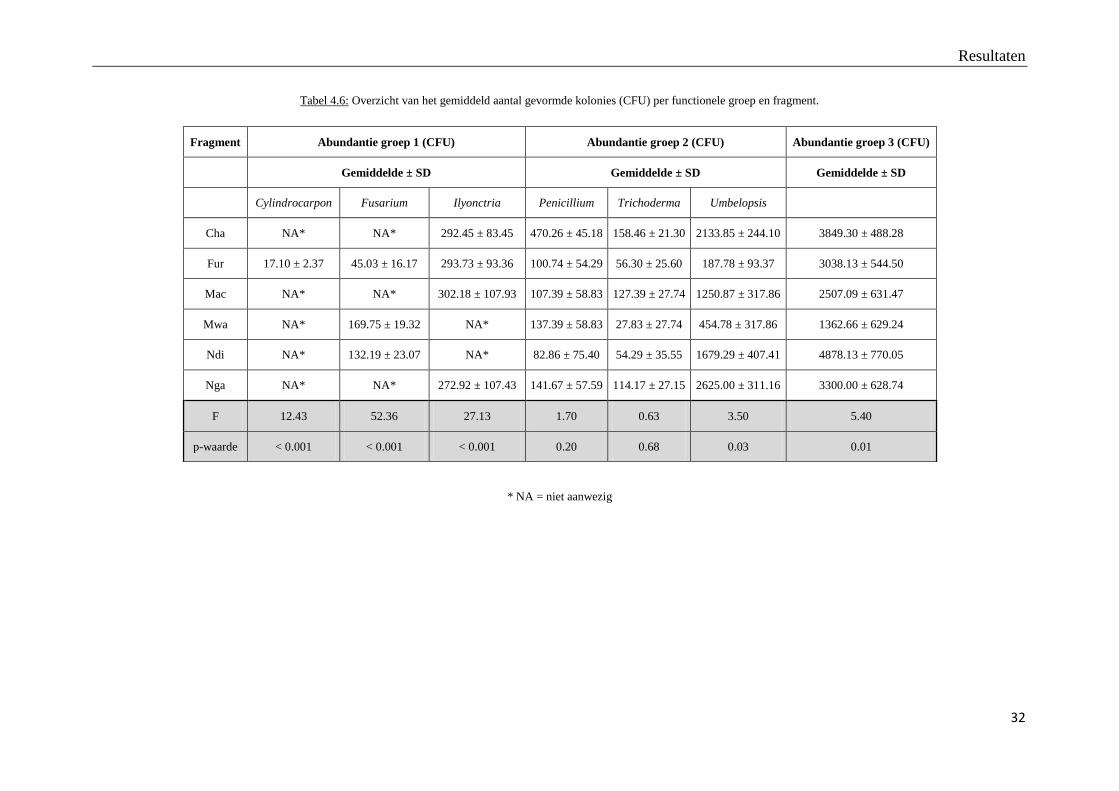
Onderstaande tabel geeft het gemiddeld aantal gevormde kolonies (CFU) van de potentieel
pathogene fungi morfotypes (groep 1), potentieel antagonistische fungi morfotypes (groep 2)
en fungi morfotypes zonder gekend pathogeen of antagonistisch effect (groep 3) en vergelijkt
deze tussen de zes bosfragmenten.
0
2
4
6
8
10
12
14
Cha Fur Mac Mwa Ndi Nga
Ab
solu
ut
aan
tal m
orf
oty
pe
s
Fragment
Potentieel pathogeeneffect
Potentieel antagonistischeffect
Geen gekend effect
Resultaten
32
Tabel 4.6: Overzicht van het gemiddeld aantal gevormde kolonies (CFU) per functionele groep en fragment.
* NA = niet aanwezig
Fragment Abundantie groep 1 (CFU) Abundantie groep 2 (CFU) Abundantie groep 3 (CFU)
Gemiddelde ± SD Gemiddelde ± SD Gemiddelde ± SD
Cylindrocarpon Fusarium Ilyonctria Penicillium Trichoderma Umbelopsis
Cha NA* NA* 292.45 ± 83.45 470.26 ± 45.18 158.46 ± 21.30 2133.85 ± 244.10 3849.30 ± 488.28
Fur 17.10 ± 2.37 45.03 ± 16.17 293.73 ± 93.36 100.74 ± 54.29 56.30 ± 25.60 187.78 ± 93.37 3038.13 ± 544.50
Mac NA* NA* 302.18 ± 107.93 107.39 ± 58.83 127.39 ± 27.74 1250.87 ± 317.86 2507.09 ± 631.47
Mwa NA* 169.75 ± 19.32 NA* 137.39 ± 58.83 27.83 ± 27.74 454.78 ± 317.86 1362.66 ± 629.24
Ndi NA* 132.19 ± 23.07 NA* 82.86 ± 75.40 54.29 ± 35.55 1679.29 ± 407.41 4878.13 ± 770.05
Nga NA* NA* 272.92 ± 107.43 141.67 ± 57.59 114.17 ± 27.15 2625.00 ± 311.16 3300.00 ± 628.74
F 12.43 52.36 27.13 1.70 0.63 3.50 5.40
p-waarde < 0.001 < 0.001 < 0.001 0.20 0.68 0.03 0.01

Resultaten
33
4.2.2.1. Variatie tussen fragmenten en plots
Figuur 4.4 geeft een NMDS plot weer waarbij de abundantie en samenstelling van alle fungi
morfotypes (groep 1, 2 en 3) wordt vergeleken tussen de zes fragmenten en de 20 plots. De
ANOSIM analyse toont significante verschillen aan tussen de fragmenten onderling (R =
0.42, p = 0.001), maar niet tussen de plots van eenzelfde
fragment. De bodemstalen clusteren dus samen op
fragmentniveau en niet op plotniveau. Tabel 4.7 geeft de
resultaten (R- en p-waarden) van de paarsgewijze
vergelijkingen tussen de zes fragmenten. Enkel tussen de
grotere fragmenten Chawia en Ngangao is er geen significant
verschil (p < 0.001). Mwachora verschilt sterk van alle andere
fragmenten met uitzondering van Macha en ook tussen Macha
en Ndiwenyi is er sprake van grote variatie. Dit kan afgeleid
worden vanuit de NMDS plot. De bodemstalen van Chawia en
Ngangao clusteren samen, terwijl deze van Mwachora en
Ndiwenyi zich min of meer afzonderen.
Tabel 4.7: R-waarden en bijhorende p-waarden van de paarsgewijze vergelijkingen tussen de zes bezochte bosfragmenten.
Figuur 4.4: NMDS plot van alle geïsoleerde fungi morfotype (groep 1, 2 en 3).
Groepen R-waarde p-waarde
Cha, Fur 0,30 0,001
Cha, Mac 0,35 0,001
Cha, Mwa 0,74 0,001
Cha, Ndi 0,23 0,001
Cha, Nga 0,08 0.047
Fur, Mac 0,32 0,001
Fur, Mwa 0,76 0.001
Fur, Ndi 0,35 0,001
Fur, Nga 0,22 0,001
Mac, Mwa 0,47 0,001
Mac, Ndi 0,72 0,001
Mac, Nga 0,32 0,001
Mwa, Ndi 0,85 0,001
Mwa, Nga 0,81 0,001
Ndi, Nga 0,32 0,001
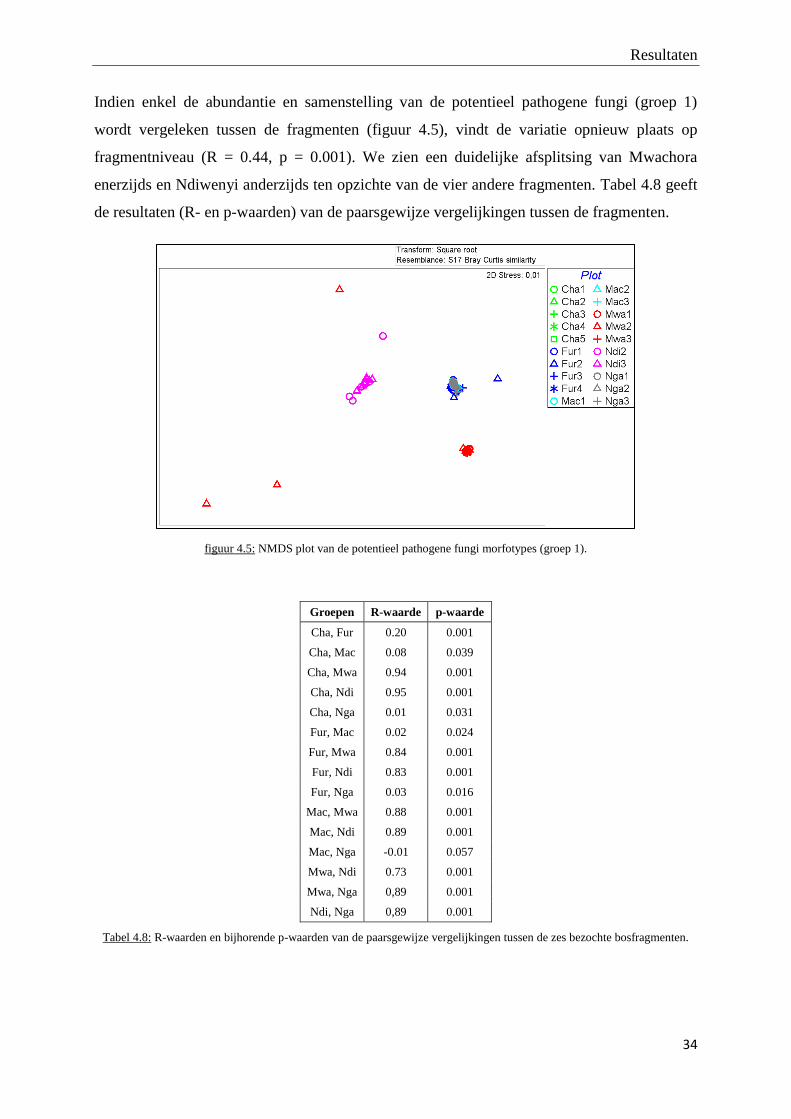
Resultaten
34
Indien enkel de abundantie en samenstelling van de potentieel pathogene fungi (groep 1)
wordt vergeleken tussen de fragmenten (figuur 4.5), vindt de variatie opnieuw plaats op
fragmentniveau (R = 0.44, p = 0.001). We zien een duidelijke afsplitsing van Mwachora
enerzijds en Ndiwenyi anderzijds ten opzichte van de vier andere fragmenten. Tabel 4.8 geeft
de resultaten (R- en p-waarden) van de paarsgewijze vergelijkingen tussen de fragmenten.
figuur 4.5: NMDS plot van de potentieel pathogene fungi morfotypes (groep 1).
Groepen R-waarde p-waarde
Cha, Fur 0.20 0.001
Cha, Mac 0.08 0.039
Cha, Mwa 0.94 0.001
Cha, Ndi 0.95 0.001
Cha, Nga 0.01 0.031
Fur, Mac 0.02 0.024
Fur, Mwa 0.84 0.001
Fur, Ndi 0.83 0.001
Fur, Nga 0.03 0.016
Mac, Mwa 0.88 0.001
Mac, Ndi 0.89 0.001
Mac, Nga -0.01 0.057
Mwa, Ndi 0.73 0.001
Mwa, Nga 0,89 0.001
Ndi, Nga 0,89 0.001
Tabel 4.8: R-waarden en bijhorende p-waarden van de paarsgewijze vergelijkingen tussen de zes bezochte bosfragmenten.

Resultaten
35
Uitsluitend de potentieel antagonistische fungi (groep 2) in beschouwing genomen (figuur
4.6), levert een significant, maar zwak verschil op tussen de fragmenten en plots (R = 0.14, p
= 0.001). De bodemstalen liggen zeer verspreid, wat eerder wijst op variatie binnen dan
tussen de fragmenten en plots. Tabel 4.9 geeft de resultaten (R- en p-waarden) van de
paarsgewijze vergelijkingen tussen de fragmenten.
Figuur 4.6: NMDS plot van de fungi morfotypes met een potentieel antagonistisch effect op de plantengemeenschap.
Groepen R-waarde p-waarde
Cha, Fur 0.34 0.001
Cha, Mac 0.13 0.012
Cha, Mwa 0.26 0.001
Cha, Ndi 0.21 0.015
Cha, Nga 0.07 0.062
Fur, Mac 0.06 0.054
Fur, Mwa 0.00 0.377
Fur, Ndi 0.11 0.036
Fur, Nga 0.23 0.001
Mac, Mwa 0.03 0.114
Mac, Ndi 0.02 0.301
Mac, Nga 0.03 0.116
Mwa, Ndi 0.06 0.095
Mwa, Nga 0,16 0.006
Ndi, Nga 0,05 0.160
Tabel 4.9: R-waarden en bijhorende p-waarden van de paarsgewijze vergelijkingen tussen de zes bezochte bosfragmenten.
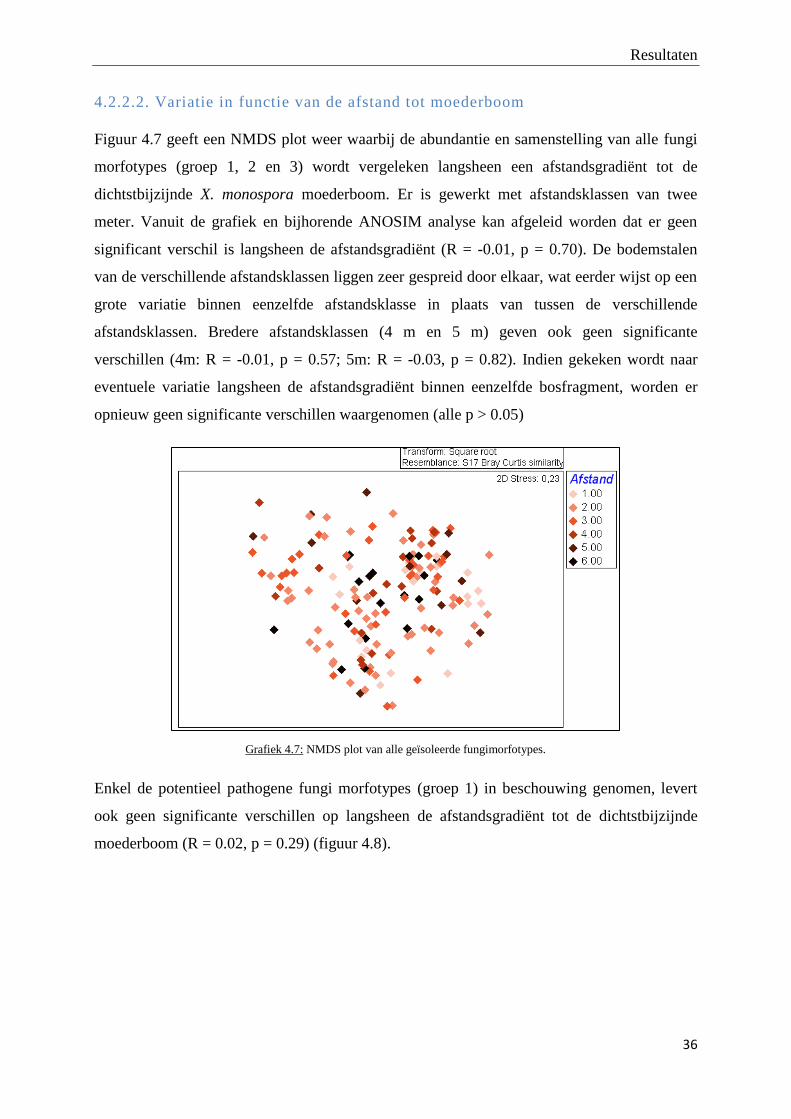
Resultaten
36
4.2.2.2. Variatie in functie van de afstand tot moederboom
Figuur 4.7 geeft een NMDS plot weer waarbij de abundantie en samenstelling van alle fungi
morfotypes (groep 1, 2 en 3) wordt vergeleken langsheen een afstandsgradiënt tot de
dichtstbijzijnde X. monospora moederboom. Er is gewerkt met afstandsklassen van twee
meter. Vanuit de grafiek en bijhorende ANOSIM analyse kan afgeleid worden dat er geen
significant verschil is langsheen de afstandsgradiënt (R = -0.01, p = 0.70). De bodemstalen
van de verschillende afstandsklassen liggen zeer gespreid door elkaar, wat eerder wijst op een
grote variatie binnen eenzelfde afstandsklasse in plaats van tussen de verschillende
afstandsklassen. Bredere afstandsklassen (4 m en 5 m) geven ook geen significante
verschillen (4m: R = -0.01, p = 0.57; 5m: R = -0.03, p = 0.82). Indien gekeken wordt naar
eventuele variatie langsheen de afstandsgradiënt binnen eenzelfde bosfragment, worden er
opnieuw geen significante verschillen waargenomen (alle p > 0.05)
Grafiek 4.7: NMDS plot van alle geïsoleerde fungimorfotypes.
Enkel de potentieel pathogene fungi morfotypes (groep 1) in beschouwing genomen, levert
ook geen significante verschillen op langsheen de afstandsgradiënt tot de dichtstbijzijnde
moederboom (R = 0.02, p = 0.29) (figuur 4.8).

Resultaten
37
Grafiek 4.8: NMDS plot van de fungi morfotypes met een potentieel pathogeen effect op de plantengemeenschap.
De potentieel antagonistische fungi morfotypes (groep 2) ten slotte vertonen ook geen
significant verschil in abundantie en samenstelling langsheen de afstandsgradiënt (R = -0.03,
p = 0.87) (figuur 4.9).
Grafiek 4.9: NMDS plot van de fungi morfotypes met een potentieel antagonistisch effect op de plantengemeenschap.
De stress-waarde van alle NMDS plots is niet groter dan 0.30, waardoor we kunnen aannemen
dat de ze een representatieve weergave van de werkelijkheid zijn.

Resultaten
38
4.2.3. Verband X. monospora en bodemfungi distributie
Tabel 4.10 en 4.11 geven de resultaten weer van de PCA analyses, waarbij per gemeten
parameter wordt weergegeven hoeveel deze van de waargenomen variatie in fungi abundantie
en samenstelling verklaart (r2) en of dit significant is of niet (p-waarde). Vanuit de PCA
analyse waarbij alle fungi morfotypes in rekening werden gebracht, blijkt dat de eerste vier
assen (PC’s) meer dan 80% van de waargenomen variatie verklaren. Indien deze gekoppeld
worden met de gemeten parameters, zien we dat geen enkele van de parameters een hoge
mate van variatie verklaard. De relatieve densiteit aan X. monospora adulten verklaart het
meest (6%) van alle parameters, maar dit is op zich nog steeds zeer laag. De PCA analyse
waarbij enkel de fungi morfotypes met een potentieel pathogeen of antagonistisch effect op de
plantengemeenschap in rekening werden gebracht, toont aan dat de eerste drie assen (PC’s)
meer dan 80% van de waargenomen variatie verklaren. Indien deze opnieuw gekoppeld
worden met de gemeten parameters, zien we dat van alle parameters de densiteit aan X.
monospora zaailingen het meeste van verklaard (8%), maar dit is opnieuw zeer weinig.
Parameter r2 p-waarde
Kroonbedekking (%) 0.02 0.29
Relatieve densiteit X. monospora adulten 0.06 0.01
Diameter X. monospora adulten (cm) 0.03 0.11
Densiteit X. monsopora zaailingen (aantal / m2) 0.04 0.07
Lengte X. monospora zaailingen (cm) 0.03 0.13
Afstand X. monospora zaailing-moederboom (m) 0.04 0.04
Tabel 4.10: Percentage verklaarde variatie per parameter en bijhorende p-waarde (alle fungi morfotypes).
Parameter r2 p-waarde
Kroonbedekking (%) 0.01 0.45
Relatieve densiteit X. monospora adulten 0.06 0.01
Diameter X. monospora adulten (cm) 0.04 0.04
Densiteit X. monsopora zaailingen (aantal / m2) 0.09 0.001
Lengte X. monospora zaailingen (cm) 0.07 0.01
Afstand X. monospora zaailing-moederboom (m) 0.06 0.08
Tabel 4.11: Percentage verklaarde variatie per parameter en bijhorende p-waarde (potentieel effect).

Discussie
39
5. Discussie
Variatie in abundantie en samenstelling van bodemfungi, aanwezig in de rhizosfeer van X.
monsopora zaailingen, werd voornamelijk waargenomen op fragmentniveau. Twee kleine en
sterk verstoorde fragmenten vertoonden opmerkelijke verschillen met de grote, minder sterk
verstoorde fragmenten. De bodemfungi gemeenschap vertoonde echter geen variatie
langsheen een afstandsgradiënt tussen X. monospora zaailingen en hun dichtstbijzijnde
moederboom.
5.1. Ruimtelijke distributie van bodemfungi in relatie tot X. monospora
5.1.1. Functionele diversiteit
Ondanks het gebruik van selectieve media voor de pathogene fungi genera Pythium,
Phytophthora, Fusarium en Rhizoctonia, werden, met uitzondering van Fusarium, vijf andere
fungi genera met een potentieel effect op X. monospora geïsoleerd. Ilyonectria en
Cylindrocarpon werden als potentieel pathogeen beschouwd, terwijl Penicillium,
Trichoderma en Umbelopis potentieel antagonistisch werden geacht. Verder werden de
saprotrofe fungi genera Mortierella en Mucor geïsoleerd, die een belangrijke rol spelen bij de
decompositie van organisch materiaal.
Het genus Ilyonectria (Ascomycota) werd recent geïdentificeerd als potentieel pathogeen en is
berucht omwille van zijn verwoestend effect op wijngaarden (Cabral et al., 2012). Recent
werd de ermee geassocieerde ziekte (black foot disease) ook waargenomen op avocadobomen
(Vitale et al., 2012) en olijfbomen (Urbez-Torres, 2012). De ziekte gaat gepaard met een
gereduceerde plantengroei, uitgesteld ontspruiten, verkorting van de internoden en chloriose
en resulteert veelal in sterfte van de plant. Het genus Cylindrocarpon (Ascomycota) herbergt
de teleomorfen van Ilyonectria en veroorzaakt wortelrot in verscheidene, economisch
belangrijke gastheerplanten (druivelaars, appelaars, beuk) (Cabral, 2012). De rol van deze
twee genera als potentiële pathogenen in natuurlijke (bos)ecosystemen dient verder
onderzocht te worden.
Penicillium kan antagonistische effecten uitoefenen op damping-off veroorzakende
fytopathogene fungi, zoals Pythium, Phytopthora, Fusarium en Rhizoctonia, en dit in zowel
natuurlijke ecosystemen als landbouwsystemen (Yamaji, 2005; Nicoletti et al., 2004). Via
mechanismen die de resistentie van de plant verhogen, de productie van antibiotische stoffen

Discussie
40
(antibiose) en de vorming van mycoparasitaire interacties, kunnen ze de negatieve gevolgen
van pathogene fungi beperken of zelfs helemaal teniet doen (Nicoletti et al., 2004). Het genus
Trichoderma (Ascomycota) herbergt de anamorfen van het genus Hypocrea en kent een zeer
brede ecologische amplitude (Harman, 2006). Deze niet-gastheerspecifieke bodemfungi
koloniseren in geen tijd gezonde plantenwortels. Het is dan ook niet verwonderlijk dat
Trichoderma tal van mechanismen heeft ontwikkeld om enerzijds de fytopathogene fungi aan
te vallen en anderzijds de wortelgroei en resistentie van de plant ten opzichte van pathogenen
te verhogen (Harman, 2006; John et al., 2010). Dankzij deze eigenschappen kunnen sommige
Trichoderma soorten een belangrijke rol spelen op zowel ecologisch als economisch vlak. Zo
kunnen ze gebruikt worden bij de biologische controle van de diverse, door pathogene fungi
veroorzaakte, plantenziektes (Harman, 2006). Umbelopsis (Zygomycota) wordt wereldwijd
teruggevonden in de bodem van diverse bosecosystemen (Meyer & Gams, 2003). Het genus
wordt veelal geassocieerd met plantenwortels (Hoff et al., 2004), maar hun ecologische rol is
tot op heden een vraagteken. Er zijn reeds aanwijzingen voor antagonistische effecten tegen
fytopathogene fungi en hun gebruik als biologische controle agentia, maar meer onderzoek is
nodig om te achterhalen hoe Umbelopsis soorten interageren met andere bosorganismen, op
welke manier ze hun gastheerplant infecteren en welke de effecten zijn op de omgevende
plantengemeenschappen (Hoff et al., 2004).
5.1.2.Variatie in bodemfungi gemeenschap tussen de fragmenten
De bodemfungi gemeenschap in de rhizosfeer van X. monospora zaailingen vertoonde sterke
verschillen tussen de fragmenten onderling. Op basis van de volledige dataset (groep 1, 2 en
3), zagen we een clustering van de twee grote fragmenten (Chawia en Ngangao), terwijl twee
kleinere fragmenten (Mwachora en Ndiwenyi) zich duidelijk afzonderden (figuur 4.4). Indien
enkel gekeken werd naar de potentieel pathogene fungi (groep 1), splitsten de bodemstalen
van Mwachora enerzijds en Ndiwenyi anderzijds zich nog duidelijker af van de andere vier
fragmenten (figuur 4.5). Louter de potentieel antagonistische fungi (groep 2) in beschouwing
genomen, gaf slechts zwakke verschillen tussen de bosfragmenten (figuur 4.6). De
waargenomen variatie tussen de fragmenten is dus voornamelijk te wijten aan verschillen in
abundantie en soortensamenstelling van potentieel pathogene bodemfungi.
We registreerden een afwezigheid van Ilyonectria in Ndiwenyi en Mwachora, terwijl deze
zeer abundant aanwezig waren in Chawia en Ngangao (en ook in Fururu en Macha). Het
genus Fusarium daarentegen was enkel aanwezig in Mwachora en Ndiwenyi (en in mindere

Discussie
41
mate in Fururu). Een belangrijke opmerking hierbij is dat in Mwachora slechts één Fusarium
soort (Fusarium culmorum) is teruggevonden, terwijl dit er in Ndiwenyi drie waren, vandaar
de clustering van Mwachora enerzijds en Ndiwenyi anderzijds. Een recente bosbrand kan
(mede)verantwoordelijk zijn voor de afsplitsing van Ndiwenyi. Destabilisering van een
ecosysteem, veroorzaakt door abiotische (brand, overstroming) of biotische (herbivorie,
ziekte, insectenplaag) verstoringen, brengt wijzigen teweeg in de bodemstructuur,
nutriëntenbeschikbaarheid en (a)biotische bodemcomponenten (Hoff et al., 2004; McMullan-
Fisher et al., 2011). De gevolgen zijn zeer variabel en afhankelijk van diverse factoren, zoals
bodem- en vegetatietype, brandintensiteit, -frequentie en -geschiedenis (McMullan-Fisher et
al., 2011). Widden en Parkinson (1970) toonden een gereduceerd aantal Trichoderma en
Penicillium soorten aan in recent afgebrande plots ten opzichte van plots waar er geen brand
had gewoed. Daarenboven constateerden ze een stimulerend effect van brand op de groei van
Cylindrocarpon. Hun hypothese luidde dan ook als volgt: brand heeft een negatief effect op
antagonistische fungi zoals Trichoderma en Penicillium, waardoor de groei van potentieel
pathogene fungi zoals Cylindrocarpon (teleomorfen van Ilyonectria) gestimuleerd wordt. Ook
Stendell et al. (1999) toonden aan dat de ectomycorrhiza biomassa tot acht keer gereduceerd
kan zijn ten gevolge van een brand. Ectomycorrhiza verhogen de absorptiecapaciteit van
zaailingwortels, bieden bescherming tegen fytopathogenen, chemische extremen of
bodemvervuiling en spelen dus een belangrijke rol bij de succesvolle vestiging van zaailingen
(Yamaji, 2001; Hoff et al., 2004). Sharma (1981) daarentegen stelde vast dat Penicillium en
Trichoderma als eerste fungi genera op het toneel verschenen na een brand. Reaves et al.
(1990) voegde hieraan toe dat Trichoderma isolaten, afkomstig van een afgebrand gebied,
sterkere antagonistische eigenschappen vertoont dan Trichoderma isolaten afkomstig van een
controlegebied. De brand in Ndiwenyi kan mogelijks de groei van Fusarium gestimuleerd
hebben en deze van potentiële antagonisten (Penicillium en Trichoderma) gereduceerd
hebben (tabel 4.6). Verder onderzoek dient de impact van externe verstoringen, zoals brand,
op de bodemfungi gemeenschap van de Taita Hills duidelijk maken. Wat ook opviel, was dat
het genus Trichoderma sterker vertegenwoordigd was in Chawia en Ngangao, dit zijn de
fragmenten waar Fusarium afwezig was. Trichoderma zou wel eens de aanwezigheid van
Fusarium onderdrukken via de eerder besproken mechanismen van mycoparasitisme,
antibiose of competitie, en hierdoor een positief effect uitoefenen op de zaadkieming,
zaailingrekrutering en -overleving X. monospora. Het genus Ilyonectria was echter wel
abundant aanwezig in deze fragmenten. Trichoderma is dus mogelijks niet antagonistisch ten
opzichte deze pathogeen, maar het kan ook zijn dat Ilyonectria geen pathogeen effect

Discussie
42
uitoefent op X. monospora. Het negatief effect van Ilyonectria werd tot nu toe namelijk enkel
geassocieerd met druivelaars, avocado- en olijfbomen.
De graad van habitatfragmentatie kan verantwoordelijk zijn voor de verschillen tussen de
grote (Chawia en Ngangao) en twee kleine fragmenten (Mwachora en Ndiwenyi). Lehouck et
al. (2009) observeerden een gereduceerde zaaddispersie in kleine en sterk verstoorde
fragmenten door de afwezigheid van enkele belangrijke zaadverbreiders. Als gevolg van
randeffecten zal de impact van antropogene activiteiten (landbouw, exotische plantages, enz.),
en de daarmee gepaard gaande veranderingen in fysische, chemische en biologische
bodemeigenschappen (Kourtev et al., 2002), op de natuurlijke dynamiek van het
bosecosysteem meer uitgesproken worden naarmate de fragmenten kleiner worden. De hoge
abundantie aan Fusarium in Mwachora en Ndiwenyi is mogelijks het gevolg van deze
randeffecten en kan, in combinatie met een gereduceerde zaaddispersie, negatieve gevolgen
hebben in termen van zaadkieming, zaailingrekrutering en -overleving van X. monospora.
Wat zeker niet over het hoofd mag gezien worden, is dat Macha en Fururu, net zoals
Mwachora en Ndiwenyi, kleine en sterk verstoorde fragmenten zijn. Toch vertoonden ze meer
gelijkenis met de grote fragmenten (Chawia en Ngangao). Er zijn dus nog andere factoren dan
de graad van habitatfragmentatie en de ermee gepaard gaande randeffecten verantwoordelijk
voor de waargenomen variatie tussen de fragmenten. Onze opgemeten parameters, waaronder
kroonbedekking, densiteit aan X. monospora adulten en zaailingen, diameter van X.
monospora adulten, lengte van X. monospora zaailingen en afstand tussen zaailing en
dichtstbijzijnde moederboom, boden echter weinig verklaring. Dit was enigszins te
verwachten aangezien de parameters op zich weinig variatie vertoonden tussen de
fragmenten. Andere, niet-gekwantificeerde omgevingsvariabelen, zoals klimatologische en
topografische kenmerken, vegetatiestructuur, chemische en fysische bodemkenmerken (pH,
dikte van de strooisellaag) bieden mogelijks meer verklaring, maar dienen verder onderzocht
te worden.
5.1.3.Variatie in bodemfungi gemeenschap tussen de plots
De bodemfungi gemeenschap verschilde niet significant tussen de plots van eenzelfde
fragment. In de kleinste fragmenten (Macha, Mwachora en Ndiwenyi) was dit enigszins te
verwachten aangezien de plots zeer dicht bij elkaar lagen. Ook in het grootste fragment
Ngangao lagen de plots zeer dicht bij elkaar omdat er enkel in het meest zuidelijke deel X.
monospora adulten en zaailingen werden aangetroffen. De plots in Chawia en Fururu lagen

Discussie
43
meer verspreid over het fragment, maar ook daar werden geen noemenswaardige verschillen
teruggevonden op plotniveau. Onze bevindingen verschillen in dit opzicht van deze van
Malmivaara-Lämsä et al. (2008). Zij registreerden variatie in de bodemfungi gemeenschap
langsheen twee afstandsgradiënten binnen eenzelfde bosfragment, één ten opzichte van de
rand van het bos en ten opzichte van de aangelegde wandelpaden in het bos. Veranderingen in
bodemkenmerken (voornamelijk pH) en vegetatiestructuur langsheen deze afstandsgradiënten
lagen aan de basis van de waargenomen variatie. De rand van een bos wordt gekenmerkt door
meer wind en lichtinval. Dit creëert warmere en drogere condities in vergelijking met het
centrum van het bos en verandert de samenstelling en activiteit van bodemorganismen (Chen
et al., 1993). Hierdoor treedt een reductie op in de snelheid van decompositie, met alle
gevolgen van dien voor de nutriëntencyclus en bovengronds levende gemeenschappen.
Randeffecten kunnen een invloed uitoefenen over enkele tientallen tot honderden meters naar
het centrum van het bos toe (Chen et al., 1993). Antropogene verstoringen in reeds
gefragmenteerde bossen, zoals het aanleggen van wandelpaden, creëren fragmentatie en dus
ook variatie in bodemgemeenschap op kleinere schaal (Malmivaara-Lämsä, 2008). Indien de
plots op verschillende plaatsen ten opzichte van de bosrand werden uitgezet, konden deze
randeffecten nagegaan worden. In sommige fragmenten (Ngangao, Mwachora) lagen alle
plots echter aan de rand van het fragment omdat enkel daar X. monsopora adulten en
zaailingen werden aangetroffen. De meeste fragmenten van de Taita Hills zijn ook zodanig
klein dat randeffecten een grote invloed uitoefenen naar het centrum toe, waardoor er niet echt
sprake kan zijn van een afstandsgradiënt.
5.1.4. Variatie in bodemfungi gemeenschap in functie van afstand tot
moederboom
Het belangrijkste onderdeel van deze masterproef was nagaan of de eerder waargenomen
gereduceerde zaadkieming, zaailingrekrutering en -overleving van X. monospora in de buurt
van conspecifieke adulten (Lehouck et al., 2009c) kan verklaard worden door de accumulatie
van gastheerspecifieke bodempathogenen in de buurt van de moederboom. Verstoorde
bosfragmenten worden gekenmerkt door de afwezigheid van enkele belangrijke
zaadverbreiders waardoor minder zaden verbreid worden en dus meer zaden blijven liggen
onder de moederboom. Dit creëert een ideale niche voor natuurlijke vijanden zoals
fytopathogenen en herbivoren. Door de bodemfungi abundantie en samenstelling te
vergelijken langsheen verschillende afstanden ten opzichte van de moederboom, trachtten we
te achterhalen of er sprake is van afstandsafhankelijke zaailingmortaliteit ten gevolge van

Discussie
44
gastheerspecifieke, fytopathogene fungi. Janzen (1970) veronderstelde bij het formuleren van
zijn hypothese dat de J-C patronen meer uitgesproken zijn indien de adulte bomen een
verspreid distributiepatroon vertonen. Deze voorwaarde was in ons geval alvast voldaan
(bijlage 1). Toch werden geen verschillen gevonden langsheen de afstandsgradiënt (figuur
4.7, 4.8 en 4.9). Dit kan te wijten zijn aan het feit dat enkel de bodemfungi, aanwezig in de
rhizosfeer van X. monospora zaailingen, werden onderzocht en niet diegene in of op de
zaailingwortels. Het kan dus zijn dat we te maken hebben met niet-gastheerspecifieke
bodemfungi. Een belangrijke voorwaarde tot het waarnemen van J-C effecten is nu net die
gastheerspecificiteit. Vervolgens moet opgemerkt worden dat er gebruik werd gemaakt van
GPS coördinaten om de positie van X. monospora zaailingen en adulten in kaart te brengen en
de euclidische afstand tussen zaailing en de dichtstbijzijnde moederboom te berekenen. Het
gebruikte GPS toestel had echter een behoorlijk grote standaardfout van 7.6 m. Hierdoor kan
het zijn dat een zaailing zich volgens de GPS op een afstand van bv. 1 m ten opzichte van de
moederboom bevindt, terwijl hij in werkelijkheid veel verder ligt. De verkregen
afstandsgradiënt is dus mogelijks onvoldoende betrouwbaar. Een andere verklaring kan zijn
dat het mechanisme van negatieve afstandsafhankelijke zaailingmortaliteit niet detecteerbaar
was op het moment van staalname. De bodemstalen werden genomen gedurende een zeer
korte periode van twee weken, terwijl de waarnemingen van zaadkieming, zaailingrekrutering
en -overleving plaatsvonden over een periode van meerdere jaren, met veel variatie doorheen
de jaren (Lehouck et al., 2009). De sterkte van J-C effecten varieert in tropische bossen dus
niet alleen op ruimtelijke schaal, maar ook tussen seizoenen en soorten (Lin et al., 2012).
Onderzoek betreffende de onderliggende mechanismen van deze temporale en interspecifieke
variatie en zijn gevolgen voor de co-existentie tussen soorten staat echter nog in zijn
kinderschoenen (Lin et al., 2012). Tot nu toe focusten de meeste studies (waaronder deze
masterproef) zich enkel op de rol van de natuurlijke vijanden, terwijl de competitie voor
limiterende bronnen, het ander mechanisme achter CNDD, over het hoofd wordt gezien
(Freckleton & Lewis, 2006). Tropische, seizoensgebonden bossen ondervinden temporele
variatie in waterbeschikbaarheid, beter gekend als de droge en natte seizoenen. Een
gereduceerde waterbeschikbaarheid verhoogt bijvoorbeeld de graad van competitie en dus
stressgevoeligheid van de plant waardoor deze laatste makkelijker wordt aangevallen door
herbivoren en pathogenen (Lin et al., 2012). Boomsoorten verschillen ook vaak in hun manier
van wateropname (bv. ten gevolge van interspecifieke variatie in wortellengte) (Meinzer et
al., 1999) en dit verhoogt de intraspecifieke competitie gedurende perioden van waterstress.
De overleving van algemeen voorkomende planten wordt hierdoor gereduceerd ten voordele

Discussie
45
van de meer zeldzame soorten, bijdragend aan het behoud van soortendiversiteit.
Gecombineerd onderzoek over de rol van zowel natuurlijke vijanden als competitie voor
limiterende bronnen wordt sterk aangeraden.
Aangezien de pathogene bodemfungi gemeenschap niet verschilde langsheen de
afstandsgradiënt ten opzichte van de moederboom, kunnen we veronderstellen dat de
overleving van X. monospora zaailingen niet gereduceerd wordt in de omgeving van deze
conspecifieke bomen. De zaailingen kunnen volwaardig deelnemen in de competitie met
andere zaailingsoorten, waardoor ze een grotere kans hebben om een plekje te veroveren
onder de moederboom. De afwezigheid van J-C effecten kan hierdoor verantwoordelijk zijn
voor een afname in plantendiversiteit aangezien een gereduceerde zaailingoverleving bij hoge
conspecifieke densiteit, in de omgeving van de moederboom, als een van de belangrijkste
mechanismen wordt beschouwd voor het verkrijgen van co-existentie tussen plantensoorten.
Onderzoek op langere termijn is noodzakelijk om meer te weten te komen over de seizoenale
variatie in J-C effecten en hoe, vaak door de mens geïnduceerde, veranderingen in
omgevingscondities een effect hebben op de mechanismen van co-existentie tussen tropische
planten.

Discussie
46
5.2. Studiedesign: moeilijkheden en mogelijke verbeteringen
5.2.1. Staalname
De veldwerkperiode vond plaats in de sterk gefragmenteerde bosfragmenten van de Taita
Hills waarbij plots (15 x 15 m) werden uitgezet op plaatsen met X. monospora moederbomen
en zaailingen. Binnen elke plot werden bodemstalen genomen in de rhizosfeer van acht van de
25 gekarteerde zaailingen. Het was nuttig geweest om in diezelfde plots alle (in plaats van
slechts 25) zaailingen te karteren en in de rhizosfeer van elke zaailing een bodemstaal te
nemen. Op deze manier kon het ruimtelijk distributiepatroon en de densiteit van X.
monospora zaailingen in elke plot bepaald worden en waren er meer replicaten ter
beschikking om de bodemfungi gemeenschap te bekijken. De voornaamste reden waarom
gekozen werd voor eerst vermelde methode is het gebrek aan tijd en de beperkte toegestane
hoeveelheid bodem dat getransporteerd mocht worden van Kenia naar België. De
veldwerkperiode nam twee weken in beslag, waardoor op korte termijn enerzijds
verschillende bosfragmenten moesten bezocht worden en anderzijds genoeg plots (replicaten)
binnen eenzelfde bosfragment moesten uitgezet worden. Een andere methode zou kunnen zijn
om in elk bosfragment op zoek te gaan naar X. monospora moederbomen, een cirkel van
bijvoorbeeld 20 m te trekken rondom de moederboom en vervolgens bodemstalen te nemen in
de rhizosfeer van de aanwezige zaailingen, op verschillende afstanden ten opzichte van de
moederboom. Deze GPS-loze manier van werken biedt het voordeel dat er een gekende
afstandsgradiënt wordt gecreëerd ten opzichte van de moederboom, waardoor de exacte
afstand waarop de bodemstalen genomen worden, gekend is. GPS toestellen bieden het
voordeel snel te kunnen werken maar zijn nadelig omwille van hun redelijke grote
standaardfout (hier 7.6 m, dus relatief groot in plots van 15 x 15 m). De fout is waarneembaar
in sommige plots (bijlage 2), waarbij te zien is dat sommige zaailingen en adulte bomen
buiten de plot liggen terwijl deze wel degelijk binnen de plot werden gekarteerd. Verder kan
in elke plot het aantal soorten gevestigde zaailingen bepaald worden, alsook het aantal
individuen per soort. Volgens de J-C hypothese zou het aantal soorten zaailingen en het aantal
zaailingen van eenzelfde soort respectievelijk toenemen en afnemen naar de moederboom toe.

Discussie
47
5.2.2. Labo analyse
De bedoeling was om enkel bodemfungi met een potentieel pathogeen effect op X. monospora
zaailingen te bestuderen. Er werd dan ook gekozen te werken met selectieve media die de
groei van reeds gekende pathogene fungi genera (Pythium, Phytopthora, Fusarium en
Rhizoctonia) stimuleren en deze van andere organismen (bacteriën, gisten, enz.) inhiberen.
Het resultaat was niet geheel zoals verwacht. Met uitzondering van Fusarium, werden
bovengenoemde genera niet teruggevonden. In plaats daarvan werden zowel potentieel
pathogene fungi (Fusarium, Ilyonectria/Cylindrocarpon) als fungi met een potentieel
antagonistisch effect op deze pathogenen (Penicillium, Trichoderma, Umbelopsis) geïsoleerd.
Ook saprotrofe bodemfungi zonder gekend pathogeen of antagonistisch effect (Mortierella,
Mucor, Trichosporon) waren dominant aanwezig. Deze resultaten staven de moeilijkheden
van de klassieke uitplaatmethoden. Sommige fungi zijn zeer gevoelig en vereisen specifieke
groeicondities die niet altijd nagebootst kunnen worden onder laboratoriumcondities. Ook de
zogenaamd ‘selectieve’ media zijn niet altijd zo effectief als gedacht. Cultuurafhankelijke
methoden selecteren vaak voor bodemorganismen die gemakkelijk in cultuur zijn op te
groeien, waardoor andere, minder competitieve maar wel in het bodemstaal aanwezige
organismen niet geïdentificeerd worden. Deze bias mag zeker niet over het hoofd gezien
worden (Hoff et al., 2004). Het kan natuurlijk ook gewoon zijn dat de genera Pythium,
Phytopthora en Rhizoctonia niet aanwezig zijn in de rhizosfeer van X. monospora zaailingen
van de Taita Hills.
Moleculaire analyses kunnen een alternatief bieden voor deze uitplaatmethode, maar kennen
ook een aantal beperkingen. De PCR methode bijvoorbeeld is zeer gevoelig en vereist
nauwgezette handelingen die toch enige ervaring vergen. Het sequeneren en vervolgens
identificeren van de organismen is ook niet altijd evident. De BLAST analyse geeft het
overeenkomstpercentage weer tussen de sequentie van het geïsoleerd organisme met een
sequentie aanwezig in de databank. Dit is echter nooit 100% en vaak worden meerdere
organismen (soms ‘unidentified species’) met hetzelfde percentage aangeduid als mogelijke
kandidaten. De fungi genome library is ook nog niet zo uitgebreid als bijvoorbeeld de
bacteriële variant, en bevat een beperkte set aan fungi geïsoleerd uit (sub)tropische bodems.
Een combinatie van deze BLAST-resultaten, morfologische (macro- en microscopie) en
fysiologische (eco-plates) eigenschappen leveren de meest betrouwbare identificaties op.

Discussie
48
5.3. Suggesties voor verder onderzoek
Deze masterproef licht slechts een klein tipje van de sluier over de ruimtelijke variatie in
bodemfungi abundantie en samenstelling tussen de sterk verstoorde bosfragmenten van de
Taita Hills en kan een aanzet zijn tot verder onderzoek binnen de wondere wereld van de
Taita bodemorganismen. Het verwerven van meer kennis over eventuele wijzigingen in de
dynamiek van de bodemgemeenschap, en hierdoor ook de bovengronds levende
gemeenschappen, ten gevolge van de alsmaar toenemende humane impact kan van belang zijn
bij het herstellen en behouden van de bosintegriteit via kleinschalige maar doeltreffende
beheersplannen. Enkele mogelijke suggesties voor verder onderzoek worden hieronder dan
ook aangeboden.
- Nagaan in welke mate de bodemfungi, geïsoleerd vanuit de rhizosfeer van X.
monospora zaailingen, overeenkomen met deze aanwezig in of op de zaailingwortels.
Hiervoor kan gebruik gemaakt worden van de wortelstalen die in deze masterproef
niet verder geanalyseerd zijn. Indien blijkt dat de geïsoleerde bodemfungi ook de
zaailingwortels koloniseren, kan met meer zekerheid bevestigd worden dat ze een
potentieel effect hebben op de plant.
- Nagaan welke effecten de geïsoleerde bodemfungi hebben op X. monospora
zaailingen. Op basis van de literatuur werd aangenomen dat Fusarium en Ilyonectria
mogelijks pathogene effecten uitoefenen en dat Penicillium, Trichoderma en
Umbelopsis mogelijks een antagonistisch effect hebben op deze pathogene fungi. De
vraag of dit ook van toepassing is voor de tropische boomsoort X. monospora blijft
echter onopgelost. Dit kan onderzocht worden met behulp van bio-essays, waarbij de
geïsoleerde morfotypes afzonderlijk geïnoculeerd worden in de steriele bodem (bodem
vrij van andere organismen) rondom opgekweekte X. monospora zaailingen. Het
effect op de plant kan dan rechtstreeks bestudeerd worden. Hierbij moet rekening
worden gehouden met het feit dat verschillende strains van eenzelfde soort een ander
effect kunnen hebben op de plant. De ene strain van Fusarium oxysporum kan
bijvoorbeeld een fytopathogeen zijn, terwijl een andere strain een positieve invloed
heeft op de plant (Augspurger, 2007). Naast de pathogeniciteit, kan ook de
gastheerspecificiteit onderzocht worden door bovenstaande bio-essay te herhalen met
zaailingen van andere boomsoorten aanwezig in de bosfragmenten van de Taita Hills.

Discussie
49
Dit vereist weliswaar het verzamelen van zaden van verschillende boomsoorten, dat
praktisch gezien niet evident is in tropische bossen wegens de onvoorspelbare
fenologie van tropische boomsoorten (Lehouck et al., 2009b).
- Nagaan of er sprake is van seizoenale variatie in J-C effecten. Meerjarige studies,
waarbij bodemstalen worden genomen op verschillende momenten van het jaar,
worden hierbij aangeraden. Omgevingsparameters zoals klimatologische en
topografische kenmerken, vegetatiestructuur, chemische en fysische bodemkenmerken
(pH, dikte van de strooisellaag) die op het moment van staalname worden opgemeten,
kunnen dan gelinkt worden met de eventueel waargenomen variatie in J-C effecten.
- Nagaan welke effecten antropogene verstoringen (klimaatsverandering,
habitatfragmentatie, enz.) hebben op de bodemfungi abundantie, samenstelling en
werking (pathogeen, antagonistisch). Het effect van klimaatsverandering bijvoorbeeld
kan opnieuw uitgevoerd worden met behulp van bio-essays waarbij de bodemfungi
gemeenschap wordt bekeken onder verschillende klimatologische condities
(verhoogde temperatuur, CO2-concentratie, veranderingen in vochtigheidsgraad, enz.).

Conclusie
50
6. Conclusie
Op basis van de verkregen resultaten, kunnen volgende conclusies gemaakt worden:
- De bodemfungi gemeenschap, aanwezig in de rhizosfeer van X. monospora zaailingen,
vertoonde variatie op fragmentniveau. Twee kleine en sterk verstoorde bosfragmenten
(Mwachora en Ndiwenyi) verschilden significant van de grote en minder sterk
verstoorde bosfragmenten (Chawia en Ngangao). Habitatfragmentatie leek de meest
voor de hand liggende verklaring te zijn voor de waargenomen variatie, ware het niet
dat de twee andere kleine fragmenten (Fururu, Macha) meer gelijkenis vertoonden met
de grote fragmenten. Onze opgemeten parameters boden echter geen verklaring voor
de deze waargenomen variatie. Andere omgevingsvariabelen, alsook externe
verstoringen (zoals een brand) bieden mogelijks meer verklaring, maar dienen verder
onderzocht te worden.
- Er werden geen verschillen waargenomen in de bodemfungi gemeenschap langsheen
een afstandsgradiënt tussen X. monsopora zaailingen hun dichtstbijzijnde
moederboom. Externe verstoringen, zoals habitatfragmentatie en de daarmee gepaard
gaande randeffecten, alsook temporele variatie in omgevingscondities kunnen
mogelijks verantwoordelijk zijn voor de afwezige variatie langsheen deze
afstandsgradiënt. Vanuit de zeer uiteenlopende literatuur betreffende de J-C
hypothese, kunnen we besluiten dat er geen eenduidig bewijs is voor een verhoogde
zaadkieming, zaailingrekrutering- en overleving verder weg van conspecifieke
moederbomen en dat deze waarneming sterk afhankelijk is van soort, ruimtelijke en
temporele schaal en context.
- De specifieke effecten van de geïsoleerde fungi morfotypes op de tropische boomsoort
X. monospora werden niet getest in de masterproef. Fusarium, Ilyonectria en
Cylindrocarpon werden beschouwd als potentieel pathogeen, terwijl Penicillium,
Trichoderma en Umbelopsis potentieel antagonistisch werden geacht. De interpretatie
van de resultaten is hierdoor eerder speculatief, maar kan een aanzet zijn tot verder
onderzoek via bio-essays om het effect van deze bodemfungi op X. monospora na te
gaan.

Samenvatting
51
7. Samenvatting
De afname in biologische diversiteit met toenemende breedtegraad is een van de meest
opmerkelijke patronen op onze planeet. Ondanks het feit dat men al tientallen jaren op zoek is
naar een universeel aanvaardbare verklaring voor deze latitudinale biodiversiteitsgradiënt,
bestaat er tot op heden nog geen eensgezindheid over de achterliggende mechanismen ervan.
Conspecifieke negatieve densiteits- of afstandsafhankelijke mortaliteit (CNDD) is één van de
mogelijke verklaringen voor de co-existentie van honderden plantensoorten per hectare
tropisch bos. CNDD wijst op een negatieve correlatie tussen de kans op overleving en de
conspecifieke (van dezelfde soort) zaailingdensiteit of afstand tussen conspecifieke zaailingen
en hun moederboom. Deze negatieve correlatie kan het gevolg zijn van een verhoogde
competitie of accumulatie van natuurlijke vijanden (herbivoren, pathogenen) in de buurt van
de moederboom. Zaden die, met behulp van (a)biotische dispersiemechanismen, verder weg
van de moederboom geraken, hebben dus een hogere kans tot overleven. De veronderstelling
dat CNDD te wijten is aan de accumulatie van gastheerspecifieke vijanden, zoals pathogene
bodemfungi, in de nabijheid van de moederboom vormt de basis van de Janzen-Connell (J-C)
hypothese, onafhankelijk beschreven door Janzen en Connell. Gastheerspecifieke pathogene
fungi die zaailingen van slechts één of enkele boomsoorten infecteren, veroorzaken een
verhoogde zaailingmortaliteit van die specifieke boomsoort. Hierdoor zien zaailingen van
andere boomsoorten hun kans te groeien op deze vrijgekomen plaatsen, resulterend in een
ensemble van zaailingen van verschillende boomsoorten. Hoe sterker de onderdrukking van
zaadkieming en zaailingrekrutering rondom de moederboom, hoe groter het aantal soorten dat
kan co-existeren in een boomgemeenschap. De soortensamenstelling en activiteit van
bodemorganismen kan variëren in tijd en ruimte en dit kan, ten gevolge van plant-bodem
terugkoppelingsmechanismen, uiteenlopende effecten hebben op de dynamiek van
bovengronds levende planten. Deze masterproef focuste zich op het ruimtelijk
distributiepatroon van pathogene bodemfungi in relatie tot de boomsoort Xymalos monsopora
en probeerde een antwoord te bieden op de vraag of de eerder waargenomen reductie in
zaadkieming, zaailingrekrutering en -overleving van X. monospora nabij conspecifieke
moederbomen (Lehouck et al., 2009) het gevolg is van een accumulatie aan
gastheerspecifieke bodempathogenen in de buurt van de moederboom.
Deze studie vond plaats in de bosfragmenten van de Taita Hills (zuidoost Kenia). Kleine en
sterk verstoorde fragmenten worden er gekenmerkt door de afwezigheid van enkele

Samenvatting
52
belangrijke zaadverbreiders, waardoor meer zaden blijven liggen onder de moederboom en
een ideale niche gecreëerd wordt voor fytopathogenen. De combinatie van een hoge
abundantie aan fytopathogenen en een gereduceerde zaaddispersie kan negatieve gevolgen
hebben voor de overleving van de bestudeerde boomsoort. Er werden in zes verschillende
bosfragmenten bodemstalen genomen in de rhizosfeer van X. monospora zaailingen,
aanwezig op verschillende afstanden ten opzichte van hun moederbomen. De bodemstalen
werden vervolgens uitgeplaat op agarmedia, selectief voor reeds gekende pathogene fungi
genera (Pythium, Phytophthora, Fusarium en Rhizoctonia), waarna het aantal gevormde
kolonies (CFU) per fungi morfotype werd geteld. Daarnaast werden enkele
omgevingsparameters opgemeten zodat een link kon gemaakt worden tussen de omgeving en
eventueel waargenomen verschillen in de ruimtelijke distributie van bodemfungi tussen de
fragmenten.
Er werden geen aanwijzingen gevonden voor de aanwezigheid van negatieve
afstandsafhankelijke zaailingmortaliteit ten gevolge van pathogene bodemfungi. De
bodemfungi gemeenschap vertoonde in geen enkel fragment variatie langsheen een
afstandsgradiënt tot de moederboom. Deze waarneming is mogelijks het gevolg van
seizoenale of interspecifieke variatie in J-C effecten, waardoor er sprake kan zijn van een
mismatch met ons moment van staalname. De bodemfungi gemeenschap vertoonde wel
variatie tussen de fragmenten onderling. Twee kleine en sterk verstoorde bosfragmenten
(Mwachora en Ndiwenyi) verschilden van de grote en minder sterk verstoorde bosfragmenten
(Chawia en Ngangao) in abundantie en samenstelling van pathogene fungi. In Mwachora en
Ndiwenyi werden het pathogene genus Fusarium aangetroffen, terwijl deze afwezig was in de
grote fragmenten. In deze laatsten werd dan weer het genus Ilyonectria teruggevonden.
Ilyonectria oefent pathogene effecten uit op druivelaars, maar het is helemaal niet zeker of
deze ook pathogeen is ten opzichte van tropische boomsoorten zoals X. monospora. De
combinatie van een gereduceerde zaaddispersie en de aanwezigheid van Fusarium in de
rhizosfeer van X. monospora zaailingen in kleine, sterk verstoorde fragmenten als Mwachora
en Ndiwenyi, kan negatieve gevolgen hebben voor de overleving van deze boomsoort.
Habitatfragmentatie en geassocieerde randeffecten lijken de meest voor de hand liggende
verklaring te zijn voor de waargenomen variatie, ware het niet dat de twee andere kleine
fragmenten (Fururu en Macha) meer gelijkenis vertonen met de grote fragmenten. Onze
opgemeten omgevingsparameters boden geen verklaring. Andere parameters, zoals de externe
verstoringen (bv. brand), klimatologische en topografische eigenschappen, bodemkenmerken

Samenvatting
53
(bv. pH, dikte van de strooisellaag) bieden mogelijks meer verklaring, maar dienen verder
onderzochte te worden.
Deze masterproef toonde duidelijke verschillen aan in bodemfungi gemeenschap op
fragmentniveau en kan een aanzet zijn tot verder onderzoek betreffende de achterliggende
mechanismen van deze waargenomen variatie. Aangezien deze studie de effecten van de
geïsoleerde bodemfungifungi op X. monospora niet getest heeft, moeten onze conclusies met
een korreltje zout genomen worden. Studies die de pathogeniciteit van deze bodemfungi
testen, worden sterk aangeraden.

Summary
54
8. Summary
The latitudinal biodiversity gradient is one of the most prominent patterns on earth. Finding a
universal explanation for the observed reduction in biodiversity with increasing latitude is a
big challenge among ecologists. Conspecific density or distance dependent mortality (CNDD)
is a possible mechanism to elucidate why hundreds of plant species can coexist on just one
hectare of tropical forest. CNDD refers to a negative correlation between seedling survival
and conspecific (of the same species) seedling density or distance between conspecific
seedlings and their mother trees. An increased competition and accumulation of natural
enemies (herbivores, pathogens) in the neighbourhood of a mother tree is assumed to be the
cause of this negative relationship. Seeds that, with the assistance of (a)biotic dispersal
mechanisms, are distributed away from the mother tree, have a higher chance to survive. The
Janzen-Connell (J-C) hypothesis, independently described by Janzen en Connell, postulates
that CNDD is driven by the accumulation of host specific pathogens (e.g. soil fungi) near the
mother tree. Host specific pathogenic soil fungi can infect seedlings belonging to just one or a
few tree species, thereby lowering the chance of seedling survival of that particular tree-
species. Seedlings of other species can grow in these free spaces, resulting in a coexistence of
seedlings of different species. Therefore we can assume that the amount of coexisting species
rises with the increasing repression of seed germination, seedling recruitment and survival.
The spatial and temporal variation in species composition and activity of soil organisms can
affect the natural dynamics of plant communities, and this can be subscribed to the
interactions between aboveground and belowground communities. The spatial variation of
pathogenic soil fungi related to the tropical tree-species Xymalos monospora, is the focus of
this thesis. The goal was to explain the observed reduction in seed germination, seedling
recruitment and survival of X. monospora near conspecific mother trees by Lehouck et al.
(2009) who assumed that this pattern is caused by the accumulation of host specific,
pathogenic soil fungi near the mother tree.
This study took place in the forest fragments of the Taita Hills (SE Kenya). Small and
strongly disturbed fragments are characterized by the absence of some important seed
dispersers. A lower amount of seeds is taken away from the mother tree, creating an ideal
niche for plant pathogens, such as pathogenic soil fungi. The combination of a high
abundance of these fungi and reduced seed dispersal can have negative consequence for the
survival of X. monospora. Soil samples were taken in the rhizosphere of X. monospora

Summary
55
seedlings, present on different distances upon the mother trees, and this in six different
fragments. The soil samples were plated on an agar media, selective against known
pathogenic fungi (Pythium, Phytophthora, Fusarium and Rhizoctonia), the amount of colonies
(CFU) of the different morphotypes were counted and identified. Furthermore, measurements
of some environmental variables were taken to make a link between the environment and
potential differences in spatial distribution patterns of soil fungi.
We found no evidence to proof the J-C hypothesis. The soil fungi community showed no
variation along a distance gradient away from the mother tree. A possible explanation is the
seasonal or interspecific variation in J-C effects, which can create a mismatch with the
moment of sampling. Besides that, this study showed a strong variation in soil fungi
abundance and composition between the fragments. Two small and strongly disturbed
fragments (Mwachora and Ndiwenyi) showed a difference in pathogenic fungi composition
with the two major fragments (Chawia and Ngangao). The genus Fusarium, known to be
pathogenic against different tropical tree species, was present in Mwachora and Ndiwenyi and
abscent in the big fragments. Instead, Chawia and Ngangao harboured the potential
pathogenic genus Ilyonectria. This genus has been recently identified as pathogenic upon
grapevines, but the effect against tropical tree species is not known yet. The lowered seed
dispersal of X. monospora in small fragments, together with the presence of Fusarium in the
rhizosphere of seedlings in these fragments can have strong negative effects on the survival of
this tree-species. Habitat fragmentation together with edge effects seemed to be the best
explanation, although the two other small fragments (Fururu and Macha) showed more
resemblance with Chawia and Ngangao. In conclusion, other, non-measured factors, such as
the degree of disturbance (e.g. forest fire), soil characteristics (e.g. pH) will explain more of
the observed variation between the fragments.
Further research can give more insight in the factors causing the variation in soil fungi
community among the forest fragments of the Taita Hills. This study didn’t test the effects of
the isolated fungi on X. monospora seedlings, so our conclusions are rather speculative.
Studies that take into account these tests are strongly recommended.

Referenties
56
9. Referenties
Aerts, R., Thijs, K.W., Lehouck, V., Beentje, H., Bytebier, B., Matthysen, E. et al. (2011). Woody plant
communities of isolated Afromontane cloud forests in Taita Hills, Kenya. Plant Ecology, 212,
639-649.
Augspurger, C.K. (1983). Seed dispersal of the tropical tree, Platypodium elegans and the escape of its
seedlings from fungal pathogens. Journal of Ecology, 71, 759-771.
Augspurger, C.K. & Wilkinson, H.T. (2007). Host specificity of pathogenic Pythium species: Implications
for tree species diversity. Biotropica, 39, 702-708.
Bardgett, R.D., Wardle, D.A. & Yeates, G.W. (1998). Linking above-ground and below-ground
interactions: How plant responses to foliar herbivory influence soil organisms. Soil Biology &
Biochemistry, 30, 1867-1878.
Beentje, H. (1988). An ecological and floristic study of the forests of the Taita Hills, Kenya. Utafiti, 1, 23-
66.
Bell, T., Freckleton, R.P. & Lewis, O.T. (2006). Plant pathogens drive density-dependent seedling mortality
in a tropical tree. Ecology Letters, 9, 569-574.
Bender, D.J., Contreras, T.A. & Fahrig, L. (1998). Habitat loss and population decline: A meta-analysis of
the patch size effect. Ecology, 79, 517-533.
Cabral, A., Rego, C., Nascimento, T., Oliveira, H., Groenewald, J.Z. & Crous, P.W. (2012). Multi-gene
analysis and morphology reveal novel Ilyonectria species associated with black foot disease of
grapevines. Fungal Biology, 116, 62-80.
Chen, J.Q., Franklin, J.F. & Spies, T.A. (1993). An empirical model for predicting diurnal air-temperature
gradients from edge into old-growth Douglas-fir forest. Ecological Modelling, 67, 179-198.
Colwell, R. K. (2009). EstimateS: statistical estimation of species richness and shared species from
samples. Version 9.0 (http://viceroy.eeb.uconn.edu/estimates) (10/05/2013).
Comita, L.S., Muller-Landau, H.C., Aguilar, S. & Hubbell, S.P. (2010). Asymmetric Density Dependence
Shapes Species Abundances in a Tropical Tree Community. Science, 329, 330-332.
Connell, J. H. (1971) On the role of natural enemies in preventing competitive exclusion in some marine
animals and in rain forest trees. Dynamics of Population, 298-312.
Cordeiro, N.J. & Howe, H.F. (2003). Forest fragmentation severs mutualism between seed dispersers and
an endemic African tree. Proceedings of the National Academy of Sciences of the United States of
America, 100, 14052-14056.
Eisenhauer, N. (2012). Aboveground-belowground interactions as a source of complementarity effects in
biodiversity experiments. Plant and Soil, 351, 1-22.
Fahrig, L. (2003). Effects of habitat fragmentation on biodiversity. Annual Review of Ecology Evolution
and Systematics, 34, 487-515.
Freckleton, R.P. & Lewis, O.T. (2006). Pathogens, density dependence and the coexistence of tropical trees.
Proceedings of the Royal Society B-Biological Sciences, 273, 2909-2916.

Referenties
57
Gilbert, G.S. (2002). Evolutionary ecology of plant diseases in natural ecosystems. Annual Review of
Phytopathology, 40, 13-43.
Giller, P.S. (1996). The diversity of soil communities, the 'poor man's tropical rainforest'. Biodiversity and
Conservation, 5, 135-168.
Harman, G.E. (2006). Overview of mechanisms and uses of Trichoderma spp. Phytopathology, 96, 190-
194.
Hatcher, M.J., Dick, J.T.A. & Dunn, A.M. (2006). How parasites affect interactions between competitors
and predators. Ecology Letters, 9, 1253-1271.
Hector, A. & Bagchi, R. (2007). Biodiversity and ecosystem multifunctionality. Nature, 448, 188-U186.
Hill, J.L. & Curran, P.J. (2001). Species composition in fragmented forests: conservation implications of
changing forest area. Applied Geography, 21, 157-174.
Hillebrand, H. (2004). On the generality of the latitudinal diversity gradient. American Naturalist, 163, 192-
211.
Hoff, J.A., Klopfenstein, N.B., McDonald, G.I., Tonn, J.R., Kim, M.S., Zambino, P.J. et al. (2004). Fungal
endophytes in woody roots of Douglas-fir (Pseudotsuga menziesii) and ponderosa pine (Pinus
ponderosa). Forest Pathology, 34, 255-271.
Hooper, D.U., Chapin, F.S., Ewel, J.J., Hector, A., Inchausti, P., Lavorel, S. et al. (2005). Effects of
biodiversity on ecosystem functioning: A consensus of current knowledge. Ecological
Monographs, 75, 3-35.
Howe, H.F. & Smallwood, J. (1982). Ecology of seed dispersal. Annual Review of Ecology and Systematics,
13, 201-228.
Hubbell, S.P. (1980). Seed predation and the coexistence of tree species in tropical forests. Oikos, 35, 214-
229.
Hyatt, L.A., Rosenberg, M.S., Howard, T.G., Bole, G., Fang, W., Anastasia, J. et al. (2003). The distance
dependence prediction of the Janzen-Connell hypothesis: a meta-analysis. Oikos, 103, 590-602.
Ingle, R.A., Carstens, M. & Denby, K.J. (2006). PAMP recognition and the plant-pathogen arms race.
Bioessays, 28, 880-889.
Janzen, D.H. (1970). Herbivores and the number of tree species in tropical forests. American Naturalist,
104, 501.
Jeffers, S.N. & Martin, S.B. (1986). Comparison of two selective media selective for Phytophthora and
Pythium. Plant Disease, 70, 1030-1043.
John, R.P., Tyagi, R.D., Prevost, D., Brar, S.K., Pouleur, S. & Surampalli, R.Y. (2010). Mycoparasitic
Trichoderma viride as a biocontrol agent against Fusarium oxysporum f. sp adzuki and Pythium
arrhenomanes and as a growth promoter of soybean. Crop Protection, 29, 1452-1459.
Johnson, D.J., Beaulieu, W.T., Bever, J.D. & Clay, K. (2012). Conspecific Negative Density Dependence
and Forest Diversity. Science, 336, 904-907.
Jost, L. (2006). Entropy and diversity. Oikos, 113, 363-375.

Referenties
58
Klironomos, J.N. (2002). Feedback with soil biota contributes to plant rarity and invasiveness in
communities. Nature, 417, 67-70.
Ko, W. & Hora, F.K. (1971). A selective medium for the quantitative determination of Rhizoctonia solani
in soil. Phytopathology, 61, 707-710.
Koh, L.P., Dunn, R.R., Sodhi, N.S., Colwell, R.K., Proctor, H.C. & Smith, V.S. (2004). Species
coextinctions and the biodiversity crisis. Science, 305, 1632-1634.
Komada, H. (1975). Development of a selective medium for quantitative isolation of Fusarium oxysporum
from natural soils. Plant Protection Research, 8, 114-125.
Kourtev, P.S., Ehrenfeld, J.G. & Haggblom, M. (2002). Exotic plant species alter the microbial community
structure and function in the soil. Ecology, 83, 3152-3166.
Kruess, A. & Tscharntke, T. (1994). Habitat fragmentation, species loss and biological control. Science,
264, 1581-1584.
Lehouck, V., Spanhove, T., Colson, L., Adringa-Davis, A., Cordeiro, N.J. & Lens, L. (2009a). Habitat
disturbance reduces seed dispersal of a forest interior tree in a fragmented African cloud forest.
Oikos, 118, 1023-1034.
Lehouck, V., Spanhove, T., Demeter, S., Groot, N.E. & Lens, L. (2009b). Complementary seed dispersal by
three avian frugivores in a fragmented Afromontane forest. Journal of Vegetation Science, 20,
1110-1120.
Lehouck, V., Spanhove, T., Gonsamo, A., Cordeiro, N. & Lens, L. (2009c). Spatial and temporal effects on
recruitment of an Afromontane forest tree in a threatened fragmented ecosystem. Biological
Conservation, 142, 518-528.
Lehouck, V., Spanhove, T. & Lens, L. (2011). Avian fruit ingestion differentially facilitates seed
germination of four fleshy-fruited plant species of an Afrotropical forest. Plant Ecology and
Evolution, 144, 96-100.
Leigh, E.G., Davidar, P., Dick, C.W., Puyravaud, J.P., Terborgh, J., ter Steege, H. et al. (2004). Why do
some tropical forests have so many species of trees? Biotropica, 36, 447-473.
Lewis, O.T. (2010). Ecology: close relatives are bad news. Nature, 466, 698-699.
Lin, L.X., Comita, L.S., Zheng, Z. & Cao, M. (2012). Seasonal differentiation in density-dependent
seedling survival in a tropical rain forest. Journal of Ecology, 100, 905-914.
Loreau, M. (2010). Linking biodiversity and ecosystems: towards a unifying ecological theory.
Philosophical Transactions of the Royal Society B-Biological Sciences, 365, 49-60.
Lovett, J.C. (1996). Elevational and latitudinal changes in tree associations and diversity in the Eastern Arc
mountains of Tanzania. Journal of Tropical Ecology, 12, 629-650.
Machon, P., Pajares, J.A., Diez, J.J. & Alves-Santos, F.M. (2009). Influence of the ectomycorrhizal fungus
Laccaria laccata on pre-emergence, post-emergence and late damping-off by Fusarium oxysporum
and F. verticillioides on Stone pine seedlings. Symbiosis, 49, 101-109.
Malmivaara-Lamsa, M., Hamberg, L., Haapamaki, E., Liski, J., Kotze, D.J., Lehvavirta, S. et al. (2008).
Edge effects and trampling in boreal urban forest fragments - impacts on the soil microbial
community. Soil Biology & Biochemistry, 40, 1612-1621.

Referenties
59
Mangan, S.A., Schnitzer, S.A., Herre, E.A., Mack, K.M.L., Valencia, M.C., Sanchez, E.I. et al. (2010).
Negative plant-soil feedback predicts tree-species relative abundance in a tropical forest. Nature,
466, 752-U710.
Mantyka-Pringle, C.S., Martin, T.G. & Rhodes, J.R. (2012). Interactions between climate and habitat loss
effects on biodiversity: a systematic review and meta-analysis. Global Change Biology, 18, 1239-
1252.
Maor, R. & Shirasu, K. (2005). The arms race continues: battle strategies between plants and fungal
pathogens. Current Opinion in Microbiology, 8, 399-404.
Matthesius A. (2006) Testing the Janzen-Connell model for species diversity in a west African montane
forest. Department of Plant Ecology, Canterbury University, 1-131.
McCallum, H. & Dobson, A. (2002). Disease, habitat fragmentation and conservation. Proceedings of the
Royal Society B-Biological Sciences, 269, 2041-2049.
McMullan-Fisher, S.J.M., May, T.W., Robinson, R.M., Bell, T.L., Lebel, T., Catcheside, P. et al. (2011).
Fungi and fire in Australian ecosystems: a review of current knowledge, management implications
and future directions. Australian Journal of Botany, 59, 70-90.
Meinzer, F.C., Andrade, J.L., Goldstein, G., Holbrook, N.M., Cavelier, J. & Wright, S.J. (1999).
Partitioning of soil water among canopy trees in a seasonally dry tropical forest. Oecologia, 121,
293-301.
Meyer, W. & Gams, W. (2003). Delimitation of Umbelopsis (Mucorales, Umbelopsidaceae fam. nov.)
based on ITS sequence and RFLP data. Mycological Research, 107, 339-350.
Mittelbach, G.G., Schemske, D.W., Cornell, H.V., Allen, A.P., Brown, J.M., Bush, M.B. et al. (2007).
Evolution and the latitudinal diversity gradient: speciation, extinction and biogeography. Ecology
Letters, 10, 315-331.
Mordecai, E.A. (2011). Pathogen impacts on plant communities: unifying theory, concepts, and empirical
work. Ecological Monographs, 81, 429-441.
Mori, A.S., Furukawa, T. & Sasaki, T. (2013). Response diversity determines the resilience of ecosystems
to environmental change. Biological Reviews, 88, 349-364.
Morris, R.J. (2010). Anthropogenic impacts on tropical forest biodiversity: a network structure and
ecosystem functioning perspective. Philosophical Transactions of the Royal Society B-Biological
Sciences, 365, 3709-3718.
Myers, N., Mittermeier, R.A., Mittermeier, C.G., da Fonseca, G.A.B. & Kent, J. (2000). Biodiversity
hotspots for conservation priorities. Nature, 403, 853-858.
Nathan, R. & Casagrandi, R. (2004). A simple mechanistic model of seed dispersal, predation and plant
establishment: Janzen-Connell and beyond. Journal of Ecology, 92, 733-746.
Nicoletti, R., De Stefano, M., De Stefano, S., Trincone, A. & Marziano, F. (2004). Antagonism against
Rhizoctonia solani and fungitoxic metabolite production by some Penicillium isolates.
Mycopathologia, 158, 465-474.
Packer, A. & Clay, K. (2000). Soil pathogens and spatial patterns of seedling mortality in a temperate tree.
Nature, 404, 278-281.
Palmer, M.W. (1994). Variation in species richness – towards a unification of hypotheses. Folia
Geobotanica & Phytotaxonomica, 29, 511-530.

Referenties
60
Pellikka, P.K.E., Lotjonen, M., Sijander, M. & Lens, L. (2009). Airborne remote sensing of spatiotemporal
change (1955-2004) in indigenous and exotic forest cover in the Taita Hills, Kenya. International
Journal of Applied Earth Observation and Geoinformation, 11, 221-232.
Reaves, J.L., Shaw, C.G. & Mayfield, J.E. (1990). The effects of Trichoderma spp isolated from burned and
nonburned forest soils on the growth and development of Armillaria ostoyae in culture. Northwest
Science, 64, 39-44.
Schemske, D.W., Mittelbach, G.G., Cornell, H.V., Sobel, J.M. & Roy, K. (2009). Is There a Latitudinal
Gradient in the Importance of Biotic Interactions? Annual Review of Ecology Evolution and
Systematics, 40, 245-269.
Schoch, C.L., Seifert, K.A., Huhndorf, S., Robert, V., Spouge, J.L., Levesque, C.A. et al. (2012). Nuclear
ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi.
Proceedings of the National Academy of Sciences of the United States of America, 109, 6241-
6246.
Schupp, E.W. (1992). The Janzen-Connell model for tropical tree diversity – population implications and
the importance of special scale. American Naturalist, 140, 526-530.
Sharma, G.D. (1981). Effect of fire on soil-microorganisms in a Meghalaya pine forest. Folia
Microbiologica, 26, 321-327.
Stendell, E.R., Horton, T.R. & Bruns, T.D. (1999). Early effects of prescribed fire on the structure of the
ectomycorrhizal fungus community in a Sierra Nevada ponderosa pine forest. Mycological
Research, 103, 1353-1359.
Swamy, V. (2008). Fruit to sapling: an ontogenetically integrated study of tree recruitment in an Amazonian
rainforest. Department of Environment, Duke University, 1-172.
Terborgh, J. (2012). Enemies Maintain Hyperdiverse Tropical Forests. American Naturalist, 179, 303-314.
Terborgh, J., Nunez-Iturri, G., Pitman, N.C.A., Valverde, F.H.C., Alvarez, P., Swamy, V. et al. (2008). Tree
recruitment in an empty forest. Ecology, 89, 1757-1768.
Thompson, S., Alvarez-Loayza, P., Terborgh, J. & Katul, G. (2010). The effects of plant pathogens on tree
recruitment in the Western Amazon under a projected future climate: a dynamical systems
analysis. Journal of Ecology, 98, 1434-1446.
Tylianakis, J.M., Didham, R.K., Bascompte, J. & Wardle, D.A. (2008). Global change and species
interactions in terrestrial ecosystems. Ecology Letters, 11, 1351-1363.
Urbez-Torres, J.R., Peduto, F. & Gubler, W.D. (2012). First Report of Ilyonectria macrodidyma Causing
Root Rot of Olive Trees (Olea europaea) in California. Plant Disease, 96, 1378-1378.
van der Heijden, M.G.A., Bardgett, R.D. & van Straalen, N.M. (2008). The unseen majority: soil microbes
as drivers of plant diversity and productivity in terrestrial ecosystems. Ecology Letters, 11, 296-
310.
van der Putten, W.H., Bardgett, R.D., Bever, J.D., Bezemer, T.M., Casper, B.B., Fukami, T. et al. (2013).
Plant-soil feedbacks: the past, the present and future challenges. Journal of Ecology, 101, 265-276.
van der Putten, W.H., Bardgett, R.D., de Ruiter, P.C., Hol, W.H.G., Meyer, K.M., Bezemer, T.M. et al.
(2009). Empirical and theoretical challenges in aboveground-belowground ecology. Oecologia,
161, 1-14.

Referenties
61
Vitale, A., Aiello, D., Guarnaccia, V., Perrone, G., Stea, G. & Polizzi, G. (2012). First Report of Root Rot
Caused by Ilyonectria (=Neonectria) macrodidyma on Avocado (Persea americana) in Italy.
Journal of Phytopathology, 160, 156-159.
Vogeli, M., Lemus, J.A., Serrano, D., Blanco, G. & Tella, J.L. (2011). An island paradigm on the mainland:
host population fragmentation impairs the community of avian pathogens. Proceedings of the
Royal Society B-Biological Sciences, 278, 2668-2676.
Wardle, D.A., Bardgett, R.D., Klironomos, J.N., Setala, H., van der Putten, W.H. & Wall, D.H. (2004).
Ecological linkages between aboveground and belowground biota. Science, 304, 1629-1633.
Willig, M.R., Kaufman, D.M. & Stevens, R.D. (2003). Latitudinal gradients of biodiversity: Pattern,
process, scale, and synthesis. Annual Review of Ecology Evolution and Systematics, 34, 273-309.
Wright, S.J. (2002). Plant diversity in tropical forests: a review of mechanisms of species coexistence.
Oecologia, 130, 1-14.
Yamaji, K., Fukushi, Y., Hashidoko, Y. & Tahara, S. (2005). Penicillium frequentans isolated from Picea
glehnii seedling roots as a possible biological control agent against damping-off. Ecological
Research, 20, 103-107.
Yamaji, K., Fukushi, Y., Hashidoko, Y., Yoshida, T. & Tahara, S. (2001). Penicillium fungi from Picea
glehnii seeds protect the seedlings from damping-off. New Phytologist, 152, 521-531.
Young, A., Boyle, T. & Brown, T. (1996). The population genetic consequences of habitat fragmentation
for plants. Trends in Ecology & Evolution, 11, 413-418.

Bijlagen
62
10. Bijlagen
Bijlage 1
Resultaten van de nearest neighbour analyse (ArcGIS), waarbij het ruimtelijk
distributiepatroon van adulte X. monospora bomen werd nagegaan op plotniveau. Indien de
nearest neighbour ratio (R-waarde) significant verschillend is van 1 (p < 0.05) is het
distributiepatroon van de X. monospora adulten binnen eenzelfde plot niet random, maar
verspreid (R > 1) of geaggregeerd (R < 1).
Plot R-waarde p-waarde Distributiepatroon
Cha1 1.32 0.02 Verspreid
Cha2 1.46 0.00 Verspreid
Cha3 2.78 0.00 Verspreid
Cha4 2.09 0.00 Verspreid
Cha5 2.70 0.00 Verspreid
Cha6 2.30 0.00 Verspreid
Fur1 2.15 0.00 Verspreid
Fur2 2.74 0.00 Verspreid
Fur3 1.71 0.00 Verspreid
Fur4 1.35 0.25 Random
Mac1 1.93 0.00 Verspreid
Mac2 1.58 0.01 Verspreid
Mac3 1.14 0.39 Random
Mwa1 1.65 0.0 Verspreid
Mwa2 2.70 0.00 Verspreid
Mwa3 1.70 0.06 Random
Ndi1 1.18 0.49 Random
Ndi2 1.01 0.98 Random
Ndi3 1.38 0.08 Random
Nga1 1.61 0.00 Verspreid
Nga2 1.94 0.01 Verspreid
Nga3 2.20 0.00 Verspreid
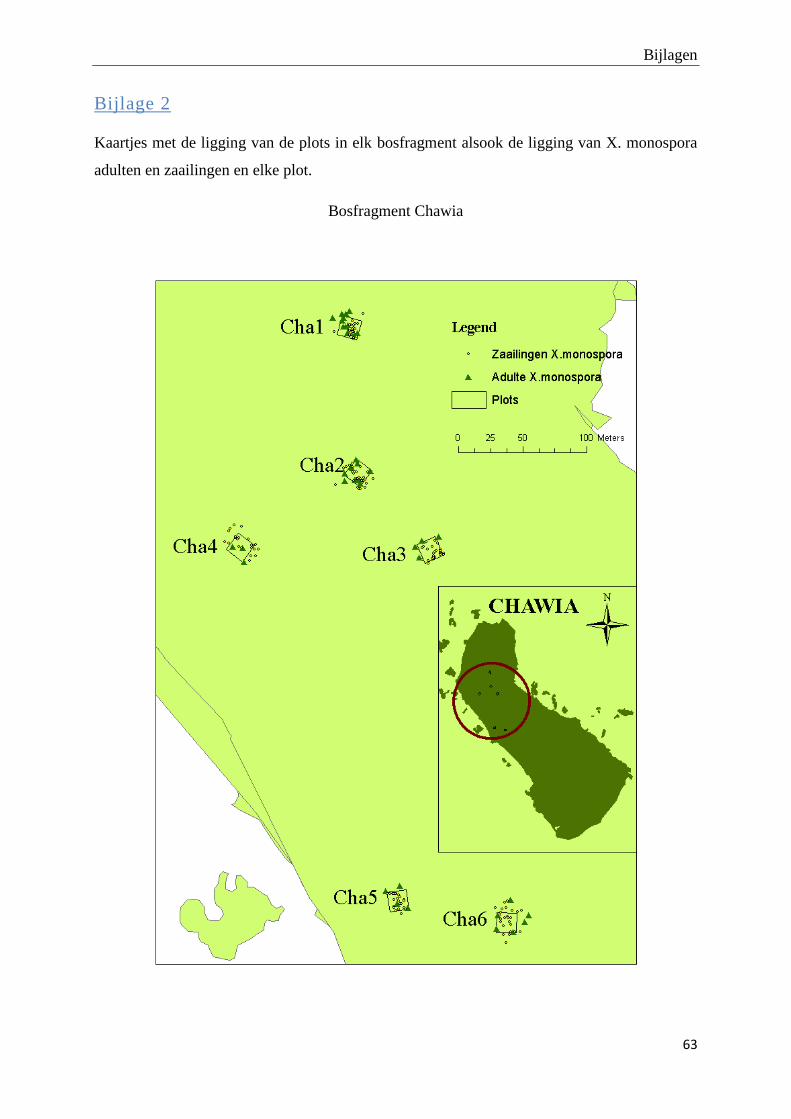
Bijlagen
63
Bijlage 2
Kaartjes met de ligging van de plots in elk bosfragment alsook de ligging van X. monospora
adulten en zaailingen en elke plot.
Bosfragment Chawia

Bijlagen
64
Bosfragment Fururu
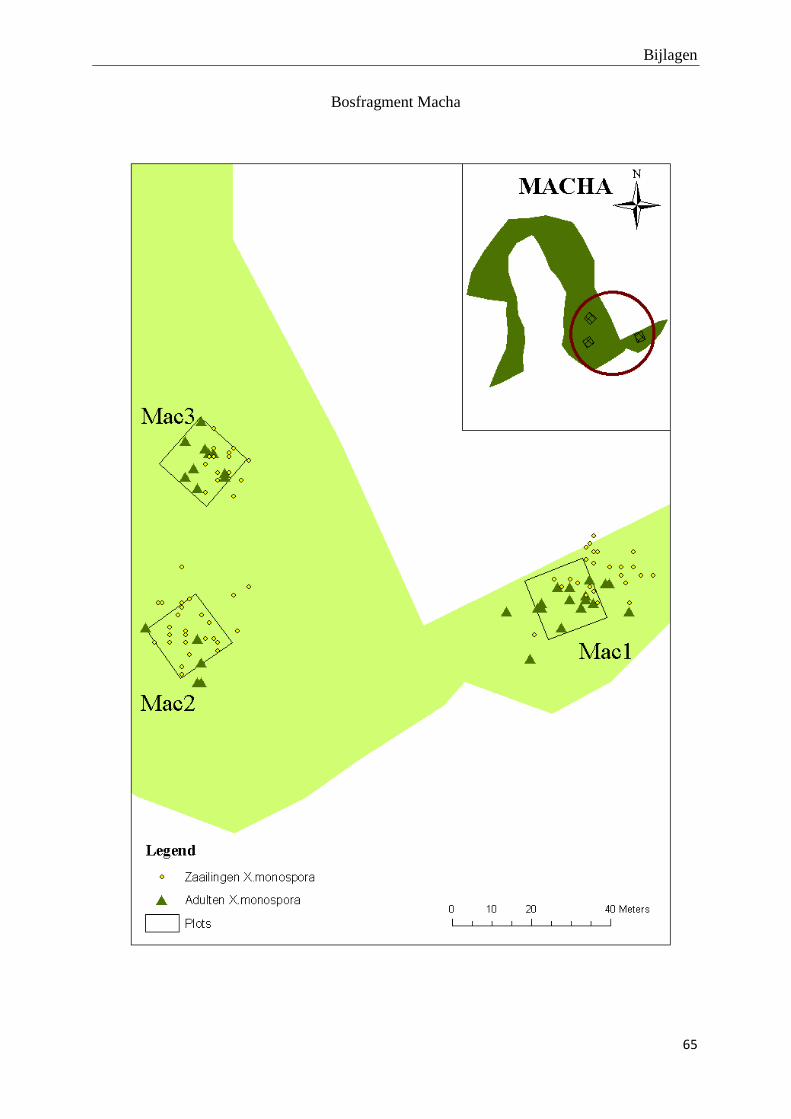
Bijlagen
65
Bosfragment Macha

Bijlagen
66
Bosfragment Mwachora
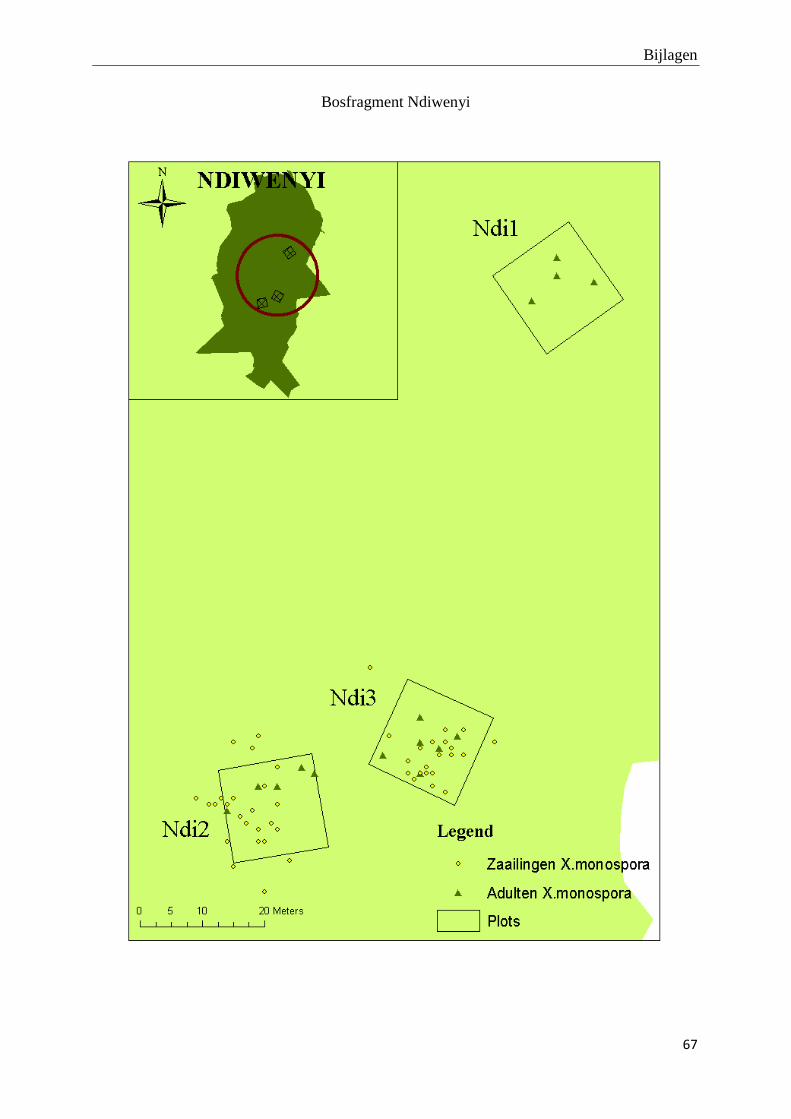
Bijlagen
67
Bosfragment Ndiwenyi
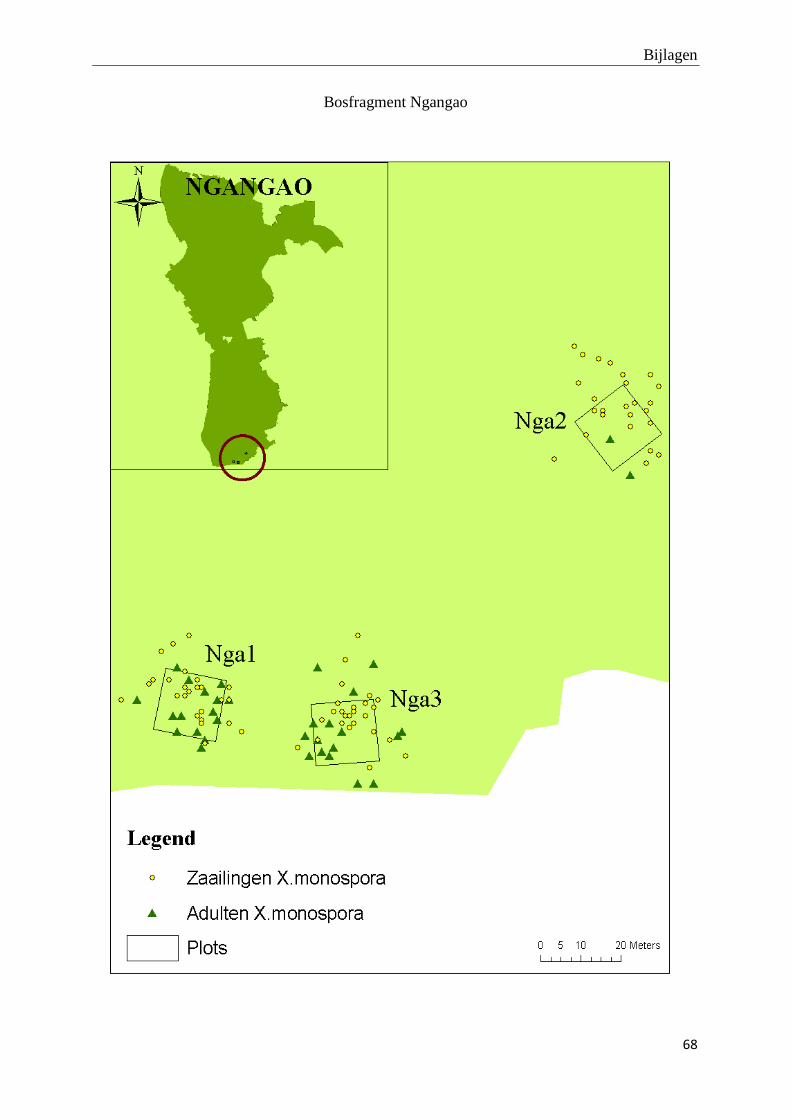
Bijlagen
68
Bosfragment Ngangao
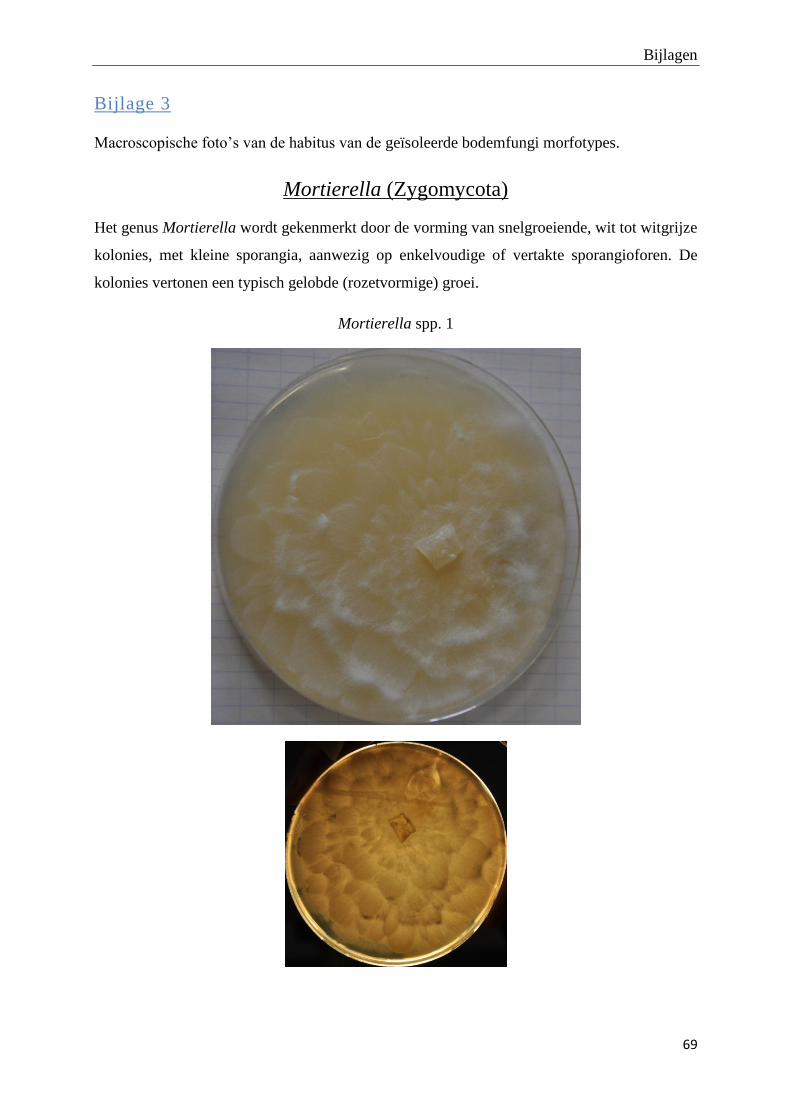
Bijlagen
69
Bijlage 3
Macroscopische foto’s van de habitus van de geïsoleerde bodemfungi morfotypes.
Mortierella (Zygomycota)
Het genus Mortierella wordt gekenmerkt door de vorming van snelgroeiende, wit tot witgrijze
kolonies, met kleine sporangia, aanwezig op enkelvoudige of vertakte sporangioforen. De
kolonies vertonen een typisch gelobde (rozetvormige) groei.
Mortierella spp. 1
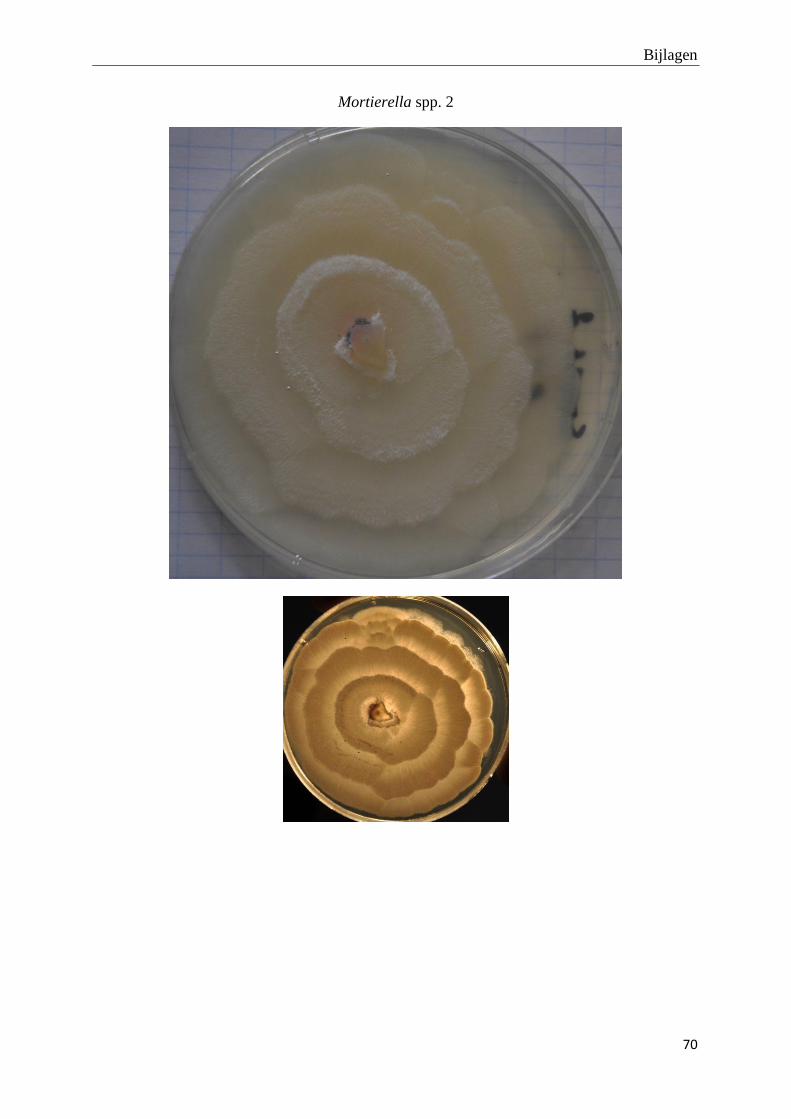
Bijlagen
70
Mortierella spp. 2
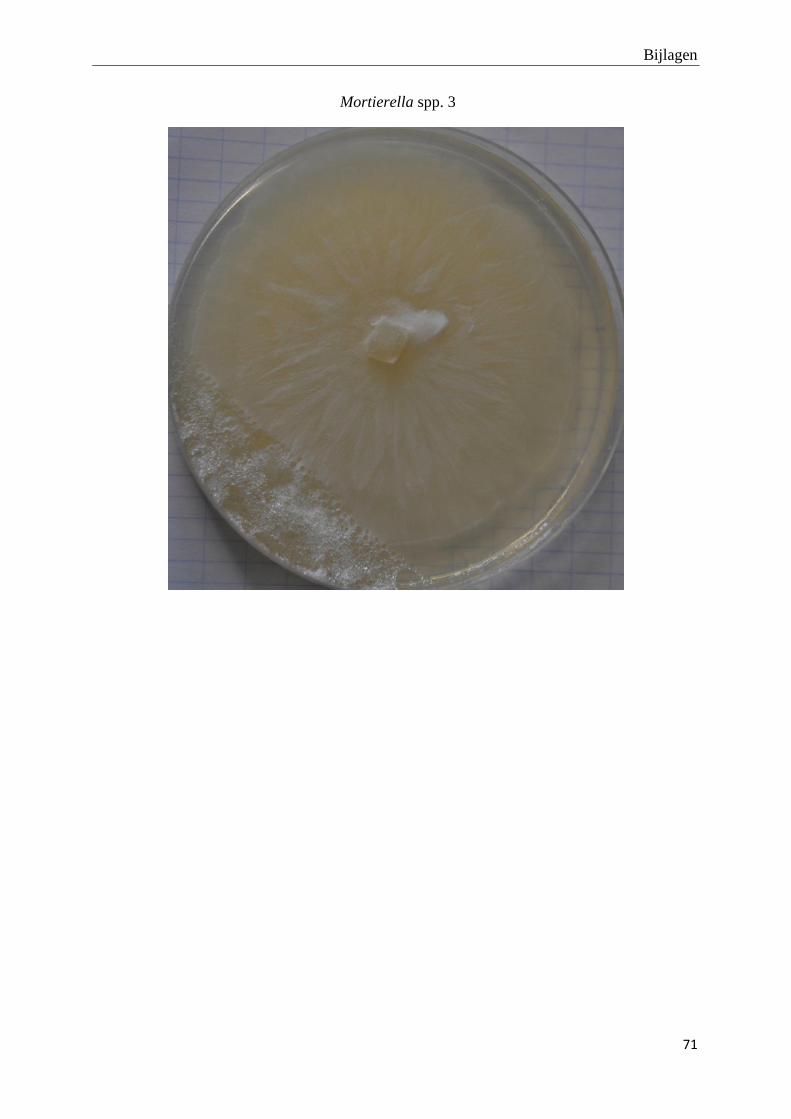
Bijlagen
71
Mortierella spp. 3
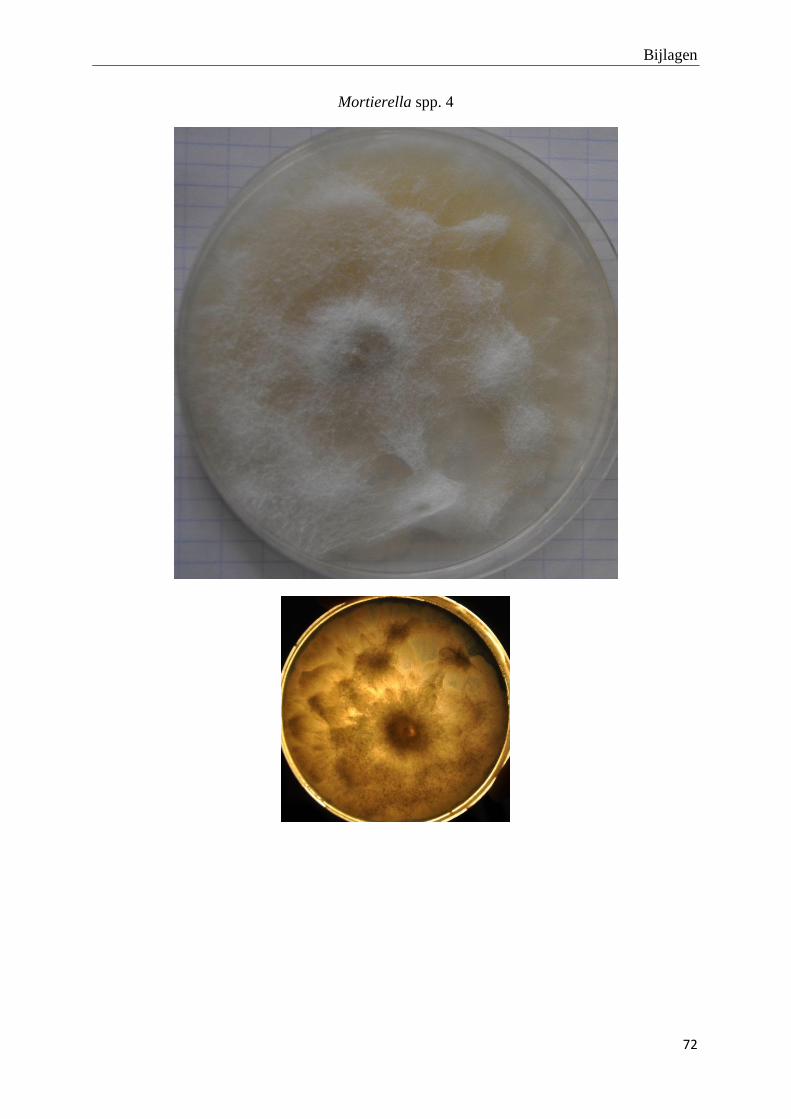
Bijlagen
72
Mortierella spp. 4

Bijlagen
73
Mortierella spp. 5

Bijlagen
74
Mucor (Zygomycota)
Het genus Mucor wordt gekenmerkt door de vorming van snelgroeiende, witachtig tot
geelgekleurde kolonies met sferische, zwartgrijze sporangia, aanwezig op opgerichte,
enkelvoudige of vertakte sporangioforen en goed ontwikkelde columnella.
Mucor fragilis
De globulaire sporangia, aanwezig op opgerichte sporangioforen (vergroting 5x).
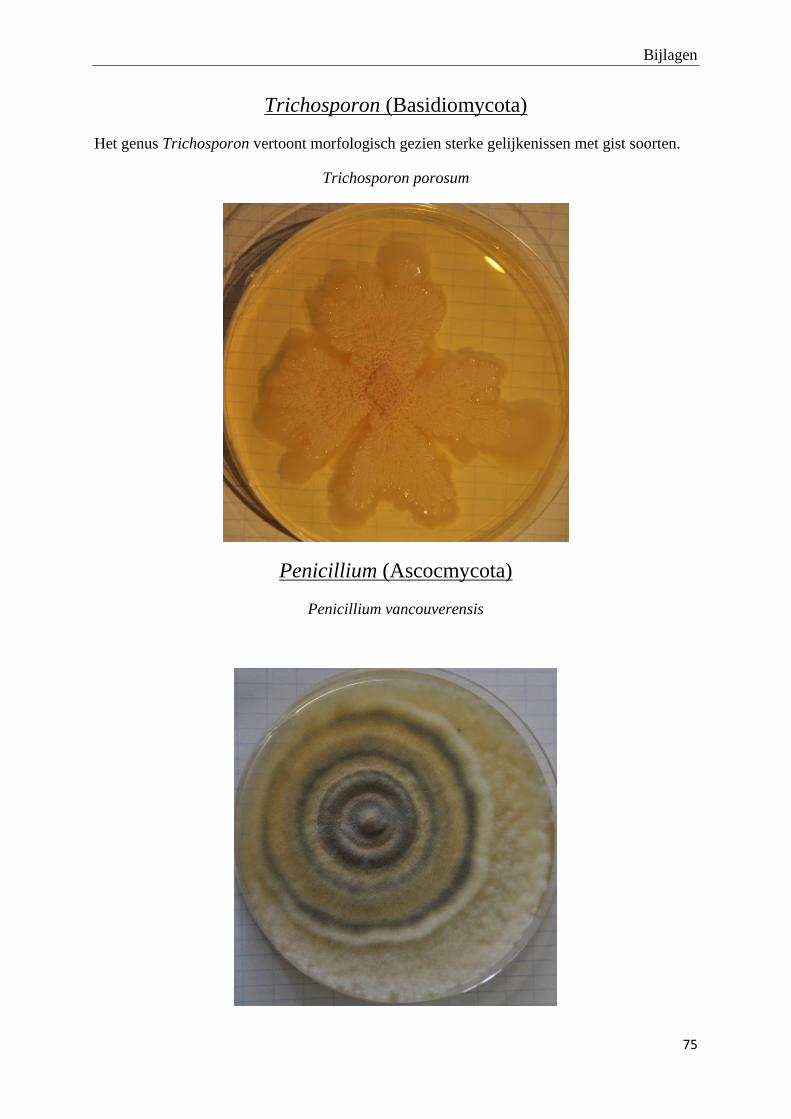
Bijlagen
75
Trichosporon (Basidiomycota)
Het genus Trichosporon vertoont morfologisch gezien sterke gelijkenissen met gist soorten.
Trichosporon porosum
Penicillium (Ascocmycota)
Penicillium vancouverensis
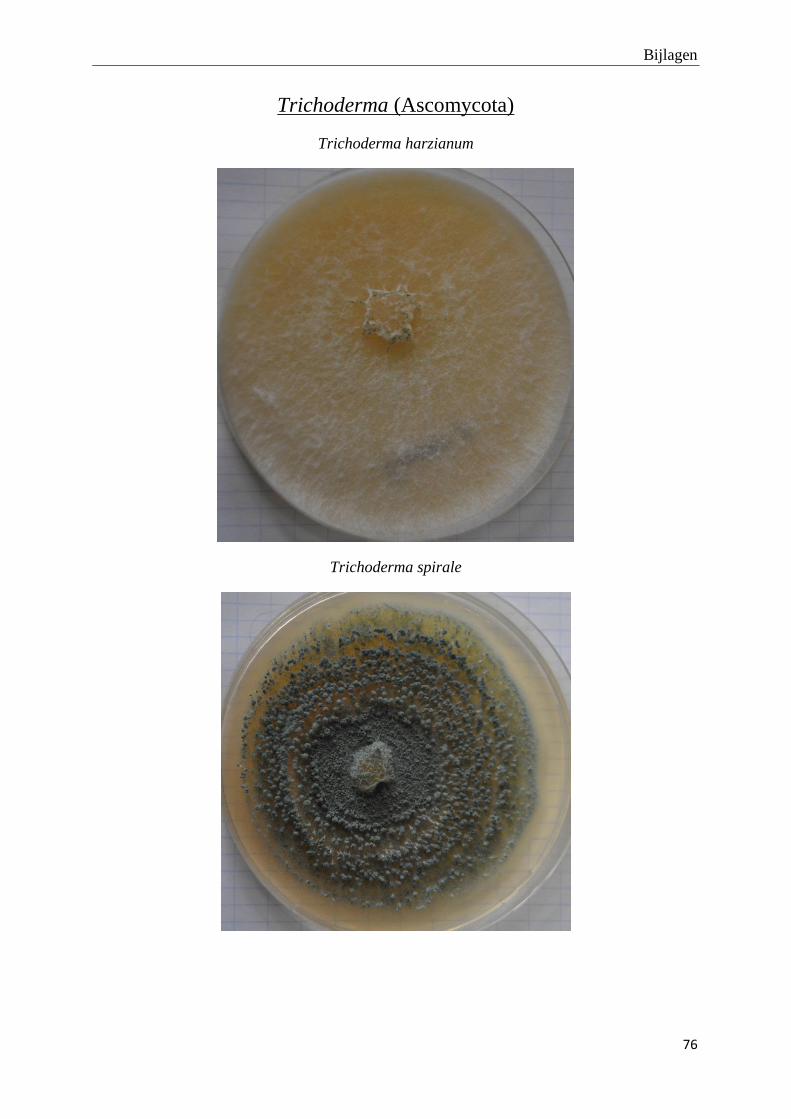
Bijlagen
76
Trichoderma (Ascomycota)
Trichoderma harzianum
Trichoderma spirale
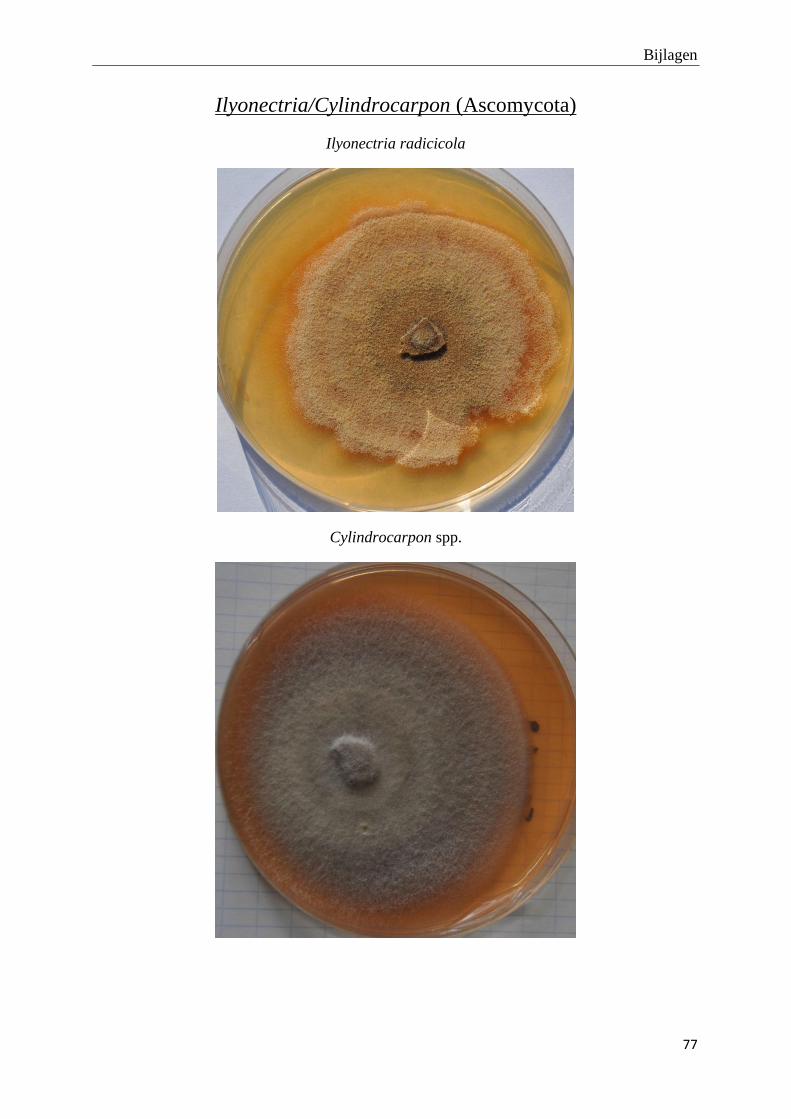
Bijlagen
77
Ilyonectria/Cylindrocarpon (Ascomycota)
Ilyonectria radicicola
Cylindrocarpon spp.
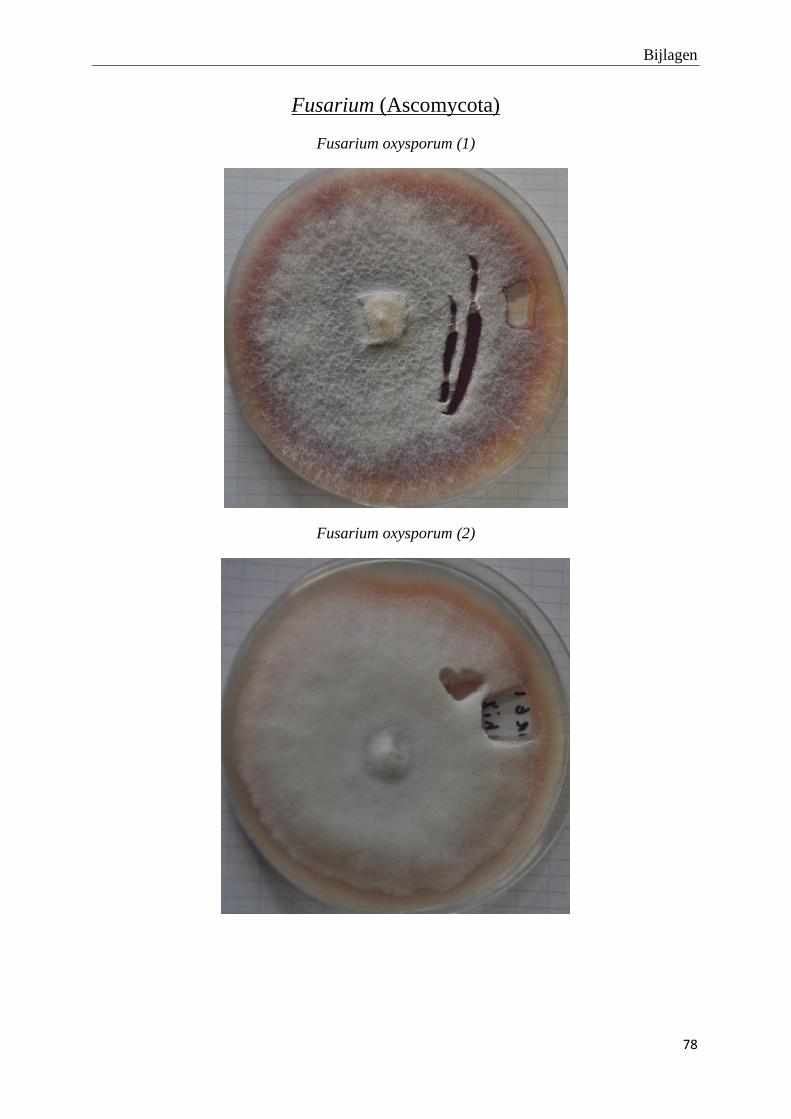
Bijlagen
78
Fusarium (Ascomycota)
Fusarium oxysporum (1)
Fusarium oxysporum (2)
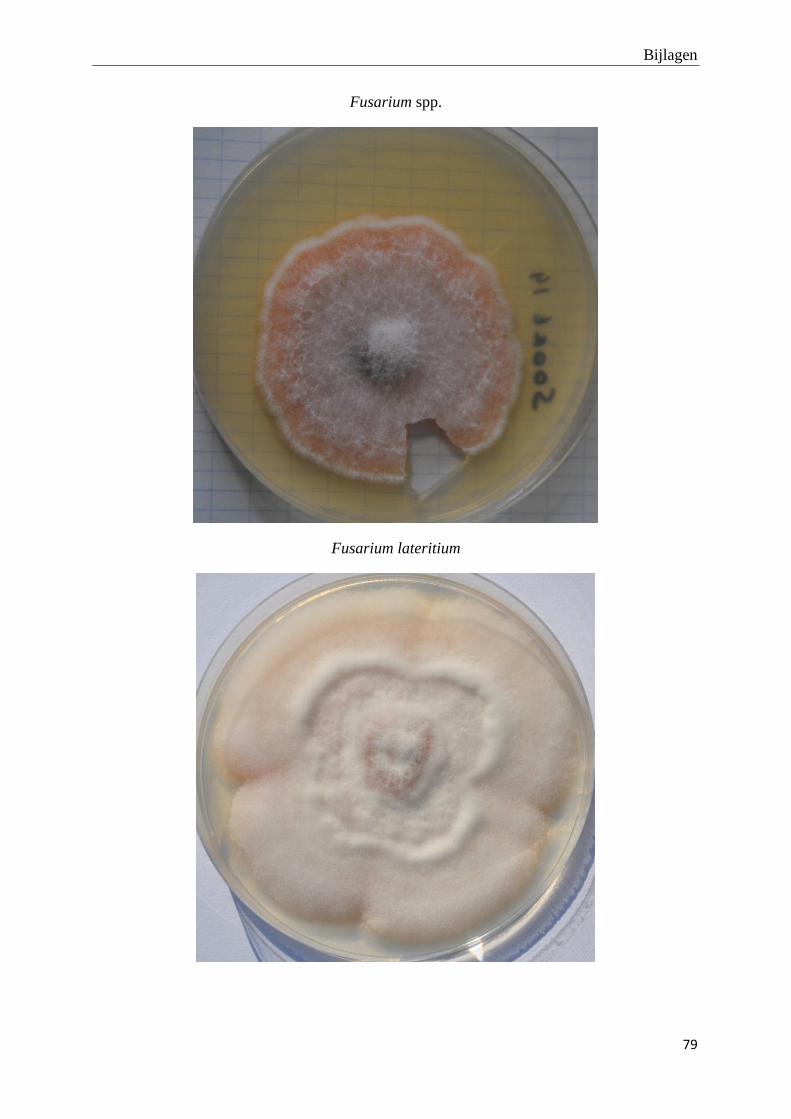
Bijlagen
79
Fusarium spp.
Fusarium lateritium

Bijlagen
80
Fusarium oxysporum (3)
Fusarium culmorum

Bijlagen
81
Bijlage 4
PCR resultaten: de verkregen amplicons vertonen, met uitzondering van één amplicon (geen
fungi), zoals verwacht een lengte van ongeveer 600 bp. Dit is de lengte van de gewenste ITS-
sequenties. In sommige rijen zijn geen bandjes waar te nemen en dit kan verschillende
oorzaken hebben, zoals de afwezigheid van DNA of interferentie van de PCR reactie door
bepaalde substanties (bv.: agar).

Bijlagen
82
Bijlage 5
Codes in R (versie 2.15)
4.1. Ruimtelijke distributie van X. monospora
Vergelijking van opgemeten parameters tussen de zes bezochte bosfragmenten onderling.
1e stap: nagaan van voorwaarden tot uitvoeren van ANOVA
- Nagaan van normaliteit van data
Shapiro-Wilk normality test
(shapiro.test(variabele))
H0: data is normaal verdeeld
p > 0.05: data is normaal verdeeld
- Nagaan van afwezigheid van correlatie tussen gemiddelde en variantie
Correlatie test
(cor(gem,var))
cor > 0.08: sterke correlatie tussen gemiddelde en variantie
- Nagaan van homogeniteit van varianties
Levene test
H0: varianties zijn homogeen
p > 0.05: varianties zijn homogeen
2e stap: uitvoeren van ANOVA (voorwaarden voldaan) of Kruskal-Wallis test (voorwaarden
niet voldaan)
Graad van kroonbedekking (%)
>library(nlme)
>lme1=lme(Canopy~fragment, random=~1|Plot)
>anova(lme1)
numDF denDF F-value p-value
(Intercept) 1 44 27772.675 <.0001
Fragment 5 16 5.934 0.0027

Bijlagen
83
Densiteit aan adulte bomen (X. monospora en andere boomsoorten)
>x=aov(log.densiteitadult~fragment)
>anova(x)
Analysis of Variance Table
Response: log.densiteitadult
Df Sum Sq Mean Sq F value Pr(>F)
Fragment 5 1.6861 0.33721 2.5323 0.07822
Residuals 14 1.8643 0.13317
Densiteit aan adulte X. monospora bomen
>x=aov(log.densiteitxym~fragment)
>anova(x)
Analysis of Variance Table
Response: log.densiteitxym
Df Sum Sq Mean Sq F value Pr(>F)
Fragment 5 1.6655 0.33311 0.7572 0.5933
Residuals 16 7.0384 0.43990
Relatieve densiteit aan adulte X. monospora bomen
>x=aov(relatievedensiteitxym~fragment)
>anova(x)
Analysis of Variance Table
Response: relatievedensiteitxym
Df Sum Sq Mean Sq F value Pr(>F)
Plot 5 0.038122 0.0076243 0.463 0.7974
Residuals 14 0.230533 0.0164667
Relatieve dominantie aan adulte X. monospora bomen
>x=aov(relatievedominantiexym~fragment)
>anova(x)
Analysis of Variance Table
Response: relatievedominantiexym

Bijlagen
84
Df Sum Sq Mean Sq F value Pr(>F)
Fragment 5 0.07779 0.015558 0.237 0.9396
Residuals 14 0.91887 0.065633
Diameter van adulte X. monospora bomen (cm)
>kruskal.test(diameter~fragment)
Kruskal-Wallis rank sum test
Data: diameter by fragment
Kruskal-Wallis chi-squared = 22.821, df = 5, p-value = 0.0003652
Densiteit aan X. monospora zaailingen
>x=aov(log.densiteitzaailing~Fragment)
>anova(x)
Analysis of Variance Table
Response: log.densiteitzaailing
Df Sum Sq Mean Sq F value Pr(>F)
Fragment 5 1.5037 0.30075 0.3406 0.8798
Residuals 14 12.3604 0.88289
Lengte X. monospora zaailingen
Kruskal-Wallis rank sum test
Data: log.lengte by fragment
Kruskal-Wallis chi-squared = 43.3142, df = 5, p-value = 3.191e-08
Afstand tussen zaailing en dichtstbijzijnde moederboom
>library(nlme)
>lme1=lme(log~Fragment, random=~1|Plot)
>anova(lme1)
numDF denDF F-value p-value
(Intercept) 1 486 220.9911 <.0001
Fragment 5 15 0.6831 0.6433

Bijlagen
85
4.2. Ruimtelijke distributie van bodemfungi gemeenschap
4.2.2. Gemeenschapsanalyse
Vergelijking van CFU tussen de zes bezochte bosfragmenten onderling.
Fungi morfotypes met potentieel effect (groep 1 en 2)
Cylindrocarpon
>Cylindr4=Cylindr^0.25
>lme3=lme(Cylindr4~Fragment, random=~1|Plot)
>anova(lme3)
numDF denDF F-value p-value
(Intercept) 1 133 15.79062 1e-04
Fragment 5 14 12.42874 1e-04
Fusarium
>Fus4=Fus^0.25
>lme2=lme(Fus4~Fragment, random=~1|Plot)
>anova(lme2)
numDF denDF F-value p-value
(Intercept) 1 133 148.12666 <.0001
Fragment 5 14 52.35896 <.0001
Ilyonectria
>Ilyon4=Ilyon^0.25
>lme1=lme(Ilyon4~Fragment, random=~1|Plot)
>anova(lme1)
numDF denDF F-value p-value
(Intercept) 1 133 413.4101 <.0001
Fragment 5 14 27.1301 <.0001

Bijlagen
86
Penicillium
>Pen4=Pen^0.25
>lme4=lme(Pen4~Fragment, random=~1|Plot)
>anova(lme4)
numDF denDF F-value p-value
(Intercept) 1 130 82.20664 <.0001
Fragment 5 14 1.70412 0.1984
Trichoderma
>Trichod4=Trichod^0.25
>lme5=lme(Trichod4~Fragment, random=~1|Plot)
>anova(lme5)
numDF denDF F-value p-value
(Intercept) 1 130 126.0504 <.0001
Fragment 5 14 0.6260 0.6829
Umbelopsis
> Umbel4=Umbel^0.25
> lme6=lme(Umbel4~Fragment, random=~1|Plot)
> anova(lme6)
numDF denDF F-value p-value
(Intercept) 1 130 142.28040 <.0001
Fragment 5 14 3.49475 0.0294
Fungi morfotypes zonder gekend effect (groep 3)
>G_effect4=G_effect^0.25
>lme7=lme(G_effect~Fragment, random=~1|Plot)
>anova(lme7)
numDF denDF F-value p-value
(Intercept) 1 138 2934.0102 <.0001
Fragment 5 14 5.4042 0.0056
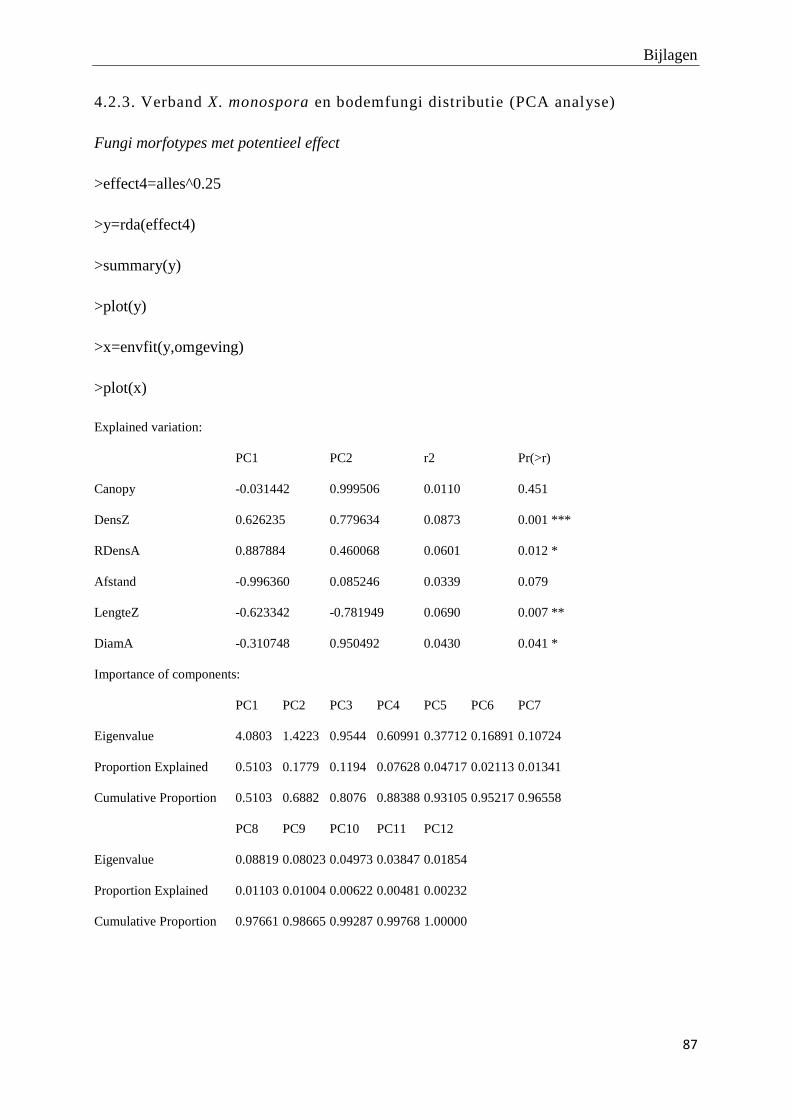
Bijlagen
87
4.2.3. Verband X. monospora en bodemfungi distributie (PCA analyse)
Fungi morfotypes met potentieel effect
>effect4=alles^0.25
>y=rda(effect4)
>summary(y)
>plot(y)
>x=envfit(y,omgeving)
>plot(x)
Explained variation:
PC1 PC2 r2 Pr(>r)
Canopy -0.031442 0.999506 0.0110 0.451
DensZ 0.626235 0.779634 0.0873 0.001 ***
RDensA 0.887884 0.460068 0.0601 0.012 *
Afstand -0.996360 0.085246 0.0339 0.079
LengteZ -0.623342 -0.781949 0.0690 0.007 **
DiamA -0.310748 0.950492 0.0430 0.041 *
Importance of components:
PC1 PC2 PC3 PC4 PC5 PC6 PC7
Eigenvalue 4.0803 1.4223 0.9544 0.60991 0.37712 0.16891 0.10724
Proportion Explained 0.5103 0.1779 0.1194 0.07628 0.04717 0.02113 0.01341
Cumulative Proportion 0.5103 0.6882 0.8076 0.88388 0.93105 0.95217 0.96558
PC8 PC9 PC10 PC11 PC12
Eigenvalue 0.08819 0.08023 0.04973 0.03847 0.01854
Proportion Explained 0.01103 0.01004 0.00622 0.00481 0.00232
Cumulative Proportion 0.97661 0.98665 0.99287 0.99768 1.00000
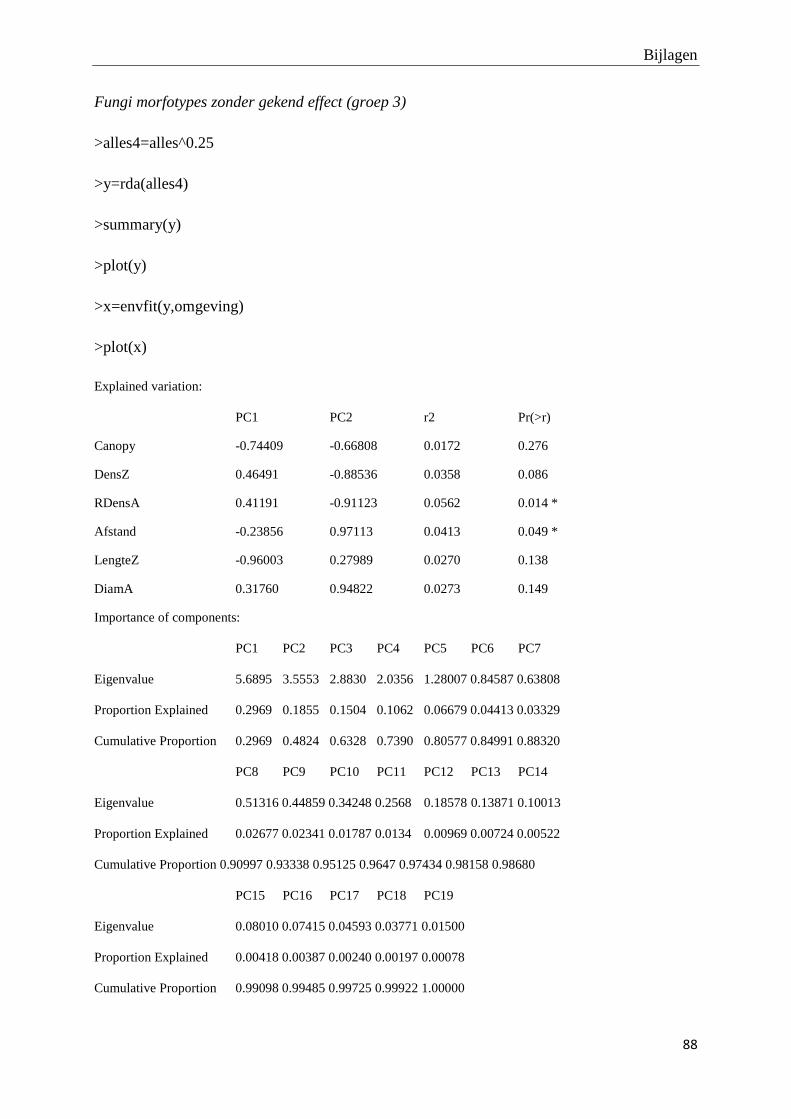
Bijlagen
88
Fungi morfotypes zonder gekend effect (groep 3)
>alles4=alles^0.25
>y=rda(alles4)
>summary(y)
>plot(y)
>x=envfit(y,omgeving)
>plot(x)
Explained variation:
PC1 PC2 r2 Pr(>r)
Canopy -0.74409 -0.66808 0.0172 0.276
DensZ 0.46491 -0.88536 0.0358 0.086
RDensA 0.41191 -0.91123 0.0562 0.014 *
Afstand -0.23856 0.97113 0.0413 0.049 *
LengteZ -0.96003 0.27989 0.0270 0.138
DiamA 0.31760 0.94822 0.0273 0.149
Importance of components:
PC1 PC2 PC3 PC4 PC5 PC6 PC7
Eigenvalue 5.6895 3.5553 2.8830 2.0356 1.28007 0.84587 0.63808
Proportion Explained 0.2969 0.1855 0.1504 0.1062 0.06679 0.04413 0.03329
Cumulative Proportion 0.2969 0.4824 0.6328 0.7390 0.80577 0.84991 0.88320
PC8 PC9 PC10 PC11 PC12 PC13 PC14
Eigenvalue 0.51316 0.44859 0.34248 0.2568 0.18578 0.13871 0.10013
Proportion Explained 0.02677 0.02341 0.01787 0.0134 0.00969 0.00724 0.00522
Cumulative Proportion 0.90997 0.93338 0.95125 0.9647 0.97434 0.98158 0.98680
PC15 PC16 PC17 PC18 PC19
Eigenvalue 0.08010 0.07415 0.04593 0.03771 0.01500
Proportion Explained 0.00418 0.00387 0.00240 0.00197 0.00078
Cumulative Proportion 0.99098 0.99485 0.99725 0.99922 1.00000


















