
ONTWIKKELING EN VALIDATIE VAN EEN …...Abstract (English) The performance of tests to determine the...
103
ONTWIKKELING EN VALIDATIE VAN EEN PREDICTIEVE MICROBIOLOGIE TOOL VOOR HALFFABRIKATEN Aantal woorden: 23 736 Simon Opsomer Stamnummer: 01270017 Promotor: Prof. dr. ir. Frank Devlieghere Tutor: Dr. ir. An Vermeulen Masterproef voorgelegd voor het behalen van de graad master in de richting Master of Science in de biowetenschappen: voedingsindustrie Academiejaar: 2016 - 2017
Transcript of ONTWIKKELING EN VALIDATIE VAN EEN …...Abstract (English) The performance of tests to determine the...

ONTWIKKELING EN VALIDATIE VAN
EEN PREDICTIEVE MICROBIOLOGIE
TOOL VOOR HALFFABRIKATEN
Aantal woorden: 23 736
Simon Opsomer Stamnummer: 01270017
Promotor: Prof. dr. ir. Frank Devlieghere
Tutor: Dr. ir. An Vermeulen
Masterproef voorgelegd voor het behalen van de graad master in de richting Master of
Science in de biowetenschappen: voedingsindustrie
Academiejaar: 2016 - 2017


“De auteur en de promotor geven de toelating deze scriptie voor consultatie beschikbaar te
stellen en delen van de scriptie te kopiëren voor persoonlijk gebruik. Elk ander gebruik valt
onder de beperkingen van het auteursrecht, in het bijzonder met betrekking tot de verplichting
de bron uitdrukkelijk te vermelden bij het aanhalen van resultaten uit deze scriptie.”
“The author and the promoter give the permission to use this thesis for consultation and to
copy parts of it for personal use. Every other use is subject to the copyright laws, more
specifically the source must be extensively specified when using the results from this thesis”.
Gent, juni 2017,
De promotor, De auteur,
Prof. dr. ir. Frank Devlieghere Simon Opsomer


V
Woord vooraf
Na maandenlang hard werken en zwoegen, kan ik met trots mijn thesis presenteren. Dit
eindresultaat zou nooit mogelijk geweest zonder de hulp van vele mensen. Daarom zou ik van
deze gelegenheid gebruik willen maken om hen te bedanken.
Om te beginnen zou ik mijn promotor, Prof. dr. ir. Frank Devlieghere willen bedanken. Bedankt
om mij dit jaar zo goed op te volgen en mij de kans te geven aan dit onderwerp te werken. U
kon altijd tijd vrij maken voor mijn vragen of opmerkingen. Ook wil ik mijn tutor, dr. ir. An
Vermeulen bedanken. Haar deur stond altijd open als ik vragen of problemen had. Bedankt ook
om mijn teksten na te lezen en te verbeteren.
Dan wil ik de CEO van CUBE, Stijn Snick bedanken voor de rondleiding in het bedrijf en het
vertrouwen in mij om aan deze thesis te werken. Lien Laurier, de kwaliteitsassistente van
CUBE, mag ik ook zeker niet vergeten. Bedankt voor de hulp tijdens het praktisch werk en de
snelle antwoorden op mijn mails.
Ik zou ook graag de labo-assistenten willen bedanken. Ze waren altijd bereid te helpen, met
name Ann Dirckx, en er ging een goede sfeer in de labo’s. Ann, je stond altijd voor mij klaar
als ik vragen had en was altijd bereid me te helpen, bedankt.
Hiernaast wil ik ook mijn vrienden bedanken. Jullie waren er altijd voor mij als het nu was om
mijn gedachten te verzetten met een fris pintje, of om mij te horen zagen. Bedankt voor de vele
mooie momenten die ik met jullie heb beleefd de afgelopen jaren en hopelijk komen er nog veel
bij. Ik kijk al uit naar de barbecues komende zomer.
Natuurlijk mag ik ook mijn vriendin, Annelies, zeker niet vergeten. Schat, bedankt om er altijd
voor mij te zijn en, met name het afgelopen jaar, en mij altijd te steunen. Veel tijd en aandacht
kropen in mijn thesis en daar was jij helaas het slachtoffer van. We gaan nu een nieuw hoofdstuk
tegemoet en ik kan niet wachten het samen met jou te doorlopen.
Vervolgens wil ik mij richten tot mijn familie. Mama en papa, bedankt voor het vertrouwen en
de vele steun tijdens mijn studentenjaren. Het ging niet altijd even gemakkelijk, maar jullie
stonden altijd klaar met advies en gaven me een duwtje in de goede richting. De raad werd
desondanks niet altijd goed ontvangen, maar zeker geapprecieerd. Ook mijn broer en zus mag
ik niet vergeten. Bedankt voor de steun en de afleiding in de weekends. Als laatste, maar zeker
niet de minst belangrijke, zou ik mijn opa willen bedanken. Opa, bedankt om mijn masterproef
na te lezen en je uitstekende kennis van de Nederlandse taal met mij te delen. Ik zeg het veel te
weinig, maar zou jullie toch graag via deze weg willen bedanken voor alle hulp tijdens mijn
studentenjaren. Bedankt!
De kans is groot dat ik nog enkele namen vergeet, dus bij deze nog een algemene dankjewel
aan iedereen die mij direct en indirect heeft geholpen met het schrijven van deze thesis.


VII
Abstract (Nederlands)
Het uitvoeren van testen om de houdbaarheid van producten na te gaan, zijn dagelijkse routines
geworden in de voedingsindustrie. Die testen zijn heel tijdrovend, zeker als het gaat over nieuwe
producten, maar ze zijn ook noodzakelijk door de stijgende vraag van de consument naar lang
houdbare producten.
Wegens deze problematiek werd een tool ontworpen gebaseerd op bestaande predictieve
modellen. Deze masterproef bouwt verder op de tool ontworpen door Margaux van Helleputte
in haar masterproef “Ontwikkelen van een evaluatietool voor de beoordeling van de microbiële
stabiliteit van halffabricaten”. De tool werd verder geoptimaliseerd en uitgebreid. De groei/niet-
groei voorstellingen zijn aangepast afhankelijk van de fysiologische staat van de pathogenen.
Verhittingscurves zijn toegevoegd voor Salmonella spp., Listeria monocytogenes, Bacillus
cereus, Yersinia enterocolitica en niet-proteolytische Clostridium botulinum. De tool bezit ook
de optie om, afhankelijk van het proces, de D-reductie te bepalen.
Vervolgens werden de groeigrafieken gevalideerd door het uitvoeren van challengetesten op
Listeria monocytogenes. De testen zijn verricht in de laboratoria van LFMFP-UGent volgens
de Europese procedure. Deze thesis is uitgevoerd in samenwerking met het bedrijf CUBE,
waarvan bepaalde producten werden gebruikt.
De tool biedt een snelle manier aan om een idee te krijgen over de houdbaarheid van een bepaald
product. Het is aangewezen dat de tool met de nodige zorg en kennis gebruikt wordt. Verdere
challengetesten zijn nodig voor het valideren van de andere pathogenen en om de bruikbaarheid
en de precisie te verhogen.
Kernwoorden: predictieve modellen, groei/niet-groei voorstellingen, verhittingscurves,
voorspellingstool

Abstract (English)
The performance of tests to determine the shelf life of products are becoming a daily routine in
the food industry. These tests are very time-consuming, especially for new products, but are
necessary because of the rising demand of consumers for products with a long shelf-life.
Because of these issues a tool was designed based on existing predictive models. This master
thesis builds on the tool designed by Margaux van Helleputte in her master thesis “Development
of an evaluation tool for the review of the microbial stability of half fabricates”. This tool was
further optimised and completed. Growth/ no growth interfaces are adapted dependent on the
physiological state of the pathogens. Heating curves are added for Salmonella spp., Listeria
monocytogenes, Bacillus cereus, Yersinia enterocolitica and the non-proteolytic Clostridium
botulinum. Besides there is an option implemented in the tool, depending on the process, to
determine the D-reduction.
The growth/ no growth conditions were validated by performing challengetests on Listeria
monocytogenes. These tests are performed in the laboratories of LMFP-UGent according to the
European technical guidance. This thesis is performed in co-operation with the company
CUBE, which delivered certain products.
The tool offers a fast method to measure the shelf life of a specific product. It is obvious that
the tool needs to be operated with certain caution and knowledge of the subject. Further
challengetests are needed to validate the other pathogens and to increase the utility and
precision.
Keywords: predictive models, growth/ no growth interfaces, heating curves, prediction tool

9
Inhoudstafel Woord vooraf ............................................................................................................................ V
Abstract (Nederlands) ............................................................................................................. VII
Abstract (English) ................................................................................................................. VIII
Inhoudstafel ................................................................................................................................ 9
Lijst van figuren ....................................................................................................................... 11
Lijst van tabellen ...................................................................................................................... 13
1. Inleiding ................................................................................................................................ 15
2. Voorspellende microbiologie ............................................................................................... 17
2.1. Theoretische modellen ................................................................................................... 17
2.1.1. Kinetische modellen ............................................................................................... 17
2.1.2. Probabilistische modellen ....................................................................................... 18
2.2. Tertiaire modellen ......................................................................................................... 18
2.2.1. Freeware modellen ................................................................................................. 19
2.2.2. Commercieel beschikbare modellen ....................................................................... 24
2.3. Europese projecten ........................................................................................................ 26
2.3.1. WebSim-MILQ....................................................................................................... 26
2.3.2. SOPHY ................................................................................................................... 26
3. Validatie van een voorspellingsmodel.................................................................................. 29
3.1. Challengetest ................................................................................................................. 29
3.2. Houdbaarheidstest ......................................................................................................... 30
3.3. Doelorganisme van deze thesis ..................................................................................... 30
3.3.1. Taxonomie en morfologie ...................................................................................... 30
3.3.2. Eigenschappen van Listeria monocytogenes .......................................................... 30
3.3.3. Voorkomen ............................................................................................................. 31
4. Materiaal en methode ........................................................................................................... 33
4.1. Aanpassingen aan het programma ................................................................................. 33
4.2. Bepalen van groei/ niet groei interfaces en verhittingscurves ....................................... 33
4.2.1. Groei/ niet groei voorstellingen met voorspelde gegevens .................................... 33
4.2.2. Groei/ niet groei interfaces bepaald via Microbial Responses Viewer .................. 34

10
4.2.3. Groei/ niet groei voostellingen bepaald via de database van ComBase ................. 34
4.2.4. Vergelijken van groei/ niet groei voorstellingen .................................................... 35
4.2.5. Verhittingscurves en decimale reductietijden ........................................................ 35
4.3. Challengetesten ............................................................................................................. 36
4.3.1. Voorbereiding van de media .................................................................................. 36
4.3.2. Opkweek van het inoculum .................................................................................... 37
4.3.3. Gebruikte levensmiddelen ...................................................................................... 38
4.3.4. VIDAS LMO2 ........................................................................................................ 39
4.3.5. Bereiding van inoculum en inoculatie .................................................................... 39
4.3.6. Uitplating en telling ................................................................................................ 39
5. Resultaten en bespreking ...................................................................................................... 41
5.1. Ontwikkelen van de softwaretool via ComBase ........................................................... 41
5.2. Groei/ geen groei voorstellingen gevormd via Microbial Responses Viewer .............. 43
5.3. Groei/ niet groei grafieken gevormd via de database van ComBase ............................ 43
5.4. Groei/ geen groei voorstellingen vergelijken ................................................................ 44
5.5. Verhittingsmodellen via Lemgo D-and z-value database for Food. ............................. 49
5.5.1. Verhittingsmodellen van Salmonella spp. .............................................................. 49
5.5.2. Verhittingsmodellen van Yersinia enterocolitica ................................................... 51
5.5.3. Verhittingsmodellen van Listeria monocytogenes ................................................. 52
5.5.4. Verhittingsmodellen van Bacillus cereus ............................................................... 53
5.5.5. Verhittingsmodellen van de niet-proteolytische Clostridium botulinum ............... 57
5.6. Validatie software tool aan de hand van challengetesten met L.monocytogenes ......... 60
5.6.1. Resultaten pH en aw meting van producten voor challengetest. ............................. 60
5.6.2. Resultaten van de VIDAS LMO2-test ................................................................... 61
5.6.3. Bepalen van groeipotentieel van Listeria monocytogenes bij 4 en 7 °C. ............... 61
5.6.4. Validatie groei/ niet groei voorstellingen ............................................................... 65
5.7. De predictieve microbiologie tool ................................................................................. 66
6. Algemeen besluit .................................................................................................................. 71
7. Bronvermelding .................................................................................................................... 71

11
Lijst van figuren
Figuur 1: algemeen verloop van de groeicurve van een micro-organisme met de vier
verschillende fasen. KVE = kolonievormende eenheden. .......................................................... 9
Figuur 2: implementatie van de verhittingscurves in de microbiologie tool via de formules INDEX en
VERGELIJKEN. ………………………………………………………………………………………28
Figuur 3: ALOA en supplementen. ……………………………………………………………………29
Figuur 4: aanmaak van de subculturen (vertaald uit EURL Lm Technical Guidance Document). …....30
Figuur 5: Half Fraser Broth met stomacherzak. ……………………………………………………….31
Figuur 6: groei/ niet groei voorstellingen van Bacillus cereus na 7 dagen bij 7 °C bij een
waarschijnlijkheid van respectievelijk 48,4% en 100% van de fysiologische staat . Met groen = geen
significante groei (< 0,45 log KVE/g); oranje = groei (0,45 < log KVE/g < 0,55); rood = groei (> 0,55
log KVE/g). ……………………………………………………………………………………………33
Figuur 7: printscreen van de optie ‘physical state uncertainty’ in de ‘ComBase Predictor’. Met links een
Gauss-curve van de waarden van de fysiologische staat en rechts de invloed van de waarde op de lag-
fase. …………………………………………………………………………………………………....34
Figuur 8: groei/ niet groei voorstelling van Bacillus cereus na 7 dagen bij 7 °C op basis van
experimentele waarden uit de database van ComBase. De waarden stellen het aantal gevallen groei voor
ten opzichte van het totaal aantal gevonden gevallen in breukvorm, of gehele getallen als groei werd
bepaald in alle gevallen. De y-as bevat de Aw-waarden en de x-as de pH waarden. Vanaf groei werd
waargenomen wordt dit met een rode kleur aangeduid, wit betekent dat er in geen enkel geval groei
werd waargenomen. ……………………………………………………………………………………36
Figuur 9: groei/ niet groei voorstelling van Bacillus cereus na 7 dagen bij 7 °C op basis van de
voorspellingen met de ‘ComBase Predictor’ met een waarschijnlijkheid van de fysiologische staat van
48,4% (Van Helleputte, 2016). ……………………………………………...…………………………36
Figuur 10: groei/ niet groei voorstellingen van Salmonella spp. na 7 dagen bij 7 °C op basis van de
database van ComBase (links) en de voorspellingen van de ‘ComBase Predictor’ met een
waarschijnlijkheid van de fysiologische staat van 48,4% (rechts; Van Helleputte, 2016). ..………….37
Figuur 11: groei/ niet groei voorstellingen van de niet-proteolytische Clostridium botulinum na 7 dagen
bij 7 °C op basis van de database van ComBase (links) en de voorspellingen van de ‘ComBase Predictor’
met een waarschijnlijkheid van de fysiologische staat van 48,4% (rechts; Van Helleputte, 2016).
………………………………………………………………………………………………..………..37
Figuur 12: groei/ niet groei voorstellingen van Yersinia enterocolitica na 7 dagen bij 4 °C op basis van
de database van ComBase (links) en de voorspellingen van de ‘ComBase Predictor’ met een
waarschijnlijkheid van de fysiologische staat van 48,4% (rechts; Van Helleputte, 2016). ……..……..38
Figuur 13: groei/ niet groei voorstellingen van Yersinia enterocolitica na 7 dagen bij 7 °C op basis van
de database van ComBase (links) en de voorspellingen van de ‘ComBase Predictor’ met een
waarschijnlijkheid van de fysiologische staat van 48,4% (rechts; Van Helleputte, 2016). ……………38
Figuur 14: groei/ niet groei voorstelling van Listeria monocytogenes na 7 dagen bij 4 °C op basis van
de voorspellingen van de ‘ComBase Predictor’ met een waarschijnlijkheid van de fysiologische staat
van 48,4% ( Van Helleputte, 2016). ………………………………………….……………………….39
Figuur 15: groei/ niet groei voorstellingen van Listeria monocytogenes na 7 dagen bij 4 °C op basis van
de database van ComBase (links) en de voorspellingen van de ‘ComBase Predictor’ met een
waarschijnlijkheid van de fysiologische staat van 82,5% (rechts). …………………………………….39

12
Figuur 16: groei/ niet groei voorstellingen van Listeria monocytogenes na 7 dagen bij 7 °C op basis van
de database van ComBase (links) en de voorspellingen van de ‘ComBase Predictor’ met een
waarschijnlijkheid van de fysiologische staat van 48,4% (rechts; Van Helleputte, 2016). ………..….40
Figuur 17: D-waarden (min) in functie van de temperatuur (°C) bij Salmonella spp. uit Lemgo. ……41
Figuur 18: Alle D-waarden in functie van de temperatuur van Salmonella spp. met een wateractiviteit ≥
0,95 uit Lemgo. ……………………………………………………………………..………….………42
Figuur 19: Thermale dodingstijdcurve van Salmonella spp. met passende trendlijn. …………………42
Figuur 20: D-waarden (min) in functie van de temperatuur (°C) bij Yersinia enterocolitica uit Lemgo +
Favier et al. (2007). …………………………………………………………………………….………43
Figuur 21: Thermale dodingstijdcurve van Yersinia enterocolitica met passende trendlijn. .…………44
Figuur 22: D-waarden (min) in functie van de temperatuur (°C) bij L. monocytogenes uit Lemgo.. …44
Figuur 23: thermale dodingstijdcurve van Listeria monocytogenes met passende trendlijn. …….……45
Figuur 24: D-waarden (min) in functie van de temperatuur (°C) bij B. cereus sporen uit Lemgo. …...46
Figuur 25: thermale dodingstijdcurve van Bacillus cereus met passende trendlijn. ……………….….48
Figuur 26: D-waarden (min) in functie van de temperatuur (°C) bij Clostridium botulinum sporen uit
Lemgo. ……………………………………………………………………………………………...…49
Figuur 27: D-waarden (min) in functie van de temperatuur (°C) bij sporen van de niet-proteolytische
Clostridium botulinum teruggevonden in de literatuur. …………………………………………….…50
Figuur 28: thermale dodingstijdcurve van de niet-proteolytische Clostridium botulinum met passende
trendlijn. …………………………………………………………………………………….…………51
Figuur 29: schimmelgroei op de drie stalen kaas saus na 21 dagen bij 7 °C. …………………………56
Figuur 30: resultaten challengetest met Listeria monocytogenes met log KVE/g in functie van het aantal
dagen (0, 7, 14 en 21) bij 4 °C. met lichtblauw = pompoen geitenkaas (p1), oranje = smokey pepper
(p2), grijs = witloof-mosterd (p3), geel = zwarte olijven (p4), donkerblauw = kaas saus (p5) en groen =
asian curry (p6). ……………………………………………………………………….………………57
Figuur 31: groei/ geen groei voorstelling van Listeria monocytogenes na 7 dagen bij 4 °C met een
waarschijnlijkheid van de fysiologische staat van 95,5%. ………………………..……………………57
Figuur 32: resultaten challengetest met Listeria monocytogenes met log KVE/g in functie van het aantal
dagen (0, 7, 14 en 21) bij 7 °C. met lichtblauw = pompoen geitenkaas (p1), oranje = smokey pepper
(p2), grijs = witloof-mosterd (p3), geel = zwarte olijven (p4), donkerblauw = kaas saus (p5) en groen =
asian curry (p6). ………………………………………………………………………….……………58
Figuur 33: groei/ geen groei voorstelling van Listeria monocytogenes na 7 dagen bij 7 °C met een
waarschijnlijkheid van de fysiologische staat van 82,5%. …………………………………………….58
Figuur 34: printscreen van de tool in Excel. Links bovenaan wordt een lijst gegeven van pathogenen die
kunnen uitgroeien aan de hand van de ingegeven T, pH en Aw. Rechts bovenaan kunnen de gekozen
stappen, waarmee de mogelijke uitgroei van de pathogenen wordt bepaald, worden teruggevonden.
Links onderaan worden de groei/ niet groei voorstellingen waargenomen en rechts onderaan de
verhittingscurves. …………………………………………………………………………..………….59

13
Lijst van tabellen
Tabel 1: Overzicht predictieve modellen met gegevens over de soort modellen (kinetisch en/of
probabilistisch), het aantal micro-organismen en welke soorten kunnen worden geselecteerd, de
verschillende producten, parameters die kunnen worden gewijzigd, de beschikbaarheid van de modellen
en of de modellen betalend zijn of niet.
gebruikte afkortingen: T = temperatuur, t = tijd, aw = wateractiviteit, i.c. = initieel celaantal. ………..19
Tabel 2: Levensmiddelen geassocieerd met de transmissie van listeriosis, vertaald uit Liu, (2008). ….24
Tabel 3: Gerapporteerde menselijke gevallen van listeriosis en meldingspercentages per 100,000 in de
EU/EEA, per land en jaar, van 2011–2015, vertaald uit Anon3. (2016). ………………………………25
Tabel 4: Gekozen producten met overeenkomstige pH- en aw-waarden uit de productenlijst van CUBE.
* IQF staat voor ‘individually quick frozen’ dit is een methode waarbij producten zeer snel
worden gekoeld met stikstof. Zo wordt de kwaliteit zeer goed behouden. ……………………………30
Tabel 5: Optimale waarschijnlijkheden van de fysiologische staat voor de vijf pathogenen bij 4 °C en /
of 7 °C. …………………………………………………………………………………………………41
Tabel 6: onderzochte D-waarden en temperaturen van de sporen van Bacillus cereus uit Lemgo met
bronvermelding. ……………………………………………………………………………………….47
Tabel 7: D- en z-waarden bekomen via de tool en opgezocht in de literatuur voor de vijf pathogenen.
* Bolton et al. (1999) werd als bron gebruikt voor het bepalen van een D-waarde................................51
Tabel 8: Eigenschappen van zes producten van CUBE. 1 = waarden uit de productenlijst van CUBE.
Vierde en vijfde kolom zijn pH en Aw waarden gemeten in het labo. 2 = waarden bepaald door CUBE bij een tweede meting. ………………………………………………52
Tabel 9: groeipotentieel van Listeria monocytogenes bij 4 °C van zes producten met de mediaan in het
vetgedrukt.
* De hoogste concentratie (log KVE/g) werd bekomen na 14 dagen en niet na 21 dagen. …………….54
Tabel 10: groeipotentieel van Listeria monocytogenes bij 4 °C van zes producten met de mediaan in het
vetgedrukt.
* De hoogste concentratie (log KVE/g) werd bekomen na 14 dagen en niet na 21 dagen. ……………..55

14

15
1. Inleiding
CUBE is een voedingsbedrijf opgericht in 2007 als spin-off van ‘Snick Ingredients’. Het bedrijf
is gelokaliseerd in Oostkamp en produceert vloeibare en pasteuze halffabrikaten, zoals sauzen,
vullingen, bouillons, soepen en pesto’s. Hiervoor worden veel verschillende grondstoffen
verwerkt, zoals groenten, fruit, zuivelproducten, vis en vleesproducten. De geproduceerde
halffabrikaten zijn meestal diepgevroren, maar kunnen ook gepekeld of gepasteuriseerd zijn.
Via een vriestunnel waarin gasvormige stikstof (-100 °C) wordt geblazen wordt op een snelle
manier diepgevroren. Sinds 2014 wordt er gebruik gemaakt van de IQF-technologie om
sommige vloeibare en pasteuze producten tot kleine druppels in te vriezen. (Verledens &
Rawoens, 2017)
Jaarlijks wordt een wisselend gamma van een vijfhonderdtal zeer uiteenlopende producten
geproduceerd (Van Helleputte, 2016). Vanwege dit groot spectrum aan geproduceerde
halffabrikaten vergt het veel tijd om de houdbaarheid van al die producten te bepalen. Vaak is
de tijdsdruk groot waardoor het moeilijk wordt om analyses uit te voeren voor elk product.
Hierdoor is het bedrijf op zoek naar een voorspellingstool om de microbiële groei mee te
bepalen.
De constructie van deze tool is vorig jaar gestart door Margaux Van Helleputte. In haar thesis
“Ontwikkelen van een evaluatietool voor de beoordeling van de microbiële stabiliteit van
halffabricaten” werden groei/ niet-groei condities voorspeld na 7, 14 en 21 dagen bij 4 en 7 °C.
Deze groeivoorspellingen werden voor vijf pathogenen bepaald, namelijk L. monocytogenes,
Salmonella spp., Y. enterocolitica, B. cereus en niet-proteolytische Cl. botulinum. De gevormde
grafieken werden geïmplementeerd in een tool die werd gemaakt in Excel®.
In deze thesis wordt er verder gewerkt op deze tool. Er worden nieuwe groei/ niet groei
interfaces bepaald voor verschillende waarden van de fysiologische staat via de ‘ComBase
Predictor’. De bekomen grafische voorstellingen van de groei worden vergeleken met de
groeivoorspellingen op basis van experimenteel bepaalde gegevens. Hieruit worden de meest
optimale voorstellingen geselecteerd en geïmplementeerd in de tool.
De groei/niet groei interfaces van Listeria monocytogenes worden vervolgens gevalideerd via
challengetesten. De producten worden bewaard bij 4 en 7 °C en groei wordt bepaald na 7, 14
en 21 dagen. De bekomen resultaten worden vergeleken met de voorspellingen van de tool.
Vervolgens wordt ook een optie voor de verhitting toegevoegd in de tool. Hiervoor wordt
gebruik gemaakt van de website ‘Lemgo D-and z-value Database for Food’. Verhittingscurves
worden opgesteld voor de hoogste gevonden D-waarden en bijpassende temperaturen voor de
vijf pathogenen. Functievoorschriften van de curves worden bepaald om de D-reductie van een
verhittingsstap te berekenen.

16
17
2. Voorspellende microbiologie
Voorspellende microbiologie is een vakgebied binnen de voedselmicrobiologie. Met
mathematische vergelijkingen worden de responsen bestudeerd van pathogene micro-
organismen naar hun omgevingsfactoren (temperatuur, zuurtegraad, etc.) in voeding. Hieraan
gaan vele stappen vooraf. Eerst wordt een proefopzet uitgedacht en data verzameld. Daarna
worden de parameters geschat, wordt het model gevalideerd en indien nodig geoptimaliseerd.
Die modellen laten wetenschappers toe het gedrag van pathogene en bederforganismen te
voorspellen in combinatie met verschillende factoren. Hierdoor kan zowel de veiligheid als de
kwaliteit van de voeding gegarandeerd worden. Predictieve modellen worden ingedeeld in twee
groepen: kinetische en probabilistische modellen. (Anon1., 2012; Pérez - Rodríguez & Valero,
2013)
2.1. Theoretische modellen
2.1.1. Kinetische modellen
Kinetische modellen beschrijven de verandering in het microbieel celaantal in functie van de
tijd. Die verandering is afhankelijk van het initieel celniveau, van de intrinsieke en extrinsieke
factoren. Intrinsieke factoren zijn de eigenschappen van het levensmiddel zelf, zoals de
wateractiviteit, de zuurtegraad, etc. De extrinsieke factoren zijn de omgevingsomstandigheden
die het product ondervindt, zoals de atmosfeer, de temperatuur, etc. Kinetische modellen
beschrijven vooral groeicurven, maar ook modellen die de overleving of inactivatie weergeven
worden teruggevonden.
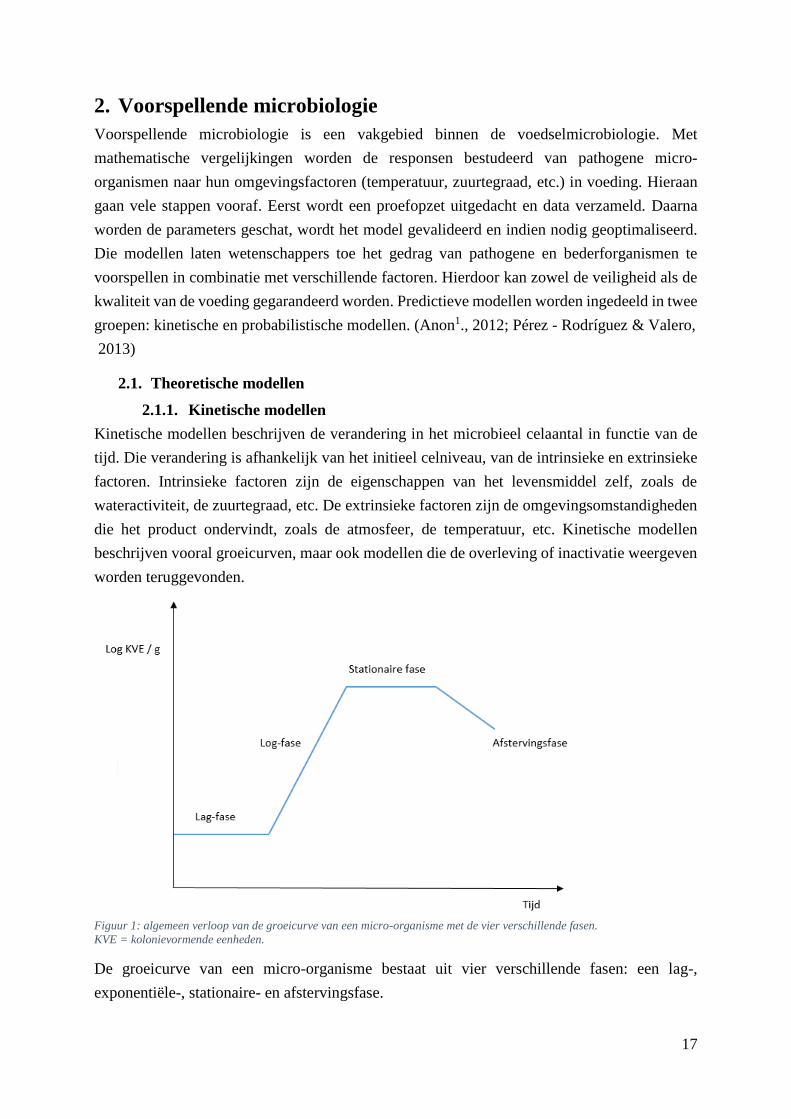
De groeicurve van een micro-organisme bestaat uit vier verschillende fasen: een lag-,
exponentiële-, stationaire- en afstervingsfase.
Figuur 1: algemeen verloop van de groeicurve van een micro-organisme met de vier verschillende fasen.
KVE = kolonievormende eenheden.

18
Tijdens de lag-fase, de eerste fase, heeft een cel tijd nodig om zich aan te passen aan zijn nieuwe
omgevingsfactoren, zoals pH, temperatuur, wateractiviteit, etc. Pas na een bepaalde tijd begint
de cel te groeien. De lag-fase verschilt per organisme en hangt af van de grootte van de
verschillen met zijn vorige omgeving.
De volgende fase wordt de exponentiële- , groei- of ook log-fase genoemd. In die periode vindt
verdubbeling van de cellen plaats. Één cel wordt twee, twee cellen worden vier enzo verder. De
log-fase kan worden gekwantificeerd via de generatietijd (GT) of de groeisnelheid (µ). De
generatietijd is de tijd dat een cel nodig heeft om zich te vermeerderen. De verandering in
celaantal over een bepaalde periode wordt gedefinieerd als de groeisnelheid. Beide zijn
afhankelijk van de extrinsieke en intrinsieke factoren die de cellen ondervinden tijdens de groei.
De generatietijd kan worden omgezet naar de groeisnelheid en vice versa met behulp van de
formule (1).
𝐺𝑇 = log 2
µ (1)
Het celaantal wordt vaak lineair voorgesteld door een semi-logaritmische schaal te gebruiken
(figuur 1).
Wanneer de populatie zijn maximale dichtheid bereikt, start de stationaire fase. Groei-
limiterende factoren, zoals de vermindering van nutriënten of de aanmaak van een inhiberend
bijproduct door het micro-organisme zijn vaak de oorzaak van dit stadium. Door die
belemmering is de celaangroei van het micro-organisme even groot als de afsterving.
Op het ogenblik dat de celdoding groter is dan de celaangroei, waardoor het celaantal dus daalt,
zit de cel in de afstervings- of inactivatiefase van de groeicurve. De groeiomstandigheden zijn
zeer ongunstig voor de bacteriën door de uitputting van nutriënten en de opeenstapeling van
inhiberende producten. (Anon1., 2009; Anon1., 2012)
2.1.2. Probabilistische modellen
Probabilistische modellen berekenen de kans dat, over een specifieke tijd of periode, een
bepaald fenomeen, zoals toxineproductie of groei, optreedt onder invloed van intrinsieke en
extrinsieke factoren. De uitkomsten van die modellen worden gecodeerd als 0 (fenomeen niet
waargenomen) en 1 (fenomeen waargenomen). Meerdere herhalingen worden onder dezelfde
condities uitgevoerd zodat een waarschijnlijkheid tussen de 0 en 100 % wordt bekomen, dat
een bepaald fenomeen voorkomt. Die probabiliteit kan worden gelinkt met de variabelen door
middel van logistische regressiemodellen. (Anon1., 2012)
2.2. Tertiaire modellen
In deze masterproef wordt een probabilistische microbiologie tool ontworpen met behulp van
gegevens bekomen uit andere modellen. Die modellen en andere veel gebruikte modellen
worden verder besproken en onderling vergeleken. Een algemeen onderscheid wordt gemaakt

19
tussen gratis (= freeware) en betalende programma’s. ComBase, FSSP, MRV, FISHMAP zijn
enkele software tools die behoren tot de freewaremodellen. FORECAST, CPMF2 en
Sym’Previus zijn betalend. Omdat het Europese projecten zijn, gericht naar de deelnemende
bedrijven, werden bepaalde modellen niet teruggevonden, waarvan WebSim-MILQ en SOPHY
twee voorbeelden zijn. (Gates, 2009)
2.2.1. Freeware modellen
2.2.1.1. ComBase
ComBase staat voor ‘Combined dataBase and models’. Oorspronkelijk begon ComBase als een
grote databank van groei- en inactivatiecurves voor de voorspellende microbiologie. IFR, FSA
en USDA verzamelden die data en de databank wordt regelmatig aangevuld. De partners
herkenden dat voedselmicrobiologen, voedselveiligheidmanagers en risicobeoordelaars in staat
moeten zijn om op een snellere manier specifieke microbiële data te lokaliseren in
gepubliceerde artikels, organisaties en in individuele onderzoekslaboratoria. (Man, 2002)
ComBase wil vooral de efficiëntie van het vinden van specifieke microbiële data verhogen,
snellere methoden voorzien om gegevens van verschillende labo’s te vergelijken en overbodige
microbiële studies te verminderen. Op de website kan de gebruiker met een interface zelf
variabelen invullen om zo specifieke voorspellingen te bekomen.
ComBase bestaat uit drie grote onderdelen: ‘Browser’, ‘ComBase Predictor’ en ‘Predictive
Models’. In de ‘Browser’ kan de gebruiker zelf criteria identificeren. Het is mogelijk om het
soort micro-organisme, soort medium, de zuurtegraad, de temperatuur, de wateractiviteit, etc.
te selecteren. Vertrekkend van die criteria kiest ComBase data uit zijn database, waaruit de
gebruiker dan studies selecteert en de data downloadt en exporteert.
De ‘ComBase Predictor’ omvat: ‘Growth’, ‘Thermal Inactivation’ en ‘Non-thermal Survival’.
Dankzij die tools kan de groei of inactivatie van verschillende pathogenen worden voorspeld
op basis van verschillende omgevingsomstandigheden. Op die manier bekomt de gebruiker
groei-, inactivatie- en overlevingscurves. Het is echter niet mogelijk om na te gaan welke
wetenschappelijke publicaties de curves voorspellen, wat een nadeel is.
‘Predictive Models’ is samengesteld uit twee modules: de ‘Perfringens Predictor’ en
‘Salmonella in egg’. Met de eerste module kan de gebruiker met een simulatie accuraat de
respons weergeven van Clostridium perfringens tijdens een koelproces van verhit vlees. De
gebruiker kan hierbij de wateractiviteit en de zuurtegraad zelf invullen. ‘Salmonella in egg’
voorspelt de groei van antibiotica-resistente Salmonella in vloeibare eiproducten tussen 10 °C
en 42 °C.
Tenslotte beschikt ComBase over de optie ‘DMFit’ die niet voor voorspellingen gebruikt wordt.
‘DMFit’ is een applicatie waarmee verschillende celaantal-versus-tijd modellen ‘gefit’ worden

20
aan eigen gegenereerde waarden, waaruit de gebruiker verschillende parameters, zoals
groeisnelheid, lag-fase, etc. kan schatten. (Anon1., 2015; Anon1., 2017)
2.2.1.2. Food Spoilage and Safety Predictor
De ‘Food Spoilage and Safety Predictor’ (FSSP) is een sterk uitgebreide versie van SSSP of
‘Seafood Spoilage and Safety Predictor’. Dit gratis programma is ontwikkeld om het praktisch
gebruik van mathematische modellen te vergemakkelijken. Deze modellen voorspellen de groei
en het bederf van pathogene micro-organismen in voeding.
Met SSSP kon het effect van de opslagtemperatuur op de ‘shelf life’ van vis bepaald worden.
Ook kon dit programma de groeimodellen van specifieke bederforganismen terugvinden, die
de ‘shelf life’ van verse vis voorspellen en modellen gebruiken die de voedselveiligheid, bv.
histaminevorming inschatten.
Aan FSSP zijn nieuwe voorspellingsmodellen toegevoegd, zoals een model dat de groei en
groeigrenzen van melkzuurbacteriën in vlees en zeevruchten bepaalt. Ook modellen waarmee
de gebruiker gelijktijdig de groei kan voorspellen van Listeria monocytogenes en
melkzuurbacteriën in zeevruchten, vleesproducten en gekoelde ongerijpte kaas (huttenkaas).
FSSP is het enige gratis programma dat die toename kan bepalen. Een algemeen groeimodel en
een model van de groeigrenzen voor elk micro-organisme/voedselcombinatie, werden ook
toegevoegd, waarbij parameters zoals de temperatuur en pH zijn bepaald. Dit soort modellen
brengen ook het effect van de opslagomstandigheden en producteigenschappen in rekening.
Voorspellingen worden verkregen voor zowel constante als dynamische temperaturen, pH-
waarden en organische zuren. FSSP biedt verder de optie om tal van parameters zelf in te
vullen, zoals T (=temperatuur), pH (=zuurtegraad), aw (=wateractiviteit), % CO2, nitriet en de
organische zuren: azijn-, benzoë-, citroen-, melk- en sorbinezuur.
FSSP vormt voor de onderzoeksgroep van de opleiding ‘voorspellende microbiologie’ van de
technische universiteit van Denemarken een belangrijk hulpmiddel voor de publieke sector bij
het consulteren, het leren en het geven van industrieel advies. (Dalgaard, 2014)
2.2.1.3. Microbial Responses Viewer
Het programma MRV of ‘Microbial Responses Viewer’ werkt met een ComBase-afgeleide
database bestaande uit microbiële groei/ geen groei data. De groei wordt gedefinieerd via
specifieke groeisnelheden, die worden gevisualiseerd door bepaalde kleuren. De gebruiker kan
zelf de assen en het bereik van de groei/ niet groei interfaces instellen om het gewenste gebied
te onderzoeken. Hierbij wordt de keuze gegeven tussen pH-aw, T-pH of T-aw als assen. Wanneer
bij een bepaalde parametercombinatie een kleur wordt aangeduid, worden alle groei/ niet groei
voorstellingen, die de kleur definiëren, weergegeven. Hier kan dezelfde opmerking worden
gemaakt als bij ComBase, namelijk dat het niet mogelijk is te achterhalen op basis van welke
wetenschappelijke publicaties de grafieken worden bepaald. (Anon2., 2017; Koseki, 2009)

21
2.2.1.4. FISHMAP-Program en Fish Shelf Life Prediction Program
Beide modellen zijn ontwikkeld door IFR (the Institute of Food Research) uit het Verenigd
Koninkrijk, in samenwerking met AZTI-TECNALIA, een technologiecentrum gelokaliseerd in
Spanje. De modellen zijn een Excel add-in dat loopt op Windows 3,1 en Excel 5 of hogere
versies. De voorspellingen kunnen worden opgeslagen in Excel. FSLP en FISHMAP vragen de
gebruiker een temperatuurprofiel in te geven waarbij de data geordend zijn, zodat de eerste
kolom de tijd bevat, uitgedrukt in uur en startend vanaf 0 en de tweede kolom de temperatuur.
Die waarden moeten zich bevinden tussen 0 en 20 °C voor FISHMAP en tussen 0 en 15 °C
voor FSLP om betrouwbare voorspellingen te bekomen. Variabelen zoals de pH en aw kunnen
niet worden aangepast in de programma’s, wat als een nadeel wordt beschouwd.
FISHMAP simuleert de groei van vier bederforganismen in visproducten, nl. C.
maltaromaticum, S. proteamaculans, S. baltica, Y. intermedia of een mix van de vier. De
gebruiker heeft de keuze om de voorspelling uit te voeren onder een specifieke
gassamenstelling (0-100% CO2) of onder luchtcondities (21% O2). De voorspelde data kunnen
ook worden vergeleken met experimentele data. Hierbij moeten de data opnieuw geordend
worden met de tijd in een eerste kolom en de groei (log KVE/g) in een tweede. De
voorspellingsmodellen zijn gebaseerd op microbiologische data verkregen door AZTI-
TECNALIA. (Alfaro et al., 2011)
Het programma FSLP of ‘Fish Shelf Life Prediction program’ voorspelt de microbiële groei en
de sensorische eigenschappen van visproducten. De microbiële groei wordt bepaald op basis
van het totaal kiemgetal; de sensorische voorspellingen zijn gebaseerd op gegevens van een
getraind panel. Het panel gaf scores van 9 (zeer vers) tot 4 (volledig bederf) aan de producten.
Een score van 6 vormt de grens van afkeuring voor de vis. FSLP biedt de mogelijkheid om de
respons van de TTI’s, tijd-temperatuurindicatoren, aan de grafiek toe te voegen bij constante
en fluctuerende temperaturen. TTI wordt beschreven als een eenvoudig, goedkoop toestel dat
een gemakkelijk berekenbare verandering toont op vlak van tijd en temperatuur. Hierdoor is
het mogelijk de volledige of gedeeltelijke temperatuurgeschiedenis en de kwaliteit van het
product te bepalen. Het programma kan ook de tijd bepalen tot de limiet van afkeuring bereikt
wordt. Hierbij worden twee tijden bepaald: één volgens het sensorisch panel en één volgens de
TTI. Het totaal kiemgetal wordt berekend voor beide tijdslimieten. De gebruiker kan ook zelf
de tijd bepalen waarop de microbiële en sensorische voorspellingen worden bepaald.
(Alfaro et al., 2007; Alfaro, 2008)
2.2.1.5. Refrigeration Index calculator – Meat & Livestock Australia
De ‘Refrigeration Index Calculator’ (RI) is een index voor de logaritmische groei van E. coli.
Het voorspellend model werd ontwikkeld door professor Tom Ross en zijn collega’s van de
universiteit van Tasmanië. RI is een vaak gebruikte term in de Australische exportcontrole van
vlees en vleesproducten. De calculator berekent de voorspelde groei van E. Coli op vlees

22
gebaseerd op de temperatuur, pH, aw en melkzuurconcentratie. De gebruiker kan zelf de data
van de temperatuur over een bepaalde tijd invullen. De waarden van de andere parameters zijn
afhankelijk van het geselecteerde producttype. Een belangrijke opmerking bij dit model is dat
RI niet dient als een voorspelling voor de groei van E. Coli in een product, maar eerder als
indicatie van de efficiëntie van het koelingsproces. (Anon1., 2016)
2.2.1.6. Risk Ranger
Met ‘Risk Ranger’ beschikt de gebruiker over een eenvoudige tool voor het berekenen van
voedselveiligheidsrisico’s. Dit programma is bedoeld als hulpmiddel bij het bepalen van
relatieve risico’s voor verschillende product-, pathogeen- en verwerkingscombinaties. ‘Risk
Ranger’ is een generiek model dat alle elementen van voedselveiligheid gebruikt om
risicoberekeningen te maken. De tool is een spreadsheet software in Excel en draagt bij om een
eerste schatting te maken van de mogelijke gevaren. De gebruiker wordt gevraagd 11 vragen te
beantwoorden over de ernst en het voorkomen van een risico in een product en de kans dat
iemand eraan wordt blootgesteld. (Anon., 2002) (Ross & Sumner, 2002)
2.2.1.7. Pathogen Modeling Program PMP Online
De ‘US Department of Agricultural Research Service’ te Wyndmoor in Pennsylvania ontwierp
een ‘Pathogen Modeling Program’ of PMP. Het programma omvat een pakket modellen dat
kan worden aangewend om de koeling, de overdracht tijdens het versnijden, de groei, de hitte-
inactivatie en de overleving te voorspellen van voedsel-overdraagbare bacteriën, voornamelijk
pathogenen. Het betreft bepaalde voeding, zoals gesneden zalm, gefermenteerde worsten,
gehakt, gekookt rundvlees, etc. De gebruiker kan zelf verschillende parameters instellen,
waaronder T, t, pH, aw, concentratie natriumnitraat, etc. specifiek bij de gekozen voeding. Op
de website kan de gebruiker naast bovenstaande vijf modellen ook vijftien bacteriën
terugvinden, waaronder vijf Salmonella-stammen. De experimentele data voor de modellen
werden onder bepaalde omstandigheden met specifieke bacteriële stammen bekomen. Hierdoor
wordt de accuraatheid van de voorspellingen niet gegarandeerd voor andere stammen of onder
andere omstandigheden. Een voordeel van PMP is dat er bij de modellen een directe link is
voorzien naar de gebruikte artikels. De groeivoorwaarden van de micro-organismen worden
echter niet goed gedefinieerd. Zo wordt bijvoorbeeld groei voorspeld voor Listeria
monocytogenes in krabsalade bij een pH van 3,7 voor de mayonaise, terwijl groei pas mogelijk
is vanaf een pH van 4,4. PMP kan online worden geraadpleegd en er worden continue nieuwe
modellen en verbeteringen aan toegevoegd. (Anon2., 2016)
2.2.1.8. Corbion Listeria Control model 2012
Corbion Purac heeft een unieke tool ontworpen, die de groei van Listeria precies kan
voorspellen. De ‘Listeria Control Model 2012’ is een gebruiksvriendelijk, kinetisch, online
groeimodel. De tool is aanpasbaar voor acht voedingsmiddelen zoals gebakken vlees, kant-en-
klare maaltijden, zachte kazen en salades en houdt rekening met het effect van organische zuren,

23
temperatuur, pH en vochtgehalte. Een nadeel is dat de pH beperkt is tussen de 5,8 en 7,2 en de
temperatuur tussen de 3,3 en 15 °C. De site biedt de keuze tussen de ‘basic trial version’, een
voorproefje van het programma en het werkelijk model. Met het volledige model kan de
gebruiker een extra parameter, zoals KCl instellen, de grafiek aanpassen, gedetailleerde
groeisnelheden verkrijgen en er is ook de optie om het verslag als PDF-bestand te downloaden.
Om die voordelen te verkrijgen vraagt de website om zich te registeren. De registratie is
volledig gratis, maar wel gericht naar bedrijven. (Anon2., 2012)
2.2.1.9. Shelf Stability Predictor
Met de online tool ‘Shelf Stability Predictor’ gaat de gebruiker na of Listeria monocytogenes
of Staphylococcus aureus uitgroeit in kant-en-klare vleesproducten op basis van pH en
wateractiviteit. Die groei wordt uitgedrukt in een waarschijnlijkheid die varieert van 0, groei
heel onwaarschijnlijk, tot 1, groei zeer waarschijnlijk. De tool is ontwikkeld door ‘the Center
for Meat Process Validation’ aan de universiteit van Wisconsin. Opmerkelijk is dat geen
rekening wordt gehouden met de temperatuur en geen periode gedefinieerd wordt, waarop het
mogelijk is de groei te bekijken. (Anon2., 2009)
2.2.1.10. THERM 2.0
‘The Center for Meat Process Validation’ ontwikkelde naast de tool ‘Shelf Stability Predictor’
ook het THERM-programma. Die tool voorspelt de groei van pathogenen in braadworst,
varkensvlees, rundvlees en gevogelte voor een korttijdige temperatuur mishandeling, tussen de
10 en 46 °C. Met ‘mishandeling’ wordt bedoeld dat door een onnauwkeurigheid, slechte
kalibratie, variaties, etc. de temperatuur tijdens het productieproces ongecontroleerd begint te
stijgen voor een korte periode. De gebruiker geeft de tijd van meten en de gemeten temperatuur
in en selecteert het soort vleesproduct. THERM bepaalt dan of Salmonella, E. coli of S. aureus
kunnen uitgroeien tot gevaarlijke hoeveelheden in die omstandigheden. Een nadeel hierbij is
dat de gebruiker geen waarden kan ingeven voor de pH en de aw van de producten. De website
beschikt over de optie om zelf een bestand te importeren waarop het programma dan de
voorspelling zal bepalen. (Anon., 2005)
2.2.1.11. GroPIN
GroPIN is een geïntegreerd tertiair model, ontwikkeld door ‘Agricultural University of Athens’
en ontworpen via VBA (Visual Basic for Application). Het programma is een
gebruiksvriendelijke database voor kinetische (groei en inactivatie) en probabilistische
modellen. Het biedt flexibiliteit met interactieve opties in het selecteren van grafische en
numerieke simulaties van modellen. Een ongelimiteerd aantal mathematische modellen kan
geïntroduceerd worden in de database via de ‘equation editor’ terwijl andere applicaties eerder
een beperkt aantal vergelijkingen bevatten. GroPIN bevat 367 gepubliceerde modellen die het
gedrag van 29 pathogenen en 43 bederforganismen in verscheidene levensmiddelen (vlees-,
zuivelproducten, groenten en fruit) bepalen. GroPIN houdt rekening met de invloed van de

24
intrinsieke en extrinsieke factoren op het microbieel bederf. Het programma bevat veel
inputsvariabelen voor de verschillende modellen, zoals de maximale groeisnelheid, de lag-fase,
de D-waarden, waarschijnlijkheid van groei, etc. die de gebruiker zelf kan invullen. GroPIN is
gratis te downloaden en wordt ondersteund door Excel 2007-2010. (Psomas & Skandamis,
2007)
2.2.2. Commercieel beschikbare modellen
2.2.2.1. FORECAST
De consultatiedienst Campden BRI ontwikkelde het systeem FORECAST. Het bevat modellen
voor bepaalde bederforganismen van specifieke voedingsmiddelen, zoals vis, vlees,
aangezuurde producten en gisten in fruit en drank . De ‘aangezuurde product’- en de ‘gist-in-
fruit-en-drank’-modellen zijn probabilistische modellen, die voorspellingen weergeven in
plaats van groeicurven. Campden BRI voert de voorspellingen uit, maakt een verslag, inclusief
groeicurves en het bepalen van de tijd om een specifiek celaantal te bereiken, en interpreteert
de informatie voor de klant. Hiervoor vraagt Campden BRI om informatie, zoals pH,
wateractiviteit, zoutgehalte en temperatuur door te geven. Wanneer die informatie niet
beschikbaar is, voert Campden BRI zelf ook analyses uit om die eigenschappen te bepalen.
De modellen, ontwikkeld voor aangezuurde producten, zijn gebaseerd op drie groepen van
organismen. Acidofiele gisten, schimmels en melkzuurbacteriën vormen de eerste categorie,
namelijk ‘bederf op koud gevulde producten’. ‘Pathogenen op koud gevulde producten’, zoals
E. coli, S. aureus en Salmonella behoren tot de tweede categorie. De derde categorie omvat
sporenvormers, zoals B. coagulans en C. pasteurianum, die warm gevulde producten
bederven. Campden BRI gebruikt ComBase en het ‘Pathogen Modeling Program’ voor het
bepalen van voedselpathogenen. (Anon1., 2009; Anon3., 2009)
2.2.2.2. CPMF2
LMFP-UGent en BioTeC-KULeuven werken samen in CPMF2 of de ‘Flemish Cluster
Predictive Microbiology in Foods’. Er wordt gebruik gemaakt van de kennis rond het wiskundig
modelleren van microbiële dynamica bij BioTeC (Afdeling Chemische en Biochemische
Procestechnologie en -regeling) en de kennis van de levensmiddelen-microbiologie bij LFMFP
(Laboratorium voor Levensmiddelenmicrobiologie en -conservering). Bij CPMF2 werden de
twee softwaremodellen ‘Listeria Meat Model’ en SWEETSHELF ontwikkeld.
Het kinetisch model ‘Listeria Meat Model’ voorspelt de groei/ niet-groei van Listeria
monocytogenes in MAP-verpakte vleeswaren. Hiervoor werden challengetesten uitgevoerd in
het laboratorium van LFMFP-UGent. Met de verkregen data en modellen, beschikbaar in de
wetenschappelijke literatuur, stelden de onderzoekers een modelaanpassingsfactor op. Via
groeipotentieel-testen werd het finale model gevalideerd.

25
SWEETSHELF voorspelt de microbiologische stabiliteit van producten met verlaagde
wateractiviteit bij verschillende temperaturen. Hierbij kan de gebruiker verschillende
parameters aanpassen, zoals aw, pH, ethanol, temperatuur, etc. Dit softwaremodel is een
probabilistisch model dat de groei of niet-groei voorspelt voor osmofiele gisten, zoals Z. rouxii
en xerofiele schimmels, zoals E. herbariorum. Meer dan dertig Belgische pralineproducenten
werkten aan dit project mee. Twee extra modellen met 1% azijnzuur en met 1500 ppm
sorbinezuur werden voor Z. rouxii ontwikkeld. (Anon1., 2012)
Er is ook een derde model, dat niet wordt beschreven op de website, namelijk het Salad and
Sauce model. Dit is een probabilistisch groei/geen groei model voor gisten, zoals
Zygosaccharomyces bailii en melkzuurbacteriën, zoals L. fructivorans, L. plantarum in zure
sauzen en voor Listeria monocoytogenes in salade op basis van mayonaise. (Vermeulen, 2008)
2.2.2.3. Sym’Previus
Sym’Previus is een commerciële software, ontwikkeld in Frankrijk. Het programma bestaat uit
drie delen: een database, simulatiesysteem en data-analysetool.
De database wordt geïnformeerd via een web browser, genaamd MIEL. De gebruiker kan
criteria, zoals de soort voeding, de micro-organismen en de omgevingsfactoren, selecteren. De
database omvat groei, overleving en hitte-inactivatie kinetiek voor bederforganismen en
pathogenen, zoals bij ComBase (Anon3., 2017).
Het simulatiesysteem houdt een grote variëteit aan primaire en secundaire modellen in om het
gedrag van micro-organismen onder bepaalde omgevingsfactoren in voeding te beschrijven. De
primaire modellen zijn gebaseerd op het ‘delay and break’ model van Rosso et al. (1995). De
secundaire modellen hebben parameters voor de minimale, maximale en optimale waarden van
de pH, aw en temperatuur. De simulatie van het gedrag van pathogenen in voeding via
secundaire modellen is gebaseerd op het gamma concept van Zwietering et al. (1993). De
primaire en secundaire modellen voor groei en hitte-inactivatie zijn gratis beschikbaar na
registratie. Het premium pakket heeft een meer uitgebreide groei en hitte-inactivatie simulatie
met daarnaast een ‘HACCP assistant’ en een ‘growth/ no growth interface’. De ‘HACCP
assistant’ (Hazard Analysis and Critical Control Point) bepaalt de kritische stappen van een
productieproces die een impact hebben op de groei van micro-organismen. De ‘growth/ no
growth interface’ helpt de karakteristieken van een product te optimaliseren en gaat de
mogelijkheid tot groei van Listeria monocytogenes na.
De data-analysetool combineert alle tools van Sym’Previus en voorziet een analyse uitgevoerd
door experten. Zo wordt een samenvatting gegeven van de kennis om de interactie tussen de
voeding en de micro-organismen beter te begrijpen. Sym’Previus is dus niet alleen een database
en een verzameling van mathematische modellen, maar meer een netwerk van
expertises. (Leporq et al., 2005; Anon3., 2012; Anon3., 2017)

26
2.3. Europese projecten
2.3.1. WebSim-MILQ
WebSim-MILQ, een web-gebaseerde modeltool, werd ontworpen om verhittingsprocessen in
de zuivelindustrie te verbeteren met respect voor de productveiligheid, de kwaliteit en de
kostprijs. Het heel gebruiksvriendelijk WebSim-MILQ is gericht naar de kleine en medium
zuivelbedrijven (KMO’s). Zestien Europese zuivelbedrijven werkten aan dit project (MILQ-
QC-TOOL) samen met ‘NIZO Food Research’ en ‘Wirelessinfo’, die de web-applicatie
ontwikkelden. De tool is op maat van de zestien bedrijven gemaakt en werd getest door de
verhittingsprocessen te optimaliseren voor yoghurt, verse room, chocolademelk en kaas.
WebSim-MILQ is een kinetisch model met gegevens over microbiële en enzymatische
inactivatie, proteïne denaturatie, componentenvorming, destructie en het biologische
rottingsproces van de producten. Hiervoor werd eerst een discussie gevoerd over de producten
en de mogelijke optimalisaties met de bedrijven. Daarna werd een audit georganiseerd om de
nodige gegevens te verzamelen en stalen af te nemen. De variabelen in het programma zijn
specifiek voor de processen die het bedrijf in kwestie uitvoert. Dankzij de tool werd de
productiekost en -afval tot 30% verminderd en werden het rendement en de kwaliteit van de
producten verbeterd. Andere bedrijven kunnen, volgens Schuytser et al. (2008), ‘NIZO Food
Research’ contacteren om ook toegang te krijgen tot het programma. ‘NIZO Food Research’
vraagt een jaarlijks bedrag voor de licentie van WebSim-MILQ. (van Dijk et al., 2008)
2.3.2. SOPHY
SOPHY staat voor ‘SOftware tool for Prediction of ready-to-eat food product sHelf life, quality
and safetY’. Voor de ontwikkeling werd een consortium opgericht van Europese bedrijven. Het
programma is gefinancierd door ‘the 7th Framework Programme of the European
Commission’. SOPHY schat het effect in van elke productiestap op de veiligheid en de
houdbaarheid, terwijl het ook rekening houdt met de kwaliteit van de producten. Vers gesneden
salades en delisalades zijn de modelproducten waarmee wordt gewerkt. Het programma omvat
zowel kinetische als probabilistische modellen en gebruikt hiervoor bestaande en zelf
gegenereerde data. De modellen zijn gebaseerd op meer dan 50 verschillende
producteigenschappen, ze omvatten algemene parameters, procesparameters, opslag- en
verpakkingsparameters en sensorische parameters. De software zal ook continu worden
geüpdatet met nieuwe producten, data, omgevingscondities, etc. Verder mikt SOPHY op het
verwerken van verschillende procestechnieken, productrecepten, omgevingsomstandigheden,
hygiëne-, kwaliteits- en veiligheidsmanagement. Geïnteresseerde bedrijven kunnen zich
registreren op de website. (Anon., 2013; Anon2., 2015)
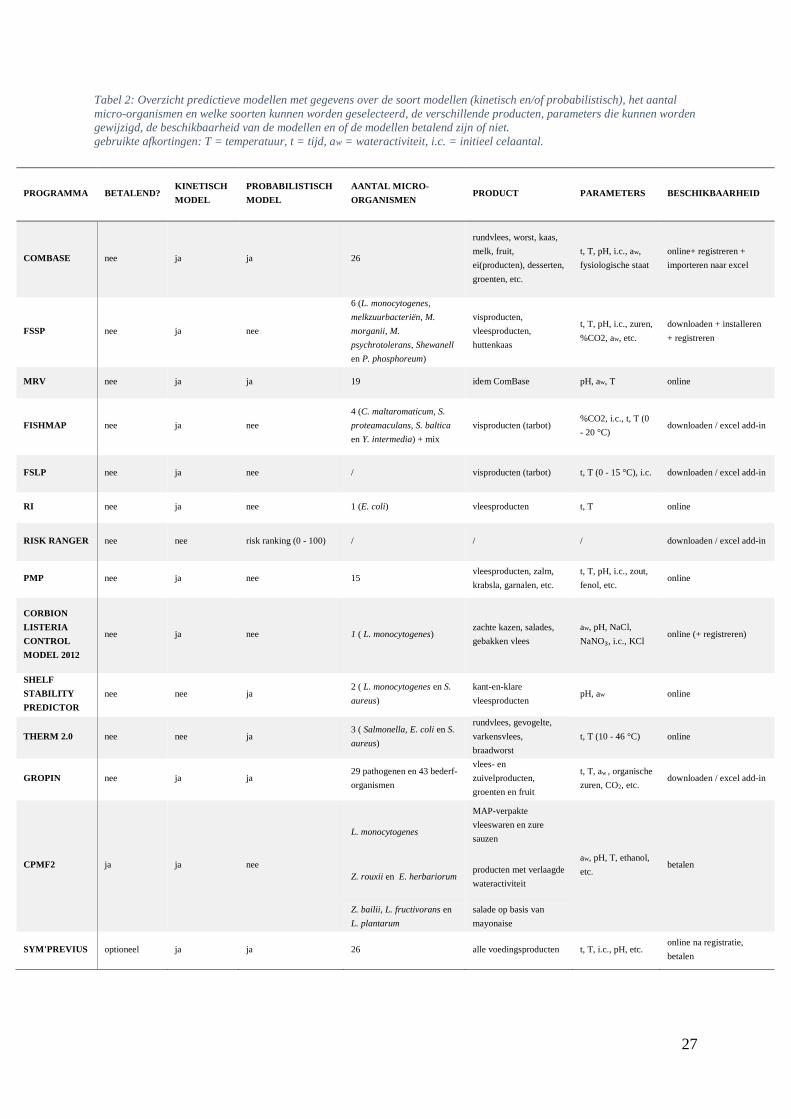
27
Tabel 2: Overzicht predictieve modellen met gegevens over de soort modellen (kinetisch en/of probabilistisch), het aantal
micro-organismen en welke soorten kunnen worden geselecteerd, de verschillende producten, parameters die kunnen worden
gewijzigd, de beschikbaarheid van de modellen en of de modellen betalend zijn of niet.
gebruikte afkortingen: T = temperatuur, t = tijd, aw = wateractiviteit, i.c. = initieel celaantal.
PROGRAMMA BETALEND? KINETISCH
MODEL
PROBABILISTISCH
MODEL
AANTAL MICRO-
ORGANISMEN PRODUCT PARAMETERS BESCHIKBAARHEID
COMBASE nee ja ja 26
rundvlees, worst, kaas,
melk, fruit,
ei(producten), desserten,
groenten, etc.
t, T, pH, i.c., aw,
fysiologische staat
online+ registreren +
importeren naar excel
FSSP nee ja nee
6 (L. monocytogenes,
melkzuurbacteriën, M.
morganii, M.
psychrotolerans, Shewanell
en P. phosphoreum)
visproducten,
vleesproducten,
huttenkaas
t, T, pH, i.c., zuren,
%CO2, aw, etc.
downloaden + installeren
+ registreren
MRV nee ja ja 19 idem ComBase pH, aw, T online
FISHMAP nee ja nee
4 (C. maltaromaticum, S.
proteamaculans, S. baltica
en Y. intermedia) + mix
visproducten (tarbot) %CO2, i.c., t, T (0
- 20 °C) downloaden / excel add-in
FSLP nee ja nee / visproducten (tarbot) t, T (0 - 15 °C), i.c. downloaden / excel add-in
RI nee ja nee 1 (E. coli) vleesproducten t, T online
RISK RANGER nee nee risk ranking (0 - 100) / / / downloaden / excel add-in
PMP nee ja nee 15 vleesproducten, zalm,
krabsla, garnalen, etc.
t, T, pH, i.c., zout,
fenol, etc. online
CORBION
LISTERIA
CONTROL
MODEL 2012
nee ja nee 1 ( L. monocytogenes) zachte kazen, salades,
gebakken vlees
aw, pH, NaCl,
NaNO₃, i.c., KCl online (+ registreren)
SHELF
STABILITY
PREDICTOR
nee nee ja 2 ( L. monocytogenes en S.
aureus)
kant-en-klare
vleesproducten pH, aw online
THERM 2.0 nee nee ja 3 ( Salmonella, E. coli en S.
aureus)
rundvlees, gevogelte,
varkensvlees,
braadworst
t, T (10 - 46 °C) online
GROPIN nee ja ja 29 pathogenen en 43 bederf-
organismen
vlees- en
zuivelproducten,
groenten en fruit
t, T, aw , organische
zuren, CO2, etc. downloaden / excel add-in
CPMF2 ja ja nee
L. monocytogenes
MAP-verpakte
vleeswaren en zure
sauzen
aw, pH, T, ethanol,
etc. betalen
Z. rouxii en E. herbariorum producten met verlaagde
wateractiviteit
Z. bailii, L. fructivorans en
L. plantarum
salade op basis van
mayonaise
SYM'PREVIUS optioneel ja ja 26 alle voedingsproducten t, T, i.c., pH, etc. online na registratie,
betalen

28

29
3. Validatie van een voorspellingsmodel
Vooraleer modellen in de praktijk gebruikt worden, moet worden aangetoond dat ze accuraat
het gedrag van een micro-organisme kunnen voorspellen in een specifiek levensmiddel. Deze
validatie van een microbiologisch model wordt bekomen door de voorspelde gegevens te
vergelijken met observaties in het product onder dezelfde omstandigheden. De waarnemingen
moeten onafhankelijk zijn van de data gebruikt bij het opstellen van het model. Zowel
challengetesten als houdbaarheidstesten zijn bruikbaar om het gedrag van een micro-organisme
in een levensmiddel te bepalen. (Ross, 1996)
3.1. Challengetest
Challengetesten bezorgen informatie over het gedrag van een micro-organisme in een
voedingsmiddel. Het product wordt geïnoculeerd met het doelorganisme onder specifieke
omstandigheden en de mogelijke groei van de bacterie wordt over een bepaalde periode
opgevolgd. De variabiliteit van de batches, de productsamples en de stammen worden hierbij
in rekening gebracht. Twee types challengetesten worden onderscheiden: een test om het
groeipotentieel te beoordelen en een test om de maximale groeisnelheid te bepalen. (Beaufort
et al., 2014)
Voor het bepalen van het groeipotentieel wordt het doelorganisme op een voedingsmiddel
geïnoculeerd. Het product wordt nadien bewaard onder omstandigheden, die de werkelijke
condities tijdens het transport, de opslag in een winkel en bij de consument benaderen. Een
microbiologische challengetest moet een representatie zijn van de mogelijke omstandigheden
die kunnen gebeuren tijdens de bescherming van de producten vanaf de producent tot bij de
consument. De bewaring start de dag van de contaminatie en duurt tot het einde van de
houdbaarheid. Het groeipotentieel, uitgedrukt in het logaritme kolonievormende eenheden per
gram (log KVE/g), is het verschil tussen de medianen, van drie herhalingen, op het einde en
op het begin van de test. Het belangrijkste voordeel van deze test is het meerekenen van de lag-
fase, want stress of adaptatie van de cellen kan namelijk de lag-fase beïnvloeden. De
mogelijkheid tot interpretatie van de gegevens is dan weer een nadeel. De resultaten zijn enkel
geldig voor het onderzochte product onder die specifieke omstandigheden van bijvoorbeeld tijd
en temperatuur. Dit type challengetest wordt ook toegepast in de masterproef. (Beaufort et al.,
2014)
Voor het bepalen van de groeisnelheid wordt het geïnoculeerde product bewaard onder een
constante temperatuur. Na uitvoering van de test kan de maximale groeisnelheid per stam en
per batch uit de groeicurves worden geschat. De maximale groeisnelheid is afhankelijk van de
initiële stam(men) en de intrinsieke en extrinsieke eigenschappen van het product. Het voordeel
van dit type challengetesten is de flexibiliteit. Het is mogelijk om via extrapolatie, van een µmax
bij een bepaalde temperatuur, de µmax op andere temperaturen te voorspellen. De formule (2)
kan hiervoor worden gebruikt, als T en Tref zich onder de 25 °C bevinden.

30
µ = µ𝑟𝑒𝑓 .(𝑇−𝑇𝑚𝑖𝑛)2
(𝑇𝑟𝑒𝑓−𝑇𝑚𝑖𝑛)2 (2)
Tmin is de minimale groeitemperatuur van het doelorganisme, hier ± -1,5 °C voor L.
monocytogenes. Tref en µref zijn de referentie waarden. Een nadeel, van de challengetesten om
de groeisnelheid te bepalen, is dat er geen rekening wordt gehouden met de lag- en stationaire
fase. (Beaufort et al., 2014)
3.2. Houdbaarheidstest
Houdbaarheidstesten maken het mogelijk een beoordeling te vormen over de houdbaarheid van
een natuurlijk gecontamineerd product tijdens de opslag onder werkelijkheidsgetrouwe
omstandigheden. Duurzaamheidstesten worden als meer realistisch gezien dan challengetesten
omdat de contaminatie natuurlijk gebeurt. Wegens een laag gehalte en een laag voorkomen van
de contaminatie zijn de testen veel meer gelimiteerd in uitvoering. (Beaufort et al., 2014)
3.3. Doelorganisme van deze thesis
De groei/ niet groei voorstellingen van Van Helleputte (2016) voorspellen de groei van de
volgende vijf micro-organismen: Salmonella spp., Listeria monocytogenes, Yersinia
enterocolitica, Bacillus cereus en de niet-proteolytische Clostridium botulinum. De groei/ niet
groei voorstellingen werden nog niet gevalideerd. Voor het bedrijf CUBE is Listeria
monocytogenes de meest interessante en belangrijkste pathogeen. Daarom wordt de nadruk in
deze thesis op Listeria monocytogenes gelegd en wordt de groei ervan onderzocht via
challengetesten.
3.3.1. Taxonomie en morfologie
L. monocytogenes is afkomstig van het geslacht Listeriae. Liu (2008) bespreekt de zes soorten
binnen het geslacht, namelijk L. innocua, L. seeligeri, L. welshimeri, L. grayi, L. ivanovii en L.
monocytogenes. De laatste twee soorten kunnen respectievelijk de ziekte Listeriosis verwekken
bij zoogdieren en mensen; de andere soorten zijn niet-pathogeen.
De micro-organismen van het geslacht Listeriae zijn niet-sporenvormend en worden
onderverdeeld onder de gram-positieve bacteriën. Ze zijn staafvormig en hebben als afmeting
0,4 – 0,5 µm bij 1 - 2 µm. De bacteriën komen alleenstaand of in korte ketens voor. Bij een
temperatuur tussen de 20 en 25 °C bezitten de zes soorten zweepstaartjes en ze zijn mobiel.
Maar vanaf 37 °C wordt de productie van flagella gereduceerd en ze worden immobiel. (Liu,
2008)
3.3.2. Eigenschappen van Listeria monocytogenes
L. monocytogenes is psychrotroof, het groeit zowel bij koude als bij hogere temperaturen. Groei
is waargenomen bij temperaturen van -1,5 tot 45 °C in levensmiddelen. De optimale groei ligt
tussen de 30 en 37 °C (Beaufort et al., 2014). Listeria groeit in een groot gamma aan producten

31
met een wateractiviteit van ten minste 0,92. Het kan concentraties van 10% NaCl en 200 ppm
NaNO2 ( = E250) overleven. Het product moet een zuurtegraad bezitten tussen de 4,3 en 9,4
om groei mogelijk te maken. De optimale pH is 7. Verder is Listeria aeroob en facultatief
anaeroob, het kan zowel bij de aan- en afwezigheid van zuurstof groeien (Liu, 2008).
Walker et al. (1990) onderzocht de aangroei van Listeria op koelkasttemperaturen (0 - 10 °C).
In de studie werden generatietijden bepaald van 13 tot 24 u bij 5 °C met een lag-fase van
maximaal drie dagen in UHT-melk en kippenbouillon. Afdoden via verhittingsprocessen is dus
nodig voor producten met een lange houdbaarheid om de veiligheid te garanderen. Hierbij moet
rekening gehouden worden met variabelen, zoals de groeitemperatuur, de pH, de aw, etc. Pagan
et al. (1997) toonde aan dat cellen van Listeria monocytogenes, die groeiden bij 37 °C, een
hogere hittetolerantie hadden dan de cellen die groeiden bij 4 °C. Door de wateractiviteit te
verlagen worden de cellen resistenter tegen verhitting. Een achtvoudige stijging in de
hitteresistentie van Listeria werd waargenomen door 1,5 mol/l NaCl toe te voegen aan het
medium (Jørgensen et al., 1995).
De ‘heat shock response’ speelt ook een rol bij de hitteresistentie. Dit fenomeen treedt op
wanneer L. monocytogenes wordt blootgesteld aan subletale temperaturen tussen de 45 en
50 °C voor het finale verhittingsproces. De cellen verkrijgen dan een verhoogde
thermotolerantie. Een studie van Shen et al. (2014) toont aan dat de tijd van blootstelling aan
subletale temperaturen ook een invloed heeft op de hitteresistentie. De hitteresistentie werd
bepaald bij een ‘heat shock’ van 48 °C voor 0, 5, 15, 30, 60 en 90 min van drie stammen Listeria
monocytogenes. De grootste hitteresistentie werd bekomen na het verhitten gedurende 30 en 60
min, overdadig verhitten voor 90 min leidde tot een verminderde resistentie.
Wanneer bij aanvang van een verhittingsproces sterk gekoeld wordt, wordt het omgekeerde
effect bekomen. Door een ‘cold shock’ zal Listeria monocytogenes veel gevoeliger worden voor
een verhittingsproces (Youssef et al., 2003).
Een pasteurisatie gericht op het afdoden van de meest hitteresistente vegetatieve pathogeen,
namelijk L. monocytogenes met een D70 = 0,25 min, wordt verkregen door het levensmiddel 2
min te verhitten bij 70 °C. Met dit pasteurisatiebarema wordt een 6D-reductie bekomen en
wordt de afwezigheid van L. monocytogenes in het levensmiddel gegarandeerd (Devlieghere et
al., 2014).
3.3.3. Voorkomen
Dankzij zijn vermogen om te overleven in talrijke stresssituaties, zoals aw, pH, temperatuur is
het geen verrassing dat L. monocytogenes wijdverspreid voorkomt in de omgeving. Op elk
gegeven moment zijn 5 tot 10% van de menselijke populatie en 1 tot 5% van de dierlijke
populatie drager van het pathogeen. Listeria wordt teruggevonden in de bodem, rottende
vegetatie, rivieren, uitwerpselen, het rioolwater en in de voeding voor dieren, maar komt ook
voor in de productieomgeving. In 99% van de menselijke gevallen van listeriosis, zowel de

32
sporadische als de epidemische ziekte, gebeurt de besmetting door de consumptie van
gecontamineerde levensmiddelen. De pathogeen komt voor in worsten, rauw vlees, salade,
rauwe melk, rauwe champignons, zachte kazen, zeedieren, etc. (Tabel 2). Voornamelijk Ready-
to-eat (RTE) levensmiddelen vormen een bron van Listeria uitbraken. Die levensmiddelen
hebben de eigenschappen de vermeerdering van Listeria te ondersteunen. Ze zijn ontwikkeld
met een verlengde houdbaarheid en worden geconsumeerd zonder verdere kookstap. (Liu,
2008)
Tabel 2: Levensmiddelen geassocieerd met de transmissie van listeriosis, vertaald uit Liu, (2008).
Nabesmetting leidt ook tot contaminaties met onder andere Listeria monocytogenes. Een
onderzoek in 1999 door de World Health Organisation (WHO) toonde aan dat nacontaminatie
de aanleiding was voor bijna 25% van de voedseluitbraken. Voornamelijk kruisbesmetting,
onhygiënische ruimtes, besmetting door het personeel en door de machines liggen aan de
oorzaak. Het opvolgen van de ‘Good Manufacturing Practices’ (GMP) en het opstellen van een
effectief HACCP-plan verminderen de kans op nabesmetting (Reij & Den Aantrekker, 2004).
Een extra korte verhittingstap van 2 min bij 70 °C volstaat ook om een veilig product te
verkrijgen (Devlieghere et al, 2014).
Volgens de wetgeving (EC Nr. 2073/2005) moet Listeria afwezig zijn in 25 g tijdens de
houdbaarheid van RTE producten bestemd voor kinderen of voor de medische zorg. In RTE-
producten, die de groei van de pathogeen ondersteunen, mag de limiet van 100 KVE/g niet
worden overschreden tijdens de houdbaarheid. Als de voorwaarde niet kan worden bewezen, is
de afwezigheid in 25 g vereist op het einde van het productieproces. Wanneer de groei niet
mogelijk is, mag tijdens de houdbaarheid de grens van 100 KVE/g niet worden overschreden.
Het EFSA, een agentschap van de Europese Unie voor voedselveiligheid, stelde in 2011
gemiddeld 0,36 incidenten vast per 100 000 Europeanen. In 2015 was dit cijfer aangegroeid tot
gemiddeld 0,46 gevallen. Ook in België is een stijging vast te stellen in het aantal gevallen
listeriosis tussen 2011 en 2015 (Tabel 3). In hetzelfde jaar veroorzaakte L. monocytogenes de
hoogste mortaliteit met 17,7% van de opgenomen zoönose, een infectieziekte waarbij mensen
door dieren besmet worden (Anon2., 2016).
Zuivelproducten Vleeswaren Visproducten Groenten Complexe levensmiddelen
zachte kazen gekookte kip vis koolsla sandwiches
(rauwe) melk worsten schelpdieren plantaardige stremsels
roomijs en softijs kalkoenworsten garnalen gezouten paddenstoelen
boter rillettes en paté gerookte vis alfalfa
varkenstong in gelatine schaaldieren rauwe groenten
kabeljauwkuit behandelde olijven
rijstsalade
gesneden fruit

33
Land
2015 2013 2011
Totaal
aantal
gevallen
Bevestigde
gevallen
Meldings-
percentages
Bevestigde
gevallen
Meldings-
percentages
Bevestigde
gevallen
Meldings-
percentages
België 83 83 0,74 66 0,59 70 -
EU
totaal 2291 2206 0,46 1883 0,45 1720 0,36
De meeste dodelijke gevallen komen voor in de YOPI-risicogroep, waartoe jongeren,
bejaarden, zwangere vrouwen en zieken behoren (Anon3., 2016). In 2010 rapporteerde het
CDC, de Amerikaanse ‘Centers for Disease Control and Prevention’, 1651
listeriosisbesmettingen tussen 2009 en 2011 met een letaliteit van 21% (Silk et al., 2013). Beide
mortaliteitpercentages liggen lager dan de gemiddelde waarde van 20 – 40%,
hospitalisatiepercentages van 90% werden ook vermeld in de studie (Liu, 2008). Wegens het
algemeen voorkomen en de grote mortaliteit vormt L. monocytogenes een grote bedreiging voor
de voedingsindustrie.
4. Materiaal en methode
4.1. Aanpassingen aan het programma
In het originele programma bevonden alle formules zich in hetzelfde tabblad waarop ook de
interface van het programma stond. De formules werden daarom verplaatst naar een apart
tabblad om verwijzingen overzichtelijker te houden en de kans op crashen te verlagen.
Lange formules die berekenden welke pathogenen uitgroeiden bij een gegeven temperatuur
werden teruggevonden. Door die lange vergelijkingen werd de maximale limiet van
verwijzingen naar eenzelfde cel bereikt op het ogenblik dat nieuwe toepassingen werden
toegevoegd. De formules werden daarom gesimplificeerd door te verwijzen naar een tabel met
de minimale en maximale temperaturen van de pathogenen. De oudere keuzelijsten werden ook
vervangen door nieuwe ActiveX keuzelijsten. Het besturingselement ActiveX biedt meer
flexibiliteit dan de traditionele formulierelementen. ActiveX heeft opties om zelf de lettertype-
eigenschappen aan te passen en lijsten geven als resultaat de waarde en niet de index, zoals bij
de formulierelementen.
4.2. Bepalen van groei/ niet groei interfaces en verhittingscurves
4.2.1. Groei/ niet groei voorstellingen met voorspelde gegevens
De grafische voorstellingen van de groei uit de masterproef “Ontwikkelen van een evaluatie-
tool voor de beoordeling van de microbiële stabiliteit van halffabricaten” werden kritisch
geëvalueerd. Ze stellen grafieken voor gecreëerd uit voorspelde data van ComBase voor vijf
Tabel 3: Gerapporteerde menselijke gevallen van listeriosis en meldingspercentages per 100,000 in de
EU/EEA, per land en jaar, van 2011–2015, vertaald uit Anon3. (2016).

34
pathogenen: Salmonella spp., Listeria monocytogenes, Bacillus cereus, Yersinia enterocolitica
en de niet-proteolytische Clostridium botulinum. De voorspellingen gebeurden na 7, 14 en 21
dagen bij 4 en 7 °C. De initiële concentratie werd op nul geplaatst en voor de fysiologische staat
werd de standaardwaarde gebruikt. Hierbij varieerde de pH van 4,5 tot en met 7,5 en de
wateractiviteit van 0,935 tot 0,995.
Voor drie verschillende waarden van de fysiologische staat werden nu groei/ niet groei
voorstellingen opgemaakt. Als eerste waarde werd de maximale waarde voor de fysiologische
staat gebruikt, nl. een waarde gelijk aan 1. Hierna werden grafische voorstellingen opgesteld
voor zowel de gemiddelde waarde plus één als twee keer de standaarddeviatie. De andere
parameters en variabelen bleven onveranderd ten opzichte van de masterproef van Van
Helleputte (2016). Een kleurcode werd ook gebruikt om de groei/ niet groei condities voor te
stellen. Een stijging kleiner dan 0,45 log KVE/g werd gedefinieerd als geen significante groei.
Dit werd in het groen aangeduid. De vakjes met een waarde tussen de 0,45 en 0,55 log KVE/g
werden in het oranje gekleurd. De rode vakjes stelden een significante groei voor, wat
overeenkomt met een stijging van meer dan 0,55 log KVE/g.
4.2.2. Groei/ niet groei interfaces bepaald via Microbial Responses Viewer
Met behulp van MRV werden de grafische voorstellingen van de groei voor experimenteel
bepaalde waarden opgesteld. Hierbij werd hetzelfde bereik voor de assen toegepast als bij de
groei/ niet groei interfaces verkregen via de ‘ComBase Predictor’. De grafieken werden ook
gevormd na 7, 14 en 21 dagen bij 4 en 7 °C. Bij sommige aw/pH-combinaties werd er zowel
groei als geen groei teruggevonden; daarom werden ze in breukvormen geschreven. Het aantal
groeiobservaties werd hierbij uitgedrukt in functie van het totaal aantal gevallen. Vanaf in één
van de observaties groei werd waargenomen, werd dit vakje, vertrekkend van een
worstcasescenario, in het rood gekleurd. Als er geen groei werd opgemerkt kreeg het vakje een
witte kleur. Groei werd gedefinieerd bij een stijging van minstens 0,50 log KVE/g.
4.2.3. Groei/ niet groei voostellingen bepaald via de database van ComBase
Vanwege de bekomen resultaten uit de groeivoorspellingen van MRV, werden groei/ niet groei
voorstellingen opgesteld via de database van ComBase. Die grafische voorstellingen van de
groei, voor experimenteel bepaalde waarden, bevatten hetzelfde bereik als de grafieken op basis
van de ‘ComBase Predictor’. Ook hier werden breuken gevormd van het aantal groeigevallen
in functie van het totaal aantal observaties bij een bepaalde aw/pH-combinatie. Dezelfde
kleurcode werd gehanteerd en vakjes werden in het rood gekleurd vanaf groei in één van de
observaties werd waargenomen. Groei werd gedefinieerd door een stijging van ten minste 0,50
log KVE/g.

35
4.2.4. Vergelijken van groei/ niet groei voorstellingen
De groei/ niet groei voorstellingen, bekomen via de ‘ComBase Predictor’, werden vergeleken
met de voorstellingen volgens de database van ComBase. Zo werden de voorspelde data
getoetst aan de experimenteel bepaalde data. Voor de groei van elke pathogeen bij 4 en/of 7 °C
werd de beste benadering bepaald, opnieuw vertrekkend van een worstcasescenario, voor de
grafieken na 7 dagen. De waarden van de fysiologische staat voor die optimale groei/ niet groei
condities werden als richtlijn genomen om de voorstellingen na 14 en 21 dagen te selecteren.
De geselecteerde groeivoorspellingen werden daarna in de tool gebracht.
4.2.5. Verhittingscurves en decimale reductietijden
‘Lemgo D- and z-value Database for Food’ is een website (http://www.hs-owl.de/fb4/ldzbase/)
die bij een bepaalde temperatuur de D- en z-waarde weergeeft van een micro-organisme. Ook
het stadium van de cel en het soort medium, dat van toepassing is in het gegeven onderzoek,
worden getoond. Alle waarden van de database komen uit 283 verschillende experimenten
(Althoff & Schwarzer, 2007).
De database van die website werd overgenomen in een Excel-document. Dit gebeurde voor
dezelfde vijf pathogenen waarvan groei/ niet groei interfaces werden bekomen. Alle waarden
werden verder uitgezet in grafieken met de D-waarde in functie van de temperatuur. Die
verhittingscurves werden geïmplementeerd in de voorspellingstool.
De implementatie gebeurde via een combinatie van de standaardformules INDEX en
VERGELIJKEN in Excel. Die combinatie werd verkozen boven de formule VERT.ZOEKEN,
omdat met VERT.ZOEKEN enkel naar een waarde kan worden gezocht en niet naar een foto
of in dit geval een verhittingscurve. Met de combinatie INDEX en VERGELIJKEN kan dit wel.
Via VERGELIJKEN wordt in een reeks van waarden de positie bepaald van de opgegeven
waarde. Via de INDEX-formule wordt uit een reeks van waarden of foto’s de gewenste waarde
of referentie geselecteerd. Figuur 2 toont de combinatie van beide formules via de zelfgemaakte
formule ‘verhittingsproces’. Eerst zal via VERGELIJKEN de positie van de opgegeven waarde
worden bepaald in het celbereik C11:C15 (blauw kader). Van de leesbare afkortingen in dit
bereik staat P.S. voor Salmonella spp., P.L.M. voor L. monocytogenes, etc. Via INDEX zal de
foto overeenkomstig met de bepaalde positie worden opgezocht in het celbereik D11:D15
(oranje kader) en worden weergegeven.

36
Naast verhittingscurves moet ook van een -proces de D-reductie kunnen worden voorspeld.
Daarom werden de hoogste D-waarden, weer vertrekkend van een worstcasescenario, en hun
corresponderende temperaturen geselecteerd en geplot. De z-waarde werd hierna bepaald voor
elk pathogeen om na te gaan of de bekomen functies betrouwbaar zijn. De bekomen z-waarden
werden vergeleken met de waarden, teruggevonden in de literatuur.
4.3. Challengetesten
De challengetesten werden uitgevoerd volgens het Europees protocol voor ‘conducting shelf-
life studies on Listeria monocytogenes in ready-to-eat foods’ (Beaufort et al., 2014).
4.3.1. Voorbereiding van de media
TSA of ‘Tryptone Soya Agar’ werd aangemaakt door 40 g op te lossen in 1 l gedestilleerd
water. De oplossing werd daarna opgekookt en dan gesteriliseerd door ze te autoclaveren voor
15 min bij 121 °C. Hierna werd het TSA in een warmwaterbad van 48 °C geplaatst om dan te
worden verdeeld over petriplaten. TSA is een niet-selectief medium, dat wordt gebruikt als
opslag van culturen, voor de isolatie van zuivere culturen en voor telling.
BHI of ‘Brain Heart Infusion’ werd bereid door 37 g op te lossen in 1 l gedestilleerd water.
Hierna werd de oplossing gemixt en verdeeld over proefbuisjes. De buisjes werden
geautoclaveerd voor 15 min bij 121 °C. BHI is een heel voedzaam medium, dat wordt toegepast
voor de opgroei van micro-organismen.
Figuur 2: implementatie van de verhittingscurves in de microbiologie tool via de formules INDEX en VERGELIJKEN.

37
ALOA werd samengesteld door 35,3 g op te lossen in
gedestilleerd water. Het mengsel werd dan opgekookt en
geautoclaveerd voor 15 min bij 121 °C. Na het autoclaveren werd
de oplossing onmiddellijk in een warmwaterbad van 48 °C
geplaatst om af te koelen. Wanneer ze de temperatuur van 48 °C
bereikte, werden de supplementen toegevoegd. De supplementen
bestaan uit een selectief en een verrijkingssupplement. Het
selectief supplement werd eerst opgelost in 5 ml ethanol 50%
vooraleer het werd toegevoegd. Het geheel werd nog eens kort
gemengd en daarna uitgegoten in petriplaten. De platen werden,
tot voor het gebruik, koel in een koelkast bewaard. ALOA wordt
gebruikt voor de detectie en telling van Listeria monocytogenes.
PPS of ‘Peptone Physical Salt’ werd bereid door 8,5 g zout te mengen met 1 g ‘Neutralised
Bacteriological Peptone’. Dit geheel werd dan opgelost in 1 l gedestilleerd water en gemengd.
Vervolgens werd de PPS voor 15 min geautoclaveerd bij 121 °C. PPS wordt benut als
verdunningsvloeistof.
‘Half fraser broth’ wordt gebruikt voor de primaire selectieve aanrijking van Listeria in voeding
en monsters. ‘Fraser broth’ wordt toegepast voor de secundaire selectieve aanrijking van
Listeria. Beide ‘broths’ maken deel uit van de VIDAS LMO2 kit om de afwezigheid van
Listeria monocytogenes in een product te controleren.
4.3.2. Opkweek van het inoculum
Drie verschillende L. monocytogenes stammen werden geselecteerd, nl. LFMFP stammen 392,
394 en 491. De stammen werden op glasparels bewaard bij -75 °C in een glyceroloplossing.
De stammen werden eerst opgekweekt met als doel het organisme in de vroege stationaire fase
te brengen. Hiervoor werd voor elke stam een parel in het BHI-medium gebracht en
geïncubeerd voor 24 uur bij 37 °C. Daarna vond voor 24 u en bij 37 °C een 4x4 uitplating op
TSA plaats om de zuiverheid van de cultuur na te gaan. Ook op ALOA werden de stammen
uitgeplaat om te controleren of het Listeria monocytogenes aanwezig was. De platen werden
omgekeerd in een plastieken zak voor 48 u en bij 37 °C geïncubeerd. Dan werd met een
oognaald één kolonie genomen van de ALOA-plaat en opnieuw 4x4 uitgeplaat op TSA voor
24 u bij 37 °C. Tenslotte werd één kolonie via een zigzagbeweging op een slant gebracht. De
slant werd in de koelkast bewaard bij 4 - 8 °C.
Figuur 3: ALOA en supplementen.
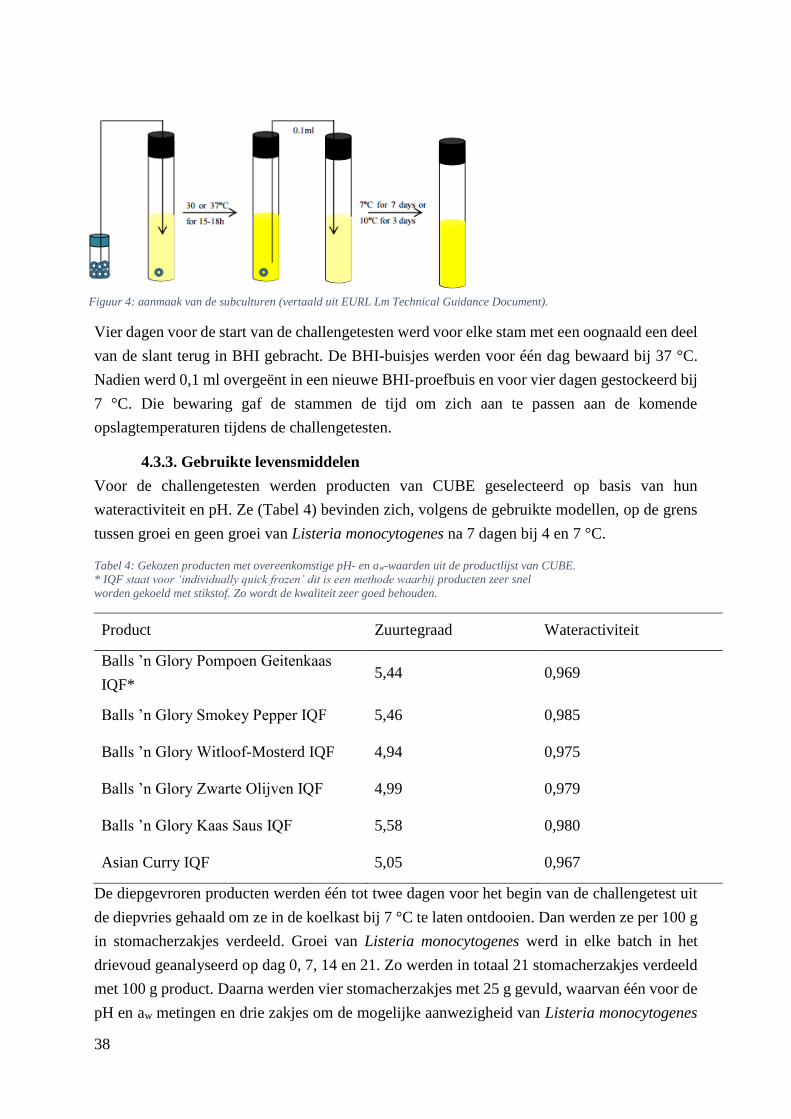
38
Vier dagen voor de start van de challengetesten werd voor elke stam met een oognaald een deel
van de slant terug in BHI gebracht. De BHI-buisjes werden voor één dag bewaard bij 37 °C.
Nadien werd 0,1 ml overgeënt in een nieuwe BHI-proefbuis en voor vier dagen gestockeerd bij
7 °C. Die bewaring gaf de stammen de tijd om zich aan te passen aan de komende
opslagtemperaturen tijdens de challengetesten.
4.3.3. Gebruikte levensmiddelen
Voor de challengetesten werden producten van CUBE geselecteerd op basis van hun
wateractiviteit en pH. Ze (Tabel 4) bevinden zich, volgens de gebruikte modellen, op de grens
tussen groei en geen groei van Listeria monocytogenes na 7 dagen bij 4 en 7 °C.
Tabel 4: Gekozen producten met overeenkomstige pH- en aw-waarden uit de productlijst van CUBE.
* IQF staat voor ‘individually quick frozen’ dit is een methode waarbij producten zeer snel
worden gekoeld met stikstof. Zo wordt de kwaliteit zeer goed behouden.
Product Zuurtegraad Wateractiviteit
Balls ’n Glory Pompoen Geitenkaas
IQF* 5,44 0,969
Balls ’n Glory Smokey Pepper IQF 5,46 0,985
Balls ’n Glory Witloof-Mosterd IQF 4,94 0,975
Balls ’n Glory Zwarte Olijven IQF 4,99 0,979
Balls ’n Glory Kaas Saus IQF 5,58 0,980
Asian Curry IQF 5,05 0,967
De diepgevroren producten werden één tot twee dagen voor het begin van de challengetest uit
de diepvries gehaald om ze in de koelkast bij 7 °C te laten ontdooien. Dan werden ze per 100 g
in stomacherzakjes verdeeld. Groei van Listeria monocytogenes werd in elke batch in het
drievoud geanalyseerd op dag 0, 7, 14 en 21. Zo werden in totaal 21 stomacherzakjes verdeeld
met 100 g product. Daarna werden vier stomacherzakjes met 25 g gevuld, waarvan één voor de
pH en aw metingen en drie zakjes om de mogelijke aanwezigheid van Listeria monocytogenes
Figuur 4: aanmaak van de subculturen (vertaald uit EURL Lm Technical Guidance Document).

39
na te gaan. Tenslotte werd een stomacherzakje van ongeveer 150 g als reserve voorzien en
bewaard in de diepvries.
4.3.4. VIDAS LMO2
VIDAS is een geautomatiseerd detectiesysteem voor
voedselpathogenen ontworpen door Biomérieux. Hierbij
werd de kit VIDAS LMO2 gebruikt, ontworpen voor de
detectie en confirmatie van Listeria monocytogenes in
voedingsmiddelen. Die test werd uitgevoerd op niet-
geïnoculeerde stalen om de afwezigheid van Listeria
monocytogenes in 25 g te bevestigen. Hierbij werd de
aanrijkingprocedure van de kit gevolgd. Eerst werd 25 g
van een levensmiddel in een stomacherzak met 225 ml
‘half fraser broth’ (Figuur 5) aangelengd. De stomacherzak werd dichtgelast en het mengsel
werd goed geschud. Daarna werden de zakjes bewaard bij 30 °C voor 24 - 26 uur. Dan werd
0,1 ml van het mengsel in buisjes van 10 ml ‘fraser broth’ gebracht en bij 37 °C bewaard voor
24 - 26 uur. Verder werd 500 µl uit de buisjes in de LMO2-strip gepipetteerd. Die werkwijze
werd per batch driemaal herhaald. Een blanco, positieve en negatieve controle werden ook
gepipetteerd in de LMO2-strips.
4.3.5. Bereiding van inoculum en inoculatie
Uitgaande van een gewenste besmetting van ongeveer 100 KVE/g werd de nodige hoeveelheid
per batch berekend. Uit ervaring met vroegere challengetesten is het gekend dat de BHI-buisjes
ongeveer 1,0 - 2,0 x 109 KVE/ml bevatten. Het inoculum werd decimaal verdund met PPS tot
105 KVE/ml werd bereikt. Een mix werd gevormd van de drie LFMFP stammen 392, 394 en
491. Hieruit werd, ook uit ervaring in het labo, 150 µl gepipetteerd om de producten in de
stomacherzakjes mee te inoculeren. De drie stammen werden decimaal verdund en uitgeplaat
om het celaantal in de mix te bepalen. De mix werd eveneens uitgeplaat om het inoculum te
kwantificeren. De inoculatie van de producten gebeurde in een verticale laminaire flow kast.
De stomacherzakjes werden na het inoculeren gemengd om de kolonies homogeen te verdelen.
Vervolgens werden de zakjes dichtgelast en op hun correcte temperatuur gestockeerd. De
stomacherzakjes van dag 0 werden hierna uitgeplaat.
4.3.6. Uitplating en telling
Voor de uitplating werden de ALOA-platen eerst 15 min gedroogd in de laminaire flow kast.
De stomacherzakjes werden handmatig gemengd en daarna werd 20 ± 0,2 g in een
stomacherzakje gebracht. Dit mengsel werd tien keer verdund door het product aan te lengen
met PPS. Hierna werden de zakjes in de stomacher gebracht en voor één minuut goed gemengd.
Vervolgens werd een decimale verdunningsreeks aangelegd.
Figuur 5: Half Fraser Broth met
stomacherzak.

40
Om de detectielimiet te verlagen werd 1 ml gelijk verdeeld over drie ALOA-platen. De vloeistof
werd met een steriele spatel uitgestreken over de gehele plaat. De platen werden voor 48 u
omgekeerd bewaard in een plastieken zak in de incubator bij 37 °C. Na twee dagen werd het
aantal Listeria monocytogenes kolonies geteld, herkenbaar aan hun blauw/groene kleur
omgeven door een halo-ring.
𝑁 = log (𝐶 . 10𝑑
𝑉 ) (3)
Het aantal kolonies kan worden berekend met de formule (3). Hierbij stelt d de verdunning
voor, V het inoculumvolume (1 of 0,1 ml), C het aantal getelde kolonies op de plaat en N de
log KVE/g. De berekening van het groeipotentieel (δ) gebeurt door het verschil te nemen tussen
de mediaan op de laatste dag van de challengetest en de mediaan op dag 0. Een verschil groter
dan 0,5 log KVE/g verwijst naar een significante groei van Listeria monocytogenes.
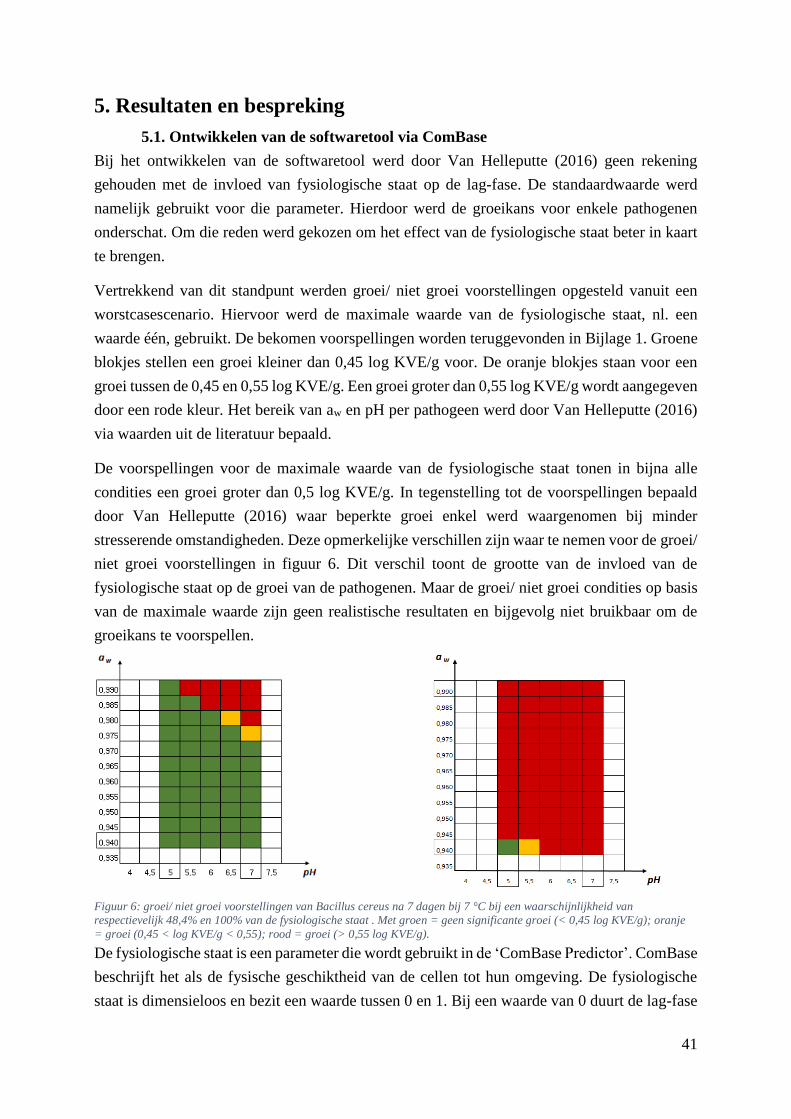
41
5. Resultaten en bespreking
5.1. Ontwikkelen van de softwaretool via ComBase
Bij het ontwikkelen van de softwaretool werd door Van Helleputte (2016) geen rekening
gehouden met de invloed van fysiologische staat op de lag-fase. De standaardwaarde werd
namelijk gebruikt voor die parameter. Hierdoor werd de groeikans voor enkele pathogenen
onderschat. Om die reden werd gekozen om het effect van de fysiologische staat beter in kaart
te brengen.
Vertrekkend van dit standpunt werden groei/ niet groei voorstellingen opgesteld vanuit een
worstcasescenario. Hiervoor werd de maximale waarde van de fysiologische staat, nl. een
waarde één, gebruikt. De bekomen voorspellingen worden teruggevonden in Bijlage 1. Groene
blokjes stellen een groei kleiner dan 0,45 log KVE/g voor. De oranje blokjes staan voor een
groei tussen de 0,45 en 0,55 log KVE/g. Een groei groter dan 0,55 log KVE/g wordt aangegeven
door een rode kleur. Het bereik van aw en pH per pathogeen werd door Van Helleputte (2016)
via waarden uit de literatuur bepaald.
De voorspellingen voor de maximale waarde van de fysiologische staat tonen in bijna alle
condities een groei groter dan 0,5 log KVE/g. In tegenstelling tot de voorspellingen bepaald
door Van Helleputte (2016) waar beperkte groei enkel werd waargenomen bij minder
stresserende omstandigheden. Deze opmerkelijke verschillen zijn waar te nemen voor de groei/
niet groei voorstellingen in figuur 6. Dit verschil toont de grootte van de invloed van de
fysiologische staat op de groei van de pathogenen. Maar de groei/ niet groei condities op basis
van de maximale waarde zijn geen realistische resultaten en bijgevolg niet bruikbaar om de
groeikans te voorspellen.
De fysiologische staat is een parameter die wordt gebruikt in de ‘ComBase Predictor’. ComBase
beschrijft het als de fysische geschiktheid van de cellen tot hun omgeving. De fysiologische
staat is dimensieloos en bezit een waarde tussen 0 en 1. Bij een waarde van 0 duurt de lag-fase
Figuur 6: groei/ niet groei voorstellingen van Bacillus cereus na 7 dagen bij 7 °C bij een waarschijnlijkheid van
respectievelijk 48,4% en 100% van de fysiologische staat . Met groen = geen significante groei (< 0,45 log KVE/g); oranje
= groei (0,45 < log KVE/g < 0,55); rood = groei (> 0,55 log KVE/g).
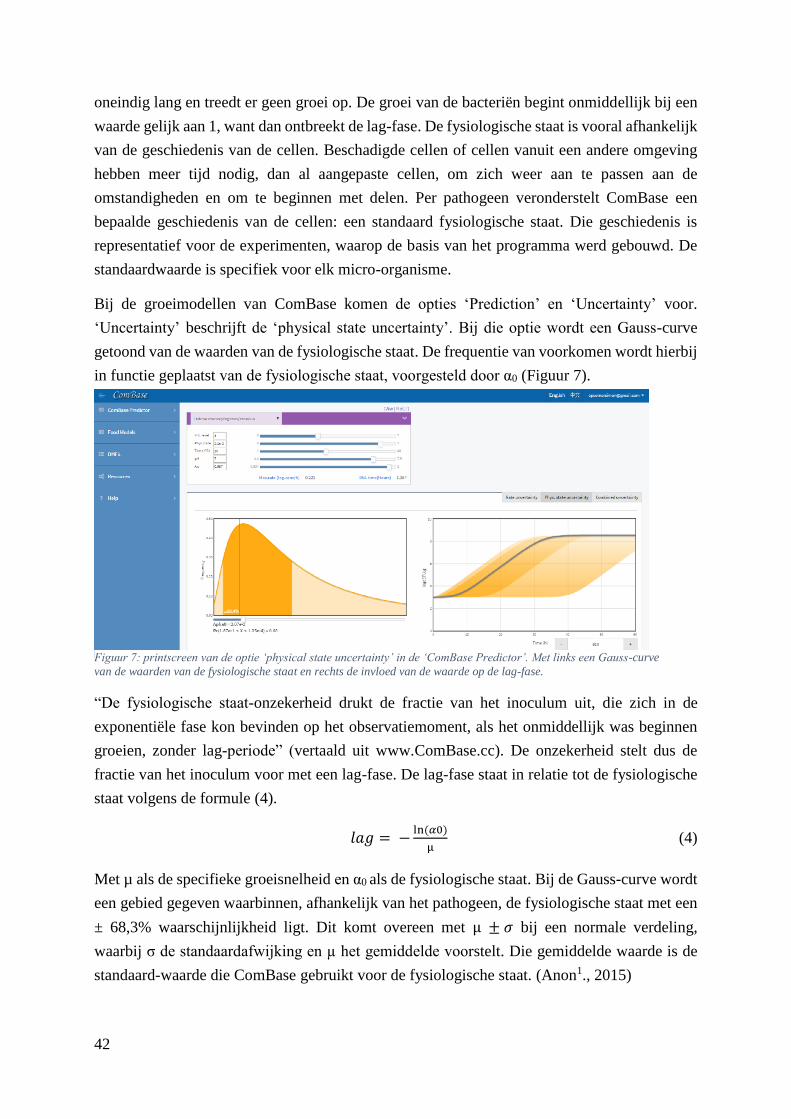
42
oneindig lang en treedt er geen groei op. De groei van de bacteriën begint onmiddellijk bij een
waarde gelijk aan 1, want dan ontbreekt de lag-fase. De fysiologische staat is vooral afhankelijk
van de geschiedenis van de cellen. Beschadigde cellen of cellen vanuit een andere omgeving
hebben meer tijd nodig, dan al aangepaste cellen, om zich weer aan te passen aan de
omstandigheden en om te beginnen met delen. Per pathogeen veronderstelt ComBase een
bepaalde geschiedenis van de cellen: een standaard fysiologische staat. Die geschiedenis is
representatief voor de experimenten, waarop de basis van het programma werd gebouwd. De
standaardwaarde is specifiek voor elk micro-organisme.
Bij de groeimodellen van ComBase komen de opties ‘Prediction’ en ‘Uncertainty’ voor.
‘Uncertainty’ beschrijft de ‘physical state uncertainty’. Bij die optie wordt een Gauss-curve
getoond van de waarden van de fysiologische staat. De frequentie van voorkomen wordt hierbij
in functie geplaatst van de fysiologische staat, voorgesteld door α0 (Figuur 7).
“De fysiologische staat-onzekerheid drukt de fractie van het inoculum uit, die zich in de
exponentiële fase kon bevinden op het observatiemoment, als het onmiddellijk was beginnen
groeien, zonder lag-periode” (vertaald uit www.ComBase.cc). De onzekerheid stelt dus de
fractie van het inoculum voor met een lag-fase. De lag-fase staat in relatie tot de fysiologische
staat volgens de formule (4).
𝑙𝑎𝑔 = −ln (𝛼0)
µ (4)
Met µ als de specifieke groeisnelheid en α0 als de fysiologische staat. Bij de Gauss-curve wordt
een gebied gegeven waarbinnen, afhankelijk van het pathogeen, de fysiologische staat met een
± 68,3% waarschijnlijkheid ligt. Dit komt overeen met µ ± 𝜎 bij een normale verdeling,
waarbij σ de standaardafwijking en µ het gemiddelde voorstelt. Die gemiddelde waarde is de
standaard-waarde die ComBase gebruikt voor de fysiologische staat. (Anon1., 2015)
Figuur 7: printscreen van de optie ‘physical state uncertainty’ in de ‘ComBase Predictor’. Met links een Gauss-curve
van de waarden van de fysiologische staat en rechts de invloed van de waarde op de lag-fase.

43
De Gauss-curve wordt cumulatief benaderd vanwege een aflopende x-as. Hierdoor kan de
fysiologische staat met een waarschijnlijkheid van ± 48,4% in het gebied van de
standaardwaarde zitten. Met een waarschijnlijkheid van ± 82,5% bevindt α0 zich in het gebied
µ + 𝜎. De fysiologische staat kan met een waarschijnlijkheid van ± 95,5% in het gebied µ ±
2𝜎 liggen. Door die benadering moeten de percentages ook anders worden gelezen. De 48,4 %,
82,5% en 95,5% stellen de grootte van de fracties ten opzichte van het inoculum voor zonder
een lag-fase. Daarbij komt 100% overeen met een fysiologische staat gelijk aan 1, wat
onmiddellijke groei betekent. De groei/ niet groei condities voor de 95,5% en 82,5% fracties
zijn respectievelijk terug te vinden in Bijlage 2 en 3.
De bekomen voorstellingen, gebaseerd op de groeimodellen van ComBase, zijn volgens
voorspelde gegevens bepaald. Zoals eerder aangehaald in de literatuur kloppen de
voorspellingen niet altijd met de resultaten bekomen in de praktijk. Daarom werden de groei/
niet groei voorstellingen vergeleken met waargenomen data.
5.2. Groei/ geen groei voorstellingen gevormd via Microbial Responses Viewer
De website MRV, afgeleid uit ComBase, bestaat uit dezelfde database. De website biedt op een
snelle manier een beeld van de mogelijke groei van een bepaald micro-organisme. Met die data
worden groei/ niet groei vaststellignen opgemaakt, die te vinden zijn in Bijlage 4. De grafieken
tonen het aantal significante groeigevallen ten opzichte van het totaal aantal gevallen.
Waargenomen significante groei voor een bepaalde combinatie, wordt in het rood aangegeven.
Als bij geen enkele observatie, significante groei wordt geobserveerd, wordt de pH/aw-
combinatie in het wit voorgesteld. Maar niet bij elke pathogeen werden veel gegevens
aangetroffen voor de gewenste pH/aw-combinaties. Hierdoor was het niet mogelijk om voor
elke pathogeen conclusies te trekken. Met de veronderstelling dat de database van ComBase
zelf meer gegevens zou bevatten, werd die verder geraadpleegd.
5.3. Groei/ niet groei grafieken gevormd via de database van ComBase
Via www.ComBase.cc kan ook de database geconsulteerd worden waarop de modellen van
ComBase berust zijn. Die database werd geraadpleegd om de groei/ geen groei interfaces op te
maken. Voor sommige pH/aw-combinaties zijn meerdere experimenten uitgevoerd, die elkaar
soms tegenspreken. De voorspellingen tonen het aantal gevallen groei ten opzichte van het
totaal aantal gevallen. Vanaf groei wordt waargenomen, bij één van de experimenten voor een
bepaalde combinatie, wordt de pH/aw-combinatie in het rood voorgesteld. Wanneer bij geen
enkele observatie significante groei wordt geobserveerd, wordt de pH/aw-combinatie in het wit
geplaatst. De bekomen groei/ niet groei grafieken worden in ‘4.4. Groei/ geen groei
voorstellingen vergelijken’ getoond en worden teruggevonden in Bijlage 5.
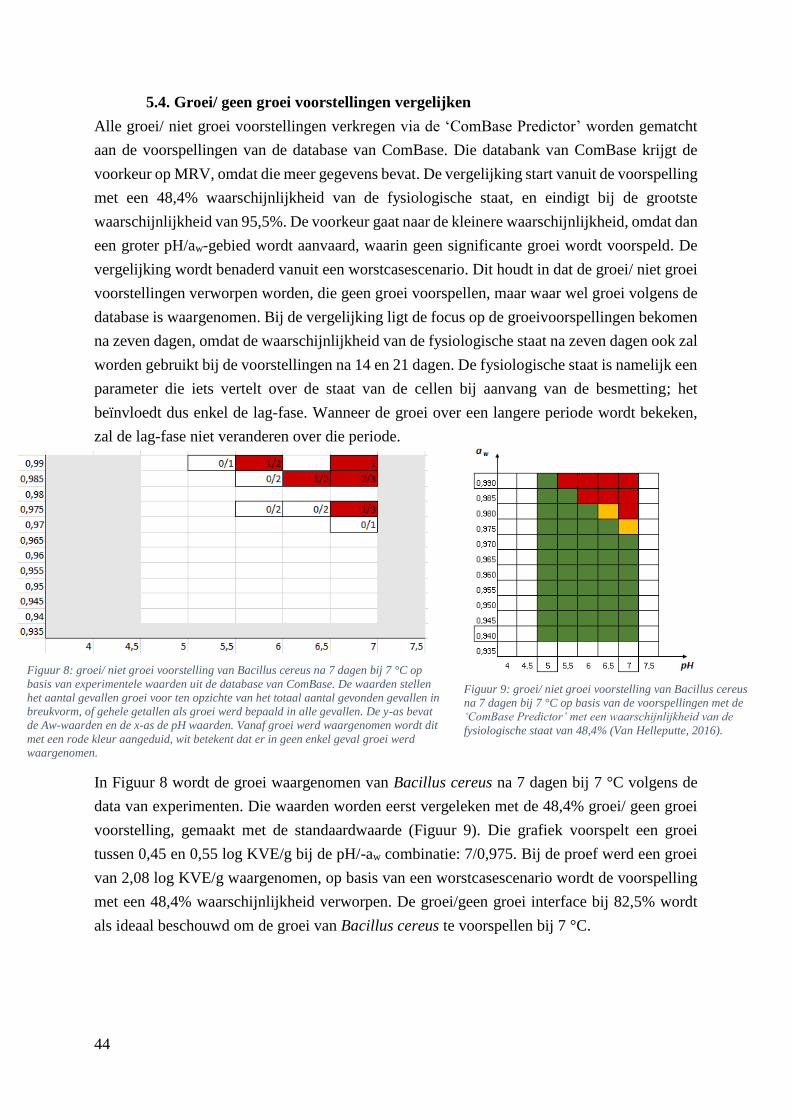
44
5.4. Groei/ geen groei voorstellingen vergelijken
Alle groei/ niet groei voorstellingen verkregen via de ‘ComBase Predictor’ worden gematcht
aan de voorspellingen van de database van ComBase. Die databank van ComBase krijgt de
voorkeur op MRV, omdat die meer gegevens bevat. De vergelijking start vanuit de voorspelling
met een 48,4% waarschijnlijkheid van de fysiologische staat, en eindigt bij de grootste
waarschijnlijkheid van 95,5%. De voorkeur gaat naar de kleinere waarschijnlijkheid, omdat dan
een groter pH/aw-gebied wordt aanvaard, waarin geen significante groei wordt voorspeld. De
vergelijking wordt benaderd vanuit een worstcasescenario. Dit houdt in dat de groei/ niet groei
voorstellingen verworpen worden, die geen groei voorspellen, maar waar wel groei volgens de
database is waargenomen. Bij de vergelijking ligt de focus op de groeivoorspellingen bekomen
na zeven dagen, omdat de waarschijnlijkheid van de fysiologische staat na zeven dagen ook zal
worden gebruikt bij de voorstellingen na 14 en 21 dagen. De fysiologische staat is namelijk een
parameter die iets vertelt over de staat van de cellen bij aanvang van de besmetting; het
beïnvloedt dus enkel de lag-fase. Wanneer de groei over een langere periode wordt bekeken,
zal de lag-fase niet veranderen over die periode.
In Figuur 8 wordt de groei waargenomen van Bacillus cereus na 7 dagen bij 7 °C volgens de
data van experimenten. Die waarden worden eerst vergeleken met de 48,4% groei/ geen groei
voorstelling, gemaakt met de standaardwaarde (Figuur 9). Die grafiek voorspelt een groei
tussen 0,45 en 0,55 log KVE/g bij de pH/-aw combinatie: 7/0,975. Bij de proef werd een groei
van 2,08 log KVE/g waargenomen, op basis van een worstcasescenario wordt de voorspelling
met een 48,4% waarschijnlijkheid verworpen. De groei/geen groei interface bij 82,5% wordt
als ideaal beschouwd om de groei van Bacillus cereus te voorspellen bij 7 °C.
Figuur 8: groei/ niet groei voorstelling van Bacillus cereus na 7 dagen bij 7 °C op
basis van experimentele waarden uit de database van ComBase. De waarden stellen
het aantal gevallen groei voor ten opzichte van het totaal aantal gevonden gevallen in
breukvorm, of gehele getallen als groei werd bepaald in alle gevallen. De y-as bevat
de Aw-waarden en de x-as de pH waarden. Vanaf groei werd waargenomen wordt dit
met een rode kleur aangeduid, wit betekent dat er in geen enkel geval groei werd
waargenomen.
Figuur 9: groei/ niet groei voorstelling van Bacillus cereus
na 7 dagen bij 7 °C op basis van de voorspellingen met de
‘ComBase Predictor’ met een waarschijnlijkheid van de
fysiologische staat van 48,4% (Van Helleputte, 2016).
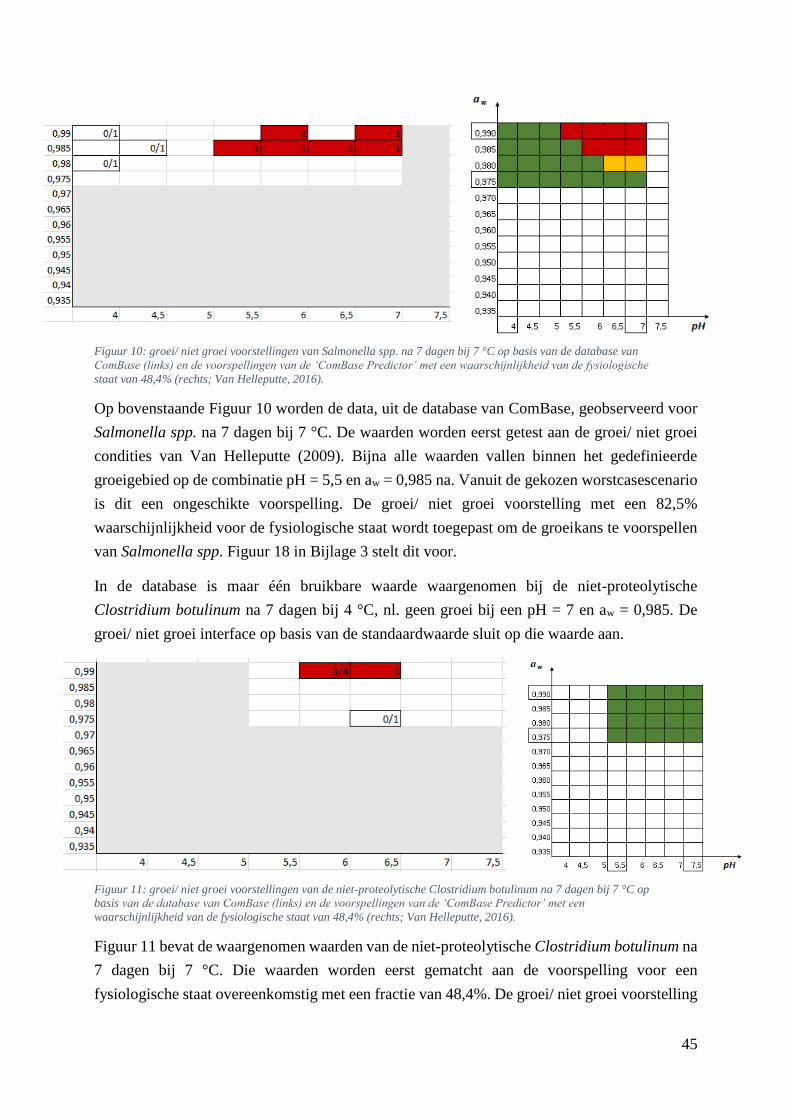
45
Op bovenstaande Figuur 10 worden de data, uit de database van ComBase, geobserveerd voor
Salmonella spp. na 7 dagen bij 7 °C. De waarden worden eerst getest aan de groei/ niet groei
condities van Van Helleputte (2009). Bijna alle waarden vallen binnen het gedefinieerde
groeigebied op de combinatie pH = 5,5 en aw = 0,985 na. Vanuit de gekozen worstcasescenario
is dit een ongeschikte voorspelling. De groei/ niet groei voorstelling met een 82,5%
waarschijnlijkheid voor de fysiologische staat wordt toegepast om de groeikans te voorspellen
van Salmonella spp. Figuur 18 in Bijlage 3 stelt dit voor.
In de database is maar één bruikbare waarde waargenomen bij de niet-proteolytische
Clostridium botulinum na 7 dagen bij 4 °C, nl. geen groei bij een pH = 7 en aw = 0,985. De
groei/ niet groei interface op basis van de standaardwaarde sluit op die waarde aan.
Figuur 11 bevat de waargenomen waarden van de niet-proteolytische Clostridium botulinum na
7 dagen bij 7 °C. Die waarden worden eerst gematcht aan de voorspelling voor een
fysiologische staat overeenkomstig met een fractie van 48,4%. De groei/ niet groei voorstelling
Figuur 10: groei/ niet groei voorstellingen van Salmonella spp. na 7 dagen bij 7 °C op basis van de database van
ComBase (links) en de voorspellingen van de ‘ComBase Predictor’ met een waarschijnlijkheid van de fysiologische
staat van 48,4% (rechts; Van Helleputte, 2016).
Figuur 11: groei/ niet groei voorstellingen van de niet-proteolytische Clostridium botulinum na 7 dagen bij 7 °C op
basis van de database van ComBase (links) en de voorspellingen van de ‘ComBase Predictor’ met een
waarschijnlijkheid van de fysiologische staat van 48,4% (rechts; Van Helleputte, 2016).
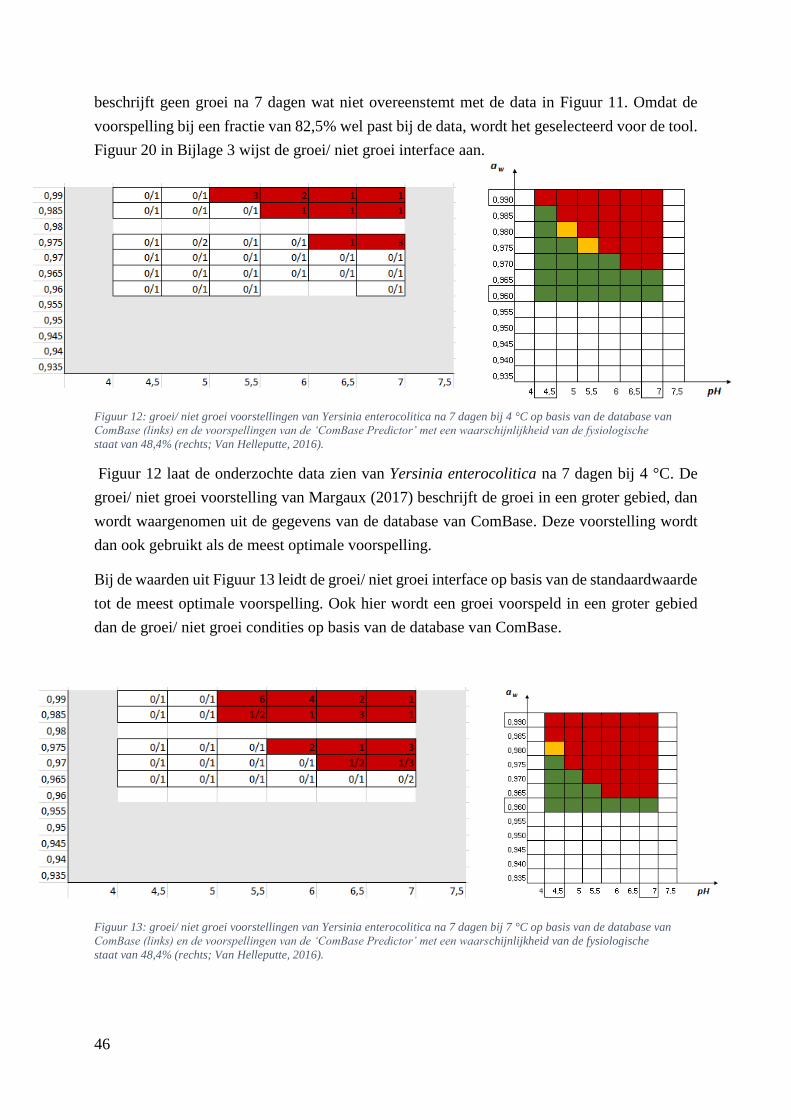
46
beschrijft geen groei na 7 dagen wat niet overeenstemt met de data in Figuur 11. Omdat de
voorspelling bij een fractie van 82,5% wel past bij de data, wordt het geselecteerd voor de tool.
Figuur 20 in Bijlage 3 wijst de groei/ niet groei interface aan.
Figuur 12 laat de onderzochte data zien van Yersinia enterocolitica na 7 dagen bij 4 °C. De
groei/ niet groei voorstelling van Margaux (2017) beschrijft de groei in een groter gebied, dan
wordt waargenomen uit de gegevens van de database van ComBase. Deze voorstelling wordt
dan ook gebruikt als de meest optimale voorspelling.
Bij de waarden uit Figuur 13 leidt de groei/ niet groei interface op basis van de standaardwaarde
tot de meest optimale voorspelling. Ook hier wordt een groei voorspeld in een groter gebied
dan de groei/ niet groei condities op basis van de database van ComBase.
Figuur 12: groei/ niet groei voorstellingen van Yersinia enterocolitica na 7 dagen bij 4 °C op basis van de database van
ComBase (links) en de voorspellingen van de ‘ComBase Predictor’ met een waarschijnlijkheid van de fysiologische
staat van 48,4% (rechts; Van Helleputte, 2016).
Figuur 13: groei/ niet groei voorstellingen van Yersinia enterocolitica na 7 dagen bij 7 °C op basis van de database van
ComBase (links) en de voorspellingen van de ‘ComBase Predictor’ met een waarschijnlijkheid van de fysiologische
staat van 48,4% (rechts; Van Helleputte, 2016).
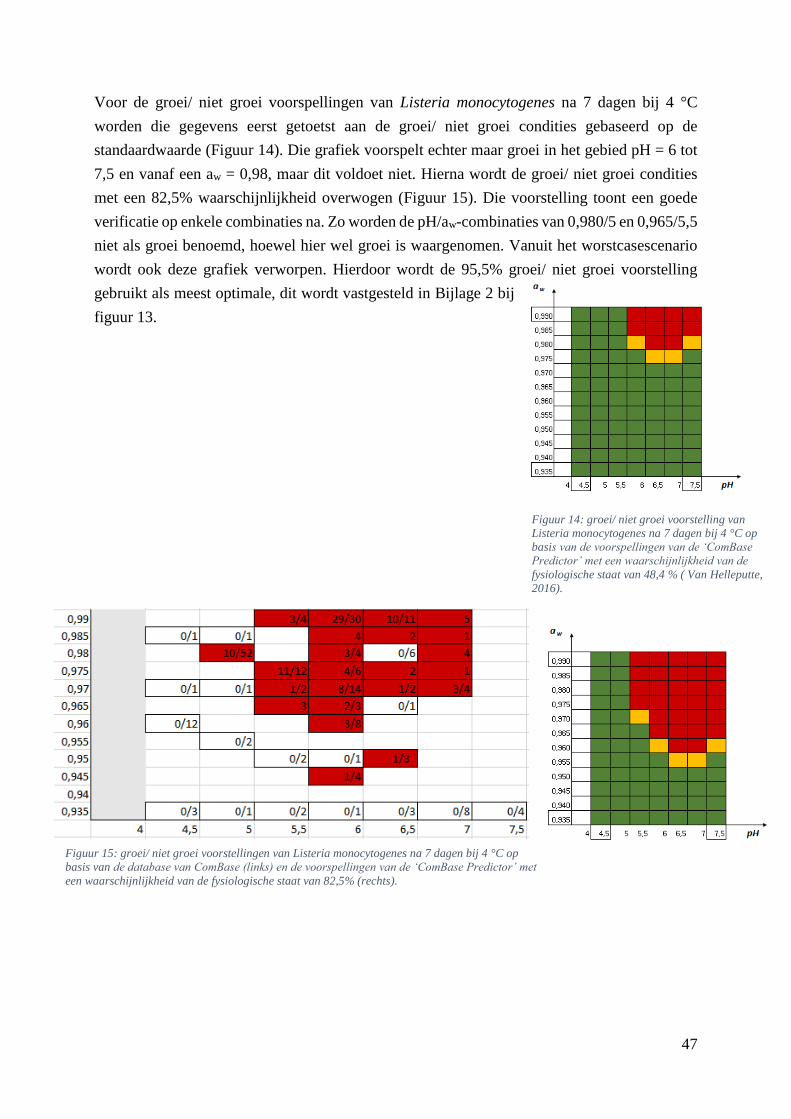
47
Voor de groei/ niet groei voorspellingen van Listeria monocytogenes na 7 dagen bij 4 °C
worden die gegevens eerst getoetst aan de groei/ niet groei condities gebaseerd op de
standaardwaarde (Figuur 14). Die grafiek voorspelt echter maar groei in het gebied pH = 6 tot
7,5 en vanaf een aw = 0,98, maar dit voldoet niet. Hierna wordt de groei/ niet groei condities
met een 82,5% waarschijnlijkheid overwogen (Figuur 15). Die voorstelling toont een goede
verificatie op enkele combinaties na. Zo worden de pH/aw-combinaties van 0,980/5 en 0,965/5,5
niet als groei benoemd, hoewel hier wel groei is waargenomen. Vanuit het worstcasescenario
wordt ook deze grafiek verworpen. Hierdoor wordt de 95,5% groei/ niet groei voorstelling
gebruikt als meest optimale, dit wordt vastgesteld in Bijlage 2 bij
figuur 13.
Figuur 15: groei/ niet groei voorstellingen van Listeria monocytogenes na 7 dagen bij 4 °C op
basis van de database van ComBase (links) en de voorspellingen van de ‘ComBase Predictor’ met
een waarschijnlijkheid van de fysiologische staat van 82,5% (rechts).
Figuur 14: groei/ niet groei voorstelling van
Listeria monocytogenes na 7 dagen bij 4 °C op
basis van de voorspellingen van de ‘ComBase
Predictor’ met een waarschijnlijkheid van de
fysiologische staat van 48,4 % ( Van Helleputte,
2016).
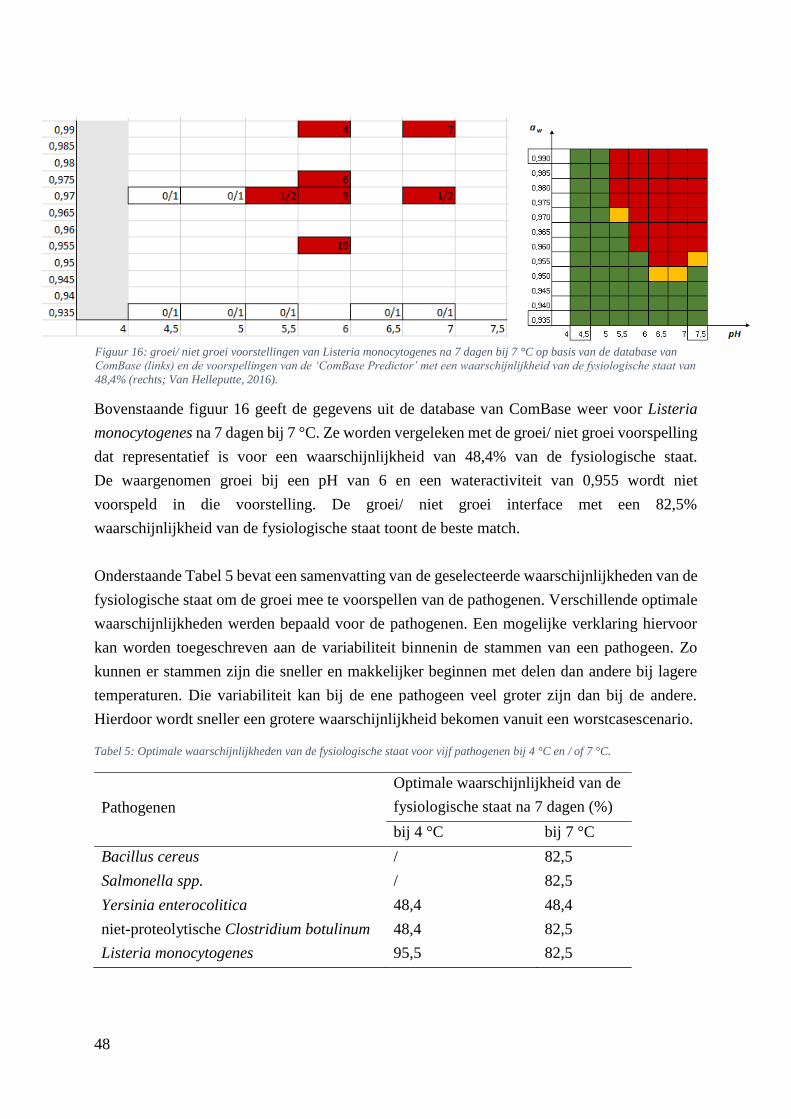
48
Bovenstaande figuur 16 geeft de gegevens uit de database van ComBase weer voor Listeria
monocytogenes na 7 dagen bij 7 °C. Ze worden vergeleken met de groei/ niet groei voorspelling
dat representatief is voor een waarschijnlijkheid van 48,4% van de fysiologische staat.
De waargenomen groei bij een pH van 6 en een wateractiviteit van 0,955 wordt niet
voorspeld in die voorstelling. De groei/ niet groei interface met een 82,5%
waarschijnlijkheid van de fysiologische staat toont de beste match.
Onderstaande Tabel 5 bevat een samenvatting van de geselecteerde waarschijnlijkheden van de
fysiologische staat om de groei mee te voorspellen van de pathogenen. Verschillende optimale
waarschijnlijkheden werden bepaald voor de pathogenen. Een mogelijke verklaring hiervoor
kan worden toegeschreven aan de variabiliteit binnenin de stammen van een pathogeen. Zo
kunnen er stammen zijn die sneller en makkelijker beginnen met delen dan andere bij lagere
temperaturen. Die variabiliteit kan bij de ene pathogeen veel groter zijn dan bij de andere.
Hierdoor wordt sneller een grotere waarschijnlijkheid bekomen vanuit een worstcasescenario.
Tabel 5: Optimale waarschijnlijkheden van de fysiologische staat voor vijf pathogenen bij 4 °C en / of 7 °C.
Pathogenen
Optimale waarschijnlijkheid van de
fysiologische staat na 7 dagen (%)
bij 4 °C bij 7 °C
Bacillus cereus / 82,5
Salmonella spp. / 82,5
Yersinia enterocolitica 48,4 48,4
niet-proteolytische Clostridium botulinum 48,4 82,5
Listeria monocytogenes 95,5 82,5
Figuur 16: groei/ niet groei voorstellingen van Listeria monocytogenes na 7 dagen bij 7 °C op basis van de database van
ComBase (links) en de voorspellingen van de ‘ComBase Predictor’ met een waarschijnlijkheid van de fysiologische staat van
48,4% (rechts; Van Helleputte, 2016).
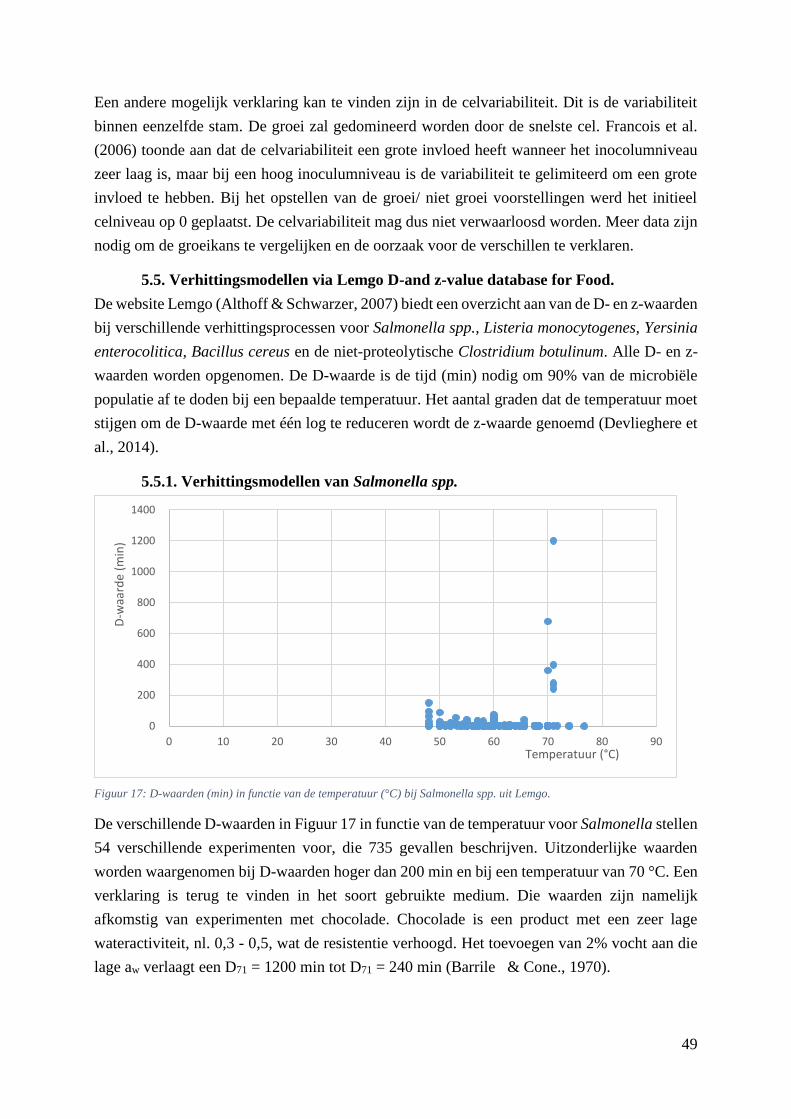
49
Een andere mogelijk verklaring kan te vinden zijn in de celvariabiliteit. Dit is de variabiliteit
binnen eenzelfde stam. De groei zal gedomineerd worden door de snelste cel. Francois et al.
(2006) toonde aan dat de celvariabiliteit een grote invloed heeft wanneer het inocolumniveau
zeer laag is, maar bij een hoog inoculumniveau is de variabiliteit te gelimiteerd om een grote
invloed te hebben. Bij het opstellen van de groei/ niet groei voorstellingen werd het initieel
celniveau op 0 geplaatst. De celvariabiliteit mag dus niet verwaarloosd worden. Meer data zijn
nodig om de groeikans te vergelijken en de oorzaak voor de verschillen te verklaren.
5.5. Verhittingsmodellen via Lemgo D-and z-value database for Food.
De website Lemgo (Althoff & Schwarzer, 2007) biedt een overzicht aan van de D- en z-waarden
bij verschillende verhittingsprocessen voor Salmonella spp., Listeria monocytogenes, Yersinia
enterocolitica, Bacillus cereus en de niet-proteolytische Clostridium botulinum. Alle D- en z-
waarden worden opgenomen. De D-waarde is de tijd (min) nodig om 90% van de microbiële
populatie af te doden bij een bepaalde temperatuur. Het aantal graden dat de temperatuur moet
stijgen om de D-waarde met één log te reduceren wordt de z-waarde genoemd (Devlieghere et
al., 2014).
5.5.1. Verhittingsmodellen van Salmonella spp.
De verschillende D-waarden in Figuur 17 in functie van de temperatuur voor Salmonella stellen
54 verschillende experimenten voor, die 735 gevallen beschrijven. Uitzonderlijke waarden
worden waargenomen bij D-waarden hoger dan 200 min en bij een temperatuur van 70 °C. Een
verklaring is terug te vinden in het soort gebruikte medium. Die waarden zijn namelijk
afkomstig van experimenten met chocolade. Chocolade is een product met een zeer lage
wateractiviteit, nl. 0,3 - 0,5, wat de resistentie verhoogd. Het toevoegen van 2% vocht aan die
lage aw verlaagt een D71 = 1200 min tot D71 = 240 min (Barrile & Cone., 1970).
0
200
400
600
800
1000
1200
1400
0 10 20 30 40 50 60 70 80 90
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 17: D-waarden (min) in functie van de temperatuur (°C) bij Salmonella spp. uit Lemgo.
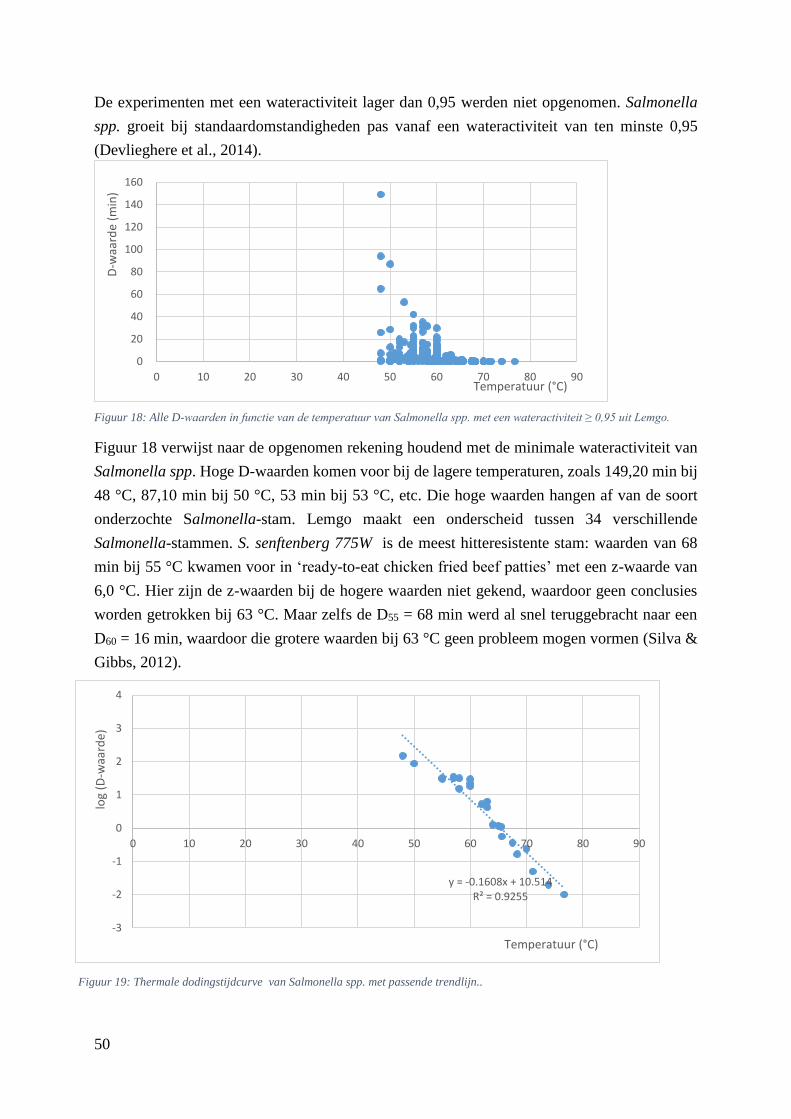
50
De experimenten met een wateractiviteit lager dan 0,95 werden niet opgenomen. Salmonella
spp. groeit bij standaardomstandigheden pas vanaf een wateractiviteit van ten minste 0,95
(Devlieghere et al., 2014).
Figuur 18 verwijst naar de opgenomen rekening houdend met de minimale wateractiviteit van
Salmonella spp. Hoge D-waarden komen voor bij de lagere temperaturen, zoals 149,20 min bij
48 °C, 87,10 min bij 50 °C, 53 min bij 53 °C, etc. Die hoge waarden hangen af van de soort
onderzochte Salmonella-stam. Lemgo maakt een onderscheid tussen 34 verschillende
Salmonella-stammen. S. senftenberg 775W is de meest hitteresistente stam: waarden van 68
min bij 55 °C kwamen voor in ‘ready-to-eat chicken fried beef patties’ met een z-waarde van
6,0 °C. Hier zijn de z-waarden bij de hogere waarden niet gekend, waardoor geen conclusies
worden getrokken bij 63 °C. Maar zelfs de D55 = 68 min werd al snel teruggebracht naar een
D60 = 16 min, waardoor die grotere waarden bij 63 °C geen probleem mogen vormen (Silva &
Gibbs, 2012).
y = -0.1608x + 10.514R² = 0.9255
-3
-2
-1
0
1
2
3
4
0 10 20 30 40 50 60 70 80 90
log
(D-w
aard
e)
Temperatuur (°C)
0
20
40
60
80
100
120
140
160
0 10 20 30 40 50 60 70 80 90
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 18: Alle D-waarden in functie van de temperatuur van Salmonella spp. met een wateractiviteit ≥ 0,95 uit Lemgo.
Figuur 19: Thermale dodingstijdcurve van Salmonella spp. met passende trendlijn..
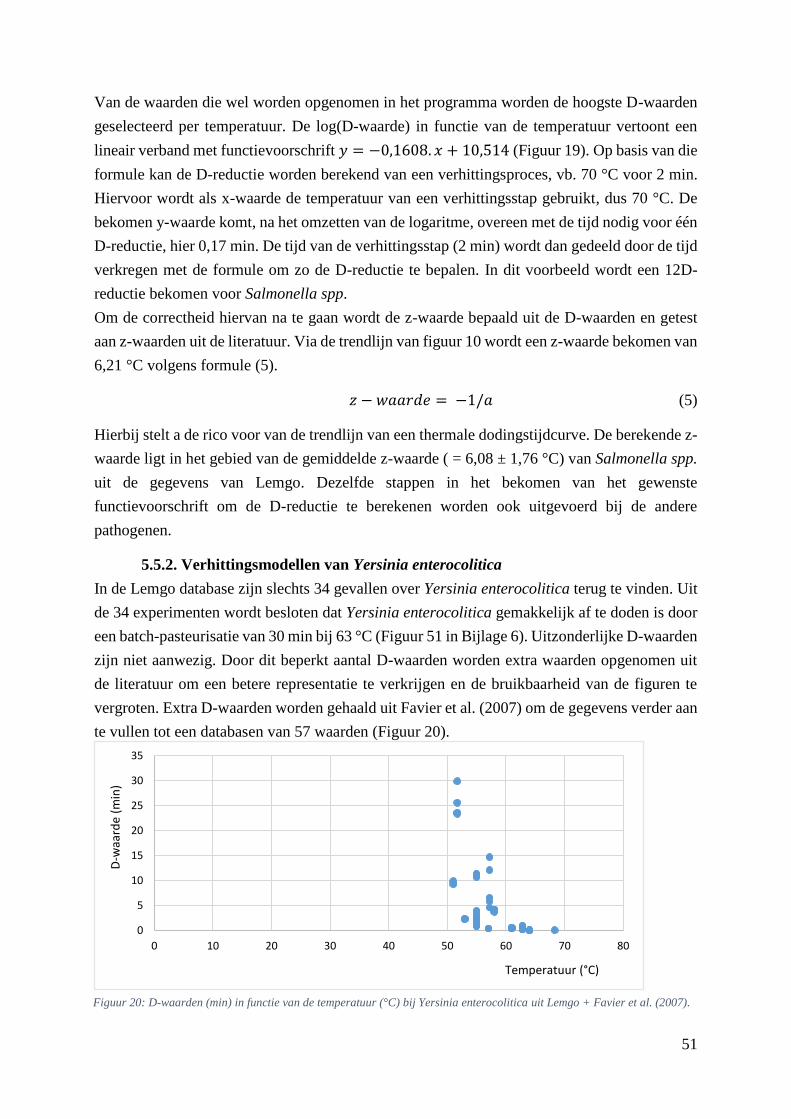
51
Van de waarden die wel worden opgenomen in het programma worden de hoogste D-waarden
geselecteerd per temperatuur. De log(D-waarde) in functie van de temperatuur vertoont een
lineair verband met functievoorschrift 𝑦 = −0,1608. 𝑥 + 10,514 (Figuur 19). Op basis van die
formule kan de D-reductie worden berekend van een verhittingsproces, vb. 70 °C voor 2 min.
Hiervoor wordt als x-waarde de temperatuur van een verhittingsstap gebruikt, dus 70 °C. De
bekomen y-waarde komt, na het omzetten van de logaritme, overeen met de tijd nodig voor één
D-reductie, hier 0,17 min. De tijd van de verhittingsstap (2 min) wordt dan gedeeld door de tijd
verkregen met de formule om zo de D-reductie te bepalen. In dit voorbeeld wordt een 12D-
reductie bekomen voor Salmonella spp.
Om de correctheid hiervan na te gaan wordt de z-waarde bepaald uit de D-waarden en getest
aan z-waarden uit de literatuur. Via de trendlijn van figuur 10 wordt een z-waarde bekomen van
6,21 °C volgens formule (5).
𝑧 − 𝑤𝑎𝑎𝑟𝑑𝑒 = −1/𝑎 (5)
Hierbij stelt a de rico voor van de trendlijn van een thermale dodingstijdcurve. De berekende z-
waarde ligt in het gebied van de gemiddelde z-waarde ( = 6,08 ± 1,76 °C) van Salmonella spp.
uit de gegevens van Lemgo. Dezelfde stappen in het bekomen van het gewenste
functievoorschrift om de D-reductie te berekenen worden ook uitgevoerd bij de andere
pathogenen.
5.5.2. Verhittingsmodellen van Yersinia enterocolitica
In de Lemgo database zijn slechts 34 gevallen over Yersinia enterocolitica terug te vinden. Uit
de 34 experimenten wordt besloten dat Yersinia enterocolitica gemakkelijk af te doden is door
een batch-pasteurisatie van 30 min bij 63 °C (Figuur 51 in Bijlage 6). Uitzonderlijke D-waarden
zijn niet aanwezig. Door dit beperkt aantal D-waarden worden extra waarden opgenomen uit
de literatuur om een betere representatie te verkrijgen en de bruikbaarheid van de figuren te
vergroten. Extra D-waarden worden gehaald uit Favier et al. (2007) om de gegevens verder aan
te vullen tot een databasen van 57 waarden (Figuur 20).
0
5
10
15
20
25
30
35
0 10 20 30 40 50 60 70 80
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 20: D-waarden (min) in functie van de temperatuur (°C) bij Yersinia enterocolitica uit Lemgo + Favier et al. (2007).
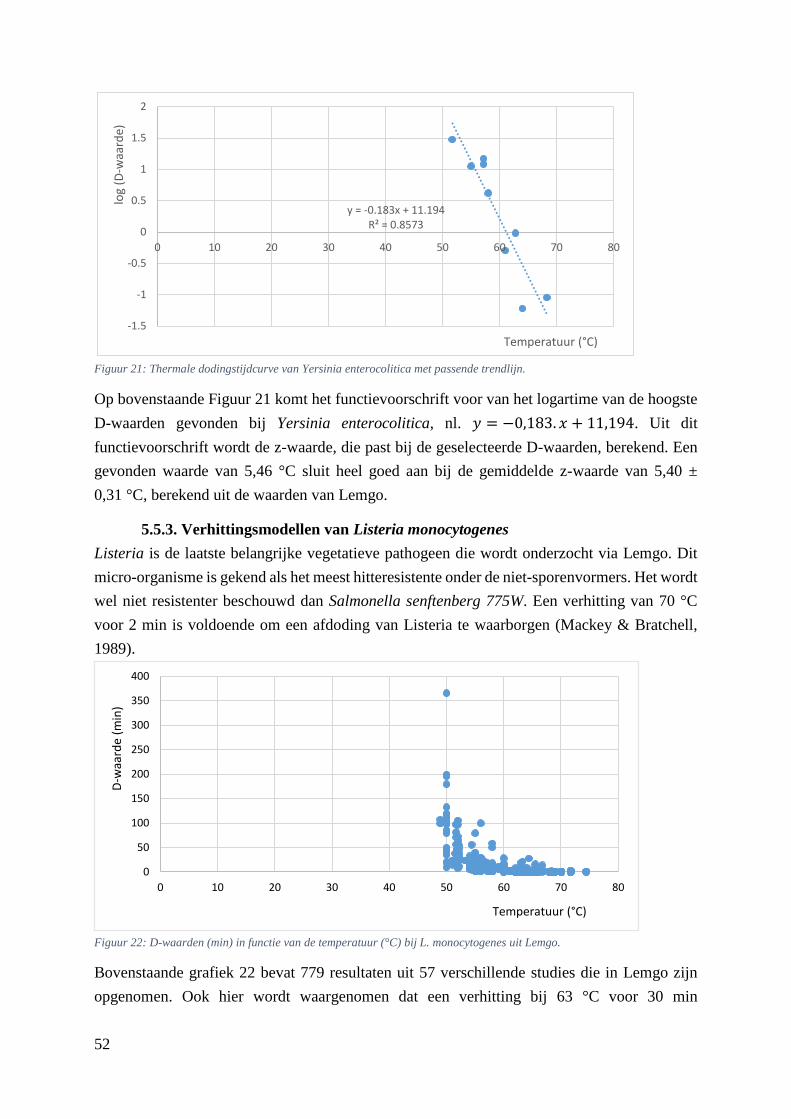
52
Op bovenstaande Figuur 21 komt het functievoorschrift voor van het logartime van de hoogste
D-waarden gevonden bij Yersinia enterocolitica, nl. 𝑦 = −0,183. 𝑥 + 11,194. Uit dit
functievoorschrift wordt de z-waarde, die past bij de geselecteerde D-waarden, berekend. Een
gevonden waarde van 5,46 °C sluit heel goed aan bij de gemiddelde z-waarde van 5,40 ±
0,31 °C, berekend uit de waarden van Lemgo.
5.5.3. Verhittingsmodellen van Listeria monocytogenes
Listeria is de laatste belangrijke vegetatieve pathogeen die wordt onderzocht via Lemgo. Dit
micro-organisme is gekend als het meest hitteresistente onder de niet-sporenvormers. Het wordt
wel niet resistenter beschouwd dan Salmonella senftenberg 775W. Een verhitting van 70 °C
voor 2 min is voldoende om een afdoding van Listeria te waarborgen (Mackey & Bratchell,
1989).
Bovenstaande grafiek 22 bevat 779 resultaten uit 57 verschillende studies die in Lemgo zijn
opgenomen. Ook hier wordt waargenomen dat een verhitting bij 63 °C voor 30 min
y = -0.183x + 11.194R² = 0.8573
-1.5
-1
-0.5
0
0.5
1
1.5
2
0 10 20 30 40 50 60 70 80
log
(D-w
aard
e)
Temperatuur (°C)
0
50
100
150
200
250
300
350
400
0 10 20 30 40 50 60 70 80
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 21: Thermale dodingstijdcurve van Yersinia enterocolitica met passende trendlijn.
Figuur 22: D-waarden (min) in functie van de temperatuur (°C) bij L. monocytogenes uit Lemgo.
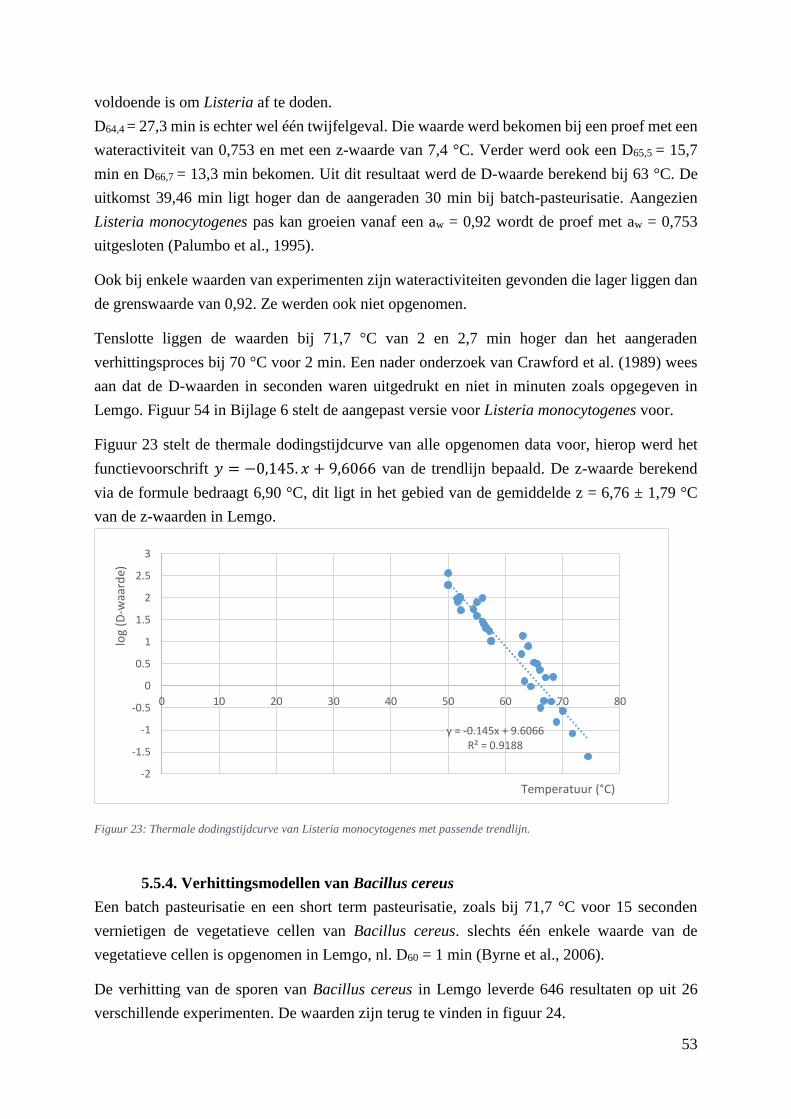
53
voldoende is om Listeria af te doden.
D64,4 = 27,3 min is echter wel één twijfelgeval. Die waarde werd bekomen bij een proef met een
wateractiviteit van 0,753 en met een z-waarde van 7,4 °C. Verder werd ook een D65,5 = 15,7
min en D66,7 = 13,3 min bekomen. Uit dit resultaat werd de D-waarde berekend bij 63 °C. De
uitkomst 39,46 min ligt hoger dan de aangeraden 30 min bij batch-pasteurisatie. Aangezien
Listeria monocytogenes pas kan groeien vanaf een aw = 0,92 wordt de proef met aw = 0,753
uitgesloten (Palumbo et al., 1995).
Ook bij enkele waarden van experimenten zijn wateractiviteiten gevonden die lager liggen dan
de grenswaarde van 0,92. Ze werden ook niet opgenomen.
Tenslotte liggen de waarden bij 71,7 °C van 2 en 2,7 min hoger dan het aangeraden
verhittingsproces bij 70 °C voor 2 min. Een nader onderzoek van Crawford et al. (1989) wees
aan dat de D-waarden in seconden waren uitgedrukt en niet in minuten zoals opgegeven in
Lemgo. Figuur 54 in Bijlage 6 stelt de aangepast versie voor Listeria monocytogenes voor.
Figuur 23 stelt de thermale dodingstijdcurve van alle opgenomen data voor, hierop werd het
functievoorschrift 𝑦 = −0,145. 𝑥 + 9,6066 van de trendlijn bepaald. De z-waarde berekend
via de formule bedraagt 6,90 °C, dit ligt in het gebied van de gemiddelde z = 6,76 ± 1,79 °C
van de z-waarden in Lemgo.
5.5.4. Verhittingsmodellen van Bacillus cereus
Een batch pasteurisatie en een short term pasteurisatie, zoals bij 71,7 °C voor 15 seconden
vernietigen de vegetatieve cellen van Bacillus cereus. slechts één enkele waarde van de
vegetatieve cellen is opgenomen in Lemgo, nl. D60 = 1 min (Byrne et al., 2006).
De verhitting van de sporen van Bacillus cereus in Lemgo leverde 646 resultaten op uit 26
verschillende experimenten. De waarden zijn terug te vinden in figuur 24.
y = -0.145x + 9.6066R² = 0.9188
-2
-1.5
-1
-0.5
0
0.5
1
1.5
2
2.5
3
0 10 20 30 40 50 60 70 80
log
(D-w
aard
e)
Temperatuur (°C)
Figuur 23: Thermale dodingstijdcurve van Listeria monocytogenes met passende trendlijn.
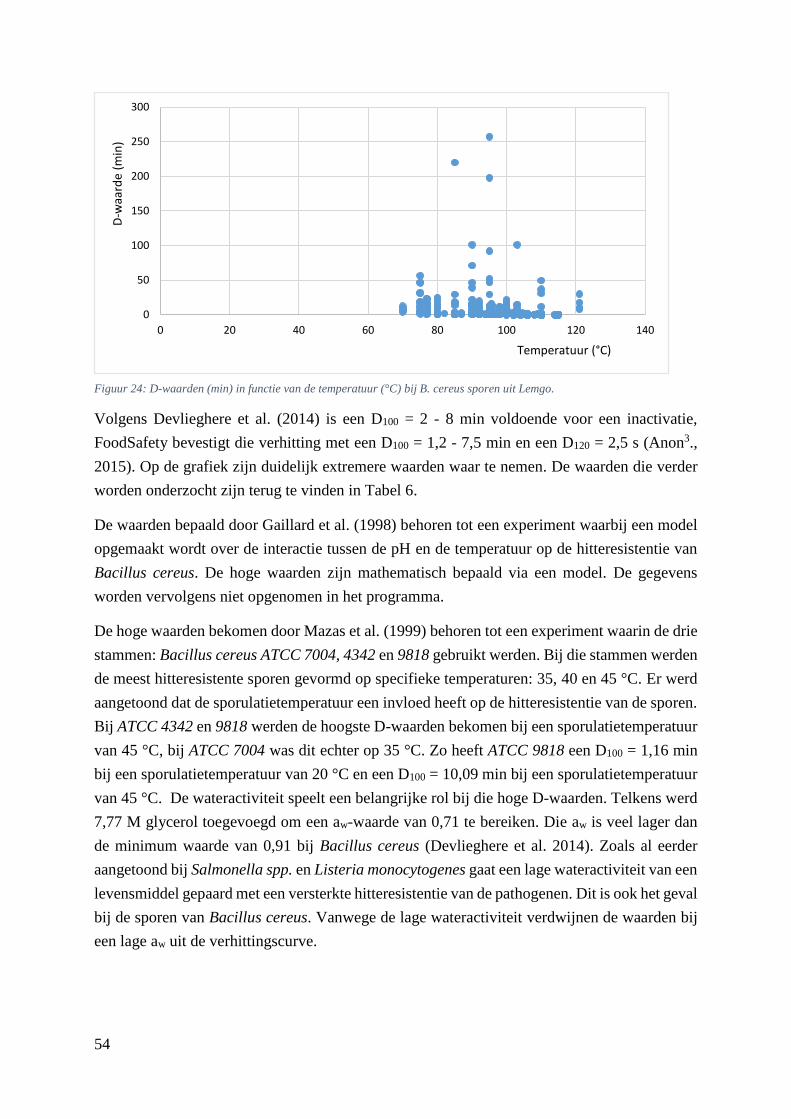
54
Volgens Devlieghere et al. (2014) is een D100 = 2 - 8 min voldoende voor een inactivatie,
FoodSafety bevestigt die verhitting met een D100 = 1,2 - 7,5 min en een D120 = 2,5 s (Anon3.,
2015). Op de grafiek zijn duidelijk extremere waarden waar te nemen. De waarden die verder
worden onderzocht zijn terug te vinden in Tabel 6.
De waarden bepaald door Gaillard et al. (1998) behoren tot een experiment waarbij een model
opgemaakt wordt over de interactie tussen de pH en de temperatuur op de hitteresistentie van
Bacillus cereus. De hoge waarden zijn mathematisch bepaald via een model. De gegevens
worden vervolgens niet opgenomen in het programma.
De hoge waarden bekomen door Mazas et al. (1999) behoren tot een experiment waarin de drie
stammen: Bacillus cereus ATCC 7004, 4342 en 9818 gebruikt werden. Bij die stammen werden
de meest hitteresistente sporen gevormd op specifieke temperaturen: 35, 40 en 45 °C. Er werd
aangetoond dat de sporulatietemperatuur een invloed heeft op de hitteresistentie van de sporen.
Bij ATCC 4342 en 9818 werden de hoogste D-waarden bekomen bij een sporulatietemperatuur
van 45 °C, bij ATCC 7004 was dit echter op 35 °C. Zo heeft ATCC 9818 een D100 = 1,16 min
bij een sporulatietemperatuur van 20 °C en een D100 = 10,09 min bij een sporulatietemperatuur
van 45 °C. De wateractiviteit speelt een belangrijke rol bij die hoge D-waarden. Telkens werd
7,77 M glycerol toegevoegd om een aw-waarde van 0,71 te bereiken. Die aw is veel lager dan
de minimum waarde van 0,91 bij Bacillus cereus (Devlieghere et al. 2014). Zoals al eerder
aangetoond bij Salmonella spp. en Listeria monocytogenes gaat een lage wateractiviteit van een
levensmiddel gepaard met een versterkte hitteresistentie van de pathogenen. Dit is ook het geval
bij de sporen van Bacillus cereus. Vanwege de lage wateractiviteit verdwijnen de waarden bij
een lage aw uit de verhittingscurve.
0
50
100
150
200
250
300
0 20 40 60 80 100 120 140
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 24: D-waarden (min) in functie van de temperatuur (°C) bij B. cereus sporen uit Lemgo.

55
Tabel 6: onderzochte D-waarden en temperaturen van de sporen van Bacillus cereus uit Lemgo met bronvermelding.
Een D100 = 11,6 min werd respectievelijke bekomen door Gonzalez et al. (1999) bij een
sporulatietemperatuur van 45 °C. Bij deze studie werd er ook gewerkt met de drie stammen van
Bacillus cereus, nl. ATCC 4342, 7004 en 9818. De hogere waarden worden toegeschreven aan
de sporulatietemperatuur en bijgevolg opgenomen in de tool.
Hoge D-waarden werden waargenomen bij Ababouch & Busta, (1987). Die hoge waarden zijn
het gevolg van het gebruikte soort medium, zoals de commerciële olie, sojaolie, olijfolie en
trioleïne waardoor hoge hitteresistenties werden verkregen. Sporen gelegen in een olieachtig
milieu bereiken namelijk dezelfde hitteresistentie als bij droge warmte (Molin & Snygg, 1967).
Die media, zoals olijfolie aw = 0,492 en commerciële olie aw = 0,479 bevatten zeer lage
wateractiviteiten. Die data worden daarom niet opgenomen in de verhittingscurve.
De waarden D100 = 19,79 min en D103 = 14,04 min zijn ook afkomstig uit de studie van Mazas
et al. (1999). Door toevoeging van NaCl werden wateractiviteiten bekomen van respectievelijk
0,84 en 0,75. Dat zorgt voor een sterkere hitteresistentie. Verdere hoge D-waarden werden in
deze studie bepaald door de aw te verlagen door de toevoeging van LiCl en sucrose. De waarden
met een te lage wateractiviteit worden niet opgenomen.
Temperatuur (°C) Tijd (min) Studie
95 257,3
Gaillard et al., (1998) 95 197,8
95 92,1
103 101
Mazas et al. (1999) 100 21,4
103 14,75
100 10,94
100 11,6 Gonzalez et al., (1999)
110 36,9
Ababouch & Busta, (1987) 110 31,17
110 49,31
121 30
100 19,79 Mazas et al. (1999)
103 14,04
100 19,1
Leguerinel et al., (2005) 100 13,32
100 8,71
110 11,5 Burgos et al. (1972)
110 1,5
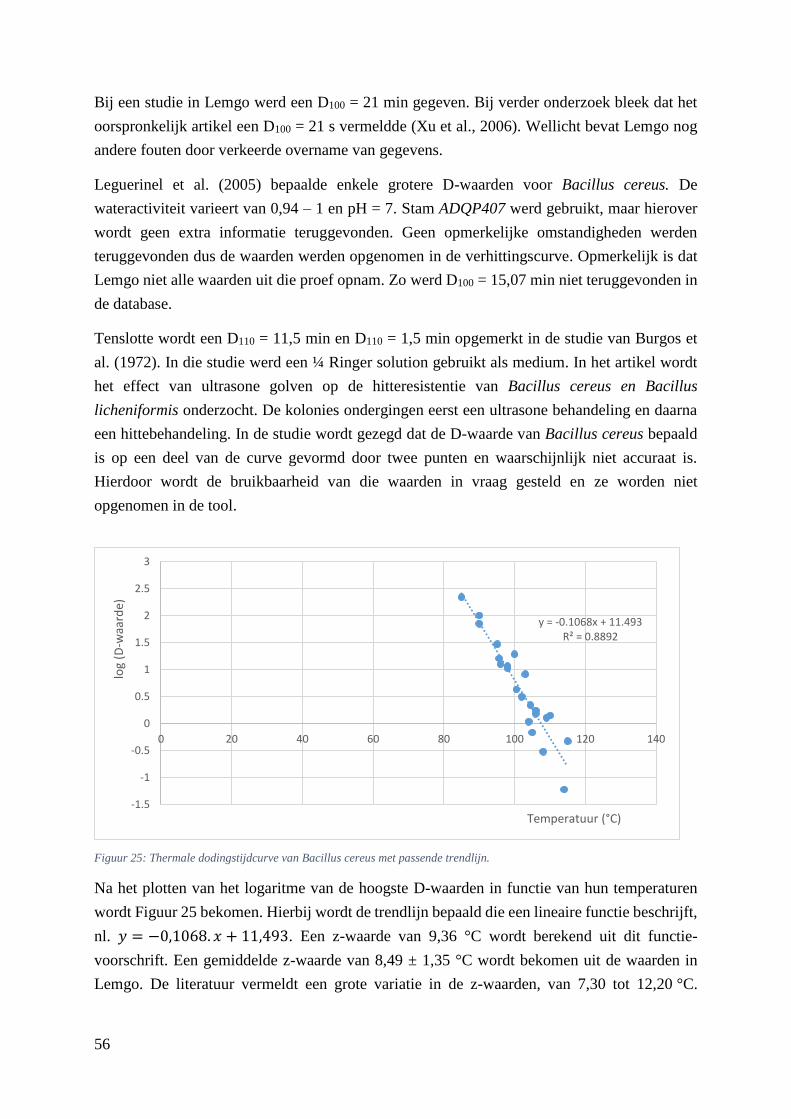
56
Bij een studie in Lemgo werd een D100 = 21 min gegeven. Bij verder onderzoek bleek dat het
oorspronkelijk artikel een D100 = 21 s vermeldde (Xu et al., 2006). Wellicht bevat Lemgo nog
andere fouten door verkeerde overname van gegevens.
Leguerinel et al. (2005) bepaalde enkele grotere D-waarden voor Bacillus cereus. De
wateractiviteit varieert van 0,94 – 1 en pH = 7. Stam ADQP407 werd gebruikt, maar hierover
wordt geen extra informatie teruggevonden. Geen opmerkelijke omstandigheden werden
teruggevonden dus de waarden werden opgenomen in de verhittingscurve. Opmerkelijk is dat
Lemgo niet alle waarden uit die proef opnam. Zo werd D100 = 15,07 min niet teruggevonden in
de database.
Tenslotte wordt een D110 = 11,5 min en D110 = 1,5 min opgemerkt in de studie van Burgos et
al. (1972). In die studie werd een ¼ Ringer solution gebruikt als medium. In het artikel wordt
het effect van ultrasone golven op de hitteresistentie van Bacillus cereus en Bacillus
licheniformis onderzocht. De kolonies ondergingen eerst een ultrasone behandeling en daarna
een hittebehandeling. In de studie wordt gezegd dat de D-waarde van Bacillus cereus bepaald
is op een deel van de curve gevormd door twee punten en waarschijnlijk niet accuraat is.
Hierdoor wordt de bruikbaarheid van die waarden in vraag gesteld en ze worden niet
opgenomen in de tool.
Na het plotten van het logaritme van de hoogste D-waarden in functie van hun temperaturen
wordt Figuur 25 bekomen. Hierbij wordt de trendlijn bepaald die een lineaire functie beschrijft,
nl. 𝑦 = −0,1068. 𝑥 + 11,493. Een z-waarde van 9,36 °C wordt berekend uit dit functie-
voorschrift. Een gemiddelde z-waarde van 8,49 ± 1,35 °C wordt bekomen uit de waarden in
Lemgo. De literatuur vermeldt een grote variatie in de z-waarden, van 7,30 tot 12,20 °C.
y = -0.1068x + 11.493R² = 0.8892
-1.5
-1
-0.5
0
0.5
1
1.5
2
2.5
3
0 20 40 60 80 100 120 140
log
(D-w
aard
e)
Temperatuur (°C)
Figuur 25: Thermale dodingstijdcurve van Bacillus cereus met passende trendlijn.
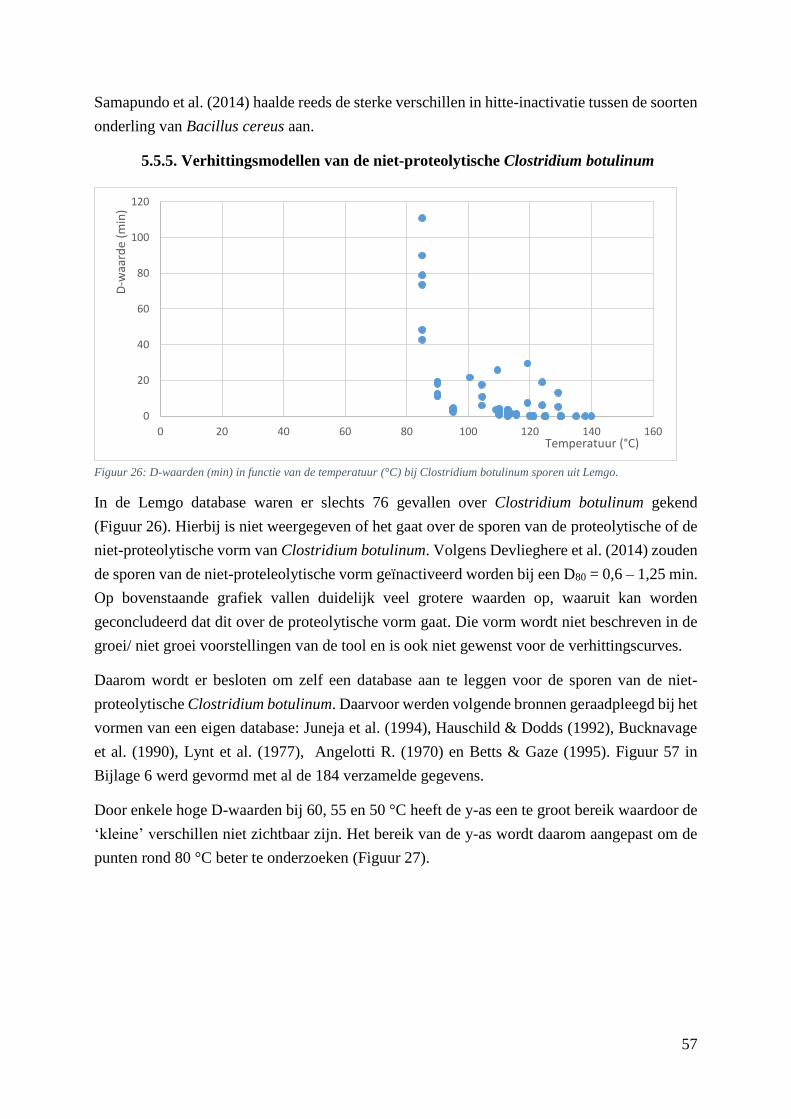
57
Samapundo et al. (2014) haalde reeds de sterke verschillen in hitte-inactivatie tussen de soorten
onderling van Bacillus cereus aan.
5.5.5. Verhittingsmodellen van de niet-proteolytische Clostridium botulinum
In de Lemgo database waren er slechts 76 gevallen over Clostridium botulinum gekend
(Figuur 26). Hierbij is niet weergegeven of het gaat over de sporen van de proteolytische of de
niet-proteolytische vorm van Clostridium botulinum. Volgens Devlieghere et al. (2014) zouden
de sporen van de niet-proteleolytische vorm geïnactiveerd worden bij een D80 = 0,6 – 1,25 min.
Op bovenstaande grafiek vallen duidelijk veel grotere waarden op, waaruit kan worden
geconcludeerd dat dit over de proteolytische vorm gaat. Die vorm wordt niet beschreven in de
groei/ niet groei voorstellingen van de tool en is ook niet gewenst voor de verhittingscurves.
Daarom wordt er besloten om zelf een database aan te leggen voor de sporen van de niet-
proteolytische Clostridium botulinum. Daarvoor werden volgende bronnen geraadpleegd bij het
vormen van een eigen database: Juneja et al. (1994), Hauschild & Dodds (1992), Bucknavage
et al. (1990), Lynt et al. (1977), Angelotti R. (1970) en Betts & Gaze (1995). Figuur 57 in
Bijlage 6 werd gevormd met al de 184 verzamelde gegevens.
Door enkele hoge D-waarden bij 60, 55 en 50 °C heeft de y-as een te groot bereik waardoor de
‘kleine’ verschillen niet zichtbaar zijn. Het bereik van de y-as wordt daarom aangepast om de
punten rond 80 °C beter te onderzoeken (Figuur 27).
0
20
40
60
80
100
120
0 20 40 60 80 100 120 140 160
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 26: D-waarden (min) in functie van de temperatuur (°C) bij Clostridium botulinum sporen uit Lemgo.
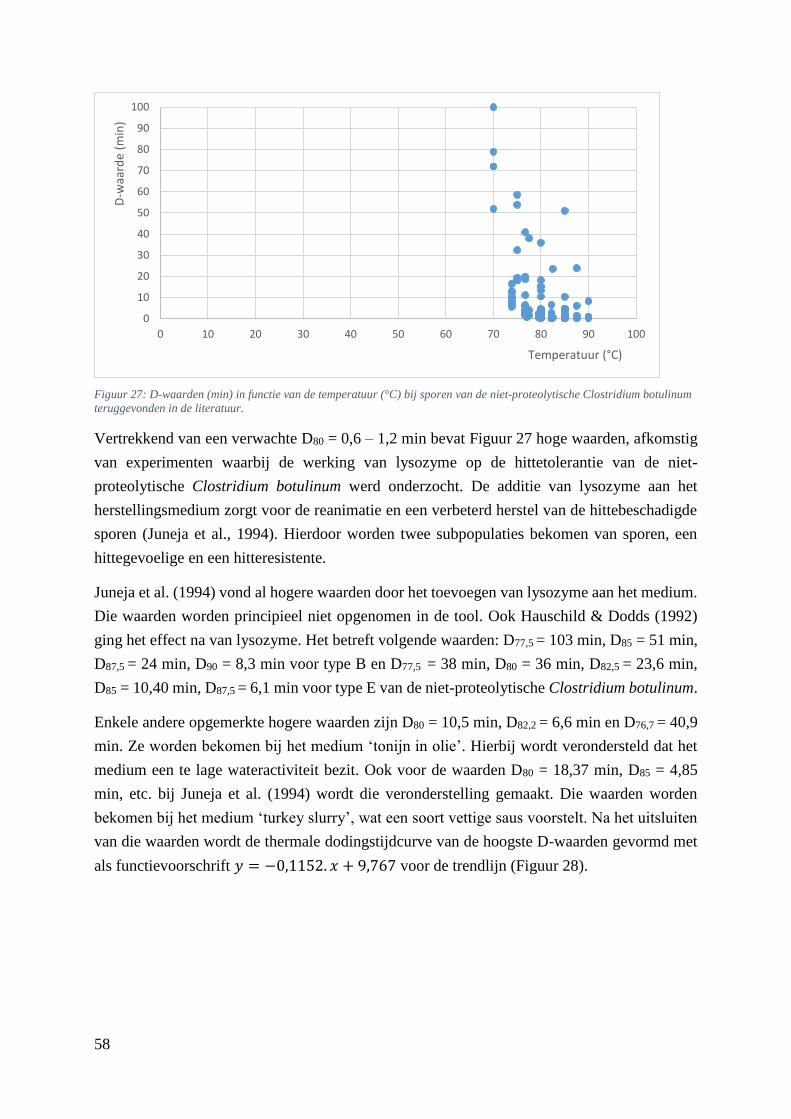
58
Vertrekkend van een verwachte D80 = 0,6 – 1,2 min bevat Figuur 27 hoge waarden, afkomstig
van experimenten waarbij de werking van lysozyme op de hittetolerantie van de niet-
proteolytische Clostridium botulinum werd onderzocht. De additie van lysozyme aan het
herstellingsmedium zorgt voor de reanimatie en een verbeterd herstel van de hittebeschadigde
sporen (Juneja et al., 1994). Hierdoor worden twee subpopulaties bekomen van sporen, een
hittegevoelige en een hitteresistente.
Juneja et al. (1994) vond al hogere waarden door het toevoegen van lysozyme aan het medium.
Die waarden worden principieel niet opgenomen in de tool. Ook Hauschild & Dodds (1992)
ging het effect na van lysozyme. Het betreft volgende waarden: D77,5 = 103 min, D85 = 51 min,
D87,5 = 24 min, D90 = 8,3 min voor type B en D77,5 = 38 min, D80 = 36 min, D82,5 = 23,6 min,
D85 = 10,40 min, D87,5 = 6,1 min voor type E van de niet-proteolytische Clostridium botulinum.
Enkele andere opgemerkte hogere waarden zijn D80 = 10,5 min, D82,2 = 6,6 min en D76,7 = 40,9
min. Ze worden bekomen bij het medium ‘tonijn in olie’. Hierbij wordt verondersteld dat het
medium een te lage wateractiviteit bezit. Ook voor de waarden D80 = 18,37 min, D85 = 4,85
min, etc. bij Juneja et al. (1994) wordt die veronderstelling gemaakt. Die waarden worden
bekomen bij het medium ‘turkey slurry’, wat een soort vettige saus voorstelt. Na het uitsluiten
van die waarden wordt de thermale dodingstijdcurve van de hoogste D-waarden gevormd met
als functievoorschrift 𝑦 = −0,1152. 𝑥 + 9,767 voor de trendlijn (Figuur 28).
0
10
20
30
40
50
60
70
80
90
100
0 10 20 30 40 50 60 70 80 90 100
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 27: D-waarden (min) in functie van de temperatuur (°C) bij sporen van de niet-proteolytische Clostridium botulinum
teruggevonden in de literatuur.
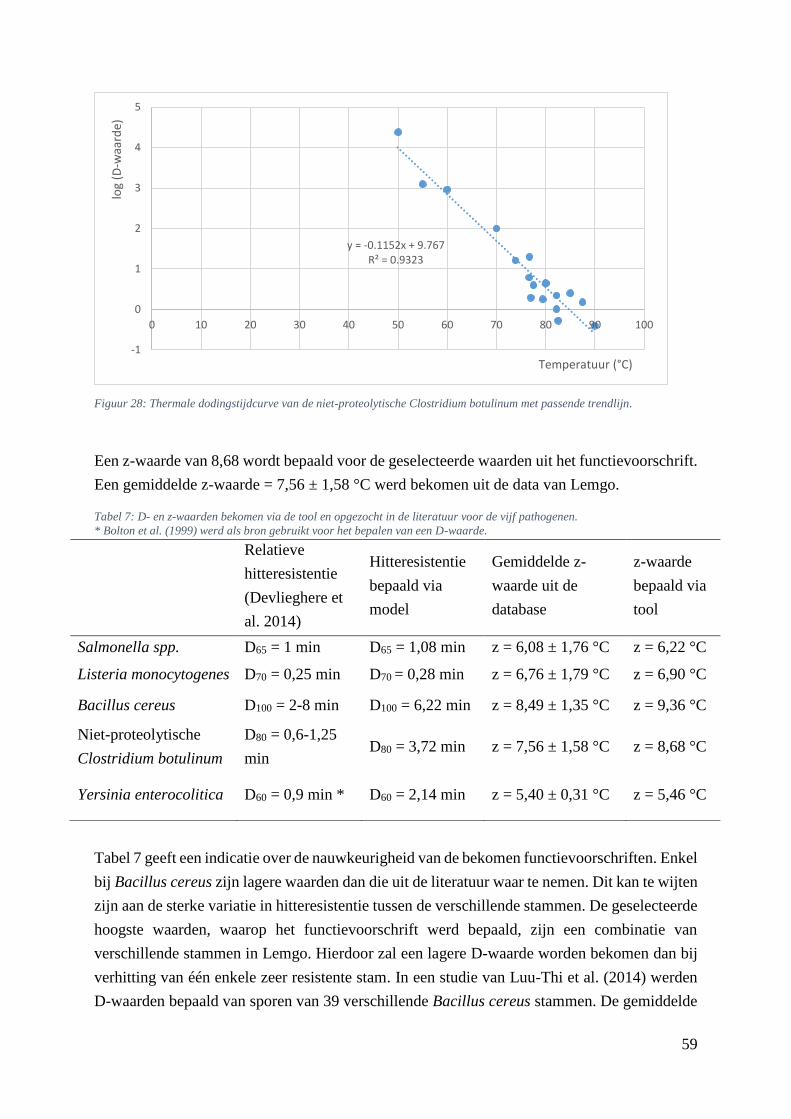
59
Een z-waarde van 8,68 wordt bepaald voor de geselecteerde waarden uit het functievoorschrift.
Een gemiddelde z-waarde = 7,56 ± 1,58 °C werd bekomen uit de data van Lemgo.
Tabel 7: D- en z-waarden bekomen via de tool en opgezocht in de literatuur voor de vijf pathogenen.
* Bolton et al. (1999) werd als bron gebruikt voor het bepalen van een D-waarde.
Tabel 7 geeft een indicatie over de nauwkeurigheid van de bekomen functievoorschriften. Enkel
bij Bacillus cereus zijn lagere waarden dan die uit de literatuur waar te nemen. Dit kan te wijten
zijn aan de sterke variatie in hitteresistentie tussen de verschillende stammen. De geselecteerde
hoogste waarden, waarop het functievoorschrift werd bepaald, zijn een combinatie van
verschillende stammen in Lemgo. Hierdoor zal een lagere D-waarde worden bekomen dan bij
verhitting van één enkele zeer resistente stam. In een studie van Luu-Thi et al. (2014) werden
D-waarden bepaald van sporen van 39 verschillende Bacillus cereus stammen. De gemiddelde
y = -0.1152x + 9.767R² = 0.9323
-1
0
1
2
3
4
5
0 10 20 30 40 50 60 70 80 90 100
log
(D-w
aard
e)
Temperatuur (°C)
Relatieve
hitteresistentie
(Devlieghere et
al. 2014)
Hitteresistentie
bepaald via
model
Gemiddelde z-
waarde uit de
database
z-waarde
bepaald via
tool
Salmonella spp. D65 = 1 min D65 = 1,08 min z = 6,08 ± 1,76 °C z = 6,22 °C
Listeria monocytogenes D70 = 0,25 min D70 = 0,28 min z = 6,76 ± 1,79 °C z = 6,90 °C
Bacillus cereus D100 = 2-8 min D100 = 6,22 min z = 8,49 ± 1,35 °C z = 9,36 °C
Niet-proteolytische
Clostridium botulinum
D80 = 0,6-1,25
min D80 = 3,72 min z = 7,56 ± 1,58 °C z = 8,68 °C
Yersinia enterocolitica D60 = 0,9 min * D60 = 2,14 min z = 5,40 ± 0,31 °C z = 5,46 °C
Figuur 28: Thermale dodingstijdcurve van de niet-proteolytische Clostridium botulinum met passende trendlijn.
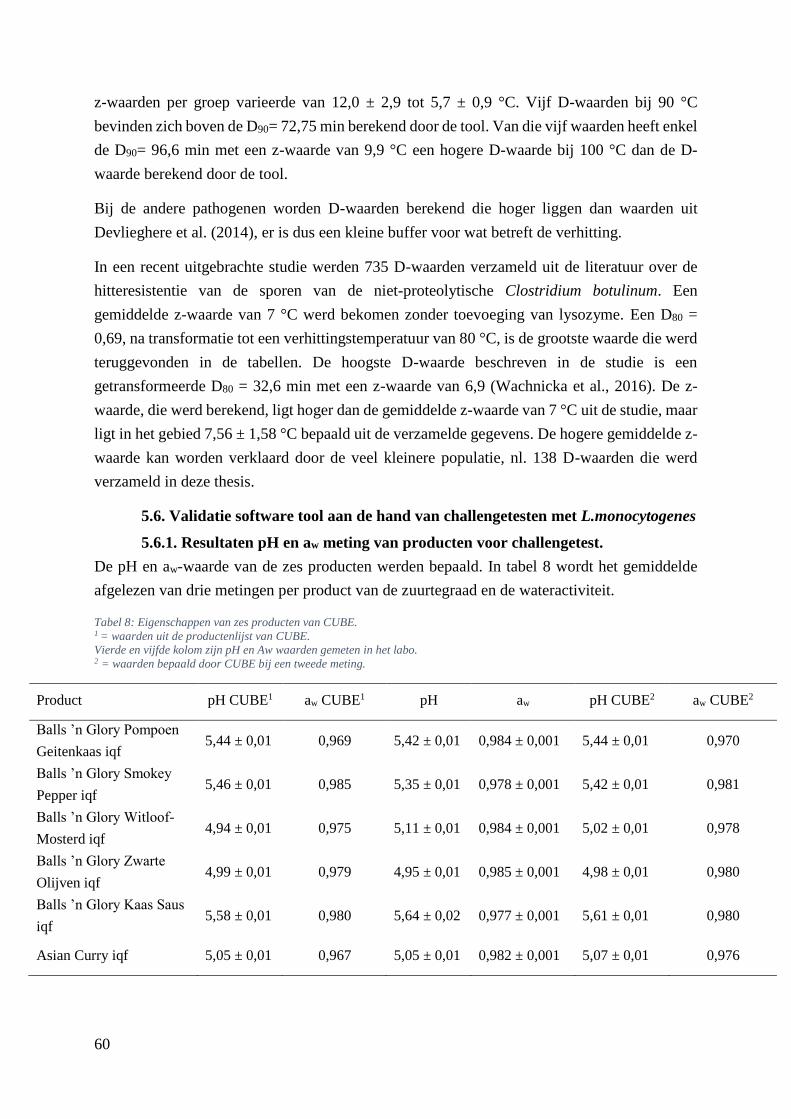
60
z-waarden per groep varieerde van 12,0 ± 2,9 tot 5,7 ± 0,9 °C. Vijf D-waarden bij 90 °C
bevinden zich boven de D90= 72,75 min berekend door de tool. Van die vijf waarden heeft enkel
de D90= 96,6 min met een z-waarde van 9,9 °C een hogere D-waarde bij 100 °C dan de D-
waarde berekend door de tool.
Bij de andere pathogenen worden D-waarden berekend die hoger liggen dan waarden uit
Devlieghere et al. (2014), er is dus een kleine buffer voor wat betreft de verhitting.
In een recent uitgebrachte studie werden 735 D-waarden verzameld uit de literatuur over de
hitteresistentie van de sporen van de niet-proteolytische Clostridium botulinum. Een
gemiddelde z-waarde van 7 °C werd bekomen zonder toevoeging van lysozyme. Een D80 =
0,69, na transformatie tot een verhittingstemperatuur van 80 °C, is de grootste waarde die werd
teruggevonden in de tabellen. De hoogste D-waarde beschreven in de studie is een
getransformeerde D80 = 32,6 min met een z-waarde van 6,9 (Wachnicka et al., 2016). De z-
waarde, die werd berekend, ligt hoger dan de gemiddelde z-waarde van 7 °C uit de studie, maar
ligt in het gebied 7,56 ± 1,58 °C bepaald uit de verzamelde gegevens. De hogere gemiddelde z-
waarde kan worden verklaard door de veel kleinere populatie, nl. 138 D-waarden die werd
verzameld in deze thesis.
5.6. Validatie software tool aan de hand van challengetesten met L.monocytogenes
5.6.1. Resultaten pH en aw meting van producten voor challengetest.
De pH en aw-waarde van de zes producten werden bepaald. In tabel 8 wordt het gemiddelde
afgelezen van drie metingen per product van de zuurtegraad en de wateractiviteit.
Tabel 8: Eigenschappen van zes producten van CUBE. 1 = waarden uit de productenlijst van CUBE.
Vierde en vijfde kolom zijn pH en Aw waarden gemeten in het labo. 2 = waarden bepaald door CUBE bij een tweede meting.
Product pH CUBE1 aw CUBE1 pH aw pH CUBE2 aw CUBE2
Balls ’n Glory Pompoen
Geitenkaas iqf 5,44 ± 0,01 0,969 5,42 ± 0,01 0,984 ± 0,001 5,44 ± 0,01 0,970
Balls ’n Glory Smokey
Pepper iqf 5,46 ± 0,01 0,985 5,35 ± 0,01 0,978 ± 0,001 5,42 ± 0,01 0,981
Balls ’n Glory Witloof-
Mosterd iqf 4,94 ± 0,01 0,975 5,11 ± 0,01 0,984 ± 0,001 5,02 ± 0,01 0,978
Balls ’n Glory Zwarte
Olijven iqf 4,99 ± 0,01 0,979 4,95 ± 0,01 0,985 ± 0,001 4,98 ± 0,01 0,980
Balls ’n Glory Kaas Saus
iqf 5,58 ± 0,01 0,980 5,64 ± 0,02 0,977 ± 0,001 5,61 ± 0,01 0,980
Asian Curry iqf 5,05 ± 0,01 0,967 5,05 ± 0,01 0,982 ± 0,001 5,07 ± 0,01 0,976

61
Volgens tabel 8 komen in het algemeen de bekomen waarden voor de pH redelijk goed overeen
met de waarden doorgegeven door CUBE. Het grootste verschil voor de pH valt op bij de
witloof-mosterd. Bij de waarden van de wateractiviteit zijn grotere relatieve verschillen op te
merken. De grootste aw-verschillen zijn terug te vinden bij de asian curry en de pompoen
geitenkaas. Dit verschil heeft een grote invloed. De groei wordt namelijk voorspeld tussen een
aw-waarde van 0,935 en 0,990 of over een bereik van 0,055 in het model. Door de verschillen
bevinden sommige producten zich niet in het gewenste grensgebied tussen groei en geen groei
van de modellen.
Een bepaalde heterogeniteit in de batch kan die afwijkingen verklaren. Hierdoor zouden er
verschillen kunnen zijn afhankelijk van het deel van het totale product waarop de aw en pH
werden bepaald. Ook de temperatuur op het ogenblik waarop de metingen zijn uitgevoerd, kan
een mogelijke verklaring zijn. In het bedrijf worden de metingen verricht bij kamertemperatuur
die sterk kan variëren, nl. van 18,9 tot 25 °C. Voor de bepaling van de wateractiviteit gebruikt
CUBE een labSwift-aw van Novasina, maar dit meetapparaat is niet altijd ideaal voor producten
met een aw > 0,9. Een standaardafwijking van ± 0,002 bij dit apparaat is enkel geldig wanneer
de meting gebeurt bij een aw-waarde tussen de 0,11 en 0,90 en bij een constante temperatuur.
De standaarddeviatie is dus niet gekend omdat alle producten een te hoge aw bezitten.
De testen worden door het bedrijf herhaald om een mogelijke variatie in de batch na te gaan.
De bekomen resultaten van een tweede meting worden teruggevonden in de laatste twee
kolommen van tabel 8. Uit het overzicht valt op dat de waarden van de pH in het algemeen
dichter aanleunen bij de waarden bepaald in het labo. Ook bij de wateractiviteit zijn de
verschillen kleiner geworden, bij de pompoen geitenkaas wordt het grootste verschil
waargenomen.
5.6.2. Resultaten van de VIDAS LMO2-test
Met de VIDAS LMO2 werd getest of Listeria monocytogenes aanwezig was in 25 g van de
producten. Elk product werd in het drievoud getest, maar geen enkel positief resultaat werd
bekomen. Hieruit kan worden geconcludeerd dat Listeria monocytogenes afwezig was in 25 g
van elk product voor aanvang van de challengetesten.
5.6.3. Bepalen van groeipotentieel van Listeria monocytogenes bij 4 en 7 °C.
In de tabellen 9 en 10 staan de groeipotentialen van Listeria monocytogenes bij respectievelijk
4 en 7 °C. Groei wordt gedefinieerd bij een significant verschil van tenminste 0,50 log KVE/g
tussen de eerste en de laatste dag. Voor de kaas saus, de pompoen geitenkaas en de zwarte
olijven wordt zowel bij 4 °C en 7 °C significante groei geobserveerd. Opvallend is dat de zwarte
olijven en de witloof-mosterd zeer gelijke producteigenschappen bezitten, beide producten
hebben een pH van ongeveer 5 en een aw-waarde van ongeveer 0,985, maar toch is er enkel
groei bij de zwarte olijven. De mogelijke antibacteriële werking van beide producten wordt in
de literatuur opgezocht.
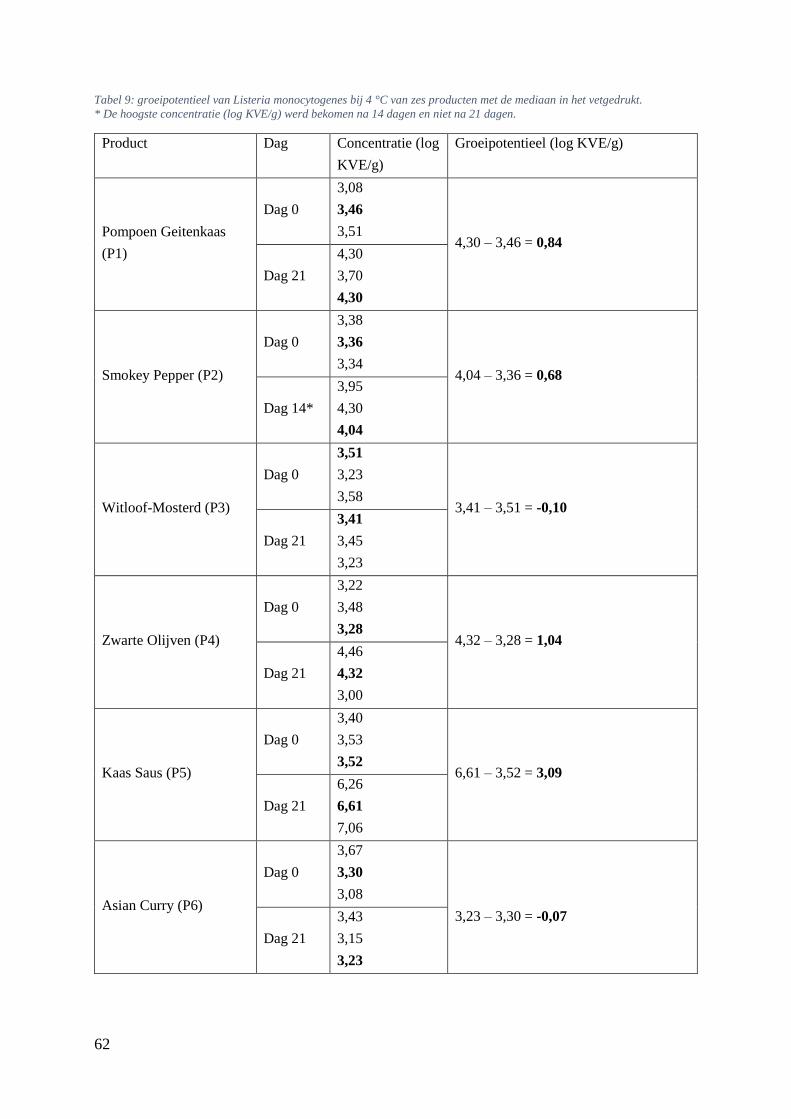
62
Tabel 9: groeipotentieel van Listeria monocytogenes bij 4 °C van zes producten met de mediaan in het vetgedrukt.
* De hoogste concentratie (log KVE/g) werd bekomen na 14 dagen en niet na 21 dagen.
Product Dag Concentratie (log
KVE/g)
Groeipotentieel (log KVE/g)
Pompoen Geitenkaas
(P1)
Dag 0
3,08
3,46
3,51 4,30 – 3,46 = 0,84
Dag 21
4,30
3,70
4,30
Smokey Pepper (P2)
Dag 0
3,38
3,36
3,34 4,04 – 3,36 = 0,68
Dag 14*
3,95
4,30
4,04
Witloof-Mosterd (P3)
Dag 0
3,51
3,23
3,58 3,41 – 3,51 = -0,10
Dag 21
3,41
3,45
3,23
Zwarte Olijven (P4)
Dag 0
3,22
3,48
3,28 4,32 – 3,28 = 1,04
Dag 21
4,46
4,32
3,00
Kaas Saus (P5)
Dag 0
3,40
3,53
3,52 6,61 – 3,52 = 3,09
Dag 21
6,26
6,61
7,06
Asian Curry (P6)
Dag 0
3,67
3,30
3,08
3,23 – 3,30 = -0,07
Dag 21
3,43
3,15
3,23
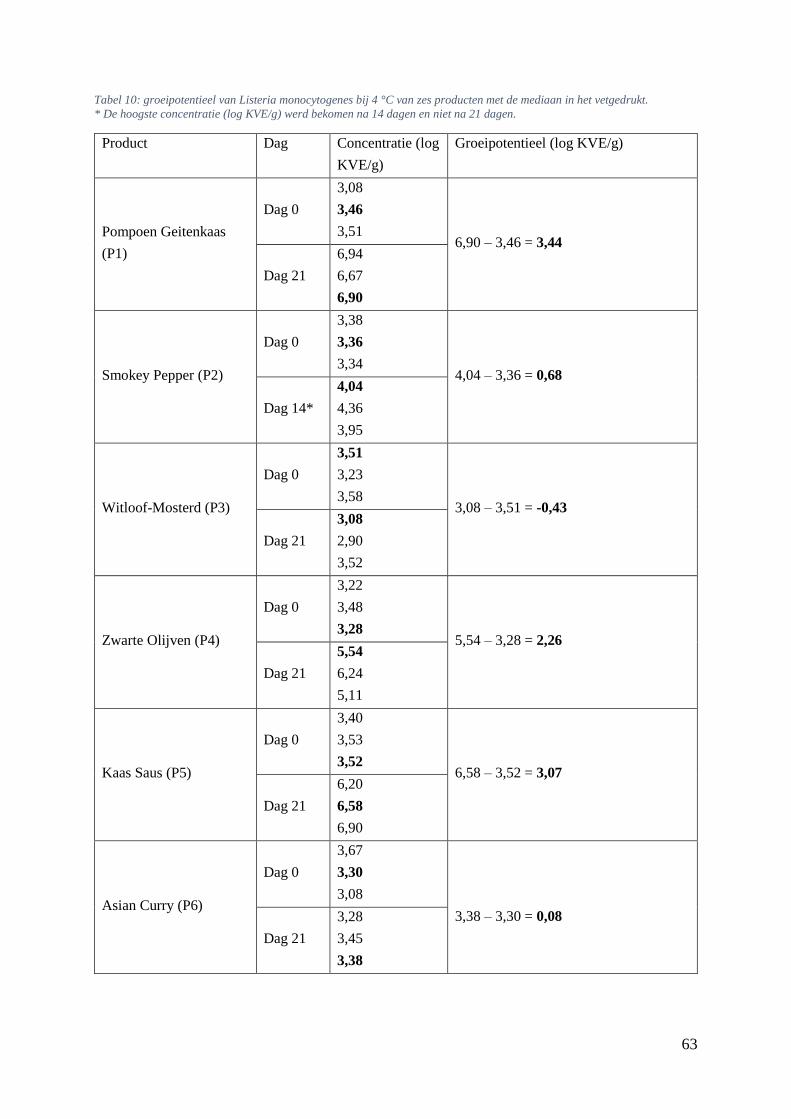
63
Tabel 10: groeipotentieel van Listeria monocytogenes bij 4 °C van zes producten met de mediaan in het vetgedrukt.
* De hoogste concentratie (log KVE/g) werd bekomen na 14 dagen en niet na 21 dagen.
Product Dag Concentratie (log
KVE/g)
Groeipotentieel (log KVE/g)
Pompoen Geitenkaas
(P1)
Dag 0
3,08
3,46
3,51 6,90 – 3,46 = 3,44
Dag 21
6,94
6,67
6,90
Smokey Pepper (P2)
Dag 0
3,38
3,36
3,34 4,04 – 3,36 = 0,68
Dag 14*
4,04
4,36
3,95
Witloof-Mosterd (P3)
Dag 0
3,51
3,23
3,58 3,08 – 3,51 = -0,43
Dag 21
3,08
2,90
3,52
Zwarte Olijven (P4)
Dag 0
3,22
3,48
3,28 5,54 – 3,28 = 2,26
Dag 21
5,54
6,24
5,11
Kaas Saus (P5)
Dag 0
3,40
3,53
3,52 6,58 – 3,52 = 3,07
Dag 21
6,20
6,58
6,90
Asian Curry (P6)
Dag 0
3,67
3,30
3,08
3,38 – 3,30 = 0,08
Dag 21
3,28
3,45
3,38

64
Rhee et al. (2003) toonde de antimicrobiële werking aan van mosterd op Listeria
monocytogenes, E. coli en Salmonella typhimirium. In het artikel werd bepaald dat mosterd op
zich (pH > 5) een betere inhiberende werking had dan een mengsel van mosterd met azijnzuur
(pH < 4). De antimicrobiële activiteit van Cichorium intybus (wilde witloof) tegen
Staphylococcus aureus, Staphylococcus aurous, Listeria monocytogenes en E. coli werd
onderzocht door Faiku et al. (2016). De fractie methanol, ethylacetaat en di-ethylether, die werd
geëxtraheerd, vertoonde een antibacteriële werking tegen Listeria monocytogenes. Het is ook
belangrijk op te merken dat mosterd veel azijn bevat. Azijn heeft een sterke anti-microbiële
werking tegen onder andere Listeria monocytogenes (Medina et al., 2007).
Olijven bezitten componenten, die ook een antimicrobiële werking hebben. Ze gelijken op
glutaaraldehyde, zoals ‘decarboxymethyl elenolic acid’ (Medina et al., 2013). Hieruit kan
worden geconcludeerd dat zowel de zwarte olijven als de witloof-mosterd antimicrobiële
componenten bezitten. Wellicht is er een groter aandeel componenten aanwezig in de Witloof-
Mosterd, waardoor in het product geen groei is opgetreden van Listeria monocytogenes.
Voor beide temperaturen kent de kaas saus de sterkste groei. Het is tevens het product met de
hoogste waarde voor pH en de laagste voor wateractiviteit. In het algemeen wordt er, zoals
verwacht, een sterkere groei waargenomen bij 7 °C dan bij 4 °C. Bij de kaas saus trad echter
schimmelgroei op vanaf dag 14 bij zowel 4 als 7 °C (Figuur 29). De schimmel werd niet verder
onderzocht, waardoor de werking van de schimmel op de groei van Listeria monocytogenes
niet gekend is.
Figuur 29: schimmelgroei op de drie stalen kaas saus na 21 dagen bij 7 °C.
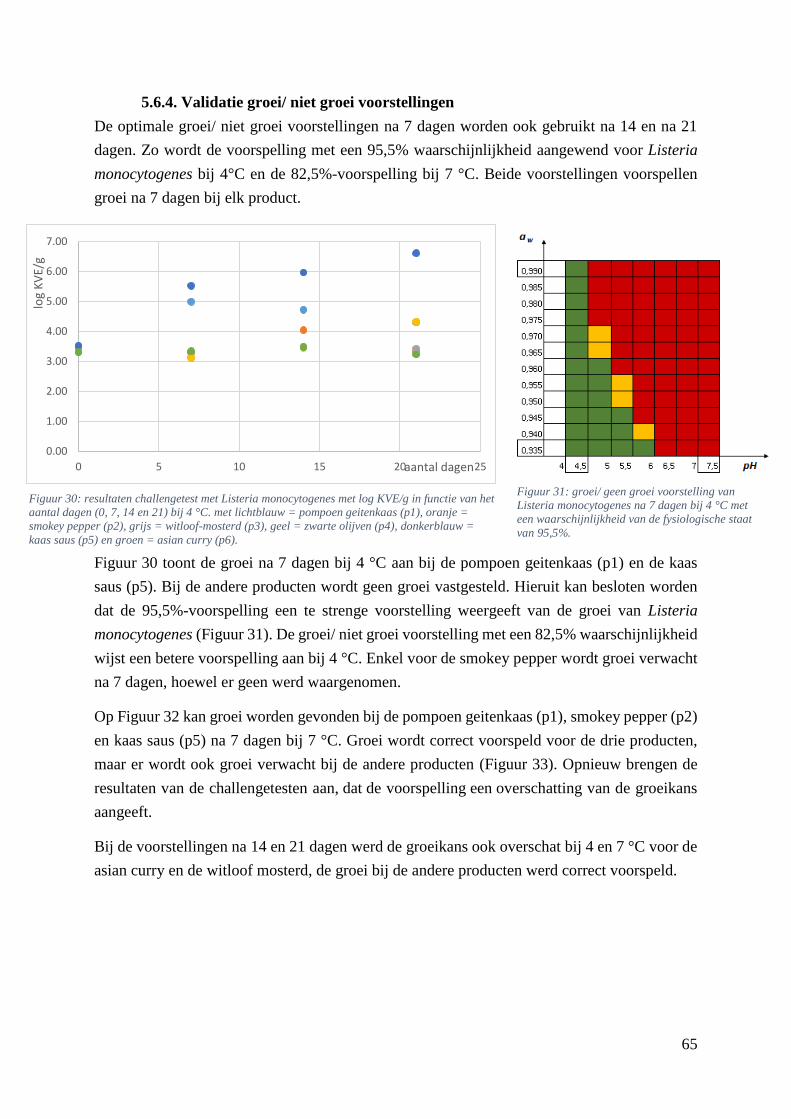
65
5.6.4. Validatie groei/ niet groei voorstellingen
De optimale groei/ niet groei voorstellingen na 7 dagen worden ook gebruikt na 14 en na 21
dagen. Zo wordt de voorspelling met een 95,5% waarschijnlijkheid aangewend voor Listeria
monocytogenes bij 4°C en de 82,5%-voorspelling bij 7 °C. Beide voorstellingen voorspellen
groei na 7 dagen bij elk product.
Figuur 30 toont de groei na 7 dagen bij 4 °C aan bij de pompoen geitenkaas (p1) en de kaas
saus (p5). Bij de andere producten wordt geen groei vastgesteld. Hieruit kan besloten worden
dat de 95,5%-voorspelling een te strenge voorstelling weergeeft van de groei van Listeria
monocytogenes (Figuur 31). De groei/ niet groei voorstelling met een 82,5% waarschijnlijkheid
wijst een betere voorspelling aan bij 4 °C. Enkel voor de smokey pepper wordt groei verwacht
na 7 dagen, hoewel er geen werd waargenomen.
Op Figuur 32 kan groei worden gevonden bij de pompoen geitenkaas (p1), smokey pepper (p2)
en kaas saus (p5) na 7 dagen bij 7 °C. Groei wordt correct voorspeld voor de drie producten,
maar er wordt ook groei verwacht bij de andere producten (Figuur 33). Opnieuw brengen de
resultaten van de challengetesten aan, dat de voorspelling een overschatting van de groeikans
aangeeft.
Bij de voorstellingen na 14 en 21 dagen werd de groeikans ook overschat bij 4 en 7 °C voor de
asian curry en de witloof mosterd, de groei bij de andere producten werd correct voorspeld.
0.00
1.00
2.00
3.00
4.00
5.00
6.00
7.00
0 5 10 15 20 25
log
KV
E/g
aantal dagen
Figuur 30: resultaten challengetest met Listeria monocytogenes met log KVE/g in functie van het
aantal dagen (0, 7, 14 en 21) bij 4 °C. met lichtblauw = pompoen geitenkaas (p1), oranje =
smokey pepper (p2), grijs = witloof-mosterd (p3), geel = zwarte olijven (p4), donkerblauw =
kaas saus (p5) en groen = asian curry (p6).
Figuur 31: groei/ geen groei voorstelling van
Listeria monocytogenes na 7 dagen bij 4 °C met
een waarschijnlijkheid van de fysiologische staat
van 95,5%.
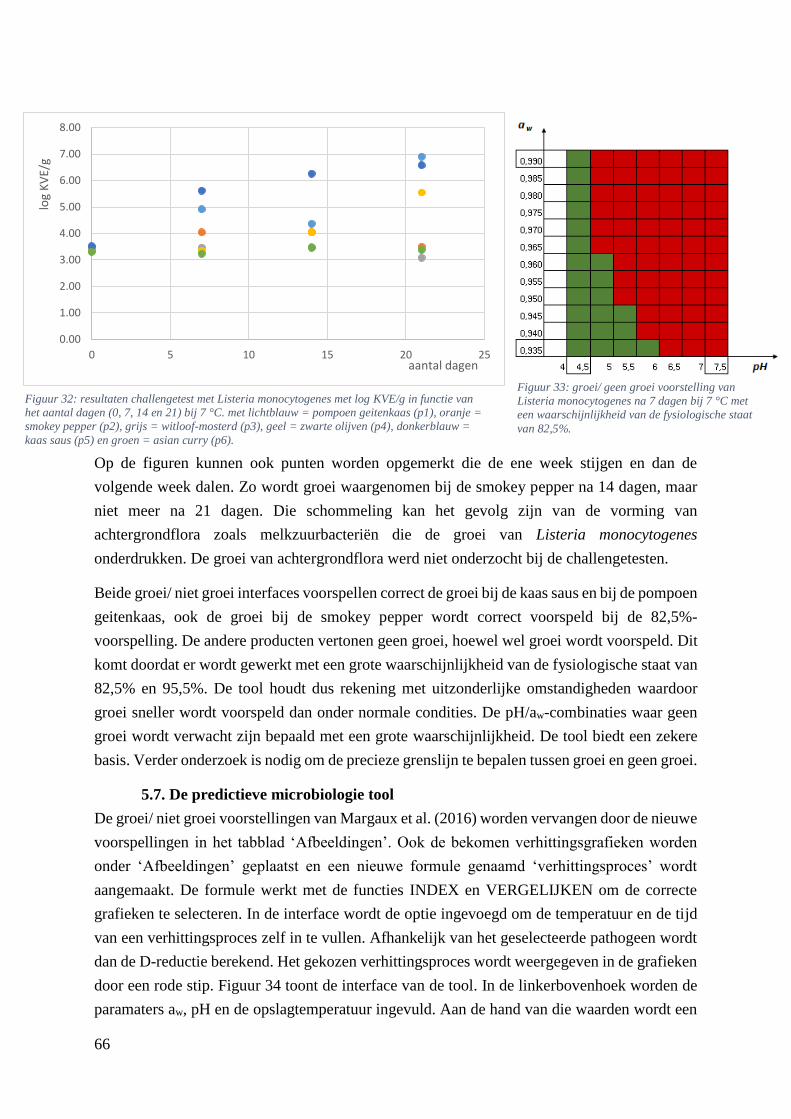
66
Op de figuren kunnen ook punten worden opgemerkt die de ene week stijgen en dan de
volgende week dalen. Zo wordt groei waargenomen bij de smokey pepper na 14 dagen, maar
niet meer na 21 dagen. Die schommeling kan het gevolg zijn van de vorming van
achtergrondflora zoals melkzuurbacteriën die de groei van Listeria monocytogenes
onderdrukken. De groei van achtergrondflora werd niet onderzocht bij de challengetesten.
Beide groei/ niet groei interfaces voorspellen correct de groei bij de kaas saus en bij de pompoen
geitenkaas, ook de groei bij de smokey pepper wordt correct voorspeld bij de 82,5%-
voorspelling. De andere producten vertonen geen groei, hoewel wel groei wordt voorspeld. Dit
komt doordat er wordt gewerkt met een grote waarschijnlijkheid van de fysiologische staat van
82,5% en 95,5%. De tool houdt dus rekening met uitzonderlijke omstandigheden waardoor
groei sneller wordt voorspeld dan onder normale condities. De pH/aw-combinaties waar geen
groei wordt verwacht zijn bepaald met een grote waarschijnlijkheid. De tool biedt een zekere
basis. Verder onderzoek is nodig om de precieze grenslijn te bepalen tussen groei en geen groei.
5.7. De predictieve microbiologie tool
De groei/ niet groei voorstellingen van Margaux et al. (2016) worden vervangen door de nieuwe
voorspellingen in het tabblad ‘Afbeeldingen’. Ook de bekomen verhittingsgrafieken worden
onder ‘Afbeeldingen’ geplaatst en een nieuwe formule genaamd ‘verhittingsproces’ wordt
aangemaakt. De formule werkt met de functies INDEX en VERGELIJKEN om de correcte
grafieken te selecteren. In de interface wordt de optie ingevoegd om de temperatuur en de tijd
van een verhittingsproces zelf in te vullen. Afhankelijk van het geselecteerde pathogeen wordt
dan de D-reductie berekend. Het gekozen verhittingsproces wordt weergegeven in de grafieken
door een rode stip. Figuur 34 toont de interface van de tool. In de linkerbovenhoek worden de
paramaters aw, pH en de opslagtemperatuur ingevuld. Aan de hand van die waarden wordt een
0.00
1.00
2.00
3.00
4.00
5.00
6.00
7.00
8.00
0 5 10 15 20 25
log
KV
E/g
aantal dagen
Figuur 32: resultaten challengetest met Listeria monocytogenes met log KVE/g in functie van
het aantal dagen (0, 7, 14 en 21) bij 7 °C. met lichtblauw = pompoen geitenkaas (p1), oranje =
smokey pepper (p2), grijs = witloof-mosterd (p3), geel = zwarte olijven (p4), donkerblauw =
kaas saus (p5) en groen = asian curry (p6).
Figuur 33: groei/ geen groei voorstelling van
Listeria monocytogenes na 7 dagen bij 7 °C met
een waarschijnlijkheid van de fysiologische staat
van 82,5%.

67
lijst weergegeven van de mogelijke uitgroei van de pathogenen. Rechts bovenaan kunnen de
gekozen stappen, waarmee de mogelijke uitgroei van de pathogenen wordt bepaald, worden
teruggevonden. In de linkerbenedenhoek kunnen de groei/ niet groei voorstellingen worden
teruggevonden voor de vijf pathogenen na 7, 14 of 21 dagen bij 4 en bij 7 °C. Rechts onderaan
worden de verhittingsgrafieken weergegeven voor één van de vijf pathogenen. Als de D-
reductie zich bevindt tussen de 6 en 12 zal het pathogeen in het lichtgroen worden aangeduid.
Een D-reductie > 12 wordt voorgesteld door een donkergroene kleur van het pathogeen. Bij
nacontaminatie verdwijnen de markeringen.
Figuur 34: printscreen van de tool in Excel. Links bovenaan wordt een lijst gegeven van pathogenen die kunnen
uitgroeien aan de hand van de ingegeven T, pH en Aw. Rechts bovenaan kunnen de gekozen stappen, waarmee de
mogelijke uitgroei van de pathogenen wordt bepaald, worden teruggevonden. Links onderaan worden de groei/ niet groei
voorstellingen waargenomen en rechts onderaan de verhittingscurves.

68

69
6. Algemeen besluit
Een tool op basis van bestaande predictieve modellen werd ontworpen. De tool biedt een visuele
voorstelling van de voorspelde groeikans van Salmonella spp., Listeria monocytogenes,
Yersinia enterocolitica, Bacillus cereus en de niet-proteolytische Clostridium botulinum na 7,
14 en 21 dagen bij 4 en 7 °C. In deze thesis werd er verder gebouwd op de resultaten van Van
Helleputte (2016). Nieuwe groei/ niet groei voorstellingen werden bepaald voor verschillende
waarschijnlijkheden van de fysiologische staat. Het effect van de fysiologische staat op de duur
van de lag-fase werd hierbij in kaart gebracht. Zo vertoonden groei/ niet groei voorstellingen
gebaseerd op de maximale waarde bijna bij elke pH/aw-combinatie groei. Terwijl groei/ niet
groei voorstellingen bepaald met de standaardwaarde beperkte groei vertoonden bij minder
stresserende omstandigheden (Van Helleputte, 2016).
De ideale waarschijnlijkheden van de fysiologische staat werden bekomen door groei/ niet groei
voorstellingen op basis van de ‘ComBase Predictor’ te vergelijken met voorspellingen op basis
van de database van ComBase. Verschillende ideale waarschijnlijkheden van de fysiologische
staat werden bekomen voor de pathogenen. Een mogelijke verklaring voor die verschillen kan
worden toegeschreven aan de variabiliteit binnenin de stammen van een pathogeen. Zo kunnen
er stammen zijn die sneller en makkelijker beginnen met delen dan andere bij lagere
temperaturen. Die variabiliteit kan bij de ene pathogeen veel groter zijn dan bij de andere.
De verschillen kunnen ook te wijten zijn aan de variabiliteit binnen eenzelfde stam, de
celvariabiliteit. De groei zal gedomineerd worden door de snelste cel. Dit is vooral van
toepassing bij een zeer laag inoculumniveau. Hierdoor wordt sneller een grotere
waarschijnlijkheid bekomen vanuit een worstcasescenario. Meer data om de groei/ niet groei
condities op basis van de ‘ComBase Predictor’ en de database van ComBase te vergelijken en
verder onderzoek zijn nodig om de oorzaak van de verschillen te verklaren.
Naast een voorspelling van de groei kan ook de D-reductie van een verhittingsstap worden
voorspeld met de tool. Hiervoor werden de hoogste D-waarden in Lemgo geselecteerd om
daarna een functievoorschrift van de trendlijn te bepalen voor elk pathogeen. D-waarden
bekomen via de voorschriften komen goed overeen met de D-waarden opgezocht in de
literatuur. Behalve bij Bacillus cereus werden in de literatuur hogere D-waarden
teruggevonden. Het verschil wordt verklaard door de grote variatie binnenin de stammen van
Bacillus cereus op vlak van hitteresistentie. De z-waarden bepaald met de functievoorschriften
liggen in de gebieden van de gemiddelde z-waarden en standaarddeviaties. De grootte van de
standaardafwijkingen wijzen op de verschillen tussen de stammen onderling op vlak van
hitteresistentie.

70
Op basis van de resultaten uit de challengetesten wordt besloten dat de tool een overschatting
van de groeikans van L. monocytogenes aangeeft. De groei werd bij 4 en 7 °C voor elk product
voorspeld, terwijl slechts in twee bij 4 °C en in drie producten bij 7 °C groei werd
waargenomen. Dit komt doordat wordt gewerkt vanuit een worstcasescenario. Een grote
waarschijnlijkheid van de fysiologische staat werd geselecteerd bij het opmaken van de
groei/niet groeivoorstellingen. Hierdoor wordt groei sneller voorspeld. Ook na 14 en 21 dagen
wordt een overschatting van de groei bekomen voor enkele producten.
Met de ontwikkelde tool gebaseerd op bestaande predictieve modellen is het mogelijk een
voorspelling te vormen over de groeikans van een pathogeen en de hitte-inactivatie van een
verhittingsstap. Het is aangewezen dat de tool met de nodige zorg en kennis gebruikt wordt. De
software biedt een zekere basis voor CUBE om verder op te werken. Verder onderzoek is nodig
om de gevonden groei/ geen groei voorspellingen van Listeria monocytogenes te verfijnen.
Challengetesten met de andere pathogenen zijn nodig om de groei/ niet groei voorstellingen te
valideren. Het aanvullen van de databasen met extra D-waarden van resistente stammen is
aangeraden, vooral voor Bacillus cereus om voorspellingen van de D-reductie te verbeteren.
De tool is gemaakt in Excel® en is dus ook toegankelijk voor andere bedrijven. Naast het
berekenen van de D-reductie van een verhittingsstap worden de verhittingscurves van de vijf
pathogenen weergegeven. Op basis van waarden voor de pH, aw en opslagtemperatuur wordt
een lijst gegeven van mogelijke pathogenen die kunnen uitgroeien bij die condities. De
gebruikte formules, condities, groei/ niet groei voorstellingen en verhittingscurves kunnen
makkelijk worden teruggevonden, uitgebreid en aangepast. Bijgevolg heeft de tool bredere
toepassingen.

71
7. Bronvermelding
Ababouch, L. & Busta, F. F. (1987). Effect on thermal treatments in oils on bacterial spore
survival. Journal of Applied Bacteriology, 62, 491-502.
Alfaro, B., Nuin, M., Le Marc, Y., Pin, C. (2007). “FISH SHELF LIFE PREDICTIONS
PROGRAM-FSLP”. user manual version 1.0. Geraadpleegd op 18 februari 2017, op
http://www.azti.es/wp-content/uploads/2013/09/Manual_FSLP.pdf
Alfaro, B. (2008). Application of “Fish Shelf life Prediction (FSLP)” software for monitoring
seafood quality in the cold chain [Powerpointpresentatie].
Alfaro, B., Escuredo, K., Pin, C. (2011). “FISHMAP-PROGRAM-”. user manual version 1.0.
Geraadpleegd op 9 februari 2017, op http://www.azti.es/wp-
content/uploads/2013/09/FISHMAP_Manual.pdf
Althoff, T., Schwarzer, K. (2007). Welcome to the Lemgo D- and z-value Database for Food
1.1. LDz-Base. Geraadpleegd op 31 oktober 2016, op http://www.hs-
owl.de/fb4/ldzbase/
Angelotti, R. (1968). The heat resistance of clostridium botulinum type E in food.
Proceedings of the First U.S.-Japan Conference on Toxic Micro-Organisms (pp. 404-
409). paper voorgedragen op U.S.-Japan cooperative program in natural resources,
Honolulu, 7 - 10 oktober 1968.
Anonymous (2002). Risk Ranger. Geraadpleegd op 11 februari 2017, op
http://www.foodsafetycentre.com.au/riskranger.php
Anonymous (2005). Center for Meat Process Validation. Geraadpleegd op 18 februari 2017,
op http://meathaccp.wisc.edu/therm/Default.aspx
Anonymous1 (2009). Predictive microbiological models. What are they and how can they be
used in the food industry?. Geraadpleegd op 8 februari 2017, op
https://www.campdenbri.co.uk/services/predictive-microbiological-models.pdf
Anonymous2 (2009). Shelf Stability Predictor. Geraadpleegd op 18 februari 2017, op
https://meathaccp.wisc.edu/ST_calc.html
Anonymous1 (2012). FAQ Listeria Control Model 2012. Geraadpleegd op 11 februari 2017,
op https://lcm.purac.com/faq.html
Anonymous2 (2012). Flemisch Cluster Predictive Microbiology in Foods. Geraadpleegd op
22 februari 2017, op http://www.cpmf2.be/index.php

72
Anonymous (2013). SOPHY: Software for fresh salads. Geraadpleegd op 18 maart 2017, op
http://www.anugafoodtec.com/redaktionell/aft/downloads_22/pdf_24/quarterly_news/
quarterly_maerz_1/Sophy.pdf
Anonymous1 (2015). ComBase-2015. Geraadpleegd op 15 oktober 2016, op
http://www.ComBase.cc/images/help/CB_Tutorial.pdf
Anonymous2 (2015). SOPHY-Development of a SOftware tool for Prediction of ready-to-eat
food product sHelf life, quality and safetY. Geraadpleegd op 18 maart 2017, op
https://biobs.jrc.ec.europa.eu/development-software-tool-prediction-ready-eat-food-
product-shelf-life-quality-and-safety
Anonymous1 (2016). Refrigeration Index Calculator. Geraadpleegd op 9 februari 2017, op
http://ricalculator.mla.com.au/
Anonymous2 (2016). Pathogen Modeling Program – Getting Started. Geraadpleegd op 11
februari 2017, op https://www.ars.usda.gov/northeast-area/wyndmoor-pa/eastern-
regional-research-center/residue-chemistry-and-predictive-microbiology-
research/docs/pathogen-modeling-program/pathogen-modeling-program-getting-
started/
Anonymous3 (2016). The European Union summary report on trends and sources of zoonoses,
zoonotic agents and food-borne outbreaks in 2015. EFSA Journal, 14, (12): 1-231.
Anonymous3 (2009). Predicting microbial spoilage. Geraadpleegd op 18 februari 2017, op
https://www.campdenbri.co.uk/news/mar09a.php
Anonymous3 (2012). Sym’Previus Software. Geraadpleegd op 4 maart 2017 op
http://www.adriafoodsafety.com/index.php?ctn=pub_aff_article.414&rch=sym%5C%
27previus#pub_aff_article.414
Anonymous3 (2015). BACILLUS CEREUS. Geraadpleegd op 8 november 2016, op
http://www.foodsafety.govt.nz/elibrary/industry/Bacillus_Cereus-Spore_Forming.pdf
Anonymous1 (2017). ComBase. A Web Resource for Quantitative and Predictive Food
Microbiology. Geraadpleegd op 7 februari 2017, op
http://www.ComBase.cc/index.php/en/
Anonymous2 (2017). Microbial Responses Viewer. Geraadpleegd op 9 februari 2017, op
http://mrviewer.info/#
Anonymous3 (2017). Software. Geraadpleegd op 4 maart 2017, op
https://symprevius.eu/software/

73
Barrile, J., Cone, F. (1970). Effect of Added Moisture on the Heat Resistance of Salmonella
anatum in Milk Chocolate. Applied Microbiology, 19, (1), 177-178.
Beaufort, A., Bergis, H., Lardeux, A.-L., Lombard, B. (2014). EURL Lm TECHNICAL
GUIDANCE DOCUMENT. for conducting shelf-life studies on Listeria
monocytogenes in ready-to-eat foods. Geraadpleegd op 12 april 2017, op
https://ec.europa.eu/food/sites/food/files/safety/docs/biosafety_fh_mc_technical_guida
nce_document_listeria_in_rte_foods.pdf
Betts, G. D., Gaze, J. E. (1995). Growth and heat resistance of psychrotrophic Clostridium
botulinum in relation to ‘sous vide’ products. Food Control, 6, (1), 57-63.
Bolton, D. J., McMahon, C. M., Doherty, A. M., Sheridan, J. J., McDowell, D. A., Blair, I. S.,
Harrington, D. (2000). Thermal inactivation of Listeria monocytogenes and Yersinia
enterocolitica in minced beef under laboratory conditions and in sous-vide prepared
minced and solid beef cooked in a commercial retort. Journal of Applied
Microbiology, 88, (4), 626-632.
Bucknavage, M. W., Pierson, M. D., Hackney, C. R., Bishop, J. R. (1990). Thermal
inactivation of Clostridium botulinum Type E Spores in Oyster Homogenates at
Minimal Processing Temperatures. Journal of Food Science, 55, (2), 372-373.
Burgos, J., Ordóñez, A. J., Sala, F. (1972). Effect of Ultrasonic Waves on the Heat Resistance
of Bacillus cereus and Bacillus licheniformis Spores. Applied Microbiology, 24, (3),
497-498.
Byrne, B., Dunne, G., Bolton, D. J. (2006). Thermal inactivation of Bacillus cereus and
Clostridium perfringens vegetative cells and spores in pork luncheon roll. Food
Microbiology, 23, (8),803 – 808.
Crawford, R. G., Beliveau, C. M., Peeler, J. T., Donelly, C. W., Bunning, K. V. (1989).
Comparative Recovery of Uninjured and Heat-injured Listeria monocytogenes Cells
from Bovine Milk. Applied and environmental microbiology, 55, (6), 1490 – 1494.
Dalgaard, P. (2014). Annual Report on Zoonoses in Denmark 2013. Food Spoilage and Safety
Predictor (FSSP) Software. Søborg: DTU Food.
Devlieghere, F., Vermeulen, A., Debevere, J., Ragaert, P., Baert, L., Uyttendaele, M. (2014)
Levensmiddelenmicrobiologie [cursustekst]. Gent: UGent.
Faiku, F., Haziri, A., Mehmeti, I., Barjrami, D., Haziri, I. (2016). Evaluation of antibacterial
activity of different solvent extracts of cichorium intybus (L.) growing wild in east part
of Kosovo. The Journal of Animal & Plant Sciences, 26, (5), 1486-1491.

74
Francois, K., Devlieghere, F., Uyttendaele, M., Debevere, J. (2006). Risk assessment of Listeria
monocytogenes: Impact of Individual Cell Variability on the Exposure Assessment Step.
Risk Analysis, 26, (1), 105-114.
Gaillard, S., Leguerinel, I., Mafart, P. (1998). Modelling combined effects of temperature and
pH on the heat resistance of spores of Bacillus cereus. Food Microbiology, 15, 625-
630.
Gates, Z. (2009). Pathogen Modeling Programs. Geraadpleegd op 8 februari 2017, op
http://ucfoodsafety.ucdavis.edu/Pathogen_Modeling_Programs/
González, I., López, M., Martínez, S., Bernardo, A., González, J. (1999). Thermal inactivation
of Bacillus cereus spores formed at different temperatures. International Journey of
Food Microbiology, 51, (1), 81-84.
Hauschild, A. H. W., Dodds, K. L. (1993). Clostridium Botulinum: Ecology and Control in
Foods. New York: Marcel Dekker Inc.
Jørgensen, F., Stephens, P. J., Knøchel, S. (1995). The effect of osmotic shock and subsequent
adaptation on the thermotolerance and cell morphology of Listeria monocytogenes.
Journal of Applied Bacteriology, 79, 274-281.
Juneja, V. K., Eblen, B. S., Marmer, B. S., Williams, A. C., Palumbo, S. A., Miller, A. J.
(1994). Thermal Resistance of Nonproteolytic Type B and Type E Clostridium
botulinum Spores in Phosphate Buffer and Turkey Slurry. Journal of Food Protection,
58, (7), 758-763.
Koseki, S. (2009). Microbial Responses Viewer (MRV): A new ComBase-derived database of
microbial responses to food environments. International Journal of Food
Microbiology, 134,(1-2), 75-82.
Leguerinel, I., Spegagne, I., Couvert, O., Gaillard, S., Mafart, P. (2005). Validation of an
overall model describing the effect of three environmental factors on the apparent D-
value of Bacillus cereus spores. International Journey of Food Microbiology, 100, (1-
3), 223-229.
Leporq, B., Membré, J-M., Dervin, C., Guyonnet, J. P. (2005). The “Sym Previus” software, a
tool to support decisions to the foodstuff safety. Internation Journal of Food
Microbiology, 100, (1-3), 231-237.
Liu, D. (2008). Handbook of Listeria monocytogenes. Boca Raton: CRC Press.
Luu-Thi, H., Khadka, D. B., Michiels, C. W. (2014). Thermal inactivation parameters of
spores from different phylogenetic groups of Bacillus cereus. International Journal of
Food Microbiology, 189, 183-188.

75
Lynt, R. K., Solomon, H. M., Lilly Jr., T., Kautter, D. A. (1977). Thermal death time of
Clostridium botulinum type E in meat of the blue crab. Journal of Food Science, 42,
(4), 1022-1025.
Mackey, B. M., Bratchell, N. (1989). The heat resistance of Listeria monocytogenes. Letters
in Applied Mircobiology, 9, (3) 89-94.
Man, D. C. M. (2015). Shelf life. Chichester , Verenigd Koninkrijk, John Wiley & Sons, Ltd.
Mazas, M., Martínez, S., López, M., Alvarez, A. B., Martin, R. (1999). Thermal inactivation
of Bacillus cereus spores affected by the solutes used to control water activity of the
heating medium. International Journal of Food Microbiology, 53, (1), 61-67.
Medina, E., de Castro, A., Romero, C., Ramírez, E., Brenes, M. (2013). Effect of
antimicrobial compounds from olive products on microorganisms related to health,
food and agriculture. Microbial pathogens and strategies for combacting them:
science, technology and education, 4, (2), Spanje, Formatex Research Center
Medina, E., Romero, C., Brenes, M., De Castro, A. (2007). Antimicrobial activity of olive oil,
vinegar, and various beverages against foodborn pathogenes. Journal of Food
Protection, 70, (5), 1194-1199.
Molin, N., Snygg, B. G. (1967). Effect of Lipid Materials on Heat Resistance of Bacterial
Spore. Applied Microbiology, 15, (6), 1422-1426.
Pagán, R., Condón, S., Sala, F. J. (1997). Effects of Several Factos on the Heat-Shock-
Induced Thermotolerance of Listeria monocytogenes. Applied and Environmental
Microbiology, 63, (8), 3225-3232.
Palumbo, M. S., Beers, S. M., Bhaduri, S., Palumbo, S. A. (1995). Thermal Resistance of
Salmonella spp. and Listeria monocytogenes in Liquid Egg Yolk and Egg Yolk
Products. Journal of food protection, 58, (9), 960-966.
Pérez-Rodríguez, F., & Valero, A. (2013). Predictive Microbiology in Foods. SpringerBriefs
in Food, Health and Nutrition(5e e). Spanje: Springer.
Psomas, A. N., Skandamis, P. N. (2007). Geraadpleegd op 29 april 2017 via
http://www.aua.gr/psomas/gropin/
Reij, M. W., Den Aantrekker, E. D. (2004). Recontamination as a source of pathogens in
processed foods. International Journal of Food Microbiology, 91, 1-11.
Rhee, M-S., Lee, S-Y., Dougherty, R. H, Kang, D-H. (2003). Applied and Environmental
Microbiology, 69, (5), 2959-2963.

76
Ross, T. (1996). Indices for performance evaluation of predictive models in food
microbiology. Journal of Applied Bacteriology, 81,(5), 501-508.
Ross, T., Sumner, J. (2002). A simple, spreadsheet-based, food safety risk assessment tool.
International Journal of Food Microbiology, 77, 39-53.
Rosso, L., Lobry, J. R., Bajard, S., Flandrois, J. P. (1995). Convenient Model to Describe the
Combined Effects of Temperature and pH on Microbial Growth. Applied and
Environmental Microbiology, 61, (2), 610-616.
Samapundo, S., Heyndrickx, M., Xhaferi, R., de Baenst, I., Devlieghere, F. (2014). The
combined effect of pasteurization intensity, water activity, pH and incubation
temperature on the survival and outgrowth of spores of Bacillus cereus and Bacillus
pumilus in artificial media and food products. International Journal of Food
Microbiology, 181, 10-18.
Schutyser, M.A.I., Straatsma, J., Keijzer, P.M., Verschueren, M., De Jong, P. (2008). A new
web-based modelling tool (Websim-MILQ) aimed at optimisation of thermal
treatments in the dairy industry. International Journal of Food Microbiology, 128,
153-157.
Shen, Q., Jangam, P. M., Soni, K. A., Nannapaneni, R., Schilling, W., Silva, J. L. (2014).
Low, Medium, and High Heat Tolerant Strains of Listeria monocytogenes and
Increased Heat Stress Resistance after Exposure to Subletal Heat. Journal of Food
Protection, 77, (8), 1298-1307.
Silk, B., Mahon, B., Griffin, P., Gould, H., Tauxe, R., Crim, S., Jackson, K. Gerner-Smidt, P.,
Herman, K., Henao, O. (2013). Vital Signs: Listeria Illness, Deaths, and Outbreaks –
United States, 2009-2011. Geraadpleegd op 15 april 2017, op
https://www.cdc.gov/mmwr/preview/mmwrhtml/mm6222a4.htm
Silva, F. V. M., Gibbs, P. A. (2012). Thermal pasteurization requirements for the inactivation
of Salmonella in foods. Food Research International, 45, (2), 695-699.
Van Dijk, J., Schuytser, M. A. I., Horak, P., Gnip, P. (2008). Final Activity Report: the
development of predictive models on the Internet for optimisation of heat treatment of
raw milk in small medium-sized dairy companies. MILQ-QC-TOOL.
Van Helleputte, M. (2016). Ontwikkelen van een evaluatietool voor de beoordeling van de
microbiële stabiliteit van halffabricaten. Thesis, Universiteit Gent.
Verledens, L., Rawoens, W. (2017). Voedingstechnologie op ambachtelijke wijze. Trends, 9,
103-105.

77
Vermeulen, A. 2008. Microbial stability and safety of acid sauces and mayonnaise-based
salads assessed through probabilistic growth/no growth models Thesis submitted in
fulfillment of the requirements for the degree of doctor (Ph.D.) in Applied Biological
Sciences. Faculty of Bioscience Engineering, University of Ghent
Wachnicka, E., Stringer, S. C., Barker, G. C., Peck, M. W. (2016). Systematic Assessment of
Nonproteolytic Clostridium botulinum Spores for Heat Resistance. Applied and
Environmental Microbiology, 82, (19), 6019-6029.
Walker, S. J., Archer, P., Banks, J. G. (1990). Growth of Listeria monocytogenes at
refrigeration temperatures. Journal of Applied Bacteriology, 68, 157-162.
Xu, S., Labuza, T. P., Diez-Gonzalez, F. (2006). Thermal Inactivation of Bacillus anthracis
Spores in Cow’s Milk. Applied and Environmental Microbiology, 72, (6), 4479-4483.
Youssef, A. E., Juneja, V. K. (2003). Microbial Stress Adaptation and Food Safety. [EPub],
Boca Raton, CRC Press.
Zwietering, M. H., Wijtzes, T., Rombouts, F. M., van’t Riet, K. (1993). A decision support
system for prediction of microbial spoilage in foods. Journal of Industrial
Microbiology, 12, 324-349.

78
79
Bijlage 1: Groeimodellen voor fysiologische staat met een waarschijnlijkheid van 100 %.
Bacillus cereus
Dag 7
Dag 14
Dag 21
Salmonella spp.
Dag 7
Dag 14
Dag 21
Figuur 2: Groei/ niet groei voorstelling van Bacillus
cereus na 7, 14 en 21 dagen. Figuur 2: Groei/ niet groei voorstelling van
Salmonella spp. na 7, 14 en 21 dagen.
80
Niet-proteolytische Clostridium
botulinum bij 4°C
Dag 7
Dag 14
Dag 21
Niet-proteolytische Clostridium
botulinum bij 7°C
Dag 7
Dag 14
Dag 21
Figuur 3: Groei/ niet groei voorstelling van de niet-
proteolytische Clostridium botulinum na 7, 14 en 21
dagen bij 4 °C.
Figuur 4: Groei/ niet groei voorstelling van de niet-
proteolytische Clostridium botulinum na 7, 14 en 21
dagen bij 4 °C.
81
Listeria monocytogenes bij 4°C
Dag 7
Dag 14
Dag 21
Listeria monocytogenes bij 7°C
Dag 7
Dag 14
Dag 21
Figuur 5: Groei/ niet groei voorstelling van Listeria
monocytogenes na 7, 14 en 21 dagen bij 4 °C. Figuur 6: Groei/ niet groei voorstelling van Listeria
monocytogenes na 7, 14 en 21 dagen bij 7 °C.
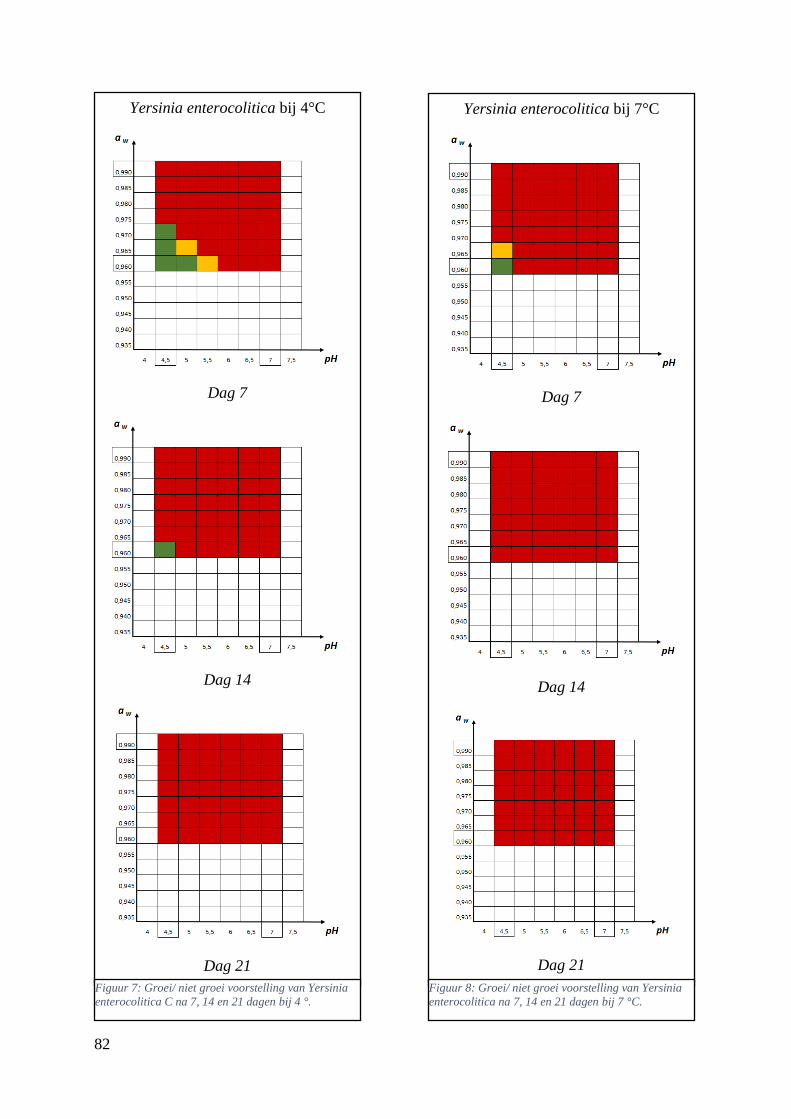
82
Yersinia enterocolitica bij 4°C
Dag 7
Dag 14
Dag 21
Yersinia enterocolitica bij 7°C
Dag 7
Dag 14
Dag 21
Figuur 7: Groei/ niet groei voorstelling van Yersinia
enterocolitica C na 7, 14 en 21 dagen bij 4 °.
Figuur 8: Groei/ niet groei voorstelling van Yersinia
enterocolitica na 7, 14 en 21 dagen bij 7 °C.
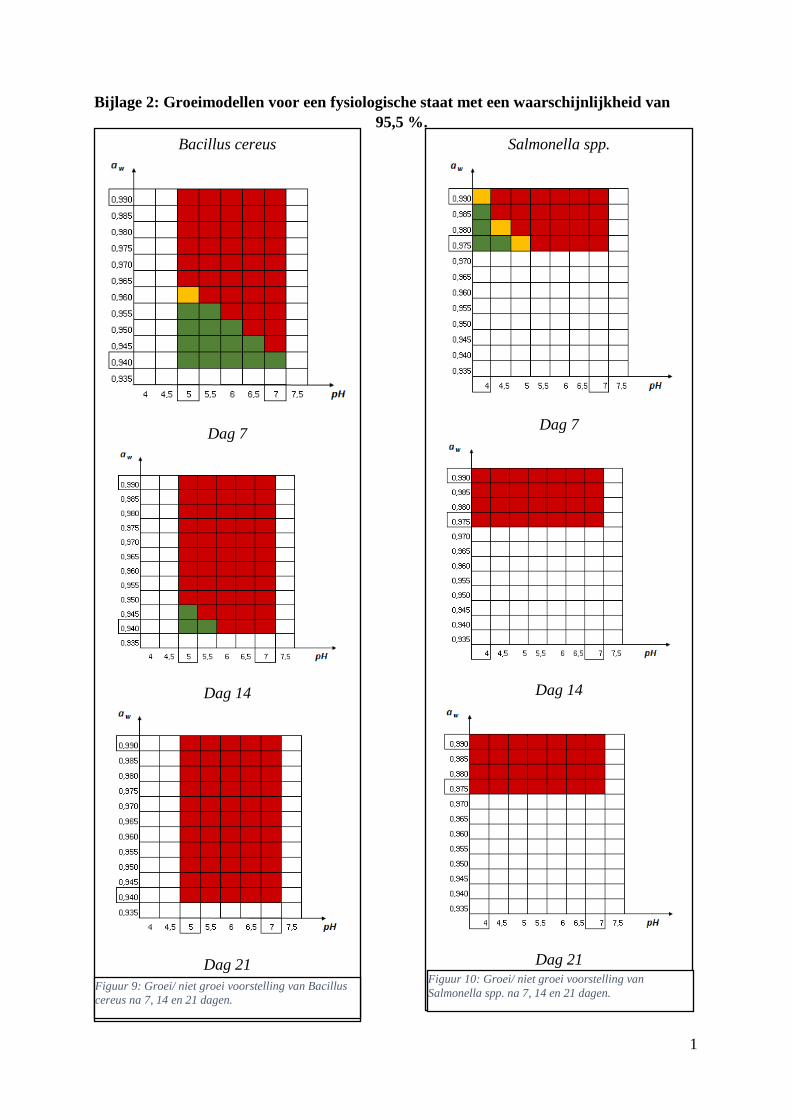
1
Bijlage 2: Groeimodellen voor een fysiologische staat met een waarschijnlijkheid van
95,5 %.
Bacillus cereus
Dag 7
Dag 14
Dag 21
Salmonella spp.
Dag 7
Dag 14
Dag 21
Figuur 9: Groei/ niet groei voorstelling van Bacillus
cereus na 7, 14 en 21 dagen.
Figuur 10: Groei/ niet groei voorstelling van
Salmonella spp. na 7, 14 en 21 dagen.
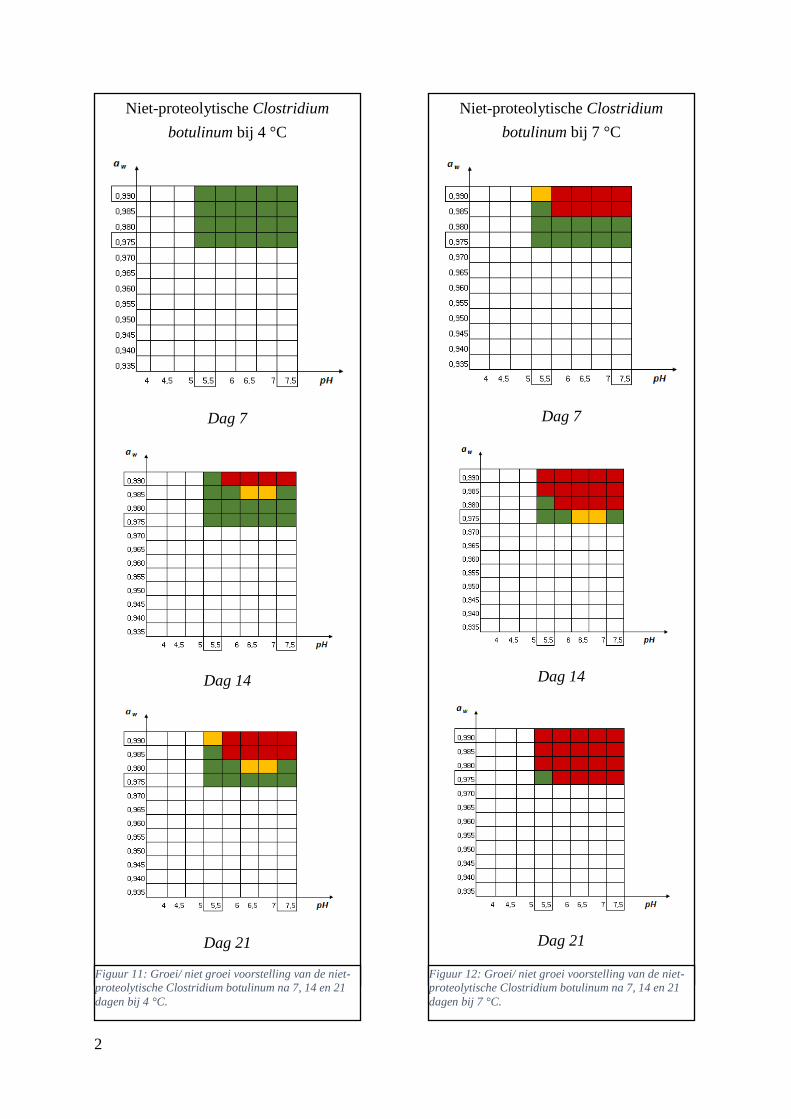
2
Niet-proteolytische Clostridium
botulinum bij 4 °C
Dag 7
Dag 14
Dag 21
Niet-proteolytische Clostridium
botulinum bij 7 °C
Dag 7
Dag 14
Dag 21
Figuur 11: Groei/ niet groei voorstelling van de niet-
proteolytische Clostridium botulinum na 7, 14 en 21
dagen bij 4 °C.
Figuur 12: Groei/ niet groei voorstelling van de niet-
proteolytische Clostridium botulinum na 7, 14 en 21
dagen bij 7 °C.
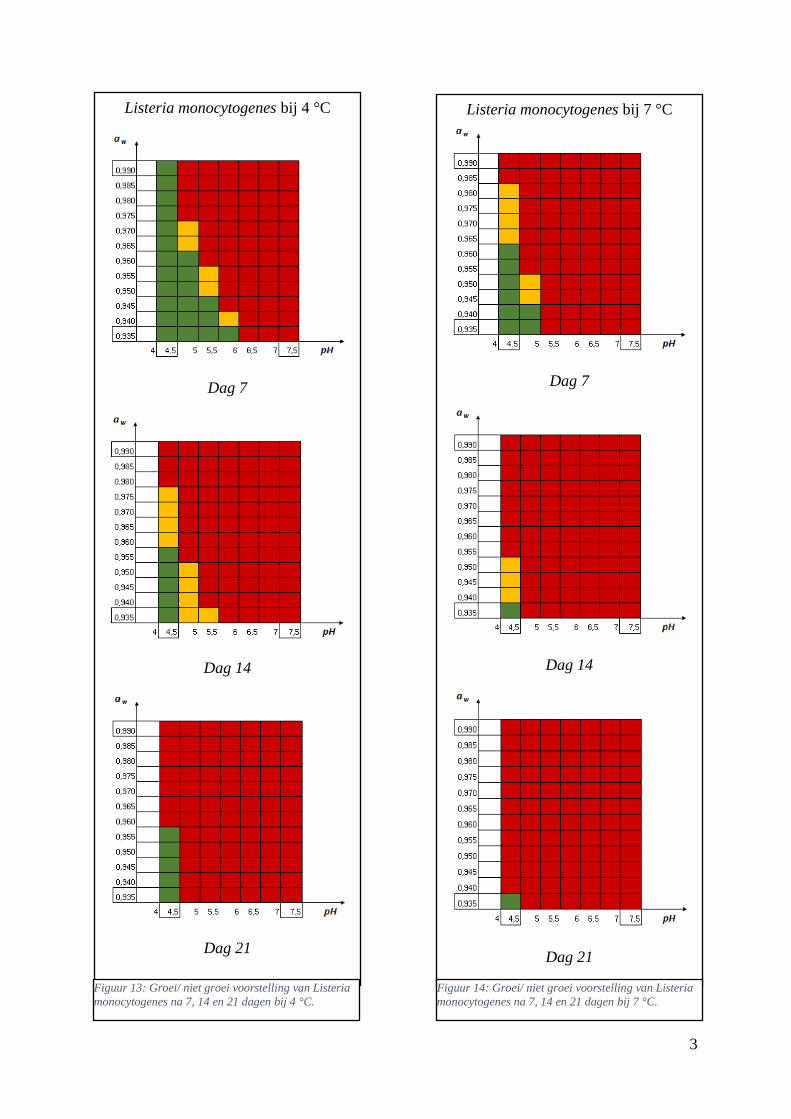
3
Listeria monocytogenes bij 4 °C
Dag 7
Dag 14
Dag 21
Listeria monocytogenes bij 7 °C
Dag 7
Dag 14
Dag 21
Figuur 13: Groei/ niet groei voorstelling van Listeria
monocytogenes na 7, 14 en 21 dagen bij 4 °C.
Figuur 14: Groei/ niet groei voorstelling van Listeria
monocytogenes na 7, 14 en 21 dagen bij 7 °C.
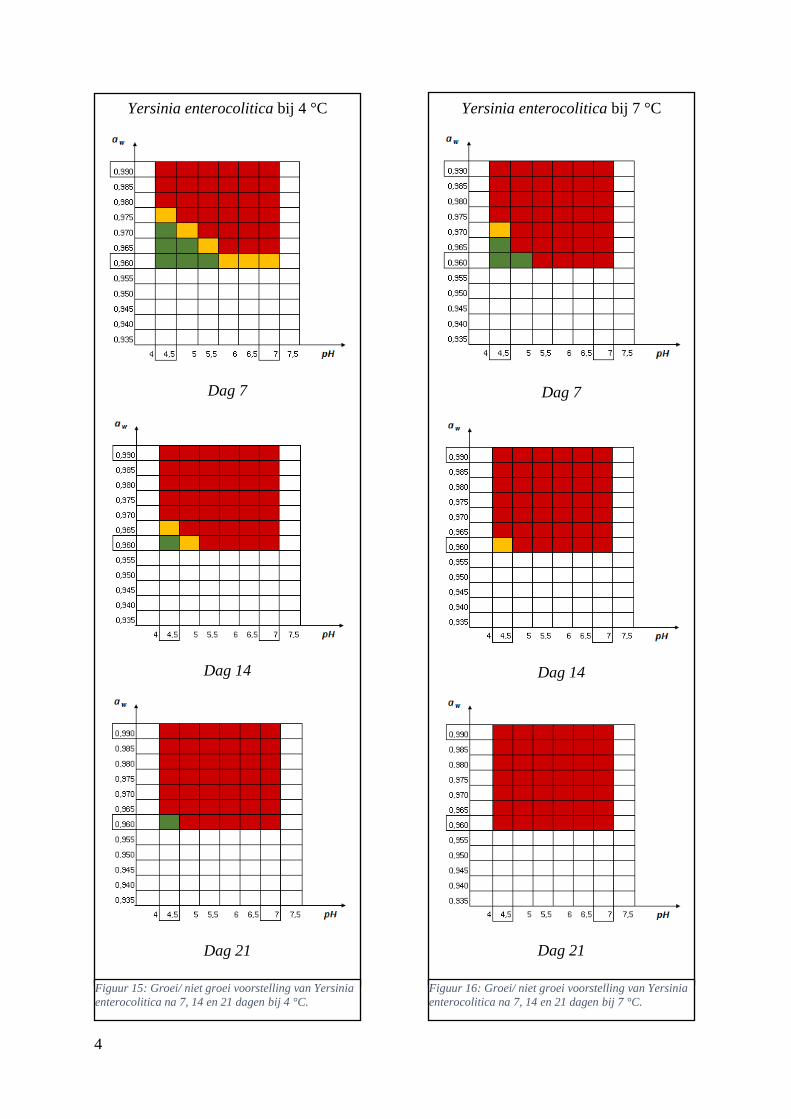
4
Yersinia enterocolitica bij 4 °C
Dag 7
Dag 14
Dag 21
Yersinia enterocolitica bij 7 °C
Dag 7
Dag 14
Dag 21
Figuur 15: Groei/ niet groei voorstelling van Yersinia
enterocolitica na 7, 14 en 21 dagen bij 4 °C.
Figuur 16: Groei/ niet groei voorstelling van Yersinia
enterocolitica na 7, 14 en 21 dagen bij 7 °C.
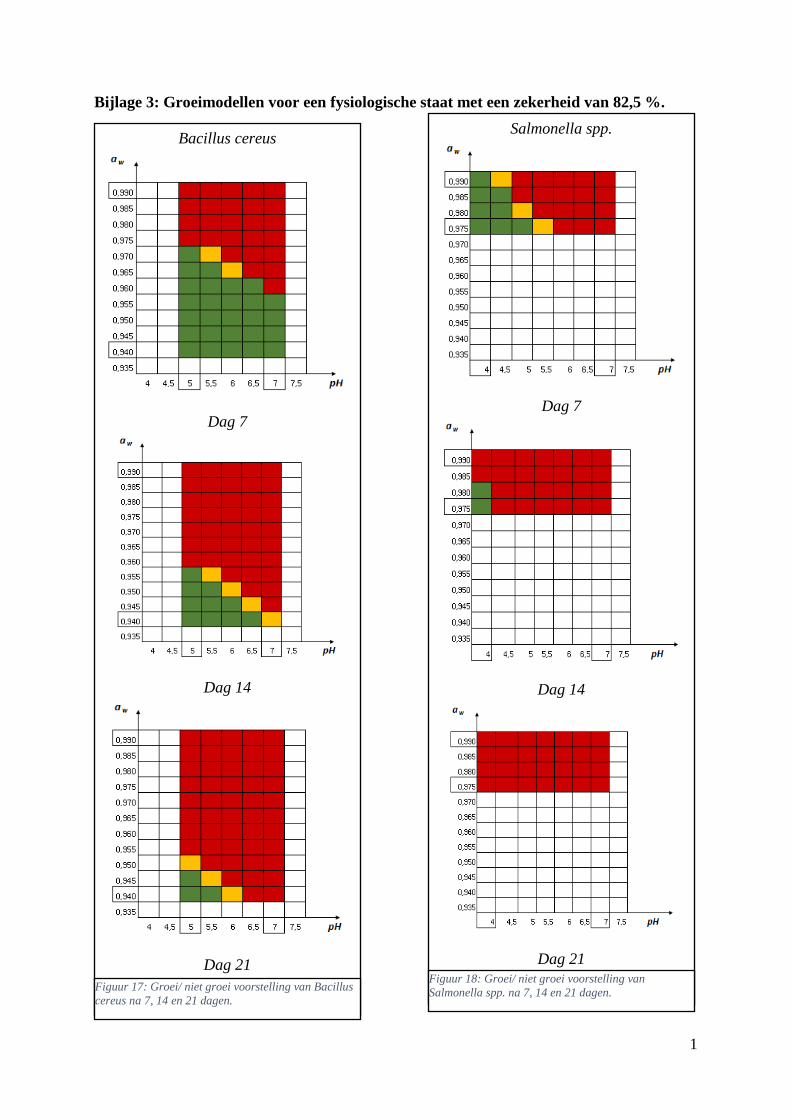
1
Bijlage 3: Groeimodellen voor een fysiologische staat met een zekerheid van 82,5 %.
Bacillus cereus
Dag 7
Dag 14
Dag 21
Salmonella spp.
Dag 7
Dag 14
Dag 21
Figuur 17: Groei/ niet groei voorstelling van Bacillus
cereus na 7, 14 en 21 dagen.
Figuur 18: Groei/ niet groei voorstelling van
Salmonella spp. na 7, 14 en 21 dagen.
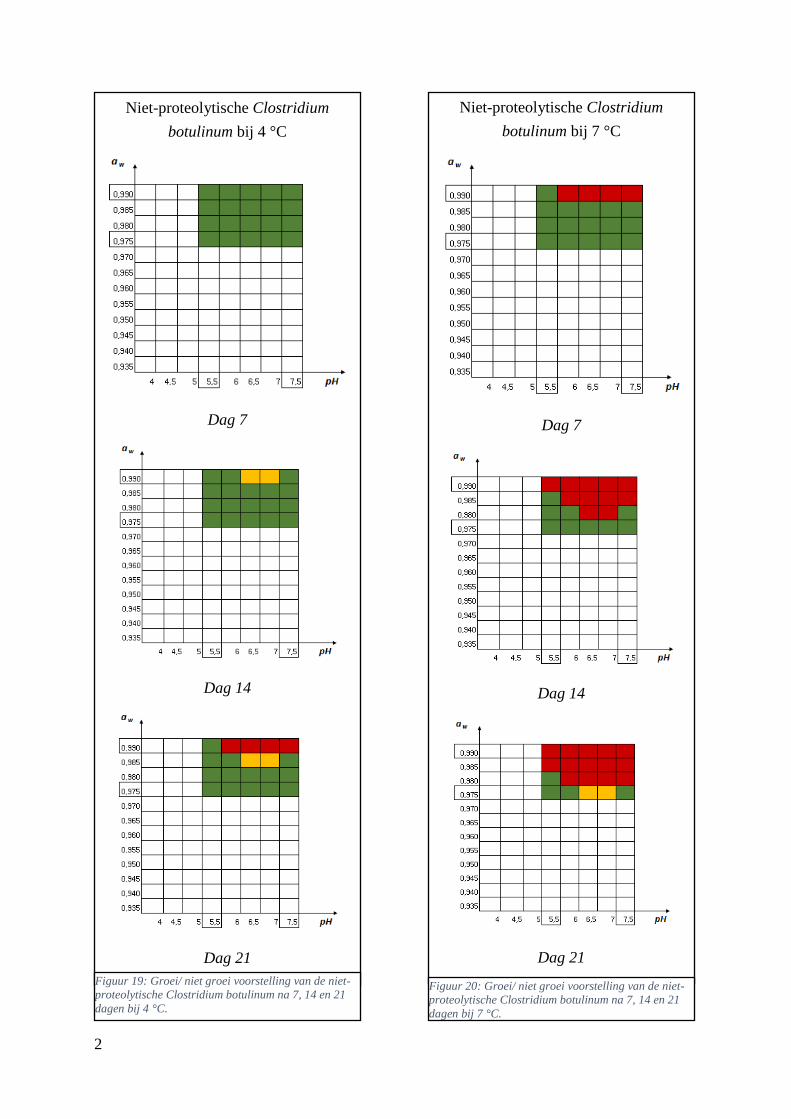
2
Niet-proteolytische Clostridium
botulinum bij 4 °C
Dag 7
Dag 14
Dag 21
Niet-proteolytische Clostridium
botulinum bij 7 °C
Dag 7
Dag 14
Dag 21
Figuur 19: Groei/ niet groei voorstelling van de niet-
proteolytische Clostridium botulinum na 7, 14 en 21
dagen bij 4 °C.
Figuur 20: Groei/ niet groei voorstelling van de niet-
proteolytische Clostridium botulinum na 7, 14 en 21
dagen bij 7 °C.
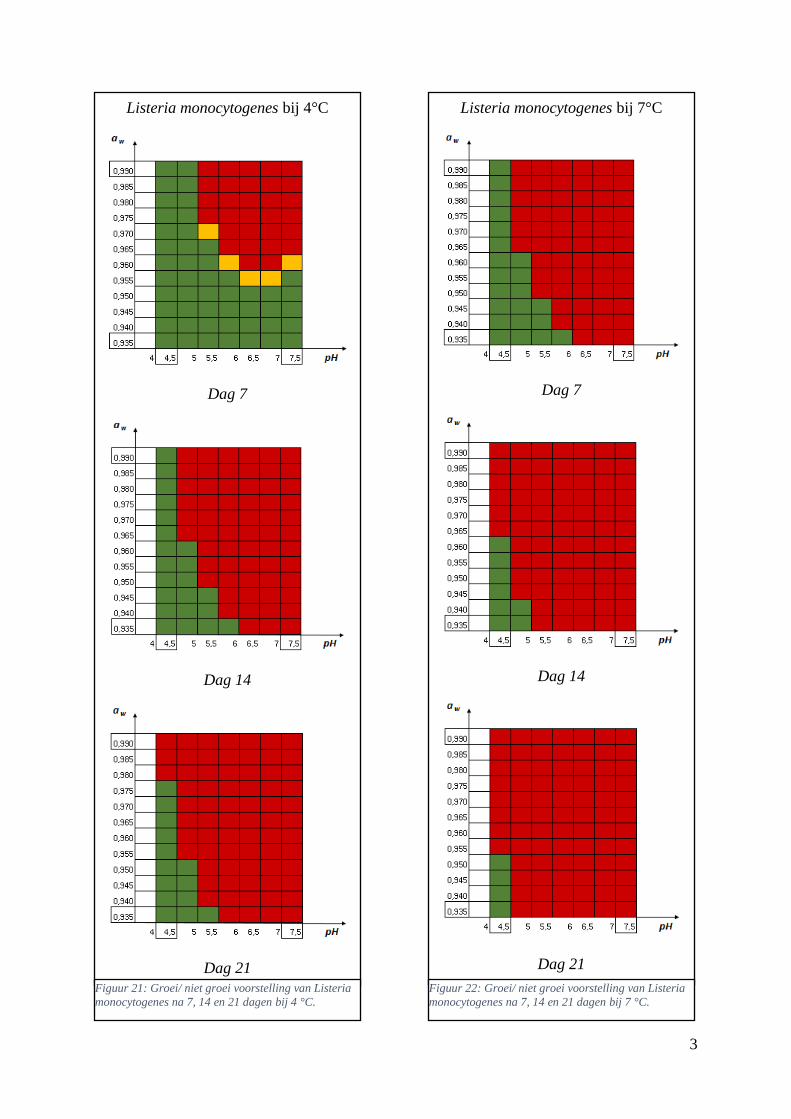
3
Listeria monocytogenes bij 4°C
Dag 7
Dag 14
Dag 21
Listeria monocytogenes bij 7°C
Dag 7
Dag 14
Dag 21
Figuur 21: Groei/ niet groei voorstelling van Listeria
monocytogenes na 7, 14 en 21 dagen bij 4 °C.
Figuur 22: Groei/ niet groei voorstelling van Listeria
monocytogenes na 7, 14 en 21 dagen bij 7 °C.
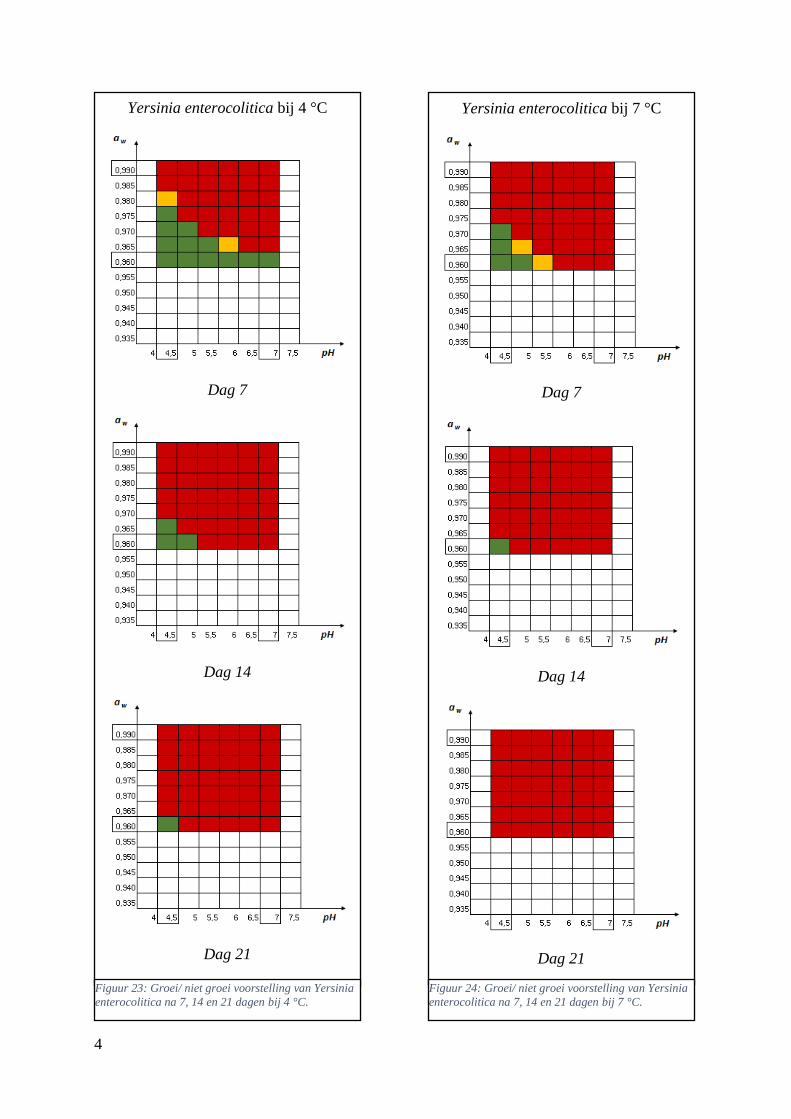
4
Yersinia enterocolitica bij 4 °C
Dag 7
Dag 14
Dag 21
Yersinia enterocolitica bij 7 °C
Dag 7
Dag 14
Dag 21
Figuur 23: Groei/ niet groei voorstelling van Yersinia
enterocolitica na 7, 14 en 21 dagen bij 4 °C.
Figuur 24: Groei/ niet groei voorstelling van Yersinia
enterocolitica na 7, 14 en 21 dagen bij 7 °C.
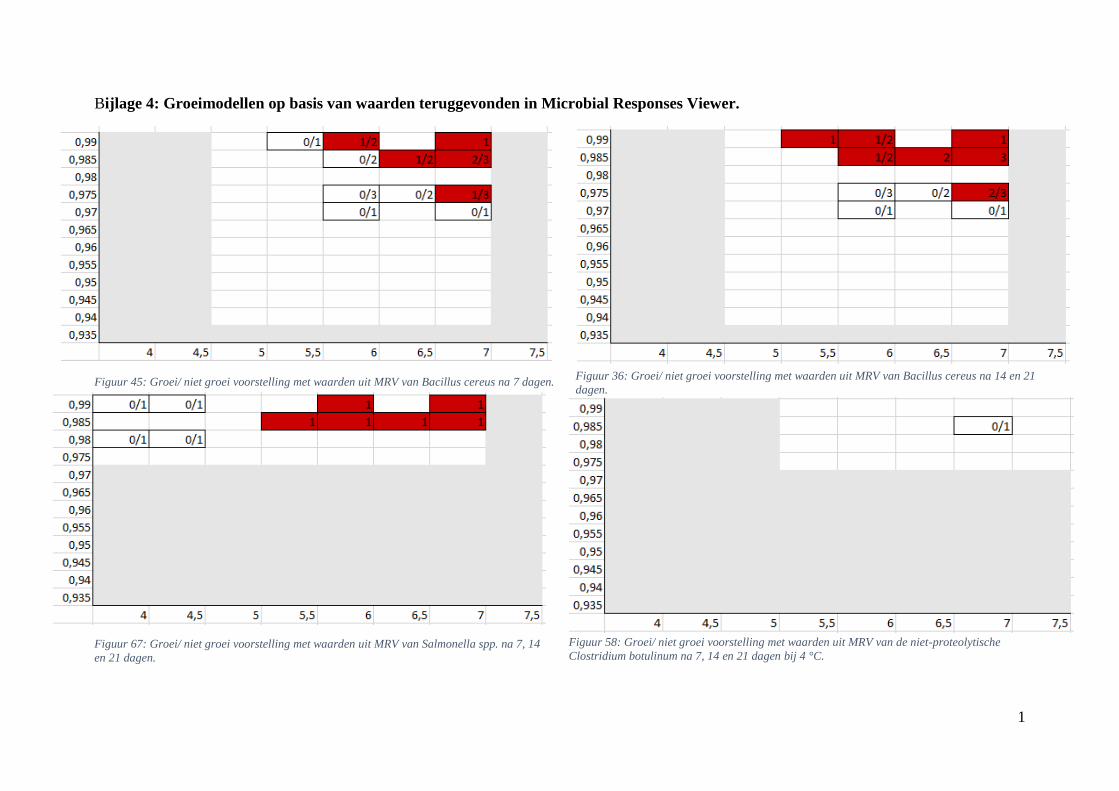
1
Bijlage 4: Groeimodellen op basis van waarden teruggevonden in Microbial Responses Viewer.
Figuur 45: Groei/ niet groei voorstelling met waarden uit MRV van Bacillus cereus na 7 dagen. Figuur 36: Groei/ niet groei voorstelling met waarden uit MRV van Bacillus cereus na 14 en 21
dagen.
Figuur 67: Groei/ niet groei voorstelling met waarden uit MRV van Salmonella spp. na 7, 14
en 21 dagen.
Figuur 58: Groei/ niet groei voorstelling met waarden uit MRV van de niet-proteolytische
Clostridium botulinum na 7, 14 en 21 dagen bij 4 °C.
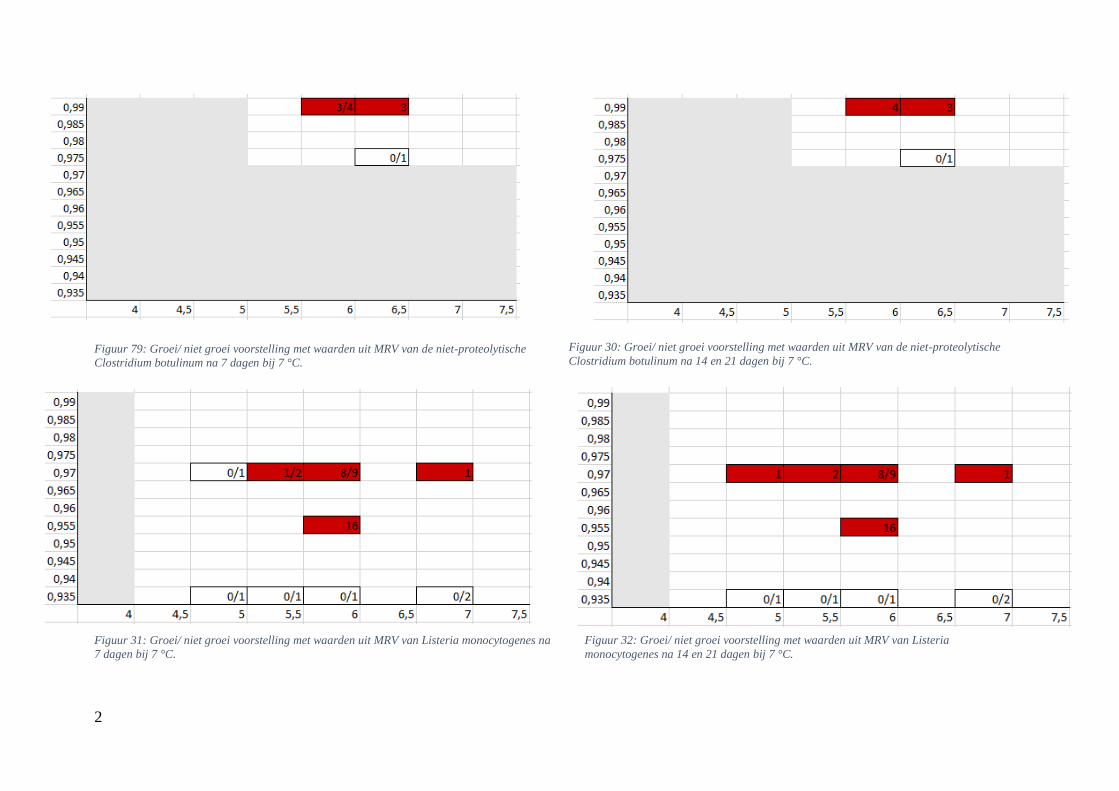
2
Figuur 79: Groei/ niet groei voorstelling met waarden uit MRV van de niet-proteolytische
Clostridium botulinum na 7 dagen bij 7 °C.
Figuur 30: Groei/ niet groei voorstelling met waarden uit MRV van de niet-proteolytische
Clostridium botulinum na 14 en 21 dagen bij 7 °C.
Figuur 31: Groei/ niet groei voorstelling met waarden uit MRV van Listeria monocytogenes na
7 dagen bij 7 °C.
Figuur 32: Groei/ niet groei voorstelling met waarden uit MRV van Listeria
monocytogenes na 14 en 21 dagen bij 7 °C.
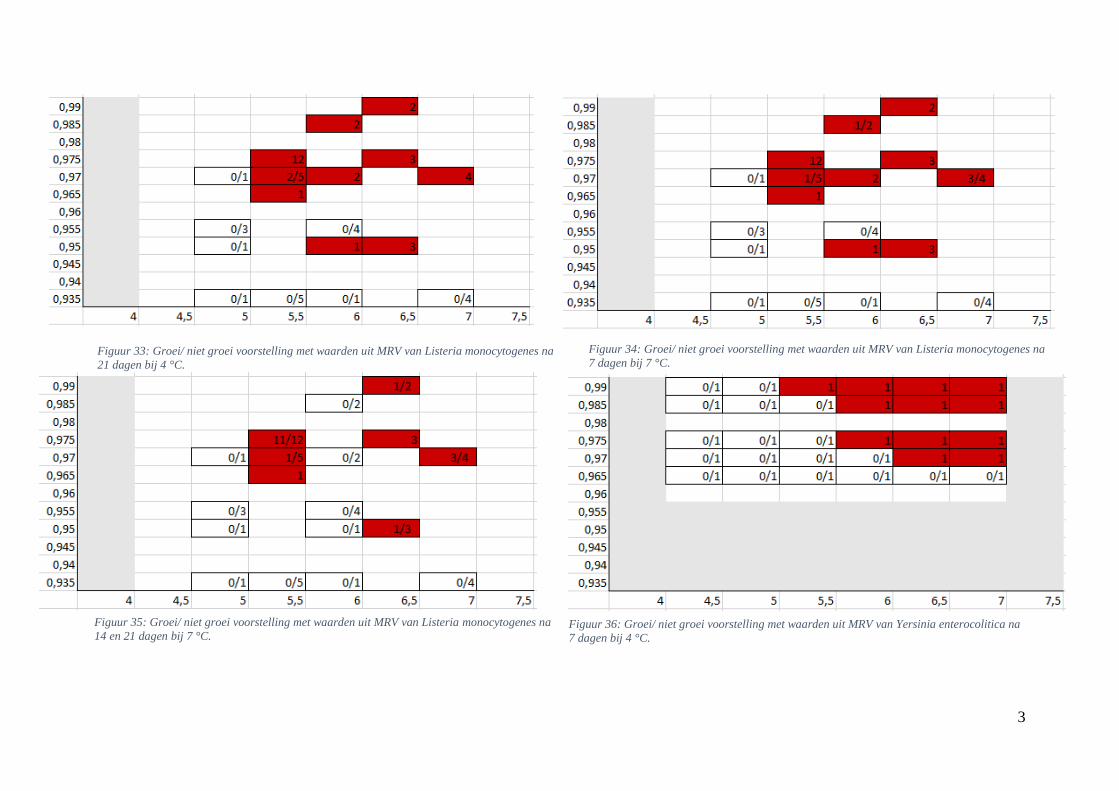
3
Figuur 33: Groei/ niet groei voorstelling met waarden uit MRV van Listeria monocytogenes na
21 dagen bij 4 °C.
Figuur 34: Groei/ niet groei voorstelling met waarden uit MRV van Listeria monocytogenes na
7 dagen bij 7 °C.
Figuur 35: Groei/ niet groei voorstelling met waarden uit MRV van Listeria monocytogenes na
14 en 21 dagen bij 7 °C. Figuur 36: Groei/ niet groei voorstelling met waarden uit MRV van Yersinia enterocolitica na
7 dagen bij 4 °C.
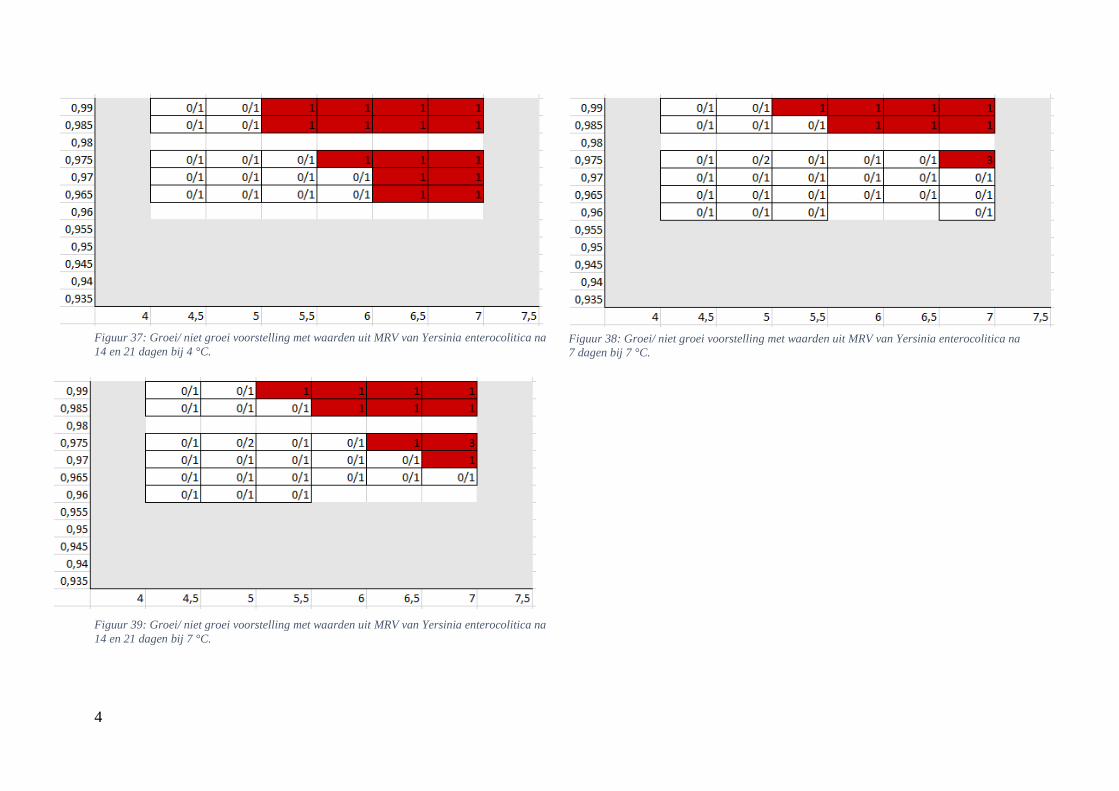
4
Figuur 37: Groei/ niet groei voorstelling met waarden uit MRV van Yersinia enterocolitica na
14 en 21 dagen bij 4 °C. Figuur 38: Groei/ niet groei voorstelling met waarden uit MRV van Yersinia enterocolitica na
7 dagen bij 7 °C.
Figuur 39: Groei/ niet groei voorstelling met waarden uit MRV van Yersinia enterocolitica na
14 en 21 dagen bij 7 °C.
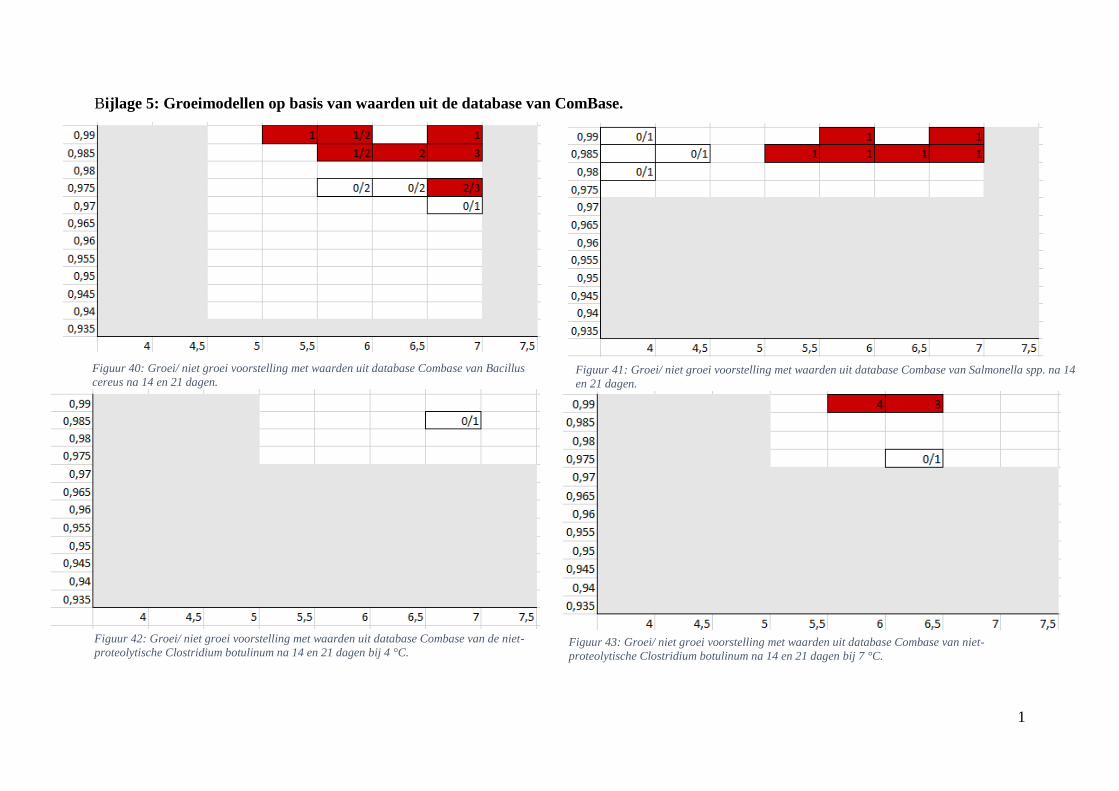
1
Bijlage 5: Groeimodellen op basis van waarden uit de database van ComBase.
Figuur 40: Groei/ niet groei voorstelling met waarden uit database Combase van Bacillus
cereus na 14 en 21 dagen. Figuur 41: Groei/ niet groei voorstelling met waarden uit database Combase van Salmonella spp. na 14
en 21 dagen.
Figuur 42: Groei/ niet groei voorstelling met waarden uit database Combase van de niet-
proteolytische Clostridium botulinum na 14 en 21 dagen bij 4 °C. Figuur 43: Groei/ niet groei voorstelling met waarden uit database Combase van niet-
proteolytische Clostridium botulinum na 14 en 21 dagen bij 7 °C.
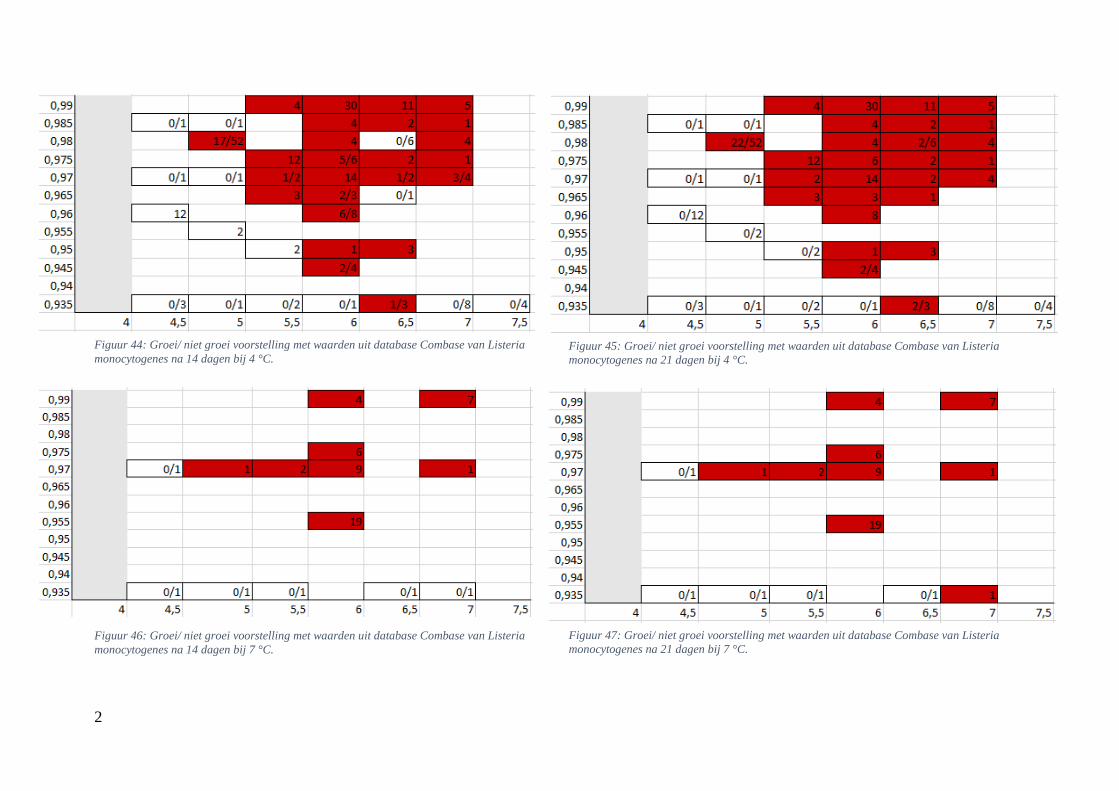
2
Figuur 45: Groei/ niet groei voorstelling met waarden uit database Combase van Listeria
monocytogenes na 21 dagen bij 4 °C.
Figuur 46: Groei/ niet groei voorstelling met waarden uit database Combase van Listeria
monocytogenes na 14 dagen bij 7 °C.
Figuur 47: Groei/ niet groei voorstelling met waarden uit database Combase van Listeria
monocytogenes na 21 dagen bij 7 °C.
Figuur 44: Groei/ niet groei voorstelling met waarden uit database Combase van Listeria
monocytogenes na 14 dagen bij 4 °C.
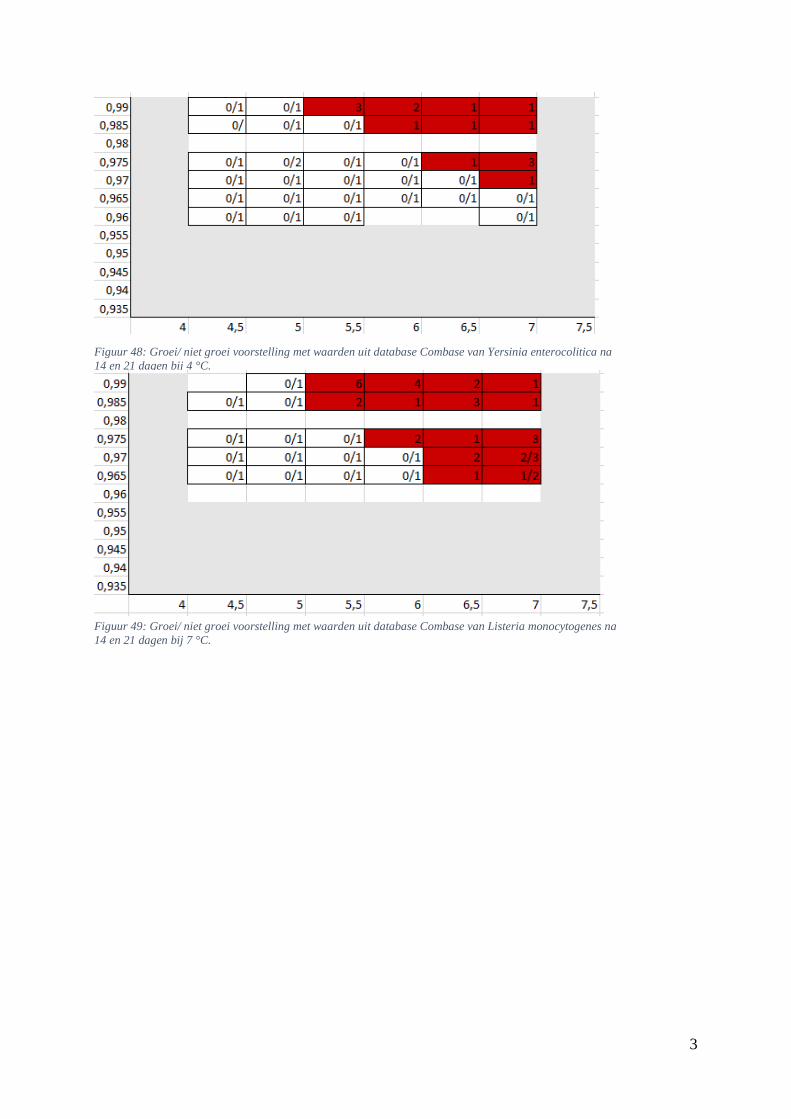
3
Figuur 48: Groei/ niet groei voorstelling met waarden uit database Combase van Yersinia enterocolitica na
14 en 21 dagen bij 4 °C.
Figuur 49: Groei/ niet groei voorstelling met waarden uit database Combase van Listeria monocytogenes na
14 en 21 dagen bij 7 °C.

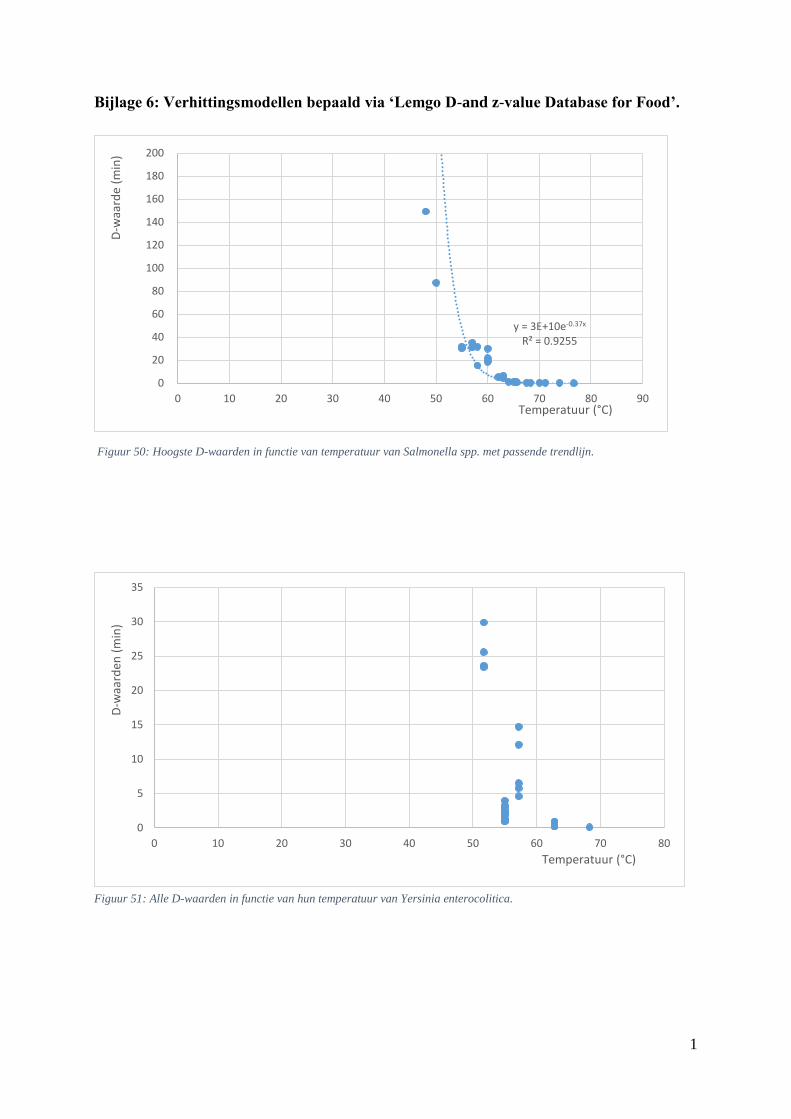
1
Bijlage 6: Verhittingsmodellen bepaald via ‘Lemgo D-and z-value Database for Food’.
y = 3E+10e-0.37x
R² = 0.9255
0
20
40
60
80
100
120
140
160
180
200
0 10 20 30 40 50 60 70 80 90
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 50: Hoogste D-waarden in functie van temperatuur van Salmonella spp. met passende trendlijn.
0
5
10
15
20
25
30
35
0 10 20 30 40 50 60 70 80
D-w
aard
en (
min
)
Temperatuur (°C)
Figuur 51: Alle D-waarden in functie van hun temperatuur van Yersinia enterocolitica.
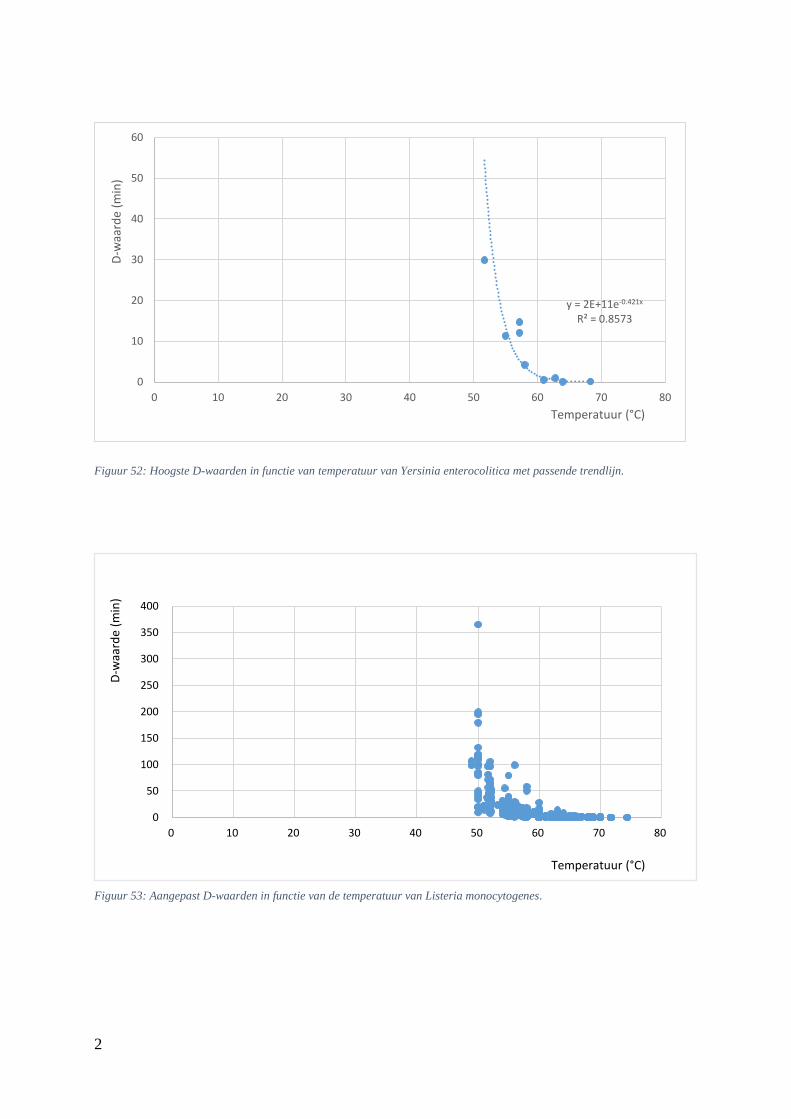
2
Figuur 52: Hoogste D-waarden in functie van temperatuur van Yersinia enterocolitica met passende trendlijn.
0
50
100
150
200
250
300
350
400
0 10 20 30 40 50 60 70 80
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 53: Aangepast D-waarden in functie van de temperatuur van Listeria monocytogenes.
y = 2E+11e-0.421x
R² = 0.8573
0
10
20
30
40
50
60
0 10 20 30 40 50 60 70 80
D-w
aard
e (m
in)
Temperatuur (°C)
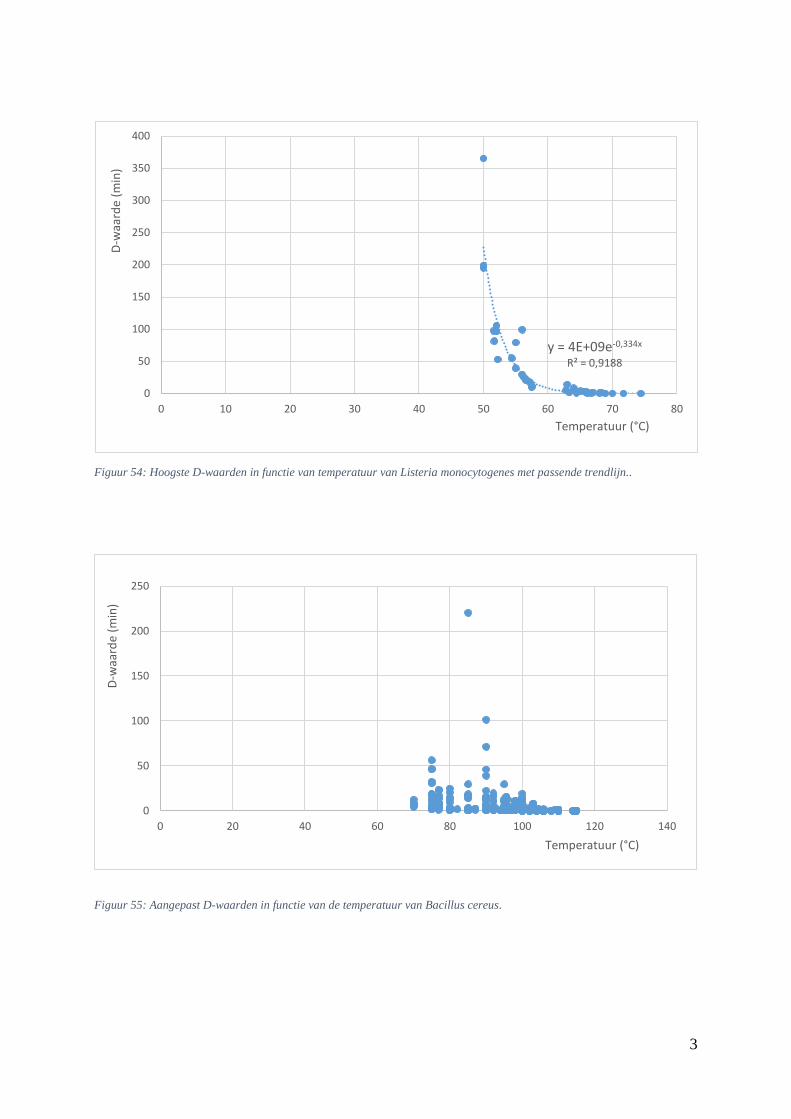
3
y = 4E+09e-0,334x
R² = 0,9188
0
50
100
150
200
250
300
350
400
0 10 20 30 40 50 60 70 80
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 54: Hoogste D-waarden in functie van temperatuur van Listeria monocytogenes met passende trendlijn..
0
50
100
150
200
250
0 20 40 60 80 100 120 140
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 55: Aangepast D-waarden in functie van de temperatuur van Bacillus cereus.
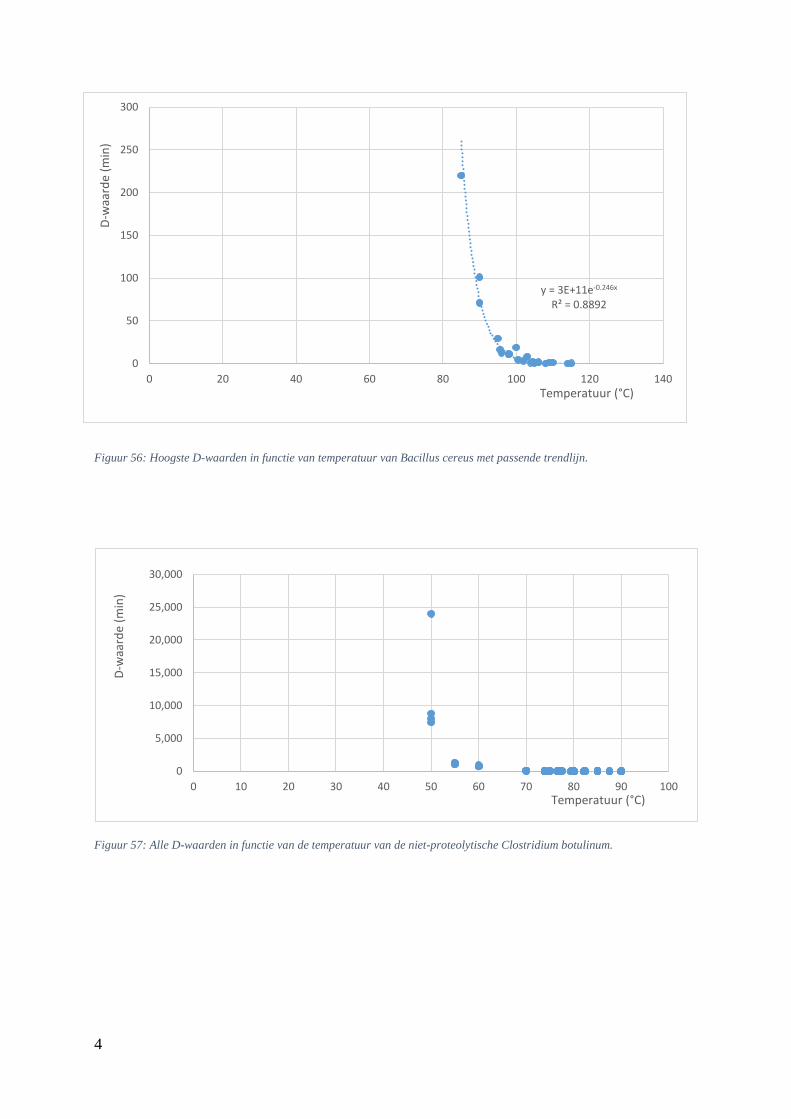
4
y = 3E+11e-0.246x
R² = 0.8892
0
50
100
150
200
250
300
0 20 40 60 80 100 120 140
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 56: Hoogste D-waarden in functie van temperatuur van Bacillus cereus met passende trendlijn.
0
5,000
10,000
15,000
20,000
25,000
30,000
0 10 20 30 40 50 60 70 80 90 100
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 57: Alle D-waarden in functie van de temperatuur van de niet-proteolytische Clostridium botulinum.
5
0
10
20
30
40
50
60
70
0 10 20 30 40 50 60 70 80 90 100
D-w
aard
e (m
in)
Temperatuur (°C)
Figuur 58: Aangepaste D-waarden in functie van de temperatuur van de niet-proteolytische Clostridium botulinum.
Figuur 59: Hoogste D-waarden in functie van temperatuur van de niet-proteolytische Clostridium botulinum met
passende trendlijn.
y = 6E+09e-0.265x
R² = 0.9323
0
20
40
60
80
100
120
140
0 10 20 30 40 50 60 70 80 90 100
D-w
aard
e (m
in)
Temperatuur (°C)


















