
ENZIME PECTINOLITICE DE ORIGINE...
63
Transcript of ENZIME PECTINOLITICE DE ORIGINE...


D. Dinu
Enzime pectinolitice de origine microbiană
D. Dinu
Universitatea din Bucureşti, Facultatea de Biologie, Spl. Independenţei nr. 91-95, Bucureşti
Introducere
Reacţiile biochimice, reacţii deosebit de complexe şi variate, sunt mediate de biocatalizatori cu proprietăţi remarcabile, cunoscuţi sub denumirea de enzime. Capacitatea enzimelor de a creşte de până la 1014ori viteza unei reacţii, condiţiile blânde de acţiune, specificitatea şi posibilitatea de reglare a activităţii, constitue un avantaj major al acestora.
Aplicaţiile practice ale enzimelor în diferite sectoare de activitate au condus la necesitatea obţinerii în cantităţi mari de preparate enzimatice cu diferite activităţi catalitice. În producţia mondială de enzime un loc de frunte îl ocupă tehnologiile care vizează obţinerea de preparate proteice cu activitate pectinolitică. Dezvoltarea biotehnologiilor care urmăresc găsirea de noi surse de izolare a enzimelor pectinolitice este o consecinţă firească a implicării şi eficienţei acestora în diferite procese. Numeroasele utilizări ale enzimelor pectinolitice în cele mai diverse domenii de activitate - industria alimentară, industria prelucrării fibrelor naturale, industria farmaceutică, industria chimică - au determinat demararea cercetărilor în vederea obţinerii acestora din surse microbiene. Orientarea către microorganisme a avut în vedere avantajele oferite de astfel de surse şi proprietăţile enzimelor pectinolitice de origine microbiană.
Larga răspândire, variabilitatea, adaptabilitatea şi potenţialul de biosinteză al microorganismelor pot fi considerate puncte de plecare pentru noi cercetări consacrate descoperirii şi manipulării unor agenţi cu proprietăţi superioare.
1.Pectinele, substraturi ale enzimelor pectinolitice
Substanţele pectice reprezintă un grup heterogen de polizaharide cu un conţinut mare de sarcini negative datorate resturilor de acid galacturonic din structură. Celulele plantelor aflate în diferite stadii de dezvoltare sunt acoperite de un perete extensibil. În structura
163

Enzime pectinolitice de origine microbiană
peretelui celular microfifrilele de celuloză sunt legate necovalent la o matrice alcătuită din hemiceluloze, pectine şi proteine structurale.
Studiile intreprinse asupra pereţilor celulari de provenienţă diferită au evidenţiat un aranjament caracteristic al principalelor componente ale acestora (Albersheim 1975, Cosgrove 1997, Capita şi alţii 2001). În acest aranjament, moleculele adiacente de celuloză aderă puternic prin legături de hidrogen şi formează agregate cristaline, numite microfibrile de celuloză. Microfibrilele de celuloză sunt principalii determinanţi ai rezistenţei peretelui celular. Seturile de microfibrile sunt aranjate în straturi sau lamele în structura cărora fiecare microfibrilă se leagă de vecina sa prin molecule de hemiceluloză. Hemicelulozele sunt heteropolizaharide care se leagă la suprafaţa microfibrilelor de celuloză, mai ales prin legături de hidrogen. Hemicelulozele ajută la legarea încrucişată a microfibrilelor într-o reţea complexă şi previn contactul direct microfibrilă-microfibrilă. Alături de această reţea alcătuită din microfibrile celulozice şi hemiceluloze, peretele celular conţine şi o altă reţea formată din molecule de pectină legate încrucişat. Pectinele formează un gel în jurul reţelei celuloză-hemiceluloză şi acţionează ca un material de umplutură care previne agregarea şi ruperea acestei reţele. Prin intermediul substanţelor pectice peretele celular îşi modulează porozitatea pentru diferite macromolecule. Datorită sarcinii lor negative, pectinele sunt puternic hidratate şi înconjurate de cationi, în special de Ca2+. Substanţele pectice se găsesc din abundenţă în lamela mijlocie, fiind agenţi de cimentare intercelulară prin intermediul legăturilor încrucişate formate între ionii de calciu şi catenele pectice din structura a doi pereţi adiacenţi.
Aranjamentul principalelor componente în cadrul unui perete celular primar este prezentat în Figura 1.
Substanţele pectice sunt macromolecule complexe în structura cărora acidul D-galacturonic este componentul principal. Catena polizaharidică centrală are unităţile de -D-galacturonat legate (14) şi conţine 2-4% unităţi de L-ramnoză (aproximativ câte una la 25 unităţi galacturonice) legate -(14) şi -(12) la resturile de acid galacturonic. Catenele laterale sunt de două feluri: lungi, alcătuite din acid galacturonic (galacturonani) sau din resturi de L-arabinoză (arabinani) şi scurte, alcătuite din resturi de D-galactoză, D-xiloză sau L-fucoză. În structura substanţelor pectice au fost evidenţiate două tipuri de regiuni: o regiune “grea“, alcătuită din catene ramnogalacturonanice puternic substituite cu zaharuri neutre şi o regiune “uşoară“ în structura căreia există, în special, catene homogalacturonanice.
Multe din grupările carboxil din structura resturilor de acid galacturonic sunt esterificate cu metanol. Gradul de esterificare variază cu sursa biologică şi cu agentul utilizat pentru extracţia substanţelor pectice. Au fost izolate şi caracterizate şi substanţe pectice care au câteva din resturile de acid galacturonic acetilate.
164

D. Dinu
Figura 1. Aranjamentul principalilor constituenţi ai peretelui celular primar.
165

Enzime pectinolitice de origine microbiană
Gradul de esterificare, proporţia de zaharuri neutre şi gradul de policondensare sunt principalele elemente ale heterogenităţii substanţelor pectice de diferite origini (Dick şi alţii 1989, MacKinnan şi alţii 2002). Deesterificarea, proces care se poate realiza chimic sau enzimatic, determină asamblarea macromoleculelor pectice prin intermediul ionilor de Ca2+. Acest proces afectează semificativ proprietăţile fizico-chimice ale pectinelor, şi, implicit, arhitectura pereţilor celulari (Braccini şi alţii 2001, Barnavon şi alţii 2001, Ralet şi alţii 2001, Camara şi alţii 2002)
Clasificarea substanţelor pectice se face după mai multe criterii. După solubilitatea lor în diferite medii, substanţele pectice au fost împărţite în trei categorii: substanţe pectice solubile în apă (pectine înalt metilate sau acizi
pectinici); substanţe pectice solubile în soluţii care complexează ionii de calciu ca
de exemplu EDTA, oxalat de amoniu (pectine slab metilate sau acizi pectici);
substanţe pectice insolubile în apă (protopectinele), considerate o formă nativă a acestor biomolecule. Insolubilitatea protopectinei se datorează atât mărimii policondensatului, cât şi legăturilor ce o caracterizează. În structura protopectinei există reţele complexe de catene poligalacturonice asociate între ele prin legături ionice stabilite între un ion bivalent (în special calciu) şi două grupări carboxil. Protopectina se leagă, prin legături de hidrogen, de celelalte componente din pereţii celulari. Protopectina poate fi solubilizată cu acizi diluaţi, la cald.
O altă clasificare a substanţelor pectice are în vedere gradul de esterificare a grupărilor carboxil din structură. După acest criteriu, substanţele pectice au fost împărţite în: pectine înalt metilate (HMP) cu au un grad de metilare de peste 50% pectine slab metilate (LMP) cu au un grad de metilare de sub 50%.
Izolarea şi caracterizarea pectinelor din diferite ţesuturi vegetale a evidenţiat prezenţa în pereţii celulari a mai multor tipuri de substanţe pectice. În funcţie de structură, substanţele pectice au fost clasificate de ONeill (1990) în: homogalacturonani - catene de resturi de acid galacturonic legate -
(14) glicozidic, metilate sau nemetilate; ramnogalacturonani de tipul I (RG-I) - o familie de polizaharide înrudite,
la care catena de bază are o structură de forma: 4)--D-Galp A-(12)- -L-Rhap-(1. Aproximativ 50% din resturile de ramnoză din structură sunt substituite la C-4 cu oligozaharide neutre (galactoză, arabinoză şi puţină fucoză);
ramnogalacturonani de tipul II (RG-II) - în structura cărora unele resturi de acid galacturonic au ataşate la C-2 sau C-3 aldo şi ceto oligozaharide;
arabinani, galactani şi arabinogalactani, care alcătuiesc catenele laterale a polizaharidelor pectice.
Substanţele pectice se găsesc aproape în toate organele plantelor. Aceste polizaharide sunt componentele majore ale pereţilor primari ai
166

D. Dinu
dicotiledonatelor şi monocotiledonatelor. Pereţii celulari ai graminaceelor conţin cantităţi mai mici de substanţe pectice. Cantităţi mari de substanţe pectice se găsesc în pulpa fructelor cărnoase (mere, pere, ananas) precum şi în sfecla de zahăr.
Cantităţi importante de substanţe pectice au fost determinate în diferite deşeuri rezultate în industria alimentară (coji de citrice, borhot de sfeclă, tărâţe de grău, resturi de pulpă de fructe). Prezenţa substanţelor pectice în aceste deşeuri justifică utilizarea acestora ca inductori ai enzimelor pectinolitice microbiene.
2.Clasificarea şi nomenclatura enzimelor pectinolitice
În general, în categoria enzimelor pectinolitice sunt incluse enzimele care acţionează asupra homogalacturonanilor şi ramnogalacturonanilor.
Enzimele pectinolitice cuprind: o esterază, şapte poligalacturonaze (nume generic preluat după Whitaker, 1990) şi patru liaze. Dintre acestea numai o parte sunt incluse în Enzyme Nomenclature şi au număr de cod. Clasificarea şi nomenclatura enzimelor pectinolitice este prezentată în Tabelul 1.
Unii cercetători au inclus -D-arabinofuranozidaza şi endoarabinaza, enzime care acţionează asupra arabinanilor din structura substanţelor pectice, în categoria enzimelor pectinolitice. Majoritatea enzimologilor şi microbiologilor nu sunt familiarizaţi cu aceste două enzime, enzime care nu au fost incluse în nici un volum din seria Methods in Enzymology.
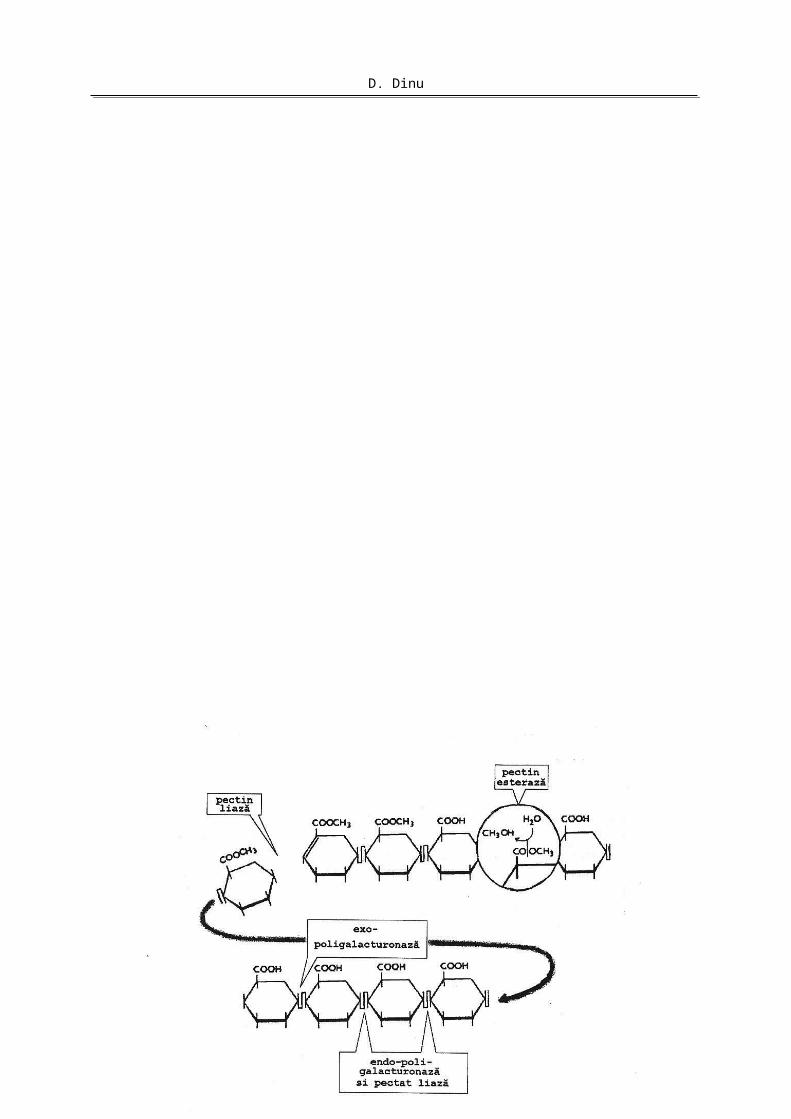
Modul de acţiune al principalelor enzime pectinolitice (endopoligalacturonaza, exopoligalacturonaza, endopectat liaza, exopectat liaza, pectin liaza şi pectinesteraza) este prezentat în Figura 2.
Cercetări recente au evidenţiat existenţa altor două enzime implicate în degradarea polimerilor pectici, ramnogalacturonaza şi -1,4-D- galactanaza. Unele tulpini de Aspergillus aculeatus biosintetizează o ramnogalacturonază, enzimă care scindează legăturile dintre resturile de acid galacturonic şi cele de ramnoză din structura ramnogalacturonanilor pectici (Beldman şi alţii 1996, Hennink şi alţii 1996). Yamaguchi (1995) a izolat din culturile de Aspergillus niger o endo--1,4-D- galactanaza, proteină cu masă moleculară de 32 000 Da, care hidrolizează polizaharidele pectice din soia cu eliminare de oligomeri galactozidici şi galactoză. O tulpină de Echerichia coli s-a dovedit a fi capabilă să biosintetizeze o pectin acetil esterază (Shevchik şi alţii 1997).
3. Surse de izolare ale enzimelor pectinoliticeEnzimele pectinolitice sunt larg răspândite la microorganisme şi la
plantele superioare. Ele joacă un rol important în procesele de creştere, în plantele
superioare permiţând elongaţia celulelor. Enzimele pectinolitice de origine 167

Enzime pectinolitice de origine microbiană
vegetală sunt implicate în procesele de înmuiere a ţesuturilor în timpul maturării şi stocării. Endopoligalacturonazele, exopoligalacturonazele, pectinesterazele apar, separat sau împreună, în diferite organe ale plantelor. Din surse vegetale nu au fost izolate proteine cu activitate pectat, sau pectin liazică.
Enzimele pectinolitice au fost separate şi caracterizate din: roşii, cireşe, piersici, portocale, grepfruit, papaia, mango, soia, in, măsline, polen.
Prezenţa enzimelor pectinolitice a fost semnalată şi la insecte. În larvele de Conotrachelus nenuphar a fost semnalată prezenţa pectinesterazei, endopoligalacturonazei, pectat liazei şi pectin liazei. Aceste enzime sunt eliberate atunci când larvele se hrănesc, fiind capabile să producă o macerare a ţesuturilor fructelor şi, împreună cu celulazele, sunt responsabile de căderea prematură a merelor şi prunelor infectate de aceste larve. Activitatea poligalacturonazică a fost decelată în saliva ploşniţei verde, Schizaphis graminum şi în cea a gărgăriţei orezului, Sitopilulus oryzae.
Un număr mare de agenţi patogeni ai plantelor, în special fungi şi bacterii, produc enzime pectinolitice. Acestea, împreună cu alte proteine, trec în mediul interior al gazdei şi joacă un rol important în procesele de colonizare şi infectare a plantelor. Majoritatea microorganismelor investigate produc cel puţin două tipuri de enzime pectinolitice cu acţiune sinergică care formează un sistem activ de degradare a polimerilor pectici. Majoritatea enzimelor pectinolitice de origine microbiană sunt inductibile, fiecare tip de enzimă având inductori specifici.
168

D. Dinu
169

Enzime pectinolitice de origine microbiană
Figura 2. Modelul unei molecule de pectină şi de pectat. Punctele de atac ale enzimelor pectinolitice.
Din clasa fungilor, speciile de Aspergillus sunt cele mai bune producătoare de enzime pectinolitice. Acestea biosintetizează în cantităţi mari endo şi exopoligalacturonaze. Poligalacturonazele de la Aspergillus sp. prezintă forme moleculare multiple cu un determinism genetic diferit (Statilova şi alţii 1993). Unele Aspergillus sp., ca de exemplu Aspergillus niger, cultivate pe medii corespunzătoare, produc şi pectinesteraze, pectat sau pectin liaze (Dinu şi alţii 1997, Dinu şi alţii 1998).
Alte specii de fungi producătoare de enzime pectinolitice sunt: Botrytis cinerea (ten Have şi alţii 2001, Rho şi alţii 2001), Fusarium sp. (Garcia-Marceira şi alţii 2001, Niture şi alţii 2001), Penicillium sp. (Chelegatti şi alţii 2000), Verticillium sp. (James şi alţii 2001).
Diferite specii de bacterii, (Cellovibrio, Clostridium, Erwinia, Lactobacillus, Pseudomonas), produc una sau mai multe tipuri de enzime pectinolitice. Echipamentul pectinolitic al acestora este diferit de cel al fungiilor. Bacteriile şi fungii secretă poligalacturonaze şi pectinesteraze. Diferenţa între cele două clase constâ în tipul de enzime care acţionează asupra polizaharidelor pectice prin mecanisme de transeliminare: pectin liazele sunt enzime caracteristice fungilor, iar pectat liazele apar cu precădere la bacterii.
Enterobacteria Erwinia chrysanthemi realizează degradarea substanţelor pectice din pereţii celulari ai plantelor invadate prin intermediul mai multor tipuri de enzime: pectin metilesteraza, pectin acetilesteraza, pectat liaza şi pectin liaza. Erwinia chrysanthemi se caracterizează prin abilitatea de a secreta mai multe izoenzime ale endopectat liazei, izoenzime care au un rol major în procesele de putrezire a plantelor infestate cu această bacterie. De la o tulpină de Erwinia chrysanthemi au fost caracterizate opt endopectat liaze, respectiv PelA, PelB, PelC, PelD, PelE, PelI, PelL şi PelZ şi o exopectat liază, PelX. Primele cinci forme aparţin familiei 1 de pectat liaze, familie care include pectat liaze bacteriene şi pectin liaze fungice, enzime cu structură primară asemănătoare. PelI aparţine familiei 2, familie în care sunt încadrate liaze de la Erwinia carotovora. PelL şi PelX fac parte din familia 4, iar PelZ din familia 5.
Diversificarea biotehnologiilor care utilizează enzime pectinolitice şi nevoia tot mai mare de preparate enzimatice cu activitate pectinolitică complexă, au determinat găsirea de noi surse microbiene care să producă astfel de enzime. Această necesitate a determinat extinderea cercetărilor şi la drojdii. Cercetări recente au arătat că unele specii de drojdii, cultivate pe medii care conţin substanţe pectice, sunt producătoare de enzime pectinolitice. Drojdia metilotrofă Candida boidinii biosintetizează poligalacturonază, pectinesterază, pectin şi pectat liază (Nakagawa şi alţii 2000), Kluyveromyces marxianus numai poligalacturonază (Jia şi alţii 2000), iar unele specii de Saccharomyces - poligalacturonază, pectinesterază şi pectin liază (Gomes şi alţii 2001).
Majoritatea enzimelor pectinolitice produse de microorganisme sunt inductibile. Fiecare tip de enzimă are inductori specifici. Penicillium
170
D. Dinu
frequentans, crescut pe medii în care sursa de carbon a fost pectina, poligalacturonatul de sodiu sau acidul galacturonic, a biosintetizat 11 poligalacturonaze şi 3 pectinesteraze. Acelaşi microorganism cultivat pe mediu de glucoză a produs 2 poligalacturonaze şi o pectinesterază (Chellegatti şi alţii 2000).
Tehnica DNA recombinat a fost utilizată pentru a obţine tulpini înalt producătoare de enzime pectinolitice. Khahn (1991) a clonat gena care codifică pectinesteraza, a expresat-o la o tulpină sălbatică de Aspergillus niger, ceea ce a condus la o creştere de douăzeci de ori a activităţii esterazice. Genele care codifică endopoligalacturonaze fungice au fost clonate, secvenţializate şi expresate la drojdii de tipul Saccharomzces cerevisiae şi Kluyveromzces marxianus (Jia şi alţii 2000, Vilanova şi alţii 2000).
4.Enzimele pectinolitice de origine microbiană: specificitate, mecanism de acţiune, structură,
proprietăţi4.1. Pectinesterazele
Pectinesterazele (EC 3.1.1.11) hidrolizează pectinele îndepărtând grupări metoxi de la grupările 6-carboxilice ale resturilor de acid galacturonic.
Aceste enzime prezintă specificitate de grup, recunoscând din structura substratului partea D-galacturonică şi legătura esterică. Elucidarea specificităăţii manifestate de pectinesteraze s-a realizat prin studii pe pectine modificate chimic. Studiile întreprinse cu pectine în care o parte din grupările carboxil sunt reduse sau pe metilesterii acidului alginic au arătat imposibilitatea pectinesterazelor de a hidroliza astfel de substraturi. Aceste experimente au evidenţiat înalta specificitate a acestor enzime faţă de structuri poli-D-galacturonanice. Specificitatea faţă de partea alcoolică este mai redusă, pectinesterazele fiind capabile să hidrolizeze cu viteze mai mici, etil, propil şi alil esterii acidului pectinic. Viteza de hidroliză a substratului este influenţată şi de gradul de policondensare al acestuia, diminuându-se odată cu scăderea acestuia (Rexová-Benková şi alţii 1976).
Mecanismele prin care acţionează pectinesterazele au fost elucidate pe baza experimentelor întreprinse de Miller şi colaboratorii (1971). Studiile în acest sens au fost realizate pe pectină cu grad de policondensare 33 şi grad de esterificare 96%. Substratul a fost supus iniţial acţiunii unei exopectat liaze purificată din Clostridium multifermentans, enzimă care acţionează secvenţial de la capătul reducător. Acţiunea liazei asupra substratelor înalt esterificate a fost neglijabilă, dar a crescut rapid la adiţia unei pectinesteraze purificate din Fusarim oxysporum. În urma acestor experimente, s-a tras concluzia că, majoritatea acţiunii esterazei decurge la capătul reducător al moleculei de pectină. Monitorizarea simultană şi continuă a celor două tipuri de activităţi pectinolitice a indicat că 57% din activitatea pectinesterazei din
171

Enzime pectinolitice de origine microbiană
Fusarim oxysporum decurge de la capătul reducător al substratului. Experimente similare întreprinse cu o proteină multienzimatică purificată din Clostridium multifermentans, care posedă activitate esterazică şi exopectat liazică, au arătat că, în cazul acestui microorganism, deesterificarea pectinei înalt metilate decurge exclusiv de la capătul reducător.
Modul de acţiune şi specificitatea pectinesterazei din Aspergillus niger au fost studiate de Kester şi colaboratorii (2000) utilizând oligogalacturonaţi total metilaţi cu grade de condensare cuprinse între 2 şi 6 (Kester şi colaboratorii 2000). Natura produşulor de reacţie a fost determinată prin cromatografie de înaltă performanţă (HPLC) pe anioniţi, iar localizarea grupărilor esterice, marcate în prealabil cu 18O , în structura produşilor de reacţie a fost posibilă prin spectrometrie de masă. Aceste studii au arătat că pectinesteraza din Aspergillus niger este capabilă să hidrolizeze şi digalacturonaţi total metilaţi. Enzima hidrolizează cu viteză mai mare legăturilre esterice localizate pe resturile de acid galacturonic din interiorul substratului. După hidroliza acestor legături sunt eliberate grupările metilice de la resturile de acid galacturonic de la capetele reducătoare ale oligogalacturonaţilor, în timp ce grupările de la capătul nereducător nu sunt hidrolizate.
Unele dintre pectinesterazele microbiene au structură glicoproteică. Rijssel (1993) a izolat un complex pectinolitic din Clostridium thermosaccharolyticum care posedă activitate poligalacturonazică şi pectinesterazică. Proteina complexă s-a dovedit a fi o glicoproteină cu resturi de N-acetil galactozamină şi galactoză.. Pectinesteraza purificată de Shevchik şi colaboratorii (1996a) din Erwinia chrysanthemi s-a dovedit a fi o lipoproteină din membrana externă, cu o secvenţă palmitat la extremitatea N-terminală.
Experimentele de mutageneză dirijată realizate de Duwe (1996) pe pectinesteraza din Aspergillus niger au vizat două resturi de histidină din structura enzimei. Substituirea restului de histidină din poziţia 137 cu un rest de alanină a avut ca efect pierderea totală a activităţii, în timp ce aceeaşi substituţie la histidina din poziţia 188 nu a avut nici un efect. Pe baza acestor rezultate s-a tras concluzia că restul de histidină din poziţia 137 este esenţială în activitatea catalitică, făcând probabil parte din centrul catalitic activ.
Markovic (1996) a evidenţiat prezenţa a şase resturi de histidină în structura pectinesterazei din Aspergillus niger. Tratamentul cu dietilpirocarbonat a arătat o inactivare a enzimei proporţională cu cantitatea de inhibitor. Aceste rezultate au sugerat că histidina nu face parte din centrul catalitic al acestei enzime, dar este implicată în stabilizarea structurilor de ordin superior.
Proprietăţile pectinesterazelor de origine microbiană sunt prezentate în Tabelul 2. Cu excepţia enzimei din Clostridium multifermentans,, pectinesterazele de origine microbiană au mase moleculare cuprinse între 27 000 şi 48 000 Da. Punctul izoelectric variază între 3,6 şi 7,4, iar pH-urile optime între 4,2 şi 9,0. Constantele Michaelis variază în limite largi, valorile acestora fiind influenţate de modul de exprimare a concentraţiei substratului.
172

D. Dinu
Acidul poligalacturonic este un inhibitor al pectinesterazelor. Datorită acestui fapt, cationii monovalenţi (în special Na+ şi K+) în concentraţii cuprinse între 50 şi 100mM, şi cationii divalenţi (în special Ca2+) în concentraţii între 5 şi 10mM, provoacă o creştere a activităţii specifice. Schejter (1988) a arătat că poligalacturonatul de sodiu este un inhibitor competitiv pentru cele două pectinesteraze din Botridis cinerea, constantele de inhibiţie având valori de 2,8 şi, respectiv, 0,11 mg/ml.
Manjou (1992) a determinat pentru o pectinesterază microbiană o constantă de inhibiţie a acidului poligalacturonic de 0,44 mg/ml. Studiile sale au arătat că această inhibiţie dispare atunci când esteraza a fost coimobilizată pe suporturi de sticlă poroasă cu o endopoligalacturonază. Un alt inhibitor competitiv al acestor enzime s-a dovedit a fi şi metanolul (Maldonado şi alţii 1994).
173

Enzime pectinolitice de origine microbiană
Pectinesteraza II, izolată de Lim (1983) din culturi de Aspergillus oryzae a fost total inhibată de HgCl2. Alţi inhibitori ai pectinesterazelor microbiene s-au dovedit a fi: CuCl2, AlCl3 (Lim şi alţii 1983, Sakelaris şi alţii 1989a), iodul (Markovic şi alţii 1989), reactivi pentru grupări SH de tipul N-etilmaleiimida, p-cloromercuribenzoatul de sodiu, iodacetamida (Lim şi alţii 1983, Cardello şi alţii 1995).
4.2. Protopectinazele
Degradarea protopectinei a fost atribuită iniţial acţiunii sinergice a pectinesterazei şi endopoligalacturonazei şi/sau pectin liazei. Studiile întreprinse de Sakai şi colaboratorii săi au pus în evidenţă capacitatea unor microorganisme de a produce o enzimă care solubilizează protopectina. Această nouă enzimă, denumită protopectinază, degradează protopectina cu eliberare de polimeri pectici solubili, fără a produce macerarea ţesuturilor plantelor. Cercetările întreprinse de Sakai şi grupul său asupra acestei enzime, au fost sumarizate şi publicate în volumul 161 din seria Methods in Enzymology (Sakai şi alţii 1988).
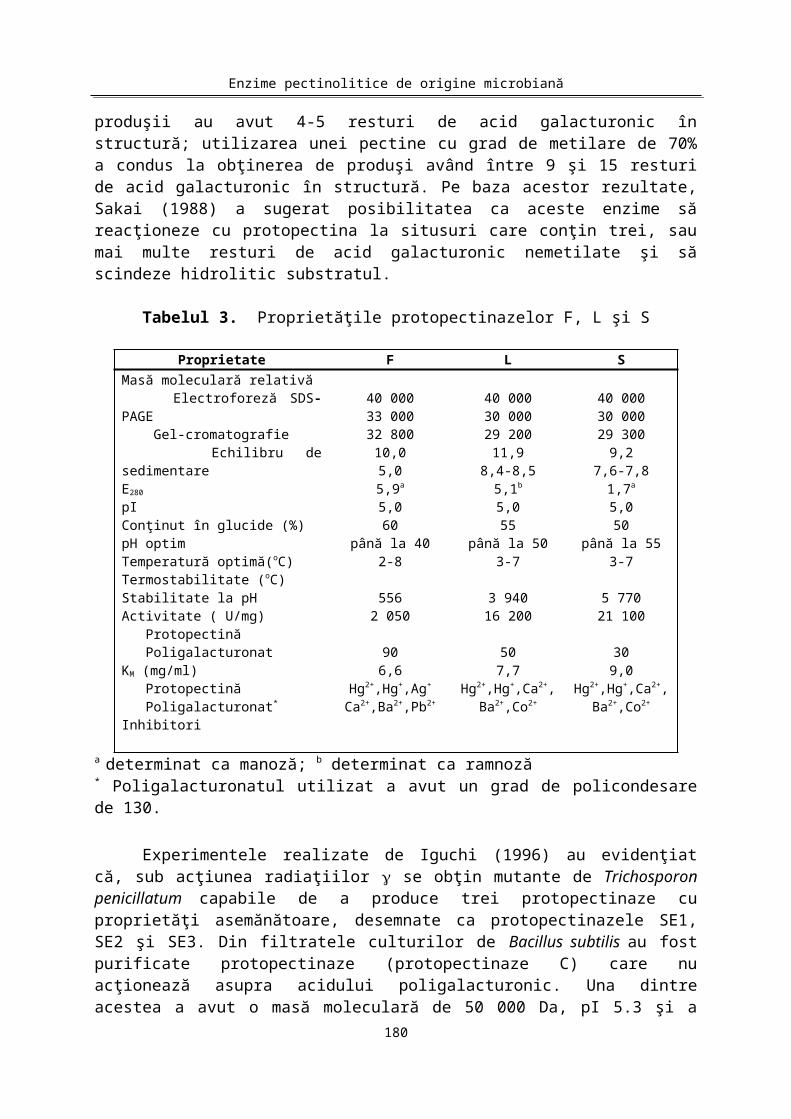
Protopectinaza a fost purificată şi cristalizată de Sakai (1988) din Kluyveromyces fragilis (protopectinaza F), Galactomyces reessii (proto-pectinaza L) şi din Trichosporon penicillatum (protopectinaza S). Proprietăţile fizice- chimice şi cinetice ale acestor enzime sunt prezentate în Tabelul 3.
Masele moleculare ale celor trei protopectinaze, estimate prin aceeaşi tehnică, au avut valori foarte apropiate. Coeficienţii de extincţie la 280nm (E280) au valori apropiate, dar nu identice, datorită diferenţelor existente între cele trei protopectinaze la nivelul compoziţiei în aminoacizi aromatici. Valorile pI indică că protopectinaza F este o proteină acidă, iar protopectinazele L şi S sunt proteine bazice. Toate trei protopectinazele sunt glicoproteine, dar cantitatea de carbohidraţi este diferită.
Compoziţia în aminoacizi, diferită pentru cele trei protopectinaze, are o caracteristică comună: lipsa metioninei (protopectinaza L) sau
174

D. Dinu
prezenţa ei în cantitate foarte mică (protopectinazele F şi S au un singur rest de metionină în structură). Protopectinazele S şi L au la capătul N-terminal un rest de glicină şi o secvenţă identică de 27 resturi de aminoacizi.
Datele prezentate în Tabelul 3 arată asemănări la nivelul unora dintre proprietăţi, mai ales între protopectinazele L şi S, enzime care s-au dovedit a fi identice din punct de vedere imunologic. Pentru toate cele trei protopectinaze poligalacturonatul de sodiu s-a dovedit a fi un substrat mai bun decât protopectina de diferite provenienţe.
Utilizând ca substrate tri-, tetra- şi pentagalacturonaţi, Sakai (1988) a determinat modul de acţiune şi constantele cinetice pentru cele trei protopectinaze (Figura 3). Rezultatele obţinute au evidenţiat o acţiune similară a protopectinazelor F, L şi S asupra substratelor cu 3 şi 4 resturi de acid galacturonic. Hidroliza trigalacturonatului decurge prin îndepărtarea unui rest de acid galacturonic de la capătul reducător, iar degradarea tetragalacturonatului decurge prin eliminarea unei molecule de acid galacturonic sau de digalacturonat de la capătul reducător (Figura 3).
Modul de acţiune asupra pentagalacturonatului a fost diferit. Astfel, protopectinazele F şi S îndepărtează o moleculă de acid galacturonic sau una de digalacturonat de la capătul reducător al moleculei pentagalacturonice, în timp ce protopectinaza L îndepărtează doar un rest de acid galacturonic din structura acestui oligogalacturonat. Pe baza raportului Vmax / KM , numit coeficient specific, Sakai (1988) a demonstrat că specificitatea celor trei enzime pentru substrate creşte în ordinea tri-, tetra- şi penta-galacturonaţi. În ceea ce priveşte acţiunea asupra poligalacturonaţilor, modul de scindare depinde şi de gradul de policondensare. Când cele trei protopectinaze au acţionat asupra acidului poligalacturonic, produşii de reacţie obţinuţi au avut 2-3 resturi de acid galacturonic în structură. Când substratul a fost o pectină cu grad de metilare de 30%, produşii au avut 4-5 resturi de acid galacturonic în structură; utilizarea unei pectine cu grad de metilare de 70% a condus la obţinerea de produşi având între 9 şi 15 resturi de acid galacturonic în structură. Pe baza acestor rezultate, Sakai (1988) a sugerat posibilitatea ca aceste enzime să reacţioneze cu protopectina la situsuri care conţin trei, sau mai multe resturi de acid galacturonic nemetilate şi să scindeze hidrolitic substratul.
Tabelul 3. Proprietăţile protopectinazelor F, L şi S
Proprietate F L S
175

Enzime pectinolitice de origine microbiană
Masă moleculară relativă Electroforeză SDS-PAGE Gel-cromatografie Echilibru de sedimentareE280pIConţinut în glucide (%)pH optimTemperatură optimă(oC)Termostabilitate (oC)Stabilitate la pHActivitate ( U/mg) Protopectină PoligalacturonatKM (mg/ml) Protopectină Poligalacturonat*
Inhibitori
40 00033 00032 800
10,05,05,9a
5,060
până la 402-8
5562 050
906,6
Hg2+,Hg+,Ag+
Ca2+,Ba2+,Pb2+
40 00030 00029 200
11,98,4-8,5
5,1b
5,055
până la 503-7
3 94016 200
507,7
Hg2+,Hg+,Ca2+,Ba2+,Co2+
40 00030 00029 300
9,27,6-7,8
1,7a
5,050
până la 553-7
5 77021 100
309,0
Hg2+,Hg+,Ca2+,Ba2+,Co2+
a determinat ca manoză; b determinat ca ramnoză * Poligalacturonatul utilizat a avut un grad de policondesare de 130.
Experimentele realizate de Iguchi (1996) au evidenţiat că, sub acţiunea radiaţiilor se obţin mutante de Trichosporon penicillatum capabile de a produce trei protopectinaze cu proprietăţi asemănătoare, desemnate ca protopectinazele SE1, SE2 şi SE3. Din filtratele culturilor de Bacillus subtilis au fost purificate protopectinaze (protopectinaze C) care nu acţionează asupra acidului poligalacturonic. Una dintre acestea a avut o masă moleculară de 50 000 Da, pI 5.3 şi a prezentat termostabilitate până la 70OC. Această enzimă a manifestat activitate protopectinazică pe fibre de bumbac şi pe protopectina din lămâie, dar şi activitate pectat liazică (Takao şi alţii 2000). Din culturile de Bacillus subtilis au fost izolate alte două protopectinaze, una cu activitate ramnogalacturonazică (Sakai şi alţii 1993), iar cealaltă cu activitate arabinazică (Sakamoto şi alţii 1995).
Protopectinazele descrise în literatură manifestă specificitate diferită faţă de protopectinele de diferite provenienţe. Astfel, protopectinaza din Trichosporon penicillatum acţionează cu viteze diferite asupra protopectinelor din portocale, grapefruit, cartofi, morcovi, pere, piersici (Sakamoto, 1995), şi o degradează cel mai bine pe cea din cartofi (Nakamura, 1995). Protopectinaza din Bacillus subtilis degradează cel mai rapid protopectinele din morcovi (Nakamura, 1995).
Existenţa unor enzime capabile să degradeze protopectina, forma insolubilă a polizaharidelor pectice, este contestată de multe cercetări din domeniu. Majoritatatea cercetărilor consideră că degradarea protopectinei se datorează acţiunii sinergice a poligalacturonazei, pectinesterazei şi pectat sau pectin liazei. Această ipoteză se bazează pe faptul că toate enzimele capabile să acţioneze asupra protopectinei izolate şi caracterizate până în prezent posedă şi o a doua funcţie catalitică. Această a doua funcţie poate fi, de la caz la caz, poligalacturonazică, pectat liazică, ramnogalacturonazică sau arabinazică.
176

D. Dinu
4.3. Endopoligalacturonazele177

Enzime pectinolitice de origine microbiană
Endopoligalacturonazele (EC 3.2.1.15) sunt enzime care hidrolizează legăturile glicozidice din structura poligalacturonaţilor într-o manieră mai mult sau mai puţin întâmplătoare, formând oligogalacturonaţi. Acţiunea acestor enzime determină scăderea pronunţată a vâscozităţii soluţiei substratului.
Endopoligalacturonazele acţionează preferenţial asupra acizilor pectici şi a pectinelor slab esterificate. Studiile întreprinse asupra endopoligalacturonazei purificate din Corticium rolfsii au arătat că această enzimă scindează, în ordine, acidul poligalacturonic, pectina 67% esterificată şi pectina 91% esterificată, cu viteze de 100%, 12% şi, respectiv, 1,5%. Utilizarea acizilor pectinici în calitate de substrate a acestor enzime a condus la obţinerea de rezultate neconcludente, în structura acestora existând porţiuni de resturi de acid galacturonic neesterificate, porţiuni suficient de mari ca să permită acţiunea endopoligalacturonazei. Studiile întreprinse de Chen şi colab. (1996) asupra produşilor de reacţie obţinuţi sub acţiunea endopoligalacturonazei din Erwinia carotovora au evidenţiat capacitatea acestei enzime de a hidroliza pectinele cu grade de metilare de 16%, 32% şi 52%, care au în structură patru resturi adiacente de acid galacturonic.
Mărimea moleculei substratului este un factor care influenţează activitatea acestor enzime. Ele acţionează cu viteză maximă asupra substratelor cu număr mare de resturi de acid galacturonic, cu viteze reduse asupra oligogalacturonaţilor şi nu scindează digalacturonatul.
Determinarea produşilor de reacţie formaţi în fazele incipiente ale reacţiei a permis diferenţierea între endo şi exo enzime: endopoligalacturonazele produc oligozaharide diferite şi cantităţi mici de mono-şi digalacturonaţi, în timp ce exopoligalacturonazele produc mono şi digalacturonaţi în cantitate mare. Lim (1980) a arătat că endopoligalacturonaza din Rhizopus arrhizus nu produce oligomeri în primele 40 minute de reacţie, iar molecule monomere de acid galacturonic apar abia dupădouăore.
Elucidarea mecanismului prin care acţionează aceste enzime s-a realizat în urma analizei produşilor de reacţie obţinuţi pe substrate polimere şi oligomere. Rezultatele obţinute au arătat că natura produşilor depinde de mărimea regiunii de contact dintre substrat şi situsul activ, mărime care afectează constantele cinetice de reacţie, KM şi Vmax.. Astfel, sub acţiunea endopoligalacturonazelor, pentru care mărimea substratului este relativ neesenţială în legare, se formează cantităţi mici de produşi intermediari, iar mecanismul după care acţionează aceste enzime a fost denumit impropriu “single chain attack“. În cazul endopoligalacturonazelor, care se leagă mai puternic de poligalacturonaţii cu mase mari decât de cei cu mase mici, concentraţia produşilor intermediari este ridicată, mecanismul de acţiune fiind denumit “multi-chain attack“. Studiind cinetica reacţiei de hidroliză a substratelor sub acţiunea endo-poligalacturonazei purificate din Saccharomyces fragilis, Demain (1954) a observat trei etape de reacţie. În prima etapă, etapă care decurge rapid, sunt scindate aproximativ 25% din legăturile glicozidice din structura substratului cu eliberarea tetra-, tri- şi digalacturonaţi. În etapa a
178

D. Dinu
doua, etapă mult mai lentă, gradul de hidroliză creşte la 50%, iar tetragalacturonaţii sunt hidrolizaţi la tri- şi monogalacturonaţi. În ultima etapă, etapă foarte lentă, trigalacturonaţii sunt scindaţi la digalacturonaţi şi acid galacturonic.
Pentru a explica diversitatea produşilor rezultaţi sub acţiunea poligalacturonazelor de provenienţe diferite, Rexová-Benková (1976) a propus trei modele care să explice modul de acţiune al poligalacturonazelor. Cele trei modele, numite A, B şi C, sunt prezentate în Figura 4. Endopoligalacturonazele din Aspergillus niger şi cele din Trichoderma reessi urmează modelul A, caracterizat prin scindarea tetramerilor la trimeri şi acid galacturonic, şi prin scindarea extrem de lentă a trimerilor. Modelul B, caracterizat printr-o scindare alternativă a tetramerilor (la trimeri şi acid galacturonic sau la două molecule de digalacturonat) şi prin degradarea rapidă a trimerilor, a fost evidenţiat la endopoligalacturonazele din roşii. Modelul C, pus în evidenţă pentru endo-enzima de la Erwinia carotovora, se deosebeşte de celelalte mecanisme printr-o scindare specifică a pentamerilor şi a hexamerilor.
Având în vedere faptul că modul de acţiune şi specificitatea de substrat a enzimelor depinde de natura situsului activ, Rexová-Benková (1976) a considerat că endopoligalacturonazele care acţionează după unul din cele trei modele au centre catalitice diferite, alcătuite dintr-un număr diferit de subsitusuri.
Studii recente (Bonnin şi alţii 2001) au evidenţiat atacul multiplu asupra substratelor şi în cazul endopoligalacturonazei din Fusarium moniliforme. Afinitatea şi viteza maximă de reacţie cresc cu creşterea lungimii catenei substratului. Situsul catalitic al enzimei este alcătuit din cinci subsitusuri, iar produşii finali de hidroliză ai homogalacturonanilor sunt monomeri şi dimeri ai acidului galacturonic.
Stereochimia acţiunii hidrolitice a două endopoligalacturonaze din Aspergillus niger, a fost investigată de Biely (1996a şi b) cu ajutorul spectroscopiei 1H-RMN. În mediu de D2O, toţi produşii de reacţie au avut noul capăt reducător în conformaţie , fapt care arată acţiunea invertivă a acestor enzime.
Model Substrat Produşi de reacţieA O-O-O O-O-O-O O-O-O-O-O O-O-O-O-O-O
O-O + OO-O-O + OO-O-O-O + OO-O-O + O-OO-O-O-O-O + OO-O-O-O + O-O O-O-O + O-O-O
B O-O-O
O-O + OO-O-O + O
179
Enzime pectinolitice de origine microbiană
O-O-O-O O-O-O-O-O
O-O + O-OO-O-O-O + OO-O-O + O-OO-O + O-O-O
C O-O-O O-O-O-O O-O-O-O-O O-O-O-O-O-O
O-O + OO + O-OO-O-O + OO-O + O-OO-O-O + O-OO-O-O + O-O-OO-O-O-O + O-O
Figura 4. Posibilităţi de acţiune a endopoligalacturonazelor pe substraturi mici
(O reprezintă o moleculă de acid galacturonic).
Pe baza studiilor de variaţie a constantelor cinetice cu pH-ul, Rexová-Benková (1976) a sugerat implicarea în mecanismul de acţiune al endopoligalacturonazei din Aspergillus niger a unei grupări carboxil şi a unei grupări imidazol protonate a histidinei. Cercetările intreprinse de Statilova şi colab. (1996b) au evidenţiat prezenţa în structura situsului activ al endopoligalacturonazei de la Aspergillus niger a unui rest de tirozină, rest ionizat la pH 10,5. Studiile de modificare chimică a unor resturi de aminoacizi din structura endopoligalacturonazei din Aspergillus ustus au indicat participarea unui rest de triptofan în legarea substratului şi a unui rest de histidină în actul catalitic (Rao şi alţii 1996 a şi b, Niture şi alţii 2001). Pe baza acestor studii, Rao şi colab. (1996b) au presupus că în mecanismul de acţiune al acestei enzime are loc transferul unui singur proton de la restul de histidină la O glicozidic din structura substratului, transfer urmat de hidroliză prin adiţia unei molecule de apă.
Studiile structurale realizate de Statilová (1998b) au pus în evidenţă caracterul glicoproteic al formelor multiple ale poligalacturonazei din Aspergillus niger. În structura celor două forme majore ale poligalacturonazei sunt prezente resturi de N-acetilgalactozamină şi manoză. Şi endopoligalacturonaza din Aspergillus aculeatus s-a dovedit a fi o glicoproteină cu o structură secundară în care motivul predominant este -structura paralelă (Cho şi alţii 2001).
Endopoligalacturonazele au fost purificate şi caracterizate din mai multe surse microbiene. Proprietăţile câtorva endopoligalacturonaze sunt prezentate în Tabelul 4. Aceste enzime au mase moleculare cuprinse între 30 000 şi 85 000 Da, pI variind în domeniul 3,7-8,3. Toate endopoligalacturonazele au pH optim acid, cuprins între 3,8 şi 5,5. Comparativ cu alte enzime pectinolitice, valorile KM pentru poligalacturonat variază pe un domeniu destul de îngust, cuprins între 0,14 şi 4,7 mg/ml.
Efecte inhibitoare asupra endopoligalacturonazelor au următoarele substanţe:
180

D. Dinu
HgCl2, cu o valoare a constantei de inhibiţie de 6,8x10-5M (Blanco şi alţii 1994);
acidul tanic (Liu 1978 , Elegado 1994); pectina, în concentraţii de ordinul 10-3 g/100ml, determină o inhibiţie
competitivă (Schejter 1988); NaCl în concentraţie de peste 1o/oo inhibă endopoligalacturonazele din
bacteriile marine (Mountfort 1995); epicatechina, în concentraţii cuprinse între 20-80 g/ml, inhibă cu 6,5%
şi respectiv 43% cele două endopoligalacturonaze din Colletotrichum gloeosporioides (posibil ca epicatechina să regleze activitatea endopoli-galacturonazelor fungice în fructele infestate) (Pruski 1989);
CaCl2 şi BaCl2, în concentraţii de 5x10-3M, au efect inhibitor datorat precipitării parţiale a acidului poligalacturonic.
4.4. Exopoligalacturonazele
Exopoligalacturonazele (EC 3.2.1.67) hidrolizează poligalacturonaţii cu eliberarea acidului galacturonic, producând o scădere mai redusă a vâscozităţii soluţiei substratului comparativ cu endopoligalacturonazele. Acţiunea exopoligalacturonazelor de origine vegetală şi microbiană începe de la capătul nereducător al moleculei. Modul de acţiune al acestor enzime a fost determinat pe baza observaţiilor experimentale care au arătat că, aceste enzime nu atacă substratele care au la extremitatea nereducătoare un rest de acid galacturonic 4:5 nesaturat sau un deoxizahar.
În literatura de specialitate au fost descrise şi exopoligalacturonaze care atacă substratul la capătul nereducător cu eliberarea digalacturonatului. Astfel de enzime au fost izolate de la Erwinia aroideae şi de la unele specii de Pseudomonas şi au denumirea corectăde poli-(1,4--D-galacturonid) digalacturonohidrolaza.
Exopoligalacturonazele nu degradeazăcomplet poligalacturonaţii, activitatea lor depinzând de gradul de policondensare al acestora şi de provenienţa lor. Astfel, exopoligalacturonaza din Aspergillus tubingensis (Kester şi alţii 1996 a şi b) a fost capabilă să hidrolizeze şi oligogalacturonaţi cu grade de condensare cuprinse între 2 şi 7, dar activitatea cea mai mare s-a înregistrat pe poligalacturonaţi cu mase moleculare mari.
181

Enzime pectinolitice de origine microbiană
182

D. Dinu
Studiile întreprinse de Biely (1996a) au arătat că exopoligalacturonaza din Aspergillus tubingensis, ca şi endopoligalacturonazele din Aspergillus niger, determină modificarea configuraţiei la nivelul grupării reducătoare a produsului de reacţie.
Proprietăţile fizico-chimice au fost stabilite pentru un număr mic de exopoligalacturonaze microbiene.
Hara (1984) a izolat două exopoligalacturonaze din culturile de Aspergillus niger. Exopoligalacturonaza I a avut masa molecularăde 66 000 Da, pH optim 3,8, temperatura optimă 600C, pI 5,6, KM pentru acid pectic de 20 mg/ml şi de 6,5 mg/ml pentru acid digalacturonic. Exopoligalacturonaza II a avut masa molecularăde 63 000 Da, pH optim
183

Enzime pectinolitice de origine microbiană
4,5, temperatura optimă 600C, pI 5,8, KM pentru acid pectic 3,85 mg/ml şi de 5,0 mg/ml pentru acid digalacturonic. Concentraţii de HgCl2, cuprinse între 0,002M şi 0,2M, au activat ambele enzime, dar mai ales forma moleculară I.
Prezenţa a două exopoligalacturonaze în culturile de Aspergillus niger a fost evidenţiatăşi de Behere (1993). Ambele forme ale exo-enzimelor au avut pH optim de 5,0, iar valorile KM raportate la poligalacturonatul de sodiu au fost de 0,24% şi 0,12%. Exopoligalacturonaza purificatăde Heinrichova (1976) din culturi de Aspergillus niger a avut un pH optim de 5,2.
Mikhailova (1995) a caracterizat două exopoligalacturonaze din Aspergillus alliaceus. Exopoligalacturonaza I a avut pH optim 3,5 şi temperatura optimă 45-500C, iar pentru forma moleculară II aceşti parametri au fost 6,8 şi 350C.
Cele două exopoligalacturonaze izolate de Devi (1996) de la Aspergillus carbonarius au avut masele moleculare de 61 000 Da şi, respectiv 47 000 Da, acţionând optim la 430C, respectiv 540C.
O altă specie producătoare de exopoligalacturonază s-a dovedit a fi Aspergillus tubingensis (Kester 1996 a şi b). Enzima din această sursă a avut masa de 70 000 Da, pI cuprins între 3,7-4,4, pH optim de 4,2 şi KM faţă de acidul poligalacturonic de 3,2 mg/ml. Enzima a fost inhibată competitiv de acidul galacturonic (Ki= 0,3mM) şi de acidul digalacturonic (Ki= 0,4mM) O tulpină de Fusarium oxysporum s-a dovedit a fi capabilă să producă patru exopoligalacturonaze cu valori pI de 9,3, 7,35, 6,85 şi 6,55. Toate cele patru forme ale enzimei au fost inhibate de ionii de calciu (Guevara, 1997).
O exopoligalacturonază a fost purificată până la omogenitate din mediul de cultură al unei tulpini de Bacillus (Kobajashi şi alţii 2001). Enzima a avut o masă moleculară de 45 000 Da, pI 5,8 şi o viteză maximă de acţiune asupra poligalacturonatului de sodiu la 600C şi pH 7,0. Spre deosebire de alte exopoligalacturonaze bacteriene, această exopoligalacturonază eliberează doar acid monogalacturonic atunci când acţionează asupra acidului poligalacturonic, di-, tri,-tetra- şi penta –galacturonaţilor.
Cercetări recente (Antov şi alţii 2001) au studiat cultivarea microorganismului Polyporus squamosus într-un sistem apos bifazic, polietilen glicol/dextran, în prezenţa melasei de sfeclă de zahăr ca sursă de pectină. În supernatantul mediului de cultură activitatea exopoligalacturonazică s-a dovedit a fi mai mare decât cea decelată în urma cultivării în sistem monofazic.
Datele prezentate arată necesitatea unor studii mai detaliate asupra aceastei categorii de enzime pectinolitice. Singura proprietate care diferenţiază clar exopoligalacturonazele de oligogalacturonat hidrolaze (enzime prezentate în paragraful 4.5) este preferinţa acestora pentru substrate cu mase moleculare mari. 4.5. Oligogalacturonat hidrolazele şi oligogalacturonat liazele
Oligogalacturonat hidrolazele şi oligogalacturonat liazele sunt enzime pectinolitice cu aplicaţii practice reduse şi destul de puţin studiate.
184

D. Dinu
Oligogalacturonat hidrolazele scindează legăturile glicozidice de la capătul nereducător al substratului. Aceste enzime diferă de exopoligalacturonaze prin faptul că manifestă activitate specifică mult mai mare faţă de oligogalacturonaţi comparativ cu cea faţă de poligalacturonaţi şi pectaţi. Oligogalacturonat liazele (EC 4.2.2.6) acţionează asupra oligogalacturonaţilor saturaţi şi nesaturaţi prin trans eliminare, îndepărtând monomeri nesaturaţi de la capătul reducător al acestora. Pentru ambele tipuri de enzime, viteza de reacţie descreşte rapid cu creşterea gradului de policondensare, fiind de până la 400 ori mai mică pe substrate macromoleculare.
În Tabelul 5 sunt prezentate tipurile de oligogalacturonaze pectice şi specificitatea manifestată de acestea faţă de diferite substraturi.
Cercetări recente (Heinrichowa, 1992) au evidenţiat biosinteza unei D-galacturonan digalacturonohidrolaze de către Selenomonas ruminantium enzimă capabilă să degradeze oligogalacturonaţi cu grade de condensare cuprinse între 3 şi 5.
Heinrichowa şi colaboratorii (1994) au evidenţiat că Clostridium lochheadii produce un complex de enzime pectinolitice în structura căruia intră o galacturonat digalacturono hidrolază (EC 3.2.1.82), o oligo-(D-galactoziduronat) liază (EC 4.2.2.6) şi o endopectat liază (EC 4.2.2.2).
Tabelul 5. Proprietăţile câtorva oligogalacturonaze microbiene după Whitaker (1990)
Microorganism Substratea,b PHoptim
Oligogalacturonat hidrolazeBacillus sp.
Bacillus spAspergillus nigerc
OligogalacturonatliazeErwinia aroideae
Erwinia carotovora
Pseudomonas sp.
trimer > tetramer > pentamer > dimer > acid pectic solubil;
n-dimer > n-trimer > n-tetramer > n-pentamer;
dimer > acid pectic solubil > n-dimer;
n-dimer > n-trimer > dimer >trimer > tetramer > acid pectic solubil;
n-dimer > dimer > trimer >tetramer > pectat;
tetramer >trimer > n-trimer >dimer > n-dimer > acid pectic
6,5
6,5
—
7,0
7,2
7,0
185

Enzime pectinolitice de origine microbiană
solubil >pectat;
asubstratele sunt prezentate în ordinea vitezei cu care sunt scindate; bterminologia utilizată: dimer, trimer, tetramer, pentamer - substrate cu 2, 3, 4, respectiv 5 resturi de acid galacturonic; n-substrate nesaturate cu resturi de acid 4,5 dehidrogalacturonic la capătul nereducător; gradul de policondensare a acidului pectic solubil este de 15-20; c proteina posedă două tipuri de activităţi hidrolazice acţionând asupra digalacturonaţilor saturaţi şi nesaturaţi.
4.6. Endopolimetilgalacturonazele
Prezenţa endopolimetilgalacturonazelelor (endopectin hidrolaze) la unele microorganisme a fost raportată de o serie de cercetători; Friedurek (1983), Wick (1982), Reddy (1983), Silley (1985). Toate aceste cercetări nu au reuşit să evidenţieze acţiunea acestor enzime asupra poligalacturonatului 100% metilat în absenţa pectinesterazei şi pectin liazei. Datorită acestui fapt, existenţa acestui tip de enzime este pusă sub semnul întrebării.
4.7. Endopectat liazele
Endopectat liazele (EC 4.2.2.2) hidrolizează poligalacturonaţi îndepărtând din structura acestora oligogalacturonaţi 4:5 nesaturaţi. Deşi endopectat liazele şi endopoligalacturo-nazele acţionează asupra aceloraşi tipuri de substrate, endopectat liazele se diferenţiază de endopoligalacturonaze prin patru trăsături caracteristice:
sunt biosintetizate numai de microorganisme, în special de bacterii şi de fungii patogeni ai plantelor şi alimentelor;
au pH-uri optime alcaline; prezenţa ionilor de calciu este absolut necesară; scindează legăturile glicozidice prin mecanism de trans
eliminare, rezultând un produs de reacţie cu o dublă legătură între atomii de carbon 4 şi 5 ai galacturonatului.
În general poligalacturonaţii şi pectinele slab metilate sunt substratele preferate ale endopectat liazelor. Există totuşi diferenţe de la o enzimă la alta. Astfel, endopectat liazele izolate din două tulpini de Arthrobacter şi dintr-o tulpină de Bacillus polymyxa, acţionează cu viteza cea mai mare asupra acizilor pectinici cu grade de esterificare de 21%, 44% şi, respectiv, 26% (Whitaker şi alţii 1990).
Studiile întreprinse de Bruhlmann (1995) au arătat că endopectat liaza, biosintetizată de o specie nesporulantă de Amycolata, degradează poligalacturonaţii prin trans eliminare întâmplătoare, produşii intermediari fiind o gamă largă de oligogalacturonaţi 4,5 nesaturaţi. Aceşti produşi intermediari sunt scindaţi în continuare sub acţiunea catalitică a enzimei, produşii finali ai reacţiei fiind de tipul dimerilor şi trimerilor.
Activitatea endoliazică scade cu scăderea lungimii catenei substratelor, majoritatea endopectat liazelor degradând extrem de lent trigalacturonaţii şi tetragalacturonaţii nesaturaţi. Pe baza acestor
186

D. Dinu
constatări experimentale, Rexová-Benková (1976) a propus două tipuri de mecanisme de acţiune diferite (modele A şi B). Endopoligalacturonazele care acţionează după modelul A au ca substrate limită trigalacturonaţii nesaturaţi, în timp ce enzimele aparţinând modelului B nu pot scinda aceşti trimeri nesaturaţi.
Un mod de acţiune diferit a fost evidenţiat de Preston (1992) în cazul formelor moleculare ale endopectat liazei din Erwinia chrysanthemi. Forma moleculară notată PLa acţionează prin mecanism random, conducând la formarea mai multor tipuri de produşi, de la dimeri la decameri. Formele PLb şi PLc generează trimeri şi tetrameri printr-o combinaţie de mecanisme endolitice şi exolitice. Forma PLd produce prioritar dimeri şi acţionează printr-un mecanism endolitic non random.
Dintre toate enzimele pectinolitice, endopectat liazele au fost cel mai mult investigate din punct de vedere al structurii. Comparând secvenţele de aminoacizi ale celor cinci pectat liaze produse de Erwinia chrysanthemi, Tardy (1997) a pus în evidenţă existenţa a două familii enzimatice cu proprietăţi diferite. Prima familie include formele moleculare de tip B şi C, forme care pot acţiona numai asupra pectinelor slab metilate şi au o afinitate de 10 ori mai mare pentru aceste substrate decât endopectat liazele aparţinând familiei a doua. Cea de a doua familie include formele A, D şi E, forme active şi pe substrate al căror grad de metilare ajunge pânăla 45%.
Studiile de difracţie cu raze X au evidenţiat prezenţa în structurile de ordin superior ale pectat liazelor C şi E din Erwinia chrysanthemi, a unui nou tip de domeniu structural care a fost numit -helix paralel (Yodler şi alţii 1993a şi b). Structura tridimensională a endopectat liazei C cuprinde numai -structuri paralele, structuri care sunt pliate într-o spirală răsucită spre dreapta (“large right handed coil“). Divizarea structurii terţiare în elemente de structură secundară a arătat prezenţa a trei -structuri paralele. Două din aceste structuri sunt pliate una faţă de cealaltă antiparalel, rezultând o structură de tip -sandwich, iar cea de a treia este perpendiculară pe acest -sandwich. Pectat liaza E din Erwinia chrysanthemi a avut o structură tridimensională asemănătoare cu cea a formei C (Lietzki şi alţii 1994). Studiile de mutageneză dirijată realizate de Jurnak (1996) au evidenţiat că pectat liazele C şi E, proteine similare din punct de vedere al plierii globale, prezintă diferenţe semnificative în cea ce priveşte mărimea şi configuraţia regiunilor “loop“ din structură. În aceste regiuni de tip “loop“ sunt localizate situsurile active ale enzimelor. Studiul structurilor tridimensionale ale endopectat liazelor C şi E a evidenţiat existenţa a două centre catalitice diferite responsabile pentru activităţi diferite (Henrissat şi alţii 1995).
Analiza structurii tridimensionale a pectat liazei din Bacillus subtilis în complex cu calciul, a evidenţiat existenţa unui domeniu -helix paralel şi a unei regiuni de tip “loop“, între care existăo cavitate la nivelul căreia ar putea fi localizat situsul catalitic activ. Situsul catalitic activ al acestei enzime are în structură un rest de arginină (Pickersgill şi alţii 1994).
Endopectat liaza din Aspergillus niger are o structură asemănătoare cu cea a pectat liazelor C şi E produse de Erwinia chrysanthemi şi este o proteină cu două funcţii catalitice (Heffron şi alţii 1995).
187

Enzime pectinolitice de origine microbiană
Studiile de modificare chimică a unor resturi de aminoacizi din structura endopectat liazei biosintetizată de Fusarium moniliforme, au evidenţiat implicarea unui rest de tirozină în legarea substratului şi a unui rest de lizină (situat în/sau lângă centrul catalitic activ) în mecanismul catalitic (Rao, 1996a).
Proprietăţile câtorva endopectat liaze microbiene sunt centralizate în Tabelul 6. Aceste enzime se caracterizează prin pH-uri optime de acţiune situate în domeniul bazic şi prin valori KM destul de mici. Studiile întreprinse de Tierny (1994) au evidenţiat modificarea proprietăţilor pectat liazei de la Bacteroides thetarotaomicron atunci când gena acesteia a fost clonată la Echeria coli.
Aspergillus niger, în culturi în fază solidă (solid-state fermentation) în care sursa de carbon a fost un amestec de tărâţe de grâu şi borhot de sfeclă, s-a dovedit a fi capabil să producă două endopectat liaze cu acţiune în domeniul acid de pH. Endopectat liaza I, enzimă cu masa de 41 700 Da şi KM pentru pectatul de sodiu 6,25 mg/ml, a acţionat cu viteza cea mai mare la pH 6,0. Endopectat liaza II, enzimă cu masa de 30 200 Da şi KM pentru pectatul de sodiu 1,35 mg/ml, a acţionat cu viteza cea mai mare la pH 4,8 (Dinu şi alţii 1998, Dinu 1997).
Endopectat liazele sunt activate de ioni de Ca2+. Studiile de spectrometrie de emisie atomică au arătat că acest ion este o parte a holoenzimei. Calciul se leagă de proteină la nivelul grupărilor carboxil din structura acesteia, dar prezenţa acestui ion nu afectează legarea substratului la enzimă (Rao şi alţii 1996a). Ionii de Ba2+, Co2+, Cu2+, Mg2+, Mn2+, Sr2+, Zn2+ nu afectează activitatea endopectat liazică (Tardy şi alţii 1997).
Inhibitori ai endopectat liazelor s-au dovedit a fi ionii bivalenţi de mercur, EDTA şi doi compuşi din plante, epicatechina şi acidul salicilic (Tardy şi alţii 1997).
188

D. Dinu
4.8. Exopectat liazele
Exopectat liazele (EC 4.2.2.9) scindează penultima legătură glicozidică din structura poli-D-galacturonaţilor cu începere de la capătul reducător al acesteia şi eliberează molecule de acid digalacturonic 4:5 nesaturate.
Prezenţa exopectat liazelor a fost semnalată doar la câteva microorganisme: Clostridium multifermentans, Erwinia aroideae, Erwinia dissolvens, Fusarium culmorum, Streptomyces nitrosporeus (citate dupâ Whitaker 1990). Guevara (1997) a pus în evidenţă existenţa a două exopectat liaze la Fusarium oxisporum sp. radicis lycopersici, proteine cu pI de 8,65 şi, respectiv, 9,0.
pH-ul optim al acestor enzime este cuprins între 8,0 şi 9,0. Cu excepţia enzimelor izolate de la speciile de Erwinia, toate celelalte au necesitate absolutăpentru ionii de calciu. Exopectat liazele nu manifestăpreferinţăfaţăde mărimea moleculei substratului, poligalacturonaţii şi oligogalacturonaţii, inclusiv trigalacturonaţii, fiind degradaţi cu aproximativ aceeaşi viteză.
189

Enzime pectinolitice de origine microbiană
Exopectat liaza din Clostridium multifermentans face parte, alături de o pectinesterază, dintr-o proteină complexă având masa de 400 000 Da. Cele două enzime din acest complex acţionează sinergic în degradarea pectinelor (Miller şi alţii 1970).
4.9. Pectin liazele
Pectin liazele (EC 4.2.2.10) manifestă specificitate pentru substrate înalt esterificate, acţiunea lor aleatoare conducând la formarea de oligo-D-galacturonaţi nesaturaţi. Majoritatea pectin liazelor purificate şi caracterizate sunt de origine fungică.
Activitatea acestui tip de liaze este afectată de distribuţia grupărilor esterificate în catena poli-D-galacturonanică. Studiile realizate pe substrate cu diferite grade de esterificare şi cu o distribuţie diferită a grupărilor esterificate, au arătat o activitate mai mare asupra substratelor deesterificate enzimatic comparativ cu cea manifestată pe substrate deesterificate în mediu alcalin. Pectinele deesterificate enzimatic au o distribuţie regulată a grupărilor esterice, în timp ce deesterificarea chimică conduce la o distribuţie statistică a acestor grupări.
Pectinele înalt esterificate sunt cele mai bune substrate pentru pectin liaze. Pectaţii, amidele acidului pectic şi glicilesterii pectinei nu sunt degradaţi de pectin liaze.
Modul de acţiune al pectin liazelor (Figura 5) a fost pus în evidenţă prin analiza produşilor rezultaţi în urma scindării tetra-, penta- şi hexa-galacturonaţilor.
Substrat Produşi de reacţie O-O-O + -O-OO-O-O-O-O-O O-O + -O-O-O
O-O-O-O-O O-O + -O-O
O-O-O-O O + -O-O
O – rest de acid galacturonic esterificat rest de acid galacturonic esterificat, cu o dublă legătură între C4 şi C5
Figura 5. Modul de acţiune al pectin liazelor.Câteva din proprietăţile pectin liazelor microbiene sunt prezentate în
Tabelul 6. Analiza datelor prezentate în acest tabel arată că majoritatea pectin liazelor au pH-ul optim în domeniul acid şi valori mici ale constantei Michaelis. Aproximativ 10% din pectin liazele izolate şi caracterizate s-au dovedit a fi enzime care acţionează la pH alcalin. O pectin liază cu afinitate mică este enzima izolată din Penicillium italicum, enzimă cu KM
190

D. Dinu
mare faţă de pectina 86% metilată (15 mg/ml), dar şi cu activitate catalitică mare. Afinitatea scăzută a acestei enzime a fost explicată de Alana (1991) prin repulsiile electrostatice care apar între enzimă şi substrat, ambele fiind încărcate negativ la pH 9,0. După depăşirea dificultăţilor de formare a complexului enzimă-substrat, activitatea catalitică este mare. Proprietăţile pectin liazei din Penicillium italicum s-au modificat în urma imobilizării covalente pe nylon (Alkorta şi alţii 1996). Creşterea stabilităţii enzimei imobilizate a permis utilizarea acesteia în 12 cicluri de degradare a pectinei.
Manifestarea activităţii catalitice a pectin liazelor nu necesită neapărat prezenţa ionilor de calciu în mediu (excepţie enzima din Fusarium oxysporum), dar ionii divalenţi de calciu, cupru, magneziu, mangan şi mercur, în concentraţie 2mM, s-au dovedit a fi activatori pentru unele pectin liaze (Lim şi alţii 1983). Activitatea pectin liazică s-a dovedit a fi inhibată de: acidul p-cloromercuri-fenil-sulfonic, anhidridă maleică, N-etilmaleimida (Manachini şi alţii1988) şi de unii hormoni ai plantelor sau analogi ai acestora.
Mikhailova (1994) a arătat că unele specii de Penicillium produc forme moleculare ale pectin liazelor care diferă din punct de vedere al controlului biosintezei. Unele dintre acestea sunt represate de glucoză şi depresate prin intermediul AMP ciclic. Fuziunea protoplaştilor de la fungii pectinolitici de tipul Aspergillus a condus la obţinerea a patru hibrizi cu activităţi endopoligalacturonazice, exopoligalacturonazice şi pectin liazice nemodificate (Solis şi alţii 1997).
O bacterie endofilică izolată de pe suprafaţa sâmburilor de cireşe s-a dovedit ca produce o pecin liază cu acţiune optimă la 400C şi pH 7,9 şi o valoare KM faţă de pectină de 4,5 mg/ml (Sakiyana şi alţii 2001).
5.Importanţa şi utilizările enzimelor pectinolitice5.1. Implicaţiile enzimelor pectinolitice în fiziologia şi patologia plantelor
Conversia protopectinei, substanţă parentală, insolubilă în apă, la pectină solubilă şi pectat şi, ulterior, la produşii lor de degradare, este un proces cu rol important în maturarea şi infectarea plantelor.
La plante, protopectina are rol de adeziv intracelular, transformarea sa în forme solubile determinând o scădere a rigidităţii ţesuturilor reflectată prin înmuierea şi, ulterior, prin lichefierea materialului plantei. Chiar dacă mecanismul acestei conversii nu este pe deplin elucidat, este evident faptul că în aceste procese un rol major îl au enzimele pectinolitice ale plantelor şi ale patogenilor, primele fiind implicate în schimbările fiziologice, iar celelalte în alterările patologice.
Paralelismul între creşterea activităţilor pectinesterazice şi endopoli-galacturonazice, pe de o parte, şi formarea pectinelor solubile în apă, pe de altă parte, paralelism observat în cursul maturării fructelor,indică o strânsă corelaţie între aceste două procese. În fructele necoapte prezenţa endopoligalacturonazei nu a fost depistată, iar activitatea pectinesterazică este foarte mică. Cele două activităţi încep să crească, existând o corelaţie
191

Enzime pectinolitice de origine microbiană
între creşterea acestora şi gradul de coacere. În fructele roşiilor transgenice, la care este expresat un RNA antisens pentru poligalacturonază, activitatea poligalacturonazică este foarte mică, ajungând până la 1% din cea depistată în fructele normale (Langley şi alţii 1994, Grierson şi alţii 1994).
192

D. Dinu
Expresia unei gene antisens pentru pectinesterază, a determinat o scădere de 10 ori a activităţii pectinesterazice în roşii (Treman şi alţii 1994). Roşiile transgenice au unele proprietăţi modificate, proprietăţi care sunt exploatate în procesele de stocare şi prelucrare a acestora.
Enzimele pectinolitice joacă un rol important în interacţia micro-organismelor cu plantele superioare, interacţie cu efecte multiple asupra ţesuturilor plantelor. Câteva dintre aceste efecte sunt:
macerarea şi distrugerea celulelor (Salinas şi alţii 1995, Federici şi alţii 2001)
apariţia reacţiilor de apărare ale plantei contra patogenului; pereţii celulari devin mai succeptibili la atacul altor polizaharidaze
microbiene; creşterea concentraţiei de etilenă, hormon care produce căderea
fructelor (Agravante şi alţii 1991).Bolile plantelor cauzate de patogeni sunt procese complexe la care
participă mai mulţi factori. Mecanismul inducerii patogenicităţii este asociat cu o interacţie între patogen şi peretele celular polizaharidic al gazdei, interacţie care influenţează producţia de enzime care degradează peretele celular (Alghisi şi alţii 1995, Yang şi alţii 1994). Rolul direct al enzimelor pectinolitice produse de patogen în procesul de infecţie este indicat prin:
capacitatea tuturor patogenilor cunoscuţi de a produce enzime pectinolitice (Rodriguezpalenzuela şi alţii 1991);
corelaţia directă între producţia acestor enzime şi virulenţa observată (Erampalli şi alţii 1995);
prezenţa enzimelor pectinolitice produse de patogen în ţesuturile plantelor infestate (Chilosi şi alţii 1997).
Producţia de enzime pectinolitice, în particular a celor cu mecanism “random“, de către patogen este una din condiţiile esenţiale pentru succesul infecţiei. Aceste enzime determină o alterare a peretelui celular, făcând posibilă penetrarea patogenului în plantele gazdă. O mutantă de Colletotrichum magna deficitară în secreţia extracelulară a pectat liazei s-a dovedit a fi nepatogenă, deşi acest microorganism este cunoscut ca fiind un patogen major al fructelor de avocado (Wattad şi alţii 1995).
Capacitatea microorganismelor de a produce enzime pectinolitice “in vitro“ nu este o dovadă a patogenicităţii lor, unele dintre ele fiind capabile să producă aceste enzime pe medii sintetice, neavând capacitatea de a le produce “in vivo“. În procesele de infecţie un rol important îl are susceptibilitatea sau rezistenţa plantei la efectele patogenului (Baayen şi alţii 1997)
Recent, un grup de cercetători canadieni (Laurent şi alţii 2000) au reuşit să obţină anticorpi policlonali faţă de o pectinesterază produsă de o genă de la Erwinia chrysanthemi expresată la de E. coli. Aceşti anticorpi nu au interacţionat cu pectinesterazele fungice sau cu cele produse de
193

Enzime pectinolitice de origine microbiană
plante, cea ce face posibilă utilizarea lor în identificarea timpurie a interacţiei plantă-patogen.
La plante au fost puse în evidenţă mai multe mecanisme de protecţie contra infecţiior microbiene. Câteva dintre acestea sunt:
conversia acidului pectic la pectatul de calciu rezistent la acţiunea enzimelor pectinolitice;
efectul inhibitor asupra enzimelor pectinolitice exercitat de unele substanţe din plante: acidul cafeic, antocianidinele, epichatechina, compuşi fenolici acidul salicilic, zaharoză (Tardy şi alţii 1997 );
inhibiţia enzimelor pectinolitice de către hormonii plantelor sau de analogi ai acestora (Chilosi şi alţii 1997);
biosinteza unor inhibitori proteici naturali ai enzimelor pectinolitice, inhibitori care formează complexe cu enzimele pectinolitice şi care contraatacă acţiunea nocivă a acestora.
Proteinele cu efect inhibitor asupra poligalacturonazelor microbiene au fost purificate şi caracterizate din fructe şi vegetale.
Din zmeură necoaptă a fost izolată o proteină cu masă moleculară de 38 500 Da, care inhibă necompetitiv două endopoligalacturonaze produse de Botrytis cinerea şi o endopoligalacturonază din Aspergillus niger (Johnston şi alţii 1993).
Perele au o glicoproteină cu masă moleculară de 43 000 Da care inhibă diferenţiat formele moleculare ale endopoligalacturonazei din Botrytis cinerea (Stotz şi alţii 1993)
Din seminţe germinate de soia a fost izolată o proteină care interacţionează diferit endopoligalacturonazele din Sclerotinia sclerotiorum şi Aspergillus niger (Flavaron şi alţii 1994).
Fructele de măr a fost pusă în evidenţă o proteină heterogenă, având grade diferite de glicozilare, care inhibă diferit cele cinci forme ale poligalacturonazelor din Botrytis cinerea (Yao şi alţii 1995).
Cu ajutorul unui biosensor, Mattei (1997) a arătat existenţa unei afinităţi diferite între inhibitorul proteic izolat din Phaseolus vulgaris şi diferite endopoligalacturonaze obţinute prin mutageneză dirijată din Fusarium moniliforme. Resturile de arginină, lizină şi histidină din structura acestui inhibitor joacă un rol esenţial în interacţii cu poligalacturonaza (Federici şi alţii 2001).
Un inhibitor proteic natural al pectinesterazelor a fost izolat şi din fructele de kiwi. Inhibitorul s-a dovedit a fi o glicoproteină care este biosintetizată sub forma unui precursor inactiv în fructele necoapte (Giovane şi alţii 1995, Camardella şi alţii 2000).
Mecanismul prin care aceste proteine inhibă activitatea enzimelor pectinolitice nu este cunoscut.
5.2. Aplicaţii biotehnologice ale enzimelor pectinolitice
Enzimele pectinolitice, singure sau în amestec cu alţi biocatalizatori, sunt utilizate pe scară largă în diferite procese biotehnologice.
Enzimele pectinolitice sunt absolut necesare în extracţia, clarificarea şi depectinizarea sucurilor de fructe (Alkorta şi alţii 1995, Chang şi alţii
194

D. Dinu
1995, Meyer şi alţii 2001). Clarificarea acestora decurge şi sub acţiunea enzimelor pectinolitice din fructe, dar procesul de autoclarificare este prea lent pentru a fi utilizat la scară industrială. În procesele industriale sunt utilizate preparate concentrate de enzime pectinolitice produse de microorganisme, preparate care realizează o degradare rapidă a polimerilor pectici. Clarificarea diferitelor sucuri necesită acţiunea sinergică a mai multor tipuri de enzime pectinolitice. Astfel, sucurile de mere au fost clarificate rapid prin combinarea activităţilor endopoligalacturonazice (din Aspergillus niger) şi pectinesterazice (din Aspergillus oryzae). Exopoligalacturonazele nu au avut efecte în astfel de procese, aceste enzime fiind adsorbite în proporţie destul de mare pe protopectina din mere (Hara şi alţii 1986). Sucurile de grapefruit, sucuri mai rezistente în procesele care implică degradarea polimerilor pectici, au fost clarificate cu ajutorul unei pectin liaze izolate din Aspergillus sojae (Ishii şi alţii 1973). Clarificarea sucurilor de citrice, sucuri cu pH foarte acid a putut fi realizată prin acţiunea combinată a pectinesterazei ce se găseşte în aceste sucuri şi a poligalacturonazei din Corticium rolfesii. Poligalacturonaza biosintetizată de Corticium rolfesii are o utilitate deosebită, deoarece ea acţionează la un pH foarte scăzut - 2,5 -, pH la care reacţia enzimatică se desfăşoară în condiţii apropiate de cele aseptice (Tagawa şi alţii 1988).
Acţiunea combinată a enzimelor pectinolitice, celulazelor şi hemicelulazelor poate conduce la lichefierea fructelor (Grassin şi alţii 1996)
Enzimele pectinolitice sunt utilizate în procesele de vinificaţie, mai ales pentru obţinerea vinurilor din soiuri de struguri bogate în substanţe pectice (Colagrande şi alţii 1994). Extracţia aromelor şi a pigmenţilor vegetali se poate face cu ajutorul acestor enzime (Solehah şi alţii 1994)
În procesele tehnologice care urmăresc prelucrarea boabelor de cafea în vederea obţinerii cafelei instant, utilizarea enzimelor pectinolitice s-a dovedit a fi eficientă (Vükari şi alţii 1993).
Enzimele pectinolitice au fost utilizate pentru macerarea fructelor şi vegetalelor, pentru obţinerea alimentelor pentru sugari şi a piureurilor (Recourt şi alţii 1996, Dijkvan şi alţii 1996).
Înmuierea mai rapidă a fibrelor vegetale s-a realizat cu ajutorul enzimelor pectinolitice (Brumaro şi alţii 1993, Baracat-Pereira şi alţii 1993). O serie de studii au evidenţiat îmbunătăţirea calităţii ţesăturilor atunci când fibrele de bumbac au fost tratate cu un amestec de hemicelulaze şi pectinaze (Csizer şi alţii 2002, Sawado şi alţii 2001).
Amestecuri de enzime pectinolitice din surse microbiene au fost utilizate în prelucrarea biomasei vegetale (Kravtchenko şi alţii 1993). Prin intermediul acestor enzime se realizează astfel curăţirea mediului şi reciclarea resurselor naturale. Deşeurile rezultate în urma prelucrării citricelor sunt bogate în glucide, fapt care permite utilizare lor pentru producerea furajelor. Tratarea acestor deşeuri cu pectinesteraze microbiene are ca rezultat formarea pectatului de calciu concomitent cu apariţia aşa numitului “fenomen de coagulare“. Acest fenomen uşurează îndepărtarea apei, realizându-se şi o economie de combustibil în procesele de uscare.
195

Enzime pectinolitice de origine microbiană
Pectinesteraza din Penicillium felutatum a fost utilizată de către Aizenberg (1996) pentru producerea pectinelor slab metilate, substanţe cu aplicaţii în medicină şi în producerea hranei pentru diabetici.
Viteza cu care amidonul din plante este hidrolizat sub acţiunea glucoamilazei produse de Aspergillus niger a fost crescută în prezenţa unui amestec de endopoligalacturonază şi pectinesterază obţinut din specii de Rhizopus (Chitradon şi alţii 1996).
Multe procese biotehnologice folosite în industria alimentară se utilizează, alături de alte tipuri de enzime pectinolitice, pectinesteraza. Acţiunea pectinesterazei conduce la eliberarea de metanol, substanţă toxică pentru organismele animale. În acest sens au fost efectuate teste toxicologice pe diferite animale. Administrarea orală, timp de 13 zile, a unei cantităţi de 10 g preparat proteic cu activitate pectinesterazică /kg corp, nu a afectat greutatea, semnele vitale, comportamentul sau parametrii biochimici ai şobolanilor (Lissau şi alţii 1998). Cu toate acestea, efectele fiziologice şi nutriţionale produse sub acţiunea metanolului trebuie avute în vedere.
Izolarea protoplaştilor din plante necesită prezenţa enzimelor pectinolitice, a celulazelor şi hemicelulazelor (Ruesink şi alţii 1988).
Lacours (1994) a arătat că enzimele pectinolitice pot fi utilizate în caracterizarea taxonomică a speciilor de Gremmenniela.
Castaldo (1996) a pus la punct o metodă enzimatică pentru dozarea concentraţiilor de pectină cu ajutorul enzimelor pectinolitice. În această tehnică, acidul D-galacturonic, rezultat din degradarea pectinelor sub acţiunea pectinesterazei şi a endopoligalacturonazei, este transformat în lactonă sub acţiunea unei glucuronolacton oxidaze. Acest proces este însoţit de o scădere a absorbanţei la 340 nm, scădere datorată oxidării NADPH, cofactor al oxidazei. O metodă asemănătoare de determinare a concentraţiei pectinelor din fructe şi vegetale a fost pusă la punct de Yoskioka (1992).
Enzimele pectinolitice au fost întrebuinţate şi în sinteza acidului ascorbic (Kulbe şi alţii 1987). În acest sens, pectina din mere a fost convertită, sub acţiunea pectinesterazei şi endopoligalacturonazei, la acid D-galacturonic. Acidul galacturonic recuperat din hidrolizat prin electrodializă, a fost convertit la acid 2-ceto-L-galacturonic. Ciclizarea catalitică, acidă sau bazică, a cetoacidului a condus la obţinerea acidului ascorbic.
Studii recente (Ruiz-Teran şi alţii 2001) au arătat o extracţie de 3 ori mai eficientă a glucovanilinei (substanţă din care se obţine vanilina) din păstăile de mazăre în prezenţa enzimelor pectinolitice.
Bibliografie
1. Agravante, J., Matsui, T., Kitagawa, H. (1991). Changes in pectin methylesterase, polygalacturonase and pectic substances of ethanol and ethylene trated bananas during ripening. Nippon Skokukim Kogyo Gakkaishi, 38 , 127-132.
2. Aizenberg, V.L., Syrchin, S.A., Sedina, S.A., Shinkarenko, L.N., Demchenko,P.I., Vitte, P.N. (1996). Properties of pectinesterase from Penicillium fellutanum Biourge and new development in pectin applications. Prog. Biotechnol., 14 [ Pectins and pectinases], 947-954.
196

D. Dinu
3. Alaña, A., Llama, M., Serra, J. (1991). Purification and some properties of the pectin lyase from Penicillium italicum. FEBS Lett., 280, 335-340.
4. Albersheim, P., Killias, U. (1962). Studies relating to the purification and properties of pectin transeliminase. Arch. Biochem.Biophys., 97, 107-115.
5. Albersheim, P. (1975). The wall of growing plants cells. Scientific American, 3, 81-95.6. Alghisi, P., Favaron, F. (1995). Pectin degrading enzymes and plant parasite
interactions. Eur. J. Plant Pathol., 101, 365-375.7. Alkorta, I., Garbisu, C., Llema, M.J., Serra, J.L. (1996). Immobilization of pectin
lyase from Penicillium italicum by covalent binding to nylon. Enzyme Microb. Technol., 18, 141-146.
8. Antov, M.G., Pericin, D.M., Dimic, G.R. (2001). Cultivation of Polyporus squamosus for pectinase production in aqueous two-phase system containing sugar beet extraction waste. J. Biotechnol., 91, 83-87.
9. Baayer, R.P., Schofffellmmeer, E.A.M., Toet, S., Elgersma, D.M. (1997). Fungal polygalacturonase activity reflects susceptibility of carnation cultivars to Fusarium wilt. Eur. J. Plant Pathol.,103, 15-23.
10. Barnavon, L., Doco, T., Terrier, N., Ageorges, A., Romieu, C., Pellerin, P. (2001). Involvement of pectin methyl-esterase during the ripening of grape berries: partial cDNA isolation, transcript expression and changes in the degree of methyl-esterification of cell wall pectins. Phytochemistry, 58, 693-701.
11. Baracat-Perreira, M.C., Santas, V.M.C., Fernandes de Aranjo,E., Olzany, S.D. (1993). Partial caracterization of Aspergillus fumigatus polygalacturonases for the degumming of natural fibers. J. Ind. Microbiol., 11, 139-140.
12. Barnby, F.M., Morpeth, F.F., Pyle, D.L. (1990). Polygalacturonases production by anaerobic fermentation of Klyveromyces maxianus. Enzyme Microb. Technol., 12, 891-897.
13. Beldman, G., Mutter, M., Searle-van Leeuwen, M.J.F., Van Den Broek, L,A.M., Schols, H.A., Voragen, A.G.J. (1996). New enzymes active towards pectic structures. Prog. Biotechnol., 14, [Pectins and pectinases],231-245.
14. Behere, A., Satyanarayan, V., Padwal-Desai, S.R. (1993). Separation and limited characterization of three polygalacturonases of Aspergillus niger. Enzyme Microb.Technol., 15, 158-161.
15. Biely, P., Benen, J.A.E., Kester H.C.M., Heinrichova, K., Visser, J. (1996a). Srereochemistry of hydrolysis of glycosidic linkage by three Aspergillus polygalacturonases. Prog. Biotechnol., 19, 706-710.
16. Biely, P., Benen, J.A.E., Heinrichova, K., Kester H.C.M., Visser, J. (1996b). Inversion of configuration during hydrolysis of -1,4-galacturonidic linkage by three Aspergillus polygalacturonases. FEBS Lett., 323, 249-255.
17. Blanco,P., Sieira, C., Diaz, A., Villa, T.G. (1994). Production and partial characterization of an endopolygalacturonase from S. Cerevisiae. Can.J. Microbiol., 40, 974-977.
18. Bonnin, E., Le Goff, A., Korner, R., Van Alebrek, G.W., Christensen, T.M., Voragen, A.G., Thibault, J. (2001). Study of the mode of action of endopolygalacturonase from Fusarium moniliforme. Biochem. Biophys. Acta, 1526, 301-309.
19. Braccini, I., Perez, S. (2001). Molecular basis of Ca 2+-induced gelation in alginates and pectins: the egg box model revisited. Biomacromolecules, 2, 1089-1096.
20. Bruhlmann, F. (1995). Purification and characterization of an extracellular pectate lyase from Amycolata sp. Appl. Environ. Microbiol., 61, 3580-3585.
21. Brumaro, M.H.N., Coelho, J.L.C., De Aranjo, E.F., Silva, D.A. (1993). Pectin lyase of P. griseoroseum related of deguming of ramie. J. Rev.Microbiol., 24, 175-178.
197

Enzime pectinolitice de origine microbiană
22. Camara Hurtado, M., Greve, L.C., Labavitch, J.M. (2002). Changes in cell wall pectins accompanying tomato (Lycopersicon esculentum Mill) paste manufacture. J. Agric. Food Chem., 50, 273-278.
23. Camardella, L., Carratore, V., Servillo, L., Giovane, A., Balestieri, C. (2000). Kivi protein inhibitor pectin methylesterase amino acid sequence and structural importance of two disulfide bridges. Eur. J. Biochem., 267, 1561-1565.
24. Capita, N.C., Defernez, M., Findlay, K., Wells, B., Shoue, D.A., Catchpole, G., Wilson, R.H., McCann, M.C. (2001). Cell wall aehitecture of the elongating maize coleoptide. Plant Physiol., 127, 551-565.
25. Cardello, L., Lourenco, E.J. (1995). Inhibitors of pectinesterases. Aliment. Nutr., 6, 25-37.
26. Chellegatli, M.A., Kawano, C.Y., Said, S., Fonseca, M.J. (2000). Sequential synthesis and secretion of pectinases by penicillium frequentans. J.Basic Microbiol., 40, 319-326.
27. Chen, E.M.W., Mort, A.J, Andrews, J., (1996). Nature of sites hydrolyzable by endopolygalacturonase in partially esterified homogalacturonans. Carbohydr. Polym., 29, 129-136.
28. Chilosi, G., Magro, P. (1997). Pectin lyase and polygalacturonase isoenzymes production by Botrytis cinerea during the early stages of infection on different host plants. J. Plant Pathol., 78, 61-69.
29. Chitradon, L., Poonpairoj, P., Mahakahan, P., Kitpreechavanich, V., Lotong, N. (1996). Pectinases from Rhizopus sp. efficient in enhancing the hydrolyzation of raw cassaya starch: purification and characterization. Prog. Biotechnol., 14 [Pectins and pectinases], 715-722.
30. Cho, S.W., Lee, S., Shin, W. (2001). The X-ray structure of Aspergillus aculeatus polygalacturonase and a modeled structure of the polygalacturonase-octopolygalacturonate complex. J.Mol.Biol., 311, 863-878
31. Colagrande, o., Silva, A., Fumi, M.D. (1994). Recent applications of biotechnology in wine production. Biotechnol. Prog., 10, 2-18.
32. Cooke, R.D., Ferber, E.M., Kanagasabapathy, H. (1976). Purification and characterization of polygalacturonase from a commercial Aspergillus niger preparation. Biochim. Biophys. Acta, 452, 440-44.
33. Cosgrove D.J. (1997). Assembly and enlargement of the primary cell wall in plants. Annu.Rev.CellDev.Bio., 13, 171-201
34. Crotti, L.B., Jorge, J.A., Terezi, H.T., Polizeli, M.L.T. (1996). Purification and characterization of galactase induced pectinase from the exo 1-mutant strain of Neurospora crassa. Prog. Biotechnol., 14 [Pectins and pectinases], 787-792.
35. Csiszar, E., Losonczi, A., Szabacs, G., Ruszuals, I., Reicher, J. (2001). Enzymes and chelating agent in cotton pretreatment. J. Biotechnol., 89, 271-279.
36. Demain, A.L., Phaff, H.J. (1954). J. Biol. Chem., 210 , 381-393, citati dupã Rexová-Benková, L., Marcovic, O. (1976). Pectic enzymes. In Advancces in carbohydrate chemistry and bichemistry, editori Tripton, R.S. si Horton, D., vol. 33, pag. 347.
37. Devi, N.A., Rao, A.G. (1996). Fractionation, purification and preliminary characterization of polygalacturonase from Aspergillus carbonarius. Enzyme Microb. Technol., 18, 59-65.
38. Dick, A.J. (1989). Cell wall metabolism in ripening fruit. Part II. Characterization of pectic polysaccharides solubilized during softening of Bartlett pear fruit. Plant Physiol.,89, 1394-1400.
39. Dijkvan, C. (1996). Pectins and pectolytic enzymes in relation to development and processing of green beans (Phaaseolus vulgaris L ). Prog. Biotechnol., 19, 399-404.
198

D. Dinu
40. Dinu, D., Dumitru, I.F. (1997). Isolation and partial characterization of a pectinesterase from Aspergillus niger. Roum. Biotechnol. Lett., 2, 489-494.
41. Dinu, D., Dumitru, I.F. (1998). Studies concerning the purification and properties of pectate lyase isolated from Aspergillus niger. Roum. Biotechnol. Lett., 3, 23-29.
42. Dinu, D. (1999). Pectic enzymes produced by Aspergillus niger MIUG 16 in solid-state fermentation on wheat bran and beet draft.. Roum. Biotechnol. Lett., 4, 287-293.
43. Di Pietro, A., Roncero, M.J.G. (1996). Purification and characterization of a pectate lyase from Fusarium oxysporum f. sp. Lycopersici produced on tomato vascular tissue. Physiol. Mol. Plant Pathol., 49 , 177-185.
44. Duwe, B., Kha, N.G., Nguyen, Q. (1996). Site-directed mutagenesis of the active site of pectin methylesterase from Aspergillus niger RH 5344. Biotechnol. Lett., 18, 621-626.
45. Elegado, F.B., Fujio, Y. (1994). Purification and some properties of endopolygalacturonase from Rhizopus sp LKN. World J. Microbiol. Biotechnol., 10, 256-259.
46. Elumalai, R.P., Mahadevan, A. (1995). Characterization of pectate lyase produced by Pseudomonas marginalis and cloning of pectate lyase genes. Physiol. Molec. Plant Pathol., 46, 109-119.
47. Errampalli, D., Kohn, L.M. (1995). Comparison of pectic zymograms produced by produced by different clones of Sclerotinia sclerotiorum in culture. Phytopathology, 85, 292-296.
48. Federici, l., Caprari, C., Mattei, B., Sanno, C., Matteo, A., De Lorenzo, G., Cervone, F., Tsernoglou, D. (2001). Structural requirements of endopolygalacturonase for interaction with PGIP (polygalacturonase-inhibiting protein). Proc. Natl. Acad. Sci., 98, 13425-13430.
49. Fiedurek, J., Ilczuk, L. (1983). Acta Alimentaria Polonica, 9, 89-92, citat din Chemical Abstract, vol. 101, 166843f (1984).
50. Flavaran, f., Davidio, R., Porcedder, F., Alghidi, P. (1994). Purification and characterization of a soybeen polygalacturonase inhibiting protein. Planta, 195, 80-87.
51. Förster, H. (1988). Pectinesterases from Phytophthora infestans. In Methods in Enzymology, vol. 161, pag. 335-350.
52. Garcia-Maceira, F.I., Di Pietro, A., Huertes-Gonzalez, M.D., Ruiz-Roldau, M.I. (2001). Molecular characterization of an endopolygalacturonase from Fusarium oxysporum expressed during early stages of infection. Appl. Envirom. Microbiol. 67, 2191-2196.
53. Gardner, J.M., Kado, C.I. (1976). Polygalacturonic acid transeliminase in osmotic shock fluid of Erwinia rubrifaciens: characterization of the purified enzyme and its effect on plant cells. J. Bacteriol., 127, 451-460.
54. Giovane, A., Balestieri, C., Quagliuolo, L., Castaldo, C., Servillo, L. (1995). A glycoprotein inhibitor of pectin methylesterase in kiwi fruits. Purification by affinity chromatography and evidence of a ripening related precursor. Eur. J. Biochem., 233, 926-929.
55. Gomes, S., Simon, G., Belarbi, A. (2001). Regulation of the expression of endopolygalacturonase gene PGU 1 in Saccharomices. Yeast, 18, 423-432.
56. Grassin, C., Fauquembergue, P. (1996). Application of pectinases in beverages. Prog. Biotechnol., 14, [Pectins and pectinases],453-462.
57. Guevara, M.A., Gonzales J.M.T., Estevez, P. (1997). Multiple forms of pectic lyase and polygalacturonase from Fusarium oxysporum f. sp. Radicis lycopersici: regulation of their synthesis by galacturonic acid. Can. J. Microbiol., 43, 245-253.
58. Hang, Y.D., Woodams, E.E. (1994). Production of fungal polygalacturonase from apple pomace. Food Sci . Technol., 27, 194-197.
199

Enzime pectinolitice de origine microbiană
59. Hara, T., Lim, J.Y., Fujio, Y. (1984). Purification and some properties of exo-polygalacturonase from Aspergillus niger cultured in the medium containg Satsuma mandarin peel. Nippon Shokuhin Kogyo Gakkaishi, 31, 581-586.
60. Hara, T., Fujio, Y., Ueda, S. (1986). Clarification of apple with polygalacturonase of Aspergillus niger and pectinesterase of Aspergillus oryzae ang their protopectin adsorption and degradation behaviour. Nippon Shokuhin Kogyo Gakkaishi, 33, 336-341.
61. Heinrichová, K., Rexová-Benková, L. (1976). Purification and characterization of an exo-D-galacturonase of Aspergillus niger. Biochim. Biophys. Acta, 422, 349-356.
62. Heinrichová, K., Dzurova, M., Rexová-Benková, L. (1992). Mechanism of action of D-galacturonan digalacturonohydrolase of Selenomonas ruminatium on oligo-galactosiduronic acids. Carbohydr. Res., 235, 269-280.
63. Heinrichová, K., Stachova, M. (1994). Pectinolytic enzymes of Clostridium lochheadii. Biologia (Bratislava), 49, 813-818.
64. Hennink, A., Ham, H., van Oort, M.G. (1996). Production, characterization and application of rhamnogalacturonase. Prog. Biotechnol., 14 [ Pectins and pectinases], 485-494.
65. Henrissat, B., Heffron, S.E., Yoder, M.D., Lietzke, S., Yurnak, F. (1995). Functional implications of structure-based sequence alignment of proteins in the extracellular pectate lyase superfamily. Plant Physiol., 107, 963-976.
66. Iguchi, K., Kishida, M., Sakai, T. (1996). Purification and characterization of three extracellular protopectinases with polygalacturonase activities from Trichosporon penicillatum. Biosci. Biotechnol. Biochem., 60, 603-607.
67. James, J.., Dubery, A. (2001). Inhibition of polygalacturonase from Verticillium dahliae by a polygalacturonase inhibiting protein from cotton. Phytochemistry, 52, 149-156.
68. Jia, J., Wheals, A. (2000). Endopolygalacturonase genes and enzymes from Saccharomices cerevisiae and Kluyveromyces maxianus. Curr. Genet., 38, 264-270.
69. Johnston, D.J., Rammanathan, V., Williamson, B. (1993). A protein from immature raspberry fruits which inhibits endopolygalacturonase from Botrytis cinerea and other microorganisms. J. Exp. Bot., 44, 971-976.
70. Jurnak, F., Kita, N., Garrett, M., Heffron, S.E., Scavetta, R., Boyd, C., Keen, N. (1996). Functional implications of the three-dimensional structures of pectate lyases. Prog. Biotechnol., 14, [Pectins and pectinases], 295-308.
71. Joyce, A.M., Fogarty, W.M. (1975). Proc.Soc. Gen. Microbiol., 3, 79-89, citaţi după Whitaker, J.R. (1990). Microbial pectolytic enzymes. In Microbial enzymes and biotechnology (editia a 2 a), editori Fogarty, W.M. si Kelly, C., Elsevier Applied Science, London, pag. 157.
72. Kester, H.C.M., Kustters-van-Someren, M.A., Margo, A., Mueller, Y., Visser, J. (1996a). Primary structure and characterization of an exopoligalacturonase from Aspergillus tubingensis. Eur. J. Biochem., 240, 738-746.
73. Kester, H.C.M., Kustters-van-Someren, M.A., Muller, Y., Visser, J. (1996b). The exo-polygalacturonase from Aspergillus tubingensis: characterization and cloning of the gene. Prog. Biotechnol., 14 [ Pectins and pectinases], 817-823.
74. Kester. H.C.M., Benen, J.A., Visser, J., Orlando, R., Bermann, C., Magaud, D., Anker, D., Douthean, A. (2000). Tandem mass spectrometric analysis of Aspergillus niger pectin methylesterase: mode of action on fully methylesterified oligogalacturonates. Biochem. J., 349(part 2), 469-474.
75. Khahn, O.N., Ruthamski, E., Leidinger, K. (1991). Characterization and expression of a genomic pectin methylesterase-encoding gene in Aspergillus niger. Gene, 106, 717-720.
200

D. Dinu
76. Kobajaschi, T., Higoki, N., Yajima, N., Suzumatsu, A., Hagilava, H., Kawai, S., Ito, S. (2001). Purification and properties of a galacturonic acid realising exopoligalacturonase from a strain of Bacillus. Biosci. Biotechnol. Biochem., 65, 842-847.
77. Koller, A., Neukom, H. (1969). Research on the degradation mecanism of a purified polygalacturonase from Aspergillus niger. Eur. J. Biochem., 7, 485-489.
78. Konno, H. (1988). Endopectate lyase from Erwinia aroideae. In Methods in Enzymology, editori Wood, W.A. si Kellogg S.T., vol. 161, pag. 381-385.
79. Kravtchenko, T.P., Penci, M., Voragen, A.G.J., Pilnik, W. (1993). Enzymic and chemical degradation of some industrial pectins. Carbohydr. Polym., 20, 195-205.
80. Kulbe, K.D. (1987). Enzymatic synthesis of L-ascorbic acid via D-uronic acids membrane-reactor integrated recovery of D-galacturonic acid from pectin hydrolysates. Ann. N.Y.Acad. Sci., 506, 543-551.
81. Lacours, N., Toti, L., Sieber, T.N., Petrini, O. (1994). Pectic enzyme patterns as a taxonomic tool for characterization of Gremmennielo sp. Isolates. Can. J. Bot., 74, 891-896.
82. Laangley, K.R., Martin,A., Stenning, R., Murray, A.J., Hobson, G.E., Schuch, W., Bird, C.R. (1994). Mecanical and optical assessment of the ripening of tomato fruit with reduced polygalacturonase activity. J. Sci. Food Agric., 66, 547-554.
83. Laurent, F., Kotoujansky A., Berteau, Y. (2000). Overproduction in E.Coli of pectin methylesterase from Erwinia chrysanthemi 3937: one-step purification, bichemical characterization, and production of polyclonal antibodies. Can. J. Microbiol., 46, 474-480.
84. Lietzki, S., Yader, K., Jurnak, F. (1994). The three-dimensional structure of pectate lyase E, a plant virulance factor from Erwinia chrysanthemi. Plant Physiol., 106, 849-862.
85. Lim, Y.K., Yamasaki, Y., Suzuki, Y., Ozawa, J. (1980). J. Agric.Biol., 44, 473-476, citati dupã Whitaker, J.R. (1990). Microbial pectinolytic enzymes. In Microbial enzymes and biotechnology (editia a 2 a), editori Fogarty, W.M. si Kelly, C., Elsevier Applied Science, London, pag. 151.
86. Lim, J.Y., Fujio, Y., Ueda, S. (1983). Purification and characterization of pectinesterase and pectin lyase from Aspergillus oryzae A-3. J. Appl. Biochem., 5, 91-98.
87. Lissau, B.G., Pedersen, P.B., Petersen, B.R., Budolfsen, G. (1998). Safety evaluation of a funga pectinesterase enzyme preparation and the use in food. Food Addit. Contam., 15, 627-636.
88. Liu, Y.K., Luh, B.S. (1978). Purification and characterization of endopolygalacturonase from Rhizopus arrhizus. J. Food Sci., 43, 721-726.
89. MacKinnen, J.M., Jardine, W.G., OKennedy, N., Renard, C.M., Jarvis, M.C. (2002). Pectin methyl and nonmethyl esters in potato cell walls. J. Agric. Food Chem., 50, 342-346.
90. MacMillan, J.D., Phaff, H.J. (1966). Exo-polygalacturonase lyase from Clostridium multifermentans. In Methods in enzymology, editori Neufeld, E.F. si Ginsburg, V., Academic Press, vol. VIII, pag 632- 645.
91. Maldonado, M.C., Shasser de Sad, A.M., Callieri, D.A.S. (1994). Purification and characterization of pectinesterase produced by a strain of Apergillus niger. Curr. Microbiol. 28, 193-196.
92. Maldonado, M.C., Shasser de Sad, A.M. (1998). Production of pectinesterase and polygalacturonase by Aspergillus niger in submerged and solide state systems. J. Ind. Microbiol. Biotechnol., 20, 34-38.
93. Manachini, P.L. (1988). Pectic enzymes from Aureobasidium pullulans LV 10. Enzyme Microbiol. Technol., 10, 682-685.
201

Enzime pectinolitice de origine microbiană
94. Manjou, A., Ibarro, J.L., Romaro, C., Canovas, M. (1992). Properties of pectinesterase and endo-D-polygalacturonase coimmobilized in a porous glass support. Appl. Biochem. Biotechnol., 32, 19-31.
95. Marcovic, O. (1989). Biologia (Bratislava), 44, 1185-1189, citat din Chemical abstracts, vol. 113, 147666g (1990).
96. Marcovic, O., Stovicková, J., Joernvall, H. (1996). Modification of tomato and Aspergillus niger pectinesterase with diethyl pyrocarbonate.. Protein Chem., 15, 127-130.
97. Martel, M.B., Letoublon, R., Fevre, M. (1998). Purification and characterization of two endopolygalacturonase secreted during the early stages of the saprophytic growth of Sclerotinia Sclerotiorum. FEMS Microbiol. Lett., 158, 133-138.
98. Mattei, B., Salvi, G., Caprari, C., Lorrenzo, G., Crescenzi, V., Cervone, F. (1996). Analysis of interaction between PGIP from Phaseoleus vulgaris L anf fungal endopolygalacturonase using biosensor technology. Prog. Biotechnol., 14 [ Pectins and pectinases],775-782.
99. Mayer, A.S., Koser, C., Adler-Nissen, J. (2001). Efficiency of enzymatic and other alternative clarification and fining treatments and haze in chery. J. Agric. Food Chem., 49, 3644-3650.
100. Mc Millan, G.P., Idnston, D.I., Perombelon, M.C. (1992). Purification to homoganity of extracellular polygalacturonase and isoenzymes of pectate lyase of Erwinia carotovora, subs. atroseptica by column chromatography. J. Appl. Bacteriol., 73, 83-86.
101. Mikhailová, R.V., Sapunová, L.I, Lobanok, A.G. (1994). Biosynthesis of pectin lyases in Penicillium adametzi, Penicillium citricum, Penicillium janthinellum. Word J. Microbiol. Biotechnol., 10, 457-461.
102. Mikhailová, R.V., Sapunová, L.I, Lobanok, A.G. (1995). The three polygalacturonases constitutively synthesized by Aspergillus alliaceus. World J. Microbiol. Biotechnol., 11, 330-332.
103. Miller, L., Macmillan, J.D. (1970). Mode of action of pectic enzymes. II. Further purification of exopolygalacturonate lyase and pectinesterase from Clostridium multifermentans. J. Bacteriology., 102, 72-78.
104. Miller, L., Macmillan, J.D. (1971). Purification and pattern action of pectinesterase from Fusarium oxysporum f.sp. vasinfectum. Biochemistry, 10, 570-576.
105. Mountfort, D.O., Atkinson,M. (1995). Evidence for the role of Na+ ang galacturonic acid in the regulation of polygalacturonase production by marine anaerobe Eubacterium sp. Strain P-1. Botanica Marina, 38, 195-201.
106. Nakagawa, T., Miyaji, T., Yurimoto, H., Sakai, H., Kato, N., Tomizuko, N. (2000). A methylotrophic pathway participates in pectin utilization by Candida boidimii. Appl. Envirom. Microbiol., 66, 4253-4257.
107. Nakamura, T., Hours, R.A., Sakai, T. (1995). Enzymatic maceration of vegetables with protopectinases. J.Food Sci., 60, 468-472.
108. Nikaidou, N., Naganuno, T., Kamio, Y., Izaki, K. (1995). Production, purification, and properties of a pectin lyase from Pseudomonas marginalis N 6301. Biosci. Biotechnol. Biochem., 59, 320-324.
109. Niture S.K., Pant, A., Kuma, A.R. (2001). Active site characterization of the single endopolygalacturonase produced by F. moniliforme NCIM 1276. Eur. J. Biochem., 268, 832-840.
110. O’Neill, M., Albersheim, P., Darvill, A. (1990). The pectic polysaccharides of primary cell wall. În Methods in Enzymology, vol. 2, pag. 415-441.
111. Pickersgill, R., Jenkins, J., Nasser, H., Robert-Boudouy, J. (1994). The structure of Bacillus subtilis pectate lyase in complex with calcium. Nat. Struct. Biol., 1, 712-723.
202

D. Dinu
112. Pitkanen, K., Heikinheimo, R., Pakkanen, R. (1992). Purification and characterization of Erwinia chtysanthemi B 374 pectin methylesterase produced by Bacillus subtilis. Enzyme Microb. Technol., 14, 832-836.
113. Preston, J.F., Rice, J.D., Jugan,O., Keen, N.T. (1992). Differential depolymerization mechanisms of pectate lyases secreted by Erwinia chrysanthemi EC 16. J. Bacteriol., 174, 2039-2042.
114. Prusky, D. (1989). Purification and characterization of an endopolygalacturonase produced by Colletotrichum gloeosporioides. Physiol. Mol. Plant Pathol., 35, 121-133.
115. Ralet, M-C, Dronnet, V., Buchholt, H.C. Thibault, J-F. (2001). Enzymatically and chemically de-esterfied lime pectins: characterization, polyelectrolyte behaviour and calcium binding properties.. Carbohydr. Res., 336, 117-125.
116. Rao, M., Narsimha, K., Asha, A., Pant, A. (1996a). Role of lysine, tryptophan and calcium in the -elimination activity of a low molecular-mass pectate lyase from Fusarium moniliforme. Biochem. J., 319, 159-164.
117. Rao, M., Narsimha, K., Asha, A., Pant, A. (1996b). Implication of tryptophan anh histidine in the active site of endo-polygalacturonase from Aspergillus ustus: elucidation of the reaction mechanism. Biochim. Biophys Acta, 1296, 167-176.
118. Recourt, K., Stolle-Smith, T., Laats J.M., Beekhuizan, J.G., Eddelaan, C.E.M., Voragen, A.G.J., Wichers, H.I. (1996). Pectins and pectolytic enzymes in relation to development and processing of green beans (Phaseolus vulgaris L). Prog. Biotechnol.,
19, 399-404.119. Reddy, S.R., Reddy, S.M. (1983). Comp. Physiol. Ecol., 8, 69, citati după Whitaker,
J.R. (1990). Microbial pectolytic enzymes. În Microbial enzymes and biotechnology (ediţia a 2 a), editori Fogarty, W.M. şi Kelly, C., Elsevier Applied Science, London, pag. 155.
120. Reignault, P., Mercier, M., Bompeix, G., Boccara, M. (1994). Pectin methylesterase from Botrytis cinerea: physiological, biochemical and immunochemical studies. Microbiology., 140, 3249-3255.
121. Rexová-Benková, L., Marcovic, O. (1976). Pectic enzymes. In Advancces in carbohydrate chemistry and bichemistry,editori Tripton, R.S. si Horton, D., vol. 33, pag. 323-381.
122. Rexová-Benková, L., Mracková, M. (1978). Active groups of extracellular endo-D-galacturonase of Aspergillus niger derived from pH effect on kinetic data. Biochim. Biophys. Acta, 523, 162-169.
123. Rho, E., Park, H.J., Kim, M.O., Chung, Y.R., Lee, C.W. (2001). Expression of exopolygalacturonase in Botrytis cinerea. FEMS Microbiol. Lett., 201, 105-109.
124. Rijssel, M., Geewig, J.G., Hansan,A.T.(1993). Isolation and characterization of an extracellular glycosylated protein complex from Clostridium thermossaccharolyticum with pectic methylesterase and polygalacturonate hydrolases activities. Appl. Environ. Microbiol., 59, 828-836.
125. Rodriguezpalenzuela, P., Burr, T.J., Collmer, A. (1991). Polygalacturonase is a vitulence factor in A. grobacterium tumefianas. J. Bacteriol., 173, 6547-6552.
126. Rombouts, F.M., Pilnik, W. (1980). Pectic enzymes.În Economic microbiology:enzymes and bioconversion, editor Ros, A.H., Academic Press, New York, vol. 5, pag. 227-282.
127. Ruiz-Teran, F., Perez-Amador, I., Lopez-Munguia, A. (2001). Enzymatic extraction and transformation of glucovanillin to vanillin from vanilla green pods. J. Agric. Chem., 49, 5207-5209.
203

Enzime pectinolitice de origine microbiană
128. Sakai, T. (1988). Protopectinase from yeasts and yeasts like fungus. In Methods in enzymology (Biomass, Part B ), editori Wood, W.A. si Kellogg, S.T., Academic Press, vol. 161,pag. 366-373.
129. Sakai, T., Sakamoto, T., Hallaert, J., Vandamme, E.J. (1993). Pectin, pectinase and protopectinase: production, properties, and applications. Adv. Appl. Microbiol. 39, 213-290.
130. Sakellaris, G. (1989a). Production, purification and characterization of extracellular pectinesterase from Lactobacillus plantarum Biotechnol. Appl. Biotechnol., 11, 503-507.
131. Sakellaris, G. (1989b). Purification and characterization of an extracellular polygalacturonaase from Lactobacillus plantarum strain B A 11. J.Appl. Bacteriol., 67, 77-85.
132. Sakiyama, C.C., Paula, E.M., Pereira, P.C., Borges, A.C., Silva, D. (2001). Characterization of pectin lyase produced by an endophylic strain isolated from cofee cherries. Lett. Appl. Microbiol., 33, 117-121.
133. Salinas, J., Verhoeff, J. (1995). Microscopical studies of the infection on gerbera flowers by Botrytis cinerea. J. Plant Pathology, 101, 377-386.
134. Sawada, K., Ueda, M. (2001). Enzyme processing of textiles in reverse micellar solution. J. Biotechnol., 89, 263-269.
135. Schejter, A., Marcus, L. (1988). Isozymes of pectinase and polygalacturonase from Botrytis cinerea Pers. In Methods in enzymology (Biomass, Part B ), editori Wood, W.A. si Kellogg, S.T., Academic Press, vol. 161,pag. 366-373.
136. Shanley, N.A., Van den Broek, L.A.M., Voragen, A.G.J., Coughlan, M.P. (1993a). Izolation and characterization of an endopolygalacturonase from phanerochaete chrysosporium. J. Biotechnol., 28, 179-197.
137. Shanley, N.A., Van den Broek, L.A.M., Voragen, A.G.J., Coughlan, M.P. (1993b). Physicochemical and catalytic properties of three endopolygalacturonase from Penicillium pinophilum. J. Biotechnol., 28, 199-218.
138. Shevchik, V.E., Condemine, G., Hugouvieux-Cotte-Pattat, N., Robert-Boudouy, J. (1996a). Characterization of pectin methylesterase B, an outer membrane lipoprotein of Erwinia chrysanthemi 3937. Mol. Microbiol., 19, 455-466.
139. Shevchik, V.E., Condemine, G., Hugouvieux-Cotte-Pattat, N., Robert-Baudouy, J. (1996b). Pectin methylesterase B of Erwinia chrysanthemi, the first pectinase characterized as a membrane lipoprotein. Prog. Biotechnol., 14, [Pectins and pectinases], 837-844.
140. Shevchik, V.E., Hugouvieux-Cotte-Pattat, N. (1997). Identification of a bacterial pectin acetyl esterase in Echerichia coli 3937. Mol. Microbiol. 24, 1285-1301.
141. Silley, P. (1985). A note on pectolytic enzymes of Lachnospira multiparus. J. Appl. Bacteriol., 58, 145-149.
142. Solehah, A., Balaumani, K.D., Amizo, M.A. (1994). Enzymes for improved extraction and stabilization of colour and flavour of orange juice. J. food Sci. Technol., 31, 508-510.
143. Solis, S., Flores, M.E., Huitron, C. (1997). Improvement of pectinase production by interspecific hybrids of Aspergillus strains. Lett. Microbiol., 24, 77-81.
144. Statilová, E., Markovic, O., Skrovinová, D., Jörnavall, H. (1993). Pectinase of Aspergillus sp. polygalacturonase: multiplicity, divergence, and structural patterns linking fungal, bacterial and plant polygalacturonases. J. Protein Chem., 12, 15-22.
145. Statilová, E., Breierová, E., Vadkertiová, R. (1996a). Candida boidinii-a new found producer of pectic enzymes complex. Prog. Biotechnol., 14, [Pectins and pectinases], 453-462.
146. Statilová, E., Dzurová, M., Markovic, O., Jörnavall, H. (1996a). An essential tyrosine rezidue of Aspergillus polygalacturonase. FEBS Lett., 382, 140-146.
204

D. Dinu
147. Statilová, E., Mislovicová, D., Kacuraková, M., Machová, E., Kolarová, M., Markovic, O., Jörnavall, H. (1998b). The glycoprotein character of multiple forms of Aspergillus polygalacturonases. J. Protein Chem., 17, 173-179.
148. Stotz,H.V., Powell, A.L.T., Damon, S.E., Greve, L.C., Bennett, A.B., Labavitch, J.M. (1993). Molecular characterization of a polygalacturonase from Pyrus communis L. cv. Bartlett. Plant Physiol., 102, 130-138.
149. Stotz,H.V., Contos, J.A., Powell, A.L.T., Bennett, A.E., Labavitch, J.M. (1994). Structure and expression of an inhibitor of fungal polygalacturonases from tomato. Plant Mol. Biol., 25, 607-617.
150. Tagawa, K., Kaji, A. (1988). Polygalacturonase from Corticium rolfsii. In Methods in enzymology (Biomass, Part B ), editori Wood, W.A. si Kellogg, S.T., Academic Press, vol. 161,pag. 361-365.
151. Takao, M., Nakaniwa, T., Yoshikawa, K., Terashita, T., Sakai, T. (2001). Purification and characterization of thermostable pectate lyase with protopectinase activity from thermophillic Bacillus sp. TS 47. Biosci. Biotechnol. Biochem., 64, 2360-2367.
152. Tierny, Y., Bechat, M., Joncquiert, J-C., Dubourguier, H-C., Guillaume, J. (1994). Molecular cloning and expression in Escherichia coli og genes encoding lyase and pectin methylesterase activities from Bacteroides thetarotaomicron. J. Appl. Bacteriol., 76, 592-602.
153. ten-Have, A., Breuil, W.O., Wublen, J., Visser, J., van Kar, J. (2001). Botrytis cinerea endopolygalacturonase genes are differentially expressed in various plant tissues. Fungal Genet. Biol., 33, 97-105.
154. Treman, D,M., Handa, A.K. (1994). Reduction of pectin methylesterase activity modifies tissue integrity and cation levels in ripening tomato fruits. Plant Physiol., 106, 429-436.
155. Ueda, S., Fujio, Y., Liu, J.V. (1982). Pectic enzymes from Aspergillus oryzae. J. Appl. Biochem., 4, 524-535.
156. Vilanova, M., Blanco, P., Cortes, S., Villa, T.G., Siliro, C. (2000). Use a PGU 1 recombinat saccharomyces cerevisiae strain in oenological fermentations. J. Appl. Microbiol., 89, 876-883.
157. Vucari, L, Tenkanen, M, Buchert, J., Rathrö, M., Barley, M., Linko, M. (1993). In bioconversion of forest and agricultural plant residues, editor Saddler, J.N., International Oxford, pag. 131-182.
158. Wattad, C., Freeman, S., Dinoor, A., Pruski, D. (1995). Colletotrichum magna is deficient in extracellular secretion of pectate lyase. Mol. Plant-Microbe Interact., 8, 621-626.
159. Whitaker, J.R. (1990). Microbial pectolytic enzymes. In Microbial enzymes and biotechnology (editia a 2 a), editori Fogarty, W.M. si Kelly, C., Elsevier Applied Science, London, pag. 133-176.
160. Yamaguchi, F., Inoue, S., Hatanaka, C. (1995). Purification and properties of endo-1,4-D-galacturonase from Aspergillus niger. Biosci. Biotechnol. Biochem., 59, 1742-1744.
161. Yang, H.A., Zhou, J., Sivasithamparam, H., Tommerup, I.C., Barton, J.E., Obrien, P.A. (1994). Genetic variability in pectic enzymes of Rhizoctonia solarii isolates causing bare-patch disease of cereals. J. Phytopathology, 141, 259-266.
162. Yao, C., Conway, W.S., Sams, C.E. (1995). Purification and characterization of a polygalacturonase produced by Penicillium expansum in apple fruit. Phytopathology, 86, 1160-1166.
163. Yoder, M.D., Keen, N.T., Jurnak, F. (1993a). New domain motif of the structure of pectate lyase C, a novel secreted plant virulance factor. Science, 260, 1903-1907.
205

Enzime pectinolitice de origine microbiană
164. Yoder, M.D., Lietzke,S.E., Jurnak, F. (1993b). Unusual structural features in the parallel -helix in pectate lyases. Structure, 1, 241-251.
165. Yoskioka, S., Ukedo, H., Matsunuoto, K., Osajimo, Y. (1992). Enzymatic determination of pectin and related lower-molecular weight compounds in fruits and vegetables using polygalacturonase, pectinesterase, glucuronolactone reductase and ascorbat axidase. Nippon Skokukin Kogyo, 39, 821-826.
206


















