
drogenesis in Hordeum vutgare L.: Physïological and...
130
drogenesis in Hordeum vutgare L.: Physïological and molecular aspects Sietske Hoekstra
Transcript of drogenesis in Hordeum vutgare L.: Physïological and...

drogenesis in Hordeum vutgare L.:
Physïological and molecular aspects
Sietske Hoekstra


}ndrogenesis in Hordeum vulgare L.:
Physiological and molecular aspects
Proefschrift
ter verkrijging van de graad van Doctoraan de Rijksuniversiteit te Leiden,
op gezag van de Rector Magnificus Dr. L. Leertouwer,hoogleraar in de faculteit der Godgeleerdheid,volgens besluit van het College van Dekanen
te verdedigen op donderdag 5 september 1996teldokke 15.15 uur
door
Sietske Hoekstrageboren te Amsterdam in 1963

romotiecommissie
Promotor Prof.dr. RA. Schilperoort
Co-promotor Dr. f. Heidekamp (TNO)
Referent Prof.dr. E. Heberle-Bors (University of Vienna, Austria)
Overige leden Prof.dr. KR UbbengaProf.dr. J.W. KijneDr. S.C. de Vries (Landbouw Universiteit Wageningen)
The research desaibed in this thesis, has been performed within the ABIN-project
and from 1992 in the strategic collaboration between the TNO department Plant
Biotechnology and the Institute of Molecular Plant Sdences section 1 of Ieiden
University, in the Center for Phytotechnology.finandal support for the publicaüon of this thesis by the TNO Nutrition and food
Research Institute is greatfully acknowledged.

Aan mijn ouders,
Voor Uli

I.bbreviations
ABA = (±)2-ds-4-trans-absdsic acidABA = intracellular ABA
ABAe = extracelküar ABABAP = 6-benzylaininopurined.a.i. days after isolation of microsporesELS = embxyo-like structuresIAA = indolacetic addLOS = lipo-oligosaccharides2,4-D = 2,4-dicfflorophenoxyaceüc acid

page
Outline of the thesis 9
Chapter 1 General introduction ii
Chapter 2 Anther and microspore culture of Hordeum vulgare 1.cv. Igri 31Plant Science 86 (1992) 89-96
Chapter 3 Microspore culture of Hordeum vulgare L.:the influence of density and osmolarity 41Plant Ceti Reports 12 (1993) 661-665
Chapter 4 The interaction of 2,4-D application and mannitolpretreatment in anther and microspore culture ofHordeum vutgare 1. cv. Igri 49Journal of Plant Physiology 14& 696-700 (1996)
Chapter 5 Androgenesis in Hordeum vulgare L.: The role of ABAduring anther pretreatment, on a solution containingmannitol and calcium 59Manuscript subnfltted
Chapter 6 Androgenesis in Hordeum vulgare 1. Apoptosis andthe effect of lipo-oligosaccharides during antherpretreatment -
75Manuscript in preparation
Chapter 7 General Discussion 89
Summary 109
Samenvatting. 113
Curriculum vitae 117
Publications 119
Nawoord 121


()utline of the
he development of pollen is an intriguing process, resulting in unicellularstructures. These ceils have a rigid ceilwall, which enables a resistance towithstand various kinds of stress. Uponappropriate conditions, these structures
are able to hydrate, gemlinate and when apoiien tube is formed, to partidpate inthe fertilization process.
This gametophytic development can bearrested and changed into the sporophyticroute. Immature pollen grains, also calledmicrospores, can be induced at the vergeof mitosis to become what are called embryogenic microspores. Under spedflcconditions, the microspore will developinto an embryo-like structure and fromthis a haploid plant can be fonned. Themechanism of this switch from thegametophytic into the sporophyticpathway has attracted the attention ofmany researchers. Moreover, the productof the switch, a homozygous plant, hassurplus value for plant breeding. By theuse of haploids from immature pollengrains, the production of new varietiescan be largely improved. The rapid andcomplete homozygosity of the offspringsimplifies phenotype selection for quanfitative inherited characters and, therefore,
breeding is more effi&nt. Today,androgenic haploïds have been produced
in more than 50 genera, and the greatesteffort has been given to economicallyimportant plants such as the cereals andvegetable crops. Although the anther ciiiture method is used successfully in breeding companies, very little is knownabout the mechanism(s) responsible forchanging microspore development intothe sporophytic pathway. What is knownis that an appropriate microsporedevelopmental stage together with a specific stress treatment is required for theproduction of microspore-derived plants.
A few model spedes have been used formicrospore-derived plant formation, namely tobacco, rapeseed and barley. forstudies presented in this thesis we usedthe crop barley, as in the TNO department of Plant Biotechnology, studies arebeing performed aiming at adaptation ofbarley for industrial needs (see Bengtsson,1992, McEfroy and Jacobsen, 1995). Theaim of the studies for this thesis was twofold: tiying to understand the mechanïsmleading to fonnation of embryogenic microspores, and optimization of plant production efflciency from androgenesis. Theimportance of several parameters in androgenesis has been described in literatu
9

Outtine of the thesis
re. We were specificafly interested in parameters which are responsïble for theinitiation and early development of thesporophytic pathway. Three preconditions were identified for androgenesis inbarley: a uninucleate developmental stage,an anther pretreatment and a growth regulator application.
eneral introduction of androgenesisfor the model spedes barley is given inchapter 1 of this thesis. In chapter 2, acomparison of the regeneration effidencybetween anther and microspore culture ismade. The basics of androgenesis is described, with an emphasis on the importance of the late uninudeate microsporedevelopmental stage. Microspore cultureproved to 5e more efficient in plant production than anther culture. Studies described in chapter 3 were focussed on thedevelopment of an effident microsporeculture protocol. After an optimized anther pretreatment, an embiyogenic subpopulation was identified resulung in theproduction of 50 green plants per anther.In chapter 4 a study on the effects of thegrowth regulator 2 ,4-dichlorophenoxyace-tic add (2,4-D) is presented. Pulse appli
cation of the synthetic awdn 2,4-D caninduce plant formation at similar frequendes as described for the cytokinine benzylaminopurine, which is generally used. Inchapter 5 and 6, the importance of antherpretreatment is demonstrated. An appropriate osmoüc level during pretreatmentis necessary for plant formation. We alsoshowed evidence for the need of specificabsdsic add levels, endogenously as wellas exogenously, during the pretreatment.In chapter 6 is shown, that these hormonal and osmotic levels might be necessaryfor induction of apoptosis in anther tissue. Also, a possible role of lipo-oligosaccha
rides in the formation of embiyogenicmicrospores was demonstrated.In the last chapter of this thesis a hypothesis is presented, based on the resuksobtained from our studies, for a mechanism by which androgenesis is initiated.
I..eferencesBengtsson B.O. Barley geneucs - not only here for
die beer. Trends 1. Genet. 8:3-5 (1992)McElroy D. and J. Jacobsen. What is brewing inbarley biotechnology? Biotechnology 13: 245-249
(1995)
10

hapter 1
General Introduction
aploidization
for vaiiety development in most cropplants, a critical step is the establishment
of truc breeding lines. A stable,homozygous for as much as possible)plant is defined as a tnie breeding line.Such plants are used as finished varietiesor as parents for hybrid seed production.Traditionally, plant breeders haveachieved homozygosity by using the timeconsuming processes of self-fertilizationor backaossing fMorrison and Evans,1988). Homozygosity in vivo arises uponabnormal fertilization by reduction of thegenetic information to the amount ofDNA present in gametes; these plants, inwhich each gene is represented once, arecalled haploids. There are several originsof in vivo haploids, i.e. parthenogenesis(autonomous growth of the ovary withoutfertilizaüon), gynogenesis (growth of theunfertilized egg in the presence of normalendospenn) and genome elimination(natural elimination of either gameticgenomes after fertilization). Naturalhaploids of higher plants are, however, ofrare occurence and restricted to only a
few spedes (Maheshwari et al., 1980).With the publication of the paper by
Guha and Maheshwari in 1966, a newhaploidization approach came intoexistence: development of haploid plantsderived from immature pollen ormicrospores in cultured anthers, what iscalled (in vitro) androgenesis. Haploidsderived from microspores opened a newdimension because of the abundance ofmicrospores produced by the plants andtheir potentially general occurrence in theplant kingdom (Heberle-Bors, 1985).Subsequently, in many laboratories agreat deal of research effort was inmtediately directed towards perfecting thetechnique so that it could routinely beused.
The plant which led to the ïniüalsuccess was a common weed, Daturainnoxia MIII., belonging to the familySolanaceae. Today, androgenic haploidshave been produced in more than 50genera, but the greatest effort has beengiven to economically important plantssuch as the vegetable crops and cereals(Cao et al., 1995; Veilleux, 1994). Viaandrogenesis new varieties have beendeveloped in a number of agricukural
11

Chapt2rl
crops such as Brassica sp., tobacco, potato,asparagus, wheat, rice, maize, barley, etc.(Bajaj, 1990).
In this thesis, the development of amethod for androgenesis in barley andthe analysis of parts thereof, aredescribed. For a good comprehension ofthe other chapters, the reproductivedevelopment, the process of microsporogenesis and various aspects ofandrogenesis are introduced.
he reproductive system andembryogenesïs
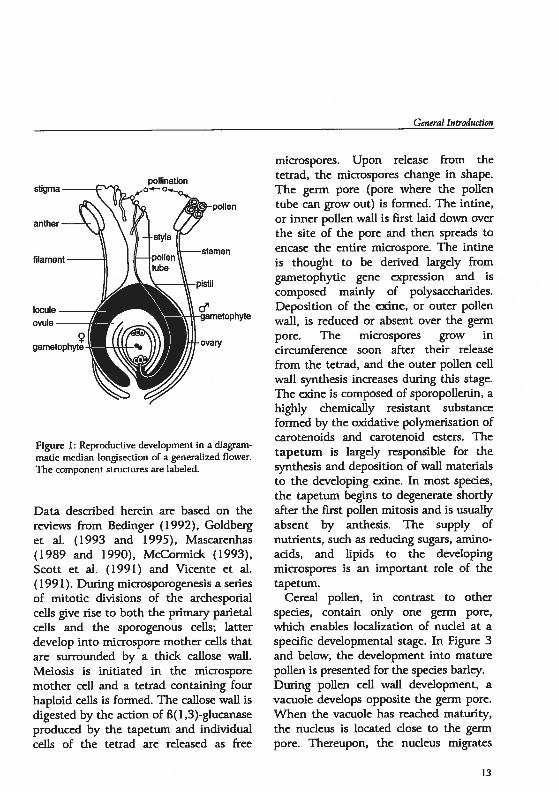
Angiosperm plants produce flowers, verybeautiful and intricate structures, inwhich their reproductive developmenttakes place. In flowering plants a sporeprodudng generation or sporophytealtemates with a gamete-produdnggeneration or gametophyte. Unlike someof the evolutionarily more primitiveplants, the male and female gametophytesof angiosperms are reduced tomicroscopic stnictures that are dependenton the tissues of the sporophyte for theirdevelopment. The flower containsspedalized structures (see Figure 1), theanthers and the pistil in which the maleand female gametophytes, respectively areformed. The role of the male gametophyte (i.e. microspores) is to producetwo generaüve celis which are transportedvia a pollen tube, through the tissues ofthe style into the ovaly. In the doublefertilization that follows, fusion of onegenerative nucleus with the egg and of thesecond generative nucleus with thecentral celi resuits in formation of
respectively the zygote and theendosperm, a nutritive tissue. The zygotecontinues to develop to the structure ofan embiyo within the ovule, and thisprocess is called zygotic embryogenesïs.In some spedes, the embryogenicpathway is well documented. Indicotyledonous spedes the stages ofoctant, globule, heart and torpedo aretypical, whereas in monocotyledonousspecies the pro-embryo, the transiüon,and the coteoptilar stage with a scutellumare formed (Meinke, 1991).
A characteristic of plant cells is theirtotipotency, which means that they havethe ability to regenerate into a plantunder the appropriate in vitro conditions.1f somaüc ceils replay in vitro adevelopmental program leading to theproduction of embryos, this process iscalled somatic embryogenesis. Theembryos arise direcily from the explant,or indirectly after a phase ofdedifferentiated growth. Altematively,somatic cells develop in vitro via organogenesis into a plant, which is mostly anadvenütious shoot (Reynolds, 1994).Regenerants of somatic cells keep thegenetic constitution of the mother plantunder appropriate conditions, so nohaploid or homozygous plants are to beexpected.
icrosporogenesis
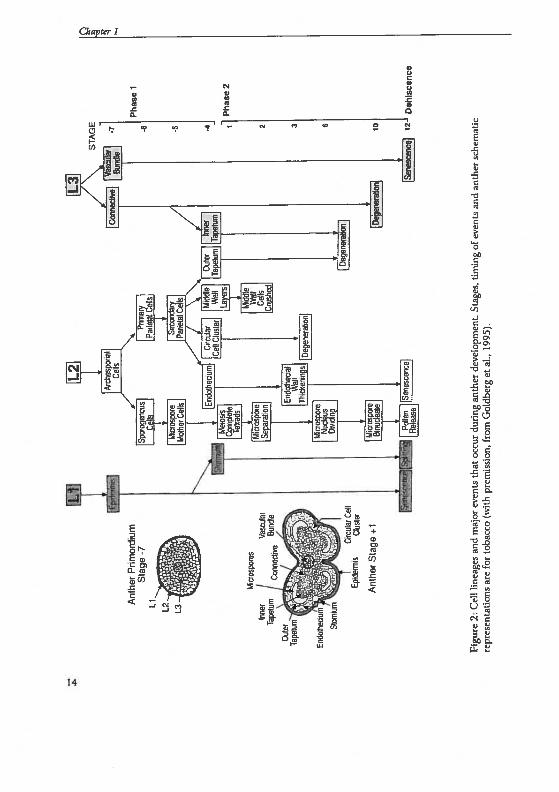
In flowering plants, the pollen is the malegametophyte. The male gametophytecompletes its development within theanther, which is illustrated in figure 2and 3 and described in the following.
12
General lutroduction
Data described herein are based on thereviews from Bedinger (1992), Goidberget al. (1993 and 1995), Mascarenhas(1989 and 1990), McCormick (1993),Scott et al. (1991) and Vicente et al.(1991). During microsporogenesis a seriesof mitotic divisions of the archesporialcelis give rise to both the primary parietalcelis and the sporogenous ceils; latterdevelop into microspore mother ceils thatare surrounded by a thick callose wall.Meiosïs is initiated in the microsporemother celi and a tetrad containing fourhaploid ceils is formed. The callose wall isdigested by the actïon of B( 1 ,3)-glucanaseproduced by the tapetum and individualcelis of the tetrad are released as free
microspores. Upon release from thetetrad, the microspores change in shape.The geini pore (pore where the pollentube can T0W Out) i5 fomied. The intine,or inner pollen wall is first laid down overthe site of the pore and then spreads toencase the entire microspore. The intineis thought to be derived largely fromgametophytic gene expression and iscomposed mainly of polysaccharides.Deposiüon of the exine, or outer pollenwall, is reduced or absent over the germpore. The microspores grow incircumference soon after their releasefrom the tetrad, and the outer pollen ceilwall synthesis increases during this stage.The exine is composed of sporopollenin, ahigNy chemically resistant substancefomied by the oxidative polymerisation ofcarotenoids and carotenoid esters. Thetapetum is largely responsible for thesynthesis and deposition of wall materialsto the developing exine. In most species,the tapetum begins to degenerate shortlyafter the first pollen mitosis and is usuallyabsent by anthesis. The supply ofnutrients, such as redudng sugars, amino
acids, and lipids to the developingmicrospores is an important role of thetapetum.
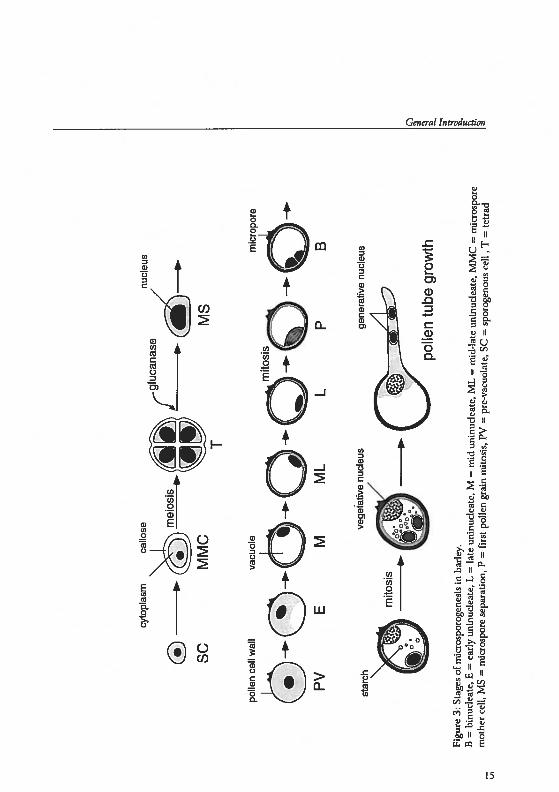
Cereal pollen, in contrast to otherspecies, contain only one germ pore,which enables localization of nudei at aspedfic developmental stage. In figure 3and below, the devdopment into maturepollen is presented for the spedes barley.During pollen cell wall development, avacuole develops opposite the genn pore.When the vacuole has reached maturity,the nudeus is located close to the gennpore. Thereupon, the nucleus migrates
figure 1 ReproducUve development in a diagrammatic median longisection of a generalized flower.The component structures are labeled.
13
Pha
se1
Pha
se2
STA
GE .71 -6 -5
Ant
her
Pri
mor
dium
Sta
ge
-7
L2
.
Mcr
ospo
res
tnne
rVa
scul
arTa
pelu
m/C
onne
ctee
Bjnd
e
Outo
r
Epid
erm
sCl
usle
r
1Micr
ospo
reA
nthe
rS
tag
e÷1
pucI
eate
RÎ
sscencc1
.4 Ii 10 i-’
Deh
isce
nce
Fig
ure
2:C
eli
line
ages
and
mat
orev
ents
that
occu
rdu
ring
anth
erde
velo
pmen
t.St
ages
,ti
min
gof
even
tsan
dan
ther
sche
mat
icre
pres
enta
tion
sar
efo
rto
bacc
o(w
ithpr
emis
sion
,fr
omG
oidb
erg
etal
.,19
95).
nucl
eus
cyto
plas
mca
llos
e mei
osis
/_Iucanase5/
SCM
MC
MS
T
poll
ence
llw
all
vacu
ole
mic
ropo
re
PVE
MM
LL
PB
vege
fati
venu
cleu
sst
arch
mito
sis
Fig
ure
3:S
tage
sof
mic
rosp
orog
enes
isin
barl
ey.
8=
binu
clea
te,
E=
earl
yun
inud
eate
,L
=ta
teun
inud
eate
,M
=m
idun
inuc
leat
e,M
L=
mid
-lat
eun
inud
eate
,M
MC
=m
icro
spor
em
othe
rce
il,M
S=
mic
rosp
ore
sepa
rati
on,
P=
firs
tpo
llen
gram
mit
osis
,PV
=pr
e-va
cuot
ate,
SC
=sp
orog
enou
see
R,T
tetr
ad
gen
erat
ive
nucl
eus
polle
ntu
begr
owth

Chapter 1
along the ceil wall til it reaches a locatÏonopposite of the genn pore, what is calledthe late uninucleate stage, to undergoan asymmetric mitotic dwision or firstpollen mitosis. This resuits in a pollencontaining a large vegetative and a smallgeneratÏve celi, that is endosed entirelywithin the vegetative ceil. The vegetativenudeus does not undergo another mitosis,and its role is to serve as a “powerhouse”to drive the further development of thepollen and the growth of the pollen tubeso that the two generative nuclei aredelivered to the ovaly. The generative cellhas a condensed nucleus and a reducedamount df cytoplasm compared to thevegetative celi. The generaüve ceil must
undergo another mitoüc divisïon.Development of the immature pollencontinues, induding the further elaboration of the outer pollen cell wall and thedeposition of reserves like starch granules.The release of the mature pollen from theanther is called dehiscence and takesplace during anthesis. In approximately70% of plant families (e.g. Solananeae, andLiÏiaceae), the pollen is released from theanther when it consïsts of just these twoceils; the second mitotic division of thegeneraüve cdl occurs while the pollentube grows through the female pistil. Inother plant families (e.g. Cruciferae andGramineae), this second mitotic dMsionoccurs before the pollen is shed from theplant. There, the mature gametophyte isfinally formed when a mitoüc division ofthe generative cel produces a pair ofgametes. Mature pollen contain a store ofpresynthesized mRNAs. It is known thata surprisingly large number of genes isexpressed in pollen. In Tradescan tja, about
60% as many genes are expressed in themale gametophyte as are expressed inshoots, that consits of several differentceli types. Most of these genes areexpressed in both pollen and sporophytictissues while about 5% are pollen-specific.The transcripts stored in mature pollenwill be translated into proteins that areneeded for processes during and afterpollen genninaüon. For a while followingdehiscence from the anther, the pollenexists as a free organism, protected under
extreme conditions by the exine, until it
is carried by wind, insects, or an otheragent to the stigma of an appropriate orcompatible flower, where the rightconditions are present for gennination ofthe pollen.
The development of the ganietophytecan be arrested and changed into the
sporophytic pathway. A plant can beproduced from immature pollen, 1f theyare cultured at the verge of mitosis in aspedfied medium and condition. Such adevelopmental switch resuks in theproduction of a plant, containing ahaploid genome, from androgenesispresumably via embryogenesis.
drogenesis in model systemsIn this paragraph, the current status ofthe sdentific knowledge concerningandrogenesis of the three model spedesBrassica napus, Nicotiana tabacum andHordeum vutgare is descrÏbed.
In Brassiar napus or rapeseed, thegametophytic development is maintainedat a temperature of 18°C and can bechanged into the sporophytic pathway byelevating the temperature to 32°C.
16

General Introductian
Microspores in the late uninudeate orearly binucleate stage are competent forplant formafion (Custers et al., 1994).The majority of vegetative nuclei re-enterthe cel cyde after 12 hours of culture at32°C (Binarova et al., 1993). About 8hours of culture at 32°C is suffident toinduce this embryogenic developmentirreversibly (Pechan et al., 1991). Such aheat shock induces a program of geneexpression in which the synthesis of afamily of proteins, that is called heatshock proteins, is higffly induced. Theseproteins result from de navo synthesis andbelong to the 70-kDa dass (Cordeweneret al., 1995). Upon heat treatment, themorphology of the ceils is changed. Thecentral vacuole becomes fragmentedallowing the nudeus to assume a centralposition within the ceil. Starch grains arenot present, the ceil develops a thickfibrillar wall, situated immediatelyadjacent to the intine, and largeaggregates of globular material (Zald andDickinson, 1990). In embryogenicmicrospores, i.e. microspores destined tobecome embryos, new microtubulararrangements are observed. The firstsymmetrical division starts in thevegetative nucleus provided the generative nudeus is arrested near the pollencell wall (Hause et al., 1993; Zaki andDickinson, 1990). The ceils dividerandomly within the exine for 4 to 7 daysfollowing heat treatment. Then multicellular structures are released from theexine and undergo peridlinal divisionsresulting in protoderm differentiation of aglobular embryo (Telmer et al., 1995).
In Nicotiana tabacum or tobacco,immature pollen develop into embryo-
genic microspores, 1f they are isolated atthe unicellular or the mid-binudeate stageand cukured in a nutritionally poormedium lacldng sucrose and nitrogen,what is called starvation. After an initialphase of gametophytic gene expression,the normally quiescent vegetafive cdlenters a phase of dedifferentiation duringwhich the bulk of its cytoplasm isdegraded. Organelles are virtually dearedfrom the ceil and little more than thenucleus and a few structurally simplifiedplastids clustered around it remain(Dunweil and Sunderland, 1974 a and b;Ganido et al., 1995). The embryogenicmicrospores have similar characterisfics asabove described for rapeseed (HeberleBors, 1989). One of the crudal eventsduring embryogenic induction is thederepression of the G1 arrest in the cdlcyde of the vegetative cell (Zarsky et al.,1992). Starvation induces de navotranscription of specific genes, a.o. atranscript for a low molecular weight heatshock protein (Garrido et al., 1993;Zarsky et al., 1993). These proteins,however, are not detected. Only recentlyTouraev et al. (1996 and in press) showthat heat shock is a potent inductivetreatment for tobacco. Apparently themRNAs are not translated in vivo butaccumulate in the embryogenicmicrospores in a translationally inactiveform (Garrido et al., 1993). Further,changes in the activity of protein ldnasehave been obsenred during stawation(Garrido et al., 1993), suggesting thatprotein phosphorylation cascades a reinvolved in the transduction of the signal.Indeed, Kyo and Harada (1 990a and b)detected phorphorylaüon of proteins
17

Qapter 1
spedflc for embryogenic microspores.
Finally in Hordeuin vutgare or barley, a
cold pretreatment or a staivation (i.e.
nutritionally poor medium as in tobacco)
induces embryogenesis in mid-late to late
uninucleate microspores (chapter 2). A
cold pretreatment for 3 to 4 weeks is
perfonned with spikes (Huang and
Sunderland, 1982), whereas for a 3 to 4
day starvation pretreatment anthers are
first dissected from the spikes (Roberts
Oehlschlager and Dunwell, 1990). Upon
cold pretreatment of spikes, degeneration
of the tapetum is observed. In fact,
changes in the morphology of the anther
resembie those observed for a matuling
anther, whereas the pollen development is
blocked before the first pollen mitosis.
Apparently, the normal sequence of
events is disrupted in the cold
(Sunderland et aL, 1984).In rapeseed and tobacco, haploid plants
are produced via androgenesis. Themajority of microspore-derived plants in
barley, however, have a doubled haploid
genome, resulting from spontaneous
diploidization. Spedflcally in barley, a
multiplicity of initial division pattems is
revealed bi vitro upon pretreatment of the
microspores. Independent dMsion of the
generative and vegetative nucleus, resuits
in the formation of chimexical units
partiüoned into distinct embryo- andendosperm- (or suspensor-) like compo
nents. Partitioning occurs only with
anther excision during the mid uninucleate microspore developmental stage
when the nuclei are probably still in the
pre-DNA replication phase (GI phase),
but anther exdsion at the early
unïnudeate stage is ineffective and leads
to rapid degeneraüon of the microspores.
With excisïon at the late uninucleate
stage, DNA replicadon is (nearly)
completed and many microspores are in
the G2 interphase. Independent contri
bution by the generative ceil is blocked
and the microspores develop into the A,
B, or C pathways. The vegetative ceil
behaves either as a haploid embryo
mother ceil without any contribution
from the generative celi fA pathway), or a
non-haploid embryo mother celi derived
from incorporation of the generative ceil
by nudear fusion (C pathway). The
microspore itself also sometimes functions
as an embryo mother-ceil, as two
chromosome complements seemingly
similar, and these might result from
fusion either in interphase or in prophase
(B pathway) (Sunderland et al., 1979;
Sunderland and Evans, 1980). This
multiple origin, from either the vegetative
or the generative ceil or both, can account
for the different ploidies found in
regenerated plantiets (Sunderland et al.,
1979; Sunderland and Evans, 1980).
Nuclear fusion has been emphasized as
the major source of ploidy increase in
barley, but endoreduplication (omission
of the G2 and M-phase; Nagi, 1995) and
nudear restitution (mitosis stops because
the spindle breaks down; Nagl, f995) are
not ruled out. fusion of the generative
and vegetative nuclei has been observed
in cultured microspores of barley by Lee
and Chen (1987), suggesting that ploidy
increases are generated veiy early in
culture.In summaly, in all three model species
there is very superfidal knowledge
available about the mechanism of
18

General Introduction
androgenesis induction. In the modelspedes barley, sofar only microscopicalobservations are reported. In contrast tothe limited number of reports on thefundaxnental analysis of androgenesis,there is vexy much empirical informationabout factors that influence the numberof plants produced through androgenesis.Information about such factors isoftensaid to be derived from “trial anderror” experiments. An oveMew of themethodology and such factors is describedbelow for the model spedes barley.
arky androgenesis in practice
Impact of methodThere are three different androgenesismethods to produce microspore-derived(doubled) haploids in barley. Haploidizaüon can be achieved by culture ofwhole anthers, what is called antherculture. The male gametes can bemechanically isolated from the anthers,what is called (isolated) mkrosporeculture. ALtematively, the microsporescan be isolated naturally from dehisdnganthers, what is cafled shed culture(Sunderland and Xu, 1982; Ziauddin etal., 1990). Comparison betweenmÎcrospore and shed culture shows thatshed culture is difficult to reproduce, asthe period required for shedding and theshed microspore density varies, due to thephysiological state of the donor plant atthe time of hanrest and the microsporedevelopmental stage. In microsporeculture on the contrasy, such variationscan be adjusted directly after isolation (atthe end of this paragraph, this statement
will be explained). Comparison betweenanther and microspore culture revealsthat former is easier to perform. Thesimplidty of performing anther cultureshas resulted in a strategy of culturing highnumbers of anthers rather than toincrease the effidency of anther culture.In many spedes the anther wall providesa physical and chemical environment formicrospore embryogenesis to a sufficientdegree in many spedes. However, theanther wall provides also inhibitosysubstances (Heberle-Bors, 1984) andbadly-defined nutrients (Sunderland andXu, 1982) for sporophytic microsporedevelopment. The technique of antherculture suffers from a large disadvantage,i.e. plants may originate not only fromthe microspores, but also from variousparts of the anther resuking in a mixedpopulation of plants derMng both fromgametic and somatic origin. Applicationof microspore or shed cukure preventsproduction of anther-derived somaticregenerants. Besides, microspore culture isfive times more effident for plantproduction than anther culture (chapter2). In the case of anther culture, it isextremely difficult to follow thedevelopment of the mkrospores withinthe anther and in general the cultureeffidency can not be assessed reiablyuntil 4 weeks after culture inïfiation. Theefficiency of the development into plantsis largely influenced by the densitythroughout culture. A significant advantage using microspore culture, is thepossibility to follow the growth of themicrospores under a light microscope anddetermine growth indices throughoutcufture. Further, in microspore culture the
19
Chapter 1
culture density can be optimized at
various time pointsduring culture. There
are two indices that can be used to
control the density during cukure,
starting from the day of microspore
isolation. Directly after pretreatment the
percentage of embryogenic microspores
(i.e. a subpopu1ation that is assumed to
develop into plant; Bolik and Koop,
1991; Otsen, 1991; chapter 4 and 7) can
be recognized. further, the densïty of
embryo-like structures (ELS, i.e. multi
cellular structures where the exine has
been disrupted; these can have the
morphology of a zygotic embryo) can lie
determined. Adjustment of both embryo
genic microspore and ELS density
significantly improves the reproducibility
of the culture system for green plant
development (chapter 4; Hoebtra et al.,
1994).The aim of androgenesis is stimulation
of microspores to form embryos, maybe
after an intennediate phase of dedif
ferentiated growth, and subsequently
plants. Conditions of culture, like culture
density, together with genetic determinants, affect the microspores in their
new developmental pathway. Identi
fication of such factors is of utmost
importance for the establishment of a
reprodudbie and effident androgenesis
protocol.
Impact of plant growth and culture conditions
The following contains an oveMew offactors of different natures whîch arereported to affect embryo fonnation in
anther or microspore cukures in barley,
and their possible modes of action. A
more detailed oveMew of cereal
microspore cukure is written by Jiihne
and Lörz (1995).The quality of the donor material such
as its vigour, has a decisive influence on
the success of androgenesis. Pest control
procedures have a detrimental effect on
microspore deveîopment (Jahne et al.,
1991). Important environmental factors
which influence the vigour of donor
plants inciude light intensity and
spectrum, photoperiod, temperature, and
nutrition. For barley high light intensity
(20,000 lux) and low temperature (12°C)
are favourable for androgenesis (foroughi
Wehr and Mix, 1979). Low temperature
increases the time of development,
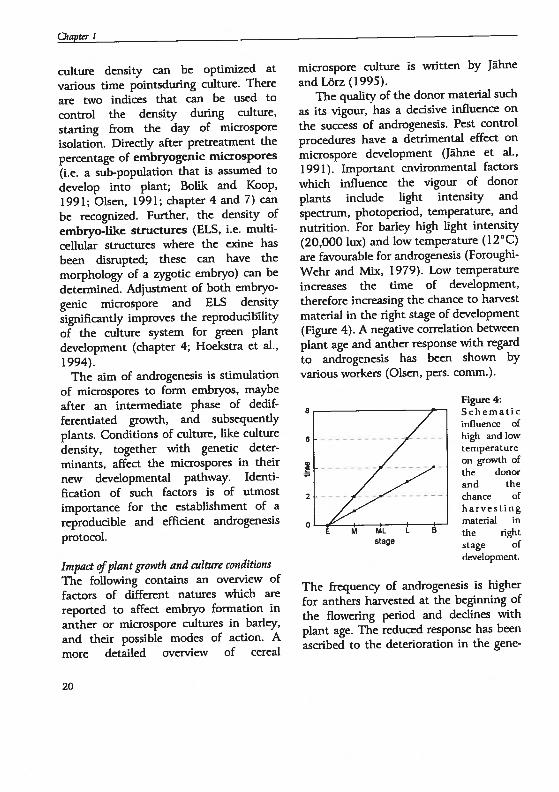
therefore increasing the chance to harvest
material in the right stage of development
(figure 4). A negative correlation between
plant age and anther response with regard
to androgenesis has been shown by
various workers (Olsen, pers. comm.).
È M P1L
stage
Figure 4:Schematicinfluence ofhigh and towtemperature0fl growth ofthe donorand thechance ofh arve stingmaterial inthe rightstage ofdevelopment.
The frequency of androgenesis is higher
for anthers harvested at the beginning of
the flowering period and declines with
plant age. The reduced response has been
ascribed to the deterioration in the gene
6
2
0
20

General Infroduction
rai condifion of the plants, particularlyduring seed set. Spikes harvested at theverge of mitosis are most responsive (Gaulet al., 1976; Wheatley et al., 1986;chapter 2). Variation in stage per spikeand spike number is found (Dunweil etal., 1987). The microspore developmentalstage is a precondftion for induction ofandrogenesis, as outlined earlier.Another precondition for gametophyticembryogenesis is applicaüon of apretreatment. Two pretreatment methodsare generally used for haploidization inbarley. Either a cold pretreatment of thespike for 3-5 weeks (Huang andSunderland, 1982) or an antherpretreatment on a mannitol containingsolution for 4 days (Roberts-Oeffischlagerand Dunweil, 1990). Preference ofmannitol pretreatment over cold periodhas been mentioned (Hoekstra et al.,1994). During pretreatment embryogenicmicrospores (chapter 4) develop, and amechanism of action is proposed inchapter 7.
The number and percentage ofembryogenic microspores obtained, islargely influenced by the method ofmicrospore isolaüon (Hoekstra et al.,1994). Methods have been described inwhich pestie maceration (chapter 3 and4), microblending (Olsen, 1991) orvortexing (Hu et al., 1995) of the anthersis used. However, observations in ourlaboratory indicated that gentie treatmentof the material is necessaiy for obtaininga good population of embryogenicmicrospores and that was, in our hands,only possibie with pestie maceraüon.
The plating effidency is influenced bythe culture medium applied. Major
improvements of medium compositionare cited. It has been shown that nitrogencomposition of the culture medium playsa significant role in androgenesis. Areduced ammonium nitrate concentrationtogether with additional glutamine as anontoxic nitrogen source, is generallyused (Olsen, 1987; Mordhorst and Lörz,1993). The importance of thecarbohydrate and its concentration resp.osmolarity is shown by finnie et al.(1989), Cistué et al. (1995) and inchapter 4. Hunter (1987) demonstratesthe benefit of giucose-based saccharides;in general, the traditional sucrose isreplaced by maltose which is degraded ata slower rate. Scott and Lyne (1994)propose that metabolization of sucroseand glucose leads to accumulation of atoxic product resulting in ceil death,whereas maltose can sustain developmentof embiyos.
A third and last precondition forinduction of androgenesis is the presenceof a growth regulator in the culturemedium. In general 4 M benzylaminopurine (BAP), sometimes combinedwith a low amount of indolacetic add(IAA), is used (Olsen, 1987; Jiihne et al.,1991; Kuhlmann and Foroughi-Wehr,1989). In chapter 5 the application of asynthetic auxin as growth regulator ispresented, which is generally used toinduce somatic embryogenesis.
A strilting phenomenon in optimizingthe physical conditions of anther cultureis, that anthers orientated in the “up”position (i.e. with one lobe in contactwith the medium) produce more greenplants (Poweil et al., 1988; Hunter, 1985;Shannon et al., 1985). An explanation for
21

Chapter 1
inferior plant effidency of anthers in the“flat’ position (with two lobes in contactwith the medium) is that they generallyshed the microspores on the solidmedium, which prevents furtherdevelopment of the sporophytic route.The shedding of microspores from anthersin the “flat” position during pretreatmenton a mannitol containing solution, on thecontrary, is positive (Roberts-Oeffischlager and Dunweil, 1990). Theseauthors further find a posidve effect onplant production if the anther density isreduced to 20 anthers; if 60 anthers areplated per dish, the number of embiyosper responding anther doubled. Köfflerand Wenzel (1985), however, prefer anincreased plating density of the anthers(40 in stead of 10 per ml). The density ofas well anthers or microspores as ELS isveiy important enabling developmentwithout an intervening dedifferenüadonphase (Datta and Potrykus, 1989; chapter4). The fonner density should berelatively high, whereas optimal embxyodevelopment requires strong ditudon.These data explain contradictory resuitsin the literature.
Spedflcally for barley, no chromosomedoubling technique is required to obtaindiploid plants after androgenesis, as about80% of the plants obtained havespontaneously doubled chromosomenumbers (see the paragraph androgenesisin model systems). The importance ofculture condiüons is demonstrated in theratio green to albino ptants, andoptimization resulted in a decrease from1:1 to 34:1 (chapter3).
plications of androgenesis
Plant breeding of barteyMethods for haploid plant production aresummarized above. In barley, haploidscan be relatively easy obtained in vivo viagenome eimination in crossings withHordeum bulbosum. Comparisons betweenthe Hordeum bulbosum method andandrogenesis have been performed.Bjömstad et al. (1993) find noconsistently negaüve impact of anther
culture in barley, and although notidentical, the methods may be consideredequivalent. Devaux (1991) describes thatandrogenesis enables doubled haploids tobe obtained from genotypes whichrespond poorly to the H. bulbosummethod. Thus, the two techniques can beused parallel to each other to improvedoubled haploid production effidency.Foroughi-Wehr and Wenzel (1989)observe that even in species whereparthenogenetic techniques had someimportance, as barley, microsporeapproaches are overtaldng.
For madmaI incorporation ofandrogenesis-derived doubled haploidsinto breeding programs, the followingcriteria should be fulfilled:
Production of targe numbers of doubledhaploids of alt genotypes should be easyand consistent, the plants shoutd lie
genetkatly nonnat and stabte and thepopulation should con tam a randomsampit of the meiotkatly recombinedgametes.
On basis of these criteria for ultimateapplicaüon of doubled haploids inbreeding, a description of advantages and
22

General Introduction
disadvantages of androgenesis is made.The major problem is that the method isvery genotype dependent, implicatingthat only from certain genotypes Jargenumbers of doubled haploids can begenerated (e.g. Maheshwari et al., 1980;foroughi- Wehr et al., 1982; Knudsen etal., 1989; Larsen et al., 1991). Anotherproblem is the unpredictability of plantproduction effidency per genotype,attrlbuting to low and fluctuatingfrequencies (e.g. Sunderland et al., 1981).for the crop barley in general, about 80%of the regenerants are spontaneousdoubled haploids (e.g. Islam et al., 1992).further, in barley there is very littieevidence of induced genetic changes. Thelevel of instability is insïgnïfcant whencompared with that which is observedfollowing meiotic recombination (finnieet al., 1991). In other crops e.g. wheatthe instable geneuc background can berecognized in the regenerants. Especiallyin cereals formation of chlorophyll deficient plants is observed due to deletionsof the plastid genome (Harada et al.,1991). Significant deviations from theexpected Mendelian ratios are observede.g. by Thompson et al. (1991) for four ofthe ten markers studied. On the otherhand, in responsive genotypes, thousandsof homozygous plants can be obtainedwithin about 3 months, and save years ofbackcrossing or seffing. Resistancesagainst nematode, viral and fungalinfection have been found amongregenerants from anther culture (e.g.Wenzel and Uhrig, 1981; foroughi-Wehrand friedt, 1984; Kintzios et al., 1994).Moreover, homogeneous parent materialcan be generated for f1 hybrid
production. Reduction of the time neededfor production of a new variety uponapplication of androgenesis is limited, asregenerants should 5e screened underfield conditions for several years. Thegenuine advantages of application ofmicrospore-derived plants are qualityimprovement by generaüon of newvariability, fixation of traits (Boppenmeler et al., 1989), superior material forgene mapping, and detenninaüon of thegeneücs of multiple disease resistance(Steffenson et al., 1995). Selection ofimproved varieües can be enhanced ifandrogenesis is combined with in vitro
selection or genetic engineering.
In vitro seÏection and genetic modflaftionAndrogenesis can be combined with invitro techniques, in order to enhance thechance on selection of the desiredgenotype. Espedally when microsporeculture is combined with such techniques,the advantage of treating haploid singleceils is favourable. Below, the mostimportant methods for combination withandrogenesîs are described.
In vitro selection is a low cost, andattractive technique for traits expressed atthe ceil biological level. This method isbeing combined with microspore culturefor selection of various traits, like low pHand heavy metal tolerance (Karsai et al.,1994). Via in vitro selection of androgenesis-derived tissue, salt tolerance hasbeen selected in rice (KrishnaRaj andSreeRangasamy, 1993), freezing tolerancein Brassica napus (Orr et al., 1990), anddisease tolerance in wheat (fadel andWenzel, 1993). In addition, Touraev etal. (1995) show that in vitro selection can

Ozapter 1
5e applied on mid-binucleate pollenduring in vitro maturation, and thereforedetennine that gametophytic selection isfeasible for enhanced transmission ofgenes to the next sporophytic generation.
Moreover, induction of genetic changeshas been performed in rapeseed in anindirect way using gamma-ray mutation(Polsoni et al., 1988). Nowadays atremendous interest is shown in moredirected, targeted methods for changes inthe (haploid) genefic background, usinggenetic transformation. for the genera
tion of stably transfonned plants, ingeneral five transfomtation methods areused. In dicotyledonous spedes themethod of DNA transfer is generallymediated by Agrobacteriurn tumefaciens. Inthe monocotyledonous species rice, coc&tivation of seed derived caflus withAgrohacteriurn tumefaciens resulted onlyrecently in stable integration of foreignDNA (Hiei et al., 1994). The othermethods for DNA transfer are electroporation, microinjection, PEG-mediateduptake and particle bombardment. An
oveMew ofthe methodsused fortransformationofuni-orbinucleatemicrospores ispresented inTable 1.There is onlyone reportfrom 1989available onAgrobacteriummediatedDNA transferto microsporesof rapeseed(Pechan); thishas proven tobe non-reprodudble.further, sofaronly transientexpression hasbeen obtainedin microsporesafter electro-
Table 1: Overview of transformaUon methods used on Ufli- or binudeate micro
spores as target tissue, the result and their reference. S = stably transfomied, T =
transient expression.
References Spedes Method
Fennel and Hauptmann, 1992 Zea rnays electroporauon T
Jardinaud et al., 1995 Zea rnays electroporation T
Joersbo et al., 1990 Hordeurn vulgare electroporation T
Olsen, 1991 Hordeurn vulgare microinjection T
Bolik and Koop, 1991 Hordeurn vulgare rnicroinjection T
Gaillard et al., 1992 Zea rnays microinjection T
Jones-ViHeneuve et al., 1995 Brassica napus microinjection T
Kuhlmann et al., 1991 Hordeurn vulgare PEG T
fennet and Hauptmann, 1992 Zea mays PEG T
Stöger et al., 1992 Nicotiana tabacum bombardment T
Jâhne et al., 1994 Hordeurn indgare bombarilment S
Kasha ei al., in press Hordeurn vutgare bombardment S
Stöger et al., 1995 Nicotiana tabacurn bombardment S
Nishihara et al., 1995 Nicotiana rustica bombardment S
Toureav et al. pers. comm. Nicohana tabacuni bombardment S
24

General Introduction
poration, microinjection and PEGmediated DNA uptake. for transformation of microspores, partidebombardment has recently proven to be asuitable method. A system for thebiolistic transfonnation of barley usingfreshly isolated microspores as the targettissue has been developed. Independenttransformaüon events led, on average, tothe reovery of 1 plant per 1 omicrospores. The transferred genes wereinherited in all progeny plants indicatingthe homozygous nature of primalyregenerants (Jhne, et al., 1994). Thismethod is repeated and similar resuitswith the same spedes are obtained(Kasha et al., in press). Recently, by usin$this approach, trangenic plants areproduced from Nicotiana sp. (Stöger etal., 1995; Nishihara et al., 1995).
In addiüon, if microspores are used astarget tissue in genetic transformation,the gametophytic developmental routecan be completed in stead of thesporophytic pathway. DNA transfer tomicrospores by partide bombardmentand subsequently in vitro maturation ofthese microspores generates maturetransgenic pollen (Akven et al., 1990).Via this elegant approach, laborious andtime consuming steps of tissue ctilture canbe omitted for the production oftransgenic plants. Recently stableinheritance of the transgene has beenachieved through bombardment oftobacco microspores, in vitro selectionduring pollen germination, and in vivofertilization; the time required frommicrospore isotation until stableexpression of the transgene is tess than 3weeks (Touraev et al., pers. comm.).
I\im of the thesis
Plant production through androgenesishas many attractive characteristics, whichare outlined in detail above. Generationof (doubled) haploid plants is of utmostimportance for breeding of improvedplant varieties. Selection of the desiredtraits can be further accelerated by in vitroselection or transformation of the haploidceils. In many spedes (doubled) haploidplants have been produced using antheror microspore culture. However, largedifferences in regeneration frequendesbetween species and genotypes, preventgeneral applicabïlity of the method. Evenin model spedes, orily one or a fewgenotypes, e.g. cv. Igri in barley and cv.Topas in rapeseed, are used infundamental studies because of theirexcellent culture effidency. As mentionedearlier, there is some empirical knowledgeavailable about factors that influence theresponse and regeneration effidendes ofmicrospores. One of them is thephysiological state of the donor materiai.Accumulation of fundamental knowledgeabout the switch from the gametophyticpathway to the sporophytic pathway isstili very limited. However, it has notbeen possible sofar, to modify cultureeffïciency in such a way that all cultivarsproduce microspore-derived green plants.
In this thesis, preconditions for theproduction of microspore-derived plantsare described using donor plants grown ina well condiüoned environment. Determination of such factors will enableimprovement of the reprodudbility of themethod. The aim of the research is to
25

Ozapter 1
identify both “direct-breeding-applicable”possibilities and more fundamentalfactors, in order to unravel part of thecomplicated mechanism necessaiy for theswitch from the gametophytic to thesporophytic pathway in barley microspores.
R.Mwen A., N. Eller, M. Kastler, RM.B. Moreno
and E. Heberte-Bors. Potential of in vitro potten
maturation for gene transfer. Physiol. Plant. 79:194-196 (1990)Bajaj Y.P.S. In vitro production of haploids andtheir use in ceil geneücs and plant breeding. In:Biotechno1o’ in agriculture and forestry vol. 12Haploids in crop improvement 1, ed. Bajaj Y.P.S.,Springer Verlag Brlin Heidelberg pp. 1-44 (1990)Bedinger P. The remarkable biology of pollen.
Plant CetI 4: 879-887 (1992)Binarova P., K Straatman, B. Hause, G. Hauseand A.A.M. VanLammeren. Nudear DNAsynthesis during the inducfion of embryogenesis incultured microspores and pollen of Bra.uica napusL. Theor.Appt. Genet. 87: 9-16 (1993)B’örnstad A., H. Sldnnes and K Thoresen.Comparisons between doubled haploid linesproduced by anther culture, the Hordeum bulbosummethod and lines produced by single seed descentin barley crosses. Euphytica 66: 135-144 (1993)Bold C.B., C. Alexopoulos, and T. Delevoras(eds.). Morphology of plants and fungi. Harperand Row Publishers, Neiv York pp. 570 (1980)Botllc M. and H.U. Koop, Identificafion ofembryogenic microspores of barley (Hordeumvutgare L) by individual selection and culture andtheir potential for transformation by microinjection. Protoplasma 162, 61-68 (1991)Boppenmeier J., S. Zuechner and B. foroughiWehr. Haploid production from barley yellowdwarf virus resistant dones of Lolium. Plant Breed.103: 216-220 (1989)Cao M.Q., Y. Li, F. flu, T. Jiang and G.S. Liu.Application of anther culture and isolatedmicrospore culture to vegetable crop improvement.Acta Hort. 392:27-38 (1995)
Cistué L., A. Ziauddin, E. Simion and ICJ.
I(asha. Effects of culture conditions on isolated
microspore response of barley cultivar Igri. Plant
Cdl Tiss. Org. Cult. 42: 163-169 (1995)Cordewener J.H.G., G. Hause, E. Goergen, RBusink, B. Hause, J.J.M. Dons, A.A.M.
VanLammeren, M.M. VanLookeren Campagne
and P. Pechan. Changes in synthesis andlocalization of members of the 70-kDa dass of
beat shock proteins accompany the induction of
embryogenesis in Brassica napus L. microspores.
Planta 196: 747-755 (1995)Custers J.B.M., J.H.G. Cordewener, Y. Noeflen,
J.J.M. Dons and M.M. VanLookeren
Campagne. Temperature controls both
gametophytic and sporophytic development in
microspore ctilturea of Brassica napus. Plant Celt
Rep. 13: 267-271 (1994)Datta S.K. and 1. Potrykus. Artificial seeds inbarley encapsulation of microspore-derived
embryos. Theor. Appi. Genet. 77: 820-824 (1989)Devaux P. Investigations to improve doubled
haploid production effidency in a winter barleybreeding programme. Cerea] Research Comm 19:51-58 (1991)Dunweil J.M. and N. Sunderland. Pollenultrastructure in anther cultures of Nicotianatabacum II. changes a.ssodated with embryogenesis.
J. Exp. Bot. 25: 363-373 (1974a)Dunweil J.M. and N. Sunderland. Pollenultrastructure in anther culttsres of Nicotiana
tabacum 111. The first sporophytic division. J. Exp.Bot. 26: 240-252 (1974b)Dunweil J.M., RJ. Francis and W. Poweil.Anther culture of Hordeum wigare L. Theor. Appl.Genet 74: 60-64 (1987)fadel F. and G. Wenzel. In vitro selecüon fortolerance to fusariurn in f1 microspore populations
of wheat. Plant Breed. 110: 89-95 (1993)Fennel A. and R Hauptmann. Etectroporationand PEG delivery of DNA into maize microspores.
Plant Celi Rep. 11:567-570(1992)Finnie S.J., W. Poweli and A.F. Dyer. The effect
of carbohydrate composition and concentration onanther culture response in barley (Hordeum vulgareL).PlantBreed. 103: 110.118(1989)finnie S.J., B.P. Foster and ICJ. Chalmers.Genetic stability of microspore-derived doubledhaploids of barley a cytological, biochemical, andmolecular study. Genome 34: 923.928 (1991)
26

General Introduction
Foroughi.Wehr B. and W. Friedt Rapid
producfion of recombinant barley yellow mosaic
virus resistant Hordeum vulgare lines by anther
culture. Theor. Appi. Genet. 67: 377-382 (1984)
foroughiWehr 3., W. Friedt and G. Wenzel.
On the geneuc improvement of androgenetic
haploid formation in Hordeum vutgare L. Theor.
AppI. Genec 62: 233-239 (1982)Foroughi.Wehr 3. and G. Mix. In vitro response
of Hordeum vulgare 1. anthers cultured from
plants grown under different environments. Env.
Exp. Bot. 19: 303-309 (1979)foroughi.Wehr B. and G. Wenzel. Androgeneüc
haploid production. IAPTC Newsletter May 11-18
(1989)GafHard A., E. MatthysRochon and C. Dumas.
Selection of microspore derived embryogenic
stmctures in maize related to transformation
potential by microinjection. Bot. Acta 105: 313-
318 (1992)Garrido D., N. EHer, E. Heberle-Bors and 0.
Vicente. De nova transcripfion of sperific mRNAs
during the induction of tobacco pollen
embsyogenesis. Sex. Plant Reprod. 6: 4045 (1993)Garrido D., 0. Vicente, E. Heberle-Bors and
M.LRodriguez-Garcia. Cellular changes during
the acquisiüon of embryogenic potential inisolated pollen grains of Nüoüana tabacum.
Protoplasma 186: 220-230 (1995)Gaul H., G. Mix, B. foroughi.Wehr and M.
Okamoto. Pollen gram development of Hordeumvulgare. Zeitschrift für Pflanzenzüchtung 76: 77-80
(1976)Goldberg B., T.P. Beals and P.M. Sanders.
Anther development: basic prindples and practical
applicaüons. Plant CelI 5: 1217-1229 (1993)
Goldberg 3., P.M. Sanders and T.P. Beals. A
noveli cell-ablafion strategy for studying plant
development. Phil. Trans. R Soc. Land. 3 350: 5-17(1995)Harada T., T. Sato, D. Asaka and 1.Matsukawa. Large-scale deletions of nce plastid
DNA in anther culture. Theor. Appi. Genet. 81:
157-161 (1991)Hause 3., G. Hause, P. Pechan and A.A.M.VanLammeren. Cytoskeletal changes and
induction of embryogenesis in microspore and
pollen cultures of Brassica napus L. CelI Biol. Int.
17: 153-168 (1993)
Heberle-Bors E. Genotypic control of pollen plantformation in Nicotiana t4bacum L. Theor. Appi
Genet. 68: 475479 (1984)Heberle-Bors E. In vitro haploid formalion from
pollen: a critical review. Theor. Appi. Genet. 71:
361-374 (1985)Heberle-Bors 3. Isolated pollen culture in
tobacco: plant reproducuve development in anutshell. Sex. Plant Reprod. 2: 1-10 (1989)
Hiei Y., S. Ohta, T. Komari and T. Kumashiro.
Effictent transformation of nce (Oryza sativa L)
mediated byAgrobacterium and sequence analysis of
the boundaries of the T-DNA. Plant J. 6: 27 1-282
(1994)Hoekstra S. M.H. VanZijdenreld, S.
VanBergen, f. VanderMark and F. Hefdekamp.
Genetic modification of barley for end use quality.In: lmprovement of cereal quality by genetic
engineering, Herny RJ., and JA Ronaids (eds.),Pl enum Press, New York. pp. 139-144 (1994)
Hu T.C., A. Ziauddin, E. Shnion and KJ.Kasha. Isolated microspore culture of wheat(Triticum agstivum L.) in a defined media 1. Effectsof pretreatment. isolation methods, and hormones.InVitroCellDev. Biol. 31: 79-83 (1995)Huang 3. and N. Sunderland. Temperature.stress in barley anther culture. Ann Bot 49: 77-88(1982)Hunter C.P. The effect of anther orientation onthe production of miaospore.derived embsyoids
and plaats of Hordeum mdgare cv. Sabarlis. PlantCeIl Rap. 4: 267-268 (1985)Hunter C.F. Plant generation method. Europeanpatent applicaüon number 872007 73.7,publication number 0245 898 (1987)Islam M.R, S. IGntzios and G. fischbeck.Anther ctilture responsiveness of Hardcumspontaneum derived spring baxley lines and agenetic analysis of plant regeneration. Plant CeilTiss. Org. Ctilt. 29: 235-239 (1992)Jâhne, A., D. Becker, R. Brettschneider and H.Lörz. Regeneration of transgenic. microspore
derived. fertile barley. Theor. Appl. Genet. 89:525-533 (1994)Jhne, A., P.A. Lazzeri, M. Jâger.Gussen and
H. Lörz. Plant regeneration from embsyogenic celisuspension derived from anther cultures of barley(Hordeum vulgare L.). Theor. Appi. Genet. 82: 74-
80(1991)
27

Ojapter 1
Jâhne A. and H Lôrz. Cereal microspore culture.PlantSd. 109: 1-12(1995)Jardinaud M.F., A. Souvre, M. Beckert and G.Mibert. OpUmisation of DNA ixansfer andtransient .glucuronidase expression inelectroporated maize (Zea mays L) microspores.Plant Ccli Rep. 15: 55.58 (1995)Joersbo M., RB. Jørgensen and P. Olesen.Transient eletropermeabitization of barley(Hordeum vulgare 1.) microspores to propidiumiodide. Plant Ceil Tiss. Org. Cult. 23: 125-129(1990)Jones-Villeneuve E., B. Huang, 1. Prudhomme,S. Bird, R ICembie, J. Hattori and 3. Miki.Assessment of microinjection for introdudng DNAinto uninuclear microspores of rapeseed. Plant CdlTiss. Org. Cult. 40: 97-100 (1995)Karsal T., Z. Bedoe, G. ICovks and 8.BarnaMs. The effect of in vivo and in vitroaluminium treatment on anther culture response oftriticale x wheat hybrids. J. Gen. Breed. 48: 353-358 (1994)Kasha et al. in pressKintzios S., RM. Islam and G. Fisckbeck.Distorted segregation for mildew resistance indoubled haploid lines of spring barley. PlantBreed. 112:248-251(1994)Knudsen S., LK. Due and S.B. Andersen.Components of response in barley anther culture.Plant Breed. 103: 24 1-246 (1989)Köhler F. And G. Wenzel. Regeneration ofisolated barley microspores in conditioned mediaand trials to characterize the responsible factor. J.Plant Physiol. 121: 181-191 (1985)l(rishnaRaj S. and S.R. SreeRangasamy. In vitrosalt tolerance screening in long-term anthercultures of nee (Oryza 5ativa L.) variety 1R50. J.Plant Physiol. 142: 754-758 (1993)Kuhlmann U. and 3. foroughi-Wehr.Production of doubled haploid lines in frequendessuffident for barley breeding programs. Plant CdlRep. 8: 78-81 (1989)Kuhlmann U., 3. ForoughiWehr, A. Granerand G. Wenzel. Improved culture system formicrospores of barley to become a target for DNAuptake. Plant Breeding 107: 165-168 (1991)Kyo M. and H. Harada. Phosphorylaion ofproteins associated wfth embryogenicdedifferenilation of immature pollen grains ofNicotiana rustica. J. Plant Phys. 136: 716-722
(1990a)Kyo M. and H. Harada. Spedficphosphoproteins in the initial period of tobaccopollen embryogenesis. Planta 182: 58-63 (1990b)Larsen EI., I.K.D. Tuvesson and 5.3.Andersen. Nudear genes affecting percentage ofgreen plants in barley (Hordeum vulgare L.) antherculture. Theor. AppI. Genec 82: 417-420 (1991)Lee F.M. and C.C. Chen. Nudear fusion incultured microspores of barley. Plant Cdl Rep.6:191-193 (1987)Maheshwari S.C., A.K Tyagi and K Malhotra.Induction of haploidy from pollen grains inangiosperms - the current status. Theor. Appl.Genet. 58: 193-206 (1980)Mascarenhas J.P. The male gametophyte ofilowening plants. Plant Ccli 1: 657-664 (1989)Mascarenhas J.P. Gene activity during pollendevelopment. Anno. Rev. Plant Physiol. Plant Mol.Biol. 41:317-338 (1990)McConsick S. Male gametophyte develop-ment.Plant Cdl 5: 1265-1275 (1993)Meinke D.W. Perspectives on genetic analysis ofplant embryogenesis. Plant Cdl 3: 857-866 (1991)Mordhorst A.P. and H. Lörz. Embxyogenesis anddevelopment of isolated barley (Hordeurn vulgare L.)microspores are influenced by the amount andcomposition of nitrogen sources in culture media.
J. Plant Physiol. 142: 485-492 (1993)Morrison RA. and D.A. Evans. Haploid plantsfrom tissue culture: new plant varieties in ashortened time frame. Biotechnology 6: 684-690(1988)NagI W. Cdc2-kinases, cydins, and the switchfrom proliferaion to polyploidizaüon.Protoplasma 188: 143-150 (1995)Nishihara M., M. Seki, M. Kyo, IC frifime andH. Morikawa Transgenic haploid plants ofNicotiana nstica produced by bombardmentmediated transformation of pollen. Transg. Res. 4:341-348 (1995)Olsen F.L. Induction of micropsore embryogenesis in cultured anthers of Hordeum vulgare. Theeffects of ammonium nitrate, glutamine andasparagine as nitorgen sources. Carlsberg Res.Commun. 52: 393-404 (1987)Olsen f.1. Isolaüon and cultivation ofembryogenic microspores from barley (Hordeumvulgare L.). Hereditas 115:255-266 (1991)Orr. W., A.M. Johnson-Flanagan, W.A. Keller
28

Geiierat Introduction
and J. Singh. Induction of freezing tolerance in
microspore-derived embryos of winter Brassica
napus. Plant Celi Rep. 8:579-581(1990)
Pechan P.M. Successful cocultivaUon of Brassica
napus microspores and proembxyos with
Agrobacfrrium. Plant Celi Rap. 8: 387-390 (1989)
Pechan P.M., D. Bartels, D.C.W. Brown and J.Scheil. Messenger-RNA and protein changes
assodated wiffi induction of Brassica microspore
embryogenesis. Planta 184: 161-165 (1991)
Polsoni L., L.S. Kott and W.D. Beversdorf.
Large-scale microspore culture technique for
mutation-selection studies in Brassica napus. Can, J.Bot. 66: 1681-1685 (1988)Poweil W., F.M. Borrino and V. Goodail: The
effect of anther orientation on microspore-derived
plant production in barley (Hordeum vulgare L).Euphytica38: 159-163 (1988)Reynolds J.F. In vitro culture of vegetable crops.
In: Plant celi and tissue culture, Vasil IK, Thorpe
T.A. eds., Kuwer Academic Publishers, Dordrecht
pp. 331-362 (1994)RobertsOeMschIager SL. and J.M Dunweil.
Barley anther culture: pretxeatmeflt 0fl manflitol
stimulates production of microspore-derived
embryos. Plant Celi, lSss. Org. Cult. 20: 235-240
(1990)Scott R., R Hodge, W. Paul and J. Draper. Themolecular biology of anther differentiation. Plant
Sd. 80: 167-191 (1991)Scott P. and RL. Lyne. Inifiatiofi of
embryogenesis from cultured barley microspores
a furter investigaüon into the toxic effects ofsucrose and glucose. Plant Ceil Tiss. Org. CulC 37:
61-65 (1994)Shannon P.RM., A.E. Nicholson, J.M.Dunweft and D.R. Davies, Effect of anther
orientation on microspore-callus production in
barley (Hordeum vulgare 1.). Plant CeIl Ttss. Org.
CuJC 4:271-280(1985)Sheridan W.f. Genes and embryo morphogenesis
1fl angiosperms. Developm. Genet. 16: 291-297
(1995)Steffenson B.J., Y. Jin, B.G. Rossnagel, J.B.
Rasmussen and IC Kao. Geneücs of multipledisease resistance in a doubled-haploid population
of barley. Plant Breed. 114: 50-54 (1995)Stôger E., RM. Benito Moreno, 8. Ylstra, 0.
Vicente and E. Heberle-Bors. Comparison of
different techniques for gene transfer into mature
and immature tobacco pollen. Transg. Res. 1: 71-
78(1992)Stöger E., C. fink, M. Pfosser and E. Heberle
Bors. Plant transfonnatiofl by partide
bombardment of embryogenic pollen. Plant Celi
Rap. 14: 273-278 (1995)Sunderland N. and L.J. Evans. Multicellular
pollen formation in cultured barley anthers II. The
A, B, and C pathways. J. Ficp. Bot. 31: 501-514
(1980)Sunderland N., 8. Huang and G.J. Huis.
Disposition of pollen in situ and its relevance toanther/pollen culture. J. Exp. Bot. 35: 521-530
(1984)Sunderland N., M. Roberts, LJ. Evans and
D.C. Wildon. Multicellular pollen formatiofl in
cultured barley anthers 1. Independent division ofthe generative and vegetative celis. J. Exp. Bot. 30:1133-1144 (1979)Sunderland N. and Z.H. Xu. Shed pollen culture
in Hordeum vulgare. 1. Exp. Bot. 33: 1086-1095
(1982)Sunderland N., Z.H. Xu and 8. Huang. Recent
advances in barley anther culture. In: Barleygenetics IV, pp. 699-703 (1981)Telmer C.A., W. Newcomb and D.H.Simmonds. Cellular changes during heat shockinduction and embryo development of culturedmicrospores of Bra5sica napus cv. Topas.
Protoplasma 185: 106-112 (1995)Telmer C.A., D.H. Simmonds and W.Newcomb. Determination of developomentalstage to obtain high frequendes of embryogenic
microspores in Brasska napus. Phys. Plant. 84: 417-
424 (1992)Thompson D.M., K Chalmers, R Waugh, 3.P.foster, W.T.B Thomas, P.D.S Caligari and W.Poweil. The inheritance of genetic markers inmicrospore-derived plants of barley Hordcumvulgare L Theor. Appi. Genet. 81: 48 7-492 (1991)Touraev A., C.S. Pink, E. Stöger and E.Heberle-Bors. Pollen selecion: a transgenic
reconstruction approach. Proc. NatI. Acad. Sd.USA92: 12165-12169 (1995)Toureav A., A. ilhain, 0. Vicente and E.Heberle-Bors. Stress-induced microsporeembtyogenesis in tobacco: an optimized system formolecular studies. Plant CeIl Rep. 15: 561-565
29

Ozapter 1
(1996)Touraev. A., M. Pfosser, 0. Vicente and E.Heberle-Bors. Stress as the major signalcontrofling the developmental fate of tobaccomiaospores: towards a unified model of inducionof microspore/pollen embryogenesis. Ptanta, inpress.Veilleux RE. Development of new cultivars viaanther culture. Hart. Sci. 29; 1238-1240 (1994)Vicente 0., RM. Benito Moreno and E.Heberle-Bors. Pollen cultures as a tool to studyplant developmenL Ceil Biol. Rev. 24: 295-305(1991)Wenzel G. and H. tlhrig. Breeding for nematodeand virus resistance in potato via anther culture.Theor. Appi. Genet. 59: 333-340 (1981)Wheatley W.G., A.A. Marsolais and KJ.Kasha. Microspore growth and anther staging inbarley anther culture. Plant Celi Reports 5: 47-49(1986)
Zaki M.A.M. and H.G. Diddnson. Structuralchanges during the first division of embiyosresulting from anther and free microspore culture
in Brassica napus. Protoplasma 156: 149-162(1990)Zarsky V., D. Garrklo, L. Rihova, J. Tupy, 0.Vicente and E. Heberle-Bors. Derepression ofthe celi cycle by starvation is involved in theinduction of tobacco pollen embsyogenesis. Sex.Plant Reprod. 5: 189-194 (1992)Zarsky V., D. Garrido, N. Eller, J. Tupy, 0.Vicente, F. Schöffl and E. Heberle-Bors. Theexpression of a small heat shock gene is activatedduring induction of tobacco pollen embryogenesisbystarvaüon. Plant Celi Env. 18: 139-147 (1995)Ziauddm A., E. Simion, and KJ. Kasha.Improved plant regenerafion from shed microsporectilture in barley (Hordeum vutgare L) cv. Igri. PlantCeil Rap. 9: 69-72 (1990)
30

hapter 2
Anther and microspore cukure ofHordeum vutgare L. cv. Igri
S. Hoeksfra, M.H. van Zijderveld, J.D. Louwerse, f. Heidekamp & F. van der Mark
SurnrnaryThe iniluence of environmental and cultural changes was investigated on both anther culture and microspore ctilture of barley cv. Igri. The highest regeneration frequency for both culture systems was obtainedwhen at least 50% of the microspore population was in the mid-late to late uninucleate stage, when theanthers had been pretreated for 4 days on mannitol and when culture was performed with oxygen supply atregular inten’als. furthermore, the supplement of vitamins and casein hydrolysate in the culture mediumimproved the isolated microspore culture, whereas these compounds had a negative effect, if any effect atall, on the culture of anthers. Anther culture is not as laborious as microspore culture, but turned out to beat least 5 Umes less efficient. When mechanically isolated microspores were cuhured, onder the condiüonsfound to be optimal in the present study, a mean of 12.4 green plants per anther was obtained.
ntroductïon
The apphcaüon of microspore-derivedplants in breeding programs is an established technique today. Advantage is provided by accelerating and simplifying procedures (Monison and Evans, 1988). Moreover, isolated microspores are unicellularand haploid and are therefore promisingtargets for in vitro manipulation with subsequent selection for dominant as well asrecessive traits (Ahmad et al., 1991; Swan
son et al., 1989; Ye et al., 1987). Microspores, microspore-derived embtyoids orpollen are also vexy useful as target ceilsin experiments aiming at stable transformation, for example by cocukivation withAgrobacterium or partide bombardment(Creissen et al., 1990; Pechan, 1989;Twelletal., 1989).
A successfiil use of microspores for suchapplications requires optimized cukureprocedures to achieve a high regenerationfrequency. In barley the techniques foranther culture are rather well established.
31

Ojapter 2
NH4NO3KNO3MgSO4.7H201012P04CaC12H,OfeNa2EDTAMnSO4.H20H3B03iuZnSO4.7H20Na2MoO4.2H20CuSO4.5H.,OCoCI,.6H20Nicotinamidel’hiarnin HCIPyridoxine HCID-Caldum
pantothenateCholine chiorideL-ascorbic acidVic B,2RibollavinPolic addBioUnCholecaldferolVit. Ap-AnünobenzoicaddMyo.inositolGlutarnineCasein
hydrolysate3MaltoseSucroseFicoil 400Agarose
HMT (Sea Kern)Benzyl
aminopurine
1 II III
1651750350200450401550.7513.40.250.0250.025
20.020.2 - -
0.40.010.010.01 - -
0.02 -
100 100 100750 750 750
1000 - -
301/502 30Y602 -
-- 10/30
100,0001 100,000’ -
6,000
Table 1: Composition of media (mg/1); 1 modified
according to Jahne et al. (1991), II and III modified
MS according to Olsen (1987). for anther culture,
for isolated-rnicrospore culture, glutamine (25%)
is the active component (Bister-Miel et al., 1985),expressed as g11.
16519003701704404022.36.20.8311.70.250.0250.025
10
16519003701704404022.36.20.8311.70.250.0250.025
10
Olsen (1987) and Jiihne et al. (1991)have reported a yield of 7 green plants peranther, but in all other studies muchtower numbers were obtained.
Few studies have been directed towardsthe cultivation of isolated barleymicrospores. Furthermore, among thesestudies the experimental approach differsconsiderably. Kuhimaun et al. (1991)used the classic method of isolaüon, i.e.pestie maceration, and reported yields ofup to 10 green plants per arnher. Thepositive effect of microblending compared to pestie maceraüon described byOlsen (1991) resulted in 9.4 green plantsper anther. A third approach for culturingisolated microspores was chosen byZiauddin et al. (1990) who purifiedmicrospores, which were naturally shedfrom floating anthers, for culture andobtained up to 3 green plants per anther.
The aim of the present study was tocompare the influence of severalparameters on the effidency of the antherand microspore culture systems. Theinvestigations include idenüficaüon ofthe stages in microsporogenesis thatrespond optimally with respect tomicrospore embryogenesis, optimizatïonof the mannïtol pretreatment, evaluationof the effect of two different media and,assessment of the effect of oxygen supplyduring culture.
aterials and Methods
Growth of tize donor plaatsDonor plants of Hordeum vulgare cv. Igri weregrowningrowth cabinetsat l2Cwitha16-h lightregime (24000 lx) as described in detail by Olsen
(1987).
32

Anther and microspore culture
Spike sekctionTillers were harvested when the interligule lengthwas about 50 mm. Before the spikes tvere removedfrom the ensheathing leaves, the surface wassterilized with an aerosol of 70% ethanol.Microspores derived from anthers of the centralfloret were stained with acetocarmine. Thedevelopmental stage of at least 100 microsporesper spike was determined. The iniluence of themicrospore developmental stage on the effidencyof culture was assessed for spikes with disünctmicrospore populafions numbered 1 to V.
Anther cult;ureSixty anthers were placed on 1 ml 0.3 M mannitoland maintained for 4 days in the dark at 25C. Atthe end of pretreatment the shed microspores werecounted and the anthers were transferred toculture medium 1 (day 0) (Table 1). The cultureswere incubated in a container sealed with Parafilmin the dark at 25C. Pure oxygen was flushedthrough the container for 15 min. at days 0, 3 and7. At days 3, 7 and 10 fresh medium was addedand from day 10 onwards the cultures wereincubated under continuous orbital shaldng at 100rpm. Proper diluüon of the developing embryo-likestructures (ELS) is important.In addifion to the culture regime outlined above,the iniluence of 3 days of mannitol starvation,culture on medium II (Table 1) and culturewithout oxygen supply was studied.
Microspore isolation and cuttureThe anthers to be used for microspore isolationwere pretreated as described above. Afterpretreatment, microspores were isolated by verygentie maceration with a tdllon rod and sieving
through a nylon mesh (pore size 100 jLm). Thefiltrate was rinsed twice in 0.3 M mannitolfollowed by centrifugation at 100 x g for 5 min.The miaospores were then resuspended in 1 mlculture medium 1 or II (Table 1). Afterdetermining the number of isolated microsporeswith a haemocytometer (modified FuchsRosenthal), the volume was adjusted with culturemedium to give a final density of l.5x104microspores per ml. from correctly staged spikes 2-lOx 10 microspores were isolated per spike. Themicrospores were cultured as described underanther culture.
Regeneration ofplantsAfter 8 weeks of culture all EIS tvith a diameterlarger than 1 mm were transferred with a forcepsto medium III (Table 1; 1% sucrose, except B on3% sucrose) and incubated at 25C for 7 daysunder 600 Ix. Then the ELS and developing plantswere grown under 3200 lx. After a total of 4 weeksof culture the plates were scored for number ofgreen and albino plants. Plants were defined asdistinct structures possessing one or more leaves atleast 5 mm in length (in previous studies wasshown, that 90% of these are able to develop invivo into a plant). The ploidy level of representative plants was determined by flow cytometricanalysis of the relative nudear DNA content.
In order to be able to compare results obtainedfrom anther and microspore culture, experiments
performed according to the split-spikeapproach described by Kao (1981). All resuits presented are the mean of two separate experiments.The general conclusions from these experimentsare supported by at least 5 addiüonal experiments.
suits
Types of inicrospores
Two different types of microspores couldbe distinguished during pre-treatment,after isolation and in culture. One typeconsisted of small, often plasmolysed,microspores with a diameter of 35-40 cm.The other type is characterized asenlarged and swollen, 40-60 m indiameter and flch in cytoplasm which ismainly of a granular appearance;moreover, a red and/or blue interferencearound the exine was seen with the lightmicroscope. The latter type of microspores can be compared with type Amicrospores described by Bolik and Koop(1991) and with the large microsporesreported by Olsen (1991). It wasapparent that only the large type ofmicrospores divided and grew.
33
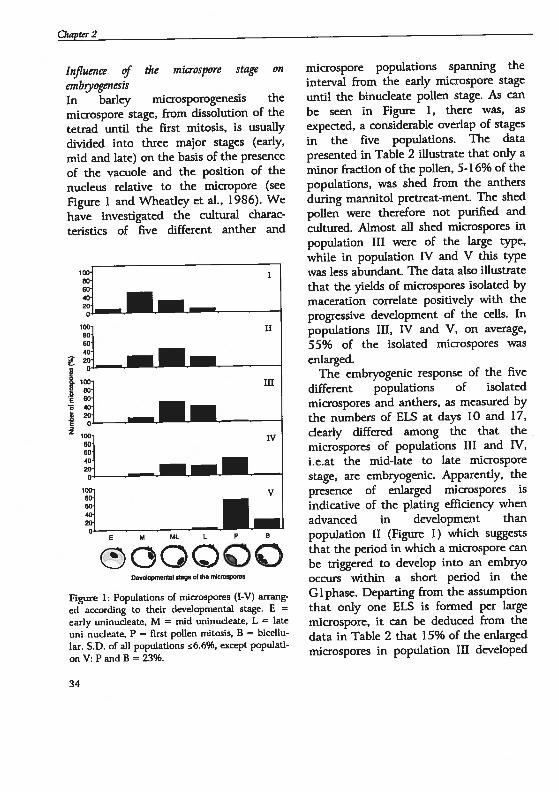
O,apter2
Infiuence of the microspore stage on
embyogenesisIn barley microsporogenesis themicrospore stage, from dissolution of thetetrad until the first mitosis, is usually
divided into three major stages (early,
mid and late) on the basis of the presence
of the vacuole and the posiüon of the
nucleus relaüve to the micropore (see
figure 1 and Wheatley et aL, 1986). We
have investigated the cultural charac
terisücs of five different anther and
v
ô66ôôDev&prn,taI stage o ue mrospores
Figure 1: Populations of microspores (I-V) arrang
ed according to their developmental stage. E =
early uninudeate, M = mid uninucleate, L = late
uni nudeate, P = first pollen mitosis, B = bicellu
lar. S.D. of all populations 6.6%, except populati
onV:PandB=23%.
microspore populations spanning the
interval from the early microspore stage
until the binucleate pollen stage. As canbe seen in figure 1, there was, as
expected, a considerable overlap of stages
in the five populations. The data
presented in Table 2 illustrate that only a
minor fraction of the pollen, 5-16% of thepopulations, was shed from the anthers
during mannitol pretreat-ment. The shed
pollen were therefore not purifled andcultured. Almost all shed microspores in
population III were of the large type,
while in population 1V and V this typewas less abundant. The data also illustratethat the yields of microspores isolated by
maceration correlate positively with theprogressive development of the ceils. In
populations III, 1V and V, on average,
55% of the isolated microspores wasenlarged.
The embryogenic response of the five
different populations of isolated
microspores and anthers, as measured by
the numbers of ELS at days 10 and 17,
clearly differed among the that themicrospores of populafions III and IV,i.e.at the mid-late to late microspore
stage, are embsyogenic. Apparently, thepresence of enlarged microspores isïndicative of the plating effldency whenadvanced in development thanpopulation II (figure 1) which suggeststhat the period in which a microspore canbe triggered to develop into an embxyooccurs within a short period in theGi pliase. Departing from the assumptionthat only one ELS is fonned per largemicrospore, it can be deduced from thedata in Table 2 that 15% of the enlargedmicrospores in population III developed
10080604020t —.
1 II
III
560
Zl9 w
10080604020
34
Anther and microspore cutture
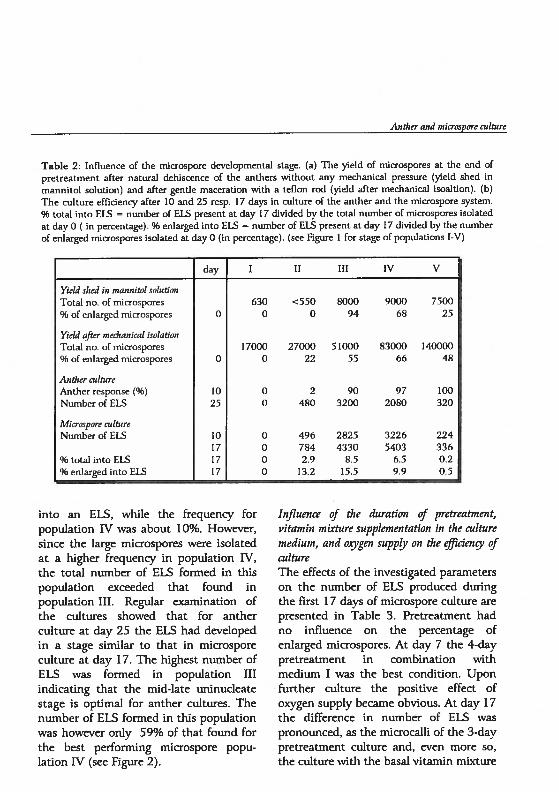
Table 2: Iniluence of the microspore developmental stage. (a) The yield of microspores at the end ofpretreatment after natural dehiscence of the anthers without any mechanical pressure (yield shed inmannitol soluüon) and after gentie maceration with a teflon rod (yield after mechanical isoaltion). (b)The culture effidency after 10 and 25 resp. 17 days in culture of the anther and the microspore system.% total into ELS = number of ELS present at day 17 divided by the total number of microspores isolatedat day 0 (in percentage). % enlarged into EIS = number of ELS present at day 17 divided by the numberof enlarged microspores isolated at day 0 (in percentage). (see Figure 1 for stage of populations I-V)
into art ELS, while the frequency forpopulation IV was about 10%. However,since the large microspores were isolatedat a higher frequency in population IV,the total number of ELS formed in thispopulation exceeded that found inpopulation III. Regular examination ofthe cukures showed that for anthercukure at day 25 the ELS had developedin a stage similar to that in microsporeculture at day 17. The highest number ofELS was formed in population IIIindicating that the mid-late uninudeatestage is optimal for anther cultures. Thenumber of ELS formed in this populationwas however only 59% of that found forthe best performing microspore population IV (see figure 2).
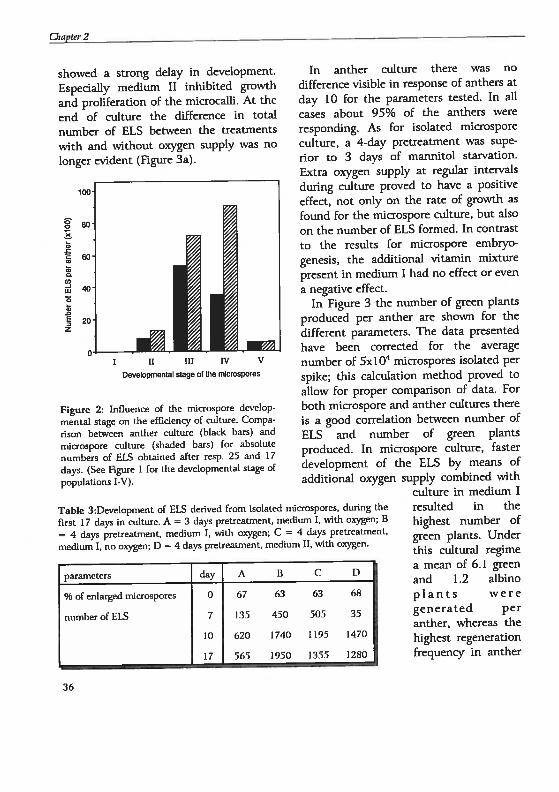
Infiuence of the duration of pretreahnent,vita,nin mixture suppiementahon in the adhiremedium, and oiygen supply on the efficiengi ofadtureThe effects of the investigated parameterson the number of ELS produced during
the first 17 days of microspore cukure arepresented in Table 3. Pretreatment hadno influence on the percentage ofenlarged microspores. At day 7 the 4-daypretreatment in combination with
medium 1 was the best condition. Uponfurther culture the positive effect ofoxygen supply became obvious. At day 17the difference in number of ELS waspronounced, as the microcalli of the 3-daypretreatment culture and, even more so,the culture with the basal vitamin mixture
Yietd shed in mannitol sotuhonTotal no. of microspores% of enlarged microspores
day 1 II III IV V
0
Yield after mechanicat isotahonTotal no. of microspores
% of enlarged microspores
630 <550 8000 9000 75000 0 94 68 25
Autlier cuttureAnther response (%)Number of ELS
17000 27000 51000 83000 1400000 22 55 66 48
1025
Microspore cultureNumber of ELS
0 2 90 97 1000 480 3200 2080 320
% total into ELS96 enlarged into ELS
10171717
0 496 2825 3226 2240 784 4330 5403 3360 2.9 8.5 6.5 0.20 13.2 15.5 9.9 0.5
Ozapter 2
showed a strong delay in development.
Espedatly medium II inhïbited growth
and proliferation of the microcalli. At the
end of culture the difference in total
number of ELS between the treatments
with and without oxygen supply was no
longer evident (Figure 3a).
100
In anther cukure there was no
difference visible in response of anthers at
day 10 for the parameters tested. In all
cases about 95% of the anthers were
responding. As for isolated microspore
culture, a 4-day pretreatment was supe
rior to 3 days of mannitol staivation.
Extra oxygen supply at regular intetvals
during culture proved to have a positive
effect, not only on the rate of growth as
found for the microspore culture, but also
on the number of ELS formed. In contrast
to the resuits for microspore embryo
genesis, the additional vitamin mixture
present in medium 1 had no effect or even
a negative effect.In Figure 3 the number of green plants
produced per anther are shown for the
different parameters. The data presented
have been conected for the average
number of 5x 1 o microspores isolated per
spike; this calculation method proved to
allow for proper comparison of data. For
both microspore and anther cultures there
is a good conelation between number of
ELS and number of green plants
produced. In microspore culture, faster
development of the ELS by means ofsupply combined with
culture in medium 1resulted in thehighest number ofgreen plants. Underthis cultural regimea meart of 6.1 greenand 1.2 albinoplants weregenerated peranther, whereas thehighest regenerationfrequency in anther
00
cl,tt0
cl,
0-JL5
0
0.0
0z
II III IV V
Developmental stage ot the microspores
Figure 2: Influence of the microspore develop
mental stage on the efficiency of culture. Compa
rison bettveen anther culture (black bars) and
microspore culture (shaded bars) for absolute
numbers of ELS obtained after resp. 25 and 17
days. (See figure 1 for the developmental stage of
populauons 1-V). addïtional oxygen
Table 3:Development of ELS derived from isolated microspores, during the
first 17 days in cukure. A = 3 days pretreatment. medium 1, with oxygen; B
= 4 days pretreatment, medium 1, with oxygen; C = 4 days pretreatment,
medium 1, no oxygen; D = 4 days pretreatment, medium II, with oxygen.
parameters day A B C D
% of enlarged microspores 0 67 63 63 68
number of ELS 7 135 450 505 35
10 620 1740 1195 1470
17 565 1950 1355 1280—
36
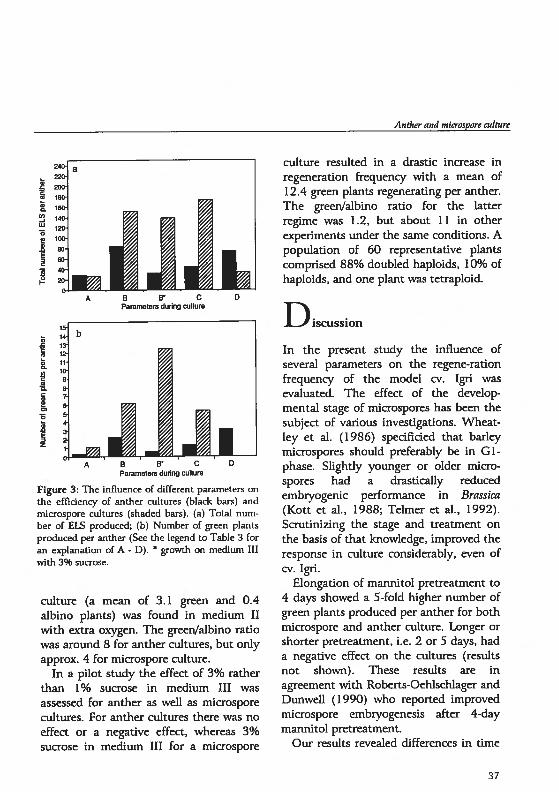
Anther and micraspore culture
24 a -
le
16
14
OBJCParameters duhng cultura
t,
t,
tt,0.t
0
0
E2
A B - B CParameters during culture
figure 3: The influence of different parameters onthe effidency of anther cultures (black bars) andmicrospore cultures (shaded bars). (a) Total numher of ELS produced; (b) Number of green plantsproduced per anther (See the legend to Table 3 foran explanation of A - D). * growth on medium IIIwith 3% sucrose.
culture fa mean of 3.1 green and 0.4albino plants) was found in medium IIwith extra oxygen. The green/albino ratiowas around 8 for anther cultures, but onlyapprox. 4 for microspore culture.
In a pilot study the effect of 3% ratherthan 1% sucrose in medium III wasassessed for anther as well as microsporecultures. for anther cultures there was noeffect or a negative effect, whereas 3%sucrose in medium III for a microspore
culture resulted in a drastic increase inregenerafion frequency with a mean of12.4 green plants regenerating per anther.The green/albino ratio for the latterregime was 1.2, but about 11 in otherexperiments under the same conditions. Apopulation of 60 representaüve plantscomprised 88% doubled haploids, 10% ofhaploids, and one plant was tetraploïd.
iscussion
In the present study the influence ofseveral parameters on the regene-rationfrequencv of the model cv. Igri wasevaluated. The effect of the developmental stage of microspores has been thesubject of various investigations. Wheatley et al. (1986) specificied that barleymicrospores should preferably be in G 1-phase. Slightly younger or older microspores had a drastically reducedembryogenic performance in Brassica(Kott et aL, 1988; Telmer et al., 1992).Scrutinizing the stage and treatment onthe basis of that knowledge, improved theresponse in culture considerably, even ofcv. Igri.
Elongation of mannitol pretreatment to4 days showed a 5-fold higher number ofgreen plants produced per anther for bothmicrospore and anther culture. Longer orshorter pretreatment, i.e. 2 or 5 days, hada negative effect on the cultures (resultsnot shown). These resuits are inagreement with Roberts-Oehischiager andDunweil (1990) who reported improvedmicrospore embyogenesis after 4-daymannitol pretreatment.
Our resuits revealed differences in time
37

Chavter 2
between microspore and anther cultureresponse. The anther wall apparentlycreates a closed environment until theintemal Pool of nutrients is depleted.Isolated pollen however, can respondimmediatety to the medium (HeberleBors, 1989) which resuits in earlierdevelopment in culture. Continuation ofpollen development within the anthercould also explain that in anther culturean earlier microspore stage (III) is optimalcompared to microspore culture.
Furthermore, both anthers and isolatedmicrospores responded differently whencultured in medium 1 and II. In barleyanther culture extra vitamins (Kao et aL,1991) and amino acids (0.1 mM) (Zhu etal, 1990) have been supplied. Xu andSunderland (1981) found an inhibitoxyeffect of g]utamine at 5.5 mM in antherculture which is consistent with ourresuits (6.8 mM glutamine). Upon cufturein medium with high concentrations ofglutamine, this compound probably isredundant in anther culture, as the antherwall and connective tissue mediate nutflents, in particular glutamine, to thepollen gram (Maheshwari et al, 1980). Inthe case of isolated microspores, the additional medium components possiblyreplaced (in part) the anther wall andtapetal tissue.
The results obtained in this studyindicate that the extra supply of oxygenhad a positive effect on the developmentof both microspore and anther cuftures.The benefit of proper aeration isdocumented by lichter (1989) who usedagitation of Brassica microspores, and byLuckett et al. (1991) and Olsen (1991)who applied membrane rafts.
In this study about 6.5 times moremicrospores were obtained when theywere isolated mechanically instead ofshed naturally. The dehiscence of anthersis dependent on free pollen present in thelumen of the anther loculi (Sunderland etal., 1984); mechanical isolation cantherefore be necessaiy to obtain sufficientmaterial. It is however crucial to applyvery gentle forces for the isolation of themicrospores, otherwise their viability willbe veiy low. From the results presented itis evident that the isolated microspore system is easier to control during cultureand provides at least 5 tïmes more greenplants than anther culture under theoptimum conditions. for Brassica napuseven a ten-fold higher efficiency ofisolated microspores as compared tocultured anthers has been reported(Siebel and Pauls, 1989). The antherculture system, however, will be a goodroutine method for the production ofdoubled-haploid barley plants.
From the resufts obtained in this studyit is dear that optimization of (1) themicrospore stage, (2) the duration ofpretreatment, (3) the medium composition and (4) the additional oxygen supplystrongly improved the number of plantsgenerated in microspore culture of barleycv. [gri.
Preliminary results show that the springbarley cuftivars Dissa and Gimpel alsodevelop optimally, when the optimizedcuftural regime is applied. Genotypespecifidty is reflected in the frequency ofregeneration, which are below thatobserved for cv. Igri.
In summary, microspore culture doesmeet the requirement of high regene
38

Anther and microsporeculture
ration frequency enabling theirapplicaüon. Experiments aiming at stabletransformaüon through partide bombardment of microspores are ïn progress.
AcknowtedgernentWe thank Dr. F.1. Olsen, Carlsberg Research
Laboratory, for the pleasant introduction in this
field and helpful discussions, Dr. P.B. Halen,
Carisberg Research Laboratory, for criticaliy
reviewing the manusaipt, C. ten Bosch and S. An
for pilot studies, J.M. Vink for growing the plants
and P. Hock for the layout of figures and tables.
This vork was supported by EUREKA grant
EU270, and is ABIN publicaion 86.
Ieferences
Ahmad 1., J.P. Day, M.V. MacDonald and D.S.Ingram. Haploid culture and UV mutagenesis inrapid-cycling Brassica napus for the generation ofresistance to chiorsulfuron and Alternaria
brassidcola.Ann. Bot. 67: 521525 (1991)Bister-Miel f., J.L. Guignard, M. Bury and C.Agier. Glutamine as an active component of caseinhydrolysate: Its balandng effect on plant cclicultured in phosphonis defident medium. PlantCcli Rep. 4: 161-163 (1985)Bolik M. and H.U. Koop. Identification ofembiyogenic microspores of barley (Hordeumvulgare 1.) by individual selection and ctilture andtheir potential for transformation bymicroin(ection. Protoplasma 162: 61-68 (1991)Creissen G., C. Smith, R Francis, H. Reynoldsand P. Muilineaux. Agrobacterium - and microprojediie - mediated viral DNA delivery intobariey microspore-derived cultures. Plant Ccli Rep.8: 680-683 (1990)Heberle-Bors E. Isolated pollen culture in tobacco: plant reproductive development in a nutsheil.Sex. Plant Reprod. 2: 1-10 (1989)Jâhne A., P.A. Lazzeri, M. Jager-Gussen and F1.Lörz. Plant regeneration from embryogenic cclisuspensions derived from anther cultures ofbariey (Hordeum vidgare 1). Theor. Appi. Genet.
82: 74-80 (1991)Kao KN. Plant fotmation from barley anther
cultures with ficoil media. Z. Pflanzenphys. 103:437-443 (1981)Kao KN., M. Saleem, S. Abrams, M. Pedras,
D. lom and C. Mallard. Culture conditions forinducüon of green plants from bariey microspores
by anther culture methods. Plant Ccli Rep. 9:
595-601 (1991)ICott LS., 1. Polsoni and W.D.Beversdorf.Cytotogicai aspects of isoiated
microspore cuiture of Brassica napus. Can. J. Bot.66: 1658-1664 (1988)Kuhlmann U., B. foroughi.Wehr, A. Graner
and G. Wenzel. Improved culture system formicrospores of bariey to become a target for DNAuptake. Plant Breed. 107: 165-16 (1991)Lichter It Efficient yield of embryoids by culture
of isolated microspores of different Brassicaceaespedes. Plant Breed 103: 119-123 (1989)Luckett D.J., S. Venkatanagappa, N.L Darvey
and RA. Smithard. Anther culture of Australianwheat germplasm using modified CI 7 medium andmembrane rafts. Aust. 1. Plant Phys. 18: 357-367
(1991)Maheshwari S.C., A.K. Tyagi and IC Malhotra.
inducüon of haploidy from pollen grains in angiosperms - the current status. Theor. Appi. Genet. 58:193-206 (1980)Momson LA. and D.A. Evans. Haploid plantsfrom tissue cuiture: new plant varieties in ashortened time frame. Biotechnology 6: 684-690(1988)Olsen F.L. Induction of microspore embryogenesisin cultured anthers of Hordeum vulgare: the effectsof ammonium nitrate, glutamine and asparagine asnitrogen sources. Carisberg Res. Comm. 52: 393--404 (1987)Olsen F.J. Isolaüon and cultivation ofembiyogenic microspores from bariey (Hordeumvulgare 1.). Hereditas 115:255-266 (1991)Pechan P.M. Succesful coctiltivation of Brassicanapus microspores and proembiyos withAgrobacteriurn. Plant Ccii Rep. 8:387-390 (1989)
Roberts-Oehischiager S.L. and J.M Dunweil.Bariey anther culture: pretreatment on mannitolstimulates production of microspore-derived
embryos. Plant Ccli Tiss. Org. Cult. 20: 235-240(1990)Siebet J. and KP. Pauis. A comparison of antherand microspore culture as a breeding taal in Brassica napus. Theor. Appi. Genet. 78: 473-479 (1989)
39

Ozapter 2
Sunderiand N., B. Huang and G.J.Hills.Disposition of pollen in situ and its relevanceto anther/pollen culture. J. Exp. Bot. 35: 52 1-530(1984)Swanson E.8., M.J. Herrgesell, M. Arnoldo,D.W. Sippeli and R.S.C. Wong. Microsporemutagenesis and selection: Canola plants with fieldtolerance to the imidazolinones. Theor. Appi.Genet. 78; 525-530 (1989)Telmer C.A., D.H. Simnionds and W. Newcomb.Determination of developmental stage to obtainhigh frequencies of embryogenic microspores inBrassica uapus. Physiol. Plant. 84: 4 17-424 (1992)Tweil D., T.M. Klein, ME. fromm and S.McCormick. Transient expression of chimericgenes delivered into pollen by micropro)ectilebombardment. Plant Phys. 91: 1270-1274 (1989)Wheatley W.G., A.A. Marsolais and K.J.Kasha. Microspore growth and anther staging in
barley anther cuiture. Plant Celi Rep. 5: 47-49(1986)Xu Z.H. and N. Sunderland. Glutamine, inositoland conditioning factor in the production of barleypollen callus in vitro. Plant Sci. LetL 23: 161-168(1981)Ye J.M., KN. Kan, B.L. Harvey and B.G.Roasnagel Screening salt-tolerant barley genotypesvia f1 anther culture in salt stress media. Theor.Appl. Genec 74: 426429 (1987)Zhu M., X. Abing, Y. Miaobao, H. Chunnong,Y. Zhilong, W. Linji and Y. Jianjun. Effects ofamino adds on callus differentiation in barleyanther culture. Plant CelI TÎss. Org. Cult. 22: 20 1--204 (1990)Ziauddin A., E. Simion and Kj. Kasha.Improved plant regeneration from shed microsporeculture in barley (Hordeum vulgare L) cv. Igri. PlantCeli Rep. 9: 69-72 (1990)
40

S
hapter 3
Microspore culture of Hordeum vulgare L.:the influence of density and osmolality
S. Hoekstra, MH. van Zijderveld, f. Heidekamp & F. van der Mark
Donor plants of Hordeum vulgare L. cv. Igri were grown in a condifioned environment to minimise
fluctualions in the composition of the microspore population. After isolation different types of rnicrospores
were identified within each population, alnongst others an embxyogenic subpopulation. It was shown that
the optimum plating density is achieved by adjusfing the density to 2x104 embryogenic microspores per ml,
wiffi a lower threshold at 5x103 per ml. By increasing the osmolality of the pretreatment solution to 440
mOs.kg’ and that of the ciilture medium to 350 mOs.kg’, up to 15% of the population developed into
embryo-like structures. When microspores of cv. Igri were cultured under the optimized conditions, the
ratio of greenMbino plants incieased from 1:1 to 34:1 and 50 green plants per anther were formed.
ntroduction
Miaospore ctilture represents a potenUal
tool for a range of biotechnological
applications. Moreover, the microspore
system ïs an attractive model to study the
molecular mechanism that controls the
switch from the gametophytic to the
sporophytic pathway. However, the
induction frequency and the effidency of
outgrowth ïnto green plants remains low
and unpredictable in the microspore
system. In order to be able to enhance the
efficiency of microspore culture, it is
necessary to identify the type of
microspore that is capable of undergoing
embryogenesis and subsequent devel
opment into a green plant. The number
of what are called embryogenic
microspores, may vaiy withïn one variety
due to the environmental condiüons in
which the donor plants are grown
(Heberle-Bors 1989). Weil-controlled
growth conditions of barley donor plants
and the use of an effident isolaüon
41

Chapter3
procedure allowed us to perfonnreprodudbie experiments throughout theyear (Hoebtra et al. 1992).
The effect of plaüng density of themicrospores on the number of developingembryo-like structures (ELS) has beenreported earlier. for rapeseed Huang et al.(1990) describe that a density of 3-4x104microspores per ml is crudal forembiyogenesis. for maize, a higherdensity viz. 6-8x104 microspores per ml,produces an optimum in culture efficiency(Gaillard et al. 1991). The effect ofdifferent types of microspores present in apopulation on the plating density andculture efficiency has not beeninvestigated before.
Besides the density, the importance ofthe medium osmolality is known foranther culture (Kao 1981; Chu et al.1990; Zhou et al. 1991). To date, onlyPesdteffi et al. (1990) applied dîfferentosmotical conditions for isolatedmicrospore culture, reporting an optimumconcentration of 7.9% sucrose for thedevelopment of embryo-like structures.
In this paper, for Hordeum vutgare L. cv.Igri a description is given of the differenttypes of microspores which are present ina population directly after isolation, inorder to recognîze the type of microsporethat has the capadty to develop into aplant. The influence of the density of thismicrospore-subpopulation on the cultureeffidency is invesügated. furthermore,the effect of the osmolality of both thepretreatment and the culture medium onELS and plant formation is studied.
aterials and Methods
The growth of the donor plants Hordeum vulgare Lcv. Igri, the selecüon of the material, thepretreatment and culture conditions wereessentially as described previously (Olsen 1991;Hoekstra et al. 1992). In short, donor plants weregrown at 12C with a 16 h light regime (24,000lux) and the anthers were harvested when at least50% of the microspores were in the Mid Late toLate Uninucleate stage. Upon pretreatment of theanthers on mannitol solution for 4 days, microspores were isolated after pestie maceration (about500 embryogenic microspores per anther) andplated as maas culture in medium 1. Under optima!conditions, at least 30% of the microsporepopulation was embryogenic (see resuits).Different types of microspores were recognized ina population by light microscopy using anOlympus inverted microscope IMT-2, on the basisof their diameter, colour interference of the exine,and appearance and colour of the cytoplasm. Theirbehaviour in culture was followed. Viability wasassessed using the viable stam fluorescein diacetate(stock soluion 5 mgfml in acetone).The iniluence of the density of embiyogenicmicrospores on the ability to develop iflto ELS wasinvestigated onder different pretreatmentconditions (viz. anthers for 2, 3 or 4 days onmannitol starvation (Hoekstra et al. 1992) orspikes incubated at 4C for 28 days (Huang andSunderland 1982)) in medium II (Hoekstra et al.1992).The effect of increased osmolality of thepretreatment solution (with mannito!) and ofmedium 1 (with maltose) on the formation ofmicrospore-derived ELS has been tested.The opilmum density of the embtyogenicmicrospores as well as the best osmotic conditionsduring pretreatment and culture have been appliedto study their effect on the ability of themicrospores to develop into ELS and on thenumber of plants subsequentiy formed. Plantformation was assessed on the Orst 1000developing ELS after careful transfer with forcepsto medium III (Hoekstra et al. 1992). The numberof plants per anther was calcsilated as the numberof plants on the first 1000 ELS, divided by the
42

Density and osmolality
number of anthers used for the microspores platedin 1 ml medium.In the present study plants were defined as distinctstructures possessing one or more leaves at least 5mm in length; previous experiments have shownthat 90% of these plants were able to develop invwo into a fertile plant.The ploidy level of representative plants wasdetermined by flow cytometric analysis of the
relative nuclear DNA content.
suits
Types of microsporesDifferent types of microspores could berecognized during the first stages ofisolation and in culture. Using fluoresceindiacetate as a viable stam, a vaiyingpercentage of each population was nonviable. This type was characterized by asmall diameter (35-40 jm), no colourinterference of the exine, and a collapsedcytoplasm (figure la). The remainingmicrospores ]ooked viable: enlargeddiameter up to 65 tm, a red and/or blueinterference of the exine and viablecytoplasm (Figure ib and c). Only afraction of these microspores, however,divided at day 3. The rest of the supposedviabte microspores did not change inmorphology or were plasmolysed. Afterthorough investigation it was evident thatmicrospores which had a diameter of 40-50 sm and showed a blue interference ofthe e,dne (figure lb), plasmolysed uponculture (Figure 1 d). They represented onaverage 15% of the isolated microspores.Visually recognizable microspores, forwhich later on many showed the capadtyto divide, had a diameter of 50-60 tm,showed red ïnterference of the exine andhad a granular cytoplasm. This group of
mÎcrospores was defined as theembryogenic subpopulaüon (figure Ic),typically 60% of the microspores at theday of isolation. It should be noted thatof this population about 25% (15% oftotal) divided at day 3 (figure le).
Influence of embryogenic microspore densiy onthe number ofELSData concerning plating density ofembiyogenic microspores are presented inFigure 2. These data are derived from 28experiments under different pretreatmentconditions, which resulted in fiuctuationsin the percentage of embryogenicmicrospores isolated. A minimum densityof 5x1 o embzyogenic microspores per mlwas necessaxy to ensure furtherdevelopment of the microspores. At thisthreshold density, even in populations
400
.300 •
cj .•wE 2000). 1
.z
.4. .•.. .
02 8
Embryogenic microspores (xl0 per ml)
figure 2: Correlation between the density ofembryogenic microspores and the number of ELSformed per iO microspores 10 days after isolagon. Different pret.reatments were appiled in 28experiments (see materials and methosds).
43
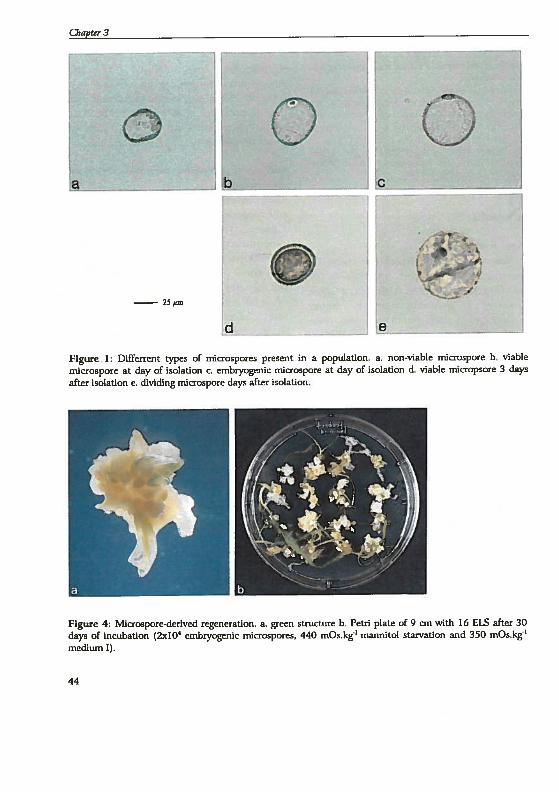
Chapter 3
0 0a
0
— 25 m
d
figure 1: Differrent types of mirrospores present in a population. a. non-viable microspore b. viable
microspore at day of isolation c. embiyogenic microspore at day of isolation d. viable micropsore 3 day’s
after isolation e. dividing microspore day’s after isolation.
Figure 4: Microspore-derived regeneration. a. green structure b. Petri piste of 9 cm with 16 ELS after 30days of incubafion (2x104 embryogenic microspores, 440 mOs.kg’ mannitol starvaion and 350 mOs.kg’medium 1).
44
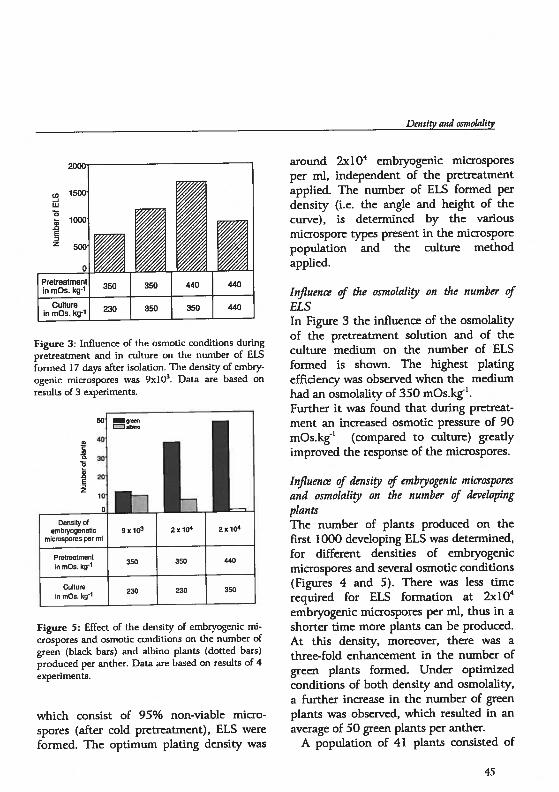
Density and osmolality
which consist of 95% non-viable microspores (after cold pretreatment), ELS wereformed. The optimum plating densïty was
around 2x1 ü embryogenic microsporesper ml, independent of the pretreatmentapplied. The number of ELS formed perdensity (i.e. the angle and height of thecurve), is determined by the vaiiousmicrospore types present in the microsporepopulaüon and the culture methodapplied.
Influenc€ of the osznola1iy on the number of
ELSIn figure 3 the influence of the osmolalityof the pretreatment solution and of theculture medium on the number of ELSformed is shown. The highest platingefficiency was observed when the mediumhad an osmotality of 350 mOs.kg’.further it was found that during pretreatment an increased osmofic pressure of 90mOs.kg’ (compared to afiture) greadyimproved the response of the microspores.
Influence of densiy of ernbiyogenic microsporesand osinola1iy on the numher of developingplantsThe number of plants produced on thefirst 1000 developing ELS was determined,for different densities of embryogenicmicrospores and several osmotic conditions(Figures 4 and 5). There was less timerequired for ELS formation at 2x104embiyogenic microspores per ml, thus in ashorter time more plants can be produced.At this density, moreover, there was athree-fold enhancement in the number ofgreen plants fonned. Under optimizedcondiüons of both density and osmolality,a further increase ïn the number of greenplants was observed, whïch resulted in anaverage of 50 green plants per anther.
A population of 41 plants consisted of
figure 3: lnfluence of the osmotic conditions duringpretreatment and in cutture on the number of ELSformed 17 days after isolation. The density of embiy
ogenic microspores was 9x103. Data are based 0fl
resuits of 3 experiments.
figure 5: Effect of the density of embryogenic microspores and osmotic conditions on liie number of
green (black bars) and albino planta (dotted bars)produced per anther. Data are based on resuits of 4experiments.
45

Chapter3
87% doubled haploids, 11% tetraploids and1 plant was triploid. The change in ratio ofgreen/albino plants from 1:1 to 34:1 underoptimized condiflons was remarkable.
T)In general, embiyogenic microspores arecharacterized by a large vacuole e.g. intobacco (Kyo and Harada 1985), rapeseed(Deslauriers et al. 1991), maize (Coumanset al. 1989) and wheat (Datta and Wenzel1987). Enlarged diameter accompanied by ared interference colour of the exine ischaractedstic for embryogenic microsporesof rice (Cho and Zapata 1990). fromprevious experiments, It was evident thatthe distinction between viable and nonviable microspores on the basis of enlargeddiameter was not reliable enough to obtainreproducible results, i.e. a fixed fraction ofthe viable population developing intoplants. In barley, Olsen (1991) describesthat microspores with a blue or red coronadivide, and that only the blue type is ableto break through the exine. The “Olsen bluetype’ is different from the blue typepresented here, as the latter was alreadyplasmolyzed after 3 days in culture. Itshould be emphasized that both diameterand colour of the exine are affected by theosmotic pressure applied (resuks notshown). Besides, the colour of the exineobserved is a resuk of the spedfic refractionof light in a given microscope. The opticalconditions and the osmotic environmentdescribed by Olsen (1992) are differentfrom our experimental design.Bolik and Koop (1991), furthennore,disfinguished 2 types of cytoplasm in barley
microspores: embryogenic type A beinggranular in appearance and nonembiyogenic type B containing less cytoplasm, which in addition is smooth andstar-like arranged. These data are based ona limited number of 5 regenerants. In ourexperiments star-like microspores wereobserved under suboptimal conditions e.g.low density of embiyogenic microspores.We observed that the presence of smallvacuole(s) or “dark-dotted” cytoplasm alsoi’ere indicative under these circumstances(resuks not shown). Only 25% of theembryogenic population divided, which isin agreement with the resuks whichGaillard et al. (1992) show for maize. fora further identification of embxyogenicmicrospores more investigaüons arerequired.
The appropriate cukure density isdependent on the quality of the materiali.e. the percentage of embryogenicmicrospores present, which is influencedby the pretreatment and the method ofmicrospore isolaüon. Therefore, differencesin optimum densities may vary betweenspecies and even laboratories. Our resuksshowed a relatively low optimum densitywhich can be explained by the use of gooddonor quality. Gaillard et al. (1991)reports 6-8x1 o microspores per ml formaize, whereas for rapeseed Huang et al.(1990) finds 3-4x104 mÏcrospores per mlto be optimal. Cho and Zapata (1990)describe for rice that non-embtyogenicmicrospores die earlïer and that toxicsubstances released from these deadmicrospores probably are deleterious to thedevelopment of embryogenic microspores.Osmotic pressure of the media bas beenshown to be an important parameter in
46

Densifyandosmolality
number of green plants together with adecrease in the number of albino plants, isin agreement with the resuits obtained foranther culture of wheat (Zhou et al. 1991).It can be assumed, that this effect is due tothe osmotic conditïons as there probablywas an abundance of carbon source. It canbe conciuded that the number of albinoplants produced is not merely genotypedependent, but is also largely determined byyet unidentified physio-logical factors in theculture conditions used. Preliminaiy resuksshowed a comparable positive effect at thelevel of ELS fonnation for cvs. Gimpel andDissa when the optimizations descijbed inthis paper were applied.
In the literature reported so far, barleymicrospore cultures of cv. Igri produce up to12 green plants per anther ( Olsen 1991;Kuhlmann et al. 1991; Hoekstra et al.1992). Our resuks show that for cv. Ign theoptÏmizations resulted in a four-foldincrease in the number of green plantsformed; it should be emphasized that thiscalculation is an underesümafion of plantformation based on only the first 1000 ELS.The fonnation of 50 green plants per antheron a routine basis affirms that applicationsfor microspore culture are more feasible inthe near future.
AcknowkdgementrWe thank Prof. RA Schilperoort for criticallyreviewing the manusaipt, J.M. Vink and J.D.LLouwerse for growing the plants and P. Hock for thelayout of the ttgures. This vork was supported byEUREKAgrant EU270 and isABIN publication 120.
I..eferences
Bolik M. and H.U. Koop. Identification ofembryogenic microspores of barley (Hordeurn vutgareL) by individual selection and culture and theirpotential for transformation by microinjection.Protoplasma 162: 61-68 (1991)Cho M.S. and f.J. Zapata. Plant regenerationfrom isolated microspores of Indica rice. Plant CeliPhysiol. 31: 881-885 (1990)Chu CC., Hili RD and AL. Brule-Babel. Highfrequency of pollen embryoid formation and plantregenerauon in Triticurn aestivnrn L. onmonosaccharide containing media. Plant Sci. 66:255-262 (1990)Coumans M.P., S. Sohota and E.B. Swanson.Plant development from isolated microspores of Zearnays L Plant Cdl Rap. 7:618.621 (1989)Datta S.K and G. Wenzel. Isolated microsporederived plant formaion via embryogenesis inTriticurn aestivun, L. Plant Sd. 48: 49-54 (1987)Deslauriers C., A.D. Poweil, K Fuchs and K.P.Pauls. How cytometric characterization and sorungof cultured Brassica napus microspores. Bioch.Biophys.Acta 1091:165-172 (1991)Gaillard A., E. Mattys.Rochon and C. Dumas.Selection of microspore derived embryogenicstructures in maize related to transformationpotenual bymicroinjecuon. Bot. Acta 105: 3 13-318(1992)Gaillard A., P. Vergne and M. Beckert.Optimization of maize micropsore isolaüon andculture conditions for reliable plant regeneraüon.Plant Ceil Rep. 10:55.58 (1991)Heberle-Bors E. Isolated pollen culture in tobacco:plant reproductive development in a nutshell. Sex.Plant Reprod. 2: 1-10(1989)Hoekstra S, M.H. VanZijderveld, J.D. I.ouwerse,F. Heidekamp and F VanderMark. Anther andmicrospore culture of Hordestm vulgare L cv. Igri.Plant Sd. 86: 89-96 (1992)Huang B., S. Bird, R Kembie, D. Simmonds,W. Ketter and B. Mild. Effects of culture densitvconditioned medium and feeder cultures onmicrospore embryogenesis in Brassica napus L. cv.Topas. Plant CeIl Rap. 8:594-597 (1990)Huang B. and N. Sunderland. Temperature-stressin barley anther culture. Ann. Bot. 49: 77-88 (1982)
anther culture (Chu et al. 1990; Kao 1981).The optimal osmotic environment from thisbarley study is comparable with the resultsdescribed for maize (Pescitelli et al. 1990).At high osmolality, an increase in the
47

O,apter 3
Kao K.N. Plant formaUon from barley anther
cultures with ficoil media. Z. Pflanzenphys. 103:
437-443 (1981)Kuhlmann U., B. Foroughi.Wehr, A. Graner and
G. Wenzel. Improved cuhure system for microspores
of barley to become a target for DNA uptake. Plant
Breed. 107: 165-168 (1991)ICyo M. And H. Harada. Studies on conditions for
cefl division and embryogenesis in isolated pollen
cultuxe of Nicotiana rustica. Plant Physiol. 79; 90-94
(1985)
Olsen F1. Isolation and cultivation of embryogenic
microspores from barley (Hardcum vulgare L.).
Hereditas 115; 255-266 (1991)Pesckelli S.M., C.D. Johnson and J.P Petolino.
Isolated microspore culture of maize; effects of
isolation technique reduced temperature and
sucrose level. Plant Celi Rep. 8; 628-631(1990)
Zhou H., Y. Zheng and C.F. IConzak Osmotic
potential of media affeding green plant percentage
in wheat anther culture. Plant Celi Rep. 10; 63-66
(1991)
48

hapter 4
The interaction of 2,4-D application andmannitol pretreatment in anther and microspore culture
of Hordeum vutgare L. cv. Igri.
S. Hoekstra, S. van Bergen, I.R. van Brouwershaven, RA. Schilperoort & f. Heidekamp
SummaryThe influence of 2,4-dichlorophenoxyacetic add (2,4-1)) on embryo-like structures (ELS) and plant
development from barley microspores was determined. Miaospores cultured on filters enabled simple
modification of growth regulator concentrations. Regeneration frequencies obtained with 2,4-D as growth
regulator were similar to the resuits achieved with the generally applied cytokinin 6-benzylaminopurine. 1f
2,4-1) was applied after a regtilar mannitol pretreatment, maximal plant regeneration was achieved 1f 10.6
moljL 2,4-D was present conünuously or for 7 days. Mtematively, maximal plant formation was induced, if
l0 or l0 moVL 2,4-D was present for 1 hour or if present resp. 3 days or 1 day. Induction of plant
regeneration by a 1 h treatment with lO mol /1. 2,4-D is a more general observed phenomenon for single
cells or small celI dusters of both dicotyledonous and monocotyledonous spedes.
Without mannitol pretreatment, in anther culture it was possible to induce plant production after 2,4-D
treatment only in anther cultures. Without mannitol pretreatment no embryogenic type of microspores can
be recognized at the moment of microspore isolation, and plating effidency never reached 1%. In anther
culture without mannitol pretreatment, a higher molarity and/or longer presence of 2.4-1) was required and
resulted only in about 1 green plant per anther. After application of a mannitol pretreatment, plant
production inaeased at least 10 times.To our knowtedge this is the first report on microspore-derived barley plants via androgenesis without
any pretreatmenc The combination of 2,4-D and anther pretreatment with mannitol as trigger for
microspore differenfiafion is discussed.
49

Chapter 4
ntroduction
Syntheüc awdns such as 2,4-dichiorophe
noxyacetic add (2,4-D), are essential
constituents of culture media that areused for production of embryogeniccultures of both dicotyledonous andmonocotyledonous plants such as theGramineae rice (Abduliah et al., 1986),wheat (Vasil et al., 1990), maize (Morocz
et al., 1990) and barley (Lührs and Lörz,1987). 2,4-D has been used to inducededifferentiafion or, if applied at a higherconcentration, for induction ofembryogenesis. Dudits et al. (1992)observed that a 10 mes higherconcentration than used for the initiationof dedifferentiation, induces embryogenesis in dedifferentiated microcallussuspensions of carrot and alfalfa.Comparable resuits are obtained forcucumber (Tabel et al., 1991) and themonocotyledonous spedes Pennisetum inflorescense-derived callus and Asparagus instem-derived callus (Talwar and Rashid,1990; 1.evi and Sink, 1991).
From carrot cel cultures, it is knownthat there are at least two phases in thedifferenüation from single ceils intoembiyos. The development of the firstphase requires exogenous auxin, whereasthe second phase is inhibited by the samegrowth regulator (Komamine et al.,1990). The response to exogenous auxinin somaüc embryo induction appears todepend on the nature of the explant usedin the experiment (Zimmerman, 1993).
Generally in microspore culture, if(synthetic) auxins are used they arepresent during the entire induction
period. However in cereal microsporeculture, auxins have rarely been used sofar
as growth regulators (Tiwari and Rahimbaev, 1991; Hassawi et al., 1990; Bali et
al., 1993). Generally 6-benzyl-
aminopurine (BAP) is used (a.o. Olsen,
1987; Hoekstra et al., 1993). Datta andPotrykus (1989) stated that low auxinwas important for the development of
embryos, because it prevents thedevelopment of unorganized cultures. Asignificant negative effect of 2,4-D, 1fapplied for a long period, on plantregeneration and green plant percentageis shown in wheat anther culture (Bali etal., 1993).
In the present paper we studied,whether 2,4-D can induce plantdevelopment in barley microspores andunder which conditions maximal plantformation was achieved. Moreover, theimportance of anther tissue inmicrospore-derived plant formation wasinvestigated upon 2,4-D treatments withand without mannitol pretreatment.
aterials and Methods
Growth of the donor plants Hurdeum vulgare L cv.Igri, selection of material, mannitol pretreatment
of anthers and culture conditions were as descrlbed
previously (Hoekstra et al. 1993) with thefoltowing modifications. Anthers (wilhout ficoil)or miaospores were cultured in medium 1 at 350
mûs.kg1 (Hoekstra et al., 1992) with 2.5%
coconut water. When the majority of ELS were atleast 1 mm in diameter, ELS were plated on
medium III (Hoekstra et al., 1992) with 0.4 mol /LIGneUn (Sigma).
Microspare cultureAfter isolation, 5x103 or 7x101 embryogenicmicrospores were plated on Transweli
50

Interaction 2,4-D and rnannïto!
polycarbonate filters with pore size 12 un(Costar). At 10 d.a.i., the filters were cut out oftheir frame, to enable culture of embryo-likestructures (ELS) in 1 mL liquid medium. Theploidy level of representative plants wasdetermined by How cytometric analysis of therelative nuclear DNA content.
Anther cultureThirty anthers (equivalent to 1/2 spike) of severalspikes were randomnized used per parameter.There was no need for the use of filters.
The effect of the concentration and the timethat 2,4-D was present in the medium, was studiedin relation to both the ability of the microspores todevelop into ELS and the number of plantssubsequently formed (result section A). Taldng thebest variables found for plant regeneration, thecapacity to develop plants of 5x103 embryogenicmicrospores was assessed on the first 500developing ELS after careful transfer wiffi forcepsto medium III (Hoebtra et al. 1992) containing0.4 mol /1. IGnetin. The regene-ration effidency ofmicrospore culture with the continuous presence of4x106 moVL 2,4-D in the above mentionedmodified medium 1 (without the use of filters), andapplicauon of 4x106 Mol /L 6-BAP according toHoekstra et al. (1993), was compared (resultsection 8). The effect of 2,4-D with omittance ofmannitol pretreatment was tested at 106 to i0mol/T. 2,4-D and different time periods of itspresence in both anther and microspore culture(result section C).
No standard deviationis given in the figures,since doe to differencesin microspore developmental stage and platingdensity, the absolutenumbers varied.However the pattems ofthe figures were reprodudbie, throughout 4independent experiments.
I&esuks
A. Effect of 2,4-D conditions after mannitolpretreatment on plant production
Microspore cultureIn order to remove 2,4-D from the culturemedium, microspores were cultured on afilter. The optimal density for cukureefficiency of microspores on filter wasassessed (Table 1). At a higher microsporedensity, a higher percentage of microspores developed into ELS. The regenerationinto plants however, was maximal when3x103 microspores were present on thefilter. The latter density was at thethreshold level enabling development ofELS (indicated by the high standarddeviations). In subsequent experimentsthe microspores were cultured at a densityof 5x103 and 7x103 per filter.
The influence of continuous presence of2,4-D on the development of microsporesinto plants was tested for theconcentration range as indicated in figure1. The size of the ELS was reduced at 1 Omoi/L or higher. At these concentrations
Table 1: The influence of various microspore densities, grmvn on a filter, onthe culture effidency. Culture condiüons were 10.6 moVL 2.4-D continuouslypresent, in microspore culture with pretreatment.
microspore density on filter
3x103 5x103 7x103 iO
(ELS/density) 0.67±0.94 3.74±1.33 4.96±1.85 5.74±1.14
xlOO
(plants’ELS) 7.50±14.9 5.50±1.06 0.15±0.54 0.09±0.61
x 100
51
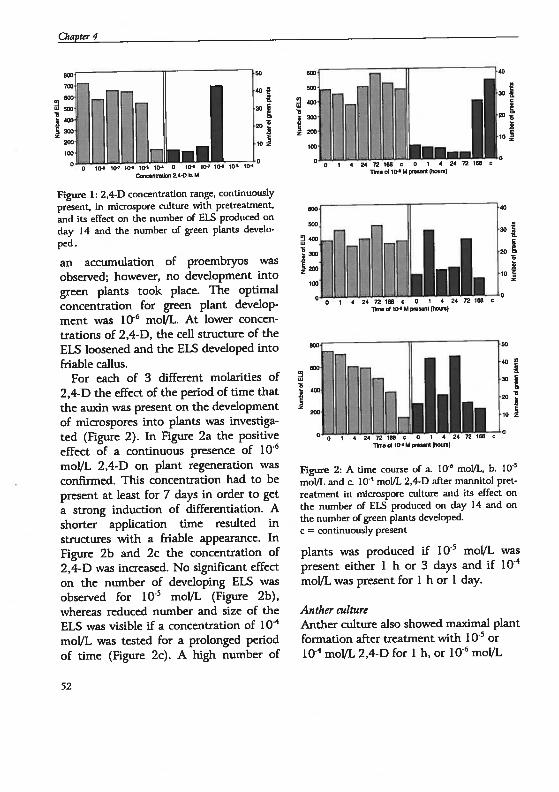
Chapter4
0001 1 50
40î
JiL..I I0 10-0 50-0 15* 15-0 55-0 0 10-0 100 10* 100 10
Conoofltmt6fl 2,4.0 in M
Figure 1: 2,4-D concentration range, continuously
present, in microspore culture with pretreatment,
and its effect on the number of ELS produced on
day 14 and the number of green plants develo
ped.
an accumulation of proembxyos wasobsewed; however, no development intogreen plants took place. The optimalconcentration for green plant development was 106 moVL. At lower concentraüons of 2,4-D, the ceil structure of the
ELS loosened and the ELS developed intofriable callus.
For each of 3 different molarities of2,4-D the effect of the period of time thatthe auxin was present on the developmentof microspores into plants tvas invesügated (Figure 2). In Figure 2a the positiveeffect of a contïnuous presence of 1 06
mol/L 2,4-D on plant regeneration wasconfirmed. This concentration had to bepresent at least for 7 days in order to geta strong induction of differentiation. Ashorter application time resulted instnictures with a friable appearance. Infigure 2b and 2c the concentration of2 ,4-D was increased. No significant effecton the number of developing ELS was
obsetved for I0 moVL (figure 2b),whereas reduced number and size of theELS was visible ïf a concentration of 1 0mol/L was tested for a prolonged periodof time (Figure 2c). A high number of
600
500
!1 400
300
100
42472100
13n,e of Is-’ M pmoont (hosro)
50
-
40
L
-—0 0 1 4 2472168
0
r.,no of 15-°U p0000flI (60010)
figure 2: A time course of a. 106 mol/L, b. iO
motiE and c. iO motiE 2.4-D after mannîtol pret
reatment in microspore cuhure and its effect on
the number of ELS produced on day 14 and on
the number of green plants developed.
c = conunuously present
plants was produced 1f iO moVL waspresent either 1 h or 3 days and if lO
moVL was present for 1 h or 1 day.
Anther cultureAnther culture also showed maximal plantformatïon after treatment with 1 o- orlO mol/L 2,4-D for 1 h, or 10.6 mol/L
1 4 0-.
Ton, of 10-0 M prooont (hoors)
II40
30 1
20 S
10
0
1 400
601
200
00
1
52

Interaction 2.4-D and mannital
resulted in similarfrequendes of bothgreen and whiteplants; dependïng onthe developmentalstage and cultureconditions used,between 25 and 50green plants peranther were obtained.
continuously. However, the time of 2 ,4-Dpresence required for plant formation, isless restricted than in microspore culture(data not shown).
B. Plant production wit); 2,4-D under optimalcondihons, after mannitol pretreatntentIn A. several conditions were clearlysuboptimal for regeneration. for the best5 parameters as obtained in A., thenumber of regenerated plants upon 2,4-Dtreatment was determined in microsporeculture under optimal regenerationconditions. No significant difference inplant production was found betweenthese various culture conditions tested(see Table 2). further, of 59 regenerantsabout 23% was haploid, 71% had thediploid level and their fertility (85%) wassimilar to the donor material. No changesin gross morphology of plants werevisible, and no event of somaclonalvariation was found by barley breeders infield-grown seed progeny from pretreatedand subsequently 2,4-D treated microspores.
A comparison of microspore culture inthe presence of 4x 10.6 mol/L 2,4-D withthe regularly applied 4x106 mol/t BAP
C. Effect of 2,4-D amditions without mannitolpretreatment on plant production
Microspore cuttttreWithout mannitol pretreatment, noembryogenic type of microspores(Hoekstra et al., 1993) can be recognisedat the moment of isolaüon. Even whenthe plating density was adjusted to 2.5-3x 1 0 microspores per filter, the plaüngeffidency did not reach 1%. This resultwas also not improved either, when theoptimal parameters for anther cukurewithout pretreatment were applied inmicrospore culture.
Anther cutturefor each of 3 different molarities of 2,4-D, the effect of the period of time thatthe auxin was present on the developmentof microspores into plants was investigated with omittance of the mannitolpretreatment (figure 3). Determinationof average ELS production was unreliable14 days after anther ctilture. Therefore,the numbers of ELS were counted afterapproximately 25 days, and werethereafter plated on plant regenerationmedium. Growth of microspores was
Table 2: Groen plant production in microspore ctilture with pretreatment,
for 5 different 2,4-D regimes. c = continuously present
Concentsation and time present of 2,4-D
lO°M 1O5M tO1M
c. 1h 3d 1h Id
n plants per anther 20.3 12.9 14.9 14.7 15.8
53
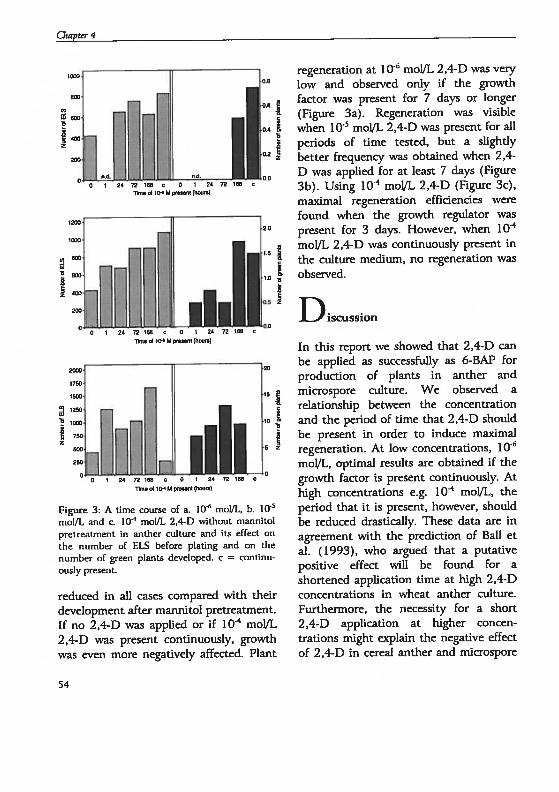
Diapter 4
00
06
04
0.2
— d 0.00 1 24 72 168 0 0 1 24 72 166 0
1)oo oI 10 U p,0680t (hoor,)
Figure 3: A time course of a. iO moVL, b. lOmol/L and c. 10° mol/L 2,4-D without mannitolpretreatment in anther culture and its effect onthe number of ELS before plating and on thenumber of green plants developed. c = continuously present.
reduced in all cases compared with theirdevelopment after mannïtol pretreatment.1f no 2,4-D was applied or 1f 10 moVL2,4-D was present continuously, growthwas even more negatively affected. Plant
regenerafion at 10° mol/L 2,4-D was veiylow and observed only if the growthfactor was present for 7 days or longer(Figure 3a). Regeneraüon was visiblewhen 1 O moVL 2 ,4-D was present for allperiods of time tested, but a slightlybetter frequency was obtained when 2,4-D was applied for at least 7 days (Figure3h). Using i0 moVL 2,4-D (Figure 3c),maximal regeneration efficiencies werefound when the growth regulator waspresent for 3 days. However, when 1 0
mol/L 2,4-D was conünuously present inthe culture medium, no regeneration wasobserved.
In this report we showed that 2,4-D canbe applled as successfully as 6-BAP forproduction of plants in anther andmicrospore culture. We observed arelationship between the concentrationand the period of time that 2,4-D shouldbe present in order to induce maximalregeneration. At low concentrations, 10.6
moVL, optimal resuks are obtxined if thegrowth factor is present conünuously. Athigh concentrations e.g. iO mo]/L, theperiod that it is present, however, shouldbe reduced drastically. These data are inagreement with the prediction of Bail etal. (1993), who argued that a putativepositive effect will be found for ashortened applicaüon time af high 2,4-Dconcentrations in wheat anther cukure.furthermore, the necessity for a short2,4-D application at higher concentrations might explain the negative effectof 2 ,4-D in cereal anther and microspore
1686
872
Sz
200
S
2.0
01.5
S1.0
S55g
_L_, iscussion00
72168 0 0 1 24 72168 0
Tooo 01 l0’U p,6840I (hors)
00 1 24
20202011750j
151500J
01250
‘hL uh :‘1680 1 24 72
Too0 0115054 p,0100l (hoor,)
S 24 72 168
54

Interaction 2,4-D and mannitol
culture, reported by Kasha et al. (1990),
Tiwari and Rahimbaev (1991) andHassawi et al. (1990). An altemativeapproach to prevent inhibition of embryo
maturaüon by prolonged awdn presence,
is the use of IAA, the easily degradable
natural auxin (Datta and Potxykus,
1989).The method of 2,4-D applicaüon for
plant production in barley anther and
microspore culture, is comparable to its
use for somaüc embryrogenesis. At low
concentrations, or if 2,4-D was not long
enough present, unorganised tissue was
generated. When app. iO moVL 2,4-D
present for several days, embryogenesis is
induced in various seed, seedling andplant tissues for carrot, cucumber, Pennise
tum and Asparagus (resp. Dudits et al.,1991; Tabei et al., 1991; Talwar and
Rashid, 1990; Levi and Sink, 1991). The1 h putse treatment with 10 mot’t 2,4-
D in dicotyledonous microcallus of affalfa
resulting in morphogenesis (Dudits et al.,1991), gave, in our hands similar resuits
in the monocotyledonous barley
microspores that were pretreated on
mannitol. When the synthetic auxin 2,4-D was applied at high concentration
during a prolonged period, hampered
growth of ELS was obsewed, togetherwith complete inhibitïon of embryo
maturation and gennination (this could
be due to high endogenous 2,4-D levels).
Dudits et al. (1991) states that embryogenesis occurs in the presence of 2,4-D, atconcentrations that already inhibit thegrowth of callus tissues, and thishypothesis is confinned by our resuits.
A comparison between anther and
mÏcrospore cukure has been perfonned
earlier in barley, and some differences aredescribed (Hoekstra et al., 1992). Upon
2,4-D application in anther andmicrospore culture, a high similarity was
observed. Furthennore in anther culture,
the presence of 2,4-D in mannitol wasinvestigated. No significant difference in
effidencv of plant production was found,
whether 2,4-D was apptied during, after,
or both durlng and after pretreatment
(data not shown).Comparison of the pattem of
regeneration with and without mannitol
pretreatment in anther culture, reveals
high similarity. There are two remarkable
differences however. The number of
plants produced was strongly reduced
without pretreatment. In addition the
pattem in the figures of 2,4-Dconcentration and time period of itspresence required for regeneration was
shifted. Without pretreatment, microspores only in the presence of anther
tissue, can to a limited extent produceplants upon application of a higher andlonger presence of 2,4-D. Non-pretreatedisolated microspores showed an extremelydeclined level of plating effidency upon2,4-D treatment, not enabling plantformation. After application of a certainshock treatment (e.g. sugar starvation,cold, high 2,4-D concentration), theanther tissue apparently acquires thecapacity to mediate a swkch from the
gametophytic to the sporophytic pathway. We suppose that in anther culture
with and without, and in microsporeculture only with anther pretreatment,another inductive trigger for differen
55

Chapter 4
tiation is active besides exogenous applied2,4-D. To our knowledge, all papers onmicrospore-derived (cereal) plant production describe a certain pretreatment. Thepretreatment of barley microspores can bea cold period of spikes (Huang andSunderland, 1982), or mannitol starvation of anthers (Roberts-Oehischiager etal., 1990). During both treatments themicrospores are surrounded by the anthertissue. We hypothesize that in antherculture and in cultiire of pretreatedmicrospores, the trigger for induction ofdifferentiation is mediated by more thanone signal. The highest efficiency of plantproduction is achieved by application of asignal strength that is optimal for aspecific celi competence. fundamentalresearch in this field will contribute tounravelingdevelopment.
induction of plant
AcknowledgementsThe authors wish to thank dr. F. VanderMark fordiscussions, M.H. VanZijderveld for theperformance of pilot studies, J.M. Vink for growingthe plants and P. Hock for the layout of thefigures.
I..eferencesAbdullah R, E.C. Cocking and J.A. Ihompson.Effident plant regeneration from rice protoplaststhrough somatic embryogenesis. Biotechnol. 4:1087-1090 (1986)Bali S.T., H.P. Zhou, and C.F. KonzakIniluence of 2,4-D, IAA, and duration of callusinducüon in anther ctiltures of spring wheat. PlantSci. 90: 195-200 (1993)Datta S.IC and 1. Potrykus. Artificial seeds inbarley: encapsulation of microspore-derivedembryos. Theor. Appi. Genet. 77: 820-824 (1989)Dudits D., L. Bogre and J. Gyorgyey. Molecularand cellular approaches to the anaiysis of plant
embiyo development from somatic cells in vitro. J.Celi Sci. 99: 475-484 (1991)Hassawi D.S., J. Qi and G.H. Liang. Effects ofgrowth regulator and genotype on production ofwheat and triticale polyhaploids from antherculture. Plant Breed. 104: 40-45 (1990)Hoekstra S., M.H. VanZijderveld, J.D.Louwerse, F. Heidekamp and F. VanderMarkAnther and microspore cutture of Hordeum vulgare
L. cv. Igri. Plant Sci. 86: 89-96 (1992)Hoekstra S., M.H. VanZijderveld, F.Heidekamp and F. VanderMark. Microsporeculture of Hordeum vulgare 1.: the influence ofdensity and osmolality. Plant Ccli Rep. 12: 661-665 t 1993)Huang 8. and N. Sunderiand. Temperaturestress in barley anther culture. Ann. Bot. 49: 77-88(1982)Kasha KJ., A. Ziauddin, E. Simion, A.A.Marsolais and C. Yu Rong. Barley and wheatnücrospore cuiture. International Assodation ofPlant flssue Culture 1990: hand-out.IComamine A., M. Matsumoto, M. Tsukahara,A. fujiwara, R ICawahara, M. Ito, J. Smith, KNomura and T. fujimura. Mechanisms ofsomaüc embnyogenesis in cdl cultures -
physiology, biochemistry and molecular biology
pp. 307. In: Progress in plant cellular andmolecular biology, eds. Nijkamp, van der Plas andvan Aartrijk, Kluwer Academic Publishers (1990)Levi A. and KC. Sink Somatic embryogenesis inasparagus: the role of explants and growffiregulators. Plant Ccli Rep. 10: 7 1-75 (1991)Lührs R. and H. Lörz. Plant regeneration in vitrofrom embryogenic cultures of spring- and winter-type barley (Hordeum vulgare L) varieties. Theor.Appl. Genet. 75: 16-25 (1987)Morocz S., G. Donn, J. Nemeth and D. Dudits.An improved system to obtain fertiie regenerantsvia maize protoplasts isolated from a highlyembryogenic suspension culture. Theor. AppI.Genet. 80: 72 1-726 (1990)Olsen F.L. Induction of microspore embiyogenesisin cuitured anthers of Hordeum vidgare: the effectsof ammonium nitrate, glutamine and asparagine asnitrogen sources. Carisberg Research Comm. 52:393-404 (1987)Roberts-Oehischiager S.L. and J.M Dunwell.Barley anther cuiture: pretreatment On mannitol
56

Interaction 2,4-1) and manniut
stimulates production of microspore-derived
embryos. Plant Cel! ‘flss. Org. Cult. 20: 235-240
(1990)Tabel Y., T. Kanno and T. Nishio. Regulaüon of
organogenesis and somatic embrogenesis by auxin
in melon, Cucumis melo L. Plant Cel! Rap. 10: 225-
229 (1991)Talwar. M. and A. Raahid. factors affecting
formaüon of somatic embiyos and embsyogenic
callus from unemerged inilorescences of a
graminaceous crop Pennisetum. Puin. Bot. 66: 17-
21(1990)
Tiwari S. and 1. Rahimbaev. Effect of 2,4-
dichloro-phenoxyacefic add on barley anther
culture in liquid and agar media. Indian J. Genet.
51: 112-117 (1991)Vasil V., F. Redway and I.IC Vasil. Regeneration
of plants from embfyogenic suspension culture
protoplasts of wheat (Triticum aestivum L).Biotechnol. 8: 429-434 (1990)Zimmerman L. Somatic embryogenesis: A model
for early development in higher plants. Plant Cel!
5: 141 1-1423 (1993)
57


hapter 5
Androgenesis in Hordeum vutgare L.:
The role of ABA during anther pretreatment,
on a solution containing mannitol and calcium
S. Hoekstra, IR. van Brouwershaven, S. van Bergen, RA. Schilperoort & M Wang
immary
Pretreatment induced androgenesis of Hordeum vulgare L cv. Igti was investigated in order to find factors
which control optimal plant production. The influence of what is called pre-medium, and of the concentra
tion of both mannitol and calcium was studied. Increasing concentrations of each of the compounds, also
resulting in a higher osmolality, improved plant production. Optimal plant production was obtained upon
anther pretreatment on 30 mM CaCI, adjusted wiffi mannitol to 440 mOs.kg’. The rather high calcium
concentration and the observaüon that calcium could be replaced by potassium ions, suested a role of
these ions as osmotic agents. Since ABA is involved in severat developmental processes and is known to be
induced by osmotic stress we studied AB% and ABA, levels during pretreatment. Optimal plant production
was observed at a lower ABA1 concentration than the initial value. This suested a negaüve correlation
between androgenetic potential and ABA level, which became even more evident when the ABA, values
were considered. Our overall data indicated that both a defined ratio ABA,/ABA1 level was required
throughout pretreatment, but in particular in a period around 24 hours. for optimal microspore-derived
plant production. The mechanism of pretreatment for induction of androgenesis is discussed.
ntroduction
Integration of microspore-derived plants
in plant breeding is hampered by Jargegenotypic differences in anther response.
Of many barley genotypes, doubled
haploids can be produced via
androgenesis if three requirements are
met. first, the developmental stage of the
microspores should be at the verge ofmitosis (Hoekstra et al., 1992). Second,the right hormone treatment is necessaryfor induction of plant producüon. For
59

Chapter 5
barley this is achieved either by 6-benzylaminopurine (BAP) in culture medium(Olsen, 1987; Hoekstra et al., 1993) orby a spedfic 2,4-dichlorophenoxyaceticacid (2,4-D) application (Hoekstra et al.,1996). And third, the right pretreatmentof the anthers is needed in order to switchthe gametophytic pathway into asporophytic development of themicrospores. for barley, a pretreatmentcan be a cold treatment of spikes (Huangand Sunderland, 1982) or anthers can bepretreated on a mannitol solution(Roberts-Oehlschlager and Dunwell,1990). The latter pretreatment enhancesdoubled-haploid production in barley; themÏcrospore viability is increased and ahigher effidency of plant production isachieved (Hoebtra et al., 1994).Optimization of culture conditions, inparticular taking into account themicrospore developmental stage and theuse of growth reguLator, reduced thegenotype dependency of androgenesis(unpublished data, Hoekstra et al.).Knowledge about biological processesinvolved in effects of pretreatment, mightresult in the development of a genotypeindependent protocol for androgenesis.
Uttle is known about the processes thatare affected by the pretreatment. HeberleBors (1989) suggests that duringpretreatment, starvation due to theabsence of useful saccharides, is the basisfor the occurence of certain degradationprocesses in pollen grains, resuking in theinduction of pollen embryogenesis.Especially in tobacco the starvation effectis pronounced, if pretreatment isperformed in nitrogen-free medium withmannitol. Pretreatment of anthers from
barley, resulting in plant production, canbe performed using similar conditions asfor pollen preculture of tobacco, i.e. onmannitol dissolved in what is called a premedium. Mannitol is thought to createosmotic stress. The presence of the sugarsmannitol during pretreatment andmaftose during culture, are important tohave proper osmotic pressure formicrospore-derived barley plant production (Hoebtra et al., 1993). Fromliterature, e.g. Davies and Jones (1991), itis known that, in response to certainstress treatments like osmotic pressureand cold, ABA ïs produced. ABA-inducedgene expression by mannitol as osmoticagent is observed in e.g. ceIl suspensioncuftures (Leonardi et al., 1995). Inaddition, relaüvely high ABA levels arecorrelated with high embiyogenicpotential of Pennisetum and carrot cells(Rajasekaran et al., 1987a, b; Kiyosue etal., 1992). In the case of tobacco androgenesis, anthers are pretreated for 2 dayson 0.5 M mannitol or by treatment with1 O M ABA for 1 to 3 days. 1f tobaccoanthers, for the induction ofandrogenesis, are pretreated on 0.5 Mmannitol for 1 day, a 2-fold increase inABA concentration is measured(Imamura and Harada, 1980).
The other part of barley pretreatment isthe pre-medium, which predominantlyconsists of CaCl2, MgSO4 and I<N03.from literature it is known that caldumions play a role in embiyogenesis(Timmers et al., 1989; Overvoorde andGrimes, 1994; Jansen et al., 1990). Nosignificant influence of magnesium ionshas been reported for gametophyticembryogenesis (Reynolds, 1990).
60

Effects ofABA, mannitol and calcium
Potassium ions, however, stimulatesomatic embxyogenesis in alfalfa (Shettyand McKersie, 1993) and have been usedfor pretreatment induced androgenesis ofseveral spedes. for the production ofdoubled haploid plants of barley, rice andwheat, 10 to 20 mM KNO3 have beenused during mannitol pretreatment (resp.Wei et al., 1986; Ogawa et al., 1992; Huet al., 1995), whereas tobaccomicrospores are pretreated in mannitolwith 20 mM KC1 (Kyo and Harada,1986).
In this paper, the role of ABA in barleyandrogenesis as well as the antherpretreatment elements mannitol and premedium, was invesügated.
aterfals and Methods
MaterialsDonor plants of Hordcuiz imigare L. cv. Igri weregrown in a phytotron under conditions describedpreviously (Hoekstra et al., 1992). Monodonalantibody to free (+)ABA was purchased fromIdetek, mc. (San Bruno, Calif., USA). Rabbit anti.mouse alkaline.phosphatase conjugate, (±)ABAand bovine senim albumin (grade suitable forenzyme-linked immunosorbent assay ELISA) wereobtained from Sigma (St. Louis, Mo., USA).fluridone was kindly provided by the BuibResearch Laboratory, lisse, Netherlands.
AndrogenesisSelection of material, mannitol pretreatment andculture condilions were as described previously(Hoekstra et al., 1992) wiffi the followingmodifications. Anthers of several spikes wererandomly distributed over the parameters to betested and per parameter 30 ant.hers vere used.Anthers were pretreated on 0.37 M mannitoldissolved in pre-medium, containing 10.2 M CaCI2,10 M MgSO4.7H,O, l0 M KNO3, 2x104 M1G{,P04, 106 M 1(1 and lO M CuSO4.2 H20 (440mOs.kg’, by Osmomat), what is called
pretreatment solution. After 4 days the antherssvere t.ransferred to medium 1 without ficoil at 350mOs.kg’. The first 100 developing ELS werecarefully transferred with forceps to medium IIIand the rest was plated directly. at the momentthat the majority of ELS were at least 1 mm indiameter. After 3 weeks plant production wasassessed per group of 30 anthers.
The osmolality of the pretreatment solution wasadjusted with mannitot to values of 350, 440 and540 mOs.kg’. MgSO4 at 0,0.01,0.1, 1 mM and aconcentraüon range of CaC12 were tested in premedium. Omittance of CaCI2 was used in stead ofEGTA to overcome the weak calcium bufferingcapadty of the latter at the pH of pre-medium.KCI, KNO3 and Ca(N03)2 were tested at 10 and20 mM in modified pre-medium (with resp. 1 mMCa(N03)2, CaC12 and KCI in stead of KNO3). Premedium with 10, 30 and 40 mM CaCI2 wasadjusted to an osmolality value of 440 mOs.kg’with mannitol.
At defined Umes the pretreatment solution wasremoved and new pretreatment solulion was addedto the anthers. (±)2.cis-4-trans-ABA (Sigma) wasadded from a stock solution (4 mM in 5% ethanol)to obtain a range of concentraüons (in a volume ofmax. 25 d) at the start and removed at 24 hours ofpretreatment. Fluridone was added at aconcentration of 40 mg4 from the beginning andremoved at 24 hours of pretreatment.
Note: Data in the Tables and figures, arepresented in percentages of regeneration on premedium since the absolute numbers varied, due todifferences in microspore developmental stage andELS density during culture. The pattems of thefigures were reprodudbie, however, throughout atleast 3 experiments. In table 1 numbers ofregenerated plants are presented, to give aasindication of the absolute numbers of plantsobtained per 30 anthers.
ABA exfzactionAfter treatment, anthers and prei.reatment soluüonwere seperately frozen in liquid N2. Pestiemaceration of the frozen anthers was not necessaryfor ABA extracüon (unpublished results, M. Wanget al.). The contents of the Eppendorf vials werefreeze dried. The samples were extracted at 150rpm, 4C with cold methanol (containing 100 mg/lbutylated hydroxytoluene and 0.5 g/l dtric acid
61

Chapter 5
monohydrate). The anther samples were extractedtwice each time in 0.5 ml for 24 hours. Thepretreatment soluUon samples were treatedsimilarly, except that the first extraction wasperformed in 1.0 ml. The vials were centrifuged at
8,000g for 15 min at 4C and the supematants ofthe samples were pooled. The samples wereconcentrated in a Speed-vac system (Savant) andthe volume adjusted to 25 sl with methanol. Thesamples were stored al .80C until assay.
ABA ELISA assayThe assay of the amount of ABA in anthers and inpretreatment soluUon was carried Out by anenzyme-linked immunosorbent assay (ELISA) asdescribed by Wang et al. (1995). No interferencein the (+)ABA ELISA was observed by commerdalmannitol, in contrast to what is noted by Belefantand fong (1989).
Table 1: The influence on plant production ofdifferent mannitol concenlrations without premedium. during pretreatment. The results of oneexperiment are shown and expressed in number ofplants per 30 anthers. from other experimenissimilar data ivere obtained. Numbers in parentheses indicate the osmolality of the pretreatmentsolution (in mOs.kg’).
mannitol n plants
0 10 (0)
0.3 65 (310)
0.37 86 (400)
0.44 89 (500)
Table 2: The iniluence of Ca2, K, C1 and NO3-ion concentrations during pretreatment in premedium, on plant production. The number ofplants obtained per 30 anthers. are expressed inpercentages.
ions in mM n plants
Ca2 K Cl NO3
10 1 20 1 100
20 1 40 1 114±33
10 1 1 20 123±49
20 1 1 40 106±37
1 10 tO 2 80±17
1 20 20 2 81±51
1 10 1 10 84±37
1 20 1 20 74±30
suits
Pretreatment solution elernentsfor optimal doubled haploid plantproduction of barley, a pretreatmentsolution consisting of mannitol and premedium, was used. The influence of theseelements on doubled haploid productionwas tested. Without pretreatment, nodoubled haploid plants can lie produced(Hoekstra et al., submitted). Antherpretreatment on water induced veiylimited plant production. The efficiencyof plant production was significantlyimproved, if mannitol was present duringpretreatment. A higher mannitol concentration accompanied by a higherosmolality of the pretreatment solution,resulted in an increase of green plantproduction upto a concentration of 0.37M mannitol. At higher mannitol concentration the number of plant producedremained constant (Table 1).
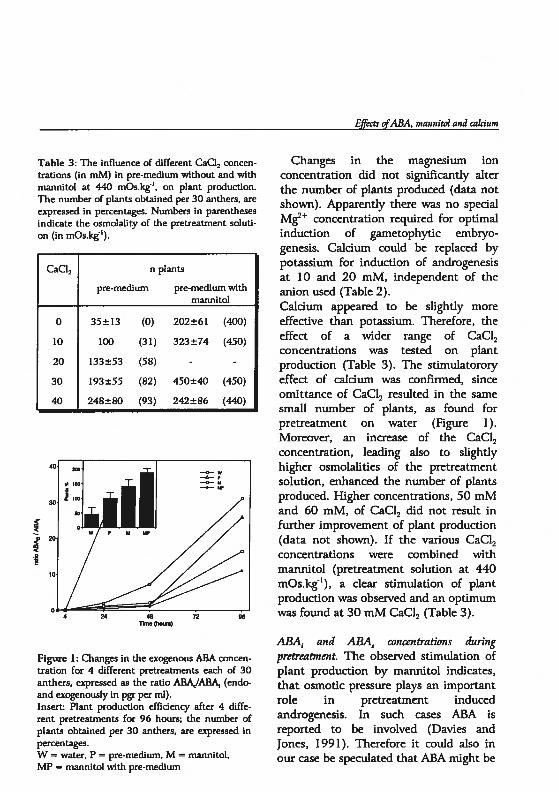
Effects ofABA. mannitol and cakzum
Table 3: The influence of different CaCI2 concenIrations (in mM) in pre-medium without and withmannitol at 440 mOs.kg’, on plant production.The number of plants obtained per 30 anthers, areexpressed in percentages. Numbers in parenthesesindicate t1e osmolality of the pretreatment solution (in m0s.kg’).
CaCI2 n plants
pre-medium pre-medium withmannitol
0 35±13 (0) 202±61 (400)
10 100 (31) 323±74 (450)
20 133±53 (58) - -
30 193±55 (82) 450±40 (450)
40 248±80 (93) 242±86 (440)—
Figure 1: Changes in the exogenous ABA concentration for 4 different pretreatments each of 30anthers, espressed as the ratio ABAJARA1 (endoand exogenously in pgr per ml).Insert: Plant production efficiency after 4 different pretreatments for 96 hours; the number ofplants obtained per 30 anthers, are expressed inpercentages.W = water, P = pre-medium, M = mannitol,MP = mannitol with pre-medium
Changes in the magnesium ionconcentration did not significantly alterthe number of plants produced (data notshowit). Apparently there was no specialMg2 concentration required for opümalinduction of gametophytic embryogenesis. Caldum could be replaced bypotassium for induction of androgenesisat 10 and 20 mM, independent of theanion used (Table 2).Calcium appeared to be slightly moreeffective than potassium. Therefore, theeffect of a wider range of CaC12concentrations was tested on plantproduction (Table 3). The stimulatororyeffect of calcium was confinned, sinceomittance of CaC12 resuked in the samesmall number of plants, as found forpretreatment on water (figure 1).Moreover, an increase of the CaCI2concentration, leading also to slightlyhigher osmolalities of the pretreatmentsolution, enhanced the number of plantsproduced. Higher concentrations, 50 mMand 60 mM, of CaCl2 did not result infurther improvement of plant production(data not shown). 1f the various CaC12concentrations were combined with
mannitol (pretreatment soluüon at 440mOs.kg’), a dear sümulation of plantproduction was observed and an optimumwas found at 30 mM CaCl2 (Table 3).
ABA and AB4 concenfrations duringpretreatment. The observed stimulation ofplant production by mannitol indicates,that osmotic pressure plays an importantrole in pretreatment inducedandrogenesis. In such cases ABA isreported to be involved (Davies andJones, 1991). Therefore it could also inour case be speculated that ABA might be
48To (howS)
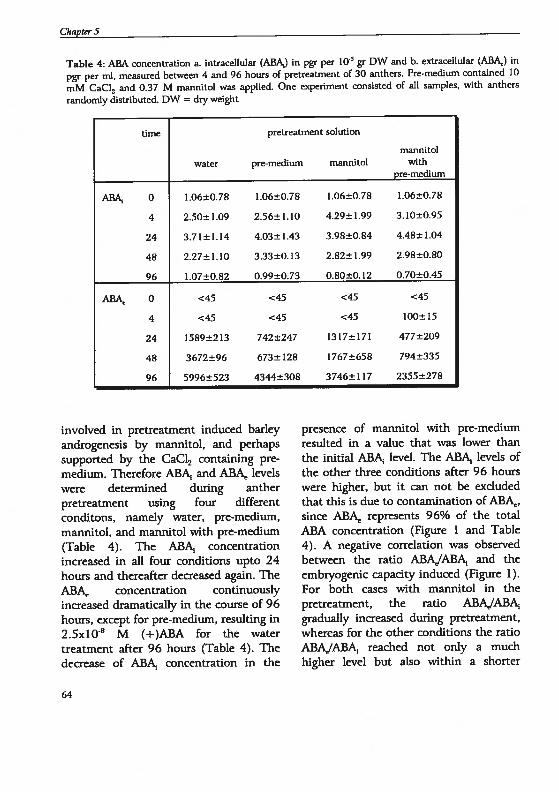
Chaprer S
Table 4: ABA concentration a. intracellular (ARA,) in pgr per i0 gr DW and Ii extracellular (ABA) in
pgr per ml, measured between 4 and 96 hours of pretreatment of 30 anthers. Pre-medium contained 10
mM CaCI2 and 037 M mannitol was applied. One experiment consisted of all samples, with anthers
randomly distributed. DW = dry weight
time pretreatment solution
mannitolwater pre-medium mannitol with
pre-medium
AllA, 0 1.06±0.78 t.06±0.78 1.06±0.78 1.06±0.78
4 2.50±1.09 2.56±1.10 4.29±1.99 3.10±0.95
24 3.71±1.14 4.03±1.43 3.98±0.84 4.48±1.04
48 2.27±1.10 3.33±0.13 2.82±1.99 2.98±0.80
96 1.07±0.82 0.99±0.73 0.80±0.12 0.70±0.45
ARk 0 <45 <45 <45 <45
4 <45 <45 <45 100±15
24 1589±213 742±247 1317±171 477±209
48 3672±96 673±128 1767±658 794±335
96 5996±523 4344±308 3746±117 2355±278
involved in pretreatment induced barleyandrogenesis by mannitol, and perhapssupported by the CaCI2 containing premedium. Therefore ABA and ABA levelswere determined during antherpretreatment using four differentconditons, namely water, pre-medium,mannitol, and mannitol with pre-medium(Table 4). The ABA concentrationincreased in all four conditïons upto 24hours and thereafter decreased again. The
ABAe concentration continuouslyincreased dramatically in the course of 96hours, except for pre-medium, resulting in2.5xlO5 M (+)ABA for the watertreatment after 96 hours (Table 4). Thedecrease of ABA1 concentration in the
presence of mannitol with pre-mediumresulted in a value that was lower thanthe initial ABA level. The ABA levels ofthe other three conditions after 96 hourswere higher, but it can not be exdudedthat this is due to contamination of ABA,since ABA,, represents 96% of the totalABA concentraüon (figure 1 and Table4). A negative correlation was observedbetween the ratio ABAJABA and theembtyogenic capadty induced (Figure 1).for both cases with mannitol in thepretreatment, the ratio ABA5/ABA5gradually increased during pretreatment,whereas for the other condkions the ratioABA,/ABA reached not only a muchhigher level but also within a shorter
64
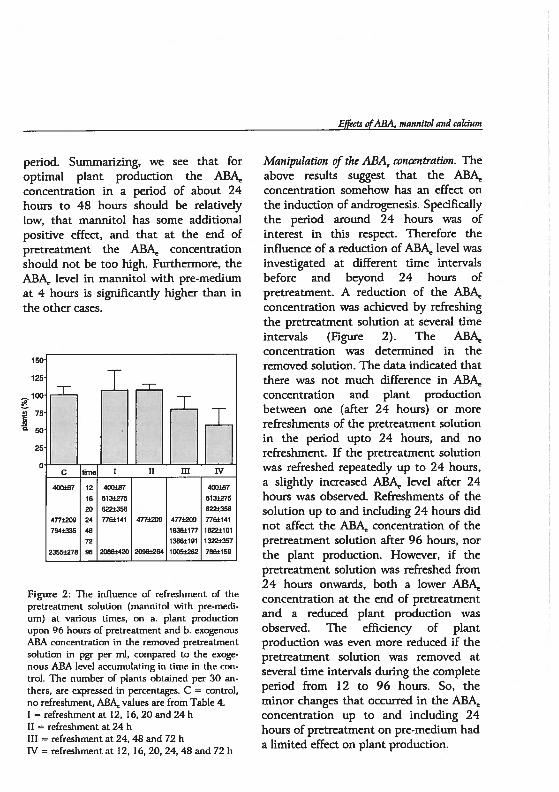
fffects ofABA, mannitol and caldurn
period. Summarizing, we see that foroptimal plant production the ABAconcentration in a period of about 24hours to 48 hours should be relativelylow, that mannitol has some additionalpositive effect, and that at the end ofpretreatment the ABA concentrationshould not be too high. Furthermore, theABAe level in mannitol with pre-mediumat 4 hours is significantly higher than inthe other cases.
400±87 12 400±87 400±87
16 613±275 613±275
20 622±356 622±356
477±209 24 776±141 477±209 477±209 770±141
754±335 48 1938±177 1822±101
72 1366±191 1322±357
2355±278 96 2086±420 2098±264 1005±262 786±159
figure 2: The influence of refreshment of thepretreatment solution (mannitot with pre-medi
um) at various Umes, on a. plant productionupon 96 hours of pretreatment and b. exogenousABA concentrauon in the removed pretreatmentsolution in pgr per ml. compared to the exogenous ABA level accumulating in time in the control. The number of plants obtained per 30 anthers. are expressed in percentages. C = control,no refreshment, ABA values are from Table 4.1 = refreshment at 12, 16, 20 and 24 hII refreshment at 24 hIII = refreshment at 24, 48 and 72 hW = refreshment at 12, 16,20,24,48 and 72 h
ManipuÏation of the AB4 contentration. Theabove results suggest that the ABAconcentration somehow has an effect onthe induction of androgenesis. Specificallythe period around 24 hours was ofinterest in this respect. Therefore theiniluence of a reduction of ABA,, level wasinvestigated at different time intervalsbefore and beyond 24 hours ofpretreatment. A reduction of the ABA,,concentraüon was achieved by refreshingthe pretreatment solution at several timeintervals (figure 2). The ABA,,concentration was determined in theremoved solution. The data indicated thatthere was not much difference in ABA,,concentration and plant productionbetween one (after 24 hours) or morerefreshments of the pretreatment soluüonin the period upto 24 hours, and norefreshment. 1f the pretreatment solutionwas refteshed repeatedly up to 24 hours,a slightly increased ABA,, level after 24hours was observed. Refteshments of thesolution up to and including 24 hours didnot affect the ABA,, concentration of thepretreatment solution after 96 hours, northe plant production. However, if thepretreatment solution was refreshed from24 hours onwards, both a lower ABA,,concentration at the end of pretreatmentand a reduced plant production wasobserved. The effidency of plantproduction was even more reduced 1f thepretreatment solution was removed atseveral time intervals during the completeperiod from 12 to 96 hours. So, theminor changes that occurred in the ABA,,concentration up to and including 24hours of pretreatment on pre-medium hada limited effect on plant production.
150
125
— 100
is 75‘0
50
25
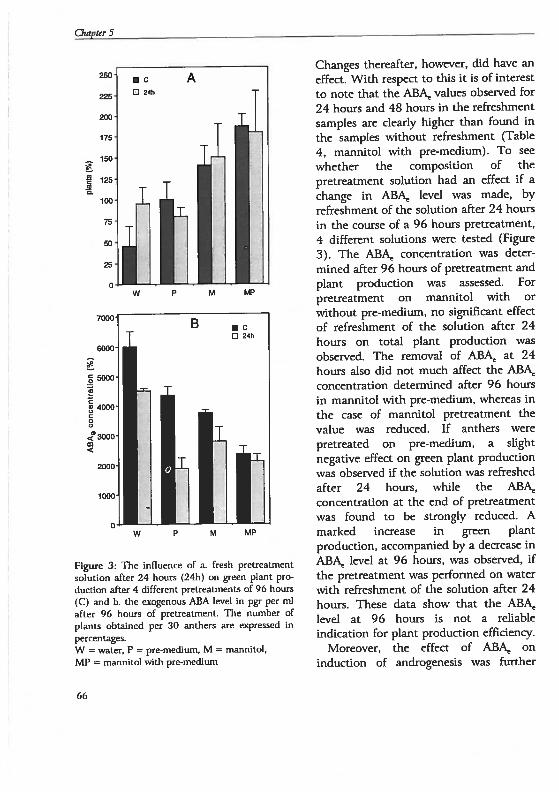
Chpkr5
Changes thereafter, however, did have an
effect. With respect to this It is of interest
to note that the ABAe values observed for
24 hours and 48 hours in the refreshment
samples are clearly higher than found in
the samples without refreshment (Table
4, mannitol with pre-medium). To see
whether the composition of the
pretreatment solutïon had an effect if a
change in ABAe level was made, byrefreshment of the solution after 24 hours
in the course of a 96 hours pretreatment,
4 different solutions were tested (figure
3). The ABAe concentration was determined after 96 hours of pretreatment andplant production was assessed. forpretreatment on mannitol tvith or
7000 without pre-medium, no significant effect• c of refreshment of the solution after 24
6000
D 245hours on total plant production wasobseived. The removal of ABA at 24
5000hours also did not much affect the ABAeconcentration determined after 96 hours
4000in mannitol with pre-mediun, whereas inthe case of mannitol pretreatment thevalue was reduced. 1f anthers werepretreated on pre-medium, a slight
2000negatïve effect on green plant productionwas observed if the solution was refreshed
1000after 24 hours, while the ABAconcentration at the end of pretreatment
0was found to be strongly reduced. A
w marked increase in green plantproduction, accompanied by a decrease inABA, level at 96 hours, was observed, ifthe pretreatment was performed on waterwith refreshment of the solution after 24hours. These data show that the ABAelevel at 96 hours is not a reliableindication for plant production efflciency.
Moreover, the effect of ABAe oninduction of androgenesis was further
250 mc
225 0 245
200
175
— 150
125t0.
100
75
50
B
figure 3: The influence of a. fresh pretreatmentsolution after 24 hours (24h) on green plant production after 4 different pretreatrnents of 96 hours(C) and b. the exogenous ABA level in pgr per ml
after 96 hours of pretreatinent. The number ofplants obtained per 30 anthers are expressed inpercentages.W = water, P = pre-medium, M = mannitol,MP = mannitol with pre-medium
66
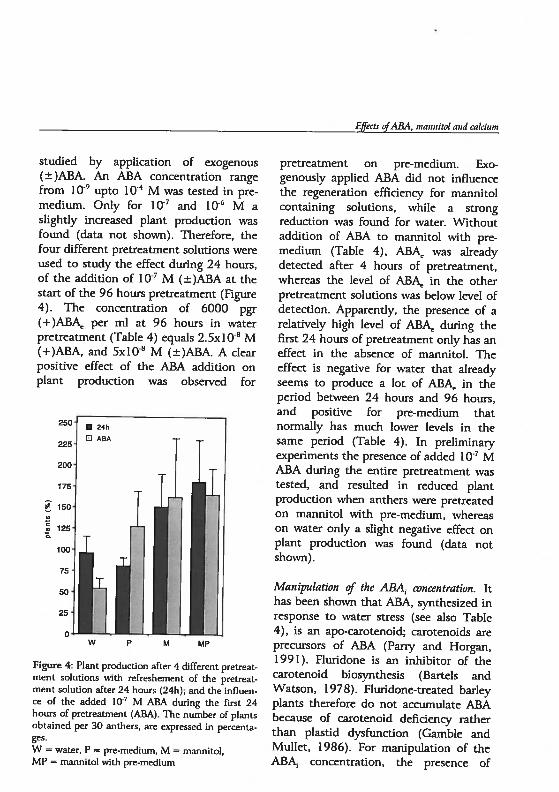
fffects ofABA, n,an,,itot and cakiurn
studied by application of exogenous(±)ABA. An ABA concentratïon rangefrom 1 o upto 1 o M was tested in premedium. Only for iO and 10b M aslightly increased plant production wasfound (data not shown). Therefore, thefour different pretreatment solutions wereused to study the effect during 24 hours,of the addition of io M (±)ABA at thestart of the 96 hours pretreatment (figure4). The concentration of 6000 pgr(+)ABAe per ml at 96 hours in waterpretreatment (Table 4) equals 2.5x108 M(+)ABA, and 5x108 M (±)ABA. A dearpositive effect of the ABA addition onplant production was observed for
Figure 4: Plant production after 4 different pretreatment solutions th refreshement of the pretreatment solution after 24 hours (24h); and the iniluence of the added io M ABA during the first 24hours of pretrealnient (ABA). The number of plantsobtained per 30 anthers, are expressed in percentages.W = water, P = pre-medium, M = mannitol,MP = mannitol with pre-medium
pretreatment on pre-medium. Exogenously applied ABA did not infiuencethe regeneration efflciency for mannitolcontaining solutions, while a strongreduction was found for water. Withoutaddition of ABA to mannitol with premedium (Table 4), ABAC was alreadydetected after 4 hours of pretreatment,whereas the level of ABAC in the otherpretreatment solutions was below level ofdetection. Apparently, the presence of arelatively high level of ABAe during thefirst 24 hours of pretreatment only has aneffect in the absence of mannitol. Theeffect is negative for water that alreadyseems to produce a lot of ABAC in theperiod between 24 hours and 96 hours,and posïtive for pre-medium thatnormally has much lower levels in thesame period (Table 4). In preliminaiyexperiments the presence of added 1 O’ MABA during the entire pretreatment wastested, and resulted in reduced plantproduction when anthers were pretreatedon mannitol with pre-medium, whereason water only a slight negative effect onplant production was found (data notshown).
Man ipulation af the ABA crnwenfration. Ithas been shown that ABA, synthesized inresponse to water stress (see also Table4), is an apo-carotenoid; carotenoids areprecursors of ABA (Pany and Horgan,1991). Fluridone is an inhibitor of thecarotenoid biosynthesis (Bartels andWatson, 1978). Fluridone-treated barleyplants therefore do not accumuiate ABAbecause of carotenoid deficiency ratherthan plastid dysfimdion (Gambie andMullet, 1986). for manipulation of theABA concentration, the presence of
w p M MP
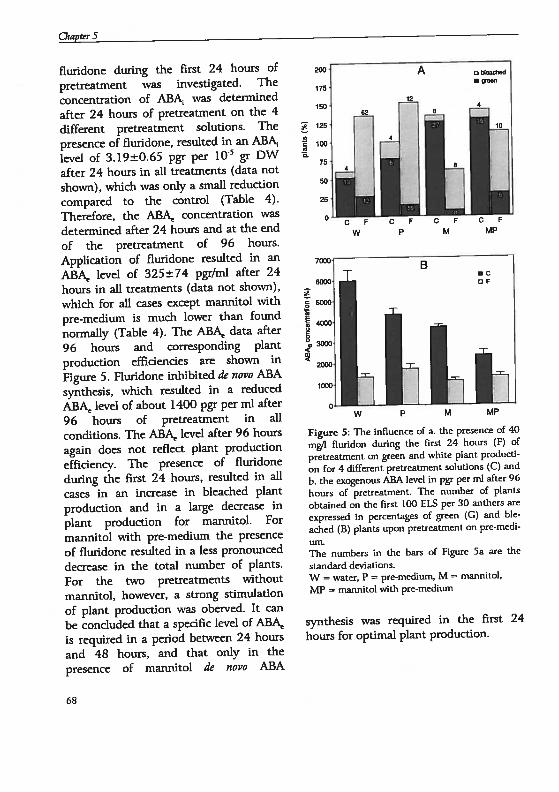
Chapter 5
fiuridone during the first 24 hours of
pretreatment was investigated. The
concentration of ABA was determined
after 24 hours of pretreatment on the 4
different pretreatment solutions. The
presence of fiuridone, resulted in an ABA
level of 3.19±0.65 pgr per iO gr DW
after 24 hours in all treatments (data not
shown), which was only a small reduction
compared to the control (Table 4).
Therefore, the ABA0 concentration was
detennined after 24 hours and at the end
of the pretreatment of 96 hours.
Application of fluridone resulted in an
ABA0 level of 325± 74 pgr/ml after 24
hours in all treatments (data not shown),
which for all cases except mannitol with
pre-medium is much lower than found
normally (Table 4). The ABA data after
96 hours and corresponding plant
production efficiencies are shown in
Figure 5. fluridone inhibited de novo ABA
synthesis, which resulted in a reduced
ABA0 level of about 1400 pgr per ml after
96 hours of pretreatment in all
conditions. The ABA0 level after 96 hours
again does not refiect plant production
effidency. The presence of flutidone
during the first 24 hours, resulted in all
cases in an increase in bleached plant
production and in a large decrease in
plant production for mannitol. for
mannitol with pre-medium the presence
of fiuridone resuked in a less pronounced
decrease in the total number of plants.
For the two pretreatments without
mannitol, however, a strong stimulation
of plant production was oberved. It can
5e conciuded that a specific level of ABA0
is required in a period between 24 hours
and 48 hours, and that only in the
presence of mannitol de navo ABA
200 A 0 bleadrod• groen
175
C F C F C t C t
W P M MP
7000 86000
5000
IEII:LIJ
Figure 5: The influence of a. the presence of 40
mg/l fluridon during the first 24 hours (F) of
pretreatment on green and white plant producti
on for 4 different pretreatment solutions (C) and
b. the exogenous ABA level in pgr per ml after 96
hours of pretreatment. The number of plants
obtained on the first 100 ELS per 30 anthers are
expressed in percentages of green (G) and bie
ached (B) piants upon pretreatment on pre-medi
05fl.
The numbers in the bars of Figtire 5a are the
standard deviations.W = water, P = pre-medium, M = mannitol,
MP = mannitol with pre-medium
synthesis was required in the first 24
hours for optimal plant production.
68

fffects OfABA, mannitol and calcium
A proper pretreatment is one of therequirements for doubled haploidproduction in barley. We identified twofactors in the pretreatment solution,mannitol and calcium, which were ofcrucial importance for efficient plantproduction. In this study, the optimalconcentration of calcium for plantproduction is 30 mM (in the presence ofmannitol), which is relatively high. Forandrogenesis in Solanum sp. the optimumcalcium concentration is 3 mM (in thepresence of 19 mM IC Reynolds, 1990).This author further reports that CaC12can be substituted by Ca(N03)2, and notby other bivalent ions such as e.g.magnesium. This is in agreement with ourresuks; our data showed moreover, thatCa2 could be replaced by K. This resultand the relaüvely high concentrationused, give an indication for a possible roleof these ions as osmotic stress agents likemannitol in the pretreatment. Howevercalcium might also have a spedficfunction in the induction of androgenesis.Other data from our laboratoiy indicatedthat programmed ceil death, also knownas apoptosis, takes place in the anthertissue during pretreatment (preliminaiyresuks). The key characteristic ofapoptosis is the systematic fragmentationand degradation of nuclear DNA (Rnff,1992). This DNA ladder is very oftencaused by the action of calciumdependent nucleases (Bowen, 1993).from our present data, we condude thatfor plant production an osmoticpretreatment of microspores (i.e. 0.3-0.4M mannitol in the presence of minimal10 mM Ca2 or K), apparently is a
general pre-condition for the speciesbarley (Wei et al., 1986), wheat (Hu etal., 1995), rice (Ogawa et al., 1992),Solanum carolinense L. (Reynolds, 1990)and tobacco (Imamura and Harada,1980; Kyo and Harada, 1986).Furthennore we assume, that differencesin optimal mannitol (and caldum)concentration per species can beexplained by differences in sensitMtytowards osmotic pressure. Our datasuggest that, the appropriate osmoticstress controls the optimal raüoABA,/ABA needed for embryogenicinduction in the microspores. Theimportance of ABA1 levels has beendetermined for morphogenic competencein Penniseturn sp. by Rajasekaran et al.(1 987a). furthermore, these authorsshow that applicaüon of ABA enhancessomatic embryogenesis, and thatinhibition of plant production is obsenredby addition of fluridone in the inductionmedium. The use of fluridone on donorplants even causes the loss of capadty forembryogenesis (Rajasekaran et al.,1 987b).
With regard to a correlation betweenandrogenetic capacity and ABA level, theliterature appears to be rathercontroversial. Johansson et al. (1982)report that appropriate pretreatment ofAnemona canadensis to induceandrogenesis, reduced the ABAconcentration at the end of pretreatmentnearly 4 times, whereas Imamura andHarada (1980) demonstrate the presenceof a peak in ABA concentration after 24hours of mannitol pretreatment intobacco anthers. Both data, nevertheless,are in agreement with our study, showinga peak in ABA level after 24 hours and a

Ojapter 5
reduction in ABA concentrafion at theend of the mannitol pretreatment after 96hours with or without pre-medium. Theseconditions were the best for plantproduction. In our study, a negativecorrelation between plant productionefficiency and ABA concentration wasmost pronounced 1f we looked at the
ABAe level and the raüo ABAJABA at theend of the pretreatment. The importancefor plant production of the ABAe leveland the ratio ABAJABA has not beenreported earlier. Imamura and Haradaalready proposed a spedfic level for ABAin 1980. These authors have shown in atime range experiment using 1 M ABA,that a 1 to 3 days period of pretreatmentstimulate plant production in tobaccoandrogenesis. for wheat androgenesis, Huet al. (1995) observe a stimulation byapplication of ABA (about iO M)throughout the pretreatment. Kyo andHarada (1985) show that applicaüon of5x10 M ABA in the second half of thepollen pretreatment stimulated embryoproduction in tobacco. from our data weconclude that, during the period between24 hours and 48 hours, specific ABAlevels both inside and outside the celimust be present.
in our investigation, the presence offiuridone during the first 24 hours ofpretreatment resulted in a large decreaseof the number of plants for mannitol andand a lower decrease for mannitol withpre-medium, indicating that normallythere is a need for de navo ABA synthesisduring the first 24 hours in the presenceof mannitol. So, a certain level of ABA1 isrequired for plant production. However,de nova ABA synthesis during the first 24hours of pretreatment was apparently not
required for optimal plant production if
pretreatment occurred in pre-medium orwater. Both data together suggest that inthe latter cases, after removal of
fluijdone, suffident ABA is synthesized to
reach the right level for optimal plantproduction. Probably this level must bereached in the period between 24 hours
and 48 hours. Fluridone is also known asa bleaching herbidde, inhibiting phytoenedesaturation in the presence of 7x 10.8 Mfluridone (Mayer et al. 1989). In ourstudy the number of bleached plantsindeed was much higher for fiuridone(about 1 O M) during pretreatment thanfor the controls.
The increase of the ratio ABAJABAduring anther pretreatment wasimportant for inductïon of androgenesis.The ratio ABAJABA can be changedeither by modification in (the rate of)ABA synthesis or by changes in themetabolism which resuk in reduction ofABA level. The presented data show thatde navo ABA synthesis is important duringthe first 24 hours of pretreatment, but itremains to be tested whether this is alsothe case at a more progressed stage ofpretreatment. In addition, changes ofABA catabolism could also be involved inthe regulation of the ratio ABAJABA andwere not investigated in this study.Differences in ABAe concentraüonsbetween treatments, also can be explainedby transport or difflision of ABi\ into thesolution. Altematively, the increase of
ABAe between 24 and 96 hours ofpretreatment, could be due to leakage ofABA from dying cells. Preliminaiy datafrom our laboratory support the lastpossibility. We found that duringpretreatment in mannitol with pre
70

Effects ofABA, mannitol and cakium
medium, anther tissue showed a highdegree of systematic DNA fragmentationand degradation after 72 hours, while theDNA had disappeared completely by 96hours. The microspores within this tissue,however, showed normal viability.Therefore we assume, that depending onthe conditons of pretreatment, ABAsynthesis is induced to a certain level thatit corresponds to the level of (osmotic)stress experienced by the ceil. Theconcentration of mannitol used in ourstudy correlates with the appropriatestress level needed for induction ofapoptosis (Cotter and Al-Rubeal, 1995)and probably protects the survivingmicrospores against a shock due to asudden too high level of ABAea such as inthe case of water pretreatment. The roleof calcium is to enable action of calciumdependent nudeases required for theprocess of apoptosis, thereby preventingthe cell’s contents (i.e. ABA in our study)spilled into the extracellular space (Raff,1992).
Our presented data indicated that ABAis also acting from outside the cefi, in casuon the microspores. There has been muchdebate as to the iniüal site of action ofABA elidting cellular responses, inparticular as to whether the ABA receptoris located in the cytosol (Assmann, 1994)or faces outward into the extracellularmatrix (Allan and Trewavas, 1994).Recently, in 2 systems evidence forextracullular action of ABA has beenreported. Wang et al. (1995) show thatdiffusion of ABA1 into the medium plays arole in germination of barley embryos. Inother morphogenetic responses as turionproduction in Spirodela polyrrhiza L., it isdemonstrated that ABA triggers a
morphogenic switch. In the latter system,ABA most likely interacts with aplasmalemma-located receptor system toinduce turion production (Smart et al.,1995). It is interesting to note that themorphogenetic switch for turionproduction, is induced by environmentaleffectors like nitrate deficiency and cold,which both also induce gametophyticembryogenesis. Apparently, the involvement of an ABA trigger is a more generalphenomenon and not restricted to themorphogenetic switch from thegametophytic to the sporophytic pathwayin pollen.
AcknowledgernentG. van Duijn (TNO, Zeist) is thanked for teachingthe Elisa, R van der Meulen for technicalassitance, J. Vink for growth of the donor planta,P. Hodc for the lay-out of the figures.
I...eferencesAllan C.A. and A.J. Trewavas. Abscisic add andgibberellin perception: inside or out? Plant Physiol.104: 1107-1108 (1994)Assmann S.M. Ins and outs of guard ccli ABAreceptors. Plant Ceil 6: 1187-1190 (1994)Bartels P.G. and C.W. Watson. Inhibition ofcarotenoid synthesis by fluridone and norfiurazon.Weed Sd. 26: 198-203 (1978)Belefant H. and F. Fong. Abscisic add ELISA:organic acid interference. Plant Physioi. 91: 1467-1470 (1989)Bowen I.D. Apoptosis or programmed ccli death?Ceil Biol. Int. 17: 365-380 (1993)Cotter T.G. and M. Al-Rubeai. Ceil death(apoptosis) in celi culture systems. Tibtech. 13:150-155 (1995)Davies W.J. and H.G. Jones (eds). In: AbscisicAcid physiology and iuiochemistry. Bio’sscientific publishers limiled (1991)Gamble P.E. and J.E. Mullet Inhibition ofcarotenoid accumulation and absdsic acidbiosynthesis in iluridone-treated dark.grown

Chapter 5
barley, Eur. J. Biochem. 160: 117-121 (1986)
Heberie-Bors E. Isolated pollen culture in
tobacco: plant reproductive development in a
nui.shell. Sex. Plant Reprod. 2: 1-10 (1989)
Hoekstra S., M.H. VanZijderveld, J.D.
1.ouwerse, f. Heidekamp and f.
VanderMark.Anther and microspore cuiture of
Hordeurn vulgare L. cv. Igri. Plant Sci. 86: 89-96
(1992)Hoekstra S., M.H. VanZijderveld, f.
Heidekamp and F. VanderMark. Microspore
culture of Hordeurn vulgare L.: the influence of
density and osmolarity. Plant CelI Rep. 12: 661-
665 (1993)Hoekstra S., S. VanBergen, I.R.
VanBrouwershaven, R.A. Schilperoort and F.
Heidekamp. The interaction of 2,4-D appilcation
and mannitol pretreatment in anther and
microspore culture of Hordeurn vulgare L. cv. Igri. J.Plant Physiol. 148: 696-700 (1996)Hoekstra S., MII. VanZijdervetd, S.
Vanflergen, f. VanderMark and F. Heidekamp.
Genetic modification of barley for end use quality.
In: Henry RJ. and JA. Ronalds (eds).Improvement of cereal quality by geneuc enginee
ring. Plenum Press pp 139-144 (1994)
Hu T.C., A. Ziauddin, E. Simion and Kj.
ICasha. Isolated microspore culture of wheat
(Trihcunt aestivurn L.) in a defined media 1. Effects
of pretreatment, isolation methods a n d
hormones. In Vitro Cell. Dev. Biol. 31: 79-83
(1995)Huang B. and N. Sunderland. Temperature
stress pretreatment in barley anther culture. Ann.Bot. 49: 77-88 (1982)Imamura J. and H. Harada Effects of absdsic
acid and water stress on the embryo and plantletproduction in anther culture of Nicotiana tabacurn
cv. Samsun. Z. Pilanzen- physio!. 100: 285-289
(1980)Ishitani M., T. Nakamura, S.Y. Han and T.
Takabe. Expression of the betaine aldehyde
dehydrogenase gene in barley in response to
osmotic stress and abseisic add. Plant Mol. Biol.
27: 307-315 (1995)Jansen M.A.I(, II. Booij, J.H.N. Schei and S.C.DeVries. Caldum increases the yield of somatic
embryos in carrot embryogenic suspension
cultuzes. Plant Celi Rep. 9:221-223(1990)
Johansson L., B. Andersson and T. Eriksson.
lmprovement of anther culture technique:
activated charcoal bound in agar medium in
combination with liquid medium and elevated C02
concentration. Physiol. Plant. 54: 24-30 (1982)
IGyosue T., M. Nakajima, I Yamaguchi, S. Satoh,
H. Kamada and H. Harada. Endogenous levels of
abscisic acid in embryogenic celis, nonembryogenic cells and somatic embryos of carrot
(Daucus carota L). Biochem. Physiol. Pilanzen 188:
343-347 (1992)Kyo M. and H. Harada. Studies on conditions
for ceil division and embzyogenesis in isolated
pollen culture of Nicotiana rustica Plant Physiol. 79:
90-94 (1985)Kyo M. and ii. Harada. Control of the
developmental pathway of tobacco pollen in vitro.
Planta 168: 427-432 (1986)Leonardi A., S. Heimovaara.Dijkstra and M.
Wang. Differential involvement of absdsic a c i d
in dehydraüon and osmotic stress in rice celi
suspension. Physiol. Plant. 93: 3 1-37 (1995)
Mayer P.M., D.L. Bartiett, P. Beyer and H.
Kleinig. The in vitro mode of action of bleaching
herbicides on the desturation of 15-ds-phytoene
and ds-C-carotene in isolated daffodilchromoplasts. Pestidde Biochem. Phys. 34: 111-
117 (1989)Monroy A.f. and R.S. Dhindsa. Lowtemperature signal transduction: induction of coid
acclimation-spedfic genes of alfalfa by caldum at
25°C. PlantCell 7:321-331(1995)Ogawa T., T. Hagio and Y. Ohkawa. Plant
regeneration from isolated pollen grains in indica
type rice (Oryza sativa L.). Japan J. Breed. 42: 675-
679 (1992)Overvoorde P.J. and H.D. Grimes. The mle of
calcium and calmodulin in carrot somaticembryogenesis. Plant CelI. Physiol. 35: 135-144
(1994)Raff MC. Social controls on celi survival and cetl
death. Nature 356: 397-400 (1992)Rajasekaran K., MII. Hein and I.I(. Vasil.
Endogenous absdsic add and indole-3-acetic add
and somaüc embryogenesis in cultured leaf
explants of Peiinisetum purpureurn Schum. PlantPhysiol. 84: 47-51 (1987a)Rajasekaran I(., M.B. Hein, G.C. Davis, MG.Cames and IK. Vasil. Endogenous growthregulators in leaves and tissue ctiltures ofPennisetum purpureum Schuin. J. Plant Physiol. 130:
13-25 (1987b)Reynolds T.L. Interactions between caldum and
72

Effects cfABA, maunitot and caldurn
auxin during pollen androgenesis in anthercultures of Solanurn carotïnense 1. Plant SU. 72: 109-114 (1990)Roberts.OehlscMager S.L and J.M. Dunweil.Barley anther cuiture: Pretreatment on mannitolstimulates production of microspore-derivedembryos. Plant Ceti Tiss. Org. Cult. 20: 235-240(1990)Shetty K and B.D. McKersie. Proline,thioproline and potassium mediated stimulation of
somatic embryogenesis in alfalfa (Medicago sativaL). Plant Sci. 88: 185-193 (1993)Smart CC., A.J. Fleming, K Chaloupkova andD.E. Hanke. The physiological role of absdsicacid in eliciting turion morphogenesis. Plant
Physiol. 108: 623-632 (1995)Timmers A.C.J., S.C. DeVries and J.H.N.Schei. Distribution of membrane-bound caldumand activated calmodulin during somaticembryogenesis of carrot (Daucus carota L).Protoplasma 153: 24-29 (1989)Wang M., S. Heimovaara-Dijkstra and B.VanDuijn. Modtilation of germination of embryosisolated from dormant and nondormant barleygrains by manipulation of endogenous absdsicadd. Planta 195: 586- 592 (1995)Wei Z.M., M. Kyo and H. Harada. Callusproduction and plant regeneraüon through direct
culture of isolated pollen of Hordeum vidgare cv.Sabarlis. Theor.Appl. Genet. 72: 252-255 (1986)


hapter 6
Androgenesis in Hordeum vutgare L.:
Apoptosis and the effect of lipo-oligosaccharidesduring anther pretreatment
S. Hoekstra, M. W. van der Heijden, B.J. Oppedijk, I.R. van Brouwerskaven,
S. van Bergen, F. Heidekainp & R.A. Schilperoort
iry
Characteristics of senescence were investigated during the pretreatment of anthers needed to induce
gametophytic embryogenesis in cultured barley microspores. Degradation of chiorophyli in the anthers and
a specific pattern of changes in the intracellular ABA were found both during and without pretreatment.
During pretreatment, different RNA transcripts which may result from the corresponding ABA-induced
genes were detected in the anther tissue as ;velt as in the microspores. Pretreatment resulted in pronounced
degradation of protein and RNA. However, breakdown of DNA was peculiar since a well-defined
fragmentauon pattern of the nuclear DNA was observed, suggesting programmed ccli death, also known as
apoptosis. furthermore, expression of the hydrolytic enzyme glucanase throughout pretreatment was
shown, whereas expression of the hydrolytic enzyme chitinase was induced during pretïeatment. A
stimuiatoy effect of lipo-oligosaccharides (LOS) appiication on microspore-derived plant production was
obsen’ed only during pretreatment of anthers. In the discussion, a mechanism for induction of
androgenesis is proposed.
ntroductionfor the production of (doubled) haploidplants from microspores Le. viagametophytic embiyogenesis, pretreatment of anthers and microspores isgenerally essential. During this pretreatment, microspores change into celis that
are called embtyogenic microspores.These embryogenic microspores can berecognized microscopically by theirincreased size (50-60 rm in diameter)and red interference of the exine(Hoekstra et al., 1993). Littie is knownabout the biologicat processes occurringduring pretreatment. In the model
75

Diapter 6
system barley, a few studies have beenperfonned to invesügate the effects ofanther pretreatment on gametophyticembryogenesis. It was found thatpretreatment on mannitol with premedium is the best condition for plantproduction (Hoekstra et aL, submitted).Recently, for plant production in barleythe importance of a certain concentraüonof endogenous as well as exogenousabscÎsic acid (ABA) during pretreatmentin the period between 24 to 48 hours wasdemonstrated (Hoekstra et al.,submitted). It is known that the honnoneABA plays a role in the process ofsenescence (Davies and Jones, 1991).Cho and Kasha (1992) suggest that themannitol pretreatment in barley isresponsible for the occurrence ofsenescence. These aüthors show that lipidperoxidation takes place in membranes ofanthers during such a pretreatment. Indusion of free radical scavengers not onlydelayed and suppressed lipid peroxidaüonand ethylene production during the earlycukure penod, but also reduced embryoidproduction. During pretreatment, alsochanges in cel wall properties areobsewed (Sunderland et al., 1984).Senescence involves synthesis ofhydrolytic enzymes, most importantlyglucanases (Salisbuxy and Ross, 1991),tvhich are responsible for cell walldegradation. Moreover, glucanasetogether with chitinase are detected inembryogenic cel suspension cukures ofbarley (Kragh et al., 1991). By the actionof chitinases, lipo-oligosaccharides (LOS)are assumed to be released from plantderived precursor molecules (probably cellwall fragments). These molectiles are
supposed to play a role in earlyembiyogenesis (de Jong et al., 1993). Inthe model system carrot de Jong et al.(1993, 1995) demonstrate a reduced levelof 32kD chitinase in a tsl 1 mutant celiline. In this mutant the embiyogeniccapadty is rescued by adding chitinase orby application of LOS, which are knownas Nod-factors. Nod-factors are secretedby Rhizobium in the process of noduleinduction and symbiotic nitrogen fixation(Spainketal., 1991).
In the present study, characteristicfeatures of senescence (Salisbuiy and Ros,1991; Smart, 1994), such as degradationof chlorophyll, proteins and RNA, aconstant DNA level, and a spedficpattem of intracellular ABA (ABA)concentration were invesügated duringpretreatment induced androgenesis ofbarley. We also studied the expression ofgenes during pretreatment by differentialdisplay using family specific domainprimers of genes that are known to be“ABA responsive” (a.o. Chandler andRobertson, 1994; Skriver and Mundy,1990) and of chitinase and glucanase.Also, the effect of LOS on androgenesiswas studied.
aterials and Methods
MaterialsDonor plants of Hordeum vulgare 1. cv. Igri veregrown in a phytotron onder conditions describedpreviously (Hoekstra et al., 1992). Monodonalantibody to free (+)ABA was purchased from[detek, mc. (San Bruno, Calif., USA). Rabbit antimouse alkaline-phosphatase conjugate, (+)ABAand bovine serum albumin (grade suitable forenzyme-linked immunosorbent assay ELISA) wereobtained from Sigma (St. Louis, Mo., USA). lipo
76

Apovtvsis and tivo-o1iosaccharides
oligo saccharides, ie. the nodABCL-dependentmetabolites of Rhiwbiurn leguminosarum bv. vidae(Spaink et aL, 1991) were kindly provided by dr.H. Spaink, Institute of Molecular Plant Sdences,Leiden University, Netherlands.
AndrogenesisSelection of material and ctilture conditions ofanthers were as described previously (Hoekstra etal,, 1996) with a few modifications. In short,anthers containing late uninudeate microspores,were pretreated on mannitol with pre-medium fora period of 4 days (these days are indicated as day-4, -3, -2, -1 and 0); at day 0 anthers weretransferred to medium 1 (Hoekstra et al., 1992)without hormones. Regeneration was assessed per30 anthers on the first 100 developing embryo-likestructures (ELS) and the remaining structures wereplated at once on medium III (Hoekstra et al.,1992) with 1 mg4 IGnetine.
As a control for possible effect(s) of antherharvest conditions or in vitro culture conditions, 30anthers were cultured without any pretreatmentdirectly on medium 1.
The number of viable microspores during antherpretreatment was determined by squashing 1 ml ofthe anther tissue and staining the squashedmaterial with about 20 l Fluoresceine Di-Acetate(stock 5 mg/ml in acetone).
A concentraüon range of lipo-oligosaccharideswas added during or after pretreatment; inaddifion the anthers were ctiltured directly withoutpretreatment on medium 1 with or without lipooligosaccharides.
Weight and protein detenninationAnther fresh weight as well as the dry weight wasmeasured after 16 hours of freeze drying of thesamples. Protein in frozen ground anthers wasdetermined using the bidnchoninic acid proteinassay. Bovine serum albumin (BSA) was used as astandard (Smith et al., 1985).
Chiorophylt measurementAnthers were ground in 80% acetone with a pestle.The debris was removed by centrifugation.Chlorophyll specific absorption was measured onsupernatan aliquots at 662.5 nm.
ABA extraction and ELISA assayAnthers were put into Eppendorf vials and frozenwith liquid Nr Pestle maceration of the frozenanthers was not necessasy for ABA extraction(unpublished resuits, M. Wang et al.). Thecontents of the Eppendorf vials were freeze dried.The samples were extracted at 150 rpm, 4°C wfthcold methanol (containing 100 mg/l butylatedhydroxytoluene and 0.5 g/l dtric acidmonohydrate) twice each time in 0.5 ml for 24hours. The vials were centrifuged at 8,000g for 15min at 4°C and the supematants of the sampleswere pooled. The samples ‘ere concentrated in aSpeed-vac system (Savant) and the volumeadjusted to 25 1.d with methanol. The samples werestored at -80°C until assay.
The assay of the amount of ABA in anthers andin pretreatment solution was carried Out by anenzyme-linked immunosorbent assay (ELISA) asdescribed by Wang et al. (1995).
RNA isolationSamples containing either anthers and/ormicrospores were used for total RNA isolation. Microspores were isolated according to Hoebtra etal. (1993) with the modification that the isolationwas performed at 4°C. The material was harvesteddirectly from the plant or after 1, 2, 3 or 4 days ofpretreatment. The material and a steel pestle, usedfor grmnding the material, were frozen in liquidnitrogen. Glass from the tips of pasteur pipets wasground and added in a ratio pollen:glass equivalentto 1:2 in an Eppendorf vial. Glass and plant matenal were gently mixed with a stainless steelspatula. Subsequently the mixture was ground for10 seconds with the cold steel pestle (manufactured to fit in the vial) mounted in an electric screwdriver. The process of chilling in liquid nitrogen,mixing and squashing was repeated three times.The degree of pollen disruption was established bymicroscopy. In general, an average of 70% of thepollen was damaged. Total RNA was isolated fromthe crushed cells using a micro RNA isolationmethod according to Chomczynski and Sacchi(1987) using 4 M gunidinium-isothiocyanate,0.02 M sodium citrate, 0.5% sarcosyl and a phenolkhloroform extraction step. A DNase treatmentwas applied to remove residual DNA from theRNA sample.Total RNA was isolated from aleuron and from
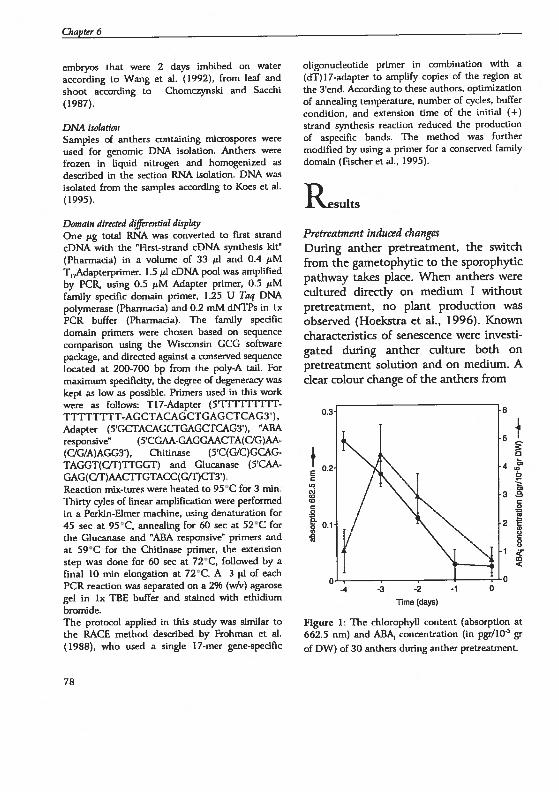
Clzapter 6
embryos that were 2 days imbibed on water
according to Wang et al. (1992), from leaf and
shoot according to Chomczynski and Sacchi
(1987).
DNA isolationSamples of anthers containing microspores svereused for genomic DNA isolation. Anthers were
frozen in liquid nitrogen and homogenized as
described in the section RNA isolation. DNA was
isolated from the samples according to Koes et al.(1995).
Domain directid dfferenflat displayOne sg total RNA was converted to first strandcDNA ;vfth the “First-strand cDNA synthesis kit”(Pharmada) in a volume of 33 1t and 0.4 MT17Adapterprimer. 1.5 i! cDNA Pool was amplified
by PCR, using 0.5 jsM Adapter primer, 0.5 Mfamily specific domain primer, 1.25 U Taq DRApolymerase (Phannacia) and 0.2 mM dNTPs in txPCR buffet (Pharmada). The family specificdomain primers vere chosen based on sequencecomparison using the Wisconsin GCG software
package, and directed against a consen’ed sequencelocated at 200-700 bp from the poly-A tal!, formaximum spec!fidty, the degree of degeneracy waskept as low as possible. Primers used in this tvorkvere as fol!ows: T17-Adapter (S’l’ITI’ITI’tY
TTTTTTTT-AGCTACAGCTGAGCTCAG3’),
Adapter (5’GCTACAGCTGAGCTCAG3’), ‘ABAresponsive” (5’CGAA-GAGGAACTA(CIG)AA-
(C/G/A)AGG3’), Chitinase (5’C(GIC)GCAG-
TAGGT(QT)TIGGT) and G!ucanase (5’CAA-GAG(C[flAACTFGTACC(G[f)CT3’).
Reaction mix-tures were heated to 95°C for 3 min.Thirty cyles of linear amplification were performedin a Perkin-Elmer machine, using denaturation for45 sec at 95C, annealing for 60 sec at 52°C forthe G!ucanase and “ABA responsive” primers andat 59°C for the Chitinase primer. the extensionstep was done for 60 sec at 72°C, fo!Iowed by afinal 10 min elongation at 72°C. A 3 d of eachPCR reaction was separated on a 2% (wA’) agarosege! in Ix TBE buffet and stained wiffi ethidiumbromide.The protocol apphed in this study was similar tothe RACE method described by Frohman et al.(1988), who used a single 17-met gene-specific
o!igonucleoude primer in combination with a
(dT) 1 7-adapter to amp!ify copies of the region at
the 3’end, According to these authors, optimization
of anneating temperature, number of cydes, buffet
condition, and extension time of the initial (+)
strand synthesis reacüon reduced the production
of aspecific bands. The method was further
modified by using a primer for a conserved fami!y
domain (Fischer et al., 1995).
l&esuits
Pretreatment induced diangesDuring anther pretreatment, the switch
from the gametophytic to the sporophyticpathway takes place. When anthers werecultured directly on medium 1 withoutpretreatment, no plant production wasobserved (Hoekstra et al., 1996). Knowncharacteristics of senescence were investigated during anther culture both onpretreatment solution and on medium. Adear colour change of the anthers from
0.3
-4 -3 -2 -1 0
Time (days)
+
03
bOi0.
t0
t
t0
Figure 1: The chiorophyti content (absorption St
662.5 om) and ABA1 concentration (in pgr/tO’5 gr
of DW) of 30 anthers during anther pretreatment.
6
78
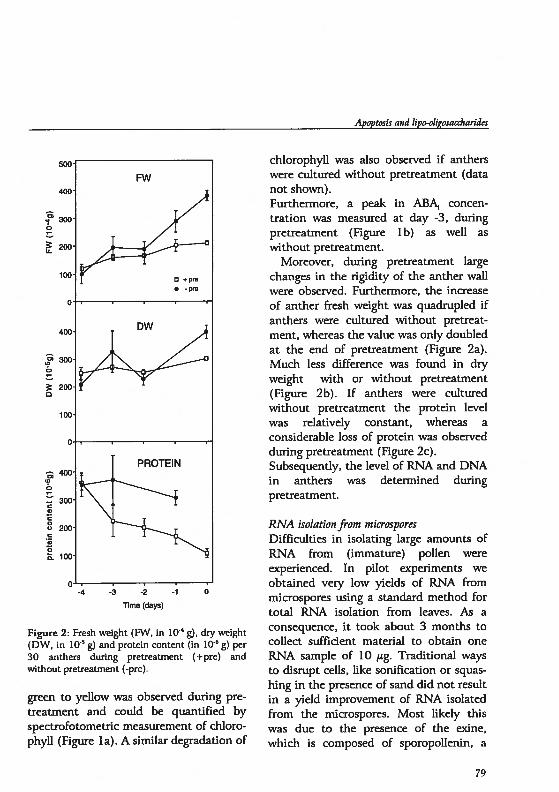
Apoptosis and 1ipo-o1iosaccharides
0,
b
0
Figure 2: fresh weight (FW, in 10 g), dry weight(DW, in lO g) and protein content (in 10:6 g) per30 anthers during pretreatment (+pre) andwithout pretreatment (-pre).
green to yellow was observed during pretreatment and could be quantffied byspectrofotometric measurement of cifiorophyll (figure 1 a). A similar degradation of
chiorophyll was also obseived if antherswere cultured without pretreatment (datanot shown).furthennore, a peak in ABA concentration was measured at day -3, duringpretreatment (figure 1h) as well aswithout pretreatment.
Moreover, dufing pretreatment largechanges in the rigidity of the anther wallwere observed. furthennore, the increaseof anther fresh weight was quadrupled ifanthers were cultured without pretreatment, whereas the value was only doubledat the end of pretreatment (figure 2a).Much less difference was found in dryweight with or without pretreatment(figure 2b). 1f anthers were cufturedwithout pretreatment the protein levelwas relatively constant, whereas aconsiderable loss of protein was observedduring pretreatment (figure 2c).Subsequently, the level of RNA and DNAin anthers was determined duringpretreatment.
RPsJA isolation from microsporesDifficufties in isolating large amounts ofRNA from (immature) pollen wereexperienced. In pilot experiments weobtained veiy low yields of RNA frommicrospores using a standard method fortotal RNA isolation from leaves. As aconsequence, it took about 3 months tocollect suffident material to obtain oneRNA sample of 10 aug. Traditional waysto disrupt ceils, like sonification or squashing in the presence of sand did not resultin a yield improvement of RNA isolatedfrom the microspores. Most likely thiswas due to the presence of the exine,which is composed of sporopollenin, a
0)
b
L6.
0,
b
0
00
0
0.
-4 -3 -2
Time (days)
79
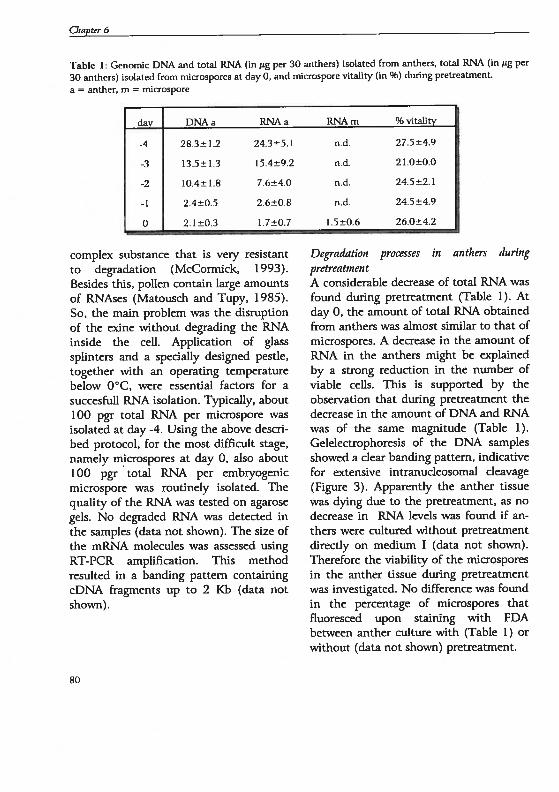
Chapter 6
complex substance that is very resistantto degradatïon (McCormidç, 1993).Besides this, pollen contain Jarge amountsof RNAses (Matousch and Tupy, 1985).So, the main problem was the dismptionof the exine without degrading the RNAinside the ceil. Application of glasssplinters and a specially designed pestie,together with an operating temperaturebelow 0°C, were essential factors for asuccesfuli RNA isolation. Typically, about100 pgr total RNA per microspore wasisolated at day -4. Using the above described protocol, for the most difficult stage,namely microspores at day 0, also about100 pgr total RNA per embtyogenicmicrospore was routinely isolated. Thequality of the RNA was tested on agarosegels. No degraded RNA was detected inthe samples (data not shown). The size ofthe mRNA molecules was assessed usingRT-PCR amplification. This methodresulted in a banding pattem containingcDNA fragments up to 2 Kb (data notshown).
Degradation processes in anthers duringpretreahnentA considerable decrease of total RNA wasfound during pretreatment (Table 1). Atday 0, the amount of total RNA obtainedfrom anthers was almost similar to that ofmicrospores. A decrease in the amount ofRNA in the anthers might be explainedby a strong reductïon in the number ofvïable celis. This is supported by theobservation that during pretreatment thedecrease in the amount of DNA and RNAwas of the same magnïtude (Table 1).Gelelectrophoresis of the DNA samplesshowed a dear banding pattem, indicativefor extensive intranucleosomal cleavage(figure 3). Apparently the anther tissuewas dying due to the pretreatment, as nodecrease in RNA levels was found if anthers were cultured without pretreatmentdirectly on medium 1 (data not shown).Therefore the viability of the microsporesin the anther üssue during pretreatmentwas investigated. No difference was foundin the percentage of microspores thatfluoresced upon staining with FDAbetween anther culture with (Table 1) orwithout (data not shown) pretreatment.
Table 1: Genomic DNA and total RNA (in jig per 30 anthers) isolated from anthers, total RNA (in pg per
30 anthers) isolated from miaospores at day 0. and microspore vita]ity (in %) during pretreatment.
a = anther, m = microspore
day DNAa RNAa RNAm %vitality
-4 28.3±1.2 24.3±5.1 n,d. 27.5±4.9
-3 13.5±1.3 15.4±9.2 n.d. 21.0±0.0
-2 10.4±1.8 7.6±4.0 n.d. 24.5±2.1
1 2.4±0.5 2.6±0.8 nU. 24.5±4.9
0 2.1±0.3 1.7±0.7 1.5±0.6 26.0±4.2—
80
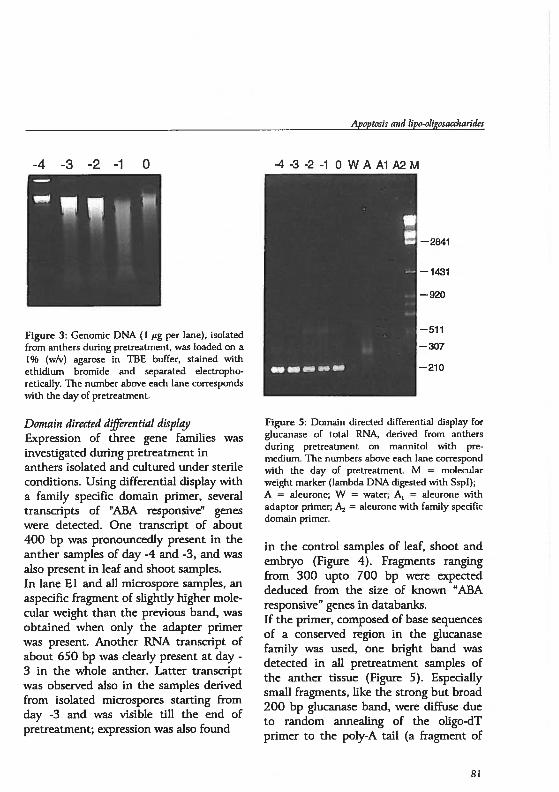
Avoptosis and livo-olieosaccharides
-4 -3 -2 -1 0
figure 3: Genomic DNA (1 jsg per lane), isolatedfrom anthers during pretreatment, was loaded on a1% (w/v) agarose in TBE buffet, stained withethidium bromide and separated electrophoreticafly. The number above each lane correspondswith the day of pretreatmenc
Dornain directed dfferential displayExpression of three gene families wasinvestigated during pretreatment inanthers isolated and cultured under sterileconditions. Using differential display witha family specific domain primer, severaltranscripts of “ABA responsive’ geneswere detected. One transcript of about400 bp was pronouncedly present in theanther samples of day -4 and -3, and wasalso present in leaf and shoot samples.In lane El and all microspore samples, anaspecific fragment of slightly higher molecular weight than the previous band, wasobtained when only the adapter primerwas present. Another RNA transcript ofabout 650 bp was clearly present at day -
3 in the whole anther. Latter transcriptwas observed also in the samples derivedfrom isolated microspores starting fromday -3 and was visible till the end ofpretreatment; expression was also found
-4 -3-2 -l 0 W A Al A2 M
Figure 5: Domain directed differential display forgtucanase of total RNA, derived from anthersduring pretreatment on mannitol with premedium. The numbers above each tane correspondwith the day of pretreatment. M = molecularweight marker (lambda DNA digested with SspI);A = aleurone; W = water; A1 = aleurone withadaptor primer; A, = aleurone with family spedficdomain primer.
in the control samples of leaf, shoot andembryo (Figure 4). fragments rangingfrom 300 upto 700 bp were expecteddeduced from the size of known “ABAresponsive” genes in databanb.1f the primer, composed of base sequencesof a consewed region in the glucanasefamily was used, one bright band wasdetected in all pretreatment samples ofthe anther tissue (Figure 5). Especiallysmall fragments, like the strong but broad200 bp glucanase band, were diffuse dueto random annealing of the oligo-dTprimer to the poly-A tail (a fragment of
81
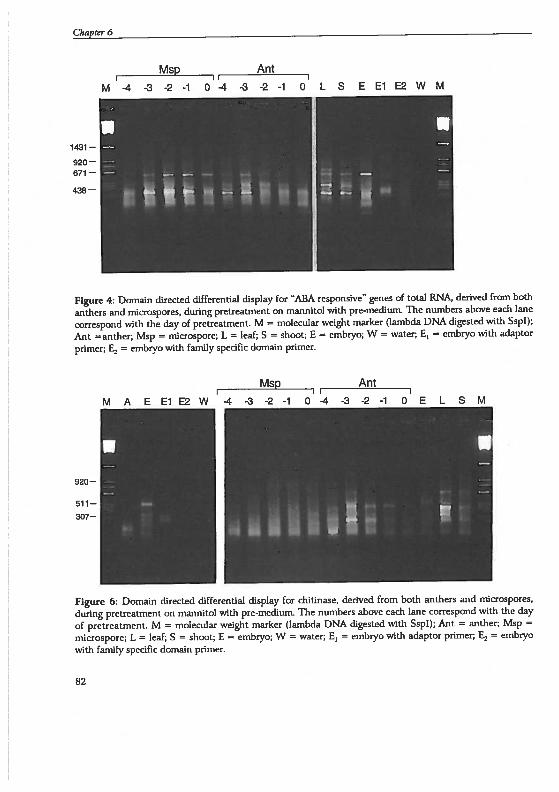
Oiapter6
1431 -
920—671—
438—
Msp Ant
M4-3-2-1 0-4-3-2-10 LS EE1E2WM
Figure 4: Domain directed differential display for “ABA responsive” genes of total RNA, derived from both
anthers and microspores, during pretreatment on mannitol with pre.medium. The numbers above each lane
correspond with the day of pretreatment. M = molecular weight marker (lambda DNA digested with SspI);
Ant =anther; Msp = microspore; L = leaf; S = shoot; E embryo; W = water; E1 = embryo with adaptor
primer; E = embryo with famity specific domain primer.
Msp Ant—ii
M A E El E2 W -4 -3 -2 -1 0 -4 -3 -2 -1 0 E L SM
figure 6: Domain directed differential display for chitinase, derived from boffi anthers and microspores,
during pretreatment on mannitol with pre-medium. The numbers above each lane correspond with the day
of pretreatment. M = molecular weight marker (Iambda DNA digested with SspT); Ant = anther; Msp =
microspore; L = leaf; S = shoot; E = embryo; W = water; E1 = embryo with adaptor primer; E2 = embryo
with family spedfic domain primer.
82
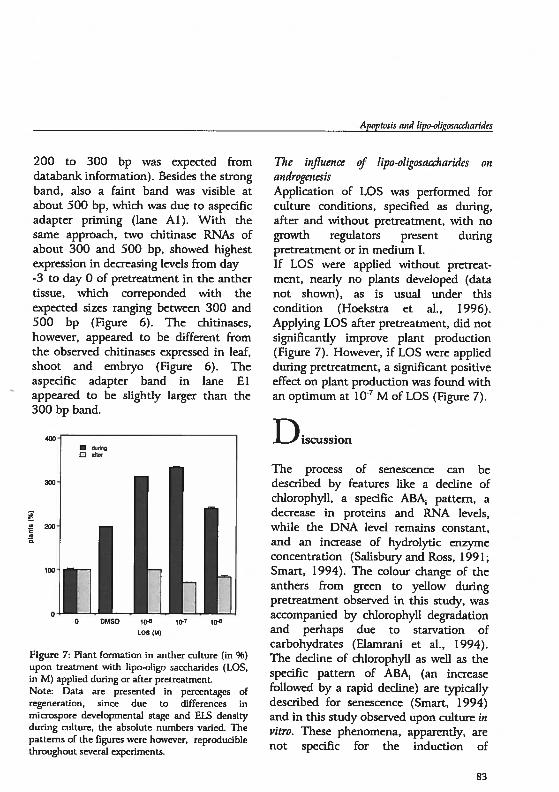
Apoptosis and 1ipo-o1iosaccharides
200 to 300 bp was expected fromdatabank information) Besides the strongband, also a faint band was visible atabout 500 bp, which was due to aspecificadapter priming (lane Al). With thesame approach, two chitinase RNAs ofabout 300 and 500 bp, showed highestexpression in decreasing levels from day-3 to day 0 of pretreatment in the anthertissue, which correponded with theexpected sizes ranging between 300 and500 bp (Figure 6). The chitinases,however, appeared to be different fromthe observed chitinases expressed in leaf,shoot and embryo (Figure 6). Theaspecific adapter band in lane Elappeared to be sllghtly larger than the300 bp band.
figure 7: Plant formation in anther culture (in %)upon treatment with lipo-oligo saccharides (LOS,in M) applied during or after pretreatment.Note: Data are presented in percentages ofregeneration, since due to differences inmicrospore developmental stage and ELS densityduring culture, the absolute numbers varied. Thepatterns of the figures were however, reproduciblethroughout several experiments.
Vie infiuence of lipo-oligosaccharides onandrogenesisApplication of LOS was performed forculture conditions, specified as during,after and without pretreatment, with nogrowth regulators present duringpretreatment or in medium 1.1f LOS were applied without pretreatment, nearly no plants developed (datanot shown), as is usual under thiscondition (Hoekstra et aL, 1996).Applying LOS after pretreatment, did notsignificantly improve plant production(figure 7). However, if LOS were appliedduring pretreatment, a significant positiveeffect on plant production was found withan optimum at lO M of LOS (figure 7).
iscussion
The process of senescence can bedescfibed by features like a decline ofchlorophyll, a spedfic ABA pattem, adecrease in proteins and RNA levels,while the DNA level remains constant,and an increase of hydrolytic enzymeconcentration (Salisbury and Ross, 1991;Smart, 1994). The colour change of theanthers from green to yellow duringpretreatment obsenred in this study, wasaccompanied by chlorophyll degradationand perhaps due to starvaüon ofcarbohydrates (Elamrani et aL, 1994).The dedine of chlorophyll as well as thespedfic pattem of ABA (an increasefollowed by a rapid decline) are typicallydescribed for senescence (Smart, 1994)and in this study obsewed upon culture invitro. These phenomena, apparently, arenot specific for the induction of
0 DM80 10-0 10-7 10-6
Los MI
83

Diapter 6
gametophytic embiyogenesis. In the
present work, under the conditions where
androgenesis was induced, in addition to
chiorophyli degradation, also protein and
RNA break down (resulting in mainly
microspore-derived RNA at day 0) was
observed. In contrast to what happens
during senescence however, we observed
DNA cleavage into well defined fragmentsbefore total degradation occurred.
Degradaüon of DNA ïnto oligomers of
oligonudeosome-sized fragments as
observed in this study, is a hailmark for
programmed cell death, which also is
called apoptosis (Raff, 1992). This author
descrlbes that another characteristic of
apoptosis is shrinkage or ftagmentauon of
the nucleus and cytoplasm, avoiding theceiPs contents being spilled in the
extracellular space and thereby preventing
accidental celi death. For further prooffor the occurence of programmed ceil
death duting pretreatment, is in progress.In many of the cases studied until now,
an increase in the cytoplasmic calciumconcentration appears to serve as acommon, early signal for initiation of
apoptosis (Ojdus et al., 1991). Calcium is
also thought to activate preexisting
endogenous endonucleases (Bowen, 1993;
Mittier and Lam, in press). Somedevelopmental pathways as well as someenvironmental insuits, as dismption of
ceil-matrix (Frisch and frands, 1994) or
celi-celI (Bates et al., 1994) interactions,or exposure to stress (Bowen, 1993),result in the activation of apoptosis.According to Greenberg et al. (1994),active oxygen and peroxidation plays a
prominent role in apoptosis. In aiilmals,apoptosis is a ubiquitous phenomenon
and play’s an important role in the
development of organs and whole
organisms (Vaux et al., 1994) and in the
control of tissue homeostasis (Raff,1992).
In plants, littie is known about the
occurrence of apoptosis beyond the
descriptive level, but apoptosis is thought
to be involved in vascular tissue
development (Chasan, 1994) and in
hypersensitive response to pathogens
(Greenberg et al., 1994; Mktler and Lam,
in press). A specific level of stress or
certain cellular conditions are needed for
induction of apoptosis (Cotter and Al
Rubeai, 1995; Mittier et al., 1995). This
could be in agreement with the specific
pretreatment condltions required forinduction of gametophytic embryo
genesis.According to Sunderland ei al, (1984) for
inducüon of androgenesis, degradation of the
tapetum as well as properties of the anther wall are
critical, and are more skin to those of maturing
anthers near to anthesis (Sunderland et al., 1984).
Comparison with literature data from microspore
development in vivo, reveal that in most spedes the
tapetum begins to degenerate shortlv after the first
pollen mitosis and is usually not present anymore
at aothesis. So, anther dehiscence in Was involves
programmed destruction of specific cell types
(Goldberg et al., 1993). The development of
microspores, starting from the slage around the
first pollen gram mitosis, into mature anthers
normally takes 10 to 11 days (Hoekstra ei al.,
unpublished). During anther pretreatment,
degradation of the tapetum probably occurred
earlier and did not have taken more than 3 days
(see DNA level in Figure 3; Cho and Kasha, 1992).
From the data presented in this study,
we propose the following mechanism ofandrogenesis induction. The anther is
exposed to several kinds of stress. Uponharvest the cetI-celi communication with
84

Aoptosis and 1io-o1iosaccharjdes
the vascular tissue is interrupted and theanther is placed on a solution containingmannitol and calcium as majorcomponents. In the anther tissuechlorophyli breaks down, and ABA issynthesized. At day -3 in the anther tissuea peak in gene expression is observed ofstress-induced genes like “ABAresponsive” genes and chitinase. Theobserved ABA1 peak at day -3 ofpretreatment coïndded with theoccurence of anther-derived ‘ABAresponsive” transcripts. Under theappropriate stress conditions, i.e. in thepresence of mannitol and 30 mM caldumions (Hoekstra et al., submïtted),nucleases that initiate programmed celideath are activated. Accidental celi deathis prevented and thereby leakage ofendogenous ABA is limited. May be LOSare freed from the anther tissue duringthe process of apoptosis. Since the anthertissue is degradïng, the microsporesbecome gradually exposed to thepretreatment solution and a specific levelof exogenous ABA. The microsporesenlarge under these osmoüc circumstances and microspore-derived “ABAresponsive” transcripts, correspondig tothe specific exogenous ABA level, areexpressed: the production of what arecalled embryogenic microspores, isestablished at the end of pretreatment.
Degradation of the anther tissue forinduction of androgenesis probablyrequires the activation of many genes thatare also inifiated during the process of in
vivo anther dehiscence. An example aregenes that encode hydrolytic enzymesrequired for ceil wall degradation, such asglucanases (Goidberg et aL, 1993). four
different glucanases have been isolatedduring in vivo anther development inLathyrus sp. and the temporal pattem ofexpression of each glucanase gene isunique (Neelam and Sexton, 1995). Inthis study, one major glucanase transcript,being expressed throughout pretreatment,was detected. The expression of bothglucanase and chitinase can be induced bypathogens (Ignaüus et al., 1994), and awide range of chemical compounds (vanLoon, 1985) like ethylene (Siefert et al.,1994). Our data suggest that theseenzymes can also be induced underaseptic condiüons by apoptosis.Moreover, these enzymes have beendetected in the medium of embryogeniccell suspension cultures of barley (Kraghet al., 1991) and are shown to play a rolein early embryo development in vitro(Dejong et al., 1993). We detected different chitinase transcripts early duringpretreatment in the anther tissue, whichis slîghtly after the highest ethyleneproduction at 12 hours of cukure (Choand Kasha, 1992). Indeed, Schmidt et al.(1993) mentïoned that chitinase genesare also expressed in the absence ofpathogens and that not all plant-producedchitinases do have an antifimgal activityin vitro. The observation that expressionof plant chitinases is correlated with plantdevelopment indicates that chitin-likemolecules occur in uninfected plants andcould play a role in plant development(Spaink and Lugtenberg, 1994). There isgrowing evidence that plants may be ableto synthesize molecules analogous to Nodfactors, which stimulate division inprotoplasts derived from legumes andnon-legumes (Schmidt et al., 1993b,
85

Diapter 6
Spaink et aL, 1993). Thus, rhizobial Nodfactors may be mimicking endogenousplant factors and consequently receptorsfor Nod factor-like molecules could bepresent in both leguminous and nonleguminous spedes, having a more generalrole than symbiosis alone (Bono et al.,1995). In our study, a positive effect ofapplicafion of Nod-like molectiles wasachieved, only when applied duringpretreatment. We speculate that underoptimal pretreatment conditions ofandrogenesis, apoptosis induces antherderived glucanases and chitinases. Chkinase subsequently releases LOS from theanther wall, which contribute to the induction of embryogenesis in microspores.
AckwwkdgementDr. A.M. Meijer (IMP, Leiden) is thanked formolecular assistance, J. Vink for growing the donorplants and P. Ho& for the lay.out of the figures.
J..eferences
Bates RC., A. Buret, D.f. Vanlielden, M.A.Horton and G.F. Bums. Apoptosis induced byinhibition of intercetlular contact. J. Celt Biol. 125:403-415 (1994)Bono J.J., J. Riond, XC. Nicolaou, N.J.8ockovich, V.A. Estevez, J.V. Cullimore and R.Ranjeva. Characterization of a binding site forchemically synthesized lipo-oligosaccharidicNodRm factors in particulate fractions preparedfrom roots. Plant J. 7:253-260 (1995)Bowen LD. Apoptosis or programmed ceil death?Celi Biol, Intern. 17: 365-380 (1993)Chandler P.M. and M. Robertson. Geneexpression regulated by abscisic add and itsrelation to stress tolerance. Annu. Rev. PlantPhysiol. Plant Mol. Bid. 45: 113-141 (1994)Chasan R Tradng tracheary elementdevelopment. Plant Celi 6: 917-919 (1994)Cho U.H. and K.J. Kasha. Relationsbip ofsenescence to androgenesis in barley (Hardeum
vulgare L. cv. Klages). J. Plant Physiol. 139: 299-
302 (1992)Chomczynski P. and N. Sacchi. Single-step
method of RNA isolation by add guanidinium
thiocyanate-phenol-chloroform extracüon. Anal.
Biochem. 162: 156-159 (1987)Cotter T.G. and M. A1-Rubeai. Ceil death
(apoptosis) in celI culture systems. Tibtech. 13:
150-155 (1995)Devies W.J. and H.G. Jones (eds) In: Abscisic
Acid physiology and biochemistry. Bio’s scienfific
publishers limited (1991)Dejong A.J., T. Hendriks, E.A. Meijer, M.
Penning, f. LoSchiavo, M. Terzi, A.
VanKammen and S.C. DeVries. Transient
reduction in secreted 32 kD chitinase prevents
somatic embryogenesis in the carrot (Daucus carota
L.)varianttsll. Dev. Genet. 16: 332-343 (1995)DeJong A.J., R Heidstra, H.P. Spaink, M.V.
Hartog, E.A. Meijer, T. Hendriks, F. I.oSchia
vo, M. Terzi, T. Bisseling, A. VanKammen and
SC. DeVries. A carrot somatic embryo mutant isrescuedbychitinase. Plant Celi 4: 425-433 (1993)
Elamrani A., 12. Gaudillere and P. Raymond.Carbohydrate starvation is a major determinant ofthe loss of greening capadty in cotyledons of darkgrown augar beet seedlings. Phys. Plant. 91: 56-64(1994)fischer A., H. Saedler and G. Theissen.Restriction fragment length potymorphism-coupled
domain-directed differential display: A highlyeffident technique for espression analysis ofmultigene families. Proc. NatI. Acad. Sci. USA 92:533 1-5335 (1995)Frisch S.M. and H. Francis. Diseuption ofepiffielial cell.matrix interactions inducesapoptosis.]. Celi Biol. 124:619-626(1994)Frohman MA., M.K Dush and G.R Martin.Rapid production of fulI-length cDNAs from raretranscripts: amplification using a single gen-spedficoligonudeoude primer. Proc. Nati. Acad. Sci. USA85: 8998-9002 (1988)Genetics Computer Groups (1991), ProgramManual for the GCG Package, Genetics ComputerGroups, MadisonGoidberg RB., T.P. Beals and P.M. Sanders.Anther development: basic prindpies and practicalapplications. Plant CelI 5: 12 17-1229 (1993)Greenberg J.T., A. Guo, D.F. IGessig and F.M.
86

Apoptosis and livo-oliosaccharides
Ausubet. Programmed ceil death in plants: apaffiogen-triggered response activated coordinatelywith multiple defense functions. Celt 77: 55 1-563(1994)Hoekstra S., M.H. VanZijderveld, J.D.Louwerse, F. Heidekamp and f. VanderMarkAnther and microspore culture of Hordeurn vulgareL. cv. Igri. Plant Sci. 86: 89-96 (1992)Hoekstra S., M.H. VanZijderveld, F.Heidekamp and F. VanderMark Microsporeculture of Hordeum vulgare L.: the influence ofdensity and osmolarity. Plant Ccli Rep. 12: 661-665 (1993)Hoekstra S., S. VanBergen, I.R. VanBrouwershaven, RA. Schilperoort and F. Heidekamp.The interaction of 2,4.D application and mannitolpretreatment in anther and microspore culture ofHordeum vutgare L. cv. Igri. J. Plant Phys. 148: 696-700 (1996)Hoekstra S., IR. VanBrouwershaven, S.VanBergen, RA. Schiiperoort and M. Wang.The role of mannitol, caldum and ABA duringpretreatment in barley androgenesis. Submitted(1996)Ignatius S.M.J., RK Chopra and S.Muthukrishnan. Effects of fungal infection andwounding on the expression of chitinases and 8-1,3glucanases in near-isogenic lines of barley. Phys.Plant. 90: 584-592 (1994)Koes R., E. Souer, A. VanHouwelingen, L.Mur, C. Spelt, F. Quattrocchio, J. Wing, B.Oppedijk, C. Ahmed, T. Maes, T. Gerats, P.Hoogeveen, M. Meesters, D. Kloos, J.N.M.Mol. Targeted gene inactivation in Pehinia by PCRbased selection of transposon insertion mutants.Proc. Nat. Acad. Sci. USA 92: 8149-8 153 (1995)Kragh 1CM., S. Jacobsen, J.D. Mikkelsen andLA. Nielsen. Purificaüon and characterization ofthree chitinases and one 8-1,3-glucanaseaccumulating in the medium of ccli suspensioncultures of barley (Hordeum vulgare L.). Plant Sci.76: 65-77 (1991)Matousch J. and J. Tupy. The release and someproperties of nudease from various pollen species.1. Plant Physiol. 119: 169-178 (1985)McCormick S. Male gametophyte development.Plant Ccli 5: 1265-1275 (1993)Mittier ii. and E. Lam. Sacrifice in the face offoes: pathogen-induced programmed ccli death in
plants. Trends Microbiol., in prees.Mittier R., V. Shulaev and E. Lam.Coordinated activation of programmed cdl deathand defense mechanisms in trangenic tobaccopiants expressing a bacterial proton pump. PlantCcli 7:29-42 (1995)Nedam A. and R. Sexton. Cellulase (endo 8-1,4glucanase) and ccli wall breakdown during antherdevelopment in the sweet pea (Latizyrus odoratusL.): isolation and characterization of partial cDNAdones. J. Plant Physiol. 146: 622-628 (1995)Ojcius D.M., A. Zychlinsky, L.M. Zheng andJ.D. Young. lonophore-induced apoptosis: Roleof DNA fragmentauon and caldum iluxes. Exp.Ccli Ree. 197: 43-49 (1991)Raff M.C. Sodal controls on ccli survival and cclideath. Nature 356: 397-400 (1992)Salisbury F.B. and C.W. Ross (cda), Plantphysiology. Wadsworth Publishing Company,Belmont California (1991)Schmidt E.D.L., A.J. de Jong and S.C. de Vries.Signal molecules involved in plant embryogenesis.Plant Mol. Biol. 26: 1305-13 13 (1993a)Schmidt J., H. Roehrig, M. John, U. Wieneke,G. Stacey, C. Koncz and J. Scheil. Mteration ofplant growth and development by Rhizobium nodAand nodB genes invoived in the synthesis ofoiigosaccharide signal molecules. Plant J. 4: 651-658 (1993b)Scott R., R Hodge, W. Paul and J. Draper. Themolecular biology of anther differentiation. PlantSci. 80: 167-191 (1991)Siefert f., C. Langebartels, T. Boiler and K.Grossmann. Are ethylene and 1-aminocydopropane-1.carboxylic acid involved inthe indudilon of chitinase and 13-1,3-glucanaseacüvity in sunilower ceil-suspension cultures?Planta 192: 43 1-440 (1994)Skriver K. And J. Mundy. Gene expression inresponse to abscisic add and osmoilc stress. PlantCcli 2:503-512(1990)Smart C.M. Gene expression during leafsenescence. New Phytol. 126: 4 19-448 (1994)Smith P.K., R.I. Krohn, G.T. Hermanson, A.IC.Mallia, f.H. Gartner, M.D. Provenzano, E.IC.fujimoto, N.M. Goeke, B.J. Olson and D.C.Klenk Measurement of protein usingbicinchoninjc add. Anal. Biochem. 150: 76-85(1985)
87

Chapter 6
Spaink H.P. and B.J.J. Lugtenberg. Role of
rhizobial lipo-chitin oligosaccharide signal
molecules in root nodule organogenesis. Plant Mol.
Biol. 26: 1413-1422 (1994)Spaink H.P., D.M. Sheeley, A.A.N.
VanBrussel, J. Glushka, W.S. York, T. Tak, 0.
Geiger, E.P. Kennedy, V.N. Reinhold and 3.J.J.Lugtenberg. A navel highly unsaturated fatty acid
moiety of lipo-oligosaccharide signals determines
host specificity of Rhizbiurn. Nature 354: 125-130
(1991)Spaink H.P., H.M. Wijfjes, T.B. VanVliet, J.W.
Kijne and B.J.J. Lugtenberg. Rhizobial lipo
oligosaccharide signals and their mle in plant
morphogenesis; are analogous lipophilic chitin
derivatives produced by the plant? Austr. J. PlantPhysiol. 20: 38 1-392 (1993)Sunderland N., B. Huang and G.J. Huis.
Disposition of pollen iu .titu and its relevance to
anther/pollen culture. 1. Exp. Bot. 35: 521-530
(1984)VanI.oon L.C. Pathogenesis.related proteins.
Plant Mol. Biol. 4: 111-116 (1985)Vaux DL., G. Haecker and A. Strasser. An
evolutionary perspecüve on apoptosis. Ceil 76:
777-779 (1994)Wang M., B. VanDuijn, R.M. VanderMeulen
and f. Heidekamp. Effect of absdsic acid
analogues on intracell&ar caldum level and gene
expression in barley aleurone protoplasts. In:
Karssen CM et al. (eds) Progress in plant growth
regulation. Klutver Academic Press, Dordrecht,
Netherlands pp. 635-642Wang M., S. Heimovaara.Dijkstra and 13.
VanDuijn. Modulation of germiantion of embryos
isolated from dormant and nondormant barley
grains by maniptilation of endogenous absdsic
add. Planta 195: 586-592 (1995)
88

hapter 7
General Discussion
n this chapter, first an overview of
efficiendes during the development frommicrospore into plant will be descrÏbed.
The tempüng question whether
androgenesis follows the process ofembryogenesis, is raised. Second a
mechanism by which androgenesis might
be induced is presented. Third, the major
parameters in the assumed model arediscussed, based on comparison with
literature.
fficiencies and embryogenesis
The effidency of microspore derived plantproduction, determines the applicability
of androgenesis in breeding. In chapter 1,2, 3, 4 and 5, factors have been describedthat infiuence the plant productionefficiency. In table 1, an oveMew ofregeneration efficiendes of anther andmicrospore culture with the model variety
of barley, cv. Igri, is presented. The
differences found in regeneration
effidendes, can be explained bydifference in the various parameters of
the method used. Data from Table 1
reveal that the regeneration efficiency of
barley androgenesis has been improved atleast five-fold (see also chapter 3), andbased on the resuits presented in chapter
5 a further improvement of about 40%
can be expected. In condusion, aftercomparison with literature data, theimportance of the major parameters
studied in the chapters 2 to 6, eg.material selection including microsporedevelopmental stage, pretreatment andgrowth regulator application, isconfinned.
With regard to the microsporedevelopmental stage, the existence of awindow for maximal plant production hasbeen described in chapter 2. In fact, therequired window of late uninucleate
microspore tumed out to be veiy small.
It is probably linked with a specific stage
in the cell cycle. Apparently celis onlybecome competent, if the indudng triggeris applied before stages of finaldifferentiation into mature pollen, withcharacteristics like starch deposition,mRNA accumulation (Goidberg et al.,1993), are commenced. Indeed,
89

Chapter 7
Table 1: Regeneration effidendes reported inliterature for androgenesis in barley cv. Igri.suboptimal conditions
Sangwan and Sangwan-Norreel (1987)hypothesize that in all androgenic species,starch accumulation only started at thelate bicellular stage, wheras in therecalcitrant species accumulation occurredthroughout pollen development.Especially as in barley a large part of themicrospore population spontaneouslydiploidizes, the optïmal stage might bedifferent compared to other species,however, does not seem to be specific forbarley. In tobacco, the application of theDNA replicatïon inhibitor hydroxyureaduring pretreatment, reveals accumulation of vegetative celis with the
nudeus in S-phase which correlates withan increased embtyo fonnation (Zarsky etal., 1992). Similar resuks are obtained byapplicaüon of the microtubulereorganizer, coichicine (Zald andDickenson, 1991; Szakacs and Bamabas,1995). These authors hypothesizesymmetrical division as being required forembryo development. Considering theefficiency during the development ofmicrospore into plant, in general highfrequencies of first divisions are observed,although only a small fraction of themicrospore population develops intoplants. To date, in the model spedesBrassica napus cv. Topas at least 40% ofthe microspores undergoes a first division,but only 1% forms a plant (Custers et al.,pers. comm.); in Nicotiana sp. upto 90% ofthe microspores start to divide, but thepercentage that develops into plant hasnever been critically determined (HeberleBors et al., pers. comm.); in Hordeumvulgare cv. Igri 15% of the microsporesprogress until the first division three daysafter isolation, and about 10% of theïsolated microspores develop into plants(Hoekstra et al., unpublished data).Apparently at most 10% of themicrospores develop into a plant and thismakes a hypothesis of many microsporesubpopulations being embiyogenic,possible. It tvould be veiy interesting toevaluate which microspores eventuallydevelop into a plant. In collaborationwith dr. T. Goids and prof.dr. H.U. Koop(University of Munich, Germany), a fewceIl traddng experiments were perfonned.Preliminary data revealed this to beextremely difficult. Mthough barleymicrospores could be embedded in
Reference(s) n plant.s
anther adtnre
Knudsenetal. 1989 2
Olsen 1987 4.6
Jâhne et al, 1991 7.2
microspore cutture
Ziauddin et al. 1990 2.9
Olsen 1991 9.4
Kuhlmanetal. 1991 10
Mordhorst et al. 1993 4.8
Cistué et al. 1995 17.4
Hoekstra et al. chapter 2 12.4
Hoekstra et al. chapter 3 50
Hoekstra et al. chapter 4 15.8
90

General Discussion
agarose (according to Goids et al., 1992)which was necessaiy for position fixaüonin a culture, the development uponisolation dramatically progressed within afew hours which hindered appropriatedetermination of the microsporemorphology directly after isolation. Uponpretreatment in barley, a mixture of bothasymmetric and symmetlic dïwsion wasobserved and conünuation of pollenmaturaüon has never been observedunder these condiüons. In condusion, celitracking could not be commenced beyondthe globular stage (about 16 days aftermicrospore isolation) and could not beperformed until and induding the plantletstage fHoekstra, Golds and Koop,unpublished data). Sofar, no dearevidence for the requirement ofsymmetric dwisions for embryogenicdevelopment has been demonstrated tomy opinion, as the complete development from a single microspore into aplant has never been followed. BesidesTouraev et al. (1995) show that uponsymmetrical division, gametophyticdevelopment can be completed intobacco, resuking in genninaüon ofpollen. These authors only recentlyobserved similar to the situation inbarley, both symmetrÏcal and asymmetricai microspore mitoses and theformation of multicellular structures fromthe products of both types of divisïon intobacco (Touraev et al., in press).
furthermore, the plane of the firstdivision is supposed to be a firstindication whether the development ofmicrospore into plant ïndeed follows theembtyogenic pathway. Comparison of thedevelopmental pattem during gameto
phytic and zygotic embiyogenesis shows,that the polarity of the zygote (Goldberget al., 1994) strongly resembles themorphology of what is cafled anembxyogenic microspore. The importanceof an asymmetrical first division inzygotic embryogenesis has beenemphasïzed by de Jong et al. (1993).West and Harada (1993) state that atransverse and asymmetlic division of thezygote is not a prerequisite forembryogenic development in all plantsand that there is no universal pattem ofearly cell deavages. Pattem fonnation inthe zygotic embryo of Arabidopsisdepends largely on cell-cell communication (Jürgens, 1995). According toMeinke (1991) the iniüat division of thezygote is often transverse, but subsequentdivision pattems clearly differ betweentaxonomic groups. for the monocotyledonous spedes barley, early phases ofzygoüc embryogenesis are largelycharacterized by irregular pattems ofearly celI divisions (SherÏdan, 1995).from our investigations we also got theimpression that, with the excepüon of thefirst division, other cell divisions withinthe exine occur at raitdom. However, arandom plane of division in earlygametophytic embryogenesis is alsoobserved in Brassica napus (Telmer et al.,1995; Yeung et al., 1996). Latter authorsfind some differences in the morphologyof zygoüc and microspore-derivedembiyos and obtain the indication fromtheir studies, that the üssues of themicrospore-derived embryo are not alwaysfully determined. Only after brealage ofthe exine, in both species a protoderm isformed, a key morphological feature of
91

Chaptgr 7
embiyogenesis in androgenesis. Moreover,from our microscopic observations noindicafion for indirect or secondaryembryogenesis was found, since embryo-like structures as a whote acquired asmooth epidermal layer. These data donot eluddate the questions when theembryogenic development of themicrospores is initiated and whether aphase specific for gametophytic embiyogenesis, that is characterized byunorganized growth, preceeds embiyogenesis. Based on the ïnterpretation ofboth literature and experimental data,here the assumption is made that inmicrospores the first (a)symmetricdivision followed by an ïrregular pattemof divisions, is part of an early directembryogenic pathway which takes placeunder spedfied in vitro conditions.
model proposed for the inductionof androgenesis
The importance of optimal pretreatmentfor maximal microspore.derived plantfonnation, has been demonstrated invarious chapters: the time of pretreatmentduration in chapter 2, the infiuence of theosmolality in chapter 3, the presence ofanther tissue during pretreatment(chapter 4) and the concentrations of aswell mannitol as calcium ions in chapter5. Based on the resuits of chapter 4, 5and 6, a model was generated by whichthe mechanism of androgenesis inductionin barley mïght be explained. Keyexperimental data have been used topropose a mechanism for optimal plantproduction from androgenesis and that is
visuatized in Figure 1 and will be outlinedbelow.Note: In the description of the model two fonts are
used in order to differentiate between experimental
data and hypothesis: data from literature or from
previons chapters of this thesis are printed in
normal font, WHEREAS HYPOTHETICAL INFOR
MATION IS DISPLAYED IN SMALL CAPITAL.
pon harvest and woundîng, theanther is subjected to stress in severalwavs. First of alt, ceil-celi communication
is interrupted, and the supply of nutrientsfrom the vascular tissue has been cut off.Secondly, under optimal conditions forandrogenesis the anther is subjected to astress treatment: the anther is placed on asolution, containing mainly mannitol andcalcium ions. The anther wall blocks theentry of these compounds into the loculecontaining microspores. Sensing thestress, the anther starts to break down thechiorophyli and to increase the ABAlevel. Around day -3 general stressinduced mRNAs, Like “ABA responsive”(Skriver and Mundy, 1990) and chitinase(Collinge et al., 1993), are synthesized inthe anther tissue. THEREUPON, proteinand RNA are degraded, and between day-3 and -2 de navo ABA synthesis takesplace. Under 11-1E APPROPRIATE stressconditions in the presence of calciumions, CA2-DEPENDENT ENDONUCLEASES
BECOME ACFIVATED RESULTING IN
NUCLEOSOMAL FRAGMENTATION, leadingto programmed celi death or apoptosis.THE PRESENCE OF MANNfTOL JN THE
PRETREATMENT SOLUTION PREVENTS
ENHANCED ANTHER CELL DEATH DUE TO
AN fl’JAPPROPRIATE OSMOTICAL LEVEL. Asa result of programmed cell death, theANTHER tissue gradually degrades
92

General Discussion
figure 1 A schematic presentation of the mechanism proposed during anther pretreatment onmannitol wiffi pre-medium leading to androgenesisin barley.
• = ABA, — pretreatment solution,= miaospore, C = anther tissue
between day -3 and -1, ABA1 DISPERSES
LIYFLE BY LIYFLE INTO THE
EXTRACELLULAR SPACES AND THE
PRETREATMENT SOLUTION, AND LIPO
OLIGO-SACCHARIDES (LOS) MIGHT 3E
FREED FROM THE CELL WALL BY CHEFINASE
ACTIVITY. An increase of the ABA level isalready detectable 4 hours after theanthers are placed on the solution. DUE
TO THE BREAK DOWN OF THE ANTHER
TISSUE, THE MICROSPORES WITHIN THE
AN3}IER COME IN CONTACT WITH thegradually increasing ABAe concentrationTOGETHER WFTH THE PRETREATMENT
SOLUTION AND MAYBE LOS, STARTING
FROM DAY -2. from that time point on,the microspores show expression ofmRNAs from “ABA responsive” genes. Ahypo-osmotical situation in 440 mOs/lcg(by calcium ions and mannitol) is createdresulting in fresh weight increase andenlargement of the microspores, andfinally in the generation of embryogenicmicrospores.
he question now tises, what happensunder suboptïmal conditions forandrogenesis, like the other threeconditions (pre-medium, mannitol andwater) analysed in chapter 5. In thepresence of pre-medium, CALCIUM IONS
PROMOTE PROGRAM MED CELL DEATH andABA is gradually released into thesolution until day -2. However, as nomannitol is present, NO APPROPRJATE
OSMOTICAL ENVONMENT IS SENSED andduring the last stages of apoptosis aroundday -1, STILL a rapid increase of ABAtakes place. During pretreatment onwater or mannitol, caldum bus areabsent and a sudden extreme increase of
QOO
93

Oapter 7
ABAe occurs AS DURING ACCEDENTAL CELL
DEATH. In the presence of mannitol aproper osmoücal environment is created,
and the rise of the ABA level is lessextreme compared to pretreatment onwater, but stil far too high for optimalinduction of androgenesis.The microspores respond OVFIMALLY to aSTIMULATORY BIJT NON-TOXIC level of
ABAe below 1000 pg/ml as in theenvironment of pre-medium withmannitol at 440 mOkg. Under thesecircumstances ABA SIGNAL
TRANSDUCFION TAKES PLACE andsignificant production of specific mRNAs,like “ABA responsive” occurs in the
microspores. THESE TLNSCRIPTS AND
ACTWATION OF OTHER GENES INVOLVED
IN STRESS RESPONSE PROBABLY ARE
REQUIRED FOR THE DEVELO?MENT INTO
EMBRYOGENIC MICROSPORES.
central part in the above describedmodel for induction of androgenesis, isactivation of programmed cell death inthe anther üssue. Whether a ceil willdivide, differentiate or undergo apoptosisis determined by controls of the cell cycle,which govem the transidons of cdlpopulations into and out of theproliferative state (Jacobs, 1992; Kingand Cidlowski, 1995; Meïkrantz andSchiegel, 1995). Certain cell cycleproteins, like cyclin-dependent kinaseshave been suggested to control apoptosis(Meikrantz and Schiegel, 1995). Furthertesting of the occurrence of apoptosisduring induction of embryogenesis wouldbe worthwhile. for example, it would beinteresting to know, whether Ca2-dependent endonuclease activity can be
detected in the anther tissue (Mittler andLam, 1995), or whether fragrnentation ofnuclei in cells from the anther wall can beobserved during different pretreatmentconditions (Earnshaw, 1995). Untit veryrecently, a correlation between activation
of programmed ceil death and inductionof embryogenesis has not been published.
Havel and Durzan (1996) show that theprocess of apoptosis occurs during diploidparthenogenesïs and early somaücembryogenesis of spmce.
In the presented model, apoptosisoccurs in the anther tissue that sunoundsthe microspores. However, in additionapoptosÏs might take place inmicrospores. Only in the species Brasska
and Nicotiana, microspores are generallypretreated without the presence of anthertissue, for development into a plant. Inother spedes only very few if any plantsare obtained when microspores arepretreated without anther tissue (Table2). Moreover, pretreatment of isolatedmicrospores more often resuks in plants,if the pollen developmental stage atpretreatment initiation is mid-binucleate(Kyo and Harada, 1986). Wei et al.(1986) report the same for barley, andmention that uninucleate pollen will diewhen pretreated without anther tissue.
Several explanations for the differencein capability of haploid plant productioncan be argued, without the presence ofanther tissue during pretreatment,considering tobacco and rapeseed versusbarley and nee. One reason might be thatsuboptimal regeneration conditions havebeen applied, comparable to the Kyo datathat are improved by Touraev; hotvever,after scrufinizing media and methods
94

General Discussion
used, this seems to be veiy unhîkely.Another reason might be that theconcentration of a mîtosis-relatedcompound or signal (required for theinduction of stress genes leading toembiyogenic microspores), and that suchmolecules are not present in unicellularmicrospores and too low abundant or notas active in rice and barley bicellularpollen; it is plausible that theconcentration or activity of suchmolecules is very dependent on thedevelopmental stage of the startingmaterial. Moreover, such molecules couldbe generated in the right molarity, ifpretreatment takes place in the presenceof the anther tissue (the developmentalstage of the microspores progresses duringpretreatment within the anther).A third reason might be that anapoptosis-related compound or signal istoo low abundant in isolated uninucleatemicrospores of rice and barley, and can be
generated by apoptosis in the anthertissue both in vitro and in vivo. Suchmolecules could be supplied in vivo by thetapetal ceils (these are present in vivo onlytili shortly after the first pollen mitosis;Goldberg et al., 1993). ABA could play arole in the model as a signal or byretardation of cell cycle completion toprovide more time for the signaltransduction (Muller et al., 1994). Thenature of such fa) signal(s) might bediverse: it could also be a cdl wallcomponent stored during this specificdevelopmental stage of the exine, or itcould be a nucleotide. Uridine e.g. isfound to act as a morphogen in plantroots (Smit et al., 1995).A last reason that can be argued is thatdifferent species have a differentmechanism for the induction ofandrogenesis, e.g. apoptosis is required inmonocotyledonous crops, whereas this isnot the case in dicotyledonous spedes.
Table 2: Plant production from isolated microspores pretreated without anther tissue presence invarious species, with the developmental stage at isolation, plant production, and the correspondingreferences tisted.* 5-10% are mitotic or bicellular pollen
Species Stage embryos plants Reference(s)
Brassica U/B* many many Gland et al. 1988; Custers et al. 1994
Nïcohana U/W many many Touraev et al. 1996
Nicotiana U 0 0 Kyo and Harada 1986B many 0
Hordeurn U 0 0 Wei et al. 1986B many 14
Hordeurn U 0 0 Hoebtra et al. chapter 4 andunpublished data
Oryza B many 3 Ogawa et al. 1994
95
Ozapter 7
P 25°C (chapter 5), if compared to cold
retreatments that induce andro- pretreatment of spikes where no extra
genesis and a comparison with caldum ions are added, can be explained
somatic embryogenesis by a 1 5-fold increased infiux of caldum at
4°C compared to 25°C (Monroy and
Activation of the ceil death process is Dhindsa, 1995).
dependent on certain cellular conditions,and a spedfic
Table 3: Various treatments that induce gametophytic embryogenesis in differentlevel of stress is specles, wth the the most important references.needed (Cotterand M-Rubeai,1995; Mittier et Species Inducer Reference(s)
al.. 1995). In Nicodana starvation Kyo and Harada 1986chapter 5, the Heberle-Bors 1989
importance ofrelatively high starvation plus heat Touraev et al. 1996
calcium C0fl Brasswa beat Pechan et al. 1991centraüons du- Custers et al. 1994
ring pretreatgamma irradlation MacDonald et al. 1988
ment ts demon Pechan and Kdller 1989strated forbarley. As ethanol Pechan and ICeller 1989
a 1 r e a d Y coichicine Zhao et al. 1995
mentionede a r 1 i e r
antherdissecUon Osolniketal. 1993
microspores are Triticum spike in water Mejza et al. 1993
e q u all vcolchidne Szakacs and Barnabas 1995
efficientlyinduced into the starvation plus ABA Hu et al. 1995
sporophyticpathway after
starvation plus beat Touraev et al. in press
c old p r e- Zea cold plus beat Genovesi 1990
treatment ofproline on donor Büter et al. 1991
barley spikes.The beneficial Hordeum cold Huang and Sunderland 1982
effect ofstarvation Roberts-Oehlschlager and Dunweil 1990
increased Hoekstra et al. chapter 3,5calcium conc e n t r a t o i 2,4-D Hoekstra et al. chapter 4
during anther Los Hoekstra et al. chapter 6
pretreatment at
96

General Discussion
In all conditions required for induction of androgenesis mentionedthroughout this thesis, single or muftipleforms of stress are involved. Duringpretreatment, osmoüc stress is evident.Further, both BAP and 2,4-D are used incombination with mannitol pretreatment,at very high concentrations (10.6 M orhigher), exceeding physiologicalconcentrations (chapter 2, 3 and 4).Moreover, in chapter 4 is demonstratedthat by application of extremely high 2,4-D concentrations (1 O M for 72 hours),also plant formation is possible withoutmannitol starvation. An overall comparison of treatments applied for inductionof gametophytic embxyogenesis indifferent spedes are listed in Table 3 andrevealed that alt treatments can becharacterized by the term ‘stress”.further, the requirement of different
stress treatments per spedes is obvious.On the contrary, recent experiments byTouraev et al. (1996 and in press)indicate that there might be a unifiedmodel for induction of androgenesis. 1fthese inducing treatments for androgenesis are compared with treatmentsapplied for inducüon of somaticembiyogenesis (Table 4), a high degree ofsimilarity can be observed.Indeed, Dudits et al. (1991) proposed asa working hypothesis that the formaflonof embryos in vitro is a stress response.The method described by the group ofKomamine (Osuga and Komamine,1995), requires levels of CaCl2 andmannitot that approach theconcentrations used in chapter 5 fresp. 30mM and 0.3M), except that the level of2,4-D is 20 times lower. Masuda et al.(1995) use in the same carrot system the
presence of 0.4 sM 2,4-Dfor 12 fl11 24 hours,which is a ten tïmesreduced concentrationcompared to our resuits(chapter 4). Zimmerman(1993) mentioned thatthe time andconcentration of 2,4-Drequired for inductiondepends on the nature ofthe explant and itscompetence for embryogenesis. The doseresponse charactetisücs ofgrowth regulators are
Table 4: Various treatments that induce somatic embryogenesis inDaucus carota with the most important references.
in Tnjotium sp., Liuurn sp. and Brassica napus.
Inducer flference(s)
005 cM 2,4.D, 15 mM CaO., Osuga and Komamine 19950.2 M mannitol
12.24 hours 0.4 jsM 2,4.D Kamada et al. 1979
BAP Merkle 1990
cold treatment donor Krul 1993
plasmolysis Wetherell 1984
wounding, pH Smith and Ifrikorian 1990a, b
heavy metals Kiyosue et al 1990 and 1992Roustan 1989
97

Diapter 7
likely to result from sensitivity variation
among the individual ceils that consfitute
the responding tissue. In addition,
biotime may be passÎng at different rates
among individual celis or tissues of a
single plant (Bradford and Trewavas,
1994). It is remarkable that somatic
embryogenesis can also be induced by
BAP in clovers, flax and oilseed rape
(Meride 1990). Besides, growth regulators
are not necessaiy for induction in carrotas mechanically wounded mature zygotic
embiyos respond in hormone-free
medium under specific pH conditions
(Smith and Kiikorian 1990a and b). Thealtemative pretreatment in barley androgenesÎs is cold incubation of spikes, andin carrot somatic embryogenesis isenhanced by cold pretreatment of stock
plants (Krul, 1993). further typical stress
treatments like heavy metals also induce
somatic embiyogenesis (Kiyosue et al.,1990, 1992; Roustan, 1989). finally It is
worth to mention that pseudomeiosis is
related to the acquisition of totipotency
in vitro (Terzi and LoSchiavo, 1990).
Their resuits provide another indication
for the existence of a general mechanism
for induction of both gametophytic and
somatic embiyogenesis.
“\Tarious stress treatments leading
to one pathway or one response?
The question is raised, whether all these
different stress treatments listed in Table
3 and 4, can activate expression of (the
same) genes required for the same cellular
response: induction of embryogenesis. In
Table 5, a few examples are presented of atranscript or protein that is inducedunder a variety of environmental stimuliused for androgenesis induction. Datafrom Table 5 indicate further that there is
a variety of transcripts or proteïnsinduced underapparently similarconditions.
B e 1 o wmolecular medianisms are outlinedthat contribute toa general stressr e 5 p o n s eaccording to Ruisand Schueller(1995), togetherwith otherrelevant literaturedata. Someproteins alreadypresent in celisnot pre-exposed
Table 5: Examples of a transcript or protein that is induced under different
environmental conditions.
Stress type References
carbon starvation, highT Tassi et al. 1993
water, lowT, highT, ABA, NaCI Pareek et al. 1995
lowT, ABA, osmoticum Wang et al. 1995
carbon starvation, highT, heavy metals Hsieh et al. 1995
water, AllA, salt,drought Ishitani et al. 1995
water, lowT, ABA Ung et al 1992
highT, ABA, heavy metals, ethylene, 2,4.D Czamecka et al. 1988
lowT, ostmicum, dehydrafion Holappa and WalkerSimmons 1995
98

General Discussion
to stress conditions, like protein ldnases,will lie activated by stress signals, asimmediate or early response in the stresssignal transduction pathway. for examplefrom the wheat abscisic acid-responsiveprotein kinase, PKABAI, is suggestedthat it may be part of generalenviromnental stress responses. Lattergene shares homology with yeast SNF 1protein ldnases that are responsive tonutrient stress (Holappa and WalkerSimmons, 1995) indicating possibleeukaryotic conservaüon of a general stressresponse. According to Ruis and Schueller(1995), such immediate or earlyresponses initiate delayed or lateresponses, e.g. the synthesis of heat shockproteins (HSP). These delayed responseswill protect ceils more permanently andeffectively by allowing an adaptation topersistent stress. Specific gene controlelements and stress-activated transcription factors, like heat shock factors (HSf)binding to them, are necessary for thisproduction of protective proteins. T h einvestigations of Pareek and co-workers(1995) on HSP, also called stressproteins, are a good example for supportof the model of androgenesis inducüonpresented in this thesis. These authorsobserved the accumulation of two HMWHSPs of 90 and 104 kDa, when riceplants were subjected to high- and lowtemperature stress, salinity, water stress,and exogenous abscisic add application.Moreover in response to hightemperature stress, the same polypepüdes were found in wheat, sorghum,pea, maize, mustard and yeast. Theseauthors suggest that both proteins areconserved stress proteins. Indeed,
conservaüon of the nucleoüde sequenceof both HSP genes and their transcriptional activators, HSF genes, has wellbeen characterized in at least certain cases(Czarnecka-Vemer et al. 1995; Vierling,1994). Different stress conditionsaccording to Ruis and Schueller (1995),might trigger a general stress response bycreating the same intracellular signal, likealterations in cytoskeletal structures. Inthe moss funaria hygrometrica, a similardisturbance of polar growth is obsewedupon exposure to either the heavy metallead or colchidne (Basile et al., 1995).Different parts of the described generalstress response mechanism can berecognized in the data available for theandrogenesis model Brassica napus:induction is accompanied by accumulation of 7OkDa HSP (Cordewener et al.,1995) and changes in the cytoskeleton,i.e. disappearance of microtubular arrays,are observed upon heat treatment (Hauseet al., 1993). By application of the microtubule dismptor colchidne, Zhao et al.(1996) show that without heat treatment,rapeseed microspores can also completethe sporophyfic pathway. These dataindicate (only) a general stress responseduring induction of embiyogenesis.
Activation of hsp genes may simplyreflect a normal stress response, but it isalso possible that spedfic HSPs aredirectly assodated with the process ofinduction of androgenesis. The latterpossibility is not unlikely, since over thelast few years it has become dear thatmany hsp genes, in animals as well as inplants, are not only induced by heat orsometimes by other stress factors, but arealso developmentally regulated, primarily
99

D,avter 7
during gametogenesis and embryogenesis
(Atldnson et al., 1993; Marrs et al., 1993;
Zarslcy et al., 1995). An alfalfa LMW Iisp
gene is transcriptionally activated in early
somatic embryos (Györgyey et aL, 1991).
Using this alfalfa cDNA done, a tobacco
homologue was isolated and its expressïon
pattem studied in tobacco. Northem
analysis reveals that the transcript is
detected in both early microspore-derived
and zygotic embryos, expression is
activated at normal temperature during
the dehydration phase of in situ pollen
development, and a dramatic increase of
the transcript level is found upon in vitro
starvation leading to embryogenic
microspores (Zarksy et aL, 1995).further, the compound 2.4-0 also
induces accumulaüon of HSPs
(Czamecka et al, 1988). Genes that are
induced upon 2,4-D treatment have been
isolated. Droog et al. (1993) demonstrate
that the encoded proteins show homology
with glutathione S-transferases. Such 2,4-
D-inducible glutathione S-transferases are
not related to auxin activity or to changes
in the endogenous auxin levels (Flwy et
al., 1995). functions proposed for the
protein by Droog et al. (1993) are
protection against dainaging effects of
oxidative reactions or induction of plant
defense genes. Besides, a glutathione S
transferase gene is found to 5e induced
during senescence (Smart et aL, 1995).
All data available on 2,4-D induced
proteîns, point to a role in certain types
of stress.After application of 2,4-0 to an
alfalfa microcallus suspension, the level of
expression of the cdc2 gene was
significantly increased (Dudits et al.,
1991). The cdc2 encodes a protein
lçinase, p34, which is assumed to be a key
regulatoty element in the cel! cyde
control (Nagl, 1995). The Arabidopsis
cdc2a gene expression is not always
coupled with celi proliferation, but may
reflect a State of competence to divide
and is induced upon 2,4-D treatment
(Hemerly et al., 1993). Comparable
accumulation of the cdc2 transcript, is
detected when LOS are applied (Spaink
et al., 1994). Such chitin derivatives also
occur in secondaiy plant cell walis of
various plant species (Benhamou and
Asselin, 1989; Spaink et al., 1993).
Expression of some plant chitinases is
correlated with plant development,
indicatïng also that chitin-like molecules
occur in uninfected plants and could play
a role in plant development (Spaink et al.,
1994). It would be interesting to
investigate which molecule specifically is
required for induction of barley
androgenesis, as a mixture of four
different motecules is applied in the LOS
experiments described in chapter 6.
Chitïn-!ike molecules could be rather
generally involved in induction of
embryogenesis, as these molecules even
might play a role during embiyogenesis in
vertebrates (Spaink et al., 1994).
enes involved in the induction of
embryogenesis
from the text above, it is dear that sofaraspecific, general stress responsive genes
and their products are found to be
assodated with induction of embryogenesis. Litde is known about spedfic
100

General Discussion
genes involved in the induction of andduring early embryogenesis, as analysis ishampered by apparent materialavailability. One gene that produces alipid transfer protein (EP2) has beenparticularly useful as a marker forepidermal celi differenüaüon duringembiyogenesis (Sterk et al., 1991). In twoother studies genes are isolated about 3days after transfer to auxin free mediumand both studies assume that the geneproducts play an important role as a ceilwall protein (Dudits et al., 1991; Sato etal., 1995). Further, a cDNA differentiallyexpressed in carrot hypocotyls after 10hours of 2,4-D treatment, shows nohomology with the EMBI nucleotidesequence data base (Nagata et al., 1993).Recently PCR methods are used such asmRNA differential display (Uang andPardee, 1992) especially suitable for smallamounts of tissue aiming at identificationof active genes. Using this method,Momiyama et al. (1995) detecteddifferential expression (of unknownfunction) in early stages of eggplantembiyogenesis. Recently Schmidt et al.(in preparation) isolated a serine ldnasegene, that is conelated with very earlystages of somatic embryogenesis in Daucuscarota. Specifically on molecular analysisof gametophytic embryogenesis, sofaronly one publication appeared. In wheat,two embryoid-abundant genes ofunknovm fimction are detected whichactivity starts 2 days after isolation(Reynolds et al., 1992). Very recently afirst confirmation of part of the model isfound in the results of Reynolds et al. (inpress). His data show accumulation of anABA-responsive, early cysteine-labeled
metallothionein transcript during the first24 to 48 hours of wheat anther culture.The transcript is absent in freshly isolatedanther fissue, but is also present indeveloping zygoüc embiyos. He furtherfinds ABA biosynthesis a requirement forthe ability of microspores to formembryoids.
he importance of the ceil wall
Studies aiming at detection ofmorphological differences betweenembryogenic and non-embryogenic tissuereveal the importance of ceil-ceilcommunication. Once the embryogenicnature is established, characteristics ofembryogenic cells are a doser adherenceand a higher number of plasmodesmata(Emons et al., 1992; Kikuchi et al.,1995). Plasmolysis of explant cells isshown to enhance induction of somaticembryogenesis in carrot. These factors arepresumed to alter the epÏgenetic state ofcells, and may be related in their abilityto disrupt the cell-cell interactionsrequired to maintain coordinated pattemsof development (Merlde, 1990).Comparison with the situation during theearly stage of zygotic embiyogenesisshows a barrier between the inner ovulecell layer and the embryo sac whichprevents the direct transfer of materialbetween these compartments (disruptionof cell-cell interaction) (Goldberg et al.,1994). Other factors found to causeenhancement of embiyogenesis induction,are arabino-galactan-proteins (AGP)(Kreuger et al., 1995; Egersdotter andVonArnold, 1995). AGPs are proteo
101

Oiapter 7
glycans occurring on ceil membranes andare supposed to play a role in a.o.pathogen defense (Kreuger and VanHoist,in press). Recently is shown, that AGPsidentify those ceils of the coleoptile whichare committed to programwed ccli death(Schindler et al., 1995). Based on themodel presented in this chapter, thesedata could be in agreement and supportthe importance of the ceil wall. In theFucus zygote it is shown that differentccli fates may be imprinted into the celiwall (Berger et al., 1994).
It might be that ceils in vitro have thepotential to produce putative maternal orgametophyuc factors under the properconditions. According to Coidberg et al.(1994) literature data suggest thatembryo morphogenesis and cclispedfication events are directed primarilyby the expression of the zygotic genomeafter the fertilizaüon process, whichwould significantly differ from animais inwhich matemafly supplied factorsinfiuence the pattems of early embryodevelopment. However, with theinvolvement of programmed ccli deathduring in vitro embryogenesis, putativeapoptosis-derived compounds, factorsfrom the ccli wall and ceil-celicommunicadon might mimick maternalfactors occurring during zygotic embryogenesis.
oncluding Remarks
The aim of this thesis was two-fold, as isdescribed in chapter 1, a ‘direct-breedingapplicable” part and a fundamental part.Parameters which are directly applicable
in breeding for the production of doubledhaploids in barley are presented inchapter 2, 3, 4 and 6. Further, thefundamental part focussed on analysîs of
the anther pretreatment (chapter 5 and
6), and resulted in the model which isdesciibed above in detail. The modelpresented provides two valuable tools foroptimization of androgenesis. Bothscreening for the occurrence of apoptosis
and determination of the extracellular
ABA concentration during pretreatment
are fast and effident methods which canbe performed within a few days in smallamounts of tissue, flirther enablingimprovement of androgenesis-derivedplants in breeding.
A research lead for the future isevaluatïon of the fit of the pretreatmentmodel for a variety of species in androgenesis as well as somatic embryogenesis.
l..eferences
Atkinson B.G., M.B. Raizada, RA. Buchard,
J.RH. Frappier and DR. Walden. Theindependent stage-spedfic expression of the 1 8kDaheat shock protein genes during microsporogenesis
in Zea map 1 Dev. Genet. 14: 15-26 (1993)Basile A., S. Giordano, V. Spagnuolo, F. Alfano
and R. Castaldo Cobianchi. Effect of lead andcolchidne on morphogenesis in protonemata ofthe moss Funana hygrometrica. Ann. Bot. 76: 597-
606 (1995)Benhamou N. and A. Asselin. Attemptedlocalization of a substrate for chiünases in plantceils reveals abundant N.acetyl-D.glucosamine
residues in seconctary wafls. Bio. CeU 67: 34 1-350
(1989)Berger Science 263: 1421-1423 (1994)Bradford K.J. and A.J. Trewavas. Sensitivity
thresholds and variable time scales in planthormone action. Plant Phys. 105: 1029-1036
(1994)
102

General Discunion
Büter 3., J.E. Schmid and P. Stamp. Effects of
L-proline and post-plating temperature treatment
on maize (Zea map L.) anther culture. Plant Ceil
Rep. 10:325(1991)Collinge D.B., 1CM. Kragh, J.D. Mikkelsen,
K.IC. Nielsen, U. Rasmussen and K. Vad. Plant
chitinases. Plant J. 3:31-40(1993)Cotter T.G. and M. Al-Rubeai. Cell death
(apoptosis) in celt culture systems. Tibtech. 13:
150-155 (1995)Cordewener J.H.G., G. Hause, E. Goergen, R
Busink, 8. Hause, J.J.M. Dons, A.A.M.
VanLammeren, M.M. VanLookeren Campagne
and P. Pechan. Changes in synthesis and
localizaUon of members of the 70-kDa class of
heat shock proteins accompany the induction of
embryogenesis in Brassica napus L. microspores.
Planta 196: 747-755 (1995)Custers J.B.M., J.H.G. Cordewener, Y. Noelien,
J.J.M. Dons and M.M. VanLookeren
Campagne. Temperature controis both
gametophytic and sporophytic development in
microspore cultures of Brassica napus. Plant Ceil
Rep. 13: 267-271 (1994)Czarnecka-Verner E., C.X. Yuan, P.C. Fox and
W.B. Gurley. Isolation and characterizalion of six
heat shock transaiption factor cDNA dones from
soybean. Plant Mol. Biol. 29: 37-51 (1995)
Czarnecke 1., RT. Nagao, J.L. ICey and W.B.
Gurley. Characterization of Gmhsp26-A, a st.ress
gene encoding a divergent heat shock protein of
soybean: heavy-metal-induced inhibition of intron
processing. Mol. Cel) Bio). 8: 1113-1122 (1988)
DeJong A.J., R. Heidstra, H.P. Spaink, M.V.
Hartog, E.A. Meijer, T. Hendriks, f. LoSchia
vo, M. Terzi, T. Bisseling, A. VanKammen and
S.C. DeVries. A carrot somatic embryo mutant is
rescued by chiünase. Plant Ccli 4: 425-433 (1993)
Droog F.N.J., P.J.J. Hooykaas, 1CR. Libbenga
and E.J. VanderZaal. Proteins encoded by anawun-regulated gene family of tobacco share
limited but significant homology with glutathione
S-transferases and one member indeed shows in
vitro GST activity. Plant Mol. Biol. 21: 965-972
(1993)Dudits D., L. Boegre and J. Gyoergyey.Molecular and cellular approaches to the analysis
of plant embxyog development from Somatic cclie
in vitro. J. Ccii Sd. 99: 475-484 (1991)
Earnshaw W.C. Nuclear changes in apoptosis.
Curr. Opinion CelI Biol. 7: 337-343 (1995)
Egersdotter U. And S. VonArnold. Importance of
arabinogalactan proteins for the devetopment
of somatic embryos of Norway spruce (Picea
abies). Physiol. Plant. 93: 334-345 (1995)
Emons A.M.C., J.W. Vos and 11. IGeft. A freeze
fracture analysis of the surface of emrbyogenic and
non-embryogenic suspension celis of Daucus carota.
Plant Sd. 87: 85-97 (1992)Hury T., D. Adam and IC Kreuz. A 2,4-D-
indudble glutathione S-transferase from soybean
(Glycine mcie). Purification, characterisation and
inducüon. Phys. Plant. 94: 312-3 18 (1995)
Genovesi A.D. Maize (Zea mays L.): In vitro
production of hapioids. In: Biotechnology in
agriculture and forestxy 12. Haploids in aop
improvement 1. Bajaj Y.P.S. cd., Springer Verlag.
pp.I7&203 (1990)Gland A., R Lichter and H.G. Schweiger.
Genetic and exogenous factors affecting
embryogenesis in isolated microspore ctiltures of
Brassica napus L. J. Plant Physiol. 132: 613-617
(1988)Gotdberg RB., 1.8. Beals and P.M. Sanders.
Anther development: basic prindples and practical
appiications. Plant Ccli 5: 12 17-1229 (1993)Goldberg R.B., G. DePaiva and R. Yadegari.
Plant embryogenesis: zygote to seed. Science 266:
605-614 (1994)Goidberg RB., P.M. Sanders and T.P. Beals. A
novel cell-ablation strategy for studying plant
development. Phil. Trans. R Soc. Lond. 3 350: 5.
17 (1995)Golds T.J., J. Babczinsky, G. Rauscher and
H.U. ICoop. Computer-controtled tracking of
single ccli development in Nicotiana tabacum L and
Hordeum vidgare 1. Protoplasts embedded in
agarose/alginate films. J. Plant Physiol. 140: 582-
587 (1992)Györgyey J. A. Gartner, K. Nemeth, Z. Magyar,
H. Hirt, E. Heberle-Bors and D. Dudits. Alfalfa
heat shock genes are differentially expressed
during somatic embryogenesis. Plant Mol. Biol. 16:
999-1007 (1991)Hause 8., G. Hause, P. Pechan and A.A.M.
VanLammeren. Cytoskeletal changes and
induction of embryogenesis in microspore and
pollen cultures of Brtzssica napus L. Ccli Bio). Int.
103

Chapfrr 7
17: 153-168 (1993)Havel L. and D.J. Durzan. Apoptosis duringparthenogenesis and early somaüc embryogenesisof norlvay spruce. InC J Plant SU. 157: 8-16(1996)Heberle-Bors E. Isolated pollen culture intobacco: plant reproductive development in anutshell. Sex. Plant Reprod. 2: 1-10 (1989)Hemerly A.S., P. ferreira P., J. DeAlmeidaEngler, M. VanMontagu, G. Engler and D.Inze. Cdc2a expression in Arabidopsis is linked withcompetence for celi division. Plant CelI 5: 1711-1723 (1993)Hotappa L.D. and M.K. Walker-Simmons. Thewheat absdsic acid-responsive protien kinasemRNA, PKABA1, is up-regulated by dehydration,cold temperature. and osmotic stress. PlantPhysiol. 108: 1203-12 10 (1995)Hsieh H.M., WIC. Liu and P.C. Huang. Anovell stress-inducible metallothionein-like genefrom nee. Plant Mol. Biol. 28: 381-389 (1995)Hu TC., A. Ziauddin, E. Simion and KJ.Kasha. Isolated rnicrospore culture of wheat(Triticum aestivum L.) in a defined media 1. Effectsof pretrealnient, isolafion methods, and hormones.In Vitro Ceil Dev. Biol. 31: 79-83 (1995)Huang 3. and N. Sunderland. Temperaturestress in barley anther culture. Mn Bot 49: 77-88(1982)Ishitani M., T. Nakamura, S.Y. Han and T.Takabe. Expression of the betaine aldehydedehydrogenase gene in barley in response toosmotic stress and absdsic acid. Plant Mol, Biol.27: 307-315 (1995)Jacobs T. Control of the celi cyde. Dcv. Biol. 153:1-15 (1992)Jâhne, A., P.A. Lazzeri, M. Jâger-Gussen andH. Lörz. Plant regeneration from embryogenic cellsuspension derived from anther cultures of barley(Hordeum vulgare 1.). Theor. Appi. Genet. 82: 74-20(199 1)Jürgens G. Axis formation in plant embryogenesis:cues and dues. Celi 81: 467-470 (1995)Kamada H. and H. Harada. Studies on theorganogenesis in carrot lissue cultures 1. Effects ofgrowth regulators on somatic embryogeneis androot formation. Z. Pflanzenphysiol. 91: 255-266(1979)Kikuchi A., S. Satoh, N. Nakamura and T.
fujli. Differences in pecüc polysaccharidesbetween carrot embiyogenic and non-embryogeniccalli. Plant Celi Rep. 14:279-284 (1995)King K.L. and J.A. Cidlowski. Ceil cycle andapoptosis: common pathways to life and death. J.Cdl. Biochem. 58: 175-180 (1995)IGyosue T., S. Satoh, H. Kamada and H.Harada. Immunological detection of anembryogenic protein (ECP3 1) during stressinduced somatic embryogenesis in carrot. Cao. J.Bot. 70: 65 1-653 (1992)Kiyosue T., K. Takano, H. ICamada and H.Harada. Inducüon of somafic embryogenesis incarrot by heavy metal lans. Cao. 1. Bot. 68: 230 1-2303 (1990)ICnudsen S., I.K. Due and S.B. Andersen.Components of response in banley anther culiure.Plant Breed. 103: 241-246 (1989)Kreuger M. and G.J. VanHoist. Arabinogalactanproteins and plant differentiation. Plant Mol. Biol.,in press.Kreuger M., E. Postma, Y. Brouwer and G.J.VanHoist. Somatic embzyogenesis of Cyclamenpersicum in liquid medium. Physiol. Plant. 94: 605-612 (1995)Krul W.R. Enhancement and repression ofsomafic embryogenesis in celi cultures of carrot bycold pretreatment of stock plants. Plant CeIl ‘flss,Org. CuIc 32: 271-276 (1993)ICuhlmann U., 3. ForoughiWehr, A. Granerand G. Wenzel. Improved culture system formicrospores of barley to become a target for DNAuptake. Plant Breeding 107: 165-168 (1991)Kyo M. and H. Harada. Control of thedevelopmental pathway of tobacco pollen in vitro.Planta 168: 427-432 (1986)Lâng V., E. Mantfyla, 8. Welin, 8. Sundbergand E.T. Palva. Alternations in water Status,
endogenous abscisic add content, and expressionof rab 16 gene during the development of freezingtolerance in Arahidopsis thaliana. Plant Physiol.104: 1341-1349 (1994)Liang P. and A.B. Pardee. Differential display ofeukaryotic messenger RNA by means of thepolymerase chain reaction. Science 257: 967.971(1992)MacDonald M.V., MA. Hadwiger, F.N. Aslaanand D.S. Ingram. The enhancement of antherculture efficiency in Brassica napus ssp. ole(fera
104

General Discussion
Metzg. (Sinsk) using low doses of gammairradiation. New Phytol. 110: 101- (1988)Marrs KA., ES. Casey, S.A. Capitant, R.A.Bouchard, P.S. Dietrich, 1.1. Mettier and R.M.Sinibaldi. Characterization of two maize HSP9Oheat shock protein genes: expression during heat
shock. embryogenesis. and pollen development.Dev. GeneC 14:27-41(1993)Masuda et al 1. Pl. Physiol. 145 53 1-534 (1995) isreference I(amada et al. In Vitro Celi Dcv. Biol. 25(89) 1163-1166)Meikrantz W. and R Schiegel. Apoptosis andthe celI cyde. J. Ceil. Biochem. 58: 160-174 (1995)Meinke D.W. Perspectives on genetic analysis of
plant embrvogenesis. Plant CelI 3: 857-866 (1991)Mejza S.J., V. Morgant, DE. DiBona and J.R.Wong. Plant regeneration from isolatedmicrospores of Triticurn aeshvurn. Plant Ceil Rep.12: 149 (1993)Merkle S.A., W.A. Parrott and E.G. Williams.Applications of somatic embiyogenesis and embryocloning. In: Plant fissue culture: appllcations andlimitations, ed. S.S. Bho)wani, Elsevier pp. 67-101(1990)Mittier R and E. Lam. Identification,characterization, and purification of a tobaccoendonuclease activity induced upon hypersensiuveresponse celI death. Plant Cdl 7: 1951-1962(1995)Mittler R., V. Shulaev and E. Lam.Coordinated activation of programmed cell deathand defense mechanisms in trangenic tobaccoplants expressing a bacterial proton pump. PlantCeil 7:29-42 (1995)Momiyama T., Afele J.C., Saito T., Kayano T.,Tabel Y., Takaiwa f., Takayanagi K,Nishimura S. Differential display identiflesdevelopmentally regulated genes during somaticembryogenesis in eggplant (Solanum melougena L).Biochem. Biophys. Res. Comm. 213: 376-382(1995)Monroy A.f. and R.S. Dhindsa. Lowtemperature signal transduction: induction of coldacclimation-specific genes of alfalfa by calcium at
25C Plant CelI 7: 321-331 (1995)Mordhorst A.P. and H. Lörz. Embrvogenesis anddevelopment of isolated barley (Hordeum vrdgare L.)microspores are iniluenced by the amount andcomposition of nitrogen sources in culture media.
J. Plant Physiol. 142: 485-492 (1993)MülIer M.L., P.W. Barlow and P.E. Pilet. Effectof absdsic add on the ceIl cyde in the growingmaize root. Planta 195: 10-16 (1994)Nagata R, Wada M., Satoh S., Kamada H.,Harada H. A cDNA differentially expressedbetsveen embryogenic and nonembryogenic carrot(Daucus carota L.) cells. Plant Physiol. 103: 102(1993)Nagi W. Cdc2-kinases, cydins, and the switchfrom proliferation to polyploidizaion.Protoplasma 188: 143-150 (1995)Ogawa T., H. Fukuoka and Y. OhkawaInducüon of ceil division of isolated pollen grainsby sugar stasvation in rice. Breed. Sti. 44: 75-77(1994)Olsen F.L. Induction of micropsore embryogenesis in cultured anthers of Hordeum vidgare. Theeffects of ammonium nitrate, glutamine andasparagine as nitorgen sources. Carlsberg Res.Commun. 52: 393-404 (1987)Olsen f.L. Isolation and csjltivation ofembryogenic microspores from barley (Hordeumvulgare L). Hereditas 115:255-266(1991)Osolnik 8., 8. Bohanec and S. Jelaska.Stimulations of androgenesis in white cabbage(Brassica oleracea var. capitata) anthers by lowtemperature and anther dissection. Plant Cdl ‘fles.Org. Cult. 32: 241(1993)Osuga I(. and A. Komamine. Synchronizaüon ofsomatic embryogenesis from carrot cells at highfrequency as a basis for the mass production ofembryos. Plant Celt Tiss. Org. Cult. 39: 125-135(1995)Pareek A., S.L. Singla and A. Grover.Immunological evidence for accumulation of twohigh-molecular-weight (104 and 90 kDa) HSPs inresponse to different stresses in nce and inresponse to high temperature stress in diverseplant genera. Plant Mol. Biol. 29: 293-3-1 (1995)Pechan P.M., D. Bartels, D.C.W. Brown and J.Scheil. Messenger-RNA and protein changesassociated with induction of Brassica microsporeembryogenesis. Planta 184: 161-165 (1991)Pechan P.M. and W.A. Keller. Induction ofmicrospore embryogenesis in Brassica napus 1. bygamma irradiation and ethanot stress. In VitroCdl. Dev. Biol. 25: 1073-74 (1989)Reynolds T.L. and RL. Crawford. Escpression of
105

Chapter 7
an absdsic acid.responsive. early cvteine-labeledmetaltothionein gene during pollen embryogenesisin bread wheat (Tndcum aestivum). Plant Mol. Biol.in press.Reynolds T.L. abd S.L. IGtto. Idenflfication ofembryoid-ahundant genes that are temporallyexpressed during pollen embryogenesis in wheratanther cultures. Plant Physiol. 100: 1744-1750(1992)Roberts-Oeffischiager S.L and J.M Dunweil.Barley anther culture: pretreatment on mannitolstimulates production of miaospore-derivedembtyos. Plant Celi, Tiss. Org. Cult. 20: 235-240(1990)Roustan J.P., A. Latche and J. falloL
Stimulation of Daucus carota somaticembryogenesis by inhibitors of ethylene synthesis:cobalt and nickel. Plant Celt Rap. 8: 182-185(1989)Ruis H. and C. Schuefler. Stress signalling inyeast. Bioessays 17: 959-965 (1995)Sangwan R.S. and B.S. Sangwan-Norreel.Ultrastructural cytology of plasfids in pollen grainsof certain androgeni and nonandrogenic plants.Protoplasma 138: 11.22 (1987)Sato S., T. Toya, R. Kawahara, RF. Whittier,H. fukuda and A. Komamine. Isolation of acorrot gene expressed spedfically during early-stage somatic embryogenesis. Plant Mol. Biol. 28:39-46 (1995)Schindler T., R Bergfeld and P. Schopfer.Arabinogalactan proteins in maize coleoptiles:developmental relationsbip to ceil death duringxylem differentiation but not to extension groMh.PlantJ. 7:25-36(1995)Sheridan W.F. Genes and embryo morphogenesisin angiosperms. Dev. Genet. 16: 29 1-297 (1995)Skrwer K And J. Mundy. Gene expression inresponse to absdsic acid and osmotic stress. PlantCeIl 2:503-512 (1990)Smart C.M., S.E. Hosken, H. Thomas, JA.Greaves, B.G. Blair and W. Schuch. The timingof maize leaf senescence and characterisation ofsenescence-related cDNAs. Phys. Plant. 93: 673-682 (1995)Smit G., C.C. DeKoster, f. Schripsema, H.P.Spaink, A.A. VanBrussel and J.W. IGjne.Uridine, a celi division factor in pea roots. PlantMol. Biol. 29: 869-873 (1995)
Smith D.L. and A. D. ICrikorian. Somaücembryogenesis of carrot in hormone-free medium:external pH control over morphogenesis. Am. J.BoL 77: 1634-1647 (1990)Smiffi D.L. and A. D. Krikorian. Somaticproembryo production from exdsed, woundedzygofic carrot embsyos on hormone-free medium:evaluation of the effects of pH, ethylene andactivated charcoal. Plant CeIl Rap. 9: 34-3 7 (1990)Spaink H.P. and B.J.J. Lugtenberg. Role ofrhizobial lipo.chiün oligosaccharide signalmolecules in root nodule organogenesis. Plant Mol.Biol. 26: 1413-1422 (1994)Spaink H.P., H.M. Wijfjes, T.B. VanVliet, J.W.IGjne and B.J.J. Lugtenberg. Rhizobial lipooligosaccharide signals and their role in plantmorphogenesis; are analogous lipophilic chitinderivatives produced by the plant? Austr. J. PlantPhysiol. 20: 38 1-392 (1993)Sterk P., H. Booij, G.A. Schellekens, A.VanKammen and S.C. DeVries. Cell-spedficexpression of the carrot EP2 lipid transfer proteingene. Plant Cell 3:907-921(1991)Szakâcs E. and B. Barnabâ.s. The effect ofcolchidne treatment on microspore division andmicrospore.derived embryo dffferenüation inwheat (Triticum aesrivum L) anther csslture.Euphytica 83: 209-2 13 (1995)Tassi f., E. Maestri, F.M. Restivo and N.Mannfroli. The effects of carbon starvation oncellular metabolism and protein and RNAsynthesis in Gerbera callus cultures. Plant Sd. 83:127 (1993)Telmer C.A., W. Newcomb and D.H.Simmonds. Cellular dianges during heat shockinduction and embryo devetopment of culturedmiaospores of Brassica uapus cv. Topas.Protoplasma 185: 106-112 (1995)Terzi M. and f. LoSchiavo. Somaticembryogenesis. In: Plant fissue culture:applications and lirnitations, ed S.S. Bho)wani.Elsevier pp. 54-66 (1990)Toureav A., A, Ilham, 0. Vicente and E.Heberle-Bors. Stress4nduced microsporeembryogenesis in tobacco: an optimized system formolectilar studies. Plant Cdl Rap. 15: 561-565(1996)Touraev A., f. I.ezin, E. Heberle-Bors and 0.Vicente. Maintenance of gametophytic
106

General Discus.çion
development after symmetrical division in tobacco
microspore culture. Sex. Plant Reprod. 8: 70-76
(1995)Vierling E. The roles of heat shock proteins in
plants. Annu. Rev. Plant Physiol. Plant Mol. Biol.
42: 579-620 (1994)Wang II., 11 Dastla, f. Georges, M. Loewen
and A.J. Cutler. Promoters from kini and cor6.6,
two homologous Arabidopsis tJ,aliana genes:
transcriptional regsilauon and gene expression
induced by low temperature, ABA, osmoticum and
dehydration. Plant Mol. Biol. 28: 605-617 (1995)
Wei Z.M., M. Kyo and H. Harada. Callus
fomiation and plant regeneration through direct
culture of isolated pollen of Hordeum vulgare cv.
Sabarlis. Theor. Appl. Genet. 72: 252-255 (1986)
West and Harada Plant Celi 5: 1361-1369
(1993)Wetherell D.f. Enhanced adventive embryogenesis
resulung from plasmolysis of cultured wild carrot
cells. Plant Celt ‘Tiss. Org. Cult. 3:221-227 (1984)
Yeung E.C., M.H. Rahman and T.A. Thorpe.
Comparative development of zygollc and
mirrospore-derived embryos in Brassica napus L.
Cv. Topas. 1. Histodifferentiation. Int J. Plant Sa.
157: 27-39 (1996)Zald M. and H. Dickinson. Modification of cell
development in vitro: the effect of colchdne 0fl
anther and isolated microspore culture in Brassica
napus. Plant Celi Tiss. Org. Cult. 40: 255 (1995)
Zaki M.A.M. and H.G. Dickenson. Microspore
derived emrbyos in Brassica: the significance of
division syrnmetry in pollen mftosis 1 to
embryogenic development. Sex. Plant Reprod. 4:
48-55 (1991)Zarsky V., D. Garrido, N. Eller, J. Tupy, 0.
Vicente, F. Schöffl and E. Heberle-Bors. The
expression of a small beat shock gene is acüvated
during induction of tobacco pollen embryogenesis
bystarvation. Plant CeIl Env. 18: 139-147 (1995)
Zarsky V., D. Garrido, L. Rihova, J. Tupy, 0.
Vicente and E. Heberle.Bors. Derepression of
the ceil cyde by starvation is involved in the
inducüon of tobacco pollen embryogenesis. Sex.
Plant Reprod. 5: 189-194 (1992)Zhao J.P., D.H. Simmonds and W. Newcomb.
Induction of embryogenesis with colchidne instead
of heat in microposres of Brassica napus 1 cv.
Topas. Planta 198: 433439 (1996)
Ziauddin A., 11. Simion, and KJ. Kasha.
Improved plant regeneration from shed microspore
culture in barley (Hordeurn vulgare L.) cv. Igri. Plant
CeIl Rep. 9: 69-72 (1990)Zimmerman L. Somatic embryogenesis: A model
for early development in higher plants. Plant CeIl
5: 141 1-1423 (1993)
107


S
he research described in this thesisdeals with the optimization ofandrogenesis, a method by which
(doubled) haploid plants from immature
pollen or microspores are created. In
addition, physiological and molecular
aspects of androgenesis induction werestudied. The method was perfonned withthe crop barley (Hordeum vulgare L.) andin all chapters mainly the model cultïvarIgri was used. Haploid plants contain halfthe genetic information of what isgenerally present in a (diploid) plant.Such haploid plants are an important toolin plant breeding for the development ofnew varieties with desired traits. Often, it
can be rather complicated to detennine
the presence of a trait in a certain plant.Desired traits, like many diseaseresistances, are encoded by multiple genesthat differ in level of dominance. The useof haploid plants enables a.o. to verify thepresence of these problematic polygenicresistances, what is calied fixation oftraits. By integration of androgenesisderived plants in breeding, the qualityand the effidency for the development ofnew variedes can be improved.
In chapter 1 the importance of areprodudbie method for the productionof doubled haploid is outlined.
Furthennore, the state of the art in theliterature and the lack of knowledge inthis field are indicated. Based on thesedata and from the resuks presented in
this thesis, three preconditïons areformulated for an effident induction ofmicrospore-derived plant production; (i)the explant should be in the lateuninucleate microspore developmental
stage, (ii) a pretreatment of anthers
should lie applied and (iii) growthregulators should be added into theculture medium. These precondiuons arethe key factors that are investigated inthe following chapters.
In chapter 2 the basic method isdesaibed, in which material is used
grown under controlled environmentalcondifions. The influence of fourparameters on plant regeneraüon, namely
(i) the microspore developmental stage,(ii) the duraüon of pretreatment, (iii) thecomposition of media and (iiii) theapplicauon of aeration, is tested. Optimalregeneration frequendes were obtained, if(i) at least 50% of the microspores werein the mid-late to late uninudeate stageat harvest of the anthers, if (ii) antherswere pretreated on a mannitol containingsolution for 4 days, and if (iii) oxygen wassupplied regularly to the ctiltures.Addition of vitamines and caseinehydrolysate to the culture mediumsümulated regeneration only in
microspore cukure. Uttle if any effect wasobserved in anther culture. Microsporeculture was at least five times moreeffident in plant production than antherculture, resuking in 12,4 green plants peranther.
109

Summaiy
In chapter 3 studies on furtheroptimization of the method formicrospore cuitwe is presented. Theimportance of the osmolality duringpretreatment and in culture wasdemonstrated. Further, it is describedhow an embryogenic subpopulation ofmicrospores can be recognized.Combination of optimal osmolality withadjustment of the microspore populationwith regards to homogenity, bystandardizing the density of embryogenicmicrospores, resulted in 50 green plantsper anther.
In chapter 4 the effect of vaiying thegrowth regulator concentration in antherand microspore cultures is shown. Instead of the generally applied cytoldninecalled benzylaminopurine (BAP), thesynthetic awdne 2,4-dicfflorophenoxyacetic add (2,4-D) was used. Both a 2,4-D concentration of 10.6 M continuouslypresent in the ctilture medium, and ahigher concentration present for ashortened period of time (1 O M for 1hour), resulted in similar regenerationfrequendes. The values observed werecomparable to those for the regenerationobtained in BAP-derived cultures (seechapter 2 and 3). Production of plants inthe absence of pretreatment also waspossible, although at lower frequendes. Itis conduded, that applicaüon of i0 M2,4-D for 1 hour might be a more generalapplicable regeneration method, sincebarley microspores as well as smalldusters of Daucus carota ceils insuspension culture produce plants undersuch inductive conditions.
Chapter 5 and 6 deal with thepretreatment conditions for the anther
tissue. In chapter 5 the influence ofmannitol and caldum ion concentraüonsduring pretreatment on regenerationfrequency is presented. Optimal resultswere obtained by anther pretreatment onpre-medium containing 30 mM CaC12dissolved in mannitol upto an osmolalityof 440 mOfkg. During pretreatment theconcentration of absdsic add (ABA), astress-related honnone, was determined.A peak of endogenous ABA after 24 hoursof pretreatment was found, which doesnot correlate with induction ofandrogenesis. The level of exogenous ABAhowever, proved to be important and wasfurhter analysed by either removal ofpretreatment solution or application ofthe ABA biosynthesis inhibitor fluridone.The resufts show that for optimaldevelopment of plants the exogenousABA concentration should be at a specificlevel between 24 and 48 hours afterinifiation of pretreatment.
In chapter 6 the question is addressedwhether senescense is involved duringpretreatment, and characteristics thereofare determined. It appeared that not theprocess of senescense, but programmedcdl death or apoptosis took place. Basedon expression pattems generated bydomain directed differential display for“ABA responsive” genes, chitinase genesand glucanase genes, a model forinduction of androgenesis in pretreatedanthers is proposed. 1fl the model, theprocess of calcium induced apoptosisplays a central role. Apoptosis is inducedin the presence of caldum ions bycalcium-depending nudeases. Since nonecrotic ceil death occurs, the exogenousABA concentration increases littie by
110

Summary
littie during pretreatment. A stimulating,but non-toxic concentration of exogenousABA ïnduces the expression of spedfic“ABA responsive” genes in themÏcrospores, resulting in the generation ofembryogenic microspores at the end ofanther pretreatment. Besides an observedstimulating effect of lipo-oligosaccharide(LOS) addition during pretreatment onplant production, is integrated in thedescribed model.
In the last chapter the model ofandrogenesis ïnduction is discussed inmore detail. Aspects like the microspore
developmental stage, the presence of theanther wall and various stress-indudngagents are reviewed. A comparison ismade for inductïon of embryogenesisderived from either gametes or somaticcells. Moreover, efficiencies during barleyandrogenesis are compared with Brasskanapus and Nicotiana tabacum. from thisinformation, the embryogenic microsporetype and the question whether thedevelopment of microspore into plantfollows the embryogenic route, isdiscussed and compared with the processin planta, zygotic embryogenesis.
111


S
it proefschrift beschrijft hetoptimaliseren van androgenese, eenmethode voor het genereren van(verdubbelde) haploide planten uitonrijpe stuifmeelkoffels, die microsporenworden genoemd. Daarnaast heeft eenstudie van fysiologische en moleculaireaspecten van androgenese plaatsgevonden. Deze methode is toegepast hijhet gewas gerst (Hordeum vulgare L.) en inalle hoofdstukken is voornamelijk demodel cultivar Igri gebruikt. Haploideplanten bevatten de helft van allegenetische informatie, die normaalgesproken in een (diploide) plantaanwezig is. Dergelijke planten zijn eenbelangrijk hulpmiddel in deplantenveredeling voor de ontwikkelingvan nieuwe rassen met gewensteeigenschappen. In veel gevallen is het nieteenvoudig om vast te stellen of een plantbepaalde nuttige eigenschappen bezit.Gewenste eigenschappen, zoals bijvoorbeeld vele soorten van ziekteresistentie, worden gecodeerd doormeerdere genen die elk verschillendemveaus van dominantie kunnen bezitten.In haploide planten is het o.a. mogelijkom de aanwezigheid van deze lastigezogenaamde polygene resistenties aan tetonen. Dit wordt het fixeren vaneigenschappen genoemd. Door integratievan via androgenese verkregen planten inde veredeling, kan de kwaliteit en deefficientie bij het ontwikkelen van nieuwe
rassen worden verbeterd.In hoofdstuk 1 wordt het belang van
een toepasbare methode voor produktievan verdubbelde haploiden nadertoegelicht. Voorts zijn in dit hoofdstuknaast de stand van zaken in de literatuurtevens de lacunes in kennis op ditvakgebiek aangegeven. Gebleken is datvoor inductie van plant produktie viamicrosporen, aan een drietal voonvaardenvoldaan moet worden: (i) hetuitgangsmateriaal moet in het laatuninucleate microsporen ontwikkelings-stadium zijn, (ii) er moet eenvoorbehandeling toegepast worden, en(iii) er moeten groeiregulatoren in hetcultuur medium worden toegediend. Dezevoorwaarden zijn in de volgendehoofdstukken nader beschreven.
In hoofdstuk 2 wordt de basis methodebeschreven, waarbij wordt gebruikgemaakt van antheren die afkomstig zijnvan planten die zijn opgegroeid ondergeconditioneerde omstandigheden. Deinvloed van een aantal factoren opregeneratie van planten, namelijk (ï) hetmicrosporen ontwikkelingsstadium in deanther, (ii) de lengte van voorbehandelingvan de anther, f iii) de samenstelling vanmedia en (liii) gebruik van aeratie,werden uitgetest. De beste regeneraüefrequenties werden verkregen, alstenminste 50% van de microsporen in hetmid-laat tot laat uninucleate stadium was,als antheren gedurende 4 dagen op eenmannitol bevattende oplossing voor-
113

Sarneuaat1ing
behandeld waren en als regelmatigzuurstof aan de cultures werd toegevoegd.Toevoeging van vitamines en caseinehydrolysaat aan het cultuunnedium heeftalleen een positief effect op microsporencultuur. Weinig of geen effect werdwaargenomen voor antheren cultuur. Bijtoepassing van microsporen cultuur inplaats van antheren cultuur, werdenminimaal vijf keer zo veel plantenverkregen, resulterend in 12,4 groeneplanten per anther.
In hoofdstuk 3 wordt het onderzoekbehandeld dat tot doel had de methodevan microsporen cultuur verder teoptimaliseren. Aangetoond werd dat deosmolaliteit tijdens de voorbehandelingen in het kiveekmedium van belang was.Verder werd vastgesteld hoe eenembryogene subpopulatie microsporenkan worden herken& In combinatie meteen optimale osmolaliteit, resulteerdestandaardlisatie van de dichtheid van desubpopulatie embryogene microsporen, in50 groene planten per anther.
In hoofdstuk 4 wordt het effect van hetvarieren van de groeistof concentratie inmicrosporen cultures getoond. In plaatsvan het algemeen gebruikte cytokinine,dat benzylaminopurine (BAP) genoemdwordt, werd het synthetische auxine 2,4-dichlorophenoxy acetic add (2,4-D)toegediend. Aangetoond werd dat zoweleen continue aanwezigheid vanconcentratie 2,4-D (10e’ M) in hetkweekmedium, als een hogereconcentratie, maar dan gedurende eenkortere tijd (10 M gedurende 1 uur),resulteerde in dezelfde regeneratiefrequenties. Er werden waarden gevondendie vergelijkbaar zijn met waarden
verkregen uit kweken met BAP (ziehoofdstuk 2 en 3). In afwezigheid van eenvoorbehandelïng van antheren is heteveneens mogelijk om, weliswaar in lagerefrequenties, plant vorming te induceren.Geconcludeerd is, dat toepassing van 1 0
M 2,4-1) gedurende 1 uur kennelijk eenmeer algemeen toepasbare methode is
voor regeneratie, aangezien deze conditiezowel bij gerst microsporen als bij kleineclusters van Daucus c.arota cellen insuspensiekiveken, leidt tot goedeplantvormïng.
Hoofdstuk 5 en 6 handelen over devoorbehandelings condities van antheren.In hoofdstuk 5 wordt de invloed vanmannitol en calcium ion concentratiestijdens de voorbehandeling op regeneratiefrequentie getoond. De beste resultatenwerden verkregen door antheren eenvoorbehandeling te geven op pre-mediumdat 30 mM CaC1, bevatte en was opgelostin mannitol tot een osmolaliteit van 440mOs/kg. Er werd tevens nagegaan inhoeverre de concentratie van abscisinezuur (ABA), een stress-gerelateerdhonnoon, een rol speelt tijdens devoorbehandeling. Een piek van endogeenABA na de eerste 24 uren vanvoorbehandeling werd waargenomen.Deze was niet gecorre]eerd met deinductie van androgenese. Met behulpvan het wegvangen van exogeen ABA ofhet toedienen van de biosynthese remmerfiuridone, werd geconstateerd dat hetniveau van exogeen ABA tussen 24 en 48uur na het begin van de voorbehandeling,een specifiek niveau moet hebben vooroptimale ontwildceling van planten.
In hoofdstuk 6 is gekeken of er sprakeis van een verouderingsproces tijdens de
114

Samenvatting
voorbehandeling van antheren en zijn dedaarvoor karakteristieke kenmerkenbestudeerd. Uit deze studie bleek dat ergeen veroudering, maar geprogrammeerde cel dood, ook wel apoptosisgenoemd, plaats vond. Op basis vanexpressie patronen verkregen met domeinspecifieke differential display’s voor “ABAresponsieve” genen, chitinase genen englucanase genen, is een model voorinductie van androgenese voorgesteld.Een centrale rol in dit model speelt hetproces van apoptosis. In aanwezigheidvan caldum ionen wordt apoptosisgeinduceerd m.b.v. caldum-afhankelijkenucleases. Doordat er geen necrotischeceldood plaats vindt, stijgt het gehaltevan exogeen ABA geleidelijk tijdens devoorbehandeling. Een stimulerende, maarniet-toxische concentratie van exogeenABA initieert de expressie van specifieke‘ABA responsieve’ genen in microsporen,dat uiteindelijk leidt tot het ontstaan vanembryogene microsporen aan het eindevan de antheren voorbehandeling.Bovendien is een waargenomen
stimulerend effect van toediening vanlipo-oligosacchariden (LOS) tijdens devoorbehandeling op plant produktie,verwerkt in het beschreven model.
In het laatste hoofdstuk wordt hetmodel van inductie van androgenese,uitgebreid bediscussieerd. Aspecten alshet microsporen ontwikkelingsstadium deaanwezigheid van de antherewand enverschillende stress-inducerende agentiaworden besproken. Tevens wordt eenvergelijking gemaakt voor inductie vanembiyogenese vanuit gameten ensomatische cellen. Daarnaast wordenefficienties tijdens gerst androgenesevergeleken met de andere modelgewassenBrassica napus en Nicotiana tahacum. Aande hand van deze informatie wordt hetembryogene type microspore en hetoptreden van het proces vanembryogenese in de ontwikkeling vanmicrosporen tot plant, bediscussieerd envergeleken met het proces dat plaats vindtaan de plant, de zygotischeembryogenese.
115


urrïcuium vitae
ietske Hoekstra werd geboren op 30juli 1963 te Amsterdam. In 1981behaalde ze haar Gymnasium- diplomaaan het Zaaxilands Lyceum te Zaandam,waarna ze in hetzelfde jaar begon met destudie biologie aan de Vrije Universiteitte Amsterdam. Tijdens haar studie werkteze hij de vakgroep Moleculaire Geneticaaan somatische cybridisatie en Agrohacterium tumefadens transformatie inLycopersiarn esculentum. Hierna onderzochtzij de invloed van ectomycorrhizae bijTilia sp. in de stad Amsterdam in eensamenwerkingsproject tussen de vak-groepen fytopathologie en Bosteelt, beidete Wageningen. Vervolgens werkte zij inhet fytopathologisch Laboratorium teBaam aan de in vitro interactie tussenLycopersiarn esculentum en Vertinitium alboatrum. Zij heeft haar hoofdvak Moleculaire Genetica in Amsterdam afgerondmet contract research naar somatischeregeneratie en cybridisatie in Brassicaoleracea; hiervoor vond een stage plaats bijprof.dr. K Glimelius te Uppsala(Zweden). In augustus 1987 verkreeg zijde titel doctorandus en tegelijkertijdondenvijsbevoegdheid voor biologie.
Het Brassica onderzoek heeft zij alsresearch medewerker van de afdelingBiotechnologie van Zaadunie B.V., teEnkhuizen voortgezet en CMS over-
dracht naar Brassica oleracea middelssomatische cybridisatie bewerkstelligt.
Vanaf april 1989 was zij in dienst bijde afdeling Planten Biotechnologie vanTNO-Voeding te Leiden. Zij is aldaarbegonnen als research medewerker enverrichtte werkzaamheden in het kadervan het ABIN-project (Adaptation ofBarley for Industrial Needs) binnen hetthema regeneratie en transformatie. Desamenwerking en een stage bij hetCarisberg Research Laboratory (Denemarken) mogen hier niet onvermeldblijven. In 1992, 1993 en de eerste helftvan 1994 heeft zij zich bezig gehoudenmet contract research en het samenwerldngsproject tussen TNO en de RULop het gebied van transformatie en demoleculaire analyse van embryogenese inOyza sativa en Hordeum vulgare. In hetkader daarvan heeft zij onderzoekverricht in de vakgroep van prof.dr. H.U.Koop in München (Duitsland). In 1993en 1994 heeft zij bovendien de functiemarktontwildelaar bekleed.
Vanaf juli 1994 was zij in dezelfdeTNO afdeling werkzaam als produkt-manager van het produkt ‘Gewasverbetering’.
Sinds april 1996 is zij werkzaam alslijnmanager ‘Gewas Transformatie’ hijMogen International N.V. te Leiden.
117


ArticlesHoekstra S., M.H. VanZijderveld, J.D.Louwerse, F. Heidekamp and F.VanderMark. Anther and microspore culiureof Hordeurn vulgare L. cv. Igri. Plant Science 86:89-96 (1992)Hoekstra S., M.H. VanZijderveld, F.Heidekamp and f. VanderMark Microsporeculture of Hôrdeum vulgare 1: the influence ofdensity and osmolarity. Plant Ceil Reports 12:661-665 (1993)Hoekatra S., M. VanZijderveld, S.VanBergen, f. VanderMark and f.Heidekamp. Genetic modification of barley forend use quality. In: Improvement of cerealquality by genetic engineering. eds. RJ. Hemyand JA Ronalds, Plenum Press, pp. 139-144(1994)Hoekstra S., S. VanBergen, IR.VanBrouwershaven, R.A. Schilperoort andf. Heidekamp. The interaction of 2,4-Dappilcalion and mannitol pretreatment inanther and microspore culture of Hordeum vulgareL. cv. Igri. J. Plant Physiol. 25: ? (1996)Hoekstra S., 1.R VauBrotxwershaven, S.Vanflergen, R.A. Schilperoort and M.Wang. Androgenesis in Hordeunt vulgare L: Therole of ABA during anther pretreatment, on asolution containing mannitol and calcium.Manuscript submitted.Hoekstra S., M. VanderHeijden, 8.Oppedijk, S. VanBergen, F. Heidekamp andR.A. Schilperoort Androgenesis in Hordeurnvulgare L: Apoptosis and the effect of lipooligosaccharides during anther pretreatment.Manuscript in preparation.
PatentsHoekstra S., A.J. Kool, M.G. Nootebos andM.M.C. Tan. Improvements in or relating toorganic systems: Method for producing CMSBrassica oleracea by protoplast fusion. PatentApplication, filed in several countries eg. GB221 1205B, fR2628601A, US5254802A
Reports and ahstractsBroerse J., K. IGngma, S. Hoekstra, f.Heidekamp, P. Sluimer, T. VandeSande and
J. Bunders. Biotechnology and cerealproduction for developing countries. EuropeanParliament, Directorate General for Research,the STOA Programme, pp. 37-51 (1994)Hoekstra S., M.H. VanZijderveld, N.J.Ruys, f. Heidekamp and F. VanderMarkMicrospore.derived cellsu.spensions andprotoplast isolation from barley. Proceedings ofEmbo workshop ‘Molecular basis of plantembryogenesis”, Doonverth, Netherlands June12-19 (1990) and of 7th IAPTC Symposium,Amsterdam, Netherlands June 24-29 (1990)
pp. A1-62Hoekstra S., M.H. VanZijderveld and F.VanderMark. Barley celisuspension culturesderived from isolated microspores and theirploidy level. Abstract of 8th InternationalProtoplast Symposium, Uppsala. Sweden June16-20(1991) pp. 216Hoekstra S., M.H. VanZijderveld, J.D.Louwerse, f.L. Olsen, f. Heidekamp and f.VanderMark. Anther and microspore cultureof Hardeurn vulgare cv. Igri. Acta BotanicaNeerlandica (1992)Hoekstra S., M.H. VanZijderveld and f.VanderMark. Microspore culture of Hordeumutilgare L. Abstract of l3th Eucarpia Congress,Angers, france July 6-11(1992) pp. 167lloekstra S., M.H. VanZijderveld, f.Heidekamp and F. VanderMark Microsporeculture of Hardcum vulgare 1. Abstract ofGordon Conference, Plant Celi and flssueCulture. Wolfeboro, NH, USA June 13-18(1993) pp.A.16Hoekstra S. Basic and applied aspects of themicrospore culture system in Hordeum vulgare 1Workshop “Gametic embryogenesis” As,NonvayJune 24-27 (1993)Hoekstra S., 1. VanBrouwershaven, S.VanBergen and f. Heidekamp. Induction ofdifferentiation in microspores of Hardcumvutgare 1 Abstracts sth International Congress
ublications
119

of Plant Tissue and Ce!! Culture, Pirenze, Italy,June 12-17, (1994) pp. S3-44Hoekstra S., M. VanderHeijden, 8.0.Oppedijk, I.R. Vanflrouwershaven, S.VanBergen, F. Heidekamp and R.A.
Schilperoort. An Apoptosis-based mode! forthe induction of androgenesis in Hordeurnvulgare L Society for In Vitro Biology, June 22-27(1996)
120

awoord
raag wil ik van deze gelegenheidgebruik maken om een aantal mensenerkentelijk te zijn.
De mensen die het TNO- lab op de2e bevolkten. Speciaal wil ik noemen:Marion met wie ik samen heel wat‘supers’ bekeken heb en enormehoeveelheden suspensies versjouwd enovergezet heb, Jeanine met wie iksamen vele discussies had ‘zou jij ditnou een late of een bi noemen’ en metwie ik vele microsporen blauw en somsook aan barrels geschoten heb, Fritsvoor zijn enthousiasme en zeer gewaardeerde kritische noot, en Jan V. dievoor continue goede kwaliteit donorenzorgde en samen met San de thripsente lijf ging.
Lok who gave me an introduction inandrogenesis by transforming aleuronprotoplasts in the middie of the night,and his colleagues Preben and Søren ofCarisberg.
Anke en Sas als de harde en gezeffigekern van de rij stgroep: eerst als burenop de 2e en later voordeurdeler op debegane grond; bij Harry zal ik tochvaak terug moeten denken aan ‘hoe zathet nou ook alweer met die ELS’...
En natuurlijk San (we hebben heelwat meegemaakt), Ilse (z666 snel),
Berry (waar een boor al niet goed vooris), Maurice (die éne gel) van demicrosporen gerstclub, en de enerverende samenwerking met Mei. Enuiteraard vele anderen, die het leven inhet Clusius tot een aangenaam toevenmaakten.
Naast ‘het Leidse’, heb ik deafgelopen jaren met veel plezier mijncontacten met de industrie onderhouden, die mij mijn theorien overhaploiden in praktijk laten zien.
De charme van het internationalenetwerk met mensen als Erwin, Alisher,Bemd, Yves, Eva, Swapan, Masaharu,
Sule, Hans Ulrich, Sasha, Ken, Debra,is een onmeetbare stimulans voor hetonderzoek.
Graag wil ik mijn ouders bedankenvoor hun eeuwige bereidheid om tehelpen en de vele vrijdagen dat jullie opNils pasten, zodat ik de handen wijhad om te schrijven.
Tot slot wil ik vooral Uh, maar ookNils bedanken voor het koffie met ietslekkers brengen in de vele “labweekenden’. Jullie gezelschap, steun,en relativeringsvermogen was onmisbaar om deze vrijetijdsbesteding zo afte kunnen ronden
121


Stellingen
1. Integratie van androgenese in de plantenveredeling wordt belemmerd door
onderschatting van complexiteit van het biologische proces en daardoor ook van
de methodiek.
2. 2,4-D is een stof, die embryogenese induceert in plantecellen, mits het in een
niet-physiologische concentratie en bepaalde tijdsduur toegepast wordt.Dit proefschrift.
3. Geprogrammeerde celdood of apoptosis ten gevolge van specifieke stresscondities, is nodig voor inductie van in vitro embryogenese.Dit proefschrift.
4. De conclusie van Zald en Dickenson (1991) en Szakacs en Bamabas (1995), dat
een symmetrische eerste deling voor de ontwildeling van microspore tot plant inBrassica napus van belang is, is niet onomstotelijk bewezen.Zaki MA.M. and H.G. Dickenson. Sex. Plant Reprod. 4:48-55 (1991)Szakcs E. and B. Barnabâs. Euphyuca 83: 209-213 (1995)
5. Microsporen transformatie in combinatie met pollen maturatie zal veelarbeidsintensieve weefseikweek overbodig maken.Ahven A., N. Efler, M. Kastler, RM. Benito Moreno and E. Heberle-Bors. Phys. Plant 79: 194-
196(1990)
6. Voor het produceren van stabiel getransformeerde planten is toepassing vanDNA overdracht m.b.v. Agrohacterium twnefaciens te prefereren boven hetdeeltjesgeweer.Hiei Y., S. Ohta, T. Komari and T. Kumashiro. Plant J. 6: 271-282 (1994)Christou P., T.L. Ford and M. Kofron. Biotechnology 9: 957-962 (1991)
7. Het aantal nïet-detecteerbare differenüals, die gegenereerd zijn met mRNAdifferenüal display middels PCR (Liang en Pardee, 1992) kan verlaagd worden,wanneer in situ hybridisatie als detectie methode gebruikt wordt.Liang P. andA.B. Pardee. Science 257: 967-971 (1992)
8. In situ hybridatie experimenten waarbij laag-moleculaire stoffen als IAAsubcellulair worden gelocaliseerd, zijn zeer discutabel.Ohmiya A. And T. Hayashi. Phys. Plant. 85: 439-445

9. Niet iedere persoon die affiniteit toont voor weefselkweek is per definitie ‘a
person sidiled in the art’.160/89 (Harvard) 0. J. EPO 268 (1992)
10. Educatie van vandaag is van essentieel belang voor een florerende economie in
de toekomst.
11. Effectief omgaan met de overvloed van data, beschikbaar via electronische
netwerken, waagt op termijn een aanpassing in de werkwijze.
12. Een onderzoeker is een eigenwijze doorzetter, die lang niet altijd open staat voor
kritiek.
Stellingen behorend bij het proefschrift: ‘Androgenesis in Hordeum vutgare L’ doorSietske Hoekstra, Leiden, 1 juli 1996.
























