De rol van viscerale adipokines bij het ontstaan van...
60
De rol van viscerale adipokines bij het ontstaan van steatohepatitis bij morbide obese patiënten Sebastiaan Colenbie Verhandeling ingediend tot het verkrijgen van de graad van Master in de Biomedische Wetenschappen Promotor: Prof. Dr. Johannes Ruige Begeleidster: drs. Marlies Bekaert Vakgroep Endocrinologie Academiejaar 2013-2014
Transcript of De rol van viscerale adipokines bij het ontstaan van...

De rol van viscerale adipokines bij het ontstaan van
steatohepatitis bij morbide obese patiënten
Sebastiaan Colenbie
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Prof. Dr. Johannes Ruige Begeleidster: drs. Marlies Bekaert Vakgroep Endocrinologie
Academiejaar 2013-2014


De rol van viscerale adipokines bij het ontstaan van
steatohepatitis bij morbide obese patiënten
Sebastiaan Colenbie
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Prof. Dr. Johannes Ruige Begeleidster: drs. Marlies Bekaert Vakgroep Endocrinologie
Academiejaar 2013-2014

“De auteur en de promotor geven de toelating deze masterproef voor consultatie
beschikbaar te stellen en delen ervan te kopiëren voor persoonlijk gebruik. Elk ander
gebruik valt onder de beperkingen van het auteursrecht, in het bijzonder met
betrekking tot de verplichting uitdrukkelijk de bron te vermelden bij het aanhalen van
resultaten uit deze masterproef.”
Datum
(handtekening student) (handtekening promotor)
(Naam student) (Naam promotor)

Voorwoord
Via dit voorwoord had ik graag een aantal mensen bedankt die het mogelijk hebben gemaakt
om deze thesis te kunnen realiseren.
Vooreerst wil ik Prof. Dr. Ruige bedanken voor de kans die hij me bood om deze thesis met
een actueel en boeiend onderwerp te mogen aanvangen. Ik heb het geluk gehad veel te mogen
leren van zijn academische ervaring en kennis.
Ook wil ik Prof. Dr. Ouwens bedanken die het mede mogelijk heeft gemaakt om dit
onderwerp, dat gebaseerd is op voorgaande klinische data, te kunnen aanvangen. Daarnaast
wil ik haar bedanken voor haar brede inzicht en kennis betreffende de insulinesignalering, die
van onmisbaar belang was.
Graag had ik ook Prof. Dr. Vanhaecke bedankt die het materiaal en de accommodatie van
departement FAFY van de Faculteit Geneeskunde en Farmacie ter beschikking heeft gesteld
voor het onderzoek van deze thesis. Alsook wil ik alle laboranten en PhD studenten van het
departement bedanken die altijd vriendelijk waren en aan wie ik altijd uitleg mocht vragen.
Mijn begeleidster drs. Marlies Bekaert wil ik bedanken voor de goede begeleiding gedurende
mijn thesis jaar. Bedankt voor het geduld waarmee je mij diverse laboratorium vaardigheden
hebt aangeleerd en bedankt voor het antwoorden op de vele vragen. Door uw kennis en goede
uitleg kon ik zelfstandig werken aan mijn thesisonderzoek.
Ten slotte wil ik mijn ouders en broer bedanken voor hun onvoorwaardelijke steun gedurende
mijn hele opleiding.

Inhoudstafel
Lijst met gebruikte afkortingen
Samenvatting ..........................................................................................................................1
Summary ................................................................................................................................2
1 Inleiding ..........................................................................................................................3
1.1 Obesitas....................................................................................................................3
1.2 Vetweefsel ...............................................................................................................3
1.2.1 Anatomie...........................................................................................................3
1.2.2 Distributie .........................................................................................................4
1.3 Energiehomeostase ...................................................................................................5
1.3.1 Lipide metabolisme ...........................................................................................5
1.3.2 Glucose metabolisme ........................................................................................5
1.3.2.1 Regulatie van de gluconeogenese ...................................................................6
1.3.3 Insulinereceptor .................................................................................................8
1.4 Metabool syndroom ..................................................................................................8
1.5 Niet alcoholische steatohepatitis (NASH) .................................................................9
1.5.1 Algemeen ..........................................................................................................9
1.5.2 Lipide metabolisme, lipotoxiciteit en oxidatieve stress .................................... 10
1.6 Adipokines ............................................................................................................. 11
1.6.1 Algemeen ........................................................................................................ 11
1.6.2 Adipokines die de Wnt signalering moduleren ................................................. 12
1.6.2.1 Wnt – β-catenine signaal pathway ................................................................ 12
1.6.2.2 Wnt signalering, gluconeogenese, insulinesignalering en lipide metabolisme:
het verband ................................................................................................................ 13
1.6.2.3 Secreted frizzled-related proteïne 4 (SFRP4) ................................................ 14
1.6.2.4 Dickkopf 1 (DKK1) ..................................................................................... 15
1.6.3 Andere adipokines verwant met obesitas, NAFLD en insulineresistentie ......... 15

1.7 Hypothese en doelstelling van dit onderzoek .......................................................... 16
2 Materialen en Methoden ................................................................................................ 18
2.1 Modelsysteem: De humane lever cellijn HepaRG® ................................................. 18
2.2 Celcultuur............................................................................................................... 18
2.2.1 Dag 0: uitplaten van de HepaRG® cellen ......................................................... 18
2.2.2 Verversen van medium op dag 1, dag 2 en dag 3 ............................................. 19
2.2.3 24 uur blootstelling aan laag glucose (0.5 mM) met of zonder insuline (100 nM)
op dag 7 ........................................................................................................................ 20
2.2.4 24 uur blootstelling aan laag glucose (0.5 mM) en adipokine met of zonder
insuline (100 nM) op dag 8 ............................................................................................ 21
2.2.4.1 Experiment met SFRP4 blootstelling ........................................................... 21
2.2.4.2 Experiment met DKK1 blootstelling ............................................................ 22
2.3 Real-time quantitative Polymerase Chain Reaction (qPCR) .................................... 23
2.3.1 qPCR, algemeen .............................................................................................. 23
2.3.2 RNA isolatie en opzuivering ............................................................................ 24
2.3.2.1 Isolatie van RNA uit HepaRG® cellijn op dag 9 ........................................... 24
2.3.2.2 Homogenisatie van het lysaat ....................................................................... 25
2.3.2.3 RNA extractie .............................................................................................. 25
2.3.2.4 DNase I behandeling .................................................................................... 25
2.3.2.5 RNA recovery .............................................................................................. 25
2.3.2.6 RNA kwaliteitscontrole en concentratie bepaling. ........................................ 26
2.3.3 cDNA aanmaak ............................................................................................... 26
2.3.3.1 cDNA clean up ............................................................................................ 27
2.3.4 qPCR meting ................................................................................................... 27
2.3.4.1 Selectie van doelwitgenen ............................................................................ 27
2.3.4.2 Selectie van referentiegenen ......................................................................... 28
2.3.4.3 Standaardcurve en bepaling van primer efficiëntie ....................................... 30
2.3.4.4 qPCR reactie ................................................................................................ 30

2.4 Statistische analyse ................................................................................................. 31
3 Resultaten...................................................................................................................... 32
3.1 Resultaten experiment SFRP4 ................................................................................ 32
3.1.1 Referentiegenen .............................................................................................. 32
3.1.2 Gluconeogenese, expressieniveaus van PCK1, G6P en FBP1 .......................... 33
3.1.3 Insulinesignalering, expressieniveaus van IR, IRS1 en IRS2 ............................ 34
3.1.4 Lipide metabolisme, expressieniveaus van MTTP, FAS en SREBF1 ............... 35
3.2 Resultaten experiment DKK1 ................................................................................. 35
3.2.1 Referentiegenen .............................................................................................. 35
3.2.2 Gluconeogenese, expressieniveaus van PCK1, G6P en FBP1 .......................... 36
4 Discussie ....................................................................................................................... 37
4.1 Belangrijkste resultaten .......................................................................................... 37
4.2 Belang van onderzoek en hypothetische verklaringen van de resultaten. ................. 37
4.3 Mogelijke toekomstperspectieven ........................................................................... 43
4.4 Besluit .................................................................................................................... 43
5 Referenties .................................................................................................................... 45

Lijst met gebruikte afkortingen
ACTB actine β
Akt/PKB proteïne kinase B
ALT alanine aminotransferase
APC adenomatous polyposis coli
apoB apolipoproteïne B
AST aspartaat aminotransferase
B2M β2-microglobuline
BMI body mass index
bp basenparen
BSA Bovine Serum Albumin
cDNA complementair desoxyribonucleïnezuur
ChREBP carbohydrate response element binding protein
CK1α caseïne kinase 1α
Ct threshold cyclus
CT computertomografie
DAG diacylglycerol
DKK1 dickkopf 1
DM 2 diabetes mellitus type 2
DMEM Dulbecco’s Modified Eagle Medium
DNA desoxyribonucleïnezuur
Dvl Dishevelled
ER endoplasmatisch reticulum
FAS Fatty Acid Synthase
FBP1 fructose-1,6-bisfosfatase
FoxO1 forkhead O box 1
Fz frizzled receptor
GAPDH glyceraldehyde-3-fosfaat dehydrogenase
G6P glucose-6-fosfatase
GS glycogeen synthase
GSK-3 glycogeen synthase kinase-3
HMBS hydroxymethylbilane synthase
HOMA-IR homeostasis model assessment for insulin resistance

HSC hepatische stellaat cellen
IGF insuline-achtige groeifactor
IL-6 interleukine-6
IR insulinereceptor
IRE insuline responsieve elementen
IRS insulinereceptor substraat
LDL low density lipoproteïne
LEF lymphoid enhancer factor
LRP6 low density lipoproteïne receptor 6
MR magnetische resonantie
mTOR mammalian target of rapamycin
MTTP microsomaal triglyceride transfer proteïne
NAFLD niet alcoholische vette leverziekte
NASH niet alcoholische steatohepatitis
NRQ genormaliseerde relatieve kwantificatie
OGTT orale glucose tolerantie test
PBS phosphate buffered saline
PC pyruvaat carboxylase
PCK1 fosfoënolpyruvaat carboxykinase 1
PDPK1 fosfoïnositide afhankelijk kinase 1
PGC-1α peroxisome proliferative activated receptor-γ coactivator 1α
PI(3)K fosfoïnositide 3-kinase
PIP2 fosfatidylinositol-(3,4,5)-trisfosfaat
PIP3 fosfatidylinositol-(4,5)-bisfosfaat
qPCR Real-time quantitative Polymerase Chain Reaction
RNA ribonucleïnezuur
ROS reactive oxygen species
SEM Standard Error of the Mean
SFRP4 secreted frizzled-related proteïne 4
SNP single-nucleotide-poylymorphism
SREBP-1
SREBF1
sterol receptor bindend proteïne-1
sterol regulatory element binding transcription factor 1
SVW subcutaan vetweefsel

TAG triacylglycerolen
TCF T cel factor
TG triglyceriden
TNF-α tumor necrose factor α
UBC ubiquitine C
UPR unfolded protein response
VLDL very low density lipoproteïnen
VVW visceraal vetweefsel
VVZ vrije vetzuren
WHO World Health Organization

1
Samenvatting
Obesitas is een wereldwijde epidemie die gekarakteriseerd wordt door een veralgemeende
insulineresistentie en een risicofactor vormt voor het ontstaan van talrijke complicaties,
waaronder diabetes mellitus type 2 (DM 2) en niet alcoholische vette leverziekte (NAFLD).
Niet alcoholische vette leverontsteking (NASH), een gevorderde vorm van NAFLD, vormt de
hepatische component van het metabool syndroom en wordt gekenmerkt door een verstoorde
insulinegevoeligheid, een verstoorde gluconeogenese en een verstoord lipide en lipoproteïne
metabolisme. Viscerale adipokines, eiwitten vrijgesteld uit visceraal vetweefsel dat portaal
gedraineerd wordt, zouden een invloed kunnen hebben op het energiemetabolisme van de
lever en aldus een rol spelen in het ontstaan van obesitas gerelateerde complicaties. Bij
obesitas is het secretiepatroon van deze adipokines namelijk sterk ontregeld. Wnt signalering
lijkt belangrijke fysiologische functies te beïnvloeden in tal van organen en weefsels die een
rol kunnen spelen in de pathofysiologie van obesitas en DM 2. Verschillende studies
suggereren daarbij een mogelijke rol van Wnt op de energiehomeostase van de lever. In nog
niet gepubliceerde data werd een verhoogde expressie gezien van de Wnt regulatoren secreted
frizzled-related proteïne 4 (SFRP4) en dickkopf 1 (DKK1), in het visceraal vetweefsel van
morbide obese DM 2 patiënten. In deze thesis werd de rol van SFRP4 en DKK1 bestudeerd
op de gluconeogenese van een humane HepaRG® lever cellijn. Parallel werd de rol van
SFRP4 op de insulinesignalering en het lipide metabolisme bestudeerd. Na een 24 uur
durende blootstelling met 100 nM insuline onder vasten condities (0.5 mM glucose), werd de
cellijn gedurende 24 uur blootgesteld aan SFRP4/DKK1. Met behulp van Real-time
quantitative Polymerase Chain Reaction (qPCR), werd de expressie bestudeerd van
sleutelgenen uit de gluconeogenese, insulinesignalering en lipide metabolisme. De resultaten
tonen dat SFRP4 (100 ng/ml), in afwezigheid van insuline, de expressie van gluconeogene
genen fosfoënolpyruvaat carboxykinase 1 (PCK1) en fructose-1,6-bisfosfatase 1 (FBP1)
reduceert. Daarnaast werd gezien dat SFRP4 (100 ng/ml) de insuline (100 nM) gemedieerde
onderdrukking van PCK1 en FBP1 grotendeels lijkt op te heffen. Deze bevindingen
bevestigen de invloed van Wnt signalering op de gluconeogenese en suggereren een
mogelijke rol van SFRP4 in de hepatische glucose productie door de insuline gemedieerde
inhibitie van de gluconeogenese te onderdrukken.

2
Summary
Background: Obesity forms an important risk factor for the onset of multiple complications
such as diabetes mellitus type 2 (DM 2) and non-alcoholic steatohepatitis (NASH). The latter
is characterized by disturbances of the gluconeogenesis, insulin signaling, lipid and
lipoprotein metabolism. Visceral adipokines or proteins secreted from the visceral adipose
tissue, which has a portal drainage system, could have an influence on the metabolism of the
liver. Several studies suggest a possible role of Wnt on the energy homeostasis of the liver.
Unpublished data have shown an increased expression of Wnt regulators secreted frizzled-
related protein 4 (SFRP4) and dickkopf 1 (DKK1), in the visceral adipose tissue of morbid
obese DM 2 patients.
Methods: The role of SFRP4 and DKK1 on the gluconeogenesis was studied in vitro by
means of a human HepaRG® liver cell line. Parallel the role of SFRP4 on the insulin signaling
and lipid metabolism was studied. After exposure with insulin (100 nM) during fasting
conditions (0.5 mM glucose), the cell line was exposed to SFRP4/ DKK1. Through qPCR, the
expression of key genes from the gluconeogenesis, insulin signaling and lipid metabolism was
studied.
Results: In the absence of insulin, SFRP4 (100 ng/ml) reduces the expression of
phosphhoenolpyruvate carboxykinase 1 (PCK1) and fructose-1,6-bisphosphatase 1 (FBP1).
Furthermore SFRP4 (100 ng/ml) seems to abolish most part of the insulin mediated repression
of PCK1 and FBP1 expression.
Conclusions: These data confirm the influence of Wnt signaling on the gluconeogenesis and
suggest a possible role of SFRP4 in the hepatic glucose production.

3
1 Inleiding
1.1 Obesitas
Obesitas, gedefinieerd door een Body mass index (BMI) groter dan 30 kg/m², is een
wereldwijde epidemie die deels het gevolg is van een genetische voorgeschiktheid, een
sedentaire levensstijl en een vetrijk of energierijk dieet waarbij de calorie inname het verbruik
overtreft (1-4). Er ontstaat een continue positieve energiebalans, wat resulteert in een
schadelijke gewichtstoename. De graad en distributie van de excessieve vetaccumulatie bij
obese individuen is hierbij onderling verschillend (1, 3, 5). De relatie tussen BMI en
lichaamsvet kan verschillen door ras, etniciteit, leeftijd en geeft geen informatie over de
vetdistributie, maakt geen onderscheid tussen vetmassa en vetvrije massa, waardoor deze
meetmethode soms bekritiseerd wordt. Alternatieve meetmethoden zijn o.a. taille omtrek,
taille-heup ratio, computertomografie (CT), dual-energy X-ray absorptiometrie en
magnetische resonantie (MR) (3, 6-8). Populatie onderzoeken toonden aan dat de prevalentie
van obesitas omstreeks 1980 dramatisch vlug is beginnen stijgen. De hoogste BMI waarden
worden waargenomen in de VS, EU en het Oostelijke Middellandse Zeegebied (1-3). Voor
2015 voorspelt de World Health Organization (WHO) dat meer dan 1.5 miljard mensen
overgewicht zullen hebben (2). Obesitas zou verantwoordelijk zijn voor 0.7 % tot 2.8 % van
de gezondheidsuitgaven van een land, wat nog steeds toeneemt, en de individuele medische
kosten voor obese personen zouden 30 % hoger zijn dan voor gezonde slanke personen (9).
Momenteel vormt obesitas wereldwijd de 5de hoogste risicofactor voor sterfte en is het
gerelateerd met complicaties zoals diabetes mellitus type 2 (DM 2), niet alcoholische vette
leverziekte (NAFLD), niet alcoholische steatohepatitis (NASH), cardiovasculaire ziekten,
osteoartritis, kanker, etc. (2, 3, 6, 8, 10, 11). De metabole complicaties als gevolg van de vet
accumulatie worden mede beïnvloed door de locatie en distributie van de overmaat aan vet in
het lichaam. Intra-abdominaal vet wordt namelijk beschouwd een belangrijke determinant te
zijn voor gezondheidsrisico’s (3, 5, 7, 8, 12, 13).
1.2 Vetweefsel
1.2.1 Anatomie
De primaire functie van het wit vetweefsel bestaat erin om de overmaat aan energie te
stockeren als triglyceriden (TG), die in tijden van tekort opnieuw kunnen aangewend worden
als energiebron voor verschillende weefsels (5, 14, 15). In de adipocyten zijn de lipiden
georganiseerd in één grote centrale druppel die het grootste deel van de intracellulaire ruimte

4
bezet, waardoor het cytoplasma en de kern herleid worden tot een dunne rand (14, 16). De
vetcellen verzamelen in lobullen die gescheiden zijn door bindweefsel septa (14). Het aantal
vetcellen, bepaald gedurende de kindertijd en de vroege adolescentie, wordt verondersteld
constant te blijven bij adulten, zowel in slanke als in obese individuen. Van de adulte
adipocyten wordt 10 % jaarlijks hernieuwd, uit mesenchymale stamcellen en of pre-
adipocyten, ongeacht de leeftijd of BMI. Overgewicht wordt gekenmerkt door een verminderd
expansie vermogen van het vetweefsel, wat leidt tot een schadelijke vergroting (hypertrofie)
van vetcellen die o.a. inflammatie en ectopische lipide accumulatie bevorderen. Dit wordt
verondersteld het gevolg te zijn van een verminderde rekrutering en differentiatie van
vetprecursor cellen. Een toename van de vetmassa bij adulten is dus eerder het gevolg van een
hypertrofie van bestaande adipocyten dan van een verhoogde vorming van nieuwe adipocyten
(hyperplasie) (13, 17, 18). Anatomisch wordt het abdominaal vetweefsel onderverdeeld in 2
regio’s die elk hun eigen, metabole, endocriene, paracriene en autocriene eigenschappen
hebben, namelijk het intra-abdominaal of visceraal vetweefsel (VVW) en subcutaan
vetweefsel (SVW) (7, 8, 13, 14). Terwijl het SVW een systemische circulatie heeft, wordt het
VVW ook portaal gedraineerd (5, 7, 8, 13). Daarenboven zouden de viscerale adipocyten
gevoeliger zijn voor lipolyse (vetafbraak) dan subcutane adipocyten (3, 7, 16).
1.2.2 Distributie
De vetdistributie wordt o.a. beïnvloed door geslacht, leeftijd, totaal lichaamsgewicht en
genotype (3, 8, 12). Mannen hebben namelijk minder “totaal” lichaamsvet en meer vetvrije
massa dan vrouwen, maar zouden over meer VVW beschikken (3, 8, 12). Daar waar mannen
een “androïde” of abdominale vetdistributie hebben, vertonen vrouwen eerder een “gynoïde”
of gluteofemorale vetdistributie (12, 16, 19). Naast de totale vetmassa dragen de verschillende
compartimenten elk apart en in verschillende mate bij tot de metabole complicaties van
obesitas (8). Bij het verouderen neemt het VVW toe en dit meer bij mannen dan bij vrouwen
(3, 7, 16). Hoewel nog controversieel zouden geslachtshormonen, zowel circulerend als lokaal
in het vetweefsel, een invloed hebben op de distributie van vet (7, 12). Testosteron zou de
adipocyt differentiatie reduceren, de lipide accumulatie inhiberen en de lipolyse verhogen,
daar waar oestradiol (oestrogeen) de adipocyt differentie en lipide opslag zou verhogen. Lage
testosteron en verhoogde oestradiol gehaltes worden frequent aangetroffen bij morbide
mannen (20). Het verschil tussen VVW en SVW inzake distributie, histologie, metabole
activiteit, associatie met metabole risico factoren lijkt dus deels een verklaring te zijn voor het
risicoverschil op complicaties tussen beide geslachten (7, 8, 12).

5
1.3 Energiehomeostase
1.3.1 Lipide metabolisme
Energie, afkomstig uit voeding, wordt enerzijds gebruikt voor verplichte biologische functies
zoals het behouden van lichaamstemperatuur, metabolisme en weefselherstel terwijl
anderzijds ook energie nodig is bij fysieke activiteit en interactie met de omgeving (1, 5). De
overschot aan energie zal tijdens de lipogenese (vetopbouw) gestockeerd worden in het
vetweefsel onder de vorm van triacylglycerolen (TAG) of TG (10, 14, 16). Deze bestaan uit 3
vetzuurmoleculen veresterd op een molecule glycerol (14, 21). De TG in vetcellen kunnen
eveneens afkomstig zijn van de novo lipogenese of de opname van vrije vetzuren (VVZ) uit
het plasma. Bij de mens vormen de chylomicronen (afkomstig van de darm) en de very low
denstiy lipoproteïnen (VLDL) (afkomstig van de lever), beiden transporteiwitten voor vetten,
een belangrijke bron van TG, die echter onder deze vorm niet kunnen penetreren door het
endotheel van de capillairen in het vetweefsel (10, 16). De adipocyten stellen daarom
lipoproteïne lipase (LPL) vrij in het lumen van de capillairen waardoor de TG hydrolyseren in
glycerol en VVZ. Deze VVZ kunnen dan wel door de vetcellen opgenomen worden, waar ze
opnieuw tot TG worden gesynthetiseerd (10, 14, 16). Een belangrijk lipogeen enzym is het
Fatty Acid Synthase (FAS) dat mede gecontroleerd wordt door het sterol receptor bindend
proteïne-1 (SREBP-1), een transcriptiefactor en regulator van de lipogenese (22-24). Bij
energie nood zullen de intracellulaire TG tijdens de lipolyse gehydrolyseerd worden door drie
belangrijke lipasen: het vetweefsel TG lipase, het hormoon sensitief lipase en het mono-acyl-
glycerol lipase (14, 16). De VVZ kunnen dan onder meer naar het spierweefsel en de lever
getransporteerd worden, waar ze via de β-oxidatie in de matrix van de mitochondriën instaan
voor energie productie met vorming van adenosine trifosfaat (ATP) (14, 21). Het glycerol kan
in de lever opnieuw gebruikt worden voor de novo synthese van TG die dan verpakt en
vrijgelaten worden onder de vorm van VLDL. Het lever VLDL bestaat uit een TG rijke kern
met een oppervlak van fosfolipiden. Het microsomaal triglyceride transfer proteïne (MTTP) is
een endoplasmatisch reticulum (ER) eiwit die de transfer van lipiden naar het hydrofobe
apolipoproteïne B (apoB) in de lever faciliteert en zo degradatie van apoB vermijdt. Dit apoB
is vereist voor de synthese van VLDL en chylomicronen. Catecholamines, natriuretische
peptiden en insuline vormen belangrijke regulatoren van de lipolyse (16, 22-25).
1.3.2 Glucose metabolisme
Koolhydraten, in het bijzonder glucose, zijn de voornaamste energiebronnen voor de mens
(10, 19, 21, 26). Zowel extreem lage als chronisch verhoogde bloedglucose waarden, zijn

6
geassocieerd met complicaties (10, 19). Om een euglycemie of normale plasma glucose
waarde (4-6.1 mmol/l) te behouden, is een strikte regulatie in opname, verbruik en productie
van glucose vereist. Onder normale fysiologische condities ontstaat er postprandiaal een
stijging van de bloedglucose spiegel (hyperglycemie), waarbij glucose via de lage affiniteit
glucose transporter, GLUT-2, de β cellen van de endocriene pancreas binnentreedt zodat de
insuline secretie gestimuleerd wordt (10, 19, 21, 24, 26, 27). Insuline zorgt voor diffusie,
opname, afbraak (glycolyse) en stockage van glucose (glycogenese) in insuline gevoelige
weefsels (19, 21, 24, 27). Hierbij vormt het skeletspierweefsel het belangrijkste postprandiale
orgaan voor glycolyse (19, 24). De glucose transporter GLUT-4 wordt eveneens gereguleerd
door insuline. In aanwezigheid van insuline zal in het vet-, skeletspier- en hartspierweefsel,
het aantal GLUT-4 moleculen in de membraan alsook de snelheid van glucose transport in de
cellen toenemen (10, 15, 19, 24, 27). Daarbij zal naast de glycolyse de conversie van glucose
in glycogeen in spieren en lever bevorderd worden, door activatie van het glycogeen synthase
(GS), terwijl de afbraak van glycogeen (glycogenolyse) afneemt. Samen met een gedaalde
vorming van glucose (gluconeogenese), uit o.a. glycerol, lactaat en aminozuren, in de lever
zal dit uiteindelijk resulteren in een daling van de bloedglucose (10, 21, 23, 24, 27). In het
vetweefsel en in mindere mate in de lever zal het opgenomen glucose ook dienen als substraat
waarbij de lipogenese gestimuleerd wordt (10, 14, 21, 24). Bij daling van de bloedglucose
(hypoglycemie) tijdens vasten en of fysieke inspanning wordt de vrijstelling van insuline
onderdrukt en de secretie van glucagon door de α cellen van de pancreas bevorderd. Glucagon
zal, na binding op zijn trimere G-eiwit gekoppelde receptor, via de levercellen de
glycogenolyse en de gluconeogenese stimuleren, terwijl de glycolyse en de glycogenese
worden geïnhibeerd. Dit leidt tot een euglycemie waarbij het hepatisch gevormde glucose
door insuline onafhankelijke weefsels zoals de hersenen kan opgenomen worden. Daar waar
de glycogenolyse reeds vlug begint, is de gluconeogenese belangrijker bij langdurig vasten
(10, 19, 21, 24, 26, 28). In het vetweefsel wordt dan eveneens de lipolyse bevorderd (10, 21,
24).
1.3.2.1 Regulatie van de gluconeogenese
Glucagon en glucorticoïden, de voornaamste activatoren van de gluconeogenese, moduleren
talrijke transcriptiefactoren, zoals Forkhead O box 1 (FoxO1), die de expressie van
gluconeogene genen reguleren (21, 28). Hierbij spelen 4 enzymen een cruciale rol:
fosfoënolpyruvaat carboxykinase 1 (PCK1), pyruvaat carboxylase (PC), fructose-1,6-
bisfosfatase (FBP1) en glucose-6-fosfatase (G6P) (15, 21, 26, 28). In de eerste stap (zie
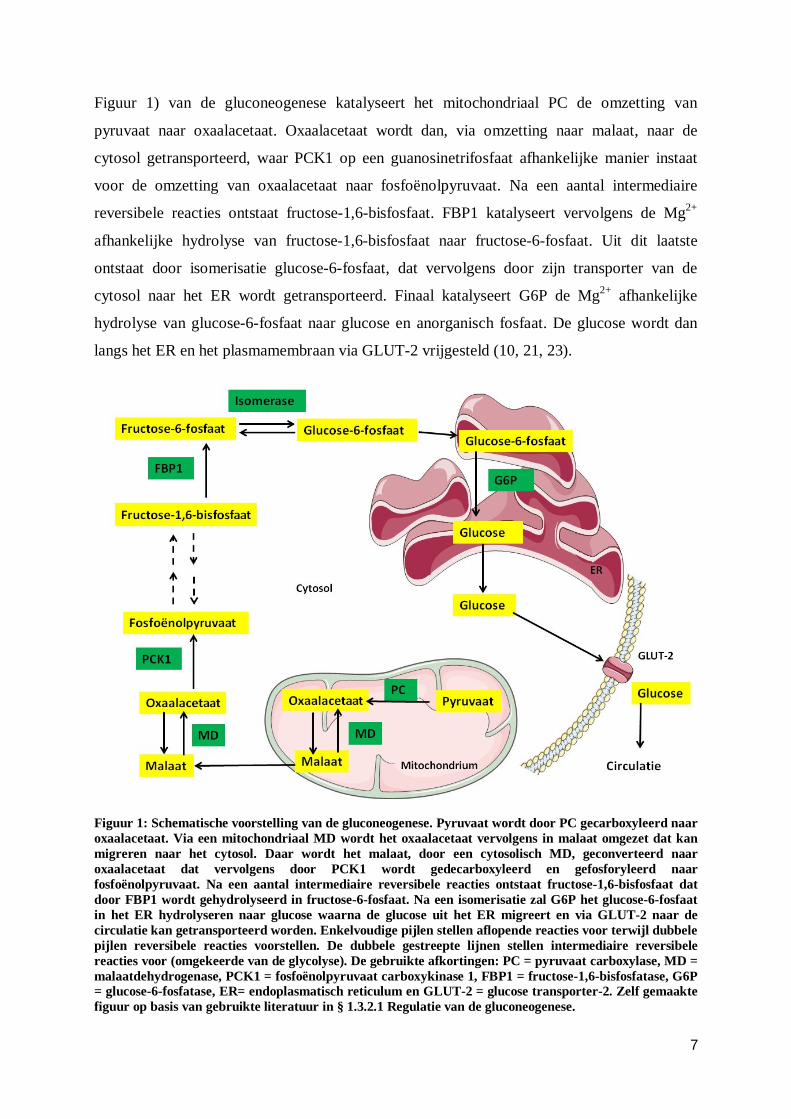
7
Figuur 1) van de gluconeogenese katalyseert het mitochondriaal PC de omzetting van
pyruvaat naar oxaalacetaat. Oxaalacetaat wordt dan, via omzetting naar malaat, naar de
cytosol getransporteerd, waar PCK1 op een guanosinetrifosfaat afhankelijke manier instaat
voor de omzetting van oxaalacetaat naar fosfoënolpyruvaat. Na een aantal intermediaire
reversibele reacties ontstaat fructose-1,6-bisfosfaat. FBP1 katalyseert vervolgens de Mg2+
afhankelijke hydrolyse van fructose-1,6-bisfosfaat naar fructose-6-fosfaat. Uit dit laatste
ontstaat door isomerisatie glucose-6-fosfaat, dat vervolgens door zijn transporter van de
cytosol naar het ER wordt getransporteerd. Finaal katalyseert G6P de Mg2+ afhankelijke
hydrolyse van glucose-6-fosfaat naar glucose en anorganisch fosfaat. De glucose wordt dan
langs het ER en het plasmamembraan via GLUT-2 vrijgesteld (10, 21, 23).
Figuur 1: Schematische voorstelling van de gluconeogenese. Pyruvaat wordt door PC gecarboxyleerd naar oxaalacetaat. Via een mitochondriaal MD wordt het oxaalacetaat vervolgens in malaat omgezet dat kan migreren naar het cytosol. Daar wordt het malaat, door een cytosolisch MD, geconverteerd naar oxaalacetaat dat vervolgens door PCK1 wordt gedecarboxyleerd en gefosforyleerd naar fosfoënolpyruvaat. Na een aantal intermediaire reversibele reacties ontstaat fructose-1,6-bisfosfaat dat door FBP1 wordt gehydrolyseerd in fructose-6-fosfaat. Na een isomerisatie zal G6P het glucose-6-fosfaat in het ER hydrolyseren naar glucose waarna de glucose uit het ER migreert en via GLUT-2 naar de circulatie kan getransporteerd worden. Enkelvoudige pijlen stellen aflopende reacties voor terwijl dubbele pijlen reversibele reacties voorstellen. De dubbele gestreepte lijnen stellen intermediaire reversibele reacties voor (omgekeerde van de glycolyse). De gebruikte afkortingen: PC = pyruvaat carboxylase, MD = malaatdehydrogenase, PCK1 = fosfoënolpyruvaat carboxykinase 1, FBP1 = fructose-1,6-bisfosfatase, G6P = glucose-6-fosfatase, ER= endoplasmatisch reticulum en GLUT-2 = glucose transporter-2. Zelf gemaakte figuur op basis van gebruikte literatuur in § 1.3.2.1 Regulatie van de gluconeogenese.

8
1.3.3 Insulinereceptor
De transmembranaire insulinereceptor (IR) komt voor in verschillende celtypes en vervult
talrijke functies in onder meer het energiemetabolisme. Na binding van insuline op zijn
tetramere α2β2 tyrosine kinase receptor zorgt een conformationele verandering voor een
tyrosine autofosforylatie (positieve regulatie) van de receptor en van zijn intracellulaire
eiwitten. Er bestaan verschillende intracellulaire substraten, zoals de IR substraat (IRS)
eiwitten 1-4, die zullen binden met de IR. De gefosforlyleerde tyrosines in deze substraten
vormen op hun beurt bindingsplaatsen voor eiwitten en enzymen die Src-homolgie-2
domeinen bevatten zoals fosfoïnositide 3-kinase (PI(3)K), een adaptor eiwit dat downstream
via het serine proteïne kinase B (Akt/PKB) onder meer het glucose metabolisme reguleert. Na
tyrosine fosforylatie ondergaan de receptor en zijn substraten na enige tijd opnieuw serine
fosforylatie (negatieve regulatie). Insulineresistentie als gevolg van genetische en of
verworven factoren wordt gekenmerkt door defecten op vele niveaus, zoals bijvoorbeeld een
daling in receptor concentratie, in kinase activiteit, in IRS-1 en 2 fosforylatie of in PI(3)K
activiteit (19, 24, 27, 29).
1.4 Metabool syndroom
Het metabool syndroom, dat gelinkt wordt aan obesitas, wordt gekenmerkt door een
veralgemeende insulineresistentie, gedefinieerd als de verminderde gevoeligheid van weefsels
aan het vermogen van insuline om euglycemie te behouden (30). De gouden standaard in het
bepalen van deze resistentie is de hyperinsulinaemic euglycemic clamp methode. Hierbij
krijgt een nuchtere patiënt een insuline infuus toegediend zodat de endogene glucose
productie wordt onderdrukt. De ontstane hypoglycemie wordt vervolgens vermeden door een
variabel glucose infuus toe te dienen dat de basale glucose concentratie behoudt. Onder
steady-state is het glucose dat toegediend wordt gelijk aan het glucose dat gemetaboliseerd
wordt. Op basis van de hoeveelheid toegediende glucose om deze steady-state te bereiken,
kan de insulinegevoeligheid bepaald worden. Daarnaast worden ook de orale glucose
tolerantie test (OGTT) en het Homeostasis model assessment for insulin resistance (HOMA-
IR) gebruikt. Dit laatste is een formule gebaseerd op nuchtere glucose en insuline waarden.
Bepaling van de prevalentie van het metabool syndroom is moeilijk doordat er nog geen
eenduidige definitie bestaat, hoewel de prevalentie toch gelijkmatig toeneemt zoals de
prevalentie van obesitas en DM 2. Enkele corresponderende kenmerken, naast de verstoorde
glycemie, zijn centrale obesiteit, dyslipidemie en hypertensie (29-36). Insulineresistent
vetweefsel kent een hogere lipolyse, dat samen met een gedaalde oxidatieve capaciteit

9
resulteert in een ectopische accumulatie van VVZ in weefsels zoals de β-cellen van de
pancreas, lever, hart en skeletspierweefsel. Gezien deze weefsels slechts over een beperkte
opslagcapaciteit beschikken, leidt deze overmaat aan VVZ tot lipotoxiciteit met cel disfunctie
en celdood tot gevolg. Vooral VVW lijkt schadelijker en wordt frequenter geassocieerd met
metabole complicaties van obesitas en het metabool syndroom zoals NAFLD en NASH (7,
11, 13, 37-40). De distributie van het VVW en de opwaartse locatie van deze adipocyten in de
portale circulatie zou hypothetisch kunnen leiden tot een verhoogde flux van VVZ naar lever
(3, 7, 13, 30). Een overmaat aan VVZ overtreft de oxidatieve capaciteit van de mitochondriën
waardoor meer lipide intermediairen zoals diacylglycerol (DAG) en ceramides worden
gevormd die mogelijks het ontstaan insulineresistentie in verschillende weefsels mediëren
(11, 24, 30). DAG zou de glucose opname verstoren via interruptie van de GLUT-4 activiteit,
alsook de insulinesignalering verminderen. De VVZ oefenen aldus een invloed uit op de
hepatische klaring van insuline, de gluconeogenese en de glycogenese (11, 19, 41). Dit leidt
samen tot een belangrijke hyperglycemie die door de compensatoir verhoogde insulinesecretie
zal resulteren in een initiële hyperinsulinemie op korte termijn. Echter, chronische
blootstelling van de β-cellen aan VVZ leidt uiteindelijk tot lipotoxiciteit en apoptose
waardoor de insuline secretie finaal opnieuw zal dalen (30). De ectopische vetaccumulatie zou
ook verantwoordelijk zijn voor een defect in de mitochondriale oxidatieve fosforylatie en het
ER (zie § 1.5 Niet alcoholische steatohepatitis (NASH)) (11, 30, 41).
1.5 Niet alcoholische steatohepatitis (NASH)
1.5.1 Algemeen
NAFLD, de meest voorkomende leverziekte, is de hepatische component van het metabool
syndroom en is geassocieerd met insulineresistentie en hyperinsulinemie, onafhankelijk van
de graad van obesiteit (11, 32, 33, 42-44). NAFLD, waarvan de prevalentie wordt geschat op
34 %, is een multifactoriële aandoening waarbij het klinisch spectrum kan variëren van
eenvoudige steatose tot NASH, cirrose, leverfalen en zelfs tot hepatocellulair carcinoma (11,
39, 43, 45). Wanneer de hepatische vetinhoud meer dan 5 % bedraagt, spreekt men van
steatose (11, 43). In aanwezigheid van obesitas verhoogt de prevalentie van NAFLD tot
ongeveer 75-95 % (32, 46, 47). Hoewel de ontwikkeling van NAFLD naar NASH niet
volledig gekend is, zou het lipide, glucose en lipoproteïne metabolisme ontregeld zijn (11, 39,
42). De pathogenese van NAFLD en evolutie naar NASH wordt frequent beschreven door een
“twee hit hypothese”. De eerste hit, bestaande uit steatose, wordt gevolgd door een tweede hit
gekenmerkt door inflammatie, inflammatoire cytokines, oxidatieve stress, vorming van

10
reactive oxygen species (ROS) en/of lipide peroxidatie (42). De producten van lipide
peroxidatie, zoals DAG en ceramide, kunnen de hepatische stellaat cellen (HSC) activeren en
de inflammatie bevorderen via activatie van de transcriptiefactor NF-κB, die de expressie van
pro-inflammatoire cytokines induceert (39, 42). Ongeveer 10–26 % van de patiënten met
NAFLD evolueert naar NASH (32, 48). Naast de gouden standaard voor diagnose, zijnde een
leverbiopsie, worden er in studies ook niet-invasieve methoden zoals ultrasonografie, CT, MR
alsook glycomerkers gebruikt. De radiologische methoden zijn echter niet in staat NASH en
NAFLD te onderscheiden en zijn net als de merkers eerder suggestief (11, 43, 45, 46). Vaak
worden ook suggestieve leverenzymen zoals alanine aminotransferase (ALT) en aspartaat
aminotransferase (AST) bepaald (33, 43-46). De histopathologie van NASH wordt
gekenmerkt door steatose, hepatocellulaire ballooning (met of zonder Mallory’s hyaline),
acute of chronisch lobulaire inflammatie alsook fibrose (45, 46, 49). De steatose en ontstane
oxidatieve stress met inflammatie vormen de trigger voor fibrose, waarbij transforming
growth factor- β vrijgesteld wordt en waardoor de HSC geactiveerd worden. De aanmaak van
extracellulaire matrix neemt hierdoor toe en littekenweefsel wordt gevormd (50).
1.5.2 Lipide metabolisme, lipotoxiciteit en oxidatieve stress
Intrahepatisch vet bij obese NAFLD patiënten is afkomstig van VVZ (59 %), van de novo
lipogenese (26 %) en van het dieet (15 %). In vergelijking tot slanke mensen (5%) is de de
novo lipogenese tijdens vasten sterk verhoogd bij obese NAFLD patiënten (26%) (44). Dit is
voornamelijk het gevolg van de hyperinsulinemie, die zorgt voor inductie van de
transcriptiefactor SREBP-1c, en van de hyperglycemie, die zorgt voor een activatie van het
carbohydrate response element binding protein (ChREBP). Samen zullen SREBP-1c en
ChREBP de activatie van lipogene genen (zoals FAS) induceren, die de overmaat aan glucose
omzetten in VVZ. Bovendien leidt dit tot de synthese van malonyl-co-enzyme A die de
oxidatie van VVZ verhindert (11, 23, 39, 44). Deze dyslipidemie wordt o.a. gekenmerkt door
een verhoogd VLDL, een verhoogd low density lipoproteïne (LDL, metaboliet van VLDL dat
cholesterol naar de cellen vervoert) en een gedaald high density lipoproteïne (HDL,
verantwoordelijk voor transport van cholesterol van de cellen naar de lever) (10, 22, 30, 33,
45, 51). Gezien de overmaat aan VVZ de cellulaire capaciteit om te oxideren, op te slaan of te
incorporeren in VLDLs overtreft, resulteert dit in lipotoxiciteit met vorming van ROS die mee
resulteren in ER stress (11, 39, 48). In het ER, een belangrijke opslagplaats van Ca2+, worden
eiwitten gemodificeerd, geassembleerd en opgevouwen (52). Accumulatie van foutief
opgevouwen eiwitten en/of toxische lipide intermediairen kunnen leiden tot ER stress, met

11
activatie van beschermende pathway “unfolded protein response” (UPR) tot gevolg. Een te
hoge ER stress en langdurige activatie van UPR zal uiteindelijk resulteren in apoptose of
geprogrammeerde celdood (48, 52). ROS, een determinant van cellulaire stress, is vaak het
gevolg van mitochondriale disfunctie en verminderde anti-oxidante capaciteit. In de
mitochondriale matrix wordt ROS gevormd door de 1 elektron reductie van superoxide (O2-).
Deze elektronen kunnen cellulaire structuren zoals desoxyribonucleïnezuur (DNA) en lipide
membranen beschadigen met apoptose tot gevolg. De mitochondriale oxidatieve activiteit
vormt aldus een bron van oxidatieve stress (11, 39, 42, 48).
1.6 Adipokines
1.6.1 Algemeen
Het vetweefsel is ook een metabool actief endocrien orgaan dat verschillende fysiologisch
actieve peptiden vrijstelt, de zogenaamde adipokines, die via autocriene, paracriene en
endocriene signalen een invloed kunnen hebben op tal van organen zoals de lever en het hart.
Daarbij kunnen ze zowel perifere als centrale processen, zoals het glucose en lipide
metabolisme, reguleren. Deze adipokines omvatten onder meer cytokines, chemokines, acute
fase eiwitten, groeifactoren en complement achtige factoren (15, 37, 40). Zowel de adipocyten
als de stromovasculaire fractie zijn in belangrijke mate verantwoordelijk voor de secretie van
adipokines (37, 53). De graad van expressie en de soort adipokines die geproduceerd worden
blijkt verschillend te zijn tussen VVW en SVW en zouden eveneens beïnvloed worden door
de geslachtshormonen. Hierbij zouden de viscerale depots meer inflammatoire adipokines
secreteren (7, 13, 20, 37, 53). Obesitas en het metabool syndroom worden gekenmerkt door
een toestand van chronische inflammatie, waarbij de adipokine secretie van het hypertrofe en
insulineresistente vetweefsel ontregeld is. Er worden dan meer pro-inflammatoire en
insulineresistentie mediërende adipokines geproduceerd zoals interleukine-6 (Il-6), resistine
en tumor necrose factor-α (TNFα), die naar de lever kunnen circuleren via de portale
circulatie. Daarnaast is er een verlaagde productie van beschermende adipokines zoals
adiponectine en omentin (13, 15, 30, 37, 54). De expansie van het vetweefsel en adipocyt
hypertrofie (zie § 1.2.1 Anatomie) lijken een stimulus te zijn voor de infiltratie van
immuuncellen. Deze geïnfiltreerde macrofagen zijn zowel lokaal als systemisch mede
verantwoordelijk voor de productie van adipokines (17, 18, 37). Macrofagen worden
eveneens aangetrokken door o.a. het monocyt chemo attractief proteïne-1, dat bij obesitas
vooral omentaal meer wordt vrijgesteld (37, 55). Door expansie van het vetweefsel geraken
adipocyten ook steeds verder verwijderd van hun vasculatuur waardoor een O2 tekort

12
(hypoxie) ontstaat, dat gekenmerkt wordt door een verhoogde productie van hypoxia-
inducible factor 1. Deze laatste is een transcriptiefactor die bij hypoxie mede
verantwoordelijk zou zijn voor o.a. de toegenomen expressie en secretie van inflammatoire en
insulineresistentie mediërende adipokines (15, 37). Het verstoorde secretiepatroon van
adipokines zou aldus mede verantwoordelijk zijn in het mediëren van inflammatie en
insulineresistentie in zowel vetweefsel als lever (15, 30, 37, 40).
1.6.2 Adipokines die de Wnt signalering moduleren
1.6.2.1 Wnt – β-catenine signaal pathway
Wnt signalering is onder meer belangrijk bij celproliferatie, celpolariteit en differentiatie
gedurende de embryogenese en weefselvorming. Mutaties in deze pathway worden daarom
frequent gerelateerd aan ziekten zoals kanker. De Wnt signaal pathway kan onderverdeeld
worden in een canonical pathway die werkt via β-catenine en een niet-canonical pathway die
onafhankelijk is van β-catenine (18, 35, 56, 57). In afwezigheid van een Wnt ligand wordt
cytoplasmatisch β-catenine afgebroken door een complex bestaande uit Axin, adenomatous
polyposis coli (APC), caseïne kinase 1α (CK1α) en glycogeen synthase kinase-3 (GSK-3). Dit
complex zorgt namelijk voor de serine fosforylatie van β-catenine, wat de bindingsplaats
vormt van het E3 ubiquitine ligase subunit, β-Trcp, die instaat voor ubiquitinatie en
proteasomale degradatie van β-catenine. Door de continue verwijdering kan β-catenine niet
diffunderen naar de nucleus, waar de Wnt doelwitgenen onderdrukt worden door DNA
gebonden eiwitten van de T cel factor (TCF) of lymphoïd enhancer factor (LEF) familie (ook
Groucho genaamd) alsook door histon deacytelasen (56, 57). Echter, bij binding van een Wnt
ligand op zijn frizzled receptor (Fz) en co-receptor, low density lipoproteïne receptor
gerelateerd proteïne 6 (LRP6), zal de Wnt–β-catenine pathway geactiveerd worden. Na
vorming van het Wnt-Fz-LRP6 complex wordt Dishevelled (Dvl) gerekruteerd, wat via CK1
en GSK-3 leidt tot fosforylatie van LRP6 en activatie van de signaalcascade. Het Axin
complex wordt dan naar de receptoren gerekruteerd, waardoor de degradatie van β-catenine
wegvalt en dit nu kan diffunderen naar de nucleus. Binding met TCF/LEF zorgt vervolgens
voor activering van de genexpressie van Wnt doelwitgenen (35, 56, 57). De Wnt signalering
zou zo een rol spelen in de regulatie van gluconeogene doelwitgenen. FoxO1 zou gereguleerd
worden door de Wnt signaalbaan en samen met β-catenine insuline responsieve elementen
(IRE) herkennen, waarbij de expressie van G6P en PCK1 gestimuleerd wordt. TCF7L2, een
highmobility group box bevattende transcriptiefactor die eveneens beïnvloed wordt door de

13
Wnt signalering, zou belangrijk zijn bij het onderdrukken van de expressie van PCK1, G6P en
FBP1 (21, 35, 36, 58-60).
1.6.2.2 Wnt signalering, gluconeogenese, insulinesignalering en lipide metabolisme: het
verband
Bij vasten lijkt er een verhoogde expressie te zijn van Wnt ligand isovormen (Wnt-5b/ Wnt-
7a) en zou β-catenine een hepatische cofactor zijn voor de FoxO1 afhankelijke
gluconeogenese respons. Zo is de verhoogde hepatische expressie van β-catenine positief
gecorreleerd met de nucleaire lokalisatie van FoxO1, de expressie van G6P en PCK1 en de
glucose output (26, 58). β-catenine zou aldus, via interactie, de lokalisatie en activiteit van
FoxO1 moduleren. Daarbij zou FoxO in competitie treden met TCF voor binding met β-
catenine (58, 61). De co-activator Peroxisome proliferative activated receptor-γ coactivator
1α (PGC-1α) zou in belangrijke mate mede verantwoordelijk zijn voor de FoxO1-afhankelijk
transcriptionele activatie van PCK1 en G6P bij vasten (21, 26, 28). Insuline zorgt via een
gedaalde expressie van gluconeogene genen voor een gedaalde glucose productie. Als gevolg
van de insuline stimulatie zorgt het actieve Akt/PKB er immers voor dat FoxO1 in de kern
gefosforyleerd wordt, met nucleaire exclusie en dus gedaalde transcriptie tot gevolg (21, 28,
51, 58, 62). Hoewel de expressie van PGC-1α in vitro niet beïnvloed lijkt te worden door
insuline, zorgt de insuline wel voor een reductie van de PGC-1α gemedieerde expressie van
gluconeogene genen. Dit is vermoedelijk een rechtstreeks gevolg van de functionele interactie
tussen FoxO1 en PGC1-α, waarbij de FoxO1 activiteit wel gevoelig is voor insuline (28). Bij
vasten daarentegen, is de insuline concentratie laag waardoor de fosforylatie van FoxO1
vermindert. Daarenboven worden PGC-1α en FoxO1, die zich op dat moment in het
cytoplasma bevinden, gedeacetyleerd door silent information regulator waardoor ze naar de
kern kunnen migreren voor transcriptie (21, 28, 51, 58, 62). Er werd reeds aangetoond dat
NASH patiënten positieve correlaties vertoonden met FoxO1 expressielevels, FoxO1
nucleaire lokalisatie, FoxO1 proteïne levels, PCK1 en G6P expressielevels, evenals een
gedaalde activiteit van Akt/PKB (51). Bovendien zou deletie van β-catenine in de lever leiden
tot een verhoogde insulinesignalering, terwijl overexpressie van β-catenine de
cytoplasmatische lokalisatie van FoxO1 en de insulinesignalering zou reduceren (58).
Paradoxaal zou de Wnt–β-catenine activatie eveneens belangrijk zijn voor binding van
TCF7L2 op het TCEF/LEF bindend motief van de IR promotor en zo de transcriptie van IR
promoten (29, 59). Varianten in TCF7L2 werden reeds geassocieerd met een verhoogd risico
op de ontwikkeling van DM 2 (34, 63). Insuline zou de hepatische gluconeogenese eveneens

14
kunnen onderdrukken, via de canonical pathway, door op-regulatie van het TCF7L2 gen. Zo
werd bij muizenstudies alsook in vitro aangetoond dat insuline de hepatische expressie van
TCF7L2 eiwitten induceert. Dit correleerde steeds met verhoogde canoncical Wnt activiteit
waarbij de actieve nucleaire vorm van β-catenine gestimuleerd werd. Daarnaast zorgde de
knockdown van TCF7L2 voor een significante verhoging van de PCK1, G6P expressie en een
verhoogde glucose output. Insuline en voeding lijken dus zowel via Wnt en TCF7L2 activatie
alsook via FoxO1 downregulatie, de gluconeogene genexpressie en glucose productie te
onderdrukken. Tenslotte zouden verhoogde TCF7L2 levels in competitie kunnen treden met
FoxO1 voor binding met β-catenine waardoor de gluconeogenese eveneens vermindert (59).
De insuline en de Wnt pathway lijken ook verbonden te zijn via GSK-3β (58). Insuline zorgt
downstream voor de activatie van Akt/PKB dat op zijn beurt het GSK-3β zal fosforyleren en
inactiveren. Hoewel insuline evenals Wnt liganden (zie § 1.6.2.1 Wnt – β-catenine signaal
pathway) beiden instaat zijn om de GSK-3β activiteit te doen dalen, lijken ze dit volgens Ding
et al. toch op een verschillende wijze te doen. Zo zou insuline geen invloed hebben op de β-
catenine accumulatie en zou de Wnt geïnduceerde daling van GSK-3β onafhankelijk zijn van
de fosforylatie door Akt/PKB (64-66). Tot slot zou de Wnt signalering ook een verband
hebben met het lipide metabolisme. Dragers van LRP6R611C, een loss-of-function mutatie
gekenmerkt door een gedaalde activiteit van LRP6, vertonen verhoogde LDL en TG waarden
(22, 29). Door de ontwikkeling van muismodellen met deze LRP6R611C mutatie konden Go et
al. het lipide metabolisme in de lever bestuderen. Hierbij hadden LRP6mut/mut muizen een
verhoogde hepatische de novo lipogenese met een gestegen synthese van TG en cholesterol.
Daarenboven werd er een verhoogde secretie van VLDL, apoB en TG vastgesteld in
vergelijking met wild type muizen. Deze verhoogde de novo lipogenese resulteerde in een
vette lever, wat bevestigd werd door verhoogde expressie- en eiwitniveaus van onder meer
MTTP en hepatische enzymen betrokken in de de novo lipogenese (o.a. FAS), TG en
cholesterol synthese alsook van hun regulatoren (o.a. SREBP1-2). Normalisatie van het
plasma LDL en TG werd bekomen door Wnt-3a administratie zowel in vivo als in vitro (22).
1.6.2.3 Secreted frizzled-related proteïne 4 (SFRP4)
Bij de mens komen secreted frizzled-related proteïnen (SFRP) 1-5 voor, die gekenmerkt
worden door een cysteïne rijk domein gelijkaardig aan dat van de Fz receptor (67). SFRPs
kunnen de canonical en niet canonical pathway inhiberen door binding met Wnt liganden of
door een inactief complex te vormen met Fz (56, 57, 67, 68). SFRPs zouden ook stimulatoren
zijn door elkaars activiteit onderling te beïnvloeden of door de simultane binding Wnt en Fz,

15
waardoor de signaalactivatie bevorderd wordt (67). SFRP4, een 40 kDa eiwit, wordt zowel
door adipocyten als de stromale fractie vrijgesteld en lijkt de Wnt signalering in verschillende
weefsels te inactiveren of te stimuleren. De secretie vanuit het VVW correleert met BMI en
HOMA-IR (68-70). Gedurende de adipogenese zou de SFRP4 expressie gradueel toenemen,
waarbij de canonical Wnt signalering geïnhibeerd wordt (70). Echter, zowel muis als humane
studies toonden aan dat SFRP4 sterk correleert met HbA1c (geglycosyleerd hemoglobine
A1c, weerspiegelt de lange termijn bloed glucose), insulineresistentie alsook DM 2 en dat
SFRP4 de insuline secretie vermindert na activatie van de canonical Wnt pathway (69).
1.6.2.4 Dickkopf 1 (DKK1)
Dickkopf 1 (DKK1) is een antagonist van de Wnt-β-catenine pathway doordat het bindt met
LRP6 en daardoor de Wnt geïnduceerde vorming van het Fz-LRP6 complex verhindert (57,
71). LRP6+/- muizen, die tot 50 % minder LRP6 activiteit bevatten, zouden beschermd zijn
tegen dieet geïnduceerde obesiteit en zowel vetweefsel- als hepatische insulineresistentie.
Daarenboven was de gluconeogenese (expressie van G6P en PCK1) alsook de activiteit van
FoxO1 verlaagd. Ook de nucleaire lokalisatie van β-catenine was significant lager in LRP6+/-
muizen, wat dus suggereert dat een verminderde β-catenine activiteit in de lever
verantwoordelijk zou zijn voor de gedaalde gluconeogenese (35). Dragers van de LRP6R611C
mutatie daarentegen, welke eveneens een gedaalde LRP6 activiteit hadden, vertoonden een
verminderde glucosetolerantie, insulinerespons, IR expressie en een verminderde
insulinesignalering in het skeletspierweefsel en huid fibroblasten. De LRP6R611C mutatie zou
hierbij de TCF7L2 gemedieerde transcriptie van IR verhinderen en de activiteit van insuline-
achtige groeifactor (IGF) 1 verhogen. Verhoogde IGF1 activiteit zou vervolgens leiden tot
verhoogde activiteit en stabiliteit van het kinase mammalian target of rapamycin (mTOR) C1,
die via IRS-1 serine fosforylatie bijdraagt tot de insulineresistentie (22, 29). Daarnaast blijkt
LRP6R611C mutatie belangrijk te zijn voor het lipide metabolisme (zie § 1.6.2.2 Wnt
signalering, gluconeogenese, insulinesignalering en lipide metabolisme: het verband). DKK1
wordt ook verondersteld een rol te spelen in de adipogenese van humane mesenchymale
stamcellen, waarbij het via zijn antagonistische werking op de Wnt signalering de
differentiatie reguleert (70).
1.6.3 Andere adipokines verwant met obesitas, NAFLD en insulineresistentie
Tal van adipokines werden reeds in verband gebracht met obesitas, in het bijzonder met
NAFLD en insulineresistentie. Adiponectine, een 30 kDa anti-inflammatoir cytokine dat
voornamelijk geproduceerd wordt door adipocyten, zou antilipogene en antifibrose

16
eigenschappen hebben en een rol spelen in de adipogenese. Adiponectine werd reeds negatief
geassocieerd met obesitas, insulineresistentie, DM 2, dyslipidemie en leververvetting (5, 7,
37, 40, 47, 72-74). In de lever zou het via de AdipoR2 receptor de gluconeogenese
verminderen maar de VVZ oxidatie bevorderen (30, 37, 40). Obese patiënten met of zonder
NASH hebben, onafhankelijk van de insulineresistentie, lagere serum adiponectine levels dan
controles en patiënten met simpele steatose (47, 72-74).
Omentin (34 kDa) wordt voornamelijk door de stromovasculaire fractie van het VVW
vrijgesteld en beïnvloed de glucose opname en insuline gevoeligheid positief. De levels van
omentin correleren met NAFLD, C-reactief proteïne (CRP) en hepatocyt ballooning (5, 75).
De expressie van pro-inflammatoire cytokines zoals TNF-α (26 kDa), door macrofagen en in
mindere mate door vetcellen, correleren met BMI, lichaamsvet en hyperinsulinemie (5, 7, 16,
37, 40, 76). Bij NASH werden zowel hogere circulerende TNF-α levels aangetoond alsook
een verhoogde expressie van TNF-α in subcutaan vet- en leverweefsel (73, 77).
Daarnaast werd ook Il-6, (22-27 kDa) reeds geassocieerd met een verminderde
insulinesignalering, verhoogde hepatische gluconeogenese, hyperglycemie en een gedaalde
glycogeensynthese (5, 13, 37, 40, 78, 79). De Il-6 expressie zou verhoogd zijn bij NASH en
correleert met inflammatie, fibrose en insulineresistentie (78).
In muismodellen werd resistine (12 kDa), voornamelijk vrijgesteld door VVW, reeds in
verband gebracht met het hepatisch glucose en lipide metabolisme alsook hepatische
insulineresistentie. Het verband met NAFLD en NASH is echter nog controversieel (5, 40, 47,
72, 74, 80).
Chemerine, een chemokine, werd geassocieerd met insulineresistentie in onder meer humane
skeletspiercellen en lijkt belangrijk in de adipogenese. Chemerine levels correleren positief
met BMI, HOMA-IR, adipocyt volume, VVW accumulatie en TG. Daarnaast is er ook een
positieve correlatie met NAFLD, portale inflammatie, fibrose, AST en ALT. De expressie van
chemerine en zijn CMKLR1 receptor is daarenboven verhoogd in de levers van mensen met
NASH (37, 75, 81-85).
1.7 Hypothese en doelstelling van dit onderzoek
Gezien de Wnt signalering bijdraagt tot de controle van weefsel homeostase, is het van
cruciaal belang het onderliggende mechanisme te ontrafelen. Er werd reeds meermaals
aangetoond dat de Wnt signalering een belangrijke rol speelt in celdifferentiatie en

17
celproliferatie, maar recent is men ook meer aandacht gaan geven naar de rol ervan in het
energiemetabolisme. In deze thesis werd in vitro, voor het eerst met een HepaRG® humane
lever cellijn, de rol van de Wnt signalering op het glucose en lipide metabolisme bestudeerd.
Gezien DKK1 en SFRP4 de Wnt signalering in hepatocyten zouden kunnen beïnvloeden, door
middel van paracriene en endocriene effecten, werden deze adipokines geselecteerd. Uit nog
niet gepubliceerde data van de onderzoeksgroep van Prof. Dr. J. Ruige werd immers gezien
dat SFRP4 en DKK1 beide verhoogd vrijgesteld worden in het VVW van obese patiënten met
DM 2. Daarnaast werd de hypothetische stelling onderzocht die de invloed van adipokines,
die meer gesecreteerd worden in het portaal gedraineerde VVW van obese DM 2 patiënten, op
het glucosemetabolisme suggereert.

18
2 Materialen en Methoden
2.1 Modelsysteem: De humane lever cellijn HepaRG®
De lever is een belangrijk regulatie orgaan van het glucose en lipide metabolisme. Hoewel
primaire humane hepatocyten het optimale celmodel vormen om het lipide en glucose
metabolisme te onderzoeken, is er een beperkte beschikbaarheid van humaan leverweefsel en
bestaat er een grote variabiliteit tussen humane donors (23). De HepaRG® cellijn,
oorspronkelijk afkomstig van de lever tumor van een vrouwelijke patiënte met
hepatocarcinoom en hepatitis C, vormt een goed alternatief voor reproduceerbaar metabool
onderzoek. De cellijn differentieert in een morfologische structuur die gekenmerkt wordt door
granulaire hepatocyten (georganiseerd in trabeculae) alsook door canaliculi en epithelium
achtige structuren (23, 86). De HepaRG® cellen zijn gevoelig voor glucose en insuline en
expresseren genen van de glycogenese, glycolyse, gluconeogenese en lipogenese. (23).
2.2 Celcultuur
2.2.1 Dag 0: uitplaten van de HepaRG® cellen
Voor dit protocol werden 3 batches van een terminaal gedifferentieerde HepaRG® cellijn
(Biopredic, Rennes, Frankrijk) aangekocht in cryovials die bewaard werden in vloeibaar N2
(Thermo Scientific™ Thermolyne™ Locator 4 Cryo Biological Storage System, Fisher
Scientific, Erembodegem, België). Batch 1 (referentie: HPR 116163-TA12) bevat cryovials
met ± 12 x 106 HepaRG® cellen terwijl batch 2 (referentie: HPR 116171-TA08) en batch 3
(referentie: HPR 116169-TA08) cryiovals bevatten met ± 8 x 106 HepaRG® cellen. Deze
batches werden gelijkwaardig gebruikt doorheen de experimenten om biologische variabiliteit
te voorkomen. Het uitplaten en alle andere manipulaties met de cellen (van dag 0 t.e.m. dag 9)
gebeurden steeds steriel onder een laminaire flow klasse 2 (Herasafe™, Fisher Scientific,
Erembodegem, België). Vers Basal Hepatic Medium (MIL600, Biopredic, 100 ml, 4 °C) en 1
vial (± 15 ml, -20 °C) HepaRG® Thaw, Seed and General Purpose Supplement (ADD670,
Life Technologies™, Gent, België) zijn nodig voor aanmaken van het “zaaimedium”. Het
supplement (ADD670, Life Technologies™) werd, na ontdooien in een warm waterbad (+37
°C), gehomogeniseerd door zachtjes te schudden en vervolgens gepipetteerd en gemengd in
100 ml Basal hepatic medium (MIL600, Biopredic). Na het verwijderen van de cryovial uit
vloeibaar N2 werd het deksel van de cryovial onder de flow een kwartslag opengedraaid om
de interne druk te verwijderen. Na ontdooien van de cryovial met HepaRG® cellen (2
minuten, +37 °C) werd de celsuspensie (1 ml) per cryovial overgebracht in 9 ml (1:10) van

19
het voorverwarmd HepaRG® Thaw, Seed and General Purpose Supplement 670 in een 50 ml
tube. De gedifferentieerde HepaRG® celsuspensie werd vervolgens gecentrifugeerd (2
minuten, 357 g, kamer temperatuur; Allegra X-15R™, Beckman Coulter®, Suarlée, België).
Na aspiratie van het supernatans werd de celpellet geresuspendeerd in 1 ml HepaRG® Thaw,
Seed and General Purpose Medium 670. Voor bepalen van de celviabiliteit en voor de
celtelling werd 10 µl van de celsuspensie en 10 µl trypaan blauw (0.40 %, kamer temperatuur;
Bio-Rad, Nazareth, België) gepipetteerd en gemengd in een eppendorf tube (1.5 ml). Viabele
cellen zijn impermeabel voor trypaanblauw en zullen dus kleurloos blijven. Vervolgens werd
10 µl van het mengsel gepipetteerd in de opening van een TC10 counting slide (Bio-Rad).
Telling gebeurde vervolgens met de TC10™ Automated Cell Counter (Bio-Rad).
De cellen werden vervolgens uitgezaaid op een 24-well plaat (4x6) gecoat met collageen
(Biopredic, +4 °C). Aan de hand van de celtelling kon de HepaRG® 1 ml celsuspensie
verdund worden met HepaRG® Thaw, Seed and General Purpose Supplement 670 naar de
geschikte celconcentratie (0.96 x 106 cellen/ml) per well (0.5 ml). Indien 12 x 106 HepaRG®
cellen werden geteld (cryovial van batch 1), diende 11.5 ml HepaRG® Thaw, Seed and
General Purpose Medium 670 gepipetteerd te worden bij de 1 ml celsuspensie (50 ml tube).
Op en neer pipetteren is hierbij heel belangrijk om clustervorming van cellen te voorkomen.
Na het pipetteren van 0.5 ml van de verdunde celsuspensie in elke well, werd de plaat lichtjes
geschud om eveneens clustervorming te voorkomen. Na visuele controle op clusters met
microscoop (Nikon eclipse Ti, lens: ∞Ph1, objectief: 10x, Nikon Instruments Europe BV,
Amsterdam, Nederland) werd de plaat in de incubator geplaatst (+37 °C, 5 % CO2, verzadigde
vochtigheid; Hera Cell150™, Fisher Scientific). Zowel voor de studie met DKK1 als met
SFRP4 blootstelling werden telkens 4 maal HepaRG® cellen uitgeplaat (= biologische
replicaten).
2.2.2 Verversen van medium op dag 1, dag 2 en dag 3
De cellen werden steeds vóór het verversen van medium gecontroleerd op viabiliteit met
behulp van de microscoop (Nikon® eclipse Ti). Vervolgens werd “onderhoudsmedium”
gemaakt door 1 vial (± 15 ml) gehomogeniseerd HepaRG® Maintenance/Metabolism Medium
supplement (ADD620, Life Technologies, -20 °C) op te lossen in 100 ml Basal hepatic
medium. Dit werd voorverwarmd in het warm waterbad (+37 °C, ± 15 minuten). Na aspiratie
van het medium van 4 wells, werd er telkens 0.5 ml HepaRG® Maintenance/Metabolism
Medium 620 medium in elke well gepipetteerd. Vervolgens werd de plaat na visuele controle
terug in dezelfde incubator geplaatst.

20
2.2.3 24 uur blootstelling aan laag glucose (0.5 mM) met of zonder insuline (100 nM) op
dag 7
Er werd telkens tweemaal 45 ml medium gemaakt met lage glucose concentratie (0.5 mM),
relevant voor vasten, waarvan 1 tevens insuline (100 nM) bevatte. Daarnaast werd ook 1 ml
penicilline–streptomycine stock oplossing (-20 °C, referentie: 15140, Gibco® Life
Technologies™) gebruikt, geschikt voor 100 ml medium, om bacteriële contaminatie door
zowel grampositieve als gramnegatieve bacteriën te verhinderen. Er werd een stock oplossing
(1 ml) van 500 mM glucose aangemaakt in een 15 ml falcon door de 90.08 g D-(+)-glucose
(180.16 g/mol, kamertemperatuur, Sigma-Aldrich®, Diegem, België) af te wegen (Kern ALJ
160-4NM, Kern®, Balingen, Duitsland) en op te lossen in 1 ml steriele phosphate buffered
saline (PBS). Er werd vervolgens 100 ml medium met antibioticum en 0.5 mM glucose
gemaakt met en zonder insuline. Gezien de gebruikte insuline stock oplossing (-20 °C, 5
mg/ml stock, 5733.49 g/mol, referentie: 16634 Sigma-Aldrich®) niet steriel was, werd het
medium telkens in een vacuüm steriele filter unit (250ml, 0.1 micron PES filter unit, Thermo
Scientific™ Nalgene™) gemaakt. Hierbij werd 98.9 ml Dulbecco’s Modified Eagle Medium
(DMEM zonder glucose, Life Technologies™), 1 ml penicilline–streptomycine (referentie:
15140, Gibco® Life Technologies™) oplossing, 0.1 ml glucose oplossing (500 mM, Sigma-
Aldrich®) en 1x met alsook 1x zonder 0.01147 ml insuline (100 nM insuline uit 5 mg/ml
stock oplossing, Sigma-Aldrich®) gepipetteerd. Na vacuüm filtratie werd telkens 45 ml van
het medium overgepipetteerd in een 50 ml tube, welke vervolgens in het warm waterbad
overgebracht werd. De rijen A en B van de 24-well plaat werden ververst met voorverwarmd
medium (0.5 ml per well) dat insuline bevat, terwijl rijen C en D ververst werden met medium
zonder insuline (zie Figuur 2). Vervolgens werd de plaat, gedurende 24 uur, terug in dezelfde
incubator geplaatst.
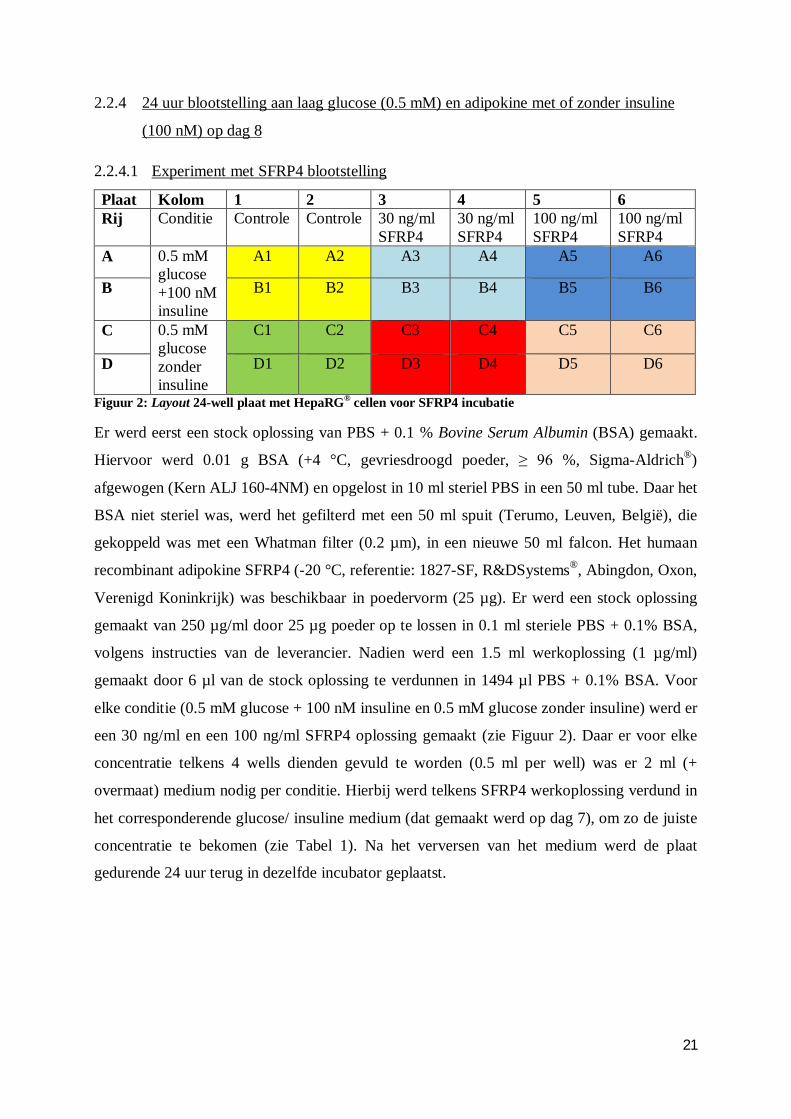
21
2.2.4 24 uur blootstelling aan laag glucose (0.5 mM) en adipokine met of zonder insuline
(100 nM) op dag 8
2.2.4.1 Experiment met SFRP4 blootstelling
Plaat Kolom 1 2 3 4 5 6 Rij Conditie Controle Controle 30 ng/ml
SFRP4 30 ng/ml SFRP4
100 ng/ml SFRP4
100 ng/ml SFRP4
A 0.5 mM glucose +100 nM insuline
A1 A2 A3 A4 A5 A6
B B1 B2 B3 B4 B5 B6
C 0.5 mM glucose zonder insuline
C1 C2 C3 C4 C5 C6
D D1 D2 D3 D4 D5 D6
Figuur 2: Layout 24-well plaat met HepaRG® cellen voor SFRP4 incubatie
Er werd eerst een stock oplossing van PBS + 0.1 % Bovine Serum Albumin (BSA) gemaakt.
Hiervoor werd 0.01 g BSA (+4 °C, gevriesdroogd poeder, ≥ 96 %, Sigma-Aldrich®)
afgewogen (Kern ALJ 160-4NM) en opgelost in 10 ml steriel PBS in een 50 ml tube. Daar het
BSA niet steriel was, werd het gefilterd met een 50 ml spuit (Terumo, Leuven, België), die
gekoppeld was met een Whatman filter (0.2 µm), in een nieuwe 50 ml falcon. Het humaan
recombinant adipokine SFRP4 (-20 °C, referentie: 1827-SF, R&DSystems®, Abingdon, Oxon,
Verenigd Koninkrijk) was beschikbaar in poedervorm (25 µg). Er werd een stock oplossing
gemaakt van 250 µg/ml door 25 µg poeder op te lossen in 0.1 ml steriele PBS + 0.1% BSA,
volgens instructies van de leverancier. Nadien werd een 1.5 ml werkoplossing (1 µg/ml)
gemaakt door 6 µl van de stock oplossing te verdunnen in 1494 µl PBS + 0.1% BSA. Voor
elke conditie (0.5 mM glucose + 100 nM insuline en 0.5 mM glucose zonder insuline) werd er
een 30 ng/ml en een 100 ng/ml SFRP4 oplossing gemaakt (zie Figuur 2). Daar er voor elke
concentratie telkens 4 wells dienden gevuld te worden (0.5 ml per well) was er 2 ml (+
overmaat) medium nodig per conditie. Hierbij werd telkens SFRP4 werkoplossing verdund in
het corresponderende glucose/ insuline medium (dat gemaakt werd op dag 7), om zo de juiste
concentratie te bekomen (zie Tabel 1). Na het verversen van het medium werd de plaat
gedurende 24 uur terug in dezelfde incubator geplaatst.
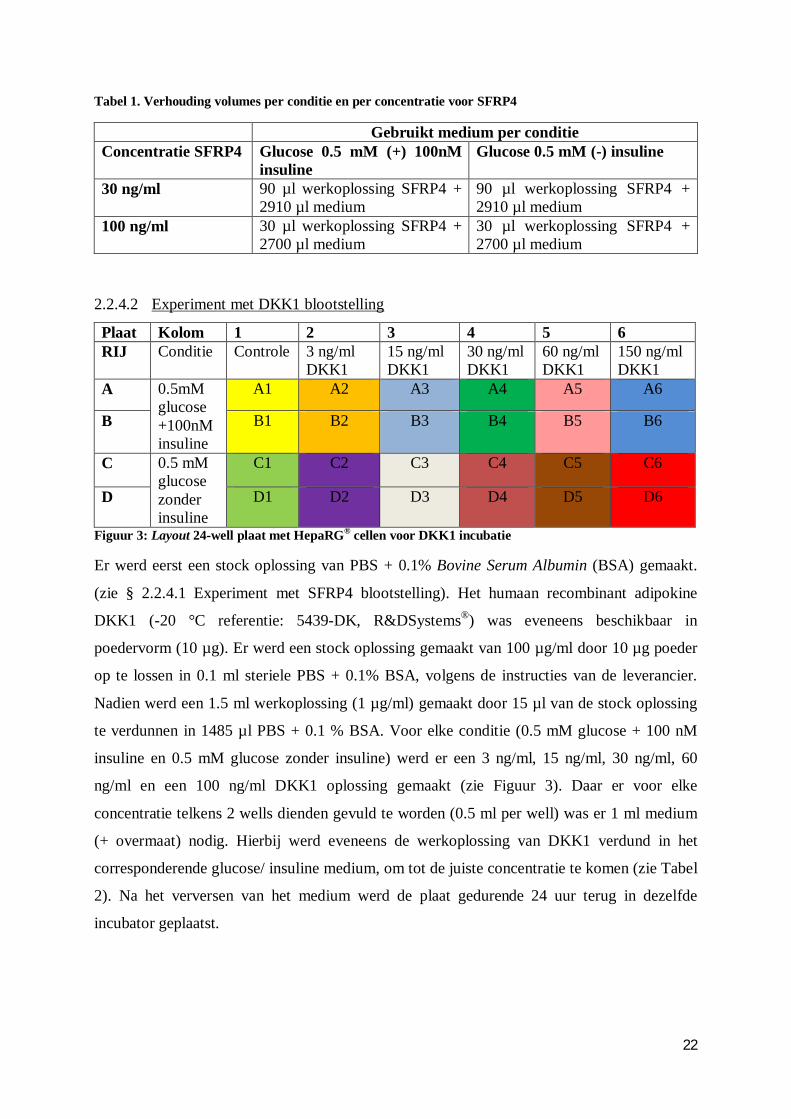
22
Tabel 1. Verhouding volumes per conditie en per concentratie voor SFRP4
Gebruikt medium per conditie Concentratie SFRP4 Glucose 0.5 mM (+) 100nM
insuline Glucose 0.5 mM (-) insuline
30 ng/ml 90 µl werkoplossing SFRP4 + 2910 µl medium
90 µl werkoplossing SFRP4 + 2910 µl medium
100 ng/ml 30 µl werkoplossing SFRP4 + 2700 µl medium
30 µl werkoplossing SFRP4 + 2700 µl medium
2.2.4.2 Experiment met DKK1 blootstelling
Plaat Kolom 1 2 3 4 5 6 RIJ Conditie Controle 3 ng/ml
DKK1 15 ng/ml DKK1
30 ng/ml DKK1
60 ng/ml DKK1
150 ng/ml DKK1
A 0.5mM glucose +100nM insuline
A1 A2 A3 A4 A5 A6
B B1 B2 B3 B4 B5 B6
C 0.5 mM glucose zonder insuline
C1 C2 C3 C4 C5 C6
D D1 D2 D3 D4 D5 D6
Figuur 3: Layout 24-well plaat met HepaRG® cellen voor DKK1 incubatie
Er werd eerst een stock oplossing van PBS + 0.1% Bovine Serum Albumin (BSA) gemaakt.
(zie § 2.2.4.1 Experiment met SFRP4 blootstelling). Het humaan recombinant adipokine
DKK1 (-20 °C referentie: 5439-DK, R&DSystems®) was eveneens beschikbaar in
poedervorm (10 µg). Er werd een stock oplossing gemaakt van 100 µg/ml door 10 µg poeder
op te lossen in 0.1 ml steriele PBS + 0.1% BSA, volgens de instructies van de leverancier.
Nadien werd een 1.5 ml werkoplossing (1 µg/ml) gemaakt door 15 µl van de stock oplossing
te verdunnen in 1485 µl PBS + 0.1 % BSA. Voor elke conditie (0.5 mM glucose + 100 nM
insuline en 0.5 mM glucose zonder insuline) werd er een 3 ng/ml, 15 ng/ml, 30 ng/ml, 60
ng/ml en een 100 ng/ml DKK1 oplossing gemaakt (zie Figuur 3). Daar er voor elke
concentratie telkens 2 wells dienden gevuld te worden (0.5 ml per well) was er 1 ml medium
(+ overmaat) nodig. Hierbij werd eveneens de werkoplossing van DKK1 verdund in het
corresponderende glucose/ insuline medium, om tot de juiste concentratie te komen (zie Tabel
2). Na het verversen van het medium werd de plaat gedurende 24 uur terug in dezelfde
incubator geplaatst.
23
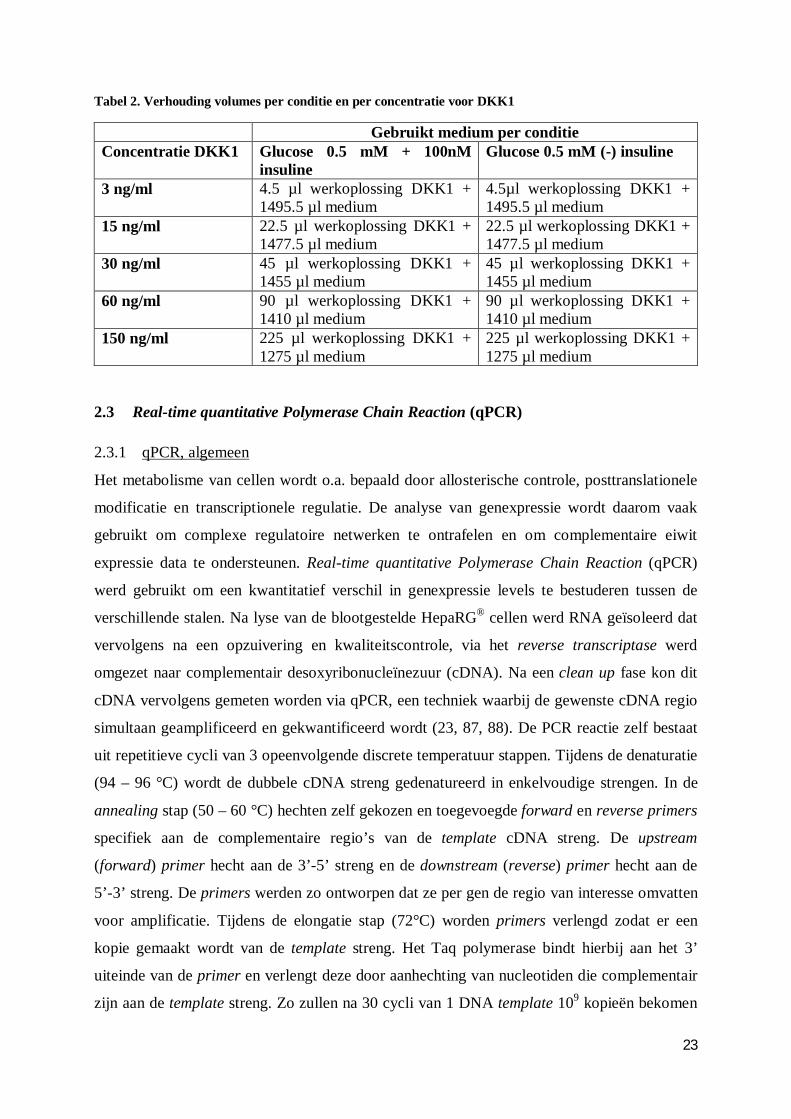
Tabel 2. Verhouding volumes per conditie en per concentratie voor DKK1
Gebruikt medium per conditie Concentratie DKK1 Glucose 0.5 mM + 100nM
insuline Glucose 0.5 mM (-) insuline
3 ng/ml 4.5 µl werkoplossing DKK1 + 1495.5 µl medium
4.5µl werkoplossing DKK1 + 1495.5 µl medium
15 ng/ml 22.5 µl werkoplossing DKK1 + 1477.5 µl medium
22.5 µl werkoplossing DKK1 + 1477.5 µl medium
30 ng/ml 45 µl werkoplossing DKK1 + 1455 µl medium
45 µl werkoplossing DKK1 + 1455 µl medium
60 ng/ml 90 µl werkoplossing DKK1 + 1410 µl medium
90 µl werkoplossing DKK1 + 1410 µl medium
150 ng/ml 225 µl werkoplossing DKK1 + 1275 µl medium
225 µl werkoplossing DKK1 + 1275 µl medium
2.3 Real-time quantitative Polymerase Chain Reaction (qPCR)
2.3.1 qPCR, algemeen
Het metabolisme van cellen wordt o.a. bepaald door allosterische controle, posttranslationele
modificatie en transcriptionele regulatie. De analyse van genexpressie wordt daarom vaak
gebruikt om complexe regulatoire netwerken te ontrafelen en om complementaire eiwit
expressie data te ondersteunen. Real-time quantitative Polymerase Chain Reaction (qPCR)
werd gebruikt om een kwantitatief verschil in genexpressie levels te bestuderen tussen de
verschillende stalen. Na lyse van de blootgestelde HepaRG® cellen werd RNA geïsoleerd dat
vervolgens na een opzuivering en kwaliteitscontrole, via het reverse transcriptase werd
omgezet naar complementair desoxyribonucleïnezuur (cDNA). Na een clean up fase kon dit
cDNA vervolgens gemeten worden via qPCR, een techniek waarbij de gewenste cDNA regio
simultaan geamplificeerd en gekwantificeerd wordt (23, 87, 88). De PCR reactie zelf bestaat
uit repetitieve cycli van 3 opeenvolgende discrete temperatuur stappen. Tijdens de denaturatie
(94 – 96 °C) wordt de dubbele cDNA streng gedenatureerd in enkelvoudige strengen. In de
annealing stap (50 – 60 °C) hechten zelf gekozen en toegevoegde forward en reverse primers
specifiek aan de complementaire regio’s van de template cDNA streng. De upstream
(forward) primer hecht aan de 3’-5’ streng en de downstream (reverse) primer hecht aan de
5’-3’ streng. De primers werden zo ontworpen dat ze per gen de regio van interesse omvatten
voor amplificatie. Tijdens de elongatie stap (72°C) worden primers verlengd zodat er een
kopie gemaakt wordt van de template streng. Het Taq polymerase bindt hierbij aan het 3’
uiteinde van de primer en verlengt deze door aanhechting van nucleotiden die complementair
zijn aan de template streng. Zo zullen na 30 cycli van 1 DNA template 109 kopieën bekomen

24
worden. Via TaqMan® Gene Expression Assays (Life Technologies™), waarbij gebruik
gemaakt wordt van een fluorogene probe, kon de hoeveelheid PCR product (amplicon) tijdens
het proces kwantitatief aangetoond worden. Bij het verlengen van de primers zal de probe
geknipt worden, waardoor deze zijn fluorescerende kleurstof kan uitzenden. De splitsing van
de probes in elke cyclus zal zorgen voor een verhoogde fluorescentie die proportioneel is met
de hoeveelheid amplicon die geproduceerd wordt. De verzamelde data zijn threshold cyclus
(Ct) waarde, de cylcus waarbij het signaal voor het eerst zichtbaar wordt na amplificatie en
dus afhankelijk is van de hoeveelheid cDNA aanwezig in het staal.
2.3.2 RNA isolatie en opzuivering
2.3.2.1 Isolatie van RNA uit HepaRG® cellijn op dag 9
Op dag 9, na een 24 uur durende blootstelling van de HepaRG® cellijn aan een bepaalde
concentratie van SFRP4/DKK1 bij een bepaalde conditie (zie § 2.2.4 24 uur blootstelling aan
glucose (0.5mM) en adipokine met of zonder insuline (100 nM) op dag 8), werd de cellijn
gelyseerd. Vooraf werden de cellen nogmaals gecontroleerd op viabiliteit met behulp van de
microscoop (Nikon eclipse Ti). Vanaf nu werd steeds RNase vrij gewerkt, waarbij alles naast
de ontsmetting met ethanol (75 %) ook met RNase Zap Wipes (Ambion® Life Technologies™)
gereinigd werd. Daarnaast werden RNase vrije filtertips en Eppendorf tubes gebruikt. De
procedure voor RNA isolatie was gebaseerd op instructies van de leverancier van de kit.
Hierbij werd de RNeasy Minikit (Qiagen®, KJ Venlo, Nederland) gebruikt. Voor de cellyse
diende er 150 µl lysebuffer per well toegevoegd te worden. Hiervoor werd 4.0 ml steriele
RNeasy lysis buffer (RLT, Qiagen®) gemengd met 40 µl β-Mercaptoethanol (≥99.0 %,
Sigma-Aldrich®) in een 15 ml tube. De RLT buffer bevat een hoge concentratie
isothiocyanaat waardoor het RNA beter bindt aan het silica membraan. Na aspiratie van
media uit elke wel (er werd gewerkt per 2 wells) werd direct 150 lysebuffer gepipetteerd in
elke well. Door lichtjes te schudden verspreidde de buffer zich egaal over oppervlak van de
wells. De lyse en homogenisatie van de cellysaten werd bevorderd door verschillende malen
op en neer te pipetteren in de well. Van rij A en B werd het cellysaat van de overeenkomstige
(zelfde kolomnummer) 2 wells (300 µl) in 1 RNase vrije tube gepipetteerd en vervolgens op
ijs gezet. Dit vormen de stalen 1 t.e.m 6. Van rij C en D werd het cellysaat van de
overeenkomstige (zelfde kolomnummer) 2 wells (300 µl) in 1 RNase vrije buisje (1.5 ml)
gepipetteerd en vervolgens direct op ijs gezet. Dit vormen de stalen 7 t.e.m.12. Op het einde
van de cellyse werden de stalen bewaard bij -80 °C tot verder gebruik voor RNA extractie.

25
2.3.2.2 Homogenisatie van het lysaat
Homogenisatie van de cellysaten werd uitgevoerd met behulp van de QIAshredder kit
(Qiagen®). De 12 tubes (stalen) met cellysaat (-80 °C) werden geleidelijk ontdooid door ze
eerst 2 minuten op ijs te zetten en vervolgens 15 minuten in een warm waterbad (+37 °C) te
plaatsen. Het lysaat van elke tube werd telkens op een aparte QIAshreder spin kolom geplaatst
in een 2 ml collectietube. De 12 kolommen werden vervolgens gedurende 2 minuten bij
10000 g gecentrifugeerd (Microfuge® 22R centrifuge, Beckman Coulter®). De QIAshreder
spin kolom werd verwijderd en 12 de collectietubes werden gesloten. Vervolgens werd in elke
tube 300 µl 70 % ethanol gepipetteerd en gemengd.
2.3.2.3 RNA extractie
Het gehomogeniseerde lysaat (± 700 µl) van elke collectietube werd direct overgebracht op
een RNeasy spin kolom in een nieuw 2 ml collectietube. Hierna werden de 12 stalen
gecentrifugeerd gedurende 15 seconden bij 10000 g (Microfuge® 22R centrifuge, Beckman
Coulter®). Het eluens werd verwijderd, waarna 350 µl RNeasy wasbuffer (RW1, Qiagen®) op
de RNeasy spin kolom gepipetteerd werd wat vervolgens opnieuw werd afgedraaid (15
seconden bij 10000 g).
2.3.2.4 DNase I behandeling
Behandeling met DNase I dient om het contaminerend genomisch DNA te verwijderen (88).
Na de centrifugatiestap (zie § 2.3.2.3 RNA extractie) werd direct 80 µl van de DNase mix
dicht tegen het membraan van de RNeasy spin kolom gepipetteerd. De DNase mix werd
aangemaakt voor 12 + 2 (overmaat) stalen, waarvoor 980 µl RNase vrije buffer (RDD,
Qiagen®) (+4 °C) vermengd werd met 140 µl DNase I (-20 °C). Na een incubatie periode van
15 minuten bij kamertemperatuur werd opnieuw 350 µl RW1 buffer op de RNeasy spin
kolom gepipetteerd. De 12 kolommen werden vervolgens afgedraaid gedurende 15 seconden
bij 10000 g (Microfuge® 22R centrifuge, Beckman Coulter®), waarna het eluens verwijderd
werd.
2.3.2.5 RNA recovery
Vervolgens werd de RNeasy spin kolom verwijderd uit elke collectietube en overgebracht
naar een nieuwe 2 ml collectietube. Na pipetteren van 500 µl RNeasy wasbuffer (RPE,
Qiagen®) op de kolom, werden de 12 kolommen afgedraaid gedurende 15 seconden bij 10000
g (Microfuge® 22R centrifuge, Beckman Coulter®) en werd het eluens verwijderd.
Vervolgens werd 500 µl RPE buffer op de kolommen gebracht die nu gedurende 2 minuten

26
bij 10000 g werden afgedraaid. Na verwijderen van het eluens werd nogmaals 15 seconden bij
10000 g gecentrifugeerd om de rest van de RPE buffer te verwijderen. Na het overbrengen
van de spin kolommen in een nieuwe 1.5 ml collectietube, werd telkens 32 µl RNase vrij
water (RNeasy minikit) dicht tegen het membraan gepipetteerd. Na 1 minuut incubatie
werden de kolommen gecentrifugeerd gedurende 1 minuut bij 10000 g waardoor het RNA
elueerde. De spin kolommen werden verwijderd en de 12 collectietubes met RNA werden
bewaard bij -80 °C.
2.3.2.6 RNA kwaliteitscontrole en concentratie bepaling.
Onzuiverheden in het RNA kunnen leiden tot inhibitie van reverse transcriptie alsook
variërende en incorrecte kwantificatie data tijdens PCR. De hoeveelheid RNA (ng / µl) werd
in elk staal bepaald met de NanoDrop® ND-100 Spectrophotometer (Thermo Scientific™). De
zuiverheid van het RNA, ten opzichte van eiwit contaminatie, werd geschat door bepaling van
de ratio van de absorbanties (A) bij een golflengte van 260 en 280 nm. Zuiver RNA, zonder
eiwit en fenol contaminatie, heeft een A260/A280 gelegen tussen 1.8 en 2.1 (88). Voor elke
meting werd telkens 2 µl RNA van het staal op het meettoestel gepipetteerd. De RNA stalen
konden vervolgens opnieuw bewaard worden bij -80 °C tot cDNA aanmaak.
2.3.3 cDNA aanmaak
De RNA stalen (-80 °C) alsook de 5x iScript™ reactie mix en het iScript™ reverse
transcriptase (-20 °C, Bio-Rad) werden ontdooid op ijs. Eerst werd de hoeveelheid RNA en
UltraPure™ DNase-RNase vrij gedestilleerd water (Gibco® Life Technologies™) berekend die
moet worden samengevoegd om 2 µg RNA te verkrijgen per staal:
Vstaal = (2 x 1000) / c (ng/µl) VDNase/RNase vrij water = 30 µl - Vstaal
De juiste hoeveelheid DNase-RNase vrij water (Gibco® Life Technologies™) werd telkens
gepipetteerd in RNase-DNase vrije PCR epjes (200 µl, Bio-Rad), geplaatst op een gekoelde
epjes houder (Eppendorf PCR- Cooler, -20 °C). Er werd vervolgens een mix gemaakt van
iScript™ reactie mix en iScript™ reverse transcriptase (Bio-Rad) in de verhouding 8:2 per
RNA staal. Nadien werd 10 µl van deze mix in elk PCR epje gepipetteerd. Ten slotte werd de
juiste hoeveelheid RNA (na vortexen en short spin met centrifugatie) toegevoegd. Na een
short spin (Beckman Coulter™ Microfuge® 22R centrifuge) van alle PCR epjes werden deze
in de iCycler Thermocycler (Bio-Rad) geplaatst voor cDNA synthese. Indien de clean up op
een ander tijdstip uitgevoerd werd, konden de PCR epjes met cDNA bewaard worden bij -20
°C.

27
2.3.3.1 cDNA clean up
Er werd gewerkt met de Gene Elute™ PCR clean up kit (Sigma-Aldrich®), volgens de
instructies van de leverancier. De PCR epjes met 40 µl cDNA (-20 °C) werden ontdooid op
ijs. Het werkoppervlak en het gebruikte materiaal werd eerst met ethanol 75 % ethanol en
vervolgens met DNA AWAY™ (Molecular BioProducts™, Fisher Scientific) gereinigd, om
contaminerend DNA te verwijderen. Voor elk PCR epje werd een GenElute Miniprep Binding
kolom in een 2 ml collectietube geplaatst. Er werd 500 µl Column Preparation Solution op
elke Miniprep kolom gepipetteerd, die vervolgens gecentrifugeerd werd gedurende 1 minuut
bij 12000 g (Microfuge® 22R centrifuge, Beckman Coulter®). Het eluens werd weggegoten.
De Column Preparation Solution maximaliseert de binding van het DNA aan het membraan,
waardoor een meer consistente opbrengst wordt bekomen. Daarnaast werd voor elk PCR epje
5 volumes Binding Solution (200 µl) gepipetteerd per volume van de PCR reactie (40 µl; na
short spin) in een RNase-DNase vrije microcentrifugatietube (1.5 ml, VWR®, Leuven,
België). Vervolgens werd het geheel (240 µl) direct overgebracht op de overeenkomstige
Binding kolom. De kolommen werden afgedraaid gedurende 1 minuut bij 12000 g en het
eluens werd verwijderd. Er werd 500 µl Wash Solution (+ ethanol) op de kolommen
gepipetteerd die nadien gedurende 1 minuut bij 12000 g werden afgedraaid. Het eluens werd
weggegoten en er werd nog eens gecentrifugeerd (2 minuten, 12000 g) opdat alle ethanol weg
zou zijn. De kolommen werden overgeplaatst in een nieuwe collectietube (2 ml). Na
pipetteren van 50 µl elution solution in het centrum van elke kolom en incubatie gedurende 1
minuut bij kamer temperatuur, werden de kolommen gecentrifugeerd (1 minuut, 12000 g). De
kolommen werden nu verwijderd en de collectietubes met geëlueerd cDNA werden op ijs
geplaatst. Het cDNA kon vervolgens bewaard worden bij -80 °C.
2.3.4 qPCR meting
2.3.4.1 Selectie van doelwitgenen
Het glucose metabolisme werd bestudeerd aan de hand van genexpressie van 3
snelheidsbepalende enzymen van het gluconeogenese proces, namelijk PCK1, FBP1 en G6P
(zie § 1.3.2.1 Regulatie van de gluconeogenese). Daarnaast werd de invloed van SFRP4 op de
insulinesignalering bestudeerd aan de hand van de expressieniveaus van IR, IRS 1 en IRS 2
(zie § 1.3.3 Insulinereceptor), alsook op het lipide metabolisme door analyse van de
expressieniveaus van MTTP, FAS en SREBF1 (zie § 1.3.1 Lipide metabolisme en § 1.6.2.2
Wnt signalering, gluconeogenese, insulinesignalering en lipide metabolisme: het verband).
Primerdesign en de keuze van de doelwitsequenties zijn belangrijk opdat de amplificatie
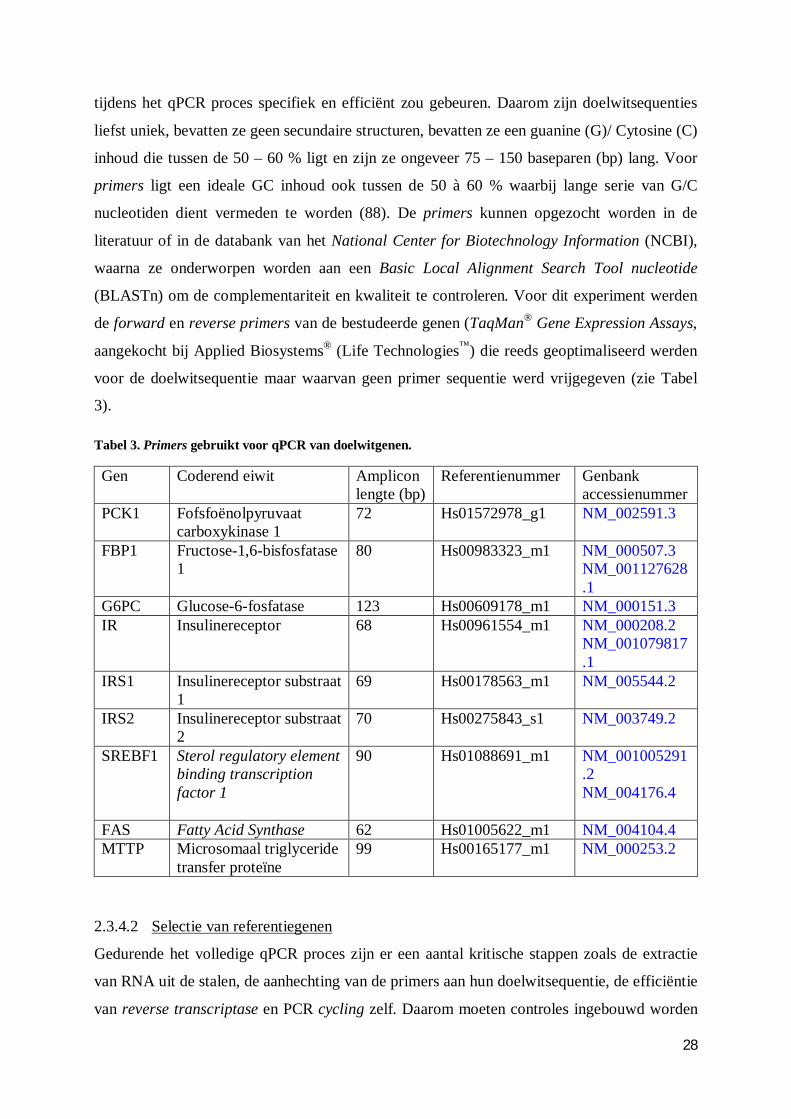
28
tijdens het qPCR proces specifiek en efficiënt zou gebeuren. Daarom zijn doelwitsequenties
liefst uniek, bevatten ze geen secundaire structuren, bevatten ze een guanine (G)/ Cytosine (C)
inhoud die tussen de 50 – 60 % ligt en zijn ze ongeveer 75 – 150 baseparen (bp) lang. Voor
primers ligt een ideale GC inhoud ook tussen de 50 à 60 % waarbij lange serie van G/C
nucleotiden dient vermeden te worden (88). De primers kunnen opgezocht worden in de
literatuur of in de databank van het National Center for Biotechnology Information (NCBI),
waarna ze onderworpen worden aan een Basic Local Alignment Search Tool nucleotide
(BLASTn) om de complementariteit en kwaliteit te controleren. Voor dit experiment werden
de forward en reverse primers van de bestudeerde genen (TaqMan® Gene Expression Assays,
aangekocht bij Applied Biosystems® (Life Technologies™) die reeds geoptimaliseerd werden
voor de doelwitsequentie maar waarvan geen primer sequentie werd vrijgegeven (zie Tabel
3).
Tabel 3. Primers gebruikt voor qPCR van doelwitgenen.
Gen Coderend eiwit Amplicon lengte (bp)
Referentienummer Genbank accessienummer
PCK1 Fofsfoënolpyruvaat carboxykinase 1
72 Hs01572978_g1 NM_002591.3
FBP1 Fructose-1,6-bisfosfatase 1
80 Hs00983323_m1 NM_000507.3 NM_001127628.1
G6PC Glucose-6-fosfatase 123 Hs00609178_m1 NM_000151.3 IR Insulinereceptor 68 Hs00961554_m1 NM_000208.2
NM_001079817.1
IRS1 Insulinereceptor substraat 1
69 Hs00178563_m1 NM_005544.2
IRS2 Insulinereceptor substraat 2
70 Hs00275843_s1 NM_003749.2
SREBF1 Sterol regulatory element binding transcription factor 1
90 Hs01088691_m1 NM_001005291.2 NM_004176.4
FAS Fatty Acid Synthase 62 Hs01005622_m1 NM_004104.4 MTTP Microsomaal triglyceride
transfer proteïne 99 Hs00165177_m1 NM_000253.2
2.3.4.2 Selectie van referentiegenen
Gedurende het volledige qPCR proces zijn er een aantal kritische stappen zoals de extractie
van RNA uit de stalen, de aanhechting van de primers aan hun doelwitsequentie, de efficiëntie
van reverse transcriptase en PCR cycling zelf. Daarom moeten controles ingebouwd worden
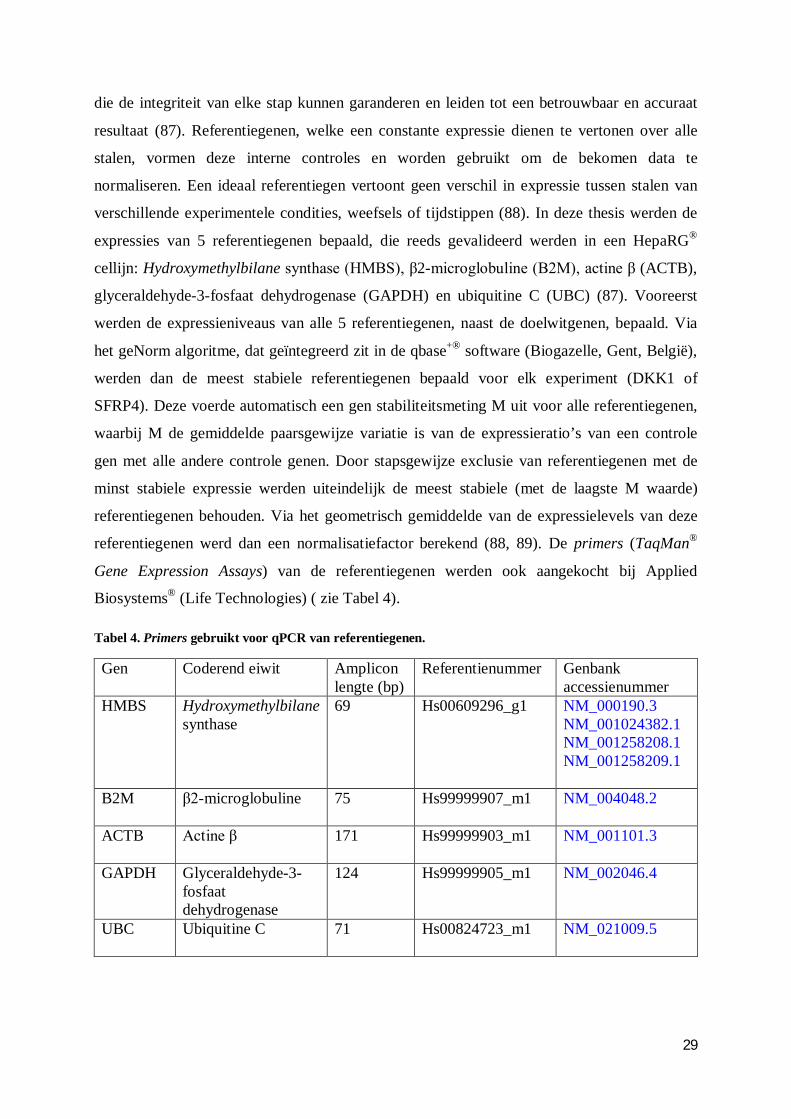
29
die de integriteit van elke stap kunnen garanderen en leiden tot een betrouwbaar en accuraat
resultaat (87). Referentiegenen, welke een constante expressie dienen te vertonen over alle
stalen, vormen deze interne controles en worden gebruikt om de bekomen data te
normaliseren. Een ideaal referentiegen vertoont geen verschil in expressie tussen stalen van
verschillende experimentele condities, weefsels of tijdstippen (88). In deze thesis werden de
expressies van 5 referentiegenen bepaald, die reeds gevalideerd werden in een HepaRG®
cellijn: Hydroxymethylbilane synthase (HMBS), β2-microglobuline (B2M), actine β (ACTB),
glyceraldehyde-3-fosfaat dehydrogenase (GAPDH) en ubiquitine C (UBC) (87). Vooreerst
werden de expressieniveaus van alle 5 referentiegenen, naast de doelwitgenen, bepaald. Via
het geNorm algoritme, dat geïntegreerd zit in de qbase+® software (Biogazelle, Gent, België),
werden dan de meest stabiele referentiegenen bepaald voor elk experiment (DKK1 of
SFRP4). Deze voerde automatisch een gen stabiliteitsmeting M uit voor alle referentiegenen,
waarbij M de gemiddelde paarsgewijze variatie is van de expressieratio’s van een controle
gen met alle andere controle genen. Door stapsgewijze exclusie van referentiegenen met de
minst stabiele expressie werden uiteindelijk de meest stabiele (met de laagste M waarde)
referentiegenen behouden. Via het geometrisch gemiddelde van de expressielevels van deze
referentiegenen werd dan een normalisatiefactor berekend (88, 89). De primers (TaqMan®
Gene Expression Assays) van de referentiegenen werden ook aangekocht bij Applied
Biosystems® (Life Technologies) ( zie Tabel 4).
Tabel 4. Primers gebruikt voor qPCR van referentiegenen.
Gen Coderend eiwit Amplicon lengte (bp)
Referentienummer Genbank accessienummer
HMBS Hydroxymethylbilane synthase
69 Hs00609296_g1 NM_000190.3 NM_001024382.1 NM_001258208.1 NM_001258209.1
B2M β2-microglobuline 75 Hs99999907_m1 NM_004048.2
ACTB Actine β 171 Hs99999903_m1 NM_001101.3
GAPDH Glyceraldehyde-3-fosfaat dehydrogenase
124 Hs99999905_m1 NM_002046.4
UBC Ubiquitine C 71 Hs00824723_m1 NM_021009.5
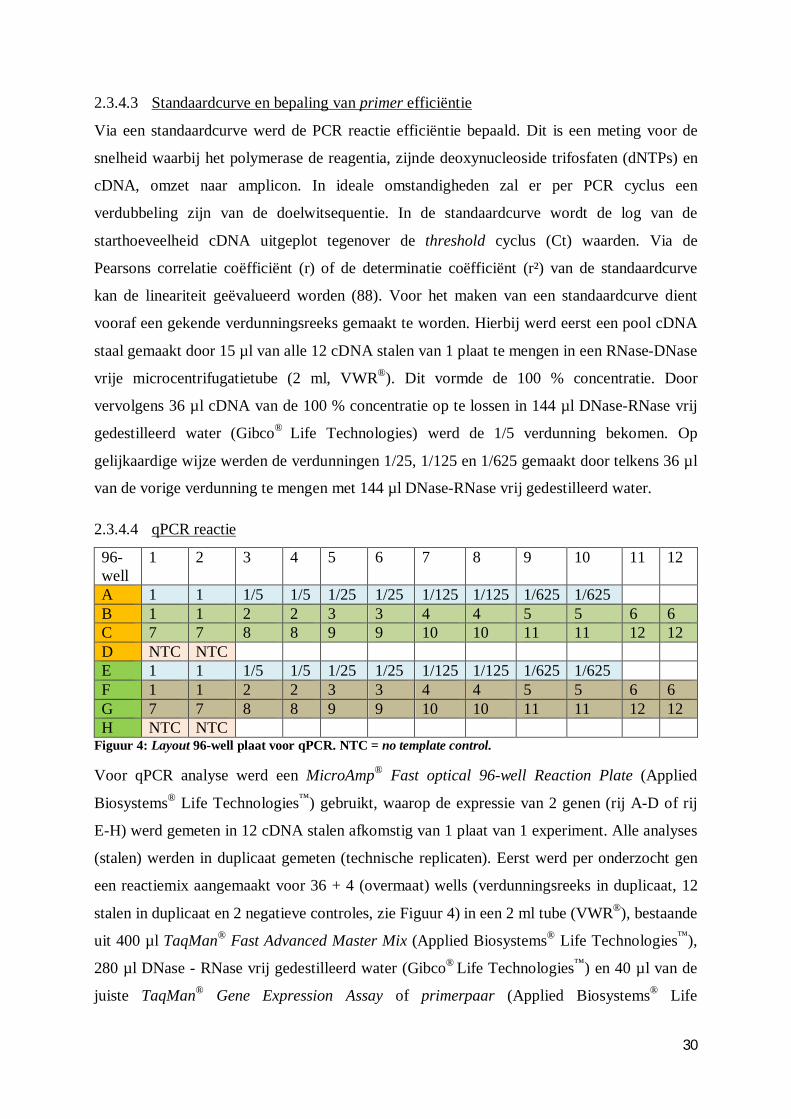
30
2.3.4.3 Standaardcurve en bepaling van primer efficiëntie
Via een standaardcurve werd de PCR reactie efficiëntie bepaald. Dit is een meting voor de
snelheid waarbij het polymerase de reagentia, zijnde deoxynucleoside trifosfaten (dNTPs) en
cDNA, omzet naar amplicon. In ideale omstandigheden zal er per PCR cyclus een
verdubbeling zijn van de doelwitsequentie. In de standaardcurve wordt de log van de
starthoeveelheid cDNA uitgeplot tegenover de threshold cyclus (Ct) waarden. Via de
Pearsons correlatie coëfficiënt (r) of de determinatie coëfficiënt (r²) van de standaardcurve
kan de lineariteit geëvalueerd worden (88). Voor het maken van een standaardcurve dient
vooraf een gekende verdunningsreeks gemaakt te worden. Hierbij werd eerst een pool cDNA
staal gemaakt door 15 µl van alle 12 cDNA stalen van 1 plaat te mengen in een RNase-DNase
vrije microcentrifugatietube (2 ml, VWR®). Dit vormde de 100 % concentratie. Door
vervolgens 36 µl cDNA van de 100 % concentratie op te lossen in 144 µl DNase-RNase vrij
gedestilleerd water (Gibco® Life Technologies) werd de 1/5 verdunning bekomen. Op
gelijkaardige wijze werden de verdunningen 1/25, 1/125 en 1/625 gemaakt door telkens 36 µl
van de vorige verdunning te mengen met 144 µl DNase-RNase vrij gedestilleerd water.
2.3.4.4 qPCR reactie
96-well
1 2 3 4 5 6 7 8 9 10 11 12
A 1 1 1/5 1/5 1/25 1/25 1/125 1/125 1/625 1/625 B 1 1 2 2 3 3 4 4 5 5 6 6 C 7 7 8 8 9 9 10 10 11 11 12 12 D NTC NTC E 1 1 1/5 1/5 1/25 1/25 1/125 1/125 1/625 1/625 F 1 1 2 2 3 3 4 4 5 5 6 6 G 7 7 8 8 9 9 10 10 11 11 12 12 H NTC NTC
Figuur 4: Layout 96-well plaat voor qPCR. NTC = no template control.
Voor qPCR analyse werd een MicroAmp® Fast optical 96-well Reaction Plate (Applied
Biosystems® Life Technologies™) gebruikt, waarop de expressie van 2 genen (rij A-D of rij
E-H) werd gemeten in 12 cDNA stalen afkomstig van 1 plaat van 1 experiment. Alle analyses
(stalen) werden in duplicaat gemeten (technische replicaten). Eerst werd per onderzocht gen
een reactiemix aangemaakt voor 36 + 4 (overmaat) wells (verdunningsreeks in duplicaat, 12
stalen in duplicaat en 2 negatieve controles, zie Figuur 4) in een 2 ml tube (VWR®), bestaande
uit 400 µl TaqMan® Fast Advanced Master Mix (Applied Biosystems® Life Technologies™),
280 µl DNase - RNase vrij gedestilleerd water (Gibco® Life Technologies™) en 40 µl van de
juiste TaqMan® Gene Expression Assay of primerpaar (Applied Biosystems® Life

31
Technologies™). Vervolgens werd 18 µl van de reactiemix in elke well gepipetteerd. De stalen
van de verdunningsreeks alsook de 12 cDNA stalen werden gevortexed en vervolgens
afgedraaid (short spin, 12000 g, Microfuge® 22R centrifuge Beckman Coulter™). Van elke
verdunning alsook elk cDNA staal werd 2 µl gepipetteerd in de overeenkomstige well (zie
Figuur 4). In no template control (NTC) werd echter 2 µl DNase-RNase vrij water (Gibco®
Life Technologies™) gepipetteerd in plaats van cDNA. Deze negatieve controles dienden om
buffers en oplossingen te controleren op DNA contaminatie en primer – dimeren. Na staal
toevoeging werd de 96-well plaat afgesloten met een MicroAmp® Optical Adhesive Film
(Applied Biosystems® Life Technologies™) en afgedraaid gedurende 1 minuut bij 1000 RPM
(rounds per minute) (Allegra X-15R™, Beckman Coulter®). De genexpressie werd bepaald op
een StepOnePlus™ Real-time PCR system (Applied Biosystems® Life Technologies™). De
cycling condities omvatten een activatie van 20 seconden bij 95 °C en 40 cycli bij 95 °C
gedurende 1 seconde en bij 60 °C gedurende 60 seconden.
2.4 Statistische analyse
Beide experimenten (SFRP4/DKK1) werden 4 maal onafhankelijk herhaald (biologische
replicaten) en in duplicaat (technische replicaten) geanalyseerd. De verzamelde data zijn Ct
waarden, de cyclus waarbij het signaal voor het eerst zichtbaar werd na amplificatie en dus
afhankelijk is van de hoeveelheid cDNA aanwezig in staal. De genormaliseerde
expressieniveaus werden berekend met de qbase+® software (Biogazelle). De kwantitatieve
resultaten voor de 4 herhalingen in duplicaat werden voor elk gen weergegeven als de
gemiddelde relatieve expressie ± de Standard Error of the Mean (SEM) ten opzichte van de
controle groep (%). De statistische analyse is gebeurd via een One-Way ANOVA met
correctie voor multiple comparisons (Holm-Sidak) en een ongepaarde t-test. Het aantal
onafhankelijke experimenten wordt weergegeven door N. De analyses alsook de figuren
werden gemaakt met Graphpad Prism® (La Jolla, Verenigde Staten) waarbij waarden met p <
0.05 als significant werden beschouwd (*, p <0.05; ** p<0.01; *** p<0.001).

32
3 Resultaten
3.1 Resultaten experiment SFRP4
3.1.1 Referentiegenen
Met behulp van geNorm uit de qbase+® software (Biogazelle) konden, voor het experiment
met SFRP4 stimulatie, de referentiegenen bepaald worden met de laagste gezamenlijke M-
waarde (meest stabiel). ACTB, GADPH en HMBS vertonen, ten opzichte van de controles,
geen verschil in expressieniveau voor de verschillende condities. Deze genen zijn aldus
onafhankelijk van insuline en SFRP4. Met deze 3 referentiegenen werden vervolgens de
genormaliseerde relatieve kwantificatie (NRQ) waarden van de expressie niveaus van de
doelwitgenen bepaald met en zonder SFRP4 stimulatie.
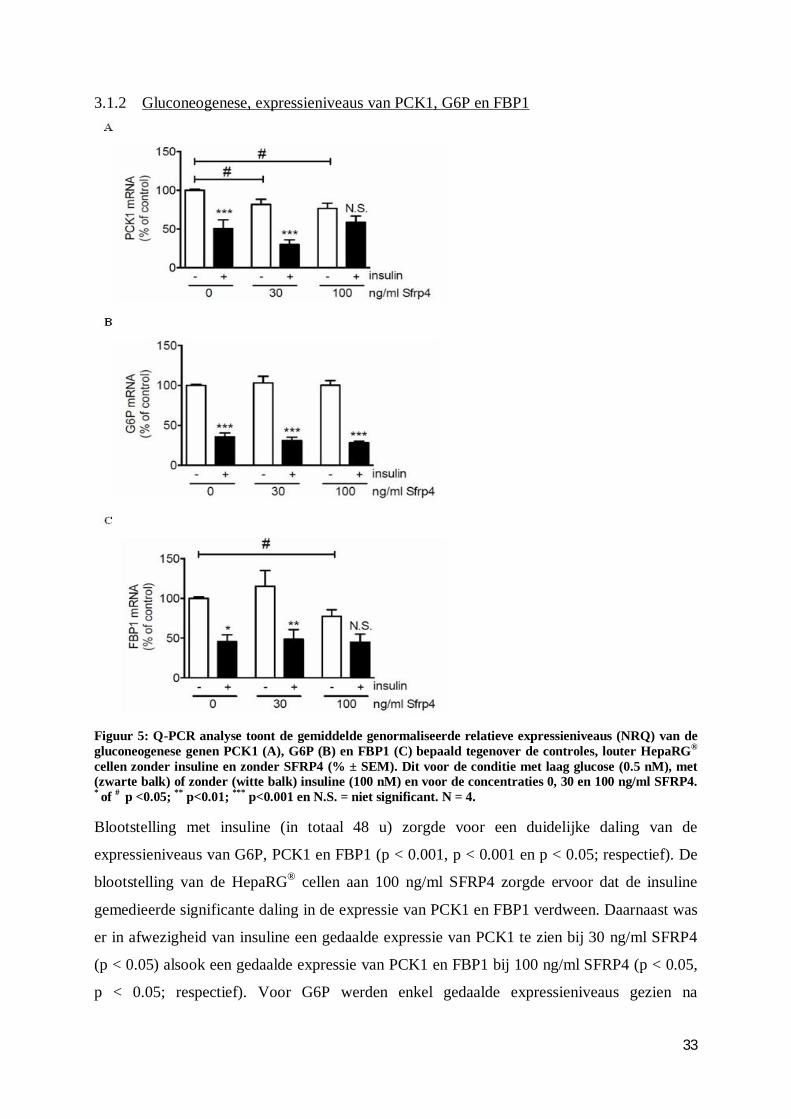
33
3.1.2 Gluconeogenese, expressieniveaus van PCK1, G6P en FBP1
Figuur 5: Q-PCR analyse toont de gemiddelde genormaliseerde relatieve expressieniveaus (NRQ) van de gluconeogenese genen PCK1 (A), G6P (B) en FBP1 (C) bepaald tegenover de controles, louter HepaRG® cellen zonder insuline en zonder SFRP4 (% ± SEM). Dit voor de conditie met laag glucose (0.5 nM), met (zwarte balk) of zonder (witte balk) insuline (100 nM) en voor de concentraties 0, 30 en 100 ng/ml SFRP4. * of # p <0.05; ** p<0.01; *** p<0.001 en N.S. = niet significant. N = 4.
Blootstelling met insuline (in totaal 48 u) zorgde voor een duidelijke daling van de
expressieniveaus van G6P, PCK1 en FBP1 (p < 0.001, p < 0.001 en p < 0.05; respectief). De
blootstelling van de HepaRG® cellen aan 100 ng/ml SFRP4 zorgde ervoor dat de insuline
gemedieerde significante daling in de expressie van PCK1 en FBP1 verdween. Daarnaast was
er in afwezigheid van insuline een gedaalde expressie van PCK1 te zien bij 30 ng/ml SFRP4
(p < 0.05) alsook een gedaalde expressie van PCK1 en FBP1 bij 100 ng/ml SFRP4 (p < 0.05,
p < 0.05; respectief). Voor G6P werden enkel gedaalde expressieniveaus gezien na
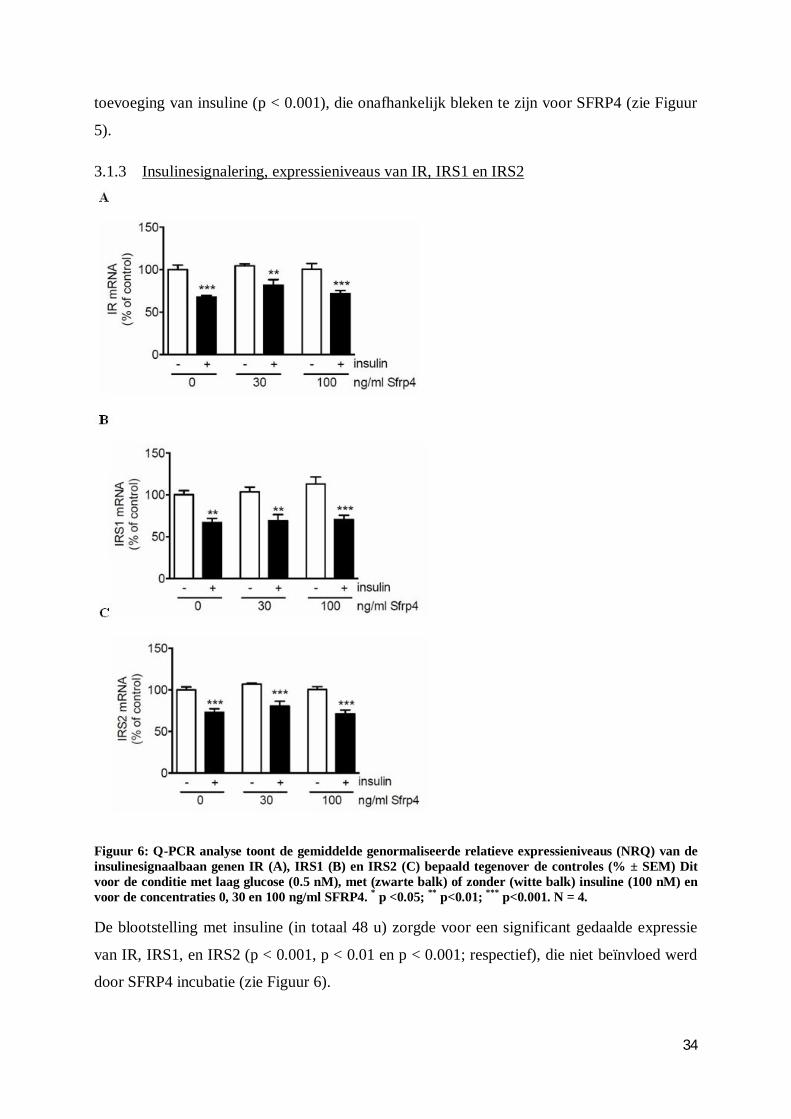
34
toevoeging van insuline (p < 0.001), die onafhankelijk bleken te zijn voor SFRP4 (zie Figuur
5).
3.1.3 Insulinesignalering, expressieniveaus van IR, IRS1 en IRS2
Figuur 6: Q-PCR analyse toont de gemiddelde genormaliseerde relatieve expressieniveaus (NRQ) van de insulinesignaalbaan genen IR (A), IRS1 (B) en IRS2 (C) bepaald tegenover de controles (% ± SEM) Dit voor de conditie met laag glucose (0.5 nM), met (zwarte balk) of zonder (witte balk) insuline (100 nM) en voor de concentraties 0, 30 en 100 ng/ml SFRP4. * p <0.05; ** p<0.01; *** p<0.001. N = 4.
De blootstelling met insuline (in totaal 48 u) zorgde voor een significant gedaalde expressie
van IR, IRS1, en IRS2 (p < 0.001, p < 0.01 en p < 0.001; respectief), die niet beïnvloed werd
door SFRP4 incubatie (zie Figuur 6).
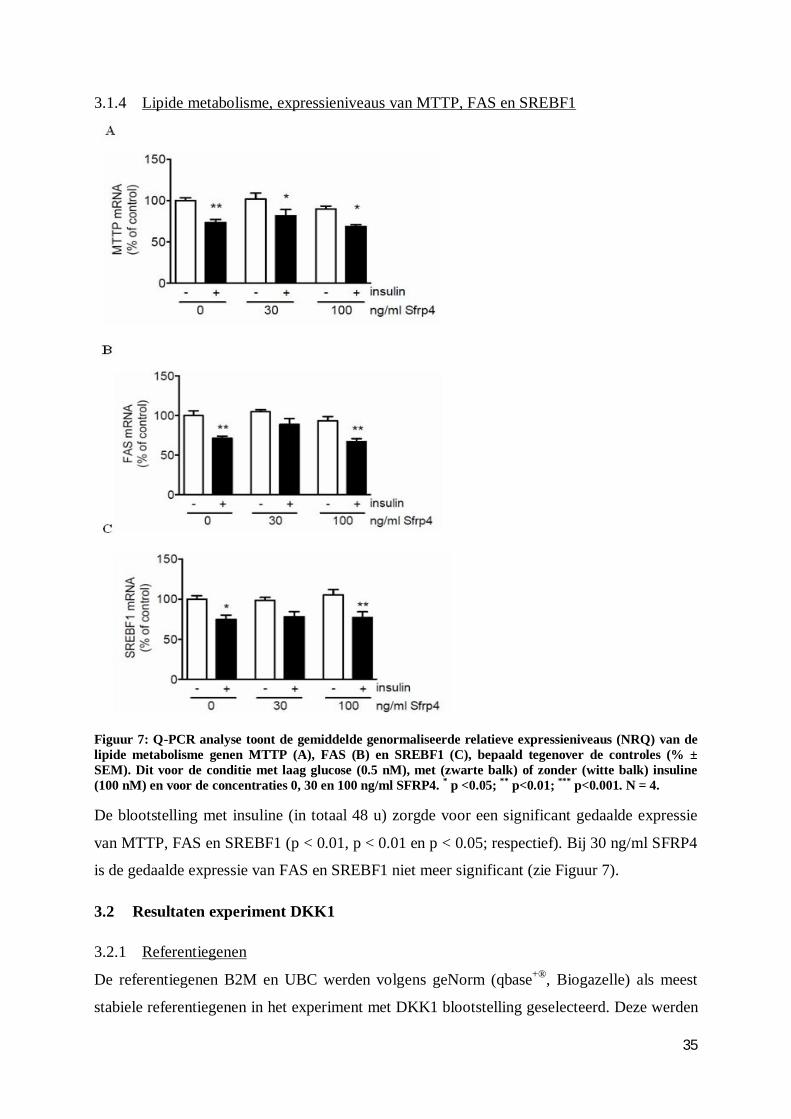
35
3.1.4 Lipide metabolisme, expressieniveaus van MTTP, FAS en SREBF1
Figuur 7: Q-PCR analyse toont de gemiddelde genormaliseerde relatieve expressieniveaus (NRQ) van de lipide metabolisme genen MTTP (A), FAS (B) en SREBF1 (C), bepaald tegenover de controles (% ± SEM). Dit voor de conditie met laag glucose (0.5 nM), met (zwarte balk) of zonder (witte balk) insuline (100 nM) en voor de concentraties 0, 30 en 100 ng/ml SFRP4. * p <0.05; ** p<0.01; *** p<0.001. N = 4.
De blootstelling met insuline (in totaal 48 u) zorgde voor een significant gedaalde expressie
van MTTP, FAS en SREBF1 (p < 0.01, p < 0.01 en p < 0.05; respectief). Bij 30 ng/ml SFRP4
is de gedaalde expressie van FAS en SREBF1 niet meer significant (zie Figuur 7).
3.2 Resultaten experiment DKK1
3.2.1 Referentiegenen
De referentiegenen B2M en UBC werden volgens geNorm (qbase+®, Biogazelle) als meest
stabiele referentiegenen in het experiment met DKK1 blootstelling geselecteerd. Deze werden
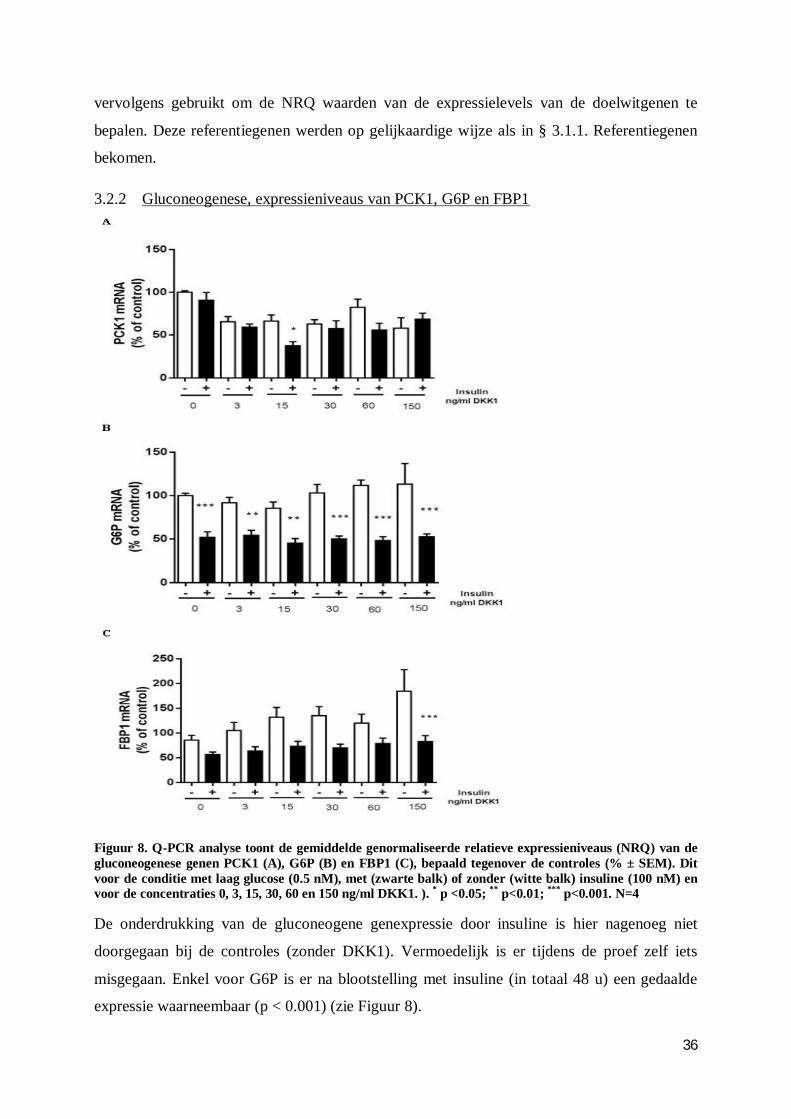
36
vervolgens gebruikt om de NRQ waarden van de expressielevels van de doelwitgenen te
bepalen. Deze referentiegenen werden op gelijkaardige wijze als in § 3.1.1. Referentiegenen
bekomen.
3.2.2 Gluconeogenese, expressieniveaus van PCK1, G6P en FBP1
Figuur 8. Q-PCR analyse toont de gemiddelde genormaliseerde relatieve expressieniveaus (NRQ) van de gluconeogenese genen PCK1 (A), G6P (B) en FBP1 (C), bepaald tegenover de controles (% ± SEM). Dit voor de conditie met laag glucose (0.5 nM), met (zwarte balk) of zonder (witte balk) insuline (100 nM) en voor de concentraties 0, 3, 15, 30, 60 en 150 ng/ml DKK1. ). * p <0.05; ** p<0.01; *** p<0.001. N=4
De onderdrukking van de gluconeogene genexpressie door insuline is hier nagenoeg niet
doorgegaan bij de controles (zonder DKK1). Vermoedelijk is er tijdens de proef zelf iets
misgegaan. Enkel voor G6P is er na blootstelling met insuline (in totaal 48 u) een gedaalde
expressie waarneembaar (p < 0.001) (zie Figuur 8).

37
4 Discussie
4.1 Belangrijkste resultaten
In deze thesis werd voor het eerst in vitro met een HepaRG® humane lever cellijn, de rol van
Wnt regulator SFRP4, die verhoogd wordt vrijgesteld in het VVW van morbide obese
patiënten met DM 2, op het glucosemetabolisme bestudeerd. Via qPCR werd gezien dat
SFRP4 (100 ng/ml) het inhiberende effect van insuline op de expressie van PCK1 en FBP1,
beide sleutelenzymen van het gluconeogenese proces, grotendeels lijkt op te heffen. Daarnaast
werd in afwezigheid van insuline een gedaalde expressie van PCK1 en FBP1 waargenomen
na blootstelling met SFRP4. Deze resultaten bevestigen de rol van de Wnt signalering op het
glucosemetabolisme en suggereren een vermoedelijke interactie tussen de Wnt signalering en
de insulinesignalering. Daarenboven dragen deze resultaten bij tot de hypothetische stelling
dat adipokines, die verhoogd worden vrijgesteld uit het portaal gedraineerde VVW, een rol
kunnen spelen in de (dis)regulatie van het glucosemetabolisme.
4.2 Belang van onderzoek en hypothetische verklaringen van de resultaten.
De canonical Wnt signaalbaan, gekenmerkt door de transcriptiefactor β-catenine (zie §1.6.2.1
Wnt – β-catenine signaal pathway), lijkt belangrijke fysiologische functies te beïnvloeden in
tal van organen en weefsels die een rol kunnen spelen bij de pathofysiologie van obesitas en
DM 2, beiden kenmerken van het metabool syndroom. Hypertrofe obesitas, gekarakteriseerd
door een verminderd expansie vermogen van het vetweefsel, lijkt het gevolg te zijn van een
gedaalde onderdrukking van de canonical Wnt pathway en een verminderd vermogen om
DKK1 te induceren, welke normaal een voorname rol speelt bij het adipogenese proces. De
hierbij sterk vergrote vetcellen promoten inflammatie en disregulatie van het vetweefsel,
alsook ectopische lipide accumulatie (18, 70). Bovendien werden reeds verschillende DM 2
geassocieerde genetische varianten gelokaliseerd in genen van de Wnt signaalbaan. Kanazawa
et al. toonden reeds de rol van Wnt signalering in DM 2 aan via een case-control studie in een
vrouwelijk Japanse cohort. Daaruit bleek een single-nucleotide-poylymorphism (SNP) locus
in het WNT5B gen (detecteerbaar in vetweefsel, pancreas en lever) sterk gecorreleerd te zijn
met DM 2 (90). De Wnt regulator SFRP4 zou eveneens verhoogd tot expressie komen in
humane pancreas eilandjes van DM 2 patiënten. Bovendien zouden systemisch verhoogde
SFRP4 concentraties de glucose tolerantie verminderen alsook de exocytose van insuline
onderdrukken (69). Door genotypering van microsatelliet merkers in een grote populatie
bestaande uit DM 2 patiënten alsook controles werd een gemeenschappelijk microsatelliet
gevonden in het intron van het TCF7L2 gen. Dragers van deze variant van het TCF7L2 gen

38
vertoonden een groter risico op DM 2 (63). Dit werd bevestigd door Florez et al. via twee
SNPs in het TCF7L2 gen, waarbij het risicovolle genotype meer geassocieerd werd met
progressie naar DM 2 en gelinkt werd met een verminderde basale insuline secretie (34). Da
Silva Xavier et al. toonden tenslotte aan dat silencing van TCF7L2 in muis pancreas eilandjes
leidde tot een daling in zowel basale als glucose gestimuleerde insuline secretie, vermoedelijk
als gevolg van een defect in een latere stap in het exocytose proces (91). De
pathofysiologische mechanismen waarmee het TCF7L2 gen systemisch bijdraagt tot DM 2
blijven echter nog onvoldoende gekend.
De lever wordt gezien als een cruciaal orgaan, waarvan geweten is dat de glucose en lipide
homeostase ontregeld is bij DM 2, obesitas en het metabool syndroom (zie § 1.4 Metabool
syndroom en § 1.5 Niet alcoholische steatohepatitis (NASH)). De hepatische gluconeogenese,
lipogenese en insulinesignalering, het bestudeerde thema van deze thesis, zouden in zekere
mate beïnvloed worden door de Wnt signalering. Liu H. et al. konden in muizen via reductie
van de hepatisch β-catenine inhoud, aantonen dat β-catenine in staat was de serum glucose
concentraties en de hepatische glucose productie te wijzigen, alsook de hepatische
insulinesignalering te moduleren. Daarnaast werd de interactie tussen β-catenine en FoxO1 in
de lever van deze muizen na vasten aangetoond, welke nodig was voor o.a. G6P en PCK1
genexpressie. Vasten induceerde bovendien de hepatische expressie van Wnt isovormen en
promootte de associatie van β-catenine met FoxO1, in plaats van associatie met TCF
transcriptiefactoren zoals TCF7L2. Daarnaast kon hepatische deletie van β-catenine de
glucose tolerantie verbeteren en de gluconeogenese reduceren (58). Liu W. et al. toonden aan
dat LRP6+/-, welke een sterk verminderde activiteit van de LRP6 co-receptor vertoonden, na
een vetrijk dieet, lagere serum glucose waarden hadden, een verhoogde insulinegevoeligheid
en insulinesignalering van de lever vertoonden, alsook minder intra-hepatisch vet en TG
bevatten. De mutatie, die gekenmerkt wordt door een gedaalde Wnt activiteit, zorgde voor een
gedaalde FoxO1 activiteit en een gedaalde gluconeogenese (verminderde expressie van G6P
en PCK1) (35). Ip et al. suggereerden met behulp van transgene muizen en hepatische
cellijnen, dat TCF7L2 opgereguleerd wordt door insuline (4 uur) en de gluconeogenese mee
helpt onderdrukken op transcriptioneel niveau. Met activators van de canonical Wnt
signaalbaan (Lithium en Wnt-3a) konden zowel in vivo als in vitro de hepatische
gluconeogenese genexpressie en/of de glucose productie gereduceerd worden. Insuline
stimuleerde in vitro de hepatische TCF7L2 expressie en β-catenine activiteit, terwijl voeding
enkel de hepatische TCF7L2 expressie verhoogde in C57BL/6 muizen. Een TCF7L2

39
knockdown leidde in vitro tot een verhoogde PCK1 en G6P genexpressie met verhoogde
glucose productie. De TCF7L2 knockdown kon de repressieve effecten van insuline op de
gluconeogenese echter niet opheffen (59). Deze bevindingen vormen een bevestiging van de
resultaten van Norton et al, welke met behulp van een rat hepatoma cellijn aantoonden dat
TCF7L2 silencing de expressie van PCK1, G6P en FBP1 alsook de glucose productie deed
toenemen. Daarenboven werd aangetoond dat TCF7L2 direct zou kunnen binden met genen
zoals PCK1, FBP1, IRS1, IRS2, die betrokken zijn in de gluconeogenese (60). Dit werd
eveneens bevestigd door Oh et al. Zij vonden ook dat TCF7L2 kan binden met
promotorregio’s van gluconeogene genen en zo de binding van onder meer FoxO1 kan
verhinderen. In tegenstelling tot Ip et al. vond de onderzoeksgroep dat het wegvallen van de
glucagon pathway, eerder dan de inductie door insuline, verantwoordelijk zou zijn voor de
regulatie van TCF7L2 expressie tijdens voeding. Met fosforylatie studies werd bovendien
verondersteld dat TCF7L2 geen directe invloed zou hebben op de insulinesignalering (36). Go
et al. toonden via een mutant LRP6 muismodel (zie § 1.6.2.2 Wnt signalering,
gluconeogenese, insulinesignalering en lipide metabolisme: het verband) aan dat Wnt-3a
administratie zowel in vivo als in vitro een normalisatie van het plasma LDL en TG gaf met
een gedaalde expressie van SREBP1 en enzymen betrokken in de de novo lipogenese, TG en
cholesterol synthese. De onderzoeksgroep suggereerde een werkingsmechanisme waarbij een
verminderde Wnt signalering resulteerde in een verhoogde activiteit van de insuline achtige
groei factor (IGF1) en mTORC2 (een nutriënt gevoelig kinase). Dit mTORC2 zou hierbij
voor een Akt/PKB fosforylatie zorgen die op zijn beurt de activiteit van SREBP1 doet
toenemen met een verhoogde expressie van lipogene enzymen tot gevolg (22). Verhoogde
IGF1 activiteit bij dragers van dezelfde LRP6 mutatie werd reeds door Singh et al. (zie §
1.6.2.4 Dickkopf 1 (DKK1)) aangetoond (29).
In niet gepubliceerde klinische data van de onderzoeksgroep van Prof. Dr. J. Ruige, werd
gezien dat de expressie van SFRP4 en DKK1 in VVW hoger was bij obese patiënten met DM
2 versus (obese) controles. Ehrlund et al. hadden reeds aangetoond dat SFRP4 verhoogd tot
expressie komt in het VVW van obese patiënten en geassocieerd was met insulineresistentie
(68). Mahdi et al. vonden eveneens verhoogde systemische SFRP4 concentraties die de
glucose tolerantie verminderden en dat de concentratie verhoogd was in het serum van DM 2
patiënten tot meerdere jaren voor diagnose (69). Vermoedelijk zouden deze adipokines en
regulatoren van de Wnt signaalbaan, die bij morbide obesitas en DM 2 een verstoord
secretieprofiel vertonen, via de portale drainage een invloed kunnen uitoefenen op het
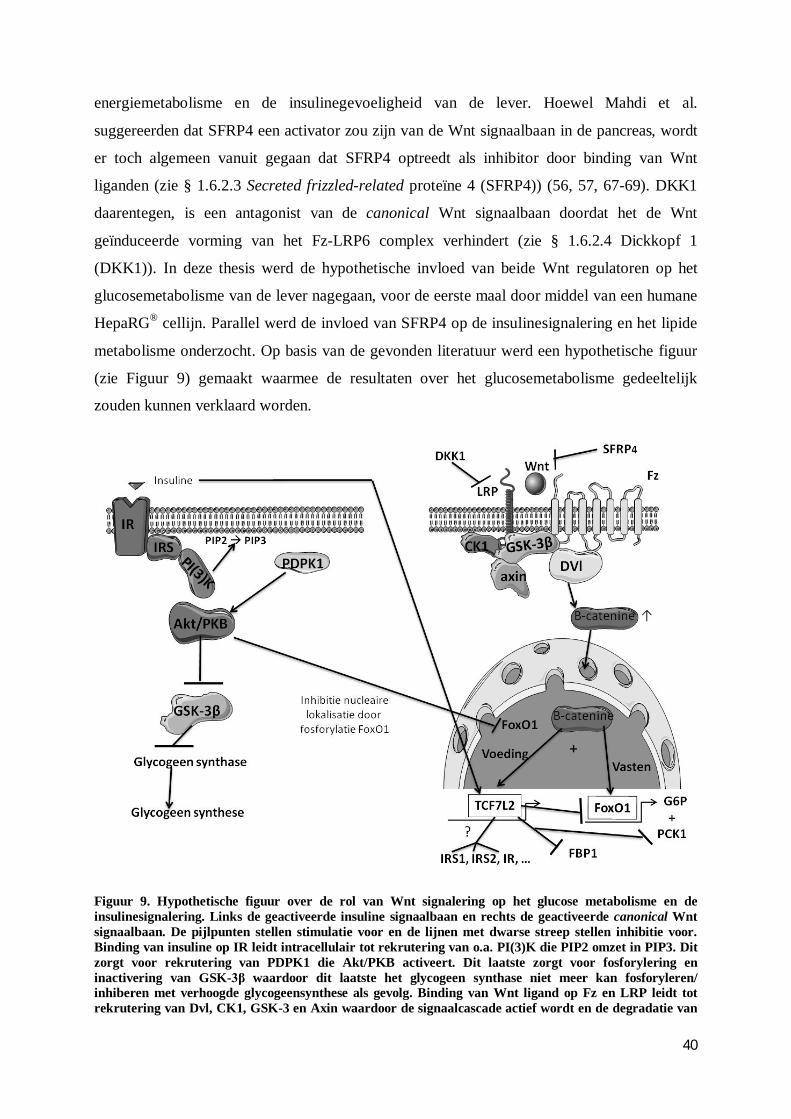
40
energiemetabolisme en de insulinegevoeligheid van de lever. Hoewel Mahdi et al.
suggereerden dat SFRP4 een activator zou zijn van de Wnt signaalbaan in de pancreas, wordt
er toch algemeen vanuit gegaan dat SFRP4 optreedt als inhibitor door binding van Wnt
liganden (zie § 1.6.2.3 Secreted frizzled-related proteïne 4 (SFRP4)) (56, 57, 67-69). DKK1
daarentegen, is een antagonist van de canonical Wnt signaalbaan doordat het de Wnt
geïnduceerde vorming van het Fz-LRP6 complex verhindert (zie § 1.6.2.4 Dickkopf 1
(DKK1)). In deze thesis werd de hypothetische invloed van beide Wnt regulatoren op het
glucosemetabolisme van de lever nagegaan, voor de eerste maal door middel van een humane
HepaRG® cellijn. Parallel werd de invloed van SFRP4 op de insulinesignalering en het lipide
metabolisme onderzocht. Op basis van de gevonden literatuur werd een hypothetische figuur
(zie Figuur 9) gemaakt waarmee de resultaten over het glucosemetabolisme gedeeltelijk
zouden kunnen verklaard worden.
Figuur 9. Hypothetische figuur over de rol van Wnt signalering op het glucose metabolisme en de insulinesignalering. Links de geactiveerde insuline signaalbaan en rechts de geactiveerde canonical Wnt signaalbaan. De pijlpunten stellen stimulatie voor en de lijnen met dwarse streep stellen inhibitie voor. Binding van insuline op IR leidt intracellulair tot rekrutering van o.a. PI(3)K die PIP2 omzet in PIP3. Dit zorgt voor rekrutering van PDPK1 die Akt/PKB activeert. Dit laatste zorgt voor fosforylering en inactivering van GSK-3β waardoor dit laatste het glycogeen synthase niet meer kan fosforyleren/ inhiberen met verhoogde glycogeensynthese als gevolg. Binding van Wnt ligand op Fz en LRP leidt tot rekrutering van Dvl, CK1, GSK-3 en Axin waardoor de signaalcascade actief wordt en de degradatie van

41
β-catenine wegvalt. Dit laatste kan nu diffunderen naar nucleus. Daar activeert het Wnt doelwitgenen door middel van transcriptiefactoren TCF7L2 en FoxO1. Gebruikte afkortingen: Akt/PKB = proteïne kinase B, CK1 = caseïne kinase 1, DKK1 = Dickkopf 1, Dvl = dishevelled, FBP1 = fructose-1,6- bisfosfatase, FoxO1 = forkhead O box 1, Fz = Frizzled receptor, G6P = glucose-6-fosfatase, GSK-3 = glycogeen synthase kinase-3, IR = insulinereceptor, IRS = insulinereceptor substraat, LRP = low density lipoproteïne receptor, PCK1 = fosfoënolpyruvaat carboxykinase 1, PDPK1 = fosfoïnositide afhankelijk kinase 1, PI(3)K = fosfoïnositide afhankelijk kinase 1, PIP2 = fosfatidylinositol-(4,5)-bisfosfaat, PIP3 = fosfatidylinositol-(3,4,5)-trisfosfaat, SFRP4 = secreted frizzled-related proteïne 4, TCF = T cel factor.
In het SFRP4 experiment zorgde blootstelling met insuline (in totaal 48 u) voor een duidelijke
daling in de expressieniveaus van PCK1, G6P en FBP1. Dit zou enerzijds verklaard kunnen
worden door het feit dat insuline het PKB/Akt activeert, dat op zijn beurt FoxO1 fosforyleert
met nucleaire exclusie als gevolg. Een andere mogelijke verklaring kan gegeven worden
doordat insuline de expressie van TCF7L2 stimuleert, die op zijn beurt in staat is om de
gluconeogenese te onderdrukken (36, 58-60). De blootstelling met SFRP4 (100 ng/ml) kon de
onderdrukkende effecten van insuline op de expressie van PCK1 en FBP1 grotendeels
opheffen (zie § 3.1.2 Gluconeogenese, expressieniveaus van PCK1, G6P en FBP1). Daar waar
Wnt activatie tijdens vasten belangrijk blijkt te zijn voor de gluconeogenese respons, zouden
insuline en voeding via de Wnt activatie de gluconeogenese onderdrukken via onder meer
TCF7L2. De bovenstaande resultaten suggereren dat SFRP4, als antagonist van de Wnt
signaalbaan, er toe in staat is om de insuline gemedieerde inhibitie van de gluconeogenese
gedeeltelijk op te heffen. Dit zou kunnen verklaard worden doordat SFRP4 de insuline
gemedieerde activatie van TCF7L2 doet reduceren, waardoor het repressieve effect van
TCF7L2 op de gluconeogene genexpressie wegvalt. Paradoxaal werden echter geen
veranderde expressieniveaus gezien bij G6P na SFRP4 blootstelling. De reden hiervoor zou
kunnen zijn dat TCF7L2 enkel bindt met PCK1 en FBP1 promotor regio’s. In afwezigheid
van insuline leidde SFRP4 (30 ng/ml) blootstelling tot een gedaalde expressie van PCK1
terwijl blootstelling met 100 ng/ml SFRP4 instaat was om zowel de expressie van PCK1 en
FBP1 te doen dalen. Daar tijdens vasten, in dit experiment vertaald door de lage glucose
concentratie (0.5 mM) zonder insuline, Wnt activatie voornamelijk belangrijk lijkt voor
stimulatie van de gluconeogenese, zou SFRP4 de Wnt gemedieerde activatie van FoxO1
kunnen inhiberen (35, 36, 58-60). De interactie tussen de Wnt signaalbaan en de
insulinesignaalbaan blijft echter nog steeds controversieel. Zo zagen Liu H. et al. dat deletie
van β-catenine in de leverstalen van muizen de activiteit (gemeten via fosforylatie studies)
van IRS1-2, Akt/PKB en GSK-3β deed toenemen, terwijl over-expressie het omgekeerde
veroorzaakte (58). Vergelijkbaar zagen Liu W. et al dat de LRP+/- muizen, eveneens
gekenmerkt door een verminderde Wnt activiteit, een verhoogde insulinegevoeligheid hadden
waarbij de IRS1 activiteit verhoogd was (35). Daarentegen konden Singh et al. een

42
verminderde IR, IRS-1, Akt/PKB en GSK-3β activiteit aantonen bij dragers van de LRP6R611C
loss-of-function mutatie. Dit zou onder meer het gevolg zijn van een gedaalde transcriptie van
IR door TCF7L2. (29). Oh et al. suggereerden, op basis van fosforylatie studies, dat TCF7L2
echter geen direct effect had op insulinesignalering (36). Via TCFL2 zouden aldus belangrijke
enzymen uit de insulinesignaalbaan kunnen beïnvloedt worden. In het SFRP4 experiment
waren de expressieniveaus van IR, IRS1 en IRS2 steeds verlaagt na insuline stimulatie, een
fenomeen dat niet beïnvloed werd door SFRP4 zelf. Dit lijkt eerder vreemd omdat een
inductie verwacht werd door insuline blootstelling. Zo konden Ip et al. aantonen dat een
insuline behandeling (4 uur) van HepG2 en Hepa1-6 cellijnen, een verhoogde fosforylatie van
onder meer Akt/PKB en GSK-3β als effect had, wat wijst op een hogere insulinesignalering
(59). Een mogelijke verklaring hiervoor zou zijn dat de 48 uur durende insuline blootstelling
gezorgd heeft voor een hyperinsulinemie-geïnduceerde insulineresistentie, waardoor de cellen
niet meer voldoende zouden reageren op de insuline blootstelling. Zo werden in de levers van
NASH patiënten met insulineresistentie reeds een reductie van Akt/PKB en van
gefosforyleerd Akt/PKB waargenomen, samen met een gedaald expressieniveau van de IR
(51). Indien de cellen insulineresistent zijn, zou volgens Oh et al. de hepatische expressie van
TCF7L2 eveneens gedaald moeten zijn (36). Het feit dat SFRP4 geen invloed vertoonde op de
expressie van sleutelgenen van de insulinesignaalbaan zou eventueel ook het gevolg kunnen
zijn van het tijdstip waarbij de HepaRG® cellen gestimuleerd werden met SFRP4 en insuline.
Naast de insulinesignalering blijft de rol van de Wnt signalering op het lipide metabolisme
ook nog onduidelijk. Terwijl Liu W. et al. een gedaalde hepatische vetinhoud vonden bij
LRP6+/- muizen, toonden Go et al. een hyperlipidemie met verhoogde lipogenese en intra
hepatische vetinhoud bij LRP6R611C muizen (22, 35). In het SFRP4 experiment zorgde de
blootstelling aan insuline voor een gedaalde significante expressie van MTTP, FAS en
SREBP1. Deze resultaten waren eerder onverwacht daar geweten is dat insuline de lipogenese
op reguleert (zie § 1.3 Energiehomeostase en § 1.5.2 Lipide metabolisme, lipotoxiciteit en
oxidatieve stress). Hoewel Donnelly et al. (44) konden aantonen dat obese NAFLD patiënten
een verhoogde de novo lipogenese vertoonden onder vasten, waren de celcultuurcondities in
deze thesis, die geoptimaliseerd waren voor studie van de gluconeogenese, wellicht niet
optimaal voor studie van het lipide metabolisme. Samanez et al. gebruikten voor de studie van
de lipogenese in de IHH cellijn, een cultuur medium met 11 mM glucose (23). Nog rijkere
condities werden gebruikt door Semenkovich et al., waarbij HepG2 cellen gedurende 12 uur
werden blootgesteld aan 25 mM glucose (92).

43
Het experiment met DKK1 lijkt foutief verlopen te zijn. Aan de expressieniveaus van de
gluconeogene genen is te zien dat de welgekende insuline gemedieerde repressie van de
gluconeogenese niet of bijna niet is doorgegaan. Zo is er bij de controles een bijna even grote
PCK1 expressie met of zonder insuline, wat niet mogelijk is. Op het eerste zicht lijkt DKK1
de gluconeogenese niet te beïnvloeden maar deze veronderstellingen kunnen niet gemaakt
worden daar de proef waarschijnlijk mislukt is. Een bevestiging van de SFRP4 data over de
gluconeogenese is dan ook niet mogelijk.
4.3 Mogelijke toekomstperspectieven
Daar de expressieniveaus van IR, IRS1-2 laag waren, waarschijnlijk door een
insulineresistentie, zou een verandering in de volgorde en duur van zowel het adipokine als de
insuline blootstelling van de cellen doorgevoerd kunnen worden. Zo kan gestart worden met
een 24 uur durende adipokine blootstelling, gevolgd door stimulatie met 100 nM insuline
gedurende 4 uur. Omwille van de cruciale rol van FoxO1 alsook de mogelijke rol van
TCF7L2, in het glucosemetabolisme zou het interessant zijn om de expressie van deze genen
mee te bepalen in de toekomst. Daarnaast lijkt het ook nuttig om de Thr-24 fosforylatie en dus
inhibitie van FoxO1 te bestuderen, alsook de nucleaire fractie van FoxO1 te gaan bepalen. Ter
bevestiging van de gluconeogenese kan de glucose productie bepaald worden in het
celcultuurmedium van de HepaRG® cellen. De studie van de insulinesignalering kan eveneens
aangevuld worden met fosforylatie studies van IR β (Tyr 1162/1163), IRS1 (Tyr1222), Akt
(Ser 473) en GSK-3β (Ser 9). Naast gebruik van de Wnt-regulatoren DKK1 en SFRP4, zou
het experiment parallel herhaald kunnen worden met een gekende activator van de canonical
pathway, zijnde Wnt-3a en Lithium, ter vergelijking. Ten slotte zou het interessant zijn om te
weten met welke eiwitten (Wnt liganden) SFRP4 bindt en dus op welke manier het de
gluconeogenese, insulinesignalering en lipide metabolisme reguleert.
4.4 Besluit
Vorige studies hebben de hypothetische rol van viscerale adipokines, in het bijzonder Wnt
regulatoren, op de pathofysiologie van obesitas en DM 2 reeds gesuggereerd. In nog niet
gepubliceerde data werd gezien dat SFRP4 hoger tot expressie komt in het visceraal
vetweefsel van morbide obese DM 2 patiënten. In deze thesis werd in vitro, voor het eerst met
een HepaRG® humane lever cellijn, de rol van de Wnt regulator en visceraal adipokine
SFRP4 op de gluconeogenese aangetoond. Door middel van Real-time quantitative
Polymerase Chain Reaction (qPCR) werd aangetoond dat SFRP4 (100 ng/ml) in staat is om
de insuline gemedieerde expressie van PCK1 en FBP1, beiden sleutelgenen uit de

44
gluconeogenese, op te heffen. Daarnaast was SFRP4 eveneens in staat om, in afwezigheid van
insuline, de expressie van PCK1 en FBP1 te onderdrukken. Deze resultaten onderbouwen het
vermoeden van de rol van Wnt signalering in de gluconeogenese van de lever en suggereren
het bestaan van een crosstalk tussen de Wnt en de insuline signalering. Daarnaast dragen deze
resultaten bij tot de hypothetische stelling waarbij viscerale adipokines, die verhoogd
vrijgesteld worden bij obese DM 2 patiënten, een rol spelen op het glucosemetabolisme in de
lever.

45
5 Referenties 1. Jeffery RW, Harnack LJ (2007). Evidence implicating eating as a primary driver for the obesity
epidemic. Diabetes 56 (11): 2673-2676. 2. The World Health Organization (2009). Global health risks: mortality and burden of disease attributable
to selected major risks. WHO Press. 3. The World Health Organization (1997). Obesity: preventing and managing the global epidemic.
WHO/NUT/NCD/981. 4. Prentice AM, Hennig BJ, Fulford AJ (2008). Evolutionary origins of the obesity epidemic: natural
selection of thrifty genes or genetic drift following predation release? International journal of obesity 32 (11): 1607-1610.
5. Rosen ED, Spiegelman BM (2006). Adipocytes as regulators of energy balance and glucose homeostasis. Nature 444 (7121): 847-853.
6. Must A, Spadano J, Coakley EH, et al. (1999). The disease burden associated with overweight and obesity. JAMA: the journal of the American Medical Association 282 (16): 1523-1529.
7. Wajchenberg BL (2000). Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome. Endocrine Reviews 21 (6): 697-738.
8. Smith SR, Lovejoy JC, Greenway F, et al. (2001). Contributions of total body fat, abdominal subcutaneous adipose tissue compartments, and visceral adipose tissue to the metabolic complications of obesity. Metabolism-Clinical and Experimental 50 (4): 425-435.
9. Withrow D, Alter DA (2011). The economic burden of obesity worldwide: a systematic review of the direct costs of obesity. Obesity Reviews 12 (2): 131-141.
10. Gerard J, Tortora BD (2006). Principles of anatomy and Physiology. Hoboken: John Wiley & Sons. 11. Van Herpen NA, Schrauwen-Hinderling VB (2008). Lipid accumulation in non-adipose tissue and
lipotoxicity. Physiology & Behavior 94 (2): 231-241. 12. Geer EB, Shen W (2009). Gender differences in insulin resistance, body composition, and energy
balance. Gender medicine 6 Supplement 1: 60-75. 13. Rytka JM, Wueest S, Schoenle EJ, et al. (2011). The portal theory supported by venous drainage-
selective fat transplantation. Diabetes 60(1): 56-63. 14. Avram AS, Avram MM, James WD (2005). Subcutaneous fat in normal and diseased states - 2.
Anatomy and physiology of white and brown adipose tissue. Journal of the American Academy of Dermatology 53 (4): 671-683.
15. Trayhurn P, Wang B, Wood IS (2008). Hypoxia in adipose tissue: a basis for the dysregulation of tissue function in obesity? The British journal of nutrition 100(2): 227-235.
16. Lafontan M, Langin D (2009). Lipolysis and lipid mobilization in human adipose tissue. Progress in Lipid Research 48 (5): 275-297.
17. Spalding KL, Arner E, Westermark PO, et al. (2008) Dynamics of fat cell turnover in humans. Nature 453 (7196): 783-787.
18. Gustafson B, Smith U (2012). The WNT inhibitor Dickkopf 1 and bone morphogenetic protein 4 rescue adipogenesis in hypertrophic obesity in humans. Diabetes 61(5): 1217-1224.
19. Shepherd PR, Kahn BB (1999). Mechanisms of disease - Glucose transporters and insulin action - Implications for insulin resistance and diabetes mellitus. The New England Journal of Medicine 341 (4): 248-57.
20. Ruige JB, Bekaert M, Lapauw B, et al. (2012). Sex Steroid-Induced Changes in Circulating Monocyte Chemoattractant Protein-1 Levels May Contribute to Metabolic Dysfunction in Obese Men. The Journal of Clinical Endocrinology & Metabolism 97(7): E1187-E1191.
21. Jitrapakdee S (2012). Transcription factors and coactivators controlling nutrient and hormonal regulation of hepatic gluconeogenesis. International Journal of Biochemistry & Cell Biology 44(1): 33-45.
22. Go GW, Srivastava R, Hernandez-Ono A, et al. (2014). The combined hyperlipidemia caused by impaired Wnt-LRP6 signaling is reversed by Wnt3a rescue. Cell Metababolism 19(2): 209-220.
23. Samanez CH, Caron S, Briand O et al. (2012). The human hepatocyte cell lines IHH and HepaRG: models to study glucose, lipid and lipoprotein metabolism. Archives of Physiology and Biochemistry 118(3): 102-111.
24. Saltiel AR, Kahn CR 2001. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 414 (6865): 799-806.
25. Shelness GS, Ingram MF, Huang XF, et al. (1999). Apolipoprotein B in the rough endoplasmic reticulum: translation, translocation and the initiation of lipoprotein assembly. The Journal of nutrition 129 (2S Supplement): 456s-462s.
26. Yoon JC, Puigserver P, Chen G, et al. (2001). Control of hepatic gluconeogenesis through the transcriptional coactivator PGC-1. Nature 413 (6852): 131-138.

46
27. Cheatham B, Kahn CR (1995). Insulin action and the insulin signalling network. Endocrine Reviews 16 (2): 117-142.
28. Puigserver P, Rhee J, Donovan J, et al (2003). Insulin-regulated hepatic gluconeogenesis through FOXO1-PGC-1 alpha interaction. Nature 423 (6939): 550-555.
29. Singh R, De Aguiar RB, Naik S et al. (2013). LRP6 enhances glucose metabolism by promoting TCF7L2-dependent insulin receptor expression and IGF receptor stabilization in humans. Cell Metabolism 17 (2): 197-209.
30. Eckel RH, Grundy SM, et al. (2005). The metabolic syndrome. The Lancet 365(9468): 1415-1428. 31. DeFronzo RA, Tobin JD, Andres R (1979). Glucose clamp technique: a method for quantifying insulin
secretion and resistance. The American journal of physiology 237(3): E214-223. 32. Dixon JB, Bhathal PS, O'Brien PE (2001). Nonalcoholic fatty liver disease: Predictors of nonalcoholic
steatohepatitis and liver fibrosis in the severely obese. Gastroenterology 121(1): 91-100. 33. Marchesini G, Brizi M, Morselli-Labate AM, et al. (1997) Association of nonalcoholic fatty liver
disease with insulin resistance. The American journal of medicine 107(5): 450-455. 34. Florez JC, Jablonski KA, Bayley N, et al. (2006). TCF7L2 polymorphisms and progression to diabetes
in the Diabetes Prevention Program. The New England Journal of Medicine 355(3): 241-250. 35. Liu WZ, Singh R, Choi CS, et al.(2012). Low Density Lipoprotein (LDL) Receptor-related Protein 6
(LRP6) Regulates Body Fat and Glucose Homeostasis by Modulating Nutrient Sensing Pathways and Mitochondrial Energy Expenditure. Journal of Biological Chemistry 287 (10): 7213-7223.
36. Oh KJ, Park J, Kim SS, et al. (2012). TCF7L2 modulates glucose homeostasis by regulating CREB- and FoxO1-dependent transcriptional pathway in the liver. Public Library of Science genetics;8(9): e1002986.
37. Karastergiou K, Mohamed-Ali V (2010). The autocrine and paracrine roles of adipokines. Molecular and Cellular Endocrinology 318 (1-2): 69-78.
38. Omagari K, Kadokawa Y, Masuda J, et al. (2002). Fatty liver in non-alcoholic non-overweight Japanese adults: incidence and clinical characteristics. Journal of gastroenterology and hepatology. 17(10): 1098-1105.
39. Browning JD, Horton JD (2004). Molecular mediators of hepatic steatosis and liver injury. The Journal of clinical investigation 114 (2): 147-152.
40. Kershaw EE, Flier JS (2004). Adipose tissue as an endocrine organ. The Journal of Clinical Endocrinology & Metababolism 89 (6): 2548-2556.
41. Samuel VT, Petersen KF, Shulman GI (2010). Lipid-induced insulin resistance: unravelling the mechanism. The Lancet 375 (9733): 2267-2277.
42. Day CP, James OFW (1998). Steatohepatitis: A tale of two "hits"? Gastroenterology 114(4): 842-845. 43. Browning JD, Szczepaniak LS, Dobbins R, et al. (2004). Prevalence of hepatic steatosis in an urban
population in the United States: Impact of ethnicity. Hepatology 40(6): 1387-1395. 44. Donnelly KL, Smith CI, Schwarzenberg SJ, et al. (2005). Sources of fatty acids stored in liver and
secreted via lipoproteins in patients with nonalcoholic fatty liver disease. The Journal of Clinical Investigation 115(5): 1343-1351.
45. Blomme B, Francque S, Trepo E, et al. (2012). N-glycan based biomarker distinguishing non-alcoholic steatohepatitis from steatosis independently of fibrosis. Digestive and liver disease: official journal of the Italian Society of Gastroenterology and the Italian Association for the Study of the Liver. 44(4): 315-322.
46. Saadeh S, Younossi ZM, Remer EM, et al. (2002). The utility of radiological imaging in nonalcoholic fatty liver disease. Gastroenterology 123(3): 745-750.
47. Argentou M, Tiniakos DG, Karanikolas M, et al. (2009). Adipokine Serum Levels Are Related to Liver Histology in Severely Obese Patients Undergoing Bariatric Surgery. Obesity Surgery 19(9): 1313-1323.
48. Leamy AK, Egnatchik RA, Young JD (2013). Molecular mechanisms and the role of saturated fatty acids in the progression of non-alcoholic fatty liver disease. Progress in Lipid Research 52(1): 165-174.
49. Kleiner DE, Brunt EM, Van Natta M, et al. (2005). Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 41(6): 1313-1321.
50. Mormone E, George J, Nieto N (2011). Molecular pathogenesis of hepatic fibrosis and current therapeutic approaches. Chemico-biological interactions 193(3): 225-231.
51. Valenti L, Rametta R, Dongiovanni P, et al (2008). Increased expression and activity of the transcription factor FOXO1 in nonalcoholic steatohepatitis. Diabetes 57 (5): 1355-1362.
52. Shen XH, Zhang KZ, Kaufman RJ (2004). The unfolded protein response - a stress signaling pathway of the endoplasmic reticulum. Journal of Chemical Neuroanatomy 28(1-2): 79-92.
53. Fain JN, Madan AK, Hiler ML, et al. (2004) Comparison of the release of adipokines by adipose tissue, adipose tissue matrix, and Adipocytes from visceral and subcutaneous abdominal adipose tissues of obese humans. Endocrinology 145 (5): 2273-2282.

47
54. Skurk T, Alberti-Huber C, Herder C et al. (2007). Relationship between adipocyte size and adipokine expression and secretion. The Journal of Clinical Endocrinology & Metababolism 92(3): 1023-1033.
55. Cancello R, Tordjman J, Poitou C, et al. (2006) Increased infiltration of macrophages in omental adipose tissue is associated with marked hepatic lesions in morbid human obesity. Diabetes. 55(6): 1554-1561.
56. Logan CY, Nusse R (2004). The Wnt signaling pathway in development and disease. Annual review of cell and developmental biology 20: 781-810.
57. MacDonald BT, Tamai K, He X (2009). Wnt/beta-Catenin Signaling: Components, Mechanisms, and Diseases. Developmental Cell 17 (1): 9-26.
58. Liu H, Fergusson MM, Wu JJ, et al. (2011) Wnt signaling regulates hepatic metabolism. Science signaling 4(158): ra6.
59. Ip W, Shao W, Chiang Y-TA, et al. (2012). The Wnt signaling pathway effector TCF7L2 is up-regulated by insulin and represses hepatic gluconeogenesis. American Journal of Physiology - Endocrinology and Metabolism 303: E1166-E1176.
60. Norton L, Fourcaudot M, Abdul-Ghani MA et al. (2011). Chromatin occupancy of transcription factor 7-like 2 (TCF7L2) and its role in hepatic glucose metabolism. Diabetologia 54(12): 3132-3142.
61. Hoogeboom D, Essers MA, Polderman PE, et al. (2008) Interaction of FOXO with beta-catenin inhibits beta-catenin/T cell factor activity. The Journal of biological chemistry 283 (14): 9224-9230.
62. Matsuzaki H, Daitoku H, Hatta M, et al. (2005). Acetylation of Foxo1 alters its DNA-binding ability and sensitivity to phosphorylation. Proceedings of the National Academy of Sciences of the United States of America 102(32): 11278-11283.
63. Grant SFA, Thorleifsson G, Reynisdottir I, et al. (2006). Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nature Genetics 38(3): 320-323.
64. Ding VW, Chen RH, McCormick F (2000). Differential regulation of glycogen synthase kinase 3 beta by insulin and Wnt signaling. Journal of Biological Chemistry 275 (42): 32475-32481.
65. Patel S, Doble B, Woodgett JR (2004). Glycogen synthase kinase-3 in insulin and Wnt signalling: a double-edged sword? Biochemical Society Transactions 32: 803-808.
66. McManus EJ, Sakamoto K, Armit LJ,et al.(2005) Role that phosphorylation of GSK3 plays in insulin. Embo Journal 24 (8): 1571-1583.
67. Bovolenta P, Esteve P, Ruiz JM, et al. (2008). Beyond Wnt inhibition: new functions of secreted Frizzled-related proteins in development and disease. Journal of Cell Science 121(6): 737-746.
68. Ehrlund A, Mejhert N, Lorente-Cebrian S, et al. (2013) Characterization of the Wnt Inhibitors Secreted Frizzled-Related Proteins (SFRPs) in Human Adipose Tissue. The jorunal of Clinical Endocrinology & Metabolism 98 (3): E503-E508.
69. Mahdi T, Hanzelmann S, Salehi A, et al. (2012). Secreted Frizzled-Related Protein 4 Reduces Insulin Secretion and Is Overexpressed in Type 2 Diabetes. Cell Metabolism 16 (5): 625-633.
70. Park JR, Jung JW, Lee YS, et al. (2008). The roles of Wnt antagonists Dkk1 and sFRP4 during adipogenesis of human adipose tissue-derived mesenchymal stem cells. Cell Proliferation 41(6): 859-874.
71. Semenov MV, Zhang X, He X (2008). DKK1 antagonizes Wnt signaling without promotion of LRP6 internalization and degradation. The Journal of biological chemistry 283 (31): 21427-21432.
72. Baranova A, Gowder SJ, Schlauch K, et al. (2006). Gene expression of leptin, resistin, and adiponectin in the white adipose tissue of obese patients with non-alcoholic fatty liver disease and insulin resistance. Obesity Surgery 16( 9): 1118-1125.
73. Hui JM, Hodge A, Farrell GC, et al. (2004) Beyond insulin resistance in NASH: TNF-alpha or adiponectin? Hepatology 40 (1): 46-54.
74. Jiang LL, Li L, Hong XF, et al. (2009). Patients with nonalcoholic fatty liver disease display increased serum resistin levels and decreased adiponectin levels. European Journal of Gastroenterology & Hepatology 21(6): 662-666.
75. Yilmaz Y, Yonal O, Kurt R et al. (2011). Serum levels of omentin, chemerin and adipsin in patients with biopsy-proven nonalcoholic fatty liver disease. Scandinavian Journal of Gastroenterology 46(1): 91-97.
76. Weisberg SP, McCann D, Desai M, et al. (2003). Obesity is associated with macrophage accumulation in adipose tissue. The Journal of clinical investigation 112 (12): 1796-1808.
77. Crespo J, Cayon A, Fernandez-Gil P, et al. (2001). Gene expression of tumor necrosis factor alpha and TNF-receptors, p55 and p75, in nonalcoholic steatohepatitis patients. Hepatology 34(6): 1158-1163.
78. Wieckowska A, Papouchado BG, Li ZZ (2008). Increased hepatic and circulating interleukin-6 levels in human nonalcoholic steatohepatitis. American Journal of Gastroenterology 103(6): 1372-1379.
79. Senn JJ, Klover PJ, Nowak IA, et al. (2002). Interleukin-6 induces cellular insulin resistance in hepatocytes. Diabetes 51(12): 3391-3399.

48
80. Steppan CM, Bailey ST, Bhat S, et al. (2001) The hormone resistin links obesity to diabetes. Nature 409(6818): 307-312.
81. Sell H, Divoux A, Poitou C, et al. (2010). Chemerin Correlates with Markers for Fatty Liver in Morbidly Obese Patients and Strongly Decreases after Weight Loss Induced by Bariatric Surgery. The Journal of Clinical Endocrinology & Metabolism 95(6): 2892-2896.
82. Shin HY, Lee DC, Chu SH, et al. (2012). Chemerin levels are positively correlated with abdominal visceral fat accumulation. Clinical Endocrinology 77(1): 47-50.
83. Sell H, Laurencikiene J, Taube A, et al. (2009). Chemerin is a novel adipocyte-derived factor inducing insulin resistance in primary human skeletal muscle cells. Diabetes 58(12): 2731-2740.
84. Docke S, Lock JF, Birkenfeld AL, et al. (2013). Elevated hepatic chemerin mRNA expression in human non-alcoholic fatty liver disease. European Journal of Endocrinology 169(5): 547-557.
85. Kukla M, Zwirska-Korczala K, Hartleb M, et al. (2010). Serum chemerin and vaspin in non-alcoholic fatty liver disease. Scandinavian Journal of Gastroenterology 45(2): 235-242.
86. Gripon P, Rumin S, Urban S, et al. (2002). Infection of a human hepatoma cell line by hepatitis B virus. Proceedings of the National Academy of Sciences of the United States of America 99(24): 15655-15660.
87. Ceelen L, De Spiegelaere W, David M, et al. (2011). Critical selection of reliable reference genes for gene expression study in the HepaRG cell line. Biochemical Pharmacology 81(10): 1255-1261.
88. Taylor S, Wakem M, Dijkman G, et al. (2010). A practical approach to RT-qPCR-Publishing data that conform to the MIQE guidelines. Methods 50(4): S1-5.
89. Vandesompele J, De Preter K, Pattyn F, et al. (2002). Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome biology 3(7): Research0034.1-0034.11.
90. Kanazawa A, Tsukada S, Sekine A, T et al. (2004). Association of the gene encoding wingless-type mammary tumor virus integration-site family member 5B (WNT5B) with type 2 diabetes. American journal of human genetics 75(5): 832-843.
91. da Silva Xavier G, Loder MK, McDonald A, et al. (2009). TCF7L2 regulates late events in insulin secretion from pancreatic islet beta-cells. Diabetes 58(4): 894-905.
92. Semenkovich CF, Coleman T, Goforth R. (1993). Physiologic concentrations of glucose regulate fatty acid synthase activity in HepG2 cells by mediating fatty acid synthase mRNA stability. The Journal of biological chemistry 268(10): 6961-6970.