A reappraisal of Eurypterus dumonti S 1917 and its …(1958, p. 1141). However, S to rm e r & W a te...
12
BULLETIN DE L'INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN SCIENCES DE LA TERRE, 76: 79-90, 2006 AARDWETENSCHAPPEN, 76: 79-90, 2006 A reappraisal of Eurypterus dumonti S tainier , 1917 and its position within the Adelophthalmidae T ollerton , 1989 by O. Erik TETLIE & Peter VAN ROY Tetlie, O.E. & Van Roy, P., 2006 — A reappraisal of Eurypterus dumonti Stainier, 1917 and its position within the Adelophthalmidae Tollerton, 1989. Bulletin de l'Institut royal des Sciences naturelles de Belgique, Sciences de la Terre 76: 79-90, 6 figs., Brussels, April 15, 2006-ISSN 0374-6291. Abstract The Carboniferous eurypterid Eurypterus dumonti Stainier, 1917 from Mechelen-aan-de-Maas (Maasmechelen), Belgium is redescribed and assigned to the genus Adelophthalmus. It is diagnosed as having a raised triangle of unknown function, dorsally on opisthosomal segment 7 and a carapace articulating laterally against the second opisthosomal segment. Earlier assignments of this species to the genus Unionopterus, based on the carapace shape figured in the original description (Stainier 1917), are incorrect and the existence of a Carboniferous eurypterid with the characteristics described for Unionopterus must be questioned. The appendages in A. dumonti are completely exposed and provide an unrivalled insight into the number of appendage podomeres in the genus; this species is interpreted to have a podomere count consistent with most other eurypterids. Small pustules previously thought to be cuticle sculpture are here interpreted as diagenetic “ dumb-bells” (see Briggs & Wilby 1996) following microbial activity on the carcase prior to fossilisation. These “ dumb-bells” are the oldest non-marine record of this diagenetic feature. A number of other features are also interpreted differently from the original descrip tion. Possible lineages within the Adelophthalmus clade are identified, A. dumonti is probably closely related to A. imhofi (Czech Republic) and A. moyseyi (United Kingdom). Key words: Eurypterida, appendages, 'dumb-bells’, Belgium, Coal Measures. Résumé L’euryptéride carbonifère de Mechelen-sur-Meuse (Maasmechelen), Belgique, originalement décrit comme Eurypterus dumonti Stainier, 1917 fait ici le sujet d’une nouvelle description, résultant de son transfert au genre Adelophthalmus. Cet euryptéride est caractérisé par la possession d’un triangle surélévé sur le segment opisthosomal 7, et par une carapace articulant latéralement contre le deuxième segment opisthosomal. L’attribution de cette espèce au genre Union opterus, basée sur la forme de la carapace comme figurée dans la description originale (Stainier 1917) s’est révélée incorrecte. L’exis tence d’un euryptéride carbonifère avec des caractéristiques comme celles décrites pour Unionopterus, est assez douteuse. Les appendices d’Adelophthalmus dumonti sont exceptionellement bien exposées et permettent d’obtenir une idée précise du nombre de podomères appen- diculaires de ce genre. Le nombre de podomères de A. dumonti est interprété comme comparable à celui d’autres euryptérides. Des petites pustules originalement considérées comme ornementales sont réinter prétées ici comme des "dumb-bells” (cfr. Briggs & Wilby 1996), résultant d’action bactérielle avant la fossilisation. Ces structures in diquent que le fossile n’est pas une exuvie. L’interprétation d’un nombre d’autres caractéristiques diffère aussi de celle de la description originale. Des lignes de descendance possible du clade Adelophthalmus sont identifiées, et la proximité d 'A. dumonti à A. imhofi (République Tchèque) et à A. moyseyi (Royaume-Uni) est suggérée. Mots-clefs: Eurypterida, appendices, 'dumb-bells’, Belgique, terrain houiller. Introduction Eurypterids are a diverse group of Palaeozoic, aquatic chelicerates ranging from the Upper Ordovician (Tollerton 2004) to the Upper Permian (Plotnick 1983). They occur most frequently in the Silurian and Lower Devonian of Europe and North America. Post- Devonian eurypterids are rare and had migrated from their earlier marginal marine environments into brackish and freshwater settings (Plotnick 1983; Tetlie 2004) - the Carboniferous Coal Measures of Europe, North America and China being classic examples. Although known from the Devonian (Tetlie et al. 2004; Tetlie & Dunlop 2005; Poschmann 2005), adelophthalmids constitute most of the post-Devonian eurypterids, both in terms of the number of species and individuals. They represent one of only two eurypterid clades to survive into the Carboniferous; the other, and more diverse clade, includes the gigantic sweep-feeding hibbertopterids (Hib- bertopterus, Cyrtoctenus, Campylocephalus, Hastimima, Dunsopterus and Vernonopterus) and the peculiar wood- wardopterids (the Devonian Borchgrevinkium and the Carboniferous Woodwardopterus, Mycterops and Mega- rachne), this latter group being allied to the hibbertopter ids (Selden et al. 2005). Adelophthalmids are small, streamlined, nektonic eur ypterids with prominent cuticle sculpture, which have previously been referred to six genera. As discussed by Tetlie & Dunlop (2005) only Adelophthalmus von M eyer, 1853 and Unionopterus Chernyshev, 1948 appear to be valid, and the four other proposed genera are synonyms of Adelophthalmus. While the morphology of Adelophthalmus is relatively well-known (Kjellesvig-
Transcript of A reappraisal of Eurypterus dumonti S 1917 and its …(1958, p. 1141). However, S to rm e r & W a te...

BULLETIN DE L'INSTITUT ROYAL DES SCIENCES NATURELLES DE BELGIQUE BULLETIN VAN HET KONINKLIJK BELGISCH INSTITUUT VOOR NATUURWETENSCHAPPEN
SCIENCES DE LA TERRE, 76: 79-90, 2006 AARDW ETENSCHAPPEN, 76: 79-90, 2006
A reappraisal o f Eurypterus dumonti S t a i n i e r , 1917 and its position within the Adelophthalmidae T o l l e r t o n , 1989
by O. Erik TETLIE & Peter VAN ROY
T e tl ie , O.E. & V an R oy , P., 2006 — A reappraisal o f Eurypterus dumonti S ta in ie r , 1917 and its position within the Adelophthalmidae T o l l e r t o n , 1989. Bulletin de l'Institut royal des Sciences naturelles de Belgique, Sciences de la Terre 76: 79-90, 6 figs., Brussels, April 15, 2 0 0 6 -IS S N 0374-6291.
A bstract
The Carboniferous eurypterid Eurypterus dumonti S ta in ie r , 1917 from Mechelen-aan-de-Maas (Maasmechelen), Belgium is redescribed and assigned to the genus Adelophthalmus. It is diagnosed as having a raised triangle o f unknown function, dorsally on opisthosomal segment 7 and a carapace articulating laterally against the second opisthosomal segment. Earlier assignments o f this species to the genus Unionopterus, based on the carapace shape figured in the original description (S ta in ie r 1917), are incorrect and the existence o f a Carboniferous eurypterid with the characteristics described for Unionopterus must be questioned. The appendages in A. dumonti are completely exposed and provide an unrivalled insight into the number o f appendage podomeres in the genus; this species is interpreted to have a podomere count consistent with most other eurypterids. Small pustules previously thought to be cuticle sculpture are here interpreted as diagenetic “ dumb-bells” (see B rig g s & W ilb y 1996) following microbial activity on the carcase prior to fossilisation. These “ dumb-bells” are the oldest non-marine record o f this diagenetic feature. A number o f other features are also interpreted differently from the original description. Possible lineages within the Adelophthalmus clade are identified, A. dumonti is probably closely related to A. imhofi (Czech Republic) and A. moyseyi (United Kingdom).
Key words: Eurypterida, appendages, 'dumb-bells’, Belgium, Coal Measures.
R ésum é
L’euryptéride carbonifère de Mechelen-sur-Meuse (Maasmechelen), Belgique, originalement décrit comme Eurypterus dumonti S ta in ie r , 1917 fait ici le sujet d ’une nouvelle description, résultant de son transfert au genre Adelophthalmus. Cet euryptéride est caractérisé par la possession d ’un triangle surélévé sur le segment opisthosomal 7, et par une carapace articulant latéralement contre le deuxième segment opisthosomal. L ’attribution de cette espèce au genre Unionopterus, basée sur la forme de la carapace comme figurée dans la description originale ( S ta in ie r 1917) s ’est révélée incorrecte. L ’existence d ’un euryptéride carbonifère avec des caractéristiques comme celles décrites pour Unionopterus, est assez douteuse. Les appendices d ’Adelophthalmus dumonti sont exceptionellement bien exposées et permettent d ’obtenir une idée précise du nombre de podomères appen- diculaires de ce genre. Le nombre de podomères de A. dumonti est interprété comme comparable à celui d ’autres euryptérides. Des petites pustules originalement considérées comme ornementales sont réinter
prétées ici comme des "dum b-bells” (cfr. B rig g s & W ilb y 1996), résultant d ’action bactérielle avant la fossilisation. Ces structures indiquent que le fossile n’est pas une exuvie. L ’interprétation d ’un nombre d ’autres caractéristiques diffère aussi de celle de la description originale. Des lignes de descendance possible du clade Adelophthalmus sont identifiées, et la proximité d 'A. dumonti à A. imhofi (République Tchèque) et à A. moyseyi (Royaume-Uni) est suggérée.
Mots-clefs: Eurypterida, appendices, 'dumb-bells’, Belgique, terrain houiller.
Introduction
E urypterids are a d iverse group o f Palaeozoic, aquatic chelicerates rang ing from the U pper O rdovician ( T o l l e r t o n 2004) to the U pper Perm ian ( P l o t n i c k 1983). They occur m ost frequently in the S ilurian and L ow er D evonian o f E urope and N orth A m erica. Post- D evonian eurypterids are rare and had m igra ted from the ir earlier m arginal m arine env ironm ents into brackish and freshw ater settings ( P l o t n i c k 1983; T e t l i e 2004) - the C arboniferous C oal M easures o f E urope, N orth A m erica and C hina being classic exam ples. A lthough know n from the D evonian ( T e t l i e et al. 2004; T e t l i e & D u n l o p 2005; P o s c h m a n n 2005), adelophthalm ids constitu te m ost o f the post-D evonian eurypterids, both in term s o f the num ber o f species and individuals. T hey represen t one o f only tw o eurypterid clades to survive into the C arboniferous; the o ther, and m ore d iverse clade, includes the g igan tic sw eep-feeding h ibbertop terids (Hib- bertopterus, Cyrtoctenus, Campylocephalus, Hastimima, Dunsopterus and Vernonopterus) and the pecu lia r w ood- w ardopterids (the D evonian Borchgrevinkium and the C arbon iferous Woodwardopterus, M ycterops and Mega- rachne), th is la tter group being allied to the h ibbertop terids ( S e l d e n et al. 2005).
A delophthalm ids are sm all, stream lined , nek ton ic eu ryp terids w ith p rom inent cu ticle scu lp ture, w hich have prev iously been referred to six genera. A s d iscussed by T e t l i e & D u n l o p (2005) only Adelophthalmus v o n M e y e r , 1853 and Unionopterus C h e r n y s h e v , 1948 appear to be valid , and the four o ther p roposed genera are synonym s o f Adelophthalmus. W hile the m orphology o f Adelophthalmus is re la tive ly w ell-know n ( K j e l l e s v i g -

80 O. Erik T E T LIE & Peter V A N ROY
W a e r i n g 1948, 1963; V a n O y e n 1956; W i l l s 1964; K u e s & K i e t z k e 1 9 8 1 ; P o s c h m a n n in press), our know ledge o f Unionopterus is ex trem ely p o o r; th is g en u s c o n ta in s o n ly o n e species a n d is k n o w n o n ly fro m a sing le sp ec im en fro m K a z a k h s ta n , d esc rib ed by C h e r
n y s h e v (1948) as U. anastasiae. T h e o rig in a l d esc r ip tio n w as in R u ss ian a n d c o n ta in e d ra th e r p o o r illu stra t io n s . B ased so le ly u p o n these figu res o f C h e r n y s h e v (1948), th e g en u s h as v a rio u s ly been in te rp re te d as allied to Adelophthalmus ( C a s t e r & K j e l l e s v i g - W a e r i n g 1964; T o l l e r t o n 1989), incertae sedis ( N o v o j i l o v 1962; S t o r m e r 1974; P l o t n i c k
1983), o r w as ig n o red a l to g e th e r ( S t o r m e r 1955). T e t l i e (2004) a n d T e t l i e & D u n l o p (2005) rea lised th a t th e sp ec im en d esc rib ed by S t a i n i e r (1917) as Eurypterus dumonti a n d la te r assig n ed to Adelophthalmus by V a n O y e n (1956), a p p e a re d d iffe re n t fro m o th e r species o f Adelophthalmus. T h ey lis ted som e s im ila ritie s , espec ia lly in te rm s o f c a ra p a c e sh a p e a n d th e w id th o f th e m a rg in a l rim , to th e specim en d esc rib ed by C h e r n y s h e v (1948) a n d suggested th a t th e B elgian species m ig h t a lso b e lo n g to Unionopterus. A s d e m o n s tra te d below , th is co n c lu s io n w as in c o rrec t; th e a n te r io r o f th e c a ra p a c e o f th e B elg ian fossil is in c o m p le te ly p rese rv ed , a n d its sh a p e is n o t tra p e z o id as f ig u red by S t a i n i e r (1917), b u t p a ra b o lic , w ith a n a r ro w m a rg in a l rim , as in o th e r species o f Adelophthalmus.
Except fo r A. dumonti, several o ther eurypterids are recorded from B elgium . T he o ldest B elgian eurypterids are Cyrtoctenus dewalquei ( F r a i p o n t , 1889) and Adelophthalmus (?) lohesti ( D e w a l q u e in F r a i p o n t , 1889) from the m iddle to late F am ennian (late D evonian) o f the C ondroz G roup at P on t de B onne M odave. T he single specim en o f E. lohesti w as first in terpreted as possib ly belong ing to Adelophthalmus by K j e l l e s v i g - W a e r i n g
(1958, p. 1141). H ow ever, S t o r m e r & W a t e r s t o n
( 1968, p. 83) in terpreted it as a possib le sty lonurid , a view supported here. M ost o f the o ther fragm entary fossils figured by F r a i p o n t (1889) w ere re in terp re ted by S t o r m e r & W a t e r s t o n (1968) as belonging to Cyrtoctenus. The probab ly related w oodw ardopterid Mycterops matthieui P r u v o s t , 1924, is know n from the M ons M br. o f the C harlero i Fm ., C harlero i, B ashkirian . A part from A. dumonti, the B elgian C arboniferous has also y ie lded th ree o ther unequivocal adelophthalm ids. A. moyseyi ( W o o d w a r d , 1907) w as recorded from the M ons M br. o f the C harlero i Fm ., in the C oal M easures o f B ernissart, B ashkirian by P r u v o s t (1930). A lso from the sam e unit com es A. corneti ( P r u v o s t , 1939), co llected from a drill core at R ieu-du-C ur, Q uaregnon. F inally , A. combieri ( P r u v o s t , 1930) is know n from the A s M br. o f the C harlero i Fm . C harbonnages R eunis coal m ine, C harleroi, B ashkirian . A ll nam ed occurrences o f eurypterids in B elgium are from the U pper D evonian or C arboniferous; the country has one o f the m ost d iverse U pper Palaeozoic eu rypterid faunas know n.
Late C arb oniferou s geology o f the C am pine Basin
T he B elg ian C arbon iferous is c lassica lly d iv ided into a fully m arine, carbonate-dom inated “ D inan tian” , over- lain by a p redom inan tly continen tal, s ilic iclastic coal- bearing “ S iles ian ” . T he in ternationally agreed chronos- tratig raphic standard scale fo r the late C arbon iferous has not found w ide accep tance in B elgium , and the trad itional d iv isions o f the “ S iles ian ” into N am urian A, B and C, and W estphalian A, B, C and D are still w idely used (Fig. 1).
W hereas the sou thern W allon ian B asin ou tcrops at the surface, the coal deposits o f the northern C am pine Basin a re en tirely covered by post-C arbon iferous deposits. The C am pine B asin can be fu rther subdiv ided into the w estern A ntw erp C am pine, only con tain ing S erpukhovian (N am urian A -C ) and B ashkirian (W estphalian A -B ) deposits p oo r in coal, and the eastern L im burg C am pine, p reserv ing coa l-rich M oscovian (W estphalian C -D ) deposits. In the C am pine B asin, the transition from the fu lly -m arine carbonate-dom inated “ D inan tian” to the continental “ S iles ian” is rep resen ted by the S ouvré Fm ., dating to the V iséan-Serpukhovian boundary. It is overla in by the B elg ian C oal M easures G roup, con tain ing all C arbon iferous silic iclastic coa l-bearing sedim ents. This group ev idences a regression, w ith a transition from a m arine p ro -delta environm ent near the base, th rough low er/upper delta p lain , to low er/upper alluv ia l p la in near the top ( L a n g e n a e k e r & D u s a r 1992; D r e e s e n et al. 1995). In the ir recen t rev iew o f the late C arbon iferous stra tig raphy o f B elgium , D e l m e r et al. (2001) subdiv ided the B elgian C oal M easures G roup into six form ations (Fig. 2).
B ecause B elgian C arbon iferous eu ryp terid d iscoveries are lim ited to the C hâtelet and C harlero i fo rm ations, only these tw o units are discussed. T he sed im en ts o f the C hâtelet Fm. are com posed o f non-m arine shales, sandstones, th in coal seam s and roo tle t beds. Tw o pervasive m arine horizons d iv ide the C hâte le t Fm. into tw o m em bers: the Sam sbank m arine band at the base o f the C hâtelet Fm. form s the base o f the R ansart M br., w hile the F inefrau N ebenbank band defines the base o f the over- lying F loriffoux M br. T he C hâte le t Fm . attains a th ick ness o f 500 m in the C am pine area. T he succeeding C harlero i Fm . con tains several th ick coal seam s, and consists o f a very characteristic rhy thm ic succession o f coa l-m udstone-sandstone sed im en tary sequences. Faint m arine bands subdiv ide the C harlero i Fm. into three m em bers: the basal M ons M br., the A s M br. and the E ikenberg M br. The C harlero i Fm. reaches a th ickness o f over 1000 m in the northeastern C am pine basin.
S tratigraph ica l assignm en t and age o f the specim en
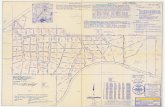
T he eurypterid fossil w as recovered from a dep th o f 435 m, from the core o f boring no. 32, carried out in the southern C am pine coalfie ld near M echelen-aan-de-M aas, w hich now adays is a borough o f M aasm echelen (Fig. 1).

Adelophthalmus from B elg ium 81
North
Campine
AntwerpB ruges
MAASMECHELEN
4-
Ghent
Hasselt
MaastrichtAachen
Namur
MonsCharleroi
ArionLuxem bourg
Fig. 1
0 10 20 30 40 50 60 70 80 90 100 km 1___I___I___I___I___I___I___ I___I___I___I
Map o f Belgium showing the geographic extent o f the Belgian Coal Measures Group. The position o f Maasmechelen, o f which Mechelen-aan-de-Maas nowadays is part, is indicated b y the cross-hairs. Adapted from R e n ie r 1 9 3 0 and P a p r o t h etal. 1983.
In his orig inal descrip tion , S t a i n i e r (1917) stated that the fossil w as found in a grey, fine-grained sandstone co n ta in ing scattered p lan t fragm ents, above a thin seam o f coal, about m idw ay through the “ great barren m easu re” ( “ grande stam pe s té rile” ) and approxim ately 5 7 0 m above the “ M illstone G rit” ( “ poud ingue” ). T he “ great barren m easu re” w as the upper subdiv ision o f the Low er W estphalian A “ assise de C h â te le t” ( S t a i n i e r 1911). T he “ M illstone G rit” w as considered by S t a i n i e r
(1911, 1917) to be a single, w idespread grit-level form ing the boundary betw een the N am urian and W estphalian stages. R e n ie r (1930), how ever, show ed there w as no such th ing as a single, con tinuous “ M illstone G rit” reference stratum , and subdivided the “ assise de C hâte- le t” into the low er “ zone d ’O u p ey e” and the upper “ zone de B eyne” . T he eurypterid described herein w as d iscovered in the “ zone de B eyne” , as recognised by P r u v o s t (1930). P a p r o t h et al. (1983) form ally estab lished the “ zone d ’O u p ey e” as the R ansart M br., and the “ zone the B ey n e” as the F loriffoux M br.. In the latest
rev iew o f B elg ian C arboniferous stratig raphy , D e l m e r et al. (2001), recogn izing the “ assise de C h â te le t” , erected the C hâtelet Fm. to contain the R ansart and F loriffoux m em bers. A ccord ing ly , the eurypterid com es from approxim ately the m iddle o f the F loriffoux M br. o f the C hâtelet Fm . in the B elgian C oal M easures G roup, and is o f early Late B ashkirian age (Fig. 2).
M aterial and m ethods
S t a i n i e r (1917) gave no repository details for the specimen he described. The single specimen o f A. dumonti ( S t a i n i e r , 1917) is now held at the Royal Belgian Institute o f Natural Sciences in Brussels, Belgium registered under number RBINS a7706. It consists o f part and counterpart; the part being relatively complete, lacking some appendages, pretelson and telson, while the counterpart is less complete, consisting o f only the carapace, partial appendages and six anterior segments. The counterpart has been repaired after breakage during original splitting o f the

82 O. Erik T E T LIE & Peter VAN ROY
(Upper) (Kasimovian)W estphalian D Neeroe teren Fm
Neerglabbeek MbrMiddle Moscovian Wasmes MbrFlenu FmW estphalian C
Meeuwen Mbr
Eikenberg MbrW estphalian B
As MbrCharleroi Fm
Mons MbrlEQM.ei B a s In ki i#i nio
i V o s t p I i . i l i a n A FI o i if f o l i X M b i 1C h n t e l e t F m
Ransart Mbr
Namurian C
A ndenne FmNamurian B
Upper Serpukhovian
Chokier FmNamurian A
Souvre Fm
Middle Visean HOYOUX GROUP
Fig. 2 — Stratigraphy o f the late Carboniferous Belgian Coal Measures Group in the Campine Basin. Ages and units relevant to A. dumonti are indicated in dark grey. Units containing other eurypterids, and that do not overlap with the occurrence of A. dumonti are in light grey. Left column shows the internationally agreed chronostratigraphical division o f the late Carboniferous. Late Pennsylvanian deposits are not represented in Belgium. Therefore, the Kasimovian is put in brackets, and the Gzhelian is omitted. Middle column shows the traditional late Carboniferous time-scale as still often used in Belgium. The Stephanian is omitted because no deposits o f this age are known in Belgium. Right column shows the lithostratigraphic division o f the late Carboniferous in the Campine Basin. Italic numbers on the right hand side o f the column denote the approximate stratigraphical distribution o f Belgian Carboniferous eurypterids: 1. A. dumonti', 2. A. corneti', 3. A. moyseyi; 4. M. mathieur, 5. A. combieri. Lithostratigraphic division adapted from D e l m e r et al. 2001.
core sample. The original orientation of the core sample is not indicated, so the way-up of the eurypterid is not known. The bedding planes intersect the core at an angle of around 70°, suggesting the beds originally had a dip o f around 20° if the core was drilled vertically. Photographs were taken using a Nikon D 100 digital SLR camera. The part was photographed initially under alcohol and subsequently with a coating of ammonium chloride, while the counterpart was photographed dry (the old glue was too brittle and fragile for alcohol) and coated with ammonium chloride. Camera lucida drawings were made using a Wild stereoscope with a drawing tube attachment. Morphological terminology follows T o l l e r t o n (1989) and higher systematics follows T e t l i e (2004). The following abbreviations are used in the text and figures: c = carapace, ch = chelicerae, ex = coxa, db = dumb-bells, e = eye, gl = genal lobe, o = ocelli, pi = plant fragment, s = spine, t = raised triangle; prosomal appendages are numbered with Roman numerals I-VI, individual podomeres in prosomal appendages 1-9 (1 is the coxa) and opisthosomal segments 1-11 (pretelson and telson not preserved). In the text, short denotation of individual podomeres of appendages is made by combining appendage numbers with podomere numbers, e.g. II1-7. All reference to left and right in the text refer to the more complete part unless otherwise stated.
S ystem atic palaeon to logy
O rder E urypterida B u r m e i s t e r , 1843 S uperfam ily A d elop hthalm oid ea superfatn . nov. D iagnosis: Sm all sw im m ing eurypterids w ith very gen eralised appearance; carapace parabo lic (possib le exception Unionopterus) w ith in tram arg inal eyes; sw im m ing leg o f Adelophthalmus type; genital spatu lae possib ly presen t in all taxa; m idsection second o rder d iffe ren tia tion present; telson lanceolate.Rem arks: T e t l i e (2004) in form ally raised th is superfam ily based on h is cladogram , and encom passing the sam e genera as suggested here. T e t l i e (2004) suggested that th is superfam ily could be d iv ided into the basal N anahughm illeridae (Nanahughmilleria K j e l l e s v i g -
W a e r i n g , 1961 and possibly PittsJ'ordipterus K j e l l e s v i g -
W a e r i n g & L e u t z e , 1966) with appendages 1I-V o f Hugh- milleria type, and the m ore derived A delophthalm idae (Parahughmilleria K j e l l e s v i g - W a e r i n g , 1961 and Adelophthalmus) w ith reduced spinosity o f the appendages. The enigm atic Unionopterus m ight represent the only post-Silurian occurrence o f the N anahughm illeridae if the

Adelophthalmus from B elg ium 83
appendage spinosity indicated by C h e r n y s h e v (1948) w as correct.
Fam ily A delop hthalm id ae T o l l e r t o n , 1989 R em arks: T o l l e r t o n (1989) raised this fam ily for Parahughmilleria, Bassipterus K j e l l e s v i g - W a e r i n g & L e u t z e , 1966, Adelophthalmus and Unionopterus, based on hav ing Adelophthalmus types o f sp in iferous (II-V ) and sw im m ing (V I) appendages respectively . T e t l i e (2004) in terpreted Bassipterus virginicus K j e l l e s v i g - W a e r i n g
& L e u t z e , 1966, the on ly know n species o f Bassipterus, as a ju n io r synonym o f Parahughmilleria bellistriata ( K j e l l e s v i g - W a e r i n g , 1950). As m entioned above, Unionopterus w as described as hav ing appendages II-V o f Hughmilleria type although its stratigraphical position w ould be m ore consisten t w ith an iden tity w ith in the A delophthalm idae. T o l l e r t o n (1989) claim ed that species w ithin Adelophthalmus w ith com pletely non-spi- n iferous appendages w ould constitu te a new genus o f a new fam ily w ith in the S lim onioidea. A new genus m ight be w arran ted , but to erect a new fam ily , and transferring it into the S lim onio idea based on a character loss, w hich c learly has happened independently in the tw o clades in question ( T e t l i e 2004), cannot be defended from a phylogenetic po in t o f view .
G enus Adelophthalmus v o n M e y e r , 1 8 5 3 D iagnosis: M edium sized stream lined eurypterid ; ca rapace parabolic w ith narrow m arginal rim and sm all, h inged triangu lar “ lock in g ” m echanism anteriorly ; in tram arg inal ren iform eyes; ocelli betw een o r slightly behind eyes; prosom al appendages II-V Adelophthalmus type; VI sw im m ing leg o f Adelophthalmus type; m etastom a oval; first op isthosom al segm ent o f reduced length and tapering in length laterally; m idsection (and usually an terio r and posterio r) second o rder opisthosom al d iffe ren tiation ; genital opercu lum w ith spatulae; te lson long and sty liform ; dense cu ticu lar scu lpture o f m inute scales (em ended from T e t l i e & D u n l o p 2 0 0 5 ) .
Adelophthalmus dumonti ( S t a i n i e r , 1 9 1 7 )
Figs. 3-5
Eurypterus sp .; S c h m itz & S t a i n i e r 1910, p p . 293, 296.V*Eurypterus dumonti', S t a i n i e r 1917, p. 646, p i. 53, fig s . 1-4. Eurypterus Dumonti S t a i n i e r ; P r u v o s t 1930, p . 191. Eurypterus dumonti S t a i n e r [s ic ] ; M o o r e 1936, p . 371. Eurypterus! dumonti S t a i n i e r ; K je l l e s v ig - W a e r i n g 1948, p. 6. Adelophthalmus dumonti S t a i n i e r ; V a n O y e n 1956, p . 49. Adelophthalmus derhiensis W o o d w a r d ; V a n O y e n 1956, p. 61. Adelophthalmus dumonti', P l o t n i c k 1983, p . 385. Unionopterus dumonti', T e t l i e 2004, p p . 183, 250, 283. Unionopterus dumonti', T e t l i e & D u n lo p 2005, p. 6.
D iagnosis: Sm all Adelophthalmus w ith th ickened bands d ista lly on podom eres o f w alk ing legs; genal lobes on carapace w hich articu la te la terally w ith second tergite, and correspond ing ly la terally reduced first terg ite; an terio r and m idsection , bu t lack ing posterio r second order
opisthosom al d ifferen tia tion ; raised triang le on op isthosom al segm ent 7.Type locality: M echelen-aan-de-M aas (= M aasm echelen), L im burg, B elgium .Type horizon: A bove a th in coal seam , approxim ately in the m iddle o f the F loriffoux M br. o f the C hâte let Fm ., in the B elgian C oal M easures G roup.Rem arks: T he susp icion that this species shou ld be assigned to the poorly know n Unionopterus ( T e t l i e &
D u n l o p 2005) w as m ain ly based on the carapace shape and the b road m arginal carapace rim figured by S t a i n i e r
(1917). A s dem onstrated herein , the ind icated shape w as in fluenced by an incom plete an terio r m arg in and w as not the true carapace shape, w hile the m arginal rim is narrow as in o ther species o f Adelophthalmus. A lso note that we do not acknow ledge the synonym y o f A. dumonti w ith A. derbiensis W o o d w a r d , 1907 as suggested by V a n O y e n (1956). A. derbiensis its e lf w as synonym ized w ith A. moyseyi by K j e l l e s v i g - W a e r i n g (1948), som eth ing w e support, bu t w e also no te that A. m oyseyi and A. dumonti are very sim ilar (see below ).
D escription
A lm ost com plete specim en w ith p reserved length 32.5 m m , m axim um opisthosom al w id th 11.5 m m . P reserved part o f carapace 7.5 m m long, 9.7 m m w ide at posterio r m argin. A n terio r and left carapace m argins are incom plete (F igs 3A , 4A). Lateral angle 95°, carapace L /W ratio restored to approx im ate ly 0.79, g iv ing a parabolic shape ( T o l l e r t o n 1989). C arapace w ith very narrow (0.1 - 0.2 m m w ide) m arginal rim on lateral m argins. R ight eye w ell-preserved and ren iform , 1.2 m m long. The ocelli are positioned betw een the eyes on the counterpart (Fig. 4B ). O n the right postero -la tera l carapace co m er o f the part (Fig. 4A ), a large rounded genal lobe projects posteriorly . A curious card io id (heart-shaped) depression (Fig. 4A ) is positioned betw een the eyes and the an terio r carapace m argin; in terpreted as rep resen ting very unusual p reserva tion o f the chelicerae (see below ). Four partial podom eres o f left appendage II m ay possib ly be observed. Poorly p reserved rem ains o f appendage II extend past the carapace m argin on the righ t and th is appendage ev idently did not ex tend far beyond the m argin . N o podom ere details can be seen on th is appendage. A ppendage III is p reserved on both sides. O n the right, podom eres III-4 to III-7 (fo r podom ere abbrev iations, see Fig. 4C ) are probably preserved , although podom ere boundaries are no t seen (Fig. 4B ). O n the left, som e o f the carapace has been broken aw ay (F igs 3A , 4A ), ex posing m ore o f the proxim al podom eres, and eigh t podom eres (coxa to III-8) can be seen (Fig. 4C ). A ppendageIV is best preserved on the left w here coxa to IV-8 can be seen; on the righ t on ly a fragm ent is p resent. A ppendageV is on ly p reserved on the left, and seven podom eres (V -2 to V -8) are preserved. V -7 has the proxim al end o f a very large spine preserved. The podom eres do not have cre- nu la ted d istal m arg ins, but a th ickened distal m argin is

84 O. Erik T E T LIE & Peter VAN ROY
Fig. 3 — Adelophthalmus dumonti ( S t a i n i e r , 1917). A) Photograph o f RBINS a7706 (part) submersed in alcohol; B) Photograph o f RBINS a7706 (part) coated with ammonium chloride; C) Photograph o f RBINS a7706 (counterpart) taken under normal conditions; D) Photograph o f RBINS a7706 (counterpart) coated with ammonium chloride. Scale bars are 5.0 mm.
observed on several podom eres on appendages IV and V. The podom ere lengths (parallel to longest axis o f appendage) and w id ths (perpend icu lar to longest axis o f appendage) in m m o f appendages III-V on the left side are: III-2 0 .5 /1 .2*; III-3 0 .8 /1 .4*; III-4 0 .8 /1 .3*; II 1-50.9/1.1 III-6 0 .7 /1 .0*; III-7 0 .7 /0 .6*; III-8 0 .9*/0 .4*; IV-2 0 .6 /1 .5 ; IV- 3 1.1/1.3; IV-4 1.0/1.3; IV-5 1.0/1.2; IV-6 0.9/1.0; IV-7 0 .9 /0 .8 / IV-8 0 .3*/0 .4 ; V -2 0 .6*/1 .4*; V-3 1.0/1.5; V-4
1.4/1.5; V -5 1.8/1.3; V -6 2.0 /1 .0 ; V -7 2.1 /0 .8 V -8 0 .9*/ 0.5 (* denotes m easurem ent o f incom ple te podom eres).
P osterio r to the eyes, on the carapace and ex tend ing onto the an terio r opisthosom al segm ent, tw o depressions m im ic the ou tline o f the posterio r parts o f coxae VI. A ppendage VI also has podom eres 7, 7a and 8 preserved on the left side, and a m inute no tch in V I-8 suggest the position o f V I-9. P roxim ally , the podom eres betw een the

Adelophthalmus from B elg ium 85
Fig. 4 — Adelophthalmus dumonti ( S t a in i e r , 1917). A ) Camera lucida drawing o f RBINS a7706 (part); B) Camera lucida drawing o f RBINS a7706 (counterpart); C ) details o f the appendages exposed on the left side of the part. Scale bars are 5.0 mm.
coxa and V I-7 are p resent, but partly covered by terg ites and the ir outlines cannot be identified. The posterio r m argin o f the m etastom a is in terpreted as rela tively straigh t o r truncated .
T he an terio r opisthosom al segm ent is strongly reduced in length com pared to the m ore posterio r ones, and is surrounded by genal lobes, a lm ost g iv ing the appearance o f being incorporated into the carapace. T he postero lateral co m er (genal lobe) o f the carapace surrounds the entire an terio r segm ent, so the carapace articu la tes w ith the second opisthosom al segm ent laterally . The second segm ent lacks lateral ep im era ju s t like the an terio r segm ent. The fo llow ing segm ents becom e w ider until the op isthosom a reaches its m axim um w idth on the fourth segm ent. G radually , these segm ents develop longer ep im era un til those o f the seventh segm ent. O n the seventh
segm ent, a m edian raised area form s a triang le w ith its apex poin ting tow ards the carapace. T here is a m oderate first o rder op isthosom al d ifferen tia tion ( T o l l e r t o n
1989). The four an terior-m ost segm ents (8-11) represen t the incom plete postabdom en. A s in m ost eurypterids, these segm ents are narrow er, but longer than the segm ents in the preabdom en. T he segm ents in the postabdom en have no ep im era preserved. The p reserved segm ent lengths (along the m id line) and w id ths in m m are: 1) 0 .6/ 9.3*; 2) 1.3/11.0*; 3) 1.8/10.9*; 4) 3 .0/11.5 ; 5) 1.7/11.2; 6) 1.4/10.4; 7) 1.9/9.5; 8) 3.9/Ó.6; 9) 3 .4 /5 .4 ; 10) 3 .7/5.2; 11) 3 .2*/4 .2 (* denotes m easurem ent o f an incom plete segm ent). The prete lson and the te lson are not preserved as they w ere ou tside o f the area covered by the core, but com parison w ith o ther species o f Adelophthalmus (e.g. A. imhofi R e u s s , 1855) suggest tha t this species p robably

86 O. Erik T E T LIE & Peter V AN ROY
m m ü r n ,
2* Ir.-;::*-
g
WiïMâÈ&iëm i fi * I i >• î
■■ y : / . p v t ô A / î i v :-î
M )
ïâièiifcï(iîto'Æ$!i; '
Fig. 5 — Dorsal reconstruction o f Adelophthalmus dumonti ( S t a i n i e r , 1917) based on RBINS a7706 and a pretelson and telson based on the presumably closely related Adelophthalmus imhofi.
also had a rela tively long, lanceolate te lson and w e have reconstructed it as such (Fig. 5).
should actua lly look like. It is therefore im prudent to p lace too m uch em phasis on illustrations w ithout re-ex am ining the fossil. As prev iously m entioned , S t a i n i e r ’s (1917) descrip tion is defective on m any counts. First, the an terio r carapace m argin w as in terpreted as com plete by S t a i n i e r (1917). This is certa in ly not the case, and the ja g g ed an terio r and left-hand m arg ins suggest that the anim al m ight have been scavenged p rio r to fossilisation . F rom the partially com plete righ t-hand m argin , it is c lear that the carapace w as m ore or less parabolic , as in o ther species o f Adelophthalmus, not trapezoid as suggested by S t a i n i e r (1917).
Second, as the carcase decom posed , sm all (0.1 - 0.2 m m ) crystal bundles, o r “ dum b -b ells” ( B r ig g s &
W il b y 1996) w ere form ed along the lateral m argins, the oral cav ity and an terio r parts o f the d igestive system . ‘D um b-bells’ are very early d iagenetic features, starting to form three days afte r death in shrim p carcases ( B r ig g s
& K h a r 1994). They are orig inally com posed o f arago nite , bu t aragon ite is less stable than ca lc ite and all ‘d um b-bells’ found in fossils have been recrysta llised to ca lc ite ( B r ig g s & K e a r 1994). W e have not analysed the com position o f the suspected ‘d um b -b ells’ in th is fossil. T hey are com m only found in associa tion w ith phospha- tization , but no th ing resem bling phosphatized m uscle tissue w as identified in the B elgian eurypterid . These ‘dum b-bells’ (F igure 6) w ere iden tified as o rnam entation by S t a i n i e r (1917). N o ‘d um b -b ells’ are seen on the postabdom en, w hich had th icker, annulate segm ents, a condition not favourab le fo r aragonite p recip ita tion as pH w ould becom e too low fo llow ing decom position o f the locally m ore abundant o rganic m ateria l ( B r ig g s & W il b y
1996). ‘D um b-bells’ are m ost com m only developed in restric ted m arine conditions, but have also been found prev iously in specim ens from a non-m arine (lacustrine) se tting at Las H oyas, Spain ( B r ig g s & W il b y 1996). The B elgian C arbon iferous occurrence is the o ldest exam ple o f ‘d um b-bells’ from a non-m arine locality . T he spec im en has ornam entation , seen on the coun terpart consisting o f fine scales as typical for the genus. T his w as not m entioned by S t a i n i e r (1917) w ho largely ignored the relevance o f the counterpart, sta ting that: “ the coun terpart o f the fossil is fractured and incom plete, and th e re fore o f little use for palaeontological pu rp o ses” ( S t a i n i e r
1917, p. 639).
T aphonom y
A num ber o f post-m ortem changes have affected this eurypterid , w hich m isled S t a i n i e r in his o rig inal descrip tion. T he iden tification o f these is a stark rem inder that taphonom ical processes w ere not understood by m ost earlier au thors and o lder publications are therefore occasionally inaccurate in the in form ation they portray. Illu strations are som etim es idealised , to the ex ten t that, rather than being a reliab le depiction o f the specim en, they reflect the au th o r’s personal v iew s on w hat the fossil
Fig. 6 — Dumb bells on right side, third and fourth segments of Adelophthalmus dumonti ( S t a in i e r , 1 917) holotype, RBINS a 7706.

Adelophthalmus from B elg ium 87
W hen the dorsal cu ticle o f the eurypterid becam e m alleable, the ou tline o f the underly ing coxae V I and m etastom a becam e v isib le th rough the carapace (Fig. 4A ), as in terpreted correctly by S t a i n i e r ( 1 9 1 7 ) , although fig ured too sym m etrically by him . W hen the d rill core w as split, another anom aly w as created. The ven tra l chelicer- ae fo llow ed the dorsal-m ost incom plete counterpart, creating a large card io id depression in the part. S t a i n i e r
(1917) acknow ledged that this w as possib ly “ accidenta l” , but failed to realise its true nature. T he size o f the depression suggests the chelicerae , like the o ther appendages, w ere rela tively robust, som eth ing also seen in o ther species o f Adelophthalmus (see W i l l s 1964).
D iscussion
T he m ain differences betw een the initial descrip tion o f S t a i n i e r (1917) and our w ork are: 1) the carapace shape; 2) the m arginal rim o f the carapace; 3) the ocelli that can be observed on the counterpart; 4) the spine on appendage V ; 5) the nature o f the c ircu lar nodes, and finally 6) the typical scaly scu lpture that can be found localised on the counterpart. A dditional m inor d ifferences include the accuracy o f the prosom al appendages includ ing the coxae o f appendage VI, the accuracy o f the ju n c tio n betw een the carapace, segm ents 1 and 2, the broken ou tline o f the segm ents o f the left side o f the part, and the lack o f ep im era on the postabdom en.
T he partial destruction o f the left side o f the carapace w as a fortu itous accident. T here has long been uncertain ty about the num ber o f appendage podom eres (here given in th is form : num ber o f podom eres in appendage I- II-III-IV -V -V I respectively) in Adelophthalmus. W i l l s
(1964, p. 486) suggested that Adelophthalmus had 7-67- 7-87-9-9, although he sta ted “ it is no t possib le to be certain about the exact num ber o f jo in ts in ea ch ” . A m ong eurypterids, th is po ten tia l podom ere count could on ly be m atched in Stylonurella (7-7-77-8-9-9?) ( P l o t n i c k
1983). This specim en, w ith w ell exposed in situ appendages, suggests there are 8, not 7 podom eres in appendage III and th is im plies fo r Adelophthalmus the sam e podom ere count (i.e. 3 -7-8-8-9-9) as m ost o ther eu ryp terids, a lthough there are still uncertain ties about the very sm all appendage II in the genus. The spine on V-P7 suggests A. dumonti had a very sim ilar appendage to the one described by S c o t t ( 1971 ) for A. mansfieldi, also figured as the Adelophthalmus type o f sp in iferous appendage by T o l l e r t o n (1989).
The only puzzling m orphology described by S t a i n i e r (1917) that cannot be assigned to taphonom ical p rocesses is the raised triangular area on the m ost posterio r p reabdom inal segm ent. This is a very regular structure although its an terio r sym m etry has been slightly d istorted by a depression continu ing onto the segm ent an terio r to the triangle. W hether this triang le had a hydrodynam ic or ano ther function is not read ily obvious. It could alternatively have been som e kind o f ‘b o ss’ fo r articulation o f the pre- and postabdom en (L. A nderson pers. com m . 2005).
T he posterio r m arg in o f the m etastom a seen th rough the carapace m ight represen t a rare m etastom a shape in Adelophthalmus, as m ost species have an oval m etastom a w ith a rounded posterio r m argin. H ow ever, since the p osterio r m argin in the specim en o f A. dumonti is very poorly exposed, the possib ility cannot be excluded that th is aberran t shape in fact is a taphonom ical artefact.
L ineages o f Adelophthalmus
A lthough this w ork does not attem pt to reso lve the phy- logeny o f the adelophthalm ids - for that our know ledge is still too lim ited - som e lineages can be iden tified since they are defined by un ique synapom orphies. O ne po tential strong synapom orphy o f certain species is the p re sence o f a raised triang le on segm ent 7. T his structure is p rev iously know n from A. imhofi ( R e u s s , 1855) as po in ted out by S t a i n i e r (1917) and A. moyseyi as no ted by M o o r e (1936, p. 371) in add ition to A. dumonti. The la tte r tw o are o f B ashkirian age, w h ile A. imhofi is G zhelian (Late C arboniferous) or A sselian (E arly Perm ian). T he triang le is not p resen t in A. lucernensis, A. sellardsi, A. granosus and A. sievertsi and probably not presen t in A. mansfieldi and A. mazonensis.
The differen tia tion o f the an terio r terg ite m igh t also be o f phylogenetic im portance in adelophthalm ids. T he sim plest type o f d ifferen tia tion o f th is segm ent is sim ply to have reduced segm ent length. This is found in A. wilsoni (see M o o r e 1936). A second type is the gradual reduction in segm ent leng th laterally found in A. sievertsi (see P o s c h m a n n in press), possib ly A. imhofi (see R e u s s
1855 or K j e l l e s v i g - W a e r i n g 1948) and A. lucernensis ( K u e s & K i e t z k e 1981). A th ird type is the one found in A. dumonti and possib ly A. moyseyi (see ind ication o f this in M o o r e 1936, fig. 63 both left and righ t sides), w here the an terio r segm ent is obscured laterally by the expanded postero latera l com ers o f the carapace tha t articu la te w ith the second segm ent laterally . The im portance o f this character is still uncertain , especially since the three species w ith a triang le on segm ent 7 appear to fall into tw o groups. A second possib le lineage is defined by having epim era on the postabdom en; these are A. mansfieldi, A. wilsoni and A. mazonensis. The la tter tw o o f these are M oscovian in age w hile A. mansfieldi is W estphalian . H ow ever, m ost adelophthalm ids canno t be assigned to any m ajo r lineage.
Is Unionopterus valid, and if so, is it an adelophthalm id?
O ne could agree unquestionab ly w ith C a s t e r & K j e l l e s v i g - W a e r i n g (1964) and T o l l e r t o n (1989) and in terpret Unionopterus anastasiae as an ade loph tha lm id. H ow ever, based only on the illustrations prov ided by C h e r n y s h e v (1948), there is scant ev idence to suggest that th is genus belongs to the adeloph tha lm ids o ther than the fact that Adelophthalmus and Unionopterus are the only know n sw im m ing eurypterids to surv ive into the

88 O. Erik T E T LIE & Peter V A N ROY
C arboniferous, and the ir sw im m ing legs are o f a sim ilar m orphology. B efore the specim en o f A. dumonti w as re studied , an assignm ent to Unionopterus seem ed likely based on sim ilarities betw een the illustrations o f S t a i n i e r (1917) and C h e r n y s h e v (1948). H ow ever, can m ore un ique synapom orphies for th is p roposed rela tionsh ip be found even w hen A. dumonti is no t a Umonopterus!
Adelophthalmus can be fairly convincing ly in terpreted as being closely re la ted to the S ilurian - D evonian genera Nanahughmilleria K j e l l e s v i g - W a e r i n g 1961, Parahughmilleria K j e l l e s v i g - W a e r i n g , 1961 and the m ono- typic Pittsfor dipterus K j e l l e s v i g - W a e r i n g & L e u t z e 1966. Pittsfordipterus is indistinguishable from Nanahughmilleria excep t for the m orphology o f its genital appendage, and is treated as a nanahughm illerid for the rem ainder o f th is d iscussion. T heir alm ost identical carapace, paddle, eye shapes and eye position support the rela tionsh ip betw een Adelophthalmus, Nanahughmilleria and Parahughmilleria in the new superfam ily A de- lophthalm oidea. M ore explicit pu ta tive synapom orphies fo r these genera are the p resence o f ep im era on segm ent 7 (= m idsection second o rder d ifferen tia tion ; T o l l e r t o n 1989) and genital spatulae, although the spatulae are very large in Adelophthalmus, m edium -sized in Parahughmilleria and very sm all in Nanahughmilleria ( S t o r m e r
1973; T e t l i e 2000). T he w alk ing legs in Nanahughmilleria p robab ly had paired spines on each podom ere ( S t o r m e r 1934; T e t l i e 2000), a condition seen for instance in Hughmilleria ( S a r l e 1903), w hile Parahughmilleria and Adelophthalmus have appendage podom eres w ith occasional iso lated sp ines and crenulated d istal p o dom ere m argins. A utapom orphies for Adelophthalmus (absen t in Nanahughmilleria and Parahughmilleria) in clude: (1) ep im era on the p re- and postabdom en, and (2) a triangular, h inged ‘lock ing ’ m echanism anterio rly on the carapace; although the p resence o f these has not been dem onstrated in all species o f Adelophthalmus. T he cuticu lar scu lpture is m uch coarser in Adelophthalmus than in Nanahughmilleria and Parahughmilleria and the form er lacks the ornam ent o f lines parallel to the segm ent m arg ins found in the la tte r tw o (see e.g. S t ü r m e r 1973). T he te lson is generally longer in Adelophthalmus than in Nanahughmilleria and Parahughmilleria, bu t the consistency o f th is character is equivocal. A final d ifference betw een Adelophthalmus versus Nanahughmilleria and
References
B r ig g s , D.E.G. & K e a r , A.J„ 1994. Decay and mineralization of shrimps. Palaios, 9 (5): 431-456.
B r ig g s , D.E.G. & W ilb y , P.R., 1996. The role o f the calcium carbonate-calcium phosphate switch in the mineralization of soft-bodied fossils. Journal o f the Geological Society, 153: 665- 668 .
B u r m e i s t e r , H., 1843 . Die Organisation der Trilobiten, aus ihren lebenden Verwandten entwickelt; nebst einer systematischen Uebersicht aller zeither beschriebenen Arten. G. Reimer, Berlin, 148 pp .
Parahughmilleria is in the m orphology o f the an terio r op isthosom al segm ent. This tapers in length tow ards the lateral m arg ins in at least som e species o f Adelophthalmus ( T e t l i e & D u n l o p 2005; see also A. dumonti), a condition not know n in any Nanahughmilleria o r Parahughmilleria. T hus, the S ilurian Nanahughmilleria and Pittsfordipterus, w ith paired spines on the appendages and none o r sm all genital spatu lae, w ere basal to the p redom inan tly D evonian Parahughmilleria, w ith reduced appendage sp inosity and enlarged spatulae. This trend is fu rther continued in the D evonian to P erm ian Adelophthalmus, w hich have also developed a triangu lar carapace “ lock in g ” m echanism , a reduced an terio r terg ite , ep im era on the pre- and (occasionally ) postabdom en and a longer telson.
So w here does this leave us in respect to the phy logenetic position o f Unionopterus? T he trapezo id carapace shape is d ifferen t to all o the r taxa in the lineage, w hich retain a parabo lic shape. Unionopterus does share som e sim ilarities to Adelophthalmus, such as ep im era on the preabdom en, and it is therefo re possib le that Unionopterus is a sister-taxon to, o r in -group of, Adelophthalmus. A lternatively , the p resence o f paired spines on the appen dages suggests that Unionopterus is the only know n post- S ilurian surv ivor o f Nanahughmilleria-Pittsfordipterus affinity .
A th ird alternative is that C h e r n y s h e v (1948), like S t a i n i e r (1917), m isin terp reted the carapace shape and appendage spinosity , and that he too w as describ ing a specim en belonging to the genus Adelophthalmus. But since the w hereabouts o f the ho lo type and on ly specim en o f U. anastasiae are p resen tly not know n, this question m ay never be answ ered.
A ck now ledgm en ts
We thank Annie D h o n d t (Brussels), for help in locating the specimen and Derek E. G. B r ig g s (Yale University, New Haven) for discussions regarding the “ dumb-bell” structures. The photographs were prepared by W ilfried M is e u r (Royal Belgian Institute o f Natural Sciences in Brussels). The manuscript was reviewed and much improved by Lyall I. A n d e r s o n (Edinburgh), Simon J. B r a d d y (Bristol), Jason A. D u n l o p (Berlin) and Rachel A. M o o r e (University o f Kansas). OET thanks Norges Forskningsrâd for financial support (grants 145565/432 and 166647/V30) into eurypterid phylogeny and palaeoecology.
C a s t e r , K.E. & K j e l l e s v ig - W a e r i n g , E.N., 1964. Upper Ordovician eurypterids from Ohio. Palaeontographica Americana, 4 (32): 300-358.C h e r n y s h e v , B .F ., 1948. New representative o f Merostomata from the Lower Carboniferous. Kievski Gos. Universitet im T. G. Shevchenko Geologisheskii Shornik, 2: 119-130. [In Russian].D e lm e r , A., D u s a r , M. & D e l c a m b r e , B., 2001. Upper Carboniferous lithostratigraphic units (Belgium). In: P. B u l t y n c k & L. D e jo n g h e (eds), Lithostratigraphic scale o f Belgium. Geológica Belgica, 4 (1-2): 95-103.

Adelophthalmus from B elg ium 89
D r e e s e n , R., B o s s i r o y , D., D u s a r , M., F l o r e s , R.M. & V e r k a e r e n , P., 1995. Overview o f the influence o f syn-sedi- mentary tectonics and palaeo-fluvial systems on coal seam and sandbody characteristics in the Westphalian C strata, Campine Basin, Belgium. In: M.K.G. W h a t e l e y & D.A. S p e a r s (eds), European Coal Geology. Geological Society Special Publication, 82: 215-232.
F r a i p o n t , J., 1889. Euryptérides nouveaux du Dévonien supérieur de Belgique (Psammites du Condroz). Annales de la Société géologique de Belgique, 17: 53-62.K j e l l e s v ig - W a e r i n g , E.N., 1948. The Mazon Creek eurypterid: A revision o f the genus Lepidoderma. In: Coal age fossils from Mazon Creek. Illinois State University Science Papers, 3(4): 48 pp.
K j e l l e s v ig - W a e r i n g , E.N., 1950. A new Silurian Hugh- milleria from West Virginia. Journal o f Paleontology, 24 (2): 226-228.
K j e l l e s v ig - W a e r i n g , E.N., 1958. The genera, species and subspecies o f the Family Eurypteridae, Burmeister 1854. Journal o f Paleontology, 32 (6): 1107-1148.K j e l l e s v ig - W a e r i n g , E.N., 1961. The Silurian Eurypterida of the Welsh Borderlands. Journal o f Paleontology, 35 (4): 789- 835.K j e l l e s v ig - W a e r i n g , E.N., 1963. Pennsylvanian invertebrates o f the Mazon Creek area, Illinois, Eurypterida. Fieldiana, Geology, 12: 85-106.K j e l l e s v ig - W a e r i n g , E.N. & L e u t z e , W.P., 1966. Eurypterids from the Silurian o f West Virginia. Journal o f Paleontology, 40(5): 1109-1122.
K u e s , B .S . & K ie tz k e , K .K ., 1981. A large assemblage o f a new eurypterid from the Red Tanks Member, Madera Formation (Late Pennsylvanian-Early Permian) o f New Mexico. Journal o f Paleontology, 55 (4): 709-729.
L a n g e n a e k e r , V. & D u s a r , M., 1992. Subsurface facies analysis o f the Namurian and earliest Westphalian in the western part o f the Campine Basin (N Belgium). Geologie en Mijnbouw, 71: 161-172.M o o r e , L.R., 1936. Some eurypterids from the English coal measures. Proceedings o f the Geologists' Association, 47 (4): 352-375.N o v o j i l o v , N.I., 1962. Otrjad Eurypterida. In: B.B. Rodendorf (ed.) Osnovy Paleontologii. Chlenistonogie, trakhejiye i khelit- serovye, Moscow: Izdatel’stvo Akademii Nauk SSSR, pp. 404- 423. [In Russian],P a p r o t h , E., D u s a r , M., B l e s s , M.J.M., B o u c k a e r t , J., D e lm e r , A., F a i r o n - D e m a r e t , M., H o u l l e b e r g h s , E., LALOUX, M., PlERART, P ., SOMERS, Y ., STREEL, M., THOREZ, J. & T r i c o t , J., 1983. Bio- and lithostratigraphic subdivisions of the Silesian in Belgium, a review. Annales de la Société géologique de Belgique, 106 (3-4): 241-283.
P l o t n i c k , R.E., 1983. Patterns in the evolution o f the eurypterids. Unpublished PhD thesis, The University o f Chicago, Chicago, 411 pp.P o s c h m a n n , M ., in press. On the Emsian (Early Devonian) arthropods o f the Rhenish slate mountains: 5. The eurypterid Adelophthalmus sievertsi from Willwerath, Germany. Palaeontology.P r u v o s t , P ., 1924. Un Euryptéride nouveau du Terrain houiller de Charleroi. Annales de la Société géologique du Nord, 48: 143-151.
P r u v o s t , P., 1930. La faune continentale du terrain houiller de la Belgique. Mémoires du Musée royal d'Histoire naturelle de Belgique, 44: 191-196.P r u v o s t , P., 1939. Eurypterus (Anthraconectes) corneti nov. sp. du Westphalien A du Couchant de Mons (Belgique). Annales de la Société scientifique de Bruxelles, 59: 56-59. R e n ie r , A., 1930. Considérations sur la stratigraphie du terrain houiller de la Belgique. Mémoires du Musée royal d'Histoire naturelle de Belgique, 44: 3-102.R e u s s , A.E., 1855. Palaeontologische Miscellen. III. Über eine neue Krusterspecies aus der Böhmischen Steinkohlenfonnation. Denkschriften der kaiserlichen Akademie der Wissenschaften zu Wien. Mathematisch-naturhistorische Classe Wien, 10, 81-83.S a r l e , C.J., 1903. A new eurypterid fauna from the base o f the Salina o f western New York. New York State Museum Bulletin, 69: 1080-1108.S c h m itz , G . & S t a i n i e r , X ., 1910 . Découverte en Campine de faunes marines et d ’un Eurypterus dans les strates inférieures du houiller. Annales de la Société géologique de Belgique, 36: 293-297.S c o t t , R.W., 1971. Eurypterids from the Dunkard Group (Pennsylvanian and Permian), southwestern Pennsylvania. Journal o f Paleontology, 45 (5): 833-837.S e l d e n , P.A., C o r r o n c a , J.A., & H ü n ic k e n , M.A., 2005. The true identity of the supposed giant fossil spider Megarachne. Biology Letters o f the Royal Society, 1 (1): 44-48.S t a i n i e r , X., 1911. Structure du bassin houiller de la province d ’Anvers. Bulletin de la Société belge de Géologie, 25: 209- 215.S t a i n i e r , X ., 1 917 . On a New Eurypterid from the Belgian Coal Measures. Quarterly Journal o f the Geological Society, 71 (4): 6 3 9 -6 4 7 .
S t o r m e r , L., 1934. Merostomata from the Downtonian Sandstone o f Ringerike, Norway. Skrifter utgitt av Det Norske Videnskaps-Akademi i Oslo I. Matematisk-Naturvidenskapelig Klasse, 10: 1-125.S t o r m e r , L., 1955. Merostomata. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology, Part P, Arthropoda 2. Geological Society o f America and University o f Kansas Press, Lawrence, Kansas, pp. 4-41.S t o r m e r , L., 1973. Arthropods from the Lower Devonian (Lower Emsian) o f Alken an der Mosel, Germany. Part 3: Eurypterida, Hughmilleriidae. Senckenbergiana lethaea, 54: 119-205.S t o r m e r , L., 1974. Arthropods from the Lower Devonian (Lower Emsian) o f Alken an der Mosel, Germany. Part 4: Eurypterida, Drepanopteridae, and other groups. Senckenbergiana lethaea, 54: 359-451.S t o r m e r , L. & W a t e r s t o n , C.D., 1968. Cyrtoctenus, gen. nov., a large late Palaeozoic arthropod with pectinate appendages. Transactions o f the Royal Society o f Edinburgh: Earth Sciences, 68 (4): 63-104.T e t l i e , O.E., 2000. Eurypterids from the Silurian o f Norway. Unpublished Cand. Scient thesis. Universitetet i Oslo, Norway, 152 pp.T e t l i e , O.E., 2004. Eurypterid phylogeny with remarks on the origin of arachnids. Unpublished PhD thesis. University o f Bristol, U.K., 320 pp.T e t l i e , O.E., B r a d d y , S.J., B u t l e r , P.D., & B r ig g s , D.E.G., 2004. A new eurypterid (Chelicerata: Eurypterida) from the

90 O. Erik T E T LIE & Peter V A N RO Y
Upper Devonian Gogo Formation o f Western Australia, with a review o f the Rhenopteridae. Palaeontology, 47 (4): 801-809. T e t l i e , O.E., & D u n lo p , J.A., 2005. A redescription o f the late Carboniferous eurypterids Adelophthalmus granosus von Meyer, 1853 and A. zadrai Pribyl, 1952. Mitteilungen aus dem Museum für Naturkunde in Berlin: Geowissenschaftliche Reihe, 8: 3-12.T o l l e r t o n J r , V.P., 1989. Morphology, taxonomy, and classification o f the Order Eurypterida Burmeister, 1843. Journal o f Paleontology, 63 (5): 642-657.T o l l e r t o n J r , V.P., 2004. Summary o f a revision o f New York State Ordovician eurypterids: implications for eurypterid palaeoecology, diversity and evolution. Transactions o f the Royal Society o f Edinburgh: Earth Sciences, 94 (3): 235-242. V a n O y e n , F.H., 1956. Contribution à la connaissance du genre Adelophthalmus Jordan et Meyer 1854. Mededelingen van de Geologische Stichting, serie C, 3 (7): 4-98. v o n M e y e r , H. 1853. Mitteilung an Prof. Bronn. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten- kunde 1853: 161-165.
W i l l s , L.J., 1964. The ventral anatomy o f the Upper Carboniferous eurypterid Anthraconectes Meek and Worthen. Palaeontology, 7: 474-507.
W o o d w a r d , H., 1907. Two new species o f Eurypterus from the Coal-Measures o f Ilkeston, Derbyshire. Geological Magazine, 4: 277-282.
O. Erik T e t l ie
Department o f Geology and Geophysics, Yale University PO Box 208109, New Haven, CT 06520-8109, U.S.A. Email: [email protected]
Peter V a n R o y
Department of Geology & Soil Science, Ghent University Krijgslaan 281 / S8, B-9000 Ghent, Belgium Email: [email protected]
T ypescrip t subm itted: June 5, 2005 R evised typescrip t received: S ep tem ber 5, 2005