![images1.loopnet.com · 2 j V Wk V d e 9 V l Y \ ] > ; @ D 9 ` 6 m g 9 V l Y \ , G > @ ] ] 9 `.j g f g U d k V d em Z Z m b g j d V g ng mW Y U f Ye b m j d ko m m b [ m ppY b [ V](https://static.fdocuments.nl/doc/165x107/5ec428f5121fe359165e2570/2-j-v-wk-v-d-e-9-v-l-y-d-9-6-m-g-9-v-l-y-g-9-j-g.jpg)
È w ¤ ï ¬ Æ @ @ ¤ ¬ Ê ñ - KAKEN · R S T P V î Õ ¤ (C) 2011 `2013 J V E É æ é e Q R...
4
科学研究費助成事業 研究成果報告書 様 式 C-19、F-19、Z-19 (共通) 機関番号: 研究種目: 課題番号: 研究課題名(和文) 研究代表者 研究課題名(英文) 交付決定額(研究期間全体):(直接経費) 34517 基盤研究(C) 2013 ~ 2011 血中カルシウムによるFgf23遺伝子発現の調節とWnt/βカテニンシグナルの関係 SerumcalciumupregulatesFgf23expressionbyDickopf-1-mediatedinhibitionofWnt/b eta-cateninsignalinginbone 70431963 研究者番号: 玉村 禎宏(TAMAMURA,Yoshihiro) 武庫川女子大学・健康・スポーツ科学部・博士研究員 研究期間: 23592696 平成 年 月 日現在 26 5 23 円 4,000,000 、(間接経費) 円 1,200,000 研究成果の概要(和文):Fgf23は骨細胞から分泌されるリン排泄促進因子である。代表的な遺伝性リン代謝疾患で血 清Fgf23濃度が上昇し低リン血症となることから、Fgf23はリン代謝において中心的な役割を果たすと考えられる。しか し、Fgf23発現調節機構は、不明な点が多い。本研究の結果から、血清カルシウムは、分泌型抑制因子Dickopf-1を介し て骨組織におけるWnt/beta-cateninシグナルを抑制するこにより、Fgf23発現を調節することが明らかとなった。これ らの知見は、抗Dickopf-1抗体を用いた新たなリン代謝性疾患の治療法の開発につながると期待される。 研究成果の概要(英文):Fgf23isaphosphaturicfactorwhichissecretedfromosteocytesinbone.Fgf23pl aysacentralroleforphosphatemetabolismasdemonstratedbyelevatedserumlevelsofFgf23inhypophosp hatemicdiseases.Post-transcriptionalmodificationofFgf23isextensivelystudied,howevermechanismsof transcriptionalregulationofFgf23arenotfullyunderstood.Inthisstudy,wedemonstratedthatserumc alciumregulatesFgf23expressionbyDickopf-1-mediatedinhibitionofWnt/beta-cateninsignalinginbone. Thisresultwillprovidethepossibilitiesofanewtherapyforhypophosphatemicdiseasesbyusinganti-Di ckpof-1antibody. 研究分野: 科研費の分科・細目: 医歯薬学 キーワード: Fgf23Wntカルシウム 歯学・形態系基礎歯科学
Transcript of È w ¤ ï ¬ Æ @ @ ¤ ¬ Ê ñ - KAKEN · R S T P V î Õ ¤ (C) 2011 `2013 J V E É æ é e Q R...

科学研究費助成事業 研究成果報告書
様 式 C-19、F-19、Z-19 (共通)
機関番号:
研究種目:
課題番号:
研究課題名(和文)
研究代表者
研究課題名(英文)
交付決定額(研究期間全体):(直接経費)
34517
基盤研究(C)
2013~2011
血中カルシウムによるFgf23遺伝子発現の調節とWnt/βカテニンシグナルの関係
Serum calcium upregulates Fgf23 expression by Dickopf-1-mediated inhibition of Wnt/beta-catenin signaling in bone
70431963研究者番号:
玉村 禎宏(TAMAMURA, Yoshihiro)
武庫川女子大学・健康・スポーツ科学部・博士研究員
研究期間:
23592696
平成 年 月 日現在26 5 23
円 4,000,000 、(間接経費) 円 1,200,000
研究成果の概要(和文):Fgf23は骨細胞から分泌されるリン排泄促進因子である。代表的な遺伝性リン代謝疾患で血清Fgf23濃度が上昇し低リン血症となることから、Fgf23はリン代謝において中心的な役割を果たすと考えられる。しかし、Fgf23発現調節機構は、不明な点が多い。本研究の結果から、血清カルシウムは、分泌型抑制因子Dickopf-1を介して骨組織におけるWnt/beta-cateninシグナルを抑制するこにより、Fgf23発現を調節することが明らかとなった。これらの知見は、抗Dickopf-1抗体を用いた新たなリン代謝性疾患の治療法の開発につながると期待される。
研究成果の概要(英文):Fgf23 is a phosphaturic factor which is secreted from osteocytes in bone. Fgf23 plays a central role for phosphate metabolism as demonstrated by elevated serum levels of Fgf23 in hypophosphatemic diseases. Post-transcriptional modification of Fgf23 is extensively studied, however mechanisms of transcriptional regulation of Fgf23 are not fully understood. In this study, we demonstrated that serum calcium regulates Fgf23 expression by Dickopf-1-mediated inhibition of Wnt/beta-catenin signaling in bone. This result will provide the possibilities of a new therapy for hypophosphatemic diseases by using anti-Dickpof-1 antibody.
研究分野:
科研費の分科・細目:
医歯薬学
キーワード: Fgf23 Wnt カルシウム
歯学・形態系基礎歯科学
様 式
1.研究開始当初の背景 Fgf23的とするホルモンとして作用する。Fgf23血中リン濃度を低下させる。血症性Fgf23異による低リン血症および同遺伝子の機能欠失型変異によるら、Fgf23な役割を持つと考えられる。しかし、遺伝子の転写調節機構に関して不明な点が多い。因子として知られアウトマウスではにおけるこれらのトマウスに対する高リン食摂取では回復されないが、高カルシウム食摂取により回復されることが報告されている。血清カルシウムによる経路を介さない存在が示唆されている 2.研究の目的 本研究では、血清カルシウムによる新たなFgf23る。 3.研(1)血清カルシウム濃度変化による骨組織での遺伝子発現の変化 4 週齢の食および高カルシウム食を摂取させ、4、および流し出した後、した。これらの法により、 (2) Wntに関して 内在性にUMR-106グナル修飾因子(Sigmaより (3) 血 清 カ ル シ ウ ム 濃 度 上 昇 に よ るDickopf コントロール食および高カルシウム食摂取後発現を1:1500)した。 (4) 血清カルシウム濃度上昇による血清Dickopf コントロール食および高カルシウム食摂取後濃 度 を
式 C-19、F-19、Z-19(共通)
1.研究開始当初の背景Fgf23 は、骨細胞から分泌され、腎臓を標
的とするホルモンとして作用する。Fgf23 は近位尿細管でのリン再吸収を抑制し、血中リン濃度を低下させる。血症性遺伝性リン代謝疾患における血清Fgf23 濃度の上昇、異による低リン血症および同遺伝子の機能欠失型変異による
Fgf23 は生体のな役割を持つと考えられる。しかし、遺伝子の転写調節機構に関して不明な点が多い。ビタミン D因子として知られアウトマウスではにおけるFgf23遺伝子発現低下が認められるこれらの Fgf23 発現の異常トマウスに対する高リン食摂取では回復されないが、高カルシウム食摂取により回復されることが報告されている。血清カルシウムによる経路を介さないFgf23存在が示唆されている
2.研究の目的 本研究では、血清カルシウムによる新たな
Fgf23 遺伝子発現調節機構の解明を目的とす
3.研究の方法 血清カルシウム濃度変化による骨組織で
の遺伝子発現の変化週齢の C57BL/6
食および高カルシウム食を摂取させ、、および 8 週間後に脛骨を採取し、骨髄を流し出した後、骨組織を粉砕し、した。これらの RNA法により、種々の
Wnt シグナルのに関して
内在性に Fgf23106 細胞に Wnt3a
グナル修飾因子(Sigma 社)処理を行い、より Fgf23 発現を調べた。
血 清 カ ル シ ウ ム 濃 度 上 昇 に よ るDickopf-1(Dkk-1)
コントロール食および高カルシウム食摂後 4 週間のマウス脛骨における
発現を抗 Dkk-11:1500)を用いたした。
血清カルシウム濃度上昇による血清Dickopf-1 濃度の変化に関して
コントロール食および高カルシウム食摂後 4 週間のマウス
濃 度 を Quantikine mouse Dkk
C-19、F-19、Z-19(共通)
1.研究開始当初の背景 は、骨細胞から分泌され、腎臓を標
的とするホルモンとして作用する。は近位尿細管でのリン再吸収を抑制し、
血中リン濃度を低下させる。代表的なリン代謝疾患における血清
濃度の上昇、Fgf23 遺伝子の異による低リン血症および同遺伝子の機能欠失型変異による高リン血症の誘発などか
は生体のリン代謝において中心的な役割を持つと考えられる。しかし、遺伝子の転写調節機構に関して不明な点が
D は、強力な因子として知られ、ビタミン Dアウトマウスでは、血清 Fgf23
遺伝子発現低下が認められる発現の異常は、同ノックアウ
トマウスに対する高リン食摂取では回復されないが、高カルシウム食摂取により回復されることが報告されている。血清カルシウムによるビタミン
Fgf23遺伝子発現調節機構の存在が示唆されている。
本研究では、血清カルシウムによる新たな
遺伝子発現調節機構の解明を目的とす
血清カルシウム濃度変化による骨組織で
の遺伝子発現の変化に関してC57BL/6 雄マウスにコントロール
食および高カルシウム食を摂取させ、週間後に脛骨を採取し、骨髄を
骨組織を粉砕し、RNA を用いて、
種々の遺伝子発現を調べた。
シグナルの Fgf23 発現
Fgf23 を発現する骨芽細胞株Wnt3a タンパク添加や
グナル修飾因子(BIO およびを行い、real-
発現を調べた。
血 清 カ ル シ ウ ム 濃 度 上 昇 に よ る1)発現に関して
コントロール食および高カルシウム食摂週間のマウス脛骨における
1 抗体(abcamを用いた免疫組織化学法により比較
血清カルシウム濃度上昇による血清濃度の変化に関して
コントロール食および高カルシウム食摂週間のマウスにおける血清
Quantikine mouse Dkk
C-19、F-19、Z-19(共通)
は、骨細胞から分泌され、腎臓を標的とするホルモンとして作用する。すなわち、
は近位尿細管でのリン再吸収を抑制し、代表的な低リン
リン代謝疾患における血清遺伝子の活性型変
異による低リン血症および同遺伝子の機能高リン血症の誘発などかリン代謝において中心的
な役割を持つと考えられる。しかし、Fgf23遺伝子の転写調節機構に関して不明な点が
は、強力な Fgf23 発現誘導D 受容体ノック
Fgf23 濃度低下と骨遺伝子発現低下が認められる
は、同ノックアウトマウスに対する高リン食摂取では回復されないが、高カルシウム食摂取により回復されることが報告されている。このことから、
ビタミン D シグナル伝子発現調節機構の
本研究では、血清カルシウムによる新たな遺伝子発現調節機構の解明を目的とす
血清カルシウム濃度変化による骨組織でに関して 雄マウスにコントロール
食および高カルシウム食を摂取させ、1、週間後に脛骨を採取し、骨髄を
骨組織を粉砕し、RNA を抽出を用いて、real-time PCR
遺伝子発現を調べた。
発現に対する作用
を発現する骨芽細胞株タンパク添加や Wntおよび XAV939、共に
-time PCR 法に
血 清 カ ル シ ウ ム 濃 度 上 昇 に よ る発現に関して
コントロール食および高カルシウム食摂週間のマウス脛骨における Dickopf
(abcam 社, ab109416, 免疫組織化学法により比較
血清カルシウム濃度上昇による血清濃度の変化に関して
コントロール食および高カルシウム食摂における血清 Dickopf
Quantikine mouse Dkk-1 Elisa
C-19、F-19、Z-19(共通)
は、骨細胞から分泌され、腎臓を標すなわち、
は近位尿細管でのリン再吸収を抑制し、低リン
リン代謝疾患における血清活性型変
異による低リン血症および同遺伝子の機能高リン血症の誘発などかリン代謝において中心的
Fgf23遺伝子の転写調節機構に関して不明な点が
発現誘導受容体ノック濃度低下と骨
遺伝子発現低下が認められる。は、同ノックアウ
トマウスに対する高リン食摂取では回復されないが、高カルシウム食摂取により回復さ
このことから、シグナル
伝子発現調節機構の
本研究では、血清カルシウムによる新たな遺伝子発現調節機構の解明を目的とす
血清カルシウム濃度変化による骨組織で
雄マウスにコントロール、3、
週間後に脛骨を採取し、骨髄をを抽出
time PCR
に対する作用
を発現する骨芽細胞株Wnt シ、共に
法に
血 清 カ ル シ ウ ム 濃 度 上 昇 に よ る
コントロール食および高カルシウム食摂Dickopf-1
, ab109416, 免疫組織化学法により比較
血清カルシウム濃度上昇による血清
コントロール食および高カルシウム食摂Dickopf-11 Elisa
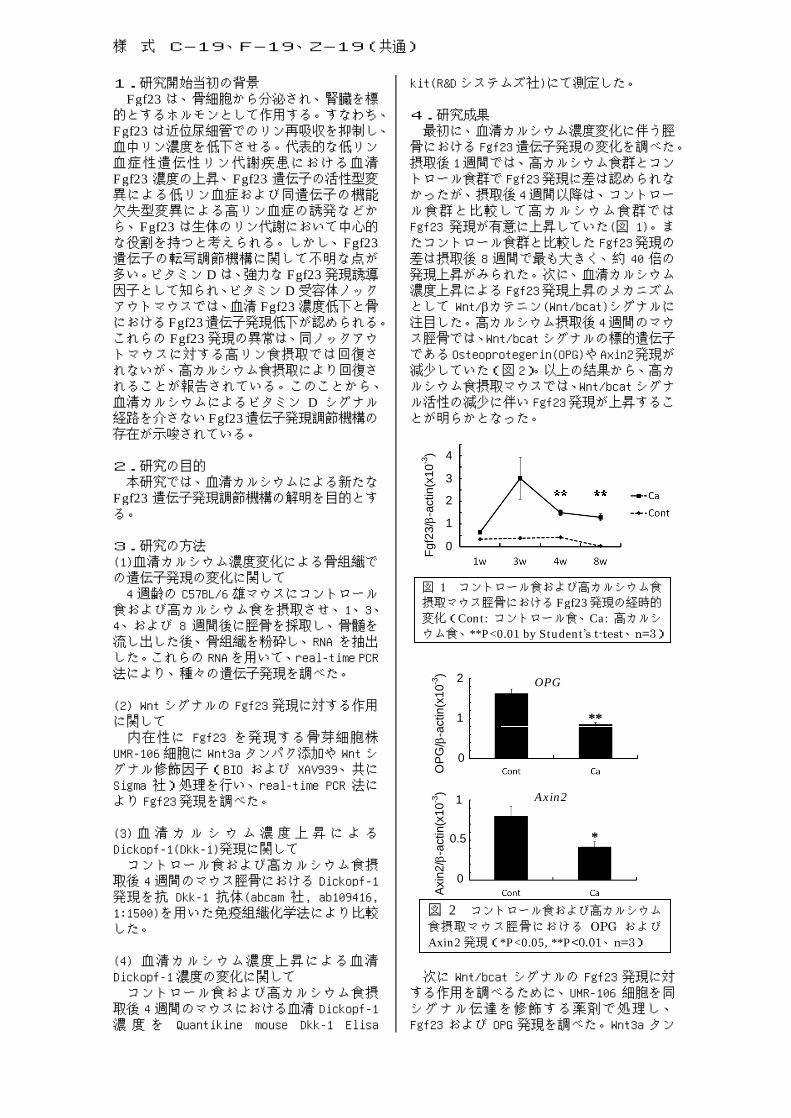
kit(R&D 4.研究成果最初に、
骨における摂取後トロール食群でかったが、摂取ル食群と比較して高カルシウム食Fgf23たコ差は摂取発現上昇がみられた。次に、血清カルシウム濃度上昇として注目した。ス脛骨では、である減少していたルシウム食摂取マウスでは、ル活性の減少に伴いとが明らかとなった。
次に
する作用を調べるために、シグナル伝達を修飾する薬剤で処理し、Fgf23
図摂取マウス脛骨における変化(ウム食、
Fgf
23/
-act
in(x
10-3
)
図食摂取マウス脛骨におけるAxin2
kit(R&D システムズ社
4.研究成果 最初に、血清カルシウム濃度変化に伴う脛
骨におけるFgf23摂取後 1週間では、トロール食群でかったが、摂取ル食群と比較して高カルシウム食Fgf23 発現が有意にたコントロール差は摂取後 8 週間発現上昇がみられた。次に、血清カルシウム濃度上昇によるとして Wnt/カテニン注目した。高カルシウム摂取ス脛骨では、Wnt/bcatであるOsteoprotegerin減少していた(図ルシウム食摂取マウスでは、ル活性の減少に伴いとが明らかとなった。
次に Wnt/bcatする作用を調べるために、シグナル伝達を修飾する薬剤で処理し、Fgf23 および OPG
0
1
2
3
4
図 1 コントロール食および高カルシウム食摂取マウス脛骨における変化(Cont: コントロール食、ウム食、**P<0.01 by Student
Fgf
23/
-act
in(x
10)
1
2
0
0.5
1
0
OP
G/
-act
in(x
10-3
) A
xin2
/-a
ctin
(x10
-3)
図 2 コントロール食および高カルシウム食摂取マウス脛骨におけるAxin2 発現(*P<0.05, **P<0.01
システムズ社)にて測定した。
血清カルシウム濃度変化に伴う脛Fgf23遺伝子発現の変化を調べた。
週間では、高カルシウムトロール食群でFgf23発現に差はかったが、摂取後 4週間以降はル食群と比較して高カルシウム食
発現が有意に上昇していたントロール食群と比較した
週間で最も大きく、約発現上昇がみられた。次に、血清カルシウム
によるFgf23発現上昇のメカニズムカテニン(Wnt/bcat)
高カルシウム摂取Wnt/bcat シグナル
Osteoprotegerin(OPG)(図 2)。以上の結果から、高カ
ルシウム食摂取マウスでは、ル活性の減少に伴いFgf23発現が上昇することが明らかとなった。
Wnt/bcat シグナルのする作用を調べるために、UMRシグナル伝達を修飾する薬剤で処理し、
OPG 発現を調べた。
コントロール食および高カルシウム食摂取マウス脛骨におけるFgf23
コントロール食、**P<0.01 by Student
**
OPG
Axin2
コントロール食および高カルシウム食摂取マウス脛骨における
*P<0.05, **P<0.01
にて測定した。
血清カルシウム濃度変化に伴う脛遺伝子発現の変化を調べた。高カルシウム食群とコン発現に差は認められな
週間以降は、コントロール食群と比較して高カルシウム食群では
していた(図 1)と比較したFgf23発現の最も大きく、約 40
発現上昇がみられた。次に、血清カルシウム発現上昇のメカニズム(Wnt/bcat)シグナルに
高カルシウム摂取後 4週間のマウシグナルの標的遺伝子(OPG)やAxin2発現が
。以上の結果から、高カルシウム食摂取マウスでは、Wnt/bcat シグナ
発現が上昇するこ
シグナルの Fgf23 発現に対UMR-106 細胞を
シグナル伝達を修飾する薬剤で処理し、発現を調べた。Wnt3a
コントロール食および高カルシウム食Fgf23発現の経時的
コントロール食、Ca: 高カルシ**P<0.01 by Student’s t-test、n=3
**
*
**
コントロール食および高カルシウム食摂取マウス脛骨における OPG および
*P<0.05, **P<0.01、n=3)
血清カルシウム濃度変化に伴う脛遺伝子発現の変化を調べた。
群とコンられな
、コントロー群では1)。ま発現の40 倍の
発現上昇がみられた。次に、血清カルシウム発現上昇のメカニズム
シグナルに週間のマウ
の標的遺伝子発現が
。以上の結果から、高カシグナ
発現が上昇するこ
発現に対細胞を同
シグナル伝達を修飾する薬剤で処理し、Wnt3a タン
コントロール食および高カルシウム食発現の経時的
高カルシn=3)
コントロール食および高カルシウムおよび)
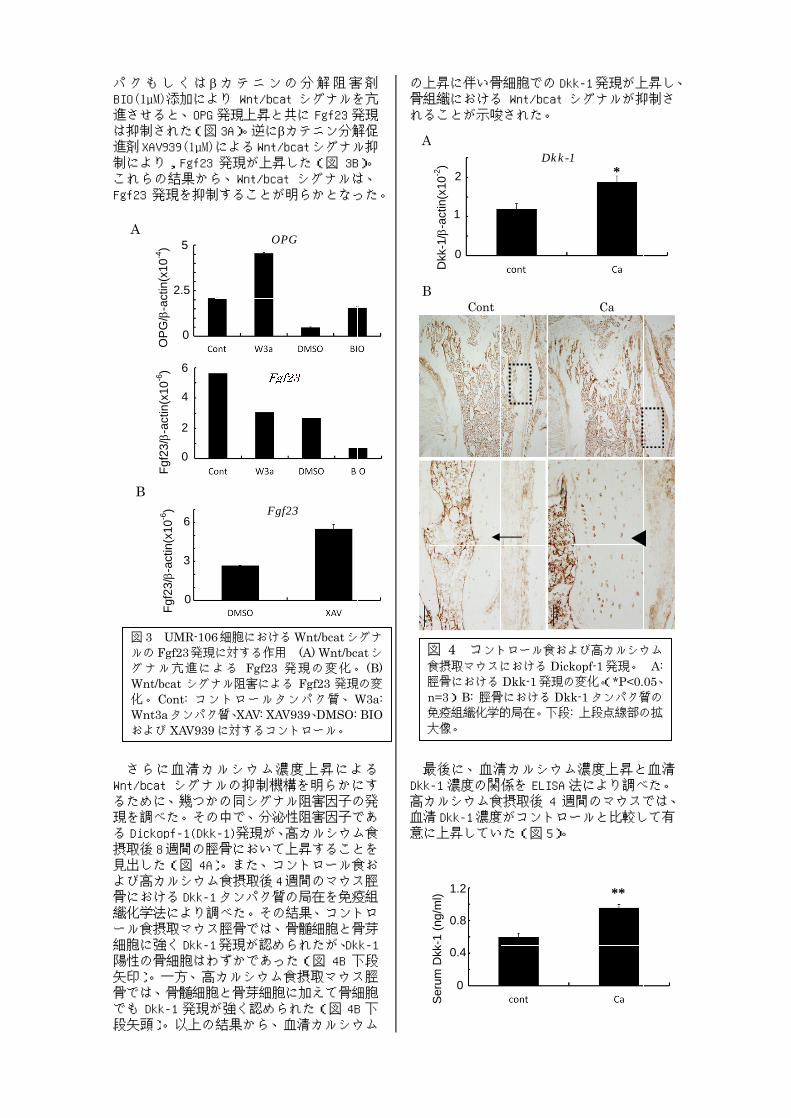
パクもしくはBIO(1µM)進させると、は抑制された進剤XAV939制によりこれらの結果から、Fgf23 さらに血清カルシウム濃度上昇による
Wnt/bcatるために、幾つかの同シグナル阻害因子の発現を調べた。その中で、る Dickopf摂取後見出したよび高カルシウム食摂取骨における織化学法により調べた。その結果、コントロール食摂取マウス脛骨では、骨髄細胞と骨芽細胞に強く陽性の骨細胞は矢印)骨では、骨髄細胞と骨芽細胞に加えて骨細胞でも段矢頭)
A
B
図ルのグナル亢進によるWnt/bcat化。Wnt3aおよび
パクもしくは (1µM)添加により
進させると、OPGは抑制された(図
XAV939(1µM)制により,Fgf23これらの結果から、Fgf23 発現を抑制することが明らかとなった。
さらに血清カルシウム濃度上昇によるWnt/bcat シグナルの抑制機構を明らかにするために、幾つかの同シグナル阻害因子の発現を調べた。その中で、
Dickopf-1(Dkk後 8週間の脛骨において上昇することを
見出した(図 4A)よび高カルシウム食摂取骨におけるDkk-1織化学法により調べた。その結果、コントロール食摂取マウス脛骨では、骨髄細胞と骨芽細胞に強くDkk-1
の骨細胞はわずか矢印)。一方、高カルシウム食摂取マウス脛骨では、骨髄細胞と骨芽細胞に加えて骨細胞でも Dkk-1 発現が強く認められた段矢頭)。以上の結果から、血清カルシウム
0
2.5
5
0
2
4
6
OP
G/
-act
in(x
10-4
) F
gf23
/-a
ctin
(x10
-6)
B
0
3
6
Fgf
23/
-act
in(x
10-6
)
図 3 UMR-106ルのFgf23発現に対する作用グナル亢進によるWnt/bcat シグナル阻害による化。Cont: コントロールタンパク質、Wnt3aタンパク質、および XAV939
カテニンの分解阻害剤添加により Wnt/bcat
OPG 発現上昇と共に(図 3A)。逆にカテニン分解促
(1µM)によるWnt/bcatFgf23 発現が上昇した
これらの結果から、Wnt/bcat発現を抑制することが明らかとなった。
さらに血清カルシウム濃度上昇によるシグナルの抑制機構を明らかにす
るために、幾つかの同シグナル阻害因子の発現を調べた。その中で、分泌性阻害因子であ
1(Dkk-1)発現が、高カルシウム食週間の脛骨において上昇することを
)。また、コントロール食および高カルシウム食摂取後4週間のマウス脛
1タンパク質の局在を免疫組織化学法により調べた。その結果、コントロール食摂取マウス脛骨では、骨髄細胞と骨芽
1発現が認められたが、わずかであった
。一方、高カルシウム食摂取マウス脛骨では、骨髄細胞と骨芽細胞に加えて骨細胞
発現が強く認められた。以上の結果から、血清カルシウム
OPG
Fgf23
Fgf23
106 細胞における発現に対する作用
グナル亢進による Fgf23 発現の変化。シグナル阻害によるコントロールタンパク質、
タンパク質、XAV: XAV939XAV939 に対するコントロール
カテニンの分解阻害剤Wnt/bcat シグナルを亢
発現上昇と共に Fgf23 発現カテニン分解促
Wnt/bcatシグナル抑発現が上昇した(図 3B
Wnt/bcat シグナルは、発現を抑制することが明らかとなった。
さらに血清カルシウム濃度上昇によるシグナルの抑制機構を明らかにす
るために、幾つかの同シグナル阻害因子の発分泌性阻害因子であ
発現が、高カルシウム食週間の脛骨において上昇することを
また、コントロール食お週間のマウス脛
タンパク質の局在を免疫組織化学法により調べた。その結果、コントロール食摂取マウス脛骨では、骨髄細胞と骨芽
発現が認められたが、Dkkであった(図 4B 下段
。一方、高カルシウム食摂取マウス脛骨では、骨髄細胞と骨芽細胞に加えて骨細胞
発現が強く認められた(図 4B。以上の結果から、血清カルシウム
OPG
Fgf23
Fgf23
細胞における Wnt/bcat シグナ発現に対する作用 (A) Wnt/bcat
発現の変化。シグナル阻害による Fgf23 発現の変コントロールタンパク質、W3a:
XAV: XAV939、DMSO: BIOに対するコントロール。
カテニンの分解阻害剤シグナルを亢
発現カテニン分解促
シグナル抑3B)。
シグナルは、発現を抑制することが明らかとなった。
さらに血清カルシウム濃度上昇によるシグナルの抑制機構を明らかにす
るために、幾つかの同シグナル阻害因子の発分泌性阻害因子であ
発現が、高カルシウム食週間の脛骨において上昇することを
また、コントロール食お週間のマウス脛
タンパク質の局在を免疫組織化学法により調べた。その結果、コントロール食摂取マウス脛骨では、骨髄細胞と骨芽
Dkk-1下段
。一方、高カルシウム食摂取マウス脛骨では、骨髄細胞と骨芽細胞に加えて骨細胞
4B 下。以上の結果から、血清カルシウム
の上昇に伴い骨細胞での骨組織におけるれることが示唆された。 最後に、血清カルシウム濃度上昇と血清
Dkk高カルシウム食摂取血清意に上昇していた
シグナWnt/bcatシ
発現の変化。 (B) 発現の変
W3a: DMSO: BIO
A
B
図食摂取マウスにおける脛骨におけるn=3免疫組織化学的大像。
の上昇に伴い骨細胞での骨組織におけるれることが示唆された。
最後に、血清カルシウム濃度上昇と血清
Dkk-1 濃度の関係を高カルシウム食摂取血清Dkk-1濃度がコントロールと比較して有意に上昇していた
A
0
1
2
Dkk
-1/
-act
in(x
10-2
)
0
0.4
0.8
1.2
Ser
um D
kk-1
(ng
/ml)
B Cont
図 4 コントロール食および高カルシウム食摂取マウスにおける脛骨におけるn=3) B: 脛骨における免疫組織化学的大像。
の上昇に伴い骨細胞でのDkk骨組織における Wnt/bcat シグナルが抑制されることが示唆された。
最後に、血清カルシウム濃度上昇と血清濃度の関係を ELISA 法により調べ
高カルシウム食摂取後 4 週間のマウスでは、濃度がコントロールと比較して有
意に上昇していた(図 5)。
Dkk-1
ントロール食および高カルシウム食摂取マウスにおける Dickopf脛骨における Dkk-1 発現の変化。(
脛骨における Dkk免疫組織化学的局在。下段:
Dkk-1発現が上昇し、シグナルが抑制さ
最後に、血清カルシウム濃度上昇と血清法により調べ週間のマウスでは、
濃度がコントロールと比較して有。
1 *
**
Ca
ントロール食および高カルシウムDickopf-1 発現。
発現の変化。(*P<0.05Dkk-1 タンパク質の
: 上段点線部の拡
発現が上昇し、シグナルが抑制さ
最後に、血清カルシウム濃度上昇と血清法により調べた。週間のマウスでは、
濃度がコントロールと比較して有
ントロール食および高カルシウム A:
*P<0.05、タンパク質の
上段点線部の拡

結論として、本研究の結果から、血清カル
シウムは、Dkk-1 発現誘導を介して Wnt/bcatシグナルを負に制御することにより、Fgf23遺伝子発現を上昇させることが示唆された。これまでに血清カルシウムとWntシグナルおよび Dkk-1 に着目した報告は無く、本研究で得られた知見は、新たな Fgf23 発現調節機構であると考えられる。また、最近ミエローマなどの腫瘍においてDkk-1発現上昇が報告されており、これらの腫瘍に対する抗 Dkk-1 抗体投与による治療法がマウスで試されている。このことから、本研究の結果は、抗 Dkk-1抗体を用いた新たなリン代謝疾患治療法開発の可能性を示唆すると考えられる。
5.主な発表論文等 (研究代表者、研究分担者及び連携研究者には下線) 〔雑誌論文〕(計 5 件) ①Matsusita Y, Sakamoto K, Tamamura Y, Shibata Y, Minamizato T, Kihara T, Ito M, Katsube K, Hiraoka S, Koseki H, Harada K, Yamaguchi A. CCN3 protein participates in bone regeneration as an inhibitory factor. J. Biol. Chem. 288(27), 19973-19985, 2013,査読有り ② Makino Y, Takahashi Y, Tanabe R, Tamamura Y, Watanabe T, Haraikawa M, Hamagaki M, Hata K, Kanno J, Yoneda T, Saga Y, Goseki-Sone M, Kaneko K, Yamaguchi A, Iimura T. Spatiotemporal disorder in the axial skeleton development of the Mesp2-null mouse: a model of spondylocostal disostosis and spondylothoracic dysostosis. Bone, 53(1), 248-258, 2013, 査読有り ③ Watanabe T, Tamamura Y, Hoshino A, Makino Y, Kamioka H, Amagasa T, Yamaguchi A, Iimura T. Increasing participation of sclerostin in postnatal bone development, revealed by three-dimensional immunofluorescence morphometry. Bone, 51(3), 447-458, 2012, 査読有り ④Tamamura Y, Yamaguchi A. Bone and tooth in calcium and phosphate metabolism. Clin. Calcium, 22(1), 11-17, 2012, 査読無し ⑤Cao L, Moriishi T, Miyazaki T, Iimura T, Hamagaki M, Nakane A, Tamamura Y, Komori T, Yamaguchi A. Comparative morphology of the osteocyte lacunocanalicular system in various vertebrates. J. Bone Miner. Metab., 29(6), 662-670, 2011, 査読有り 6.研究組織
(1)研究代表者 玉村 禎宏 (TAMAMURA, Yoshihiro)
武庫川女子大学・健康スポーツ科学部・博士研究員
研究者番号:70431963 (2)研究分担者 (3)連携研究者 山口 朗 (YAMAGUCHI, Akira)
東京医科歯科大学・口腔病理学講座・教授 研究者番号: 00142430 飯村 忠浩 (IIMURA, Tadahiro) 愛媛大学・プロテオサイエンスセンター バイオイメージング部門・准教授
研究者番号:20282775 坂本 啓 (SAKAMOTO, Kei) 東京医科歯科大学・口腔病理学講座・助教 研究者番号:00302886
図 5 コントロール食および高カルシウム食摂取マウスにおける血清 Dkk-1 濃度(**P<0.01、n=3)



![Ë ' 0 129 · 2020-01-17 · P \ü A Ò B ]B]d] Þ Ã C\¤ No.129 Ë ' 0 ! H Æ H | ! H\Ø ¿ Ø\² Ë ' ¨ 0 O N P O { E J Æ ¥ Á { ù T S Å ¼ v < Y ù ß w 8 J 0 x § Ï u °](https://static.fdocuments.nl/doc/165x107/5ec3cd536858954c363a3424/-0-129-2020-01-17-p-a-b-bd-f-c-no129-0-h-h-.jpg)






![U ²aA æ æ AIF :æ 3 - Visit 1066 Country Flemish.pdf · 2 $uxqgho (o $pehu h\ &dvw h 35$.7,6&+ +dvwlqjv 7rzq &hqwuh 3ulru\ 0hdgrz]hh ohhj ,v +dvwlqjv hhq ohyhqgljh vwdg phw hhq4xhhqv](https://static.fdocuments.nl/doc/165x107/5a78a2407f8b9a87198e3dd5/u-aa-aif-3-visit-1066-country-flemishpdf2-uxqgho-o-pehu-h-dvw-h-3576.jpg)







