
· Web viewstraling kan aanrichten aan "bloed" dat zo nauw aan UV Stralen word blootgesteld-in...
40
Ik heb eens gelezen dat de ganglioncellen en bipolaire cellen (de 'bedrading') die bovenop de dieper gelegen staafjes en kegeltjes liggen, de functie hebben dat ze ultraviolet licht tegenhouden, hetgeen kanker zou kunnen veroorzaken. Inktvissen hebben dit niet van node, omdat ze in de diepzee leven Kanker (3) als gevolg van UV-straling onstaat( ondermeer ) door het gebrek aan melanine(of, gebreken aan de pigmentcellen ) in de IRIS : REGENBOOGVLIES( en ook op de huid --> zonneslag en huidkanker ) ,en waardoor vooral het regenboogvlies "verbrand" en niet meer in staat is de lichtinval te regelen ..( zoals bijvoorbeeld bij http://nl.wikipedia.org/wiki/Albinisme ) Bovendien liggen bedradingen in het "verkeerd gewonden "(vertebraten)-oog bovenop de staafjes en kegeltjes dus dichter bij de lichtinval -bron ( de iris in camera ogen ) dan dezen die zoals bij de inktvis onder deze fotocellen laag liggen .... (1)(2) Het is allemaal het gevolg van zeer oude evolutionaire prutserij-paden die werden bewandeld zoals in een "drunkards walk " en met rude goldberg design tot gevolg (in het oog van vertrebraten ....) Maw ---> een anatomische miskleun ... -Bovendien zijn later (vervang)oplossingen gebracht die dit euvel moesten omzeilen en toch scherp zicht bij vertebraten hebben mogelijk gemaakt ; o.a. de fovea en de Muller cellen .... (Net zoals men oude electrische leidingen afkoppeld , maar wel in de muur laat zitten , en men nieuwe leidingen gaat installeren ) IP Theorie (1)(2) Het gaat voornamelijk over: het " binnenste buiten" geplaatst zijn van die oogstructuren rond het vertebraten -netvlies en in vergelijking met de configuratiesd rond het netvliesepitheel van bijvoorbeeld, inktvissen ( ---> bij de mens is die bedoelde netvlies-structuur : de binnenste derde laag van het oog )die betaat uit fotocellen ( staafjes en kegeltjes ) en de erbovenop liggende ( dichter bij de lichtinvalbron ) "bedradingen" ... ) http://cas.bellarmine.edu/tietjen/Laboratories/Eye02.gif
Transcript of · Web viewstraling kan aanrichten aan "bloed" dat zo nauw aan UV Stralen word blootgesteld-in...

Ik heb eens gelezen dat de ganglioncellen en bipolaire cellen (de 'bedrading') die bovenop de dieper gelegen staafjes en kegeltjes liggen, de functie hebben dat ze ultraviolet licht tegenhouden, hetgeen kanker zou kunnen veroorzaken. Inktvissen hebben dit niet van node, omdat ze in de diepzee leven
Kanker (3) als gevolg van UV-straling onstaat( ondermeer ) door het gebrek aan melanine(of, gebreken aan de pigmentcellen ) in de IRIS : REGENBOOGVLIES( en ook op de huid --> zonneslag en huidkanker ) ,en waardoor vooral het regenboogvlies "verbrand" en niet meer in staat is de lichtinval te regelen ..( zoals bijvoorbeeld bij http://nl.wikipedia.org/wiki/Albinisme )
Bovendien liggen bedradingen in het "verkeerd gewonden "(vertebraten)-oog bovenop de staafjes en kegeltjes dus dichter bij de lichtinval -bron ( de iris in camera ogen ) dan dezen die zoals bij de inktvis onder deze fotocellen laag liggen .... (1)(2)
Het is allemaal het gevolg van zeer oude evolutionaire prutserij-paden die werden bewandeld zoals in een "drunkards walk " en met rude goldberg design tot gevolg (in het oog van vertrebraten ....) Maw ---> een anatomische miskleun ...-Bovendien zijn later (vervang)oplossingen gebracht die dit euvel moesten omzeilen en toch scherp zicht bij vertebraten hebben mogelijk gemaakt ; o.a. de fovea en de Muller cellen ....(Net zoals men oude electrische leidingen afkoppeld , maar wel in de muur laat zitten , en men nieuwe leidingen gaat installeren )
IP Theorie(1)(2)Het gaat voornamelijk over: het " binnenste buiten" geplaatst zijn van die oogstructuren rond het vertebraten -netvlies en in vergelijking met de configuratiesd rond het netvliesepitheel van bijvoorbeeld, inktvissen( ---> bij de mens is die bedoelde netvlies-structuur : de binnenste derde laag van het oog )die betaat uit fotocellen ( staafjes en kegeltjes ) en de erbovenop liggende ( dichter bij de lichtinvalbron ) "bedradingen" ... )
http://cas.bellarmine.edu/tietjen/Laboratories/Eye02.gif

fig ; het menselijke netvlies met bedradingen(links ) en fotocellen ( rechts ; gekleurd ; rods & cones ) het vaatvlies volgt NA de fotocellen ( gaande van binnen naar buiten in de dwars-dorsnede van de oogbol )-->niet op de afbeelding /uiterst rechts buiten beeld ig ; het
zie ook --> voor het blog artikel van Tomaso AgricolaIP Theorie
De voedende bloedvaten bevinden zich in de tweede ( midden)laag van het oog ---> het vaatvlies (2)
NOTEN(1)Bij inkvissen zit de bedradigs eronder .....Op de fovea-put ( de gele vlek ) bij de mens ( het punt waarmee we scherp zien ) is die bedradingslaag erg dun ...zoals je hieronder kunt zien

(2) De lichtinval is aan de voorkant daar waar ook de bedrading zit,als de bedrading aan de achterkant of ergens anders zit kan het geen licht impulsen opvangen en ook niet omzetten in info.De allereerste initieele omzetting van invallende fotonen (en de beeldvorming dpoor de diverse inval op verschillende fotocellen ) gebeurt in
de staafjes en kegeltjes( de primaire foto sensoren ) ; het aldus bekomen geheel aan signalen wordt verder geleid , doorgegeven en bewerkt door de zenuwbedrading die bovenop de fotocellen- laag zit ....Dat doen zintuigelijke zenuwen-bedradingen altijd .... diverse detecteerbare binnenkomende fysische en chemische signalen vanuit de primaire sensoren ( in dit geval de staafjes en kegeltjes )omzetten en coderen( door de synapsen werking ) in elektro-chemische puls-treinen die uiteindelijk de (in dit geval) visuele cortex bereiken ....(3)Wat betreft oogkanker heb ik het vooral over oog melanomen ; http://www.kankerwiehelpt.nl/default.asp?V_ITEM_ID=355 Dat is een kankertype dat onstaat bij een onstsporing in pigment( melanime )cellen die erg gevoelig zijn aan UV-stralingen ...Er bestaan natuurlijk ook andere oogkankers maar die manifesteren zich reeds op zeer jonge leeftijd en zijn NIET onmiddelijk UV -gebonden ( voor zover is geweten ) ; van melanomen wordt dat laatste wel vermoed .....
Creationistishe claim ....Onze ogen zijn verkeerd gewonden ontworpen , omdat we met inkvissenogen blind zouden worden door de UV-straling die nu door de bloedvaten tegengehouden wordt. ...
Is het net-ontwerp van bloed en zenuw leidingen en bedradingen "in het menselijke oog een UV filter ? "1.- (menselijke ALBINO's ( voorkomend bij alle gewervelde /ongewervelde dieren )kunnen nog steeds niet ongestraft in de zon lopen , zonder allerlei zonnebrillen en bedekkingen ; Ondanks het feit dat ze ook over zo'n zelfde soortspecifieke "verkeerde " beshermende zenuwwindingen en bloedvaten -filter (vertebraten oog ) systeem bechikken die hun ogen zogezegd "beschermen " tegen UV : Wat ze missen is NIET die netwerken of beschermende binnenste buiten windingen , wel de nodige hoeveelheid geschikte melanine in hun regenboogvlies ( iris )waardoor ze " rode " of een zeer lichtgekleurde ogen [/b]en ee "witte" huid bezitten .... ---->ALBINISME<KLIK)
</KLIK)2.- En nee , die oog- melamine in het oog word NIET aangevoerd of vervangen door het "bloed" ...

Je hebt gewoon van bij de geboorte :Vanaf de 20 ste embryonala week (en daarna nog een aantal jaren een beetje verder ontwikkeld ) een voorraadje melanine in het regenboogvlies , in bezit ... je zingt er de rest van je levenstijd wel mee uit ( of toch lang genoeg om je partner te vinden en vlug wat kindjes te maken ) ...
Zenuw- en haarvaten netwerken als filter ? M-E-N-A-L-I-N-E , is de voornaamste en belangrijkste filter , ( in het oog ) samen met de gekleurde iris ( een te weinig gekleurde iris " verbrand " ) die de lichtintensiteit naar het binnenoog regelt (= Het reflexmatig vernauwen en verbreden van de lichtinval wordt aangestuurd door een sensor = de "derde" fotocel (een zogenaamde ganglioncel) die ook de biologische rythmes lijkt te regelen en bij "blinde muizen " -mutanten nog steeds actief is http://www.scientificamerican.com/article.cfm?id=the-hidden-organ-in-our-eyeshttp://en.wikipedia.org/wiki/Ignacio_Provencio )--->IRIS : REGENBOOGVLIES<KLIK)</KLIK)(en misschien toch een " heel klein beetje" die netwerken ??? ... )Zoek maar eens op wat de schade kan zijn die de UV straling kan aanrichten aan "bloed" dat zo nauw aan UV Stralen word blootgesteld-in dat oog - Of vraag het eens aan een oogarts ; ik ben er geen , ik zal me dus ook niet wagen aan een uitspraak daarover .... )
3.- verder :
---->Er zijn vertebraten die uitsluitend in het water leven en toch zo'n verkeerd gewonden oog hebben ( vissen / walvisachtigen ZELFS die "verkeerde windingen "in het vertebraten oog van( de naar het water terug gekeerde ,) zoogdieren zijn dus wèl behouden door de evolutie ondanks hun onnut ? ) en ---> Volgens de geologen en paleontologen zijn vissen ouder dan de tetrapode landvertebraten ( die er met enkelen onder hen een gemeenschappelijke voorouder op nahouden ? ---> ) .... -- HET WATER filtert toch voor die vis- gasten eveneens UV stralen weg , net zoals dat het geval is voor de intvis ? ... Vanwaar dan het "design verschil ?
---> bovendien zijn er veel LANDslakken die net als inktvissen de" juiste "gewonden ogen hebben ( ja t'is wel verre familie hé ) ; alleen zitten die voyeurs niet in het "beshermende" water maar wel op het land ---> en zelfs spinnen hebben camera-lens -ogen die ook al tegen UV kunnen ( ze zien wel niet zo goed , schijnt het ) .... ---> Daarentegen bestaan er in de zee levende platwormen met ogen die eveneens "verkeerd " zijn gewonden ... overbodig dus als het water een uv filter is ... toch ?

http://library.thinkquest.org/28030/eyeevo.htmhttp://www.scholarpedia.org/article/Planaria_nervous_systemhttp://www.pnas.org/content/100/24/14046.full
http://cmirt.wcupa.edu/PDF_Files/secret-pdfs/Planaria/EM/511.pdf
http://jeb.biologists.org/content/201/9/1263.full.pdfhttp://web1.stmaryssen-h.schools.nsw.edu.au/SMSHS/ricks%20sites/Biology%20web%20site/HSC_9_5%20communications/eyes_compared/Eyes.html
-- Bovendien hebben alle modellen van ogen die er maar bestaan ( er bestaan zo'n 40 a 60 verschillend gebouwde kijkers , geloof ik )allemaal pigmenten of beter een pigmentlaag .( melanine dus ? ).. http://tsjok45.multiply.com/photos/album/1938
- Inktvissen kan je zelfs in de " mooie blauwe ogen " kijken ,ze hebben ook een regenboogvlies of iris ..... ( blonde haren hebben ze niet .... )
oog reuzenintvis
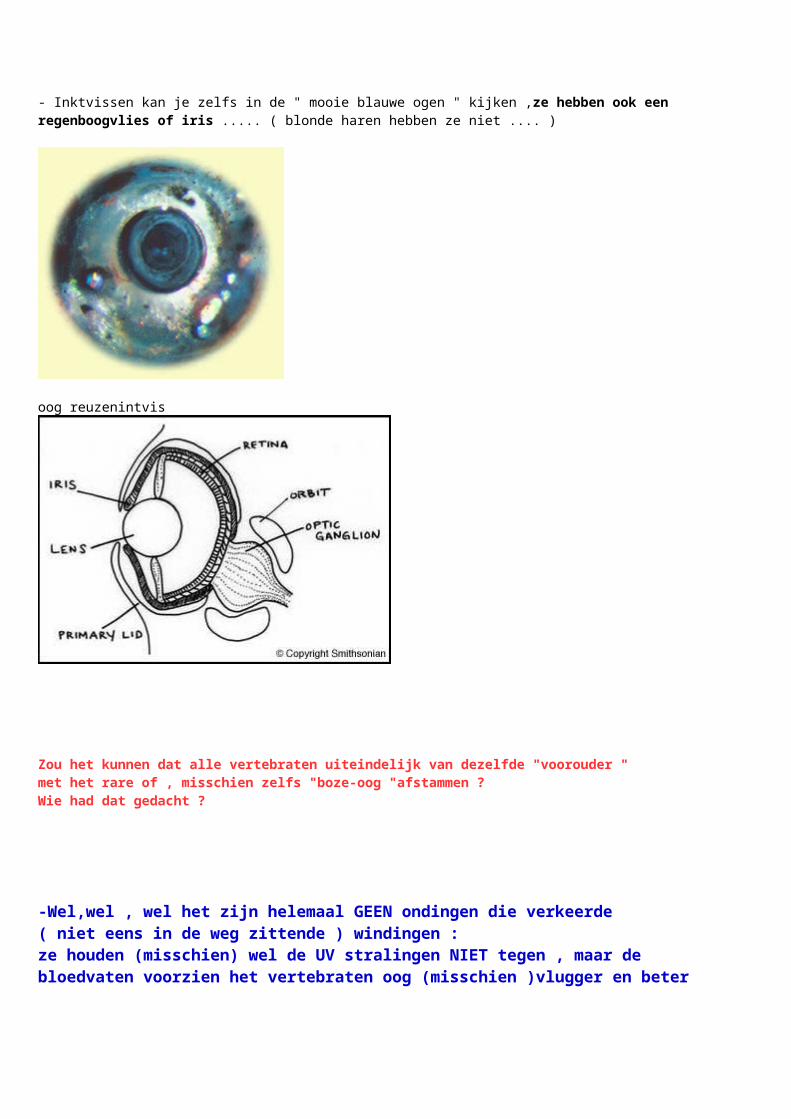
Zou het kunnen dat alle vertebraten uiteindelijk van dezelfde "voorouder "met het rare of , misschien zelfs "boze-oog "afstammen ? Wie had dat gedacht ?
-Wel,wel , wel het zijn helemaal GEEN ondingen die verkeerde ( niet eens in de weg zittende ) windingen : ze houden (misschien) wel de UV stralingen NIET tegen , maar de bloedvaten voorzien het vertebraten oog (misschien )vlugger en beter van voeding zodat een beter zicht mogelijk is : vogels hebben dat zelfs nog uitgebreid - maar dat is een ander verhaaltje ;(namelijk = Over de fovea )
alvast ook dit : ".....The possibility that the vitreous humour plays an important role in transporting nutrients is supported by existence of a structure in the eye in certain birds. Birds, like us, have a back to front retina. The special structure in avian eyes is called ‘a pecten’ or ‘comb’ and it has an excellent blood supply. ...."
Zie meer daarover -- http://www.catalase.com/retina.htm
Toch zitten we met een blinde vlek he ? en met een zeer kwetsbare risico -volle inrichting .... de zenuwbedrading ligt immers dichter bij de buitenwereld ( en kan dus veel makkelijker in de brokken delen bij een ongevalletje ) dan in het invertebraten oog ... Kleinigheid ?
Dat daarbij ;ook nog de hersenen beter moeten worden ontworpen om de handicaps die door het speciale vertebraten-oog ( blinde vlek ) onstaan te compenseren is een andere kleinigheid : echter een uiterst "raar" en verspilziek dom - ontwerp principe ;
--->De fouten in de oorspronkelijke design , oplossen door een gesofistikeerd apparaat dat de resultaten moet oplappen is gewoon te gek : - je kan toch veel goedkoper direkt de foute boel herbouwen ; zodat de resultaten van die broodnodige "processor"w 챕 l meteen correct zijn ; --->maar daar heb je natuurlijk wel ingenieurs voor nodig :

die was (waren) er dus even niet ? ( misschien op de boemel ?op stap in Antwerpen ? )
zie verder - http://wiki.cotch.net/index.php/Vertebrate_eye
We moeten toch wat voorzichtiger zijn vooraleer we " slecht ontwerp" schreeuwen ;
Echter ; knoei en knutselwerk en "roeien met de riemen" die er zijn is een veel betere verklaring voor een door het evolutie-proces ineengeflanst tussen"resultaat" Wat betekent ; de windingen in vertebraten ogen (retina) zijn omgekeerd ?
Quote:In vertebrates the eye forms by invagination of the optic vesicle, and the inner layer of the double walled optic cup becomes the retina. Therefore, the retina is "reversed" with the light-sensitive portion (the rods and cones) on the outside. This means that incoming light has to pass through all of the layers before it reaches the rods and cones. There appears to be no selective advantage or disadvantage to this reversed setup; it's just the way it turned out in vertebrates because of the other engineering considerations related to eye development.
Daarom ; De samenstelling van het oog laat geen doordacht concept zien, maar iets dat nou eenmaal zo tot stand is gekomen.
Maar zelfs vertrekkend van het zeer oude bouwprincipe geldend in vertebraten ogen ( en in de trilhaarwormen (stam plathelminthes- Turbellaria/ planaria ) is de evolutie er toch in geslaagd een paar van de meest efficiente en scherpe ogen te ontwikkelen die er bestaan ... de roofvogel en primaten ogen ....

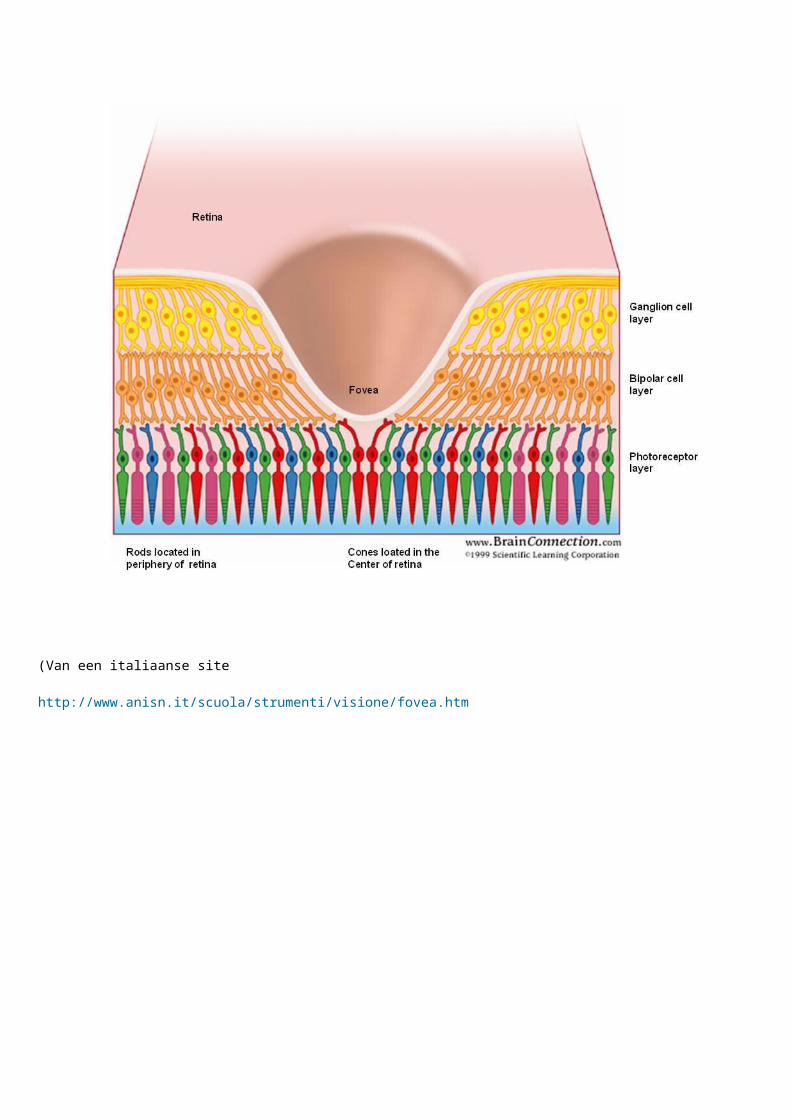
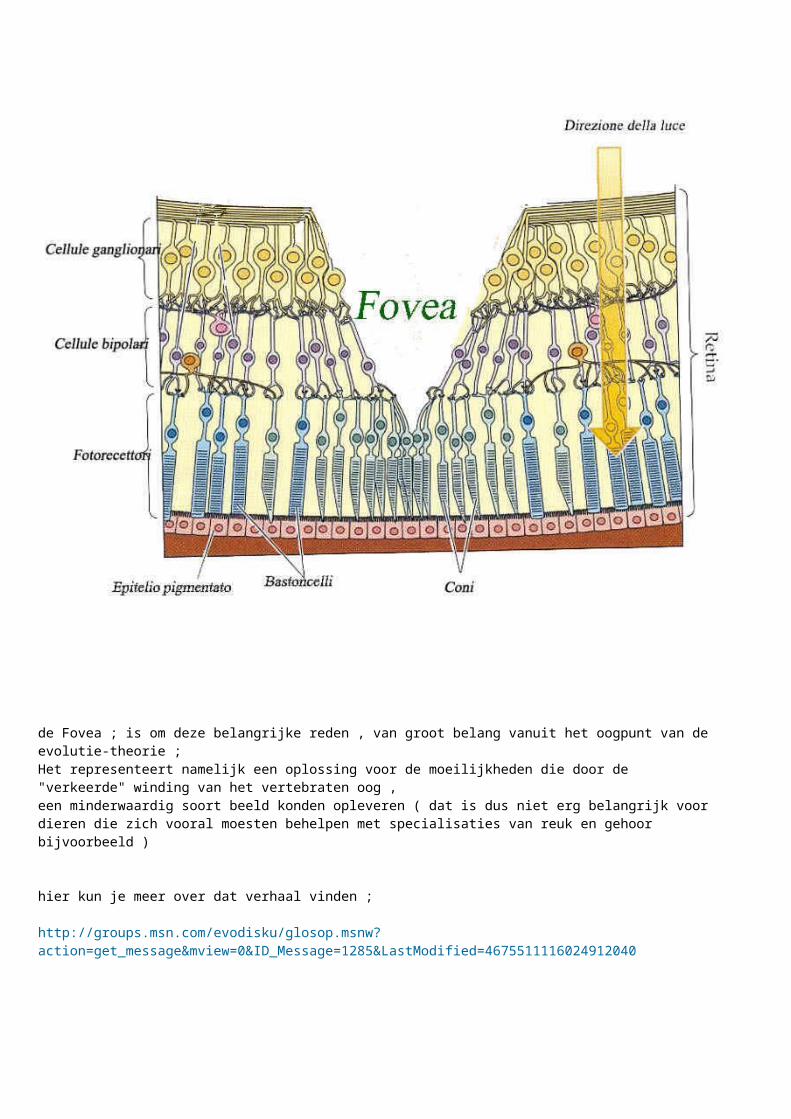
En dat is gebeurt door de ontwikkeling van de fovea ( bij roofvogels meerdere fovea"s http://ebiomedia.com/gall/eyes/sharp.html )
De fovea is een plaats in het oog waar de normale zenuwbedrading - laag die het inkomende licht plaatselijk hinderd , drastisch is verminderd in dikte , en met een hoge koncenytratie aan photocellen met als gevolg een grotere helderheid en scherpte op dat lichtinvals- "punt" .... http://brainconnection.positscience.com/topics/?main=gal/vision-home
(Van een italiaanse site
http://www.anisn.it/scuola/strumenti/visione/fovea.htm
de Fovea ; is om deze belangrijke reden , van groot belang vanuit het oogpunt van de evolutie-theorie ; Het representeert namelijk een oplossing voor de moeilijkheden die door de "verkeerde" winding van het vertebraten oog , een minderwaardig soort beeld konden opleveren ( dat is dus niet erg belangrijk voor dieren die zich vooral moesten behelpen met specialisaties van reuk en gehoor bijvoorbeeld )
hier kun je meer over dat verhaal vinden ;
http://groups.msn.com/evodisku/glosop.msnw?action=get_message&mview=0&ID_Message=1285&LastModified=4675511116024912040

Het gebeurt steeds weer ; de evolutie optimaliseerd bij voortduring de inrichtingen van de organismen die overleven ; zelfs wanneer er wordt vertrokken met een prutsonderdeel ___ net zoals dat gebeurt dmv evolutionaire algoritmes in bijvoorbeeld A(rtificial) L(ife) settings ____....Sommigen raken zelf zodanig goed ingespeeld op hun stabiele omgeving dat ze gedurende miljoenen jaren bijna niet meer schijnen te veranderen van globaal ( fossiel te konstateren ) ontwerp ... (- haaien , levende fossielen )
Misschien kunnen creationisten eens uitleggen waarom een GROTE INGENIEUR moest vertrekkken van een duidelijk slecht basisgegeven ( het omgekeerd gewonden vertedraten oog ) waar een paar hulpjes /acessoires moesten worden aan toegevoegd of een paar correcties aan uitgevoerd ( de fovea ) om weer toch maar weer iets bruikbaars te krijgen in het nieuwe model ... ipv meteen maar een GOED ontwerp in te bouwen in het nieuwe schepsel ..... Moest de Schepper gewoon ook alles uitproberen en uittesten door wat te prutsen aan zijn uitgangsmodellen .... en allerlei rude goldberg
oplossingen te bedenken , waarbij zelfs '"overschotjes " van vroegere pogingen bleven rondslingeren ?
( rood van mij ) Quote:tenzij zo'n complexe structuur ( hier worden dus in dit speciale geval de hersenen bedoeld ... toch ? ) aanwezig was voordat het vertebraten-oog evolueerde, zullen individueen met zo'n oog ondergegaan zijn in de selectiedruk
terloops ; er bestaan ook met vertebraten te vergelijken camera-ogen ZONDER de steun van / want niet- aanwezige hersenen : bij ...... kwallen

Moesten die " niet efficient " zijn ontworpen dan zouden ze waarschijnlijk wel moeten worden gecorrigeerd door een bijsturende hersenstructuur .... anders zouden ze verdwijnen ... weggeselecteerd Misschien heeft de ID-er wel een voorkeur voor kwallen met ogen ... of zijn misschien eerst de ogen ( en andere zintuigen ? ) onstaan en daarna de hersenen toegevoegd om ze te besturen en bij te sturen ?
P.S.
1.- Het valt me steeds weer op dat anti-evolutionisten , nogal graag uitspraken doen over "wetmatigheden " die altijd en overal gelden , ook wanneer ze het hebben over "eigenschappen " die voortdurend varieeren ook binnen de eigen extante populaties van de besproken soorten ( species ) .... en over ontwerpen voor "oplossingen van efficientie problemen " waarbij er blijkbaar niet wordt van uitgegaan dat een probleem op verschillende manieren en op grond van andere principes /uitgangsgegevens/ aapakken en/of beschikbare componenten ( en coincidenteele emergente van lokale wetten en interacties )kan worden opgelost ....
In werkelijkheid is dat allemaal veel minder strak , maar vloeiender en vooral veelvuldig en meervoudig op te lossen , startend vanuit verschillende principes en " fine tunings " (Net zoals ( naar analogie ) dezelfde technische functies en verlangde wensen kunnen worden vervuld door apparaten gebouwd met verschillende patenten en octrooien : diesel en bezineauto's , digitale en analoge opname apparatuur ; Pal en secam tV , microsoft en linux besturingssystemen ; om maar een paar voorbeelden te noemen )
Bovendien wordt daardoor het keuzeaanbod aan oplossingen plastischer , groter en geschikter onder diverse omgevings-omstandigheden .... Ogen zijn oeroud en minstens tussen de 40 a 60 x onhafhankelijk geevolueerd ...
Eveneens zijn ook lichtgevoelige cellen ( photocellen ) aan te treffen binnen het plantenrijk en verwacht ik ook biochemische overeenkomsten ( homeobox ? genetische coderingen ?)- tussen dieren en planten , te vinden
2.- Misschien is alles wel in elkaar geknutseld door een grote groep ID-ers en/of buitenaardse concurerende gelukszoekers en projectontwikkelaars ? een stam kabouters en elfen bijvoorbeeld ? Zou toch ook goed kunnen ? Zouden ze ergens hun handtekeningen en merknamen hebben gezet ? of door hun bestaan hun eigenste " morfogenetische veld " hebben ontwikkeld ?
Waarbij ik bij deze zo'n ( bovenstaande ) alternatieve" theorieen" ook graag zou zie opgenomen worden in het lespakket van de aankomende scholieren ... dat kan alleen maar een " open mind" instelling bevorderen .... en de verkoop van station-literatuur bevorderen ...bron (O;a.) ; (verdwenen) NaDarwin forum Zie ook volgende updates : More creationists misconception about eye-evolution http://pandasthumb.org/archives/2010/06/more-creationis.htmlOOG http://en.wikipedia.org/wiki/Eyehttp://cas.bellarmine.edu/tietjen/images/Eyes!.htmhttp://www.mun.ca/biology/dinnes/B2900/Jan14_files/frame.htmhttp://www.manandmollusc.net/advanced_introduction/gastro_eye.html
A: Simple pigment-cup style of Molluscan eye

B: Molluscan Lens Containing Eye
http://www.manandmollusc.net/advanced_introduction/cepheye.html
Cephalopod EyeSource: Brusca & Brusca 1990

http://cas.bellarmine.edu/tietjen/Ethology/simplenervoussystpix.htm http://cas.bellarmine.edu/tietjen/Ethology/light.gif
Turbellaria ogen
Vertebraten oog ---> http://wiki.cotch.net/index.php/Vertebrate_eyeOog-evolutie http://wiki.cotch.net/index.php/Eye_evolutionhttp://wiki.cotch.net/index.php/What_use_is_half_an_eye%3Fhttp://wiki.cotch.net/index.php/The_eye_is_too_complex_to_have_evolved

gradualisme en "half "-oog ZWEEPSTAARTJE en oogDr Koen Martens ( Eos juni 2006 )
Vele argumenten waarmee crearionisten evolutie betwisten komen terug in het design-discours.
Zo wordt aangevoerd dat het flagellum-systeem van baccerien te complex is om via natuurlijke selectie te ontstaan.
Dit argument moet eigenlijk gelezen worden als:
(1) het is te complex om in een keer en de novo (uit het niets) te ontstaan en (2) tussenstadia zullen ofwel niet functioneel zijn ofminstens selectief inferieur aan bestaande systemen zodat K niet kunnen overleven. Daarom, is dan de conclusie, moet het wel in een keer gecreeerd zijn. Door een intelligent wezen, uiteraard.Toen ik dit hoorde kreeg ik een deja vu. Jarenlang hebben creationisten exact hetzelfde argument gebruikt in verband met ons oog.
Het is zeer complex (dat is juist) en het is onmogelijk dat de tussenstadia nodig voor graduele evolutie van het oog functioneel en selectief voordelig zouden zijn. Dat laatste wcrd intussen volledig en grondig weerlegd. Computersimulaties hebben aangetoond dat graduele evolutie wel degelijk tot functionele tussenstadia kan leiden. --->Attachment: Nilson & Pelger.doc<--klikHer en der werden zelfs tussenstadia in verschillende groepen ontdekt. Maar wat het creationistisch argument voorgoed naar dromenland verwees, was de analyse van de oogstructuur in de verschillende groepen van de stam Mollusca (slakken, mossels, maar ook inktvissen). Alle stadia in de graduele evolutie naar een zo complex oog als dat van de mens kunnen hier teruggevonden worden. Van een eenvoudig lichtgevoelig pigmentlaagje, over een pigmentlaagje in een invaginatie van de huid (zodat de richting van licht kan worden waargenomen), naar een cenvoudige lensstructuur en ten slotte naar een complexe lensstructuur met spiertjes om verschillende focale velden te bereiken.
Al deze stadia komen bij bepaalde groepen van de Mollusca voor (zie figuur). http://bio1151.nicerweb.com/doc/class/bio1151/Locked/media/ch24/24_14MolluscEyes.jpg

Uiteindelijk was de bewijsvoering van de biologen zo compleet, dat de creationisten het argument volledig lieten vallen.
En dus komen de designers met iets anders aanzetten. Erg creatiefzijn ze niet (de woordspeling is opzettelijk).
Evolutie van het oog -argument http://134.184.200.76/forum/viewtopic.php?p=2088&highlight=#2088
Vreemd dat net ( ook tegenwoordige ) creationisten altijd het argument van de evolutie van het oog aanhalen. ( eigenlijk het onherleidbaarheids-argument ( IC) van Behe )Darwin had het probleem erkend en vanuit de evolutieleer verklaard(ja, toen al). Slakken==>gepigmenteerde vlek, inktvis==>holte met gepigmenteerde vlek, vis==>volledig afgesloten oog, talloze variaties op ogen bij reptielen, amfibieên

en zoogdieren... Er zijn tientallen tussenvormen van het oog bekend.
Zoöloog Dan-Eriik Nilsson schreef:
"When evolution skeptics want to attack Darwin's theory, they often point to the human eye. How could something so complex, they argue, have developed through random mutations and natural selection, even over millions of years? If evolution occurs through gradations, the critics say, how could it have created the separate parts of the eye -- the lens, the retina, the pupil, and so forth -- since none of these structures by themselves would make vision possible? In other words, what good is five percent of an eye? Darwin acknowledged from the start that the eye would be a difficult case for his new theory to explain. Difficult, but not impossible. Scientists have come up with scenarios through which the first eye-like structure, a light-sensitive pigmented spot on the skin, could have gone through changes and complexities to form the human eye, with its many parts and astounding abilities. Through natural selection, different types of eyes have emerged in evolutionary history -- and the human eye isn't even the best one, from some standpoints. Because blood vessels run across the surface of the retina instead of beneath it, it's easy for the vessels to proliferate or leak and impair vision. So, the evolution theorists say, the anti-evolution argument that life was created by an "intelligent designer" doesn't hold water: If God or some other omnipotent force was responsible for the human eye, it was something of a botched design. Biologists use the range of less complex light sensitive structures that exist in living species today to hypothesize the various evolutionary stages eyes may have gone through. Here's how some scientists think some eyes may have evolved: The simple light-sensitive spot on the skin of some ancestral creature gave it some tiny survival advantage, perhaps allowing it to evade a predator. Random changes then created a depression in the light-sensitive patch, a deepening pit that made "vision" a little sharper. At the same time, the pit's opening gradually narrowed, so light entered through a small aperture, like a pinhole camera. Every change had to confer a survival advantage, no matter how slight. Eventually, the light-sensitive spot evolved into a retina, the layer of cells and pigment at the back of the human eye. Over time a lens formed at the front of the eye. It could have arisen as a double-layered transparent tissue containing increasing amounts of liquid that gave it the convex curvature of the human eye. In fact, eyes corresponding to every stage in this sequence have been found in existing living species. The existence of this range of less complex light-sensitive structures supports scientists' hypotheses about how complex eyes like ours could evolve. The first animals with anything resembling an eye lived about 550 million years ago. And, according to one scientist's calculations, only 364,000 years would have been needed for a camera-like eye to evolve from a light-sensitive patch. "
Zien en Brein “Alhoewel de prestaties van de huidige computer hardware buitengewoon indrukwekkend zijn, is het duidelijk dat de specificaties van de menselijke retina ongeëvenaard zijn. Om 10 milliseconden (ms) van de volledige verwerking van zelfs maar een enkele zenuwcel van de retina te simuleren, moeten ongeveer 500 simultane niet-lineaire differentiaal vergelijkingen 100 voudig opgelost worden, hetgeen minstens enkele minuten rekentijd kost op een Cray supercomputer. Als we bedenken dat er 10 miljoen of meer van dergelijke cellen op complexe wijze met elkaar verweven zijn, dan zou het tenminste 100 jaar rekentijd op een Cray kosten om te simuleren wat er in het oog iedere seconde plaatsvindt.” (John K. Stevens, “Reverse Engineering the Brain,” Byte, April 1985, blz. 287.)
Vreemd dat creationisten zich analoog hiermee niet afvragen hoeveel computerrekenkracht het vraagt om het geluid van een scheet na te bootsen. Volgens mij lukt dat ook niet met een rechttoe-rechtaan lineaire vergelijking. Het nabootsen van een flatus gelatus is redelijk te doen, echter niet door eerst alle differentiaalvergelijkingen op te lossen en die vervolgens te programmeren. Analyseer het geluid en baseer daar je simulatie op. Evenzo is de creationistenvergelijking van de retinasignaalverwerking met een computersimulatie volkomen misplaatst.

Het zijn overigens niet de specificaties van de retina die zo indrukwekkend zijn, maar de hersenfuncties die de benodigde patroonherkenning leveren.Hoe de hersenen het presteren om te zien is nog altijd groot onbekend terrein.
Is het brein een computer? In de jaren zeventig pioneerde David Marr met die opvatting in zijn boek"Vision: A Computational Investigation into the Human Representation and Processing of Visual Information".
Niet geheel toevallig loopt dit idee parallel met de vooruitgang die men begon te boeken op het gebied van de robotica. Om die reden is een afwachtende houding misschien aangewezen, want parallelle overeenkomsten zouden wel eens het zicht kunnen belemmeren op wat er op biologisch vlak wérkelijk gebeurt.
Van één ding zijn we wel zeker. Zien is ook een leerproces. Wie door oogschade blind geboren is, zal, wanneer na een bepaalde leeftijd de oogfunctie is hersteld, toch blind blijven ook al scheelt er aan de hersenen verder niets.
Zie ik een dier of niet?http://noorderlicht.vpro.nl/noorderlog/
Een voorbeeld van de afbeeldingen die de computer kreeg voorgeschoteld (Thomas Serre, McGovern Institute for Brain Research at MIT). Dossiers
Het bizarre brein
Links
Link naar de originele publicatie van Tomaso Poggio en Aude Olivia in het tijdschrift PNAS. Lees ook: "Pakkende aandacht" (20 maart 2003)
Een nieuw computerprogramma kan net zo goed plaatjes sorteren als een mens. Niet zo gek? Toch wel, want zo makkelijk is dit kunstje niet voor een computer.

Een computerprogramma dat kan zeggen of er een dier op een foto staat of niet. Knap hoor, dat kan mijn kleine zusje ook. Maar computers vinden zo’n taak heel moeilijk. Mensen kunnen in een flits een beeld herkennen en erop reageren. Maar hoe doen de hersenen dit nou? Neurowetenschappers Tomaso Paggio en Aude Oliva van het McGovern Institute for Brain Research in Cambridge probeerden erachter te komen door een computerprogramma te schrijven dat ook dieren kan herkennen op foto’s. Daarvoor gebruikten ze informatie over de visuele cortex, het hersengebied waar alle informatie over beelden in de mens wordt verwerkt.
Het programma werd getraind met enkele honderden afbeeldingen van dieren en niet-dieren melden de wetenschappers in de online versie van het tijdschrift PNAS. Daarna moest de computer kunnen zeggen of er op de foto een dier stond of niet. En die taak was niet makkelijk. Dieren zijn natuurlijk erg verschillend. Sommige zijn klein, andere groot, soms staan ze in groepjes of vliegen ze in de lucht. Ook kon, net als in het echt, een dier ver weg of dichtbij staan, of was de foto van de zijkant genomen. Bovendien was de foto maar vijftig milliseconden te zien. In die tijd kan de mens wel zeggen of er een dier op staat of niet, maar is hij zich niet bewust van wat hij gezien heeft. Maar de computer kon het. Sterker nog, tijdens de test kon de computer net zo goed zien of er op een foto een dier stond, of niet, als een menselijk proefpersoon. Maar met dit computermodel is het raadsel rondom hoe mensen beelden verwerken niet ontrafeld.
Maar het geeft wel aan dat de onderzoekers op de goede weg zijn
Foto-cellen Alle planten bezitten minstens licht gevoelige cellen Niet alleen is fotosynthese afhankelijk van lichtgevoeligheid maar ook bepaalde bewegingen van planten zijn lichtgestuurd ---> Zonnebloemen zijn een zeer triviaal voorbeeld van bewegingen die zijn gestuurd door lichtgevoelige cellen (= detectie-apparatuur ) Müller cells transmitted light efficiently across an .... http://neurophilosophy.wordpress.com/2007/05/09/muller-cells-natures-fibre ...
Lichttropisme is trouwens ook een eigenschap van allerlei eencelligen
Denton vs Squid; the eye as suboptimal design.http://www.pandasthumb.org/archives/2006/11/denton_vs_squid.html
What Good is Half a Wing?Steven Dutch, Natural and Applied Sciences, University of Wisconsin - Green BayFirst-time Visitors: Please visit Site Map and Disclaimer. Use "Back" to return here.
http://www.uwgb.edu/dutchs/PSEUDOSC/HalfaWing.H
LINK Meer over oog- evolutie en ontwikkeling
Where d'you get those peepers http://www.firebirdmarchingband.com/~jmagee/APBiologySite/AP/Readings/eye%20evolution.pdf
Eye Morphogenesis http://dragon.zoo.utoronto.ca/~B03T0801B/eye.html

Moderne (ID)versies van "Mivart's probleem "
MIVART'S Problem1.- Quote mining het artikel van Gould, S. J. -
"Not Necessarily a Wing"
http://www.arn.org: http://www.arn.org/blogsq/index.php?title=gould_s_j_not_necessarily_a_wing&more=1&c=1&tb=1&pb=1
Voor creationisten en Id-ers gaat het bij het quoten uit dit artikel nu juist om het opzettelijke weglaten van de context van het citaat van Gould. Er wordt geciteerd uit de context: daarom is het een quote-mine i.p.v. een quote.
Zoals het hier wordt beschreven in het grote quotes-boek van de ARN org (ID-ots ) , lijkt Gould, “de beroemde paleontoloog”, het met St Georges Mivart eens te zijn.
Dat SJ GOULD dat niet is blijkt uit wat er vlak na het citaat in de oorspronkelijke tekst staat.
(Hier volgt het integrale artikel van Gould )
2.- http://www.stephenjaygould.org/library/gould_functionalshift.html
Not Necessarily a Wing
by Stephen Jay Gould
rom Flesh Gordon to Alex in Wonderland, title parodies have been a stock-in-trade of low comedy. We may not anticipate a tactical similarity between the mayhem of Mad magazine's movie reviews and the titles of
major scientific works, yet two important nineteenth-century critiques of Darwin parodied his most famous phrases in their headings.
In 1887, E. D. Cope, the American paleontologist known best for his fossil feud with O. C. Marsh but a celebrated evolutionary theorist in his own right, published The Origin of the Fittest—a takeoff on Herbert Spencer's phrase, borrowed by Darwin as the epigram for natural selection: survival of the fittest. (Natural selection, Cope argued, could only preserve favorable traits that must arise in some other manner, unknown to Darwin. The fundamental issue of evolution cannot be the differential survival of adaptive traits, but their unexplained origin-hence the title parody.)
St. George Mivart (1817-1900), a fine British zoologist, tried to reconcile his unconventional views on religion and biology but ended his life in tragedy, rejected by both camps. At age seventeen, he abandoned his Anglican upbringing, became a Roman Catholic, and consequently (in a less tolerant age of state religion) lost his opportunity for training in natural history at Oxford or Cambridge. He became a lawyer but managed to carve out a distinguished career as an anatomist nonetheless. He embraced evolution and won firm support from the powerful T. H. Huxley, but his strongly expressed and idiosyncratic anti-Darwinian views led to his rejection by the biological establishment of Britain. He tried to unite his biology with his religion in a series of books and essays, and ended up excommunicated for his trouble six weeks before his death.
Cope and Mivart shared the same major criticism of Darwin—that natural selection could explain the preservation and increase of favored traits but not their origin. Mivart, however, went gunning for a higher target than Darwin's epigram. He shot for the title itself, naming his major book (1871) On the Genesis of Species. (Darwin, of course, had called his classic On the Origin of Species.)
Mivart's life may have ended in sadness and rejection thirty years later, but his Genesis of Species had a major impact in its time. Darwin himself offered strong, if grudging, praise and took Mirvart far more seriously than

any other critic, even adding a chapter to later editions of the Origin of Species primarily to counter Mivart's attack. Mivart gathered, and illustrated "with admirable art and force" (Darwin's words), all objections to the theory of natural selection—"a formidable array" (Darwin's words again). Yet one particular theme, urged with special attention by Mivart, stood out as the centerpiece of his criticism. This argument continues to rank as the primary stumbling block among thoughtful and friendly scrutinizers of Darwinism today. No other criticism seems so troubling, so obviously and evidently "right" (against a Darwinian claim that seems intuitively paradoxical and improbable).
Mivart awarded this argument a separate chapter in his book, right after the introduction. He also gave it a name, remembered ever since. He called his objection "The Incompetency of 'Natural Selection' to Account for the Incipient Stages of Useful Structures." If this phrase sounds like a mouthful, consider the easy translation: We can readily understand how complex and fully developed structures work and how their maintenance and preservation may rely upon natural selection—a wing, an eye, the resemblance of a bittern to a branch or of an insect to a stick or dead leaf. But how do you get from nothing to such an elaborate something if evolution must proceed through a long sequence of intermediate stages, each favored by natural selection? You can't fly with 2 percent of a wing or gain much protection from an iota's similarity with a potentially concealing piece of vegetation. How, in other words, can natural selection explain the incipient stages of structures that can only be used in much more elaborated form?
I take up this old subject for two reasons. First, I believe that Darwinism has, and has long had, an adequate and interesting resolution to Mivart's challenge (although we have obviously been mightily unsuccessful in getting it across). Second, a paper recently published in the technical journal Evolution has provided compelling experimental evidence for this resolution applied to its most famous case—the origin of wings.
The dilemma of wings—the standard illustration of Mivart's telling point about incipient stages—is set forth particularly well in a perceptive letter that I recently received from a reader, a medical doctor in California. He writes:
How does evolutionary theory as understood by Darwin explain the emergence of items such as wings, since a small move toward a wing could hardly promote survival? I seem to be stuck with the idea that a significant quality of wing would have to spring forth all at once to have any survival value.
Interestingly, my reader's proposal that much or most of the wing must arise all at once (because incipient stages could have no adaptive value) follows Mivart's own resolution. Mivart first enunciated the general dilemma (1871, p. 23):
Natural selection utterly fails to account for the conservation and development of the minute and rudimentary beginnings, the slight and infinitesimal commencements of structures, however useful those structures may afterwards become.
After fifty pages of illustration, he concludes: "Arguments may yet be advanced in favor of the view that new species have from time to time manifested themselves with suddenness, and by modifications appearing at once." Advocating this general solution for wings in particular, he concludes (p. 107): "It is difficult, then, to believe that the Avian limb was developed in any other way than by a comparatively sudden modification of a marked and important kind."
Darwin's theory is rooted in the proposition that natural selection acts as the primary creative force in evolutionary change. This creativity will be expressed only if the fortuitous variation forming the raw material of evolutionary change can be accumulated sequentially in tiny doses, with natural selection acting as the sieve of acceptance. If new species arise all at once in an occasional lucky gulp, then selection has no creative role. Selection, at best, becomes an executioner, eliminating the unfit following this burst of good fortune. Thus, Mivart's solution—bypassing incipient stages entirely in a grand evolutionary leap—has always been viewed, quite rightly, as an anti-Darwinian version of evolutionary theory.
Darwin well appreciated the force, and potentially devastating extent, of Mivart's critique about incipient stages. He counterattacked with gusto, invoking the standard example of wings and arguing that Mivart's solution of sudden change presented more problems than it solved—for how can we believe that so complex a structure

as a wing, made of so many coordinated and co-adapted parts, could arise all at once:
He who believes that some ancient form was transformed suddenly through an internal force or tendency into, for instance, one furnished with wings, will be…compelled to believe that many structures beautifully adapted to all the other parts of the same creature and to the surrounding conditions, have been suddenly produced; and of such complex and wonderful co-adaptations, he will not be able to assign a shadow of an explanation.…To admit all this is, as it seems to me, to enter into the realms of miracle, and to leave those of Science.
(This essay must now go in other directions but not without a small, tangential word in Mivart's defense. Mivart did appreciate the problem of complexity and coordination in sudden origins. He did not think that any old complex set of changes could arise all at once when needed—that would be tantamount to miracle. Most of Mivart's book studies the regularities of embryology and comparative anatomy to learn which kinds of complex changes might be possible as expressions and elaborations of developmental programs already present in ancestors. He advocates these changes as possible and eliminates others as fanciful.)
Darwin then faced his dilemma and developed the interestingly paradoxical resolution that has been orthodox ever since (but more poorly understood and appreciated than any other principle in evolutionary theory). If complexity precludes sudden origin, and the dilemma of incipient stages forbids gradual development in functional continuity, then how can we ever get from here to there? Darwin replies that we must reject an unnecessary hidden assumption in this argument—the notion of functional continuity. We will all freely grant that no creature can fly with 2 percent of a wing, but why must the incipient stages be used for flight? If incipient stages originally performed a different function suited to their small size and minimal development, natural selection might superintend their increase as adaptations for this original role until they reached a stage suitable for their current use. In other words, the problem of incipient stages disappears because these early steps were not inadequate wings but well-adapted something-elses. This principle of functional change in structural continuity represents Darwin's elegant solution to the dilemma of incipient stages.
Darwin, in a beau geste of argument, even thanked Mivart for characterizing the dilemma so well—all the better to grant Darwin a chance to elaborate his solution. Darwin writes: "A good opportunity has thus been afforded [by Mivart] for enlarging a little on gradations of structure, often associated with changed functions—an important subject, which was not treated at sufficient length in the former editions of this work." Darwin, who rarely added intensifiers to his prose, felt so strongly about this principle of functional shift that he wrote: "In considering transitions of organs, it is so important to bear in mind the probability of conversion from one function to another."
Darwin presented numerous examples in Chapters 5 and 7 of the final edition of the Origin of Species. He discussed organs that perform two functions, one primary, the other subsidiary, then relinquish the main use and elaborate the formerly inconspicuous operation. He then examined the flip side of this phenomenon—functions performed by two separate organs (fishes breathing with both lungs and gills). He argues that one organ may assume the entire function, leaving the other free for evolution to some other role (lungs for conversion to air bladders, for example, with respiration maintained entirely by gills). He does not, of course, neglect the classic example of wings, arguing that insects evolved their organs of flight from tracheae (or breathing organs—a minority theory today, but not without supporters). He writes: "It is therefore highly probable that in this great class organs which once served for respiration have been actually converted into organs of flight."
Darwin's critical theory of functional shift, usually (and most unfortunately) called the principle of "preadaptation,"[1] has been with us for a century. I believe that this principle has made so little headway not only because the basic formulation seems paradoxical and difficult, but mainly because we have so little firm, direct evidence for such functional shifts. Our technical literature contains many facile verbal arguments—little more than plausible "just-so" stories. The fossil record also presents some excellent examples of sequential development through intermediary stages that could not work as modern organs do—but we lack a rigorous mechanical analysis of function at the various stages.
Let us return, as we must, to the classic case of wings. Archaeopteryx, the first bird, is as pretty an intermediate as paleontology could ever hope to find—a complex mélange of reptilian and avian features. Scientists are still debating whether or not it could fly. If so, Archaeopteryx worked like the Wrights' biplane to a modern eagle's Concorde. But what did the undiscovered ancestors of Archaeopteryx do with wing rudiments that surely could

not produce flight? Evolutionists have been invoking Darwin's principle of functional shift for more than 100 years, and the list of proposals is long. Proto-wings have been reconstructed as stabilizers, sexual attractors, or insect catchers. But the most popular hypothesis identifies thermoregulation as the original function of incipient stages that later evolved into feathered wings. Feathers are modified reptilian scales, and they work very well as insulating devices. Moreover, if birds evolved from dinosaurs (as most paleontologists now believe), they arose from a lineage particularly subject to problems with temperature control. Archaeopteryx is smaller than any dinosaur and probably arose from the tiniest of dinosaur lineages. Small animals, with high ratios of surface area to volume, lose heat rapidly and may require supplementary devices for thermoregulation. Most dinosaurs could probably keep warm enough just by being large. Surface area (length length, or length squared) increases more slowly than volume (length length length, or length cubed) as objects grow. Since animals generate heat over their volumes and lose it through their surfaces, small animals (with their relatively large surface areas) have most trouble keeping warm.
There I go again—doing what I just criticized. I have presented a plausible story about thermoregulation as the original function of organs that later evolved into wings. But science is tested evidence, not tall tales. This lamentable mode of storytelling has been used to illustrate Darwin's principle of functional shift only faute de mieux—because we didn't have the goods so ardently desired. At least until recently, when my colleagues Joel G. Kingsolver and M. A. R. Koehl published the first hard evidence to support a shift from thermoregulation to flight as a scenario for the evolution of wings. They studied insects, not birds—but the same argument has long been favored for nature's smaller and far more abundant wings (see their article, "Aerodynamics, Thermoregulation, and the Evolution of Insect Wings: Differential Scaling and Evolutionary Change," in Evolution, 1985).
In preparing this essay, I spent several days reading the classical literature on the evolution of insect flight—and emerged with a deeper understanding of just how difficult Darwin's principle of functional shift can be, even for professionals. Most of the literature hasn't even made the first step of applying functional shift at all; not to mention the later reform of substituting direct evidence for verbal speculation. Most reconstructions are still trying to explain the incipient stages of insect wings as somehow involved in airborne performance from the start—not for flapping flight, of course, but still for some aspect of motion aloft rather than, as Darwin's principle would suggest, for some quite different function.
To appreciate the dilemma of such a position (so well grasped by Mivart more than 100 years ago), consider just one recent study (probably the best and most widely cited) and the logical quandaries that a claim of functional continuity entails. In 1964, J. W. Flower presented aerodynamic arguments or wings evolved from tiniest rudiment to elaborate final form in the interest of airborne motion. Flower argues, supporting an orthodox view, that wings evolved from tiny outgrowths of the body used for gliding prior to elaboration for sustained flight. But Flower recognizes that these incipient structures must themselves evolve from antecedents too small to function as gliding planes. What could these very first, slight outgrowths of the body be for? Ignoring Darwin's principle of functional shift, Flower searches for an aerodynamic meaning even at this very outset. He tries to test two suggestions: E. H. Hinton's argument that initial outgrowths served for "attitude control," permitting a falling insect to land in a suitable position for quick escape from predators; and a proposal of the great British entomologist Sir Vincent Wigglesworth (wonderful name for an insect man, I always thought) that such first stages might act as stabilizing or controlling devices during takeoff in small, passively aerial insects.
Flower proceeded by performing aerodynamic calculations on consequences of incipient wings for simple body shapes when dropped—and he quickly argued himself into an inextricable logical corner. He found, first of all, that tiny outgrowths might help, as Wigglesworth, Hinton, and others had suggested. But the argument foundered on another observation: The same advantages could be gained far more easily and effectively by another, readily available alternative route—evolution to small size (where increased surface/volume ratios retard failing and enhance the probability of takeoff). Flower then realized that he would have to specify a reasonably large body size for incipient wings to have any aerodynamic effect. But he then encountered another problem: At such sizes, legs work just as well as, if not better than, proto-wings for any suggested aerodynamic function. Flower admitted:
The first conclusion to be drawn from these calculations is that the selective pressure in small insects is towards smaller insects, which would have no reason to evolve wings.
I would have stopped and searched elsewhere (in Darwin's principle of functional shift) at this point, but Flower

bravely continued along an improbable path:
The main conclusions, however, are that attitude control of insects would be by the use of legs or by very small changes in body shape [i.e., by evolving small outgrowths, or proto-wings].
Flower, in short, never considered an alternative to his assumption of functional continuity based upon some aspect of aerial locomotion. He concluded
At first they [proto-wings] would affect attitude; later they could increase to a larger size and act as a true wing, providing lift in their own right. Eventually they could move, giving the insect greater maneuverability during descent, and finally they could "flap," achieving sustained flight.
As an alternative to such speculative reconstructions that work, in their own terms, only by uncomfortable special pleading, may I suggest Darwin's old principle of functional shift (preadaptation—ugh—for something else). The physiological literature contains voluminous testimony to the thermodynamic efficiency of modern insect wings: in presenting, for example, a large surface area to the sun for quick heating (see B. Heinrich, 1981). If wings can perform this subsidiary function now, why not suspect thermoregulation as a primary role at the outset? M. M. Douglas (1981), for example, showed that, in Colias butterflies, only the basal one-third of the wing operates in thermoregulation—an area approximately equal to the thoracic lobes (proto-wings) of fossil insects considered ancestral to modern forms.
Douglas then cut down some Colias wings to the actual size of these fossil ancestral lobes and found that insects so bedecked showed a 55 percent greater increase in body temperature than bodies deprived of wings entirely. These manufactured proto-wings measured 5 by 3 millimeters on a body 15 millimeters long. Finally, Douglas determined that no further thermoregulatory advantage could be gained by wings longer than 10 millimeters on a 15-millimeter body.
Kingsolver and Koehl performed a host of elaborate and elegant experiments to support a thermoregulatory origin of insect proto-wings. As with so many examples of excellent science producing clear and interesting outcomes, the results can be summarized briefly and cleanly.
Kingsolver and Koehl begin by tabulating all the aerodynamic hypotheses usually presented in the literature as purely verbal speculations. They arrange these proposals of functional continuity (the explanations that do not follow Darwin's solution of Mivart's dilemma) into three basic categories: proto-wings for gliding (aerofoils for steady-state motion), for parachuting (slowing the rate of descent in a falling insect), and attitude stability (helping an insect to land right side up). They then transcended the purely verbal tradition by developing aerodynamic equations for exactly how proto-wings should help an insect under these three hypotheses of continuity in adaptation (increasing the lift/drag ratio as the major boost to gliding, increasing drag to slow the descent rate in parachuting, measuring the moment about the body axis produced by wings for the hypothesis of attitude stability).
They then constructed insect models made of wire, epoxy, and other appropriate materials to match the sizes and body shapes of flying and nonflying forms among early insect fossils. To these models, they attached wings (made of copper wire enclosing thin, plastic membranes) of various lengths and measured the actual aerodynamic effects for properties predicted by various hypotheses of functional continuity. The results of many experiments in wind tunnels are consistent and consonant: Aerodynamic benefits begin for wings above a certain size, and they increase as wings get larger. But at the small sizes of insect proto-wings, aerodynamic advantages are absent or insignificant and do not increase with growing wing length. These results are independent of body shape, wind velocity, presence or placement of legs, and mounting position of wings. In other words, large wings work well and larger wings work better—but small wings (at the undoubted sizes of Mivart's troubling incipient stages) provide no aerodynamic edge.

The thermoregulatory (upper curve) and aerodynamic (lower curve) Advantages for increasing wing length in insects. Note that thermodynamic benefits accrue rapidly when the wing is very small (too small for flight), but scarcely increase at all for wings of larger size. Aerodynamic advantages, on the other hand, are insignificant for small size, but increase rapidly at larger wing dimensions, just as the thermodynamic benefits cease. Ben Gamit. Adapted from Joe Lemonnier. Courtesy of Natural History.
Kingsolver and Koehl then tested their models for thermoregulatory effects, constructing wings from two materials with different thermal conductivities (construction paper and aluminum foil) and measuring the increased temperature of bodies supplied with wings of various lengths versus wingless models. They achieved results symmetrically opposite to the aerodynamic experiments. For thermoregulation, wings work well at the smallest sizes, with benefits increasing as the wing grows. However, beyond a measured length, further increase of the wing confers no additional effect. Kingsolver and Koehl conclude:
At any body size, there is a relative wing length above which there is no additional thermal effect, and below which there is no significant aerodynamic effect.
The accompanying chart illustrates these combined results. Note how the thermoregulatory effect of excess body temperature due to wings (solid line) increases rapidly at small wing sizes but not at all above an intermediate wing length. Conversely, the aerodynamic effect of lift/drag ratio does not increase at all until intermediate wing length, but grows rapidly thereafter.
We could not hope for a more elegant experimental confirmation of Darwin's solution to Mivart's challenge. Kingsolver and Koehl have actually measured the functional shift by showing that incipient wings aid thermoregulation but provide no aerodynamic benefit—while larger wings provide no further thermoregulatory oomph but initiate aerodynamic advantage and increase the benefits steadily thereafter. The crucial intermediate wing length, where thermoregulatory gain ceases and aerodynamic benefits begin, represents a domain of functional shift, as aerodynamic advantages pick up the relay from waning thermoregulation to continue the evolutionary race to increasing wing size.
But what might push an insect across the transition? Why reach this crucial domain at all? If wings originally worked primarily for thermoregulation, why not just stop as the length of maximum benefit approached? Here, Kingsolver and Koehl present an interesting speculation based on another aspect of their data. They found that the domain of transition between thermal and aerial effects varied systematically with body size: The larger the body, the sooner the transition (in terms of relative wing length). For a body 2 centimeters long, the transition occurred with wings 40 to 60 percent of body length; but a 10-centimeter body switches to aerodynamic advantage at only 10 percent of body length.
Now suppose that incipient ancestral wings worked primarily for thermoregulation, and had reached a stable, optimum size for greatest benefit. Natural selection would not favor larger wings and a transition to the available domain of aerodynamic advantage. But if body size increased for other reasons, an insect might reach the realm of aerial effects simply by growing larger, without any accompanying change of body shape or relative wing length.
We often think, naively, that size itself should make no profound difference. Why should just more of the same have any major effect beyond simple accumulation? Surely, any major improvement or alteration must require an extensive and explicit redesign, a complex reordering of parts with invention of new items.
Nature does not always match our faulty intuitions. Complex objects often display the interesting and paradoxical property of major effect for apparently trifling input. Internal complexity can translate a simple

quantitative change into a wondrous alteration of quality. Perhaps that greatest and most effective of all evolutionary inventions, the origin of human consciousness, required little more than an increase of brain power to a level where internal connections became rich and varied enough to force this seminal transition. The story may be much more complex, but we have no proof that it must be.
Voltaire quipped that "God is always for the big battalions." More is not always better, but more can be very different.
Notes
1. This dreadful name has made a difficult principle even harder to grasp and understand. Preadaptation seems to imply that the proto-wing, while doing something else in its incipient stages, knew where it was going—predestined for a later conversion to flight. Textbooks usually introduce the word and then quickly disclaim any odor of foreordination. (But a name is obviously ill-chosen if it cannot be used without denying its literal meaning.) Of course, by "preadaptation" we only mean that some structures are fortuitously suited to other roles if elaborated, not that they arise with a different future use in view-now there I go with the standard disclaimer. As another important limitation, preadaptation does not cover the important class of features that arise without functions (as developmental consequences of other primary adaptations, for example) but remain available for later co-optation. I suspect, for example, that many important functions of the human brain are co-opted consequences of building such a large computer for a limited set of adaptive uses. For these reasons, Elizabeth Vrba and I have proposed that the restrictive and confusing word "preadaptation" be dropped in favor of the more inclusive term "exaptation"—for any organ not evolved under natural selection for its current use—either because it performed a different function in ancestors (classical preadaptation) or because it represented a nonfunctional part available for later co-optation. See our technical article, "Exaptation: A Missing Term in the Science of Form," Paleobiology, 1981.
[ Stephen Jay Gould "Not Necessarily a Wing," Natural History 94 (October 1985): 12-25; Reprinted here with permission from Bully for Brontosaurus, New York: W. W. Norton & Co., 1991, pp. 139-151. ]
CONCLUSIE ?De quote miner noemt een probleem, maar hij vermeld de aangedragen oplossingen niet .
Gould beschrijft een theoretisch kader dat het probleem van Mivart oplost en geeft er een praktische invulling bij. Daar kan een creaqionist het mee oneens zijn, maar dan moet die ook laten zien waarom.
Teksten kunnen taalkundig kloppen maar inhoudelijk (of journalistiek) over de grens gaan. Misquoted tekststen zijn meestal suggestief en onthouden de lezer belangrijke informatie (nl. dat er tegengeluiden zijn).
adaptaties
Het perfectie-probleem : het verklaren van adaptatieshttp://www.uia.ac.be/u/rvdamme/evobio/h32.doc
To suppose that the eye, with all its inimitable contrivances ... could have been formed by natural selection, seems, I freely confess, absurd in the highest degree. - C. R. Darwin, The Origin.(1*)
Eén van de eerste en belangrijkste kritieken op Darwin’s theorie van evolutie door natuurlijke selectie was het zogenaamde “Argument from design” .

Invloedrijke Geologen / biologen als Georges Cuvier (1769-1832, directeur van het Natuurhistorisch Museum te Parijs en zowat de grondlegger van de vergelijkende anatomie en de paleontologie = en tevens grondlegger van het catastrofisme ), zijn Engelse leerling Richard Owen(1804-1892) en St. George Jackson Mivart(1827-1900) voerden aan dat
.....de fundamentele eenheid van functie en vorm in de anatomie van dieren zo subtiel was, en zo’n stabiliteit vereiste, dat het elke vorm van evolutie uitsloot. ....
* ( In feite in moderne vorm --> het onherleidbaarheids- argument ( IC) van Behe die ook beweerd dat complexe organismen niet kunnen geevolueerd zijn )
De inwendige complexiteit van organismen waren voor hen een bewijs dat er een Ontwerper aan het werk geweest was.
In een vaak geciteerd voorbeeld maakt de priester-wetenschapper William Paley de vergelijking tussen de complexiteit van organismen en die van uurwerken.
Het vinden van een uurwerk op de grond impliceert ook het bestaan van een horlogemaker, zo gaat het argument.
---> Het zou belachelijk zijn te veronderstellen dat de horloge-onderdelen zich spontaan zouden verzameld hebben en aanleiding gegeven zouden hebben tot de exacte configuratie nodig om een uurwerk te laten werken.
----> Op dezelfde manier vormt de verregaande complexiteit van organismen het bewijs voor het bestaan van een Creator.
(1*) Darwin was van origine/opleiding theoloog en was zeker wél bekend met de"natural theology "en het argument van design van Paley ... Darwin onwikkelde juist een theorie die natuurlijke evolutie en een natuurlijk mechanisme(n)introduceerde en zodoende een antwoord vormde op dat argument

......Het verklaren van adaptaties, vooral van ingewikkelde adaptaties die de gecoördineerde evolutie van vele verschillende onderdelen impliceert, stond en staat bovenaan de agenda van evolutionaire biologen.
Toeval én selectie
Toeval neemt in de theorie van evolutie door natuurlijke selectie een belangrijke plaats in. Darwin’s suggestie dat de toevallige variaties tussen organismen aan de basis liggen van alle evolutionaire gebeurtenissen, stuitte aanvankelijk op groot ongeloof en een storm van protest. Hoewel de ontwikkelingen binnen de genetica Darwins vermoeden ondertussen bevestigden, blijft hetzelfde argument regelmatig opduiken.
Een molecule als hemoglobine bestaat uit 146 aminozuren. De kans dat in een soepje, samengesteld uit een mengsel van de 20 bestaande aminozuren, door toeval hemoglobine zou ontstaan, is ontzaglijk klein (ongeveer 1 kans op 10190).
Een dergelijke voorstelling van evolutie door natuurlijke selectie verliest echter de tweede component van het mechanisme uit het oog. Na het ontstaan van de variatie -inderdaad een puur toevallig gebeuren- start het (hersenloos proces van ) onverbiddelijke trieëren en uitkiezen van de variaties - en in dit onderdeel van het proces speelt toeval weinig of geen rol.
In het voorbeeld van de hemoglobine-molecule wordt het selectieproces voorgesteld als een enkele zeef; men blijft het ruwe product uitzeven tot "per toeval " de adaptatie, in al zijn glorie, tevoorschijn komt.
Maar natuurlijke selectie werkt niet als een enkelvoudige zeef, het werkt cumulatief. Het eindproduct van één sessie is het beginproduct van de volgende selectiecyclus. Hieronder volgt een beroemd geworden parallel van Richard Dawkins (1987).
Me thinks it is like a weaselVeronderstellen dat toeval zal leiden tot complexe adaptaties als het oog, zo beweren de tegenstanders van natuurlijke selectie smalend, is even dom als geloven dat een aap op een typemachine de werken

van Shakespeare kan reproduceren.
De kans dat puur toevallige toetsaanslagen zullen leiden tot zelfs maar 챕챕 n vers is inderdaad al zéér klein. Stel je voor dat we zullen wachten tot de aap de volgende regel uit Hamlet heeft geproduceerd :
“Methinks it is like a weasel”
(Hamlet vindt dat een voorbijdrijvende wolk meer op een wezel gelijkt dan op een kameel, zoals de hofmaarschalk Polonius beweert). Het zinnetje omvat 28 in te vullen plaatsen, en met 27 toetsen (alle letters plus de spatiebalk), bedraagt de kans dat deze zin perfect gereproduceerd wordt (1/27)28. Zelfs met een zeer snel typende aap zullen we bijna oneindig lang moeten wachten.
Veranderen we nu echter het experiment een beetje, zodat het beter het proces van cumulatieve selectie imiteert. We starten met een compleet willekeurige sequentie van 28 letters en spaties, bijvoorbeeld
wdlmnlt dtjbkwirzrezlmqco P
Nu kweken we verder met dit zinnetje, door er massa’s kopietjes van te maken, met een zekere kans op een willekeurig foutje (een mutatie). Uit al deze kopietjes kiezen we die kopie eruit die het best op de uiteindelijk te bekomen zin lijkt. In onze simulatie was het winnende zinnetje
wdltmnt dtjbswirzrezlmqco p
Niet echt een spectaculaire verbetering, maar kijk hoe snel evolutie door selectie voortsnelt :
na 10 generaties : mdldmns itjiswhrzRez mecs
na 20 generaties : meldinls it iswprke z wecse
na 30 generaties : methings it iswlike b wecse
na 40 generaties : methinks it is like i weasel
Na 43 generaties werd Hamlet’s repliek perfect gereproduceerd. Dit voorbeeld illustreert het verschil tussen cumulatieve selectie (waarbij elke kleine verbetering gebruikt wordt als basis voor verdere constructie) en eenstaps-selectie (waarbij telkens van nul herbegonnen wordt). Bovengenoemde critici verloren uit het oog dat toevallige variatie slechts één onderdeel is in de theorie van natuurlijke selectie. Na het ontstaan van die variatie grijpt telkens selectie plaats, die bij uitstek niet willekeurig is. Adaptaties zijn het gecumuleerde effect van die selectie.
LINKS
APPLET ---> http://home.pacbell.net/s-max/scott/weasel.html ---> http://home.pacbell.net/s-max/scott/weasel_alt.html
http://www.talkorigins.org/indexcc/CF/CF011_1.html
Complexity/ AIhttp://myxo.css.msu.edu/papers/nature2003/
The evolutionary origin of complex features

Richard E. Lenski
*, Charles Ofria†, Robert T. Pennock‡ & Christoph AdamiëDepartment of Microbiology & Molecular Genetics, † Department of Computer Science & Engineering, and ‡ Lyman Briggs School & Department of Philosophy,
Michigan State University, East Lansing, Michigan 48824, USA
Digital Life Laboratory, California Institute of Technology, Pasadena, California 91125, USAhttp://myxo.css.msu.edu/papers/nature2003/Nature03_Complex.pdfArtificial Life Experiments Show How Complex Functions Can Evolve http://www.sciencedaily.com/releases/2003/05/030508075843.htm
Vroege kritiek op DARWIN's Theorie
St George Mivart, (1827-1900) een zoöloog beargumenteerde dat organen onmogelijk zouden kunnen zijn geêvolueerd. Welk nut, immers, zou men kunnen halen uit een orgaan dat maar half gevormd was? Darwins antwoord hierop was tweeledig. *Enerzijds kunnen organen een andere functie vervuld hebben (bv wordt er gesuggereerd dat vleugels eerst zouden gevormd zijn om afkoeling te bezorgen en dat de vliegfunctie pas later kwam). *Anderzijds dat niet zozeer het nut van een orgaan bepalend is voor het overleven, maar wel het extra voordeel dat het biedt ten opzichte van organismen waar het nog slechter ontwikkeld is. Nemen we als voorbeeld het oog. Het moderne oog van zoogdieren is in staat om een perfect scherp beeld te geven. Wat nut heeft een oog dat onscherpe beelden geeft? Of moet het oog ineens in zijn volmaaktheid gevormd zijn? Dus toch door een creator? Het darwinistisch antwoord hierop luidt als volgt: het oog is lang geleden ontstaan als een groepje cellen dat toevallig lichtgevoelig werd. Deze lichtgevoeligheid bood een voordeel tegenover organisme die deze eigenschap nog niet hadden. Later evolueerde dit naar de mogelijkheid om lichtnuances vanuit een bepaalde richting te kunnen opmerken. Hiervoor moest het laagje lichtgevoelige cellen een lichte holling vormen. Naarmate deze uitholling meer uitgesproken werd, verbeterde de scherpte van het zicht. Dit bood alsmaar meer overlevingskansen en werd dus door gegeven aan de volgende generaties. We zouden het ook als volgt kunnen samenvatten. Hoe snel moet een konijn kunnen lopen? Sneller dan de vos? Neen, gewoon sneller dan het konijn naast hem!!
(Creationist )
EvoTheorie laat (een individuele soort-vertegenwoordiger )niet toe om te muteren op het moment dat het nodig is.Zodra er zich een nood-situatie voordoet moet de juiste "tool" al "per ongeluk "(= toevallig ) aanwezig zijn."

m.a.w. De tool moet al ( in aanleg )aanwezig zijn in de populatie anders kun je het niet gaan selecteren.
In de meeste gevallen ontstaat een geperfectioneerde "tool" pas wanneer er langdurig zwakke selektiedruk was die een bepaalde tendens of eigenschap optimaliseerde Echter wel als een continu proces, dus in kleine stapjes, en gestuurd door de omgevingscondities (in de brede zin van het woord)... en eigenlijk dus als voortdurende optimalisatie van een eigenschap die een zwakke maar continu bedreiging beter aankan , wat dus een geleidelijk fitnes voordeel bied tijdens de opeenvolging van generaties . Kleine stapjes dus en gestuurd door omgevingscondities
Kan het complexe menselijke oog stapsgewijs zijn geëvolueerd, of moet het zijn ontworpen door een Intelligente Ontwerper? http://noorderlicht.vpro.nl/afleveringen/23314456/
Larven van de glaszakpijp doen nog wel aan gewervelde dieren denken, maar volwassen zakpijpen zijn sterk gedegenereerd.
Lichtsensor-gen van zakglaspijp actief in oogzenuw kikker. (foto Ron Dirks, Radboud Universiteit Nijmegen)
*** De Nijmeegse onderzoekster Lettie Lubsen over de zakpijp en de geheimzinnige evolutie van het menselijk oog.
Audio
Noorderlicht Radio (2005) 27 sept 2005
HET OOG VAN DE ZAKPIJP
Het menselijk oog zit zo ingewikkeld in elkaar, dat het pas functioneert als alle onderdelen op hun plek zitten. Aan een half oog heb je immers niks en daarom, zeggen aanhangers van de 'Intelligent Design theorie', kan het oog

nooit stapsgewijs zijn ge 챘 volueerd. Maar zij rekenden buiten de zakpijp. Dit ongewervelde, blinde zeedier bevat de evolutionaire voorloper van onze eigen complexe ooglens-eiwitten.
Deze ontdekking komt van een internationaal onderzoeksteam onder leiding van dr. Lettie Lubsen, biochemica aan de Radboud Universiteit Nijmegen met een passie voor het oog. Jair Stein praat met haar.
Links
Uitgebreid persbericht met foto's over de zakpijp en de evolutie van het menselijk oog (Radboud Universiteit Nijmegen, sep 2005).
Site van Arjan Gittenberger met een overzicht van de talloze soorten zakpijpen die er bestaan.
Oog evolutie →
(PM) ..de " tool" is wellicht reeds aanwezig in de populatie en wel om een heel andere reden dan omwille van haar " toepasbaarheid " in de noodsituatie .
Haal de 'Draco's nog maar eens voor de geest.. Hagedisjes die als ( sexueel) "signaal" de huid tussen hun ribben tonen, blijken die huid ook te kunnen gebruiken als zweef-vleugels. Eenmaal in die toepassing gebruikt, wordt de zweeffunctie belangrijker dan de signaalfunctie. Zo zijn er meer voorbeelden.
(crea)
Dat een bepaalde eigenschap een dubbele functie heeft ? Kan best .Zo kan ik ook een spijker in de muur slaan met een beitel of een steeksleutel.Het zegt alleen nog steeds helemaal niets over het ontstaan van de hamer de beitel of de steeksleutel.
(PM) ( antwoord )
Het zegt wel degelijk iets over (bijvoorbeeld) het onstaan van de hamer .. Als je een steeksleutel beschikbaar hebt en je gebruikt die voor hameren, zul je in de verdere evolutie ervoor toch steeds voor de meest hamerachtige steeksleutel kiezen? En zo kom je tot een hamer :.... de steeksleutel wordt omgebouwd tot een hamerachtig ding dat verder worcdt geoptimaliseerd ...Evolutie bouwt( knutsel ) altijd verder met het aanwezige materiaal en komt in vele gevallen met "rude -goldberg " oplossingen
(crea ) Dan heb je ook nog te maken met selectiedruk.Als iets is gefixeerd dan moet er grote selectiedruk op hebben gestaan.Daarom is het ook onmogelijk om dingen als wenkbrauwen, mullercellen, gehoorbereiken van 20 - 20.000 hertz en dergelijke te fixeren.
(PM)

Hier ontgaat me de onderlinggende logica volkomen van..Als een gehoorbereik van 20 - 20k Hz dat stuk is en dat je als "mensen" het meest nodig hebt, is het toch voor de hand liggend dat daarop geselecteerd wordt?
Uiteindelijk gaat het niet om "grote" selektiedruk maar om langdurige selektiedruk die op elk ogenblik van haar aanwezigheid geen beletsel vormt voor een populatie om zich gedurende al die tijd te blijven voort planten en zodoende te overleven ...Te" grote " ( hevige en "geologisch" kortstondig ) selektiedruk onstaat meestal bij grote veranderingen in de biosfeer (en de fysisch-chemische omgeving van het betrokken aan de druk onderworpen levende systeem : bijvoorbeeld "klimaat" , neerstortende grotere meteoren en planetoiden-brokstukken )___althans dat is het nieuwe inzicht ___ het is meestal het sein van uitstervingen ( zeker voor gespecialiseerde en sexuele soorten met zeer lage generatiewisseling ...)
(crea) Het verhaal blijft hetzelfde.De eerste organismen die absoluut aan fotosynsthese moesten kunnen doen, moesten dat al hebben ontwikkeld voordat ze het nodig hadden.
Er is geen sprake van moeten .... Op het moment dat het eerste organisme door een mutatie in staat blijkt lichtenergie vast te leggen (en dus te "leven" als autotrofen van chemische en fysische eenvoudige (niet levende ) componenten uit de licht- omgeving ) voor later heeft die daar acuut een voordeel aan.. -in het licht tenminste -.
Moest het lichtenergie-vastleg-systeem in het donker ontwikkeld(= verder geoptimaliseerd ) zijn .. Dan is het een reden om aan evolutie te gaan twijfelen.. ...immers dat zou alle logica (?)van evolutie tegenspreken. Alle fasen van de ontwikkeling hebben meteen een voordeel..( in de " licht-niche" tenminste )Maar het betekent niet dat de eerste ( aarzelend ) fotosynthiserende mutanten meteen moesten sterven zonder die eigenschap
De grotere procenten "zuurstof"in de atmosfeer zijn trouwens van oorsprong een afvalprodukt van de eerste fotosynthetiserende systemen ...Elk nadeel heb zijn voordeel( ern vice versa) ; het maakte het onstaan van complexere en bewegelijke zuurstof gebruikers mogelijk Maar die aangroei naar grotere procenten zuurstof bleken (waarschijnlijk ) giftig voor de vele anaerobe( en anders -chemo-trofen ) concurenten van het eerste uur ....
Teveel zuursofpercentage( dan het huidige ) is trouwens nog altijd giftig ook voor de obligate zuurstof-ademhalers :
--> zuurstof toedienen in ziekenhuizen( bij ademhalingsziekten ) is vermoedelijk nog altijd een kantje- boortje ingreep ( dat slechts in de laatste terminale fasen van dergelijke chronische aandoeningen een permanent" uitstel van executie " wordt... = het nog eventjes rekken van de de levensverwachting )
( Crea ) Zo kun je alle voorbeelden wel weer af gaan.Maar daaronder zitten ook dingen als CytochroomC Cytochroom C is zo onmisbaar , dat het eigenlijk niet kon evolueren.
PM : In tegendeel. Cytochroom C is een voorbeeld van een stof die kan veranderen in haar chermische structuur en toch z'n werking behouden.. Er zijn heel veel verschillende cytochroom-c's.. Je kunt aan de hand van de verschillen de stamboom van het leven achterhalen.
Dat cytochroom-c nu onmisbaar is -omdat allerlei 'pathway's er gebruik van maken - zegt niets over het verleden.
Neem de 'afsluitdijk-analogie'.. Ook voor 1932 was er verkeer mogelijk tussen Leeuwarden en Den Helder. Nu zijn er mensen die werken in Leeuwarden en wonen in Den helder en andersom..

Er is intensief verkeer tussen Friesland en West-Friesland. Daardoor is de Afsluitdijk in de tussentijd onmisbaar geworden.. Maar omdat die afsluitdijk nu onmisbaar is, betekent niet dat hij altijd al onmisbaar is geweest..
cytochrome C <--


![g]kfn /fi^« a}+s sd{rf/L ;]jf ljlgodfjnL, 2059 · PDF file161= k"g/fj]bgsf] lg)f{oM..... 128 162= k"g/fj]bgsf] vf/]hLM ... 174= bfx;+:sf/ vr{](https://static.fdocuments.nl/doc/165x107/5ab7dbb77f8b9ac1058bf364/gkfn-fi-as-sdrfl-jf-ljlgodfjnl-2059-kgfjbgsf-lgfom-128-162.jpg)











![Voor DDD geselecteerde titels 1940-1945 Illegale pers · PDF file88 De Vonk – (Haarlem) 89 De "V 3" [De V III, De V3] - (Leeuwarden) ... 178 De Laatste ronde : dagelijksche editie](https://static.fdocuments.nl/doc/165x107/5a81a2f67f8b9a571e8d9073/voor-ddd-geselecteerde-titels-1940-1945-illegale-pers-de-vonk-haarlem-89-de.jpg)



