VERTROUWELIJK NIET KOPIEREN, VERDELEN OF ......VERTROUWELIJK NIET KOPIEREN, VERDELEN OF PUBLIEK...
67
VERTROUWELIJK NIET KOPIEREN, VERDELEN OF PUBLIEK BEKEND MAKEN UNIVERSITEIT GENT FACULTEIT DIERGENEESKUNDE Academiejaar 2016 – 2017 INVLOED VAN EEN HELICOBACTER PYLORI EN HELICOBACTER HEILMANNII INFECTIE OP HET VERLOOP VAN DE ZIEKTE VAN PARKINSON Door Céline RIEMER Promotoren: Eva Bauwens Onderzoek uitgevoerd in het Prof. dr. Freddy Haesebrouck kader van de Masterproef © 2017 Céline Riemer
Transcript of VERTROUWELIJK NIET KOPIEREN, VERDELEN OF ......VERTROUWELIJK NIET KOPIEREN, VERDELEN OF PUBLIEK...

VERTROUWELIJK
NIET KOPIEREN, VERDELEN OF PUBLIEK BEKEND MAKEN
UNIVERSITEIT GENT
FACULTEIT DIERGENEESKUNDE
Academiejaar 2016 – 2017
INVLOED VAN EEN HELICOBACTER PYLORI EN HELICOBACTER HEILMANNII INFECTIE OP
HET VERLOOP VAN DE ZIEKTE VAN PARKINSON
Door
Céline RIEMER
Promotoren: Eva Bauwens Onderzoek uitgevoerd in het
Prof. dr. Freddy Haesebrouck kader van de Masterproef
© 2017 Céline Riemer


Vrijwaringsclausule Universiteit Gent, haar werknemers of studenten bieden geen enkele garantie met betrekking tot de juistheid of
volledigheid van de gegevens vervat in deze masterproef, noch dat de inhoud van deze masterproef geen inbreuk
uitmaakt op of aanleiding kan geven tot inbreuken op de rechten van derden.
Universiteit Gent, haar werknemers of studenten aanvaarden geen aansprakelijkheid of verantwoordelijkheid voor
enig gebruik dat door iemand anders wordt gemaakt van de inhoud van de masterproef, noch voor enig vertrouwen
dat wordt gesteld in een advies of informatie vervat in de masterproef.
VERTROUWELIJK – BELANGRIJK
Deze masterproef bevat vertrouwelijke informatie en/of vertrouwelijke
onderzoeksresultaten die toebehoren aan de Universiteit Gent of aan derden.
Deze masterproef of enig onderdeel ervan mag op geen enkele wijze publiek
gemaakt worden zonder uitdrukkelijke schriftelijke voorafgaande toestemming
vanwege de Universiteit Gent. Zo mag de masterproef onder geen voorwaarde
door derden worden ingekeken of aan derden worden meegedeeld. Het nemen
van kopieën of het op eender welke wijze dupliceren van de masterproef is
verboden. Het niet respecteren van de vertrouwelijke aard van de masterproef
kan onherstelbare schade veroorzaken aan de Universiteit Gent.

VERTROUWELIJK
NIET KOPIEREN, VERDELEN OF PUBLIEK BEKEND MAKEN
UNIVERSITEIT GENT
FACULTEIT DIERGENEESKUNDE
Academiejaar 2016 – 2017
INVLOED VAN EEN HELICOBACTER PYLORI EN HELICOBACTER HEILMANNII INFECTIE OP
HET VERLOOP VAN DE ZIEKTE VAN PARKINSON
Door
Céline RIEMER
Promotoren: Eva Bauwens Onderzoek uitgevoerd in het
Prof. dr. Freddy Haesebrouck kader van de Masterproef
© 2017 Céline Riemer

VOORWOORD
Na een jaar intensief gewerkt te hebben aan dit onderzoek in het kader van de masterthesis voor het
derde masterjaar Diergeneeskunde is deze Proeve van Bekwaamheid ontstaan. Als studente
Diergeneeskunde aan de Universiteit Gent is mijn keuze uitgegaan naar een onderzoek over de invloed
van Helicobacter pylori en Helicobacter heilmannii infecties op het verloop van de ziekte van Parkinson.
Deze bacteriën komen regelmatig voor bij de mens en er is nog volop onderzoek naar de gevolgen van
een besmetting met deze bacteriën. Tevens ben ik erg geïnteresseerd in de ziekte van Parkinson en
door deze studie was het mogelijk om een van de mogelijke oorzaken te onderzoeken.
Tijdens het uitvoeren van het onderzoek en het schrijven van deze masterthesis heb ik veel hulp gehad
van mijn promotor Eva Bauwens, waarvoor mijn hartelijke dank. Zij heeft mij zeer veel geleerd over de
technieken in het laboratorium en over de verschillende onderzoeksmogelijkheden. Met vragen kon ik
altijd bij haar terecht en het was erg fijn dat ik zelf ook al meteen mee mocht helpen. Ook wilde ik Eva
Bauwens bedanken, dat zij het vertrouwen in mij had, om mij zelfstandig dingen te laten doen in het
laboratorium. Tevens wil ik Dr. Caroline Blaecher heel erg bedanken voor de uitleg over het pipetteren
en over de verzorging van de muizen die gebruikt zijn bij dit onderzoek. Als derde wil ik Sofie De
Bruyckere bedanken om mij te laten zien hoe men een DNA-extractie uitvoert. Tevens wil ik mijn co-
promotor Prof. dr. Freddy Haesebrouck bedanken voor het controleren van mijn thesis en zijn
waardevolle opmerkingen. Als laatste wil ik Jason Schoukens bedanken voor alle hulp bij het uitvoeren
van de RNA-extracties en het maken van cDNA.
Ik wens u veel plezier met het lezen van deze masterthesis.
Plaats: Merelbeke (België)
Datum afronding: 8 mei 2017
Handtekening:

INHOUDSOPGAVE
SAMENVATTING………………………………………………………..………………………………... p. 1
LIJST MET AFKORTINGEN………………………………………….………………………………….. p. 2
INLEIDING………………………………………………………………………………………………..... p. 4
DEEL 1: LITERATUUR…………………………………………………………………………………… p. 5
1. Gastrale Helicobacter species bij de mens…………………………………………………............. p. 5
1.1. Helicobacter pylori………………………………………………………………………………… p. 5
1.1.1. Transmissie…………………………………………………………………………………. p. 6
1.1.2. Virulentiefactoren………………………………………………………….……….……….. p. 6
1.1.2.1. Virulentiefactoren verantwoordelijk voor de kolonisatie van de gastrale
mucosa…………………………………………………………………………….. p. 6
1.1.2.2. Virulentiefactoren verantwoordelijk voor de inductie van gastrale lesies…… p. 7
1.1.3. Symptomen………………………………………………………………………................ p. 8
1.1.4. Diagnose…………………………………………………………………………….………. p. 9
1.1.5. Behandeling…………………………………………………………………………………. p. 10
1.2. Helicobacter heilmannii…………..………………………………………………………………. p. 11
1.2.1. Transmissie………………….……………………………………………………………… p. 14
1.2.2. Virulentiefactoren…………………………………………………………………………… p. 14
1.2.2.1. Virulentiefactoren verantwoordelijk voor de kolonisatie van de gastrale
mucosa…………………………………………………………………………….. p. 14
1.2.2.2. Virulentiefactoren verantwoordelijk voor de inductie van gastrale lesies…… p. 15
1.2.3. Symptomen………………………………………………………………………………….. p. 15
1.2.4. Diagnose…………………………………………………………………………………….. p. 15
1.2.5. Behandeling…………………………………………………………………………………. p. 16
2. De ziekte van Parkinson………………………………………………………………………………..p. 17
2.1. Inleiding…………………………………………………………………………………………….. p. 17
2.2. Oorzaken…………………………………………………………………………………………… p. 17
2.2.1. Genetische factoren ………………………………………………………………………. p. 18
2.2.2. Milieufactoren ……………………………………………………………………………… p. 18
2.3. Pathogenese………………………………………………………………………………………. p. 19
2.3.1. Dysfunctie van de hersenbarrières ……………………………………………………… p. 20
2.3.2. Bloed-hersen barrière……………………………………………………………………... p. 21
2.3.3. Bloed-cerebrospinaalvocht barrière……………………………………………………… p. 22
2.3.4. Junctions in de hersenbarrières………………………………………………………….. p. 22
2.4. Symptomen………………………………………………………………………………………… p. 23
2.5. Diagnose…………………………………………………………………………………………… p. 24
2.6. Prevalentie…………………………………………………………………………………………. p. 24
2.7. Behandeling………………………………………………………………………………………... p. 25
3. Betrokkenheid van Helicobacter infecties in het verloop van de ziekte van Parkinson………… p. 26
DEEL 2: DOELSTELLING…..…..………………………………………………………………............. p. 27
DEEL 3: MATERIAAL EN METHODEN….……………….…………………………………………….. p. 29
1. Muizen......................................................................................................................................... p. 29
2. Cultuur van de H. pylori en H. heilmannii bacteriën.................................................................... p. 29
3. In vivo infectieprocedure………………........................................................................................ p. 29
4. Weefselcollectie…………………………………............................................................................ p. 30

5. RNA-extractie en RT-qPCR voor genexpressie.......................................................................... p. 31
6. DNA -extractie en kwantificatie van H. pylori en H. heilmannii in de maag in het gastro-
Intestinaal stelsel……………………………………………………………………………………….. p. 31
7. Histopathologie en immunohistochemie……………………………………………………………... p. 32
8. Statistiek…………………………………………………………………………………………………. p. 32
DEEL 4: RESULTATEN……..………………………………………………………..………………….. p. 33
1. Kwantificatie……………………………………………………………………................................... p. 33
1.1. Helicobacter heilmannii........................................................................................................ p. 33
1.2. Helicobacter pylori………………………………………………………………………………… p. 33
2. Helicobacter heilmannii: integriteit van het gastro-intestinaal stelsel.......................................... p. 34
2.1. Relatieve mRNA expressie: Algemeen……………………………………..…………………... p. 34
2.2. Relatieve mRNA expressie van genen die coderen voor tight junctions……….…………… p. 34
2.3. Relatieve mRNA expressie van genen die coderen voor ontstekingsmediatoren………..... p. 35
2.4. Relatieve mRNA expressie van genen die coderen voor Vip, VipR1 en VipR2……............ p. 36
2.5. Relatieve mRNA expressie van genen die coderen voor Par3, Par6 en aPKC…................ p. 37
2.6. Relatieve mRNA expressie van genen die coderen voor mucines……………….…………. p. 38
3. Helicobacter pylori: integriteit van het gastro-intestinaal stelsel.................................................. p. 39
3.1. Relatieve mRNA expressie: Algemeen……………………………………..…………………... p. 39
3.2. Relatieve mRNA expressie van genen die coderen voor tight junctions……….…………… p. 39
3.3. Relatieve mRNA expressie van genen die coderen voor ontstekingsmediatoren………..... p. 40
3.4. Relatieve mRNA expressie van genen die coderen voor Vip, VipR1 en VipR2……............ p. 41
3.5. Relatieve mRNA expressie van genen die coderen voor Par3, Par6 en aPKC...…............. p. 41
3.6. Relatieve mRNA expressie van genen die coderen voor mucines……………….…………. p. 42
4. Integriteit van de bloed-hersen barrière en de bloed-cerebrospinaalvocht barrière…………….. p. 43
4.1. MRNA expressie: Algemeen……………………………………………………………………... p. 44
4.2. Helicobacter heilmannii: Relatieve mRNA expressie van genen die coderen voor tight
junctions……………………………………………………………………………………………. p. 44
4.3. Helicobacter heilmannii: Relatieve mRNA expressie van genen die coderen voor tight
junctions……………………………………………………………………………………………. p. 45
DEEL 5: DISCUSSIE....................................................................................................................... p. 46
1. Het onderzoek…………………………………………………………………………………………... p. 46
2. De spijsverteringsstelsel-hersenen as in relatie met een H. pylori en H. heilmannii infectie…... p. 46
3. Invloed van een Helicobacter infectie op de integriteit van het gastro-intestinaalstelsel……….. p. 47
3.1. Kwantificatie……………………………………………………………………………………….. p. 47
3.2. Relatieve mRNA expressie van genen die coderen voor tight junctions…….……………… p. 48
3.3. Relatieve mRNA expressie van genen die coderen voor ontstekingsmediatoren………..... p. 48
3.4. Relatieve mRNA expressie van genen die coderen voor Vip, VipR1 en VipR2………..…... p. 48
3.5. Relatieve mRNA expressie van genen die coderen voor mucines……………….…………. p. 49
4. Invloed van een Helicobacter infectie op de hersenbarrières……………………………………... p. 49
5. Conclusie en aanbevelingen………………………………………………………………………….. p. 49
LITERATUURLIJST........................................................................................................................ p. 51
BIJLAGEN....................................................................................................................................... p. 60

1
SAMENVATTING
Deze thesis gaat over de invloed van een Helicobacter (H.) pylori en H. heilmannii infectie op het verloop
van de ziekte van Parkinson (Parkinson‘s Disease; PD). H. pylori is een gram negatieve, spiraalvormige
bacterie die de maag van ongeveer vijftig procent van de wereldwijde humane populatie koloniseert.
Een besmetting met H. pylori wordt hoogstwaarschijnlijk reeds verworven tijdens de eerste levensjaren.
H. heilmannii infecteert van nature uit de maag van katten en kan overgedragen worden op de mens.
Infecties met deze micro-organismen worden geassocieerd met gastrale ziekten bij de mens. In deel 1
van deze thesis zal er een overzicht gegeven worden van het genus Helicobacter waarbij er dieper
wordt ingegaan op de transmissie, virulentiefactoren en symptomen van een infectie met H. heilmannii
of H. pylori. Vervolgens zullen verschillende diagnostische technieken voor de detectie van een gastrale
Helicobacter infectie worden besproken en de behandeling van een infectie met deze micro-
organismen. Daarna zal de link gelegd worden tussen een infectie met een van deze species en PD.
Vervolgens zullen de pathogenese, oorzaken, symptomen, diagnose, prevalentie en behandeling van
PD besproken worden.
In deel 2 zal de doelstelling van deze masterthesis besproken worden. Deel 3 beschrijft het materiaal
en de methoden van dit onderzoek en deel 4 bevat de resultaten waaruit blijkt dat een gastrale infectie
met H. heilmannii en H. pylori mogelijks de communicatie tussen het gastro-intestinaal stelsel en de
hersenen beïnvloedt. Als laatste zal er een discussie volgen.
Sleutelwoorden: Helicobacter heilmannii, Helicobacter pylori, Infectie, Ziekte van Parkinson
2
LIJST MET AFKORTINGEN
Afkorting Engelse benaming Nederlandse benaming
AJ Adherens Junctions -
aPKC Atypical protein kinase C Atypisch proteïne kinase C
BBB Blood-Brain Barrier Bloed-hersen barrière
BCSFB Blood-Cerebrospinal Fluid
Barrier
Bloed-cerebrospinaal vocht
barrière
C Carbon Koolstof
CagPaI Cytotoxin-associated gene
pathogenicity island
Cytotoxine geassocieerd gen
pathogeniciteitseiland
Cldn Claudin Claudine
CNS Central Nervous System Centraal zenuwstelsel
CP Choroid plexus Choroïdale plexus
CSF Cerebrospinal Fluid Cerebrospinaal vocht
CT Computer Tomography Computer Tomografie
DBS Deep Brain Stimulation Diepe hersenstimulatie
DNA Desoxyribonuclein acid Desoxyribonucleïnezuur
dPBS Dulbecco's phosphate-buffered
saline
Dulbecco’s fosfaat gebufferde
zoutoplossing
E-cadh E-cadherin E-cadherine
ELISA Enzym-linked Immuno Sorbent-
Assay
Enzymgekoppelde
Immunosorbent Test
ErbB Epidermal growth factor
receptor
Epidermale groeifactor receptor
FITC Fluorescein-isothiocyanate -
FldA Flavodexine protein Flavodexine proteïne
GABA γ-Amino Butyric Acid Gamma-Aminoboterzuur
GGT Gamma-
glutamyltranspeptidase
Gamma-
glutamyltranspeptidase
HBSS Hanks balanced salt solution Hanks gebufferde
zoutoplossing
HCL Hydrochloric acid Waterstofchloride
H. Helicobacter Helicobacter
Hof Helicobacter OMP family Helicobacter OMP familie
Hom Helicobacter outer membrane Helicobacter buitenste
membraan
Hop Helicobacter outer membrane
porin
Helicobacter buitenste
membraan porine
3
Hor Hop related Hop gerelateerd
Ig Immunoglobulin Immunoglobuline
Il Interleukin Interleukine
Kc Keratinocyte chemoattractant -
Lix Lipopolysaccharide-induced
CXC chemokine
Lipopolysaccharide-
geïnduceerd CXC chemokine
LRRK2 Leuncin-Rich Repeat Kinase 2 -
MALT mucosa-associated lymphoid
tissue
Mucosa-geassocieerd lymfoïd
weefsel
MIC Minimal Inhibitory
Concentration
Minimale inhibitorische
concentratie
MIP Macrophage inflammatory
protein
Macrofaag inflammatoir
proteïne
Muc Mucin Mucine
NHPH Non-Helicobacter Pylori
Helicobacter
Niet-Helicobacter Pylori
Helicobacter
Ocln Occludin Occludine
OipA Outer inflammatory protein A -
OMP Outer membrane protein Buitenste membraan proteïne
Par Pseudoautosomal region Pseudo-autosomale regio
PCR Polymerase Chain Reaction Polymerase-kettingreactie
PD Parkinson‘s Disease Ziekte van Parkinson
PET Positron Emission Tomography Positron Emissie Tomografie
PINK Putative Kinase 1 -
RNA Ribonuclein acid Ribonucleïnezuur
SEM Standard Error of the Mean Standaard fout van het
gemiddelde
Th T-helper cell T-helper cel
TJ Tight Junctions Tight Junctions
TNF-a Tumor Necrosis Factor-alpha Tumornecrosefactor-alfa
VacA Vacuolating Cytotoxine A Vacuoliserend Cytotoxine A
Vip vasoactive intestinal peptid vaso-actieve intestinale peptide
wt - wildtype
Zo Zonula occludens Zonula occludens

4
INLEIDING
Meer dan honderd jaar geleden beschreef de Poolse wetenschapper W. Jaworski voor het eerst
spiraalvormige micro-organismen die voorkwamen in de maag van de mens. De daaropvolgende jaren
werden deze species nog vaak ontdekt, gerapporteerd, maar dan weer vergeten. In 1975, mede door
de opkomst van biopsie technieken, toonden Steer en Colin-Jones in 80% van de patiënten, die te
kampen hadden met maagzweren, Gram-negatieve bacteriën aan (Steer en Colin-Jones,1975; Marshall
en Warren, 1984). Ook hier werd echter geen aandacht aan besteed. Enkele jaren later, in 1982
slaagden de Australische arts B.J. Marshall en patholoog J.R. Warren erin om deze bacteriën te isoleren
uit maagbiopten van elf patiënten met gastritis (Marshall en Warren, 1984; Brown, 2000; Konturek,
2003).
De geïsoleerde micro-organismen werden eerst ingedeeld in het geslacht Campylobacter, maar werden
al snel hertoegewezen aan het geslacht Helicobacter. Deze herclassificatie was te wijten aan verschillen
in onder andere de cellulaire vetzuren, de antibiotica resistentie en de 5S en/of 16srRNA gen sequenties
tussen de ontdekte species en leden van het geslacht Campylobacter (Goodwin 1989; Konturek, 2003;
Kusters et el., 2006).
Op dit moment zijn er reeds 46 verschillende Helicobacter soorten gekend, waarvan de humane
pathogeen H. pylori als eerste werd ontdekt (Flahou et al., 2016).
Deze verschillende bacteriën kunnen grofweg ingedeeld worden in twee groepen, namelijk de gastrale
en de enterohepatische Helicobacter species (Solnick en Schauer, 2001; Flahou et al., 2016). De
gastrale helicobacters zijn in staat om te overleven in het zure milieu van de maag, omdat ze in hoge
mate urease produceren (Weeks et al., 2000; Flahou et al., 2016). Dit in tegenstelling tot de
enterohepatische soorten die voornamelijk terug te vinden zijn in de mucosale oppervlakten van het
intestinale stelsel en/of in de lever (Solnick en Schauer, 2001; Sterzenbach et al., 2007). Vele, zo niet
alle species zijn aangetoond in dieren en enkele van deze dier-gerelateerde micro-organismen zijn in
staat om ziekte te veroorzaken bij de mens (Haesebrouck et al., 2009; Flahou et al., 2016).
Na de ontdekking van H. pylori in de maag van mensen die leden aan gastritis en maagzweren, werd
de mogelijke link gelegd tussen PD en een infectie met H. pylori (Dukowicz et al., 2007; Dobbs et al.,
2008; Nielsen et al., 2012; Tan et al., 2015; Csoti et al., 2016; Dobbs et al., 2016).
PD is, op de ziekte van Alzheimer na, de tweede meest voorkomende neurodegeneratieve aandoening
bij de huidige humane populatie. De aandoening kan gepaard gaan met zowel motorische als non-
motorische symptomen. Het neurologische proces wordt gekarakteriseerd door de degeneratie van
dopamine producerende neuronen in de pars compacta van de substantia nigra. Op dit moment is
echter de exacte etiologie van PD bij 90 tot 95 procent van de gevallen nog niet bekend. De ziekte wordt
beschouwd als een multifactoriële aandoening waarbij zowel genetische als milieufactoren een rol
spelen (Wirdefelt et al., 2011; Mhyre et al., 2012). In deze thesis zal de invloed van een infectie met H.
pylori en H. heilmannii op het verloop van PD verder onderzocht worden.

5
DEEL 1: LITERATUUR 1. GASTRALE HELICOBACTER SPECIES BIJ DE MENS
H. pylori is de meest bestudeerde Helicobacter species, die bij mensen aanleiding kan geven tot
verschillende maagaandoeningen zoals gastritis, maagulcera, gastrale adenocarcinomen en gastrale
MALT-lymfomen (“mucosa-associated lymphoid tissue lymphoma”). Bovendien worden deze gastrale
ziekten ook geassocieerd met andere lange spiraalvormige bacteriën, namelijk niet-Helicobacter pylori
Helicobacter (NHPH) species. Deze worden teruggevonden in de maag van gedomesticeerde dieren
zoals honden, katten, varkens en niet-menselijke primaten (Haesebrouck et al., 2009; Vermoote et al.,
2011; Joosten et al., 2013; Flahou et al., 2016). Ook worden de NHPHs teruggevonden in 0,2 tot 6
procent van de gastrale biopsies genomen bij mensen met erge maagklachten (Heilmann en Borchard,
1991; Solnick en Goldstein, 2003). Deze percentages zijn echter een onderschatting aangezien infecties
met NHPHs soms asymptomatisch verlopen of slechts leiden tot milde symptomen waardoor dit vaak
niet verder onderzocht wordt (Parsonnet 1995; Brown 2000). Bovendien komen de bacteriën slechts in
bepaalde delen van de maagmucosa voor en worden ze bij het nemen van een biopsie vaak gemist
door hun verspreide distributie (Haesebrouck et al., 2009).
Bacteriën die tot de groep van de NHPHs behoren zijn onder andere H. suis, afkomstig van varkens, H.
felis, H. bizzozeronii, H. salomonis en H. heilmannii afkomstig van honden en katten (Haesebrouck et
al., 2009; Flahou et al., 2016). Infecties met deze micro-organismen worden bij de mens geassocieerd
met gastritis, maag- en duodenale zweren, maar ook met ergere maag-pathologieën zoals MALT-
lymfomen (Brown, 2000; Kusters et al., 2006; Vermoote et al., 2011).
In deze masterthesis zal er voornamelijk dieper ingegaan worden op H. pylori en H. heilmannii.
1.1. HELICOBACTER PYLORI
H. pylori, een gram negatieve, spiraalvormige bacterie, koloniseert de maag (Sgouras et al., 2015; Singh
et al., 2015) van ongeveer vijftig procent van de wereldwijde humane populatie (Kusters et al., 2006;
Joosten et al., 2013; Sgouras et al., 2015; Singh et al., 2015). In ontwikkelingslanden kan het percentage
H. pylori geïnfecteerde mensen zelfs oplopen tot meer dan tachtig procent. Dit in tegenstelling tot de
meer ontwikkelde landen, waar de prevalentie varieert van twintig tot vijftig procent (Suerbaum en
Michetti, 2002; Kusters et al., 2006). Deze grote geografische variaties kunnen te wijten zijn aan een
combinatie van verschillende factoren zoals de H. pylori stam, de leeftijd waarop men geïnfecteerd
raakt, genetische factoren en de socio-economische status van de gastheer als ook milieufactoren
(Kusters et al., 2006; Conteduca et al., 2012).
H. pylori is dus een van meest succesvolle pathogenen bij de mens en zonder behandeling zal de kiem
levenslang in de gastheer aanwezig blijven (Suerbaum en Michetti, 2002; Kusters et al., 2006; Sgouras
et al., 2015).

6
1.1.1. Transmissie Een besmetting met H. pylori wordt hoogstwaarschijnlijk reeds verworven tijdens de eerste levensjaren
via oraal-oraal contact. De exacte wijze van overdracht werd echter nog steeds niet volledig
opgehelderd. Verder is het ook moeilijk om het tijdstip van besmetting te bepalen aangezien een infectie
met H. pylori vaak asymptomatisch verloopt (Suerbaum en Michetti; 2002; Suerbaum en Josenhans,
2007). In geïndustrialiseerde landen is het aannemelijk dat de directe transmissieroute van de ene
persoon naar de andere door middel van braaksel, speeksel of faeces de belangrijkste is. Bijkomende
transmissieroutes zoals via water zijn vooral belangrijk in ontwikkelingslanden (Goodman et al., 1996;
Parsonnet et al., 1999; Suerbaum en Michetti, 2002).
1.1.2. Virulentiefactoren
1.1.2.1. Virulentiefactoren verantwoordelijk voor de kolonisatie van de gastrale mucosa
De specifieke helix vorm van H. pylori is cruciaal
voor de beweeglijkheid en een vereiste voor een
succesvolle kolonisatie (Brown, 2000; Sgouras et
al., 2015).
De beweeglijkheid van de bacterie wordt
gegarandeerd door 2 tot 6 omhulde flagellen, die
zich aan één uiteinde van de bacterie bevinden.
Door deze flagellen kan de bacterie standhouden
tijdens de ritmische contracties van de maag. Ook
kan de kiem door middel van deze flagellen
doorheen de mucosa van de maag penetreren
(Brown, 2000).
Verder maakt H. pylori gebruik van chemotaxis,
wat wil zeggen dat de bacteriën hun bewegingen
aanpassen aan extracellulaire chemische
signalen. Dit is essentieel voor een succesvolle
kolonisatie en voor het in stand houden van de infectie. In recente studies werd aangetoond dat
gemuteerde bacteriën met defecten in de chemotaxis enkel in staat waren om de maagmucosa, maar
niet de antrale klieren van mensen en muizen te koloniseren (Sgouras et al., 2015; Sigal et al., 2015).
Ter hoogte van de buitenste membraan van de celwand of in het cytoplasma van de bacterie komen
transducer-like eiwitten voor. Dit zijn moleculen die zich gedragen als chemoreceptoren voor
extracellulaire chemische signalen en voor interactie met de respectievelijke liganden. Deze
wisselwerking veroorzaakt een moleculaire signaaltransductie-cascade die er uiteindelijk voor zorgt dat
de rotatie richting van de motors van de flagellen verandert (Liu en Roujeinikova, 2015; Sgouras et al.,
2015).
Figuur. 1. Structuur van Helicobacter pylori
Bron: https://www.dreamstime.com/stock-illustration-
structure-helicobacter-pylori-infographics-vector-
illustration-isolated-background-image74057637

7
Veder kan de immuunrespons van de gastheer aanleiding geven tot oxidatieve stress waartegen de
kiem zich moet verweren. H. pylori is in staat om zich hiertegen te beschermen door middel van
verscheidene componenten zoals katalase (gecodeerd door het KatA gen), superoxide dismutase
(gecodeerd door het SodB gen) en alkyl hydroperoxide reductase (gecodeerd door het HhpC gen)
(Seyler et al., 2001; Harris et al., 2002; Olczak et al., 2003; Ernst et al., 2005, Arnold et al., 2011; Schott
et al., 2011; Smet et al., 2011; Vermoote et al., 2011).
Een van de eerste stappen in de pathogenese van gastrale infecties veroorzaakt door Helicobacter is
de kolonisatie en daarbij vooral de adhesie van deze bacteriën aan de gastrale mucosa (McGuckin et
al., 2011). H. pylori is hiervoor voorzien van ongeveer 64 OMPs die onderverdeeld kunnen worden in 5
groepen. De grootste familie (familie 1) bevat de Hop (Helicobacter outer membrane porin) en Hor (Hop
related) eiwitten. Families 2 en 3 omvatten de Hof (Helicobacter OMP familly) en Hom (Helicobacter
outer membrane) eiwitten. Families 4 en 5 zijn samengesteld uit ijzer-regulerende OMPs en efflux-pomp
OMPs (Alm et al., 2000; Oleastro en Ménard, 2013; Roesler et al., 2014; Teymournejad et al., 2014;
Sgouras et al., 2015).
1.1.2.2. Virulentiefactoren verantwoordelijk voor de inductie van gastrale lesies
Een belangrijke virulentiefactor is het cytotoxine geassocieerd gen pathogeniciteitseiland (Cytotoxin-
associated gene pathogenicity island: cagPAI) dewelke het risico op maagkanker verhoogt. Hanada et
al. (2014) beschreven dat patiënten die geïnfecteerd waren met cagPAI-positieve stammen een twee
keer zo groot risico hebben op gastrale carcinogenese dan de patiënten geïnfecteerd met cagPAI-
eiland-negatieve stammen.
Verder scheidt H. pylori het Vacuolating Cytotoxine A (VacA), een porie vormend toxine, uit. Eenmaal
VacA is binnengedrongen in de cellen, kan het veel cel veranderingen veroorzaken, inclusief celdood.
Het mechanisme dat verantwoordelijk is voor deze celdood is nog niet volledig gekend (Radin et al.,
2014).
Een andere belangrijke virulentiefactor van H. pylori is het membraan-geassocieerde gamma-
glutamyltranspeptidase (GGT) (Haesebrouck et al., 2009; Wachino et al., 2010; Arnold et al., 2011;
Schott et al., 2011; Smet et al., 2011; Vermoote et al., 2011; Rossi et al., 2012), dat een belangrijke rol
speelt in het metabolisme van het antioxidans gluthation (Orlowski en Meister, 1970). Het GGT gebruikt
zowel glutamine als glutathion, afkomstig van epitheelcellen, als bron voor de productie van glutamaat
(Shibayama et al., 2003; Flahou et al., 2011). Aangezien glutamine en glutathion belangrijke
voedingsstoffen zijn voor het onderhouden van een gezonde maagmucosa, speelt hun verbruik door
GGT mogelijks een rol in de beschadiging van de epitheelcellen van de maag (Shibayama et al., 2001;
Shibayama et al., 2003; Kim et al., 2007). Bijkomend is GGT ook verantwoordelijk voor de inhibitie van
T-cel proliferatie, waardoor de immuniteit van de gastheer wordt aangetast (Schmees et al., 2007;
Zhang et al., 2015).
Naast het cagPAI, VacA, en GGT zijn er nog andere virulentiefactoren betrokken bij de inductie van
gastrale ziektes zoals FldA en IceA (Arnold et al., 2011; Schott et al., 2011; Smet et al., 2011; Vermoote

8
et al., 2011; Kersulyte et al., 2013; Joosten et al., 2015). Het flavodoxine proteïne (FldA) wordt
bijvoorbeeld geassocieerd met MALT lymphoma (Chang et al., 1999).
Het ulcer-geassocieerde proteïne restrictieve endonuclease (IceA) is bijna bij alle helicobacters
aanwezig en geeft aanleiding tot maagzweren (Peek et al., 1998; Donahue et al., 2000).
1.1.3. Symptomen
Het klinische verloop van een infectie met H. pylori is afhankelijk van meerdere factoren van zowel de
gastheer als de bacterie en hun complexe interactie (Kusters et al., 2006; Conteduca et al., 2012).
Slechts 10 tot 20 procent van de geïnfecteerde patiënten ontwikkelen daadwerkelijk ziekte (Konturek
2003; Kusters et al., 2006; Suerbaum en Josenhans, 2007; Conteduca et al., 2012). De ziekte kan zeer
verschillend verlopen, gaande van een chronische actieve gastritis zonder klinische tekens tot
maagulcera, gastrale adenocarcinomen en gastrale MALT-lymfomen. Geïnfecteerde mensen hebben
een 2 tot 6 keer verhoogd risico op het ontwikkelen van maagkanker en gastrale MALT-lymfomen
(Konturek 2003; Kusters et al., 2006; Vermoote et al., 2011; Conteduca et al., 2012).
Het polymorfisme van de immuniteitsgenen en de maagzuursecretie van de gastheer bepalen
grotendeels of de kiem stand kan houden (Kusters et al., 2006). Uit een studie van Conteduca et al.
(2012) blijkt dat de kans op het ontwikkelen van antrum-predominante gastritis die vervolgens aanleiding
kan geven tot duodenale zweren groter is bij patiënten met een verhoogde maagzuur secretie. Patiënten
met een lage zuursecretie hebben een grotere kans op het genereren van een gastritis ter hoogte van
het corpus van de maag, waardoor zij eerder maagzweren kunnen ontwikkelen. Dit kan vervolgens
leiden tot maagatrofie, intestinale metaplasie en dysplasie (Conteduca et al., 2012). Hieronder volgt een
overzicht van de verschillende ziekten die kunnen ontstaan bij een infectie met H. pylori (figuur 2).
Fig. 2. Overzicht van de verschillende met Helicobacter pylori geassocieerde gastro-intestinale
ziekten bij humane patiënten
Bron: Conteduca et al., 2012

9
1.1.4. Diagnose
Aangezien een acute infectie met H. pylori meestal asymptomatisch verloopt, is het moeilijk om aan de
hand van klinische symptomen vast te stellen wanneer een individu geïnfecteerd is geraakt (Parsonnet
1995; Brown 2000). Vroeger stelde men de diagnose van een infectie met H. pylori door de kiem te
isoleren vanuit een maagbiopt, dat men verkreeg via endoscopie, of door de bacterie te identificeren op
aangekleurde weefselcoupes (Perez-Perez et al.,1982; Marshall en Warren 1984). Een nadeel hiervan
is, dat een biopt of cultuur wel 100 procent specifiek is, maar niet 100 procent sensitief aangezien men
op deze manier slechts een klein deel van de maag kan beoordelen. Een ander nadeel van deze
methoden is, dat ze erg invasief en redelijk duur zijn.
Tegenwoordig kan men de diagnose stellen door middel van serologische testen. Hierbij wordt gebruik
gemaakt van een niet-kwantitatieve enzymgekoppelde immunosorbent test (Enzym-Linked Immuno
Sorbent Assay; ELISA). Hierbij tracht men H. pylori antilichamen te detecteren in het serum van de
patiënt. Men gebruikt hierbij immunoglobuline (Ig)G antilichamen, alhoewel er ook gebruik kan worden
gemaakt van IgA en IgM antilichamen (Andersen et al., 1996). Een groot voordeel van deze test is dat
men veel patiënten binnen een korte tijdspanne kan screenen. Bovendien is dit onderzoek relatief
goedkoop, wat dit een goede test maakt voor het uitvoeren van epidemiologische studies (Taylor en
Blaser; 1991). Er zijn natuurlijk ook nadelen verbonden aan deze immunologische test. Het eerste
nadeel is dat er geen enkele test bestaat die in staat is om elke H. pylori stam te herkennen (Atherton
1997; Hook-Nikanne et al., 1997). Als tweede is het moeilijk om een cut-off waarde te bepalen die de
positief geteste patiënten van de negatieve onderscheidt. Als derde is de test gevoelig aan
veranderingen in het laboratorium. Sera die gecollecteerd worden op verschillende momenten,
bijvoorbeeld voor en na de behandeling van de patiënt, worden dus het beste in dezelfde
omstandigheden getest. Het laatste nadeel is dat men met serologie niet kan bepalen of een
behandeling tegen H. pylori aanslaat aangezien het enkele maanden duurt vooraleer de verhoogde
antilichaam titers zullen dalen (Brown, 2000).
Een andere test waarvan tegenwoordig gebruik gemaakt wordt, is de ureum ademtest waarbij men 13
koolstof (carbon; C) of 14C gebruikt. Dit is een niet-invasieve, niet-kwantitatieve test die de huidige H.
pylori status in de maag bepaalt door detectie van de urease activiteit van de bacteriën. De sensitiviteit
van de ureum ademtest is 88 tot 95 procent en de specificiteit 95 tot 100 procent (Malfertheiner et al.,
2012). Een groot voordeel van deze test, ten opzichte van serologische testen, is dat deze test wel te
gebruiken is om een behandeling met antibiotica op te volgen. Urease is namelijk enkel aanwezig als
de bacteriën, die dit urease produceren, ook daadwerkelijk voorkomen in de maag. Een nadeel van
deze test is dat er ook vals negatieve resultaten kunnen voorkomen bij behandelde patiënten als er te
weinig bacteriën aanwezig zijn in de maag om meetbare urease spiegels te produceren. Bij minder dan
5 procent van de populatie kunnen er vals positieve resultaten voorkomen, doordat bepaalde andere
bacteriën ook in staat zijn urease te produceren. Een ander nadeel van deze test is dat de ureum
ademtest duurder is dan serologie. Toch neemt het gebruik ervan in epidemiologische studies toe,
vooral als er kinderen betrokken zijn, aangezien deze test niet invasief is (Brown, 2000).

10
Er bestaat ook een faecale antigen test. Dit is een niet-invasieve test die de uitscheiding van H. pylori
antigenen in de faeces van de patiënt meet en zo de aanwezigheid van een infectie bepaalt (Vaira et
al., 2002; Pajares en Gisbert, 2006). Deze test heeft een sensitiviteit van 94 procent en een specificiteit
van 92 procent. Het nadeel van deze test is echter dat dit voor patiënten van bepaalde culturen minder
acceptabel is doordat men een staal van de ontlasting moet nemen (Malfertheiner et al., 2012).
Een recente techniek om H. pylori en andere Helicobacter species te detecteren is real-time-
Polymerase-kettingreactie (Polymerase Chain Reaction; PCR) (Mentis et al.; 2015). In recente studies
werd er gebruik gemaakt van real-time-PCR op maagbiopten van Franse kinderen om de diagnose van
een H. pylori infectie te stellen. Bij dit onderzoek kwam men tot de conclusie dat dit een nauwkeurigere
test is dan enkel een routinematige cultuur of histologie. Een ander voordeel van deze methode is dat
men hiermee ook kleine aantallen bacteriën kan detecteren (Kalach et al., 2015; Mentis et al.; 2015).
1.1.5. Behandeling
Jarenlang werd er gebruik gemaakt van een drievoudige combinatietherapie als standaard behandeling
van een H. pylori infectie bij de mens. Deze combinatietherapie bestond uit de toediening van twee
antibiotica zoals amoxicilline met clarithromycine of metronidazole samen met een protonpomp inhibitor
zoals omeprazole of pantoprazole gedurende tenminste één week (Graham en Fischbach 2010; Flahou
et al., 2013; Joosten et al., 2013). Tegenwoordig blijkt deze behandelingsmethode wereldwijd niet meer
zo succesvol te zijn aangezien de eradicatie ratio van H. pylori in veel landen afgenomen is tot minder
dan tachtig procent, door de toegenomen antibioticaresistentie (Suerbaum en Michetti, 2002; Chey en
Wong, 2007; Graham en Shiotani, 2008; Talley et al., 2008; Horiki et al., 2009; Graham en Fischbach,
2010; Kivistö et al., 2010; Malfertheiner et al., 2012). Men probeert nu op verschillende manieren deze
ratio weer te verhogen. Volgens Graham en Fischbach zou bijvoorbeeld de hierboven beschreven
drievoudige combinatietherapie moeten verlengd worden tot 14 dagen. Ook vinden zij dat artsen enkel
een therapie moeten toepassen die bij de lokale bevolking goede resultaten geeft (Graham en
Fischbach 2010).
Men heeft ook geprobeerd om opeenvolgende combinatietherapieën toe te passen in plaats van een
drievoudige combinatietherapie. De eerste 14 dagen van deze opeenvolgende therapie wordt een
combinatie van twee antibiotica zoals amoxicilline met clarithromycine of nitroimidazole samen met een
protonpomp inhibitor gegeven. De 5 daaropvolgende dagen bestaan uit een protonpomp inhibitor in
combinatie met amoxicilline en de laatste 5 dagen uit een protonpomp inhibitor samen met
clarithromycine en metronidazole of nitroimidazole (Fuccio et al., 2007; Vaira et al., 2007; Zullo et al.,
2007; Jafri et al., 2008; Graham en Fischbach, 2010; Greenberg et al., 2011; Malfertheiner et al., 2011).
Deze opeenvolgende behandeling blijkt effectiever te zijn dan de drievoudige combinatietherapie (Jafri
et al., 2008; Vaira et al., 2007; Zullo et al., 2007). Een mogelijke verklaring hiervoor is dat de
werkzaamheid van opeenvolgende behandelingen minder wordt beïnvloed door de resistentie van H.
pylori voor clarithromycine dan bij de drievoudige combinatietherapie. Deze behandeling zou de
standaard therapie kunnen worden voor de behandeling van een infectie met H. pylori bij de mens
(Moayyedi, 2007; Vakil en Vaira, 2008).

11
1.2. HELICOBACTER HEILMANNII
Niet enkel H. pylori kan maagproblemen veroorzaken bij de mens. Ook de NHPH-bacteriën zoals H.
heilmannii worden hiermee geassocieerd (Haesebrouck et al., 2009; Vermoote et al., 2011; Joosten et
al., 2013; Flahou et al., 2016). Deze spiraalvormige micro-organismen werden eerst “Gastrospirillum
hominis” genoemd (McNulty et al., 1989). Analyse van hun 16S rRNA gen leidde echter tot de
herclassificatie van deze NHPHs in het geslacht Helicobacter, waarna ze H. heilmannii genoemd
werden, verwijzend naar de Duitse partholoog Konrad Heilmann die als eerste de pathologie
geassocieerd met deze spiraalvormige micro-organismen bestudeerd had (Heilmann en Borchard,
1991). H. heilmannii werd jarenlang gebruikt om te refereren naar de volledige groep van NHPHs die
aanwezig zijn in de maag van mensen, alhoewel er eigenlijk meerdere species hiertoe behoren.
Verdere ontleding van het 16S rRNA gen resulteerde in een herverdeling van deze gastrale
helicobacters in “H. heilmannii type 1” en “H. heilmannii type 2”. Deze twee types verschillen meer dan
3 procent in hun nucleotide sequentie van het 16S rRNA gen. “H. heilmannii” type 1 is identiek aan H.
suis, dewelke van nature uit de maagmucosa van varkens koloniseert. Het is tevens ook de meest
voorkomende NHPH soort bij de mens (O'Rourke et al., 2004; Baele et al., 2008; Vermoote et al., 2011).
H. heilmannii type 2 bestaat niet uit één enkele species, maar uit een groep bacteriën die van nature uit
de maagmucosa van honden en katten koloniseren. Deze groep bacteriën omvat onder andere: H.
bizzozeronii, H. heilmannii, H. felis, H. salomonis, H. cynogastricus, H. baculiformis en H. ailurogastricus
(Haesebrouck et al., 2009; Smet et al., 2011). Om verwarring in verband met de nomenclatuur te
voorkomen spreekt men over H. heilmannii sensu stricto, waarmee men de bacterie species zelf
bedoelt, of H. heilmannii sensu lato, waarmee men refereert naar de groep van gastrale NHPHs.
(Haesebrouck et al., 2011). Naast de gastrale Helicobacter bacteriën afkomstig van katachtigen,
varkens en honden zijn er ook nog 3 andere dier-geassocieerde Helicobacter species die behoren tot
de groep van H. heilmannii sensu lato, namelijk H. acinonychis, afkomstig van wilde katachtigen, H.
cetorum, afkomstig van zeezoogdieren en H. mustelae, afkomstig van fretten. De morfologie van deze
Helicobacter species lijkt meer op die van H. pylori. Deze drie bacteriën zijn voor het eerst beschreven
in de vroege jaren ’90 en in het begin van de 21e eeuw, maar ze werden tot nu toe niet aangetoond bij
de mens (Paster et al., 1991; Harper et al., 2002).

12
Fig. 3. Fylogenetische stamboom van de gastrale Helicobacter soorten gebaseerd op uitgelijnde
en aaneengeschakelde hoofdgenen, uit Liu C, 2016.
Er zijn 3 gastrale clusters te zien. Het eerste cluster bevat de porciene, caniene en feliene helicobacters.
Het tweede cluster bevat H. mustelae en het laatste cluster H. pylori, H. acinonychis en H. cetorum.
Ondanks het feit dat al deze species een typische spiraalvorm hebben, zijn er toch verschillen
beschreven in morfologie tussen deze NHPHs. Een overzicht hiervan wordt weergegeven in tabel 1.
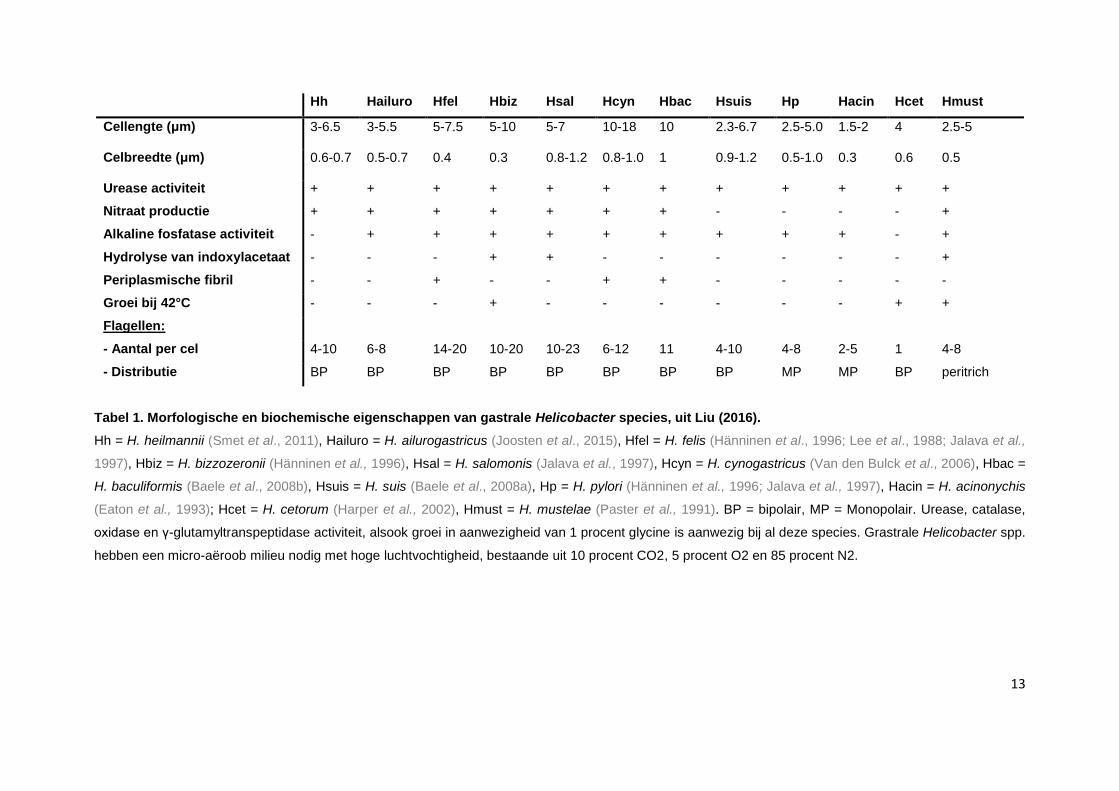
13
Hh Hailuro Hfel Hbiz Hsal Hcyn Hbac Hsuis Hp Hacin Hcet Hmust
Cellengte (μm) 3-6.5 3-5.5 5-7.5 5-10 5-7 10-18 10 2.3-6.7 2.5-5.0 1.5-2 4 2.5-5
Celbreedte (μm) 0.6-0.7 0.5-0.7 0.4 0.3 0.8-1.2 0.8-1.0 1 0.9-1.2 0.5-1.0 0.3 0.6 0.5
Urease activiteit + + + + + + + + + + + +
Nitraat productie + + + + + + + - - - - +
Alkaline fosfatase activiteit - + + + + + + + + + - +
Hydrolyse van indoxylacetaat - - - + + - - - - - - +
Periplasmische fibril - - + - - + + - - - - -
Groei bij 42°C - - - + - - - - - - + +
Flagellen:
- Aantal per cel 4-10 6-8 14-20 10-20 10-23 6-12 11 4-10 4-8 2-5 1 4-8
- Distributie BP BP BP BP BP BP BP BP MP MP BP peritrich
Tabel 1. Morfologische en biochemische eigenschappen van gastrale Helicobacter species, uit Liu (2016).
Hh = H. heilmannii (Smet et al., 2011), Hailuro = H. ailurogastricus (Joosten et al., 2015), Hfel = H. felis (Hänninen et al., 1996; Lee et al., 1988; Jalava et al.,
1997), Hbiz = H. bizzozeronii (Hänninen et al., 1996), Hsal = H. salomonis (Jalava et al., 1997), Hcyn = H. cynogastricus (Van den Bulck et al., 2006), Hbac =
H. baculiformis (Baele et al., 2008b), Hsuis = H. suis (Baele et al., 2008a), Hp = H. pylori (Hänninen et al., 1996; Jalava et al., 1997), Hacin = H. acinonychis
(Eaton et al., 1993); Hcet = H. cetorum (Harper et al., 2002), Hmust = H. mustelae (Paster et al., 1991). BP = bipolair, MP = Monopolair. Urease, catalase,
oxidase en γ-glutamyltranspeptidase activiteit, alsook groei in aanwezigheid van 1 procent glycine is aanwezig bij al deze species. Grastrale Helicobacter spp.
hebben een micro-aëroob milieu nodig met hoge luchtvochtigheid, bestaande uit 10 procent CO2, 5 procent O2 en 85 procent N2.

14
1.2.1. Transmissie
Het blijkt dat mensen die nauw contact hebben met katten, honden of varkens meer kans hebben om
geïnfecteerd te raken met een dier-geassocieerd Helicobacter species (De Bock et al., 2007; Joosten
et al., 2013). Er werd reeds Helicobacter DNA aangetroffen in het speeksel van katten, honden en
varkens. De mondholte van deze dieren kan dus dienen als infectiebron van NHPHs voor de mens
(Casagrande Proietti et al., 2010; Ekman et al., 2013).
Naast direct contact, werden ook andere infectieroutes beschreven. H. felis blijkt bijvoorbeeld in staat
te zijn om een paar dagen in water te overleven, wat de mogelijke rol van water als transmissieroute
benadrukt (Azevedo et al., 2008). De overdracht van H. suis naar de mens zou ook mogelijk kunnen
zijn via consumptie van rauw of niet volledig gaar varkensgehakt aangezien de bacterie hierop aanwezig
kan zijn en in staat is hierin te overleven (De Cooman et al., 2013).
1.2.2. Virulentiefactoren
1.2.2.1. Virulentiefactoren verantwoordelijk voor de kolonisatie van de gastrale mucosa
H. heilmannii is net zoals H. pylori in staat om te overleven in het zure milieu van de maag (Weeks et
al., 2000; Pot et al., 2007). Dit komt doordat deze kiem ook genen heeft die coderen voor de productie
en uitscheiding van urease (Brown, 2000; Smet et al., 2011; Vermoote et al., 2011; Sgouras et al.,
2015).
H. heilmannii is eveneens in staat om met zijn sterk beweeglijke flagellen de mucosa van de maag te
penetreren (Smet et al., 2011; Vermoote et al., 2011). Deze flagellen bevinden zich aan beide uiteinden
van de kiem en zijn opgebouwd uit een lichaam, haak en flagellair filament (O’Toole et al., 1994; Kim et
al., 1999).
Tevens maken deze micro-organismen net zoals H. pylori gebruik van chemotaxis voor een goede
kolonisatie (Foynes et al., 2000; McGee et al., 2005; Terry et al., 2005). Ze bewegen zich namelijk voort
in de richting van ureum, bicarbonaat, cholesterol, arginine en andere aminozuren, maar distantiëren
zich van zoutzuur (O’Toole et al., 1994; Kim et al., 1999). In vergelijking met H. pylori bevatten ze een
groter aantal methyl-accepterende chemotaxis eiwitten, waardoor ze op een bredere range van signalen
uit het milieu kunnen reageren (Arnold et al., 2011; Schott et al., 2011; Smet et al., 2011; Vermoote et
al., 2011). Verder produceert H. heilmanni ook katalase (KatA), superoxide dismutase (SodB) en alkyl
hydroperoxide reductase (HhpC) die de kiem beschermen tegenover zuurstofradicalen (Seyler et al.,
2001; Harris et al., 2002; Olczak et al., 2003; Ernst et al., 2005, Arnold et al., 2011; Schott et al., 2011;
Smet et al., 2011; Vermoote et al., 2011).
De gastrale NHPHs bevatten ongeveer 55 tot 60 OMP’s (Alm et al., 2000; Joosten et al., 2015).
Vergelijkend genetisch onderzoek heeft onthuld dat alle caniene, feliene en porciene helicobacters de
H. pylori Hop en Hom adhesines missen (Joosten et al., 2015). Welke OMPs dan wel betrokken zijn bij
de binding van H. heilmannii, H. ailurogastricus, H. felis, H. bizzozeronii en H. suis aan de gastrale
mucosa is tot nu toe onbekend. Opvallend is wel dat bij de gastrale NHPHs homologen van alle leden
van de H. pylori Hof familie aanwezig zijn, behalve bij H. mustelae (Smet et al., 2014).

15
1.2.2.2. Virulentiefactoren verantwoordelijk voor de inductie van gastrale lesies
Het VacA (zie 1.1.2.2.), dat een belangrijke virulentiefactor is van H. pylori, is afwezig bij H. heilmannii,
H. ailurogastricus, H. felis, H. bizzozeronii, H. suis en H. acynonichis. Homologen van H. pylori VacA
zijn enkel teruggevonden in H. cetorum (Kersulyte et al., 2013; Joosten et al., 2015). Verder is ook het
cagPAI afwezig bij de NHPHs.
Het hierboven reeds besproken GGT (1.1.2.2) is daarentegen wel goed bewaard gebleven binnen het
geslacht Helicobacter. Homologen van andere virulentiefactoren zoals FldA en IceA zijn ook
gedetecteerd in het genoom van H. heilmannii, H. felis, H. bizzozeronnii, H. ailurogastricus, H. suis, H.
acinonychis en H. cetorum (Arnold et al., 2011; Schott et al., 2011; Smet et al., 2011; Vermoote et al.,
2011; Kersulyte et al., 2013; Joosten et al., 2015). Ondanks het feit dat een groot aantal genen die
coderen voor virulentiefactoren beschreven zijn in NHPHs, is voor de meerderheid de rol in de
pathogenese nog steeds onduidelijk.
1.2.3. Symptomen
H. bizzozeronii, H. felis, H. heilmannii, H. salomonis en H. suis zijn in staat om gastritis, gastrale zweren,
duodenale zweren en laaggradige MALT-lymfomen te veroorzaken bij de mens (Haesebrouck et al.,
2009; Iwanczak et al., 2012). In tegenstelling tot H. pylori is er nog geen verband aangetoond tussen
deze micro-organismen en het ontstaan van adenocarcinomen. Verder veroorzaken deze NHPH-
bacteriën een minder erge en minder actieve gastritis dan H. pylori. Er is echter wel een veel groter
risico op het ontwikkelen van een MALT-lymfoom na infectie met NHPHs.
De klinische tekens bij mensen met een NHPH-infectie omvatten onder meer: misselijkheid, braken,
zure oprispingen, haematemesis, epigastrische pijn, onregelmatige defecatie frequentie, onregelmatige
consistentie van de stoelgang, terugkerende spijsverteringsstoornissen, dysfagie en een afgenomen
eetlust (Heilmann en Borchard, 1991; Dieterich et al., 1998; Kaklikkaya et al., 2002; Yoshimura et al.,
2002; Sykora et al., 2003; Van Loon et al., 2003).
Bij geïnfecteerde patiënten kunnen er door middel van endoscopie meerdere laesies waargenomen
worden gaande van een normale tot licht hyperemische mucosa, mucosaal oedeem, nodulaire
inflammatie en de aanwezigheid van zweren in het antrum van de maag of ter hoogte van het duodenum
(Yoshimura et al., 2002; Sykora et al., 2003; Van Loon et al., 2003; Haesebrouck et al., 2009).
Histologisch onderzoek van maagbiopten van geïnfecteerde patiënten wordt gekenmerkt door infiltratie
van lymfocyten en plasmacellen. In enkele gevallen was er zelfs sprake van aggregaten van lymfocyten
of intestinale metaplasieën (Yakoob et al., 2012; Joosten et al., 2013; Matsumoto et al., 2014).
1.2.4. Diagnose
Op dit moment zijn er geen commerciële testen voor handen om een diagnose te stellen van een
gastrale infectie met NHPHs. De ureum ademtest, die vaak als standaard test gebruikt wordt om een
infectie met H. pylori op te sporen, is meestal negatief bij patiënten geïnfecteerd met dier-geassocieerde
gastrale Helicobacter species (Matsumoto et al., 2014). Dit kan verklaard worden door het feit dat de

16
kolonisatie door deze kiemen, in tegenstelling tot H. pylori, zeer focaal is en het aantal kiemen meestal
relatief laag is (Solnick en Schauer, 2001). Deze kiemen zijn bijzonder moeilijk te kweken in het
laboratorium en kiemisolatie uit een maagbiopt is geen optie voor routinematige diagnostiek. Tot nu toe
heeft men trouwens enkel H. bizzozeronii (Andersen et al., 1999; Kivistö et al., 2010) en H. felis
(Wüppenhorst et al., 2013) kunnen isoleren uit maagbiopten van geïnfecteerde mensen. Daarom is
analyse van gastrale biopten met behulp van histologie en moleculaire methodes op dit moment de
enige manier om infecties met deze pathogenen vast te stellen (Haesebrouck et al., 2009).
1.2.5. Behandeling
Voor patiënten met erge maagklachten, waarbij een infectie met NHPHs werd aangetoond, is een
behandeling noodzakelijk. Op dit moment is er echter een tekort aan klinische onderzoeken en slechts
enkele rapporten beschrijven de antimicrobiële gevoeligheid van deze NHPHs (Vermoote et al., 2011;
Blaecher et al., 2013). Hierdoor wordt er nu, net zoals voor de behandeling van infectie met H. pylori,
gebruik gemaakt van een protonpomp inhibitor in combinatie met twee verschillende soorten antibiotica,
waarbij de voorkeur uitgaat naar clarithromycine en amoxicilline. Desalniettemin lijkt de behandeling van
zowel infecties met H. pylori als met NHPHs meer en meer te falen door de toenemende
antibioticaresistentie (Horiki et al., 2009; Graham en Fischbach, 2010; Kivistö et al., 2010). Vermoote et
al. (2011) toonden bijvoorbeeld aan dat een H. suis bacterie, geïsoleerd uit de maag van een varken,
verworven resistentie vertoonde tegenover fluoroquinolones. Ook zagen ze dat bij alle geteste H. suis
stammen de minimale inhibitorische concentratie (Minimal Inhibitory Concentration; MIC) voor
ampicilline hoger was dan bij H. felis, H. bizzozeronii, H. salomonis en H. pylori. Dit zou erop kunnen
wijzen dat H. suis intrinsiek minder gevoelig is voor aminopenicillines dan andere gastrale Helicobacter
species. De significantie van deze vaststelling voor de behandeling van H. suis infecties is momenteel
nog niet gekend (Vermoote et al., 2011).

17
2. DE ZIEKTE VAN PARKINSON
2.1. INLEIDING
Het idee dat darmbacteriën een drijvende kracht zijn voor de rijping van het immuunsysteem van de
gastheer is al langer bekend. Het feit dat bacteriën ook de hersenfunctie en het gedrag zouden kunnen
beïnvloeden werd eerst als minder waarschijnlijk beschouwd. Er zijn echter wel verscheidene publicaties
die aantonen dat patiënten na antibiotica gebruik voor de eradicatie van darmbacteriën een psychose
ontwikkelden (Plana et al., 2006; Weis et al., 2006; Grimm en Alm, 2007; Ozsoylar et al., 2007; Bell et
al.,2008; Murphy et al., 2008; Mehdi, 2010). Tevens hebben enkele klinische studies aangetoond dat
mensen met autisme een andere darmmicrobiota samenstelling hebben (Shaw et al.,1995; Bolte,1998;
Parracho et al., 2005).
Op dit ogenblik wordt het bewijs dat er een verband zou kunnen zijn tussen micro-organismen en de
hersen-biochemie uit dierproeven gehaald. Wat de Helicobacter species betreft werd er tot nu toe enkel
een verband aangetoond tussen H. pylori en enkele extragastrale ziekten. Bercik et al. toonden
bijvoorbeeld aan dat een chronische infectie met H. pylori bij muizen de fysiologie van de maag kan
veranderen wat aanleiding gaf tot een vertraagde maaglediging en een toegenomen viscerale
sensitiviteit. Verder zag men dat niet enkel de maag werd aangetast. Er werden namelijk ook
veranderingen ter hoogte van het centraal zenuwstelsel waargenomen bij de geïnfecteerde muizen,
waardoor ze frequenter kleinere hoeveelheden voedsel opnamen, tegenover de controlegroep (Bercik
et al, 2012). Na eradicatie van de kiem hield het afwijkende gedrag van de geïnfecteerde muizen voor
minstens twee maanden aan. Dit doet vermoeden dat veranderingen geïnduceerd door chronische
infecties met H. pylori in het centraal zenuwstelsel langdurig tot zelfs permanent aanwezig kunnen zijn
(Dobbs et al., 2010; Bercik et al, 2012; Alvarez-Arellano en Maldonado-Bernal, 2014).
Op dit moment werd er, zoals hierboven beschreven, reeds een mogelijke link aangetoond tussen H.
pylori infecties en verscheidene neurodegeneratieve aandoeningen. In deze thesis zal enkel de relatie
tussen een infectie met Helicobacter en PD uitgebreid behandeld worden.
In 1817 werd PD voor het eerst beschreven door James Parkinson (Parkinson, 1817).
PD is een algemeen voorkomende ouderdoms gerelateerde neurodegeneratieve aandoening die zelden
voorkomt bij mensen jonger dan vijftig jaar (Lau en Breteler, 2006; Hirsch et al., 2013; Kleinman en
Frank, 2013). De ziekte wordt gekarakteriseerd door de redelijk selectieve celdood van voornamelijk de
dopamine producerende neuronen. Mensen met PD kunnen zowel motorische als non-motorisch
symptomen vertonen (Parkinson, 1817; Fahn en Przedborski, 2010; Mhyre et al., 2012; Hirsch et al.,
2013).
2.2. OORZAKEN Op dit moment is de etiologie van PD bij 90 tot 95 procent van de gevallen nog niet bekend. Deze is
allicht multifactorieel, waarbij zowel genetische als milieufactoren een rol spelen (Wirdefelt et al., 2011;
Mhyre et al., 2012). Slechts bij 5 tot 10 procent van de mensen die leiden aan PD, is de ziekte het
resultaat van een specifieke monogenetische mutatie (Lau en Breteler, 2006; Tansey et al., 2007;

18
Kleinman en Frank, 2013). Voorbeelden van deze mutaties zijn: mutaties in het gen coderend voor alfa-
synucleïne (PAK1/4, SNCA), mutaties in het Leuncin-Rich Repeat Kinase 2 (LRRK2) coderende gen,
mutaties in de Parkin (PARK2) locus en in het fosfaat en Tensine Homologe (PTEN)-geïnduceerde
Putative Kinase 1 (PINK1) gen (Wirdefelt et al., 2011; Mhyre et al., 2012). De mutaties in het gen
coderend voor LRRK2 worden geassocieerd met de dominante overerfbare vorm van PD, terwijl de
mutaties in genen coderend voor PINK en parkin geassocieerd worden met de recessieve erfelijke vorm
van PD (Tansey et al., 2007). De neuronale degeneratie die optreedt bij patiënten met PD kan ontstaan
als respons op een mengeling van schadelijke mechanismen die plaatsvinden zowel binnenin (cel
autonome processen) als buiten de degeneratieve neuronen (niet-cel autonome processen) (Hirsch et
al., 2013). Hiertoe behoren inflammatoire processen, die gekarakteriseerd worden door oxidatieve
stress, accumulatie van cytokines en activatie van microgliacellen, wat aanleiding kan geven tot afbraak
van de nigrostriatale banen en een versneld ziekteverloop. Er wordt aangenomen dat dit voornamelijk
kan leiden tot PD bij patiënten met de idiopathische vorm, aangezien de etiologie bij hen niet gekend is
(Lau en Breteler, 2006; Dobbs et al., 2008; Kleinman en Frank, 2013).
2.2.1. Genetische factoren De meest frequent bestudeerde genen zijn deze betrokken bij het mitochondriale- en dopaminerge
metabolisme, detoxificatie en andere neurodegeneratieve ziekten. Ook is er onderzoek gedaan naar
genen die geassocieerd worden met vermoedelijke risicofactoren voor PD, zoals lipoproteïne,
hormonen en het homocysteïne metabolisme (Tan et al., 2000; Maraganore et al., 2002; Healy et al.,
2004; Huang et al., 2004; Tan et al., 2004; Lau et al., 2005). Op dit moment is het grootste nadeel van
deze studies hun ontoereikende statistieke kracht (Lau en Breteler, 2006).
Wanneer PD voorkomt in de familie, evenals dementie, essentiële tremor en psychiatrische stoornissen,
wordt dit geassocieerd met een groter risico op het ontwikkelen van PD. Uit bepaalde studies blijkt dat
mannen een 1,5 keer groter risico lopen dan vrouwen (Twelves et al., 2003; Hirtz et al., 2007; Wirdefelt
et al., 2011; Kleinman en Frank, 2013). Dit verhoogde risico zou kunnen verklaard worden doordat
oestrogenen mogelijks een beschermende functie bieden aan zenuwen, maar op dit moment is de rol
van oestrogenen nog steeds controversieel. Volgens andere studies is er namelijk geen significant
verschil tussen vrouwen of mannen (Morgante et al., 1992; Tison et al., 1994; De Rijk et al., 1995; De
Rijk et al., 1997; Lau en Breteler, 2006).
Een studie met ongeveer 20.000 mannelijke tweelingen heeft aangetoond dat er een significant effect
was van genetische factoren op het ontwikkelen van PD. Dit geldt vooral voor PD waarbij de symptomen
al voor de leeftijd van vijftig optreden (Tanner et al., 1999).
2.2.2. Milieufactoren Het ontwikkelen van PD kan onder meer te wijten zijn aan een eerder opgelopen hoofdtrauma,
blootstelling aan zware metalen, veelvuldig contact met pesticiden of herbiciden (Lau en Breteler, 2006;
Wirdefelt et al., 2011; Kleinman en Frank, 2013). Blootstelling aan zware metalen zoals ijzer, mangaan,
koper, lood, amalgaam, aluminium of zink zou het risico op PD kunnen verhogen omdat de accumulatie

19
van metalen in de substantia nigra aanleiding kan geven tot oxidatieve stress. Er zou echter nog meer
onderzoek hieromtrent moeten gedaan worden om dit te bevestigen (Wirdefelt et al., 2011; Mhyre et al.,
2012).
Bepaalde studies hebben de consumptie van koffie in verband gebracht met een verminderd risico op
het ontwikkelen van PD (Ross et al., 2000; Wirdefelt et al., 2011; Mhyre et al., 2012). Dit zou
voornamelijk te wijten zijn aan de cafeïne wat een inhibitor is van de adenosine A2 receptor. Het roken
van sigaretten blijkt ook het risico op PD te verminderen (Lau en Breteler, 2006). De meest logische
verklaring hiervoor is dat nicotine de vrijstelling van dopamine stimuleert en zo werkt als een antioxidant
(Quik, 2004). Tevens blijkt het chronisch gebruiken van niet-steroïdale anti-inflammatoire
geneesmiddelen (Non-Steroidal Anti-Inflammatory Drugs; NSAID) het risico op het ontwikkelen van PD
te verminderen (Tansey et al., 2007). Er is echter nog meer inzicht nodig in deze factoren om de
pathogenese beter te kunnen begrijpen en om effectieve behandelingen te kunnen ontwikkelen. Op dit
moment zijn er namelijk nog veel tegenstrijdigheden over deze niet genetische oorzaken en hun rol in
de ontwikkeling van PD (Lau en Breteler, 2006).
2.3. PATHOGENENSE
PD is, op de ziekte van Alzheimer na, de tweede meest voorkomende neurodegeneratieve aandoening
in de huidige humane populatie. Het neurologische proces wordt gekarakteriseerd door de degeneratie
van dopamine producerende neuronen ter hoogte van de pars compacta in de substantia nigra
(Wirdefeldt et al., 2011; Mhyre et al., 2012). Deze degeneratie kan leiden tot activatie van de
microgliacellen, de macrofagen van het centraal zenuwstelsel (Central Nervous System; CNS) (Mhyre
et al., 2012). Verder zou perifere inflammatie ook kunnen bijdragen tot de activatie van de
microgliacellen, vooral wanneer de hersenbarrières verstoord zijn (Alvarez-Arellano en Maldonado-
Bernal, 2014).
Fig. 4. De neurotransmitter dopamine zendt signalen uit tussen de neuronen in de hersenen die
gerelateerd zijn aan plezier, beloning en motorische functies. Een tekort aan dopamine wordt in
verband gebracht met bewegingsstoornissen zoals bij de ziekte van Parkinson.
Bron: Cohn, 2013
20
Ook wordt PD gekenmerkt door
zogenaamde Lewy bodies. Deze worden
geïdentificeerd als cytoplasmatische,
eosinofiele inclusielichaampjes in de
neuronen en dan vooral in de overlevende
neuronen. Ze ontstaan door de accumulatie
van intracellulaire eiwit aggregaten,
waaronder het alfa-synucleïne eiwit (Gibb
en Lees, 1988; Wirdefeldt et al., 2011;
Mhyre et al., 2012; Kannarkat et al., 2013).
Mutaties in genen coderend voor dit eiwit
worden geassocieerd met een dominante
vorm van erfelijke PD (Tansey et al.,
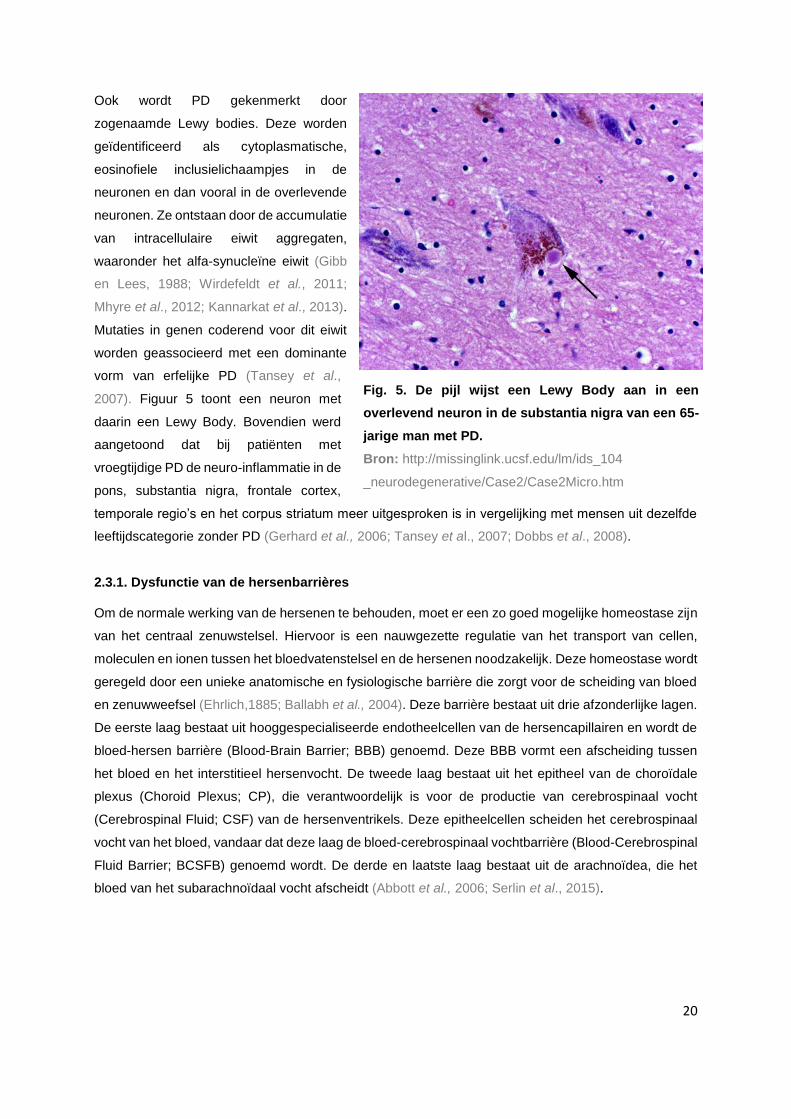
2007). Figuur 5 toont een neuron met
daarin een Lewy Body. Bovendien werd
aangetoond dat bij patiënten met
vroegtijdige PD de neuro-inflammatie in de
pons, substantia nigra, frontale cortex,
temporale regio’s en het corpus striatum meer uitgesproken is in vergelijking met mensen uit dezelfde
leeftijdscategorie zonder PD (Gerhard et al., 2006; Tansey et al., 2007; Dobbs et al., 2008).
2.3.1. Dysfunctie van de hersenbarrières Om de normale werking van de hersenen te behouden, moet er een zo goed mogelijke homeostase zijn
van het centraal zenuwstelsel. Hiervoor is een nauwgezette regulatie van het transport van cellen,
moleculen en ionen tussen het bloedvatenstelsel en de hersenen noodzakelijk. Deze homeostase wordt
geregeld door een unieke anatomische en fysiologische barrière die zorgt voor de scheiding van bloed
en zenuwweefsel (Ehrlich,1885; Ballabh et al., 2004). Deze barrière bestaat uit drie afzonderlijke lagen.
De eerste laag bestaat uit hooggespecialiseerde endotheelcellen van de hersencapillairen en wordt de
bloed-hersen barrière (Blood-Brain Barrier; BBB) genoemd. Deze BBB vormt een afscheiding tussen
het bloed en het interstitieel hersenvocht. De tweede laag bestaat uit het epitheel van de choroïdale
plexus (Choroid Plexus; CP), die verantwoordelijk is voor de productie van cerebrospinaal vocht
(Cerebrospinal Fluid; CSF) van de hersenventrikels. Deze epitheelcellen scheiden het cerebrospinaal
vocht van het bloed, vandaar dat deze laag de bloed-cerebrospinaal vochtbarrière (Blood-Cerebrospinal
Fluid Barrier; BCSFB) genoemd wordt. De derde en laatste laag bestaat uit de arachnoïdea, die het
bloed van het subarachnoïdaal vocht afscheidt (Abbott et al., 2006; Serlin et al., 2015).
Fig. 5. De pijl wijst een Lewy Body aan in een
overlevend neuron in de substantia nigra van een 65-
jarige man met PD.
Bron: http://missinglink.ucsf.edu/lm/ids_104
_neurodegenerative/Case2/Case2Micro.htm

21
2.3.2. Bloed-hersen barrière De BBB vormt een soort filter tussen de
systemische circulatie en het
hersenparenchym. Op die manier wordt er
bepaald wat er wel en niet mag
binnentreden in de hersenen. Hierdoor blijft
de gespecialiseerde samenstelling van het
interstitiële hersenvocht behouden (Ballabh
et al., 2004).
De BBB bestaat uit een enkelvoudige laag
endotheelcellen afkomstig van de
hersencapillairen. Deze endotheelcellen
onderscheiden zich van andere
endotheelcellen in de rest van het
lichaam doordat ze geen fenestraties
hebben. Verder hebben ze meer tight
junctions (Tight Junctions; TJ) en doen ze
minder aan vesiculaire pinocytose. Dit
samen zorgt voor de restrictieve functie van de BBB. De vele TJ van de endotheelcellen rondom de
capillairen sluiten de intercellulaire openingen af en gaan zo paracellulaire diffusie van ionen, opgeloste
stoffen, proteïnen, water en bloedcellen tegen. Zuurstof en koolstofdioxide zijn kleine lipofiele
substanties die via de plasmamembranen wel vrij heen en weer kunnen bewegen aan de hand van een
concentratiegradiënt (Grieb et al., 1985; Ballabh et al., 2004; Abbott, 2014; Serlin et al., 2015; De Bock
et al., 2016). Voedingsstoffen zoals glucose en aminozuren kunnen de hersenen binnentreden via
transportsystemen, terwijl grotere moleculen inclusief insuline, leptine en ijzer-transferrine worden
opgenomen via receptor gemedieerde endocytose (Pardridge et al., 1985; Zhang en Pardridge, 2001).
De BBB bestaat naast de endotheelcellen ook nog uit astrocytenvoetjes, die de bloedvaten afschermen,
en de basaalmembraan met daarin de pericyten. Deze laatste spelen een hoofdrol in de samenstelling
van de endotheliale TJ en de angiogenese, de structurele volledigheid en de differentiatie van de
bloedvaten (Ballabh et al., 2004; Abbott, 2014; Serlin et al., 2015).
Bij patiënten die leiden aan PD werd een toegenomen doorlaatbaarheid van de BBB waargenomen
(Stolp en Dziegielewska, 2009; Nielsen et al., 2012). Hierbij zouden zowel lokale als systemische
inflammatie een rol kunnen spelen. Gedurende het inflammatie proces worden er bepaalde cytokines
vrijgesteld zoals interleukine-1-beta (Interleukin; Il), Il-6 en tumornecrosefactor-alfa (Tumor Necrosis
Factor-Alpha; TNF-α) die kunnen zorgen voor veranderingen in de TJ, waardoor de doorlaatbaarheid
van de BBB toeneemt. Deze toegenomen permeabiliteit, samen met het verhoogde expressieniveau
van de cytokines en andere inflammatiemediatoren, kan vervolgens aanleiding geven tot neuro-
inflammatie en oxidatieve stress. Hierdoor vindt er verstoring van de BBB plaats wat kan resulteren in
hersenbeschadiging (Stolp en Dziegielewska, 2009).
Fig. 6. Geïllustreerde weergave van de bloed-hersen
barrière, een multicellulaire structuur bestaande uit
endotheelcellen afkomstig van de hersencapillairen,
tight junctions, basaalmembraan, astrocytenvoetjes
en pericyten.
Bron: http://home.szbk.u-szeged.hu/~krizbai/index.html

22
2.3.3. Bloed-cerebrospinaalvocht barrière Deze barrière bestaat uit twee delen. Het eerste deel is de CP, gevasculariseerd weefsel, dat in alle
hersenventrikels aanwezig is (Laterra et al., 1999; Serlin et al., 2015). Verder bestaat het functionele
deel van de CP uit een bloedvat, omgeven door een laag van gedifferentieerd ependym epitheelweefsel.
De bloedvaten van de CP zijn gefenestreerd en de endotheelcellen hebben geen TJ in vergelijking met
deze van de BBB. Het endotheel vormt dus geen barrière voor verplaatsing van kleine moleculen uit het
bloed. Om dit transport tegen te gaan wordt de BCSFB afgesloten door epitheelcellen die onderling
verbonden zijn door TJ. Het tweede deel van de BCSFB is de arachnoïdale membraan, die de hersenen
bedekt. De cellen van deze membraan zijn ook verbonden door TJ (Laterra et al., 1999).
Aangezien de barrière, gevormd door de epitheelcellen van de CP, minder sterk is dan de BBB, is de
BCSFB gevoeliger voor systematische veranderingen en inflammatie (De Bock et al., 2014). Daarom
wordt PD ook geassocieerd met een toegenomen doorlaatbaarheid van de BCSFB (Pisani et al., 2012).
2.3.4. Junctions in de hersenbarrières Het junctionele complex van de BBB bevat TJ en adherens junctions (Adherens Junctions; AJ). De TJ
komen structureel voor ter hoogte van de buitenzijden van de plasmamembranen van aangrenzende
endotheelcellen om deze met elkaar te verbinden. TJ bestaan uit een variabel aantal intercellulaire
vertakte strengen en zijn opgebouwd uit drie integrale membraaneiwitten namelijk claudine, occludine
en junctionale adhesie moleculen. Verder bestaan TJ nog uit een groot aantal bijkomende
cytoplasmatische eiwitten zoals zonula occludens-1 (Zonula occludens-1; Zo-1), Zo-2, Zo-3 en
cinguline. Deze cytoplasmatische eiwitten koppelen de membraaneiwitten (claudine en occludine) aan
actine. Actine is het primaire proteïne van het cytoskelet dat de structurele en functionele integriteit van
het endotheel onderhoudt (Ballabh et al., 2004). Claudine eiwitten van aangrenzende endotheelcellen
binden aan elkaar om zo een eerste afsluiting te vormen. Verder hechten de carboxy uiteinden van de
claudine eiwitten zich vast aan de cytoplasmatisch eiwitten (Furuse et al., 1999).
Occludine is significant groter dan claudine waardoor het cytoplasmatische deel van occludine
rechtstreeks verbonden is met de Zo-eiwitten. De expressie van occludine is veel hoger in het endotheel
van de hersenen dan in ander endotheel. Occludine is een regulerend eiwit dat de paracellulaire
doorlaatbaarheid kan wijzigen (Hirase et al., 1997).
Occludine en claudine vormen samen heteropolymeren en zo ontstaan intramembraneuze strengen die
doorlaatbare kanalen bevatten om zo selectieve diffusie van ionen en hydrofiele moleculen toe te laten
(Matter en Balda, 2003).
AJ daarentegen bevatten het membraaneiwit cadherine dat in verbinding staat met het actine cytoskelet
via intermediaire eiwitten, namelijk de catenines. Op die manier worden nauwe contacten gevormd
tussen de aangrenzende cellen. AJ verbinden zich onderling tussen de extracellulaire calcium-
afhankelijke domeinen van cadherine aan de oppervlakte van aangrenzende cellen (Matter en Balda,
2003; Ballabh et al., 2004).

23
Fig. 7. Schematische weergave van de eiwitinteractie van de TJ en AJ van de bloed-hersen
barrière.
Bron: Ballabh et al., 2004
2.4. SYMPTOMEN
Er zijn zowel motorische als non-motorische symptomen die kunnen voorkomen bij PD. De vier
motorische hoofdsymptomen van PD zijn: tremor tijdens rust, rigiditeit, akinesie (of bradykinesie) en
houdingsinstabiliteit. Er kunnen echter ook bijkomende symptomen optreden, namelijk een gebogen
houding en het zogenaamde bevriezen door blokkade van de motorneuronen (Parkinson, 1817;
Jankovic, 2008; Fahn en Przedborski, 2010; Wirdefelt et al., 2011, Hirsch et al., 2013; Kannarkat et al.,
2013). Een aanhoudende tremor is het meest voorkomende symptoom. De tremors kunnen voorkomen
ter hoogte van de handen, lippen, kin, kaken en benen (Jankovic, 2008). Bradykinesie betekent een
vertraagde beweging en is het meest karakteristieke klinische symptoom van PD. De pathofysiologie
van bradykinesie is nog niet goed gekend, maar het hangt waarschijnlijk samen met de graad van
dopamine deficiëntie (Vingerhoets et al., 1997). Bradykinesie omvat onder andere moeilijkheden met
plannen, starten en uitvoeren van een beweging en moeite om achtereenvolgende of gelijktijdige taken
uit te voeren (Berardelli et al., 2001). Andere uitingen van bradykinesie zijn het verlies van het uitvoeren
van spontane bewegingen en gebaren. Tevens kan men gaan kwijlen door een verminderd
slikvermogen (Bagheri et al., 1999). Ook monotonische en hypotonische spraakstoornissen, verlies van
gezichtsuitdrukking, verminderd knipperen en minder bewegen met de armen tijdens het lopen kunnen
een gevolg zijn van PD (Jankovic, 2008).
Bij mensen met PD kunnen ook non-motorische symptomen voorkomen. Dertig tot veertig procent heeft
te kampen met dementie, apathie, angststoornissen, psychosen en depressie. Tachtig tot negentig
procent leidt aan slaapstoornissen en tot negentig procent van de patiënten ondervindt een veranderde

24
olfactorische perceptie. Verder worden er ook frequent autonome afwijkingen waargenomen bij PD
patiënten, zoals cardiovasculaire, gastro-intestinale en urogenitale stoornissen (Mhyre et al., 2012).
2.5. DIAGNOSE
Op dit moment is er nog geen betrouwbare diagnostische test om PD aan te tonen (Lau en Breteler,
2006; Jankovic, 2008). Momenteel wordt de diagnose gesteld aan de hand van de klinische tekens:
aanhoudende tremor, bradykinesie, rigiditeit of houdingsinstabiliteit. Wanneer iemand met minstens 2
van deze symptomen te kampen heeft, spreekt men over PD (Gelb et al., 1999; Litvan et al., 2003; Lau
en Breteler, 2006; Jankovic, 2008). Verder dient men ook andere potentiële oorzaken voor secundaire
PD uit te sluiten. Een asymmetrische start van de symptomen en een goede reactie op behandeling met
levodopa zijn ondersteunend voor het stellen van de diagnose en worden gezien als de belangrijkste
criteria om PD te onderscheiden van andere aandoeningen (Litvan et al., 2003; Lau en Breteler, 2006;
Tolosa et al., 2006; Jankovic, 2008).
Geavanceerde beeldvorming met enkelvoudige-proton-emissie computer tomografie (Computer
Tomography; CT) of positron emissie tomografie (Positron Emission Tomography; PET) is eventueel
waardevol voor het stellen van de diagnose van PD in gespecialiseerde omstandigheden. De
bruikbaarheid van deze technieken voor populatie gebaseerd epidemiologisch onderzoek is echter nog
steeds beperkt, alhoewel ze steeds meer beschikbaar en makkelijker in gebruik worden (Litvan et al.,
2003; Lau en Breteler, 2006).
Langdurige opvolging van patiënten zal de diagnostische nauwkeurigheid verbeteren aan de hand van
informatie die verworven wordt gedurende het verloop van de ziekte, zoals het verschijnen van
bijkomende symptomen en de respons op levodopa therapie (Litvan et al., 2003; Lau en Breteler, 2006).
Klinische criteria leiden slechts tot een waarschijnlijke diagnose van PD. Volledige zekerheid kan slechts
bekomen worden na het uitvoeren van een post-mortem onderzoek. Klinische pathologische studies
toonden aan dat bij tachtig tot negentig procent van de mensen die al een klinische diagnose hadden,
PD bevestigd werd na autopsie (Litvan et al., 2003; Lau en Breteler, 2006).
2.6. PREVALENTIE
PD is een leeftijd gerelateerde ziekte die zelden voorkomt bij mensen jonger dan vijftig jaar (Schoenberg
et al., 1988; Mayeux et al.,1995; Lau en Breteler, 2006; Hirsch et al., 2013; Kleinman en Frank, 2013;
Pringsheim et al., 2014). Aangezien de levensverwachting van mensen in de Westerse wereld over het
algemeen aan het toenemen is, kan men ook een toename van patiënten met PD verwachten (Dorsey
et al., 2007).
Het voorkomen van PD in geïndustrialiseerde landen wordt geschat op 0,3 procent bij de hele populatie,
gaande van 1 procent bij mensen boven de zestig jaar (Fahn, 2003; Nussbaum en Ellis, 2003; Lau en
Breteler, 2006) tot 4-5 procent bij de populatie boven de 85 jaar (Fahn, 2003; Lau en Breteler, 2006).
De prevalentie van PD werd ook reeds bepaald in cross culturele studies om zo te bepalen of verschillen
in milieu of in genen geassocieerd met de ziekte het risico beïnvloeden (Van Den Eeden et al., 2003).
Hieruit blijkt dat PD mogelijks minder voorkomt bij mensen uit Afrika en Azië. Echter zijn de resultaten

25
vaak niet betrouwbaar omdat er grote verschillen zijn in respons ratio’s, overleving en omschrijving van
de oorzaak. Hierdoor weet men niet zeker of er echt een verschil is in het voorkomen van de ziekte
tussen verschillende etnische groepen (Schoenberg et al., 1988; Mayeux et al.,1995; Morens et al.,
1996; Van Den Eeden et al., 2003).
2.7. BEHANDELING
Op dit moment bestaat er nog geen geneesmiddel of chirurgische methode om de progressie van de
ziekte te vertragen. Er kan enkel op verschillende manieren geprobeerd worden om de symptomen te
verminderen (Fahn, 2003).
De meest gebruikte medische behandeling is momenteel de dopamine substitutie therapie met het
catecholamine levodopa. Dit is een sterke dopamine-agonist die wordt toegediend in combinatie met
een perifere decarboxylase inhibitor om de vroegtijdige vorming van dopamine in de perifere weefsels
te verhinderen. Alhoewel levodopa het meest effectieve geneesmiddel is om de symptomen te
onderdrukken, krijgt ongeveer zestig procent van de patiënten te maken met complicaties zoals het
intermitterend niet reageren op de therapie, wat ook wel het “wearing-off” effect genoemd wordt. Tevens
ontwikkelen sommige patiënten dyskinesie wanneer ze al meer dan 5 jaar levodopa gebruiken.
Patiënten jonger dan zestig jaar zijn gepredisponeerd om deze problemen al eerder te ontwikkelen
(Fahn, 2003).
Er zijn ook nog andere dopamine-agonisten die men kan gebruiken als behandeling van PD. De sterkste
dopamine-agonist is apomorfine. Een nadeel van deze agonist is dat deze sublinguaal dient ingenomen
te worden of ingespoten moet worden. De andere agonisten kunnen per os genuttigd worden. Pergolide,
pramipexole en ropinirole zijn allemaal even effectief en ze hebben een betere werking dan
bromocriptine. Cabergoline heeft de langste halfwaardetijd en is daardoor handig in gebruik. Een nadeel
van deze agonisten, in vergelijking met levodopa, is dat ze voornamelijk bij ouderen sneller hallucinaties,
verwarring en psychosen veroorzaken. Daarom is het veiliger om patiënten die ouder zijn dan zeventig
jaar enkel te behandelen met levodopa. Bovendien kunnen de dopamine agonisten na langdurig gebruik
ook aanleiding geven tot slaperigheid en oedeem ter hoogte van de benen. Anderzijds blijkt uit
gecontroleerde klinische studies dat dopamine agonisten minder kans op slaperigheid geven en het
“wearing-off” fenomeen minder vaak veroorzaken dan levodopa (Parkinson Study Group, 2000; Rascol
et al., 2000; Fahn, 2003).
Er wordt ook steeds vaker gebruik gemaakt van een chirurgische behandeling aangezien er steeds
meer nieuwe technieken op het gebied van elektrostimulatie zijn ontwikkeld en de fysiologie van de
basale ganglia steeds beter begrepen wordt. Een methode waarvan gebruik gemaakt wordt, is de
stereotaxische diepe hersenstimulatie (Deep brain stimulation; DBS), waarbij stimulerende elektrodes
op het hoofd van de patiënt geplaatst worden. De locatie in de hersenen waarop de behandeling wordt
toegepast, verschilt van patiënt tot patiënt. DBS van de subthalamische nucleus lijkt de meest belovende
resultaten te geven, aangezien dit zorgt voor verminderen van de bradykinesie, waardoor de dosering
van levodopa gereduceerd kan worden. Deze behandeling verlaagt tevens de mate van de dyskinesie
(Fahn, 2003).

26
3. BETROKKENHEID VAN HELICOBACTER INFECTIES IN HET VERLOOP VAN DE ZIEKTE VAN
PARKINSON
Constipatie als gevolg van niet frequente darmbewegingen en trage darmpassage (Abbott et al., 2001;
Dobbs et al., 2016) is beschreven als een van de meest bijkomende problemen van PD (Parkinson,
1817; Dobbs et al., 2016).
Recent is er een hoge prevalentie van H. pylori infecties beschreven in PD patiënten (Rees et al., 2011;
Nielsen et al., 2012). Bijkomend werd een drie keer hogere seropositiviteit voor H. pylori waargenomen
onder PD patiënten in vergelijking met controle patiënten (Charlett et al., 1999). Dezelfde studie laat
zien dat broers en zussen die allebei leiden aan PD, meer kans hebben op het vertonen van PD achtige
symptomen als ze bijkomend ook positief testen op H. pylori serologie. Verder zijn de titers van H. pylori
antilichamen vijf maal hoger bij patiënten ouder dan tachtig jaar die PD symptomen vertonen, in
vergelijking met een controlegroep van dezelfde leeftijd, zonder PD symptomen (Weller et al., 2005).
Na een eradicatie-therapie van H. pylori bij patiënten met PD is er over het algemeen een verbetering
van de motorische symptomen en levenskwaliteit (Dobbs et al., 2010). Dergelijke patiënten reageren
ook beter op de levodopa therapie.
In een recente studie werd een tien maal hogere frequentie van H. suis infecties gevonden bij patiënten
die leden aan PD dan bij controle patiënten, zonder symptomen van deze neurologische aandoening
(Blaecher et al., 2013).
Het blijkt dus nuttig om de invloed van een infectie met Helicobacter op het verloop van PD verder te
bestuderen en het mechanisme hierachter verder te ontrafelen.

27
DEEL 2: DOELSTELLING Ons microbioom vervult bepaalde belangrijke functies in het lichaam. De grote bacteriële diversiteit van
het spijsverteringsstelsel kan bijvoorbeeld een invloed hebben op onze gezondheid, ons algemeen
welzijn en onze gemoedstoestand. Mogelijks speelt deze bacteriële diversiteit ook een grote rol in allerlei
ziektes, waaronder neurodegeneratieve aandoeningen. De spijsverteringsstelsel-hersenen as kreeg
dan ook recent veel aandacht. De term spijsverteringsstelsel-hersenen as verwijst naar een
bidirectionele communicatie route tussen het gastro-intestinaal stelsel en de hersenen. De verstoring
van deze as kan pathofysiologische gevolgen hebben. Zo werd bijvoorbeeld bij 27% van de patiënten
met PD een H. suis infectie teruggevonden, ten opzichte van slechts 2% bij de controlegroep. In
hoeverre een infectie met een Helicobacter bacterie invloed heeft op de pathologie van deze
neurodegeneratieve aandoening werd reeds gedeeltelijk onderzocht door Blaecher et al. (niet
gepubliceerde data). Men onderzocht of een gastrale infectie met H. suis een impact kan hebben op de
homeostase van de hersenen bij muizen. Eén maand na infectie zag men dat H. suis de hersenen
mogelijks beïnvloedde via de inductie van systemische inflammatie. Dit ging gepaard met een gestegen
expressie van het Il1β in de maag, het serum en de CP. Verder was de integriteit van de BCSFB sterk
verstoord wat gepaard ging met een gedaalde expressie van de TJ Claudine-1 en Occludine. Hieruit
blijkt dat systemische inflammatie mogelijks bijdraagt tot de ontwikkeling van de ziekte. De ontsteking
van de hersenen kan mede veroorzaakt zijn doordat de integriteit van de verschillende hersenbarrières,
waaronder de BBB en de BCSFB, aangetast werd. Het achterhalen van de onderliggende oorzaken van
deze inflammatie kan de basis vormen van onderzoek naar een effectieve behandeling van
neurodegeneratieve aandoeningen zoals PD.
In deze masterproef zal worden nagegaan wat het effect is van een infectie met bepaalde Helicobacter
species op de integriteit van de maag- en hersenbarrières en de daaruit volgende invloed op het verloop
van PD.
Er zal gewerkt worden met het wildtype H. heilmannii sensu stricto en het wildtype H. pylori. Daarnaast
zal er ook gebruik gemaakt worden van gemuteerde varianten van deze Helicobacter species. Deze
missen een specifiek virulentie-gen, coderend voor één van de Hof OMPs en zijn bijgevolg niet meer in
staat de maag goed te koloniseren. In alle onderzochte opstellingen zal nagegaan worden of er een
verschil is tussen de controlegroep en de geïnfecteerde groepen. Ook zal er worden nagegaan of er
een verschil is tussen de muizen die geïnfecteerd zijn met het wildtype species en deze geïnfecteerd
met de gemuteerde varianten.
Als eerste zal de invloed van de infectie op de integriteit van de BBB en de BCSFB onderzocht worden.
Hiervoor wordt eerst kwantitatief de fluorescentie in het CSF en in het hersenvocht bepaald. Daarna zal
de expressie van de genen coderend voor TJ eiwitten onderzocht worden ter hoogte van deze
hersenbarrières.
Vervolgens zal de expressie van bovengenoemde genen samen met genen die coderen voor
verschillende inflammatoire mediatoren bestudeerd worden ter hoogte van het maagdarmstelsel.
Ten slotte zal ook de genexpressie van het vasoactief intestinale peptide (Vip) en diens receptoren
worden onderzocht, aangezien deze mogelijks bijdragen tot de communicatie tussen de hersenen en

28
het gastro-intestinaal stelsel. Vip reactieve neuronen zijn namelijk zowel in de maag als in de hersenen
veelvuldig aanwezig en bieden bescherming aan de intestinale barrière.

29
DEEL 3: MATERIAAL EN METHODEN 1. MUIZEN
Er werd gebruik gemaakt van 60 vrouwelijke C57BL/6 muizen, die op de leeftijd van 5 weken werden
aangeschaft via Harlan (Harlan Laboratoria, Nederland). De muizen werden gehuisvest in filtertop
kooien in groepen van 5 waarin ze onbeperkt vrije toegang hadden tot voer en water en blootgesteld
werden aan een 12 uur licht/12 uur donker cyclus. Alle experimenten werden vooraf goedgekeurd door
de ethische commissie van de faculteit Diergeneeskunde van de Universiteit Gent.
2. CULTUUR VAN DE H. PYLORI EN H. HEILMANNII BACTERIEN
De H. pylori bacteriën werden uitgeplaat op bloedagar. De H. heilmannii bacteriën groeiden op een
bifasisch 1.5% Brucella agar (Immunosource, België) aangevuld met 1 ml skirrow (Oxoid, Verenigd
koninkrijk), 3 ml vitox (Oxoid, verenigd Koninkrijk), 5 µg/ml amfotericine B (Sigma-Aldrich, Duitsland),
0.05% waterstofchloride (HCL; hydrochloric acid) (Sigma-Aldrich, Duitsland) en 20% geïnactiveerd
foetaal boviene serum (Perbio, Thermo Scientific Hyclone, Verenigd koninkrijk) met daarbovenop
Brucella bouillon (Immunosource, België) aangevuld met 0.05% HCL (Sigma-Aldrich, Duitsland). Zowel
de platen met H. pylori als met H. heilmannii werden geïncubeerd bij 37°C onder micro-aërobe
omstandigheden (10% CO2, 5% O2 en 85% N2).
3. IN VIVO INFECTIEPROCEDURE
De dag voor inoculatie werden de H. pylori bacteriën met een swab van de agarplaat gehaald en
gesuspendeerd in 23 ml Brucella bouillon (Immunosource, België), aangevuld met 58,5 µl skirrow
(Oxoid, Verenigd koninkrijk) en 1,25 ml geïnactiveerd foetaal boviene serum (Perbio, Thermo Scientific
Hyclone, Verenigd koninkrijk). Overnacht werden ze dan bij 37 °C onder micro-aërobe omstandigheden
op de schudder gezet.
Vlak voordat de muizen geïnoculeerd werden, werden de bacteriën geteld door middel van een telkamer
(Neubauwer Improved Assistant) en vervolgens verdund tot 108 bacteriën/ml.
Vervolgens werden de muizen op de leeftijd van 6 weken intragastraal geïnoculeerd. De inoculatie
gebeurde onder lichte anesthesie met behulp van 2,5% isofluraan. De 60 muizen werden ingedeeld in
6 verschillende groepen van 10 muizen per groep. De eerste groep was de controlegroep. Deze werden
geïnoculeerd met 300 µl van het groeimedium voor de bacteriën. De andere 5 groepen werden
geïnoculeerd met 300 µl van de suspensie met daarin 108 levende bacteriën/ml. De muizen werden
geïnoculeerd met verschillende wildtype Helicobacter species of mutanten waarbij een OMP-gen van
de Hof familie werd gemuteerd.
De tweede groep werd met H. heilmannii ASB1ΔHofE (mutant) bacteriën geïnoculeerd. De derde groep
werd met H. heilmannii ASB1 (wildtype) geïnoculeerd. De vierde groep werd met H. pylori SS1 (wildtype)
geïnoculeerd. De vijfde groep werd met H. pylori PMSS1 (Pre-mouse SS1; niet muis-geadapteerd
wildtype) geïnoculeerd. De zesde en laatste groep werd met H. pylori STM138ΔHofF geïnoculeerd
(mutant).

30
De muizen werden een maand na de inoculatie geëuthanaseerd.
4. WEEFSELCOLLECTIE
Om verschillende analyses uit te voeren, werden de muizen in 2 verschillende groepen van 30 muizen
per groep ingedeeld. De eerste groep werd gebruikt om de integriteit van de BCSFB en de BBB te
bepalen en voor immunohistochemische analyse van het maagdarmstelsel en de hersenen. Verder
werden hiervan ook stalen genomen van het gastro-intestinaal stelsel om genexpressie na te gaan. De
tweede groep muizen werd gebruikt voor immunohistochemische analyse, eiwitextractie, RNA- en DNA-
analyse van de maag en de hersenen.
Allereerst werden de muizen intraveneus, via de straatvene, geïnjecteerd met 4 kDa Fluorescein-
isothiocyanate-dextran (FITC) (Sigma). Vervolgens werden de muizen verdoofd door middel van een
intraperitoneale injectie met een ketamine/xylazine oplossing (Ceva/ Rompun Bayer), bestaande uit
20% ketamine (100 mg/ml) en 20% xylazine (23,32 mg/ml) opgelost in Dulbecco’s fosfaat gebufferde
zoutoplossing (Dulbecco's phosphate-buffered saline; dPBS) (Life Technologies) (zie bijlage I).
Als eerste werd dan CSF verkregen uit het vierde hersenventrikel door gebruik te maken van de cisterna
magna punctie methode (zie bijlage I). Daarna werd de eerste groep transcardiaal geperfuseerd met D-
PBS/heparine (0.2% heparine). De tweede groep werd ook transcardiaal geperfuseerd met D-
PBS/heparine (0.2% heparine), maar dan aangevuld met 0.5% broomfenolblauw (zie bijlage I). Het
hersenweefsel werd vervolgens geïsoleerd. Bij de tweede groep werd de CP van alle 4 de
hersenventrikels gecollecteerd (zie bijlage I) en bewaard in vloeibare stikstof. Bij groep 1 werden de
hersenen na isolatie in twee hersenhelften verdeeld. De rechter hersenhelft werd in kleine stukjes
geknipt en overnacht geïncubeerd in formaldehyde bij 37°C en op een schudder gezet. Het supernatans
(100 microliter onverdund) werd de volgende dag gecollecteerd, nadat het 15 minuten gecentrifugeerd
werd op maximale snelheid. Hiervan werd dan de fluorescentie gemeten wat een maat is voor de
lekkage van de BBB. De linker hemisfeer werd gefixeerd in 4% PFA en ingebed in paraffine om er
vervolgens paraffine coupes van te maken voor immunohistochemische kleuring. Bij beide groepen
werd voor de analyse van RNA en eiwitten de maag gecollecteerd en geopend ter hoogte van de
curvatura major. Vervolgens werd de maag gespoeld met steriele Hanks gebufferde zoutoplossing
(Hanks balanced salt solution; HBSS) (Gibco by Thermo Fisher Scientific, Verenigde Staten). De stalen
van het antrum, corpus, duodenum en colon werden gecollecteerd (zie bijlage I) en opgeslagen in RNA
later (Qiagen) bij -70°C voor RNA extractie of bevroren in vloeibare stikstof voor eiwitbepaling.
Bij de eerste groep werd bijkomend een overlangse doorsnede genomen van de maag, die overnacht
gefixeerd werd in formol voor immunohistochemische analyses.
De maag van groep 1 werd ten slotte ook nog gebruikt voor de bepaling van het microbioom. Hiervoor
isoleerde men het antrum, corpus, duodenum, ileum en colon, zonder vooraf de maag te spoelen.
Onmiddellijk na de staalname werd het CSF 100 keer verdund in steriel D-PBS. De lekkage van de
BCSF-barrière werd bepaald door de fluorescentie te meten bij λex/λem = 488/520 nm.

31
5. RNA-EXTRACTIE EN RT-QPCR VOOR GENEXPRESSIE
Er werd RNA geïsoleerd uit de CP met behulp van de Tissuelyser (Qiagen) en de Aurum Total RNA
Mini kit (Bio-Rad). Om de kwaliteit en de concentratie van het RNA na te gaan werd gebruikt gemaakt
van de NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific). Indien nodig werd het RNA
opgezuiverd via ethanol precipitatie.
De stalen van de maag, het duodenum en het colon die opgeslagen waren in RNA later werden
gehomogeniseerd door gebruik te maken van de Tissuelyser (Qiagen). Het RNA en DNA werd
gescheiden waarna het DNA terug ingevroren werd bij -80°C, zodat het vervolgens nog gebruikt kon
worden voor verdere DNA isolatie. De RNA-extractie van het antrum en het corpus van de maag, het
duodenum en het colon werd uitgevoerd met behulp van de RNeasy mini kit (Qiagen). De RNA-extractie
werd uitgevoerd volgens de fabrieksinstructies. Vervolgens werd het RNA net zoals voor de CP gemeten
via de NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific). Ook hier werd indien nodig het
RNA opgezuiverd via ethanol precipitatie.
Voor de bereiding van het cDNA van de CP werd respectievelijk 125 ng/µL startmateriaal gebruikt, wat
nadien 2.5 keer verdund werd met nuclease-vrij water (Ambion). De cDNA synthese gebeurde met
behulp van de SensiFAST™ cDNA Synthesis Kit (Bioline).
Voor de bereiding van het cDNA van het antrum en het corpus van de maag, het duodenum en het
colon werd 500 of 1000 ng/µL startmateriaal gebruikt, wat nadien 10 keer verdund werd met nuclease-
vrij water (Ambion). Het cDNA werd vervolgens gesynthetiseerd door middel van een cDNA Synthese
Kit (Bio-Rad). De RT-qPCR voor analyse van alle stalen werd uitgevoerd op een CFX384TM RT-qPCR
System met een C1000 Thermal Cycler (Roche VIB, Hercules), gebruikmakend van de Sensifast SYBR
reactiemix (Bioline). Expressie levels werden genormaliseerd aan de hand van de expressie van 3
referentie genen: Hprt, H2afz en Ppia voor de stalen van de maag, het duodenum en het colon. Voor
de stalen van de CP werden Gapdh, Rpl en Ubc gebruikt.
6. DNA-EXTRACTIE EN KWANTIFICATIE VAN H. PYLORI EN H. HEILMANNII IN HET GASTRO-
INTESTINAAL STELSEL
Het DNA werd geïsoleerd uit de heringevroren fractie (zie paragraaf 5). Vervolgens werd het aantal
aanwezige H. pylori en H. heilmannii bacteriën per milligram weefsel bepaald in de verkregen DNA
stalen door middel van Real-Time (RT)-kwantitatieve PCR. Om vooraf de kwaliteit en concentratie van
het DNA na te gaan werd gebruikt gemaakt van de NanoDrop 1000 Spectrophotometer (Thermo Fisher
Scientific). RT-qPCR detectie van H. pylori en H. heilmannii DNA werd gedaan door middel van ureA-
gebaseerde primers en de standaard werd gemaakt zoals reeds vroeger beschreven werd (O’rourke et
al., 2004). Vervolgens werd de RT-qPCR voor analyse van de standaard en alle stalen in duplicaat
uitgevoerd op een CFX384TM RT-qPCR systeem met een C1000 Thermal Cycler (Bio-Rad, Temse,
Belgium) volgens fabrieksinstructies.

32
7. HISTOPATHOLOGIE EN IMMUNOHISTOCHEMIE
Om de inflammatie van de maag en het duodenum te beoordelen werden er paraffine coupes van 5 µm
gesneden, gebruik makend van een microtoom (Microm, Prosan, Thermo-Fisher). De snedes werden
aangekleurd met Hematoxylin Gill III Prosan (Merck) en Eosine Yellow (VWR) volgens standaard
protocollen. De HE-kleuringen van de maag werden aangewend om de uitgebreidheid van de totale
gastritis te bepalen, waarbij men gebruik maakte van de gemodificeerde visuele analoge schaal gelijk
aan het aangepaste Updated Sydney System (Flahou et al., 2010). Maagmucosa werd gevisualiseerd
door gebruik te maken van een lichtmicroscoop (Leica DM2000).
Voor de MUC13 kleuring van de maag, werden er met behulp van een microtoom (Microm, Prosan,
Thermo-Fisher) 5 µm dikke paraffine sneden gemaakt. Vervolgens werden de coupes ontdaan van de
paraffine door achtereenvolgens xylene voor 10 minuten, isopropyl alcohol voor 5 minuten, 95% ethanol
voor 5 minuten, 50% ethanol voor 5 minuten en gedestilleerd water voor 5 minuten toe te voegen. Het
antigen werd teruggekregen door een citraatbuffer (10 mM, pH6) toe te voegen, gebruik makend van
een snelkookpan, waarin de stalen werden gespoeld met een was buffer (Dako). De coupes werden
vervolgens gedurende 5 minuten ingevroren met peroxidase blocking reagent (Dako) en daarna
gespoeld met gedestilleerd water en was buffer (Dako). Daarna werden ze gedurende 30 minuten
geïncubeerd met het primaire in-house polyklonale muis MUC13 antilichaam (1/1000, Dako, S3022).
Na het spoelen met de was buffer (Dako), werden de stalen op kamertemperatuur voor 30 minuten
geïncubeerd met een geëtiketteerde polymeer-HRP anti-muis (Evision+ system-HRP (DAB) (K4007)).
Vervolgens werd er 2 keer gespoeld met de was buffer (Dako), waarna DAB + substraat (Dako) en DAB
+ chromogen (Dako) werden toegevoegd gedurende 5 minuten. Tenslotte werd er een tegenkleuring
gedaan met hematoxylline gedurende 10 minuten, gevolgd door een 1 minuut durende spoelstap met
behulp van stromend kraanwater en een dehydratatie stap. De maagmucosa werd vervolgens
gevisualiseerd door gebruik te maken van een lichtmicroscoop (Leica DM2000).
Voor de IBA1 en GFAP immunohistochemische kleuring werden 5 µM dikke paraffine coupes van de
hersenen gemaakt. De secties werden gekleurd gebruik makend van het anti-IBA1 antilichaam (1/1000,
konijn, Wako, 019-19741) en het anti-GFAP antilichaam (1/200.000, konijn, Dako) gelijkaardig aan de
hierboven beschreven methode. De snedes gekleurd voor IBA1 en GFAP werden gescand met behulp
van de Actioscanner en de verkregen afbeeldingen werden verwerkt en geconverteerd naar JPEG met
behulp van ZEN 2 (blue edition, Carl Zeiss). De hoeveelheid bruine kleur wordt gekwantificeerd met
behulp van een script in Fiji Just ImagJ dat de oppervlakte met bruine kleur berekent ten opzichte van
de volledige weefseloppervlakte, hierbij worden gaten in het weefsel buiten beschouwing gelaten.
8. STATISTIEK Data werden geanalyseerd door middel van de Mann-Whitney test gebruik makend van GraphPad
Prism en worden gepresenteerd als gemiddelden standard error of mean (SEM), tenzij anders
bepaald. Significantie levels zijn aangeduid op de figuren: * indien 0.01 ≤ P < 0.05; ** indien 0.001 ≤ P
< 0.01; ***indien 0.0001 ≤ P < 0.001; **** indien P < 0.0001.
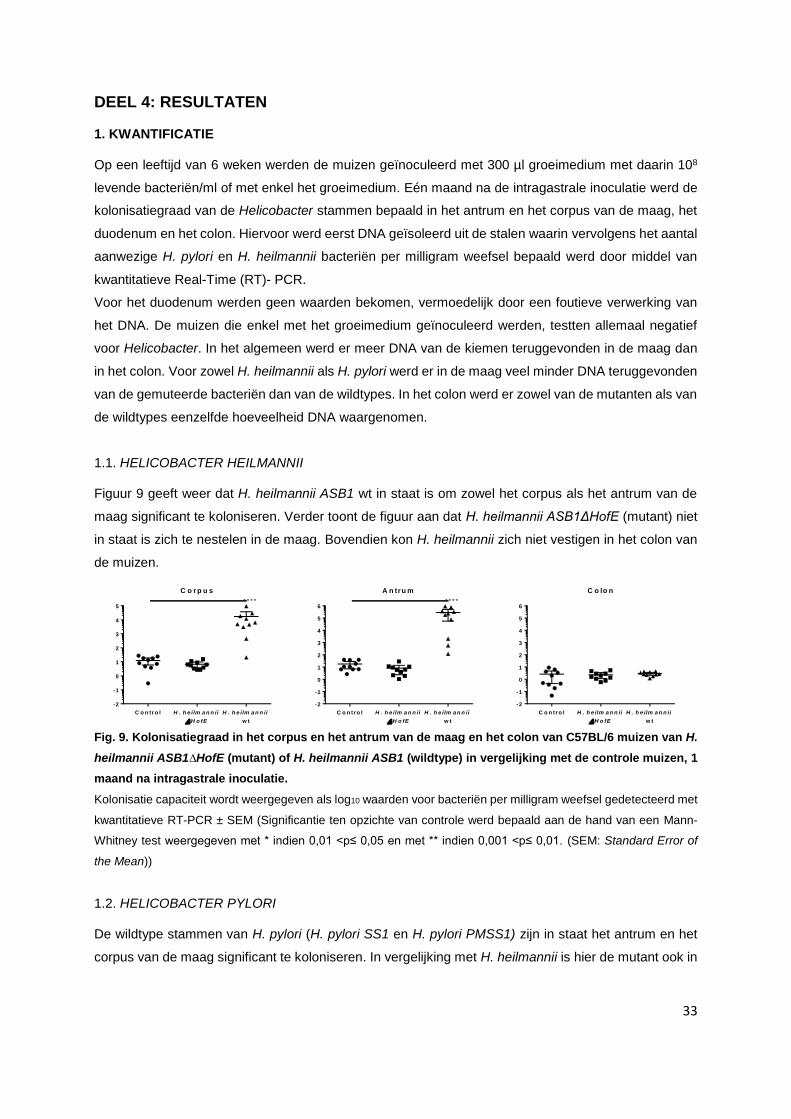
33
DEEL 4: RESULTATEN 1. KWANTIFICATIE Op een leeftijd van 6 weken werden de muizen geïnoculeerd met 300 µl groeimedium met daarin 108
levende bacteriën/ml of met enkel het groeimedium. Eén maand na de intragastrale inoculatie werd de
kolonisatiegraad van de Helicobacter stammen bepaald in het antrum en het corpus van de maag, het
duodenum en het colon. Hiervoor werd eerst DNA geïsoleerd uit de stalen waarin vervolgens het aantal
aanwezige H. pylori en H. heilmannii bacteriën per milligram weefsel bepaald werd door middel van
kwantitatieve Real-Time (RT)- PCR.
Voor het duodenum werden geen waarden bekomen, vermoedelijk door een foutieve verwerking van
het DNA. De muizen die enkel met het groeimedium geïnoculeerd werden, testten allemaal negatief
voor Helicobacter. In het algemeen werd er meer DNA van de kiemen teruggevonden in de maag dan
in het colon. Voor zowel H. heilmannii als H. pylori werd er in de maag veel minder DNA teruggevonden
van de gemuteerde bacteriën dan van de wildtypes. In het colon werd er zowel van de mutanten als van
de wildtypes eenzelfde hoeveelheid DNA waargenomen.
1.1. HELICOBACTER HEILMANNII Figuur 9 geeft weer dat H. heilmannii ASB1 wt in staat is om zowel het corpus als het antrum van de
maag significant te koloniseren. Verder toont de figuur aan dat H. heilmannii ASB1ΔHofE (mutant) niet
in staat is zich te nestelen in de maag. Bovendien kon H. heilmannii zich niet vestigen in het colon van
de muizen.
Fig. 9. Kolonisatiegraad in het corpus en het antrum van de maag en het colon van C57BL/6 muizen van H.
heilmannii ASB1∆HofE (mutant) of H. heilmannii ASB1 (wildtype) in vergelijking met de controle muizen, 1
maand na intragastrale inoculatie.
Kolonisatie capaciteit wordt weergegeven als log10 waarden voor bacteriën per milligram weefsel gedetecteerd met
kwantitatieve RT-PCR ± SEM (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-
Whitney test weergegeven met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of
the Mean))
1.2. HELICOBACTER PYLORI De wildtype stammen van H. pylori (H. pylori SS1 en H. pylori PMSS1) zijn in staat het antrum en het
corpus van de maag significant te koloniseren. In vergelijking met H. heilmannii is hier de mutant ook in
C o n tr o l H . h e ilm a n n ii
H o fE
H . h e ilm a n n ii
w t
-2
-1
0
1
2
3
4
5
C o r p u s
* * * *
C o n tr o l H . h e ilm a n n ii
H o fE
H . h e ilm a n n ii
w t
-2
-1
0
1
2
3
4
5
6
A n tru m
* * * *
C o n tr o l H . h e ilm a n n ii
H o fE
H . h e ilm a n n ii
w t
-2
-1
0
1
2
3
4
5
6
C o lo n
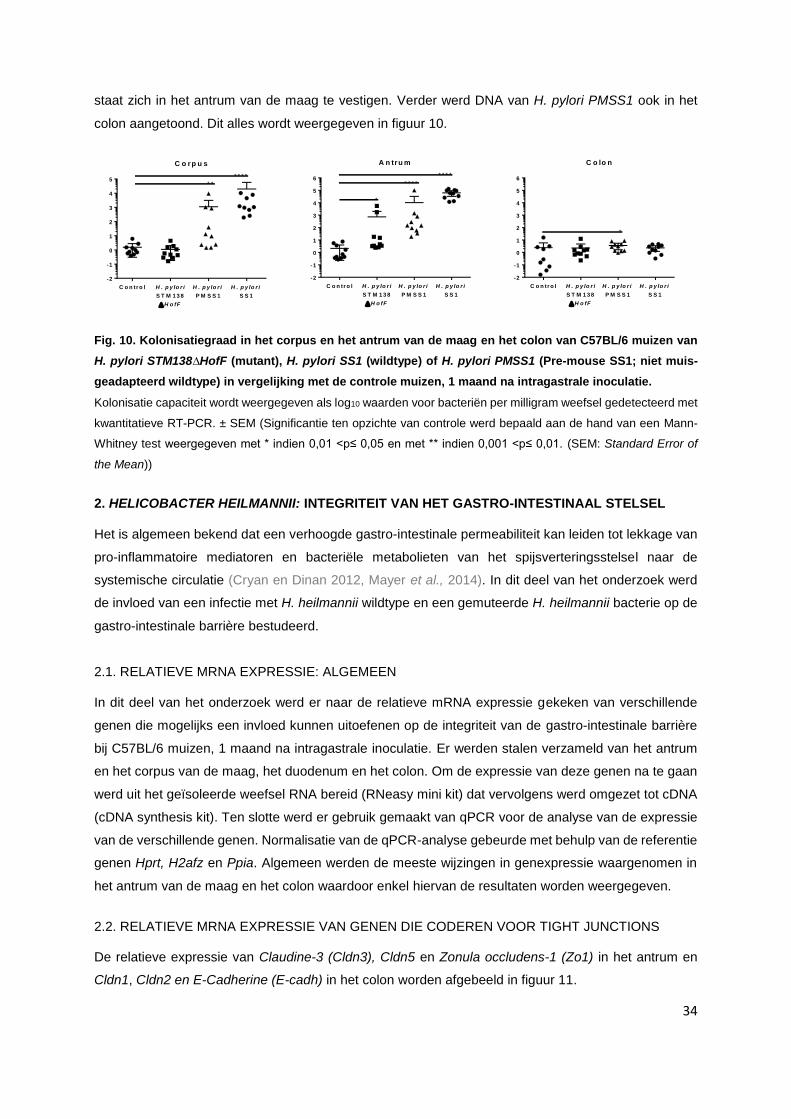
34
staat zich in het antrum van de maag te vestigen. Verder werd DNA van H. pylori PMSS1 ook in het
colon aangetoond. Dit alles wordt weergegeven in figuur 10.
Fig. 10. Kolonisatiegraad in het corpus en het antrum van de maag en het colon van C57BL/6 muizen van
H. pylori STM138∆HofF (mutant), H. pylori SS1 (wildtype) of H. pylori PMSS1 (Pre-mouse SS1; niet muis-
geadapteerd wildtype) in vergelijking met de controle muizen, 1 maand na intragastrale inoculatie.
Kolonisatie capaciteit wordt weergegeven als log10 waarden voor bacteriën per milligram weefsel gedetecteerd met
kwantitatieve RT-PCR. ± SEM (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-
Whitney test weergegeven met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of
the Mean))
2. HELICOBACTER HEILMANNII: INTEGRITEIT VAN HET GASTRO-INTESTINAAL STELSEL Het is algemeen bekend dat een verhoogde gastro-intestinale permeabiliteit kan leiden tot lekkage van
pro-inflammatoire mediatoren en bacteriële metabolieten van het spijsverteringsstelsel naar de
systemische circulatie (Cryan en Dinan 2012, Mayer et al., 2014). In dit deel van het onderzoek werd
de invloed van een infectie met H. heilmannii wildtype en een gemuteerde H. heilmannii bacterie op de
gastro-intestinale barrière bestudeerd.
2.1. RELATIEVE MRNA EXPRESSIE: ALGEMEEN In dit deel van het onderzoek werd er naar de relatieve mRNA expressie gekeken van verschillende
genen die mogelijks een invloed kunnen uitoefenen op de integriteit van de gastro-intestinale barrière
bij C57BL/6 muizen, 1 maand na intragastrale inoculatie. Er werden stalen verzameld van het antrum
en het corpus van de maag, het duodenum en het colon. Om de expressie van deze genen na te gaan
werd uit het geïsoleerde weefsel RNA bereid (RNeasy mini kit) dat vervolgens werd omgezet tot cDNA
(cDNA synthesis kit). Ten slotte werd er gebruik gemaakt van qPCR voor de analyse van de expressie
van de verschillende genen. Normalisatie van de qPCR-analyse gebeurde met behulp van de referentie
genen Hprt, H2afz en Ppia. Algemeen werden de meeste wijzingen in genexpressie waargenomen in
het antrum van de maag en het colon waardoor enkel hiervan de resultaten worden weergegeven.
2.2. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR TIGHT JUNCTIONS De relatieve expressie van Claudine-3 (Cldn3), Cldn5 en Zonula occludens-1 (Zo1) in het antrum en
Cldn1, Cldn2 en E-Cadherine (E-cadh) in het colon worden afgebeeld in figuur 11.
C o n tr o l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
-2
-1
0
1
2
3
4
5
C o r p u s
* *
* * * *
C o n tr o l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
-2
-1
0
1
2
3
4
5
6
A n tru m
* * * *
* * * *
*
C o n tr o l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
-2
-1
0
1
2
3
4
5
6
C o lo n
*
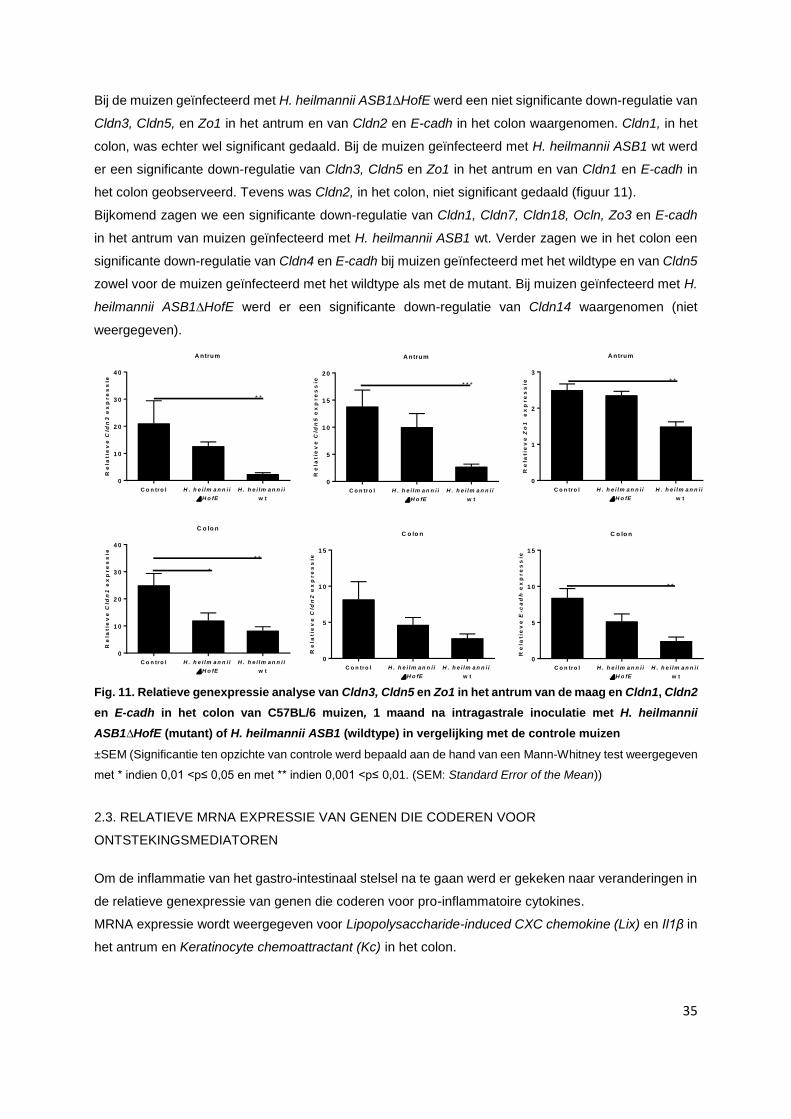
35
Bij de muizen geïnfecteerd met H. heilmannii ASB1∆HofE werd een niet significante down-regulatie van
Cldn3, Cldn5, en Zo1 in het antrum en van Cldn2 en E-cadh in het colon waargenomen. Cldn1, in het
colon, was echter wel significant gedaald. Bij de muizen geïnfecteerd met H. heilmannii ASB1 wt werd
er een significante down-regulatie van Cldn3, Cldn5 en Zo1 in het antrum en van Cldn1 en E-cadh in
het colon geobserveerd. Tevens was Cldn2, in het colon, niet significant gedaald (figuur 11).
Bijkomend zagen we een significante down-regulatie van Cldn1, Cldn7, Cldn18, Ocln, Zo3 en E-cadh
in het antrum van muizen geïnfecteerd met H. heilmannii ASB1 wt. Verder zagen we in het colon een
significante down-regulatie van Cldn4 en E-cadh bij muizen geïnfecteerd met het wildtype en van Cldn5
zowel voor de muizen geïnfecteerd met het wildtype als met de mutant. Bij muizen geïnfecteerd met H.
heilmannii ASB1∆HofE werd er een significante down-regulatie van Cldn14 waargenomen (niet
weergegeven).
Fig. 11. Relatieve genexpressie analyse van Cldn3, Cldn5 en Zo1 in het antrum van de maag en Cldn1, Cldn2
en E-cadh in het colon van C57BL/6 muizen, 1 maand na intragastrale inoculatie met H. heilmannii
ASB1∆HofE (mutant) of H. heilmannii ASB1 (wildtype) in vergelijking met de controle muizen
±SEM (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitney test weergegeven
met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of the Mean))
2.3. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR
ONTSTEKINGSMEDIATOREN
Om de inflammatie van het gastro-intestinaal stelsel na te gaan werd er gekeken naar veranderingen in
de relatieve genexpressie van genen die coderen voor pro-inflammatoire cytokines.
MRNA expressie wordt weergegeven voor Lipopolysaccharide-induced CXC chemokine (Lix) en Il1β in
het antrum en Keratinocyte chemoattractant (Kc) in het colon.
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
1 0
2 0
3 0
4 0
A ntrum
Re
latie
ve
Cld
n3
ex
pr
es
sie
* *
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
5
1 0
1 5
2 0
A ntrum
Re
latie
ve
Cld
n5
ex
pr
es
sie
* * *
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
1
2
3
A ntrum
Re
latie
ve
Zo
1e
xp
re
ss
ie * *
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
1 0
2 0
3 0
4 0
C o lo n
Re
latie
ve
Cld
n1
ex
pr
es
sie
*
* *
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
5
1 0
1 5
C o lo n
Re
latie
ve
Cld
n2
ex
pr
es
sie
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
5
1 0
1 5
C o lo n
Re
latie
ve
E-c
ad
he
xp
re
ss
ie
* *
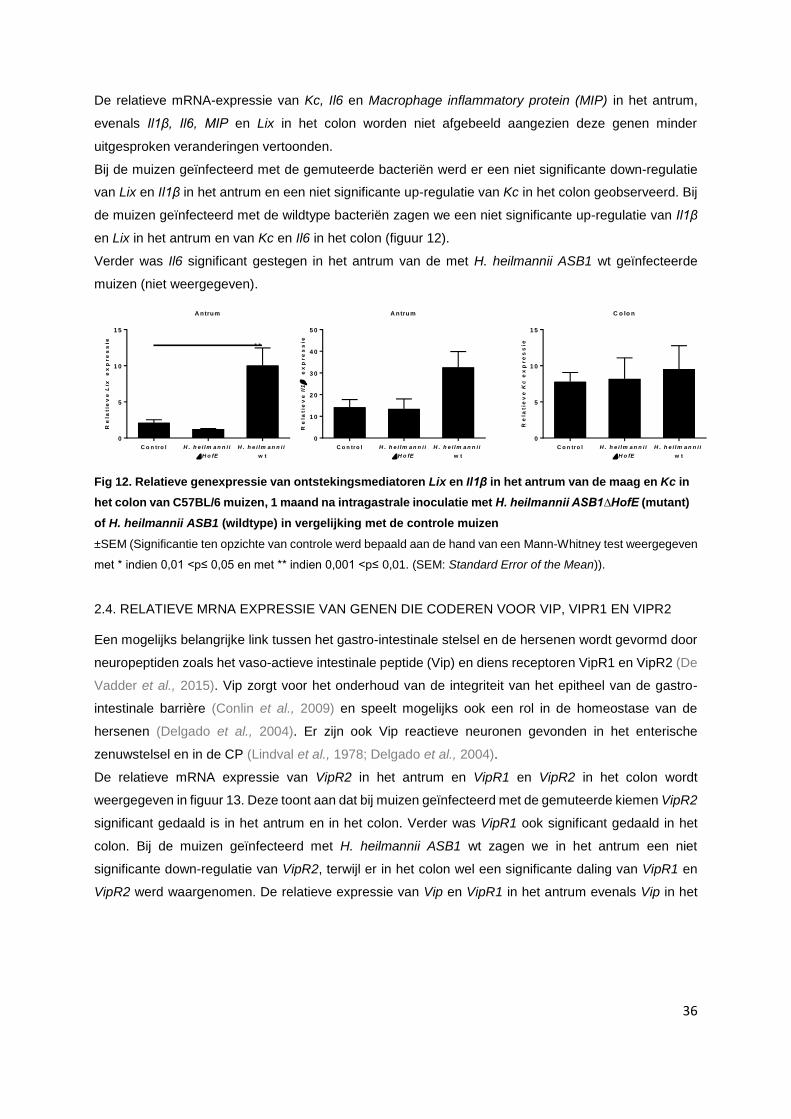
36
De relatieve mRNA-expressie van Kc, Il6 en Macrophage inflammatory protein (MIP) in het antrum,
evenals Il1β, Il6, MIP en Lix in het colon worden niet afgebeeld aangezien deze genen minder
uitgesproken veranderingen vertoonden.
Bij de muizen geïnfecteerd met de gemuteerde bacteriën werd er een niet significante down-regulatie
van Lix en Il1β in het antrum en een niet significante up-regulatie van Kc in het colon geobserveerd. Bij
de muizen geïnfecteerd met de wildtype bacteriën zagen we een niet significante up-regulatie van Il1β
en Lix in het antrum en van Kc en Il6 in het colon (figuur 12).
Verder was Il6 significant gestegen in het antrum van de met H. heilmannii ASB1 wt geïnfecteerde
muizen (niet weergegeven).
Fig 12. Relatieve genexpressie van ontstekingsmediatoren Lix en Il1β in het antrum van de maag en Kc in
het colon van C57BL/6 muizen, 1 maand na intragastrale inoculatie met H. heilmannii ASB1∆HofE (mutant)
of H. heilmannii ASB1 (wildtype) in vergelijking met de controle muizen
±SEM (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitney test weergegeven
met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of the Mean)).
2.4. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR VIP, VIPR1 EN VIPR2 Een mogelijks belangrijke link tussen het gastro-intestinale stelsel en de hersenen wordt gevormd door
neuropeptiden zoals het vaso-actieve intestinale peptide (Vip) en diens receptoren VipR1 en VipR2 (De
Vadder et al., 2015). Vip zorgt voor het onderhoud van de integriteit van het epitheel van de gastro-
intestinale barrière (Conlin et al., 2009) en speelt mogelijks ook een rol in de homeostase van de
hersenen (Delgado et al., 2004). Er zijn ook Vip reactieve neuronen gevonden in het enterische
zenuwstelsel en in de CP (Lindval et al., 1978; Delgado et al., 2004).
De relatieve mRNA expressie van VipR2 in het antrum en VipR1 en VipR2 in het colon wordt
weergegeven in figuur 13. Deze toont aan dat bij muizen geïnfecteerd met de gemuteerde kiemen VipR2
significant gedaald is in het antrum en in het colon. Verder was VipR1 ook significant gedaald in het
colon. Bij de muizen geïnfecteerd met H. heilmannii ASB1 wt zagen we in het antrum een niet
significante down-regulatie van VipR2, terwijl er in het colon wel een significante daling van VipR1 en
VipR2 werd waargenomen. De relatieve expressie van Vip en VipR1 in het antrum evenals Vip in het
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
5
1 0
1 5
A ntrum
Re
latie
ve
Lix
ex
pr
es
sie
* *
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
5
1 0
1 5
C o lo n
Re
latie
ve
Kc
ex
pr
es
sie
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
1 0
2 0
3 0
4 0
5 0
A ntrum
Re
latie
ve
Il1
ex
pr
es
sie
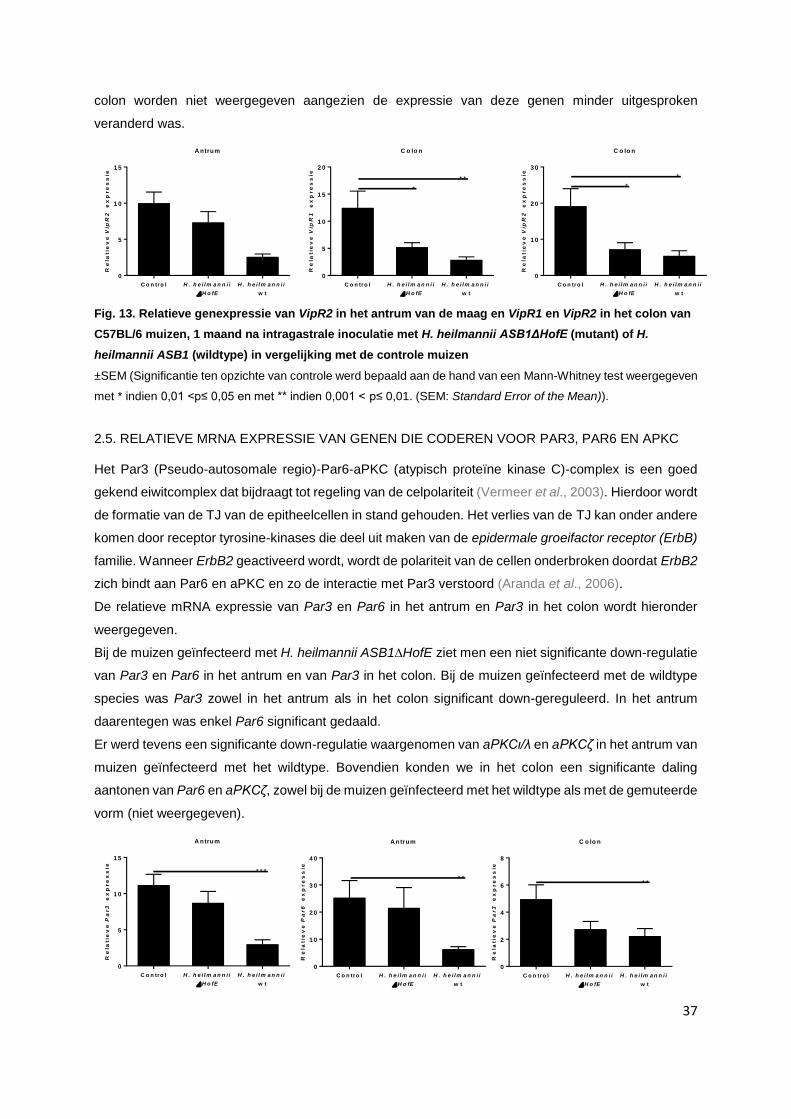
37
colon worden niet weergegeven aangezien de expressie van deze genen minder uitgesproken
veranderd was.
Fig. 13. Relatieve genexpressie van VipR2 in het antrum van de maag en VipR1 en VipR2 in het colon van
C57BL/6 muizen, 1 maand na intragastrale inoculatie met H. heilmannii ASB1ΔHofE (mutant) of H.
heilmannii ASB1 (wildtype) in vergelijking met de controle muizen
±SEM (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitney test weergegeven
met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 < p≤ 0,01. (SEM: Standard Error of the Mean)).
2.5. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR PAR3, PAR6 EN APKC Het Par3 (Pseudo-autosomale regio)-Par6-aPKC (atypisch proteïne kinase C)-complex is een goed
gekend eiwitcomplex dat bijdraagt tot regeling van de celpolariteit (Vermeer et al., 2003). Hierdoor wordt
de formatie van de TJ van de epitheelcellen in stand gehouden. Het verlies van de TJ kan onder andere
komen door receptor tyrosine-kinases die deel uit maken van de epidermale groeifactor receptor (ErbB)
familie. Wanneer ErbB2 geactiveerd wordt, wordt de polariteit van de cellen onderbroken doordat ErbB2
zich bindt aan Par6 en aPKC en zo de interactie met Par3 verstoord (Aranda et al., 2006).
De relatieve mRNA expressie van Par3 en Par6 in het antrum en Par3 in het colon wordt hieronder
weergegeven.
Bij de muizen geïnfecteerd met H. heilmannii ASB1∆HofE ziet men een niet significante down-regulatie
van Par3 en Par6 in het antrum en van Par3 in het colon. Bij de muizen geïnfecteerd met de wildtype
species was Par3 zowel in het antrum als in het colon significant down-gereguleerd. In het antrum
daarentegen was enkel Par6 significant gedaald.
Er werd tevens een significante down-regulatie waargenomen van aPKCι/λ en aPKCζ in het antrum van
muizen geïnfecteerd met het wildtype. Bovendien konden we in het colon een significante daling
aantonen van Par6 en aPKCζ, zowel bij de muizen geïnfecteerd met het wildtype als met de gemuteerde
vorm (niet weergegeven).
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
5
1 0
1 5
A ntrum
Re
latie
ve
Vip
R2
ex
pr
es
sie
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
5
1 0
1 5
2 0
C o lo n
Re
latie
ve
Vip
R1
ex
pr
es
sie
*
* *
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
1 0
2 0
3 0
C o lo n
Re
latie
ve
Vip
R2
ex
pr
es
sie
*
*
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
5
1 0
1 5
A ntrum
Re
latie
ve
Pa
r3
ex
pr
es
sie
* * *
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
1 0
2 0
3 0
4 0
A ntrum
Re
latie
ve
Pa
r6
ex
pr
es
sie
* *
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
2
4
6
8
C o lo n
Re
latie
ve
Pa
r3
ex
pr
es
sie
* *
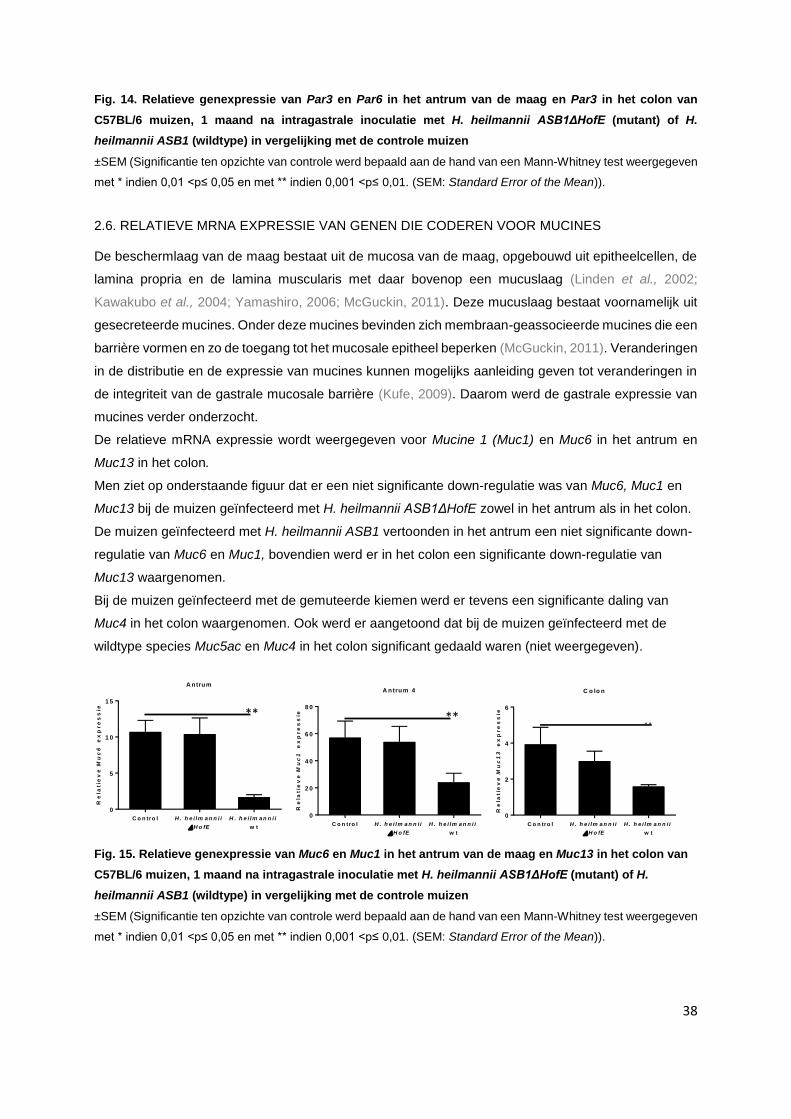
38
Fig. 14. Relatieve genexpressie van Par3 en Par6 in het antrum van de maag en Par3 in het colon van
C57BL/6 muizen, 1 maand na intragastrale inoculatie met H. heilmannii ASB1ΔHofE (mutant) of H.
heilmannii ASB1 (wildtype) in vergelijking met de controle muizen
±SEM (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitney test weergegeven
met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of the Mean)).
2.6. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR MUCINES De beschermlaag van de maag bestaat uit de mucosa van de maag, opgebouwd uit epitheelcellen, de
lamina propria en de lamina muscularis met daar bovenop een mucuslaag (Linden et al., 2002;
Kawakubo et al., 2004; Yamashiro, 2006; McGuckin, 2011). Deze mucuslaag bestaat voornamelijk uit
gesecreteerde mucines. Onder deze mucines bevinden zich membraan-geassocieerde mucines die een
barrière vormen en zo de toegang tot het mucosale epitheel beperken (McGuckin, 2011). Veranderingen
in de distributie en de expressie van mucines kunnen mogelijks aanleiding geven tot veranderingen in
de integriteit van de gastrale mucosale barrière (Kufe, 2009). Daarom werd de gastrale expressie van
mucines verder onderzocht.
De relatieve mRNA expressie wordt weergegeven voor Mucine 1 (Muc1) en Muc6 in het antrum en
Muc13 in het colon.
Men ziet op onderstaande figuur dat er een niet significante down-regulatie was van Muc6, Muc1 en
Muc13 bij de muizen geïnfecteerd met H. heilmannii ASB1ΔHofE zowel in het antrum als in het colon.
De muizen geïnfecteerd met H. heilmannii ASB1 vertoonden in het antrum een niet significante down-
regulatie van Muc6 en Muc1, bovendien werd er in het colon een significante down-regulatie van
Muc13 waargenomen.
Bij de muizen geïnfecteerd met de gemuteerde kiemen werd er tevens een significante daling van
Muc4 in het colon waargenomen. Ook werd er aangetoond dat bij de muizen geïnfecteerd met de
wildtype species Muc5ac en Muc4 in het colon significant gedaald waren (niet weergegeven).
Fig. 15. Relatieve genexpressie van Muc6 en Muc1 in het antrum van de maag en Muc13 in het colon van
C57BL/6 muizen, 1 maand na intragastrale inoculatie met H. heilmannii ASB1ΔHofE (mutant) of H.
heilmannii ASB1 (wildtype) in vergelijking met de controle muizen
±SEM (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitney test weergegeven
met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of the Mean)).
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
5
1 0
1 5
A ntrum
Re
latie
ve
Mu
c6
ex
pr
es
sie **
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
2 0
4 0
6 0
8 0
A ntrum 4
Re
latie
ve
Mu
c1
ex
pr
es
sie **
C o n tro l H . h e i lm a n n i i
H o fE
H . h e i lm a n n i i
w t
0
2
4
6
C o lo n
Re
latie
ve
Mu
c1
3e
xp
re
ss
ie
* *
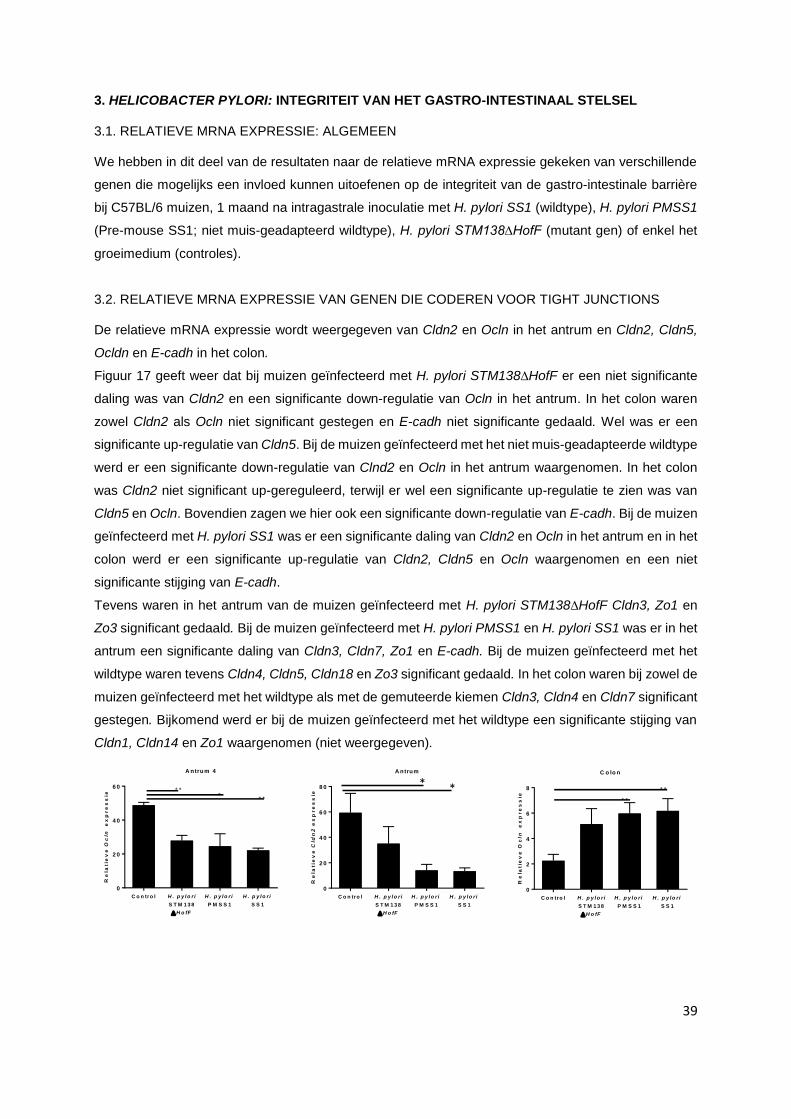
39
3. HELICOBACTER PYLORI: INTEGRITEIT VAN HET GASTRO-INTESTINAAL STELSEL 3.1. RELATIEVE MRNA EXPRESSIE: ALGEMEEN We hebben in dit deel van de resultaten naar de relatieve mRNA expressie gekeken van verschillende
genen die mogelijks een invloed kunnen uitoefenen op de integriteit van de gastro-intestinale barrière
bij C57BL/6 muizen, 1 maand na intragastrale inoculatie met H. pylori SS1 (wildtype), H. pylori PMSS1
(Pre-mouse SS1; niet muis-geadapteerd wildtype), H. pylori STM138∆HofF (mutant gen) of enkel het
groeimedium (controles).
3.2. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR TIGHT JUNCTIONS De relatieve mRNA expressie wordt weergegeven van Cldn2 en Ocln in het antrum en Cldn2, Cldn5,
Ocldn en E-cadh in het colon.
Figuur 17 geeft weer dat bij muizen geïnfecteerd met H. pylori STM138∆HofF er een niet significante
daling was van Cldn2 en een significante down-regulatie van Ocln in het antrum. In het colon waren
zowel Cldn2 als Ocln niet significant gestegen en E-cadh niet significante gedaald. Wel was er een
significante up-regulatie van Cldn5. Bij de muizen geïnfecteerd met het niet muis-geadapteerde wildtype
werd er een significante down-regulatie van Clnd2 en Ocln in het antrum waargenomen. In het colon
was Cldn2 niet significant up-gereguleerd, terwijl er wel een significante up-regulatie te zien was van
Cldn5 en Ocln. Bovendien zagen we hier ook een significante down-regulatie van E-cadh. Bij de muizen
geïnfecteerd met H. pylori SS1 was er een significante daling van Cldn2 en Ocln in het antrum en in het
colon werd er een significante up-regulatie van Cldn2, Cldn5 en Ocln waargenomen en een niet
significante stijging van E-cadh.
Tevens waren in het antrum van de muizen geïnfecteerd met H. pylori STM138∆HofF Cldn3, Zo1 en
Zo3 significant gedaald. Bij de muizen geïnfecteerd met H. pylori PMSS1 en H. pylori SS1 was er in het
antrum een significante daling van Cldn3, Cldn7, Zo1 en E-cadh. Bij de muizen geïnfecteerd met het
wildtype waren tevens Cldn4, Cldn5, Cldn18 en Zo3 significant gedaald. In het colon waren bij zowel de
muizen geïnfecteerd met het wildtype als met de gemuteerde kiemen Cldn3, Cldn4 en Cldn7 significant
gestegen. Bijkomend werd er bij de muizen geïnfecteerd met het wildtype een significante stijging van
Cldn1, Cldn14 en Zo1 waargenomen (niet weergegeven).
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
2 0
4 0
6 0
A ntrum 4
Re
latie
ve
Oc
lne
xp
re
ss
ie
* *
* **
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
2 0
4 0
6 0
8 0
A ntrum
Re
latie
ve
Cld
n2
ex
pr
es
sie
**
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
2
4
6
8
C o lo n
Re
latie
ve
Oc
lne
xp
re
ss
ie
* *
* *
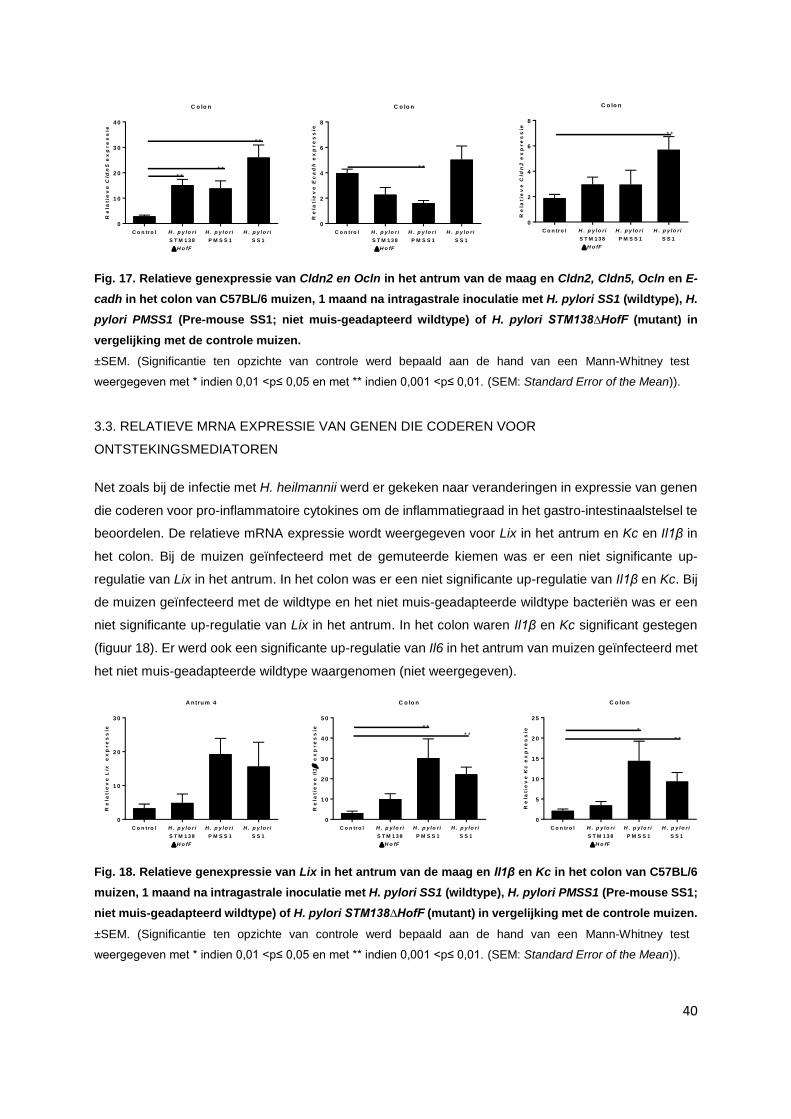
40
Fig. 17. Relatieve genexpressie van Cldn2 en Ocln in het antrum van de maag en Cldn2, Cldn5, Ocln en E-
cadh in het colon van C57BL/6 muizen, 1 maand na intragastrale inoculatie met H. pylori SS1 (wildtype), H.
pylori PMSS1 (Pre-mouse SS1; niet muis-geadapteerd wildtype) of H. pylori STM138∆HofF (mutant) in
vergelijking met de controle muizen.
±SEM. (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitney test
weergegeven met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of the Mean)).
3.3. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR
ONTSTEKINGSMEDIATOREN
Net zoals bij de infectie met H. heilmannii werd er gekeken naar veranderingen in expressie van genen
die coderen voor pro-inflammatoire cytokines om de inflammatiegraad in het gastro-intestinaalstelsel te
beoordelen. De relatieve mRNA expressie wordt weergegeven voor Lix in het antrum en Kc en Il1β in
het colon. Bij de muizen geïnfecteerd met de gemuteerde kiemen was er een niet significante up-
regulatie van Lix in het antrum. In het colon was er een niet significante up-regulatie van Il1β en Kc. Bij
de muizen geïnfecteerd met de wildtype en het niet muis-geadapteerde wildtype bacteriën was er een
niet significante up-regulatie van Lix in het antrum. In het colon waren Il1β en Kc significant gestegen
(figuur 18). Er werd ook een significante up-regulatie van Il6 in het antrum van muizen geïnfecteerd met
het niet muis-geadapteerde wildtype waargenomen (niet weergegeven).
Fig. 18. Relatieve genexpressie van Lix in het antrum van de maag en Il1β en Kc in het colon van C57BL/6
muizen, 1 maand na intragastrale inoculatie met H. pylori SS1 (wildtype), H. pylori PMSS1 (Pre-mouse SS1;
niet muis-geadapteerd wildtype) of H. pylori STM138∆HofF (mutant) in vergelijking met de controle muizen.
±SEM. (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitney test
weergegeven met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of the Mean)).
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
1 0
2 0
3 0
4 0
C o lo nR
ela
tie
ve
Cld
n5
ex
pr
es
sie
* *
* *
* *
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
2
4
6
8
C o lo n
Re
latie
ve
Ec
ad
h e
xp
re
ss
ie
* *
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
1 0
2 0
3 0
A ntrum 4
Re
latie
ve
Lix
ex
pr
es
sie
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
5
1 0
1 5
2 0
2 5
C o lo n
Re
latie
ve
Kc
ex
pr
es
sie
* *
*
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
2
4
6
8
C o lo n
Re
latie
ve
Cld
n2
ex
pr
es
sie
* *
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
1 0
2 0
3 0
4 0
5 0
C o lo n
Re
latie
ve
Il1
ex
pr
es
sie
* *
* *
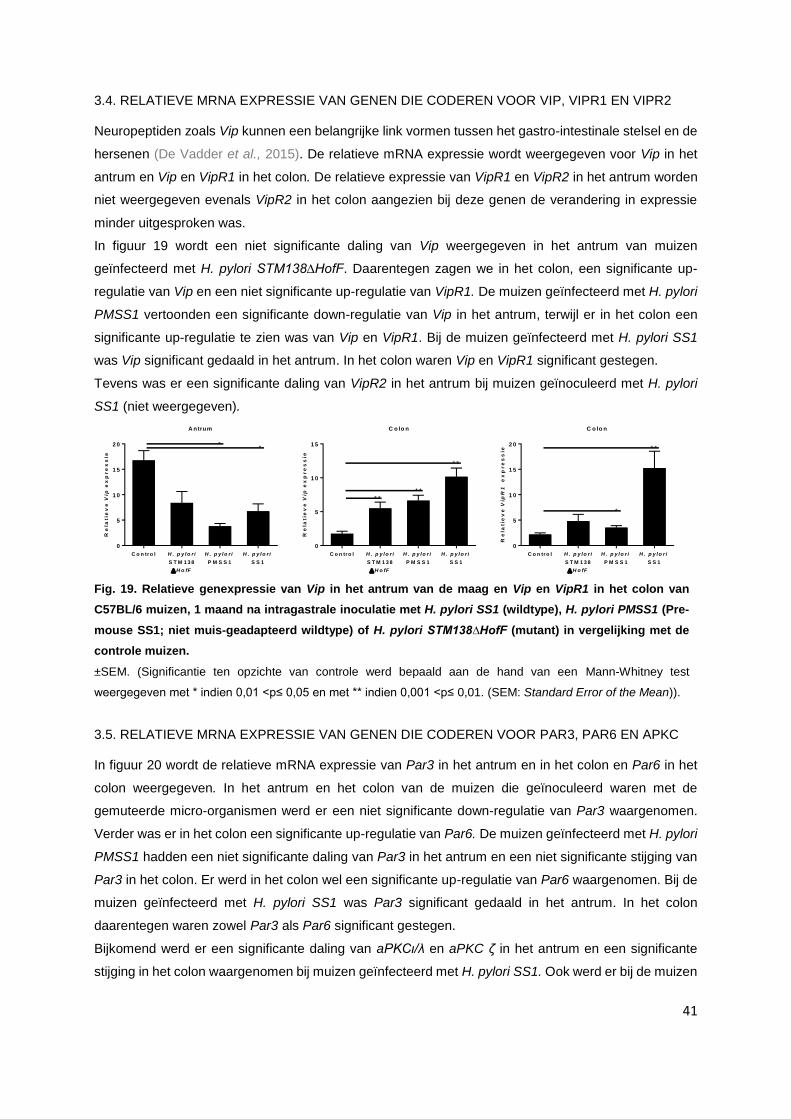
41
3.4. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR VIP, VIPR1 EN VIPR2 Neuropeptiden zoals Vip kunnen een belangrijke link vormen tussen het gastro-intestinale stelsel en de
hersenen (De Vadder et al., 2015). De relatieve mRNA expressie wordt weergegeven voor Vip in het
antrum en Vip en VipR1 in het colon. De relatieve expressie van VipR1 en VipR2 in het antrum worden
niet weergegeven evenals VipR2 in het colon aangezien bij deze genen de verandering in expressie
minder uitgesproken was.
In figuur 19 wordt een niet significante daling van Vip weergegeven in het antrum van muizen
geïnfecteerd met H. pylori STM138∆HofF. Daarentegen zagen we in het colon, een significante up-
regulatie van Vip en een niet significante up-regulatie van VipR1. De muizen geïnfecteerd met H. pylori
PMSS1 vertoonden een significante down-regulatie van Vip in het antrum, terwijl er in het colon een
significante up-regulatie te zien was van Vip en VipR1. Bij de muizen geïnfecteerd met H. pylori SS1
was Vip significant gedaald in het antrum. In het colon waren Vip en VipR1 significant gestegen.
Tevens was er een significante daling van VipR2 in het antrum bij muizen geïnoculeerd met H. pylori
SS1 (niet weergegeven).
Fig. 19. Relatieve genexpressie van Vip in het antrum van de maag en Vip en VipR1 in het colon van
C57BL/6 muizen, 1 maand na intragastrale inoculatie met H. pylori SS1 (wildtype), H. pylori PMSS1 (Pre-
mouse SS1; niet muis-geadapteerd wildtype) of H. pylori STM138∆HofF (mutant) in vergelijking met de
controle muizen.
±SEM. (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitney test
weergegeven met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of the Mean)).
3.5. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR PAR3, PAR6 EN APKC In figuur 20 wordt de relatieve mRNA expressie van Par3 in het antrum en in het colon en Par6 in het
colon weergegeven. In het antrum en het colon van de muizen die geïnoculeerd waren met de
gemuteerde micro-organismen werd er een niet significante down-regulatie van Par3 waargenomen.
Verder was er in het colon een significante up-regulatie van Par6. De muizen geïnfecteerd met H. pylori
PMSS1 hadden een niet significante daling van Par3 in het antrum en een niet significante stijging van
Par3 in het colon. Er werd in het colon wel een significante up-regulatie van Par6 waargenomen. Bij de
muizen geïnfecteerd met H. pylori SS1 was Par3 significant gedaald in het antrum. In het colon
daarentegen waren zowel Par3 als Par6 significant gestegen.
Bijkomend werd er een significante daling van aPKCι/λ en aPKC ζ in het antrum en een significante
stijging in het colon waargenomen bij muizen geïnfecteerd met H. pylori SS1. Ook werd er bij de muizen
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
5
1 0
1 5
2 0
A ntrum
Re
latie
ve
Vip
ex
pr
es
sie
**
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
5
1 0
1 5
C o lo n
Re
latie
ve
Vip
ex
pr
es
sie
* *
* *
* *
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
5
1 0
1 5
2 0
C o lo n
Re
latie
ve
Vip
R1
ex
pr
es
sie
*
* *
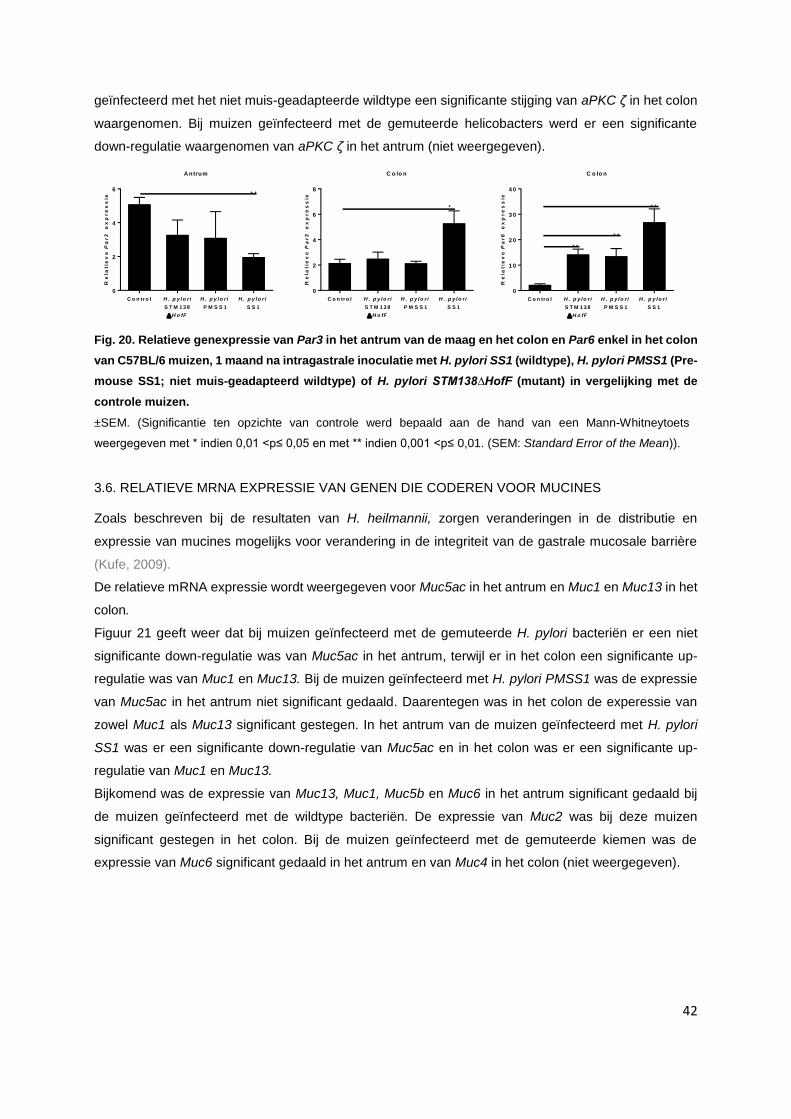
42
geïnfecteerd met het niet muis-geadapteerde wildtype een significante stijging van aPKC ζ in het colon
waargenomen. Bij muizen geïnfecteerd met de gemuteerde helicobacters werd er een significante
down-regulatie waargenomen van aPKC ζ in het antrum (niet weergegeven).
Fig. 20. Relatieve genexpressie van Par3 in het antrum van de maag en het colon en Par6 enkel in het colon
van C57BL/6 muizen, 1 maand na intragastrale inoculatie met H. pylori SS1 (wildtype), H. pylori PMSS1 (Pre-
mouse SS1; niet muis-geadapteerd wildtype) of H. pylori STM138∆HofF (mutant) in vergelijking met de
controle muizen.
±SEM. (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitneytoets
weergegeven met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of the Mean)).
3.6. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR MUCINES Zoals beschreven bij de resultaten van H. heilmannii, zorgen veranderingen in de distributie en
expressie van mucines mogelijks voor verandering in de integriteit van de gastrale mucosale barrière
(Kufe, 2009).
De relatieve mRNA expressie wordt weergegeven voor Muc5ac in het antrum en Muc1 en Muc13 in het
colon.
Figuur 21 geeft weer dat bij muizen geïnfecteerd met de gemuteerde H. pylori bacteriën er een niet
significante down-regulatie was van Muc5ac in het antrum, terwijl er in het colon een significante up-
regulatie was van Muc1 en Muc13. Bij de muizen geïnfecteerd met H. pylori PMSS1 was de expressie
van Muc5ac in het antrum niet significant gedaald. Daarentegen was in het colon de experessie van
zowel Muc1 als Muc13 significant gestegen. In het antrum van de muizen geïnfecteerd met H. pylori
SS1 was er een significante down-regulatie van Muc5ac en in het colon was er een significante up-
regulatie van Muc1 en Muc13.
Bijkomend was de expressie van Muc13, Muc1, Muc5b en Muc6 in het antrum significant gedaald bij
de muizen geïnfecteerd met de wildtype bacteriën. De expressie van Muc2 was bij deze muizen
significant gestegen in het colon. Bij de muizen geïnfecteerd met de gemuteerde kiemen was de
expressie van Muc6 significant gedaald in het antrum en van Muc4 in het colon (niet weergegeven).
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
2
4
6
A ntrum
Re
latie
ve
Pa
r3
ex
pr
es
sie
* *
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
2
4
6
8
C o lo n
Re
latie
ve
Pa
r3
ex
pr
es
sie
*
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
1 0
2 0
3 0
4 0
C o lo n
Re
latie
ve
Pa
r6
ex
pr
es
sie
* *
* *
* *
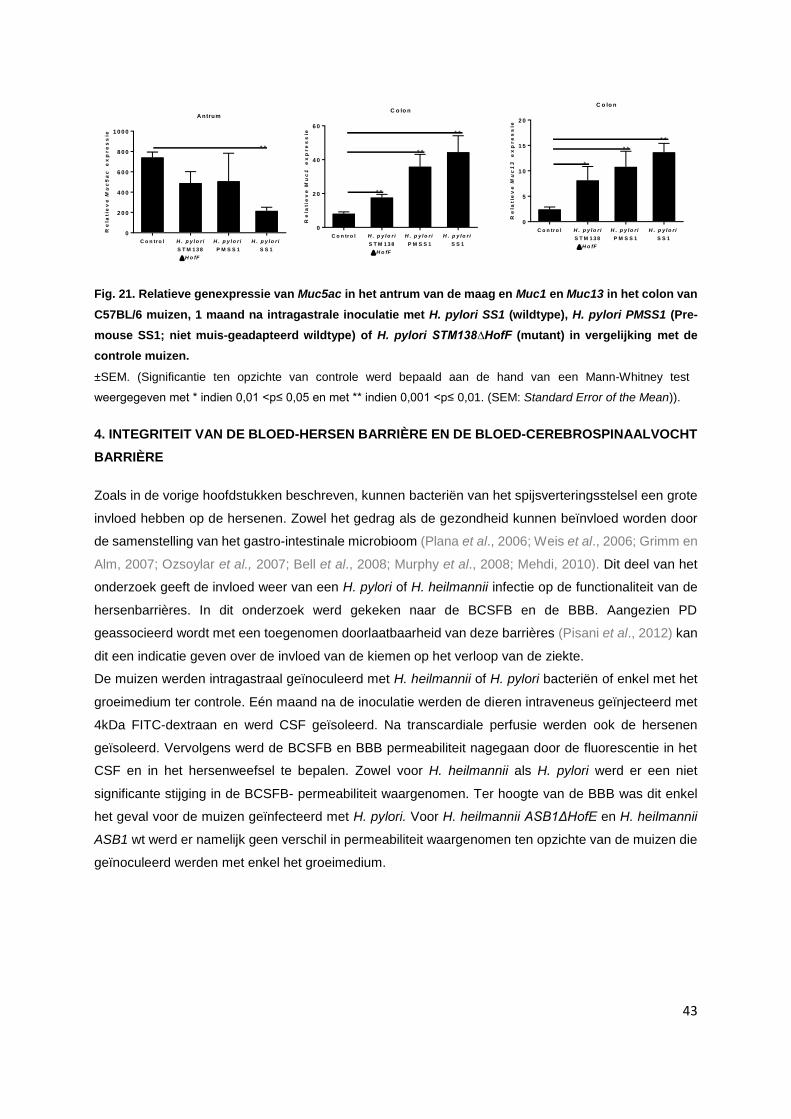
43
Fig. 21. Relatieve genexpressie van Muc5ac in het antrum van de maag en Muc1 en Muc13 in het colon van
C57BL/6 muizen, 1 maand na intragastrale inoculatie met H. pylori SS1 (wildtype), H. pylori PMSS1 (Pre-
mouse SS1; niet muis-geadapteerd wildtype) of H. pylori STM138∆HofF (mutant) in vergelijking met de
controle muizen.
±SEM. (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitney test
weergegeven met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of the Mean)).
4. INTEGRITEIT VAN DE BLOED-HERSEN BARRIÈRE EN DE BLOED-CEREBROSPINAALVOCHT
BARRIÈRE
Zoals in de vorige hoofdstukken beschreven, kunnen bacteriën van het spijsverteringsstelsel een grote
invloed hebben op de hersenen. Zowel het gedrag als de gezondheid kunnen beïnvloed worden door
de samenstelling van het gastro-intestinale microbioom (Plana et al., 2006; Weis et al., 2006; Grimm en
Alm, 2007; Ozsoylar et al., 2007; Bell et al., 2008; Murphy et al., 2008; Mehdi, 2010). Dit deel van het
onderzoek geeft de invloed weer van een H. pylori of H. heilmannii infectie op de functionaliteit van de
hersenbarrières. In dit onderzoek werd gekeken naar de BCSFB en de BBB. Aangezien PD
geassocieerd wordt met een toegenomen doorlaatbaarheid van deze barrières (Pisani et al., 2012) kan
dit een indicatie geven over de invloed van de kiemen op het verloop van de ziekte.
De muizen werden intragastraal geïnoculeerd met H. heilmannii of H. pylori bacteriën of enkel met het
groeimedium ter controle. Eén maand na de inoculatie werden de dieren intraveneus geïnjecteerd met
4kDa FITC-dextraan en werd CSF geïsoleerd. Na transcardiale perfusie werden ook de hersenen
geïsoleerd. Vervolgens werd de BCSFB en BBB permeabiliteit nagegaan door de fluorescentie in het
CSF en in het hersenweefsel te bepalen. Zowel voor H. heilmannii als H. pylori werd er een niet
significante stijging in de BCSFB- permeabiliteit waargenomen. Ter hoogte van de BBB was dit enkel
het geval voor de muizen geïnfecteerd met H. pylori. Voor H. heilmannii ASB1ΔHofE en H. heilmannii
ASB1 wt werd er namelijk geen verschil in permeabiliteit waargenomen ten opzichte van de muizen die
geïnoculeerd werden met enkel het groeimedium.
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
2 0 0
4 0 0
6 0 0
8 0 0
1 0 0 0
A ntrum
Re
latie
ve
Mu
c5
ac
ex
pr
es
sie
* *
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
2 0
4 0
6 0
C o lo n
Re
latie
ve
Mu
c1
ex
pr
es
sie
* *
* *
* *
C o n tro l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
5
1 0
1 5
2 0
C o lo n
Re
latie
ve
Mu
c1
3e
xp
re
ss
ie
*
* *
* *
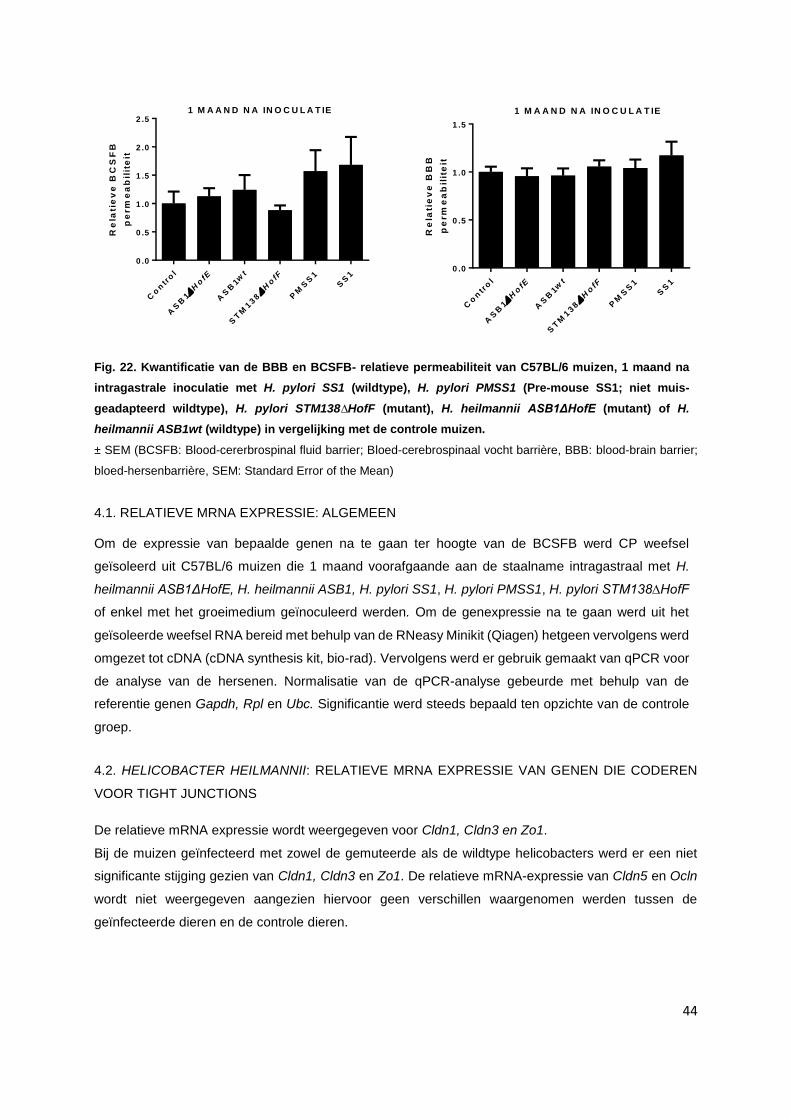
44
Fig. 22. Kwantificatie van de BBB en BCSFB- relatieve permeabiliteit van C57BL/6 muizen, 1 maand na
intragastrale inoculatie met H. pylori SS1 (wildtype), H. pylori PMSS1 (Pre-mouse SS1; niet muis-
geadapteerd wildtype), H. pylori STM138∆HofF (mutant), H. heilmannii ASB1ΔHofE (mutant) of H.
heilmannii ASB1wt (wildtype) in vergelijking met de controle muizen.
± SEM (BCSFB: Blood-cererbrospinal fluid barrier; Bloed-cerebrospinaal vocht barrière, BBB: blood-brain barrier;
bloed-hersenbarrière, SEM: Standard Error of the Mean)
4.1. RELATIEVE MRNA EXPRESSIE: ALGEMEEN Om de expressie van bepaalde genen na te gaan ter hoogte van de BCSFB werd CP weefsel
geïsoleerd uit C57BL/6 muizen die 1 maand voorafgaande aan de staalname intragastraal met H.
heilmannii ASB1ΔHofE, H. heilmannii ASB1, H. pylori SS1, H. pylori PMSS1, H. pylori STM138∆HofF
of enkel met het groeimedium geïnoculeerd werden. Om de genexpressie na te gaan werd uit het
geïsoleerde weefsel RNA bereid met behulp van de RNeasy Minikit (Qiagen) hetgeen vervolgens werd
omgezet tot cDNA (cDNA synthesis kit, bio-rad). Vervolgens werd er gebruik gemaakt van qPCR voor
de analyse van de hersenen. Normalisatie van de qPCR-analyse gebeurde met behulp van de
referentie genen Gapdh, Rpl en Ubc. Significantie werd steeds bepaald ten opzichte van de controle
groep.
4.2. HELICOBACTER HEILMANNII: RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN
VOOR TIGHT JUNCTIONS
De relatieve mRNA expressie wordt weergegeven voor Cldn1, Cldn3 en Zo1.
Bij de muizen geïnfecteerd met zowel de gemuteerde als de wildtype helicobacters werd er een niet
significante stijging gezien van Cldn1, Cldn3 en Zo1. De relatieve mRNA-expressie van Cldn5 en Ocln
wordt niet weergegeven aangezien hiervoor geen verschillen waargenomen werden tussen de
geïnfecteerde dieren en de controle dieren.
Co
ntr
ol
AS
B1
Ho
fE
AS
B1w
t
ST
M138
Ho
fF
PM
SS
1
SS
1
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5R
ela
tie
ve
BC
SF
B
pe
rm
ea
bil
ite
it
1 M A A N D N A IN O C U L A T IE
Co
ntr
ol
AS
B1
Ho
fE
AS
B1w
t
ST
M138
Ho
fF
PM
SS
1
SS
1
0 .0
0 .5
1 .0
1 .5
Re
lati
ev
e B
BB
pe
rm
ea
bil
ite
it
1 M A A N D N A IN O C U L A T IE
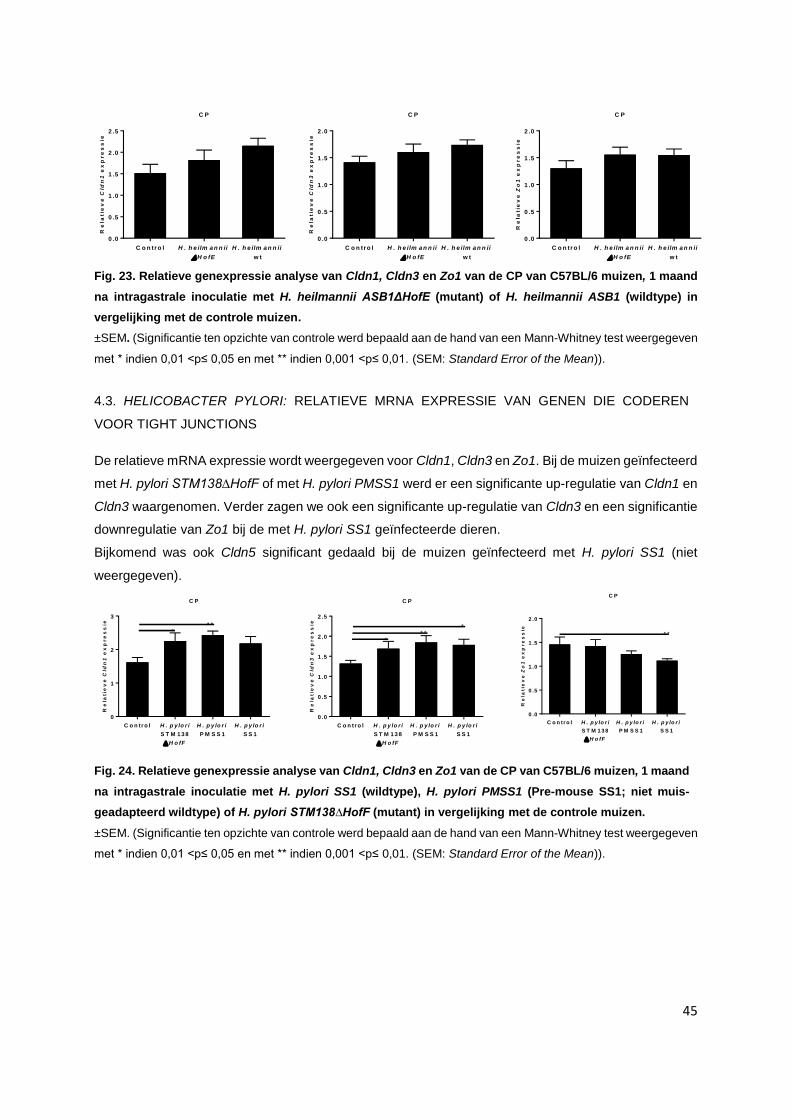
45
Fig. 23. Relatieve genexpressie analyse van Cldn1, Cldn3 en Zo1 van de CP van C57BL/6 muizen, 1 maand
na intragastrale inoculatie met H. heilmannii ASB1ΔHofE (mutant) of H. heilmannii ASB1 (wildtype) in
vergelijking met de controle muizen.
±SEM. (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitney test weergegeven
met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of the Mean)).
4.3. HELICOBACTER PYLORI: RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN
VOOR TIGHT JUNCTIONS
De relatieve mRNA expressie wordt weergegeven voor Cldn1, Cldn3 en Zo1. Bij de muizen geïnfecteerd
met H. pylori STM138∆HofF of met H. pylori PMSS1 werd er een significante up-regulatie van Cldn1 en
Cldn3 waargenomen. Verder zagen we ook een significante up-regulatie van Cldn3 en een significantie
downregulatie van Zo1 bij de met H. pylori SS1 geïnfecteerde dieren.
Bijkomend was ook Cldn5 significant gedaald bij de muizen geïnfecteerd met H. pylori SS1 (niet
weergegeven).
Fig. 24. Relatieve genexpressie analyse van Cldn1, Cldn3 en Zo1 van de CP van C57BL/6 muizen, 1 maand
na intragastrale inoculatie met H. pylori SS1 (wildtype), H. pylori PMSS1 (Pre-mouse SS1; niet muis-
geadapteerd wildtype) of H. pylori STM138∆HofF (mutant) in vergelijking met de controle muizen.
±SEM. (Significantie ten opzichte van controle werd bepaald aan de hand van een Mann-Whitney test weergegeven
met * indien 0,01 <p≤ 0,05 en met ** indien 0,001 <p≤ 0,01. (SEM: Standard Error of the Mean)).
C o n tr o l H . h e ilm a n n ii
H o fE
H . h e ilm a n n ii
w t
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
C P
Re
latie
ve
Cld
n1
ex
pr
es
sie
C o n tr o l H . h e ilm a n n ii
H o fE
H . h e ilm a n n ii
w t
0 .0
0 .5
1 .0
1 .5
2 .0
C P
Re
latie
ve
Cld
n3
ex
pr
es
sie
C o n tr o l H . h e ilm a n n ii
H o fE
H . h e ilm a n n ii
w t
0 .0
0 .5
1 .0
1 .5
2 .0
C P
Re
latie
ve
Zo
1e
xp
re
ss
ie
C o n tr o l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0
1
2
3
C P
Re
latie
ve
Cld
n1
ex
pr
es
sie
** *
C o n tr o l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
C P
Re
latie
ve
Cld
n3
ex
pr
es
sie
** *
*
C o n tr o l H . p y lo r i
S T M 1 3 8
H o fF
H . p y lo r i
P M S S 1
H . p y lo r i
S S 1
0 .0
0 .5
1 .0
1 .5
2 .0
C P
Re
latie
ve
Zo
1e
xp
re
ss
ie
* *

46
DEEL 5: DISCUSSIE
1. HET ONDERZOEK Tijdens dit onderzoek werd de invloed van een infectie met H. heilmannii en H. pylori op het gastro-
intestinaal stelsel en de BBB en BCSFB nagegaan. Er werd hierbij niet enkel gebruik gemaakt van de
wildtype stammen, maar ook van een mutant type voor zowel H. heilmannii als H. pylori. Door de infectie
met deze gemuteerde kiemen te onderzoeken, kon er worden nagegaan of deze mutaties het effect van
de infectie ter hoogte van de maag en hersenen beïnvloedden. H. pylori en NHPHs werden in het
verleden reeds geassocieerd met PD. Ze werden reeds teruggevonden bij patiënten die te kampen
hadden met een chronische maagontsteking (Dukowicz et al., 2007; Dobbs et al., 2008; Nielsen et al.,
2012; Tan et al., 2015; Csoti et al., 2016; Dobbs et al., 2016).
Op dit moment is de oorzaak van PD in 90 tot 95 procent van de gevallen nog onbekend en kan een
infectie met Helicobacter mogelijks een rol spelen in de pathogenese ervan (Wirdefelt et al., 2011; Mhyre
et al., 2012). De activatie van microgliacellen, resulterend in neuro-inflammatie en apoptose van
zenuwcellen als een gevolg van chronische gastrale inflammatie, zou een mogelijke verklaring kunnen
zijn voor het verband tussen een Helicobacter infectie en neurodegeneratieve aandoeningen (Dobbs et
al., 2000; Dobbs et al., 2008; Dobbs et al., 2010; Dobbs et al., 2012; Kountouras et al., 2012; Dobbs et
al., 2016). Er is echter wel nog verder onderzoek vereist naar de rol van een Helicobacter infectie op
het verloop van PD.
Een eerder onderzoek bestudeerde de invloed van een acute infectie met H. pylori en H. heilmannii en
hun bijhorende mutanten op het gastro-intestinaal stelsel in BALB/c muizen. Deze muizen bleken ten
opzichte van de C57BL/6 muizen een meer uitgesproken inflammatiebeeld te vertonen na infectie met
deze kiemen (Liu, 2016). Deze verhoogde inflammatie kan verklaard worden door het verschil in
immuunrespons. De C57BL/6 muizen zijn beschreven als genetisch predominante Th1 responders,
terwijl de BALB/c muizen voornamelijk Th2 responders zijn (Flahou et al., 2010). Verder werd in deze
studie ook aangetoond dat er een meer uitgesproken inflammatie was in de maag van BALB/c muizen
ten opzichte van C57BL/6 muizen na een infectie met H. suis. Een reden hiervoor zou kunnen zijn dat
een infectie met H. suis een predominante Th17/Th2 immuunrespons veroorzaakt, net zoals H.
heilmannii, in BALB/c en C57BL/6. Aangezien C57BL/6 muizen Th1 responders zijn zou dat dus de
minder uitgesproken inflammatie kunnen verklaren (Flahou et al., 2012; Joosten et al., 2013).
2. DE SPIJSVERTERINGSSTELSEL-HERSENEN AS IN RELATIE MET EEN H. PYLORI EN H.
HEILMANNII INFECTIE
Bewijs is zich aan het ophopen dat neuro-inflammatie een belangrijke rol speelt in de pathofysiologie
van PD. Gespecialiseerde centraal zenuwstelsel barrières, zoals de BBB en de BCSFB, spelen een
cruciale rol in het onderhouden van de homeostase van de hersenen, aangezien deze barrières bepalen
wat er wel en niet de hersenen mag binnendringen (De Bock et al., 2014). Dysfunctie van deze barrières
zou een rol kunnen spelen in de pathogenese van PD (Vawter et al., 1996; Stolp et al., 2009; Pisani et
al., 2012; Lee et al., 2014).

47
Mogelijke interacties tussen het gastro-intestinale stelsel en de hersenen zijn reeds goed gekend (Cryan
et al., 2012). Er wordt gesuggereerd dat neuro-inflammatie zou kunnen voortkomen uit aanwezige
infecties van het gastro-intestinaal stelsel (Dobbs et al., 2000; Dobbs et al., 2008; Dobbs et al., 2010;
Ferrari en Tarelli, 2011; Dobbs et al., 2013; Dobbs et al., 2016). Er is op dit moment reeds aangetoond
dat een intestinale infectie of dysbiose van het gastro-intestinaal stelsel mogelijks een rol speelt in de
pathogenese van PD. Gastrale Helicobacter spp. veroorzaken, indien onbehandeld, over het algemeen
een levenslange infectie aangezien de immuniteit van de gastheer niet in staat is om de infectie onder
controle te krijgen. Dit kan resulteren in een persisterende inflammatie en beschadiging van de
epitheelcellen van de maag (Montecucco en Rapuoli, 2001; Flahou et al., 2011; Flahou et al., 2012).
Beschadiging van de gastrale mucosa zou kunnen leiden tot lekkage van bacteriële componenten,
bacteriële metabolieten en pro-inflammatoire factoren in de bloedcirculatie, resulterend in een
systemische inflammatie (Fukuda et al., 2001; Sun et al., 2004; Fedwick et al., 2005; Guttman en Finlay,
2009; Wroblewski et al, 2011; Mayer et al., 2014; Caron et al., 2015). Deze inflammatoire mediatoren
dragen mogelijks bij aan de verstoring van de homeostase van de hersenen wat aanleiding kan geven
tot neurodegeneratieve ziekten zoals PD (Charlett et al. 1998; Dobbs et al. 1999; Dobbs et al., 2000;
Dobbs et al., 2008; Reale et al, 2009; Dobbs et al., 2010; Dobbs et al., 2012; Kontouras et al., 2015;
Dobbs et al., 2016).
3. INVLOED VAN EEN HELICOBACTER INFECTIE OP DE INTEGRITEIT VAN HET GASTRO-
INTESTINAAL STELSEL
3.1. KWANTIFICATIE De bekomen resultaten tonen aan dat de intragastrale infectie met de kiemen gelukt was. Verder was
er duidelijk te zien dat de kiemen enkel in staat zijn zich te handhaven in de maag en niet in de darm.
Dit werd ook reeds aangetoond in studies van Stolte et al. (1997) en Flahou et al. (2010). Gastrale
Helicobacter spp. zijn in staat zich in de maag te vestigen, onder andere omdat ze grote hoeveelheden
urease vormen die hen via vorming van ammoniak toelaten om het zure milieu van de maag te trotseren
(Weeks et al., 2000; Flahou et al., 2016; deel 1, paragraaf 1.1.2 en 1.2.2). Dit in tegenstelling tot de
enterohepatische soorten die voornamelijk terug te vinden zijn in de mucosale oppervlakten van het
intestinale stelsel en/of in de lever (Solnick en Schauer, 2001; Sterzenbach et al., 2007).
Als men kijkt naar de kwantificatie data dan ziet men dat er een hogere kolonisatiegraad is in de maag
voor de wildtypes van zowel H. pylori als H. heilmannii ten opzichte van de mutant. Dit is te verklaren
doordat de mutant een belangrijk gen, dat codeert voor een buitenste membraan proteïne, mist dat
noodzakelijk is voor de kolonisatie van de kiem. In een studie van Liu et al. (2016), met BALB/c muizen,
werd reeds aangetoond dat er een lagere kolonisatiegraad was in de maag van H. heilmannii stammen
die een gemuteerd HofE of HofF gen hadden ten opzichte van de wildtype bacteriën. Wij konden tijdens
dit onderzoek ook een lagere kolonisatiegraad waarnemen van de mutante kiemen ten opzichte van de
wildtypes, maar dan bij C57BL/6 muizen.

48
3.2. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR TIGHT JUNCTIONS Uit de genexpressie data bleek dat een infectie met Helicobacter het grootste effect uitoefent op de TJ
ter hoogte van het antrum en het colon. Er werd aangetoond dat er ter hoogte van het colon zeer weinig
tot geen kiemen aanwezig waren, zoals te zien was bij de kwantificatie data en toch is de expressie van
genen die coderen voor TJ sterk gewijzigd ten opzichte van de controle groep. Dit is mogelijks te wijten
aan een verstoring in het microbioom van het colon door de aanwezige infectie ter hoogte van de maag.
Veel recente studies hebben aangetoond dat de veranderingen in het gastro-intestinale microbioom
mogelijks een significant effect hebben op de gezondheid van de gastheer (Ochoa-Reparaz en Kasper,
2014; Walters et al., 2014; Zhang et al., 2015). Verder werd ook reeds gestaafd dat dysbiose
geassocieerd wordt met verschillende gezondheidsproblemen bij de mens (Clemente et al., 2012;
Zhang et al., 2015; Boulange et al., 2016).
3.3. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR
ONTSTEKINGSMEDIATOREN
De inflammatoire respons van de gastheer na een infectie met Helicobacter bacteriën wordt
gekarakteriseerd door een infiltratie van immuuncellen in de gastrale mucosa en submucosa en door
de secretie van ontstekingsmediatoren zoals Il1β, Lix en Kc ter hoogte van de infectieplaats (Flahou et
al., 2010; Peek et al., 2010; Flahou et al., 2012; Alvarez-Arellano et al., 2014; Hashim et al., 2014). Om
ook in deze studie de inflammatie in beeld te kunnen brengen werden er HE-kleuringen van de maag
aangewend om de uitgebreidheid van de totale gastritis te bepalen, waarbij er gebruik gemaakt werd
van de gemodificeerde visuele analoge schaal gelijk aan het aangepaste Updated Sydney System
(Flahou et al., 2010). Hiervan zijn op dit moment nog geen data voorhanden, maar dit is zeker
noodzakelijk om de, op basis van genexpressie aangetoonde, inflammatie in het gastro-intestinaal
stelsel, veroorzaakt door de infectie met de kiemen, te bevestigen.
3.4. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR VIP, VIPR1 EN VIPR2 Een opmerkelijke bevinding tijdens dit onderzoek was de gewijzigde expressie van Vip en zijn
receptoren VipR1 en VipR2 bij de muizen geïnfecteerd met H. pylori of H. heilmannii in vergelijking met
de controle dieren. Neuropeptiden zoals Vip zorgen normaal gesproken voor het behoud van de
homeostase van de mucosale barrière van het gastro-intestinaal stelsel (Anlauf et al., 2003; Chen et al.,
2015; Mulak et al., 2015). Een significante daling van de mucosale Vip levels werd ook reeds
teruggevonden in biopten van humane patiënten met chronische gastritis en ulcers (Bercik et al., 2002;
Li et al., 2009; Erin et al., 2012; Chen et al., 2015; Islek et al., 2016). Veranderingen in Vip expressie
zorgen mogelijks voor dysfunctie van de gastro-intestinale barrière en zouden op die manier
verantwoordelijk kunnen zijn voor het ontstaan van gastrale pathologieën.
Vip is zowel aanwezig in het CNS als in het gastro-intestinaal stelsel en zorgt daar voor de integriteit
van de barrières. Het gebruik van een “Vip knock-out” muismodel zou eventueel nieuwe informatie

49
kunnen geven omtrent de rol van Vip in het behoud van de integriteit van de gastro-intestinale barrières
en de hersenbarrières bij een infectie met H. pylori en H. heilmannii.
3.5. RELATIEVE MRNA EXPRESSIE VAN GENEN DIE CODEREN VOOR MUCINES In dit onderzoek werd gekeken naar veranderingen in genexpressie van Muc13 in het gastro-intestinaal
stelsel. Hierbij zagen we dat Muc13 in het colon significant gestegen was na een infectie met H.
heilmannii ASB1 (wildtype) en H. pylori (wildtype + mutant) terwijl er in het antrum geen significante
stijging werd waargenomen. Een onderzoek van Liu et al. (2014) toonde aan dat bij BALB/c muizen
geïnfecteerd met zowel H. heilmannii ASB1 (wildtype) als met H. pylori SS1, Muc13 wel significant
gestegen was in het antrum. Als men die resultaten vergelijkt met de data van dit onderzoek, dan is een
infectie met deze kiemen dus meer uitgesproken bij de BALB/c muizen dan bij de C57BL/6 muizen (Liu,
2016).
4. INVLOED VAN EEN HELICOBACTER INFECTIE OP DE HERSENBARRIÈRES Bij patiënten met PD werd reeds aangetoond dat de permeabiliteit van de hersenbarrières kan
toegenomen zijn en dat de morfologie van de hersenen ook grondig gewijzigd kan zijn (Stolp en
Dziegielewska, 2009; Nielsen et al., 2012). Om na te gaan welke invloed een infectie met H. heilmannii
of H. pylori heeft op de hersenbarrières, werd de relatieve permeabiliteit van de BCSFB en de BBB
onderzocht, 1 maand na een infectie met H. heilmannii of H. pylori (wildtype en mutant). Hieruit bleek
dat zowel de integriteit van de BCSFB als van de BBB niet significant aangetast was. Bij een onderzoek
naar een infectie met H. suis werd er wel een significante aantasting van de BCSFB aangetoond
(ongepubliceerde data). Blaecher et al. (2013) vonden een tienmaal hogere frequentie van gastrale
infecties met H. suis bij patiënten met PD dan bij een controle groep zonder dergelijke symptomen
(Blaecher et al., 2013). Dit was niet het geval voor H. heilmannii (F. Haesebrouck, persoonlijke
mededeling 2017). Waarom een gastrale infectie met H. suis in sterkere mate een effect heeft op de
BCSFB dan H. heilmannii is momenteel niet gekend.
5. CONCLUSIE EN AANBEVELINGEN Tijdens dit onderzoek is er gekeken naar veranderingen in genexpressie. Deze waargenomen
veranderingen gaan niet noodzakelijk gepaard met veranderingen op eiwitniveau. Ook al is er een
significante stijging of daling van een bepaald gen, dan weet men nog niet wat dit voor effect heeft op
eiwitniveau. Pas als men weet welke invloed een verandering in genexpressie heeft op eiwitniveau weet
men of een infectie met deze kiemen ook daadwerkelijk invloed heeft op het weefsel van de gastheer.
Om veranderingen op eiwitniveau te kunnen bepalen zou men eiwitkleuringen kunnen doen. Ook zou
men proteomics kunnen toepassen. Hiermee kan men een beeld krijgen van de eiwitten die op een
bepaald moment in een organisme aanwezig zijn. Om de cytokines in het serum te bepalen zou er
gebruik kunnen gemaakt worden van bioplex analyse. In een verder onderzoek zou kunnen nagegaan
worden of een Helicobacter infectie het verloop van PD beïnvloedt. Hierbij kan gebruik gemaakt worden

50
van muismodellen waarin de ziekte kan gereproduceerd worden. In dergelijke modellen kan men
onderzoeken of muizen geïnfecteerd met Helicobacter kiemen sneller PD ontwikkelen dan de controle
muizen (Sampson et al., 2016). Naast het onderzoeken van de integriteit van de spijsverteringsstelsel-
hersenen as zou men verlies van dopamine producerende neuronen in de substantia nigra van de
hersenen verder moeten onderzoeken. Dit kan gedaan worden door immunohistochemische kleuringen
zoals de α-synucleïne kleuring (Stefanis, 2012) of de ubiquitine kleuring (Chu et al., 2000).

51
LITERATUURLIJST
Abbott N. (2014). Anatomy and physiology of the blood–brain barriers. New York: Springer, Volume 10; p. 3–21. Abbott N.; Rönnbäck L.; Hansson E. (2006). Astrocyte-endothelial interactions at the blood–brain barrier. Nat Rev
Neurosci, 7, p. 41–53 Alm R.; Bina J.; Andrews B.; Doig P.; Hancock R.; Trust T. (2000). Comparative Genomics of Helicobacter pylori:
Analysis of the Outer Membrane Protein Families. Infect. Immun., volume 68, no. 7, p. 4155-4168 Álvarez-Arellano L.; Maldonado-Bernal C. (2014). Helicobacter pylori and neurological diseases: Married by the
laws of inflammation. World J Gastrointest Pathophysiol, volume 5, p. 400–404 Andersen L.; Boye K.; Blom J.; Holck S.; Nørgaard A.; Elsborg L. (1999). Characterization of a Culturable
“Gastrospirillum hominis” (Helicobacter heilmannii) Strain Isolated from Human Gastric Mucosa, J. Clin. Microbiol., volume 37, p.1069-1076
Andersen L.; Rosenstock S.; Bonnevie O.; et al. (1996). Seroprevalence of immunoglobulin G, M, and A antibodies to Helicobacter pylori in an unselected Danish population. Am J Epidemiol., volume 143, p. 1157-1164
Anlauf M.; Schäfer M.; Eiden L.; Weihe E. (2003). Chemical coding of the human gastrointestinal nervous system: cholinergic, VIPergic, and catecholaminergic phenotypes. J Comp Neurol., volume 459, p.90-111
Aranda V.; Haire T.; Nolan M.; Calarco J.; Rosenberg A.; Fawcett J.; Pawson T.; Muthuswamy S. (2006). Par6-aPKC uncouples ErbB2 induced disruption of polarized epithelial organization from proliferation control. Nat Cell Biol., volume 8, p. 1235-1245
Arnold I.; Zigova Z.; Holden M.; Lawley T.; Rad R.; Dougan G.; Falkow S.; Bentley S.; Müller A. (2011). Comparative Whole Genome Sequence Analysis of the Carcinogenic Bacterial Model Pathogen Helicobacter felis. Genome Biol Evol., volume 3, p. 302-308
Atherton J. (1997). Non-endoscopic tests in the diagnosis of Helicobacter pylori infection. Aliment Pharmacol Ther., volume 2, p. 2-20
Azevedo N.; Almeida C.; Fernandes I.; Cerqueira I.; Dias S.; Keevil C.; Vieira M. (2008). Survival of Gastric and Enterohepatic Helicobacter spp. in Water: Implications for Transmission. Appl. Environ. Microbiol., volume 74, p. 1805-1811
Baele M.; Decostere A.; Vandamme P.; Ceelen L.; Hellemans A.; Chiers K.; Ducatelle R.; Haesebrouck F. (2008a). Isolation and characterization of Helicobacter suis sp. nov. from pig stomachs. International Journal of Systematic and Evolutionary Microbiology, volume 58, p. 1350-1358
Baele M.; Haesebrouck F.; Vandamme P.; Van den Bulck K.; Gruntar I.; Mehle J.; Mast J.; Ducatelle R.; Decostere A. (2008b). Helicobacter baculiformis sp. nov. isolated from feline stomach mucosa. International Journal of Systematic and Evolutionary Microbiology, volume 58, p. 357-364
Bagheri H.; Damase-Michel C.; Lapeyre-Mestre M.; et al. (1999). A study of salivary secretion in Parkinson’s disease. Clin Neuropharmacol, 22, p. 213–215
Ballabh P.; Braun A.; Nedergaard M. (2004). The blood–brain barrier: an overview Structure, regulation, and clinical implications. Neurobiology of Disease, Volume 16, Issue 1, p. 1-13
Bell C.; Watson B.; Waring W. (2008). Acute psychosis caused by co-amoxiclav. BMJ., volume 337, p.996 Berardelli A.; Rothwell J.; Thompson P.; et al. (2001). Pathophysiology of bradykinesia in Parkinson’s disease.
Brain, 124, p. 2131–2146 Bercik P.; Collins S.; Verdu E. (2012). Microbes and the gut-brain axis. Neurogastroenterol Motil., Volume 24, issue
5, p. 405-413 Bercik P.; De Giorgio R.; Blennerhassett P.; Verdú E.; Barbara G.; Collins S.; (2002). Immune-mediated neural
dysfunction in a murine model of chronic Helicobacter pylori infection. Gastroenterol., volume 123, p. 1205-1215
Blaecher C.; Smet A.; Flahou B.; Pasmans F.; Ducatelle R.; Taylor D.; Weller C.; BjarnasoncI.; Charlett A.; Lawson A.; Dobbs R.; Dobbs S.; Haesebrouck F. (2013). Significantly higher frequency of Helicobacter suis in patients with idiopathic parkinsonism than in control patients. Helicobacter, volume 38, Issue 11-12, p. 1347-1353
Bolte E. (1998). Autism and Clostridium tetani. Med Hypotheses, volume 51, p. 133–144 Boulange C.; Neves A.; Chilloux J.; Nicholson J.; Dumas M. (2016). Impact of the gut microbiota on inflammation,
obesity, and metabolic disease. Genome Med., volume 8, p.42 Brown L. (2000). Helicobacter pylori: Epidemiology and Routes of Transmission. Epidemiologic Reviews, volume
22, p. 283-297 Caron T.; Scott K.; Fox J.; Hagen S. (2015). Tight junction disruption: Helicobacter pylori and dysregulation of the
gastric mucosal barrier. World J Gastroenterol., volume 21, p.11411–11427 Casagrande Proietti P.; Bietta A.; Brachelente C.; Lepri E.; Davidson I., Franciosini M. (2010). Detection of
Helicobacte spp. in gastric, fecal and saliva samples from swine affected by gastric ulceration. J Vet Sci., volume 11, p. 221–225
Chang C.; Chen L.; Yang J.; Lin J.; Chang K.; Wang J. (1999). Isolation of a Helicobacter pylori protein, FldA, associated with mucosa-associated lymphoid tissue lymphoma of the stomach. Gastroenterology, volume 117, p. 82-88
Charlett A.; Dobbs R.; Purkiss A.; Wright D.; Peterson D.; Weller C.; Dobbs S. (1998). Cortisol is higher in parkinsonism and associated with gait deficit. Acta Neurol Scand., volume 97, p. 77-85
https://www.ncbi.nlm.nih.gov/pubmed/?term=Calarco%20JP%5BAuthor%5D&cauthor=true&cauthor_uid=17060907

52
Chen L.; Yuan W.; Chen Z.; Wu S.; Ge J.; Chen J.; Chen Z. (2015). Vasoactive intestinal peptide represses activation of tumor-associated macrophages in gastric cancer via regulation of TNFα, IL-6, IL-12 and iNos. Internat Journal of Oncology, volume 47, p. 1360-1370
Chey W.; Wong B. (2007). Practice Parameters Committee of the American College of Gastroenterology. American college of gastroenterology guideline on the management of Helicobacter pylori infection. Am J Gastroenterol., volume 102, p. 1808–1825
Chu C.; Caruso J.; Cummings T.; Ervin J.; Rosenberg C.; Hulette C.; (2000). Ubiquitin immunochemistry as a diagnostic aid for community pathologists evaluating patients who have dementia. Mod Pathol., volume 13, p. 420-426
Clemente J.; Ursell L.; Parfrey L.; Knight R (2012). The impact of the gut microbiota on human health: an integrative view. Cell., volume 148, p. 1258-1270
Conlin V.; Wu X.; Nguyen C.; Dai C.; Vallance B.; Buchan A.; Boyer L.; Jacobson K. (2009). Vasoactive intestinal peptide ameliorates intestinal barrier disruption associated with Citrobacter rodentium-induced colitis. Am J Physiol Gastrointest Liver Physiol., volume 297, p.735-750
Conteduca V.; Sansonno D.; Lauletta G.; Russi S.; Ingravallo G.; Dammacco F. (2012). H. pylori infection and gastric cancer: State of the art (Review). International Journal of Oncology, volume 42, issue 1, p. 5-18
Cryan J.; Dinan T. (2012) Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci., volume 13, p. 701-712
Csoti I.; Jost W.; Reichmann H. (2016). Parkinson's disease between internal medicine and neurology. J Neural Transm., volume 123, p. 3-17
De Bock M.; Van den Bulck K.; Hellemans A.; Daminet S.; Coche JC.; Debongnie JC.; Decostere A.; Haesebrouck F.; Ducatelle R. (2007). Peptic ulcer disease associated with Helicobacter felis in a dog owner. European Journal of Gastroenterology & Hepatology, volume 19, issue 1, p. 79-82
De Bock M.; Van Haver V.; Vandenbroucke R.; Decrock E.; Wang N.; Leybaert L. (2016). Into Rather Unexplored Terrain – Transcellular Transport Across the Blood-Brain Barrier. GLIA 2016, volume 64, p. 1097-1123
De Bock M.; Vandenbroucke R.; Decrock E.; Culot M.; Cecchelli R.; Leybaert L. (2014). A new angle on blood–CNS interfaces: A role for connexins? FEB Letters, volume 588, Issue 8, p. 1259–1270
De Cooman L.; Flahou B.; Houf K.; Smet A.; Ducatelle R.; Pasmans F.; Haesebrouck F. (2013). Survival of Helicobacter suis bacteria in retail pig meat. International Journal of Food Microbiology, volume 166, issue 1, p. 164-167
De Rijk M.; Breteler M.; Graveland G.; et al. (1995). Prevalence of Parkinson’s disease in the elderly: the Rotterdam Study. Neurology, volume 45, p. 2143–2146
De Rijk M.; Tzourio C.; Breteler M.; et al. (1997). Prevalence of parkinsonism and Parkinson’s disease in Europe: the EUROPARKINSON Collaborative Study. European Community Concerted Action on the Epidemiology of Parkinson’s disease. J Neurol Neurosurg Psychiatry, volume 62, p. 10–15
De Vadder F.; Plessier F.; Gautier-Stein A.; Mithieux G. (2015). Vasoactive intestinal peptide is a local mediator in a gut-brain neural axis activating intestinal gluconeogenesis. Neurogastroenterol Motil., volume 27, p. 443-448
Delgado M.; Ganea D. (2003). Neuroprotective effect of vasoactive intestinal peptide (VIP) in a mouse model of Parkinson's disease by blocking microglial activation. FASEB J., volume 17, p. 944-946
Dieterich C.; Wiesel P.; Neiger R.; Blum A.; Corthésy-Theulaz I. (1998). Presence of Multiple “Helicobacter heilmannii” Strains in an Individual Suffering from Ulcers and in His Two Cats. J. Clin. Microbiol., volume 36,
p.1366-1370 Dobbs R.; Charlett A.; Dobbs S.; Weller C.; Ibrahim M.; Iguodala O.; Smee C.; Plant J.; Lawson A.; Taylor D.;
Bjarnason I. (2012). Leukocyte-subset counts in idiopathic parkinsonism provide clues to a pathogenic pathway involving small intestinal bacterial overgrowth. A surveillance study. Gut Pathogens, volume 4, p.
12
Dobbs R.; Charlett A.; Purkiss A.; Dobbs S.; Weller C.; Peterson D. (1999). Association of circulating TNF- and IL-6 with ageing and parkinsonism. Acta Neurol Scand., volume 100, p. 34-41
Dobbs R.; Dobbs S.; Weller C.; Charlett A.; Bjarnason I.; Curry A.; Ellis D.; Ibrahim M.; McCrossan M.; O'Donohue J.; Owen R.; Oxlade N.; Price A.; Sanderson J.; Sudhanva M.; Williams J. (2008). Helicobacter hypothesis for idiopathic parkinsonism: before and beyond. Helicobacter, volume 13, Issue 5, p. 309-322
Dobbs S.; Charlett A.; Dobbs R.; Weller C.; Iguodala O.; Smee C.; Lawson A.; Taylor D.; Bjarnason I. (2013). Antimicrobial surveillance in idiopathic parkinsonism: indication –specific improvement in hypokinesia following Helicobacter pylori eradication and non –specific effect of antimicrobials for other indications in worsening rigidity. Helicobacter, volume 18, p. 187-196
Dobbs S.; Dobbs R.; Weller C.; Charlett A. (2000). Link between Helicobacter pylori infection and idiopathic parkinsonism. Medical Hypothesis, volume 55, p. 93-98
Dobbs S.; Dobbs R.; Weller C.; Charlett A.; Augustin A.; Taylor D.; Ibrahim M.; Bjarnason I. (2016). Peripheral aetiopathogenic drivers and mediators of Parkinson’s disease and co-morbidities: role of gastrointestinal microbiota. Journal of NeuroVirology, volume 22, p.22–32
Dobbs S.; Dobbs RJ.; Weller C.; Charlett A.; Bjarnason I.; Lawson A.; Letley D.; Harbin L.; Price A.; Ibrahim M.; Oxlade N.; Bowthorpe J.; Leckstroem D.; Smee C.; Plant J.; Peterson D. (2010). Differential effect of Helicobacter pylori eradication on time-trends in brady/hypokinesia and rigidity in idiopathic parkinsonism. Helicobacter, volume 15, p. 279-294
https://www.ncbi.nlm.nih.gov/pubmed/?term=Hulette%20CM%5BAuthor%5D&cauthor=true&cauthor_uid=10786809
http://www.ncbi.nlm.nih.gov/pubmed/?term=Bjarnason%20I%5BAuthor%5D&cauthor=true&cauthor_uid=23336966
https://www.ncbi.nlm.nih.gov/pubmed/?term=Charlett%20A%5BAuthor%5D&cauthor=true&cauthor_uid=20633189

53
Donahue J.; Peek R.; Van Doorn L.; Thompson S.; Xu Q.; Blaser M.; Miller G. (2000). Analysis of iceA1 transcription in Helicobacter pylori. Helicobacter, volume 5, p. 1-12
Dorsey E.; Constantinescu R.; Thompson J.; Biglan K.; Holloway R.; Kieburtz K.; Marshall F.; Ravina B.; Schifitto G.; Siderowf A.; Tanner C. (2007). Projected number of people with Parkinson disease in the most populous nations, 2005 through 2030.Neurology, volume 68, p. 384-386
Dukowicz A.; Lacy B.; Levine G. (2007). Small Intestinal Bacterial Overgrowth. Gastroenterol Hepatol (N Y), volume 3, p.112–122
Eaton K.; Dewhirst F.; Radin.; Fox J.; Paster B.; Krakowka S.; Morgan D. (1993). Helicobacter acinonyx sp. nov., Isolated from Cheetahs with Gastritis. International Journal of Systematic and Evolutionary Microbiology, volume 43, p. 99-106
Ehrlich P. (1885). Das sauerstufbudurfnis des organismus. In: Eine Farbenanalytische Studie. Berlin: Hirschwald, p. 9
Ekman E.; Fredriksson M.; Trowald-Wigh G. (2013). Helicobacter spp. in the saliva, stomach, duodenum and faeces of colony dogs. The Veterinary Journal, volume 195, issue 1, p. 127-129
Erin N.; Türker S.; Elpek O.; Yildirim B. (2012). Differential changes in Substance P, VIP as well as neprilysin levels in patients with gastritis or ulcer. Peptides, volume 35, p. 218-224
Ernst F.; Homuth G.; Stoof J.; Mäder U.; Waidner B.; Kuipers E.; Kist M.; Kusters J.; Bereswill S.; van Vliet A. (2005). Iron-responsive regulation of the Helicobacter pylori iron-cofactored superoxide dismutase SodB is mediated by Fur. J Bacteriol., volume 187, p. 3687-3692
Fahn S. (2003). Description of Parkinson's Disease as a Clinical Syndrome. Annals of the New York Academy of Sciences, volume 991, p. 1–14
Fahn S.; Przedborski S. (2010). Parkinsonism. In: Rowland LP, Pedley TA, eds. Merritt's Neurology. New York: Lippincott Williams & Wilkins, p. 751–769
Fedwick J.; Lapointe T.; Meddings J.; Sherman P.; Buret A. (2005). Helicobacter pylori Activates Myosin Light-Chain Kinase To Disrupt Claudin-4 and Claudin-5 and Increase Epithelial Permeability. Infect Immun., volume 73, p. 7844–7852
Ferrari C.; Tarelli R. (2011). Parkinson’s disease and systemic inflammation. Parkinson’s Disease, volume 2011, p. 1-9
Ferrari C.; Tarelli R. (2011). Parkinson’s disease and systemic inflammation. Parkinson’s Disease Flahou B.; Haesebrouck F.; Chiers K.; Van Deun K.; De Smet L.; Devreese B.; Vandenberghe I.; Favoreel H.; Smet
A.; Pasmans F.; D'Herde K.; Ducatelle R. (2011). Gastric epithelial cell death caused by Helicobacter suis and Helicobacter pylori γ-glutamyl transpeptidase is mainly glutathione degradation-dependent. Cell Microbiol., volume 13, p.1933-1955
Flahou B.; Haesebrouck F.; Pasmans F.; D'Herde K.; Driessen A.; Van Deun K.; Smet A.; Duchateau L.; Chiers K.; Ducatelle R. (2010). Helicobacter suis causes severe gastric pathology in mouse and mongolian gerbil models of human gastric disease. PLoS One 5, e14083
Flahou B.; Haesebrouck F.; Smet A. (2016). Non-Helicobacter pylori Helicobacter Infections in Humans and Animals. Helicobacter pylori Research, p. 233-269
Flahou B.; Haesebrouck F.; Smet A.; Yonezawa H.; Osaki T.; Kamiya S. (2013). Gastric and Enterohepatic Non-Helicobacter pylori Helicobacters. Helicobacter, volume 18, Issue s1, p. 66-72
Flahou B.; Van Deun K.; Pasmans F.; Smet A.; Volf J.; Rychlik I.; Ducatelle R.; Haesebrouck F. (2012). The local immune response of mice after Helicobacter suis infection: strain differences and distinction with H. pylori. Vet Research, volume 43, p. 75
Foynes S.; Dorrell N.; Ward S.; Stabler R.; McColm A.; Rycroft A.; Wren B. (2000). Helicobacter pylori Possesses Two CheY Response Regulators and a Histidine Kinase Sensor, CheA, Which Are Essential for Chemotaxis and Colonization of the Gastric Mucosa. Infect. Immun., volume 68 no. 4, p. 2016-2023
Fuccio L.; Minardi M.; Zagari R.; Grilli D.; Magrini N.; Bazzoli F. (2007). Meta-analysis: duration of first-line proton-pump inhibitor based triple therapy for Helicobacter pylori eradication. Ann Intern Med, volume 147, p. 553–562
Fukuda Y.; Bamba H.; Okui M.; Tamura K.; Tanida N.; Satomi M.; Shimoyama T.; Nishigami T. (2001). Helicobacter pylori infection increases mucosal permeability of the stomach and the intestine. Digestion, volume 63, p. 93-96
Furuse M.; Sasaki H.; Tsukita S. (1999). Manner of interaction of heterogeneous claudin species within and between tight junction strands. J. Cell Biol., volume 147, p. 891–903
Gelb D.; Oliver E.; Gilman S. (1999). Diagnostic Criteria for Parkinson Disease. Arch Neurol., volume 56, p. 33-39 Gerhard A.; Pavese N.; Hotton G.; Turkheimer F.; Es M.; Hammers A.; Eggert K.; Oertel W.; Banati R.; Brooks D.
(2006). In vivo imaging of microglial activation with C(R)-PK11195 PET in idiopathic Parkinson’s disease. Neurobiology of Disease, Volume 21, Issue 2, p. 404–412
Gibb W.; Lees A. (1988). The relevance of the Lewy body to the pathogenesis of idiopathic Parkinson's disease. Journal of Neurology, Neurosurgery, and Psychiatry, volume 51, p. 745-752
Goodman KJ.; Correa P.; Tengana Aux HJ.; et al. (1996). Helicobacter pylori infection in the Colombian Andes: a population-based study of transmission pathways. Am J Epidemiol., volume 144, p. 290-299
Goodwin C.; Armstrong J.; Chilvers T.; Peters M.; Collins M.; Sly L.; McConnell W.; Harper W. (1989). Transfer of Campylobacter pylori and Campylobacter mustelae to Helicobacter gen. nov. as Helicobacter pylori comb. nov. and Helicobacter mustelae comb. nov., Respectively. Int J Syst Evol Microbiol, volume 39, p. 397-405

54
Graham D., Fischbach L. (2010). Helicobacter pylori treatment in the era of increasing antibiotic resistance. Gut, volume 59, p. 1143-1153
Graham D.; Shiotani A. (2008). New concepts of resistance in the treatment of Helicobacter pylori infections. Nat Clin Pract Gastroenterol Hepatol., volume 5, p. 321–331.
Greenberg E.; Anderson G.; Morgan D.; et al. (2011). 14-day triple, 5-day concomitant, and 10-day sequential therapies for Helicobacter pylori infection in seven Latin American sites: a randomised trial. Lancet, volume 378, p. 507–514
Grieb P.; Forster R.; Strome D.; Goodwin C.; Pape P. (1985). O2 exchange between blood and brain tissues studied with 18O2 indicator- dilution technique. J. Appl. Physiol., volume 58, p. 1929–1941
Grimm O.; Alm B. (2007). A case of ciprofloxacin-induced acute polymorphic psychosis with a distinct deficit in executive functions. Psychosomatics, volume 48, p. 269
Guttman J.; Finlay B. (2009). Tight junctions as targets of infectious agents. Biochimica et Biophysica Acta, volume 1788, p. 832-841
Haesebrouck F.; Pasmans F.; Flahou B.; Chiers K.; Baele M.; Meyns T.; Decostere A.; Ducatelle R. (2009). Gastric Helicobacters in Domestic Animals and Nonhuman Primates and Their Significance for Human Health. Clin Microbiol Rev, volume 22, p. 202–223
Haesebrouck F.; Pasmans F.; Flahou B.; Smet A.; Vandamme P.; Ducatelle R. (2011). Non-Helicobacter pylori Helicobacter Species in the Human Gastric Mucosa: A Proposal to Introduce the Terms H. heilmannii Sensu Lato and Sensu Stricto. Helicobacter, volume 16, p. 339–340
Hanada K.; Uchida T.; Tsukamoto Y.; Watada M.; Yamaguchi N.; Yamamoto K.; Shiota S.; Moriyama M.; Graham D.; Yamaoka Y. (2014). Helicobacter pylori infection introduces DNA double-strand breaks in host cells. Infect Immun., volume 82, p. 4182–4189
Hänninen M.; Happonen I.; Saari S.; Jalava K. (1996). Culture and characteristics of Helicobacter bizzozeronii, a new canine gastric Helicobacter sp.. International Journal of Systematic Bacteriology, volume 46, 160-166
Harper C.; Feng Y.; Xu S.; Taylor N.; Kinsel M.; Dewhirst F.; Paster B.; Greenwell M.; Levine G.; Rogers A.; Fox J. (2002). Helicobacter cetorum sp. nov., a Urease-Positive Helicobacter Species Isolated from Dolphins and Whales. J. Clin. Microbiol., volume 40, p. 4536-4543
Harper C.; Xu S.; Feng Y.; Lawrence Dunn J.; Taylor N.; Dewhirst F.; Fox J. (2002). Identification of Novel Helicobacter spp. from a Beluga Whale. Appl. Environ. Microbiol., volume 68, p. 2040-2043
Harris A.; Hinds F.; Beckhouse A.; Kolesnikow T.; Hazell S. (2002). Resistance to hydrogen peroxide in Helicobacter pylori: role of catalase (KatA) and Fur, and functional analysis of a novel gene product designated ‘KatA-associated protein’, KapA (HP 0874). Microbiology, volume 148, p. 3813-3825
Hashim H.; Azmin S.; Razlan H.;Yahya N.; Tan H.; Abdul Manaf M.; Ibrahim N. (2014). Eradication of Helicobacter pylori Infection Improves Levodopa Action, Clinical Symptoms and Quality of Life in Patients with Parkinson's Disease. PLoS One. 9, e112330
Healy D.; Abou-Sleiman P.; Lees A.; et al. (2004). Tau gene and Parkinson’s disease: a case-control study and meta-analysis. J Neurol Neurosurg Psychiatry, volume 75, p. 962–965
Heilmann K.; Borchard F. (1991). Gastritis due to spiral shaped bacteria other than Helicobacter pylori: clinical, histological, and ultrastructural findings. Gut, volume 32, p. 137-140
Hirase T.; Staddon J.; Saitou M.; Ando-Akatsuka Y.; Itoh M.; Furuse M.; Fujimoto K.; Tsukita S.; Rubin L. (1997). Occludin as a possible determinant of tight junction permeability in endothelial cells. J. Cell. Sci., volume 110, p. 1603–1613
Hirsch E; Jenner P; Przedborski S. (2013). Pathogenesis of Parkinsons’s Disease. Movement Disorders, volume 28, No 1, p. 24-30
Hirtz D.; Thurman D.; Gwinn-Hardy K.; Mohamed M.; Chaudhuri A.; Zalutsky R. (2007). How common are the “common” neurologic disorders? NEUROLOGY, volume 68, p. 326-337
Hook-Nikanne J.; Perez-Perez GI.; Blaser M. (1997). Antigenic characterization of Helicobacter pylori strains from different parts of the world. Clin Diagn Lab Immunol., volume 4, p. 592-597
Horiki N.; Omata F.; Uemura M.; Suzuki S.; Ishii N.; Iizuka Y.; Fukuda K.; Fujita Y.; Katsurahara M.; Ito T.; Cesar G.; Imoto I.; Takei Y. (2009). Annual Change of Primary Resistance to Clarithromycin among Helicobacter pylori Isolates from 1996 through 2008 in Japan. Helicobacter, volume 14, Issue 5, p. 438–442
Huang X.; Chen P.; Poole C. (2004). APOE-epsilon2 allele associated with higher prevalence of sporadic Parkinson disease. Neurology, volume 62, p. 2198–2202
Islek A.; Yilmaz A.; Elpek G.; Erin N. (2016). Childhood chronic gastritis and duodenitis: Role of altered sensory neuromediators. World Journal Gastroenterol., volume 22, p. 8349-8360
Iwanczak B.; Biernat M.; Iwanczak F.; Grabinska J.; Matusiewicz K.; Gosciniak G. (2012). the clinical aspects of helicobacter heilmannii infection in children with dyspeptic symptoms. Journal of physiology and pharmacology, volume 63, p. 133-136
Jafri N.; Hornung C.; Howden C. (2008). Meta-analysis: sequential therapy appears superior to standard therapy for Helicobacter pylori infection in patients naive to treatment. Ann Intern Med., volume 148, p. 923–931
Jalava K.; Kaartinen M.; Ultriainen M.; Happonen I.; Hänninen M. (1997). Helicobacter salomonis sp. nov., a canine gastric Helicobacter sp. related to Helicobacter felis and Helicobacter bizzozeronii. International Journal of Systematic Bacteriology, volume 4, p. 975-982
Jankovic J. (2008). Parkinson’s disease: clinical features and diagnosis. J Neurol Neurosurg Psychiatry, volume 79, p. 368-376

55
Joosten M.; Blaecher C.; Flahou B.; Ducatelle R.; Haesebrouck F.; Smet A. (2013). Diversity in bacterium-host interactions within the species Helicobacter heilmannii sensu stricto. Vet. Res., volume 44, p.65
Joosten M.; Flahou B.; Meyns T.; Smet A.; Arts J.; De Cooman L.; Pasmans F.; Ducatelle R.; Haesebrouck F. (2013). Case Report: Helicobacter suis infection in a Pig Veterinarian. Helicobacter, volume 18, Issue 5, p. 392-396
Joosten M.; Lindén S.; Rossi M.; Tay C.; Skoog E.; Padra M.; Peters F.; Perkins T.; Vandamme P.; Van Nieuwerburgh F.; D’Herde K.; Van den Broeck W.; Flahou B.; Deforce D.; Ducatelle R.; Marshall B.; Haesebrouck F.; Smet A. (2015). Divergence between the Highly Virulent Zoonotic Pathogen Helicobacter heilmannii and Its Closest Relative, the Low-Virulence "Helicobacter ailurogastricus" sp. nov. Infect Immun., volume 84, p.293-306
Kaklikkaya N.; Ozgur O.; Aydin F.; Cobanoglu U. (2002). Helicobacter heilmannii as causative agent of chronic active gastritis. Scandinavian Journal of Infectious Diseases, volume 34, p. 768-770
Kalach N.; Gosset P.; Dehecq E.; Decoster A.; Spyckerelle C. (2015). Usefulness of gastric biopsy-based real time-PCR for the diagnosis of Helicobacter pylori Infection in children. J Pediatr Gastroenterol Nutr, volume 61, p. 307-312
Kannarkat G.; Boss J.; Tansey M. (2013). The role of innate and adaptive immunity in Parkinson's disease. J Parkinsons Dis., volume 3, p. 493-514
Kawakubo M.; Ito Y.; Okimura Y.; Kobayashi M.; Sakura K.; Kasama S.; Fukuda M.; Fukuda M.; Katsuyama T.; Nakayama J. (2004). Natural antibiotic function of a human gastric mucin against Helicobacter pylori infection. Science, volume 305, p. 1003-1006
Kersulyte D.; Rossi M.; Berg D. (2013). Sequence Divergence and Conservation in Genomes of Helicobacter cetorum Strains from a Dolphin and a Whale. PLOS, volume 8, issue 12, e83177
Kim H.; Seo J.; Kim K. (2003). The effect of p38 mitogen-activated protein kinase on mucin gene expression and apoptosis in Helicobacter pylori-infected gastric epithelial cells. Ann N Y Acad Sci., volume 1010, p. 90-94
Kim J.; Chang J.; Chung S.; Yum J. (1999). Molecular Cloning and Characterization of theHelicobacter pylori fliD Gene, an Essential Factor in Flagellar Structure and Motility. J. Bacteriol. volume 181, no. 22, p. 6969-6976
Kivistö R.; Linros J.; Rossi M.; Rautelin H.; Hänninen M. (2010). Characterization of Multiple Helicobacter bizzozeronii Isolates From a Finnish Patient With Severe Dyspeptic Symptoms and Chronic Active Gastritis. Helicobacter, volume 15, Issue 1, p. 58–66
Kleinman M.; Frank S. (2013). Epidemiology and Clinical Diagnosis of Parkinson Disease. PET Clinics, volume 8, Issue 4, p. 447-458
Konturek J. (2003). Discovery by Jaworski of helicobacter pylori and its pathogenetic role in peptic ulcer, gastritis and gastric cancer. Journal of physiology and pharmacology, volume 54, p. 23-41
Kountouras J.; Zavos C.; Polyzos S.; Deretzi G. (2015). The gut-brain axis: interactions between Helicobacter pylori and enteric and central nervous systems. Ann Gastroenterol., volume 28, p. 506
Kufe D. (2009). Mucins in cancer: function, prognosis and therapy. Nat Rev Cancer, volume 9, p. 874-885 Kusters J.; van Vliet A.; Kuipers E. (2006). Pathogenesis of Helicobacter pylori Infection. Clin. Microbiol., volume
19, p. 449-490
Laterra J.; Keep R.; Betz L.; Goldstein G. (1999). Basic Neurochemistry: Molecular, Cellular and Medical Aspects. 6th edition. Bookshelf ID: NBK27998
Lau de L.; Breteler M. (2006). Epidemiology of Parkinson’s disease. volume 5, Issue 6, p. 525-535 Lau de L.; Koudstaal P.; Meurs van J.; Uitterlinden A.; Hofman A.; Breteler M.; (2005). Methylenetetrahydrofolate
reductase C677T genotype and PD. Ann Neurol., volume 57, p. 927–930 Lee A.; Hazell S.; O'Rourke J.; Kouprach S. (1988). Isolation of a spiral-shaped bacterium from the cat stomach.
Infection and Immunity, volume 56, p. 2843–2850 Lee H.; Pienaar I. (2014). Disruption of the blood-brain barrier in Parkinson’s disease: curse or route to cure? Front
Biosc (Landmark Ed), volume 19, p. 272-280 Li X.; Chen H.; Lu H.; Zheng Q.; Chen X.; Peng Y.; Ge Z.; Liu W. (2009). Role of Helicobacter pylori infection on
neuronal expression in the stomach and the spinal cord of a murine model. J. Dig. Dis., volume 10, p. 286-292
Linden S.; Nordman H.; Hedenbro J.; Hurtig M.; Boren T.; Carlstedt I. (2002). Strain- and blood group-dependent binding of Helicobacter pylori to human gastric MUC5AC glycoforms. Gastroenterology, volume 123, p. 1923-1930
Lindvall M.; Alumets J.; Edvinsson L.; Fahrenkrug J.; Hakanson R.; Hanko J.; Owman C.; Schaffalitzky de Muckadell O.; Sundler F. (1978). Peptidergic (VIP) nerves in the mammalian choroid plexus. Neurosci Lett., volume 9, p. 77-82
Liou JM.; Chen CC.; Chen MJ.; Chen CC.; Chang CY.; Fang YJ.; Lee JY.; Hsu SJ.; Luo JC.; Chang WH.; Hsu YC.; Tseng CH.; Tseng PH.; Wang HP.; Yang UC.; Shun CT.; Lin JT.; Lee YC.; Wu MS. (2013). Sequential versus triple therapy for the first-line treatment of Helicobacter pylori: a multicentre, open-label, randomised trial. The Lancet, volume 381, Issue 9862, p. 205–213
Litvan I.; Bhatia K.; Burn D.; et al. (2003). Movement Disorders Society Scientifi c Issues Committee report: SIC Task Force appraisal of clinical diagnostic criteria for Parkinsonian disorders. Mov Disord.; volume 18, p. 467–486
Liu C. (2016). PhD Thesis: Bacterial outer membrane proteins and host mucins involved in colonization of the gastric mucosa by the zoonotic pathogen Helicobacter heilmannii.

56
Liu C.; Rossi M.; Linden S.; Medae P.; Blaecher C.; Bauwens E.; Joosten M.; Flahou B.; Van den Broeck W.; Ducatelle R.; Haesebrouck F.; Smet A. (2016). The Helicobacter heilmannii hofE and hofF Genes are Essential for Colonization of the Gastric Mucosa and Play a Role in IL-1β-Induced Gastric MUC13 Expression, volume 21, p. 504-522
Liu C.; Smet A.; Blaecher C.; Flahou B.; Ducatelle R.; Linden S.; Haesebrouck F. (2014). Gastric de novo expression and spasmolytic polypeptide-expressing metaplasia during Helicobacter heilmannii infection. Infect. and Immun., volume 82, p. 3227-3239
Liu Y.; Roujeinikova A. (2015). Expression, refolding, purification and crystallization of the sensory domain of the TlpC chemoreceptor from Helicobacter pylori for structural studies. Protein Expr Purif., volume 107, p. 29–
34 Malfertheiner P.; Bazzoli F.; Delchier J.; et al. (2011). Helicobacter pylori eradication with a capsule containing
bismuth subcitrate potassium, metronidazole, and tetracycline given with omeprazole versus clarithromycin-based triple therapy: a randomised, open-label, non-inferiority, phase 3 trial.Lancet, volume 377, p. 905–
913 Malfertheiner P.; Megraud F.; O’Morain C.; Atherton J.; Axon A.; Bazzoli F.; Gensini G.; Gisbert J.; Graham D.;
Rokkas T.; El-Omar E.; Kuipers E.; The European Helicobacter Study Group (EHSG). (2012). Management of Helicobacter pylori infection – the Maastricht IV/ Florence Consensus Report. Gut, volume 61, p. 646 -
664 Maraganore D.; Farrer M.; McDonnell S.; et al. (2002). Case-control study of estrogen receptor gene polymorphisms
in Parkinson’s disease. Mov Disord, volume 17, p.509–512 Marshall B.; Warren J. (1984). Unidentified curved bacilli in the stomach of patients with gastritis and peptic
ulceration. The Lancet, volume 323, Issue 8390, p. 1311–1315 Matsumoto T.; Kawakubo M.; Akamatsu T.; Koide N.; Ogiwara N.; Kubota S.; Sugano M.; Kawakami Y.; Katsuyama
T.; Ota H. (2014). Helicobacter heilmannii sensu stricto-related gastric ulcers: A case report. World J Gastroenterol, volume 20, p. 3376-3382
Matter K.; Balda M. (2003). Signalling to and from tight junctions. Nat. Rev., Mol. Cell Biol., volume 4, p. 225–236
Mayer E.; Knight R.; Mazmanian S.; Cryan J.; Tillisch K. (2014) Gut microbes and the brain: paradigm shift in neuroscience. J Neurosci., volume 34, p.15490-15496
Mayeux R.; Marder K.; Cote L.; et al. (1995). The frequency of idiopathic Parkinson’s disease by age, ethnic group, and sex in northern Manhattan, 1988-1993. Am J Epidemiol., volume 142, p.820–827
McGee D.; Langford M.; Watson E.; Carter J.; Chen Y.; Ottemann K. (2005). Colonization and Inflammation Deficiencies in Mongolian Gerbils Infected by Helicobacter pylori Chemotaxis Mutants. Infect. Immun., volume 73, p. 1820-1827
McGuckin M.; Lindén S.; Sutton P.; Florin T. (2011). Mucin dynamics and enteric pathogens. Nature Reviews Microbiology, volume 9, p. 265-278
McNulty C.; Dent J.; Curry A.; Uff J.; Ford G.; Gear M.; Wilkinson S. (1989). New spiral bacterium in gastric mucosa. J Clin Pathol., volume 42, p. 585-591
Mehdi S. (2010). Antibiotic-induced psychosis: a link to D-alanine? Med Hypotheses, volume 75, p. 676–677 Mentis A.; Lehours P.; Megraud F. (2015). Epidemiology and Diagnosis of Helicobacter pylori infection Helicobacter,
volume 20, issue s1, p. 1-7 Mhyre T.; Boyd J.; Hamill R.; Maguire-Zeiss K. (2012). Parkinson’s Disease. Subcell Biochem., volume 65, p. 389–
455 Moayyedi P. (2007). Sequential regimens for Helicobacter pylori eradication. Lancet, volume 370, p. 1010–1012 Montecucco C, Rappuoli R. (2001). Living dangerously: how Helicobacter pylori survives in the human stomach.
Nature Reviews Molecular Cell Biology, volume 2, p. 457-466 Morens D.; Davis J.; Grandinetti A.; Ross G.; Popper J.; White L. (1996). Epidemiologic observations on Parkinson’s
disease: incidence and mortality in a prospective study of middle-aged men. Neurology, volume 46, p.1044–1050
Morgante L.; Rocca W.; Di Rosa A.; et al. (1992). Prevalence of Parkinson’s disease and other types of parkinsonism: a door-to-door survey in three Sicilian municipalities. Neurology, volume 42, p. 1901–1907
Mulak A.; Bonaz B. (2015). Brain-gut-microbiota axis in Parkinson’s disease. World Journal Gastroenterol, volume 21, p. 10609-10620
Murphy M.; Alcera L.; Gill J.; et al. (2008). The inexplicably suicidal patient. Curr Psychiatry, volume 7, p.73 Nielsen H.; Qiu J.; Friis S.; Wermuth L.; Ritz B. (2012). Treatment for Helicobacter pylori infection and risk of
parkinson’s disease in Denmark. European Journal of Neurology, volume 19, Issue 6, p. 864–869 Nussbaum R.; Ellis C. (2003). Alzheimer’s disease and Parkinson’s disease. N Engl J Med., volume 348, p.1356–
1364 Ochoa-Reparaz J.; Kasper L. (2014). Gut microbiome and the risk factors in central nervous system autoimmunity.
FEBS Lett., volume 588, p. 4214-4222 Olczak A.; Seyler R.; Olson J.; Maier R. (2003). Association of Helicobacter pylori antioxidant activities with host
colonization proficiency. Infect Immun., volume 71, p. 580-583 Oleastro M.; Ménard A. (2013). The Role of Helicobacter pylori Outer Membrane Proteins in Adherence and
Pathogenesis. Biology (Basel), volume 2, p. 1110-1134
Orlowski M.; Meister A. (1970). The gamma-glutamyl cycle: a possible transport system for amino acids. Proc Natl Acad Sci USA, volume 67, p.1248-1255

57
O'rourke J.; Solnick J.; Neilan B.; Seidel K.; Hayter R.; Hansen L.; Lee A. (2004). Description of ‘Candidatus Helicobacter heilmannii’ based on DNA sequence analysis of 16S rRNA and urease genes. Int J Syst Evol Microbiol, volume 54, p. 2203-2211
O'toole p.; Kostrzynska M.; Trust T. (1994). Non-motile mutants of Helicobacter pylori and Helicobacter mustelae defective in flagellar hook production. Molecular Microbiology, volume 14, Issue 4, p. 691–703
Ozsoylar G.; Sayin A.; Bolay H. (2007). Clarithromycin monotherapy-induced delirium. J Antimicrob Chemother., volume 59, p. 331
Pajares J.; Gisbert J. (2006). Helicobacter pylori: its discovery and relevance for medicine. Rev Esp Enferm Dig., volume 98, p. 770-785
Pardridge W.; Eisenberg J.; Yang J. (1985). Human blood–brain barrier insulin receptor. J. Neurochem., volume 44, p. 1771–1778
Parkinson J. (1817). An essay on the shaking palsy. J Neuropsychiatry Clin Neurosci., 2002 Spring, volume 14, p. 223-236
Parkinson Study Group. (2000). Pramipexole versus levodopa as the initial treatment for Parkinson's disease: a randomized controlled trial. JAMA, volume 284, p. 1931–1938
Parracho H.; Bingham M.; Gibson G.; McCartney A. (2005). Differences between the gut microflora of children with autistic spectrum disorders and that of healthy children. J Med Microbiol., volume 54, p. 987–991
Parsonnet J. (1995). The incidence of Helicobacter pylori infection. Aliment Pharmacol Ther., volume 9, p. 45-51 Parsonnet J.; Shmuely H.; Haggerty T. (1999). Fecal and oral shedding of Helicobacter pylori from healthy infected
adults. JAMA, volume 282, p. 2240-2245 Paster B.; Lee A.; Fox G.; Dewhirst F.; Tordoff L.; Fraser G.; O’Rourke J.; Taylor N.; Ferrero R. (1991). Phylogeny
of Helicobacter felis sp. nov., Helicobacter mustelae, and Related Bacteria. Int J Syst Evol Microbiol., volume 41, p. 31-38
Peek R.; J.; Fiske C.; Wilson K. (2010). Role of innate immunity in Helicobacter pylori-induced gastric malignancy. Physiol Rev., volume 90, p. 831-858
Peek R.; Thompson S.; Donahue J.; Tham K.; Atherton J.; Blaser M.; Miller G. (1998). Adherence to gastric epithelial cells induces expression of a Helicobacter pylori gene, iceA, that is associated with clinical outcome. Proc Assoc Am Physicians, volume 110, p. 531-544
Perez-Perez G.; Dworkin B.; Chodos J.; et al. (1988). Campylobacter pylori antibodies in humans. Ann Intern Med. Volume 109, p. 2-17
Pisani V.; Stefani A.; Pierantozzi M.; Natoli S.; Stanzione P.; Franciotta D.; Pisani A. (2012). Increased blood-cerebrospinal fluid transfer of albumin in advanced Parkinson’s disease. Journal of Neuroinflammation, volume 9, p.188
Plana M.; Blanch J.; Romero S. et al. (2006). Toxic catatonia secondary to azithromycin. J. Clin. Psychiatry, volume 67, p. 492–493
Pot R.; Stoof J.; Nuijten P.; De Haan L.; Loeffen P.; Kuipers E.; van Vliet A.; Kusters J. (2007). UreA2B2: a second urease system in the gastric pathogen Helicobacter felis. FEMS Immunol Med Microbiol, volume 50, p. 273–279
Pringsheim T.; Jette N.; Frolkis A.; Steeves T. (2014). The prevalence of Parkinson's disease: a systematic review and meta-analysis. Mov Disord., volume 29, p.1583-1590
Quik M. (2004). Smoking, nicotine and Parkinson’s disease. Trends Neurosci., volume 27, p. 561–568 Radin J.; Gonzalez-Rivera C.; Frick-Cheng A.; Sheng J.; Gaddy J.; Rubin D.; Algood H.; McClain M.; Cover T.
(2014). Role of connexin 43 in Helicobacter pylori VacA-induced cell death. Infect Immun., volume 82, p. 423–432
Rascol O.; Brooks D.; Korczyn A.; et al. (2000). A five-year study of the incidence of dyskinesia in patients with early Parkinson's disease who were treated with ropinirole or levodopa. N. Engl. J. Med., volume 342, p.
1484–1491 Reale M.; Iarlori C.; Thomas A.; Gambi D.; Perfetti B.; Nicola M.; Onofrj M. (2009). Peripheral cytokines profile in
Parkinson’s disease. Brain Behaviour Immunity, volume 23, p. 55-63 Roesler B.; Rabelo-Gonçalves E.; Zeitune J. (2014). Virulence Factors of Helicobacter pylori: A Review. Clin Med
Insights Gastroenterol., volume 7, p. 9–17 Ross G.; Abbott R.; Petrovitch H.; et al. (2000). Association of coffee and caffeine intake with the risk of Parkinson
disease. JAMA, volume 283, p. 2674– 2679 Rossi M.; Bolz C.; Revez J.; Javed S.; El-Najjar N.; Anderl F.; Hyytiäinen H.; Vuorela P.; Gerhard M.; Hänninen M.
(2012). Evidence for conserved function of γ-glutamyl transpeptidase in Helicobacter genus. PLoS One, volume 7, e30543
Sampson T.; Debelius J.; Thron T.; Janssen S.; Shastri G.; Ilhan Z.; Challis C.; Schretter C.; Rocha S.; Gradinaru V.; Chesselet M.; Keshavarzian A.; Shannon K.; Krajmalnik-Brown R.; Wittung-Stafshede P.; Knight R.; Mazmanian S. (2016). Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson's disease. Cell., volume 167, p. 1469-1480
Schmees C.; Prinz C.; Treptau T.; Rad R.; Hengst L.; Voland P.; Bauer S.; Brenner L.; Schmid R.; Gerhard M. (2007). Inhibition of T-cell proliferation by Helicobacter pylori gamma- glutamyl transpeptidase. Gastroenterology, volume 132, p.1820-1833
Schoenberg B.; Osuntokun B.; Adeuja A.; et al. (1988). Comparison of the prevalence of Parkinson’s disease in black populations in the rural United States and in rural Nigeria: door-to-door community studies. Neurology, volume 38, p. 645–646
https://www.ncbi.nlm.nih.gov/pubmed/?term=Sampson%20TR%5BAuthor%5D&cauthor=true&cauthor_uid=27912057
https://www.ncbi.nlm.nih.gov/pubmed/?term=Shastri%20GG%5BAuthor%5D&cauthor=true&cauthor_uid=27912057

58
Schott T.; Kondadi P.; Hänninen M.; Rossi M. (2011). Comparative Genomics of Helicobacter pylori and the human-derived Helicobacter bizzozeronii CIII-1 strain reveal the molecular basis of the zoonotic nature of non-pylori gastric Helicobacter infections in humans. BMC Genomics, volume 12, p. 534
Schwab R. (1964). Symptomatology and medical treatment of Parkinson's disease. Int J Neurol., volume 2, p. 61 Serlin Y.; Shelef I.; Knyazer B.; Friedman A. (2015). Anatomy and Physiology of the Blood-Brain Barrier. Semin
Cell Dev Biol., volume 38, p. 2-6 Seyler R.; Olson J.; Maier R. (2001). Superoxide dismutase-deficient mutants of Helicobacter pylori are
hypersensitive to oxidative stress and defective in host colonization. Infect Immun., volume 69, p. 4034-4040 Sgouras D.; Trang T.; Yamaoka Y. (2015). Pathogenesis of Helicobacter pylori Infection. Helicobacter, volume 20,
Issue S1, p. 8-16 Shaw W.; Kassen E.; Chaves E. (1995). Increased urinary excretion of analogs of Krebs cycle metabolites and
arabinose in two brothers with autistic features. Clin Chem., volume 41, p.1094–1104 Shibayama K.; Doi Y.; Shibata N.; Yagi T.; Nada T.; Iinuma Y.; Arakawa Y. (2001). Apoptotic signaling pathway
activated by Helicobacter pylori infection and increase of apoptosis-inducing activity under serum-starved conditions. Infect Immun., volume 69, p. 3181- 3189
Shibayama K.; Kamachi K.; Nagata N.; Yagi T.; Nada T.; Doi Y.; Shibata N.; Yokoyama K.; Yamane K.; Kato H.; Iinuma Y.; Arakawa Y. (2003). A novel apoptosis-inducing protein from Helicobacter pylori. Mol Microbiol., volume 47, p. 443-451
Sigal M.; Rothenberg M.; Logan C.; et al. (2015). Helicobacter pylori activates and expands Lgr5 stem cells through direct colonization of the gastric glands. Gastroenterology, volume 148, p. 1392–1404
Singh S.; Zambon C.; Basso D.; Pelloso M.; Tessari A.; Plebani M. (2015). Pathogenesis of Helicobacter pylori in Humans. Human Emerging and Re-emerging Infections: Viral and Parasitic Infections, Volume 1, chapter 1
Smet A.; Flahou B.; Mukhopadhya I.; Ducatelle R.; Pasmans F.; Haesebrouck F.; Hold G. (2011). The Other Helicobacters. Helicobacter, volume 16, Issue s1, p. 70–75
Solnick J.; Goldstein E. (2003). Clinical Significance of Helicobacter Species Other than Helicobacter pylori. Clin. Inf. Dis., volume 36, issue 3, p. 349-354
Solnick J.; Schauer D. (2001). Emergence of Diverse Helicobacter Species in the Pathogenesis of Gastric and Enterohepatic Diseases. Clin Microbiol Rev, volume 14, p. 59–97
Steer H.; Colin-Jones D. (1975). Mucosal changes in gastric ulceration and their response to carbenoxolone sodium. Gut, volume 16, p. 590-597
Stefanis L. (2012). α-Synuclein in Parkinson's Disease. Cold Spring Harb Perspect Med. volume 4, p. 1-23 Sterzenbach T.; Lee S.; Brenneke B.; von Goetz F.; Schauer D.; Fox J.; Suerbaum S.; Josenhans C. (2007).
Inhibitory effect of enterohepatic Helicobacter hepaticus on innate immune responses of mouse intestinal epithelial cells. Infect Immun., volume 75, p. 271-272
Stolp H.; Dziegielewska.K. (2009). Review: Role of developmental inflammation and blood–brain barrier dysfunction in neurodevelopmental and neurodegenerative diseases. Neuropathology and Applied Neurobiology, volume 35, Issue 2, p. 132–146
Stolte M.; Kroger G.; Meining A.; Morgner A.; Bayerdörffer E.; et al. (1997) A comparison of Helicobacter pylori and H. heilmannii gastritis. A matched control study involving 404 patients. Scand. J. Gastroenterol., volume 32, p. 28–33
Suerbaum S.; Josenhans C. (2007). Helicobacter pylori evolution and phenotypic diversification in a changing host. Nature rev. microbiol., volume 5, p. 441-452
Suerbaum S.; Michetti P. (2002). Helicobacter pylori Infection. N Engl J Med., volume 347, p. 1175-1186 Sun Y.; Söderholm J.; Petersson F.; Borch K. (2004). Long-standing gastric mucosal barrier dysfunction in
Helicobacter pylori-induced gastritis in Mongolian gerbils. Helicobacter, volume 9, p. 217-227 Sykora J.; Hejda V.; Varvarovská, J.; Stozicky, F.; Gottrand F.; Siala K. (2003). Helicobacter heilmannii Related
Gastric Ulcer in Childhood. Journal of Pediatric Gastroenterology & Nutrition, volume 36, p. 410-414 Talley N.; Fock K.; Moayyedi P. (2008). Gastric cancer consensus conference recommends Helicobacter pylori
screening and treatment in asymptomatic persons from high-risk populations to prevent gastric cancer. Am J Gastroenterol., volume 103, p. 510–14
Tan A.; Mahadeva S.; Marras C.; Thalha A.; Kiew C.; Yeat C.; et al. (2015). Helicobacter pylori infection is associated with worse severity of Parkinson's disease. Parkinsonism Relat Disord., volume 21, p. 221–225
Tan E.; Drozdzik M.; Bialecka M.; et al. (2004). Analysis of MDR1 haplotypes in Parkinson’s disease in a white population. Neurosci Lett., volume 372, p. 240–244
Tan E.; Khajavi M.; Thornby J.; Nagamitsu S.; Jankovic J.; Ashizawa T. (2000). Variability and validity of polymorphism association studies in Parkinson’s disease. Neurology, volume 55, p. 533–538
Tanner C.; Ottman R.; Goldman S.; et al. (1999). Parkinson disease in twins: an etiologic study. JAMA, volume 281, p. 341–346
Tansey M.; McCoy M.; Frank-Cannon T. (2007). Neuroinflammatory mechanisms in Parkinson's disease: Potential environmental triggers, pathways, and targets for early therapeutic intervention. Experimental Neurology, volume 208, Issue 1, p. 1–25
Taylor D.; Blaser M. (1991). The epidemiology of Helicobacter pylori infection. Epidemiol Rev., volume 13, p. 42-59 Terry K.; Williams S. Connolly L.; Ottemann K. (2005). Chemotaxis Plays Multiple Roles during Helicobacter pylori
Animal Infection. Infect. Immun., volume 73, no. 2, p. 803-811 Teymournejad O.; Mobarez A.; Hassan Z.; Moazzeni S.; Ahmadabad H. (2014). In vitro suppression of dendritic
cells by Helicobacter pylori OipA. Helicobacter, volume 19, p. 136–143

59
Tison F.; Dartigues J.; Dubes L.; Zuber M.; Alperovitch A.; Henry P. (1994). Prevalence of Parkinson’s disease in the elderly: a population study in Gironde, France. Acta Neurol Scand., volume 90, p.111–115
Tolosa E.; Wenning G.; Poewe W. (2006). The diagnosis of Parkinson’s disease. Lancet Neurol., volume 5, p.75–86
Twelves D.; Perkins K.; Counsell C. (2003). Systematic review of incidence studies of Parkinson's disease. Movement Disorders, volume 18, Issue 1, p. 19–31
Vaira D.; Vakil N; Menegatti M.; van’t Hoff B.; Ricci C.; Gatta L.; Gasbarrini G.; Quina M.; Pajares Garcia G.; van der Ende A.; van der Hulst R.; Anti M.; Duarte C.; Gisbert J.; Miglioli M.; Tytgat G. (2002). The Stool Antigen Test for Detection of Helicobacter pylori after Eradication Therapy. Annals of Internal Medicine, volume 136,
p. 280-287 Vaira D.; Zullo A.; Vakil N.; et al. (2007). Sequential therapy versus standard triple-drug therapy for Helicobacter
pylori eradication: a randomized trial. Ann Intern Med., volume 146, p. 556–563 Vakil N.; Vaira D. (2008). Sequential therapy for Helicobacter pylori: time to consider making the switch? JAMA,
volume 300, p.1346–1347 Van den Bulck K.; Decostere A.; Baele M.; Vandamme P.; Mast J.; Ducatelle R.; Haesebrouck F. (2006).
Helicobacter cynogastricus sp. nov., a Helicobacter species isolated from the canine gastric mucosa. International Journal of Systematic and Evolutionary Microbiology, volume 56, 1559-1564
Van Den Eeden S.; Tanner C.; Bernstein A.; et al. (2003). Incidence of Parkinson’s disease: variation by age, gender, and race/ethnicity. Am J Epidemiol., volume 157, p. 1015–1022
Van Loon S.; Aldert B.; den Hertog E.; Nikkels P.; Houwen R.; De Schryver J.; Oudshoorn J. (2003). Helicobacter heilmannii Gastritis Caused by Cat to Child Transmission. Journal of Pediatric Gastroenterology & Nutrition, volume 36, p. 407-40
Vawter M.; Dillon-Carter O.; Tourtellotte W.; Carvey P.; Freed W. (1996). TGFbeta1 and TGFbeta2 concentrations are elevated in Parkinson's disease in ventricular cerebrospinal fluid. Exp Neurol., volume 142, p. 313-322
Vermeer P.; Einwalter L.; Moninger T.; Rokhlina T.; Kern J.; Zabner J.; Welsh M. (2003). Segregation of receptor and ligand regulates activation of epithelial growth factor receptor. Nature. volume 422, p.322-326
Vermoote M.; Vandekerckhove T.; Flahou B.; Pasmans F.; Smet A.; De Groote D.; Van Criekinge W.; Ducatelle R.; Haesebrouck F. (2011). Genome sequence of Helicobacter suis supports its role in gastric pathology. Veterinary Research 2011, volume 42, p. 51
Vingerhoets F.; Schulzer M.; Calne D.; et al. (1997). Which clinical sign of Parkinson’s disease best reflects the nigrostriatal lesion? Ann Neurol., volume 41, p.58–64
Wachino J.; Shibayama K.; Suzuki S.; Yamane K.; Mori S.; Arakawa Y. (2010). Profile of Expression of Helicobacter pylori gamma-glutamyltranspeptidase. Helicobacter, volume 15, p 184-192
Walters W.; Xu Z.; Knight R. (2014). Meta-analyses of human gut microbes associated with obesity and IBD. FEBS Lett., volume 588, p. 4223-4233
Weeks D.; Eskandari S.; Scott D.; Sachs G. (2000). A H1-Gated Urea Channel: TheLink Between Helicobacter pylori Urease and Gastric Colonization. Science, volume 287, p. 482-485
Weis S.; Karagulle D.; Kornhuber J.; et al. (2006). Cotrimoxazole-induced psychosis: a case report and review of the literature. Pharmacopsychiatry, volume 39, p. 236–238
Wirdefeldt K.; Adami H.; Cole P.; Trichopoulos D.; Mandel J. (2011). Epidemiology and etiology of Parkinson's disease: a review of the evidence. Eur J Epidemiol., volume 26, p. 1-58
Wroblewski L.; Peek R. (2011). Targeted disruption of the epithelial-barrier by Helicobacter pylori. Cell Commun Signal., volume 9, p. 29
Wüppenhorst N.; von Loewenich F.; Hobmaier B.; Vetter-Knoll M.; Mohadjer S.; Kist M. (2013). Culture of a gastric non-Helicobacter pylori Helicobacter from the stomach of a 14-year-old girl. Helicobacter, volume 18, p. 1-5
Yakoob J.; Abbas Z.; Khan R.; Naz S.; Ahmad Z.; Islam M.; Awan S.; Jafri F.; Jafri W. (2012). Prevalence of non Helicobacter pylori species in patients presenting with dyspepsia. BMC Gastroenterology, volume 12, p 1-8
Yamashiro S. (2006). Dellmann's Textbook of Veterinary Histology. The Canadian Veterinary Journal, volume 48, p. 414
Yoshimura M.; Isomoto H.; Shikuwa S.; Osabe M.; Matsunaga K.; Omagari K.; Mizuta Y.; Murase K.; Murata I.; Kohno S. (2002). A Case of Acute Gastric Mucosal Lesions Associated with Helicobacter heilmannii Infection. Helicobacter, volume 7, Issue 5, p 322–326
Zhang G.; Ducatelle R.; De Bruyne E.; Joosten M.; Bosschem I.; Smet A.; Haesebrouck F.; Flahou B. (2015). Role of γ-glutamyltranspeptidase in the pathogenesis of Helicobacter suis and Helicobacter pylori infections. Vet Res., volume 46, p. 31
Zhang Y.; Pardridge W. (2001). Rapid transferrin efflux from brain to blood across the blood–brain barrier. J. Neurochem., volume 76, p. 1597–1600
Zullo A.; De Francesco V.; Hassan C.; et al. (2007). The sequential therapy regimen for Helicobacter pylori eradication: a pooled-data analysis. Gut, volume 56, p.1353–1357
https://www.ncbi.nlm.nih.gov/pubmed/?term=Vermeer%20PD%5BAuthor%5D&cauthor=true&cauthor_uid=12646923

60
BIJLAGE I: FOTO’S VAN DE WEEFSELCOLLECTIE
Fig. F. Collectie darmstalen Fig. E. Collectie choroïdale plexus van alle 4
de hersenventrikels
Fig D. Einde transcardiale perfusie met D-
PBS/heparine (0.2% heparine) aangevuld met
0.5% broomfenolblauw.
Fig. C. Begin transcardiale perfusie met D-
PBS/heparine (0.2% heparine) aangevuld met
0.5% broomfenolblauw.
Fig. A. Intraperitoneale injectie met een
ketamine/xylazine oplossing (Ceva/ Rompun
Bayer), voorafgaande aan de weefselcollectie.
Fig. B. Cisterna magna punctie om
cerebrospinaal vocht te verkrijgen uit het
vierde hersenventrikel