
The importance of being connected - Tufts...
135
Sara Gómez The importance of being connected Induced resistance to herbivory in Trifolium repens networks
Transcript of The importance of being connected - Tufts...

Sara Gómez
The importance of being connectedInduced resistance to herbivory in Trifolium repens networks

The importance of being connected Induced resistance to herbivory in Trifolium repens networks
Sara Gómez


The importance of being connected Induced resistance to herbivory in Trifolium repens networks
Een wetenschappelijke proeve op het gebied van de Natuurwetenschappen, Wiskunde en Informatica
Proefschrift
Ter verkrijging van de graad van doctor aan de Raadboud Universiteit Nijmegen
op gezag van de Rector Magnificus prof. mr. S.C.J.J. Kortmann, volgens besluit van het College van Decanen
in het openbaar te verdedigen op donderdag 22 mei 2008 om 13:30 uur precies
door
Sara Gómez geboren op 22 Augustus 1978
te Córdoba (Spanje)

Promotor: Prof. dr. J.C.J.M. de Kroon
Copromotor: Dr. J.F. Stuefer
Manuscriptcommissie: Prof. dr. J.M. van Groenendael
Prof. dr. Martin Heil (Politechnical Institute of Mexico, CINVESTAV)
Dr. Nicole M. van Dam (Netherlands Institute of Ecology, NIOO-KNAW)
ISBN: 978-90-9023039-9 Cover photo: Josef F. Stuefer

Para mis padres (To my parents)
“Writing a book is an adventure.
To begin with, it is a toy and an amusement;
then it becomes a mistress,
and then it becomes a master, and then a tyrant.
The last phase is that just as you are about to be reconciled to your servitude,
you kill the monster and fling him out to the public.”
~ Winston Churchill ~


TABLE O
F CO
NTEN
TS
Chapter 1 General introduction 9
Chapter 2 Clonal integration beyond resource sharing: 17 implications for defense signaling and disease transmission in clonal plant networks
Chapter 3 Members only: induced systemic resistance in a clonal plant network 33
Chapter 4 Time is precious: dynamics of induced resistance 47 in a clonal plant network
Chapter 5 Systemic induced resistance: 57 a risk-spreading strategy in clonal plant networks?
Chapter 6 Costs and benefits of induced resistance in a clonal plant network 75
Chapter 7 Summarizing discussion 91
References 101
Dutch Summary 123
Acknowledgements 129
List of publications 133
Curriculum vitae 134


Sara Gómez
CHA
PTER1
General Introduction

Chapter 1
10
CLONAL PLANT NETWORKS
Horizontal and vertical growth Clonality is a common form of plant growth in nature, being present in a wide array of forms and
constituting up to 70% of all plant species found in temperate Europe (van Groenendael and de Kroon
1990). In the case of stoloniferous plants, clonal growth is represented by the production of horizontal
stems above ground (stolons), which connect multiple genetically identical individuals (ramets), leading to
the formation of clonal plant networks (also referred to as clonal fragments). One of the most obvious
differences between clonal and non-clonal plants is that the former are able to transport resources, signals
and pathogens both vertically and horizontally (Figure 1).
The implications of horizontal transport, as explained below, confer clonal plants with remarkable
advantages in heterogeneous (Evans 1992, Stuefer 1997, Alpert et al. 2003) and harsh environments
(Shumway 1995). Horizontal transport allows for information exchange between interconnected
individuals, minimizing the impact of competition and optimizing the uptake of resources (Holzapfel and
Alpert 2003, Gruntman and Novoplansky 2004). Furthermore, horizontal transport also allows for the
transport of non-resource signals such as defense signals between interconnected ramets (Gómez and
Stuefer 2006).
FIGURE 1. Vascular flows in clonal and non-clonal plants. Clonal plants are able to share resources and defense signals both within
ramets (vertical transport) and between ramets (horizontal transport) through stolons or rhizomes by means of physiological
integration. This is a potential advantage over non-clonal plants, which lack the physical channels for the exchange of resources and
information between plants. Grey arrows indicate the direction of transport.
CLONAL PLANTS
NON-CLONAL PLANTS
CLONAL PLANTS
NON-CLONAL PLANTS

General introduction
11
In dense canopies with severe above and belowground competition, seed germination and seedling
survival is very limited, making vegetative propagation a considerable advantage. Clonal plants may
express different architectures and morphologies in response to neighbors (Solangaarachchi and Harper
1989, Turkington et al. 1991, Weijschedé et al., in press) or in response to environmental conditions
(Slade and Hutchings 1987a; 1987b, Sutherland 1987, Cain 1994, de Kroon & Hutchings 1995).
Large spatial scale The size of clonal plant networks ranges from few to very many connected ramets belonging to the same
network. In some cases individual clones can cover very large extensions. Examples of clonal networks
are not restricted to herbaceous species (e.g. strawberries, clovers). The largest clonal plant networks are
formed by woody species such as aspens (Populus tremuloides), where clones may cover several
hectares (reviewed in Cook 1985).
Inside the network: “what is mine is yours” Each ramet can produce leaves, roots and side meristems, providing them with all the structures
necessary for the uptake and production of their own resources (i.e. water, minerals, assimilates). This
confers the potential for functional independence. However, providing a ramet remains connected it can
share resources by means of physiological integration with other ramets (Pitelka and Ashmun 1985). In
heterogeneous environments, physiological integration among members of the clone confers great
benefits in terms of whole clone performance (Marshall 1990, Marshall and Price 1997). In cases where
there is heterogeneity in the availability of two resources (e.g. water and light) which spatially co-vary in a
complementary way (e.g. dry and high light vs. wet and low light), individual ramets may locally specialize
by investing larger biomass in the organ responsible for the uptake of the abundant resource. The surplus
of the abundant resource is transferred to other ramets in less favorable environmental conditions or in an
early developmental stage. The phenomenon of specialization and subsequent transport of resources
through physiological integration is known as division of labor, and has been shown to enhance the clone
productivity by up to 70% (Stuefer 1997).
The intranet transport of resources among ramets follows vascular source-sink dynamics (Marshall
1990). Mature, fully developed ramets (sources) support young developing ramets (sinks) by directing the
transport of resources towards them (acropetal transport). However, there is a degree of flexibility in the
dynamics of this dominant vascular flow, which allows for temporal changes in response to alterations in
source-sink patterns caused by a local change in the environmental conditions (e.g. shading). The extent
of flexibility in the vascular dynamics is limited by physical constraints imposed by the integration pattern
of each species (i.e. sectoriality). This refers to the degree of vascular connections shared among the
members of the clonal network. Some species are highly sectorial, having no or limited cross-transport of
resources between ramets from different orthostichies, while others are more fully integrated, sharing a
common pool of resources among all members of the clone (Hay and Sackville-Hamilton 1996; Price et al.
1996; Marshall and Price 1997). In contrast to the basic source-sink patterns, which can be temporally

Chapter 1
12
modified, sectoriality is a permanent constraint on resource and signal exchange within clonal plant
networks.
Individual ramets of clonal plants are not only able to respond to their changing environment in a
plastic way, but they can also coordinate their response with other members of the network (de Kroon et
al. 2005), taking the concept of plant plasticity to a higher level with additional possibilities to overcome
and exploit environmental difficulties.
PLANT DEFENSE AGAINST HERBIVORES
As sessile organisms, plants need to develop specific defense mechanisms to cope with herbivore
attacks. Plant defense can be expressed permanently (constitutive defense) in the form of morphological
traits or chemical compounds which reduce the performance or survival of the attacker. However, defense
is a costly process and the investment of resources in that function may result in an allocation trade-off to
other plant functions such as growth and reproduction (Herms and Mattson 1992). Plant defense theory
suggests that, plants have evolved a form of defense that is only activated when it is needed (induced
defense) in order to optimize resource allocation and minimize defense costs. This type of defense has
been described for over 100 plant-herbivore systems (Karban and Baldwin 1997). After local attack, plants
may enhance their protection by producing chemical compounds (e.g. phenolics: Arnold et al. 2004, Van
Dam et al. 2005) or developing physical structures (e.g. trichomes: Agrawal 2000; Traw and Dawson
2002) to reduce the negative impact of the attacker at the site of wounding.
After damage, there is a time lag until defense becomes active. This activation phase may take
from hours (Baldwin et al. 1994; Alborn et al. 1996) to days (Underwood 1998; McAuslane et al. 1997;
Agrell et al. 2003) or even years (Zvereva et al. 1997). Defense levels gradually decay over time back to
base levels (Schultz 1988, Underwood 1998).
Local activation of defense can then be further expressed in other undamaged parts of the plant
(systemic defense) preventing or reducing future damage (Agrawal et al. 1999a, Tollrian and Harvell
1999). The path followed by the defense agents within a plant is usually dictated by the source-sink
relationships and vascular flow. Only parts of the plant connected by phyllotaxy will be systemically
induced after a local attack (Jones et al. 1993, Orians et al. 2000, Viswanathan and Thaler 2004).
However, plants might be able to overcome constraints associated with internal defense signaling (e.g.
sectoriality) by using alternative external channels for information exchange about herbivory risks, such as
the release of airborne infochemicals into the environment (Karban et al. 2006).
Plant communication Communication is “a process by which information is exchanged between individuals through a common
system of symbols, signs, or behavior” (Merrian Webster dictionary). After herbivory, plants elicit signals
that may be used as cues by other neighboring plants. These signals can be released in the form of
airborne substances (i.e. volatiles organic compounds, VOCs; Karban and Baldwin 1997, Dicke and Bruin

General introduction
13
2001, Karban et al. 2003) and root exudates (Chamberlain et al. 2001, Dicke and Dijkman 2001, Guerrieri
et al. 2002) by the attacked plant. These may subsequently prompt a response in other undamaged plants
or parts of the damaged plant (Heil and Silva Bueno 2007; Frost et al. 2007). However, passing on
information through external channels may pose risks for the focal plant if the information about the
herbivory threat is used by competing neighboring plants, (i.e. eavesdropping; Karban et al. 2003; 2004)
as they can activate their defense system and become ready to counterattack before the actual herbivore
arrives. In the case of a subsequent herbivore attack, this confers an advantage for the eavesdropper by
reducing the damage received and consequently increasing their competitive ability with respect to the
sender plant.
Communication in clonal plants: members only Resource sharing between ramets of clonal plants has been extensively described in the literature
(Pitelka and Ashmun 1985; Alpert 1996; Hutchings and Wijesinghe 1997). However, besides resources,
considerably less is known about the transport of other substances such as defense compounds,
signaling molecules or pathogens (Stuefer et al. 2004). Stolons provide clonal plant networks an
advantageous feature over non-clonal plants. They provide a secure means to transfer information about
herbivore threats, avoiding the risk of eavesdropping and other problems linked to external information
transport (e.g., air current direction, distance between sender and receiver, etc). Sharing information
about herbivory risks in clonal plant networks can be considered as a special case of plant-plant
communication where the sender(s) and the receiver(s) are connected. Once a ramet is damaged by a
herbivore, it may pass on this information to other adjacent ramets, resulting in the induction of defensive
mechanisms in those undamaged ramets before the actual attack takes place. Damage inferred to a
single ramet might be a reliable cue predicting future damage, particularly since stolons may also act as
physical bridges from ramet to ramet, allowing some small herbivores (e.g. mites) to walk on them during
foraging.
The dark side of intranet communication Connections between ramets may incur large benefits for clonal plants in terms of support and defense,
especially for vulnerable developing ramets. However, there are some inherent risks of being connected.
The stolons do not only provide a physical bridge from ramet to ramet for small herbivores, they also
provide excellent means for the rapid internal spreading of systemic pathogens (Stuefer et al. 2004). Once
a ramet becomes infected, pathogens such as viruses can enter the vascular flow, resulting in the
infection of other members of the network (T. van Mölken & J.F. Stuefer, unpublished data). Where
virulent pathogens are concerned, this will result in the immediate extermination of (part of) the clone. In
cases of milder pathogens there are a number of strategies plants might use to counterattack the
infection, but once the clone is successfully infected, pathogens will remain in their host. The inherent

Chapter 1
14
longevity of clonal plants (Cook 1985) makes them long-term pathogen stores (Stuefer et al. 2004) and
sources of infection for neighboring plants.
AIM AND OUTLINE OF THE THESIS
The main aim of this thesis was to test the general concept that ramets of stoloniferous plants use their
internode connections for more than just resource sharing. In particular I wanted to investigate the use of
internode connections for defense signaling, and to explore the associated ecological implications. I used
the stoloniferous herb Trifolium repens as the model plant for all my experiments. T. repens is a classic
plant in the field of clonal plants research. Its morphological simplicity and the vast literature available on
intraclonal resource transport makes T. repens an ideal candidate for testing questions related to the
spatio-temporal dynamics of systemic induced resistance against herbivores in clonal plant networks. I
used larvae of the generalist herbivores Spodoptera exigua and Mamestra brassicae as herbivores to
induce and test for defense induction in the plants.
This thesis starts by making predictions about the ecological and evolutionary implications of
physiological integration between ramets (chapter 2). We predict that the activation of an early warning
system after local herbivore attack confers protection to other members of the network by means of
activation of direct (negative effect on herbivore’s performance and/or preference) and indirect (attraction
of natural enemies of the herbivore) defense traits. Additionally, in chapter 2 we also make predictions
regarding the risks of physiological integration, focusing in particular on virus transmission in clonal plant
networks. In the following chapters, I focus exclusively on the transport of defense agents in response to
attack by herbivores.
In chapter 3, we tested the early warning system hypothesis proposed in chapter 2. Specifically, we
tested whether ramets of T. repens are able to switch on their defense machinery in other parts of the
clone in response to local herbivory. Furthermore, we investigated the path and transport direction of the
defense agents in the network by experimentally manipulating phloem flows.
Once the internal transport of defense agents was confirmed, we investigated temporal aspects of
the induced response in clonal fragments (chapter 4). I studied the time necessary for the defense signal
to be transported and expressed from ramet to ramet after local attack and the time needed for the
induced resistance to decay after activation.
From chapter 3 it was concluded that defense induction had a predominantly acropetal expression
(i.e. transport from mature to young ramets). This unidirectional transport suggests that the initial point of
attack is crucial for the safety of other network members. Young ramets are strong sinks and if they are
attacked first, the signal will not be transported to other members of the network. In order to asses the
ecological viability of systemic resistance in clonal plant networks, I investigated herbivore preference
associated with ramet age and defense induction (chapter 5). A second aim of this chapter was to
elucidate the traits and mechanisms explaining observed preferences. To measure tissue quality, three
groups of traits were selected: biomechanics, nutrition and chemical defense. Traits were measured in

General introduction
15
young and mature ramets in control and defense-induced plants and correlated with herbivores
preference in dual-choice tests.
Induced resistance is generally a costly process (Bergelson and Purlington 1996; Strauss et al.
2002; Heil and Baldwin 2002) and in order for it to be an ecologically viable defense strategy, benefits
must outweigh or equal the costs of defense. In clonal plants, costs of defense expression may be
exceptionally large if all the members of the clone are defense-induced after local herbivory and there is
no subsequent herbivore attack. Alternatively, early warning after damage among members of the network
might confer large benefits by decreasing the damage inflicted by herbivores in case of subsequent attack
of other ramets. In order to assess the costs and benefits of induced resistance in a clonal network, we
grew defense-induced and control plants together in the presence and absence of herbivores respectively
and assessed plant performance and fitness related traits (chapter 6).
Finally, this thesis ends with a summarizing discussion (chapter 7) of the main results of this project
and the potential implications of early warning systems in clonal plant networks.

Chapter 1
16

JF Stuefer, S Gómez & T van Mölken. Evolutionary Ecology (2004)
CHA
PTER2
§ All authors have contributed equally to the ideas and preparation of this manuscript
Clonal integration beyond resource sharing: Implications for defense signaling and
disease transmission in clonal plant networks (§)

Chapter 2
18
ABSTRACT
Resource sharing between ramets of clonal plants is a well-known phenomenon, which allows stoloniferous
and rhizomatous species to internally translocate water, mineral nutrients and carbohydrates from sites of high
supply to sites of high demand. The mechanisms and implications of resource integration in clonal plants have
extensively been studied in the past. Vascular ramet connections are likely to provide an excellent means to
share substances other than resources, such as systemic defense signals and pathogens. The aim of this
paper is to propose the idea that physical ramet connections of clonal plants can be used (1) to transmit
signals, which enable members of clonal plant networks to share information about their biotic and abiotic
environments, and (2) to facilitate the internal distribution of systemic pathogens in clonal plant networks and
populations. We will focus on possible mechanisms as well as on potential ecological and evolutionary
implications of clonal integration beyond resource sharing. More specifically, we will explore the role of
physiological integration in clonal plant networks for the systemic transmission of direct and indirect defense
signals after localized herbivore attack. We propose that sharing defense induction signals among ramets may
be the basis for an efficient early warning system, and it may allow for effective indirect defense signalling to
herbivore enemies through a systemic release of volatiles from entire clonal fragments. In addition, we will
examine the role of clonal integration for the internal spread of systemic pathogens and pathogen defense
signals within clonal plants. Clonal plants may use developmental mechanisms such as increased flowering
and clone fragmentation, but also specific biochemical defense strategies to fight pathogens. We propose that
clonal plant networks can act as stores and vectors of diseases in plant populations and communities and that
clonal life histories favor the evolution of pathogens with a low virulence.

Non-resource sharing in clonal plant networks
19
INTRODUCTION
Many clonal plant species form horizontal stems (rhizomes or stolons), which expand laterally from the parent
plant and grow roots and new shoots at each of their nodes. This type of clonal growth leads to the formation of
networks of physically interconnected, genetically identical, and functionally autonomous offspring individuals,
called ramets. Physical connections between ramets allow for the internal transport of substances such as
water, carbohydrates and mineral nutrients between different parts of the clonal network. This phenomenon,
termed physiological integration or resource sharing, has received rather broad attention in the last few
decades, resulting in an extensive body of literature dealing with the physiological mechanisms as well as the
ecological implications of resource integration in clonal plants. Since the pioneering work of Qureshi and
Spanner (1971, 1973), and the seminal review by Pitelka and Ashmun (1985), clonal integration has attracted
considerable attention from (eco)-physiologists (e.g. Chapman et al. 1991, 1992a; Kemball and Marshall 1995),
ecologists (e.g. Shumway 1995; Alpert 1991, 1996; Evans 1991, 1992; Stuefer et al. 1994, 1996) and
theoretical biologists (Caraco and Kelly 1991; Oborny et al. 2000, 2001; Magori et al. 2003). These and
numerous other studies have helped to elucidate the mechanisms, preconditions and constraints of
carbohydrate, water and mineral nutrient sharing in clonal plants. They have also provided clear evidence for
the ecological importance of clonal integration for enabling plant species to provide their (clonal) offspring with
post-natal care, for avoiding the vulnerable life-cycle processes of seed germination and seedling establish-
ment, and for allowing an efficient resource extraction from heterogeneous environments and the provisioning
of internal support to damaged or resource-stressed ramets (Pitelka and Ashmun 1985; Marshall 1990;
Marshall and Price 1997).
While it is clear that physical connections between ramets can be used to transport resources within
clonal plants, much less is known about the possible function of stolon and rhizome connections for the internal
distribution of non-resource substances and agents, such as plant hormones, defense signals, toxins,
pathogens, and others. In spite of early suggestions that clonal plant networks may be used for sharing
substances other than water, carbohydrates and mineral nutrients (Cook 1978, Pitelka and Ashmun 1985), the
mechanisms, dynamics and implications of clonal integration beyond resource sharing has never been
considered in any comprehensive way so far. Nonetheless, it seems likely that vascular ramet connections are
an efficient and suitable means to distribute information among interlinked members of a clonal network, and
that clonal fragments bear an inherent risk of intrusion and possible rapid internal spread of systemic diseases.
Since any form of clonal integration (i.e., irrespective of what substance or agent is shared among connected
ramets) makes use of the vascular system for long-distance transport within clonal plant networks, non-
resource integration is likely to follow the same (or very similar) principles as ordinary resource integration,
including known constraints and limits on clone-internal movement of substances along vascular pathways.
Irrespective of their similarity with respect to mechanisms and basic principles, however, resource and non-
resource sharing are likely to have very different implications for plant functioning, and for plant responses to
the biotic and abiotic environment.

Chapter 2
20
The existing knowledge and broad experimental experience with resource sharing in clonal plants can
serve as an excellent basis for studying and predicting aspects of information sharing and disease spread in
clonal plants. Plant traits and environmental features, which are known to be of major importance for the
dynamics and the extent of resource integration, can also be expected to play a prominent role in non-resource
sharing. Architectural and physiological features, such as the branching structure, the average distance
between connected ramets, vascular sectoriality, source-sink dynamics, plant-internal water potential
gradients, as well as life-history traits, such as the longevity of ramets and ramet connections, are all likely to
affect processes and phenomena that rely on resource as well as non-resource integration. Sectoriality for
instance, can prevent resources and anything else transported in the vascular system from reaching all parts of
a plant (Watson and Casper 1984; Vuorisalo and Hutchings 1996; Stuefer 1996). In clonal plants, sectoriality
can effectively isolate ramets or larger clone parts in terms of physiological integration (Hay and Sackville-
Hamilton 1996; Price et al. 1996). Sectoriality can be regarded as “a structural constraint on the transport of
resources in both the phloem and xylem that results in their distribution to restricted regions of the plant”
(Marshall and Price 1997).
In addition, aspects of the (biotic and abiotic) environment can promote and constrain physiological
integration in clonal plants. Spatial habitat heterogeneity (e.g. in the availability of resources or other abiotic
and biotic factors) has often been shown to promote or constrain the exchange of resources between
interconnected clone parts of clonal plants (Stuefer and Hutchings 1994; Evans 1991; 1992; Shumway 1995).
The longevity and transport capacity of connecting stolon or rhizome internodes is an example for plant-internal
features that strongly affect the spatio-temporal degree and the quantitative amount of physiological integration
and related processes in clonal plants (Stuefer et al. 1998; Oborny et al. 2001). These and other plant-internal
and external features can thus be expected to critically affect non-resource integration and its implications in
clonal plants.
The aims of this paper are (1) to explore processes and phenomena which rely on the transmission of
signals and other non-resource agents and substances through physical connections between ramets of a
clonal plant, (2) to draw attention to possible ecological implications of non-resource integration, and (3) to put
forward expectations, predictions and hypotheses which can be tested in future research initiatives. Due to the
scarcity of specific background information, most of our predictions will be speculative in nature. We will focus
on two main topics, namely the role of clonal integration (a) for distributing defense induction signals in
response to herbivory and (b) on mechanisms and implications of the systemic spread of pathogens and
pathogen defense signals in clonal plant networks.
PLANT HERBIVORE DEFENSE
Plants have developed a broad array of mechanisms to cope with herbivores. Plant defense traits can be
constitutive or inducible (i.e. plastic), and defense strategies may be direct or indirect. Constitutive defense
traits are always expressed, even at times and in environments where they are not needed (Wittstock and

Non-resource sharing in clonal plant networks
21
Gershenzon 2002). Plastic defense traits are only expressed after initial damage or they may be induced by
external signals such as volatiles (Bruin et al. 1991; Karban and Baldwin 1997; Dicke and Hilker 2003).
Defense mechanisms can have direct or indirect effects on herbivores. Direct defenses consist of inducible
changes in tissue quality, plant palatability and toxicity, or in plastic alterations of anatomical and morphological
traits that reduce the herbivores’ preference for, or performance on the host plant (Karban and Baldwin 1997).
Many plants can also make use of indirect defenses against herbivores (e.g. damaged plants may release
specific info-chemicals to attract the natural enemies of the herbivore; Karban and Baldwin 1997; Takabayashi
and Dicke 1996).
Inducible defenses can either be exclusively expressed at the site of damage or they can also be
activated in other, undamaged parts of the plant. The latter phenomenon, termed in this text as induced
systemic resistance (ISR) is a common defense mechanism of plants against herbivores (Agrawal et al. 1999d;
Tollrian and Harvell 1999). The complex signaling cascade leading to the local induction, systemic spread of
the signal and non-local activation of defense traits is not yet fully understood (Roda and Baldwin 2003).
Nevertheless it seems clear that the jasmonic acid pathway and phloem-transmitted signals play a key role in
the systemic induction of defense genes after herbivore damage (e.g. proteinase inhibitor genes in
Solanaceae; Stastwick and Lehman 1999; Stratmann 2003; Thaler et al. 2002a; 2002b). ISR, though
mechanistically not fully understood, has been described for numerous plant-herbivore systems (Karban and
Baldwin 1997; Agrawal et al. 1999d; Tollrian and Harvell 1999).
A) EARLY WARNING SYSTEM?
Herbivory triggers defensive responses in host plants showing systemic inducible resistance. Herbivore feeding
(and in some cases also mechanical leaf damage) elicits a specific response, which leads to the production of
a defense induction signal at the site of attack (Karban and Baldwin 1997). This alerting signal then travels
through vascular pathways (i.e. phloem vessels) from the site of damage to other parts of the plant. As a
consequence undamaged plant parts will also activate their defensive machinery. Though never tested
explicitly, stolon and rhizome connections between ramets of clonal plants are most likely to act as pathways
for systemic defense induction signals (Haukioja et al.1990, Haukioja 1991).
If herbivores attack one or a few ramets of a clonal plant, a warning signal may be produced at the site
of damage and sent to other uninfested ramets through physiological clonal integration. This may allow
undamaged ramets to receive a defense induction signal even if they are rather distant from the site of the first
herbivore attack, and to activate direct or indirect defense mechanisms before the herbivores actually arrive.
ISR in clonal plants can be seen as a preemptive defense strategy of connected, uninfested ramets against
impending herbivore attacks. Such a spatial alerting strategy is beneficial whenever the induction signal
spreads faster than the herbivore, and if the initial herbivore attack is a reliable cue for future damage in
connected ramets of a clonal plant (Karban et al. 1999). Due to the potentially large size of clonal plant
networks, they may constitute an ideal system to study the complex space-time relationships between benefits
and costs of ISR on one hand, and cue reliability and the spatio-temporal dynamics of herbivore spread in

Chapter 2
22
relation to the speed and extent of systemic signal transduction, on the other hand. The rate and spatial
dimensions of internal signal transmission can be expected to depend crucially on architectural plant
characteristics such as average ramet distance, vascular sectoriality, source-sink relationships, and on the
transport capacity of connecting internodes between ramets.
In most cases herbivore damage on a given ramet is likely to entail a considerable risk of attack for
adjacent ramets. However, the risk of attack by an herbivore present on a connected sibling ramet should
decrease with increasing distance from the point of initial damage. In other words, the information content of
the systemic warning signal is very likely to decrease with increasing distance between sender and receiver
ramets in a clonal plant network. The rate of decrease (i.e., the exact decay profile of spatial autocorrelation in
attack risk) depends on the behavior, mobility and population size of the herbivore, the size and architecture
(e.g. average ramet distances) of the clonal plant under attack, and on general canopy characteristics (e.g.
presence of alternative host plants). To be beneficial in the long run, the physiological and ecological costs of
inducible defenses should be equaled or outweighed by benefits in terms of enhanced plant performance and
fitness (Karban and Baldwin 1997; Heil 2001; 2002; Cipollini et al. 2003). This implies that uninfested ramets of
a clonal plant should only respond to the systemic signal and switch on their inducible defenses if the danger of
being attacked is high enough to justify the costs of inducing and temporarily expressing defense traits (Karban
et el. 1999). Any mismatch in time or space between systemic defense induction and herbivore attack is likely
to lead to a costly misallocation of resources. Examples for such mismatches include cases in which
herbivores disperse faster than the induction signal and cases in which the herbivore does not spread to
adjacent ramets (no attack of systemically induced ramets).
Intermediate levels of spatio-temporal spread of the induction signal may be the most appropriate
response of many clonal plant networks to localized herbivore attack. If the spatial scale of the systemic
defense induction is too local (e.g. induction occurs only in the nearest neighbor ramets) or if the induction
does not spread fast enough within connected ramets, the herbivores may be able to disperse to surrounding
ramets before systemic induction has occurred there. If, however, the scale of systemic induction is too large,
distant ramets of a clonal fragment may be induced even though the risk of being reached by the herbivore is
rather low. A graded response of ramets according to their distance to the site of attack, possibly mediated by
a decrease in induction over space and time, may also allow optimizing the cost-benefit balance of systemically
induced defense mechanisms in response to local herbivore damage. No data are currently available to
confirm or to reject this and other predictions about the mechanisms and implications of ISR in clonal plant
networks.
Although the same induction and defense mechanisms are expected to operate in clonal and non-clonal
plants, sharing ISR elicitors between integrated ramets of clonal plants could have important population-level
consequences for plant performance, plant-plant and plant-herbivore interactions. The systemic induction of
direct defense traits in clonal ramet populations could act as an effective spatial information and early warning
system for spatially scattered network members in case of local herbivore attack. Such an early warning
system would be particularly beneficial for clonal plants by conferring integrated ramets faster and better
protection from herbivore damage (through reduced palatability or other direct defenses) than their

Non-resource sharing in clonal plant networks
23
surrounding, uninduced competitors. This effect may be enforced by the selectivity of herbivores which may be
discouraged from feeding on the induced parts of a clonal fragment and move preferentially to neighboring host
plantsto feed. In herbivore-prone environments the ability of clonal fragments to share ISR signals over
considerable distances may critically affect competitive relations between clonal and non-clonal plants.
Physical inter-ramet connections may not only reduce, but can also increase the risk and intensity of
future herbivore damage. In many clonal plant species, ramets are connected by aboveground structures (i.e.
stolon internodes). They can be used as bridges by foraging herbivores, guiding them to uninfested ramets.
This negative effect of physical integration should be strongest if clonal plants with a sparse growth habit
(relatively long inter-ramet distances) are attacked by specialist herbivores with limited mobility. Except for
enhancing protection through ISR, clonal connections between ramets might thus also increase the chance of
being located by herbivores in specific cases.
Resource flows and transport pathways within clonal plant networks allow for, but also constrain the
distribution of ISR signals among interconnected ramets. Resource flows in the xylem and in the phloem are
largely governed by plant-internal source-sink relationships and water potential gradients, which are a function
of environmental factors (such as heterogeneity in resource availability) and of the developmental relations
between connected ramets and branches (Marshall 1990). Transport of resources and signals can be severely
constrained by the physical construction of the vascular system (sectoriality) and/or by predominantly
unidirectional flows of organic resources in several clonal species (Marshall and Price 1997). In addition,
herbivore attacks themselves, may change the source-sink relationships and resource movement patterns
within interconnected groups of ramets (Haukioja 1991; Honkanen and Haukioja 1998; Honkanen et al. 1998),
thereby affecting the direction and/or speed of information sharing within the network.
Generally speaking, the effects of induced systemic resistance in clonal plants should be strongest in
plants with large-scale phloem integration among connected ramets and little directional constraints on
carbohydrate movement (e.g. Potentilla spp.; Stuefer et al. 1994; van Kleunen and Stuefer 1999; Hydrocotyle
bonariensis; Evans 1991; 1992). Strongly sectorial species, such as Glechoma hederacea, Trifolium repens
(Price et al. 1992a; 1992b; Price et al. 1996; Hay and Sackville-Hamilton 1996; Lötscher and Hay 1997) and
possibly most other species with monopodial stem growth and so-called leaf ramets (i.e. ramets that consist of
only one leaf, an axillary meristem, and a root system) are likely to be constrained in the internal transmission
of signals. Sectoriality, however, may not strongly affect the effectiveness of ISR in clonal plants, if herbivore
damage and defense induction occurs not on a single but on several adjacent ramets. Due to the basic
differences in vasculature, monocotyledonous species are less likely to be constrained by sectoriality than
dicots (Stuefer 1996).
Based on knowledge of clone-internal resource sharing, we can expect that young ramets and growing
parts of the clonal network in general will be most likely to receive induction signals shortly after an attack has
happened, because they usually represent strong sinks for carbohydrates (Marshall 1990). Environmental
heterogeneity, such as partial shading of a clonal plant network, may enhance the transmission rate and
distance of ISR signals by promoting the transport of phloem based resources to stressed clone parts.
Environmental heterogeneity imposing source-sink gradients is also likely to block or constrain the dispersal of

Chapter 2
24
A. Clonal plant network B. Non-clonal plants
defense signals from ramets that act as strong resource sinks. Whether herbivores can make use of this fact
(e.g. by preferentially feeding on shaded ramets in habitats with small-scale heterogeneity in light conditions) is
unknown to date.
B) MASSIVE PERFUME ATTRACTION
Many plant species emit a specific blend of gaseous substances (volatiles) when they are damaged by
herbivores. The volatile mixture and concentration profile of constituent compounds can be species-specific
and may differ for different herbivores. In various systems damage-induced volatiles attract natural enemies of
the herbivores (such as carnivores or parasitoids) that are causing the damage through feeding (Pichersky and
Gershenzon 2002; Dicke et al. 1993a). Strong evidence from the scientific literature and the successful
application of such triangular interactions for the biological control of herbivores (Karban and Baldwin 1997 and
studies quoted there) confirm the effectiveness of this indirect defense mechanism in several plant-animal
systems. Experimental studies have shown that recruiting volatiles may not only be emitted from the site of
damage, but systemic induction of volatile emission can lead to the production and release of info-chemicals
from undamaged parts of a plant (Dicke et al. 1993b; Takabayashi and Dicke 1997; Dicke and Dijkman 2001).
In the case of large clonal plant networks such as those formed by many stoloniferous and rhizomatous
species, a systemic emission of volatiles could lead to a significant amplification (in terms of air volume
containing the info-chemical) of the indirect defense signal, and a highly increased chance of attracting natural
enemies of the herbivore that caused the defense induction (Figure 1).
Figure 1. Emission of volatiles after initial herbivory in one ramet (black). (A) The attacked ramet sends a signal to induce volatile emission in adjacent ramets to attract enemies of the herbivores. To help predators locate their prey, there should be a concentrationgradient in relation to the distance from the attack point (fading colored arrows). (B) In non-clonal plants, only the attacked individual can systemically induce volatiles in other parts of the plant. Only very few and partly controversial studies (Dicke and Bruin 2001 and studies mentioned therein) have reported volatile emission of neighboring plants after contact with volatiles from infested plants.
The release of volatiles from numerous ramets after systemic induction would facilitate the attraction of
predators and parasitoids. However, the spatial de-coupling of herbivore position and volatile release due to
the systemic transmission of induction signals among scattered ramets of a clonal network may reduce the
information content of the volatile signal, thereby potentially jeopardizing the ability of predators and parasitoids

Non-resource sharing in clonal plant networks
25
to locate their prey. This may occur if infested and uninfested ramets emit volatiles in similar concentrations
after an herbivore attack. According to Dicke (1994/1995), however, the emission of volatiles from uninfested
plant parts may be weaker than the emission from the site of wounding. If so, the strength of the volatile signal
produced by interconnected ramets may decrease with increasing distance from the attacked ramet, thereby
creating a concentration gradient which can facilitate the predator or parasitoids to locate the herbivores
among infested and uninfested ramets of a clonal plant. Experimental studies are needed to clarify the
existence and action of indirect defense signaling from clonal plant networks to enemies of their herbivores.
Clonal architecture can also affect the total volume and concentration of emitted volatiles. One can
expect that species with a spatially scattered distribution of ramets (“guerrilla”-type of clonal growth) produce
bigger and less concentrated volatile emissions than species with an aggregated ramet distribution (“phalanx”-
type). In both cases, however, clonal plants are likely to release a bigger amount of volatiles than infested
individuals of comparable non-clonal plants or non-integrated ramets of clonal species. Indirect defenses of this
type could provide clonal plants with an increased protection, which could result in an enhanced competitive
strength of clonal versus non-clonal plants in herbaceous canopies. This prediction can be tested in experi-
mental studies by comparing volatile emissions and carnivore attraction between integrated and non-integrated
ramets of clonal plants, or by competition experiments between clonal and non-clonal plants grown in the
presence of herbivores and a natural enemy that can perceive volatile signals. Such studies would also allow
for the measurement of volatile concentration gradients around attacked ramets, and the assessment of the
foraging precision of attracted enemies.
The beneficial effects of a massive emission of volatiles from interconnected ramets of clonal plants may
also depend on whether or not neighboring plants are able to perceive and respond to these info-chemicals
(Bruin and Dicke 2001). Benefits to neighbors can arise from the direct perception of info-chemicals, or they
may stem from an increased protection against herbivores by attracted carnivores or parasitoids. The net effect
of information sharing with unconnected (and possibly unrelated) neighbors can be positive (through increased
protection of a larger area) or negative (through increased performance and competitive strength of the
neighbors). In other words eavesdropping (in the sense of an activation of defensive phenotypes after being
exposed to info-chemicals produced by other damaged plants; Karban et al. 2000; Dolch and Tscharntke 2000;
Karban and Baldwin 1997) as well as mechanisms of group selection could favor or disfavor the massive
emission of defense signals from clonal plant networks, depending on the competitive relationship and spatial
arrangement of clonal fragments, genets and other species in natural populations and communities. These
unresolved questions call for specific studies into the proximate mechanisms and ultimate implications of multi-
trophic interactions between clonal plants, herbivores and their enemies.
It should be noted, that the information content, reliability and specificity of gaseous info-chemicals
released to the air by damaged plants or ramets is probably lower than that of internally transmitted direct
defense signals. The effectiveness of an alerting system based on volatile emissions is hence likely to be lower
than the early warning system based on the systemic induction of direct defense traits in interconnected ramets
of clonal fragments. In most of the studies reporting plant-plant communication, the activation is thought to be
mediated by airborne info-chemicals (Karban and Baldwin 1997 and studies therein, Dicke and Bruin 2001 and

Chapter 2
26
A
B
studies therein; Karban et al. 2003) and also in some cases by root-exudates (Chamberlain et al. 2001, Dicke
and Dijkman 2001, Guerrieri et al. 2002). However, in these cases of “external warning” the distance between
infested and uninfested plants should be small in order to perceive the warning signal (both via air and soil)
and activate their defensive phenotype. Normally in these studies the distance between infested and
undamaged plants is about 15 cm or less (e.g. 15 cm for volatiles perception in Karban et al. 2000, less than
15 cm for root exudates perception in Chamberlain et al. 2001 and Guerrieri et al. 2002). Another factor that
could interfere in the perception of the volatiles emitted by infested plants is the direction of the wind. In clonal
networks, the members would be able to overcome these constraints by the systemic transmission of the
warning signal through physiological integration between ramets.
SYSTEMIC PATHOGENS: THE DARK SIDE OF NETWORK INTEGRATION?In spite of all obvious advantages, resource and information sharing has its risks. Modern, man-made
information networks are well known for their vulnerability to viruses and worms. In direct analogy, populations
of interconnected ramets of clonal plants may be especially susceptible to infections by systemic diseases, as
physical links between clone members can be (ab-)used as internal dispersal highways, enabling pathogens to
spread among connected ramets and to disperse rapidly within clonal ramet populations (Cook 1985; Eriksson
and Jerling 1990). This risk might create selection pressures against communication and resource integration
in clonal plants, and/or it may prompt a co-evolutionary arms race between networks and intruders (such as
seen in electronic information networks). Although interconnected ramets of clonal plants can be functionally
independent, they may not have independent risks of pathogen infection. After an initial infection, pathogens
can trace uninfected ramets by following vascular connections through the use of the plant internal transport
system for water and carbohydrates. Specialist pathogens may particularly benefit from internal spread,
because they can successfully locate and infect genetically identical hosts without the help of external vectors.
From this perspective, clonal plants seem ideal hosts for systemic pathogens. In non-clonal plants, infections
by internal disease transmission are restricted to one individual (Figure 2).
Figure 2. Systemic spread of a pathogen in clonal and non- clonal plants. In clonal plants (A) the pathogen can spread through many potential individuals. The spread of a pathogen in a non- clonal plant (B) is restricted to one individual. Black filling indicates the infected plant parts.

Non-resource sharing in clonal plant networks
27
Pathogen effects on host plants are extremely diverse, ranging from lethal or severely damaging effects,
to symptom-less infections, and positive impacts on plant growth and performance. A wide range of
qualitatively and quantitatively different pathogen actions is described in the literature: many pathogens cause
a decrease in photosynthetic activity (Chia and He 1999, Funayama and Terashima 1999, Sampol et al. 2003),
while others affect hormone levels of their hosts (Pan and Clay 2002) or alter resource allocation patterns to
different organs and functions (García-Guzmán and Burdon 1997). Pathogen infection may lead to a
deformation of leaves (Gibbs and Harrison 1976), reduced growth rates (Jones 1992, Piqueras 1999, Potter
1993) and changes in the growth form of plants (Piqueras 1999, Wennström and Ericson 1992). In some
cases, pathogens can effectively castrate (impede sexual reproduction) the host plant (García-Guzmán and
Burdon 1997, Groppe et al. 1999, Pan and Clay 2002). In rather rare cases, host plants may actually benefit
from pathogen infections in terms of increased biomass production (Pan and Clay 2002; Groppe et al. 1999) or
enhanced levels of allelopathy (Mattner and Parbery 2001).
However, plants are by no means defenseless against pathogens. On the contrary, they have evolved
an impressive array of mechanisms and strategies to tolerate, avoid or fight pathogens. In the following
sections, we will focus on systemic pathogen spread and various defense mechanisms that (clonal) host plants
may exhibit in response to disease infection and pathogen spread. We will concentrate on possible ecological
implications of clonality in relation to systemic pathogens.
A) RACE AGAINST TIME: SYSTEMIC SPREAD OF PATHOGENS AND DEFENSE SIGNALS
Pathogens can be either systemic or non-systemic. Systemic pathogens are able to move away from the initial
site of infection and can contaminate other parts of the plant. In most cases they live perennially in the host
plant. In contrast, non-systemic pathogens are restricted to the initial site of infection. They are often annuals
that re-infect their host plants every year (Wennström 1999). In the following sections, we will focus exclusively
on systemic pathogens, which can spread through the vascular system of their host plants. Fungi, for example,
may grow along vascular vessels or sporulate directly into the xylem sap. Viruses can be transported in the
phloem. They can directly be entered into the vascular system by feeding aphids or other animal vectors, or
they can use plasmodesmata to enter and exit the phloem. Once inside the phloem, viruses usually follow
plant-internal source-sink flows (Cheng et al. 2000; Leisner and Turgeon 1993; Thompson and Schulz 1999),
thereby predominantly ending up at sites with high sink strengths (e.g. young, developing plant parts, resource
deficient and damaged plant parts or ramets). In analogy, pathogens and pathogen propagules present in the
xylem sap of plants are likely to move along water potential gradients from sites of water uptake to sites of high
water loss through transpiration (Marshall 1990; Stuefer 1996).
Plants have developed different defense strategies to cope with pathogens. Pathogen defense
mechanisms can act on morphological, developmental and biochemical levels of the plant. An infected host
plant may, for instance, escape the pathogen by a (partial or full) developmental switch from vegetative growth
to flowering. Korves and Bergelson (2003) have recently shown that the time to flowering can be significantly
shortened by pathogen infection in Arabidopsis. Several studies have shown that clonal plants may be able to

Chapter 2
28
escape their systemic pathogens by fast vegetative growth (Frantzen 1994, García-Guzmán and Burdon 1997,
Wennström and Ericson 1992) and by clone fragmentation (McCrea and Abrahamson 1985; Kelly 1995).
It is still unknown however, whether the timing and the extent of ramet isolation through clone
fragmentation is a pathogen-inducible trait in clonal plant species. It is known for several groups of clonal
plants (e.g. pseudo-annuals, Jerling 1988; tussock-forming grasses, Wilhalm 1995) that the connections
between ramets are short-lived and that clones fragment into individual ramets or small ramet groups as part of
their regular development. These so-called genet splitters (as opposed to the group of genet integrators)
exhibit spontaneous clonal fragmentation. Their ramets do not stay interconnected for extended periods of
time, but become physiologically and physically independent after a short offspring production and establish-
ment phase (Eriksson and Jerling 1990; Piqueras and Klimes 1998; Piqueras 1999; Verburg and During 1998).
This seems counterintuitive because of the apparently low costs of maintaining connections and the broad
evidence for positive effects of clonal integration on plant performance and fitness. It has been suggested that
genet splitters may give up physical ramet connections to spread the risk of mortality (e.g. generated for
instance by pathogens; Eriksson and Jerling 1990) among independent ramets. According to this hypothesis,
genet splitters could have lost their ability for prolonged integration due to strong past and/or current selection
pressures created by systemic pathogens. This idea proved difficult to verify as most clonal species are either
obligate splitters or integrators, and genetic variation for clone fragmentation is usually small or absent in most
clonal species. Direct experimentation with pathogens, intact and artificially severed clonal fragments may
provide more insight into this topic.
Plants can also defend themselves against pathogens by a mechanism known as post-transcriptional
gene silencing (PTGS). PTGS is an effective defense mechanism targeted specifically at viruses, which
protects plant cells by degrading the nucleic acid of RNA viruses (Waterhouse et al. 2001). PTGS can spread
through the plant by an unknown signal that is capable of traveling both between cells (through
plasmodesmata) and through the phloem. Infected plant cells can use this system to send a warning message
to uninfected parts of the plant. These parts can then prepare their virus degradation machinery, in order to
stop the infection (Waterhouse et al. 2001 and references therein). This mechanism is induced whenever a
pathogen carrying an avirulence (Avr) gene challenges a host plant with the matching resistance (R) gene. A
so-called hypersensitive response is usually induced after infection. This process is mediated by salicylic acid
(SA) and is commonly referred to as systemic acquired resistance (SAR; Gozzo 2003). Through this
mechanism, the host plant may temporarily exhibit a stronger resistance to following challenges by the same or
in some cases also by other pathogens. SAR is active against viruses, bacteria and fungi (Conrath et al. 2002;
Gozzo 2003; Maleck and Dietrich 2003).
PTGS and SAR can systemically protect plants against invading pathogens. Both defense mechanisms
may be of considerable importance in clonal plant networks, because they can internally spread to many (or all)
functional individuals on a clonal fragment. In analogy to the early warning system against herbivores (see
above) PTGS and SAR may be an effective means to save connected sibling ramets from getting infected. The
costs-benefit balance of PTGS and SAR will most likely depend on the effectiveness of protection against
further pathogen damage, which is in turn a function of the relative speed with which pathogens and pathogen

Non-resource sharing in clonal plant networks
29
defense signals can travel within clonal plants.
The spatio-temporal extent and the dynamics of internal spread of systemic pathogens and appropriate
defense mechanisms (SAR, PTGS, fragmentation) in clonal plants are largely unknown. Both the speed and
the spatial extent of this spread may be system-specific and depend strongly on environmental conditions and
on source-sink relationships between connected ramets. The outcome of the race between pathogens and
specific defense signals may vary according to circumstances at the time and place of infection. In the absence
of experimental data, any prediction as to whether systemic pathogens or systemic defense signals may win
this race against time seems futile and excessively speculative. However, if pathogens could on average
spread faster than the defense mechanism, clonality would be a major disadvantage for plants, and systemic
pathogens should exert selection pressures against the prolonged maintenance of physical inter-ramet
connections. If, on the other hand, the defense mechanisms were usually effective, clonal growth and
physiological integration can be a great benefit for plants, because it allows for an effective protection of
spatially scattered, yet functionally independent individuals. In non-clonal plants systemic defense mechanisms
are restricted to a single individual (Figure 3).
Figure 3. Race between the pathogen and the defense signals, in clonal and non-clonal plants. After infection by a pathogen (indicated with the left black arrows), PTGS and/ or SAR are induced. If these defense mechanism (indicated by the grey cross) can be establishedfaster than the infection, the defense signals will spread through many potential individual in clonal plants (A), thereby protecting them from further infections. Whereas in non- clonal plants (B) only one individual is protected by these defense mechanisms and other individuals are still susceptible to infection. On the other hand if the pathogen wins, many potential individuals are infected in clonal plants (A1), whereas only one individual is infected in non- clonal plants (B1). Black filling indicates the infected plant parts; grey filling indicates the plant parts protected by SAR and/ or PTGS. Arrows indicate (possible) infection sites.
B) CLONAL PLANTS: STORES AND VECTORS FOR DISEASES?The presence of systemic pathogens in clonal plants may have serious repercussions on the population and
community level, because clonal plant networks could serve as vectors for diseases and provide ideal long-
term storage space for pathogens. The survival chances and the longevity of clonal plant genets are generally
expected to be higher than those of non-clonal plants. Clonal plants can persist as long as the rate of clonal
proliferation by initiating new meristems is higher or equal than the rate at which old plant parts die off (Thomas
A A1
B B1

Chapter 2
30
2002). Clonal plants can circumvent senescence and avoid the developmentally programmed death of the
genetic individual by repeated rejuvenation from newly activated meristems (i.e. by spontaneous self-cloning).
Therefore, clonal genets can be extremely long-lived (Oinonen 1967; Kemperman and Barnes 1976; Cook
1985; Steinger et al. 1996). Systemic pathogens that can persist in plants during the whole lifetime of the host
may be preserved in clonal plants for very long, potentially endless periods of time. Specialist pathogens
(which can only infect few species), could especially benefit from storage in clonal plants, since the need to
transmit to new, maybe difficult to find, hosts diminishes. In general the availability of suitable hosts can be
expected to be higher for generalist pathogens, therefore storage in clonal plants may be less necessary for
these pathogens. Non-clonal plants are less likely to play such a long-term storage role, because their lifetime
is generally more restricted than that of clonal plant individuals.
Clonal plants could function as spatial vectors for pathogens in natural plant populations and
communities. The spread of systemic diseases within populations and communities may be significantly
facilitated by the presence of clonal plant networks, because they allow pathogens to move between plants in
the absence of suitable external vectors, and without the production of specialized dispersal units. A pathogen
that can persist in a clonal plant can use its host as a long-term basis and spatial vector to spread to other
plants within the system. Non-clonal plants are more likely to die of senescence before the pathogen can
spread to other plants. From this perspective, clonal plant networks may represent spatio-temporal stepping-
stones facilitating the spread of systemic diseases within populations and communities. Specialist as well as
generalist pathogens are likely to use clonal plants as vectors and storage medium. However, generalist
pathogens probably use clonal plants in particular as spatial vectors. Because they can spread to many host
species, clonal plants may provide a perfect starting-point for generalist pathogens to (re-) infect surrounding
plants. The presence of generalist pathogens in long-lived, spatially extensive clonal networks may pose a
greater threat to neighboring plants because they are more likely to infect individuals of other genets and
species in a plant community. Specialist pathogens may benefit predominantly from clonal plants as temporal
vectors. Clonal plants could give specialist pathogens the time to “wait” for suitable host species that may not
be present at all times. This notion of clonal plants as possible stores and vectors of diseases would predict
that, in the long run and under comparable environmental conditions, populations with a high frequency of
clonal plants might accumulate more resident pathogens and therefore suffer from higher disease loads than
populations with a lower presence of clonal species. Specific data to test this prediction are not currently
available.
High levels of virulence are likely to preclude systemic pathogens from using clonal host plants as long-
term storage space and spatio-temporal vectors. Highly virulent pathogens are likely to kill or seriously damage
the host plant in short periods of time, thereby creating the need to spread quickly between hosts (Lively 2001).
Highly virulent, systemic pathogens should be able to rapidly eradicate entire clonal networks. To be
ecologically and evolutionarily feasible, however, high levels of virulence must be coupled to very fast and
efficient between-plant dispersal (Day 2003), and is therefore expected to be more common for generalist than
for specialist pathogens. To date we do not have any compelling evidence for the existence or common
occurrence of highly virulent killer-pathogens in clonal plants. If they existed, they should cause very strong

Non-resource sharing in clonal plant networks
31
selection pressures against clonality. We suggest that clonal plant life histories selectively favor pathogens with
a low virulence. The benefits conferred to the pathogen by the potentially unrestricted lifetime of clonal host
plants might strongly select against high levels of pathogen virulence. This expectation is in concordance with
general dispersion-virulence models that predict a positive relation between transmission rates and pathogen
virulence (Day 2003; Lipsitch and Moxon 1997; Lively 2001).
In specific cases clonal plants can actually benefit from pathogen infections. Groppe et al. (1999) have
shown that the internal concentration of the endophytic fungus Epichloë bromicola is positively correlated with
the vegetative vigor of the clonal host plant Bromus erectus. Although infected plants showed a significant
increase in vegetative growth and performance, endophyte infection also had strongly negative impacts on
sexual reproduction. Seed output of the host plant was negatively correlated with fungal concentration. Pan
and Clay (2002) reported a similar pathogen-mediated trade-off between vegetative growth and sexual
reproduction in their system. Epichloë glyceria infections enhance stolon production and accumulation of clonal
growth biomass of the clonal host plant Glyceria striata, and at the same time E. glyceria effectively castrates
its host. By doing so, the fungal endophyte blocks the possible escape route for the host to dispose of the
pathogen by flowering and sexual reproduction. In terms of vegetative growth and competitive ability of the
host plant these fungi-plant associations can be considered mutualistic: the fungal endophyte enhances host
performance and the host plant provides a suitable environment for the pathogen. In terms of life-history
evolution of these pathogens, the lack of sexual reproduction of the host in combination with the virtual
absence of genet senescence in clonal plants removes the necessity to disperse after successfully infecting a
host. A very low pressure to disperse might eventually lead to the evolution of low virulence in these specific
clonal plant pathogens, possibly generating a basis for the evolution of (partially) mutualistic plant-pathogen
systems (Clay 1990).
CONCLUSION
Based on the information and arguments provided above, we conclude that sharing substances and agents
other than resources between ramets of clonal plants may have far-reaching consequences for the functioning
of plant individuals, populations and communities, as well as for interactions between clonal plants on one
hand and pathogens, herbivores and the natural enemies of herbivores on the other hand. We are currently
only at the beginning of research that will hopefully elucidate the various ecological roles, proximate
mechanisms and ultimate implications of clonal integration beyond resource sharing. Future studies may shed
light on these complex yet fascinating interactions.
Acknowledgements. We thank Hans de Kroon for comments on a previous version of the manuscript.

Chapter 2
32

S Gómez & JF Stuefer. Oecologia (2006)
CHA
PTER3
Members only: Induced systemic resistance
in a clonal plant network

Chapter 3
34
ABSTRACT
We used the stoloniferous herb Trifolium repens to study the expression of induced systemic resistance
(ISR) to the generalist caterpillar Spodoptera exigua in interconnected ramets of clonal fragments. ISR
was assessed as caterpillar preference in dual choice tests between control and systemically induced
plants. ISR was detected in young ramets, after inducing older sibling ramets on the same stolon by a
controlled herbivore attack. However, developmentally older ramets did not receive a defense induction
signal from younger ramets unless the predominant phloem flow was inverted by means of basal shading.
This provides evidence for the notion that in T. repens the clone-internal expression of ISR is coupled to
phloem transport and follows source-sink gradients. The inducibility of the genotypes was not linked to
their constitutive ability to produce cyanide, implying the absence of a trade-off between these two
defense traits. To our knowledge this is the first study that explores induced systemic resistance to
herbivory in the context of physiological integration in potentially extensive clonal plant networks.

Members only: Induced resistance in a clonal plant network
35
INTRODUCTION
Clonal plants form networks of interconnected, semi-autonomous and genetically identical individuals
(ramets) that are produced at the nodes of horizontal stems. Physiological integration is an important
feature of most clonal plants as it allows ramets to share resources like water, photo-assimilates and
nutrients following source-sink principles. The mechanisms and ecological implications of resource
translocation within clonal plants have been extensively described in the literature (Pitelka and Ashmun
1985; Marshall 1990; Alpert 1996; Alpert and Stuefer 1997).
However, much less is known about the intra-clonal transport of non-resource agents like defense
compounds, signaling molecules or pathogens (Stuefer et al. 2004), even though stolon and rhizome
connections are likely to provide excellent means for sharing such compounds among integrated
members of clonal plant networks. The systemic induction of resistance against herbivores (ISR) by
means of clonal integration may enable spatially scattered ramets to alert each other about the presence
of herbivores. Such an early warning system could allow undamaged members of a clonal fragment to
prepare themselves for impending herbivore attacks by up-regulating the expression of chemical and
other defense traits. The internal distribution of defense induction signals among interconnected ramets
might be a fast and efficient way of responding to local herbivore damage. Moreover, the high specificity
of a spatial alerting system based on clonal integration would also minimize the risk of eavesdropping by
neighboring competitors of the same and of other species. To date, however, no published information
exists about the possible role of physiological integration in the internal transmission of substances
involved in defense induction in clonal plants (Stuefer et al. 2004).
Between-ramet transport of signals or substances involved in the systemic expression of resistance
may take place in the phloem as shown for a number of non-clonal species such as Populus species
(Davis et al. 1991, Jones et al. 1993), Lycopersicon esculentum (Orians 2000), and Arabidopsis thaliana
(Chen et al. 2001). As a consequence, plant-internal featured (e.g., vascular architecture, source-sink
dynamics; Marshall 1990, Marshall and Price 1997) as well as environmental features (e.g., heterogeneity
in resource supply; Stuefer 1996) affecting the degree and pattern of phloem integration between ramets
are also likely to interfere with the systemic expression of induced defense traits. The large amount of
existing knowledge on inter-ramet resource sharing can thus be used to predict both the potential and
limitations of systemic expression of induced resistance in clonal plants.
Trifolium repens (White Clover) is one of the best-studied clonal herbs with respect to physiological
integration and the internal movement of photo-assimilates (Chapman et al. 1990a; Chapman et al.1992a,
Kemball et al. 1992; Kemball and Marshall 1995; Marshall and Price 1997; Hamilton and Hay 1998). In
addition, a considerable number of studies has been devoted to the interaction between T. repens and
different herbivores, many of them focusing on the fact that some genotypes of this species can produce
hydrogen cyanide (HCN) upon tissue damage (Dirzo and Harper 1982a; 1982b; Puustinen and Mutikainen
2001). Cyanogenesis in T. repens is a constitutive defense trait (Hayden and Parker 2002) that has been
shown to confer resistance to a number of leaf-chewing insects and mollusks (Pederson and Brink 1998).

Chapter 3
36
This feature provides the opportunity to test for possible trade-offs in the expression of constitutive and
inducible defenses (Herms and Mattson 1992; Karban and Baldwin 1997; Underwood et at. 2000) and
adds to the suitability of T. repens as a model system for the study of induced systemic resistance to
herbivory in clonal plant networks.
We specifically tested the following hypotheses: (1) Local damage by herbivores induces systemic
resistance in undamaged ramets of the same clonal fragment. (2) The signals or compounds involved in
the systemic induction of resistance are linked to photo-assimilate flows in the phloem of stolon
connections between ramets. (3) The systemic expression of the defense response within clonal
fragments is constrained by the directionality of predominant phloem flows, which is in turn a function of
plastic source-sink relationships. (4) Cyanogenesis has a negative effect on the level of systemic defense
induction.
MATERIALS AND METHODS
We used Trifolium repens (White Clover) and Spodoptera exigua (Beet Armyworm) as a model system for
all experiments.
The beet armyworm, Spodoptera exigua (Hübner), is a generalist caterpillar with a broad host
range. Egg batches were obtained from the Virology Laboratory at Wageningen University, The
Netherlands. The caterpillars were reared on the same artificial diet as described in Biere et al. (2004).
Trifolium repens L. is a perennial, stoloniferous herb, which can form extensive clonal fragments by
vegetative growth. Stolon-based transport of carbohydrates is potentially bi-directional in T. repens
(Marshall and Price 1997). In undisturbed plants grown in homogeneous environments the bulk phloem
flow is directed from developmentally older ramets to younger ramets. This acropetal flow of photo-
assimilates is driven by internal source-sink relationships between ramets with adult leaves (source sites)
on one hand and ramets with developing leaves and growing stolon apices (sink sites) on the other hand
(Marshall 1990; Kemball and Marshall 1995). In T. repens, factors like shading (Chapman et al. 1990a,
Kemball et al. 1992, Stuefer et al. 1996) or defoliation (Chapman et al. 1992a; 1992b) can strongly alter
and partially reverse the flow of photo-assimilates (i.e., induce basipetal phloem flows) by creating carbon
sinks in the shaded or defoliated parts of the clonal fragment.
Some genotypes of Trifolium repens are capable of producing hydrogen cyanide, depending on
their genetically determined ability to produce two indispensable precursors of HCN. This ability is
controlled by two independent loci termed Ac and Li (Till 1987). All genotypes used in these experiments
were screened for cyanogenesis using the Feigl-Anger test modified by Hayden and Parker (2002). This
test discriminates between four different cyanide classes according to the ability or inability of genotypes
to synthesize both, one or none of the two biochemical precursors of cyanide. Ten genotypes were used
in the experiments, five of them cyanogenic (AcLi) and five acyanogenic (acli).
While cyanogenesis is well studied for T. repens, almost nothing is known about the ecology,
physiology and biochemistry of induced resistance to herbivores in this species. To our knowledge, only

Members only: Induced resistance in a clonal plant network
37
one study has so far attempted to identify some compounds which may be involved in systemic cross-
resistance of T. repens seedlings to the blue-green aphid Acyrthosyphon kondoi (Kempster et al. 2002). It
is hence unknown what traits might confer inducible resistance to herbivores in T. repens and how these
traits are activated after herbivory.
The ten genotypes of T. repens used in the experiments (see below) were collected from semi-
natural pastures along the floodplains of the river Waal (The Netherlands). The experiments were
conducted during spring and summer of 2004 in the greenhouse complex of the Radboud University
Nijmegen, The Netherlands. We conducted two experiments to investigate the capability of herbivore-
damaged ramets to induce resistance in connected, undamaged sibling ramets and to investigate the
direction of the systemic defense induction within clonal fragments.
Throughout this paper the term induced systemic resistance (ISR) is used to describe the plant-
internal spread of compounds and/or signals which confer increased levels of herbivore resistance to
undamaged ramets after a local herbivore attack on connected ramets. The internal systemic spread of
resistance agents can be either partial or full, meaning that only a part of the clonal fragment or all ramets
of the clonal fragment up-regulate defense traits after local herbivore damage. The term clonal fragment
refers to an assemblage of ramets which are all inter-connected by stolon internodes.
Experiment 1: Acropetally Induced Systemic Resistance
In this experiment we tested for the presence or absence of induced systemic resistance in
developmentally younger ramets after exposing developmentally older ramets on the same clonal
fragment to controlled herbivore damage. Since the induction of defense was expected to move from older
(induced) to younger ramets (uninduced) in this set-up, we call the hypothesized phenomenon acropetal
expression of ISR. For each of the ten genotypes, we placed 14 apical cuttings, each consisting of 5 to 6
ramets, into individual pots (12cm x 8.5cm x 8.5cm) filled with sieved loamy sand and 3g/l Osmocote
Exact (The Scotts Company, The Netherlands) to avoid nutrient limitation. We randomly distributed the
pots of each genotype on greenhouse benches and grew the cuttings under homogenous conditions for
four weeks before the start of the experiment. The experiment was conducted in two consecutive time
blocks, each of them lasting for one week.
The induction consisted of a controlled herbivore attack on the experimental plants with early instar
caterpillars to induce a defense response in undamaged sibling ramets on the same clonal fragment. At
the start of the experiment each plant had several stolons. The treatment was carried out on one of the
stolons that had at least four fully developed, unrooted ramets. Two second instar caterpillars were
confined inside a plastic petri dish mounted on the two ramets situated basally to the youngest fully
developed ramet of the target stolon. Empty petri dishes were placed in the same manner on the control
plants (Figure 1a). The caterpillars were allowed to feed for six days to ensure the presence of
considerable damage on the leaves.

Chapter 3
38
Figure 1. Schematic representation of the experimental design to test for induced systemic resistance in (A) younger ramets and (B) mature ramets of Trifolium repens. The circles represent petri dishes where control (no caterpillars) and induction treatment (2caterpillars; represented by two gray wavy lines) were conducted. After the induction treatment on two ramets, a dual choice test(square box) was done between fully expanded ramets from the induced and the control plant. The arrows point the direction of growth of the target stolon. In figure C, the dotted box represents a shade treatment imposed to create a carbon sink at the base of the plant to reverse the phloem flow.
The petri dishes were 9 cm in diameter with an opening on the side to allow the non-destructive
insertion of petioles. The ramets remained attached to the plants during the treatment and subsequent
preference assay. The lid of the petri dish was pierced and covered with a fine mesh to avoid water
condensation and gas accumulation inside the petri dish.
After the induction period, we conducted in-situ dual choice tests to study the preference of S.
exigua caterpillars for systemically induced or uninduced leaves of the same genotype. A consistent
preference for the control leaf suggests the presence of a systemic response as a consequence of the
herbivory attack on a connected ramet. The choice tests were conducted between the youngest fully
expanded ramet on the induced and the uninduced control stolon, respectively. We placed both ramets
into a petri dish opposing each other and placed a third instar caterpillar in the middle (Figure 1a). The
ControlChoice
test
Induction
InductionControl
Choice test
InductionControl
Choice test
A
B
C

Members only: Induced resistance in a clonal plant network
39
caterpillar was allowed to feed for 24 hours. In the second time block, we used fourth instar caterpillars to
increase the amount of leaf area consumed. Due to the higher intake rate of fourth instar caterpillars, we
stopped the choice test when approximately 30% of one of the leaves was consumed to avoid obscuring
the preference by food shortage (see Akhtar and Isman 2004). After 24 hours all choice tests were
stopped irrespective of the amount of damage. Each choice test was replicated seven times for each
genotype. At the end of each choice test we took digital pictures of the leaves and the leaf area consumed
was measured with the image analysis software Image-Pro Plus, version 1.1 (Media Cybernetics, Silver
spring, Maryland, USA).
Experiment 2: Basipetally Induced Systemic Resistance
a) Full light
This experiment was conducted to test for the presence or absence of induced systemic resistance in
developmentally older ramets after herbivore damage had taken place in developmentally younger ramets
(basipetal expression of ISR). The experimental set-up was the same as in Experiment 1 (see description
above), but the locations of the induction treatment and choice test were inverted (Figure 1b). Thus, the
induction treatment was imposed on the youngest and second-youngest fully developed ramets of a
stolon and the in- situ dual choice test was conducted between the third-youngest fully developed ramets
on the induced and the control plants, respectively. Each choice test was replicated six times for each of
the ten genotypes. We used second instar caterpillars for the induction treatment and fourth instar
caterpillars for the choice test. The induction treatment lasted for five days.
b) Partial shade
To test whether the expression of induced systemic resistance is coupled to phloem flows, we imposed
partial shading on the basal ramets of the target stolon to create a carbon sink, thereby partially reversing
the phloem flow (Chapman et al. 1990a, Stuefer et al. 1996). The experimental set-up was similar to the
one described for experiment 2a with the addition of shading (Figure 1c). For each genotype we placed
six randomly selected pairs of plants under shade cages which covered the developmentally older parts of
the clonal fragment and which left the 4-6 terminal unrooted ramets of the target stolons unshaded. The
shade treatment lasted for 11 days. The shade cages consisted of metal frames covered by a fabric that
reduced light intensity by 75%. The induction treatment was imposed in ramets outside the shade cage
and the choice tests were done on ramets inside the shade cage. The plants remained under the shade
cages until the end of the experiment.
Statistical Analysis
The effect of systemic defense induction on caterpillar preference (i.e., leaf area consumed) within choice
tests was assessed with repeated measures ANOVA (SAS GLM procedure) considering each choice test
(petri dish) an independent subject. We used repeated reasures ANOVA to account for the fact that
measurements on leaves of the same choice test cannot be considered independent from each other
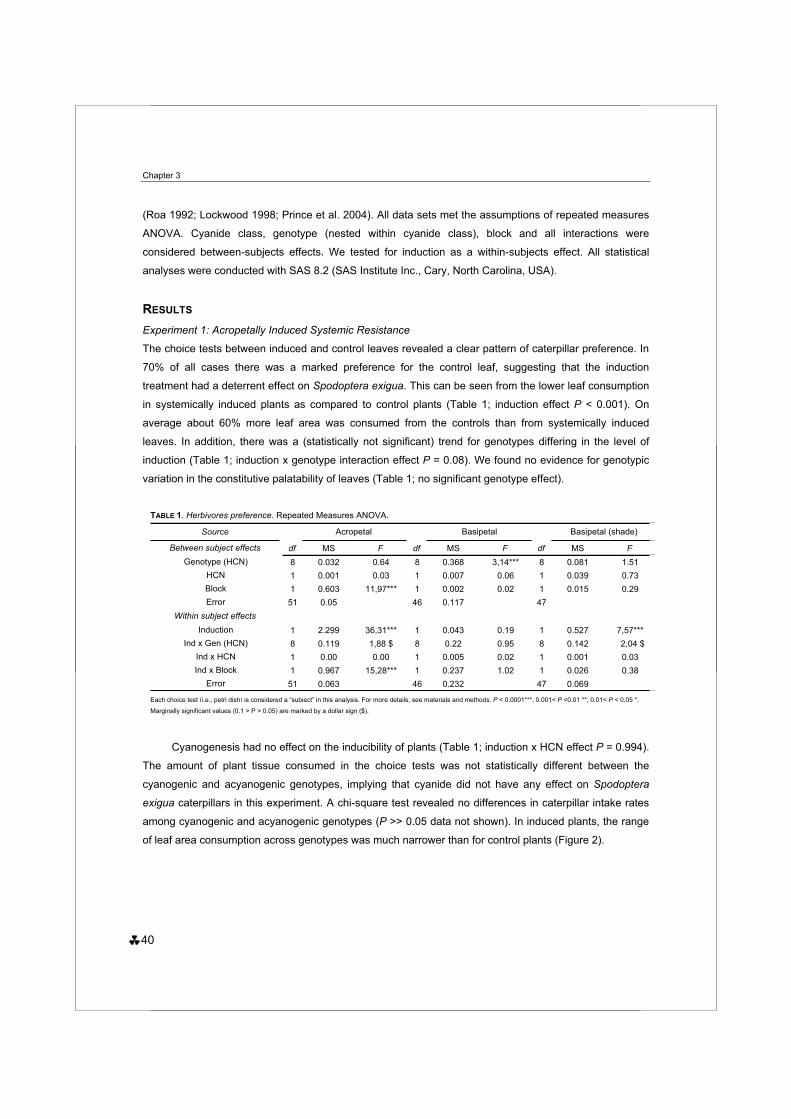
Chapter 3
40
(Roa 1992; Lockwood 1998; Prince et al. 2004). All data sets met the assumptions of repeated measures
ANOVA. Cyanide class, genotype (nested within cyanide class), block and all interactions were
considered between-subjects effects. We tested for induction as a within-subjects effect. All statistical
analyses were conducted with SAS 8.2 (SAS Institute Inc., Cary, North Carolina, USA).
RESULTS
Experiment 1: Acropetally Induced Systemic Resistance
The choice tests between induced and control leaves revealed a clear pattern of caterpillar preference. In
70% of all cases there was a marked preference for the control leaf, suggesting that the induction
treatment had a deterrent effect on Spodoptera exigua. This can be seen from the lower leaf consumption
in systemically induced plants as compared to control plants (Table 1; induction effect P < 0.001). On
average about 60% more leaf area was consumed from the controls than from systemically induced
leaves. In addition, there was a (statistically not significant) trend for genotypes differing in the level of
induction (Table 1; induction x genotype interaction effect P = 0.08). We found no evidence for genotypic
variation in the constitutive palatability of leaves (Table 1; no significant genotype effect).
Cyanogenesis had no effect on the inducibility of plants (Table 1; induction x HCN effect P = 0.994).
The amount of plant tissue consumed in the choice tests was not statistically different between the
cyanogenic and acyanogenic genotypes, implying that cyanide did not have any effect on Spodoptera
exigua caterpillars in this experiment. A chi-square test revealed no differences in caterpillar intake rates
among cyanogenic and acyanogenic genotypes (P >> 0.05 data not shown). In induced plants, the range
of leaf area consumption across genotypes was much narrower than for control plants (Figure 2).
TABLE 1. Herbivores preference. Repeated Measures ANOVA.
df MS F df MS F df MS F8 0.032 0.64 8 0.368 3,14*** 8 0.081 1.511 0.001 0.03 1 0.007 0.06 1 0.039 0.731 0.603 11,97*** 1 0.002 0.02 1 0.015 0.29
51 0.05 46 0.117 47
1 2.299 36,31*** 1 0.043 0.19 1 0.527 7,57***8 0.119 1,88 $ 8 0.22 0.95 8 0.142 2,04 $1 0.00 0.00 1 0.005 0.02 1 0.001 0.031 0.967 15,28*** 1 0.237 1.02 1 0.026 0.38
51 0.063 46 0.232 47 0.069
Basipetal (shade)
Between subject effectsGenotype (HCN)
Acropetal Basipetal
Ind x BlockError
Source
Within subject effectsInduction
Ind x Gen (HCN)Ind x HCN
HCNBlockError
Each choice test (i.e., petri dish) is considered a “subject” in this analysis. For more details, see materials and methods. P < 0.0001***, 0.001< P <0.01 **, 0.01< P < 0.05 *.
Marginally significant values (0.1 > P > 0.05) are marked by a dollar sign ($).

Members only: Induced resistance in a clonal plant network
41
1.2 0.8 0.4 0 0.4 0.8 1.2
A15
A23
A42
A25
A13
B11
C28
B42
D28
B36
Gen
otyp
es
Leaf area consumed (cm2)
Control Induction
HCN -HCN +
1.2 0.8 0.4 0 0.4 0.8 1.2
A15
A23
A42
A25
A13
B11
C28
B42
D28
B36
Gen
otyp
es
Leaf area consumed (cm2)
Control Induction
1.2 0.8 0.4 0 0.4 0.8 1.2
A15
A23
A42
A25
A13
B11
C28
B42
D28
B36
Gen
otyp
es
Leaf area consumed (cm2)
Control Induction
HCN -HCN +HCN -HCN +
Figure 2. Average area (and SE) consumed by caterpillars in the choice tests from control (gray) and systemically induced leaves (black) in 10 genotypes of T. repens. The choice test was done between the youngest fully expanded ramets of control and systemically induced plants. The induction was conducted on older ramets. The dash line separates acyanogenic genotypes (upper half) from cyanogenic ones (lower half).
Experiment 2: Basipetally Induced Systemic Resistance
In the experiment under full light conditions (2a), we found no evidence for systemic defense induction
(Table 1) in ramets situated basally to the point of controlled herbivore damage. The amount of leaf tissue
consumed from the control and the induced plants was not significantly different from each other (Figure
3), indicating that the choice of leaves by feeding caterpillars was due to a random decision. Genotypes
differed significantly with respect to leaf consumption by caterpillars (Table 1), suggesting genotypic
differences in leaf palatability.
When basal shading was imposed, however, we found a significant induction effect (Table 1; P =
0.008; Figure 4). The average leaf area consumed from the control leaves was considerably larger (about
30%) than that from the systemically induced leaves. The strength of induction was weaker than in
experiment 1 (acropetal expression of ISR). There was a non-significant trend (Table 1; induction x
genotype effect P = 0.08) indicating variation in the level of induction among genotypes. In accordance
with the previous experiment there was no effect of cyanide on inducibility and on the total amount of leaf
area ingested by the caterpillars.

Chapter 3
42
1.2 0.8 0.4 0 0.4 0.8 1.2
A15
A42
A13
A25
A23
C28
B11
B42
D28
B36
Gen
otyp
es
Leaf area consumed (cm2)
Control Induction
HCN -HCN +
1.2 0.8 0.4 0 0.4 0.8 1.2
A15
A42
A13
A25
A23
C28
B11
B42
D28
B36
Gen
otyp
es
Leaf area consumed (cm2)
Control Induction
1.2 0.8 0.4 0 0.4 0.8 1.2
A15
A42
A13
A25
A23
C28
B11
B42
D28
B36
Gen
otyp
es
Leaf area consumed (cm2)
Control Induction
HCN -HCN +HCN -HCN +
Figure 3. Average area (and SE) consumed by caterpillars in the choice tests from control (gray) and systemically induced leaves (black) in 10 genotypes of T. repens. The choice test was done between mature ramets of control and systemically induced plants.The induction was conducted on younger ramets. The dash line separates acyanogenic genotypes (upper half) from cyanogenic
ones (lower half).
Figure 4. Average area (and SE) consumed by caterpillars in the choice tests from control (gray) and systemically induced leaves (black) in 10 genotypes of T. repens. The choice test was done between mature ramets of control and systemically induced plantsunder a shade cage. The induction was conducted on younger ramets outside the shade cage. The dash line separates acyanogenic genotypes (upper half) from cyanogenic ones (lower half).
1.2 0.8 0.4 0 0.4 0.8 1.2
A42
A15
A25
A13
A23
B42
C28
B36
B11
D28
Gen
otyp
es
Leaf area consumed (cm2)
Control Induction
HCN -HCN +
1.2 0.8 0.4 0 0.4 0.8 1.2
A42
A15
A25
A13
A23
B42
C28
B36
B11
D28
Gen
otyp
es
Leaf area consumed (cm2)
Control Induction
1.2 0.8 0.4 0 0.4 0.8 1.2
A42
A15
A25
A13
A23
B42
C28
B36
B11
D28
Gen
otyp
es
Leaf area consumed (cm2)
Control Induction
HCN -HCN +HCN -HCN +

Members only: Induced resistance in a clonal plant network
43
DISCUSSION
The results of our experiments provide clear evidence that interconnected ramets of a clonal fragment are
able to systemically induce resistance upon herbivore attack. Our in situ, dual choice tests show a clear
reduction in herbivore preference for systemically induced ramets as compared to uninduced ramets,
indicating a plastic decrease in leaf palatability of induced plants. Moreover, our results strongly suggest a
functional coupling between dominant phloem flows and the spatial expression of ISR in clonal plant
networks. To our knowledge this is the first study to report that physiological integration of ramets allows
for, as well as, constrains the expression of systemic induced resistance to herbivores in interconnected
ramets of clonal plants.
The set up of our experiment ensured that the systemic induction effect was not caused by volatiles
or root exudates. The plants assigned to control and induction treatments were spatially randomized
without physical barriers between them. If defense induction was caused by airborne info-chemicals
consistent differences between control and induced plants, as shown in our experiments, would be very
unlikely to detect. Ramet induction through substances released by roots can also be excluded because
all ramets on the target stolons used for the induction treatment and choice tests were unrooted.
Our experiments strongly suggest a phloem-based, source-sink driven transport of the signal
responsible for the expression of induced systemic resistance in clonal fragments of T. repens. This notion
can be inferred from the presence of an otherwise absent (experiment 2a) basipetal induction effect after
creating a local photo-assimilate sink, which can be assumed to partially reverse the phloem transport
(experiment 2b). This basipetal induction effect, however, was weaker than the acropetal induction
(compare experiments 1 and 2b), which may be due to the fact that basal shading does not completely
invert phloem flows. A considerable portion of exported photo-assimilates by source leaves continues to
move acropetally to support strong sinks in the developing and apical region of stolons (Chapman et al.
1990a). The xylem transport was not manipulated in any of our experiments. Since only the older ramets
of our system were rooted (see Material and Methods), the mass flow of water in the xylem was directed
from the basal ramets (source sites for water) towards the unrooted ramets (sink sites for water) on the
stolon. This was also true for the shading treatment, implying that we can rule out xylem integration as a
transport pathway for defense induction signals in our species.
Studies on poplars (Populus spp.) have shown that assimilate movement controls the systemic
expression of wound-induced genes after mechanical damage (Davis et al. 1991) and that the expression
of systemically induced resistance can be predicted by plant vascular architecture (Jones et al. 1993)
which has recently also been shown for the bittersweet nightshade (Solanum dulcamara; Viswanathan
and Thaler 2004). Orians et al. (2000) also showed that vascular architecture in tomato plants explains at
least partly the systemic induction of proteinase inhibitors after mechanical damage.
If induced systemic resistance in T. repens relies on the spatial pattern and temporal dynamics of
predominant phloem flows, its expression will be shaped by plant-internal and environmental factors which
influence the bulk movement of photo-assimilates in the phloem. Under homogeneous environmental

Chapter 3
44
conditions, source-sinks patterns within clonal fragments are mainly a function of developmental
differences among ramets (Marshall 1990), implying that most phloem-based resources move from
developmentally older sink sites to developing source sites. On one hand, such a strong acropetal
transport of phloem-based compounds could be considered a constraint, as it is likely to hinder the
expression of inducible defense in clonal ramet populations. As shown in our experiments ramets located
basally to the herbivore-damaged leaves cannot be alerted of the risk of subsequent herbivore attacks. On
the other hand, however, a predominantly acropetal phloem flow is likely to contribute to a better
protection of young ramets, which are both the most valuable parts in terms of future growth and
(vegetative and sexual) reproduction of the clonal fragment, and they are often the most attractive and
most vulnerable targets for herbivores (Fenner et al. 1999). From this perspective, the disadvantages of
predominant acropetal phloem flows may be outweighed by the advantages of specifically protecting
valuable tissue.
Cyanide, although known for its herbivore deterrent properties, did not play any role as a defense
compound against Spodoptera exigua. No evidence was found to support the trade-off hypothesis
between cyanogenesis (i.e., the constitutive ability of genotypes to produce HCN) and inducible
resistance. Moreover, the level of induction was independent of the presence or absence of cyanogenesis
(Table 1; non-significant induction x HCN effect for all experiments), implying that we can rule out any
possible involvement of cyanide in the expression of induced resistance in T. repens.
Our experiments show that inducible resistance in Trifolium repens serves as an effective defense
component against this generalist herbivore. There was a marginal but consistent trend (experiment 1 and
experiment 2.b) showing genotypic variability in the magnitude of the induced response against
Spodoptera exigua. This variability could be due to genotypic differences in vascular architecture between
genotypes (Lotscher and Hay 1996), which is known to play an important role in the translocation of
resources (Arnold et al. 2004) and systemic signals (Schittko and Baldwin 2003). In the absence of
induced systemic resistance (experiment 2a) genotypic differences in plant tissue palatability (see
genotype effects in ANOVA tables) appeared to be much stronger than when systemic induction took
place (experiments 1 and 2b). This suggests that defense induction may at least partly override
differences between genotypes in constitutive leaf palatability.
Our study provides clear evidence for the notion that stolons of clonal herbs can serve as pathways
for the transmission of systemic defense induction signals in physiologically integrated clonal plant
species. They confirm the largely neglected idea that clonal integration may not only allow for an efficient
transport of resources such as water, mineral nutrients and carbohydrates, but that it can also operate as
an information sharing system among members of clonal plant networks (Stuefer et al. 2004). Since
ramets of clonal plants are functionally semi- or fully autonomous individuals, this can be seen as a
special case of plant-plant communication where the sender(s) and the receiver(s) are physically
connected to each other. Transport of systemic defense induction signals through stolons or rhizomes of
clonal plants may be an effective way to pass on information to a specific group of receivers (i.e., all
network members) and to avoid the interception and exploitation of information by (opportunistic)

Members only: Induced resistance in a clonal plant network
45
neighbors (eavesdropping), as reported for some systems where the signal is transmitted externally
(Dolch and Tscharntke 2000, Dicke and Bruin 2001, Chamberlain et al. 2001; Guerrieri et al. 2002). In
populations of non-clonal plants, undamaged individuals can only be induced via external pathways such
as the emission of volatile info-chemicals or the release of root exudates, which might result in a
competitive disadvantage with respect to clonal ramet populations in environments with a high incidence
of herbivore damage.
To get a better insight into the ecological and evolutionary implications of ISR in clonal plants, future
studies should explicitly take into account the spatio-temporal scale at which induced systemic resistance
is expressed within clonal plant networks. The spatial alert function of ISR may be most beneficial for
clonal fragments interacting with relatively small, moderately mobile herbivores. In such a system a small
initial damage is a good predictor of a more wide-spread attack at the spatial scale of entire clonal
fragments, and at the temporal scale which allows systemic defense induction to take place (Karban et al.
1999, Järemo et al. 1999; Stuefer et al. 2004). In other cases between-ramet ISR should be ineffective
and costly. Mismatches between the scale and dynamics of herbivore damage and the induced responses
are likely to result in a misallocation of resources that are necessary to up-regulate defense traits in
undamaged and unthreatened ramets. The spatio-temporal dynamics of damage and induced responses
seem hence the key element determining the costs and benefits, and the potential adaptive nature of
information sharing through physiological integration among interconnected ramets in response to
herbivore attacks.
Acknowledgements: We are especially grateful to Y. Verhulst and S. van Nieuwkerk for practical
assistance and J. Reudler Talsma for supplying Spodoptera exigua cocoons. We thank T. van Mölken, E.
Visser, H. Huber, H. de Kroon and two anonymous reviewers for useful comments on a previous version
of this manuscript.

Chapter 3
46

S Gómez, W van Dijk & JF Stuefer. Submitted.
CHA
PTER4
Time is precious: Dynamics of induced resistance
in a clonal plant network

Chapter 4
48
ABSTRACT
After local herbivory, plants can activate defense traits both on the damaged site and in undamaged plant
parts such as connected ramets in clonal plants. The time lag between the first herbivore attack and
defense activation is important to ensure an efficient protection of the plant. Since defense induction has
costs, a mismatch in time and space between defense activation and herbivore feeding might result in
negative consequences for plant fitness. In this study we quantified the timing of the induced responses in
ramet networks of the stoloniferous herb Trifolium repens after local herbivory. We studied the activation
time of systemic defense induction in undamaged ramets and the decay time of the response after local
attack. Undamaged ramets became defense-induced 1.5 to 2 days after the initial attack. Defense
induction was measured as a reduction in leaf palatability. Defense induction lasted at least 28 days, and
there was strong genotypic variation in the duration of this response. Ramets which were formed after the
initial attack were also defense-induced, implying that induced defense can be extended to new ramet
generations, thereby contributing to the protection of plant tissue which is both very vulnerable to
herbivores and most valuable in terms of future plant growth and fitness.

Timing of induced resistance in a clonal plant network
49
INTRODUCTION
Plant defense against herbivores is a costly process (Gershenzon 1994; Sagers and Coley 1995; Elle et
al. 1999). Defense costs can be a direct consequence of diverting resources from growth and
reproduction, and they can indirectly stem from ecological phenomena such as reduced competitive
ability, auto-toxicity and possible decreases in pollinator visits (Strauss et al. 2002; Heil and Baldwin
2002). Inducible defenses may have evolved to reduce the costs of constitutive defense expression by
reducing the time during which defense is active (Herms and Mattson 1992; Karban and Baldwin 1997;
Agrawal et al. 1999c).
After local herbivore damage many plants increase the production of defensive compounds to deter
or discourage attackers from continued feeding. Defense levels show bell-shaped intensity curves over
time. After an initial build-up phase, inducible defense levels reach a peak and then gradually decrease to
base levels (Schultz 1988; but see Underwood 1998). The duration of the different phases determines the
effectiveness of induced defenses against herbivores. There are two key aspects to consider in the study
of temporal dynamics of systemically induced responses. First, the time it takes for the defense signal to
leave the damaged site and activate defense mechanisms in undamaged parts of the plant. This period is
known as the systemic activation time. Second, the decay time of the induced response, which indicates
how long plastically enhanced defense traits are expressed in the plant.
In order to optimize resource investment in induced defense and reduce allocation costs, the
temporal expression of defense should match the average time span of transitory herbivore attacks. This
can only be achieved by fine-tuning the periods of defense activation and decay. The plastic induction of
defense is associated with a time lag between information acquisition (i.e. initial herbivore damage) and
plastic plant response (i.e. activation of defense), potentially resulting in an ineffective protection of the
plant under attack (DeWitt et al. 1998). Depending on the study system, this time lag can range from
hours (Baldwin et al. 1994; Alborn et al. 1996) and days (Underwood 1998; McAuslane et al. 1997; Agrell
et al. 2003) to whole seasons (Zvereva et al. 1997). Nevertheless, initial damage is usually a reliable cue
for future herbivory risks (Karban and Adler 1996; Karban et al. 1999), and can hence be used as a
predictive cue for subsequent damage. Fast activation and decay of induced defenses might be directed
against current herbivores, while longer response times are likely to affect future herbivory (Karban and
Baldwin 1997).
Clonal plant networks are formed by assemblages of interconnected individuals (ramets) that share
vascular connections through which resources and defense signals can be transported (Stuefer et al.
2004). In the stoloniferous herb Trifolium repens, defense is systemically induced after local herbivore
attacks (Gómez and Stuefer 2006). This response is based on the internal transmission of unknown
signals through stolon connections between members of the ramet network (Stuefer et al. 2004). In spite
of obvious benefits of having private channels for information transfer, the systemic induction of resistance
in a clonal plant network can also have negative consequences. Considerable costs of defense induction

Chapter 4
50
may be incurred if defense activation within the network does not match the spatio-temporal patterns of
herbivore attack (Gómez et al. 2007).
We aimed at studying the temporal aspects of plastic defense expression between members of
clonal plant networks of Trifolium repens, thereby addressing the following research questions: (a) how
long does it take for a ramet to become systemically induced after local attack in an adjacent ramet
(systemic activation time), and (b) how long does a clonal network remain defense-induced (decay time).
MATERIALS AND METHODS
Study system
We used four genotypes of the stoloniferous herb Trifolium repens (white clover), which had previously
been collected from natural populations growing on the flood plains of the river Waal, The Netherlands.
The same genotypes (A13, A23, B11 and D28) were previously used in other studies (Gómez and Stuefer
2006; Gómez et al. 2007). Larvae of the generalist moth Mamestra brassicae (cabbage army moth) were
used as herbivores. Egg batches were obtained from the Entomology Laboratory at Wageningen
University, the Netherlands. The colony was reared on cabbage and maintained at 23 °C and 16 h/ 8 h
photoperiod.
Pre-growth conditions
We made apical cuttings from each genotype, consisting of three unrooted ramets on a single stolon. The
cuttings were placed in trays with wet potting soil and covered with a plastic foil during five days to
promote rooting. The rooted cuttings were then individually transplanted into plastic trays (16cm x 12cm x
5cm) with 1 sand: 3 potting soil substrate. 20μl of a solution containing rhizobium was added to each tray
to promote nodulation. The experiment started one week after the cuttings had been transplanted.
Defense induction
To systemically induce defense, we performed controlled herbivory attacks on the 2nd and 3rd youngest
ramets on the main stolon of each cutting (Figure 1). This was achieved by confining either two 4-weeks
old (experiment 1) or three 1-week old caterpillars (experiment 2) in a Petri dish mounted on the plant.
The larvae were starved for the night preceding the start of the experiment to promote immediate
consumption of leaf tissue. An empty Petri dish was placed on control plants (Gómez and Stuefer 2006).
Choice tests
We used dual-choice tests as a tool to measure the presence or absence of systemically induced defense
in undamaged ramets (defense-induction treatment). For each choice test we collected the youngest fully
expanded leaf of the control and the corresponding defense-induced plant, respectively. Both leaves were
placed on moist filter paper in a Petri dish. A 3-4 weeks old caterpillar was released in the middle of the
Petri dish and allowed to feed until at least 30% of one of the leaves was consumed or for a maximum

Timing of induced resistance in a clonal plant network
51
period of 48 hours. At the end of each choice test, digital pictures were taken and the remaining leaf area
was measured using the image analysis software Image Pro Plus, version 1.1 (Media Cybernetics,
Bethesda, MD, USA). Regardless of the compounds and processes involved in the induction of
resistance, a reduced herbivore preference for induced as compared to uninduced plants can be seen as
a sign of induction of resistance (sensu Karban and Baldwin 1997), defined as a decrease in the
herbivore’s preference or performance feeding on a plant that has been previously damaged.
T. repens can show strong sectoriality (Lötscher and Hay 1996; Marshall and Price 1997), which
results in limited vascular contact between ramets belonging to different orthostichies on the stolon. For
this reason all choice tests were carried out using the youngest fully expanded leaves from the same
orthostichy as the most heavily damaged ramet used for defense-induction. A comparable aged leaf was
selected from the control plant.
Systemic activation time
To determine the window of time during which the induction response was activated in undamaged
ramets, we carried out three herbivory treatments differing in the duration of the attack (24, 38 and 51
hours, respectively). Twenty apical cuttings (i.e., 4-5 interconnected ramets) of each genotype were used
per herbivory treatments. Half of the replicates were randomly assigned to the control and defense-
induction group, respectively. At the end of each treatment, we performed ten dual choice tests as
Choice test 7 d
Choice test 28 d
Choice test 14 d
Control
Defense induction
Systemic activation time Decay time
Choice test Choice test 7 d
Choice test 28 d
Choice test 14 d
ControlControl
Defense inductionDefense induction
Systemic activation time Decay time
Choice test
FIGURE 1. Schematic representation of the experimental set-up. Pairs of plants assigned to control (white) and defense-induction treatments (gray) were used to study (a) the systemic activation time (left panel) of defense after local damage (defense induction) and (b) the decay time (right panel) of the defense expression after 7, 14, and 28 days, respectively. Dashed lines represent ramets that were not present at the time of the defense-induction treatment.
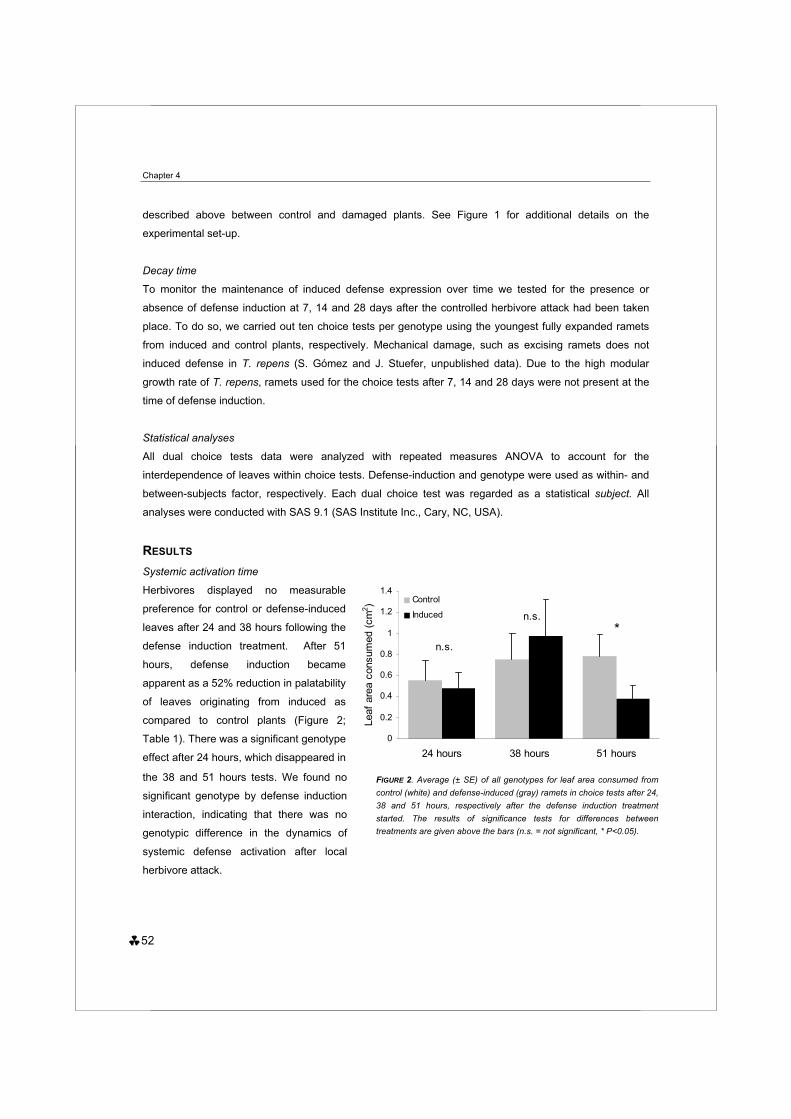
Chapter 4
52
described above between control and damaged plants. See Figure 1 for additional details on the
experimental set-up.
Decay time
To monitor the maintenance of induced defense expression over time we tested for the presence or
absence of defense induction at 7, 14 and 28 days after the controlled herbivore attack had been taken
place. To do so, we carried out ten choice tests per genotype using the youngest fully expanded ramets
from induced and control plants, respectively. Mechanical damage, such as excising ramets does not
induced defense in T. repens (S. Gómez and J. Stuefer, unpublished data). Due to the high modular
growth rate of T. repens, ramets used for the choice tests after 7, 14 and 28 days were not present at the
time of defense induction.
Statistical analyses
All dual choice tests data were analyzed with repeated measures ANOVA to account for the
interdependence of leaves within choice tests. Defense-induction and genotype were used as within- and
between-subjects factor, respectively. Each dual choice test was regarded as a statistical subject. All
analyses were conducted with SAS 9.1 (SAS Institute Inc., Cary, NC, USA).
RESULTS
Systemic activation time
Herbivores displayed no measurable
preference for control or defense-induced
leaves after 24 and 38 hours following the
defense induction treatment. After 51
hours, defense induction became
apparent as a 52% reduction in palatability
of leaves originating from induced as
compared to control plants (Figure 2;
Table 1). There was a significant genotype
effect after 24 hours, which disappeared in
the 38 and 51 hours tests. We found no
significant genotype by defense induction
interaction, indicating that there was no
genotypic difference in the dynamics of
systemic defense activation after local
herbivore attack.
FIGURE 2. Average (± SE) of all genotypes for leaf area consumed from control (white) and defense-induced (gray) ramets in choice tests after 24, 38 and 51 hours, respectively after the defense induction treatment started. The results of significance tests for differences between treatments are given above the bars (n.s. = not significant, * P<0.05).
0
0.2
0.4
0.6
0.8
1
1.2
1.4
24 hours 38 hours 51 hours
Leaf
are
a co
nsum
ed (c
m2 ) Control
Induced
* n.s.
n.s.

Timing of induced resistance in a clonal plant network
53
df MS F MS F MS F3 1.84 4.69** 1.32 0.88 0.39 1.63
36 0.39 1.50(35) 0.24
1 0.11 0.47 0.95 1.76 3.21 6.26*3 0.13 0.52 0.65 1.2 0.92 1.79
36 0.24 0.54(35) 0.51Error
Source
Between subjects effectsGenotype (Gen)
Within subjects effects
Ind x Gen
Error
Induction (Ind)
24 hours 38 hours 51 hours
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
7 days 14 days 28 days
Leaf
are
a co
nsum
ed (c
m2 ) Control
Induced
*** ** **
FIGURE 3. Average (± SE) leaf area consumed from control (white) and defense-induced (gray) ramets in choice tests after 7, 14 and 28 days, respectively after the defense induction treatment stopped. Asterisks indicate significant differences between defense-induction and control leaves (** P<0.01, P<0.001). After 28 days, there was a significant genotype x defense-induction effect (table 2).
Decay time
Our dual choice tests indicated that
defense induction persisted for at
least 28 days (Table 2) in one of the
genotypes. However, the strength of
the defense expression decreased
with time. Seven days after the end of
the defense induction treatment,
control ramets suffered 80% more
damage than defense-induced ramets
(Figure 3). After 28 days, the overall
effect of induction was still significant,
but the difference in damage between
control and induced ramets was
reduced to 50%.
Values in brackets indicate degrees of freedom. Statistically significant effects are marked with asterisks. 0.001< P <0.01 **, 0.01< P
< 0.05 *
TABLE 1. Induction time. Repeated measures ANOVA.
df MS F MS F MS F3 0.22 1.69 0.30 2.17 0.10 0.69
36 0.13 0.14 0.14
1 5.05 28.38*** 1.70 8.84** 2.78 9.75**3 0.12 0.68 0.29 1.5 1.17 4.11*
36 0.18 0.19 0.29Error
Source
Between subjects effectsGenotype (Gen)
Within subjects effects
Ind x Gen
Error
Induction (Ind)
7 days 14 days 28 days
Statistically significant effects are marked with asterisks. P < 0.0001***, 0.001< P <0.01 **, 0.01< P < 0.05 *
TABLE 2. Decay time. Repeated measures ANOVA.
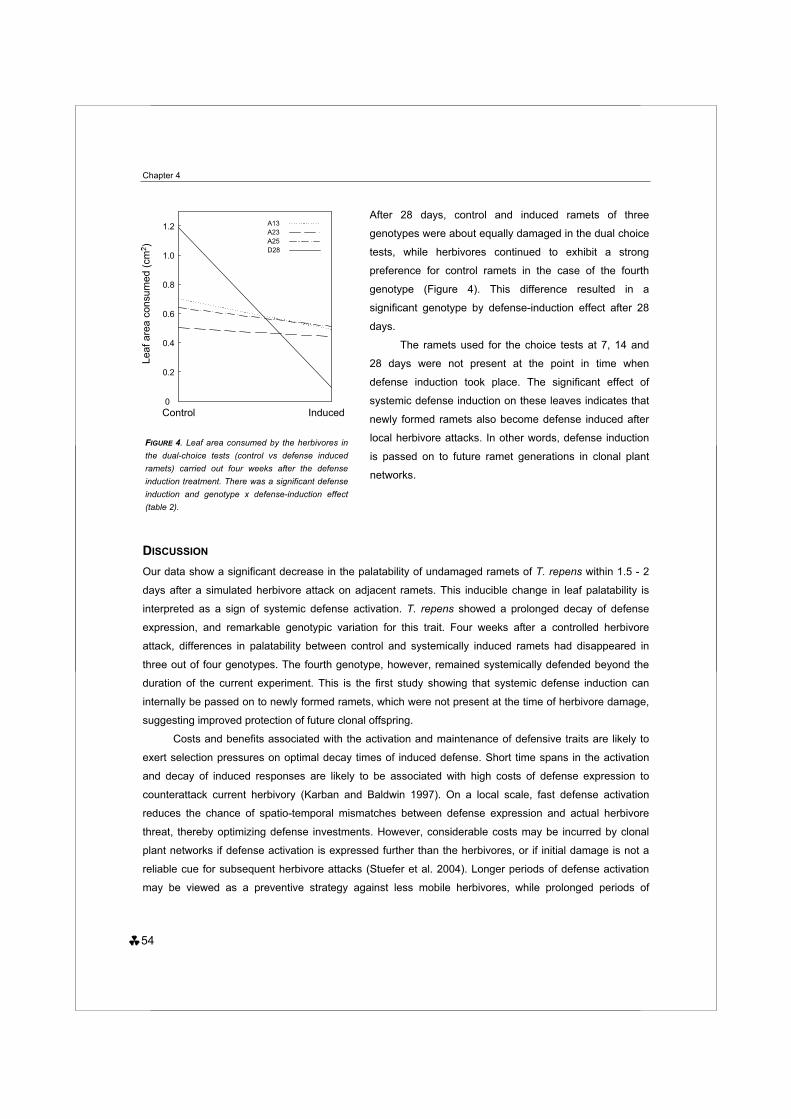
Chapter 4
54
Control Induced
Leaf
are
a co
nsum
ed (c
m2 )
0
0.2
0.4
0.6
0.8
1.0
1.2 A13 A23 A25 D28
Control Induced
Leaf
are
a co
nsum
ed (c
m2 )
0
0.2
0.4
0.6
0.8
1.0
1.2
Control Induced
Leaf
are
a co
nsum
ed (c
m2 )
0
0.2
0.4
0.6
0.8
1.0
1.2 A13 A23 A25 D28
After 28 days, control and induced ramets of three
genotypes were about equally damaged in the dual choice
tests, while herbivores continued to exhibit a strong
preference for control ramets in the case of the fourth
genotype (Figure 4). This difference resulted in a
significant genotype by defense-induction effect after 28
days.
The ramets used for the choice tests at 7, 14 and
28 days were not present at the point in time when
defense induction took place. The significant effect of
systemic defense induction on these leaves indicates that
newly formed ramets also become defense induced after
local herbivore attacks. In other words, defense induction
is passed on to future ramet generations in clonal plant
networks.
DISCUSSION
Our data show a significant decrease in the palatability of undamaged ramets of T. repens within 1.5 - 2
days after a simulated herbivore attack on adjacent ramets. This inducible change in leaf palatability is
interpreted as a sign of systemic defense activation. T. repens showed a prolonged decay of defense
expression, and remarkable genotypic variation for this trait. Four weeks after a controlled herbivore
attack, differences in palatability between control and systemically induced ramets had disappeared in
three out of four genotypes. The fourth genotype, however, remained systemically defended beyond the
duration of the current experiment. This is the first study showing that systemic defense induction can
internally be passed on to newly formed ramets, which were not present at the time of herbivore damage,
suggesting improved protection of future clonal offspring.
Costs and benefits associated with the activation and maintenance of defensive traits are likely to
exert selection pressures on optimal decay times of induced defense. Short time spans in the activation
and decay of induced responses are likely to be associated with high costs of defense expression to
counterattack current herbivory (Karban and Baldwin 1997). On a local scale, fast defense activation
reduces the chance of spatio-temporal mismatches between defense expression and actual herbivore
threat, thereby optimizing defense investments. However, considerable costs may be incurred by clonal
plant networks if defense activation is expressed further than the herbivores, or if initial damage is not a
reliable cue for subsequent herbivore attacks (Stuefer et al. 2004). Longer periods of defense activation
may be viewed as a preventive strategy against less mobile herbivores, while prolonged periods of
FIGURE 4. Leaf area consumed by the herbivores in the dual-choice tests (control vs defense induced ramets) carried out four weeks after the defense induction treatment. There was a significant defense induction and genotype x defense-induction effect (table 2).

Timing of induced resistance in a clonal plant network
55
maintained induction should be especially effective against herbivores with predominantly local feeding
behavior.
The time lag between localized herbivore damage and the systemic activation of defense is mainly
a function of three processes: (1) within-leaf processes such as phloem loading, which affects the signal
transport out of the damaged leaf (Babst et al. 2005), (2) long-distance transport between sites of damage
and sites of defense activation, and (3) the induced expression of defense traits in undamaged leaf
tissues (Baldwin et al. 1994). Since phloem sap moves at rates of up to 1-2 cm*min-1 in herbaceous plants
(P. Minchin, personal communication), the initial and final phases can be held responsible for the
prolonged defense activation time, such as reported in this study. The dynamics of these processes can
likely constrain adaptive responses to mobile herbivores by hindering a fast and efficient spread and
expression of induction signals within clonal plant networks.
In accordance with previously published studies on non-clonal plants (Stout et al. 1996; Anderson
et al. 2001; Agrell et al. 2003; Alves et al. 2007), the strength of induced resistance decreased gradually
with time in our system. One of the four genotypes used in this study, however, showed a significantly
prolonged decay time, suggesting the potential for considerable variation in defense timing in this species.
Such variation may translate into ecologically important changes in genotype frequencies over time. In the
case of prolonged herbivore attacks, genotypes with long decay times are likely to gain relative
advantages over genets with short decay times, owing to the selectivity of foraging herbivores. We have
recently shown for the same genotypes used in this study (Gómez et al. 2007) that maintaining
systemically induced defense over a period of three weeks has rather low costs in terms of plant
performance, while prolonged defense expression confers very considerable benefits in case of
subsequent herbivore attacks. These results are indicative of thus far unexplored links between genetic
variation in temporal defense expression and the feeding dynamics of insect herbivores and their impact
on competitive interactions between genotypes in natural populations of clonal plants.
The current study points at an additional benefit resulting from prolonged defense induction.
Ramets developing after the end of the inducing herbivore attack experienced increased protection
against herbivores. This finding implies that induced defense can be extended to new ramet generations,
thereby contributing to the protection of plant tissue which is both very vulnerable to herbivores (Bråthen
et al. 2004, Gómez et al. 2007) and most valuable in terms of future plant growth and fitness (Beinhart
1963). Protection of developing ramets through plastic defense induction may be a crucial element in the
defense strategy of T. repens and similar clonal plant species. Since defense induction signals follow
phloem source-sink gradients (Gómez and Stuefer 2006), systemically induced defense traits are likely to
be unidirectionally expressed in clonal plants. In other words, plastic defenses are mainly expressed in
sink regions for carbohydrates, but cannot easily reach source regions within the network. Because young
ramets represent both strong sinks and are usually preferred by herbivores, systemic defense signaling
may often be ineffective in preventing damage on older (source) ramets. However, the enhanced
protection of newly formed ramets, as shown here, may last long enough to bridge the period of time
during which ramets are especially vulnerable to herbivory (Gómez et al. 2007). Protection during the

Chapter 4
56
early stage of development can be crucial for reducing herbivore damage (Aide and Londoño 1989). An
early warning system such as present in clonal plant networks (Gómez and Stuefer 2006, Gómez et al.
2007) will increase their chance of establishment and survival in presence of future herbivore bouts.
Acknowledgments: The authors thank H. de Kroon and T. van Mölken for their comments on a previous
version of the manuscript. This study was funded by the Netherlands Organization for Scientific Research
(NWO-VIDI fellowship to JFS)

S Gómez, Y Onoda, V Ossipov & JF Stuefer. Submitted.
CHA
PTER5
Systemic induced resistance: a risk-spreading strategyin clonal plant networks?

Chapter 5
58
ABSTRACT
Clonal plant networks consist of interconnected individuals (ramets) of different developmental stages that
differ in nutritional values, biomechanical properties and chemical tissue composition. Damage by
herbivores cause additional changes in plant traits. Clonal plant networks thus represent heterogeneous,
in space and time, ramet assemblages that differ markedly in quality and attractiveness for herbivores.
This study aims to investigate plastic plant responses to herbivore damage which may reduce prolonged,
selective feeding on valuable plant parts. We studied the effect of systemic defense induction on variation
in nutritional, biomechanical and chemical leaf quality between ramets of differently age of the
stoloniferous herb Trifolium repens. Dual choice tests were used to assess the preference of insect
herbivores for young versus mature ramets in induced and uninduced control plants, respectively. In
parallel, we measured leaf traits related to nutrition, biomechanics and chemical defense as variation in
these traits may explain herbivore preference. Prior to damage, young ramets were damaged more than
mature leaves. After systemic defense induction, damage on young ramets was greatly reduced, whilst
leaf consumption on mature ramets increased slightly after induction. Four days of controlled herbivore
attack resulted in a significant increase in leaf strength and leaf thickness, a decrease in total soluble
carbohydrates and a substantial change in the phenolic profile of undamaged ramets connected to
attacked individuals. Systemic induced resistance led to a reduced and more dispersed feeding pattern
among ramets of different ages. We propose that inducible defense acts as a risk-spreading strategy in
clonal plant networks by equalizing herbivore preference within host plants, thereby avoiding extended,
selective feeding on valuable plant tissues. The results are discussed in terms of optimal defense theory.

A risk-spreading strategy in clonal plants?
59
INTRODUCTION Plant individuals are formed by a fine-scale mosaic of tissues differing in structure and quality. Variation in
tissue quality is present both within and between plant organs, making plants very heterogeneous food
sources. This can negatively affect small insect herbivores, whose performance and fitness can be
influenced even by subtle within-leaf changes in herbivore-relevant traits (Shelton 2004). Negative effects
of tissue heterogeneity on herbivore performance may manifest as reduced detoxification efficiency of
feeding animals (Berenbaum and Zangerl 1993), as prolonged larval development periods (Stockhoff
1993) or as altered dispersal behavior of feeding animals (Anderson and Agrell 2005; Rodriguez-Saona
and Thaler 2005).
Small-scale heterogeneity in food quality is virtually ubiquitous and herbivores have therefore
developed intricate abilities to avoid low-quality tissues and search selectively for more nutritious and less
well-defended food (Bernays 1998). In turn, efficient foraging and selective feeding of herbivores can
strongly impair plant performance, because the most attractive tissues for herbivores, such as young
leaves, fruits and seeds, are often the most valuable parts in terms of plant functioning and fitness.
Selective feeding on valuable tissues may prompt the evolution of plant defense strategies which act to
mislead foraging herbivores by disguising plant organs whose functioning is closely linked to fitness.
Consequently, plants may not increase but decrease within-host variation in food quality upon herbivore
attacks. Coordinated modular responses (de Kroon et al. 2005) may allow fine-tuning of within-plant
defense levels in order to compensate for variation in tissue attractiveness and vulnerability, thereby
spreading the risk of herbivore attacks on valuable tissues over a larger part of the whole plant.
Clonal plants often form networks consisting of many interconnected individuals (i.e. ramets) which
are successively produced on the nodes of laterally extending stolons and rhizomes. Due to this growth
pattern, assemblages of ramets originating from the same clone differ consistently in their developmental
stages, and thus represent plants with a potentially high within-clone variation in tissue quality for feeding
insects.
From an herbivore’s perspective leaf quality is mainly a function of the nutritional value of tissues,
biomechanical leaf properties and the presence of defense-related metabolites. Consequently, within-host
variation in tissue quality is largely driven by constitutive traits (generated by ontogeny) and inducible
defense expression. Young leaves tend to have higher nutritional values than older leaves (e.g., higher
amounts of nitrogen; Coley 1983), and before completion of their structural development juvenile leaves
often show low degrees of biomechanical resistance (e.g. leaf strength; Coley 1983). Both of these factors
act to increase quality differences between tissues of different ages potentially resulting in major within-
plant differences in herbivory pressures (Coley and Barone 1996; Anderson and Agrell 2005). Continued
selective feeding on certain plant parts can seriously impair plant performance, and may ultimately affect
population and community dynamics (reviewed in Halpern and Underwood 2006).
To compensate and counteract selective damage by herbivores, plants may unevenly distribute
defense components. Optimal defense theory (McKey 1974) postulates that within-host variation in

Chapter 5
60
defense expression confers improved protection of tissues with a relatively high contribution to plant
fitness and under high risk of attack. Accordingly, young and reproductive tissues often contain
significantly enhanced amounts of chemical defense compounds (van Dam et al. 1994; de Boer 1999;
Bråthen et al. 2004; but see Cronin and Hay 1996; Bluthgen and Metzner 2007).
Many clonally growing plants form networks of interconnected individuals, called ramets, which are
produced on rooting nodes of laterally extending stolons and rhizomes. Due to this growth pattern, clonal
plants are characterized by potentially large spatial extents (van Groenendael and de Kroon 1990) and by
high degrees of modularity and module autonomy (de Kroon et al. 2005; Magyar et al. 2007; Oborny
2003). In terms of host-plant heterogeneity for feeding insects, clonal plant networks can be regarded as
assemblages of genetically identical ramets, which differ consistently in tissue age and developmental
stage (Huber and Stuefer 1997), leading to potentially high degrees of within-clone variation in tissue
quality. Ontogenetic differences between ramets may drive herbivore preference for young ramets in
uninduced clonal plants (Bråthen et al. 2004; Gómez et al. 2007). Selective feeding on young ramets can
have detrimental effects on the performance and fitness of clones since young ramets are primarily
responsible for future vegetative growth and reproductive success of stoloniferous plant individuals
(Beinhart 1963). Systemic defense induction after localized herbivore damage, may act as a modular risk-
spreading strategy within groups of unevenly-aged ramets of clonal plants by overriding ontogenetic
differences in tissue quality, and by making both mature and young ramets equally (un)palatable, and
thereby selectively enhancing the protection of more vulnerable young ramets.
Herbivory can prompt plastic responses in the attacked plant that extend considerably beyond the
site of damage (Karban and Baldwin 1997). Inducible systemic defense responses have the potential to
protect undamaged plant tissues by spreading defense activation signals to unaffected plant parts thereby
alerting them of impending herbivore threats (Gómez and Stuefer 2006; Heil and Silva Bueno 2007).
Induced defense responses can hence affect within-plant variation in tissue quality (Stout et al. 1996;
Shelton 2005) by altering biomechanical, nutritional and chemical leaf traits. Most studies on induced
defenses have focused on the production of chemical defense compounds (Karban and Baldwin 1997).
Nevertheless, nutritional and biomechanical tissue properties should not be disregarded in the context of
inducible defense strategies as they can strongly reduce herbivores preference and/or performance (Levin
1973; Coley 1983), and because they are at least partially inducible by herbivore feeding (Baur et al.
1991; Rautio et al. 2002; Xiang and Chen 2004; Massey and Hartley 2006; Massey et al. 2007; Redak
and Capinera 1994).
We aimed at answering the following research questions. (1) Is there consistent within-plant
variation in herbivore preference in clonal plant networks of the stoloniferous species Trifolium repens? In
other words: are younger ramets more attractive for feeding insects than older ramets? (2) Does systemic
defense induction alter within-plant herbivore preference? (3) Do young and mature ramets of the same
clonal plant network differ in their nutritional, biomechanical and chemical defense traits, before and after
controlled herbivore damage? (4) How does ramet variation in nutritional, biomechanical and chemical
defense traits affect herbivore preference for young and mature ramets within clonal plant networks?

A risk-spreading strategy in clonal plants?
61
MATERIAL AND METHODS
Study organisms
We used four genotypes (labeled A13, A23, B11, D28) of the stoloniferous herb Trifolium repens L. based
on their ability to become defense-induced after herbivory damage as shown in previous studies (Gómez
and Stuefer 2006; Gómez et al. 2007). The genotypes originated from natural riverine grasslands situated
along the river Waal, The Netherlands.
Larvae of Spodoptera exigua H. were used as herbivore in this study. S. exigua is a generalist
caterpillar with a broad host range. The caterpillar colony was maintained at a constant temperature of 24
ºC and 8h dark : 16h light photoperiod. Larvae were reared on an artificial diet described in Biere et al.
(2004).
Experimental procedure
A series of four studies with a similar set-up were conducted to study effects of systemic defense
induction on nutritional, biomechanical and chemical plant traits. The stock plant material was vegetatively
propagated in a greenhouse at a mean temperature of 23.9 ± 0.1°C and at a 8h dark : 16h light photo-
period. For each study we used 16 to 20 apical cuttings per genotype. Cuttings consisted of three to five
ramets (T. repens has one leaf per ramet) and were transplanted individually into plastic trays (16cm x
12cm x 5cm) at least one week before the start of the induction treatment. Half of the cuttings of each
genotype were randomly assigned to the defense-induction treatment and the other half to the control
group. We used the same procedure to induce defense in each of the studies described below. We
confined two 3 P
rdP-instar caterpillars in a plastic cage mounted on the 6P
thP and 7 P
thP youngest ramet (Gómez
and Stuefer 2006). An empty cage was mounted on the control plants in a similar manner. The caterpillars
fed on the plants for four days. After the defense induction period, a number of traits (see below) were
measured on the first fully unfolded ramet (hereafter referred to as young ramet) and the fifth-youngest
ramet (hereafter referred to as mature ramet) from the same plant. Ramets of Trifolium repens transition
from sink to source tissues once they have expanded about 25-45% of their final surface area and
reached 45-55% of their maximum weight (Chapman et al. 1990b). In our experiments, we started
counting from the first fully unfolded ramet (young ramet) and selected ramets in the 4P
thP or 5 P
thP position as
mature ramets to ensure that they were fully developed. Due to the small size of T. repens leaves, the
experimental procedure was repeated in four different occasions to collect enough material for the C and
N analysis (May 2005), biomechanical traits and carbohydrate measurements (May 2006), herbivore
preference (September 2006) and phenolic profiling (February 2007), respectively.
Herbivore preference
To determine the preference of herbivores for either young or mature ramets, and to investigate whether
this preference is influenced by defense induction, we conducted 20 dual-choice tests per genotype
between young and mature leaves of defense-induced and control plants, respectively (Figure 1). After

Chapter 5
62
FIGURE 1. Schematic representation of the experimental design. Defense induced plants (black) were submitted to a controlled herbivore attack on the 6P
thP and 7 P
thP ramets by
confining two S. exigua larvae in a petri dish mounted on the ramets. An empty petri dish was placed on the 6 P
thP and 7 P
thP ramets of the
control plants (white). After the defense induction treatment, dual choice tests were performed between the 1 P
stP (young) and 5 P
thP
ramet (mature) of each plant.
the defense induction treatment described above, we collected the young and the mature ramet from each
plant. The excision of leaves does not alter caterpillar preference (Gómez and Stuefer, unpublished data).
Both leaves (without the petioles) were paired in a petri dish on moist filter paper. A 3P
rdP-instar caterpillar
was released in the middle between the leaves and allowed to feed until 30% of one of the leaves was
consumed or after 48 hours had passed. At the end of the bioassays, digital photographs were taken and
the area consumed by the herbivores was measured with the image analysis software Image Pro Plus,
version 1.1 (Media Cybernetics, Bethesda, Maryland, USA)
Leaf quality
We defined tissue quality in terms of three categories of traits known to have an effect on herbivore
preference and/or performance: (a) nutrition: nitrogen and carbon percentage, C/N ratio and total
concentration of soluble carbohydrate; (b) biomechanics and morphology: thickness, toughness, strength,
fresh and dry leaf mass, area and leaf mass per area (LMA); and c) chemical defense: phenolic
compounds of leaf tissue (phenolic profile).
Leaf carbon and nitrogen percentage
We determined the carbon (C) and nitrogen (N) percentage and calculated C/N ratios for the young and
mature ramets of defense-induced and control plants, respectively. We analyzed the first fully expanded
ramet (young) and the fourth-youngest ramet (mature) from the same plant. Leaves were oven dried at
75˚C for two days, weighed, and ground. 1-2 mg per sample were used to determine carbon and nitrogen
percentage in a nitrogen/carbon/sulphur analyzer (NA1500, Carlo Elba Instruments, Milan, Italy).
Defense induction
5th ramet 1st ramet
Choice test
Choice test
Control
Defense induction
5th ramet 1st ramet
Choice test
Choice test
Control
Defense induction
5th ramet 1st ramet
Choice test
Choice testDefense induction
5th ramet 1st ramet
Choice test
Choice test
Control

A risk-spreading strategy in clonal plants?
63
Leaf morphology, biomechanics and anatomy
Four days after the defense induction treatment started, the young and mature ramets were cut at the
base of the petiole and placed in a wet tissue to minimize water loss and transported in a cooler to the
laboratory for measurements. We used a punch-and-die test to measure leaf strength and leaf toughness
(Aranwela et al. 1999) on the middle leaflet from each trifoliate leaf by penetrating the tissue with a 1.345
mm diameter steal punch. Force and displacement were measured simultaneously with a general testing
machine (5542, Instron, Canton, MA, USA). From the force-displacement curve, we calculated punch
strength (the maximum force per unit punch area to fracture a leaf, MN m P
-2P)) and punch toughness (total
work required to fracture a leaf per unit punch area, KJ m P
-2P) (Aranwela et al. 1999). As these parameters
depend on thickness and material property, we also calculated thickness adjusted mechanical properties
namely specific punch strength (punch strength/thickness, MN mP
-2P mmP
-1P) and specific punch toughness
(punch toughness/thickness, KJ mP
-2P mmP
-1P) (Read & Sanson 2003). Thickness, area, fresh and dry mass of
one of the leaflets was also measured.
After performing the punch-and-die test, we cut off two pieces (~ 2 mmP
2P) of each leaf for an
anatomical analysis. The pieces were chemically fixed and embedded in resin (LR-white, London Resin
Company, Berkshire, England). Slices of 1 m thickness were made from the embedded material using a
microtome (OMU-3, Leica Rijswik, The Netherlands) and stained with toluidine blue for light microscopy.
Digital photographs were taken of each sample. To have a better understanding of the mechanism
underlying changes linked to biomechanical traits, we counted the average number of cells per cross
section and measured thickness of different tissue layers. We measured the average thickness of the
upper epidermis, the spongy and the palisade parenchyma and the lower epidermis on three line
transects running perpendicularly to the leaf surface through each cutting.
Leaf carbohydrate concentration
The area of the remaining two leaflets was measured and stored at -80˚C. Each leaf sample (5-7 mg) was
frozen in liquid nitrogen and homogenized using a ball-mill (MM 300, Retsch GmbH and Co. KG). The
homogenate was suspended in 1 ml of 80% ethanol for 30 min at 80°C, centrifuged for 10 min at 10000 g
and the resulting supernatant was used to measure total content of soluble carbohydrates using anthrone
reagent (Yemm and Willis 1954). To remove the starch fraction, the remaining pellet was washed with
70% ethanol, suspended in 1 ml of 0.2M KOH and incubated for 30 min at 90°C. After cooling, 0.2 ml of
1M acetic acid and 1 ml of amyloglucosidase (35 units per ml in 50mM acetate buffer, pH 4.5) were added
and incubated for 30 min at 55°C. The extract was centrifuged 10 min at 10000 g. The pellet was washed
with water and ethanol two times and centrifuged. The remaining pellet was dried and the weight used as
an estimate of the cell wall content (structural carbohydrates).

Chapter 5
64
Leaf phenolics
Due to the small size of T. repens leaves, two leaves of the same age were combined for each sample.
Young and mature leaves of induced and control plants were collected, frozen in liquid nitrogen and
transported to the laboratory. The plant material was freeze-dried for 72 h. The dried samples were
homogenized into a powder using a ball-mill (Retsch GmbH and Co. KG) and stored at -20 ºC. The
homogenized sample (15 - 20 mg) was extracted with 1 ml of 80% methanol for 40 min at room
temperature with continuous stirring (1.5 units, Vortex, Genie 2). The homogenate was centrifuged for 20
min at 2900 g, the extract was transferred into a 1.5 ml plastic Eppendorf tube and methanol removed in
vacuum concentrator (Concentrator 5301, Eppendorf AG, Germany). The resulting aqueous phase was
frozen and lyophilized for 24 h. Dry residue of T. repens metabolites was dissolved in 0.2 ml of 50%
methanol, and then centrifuged for 20 min at 2900 g. The clean extract was transferred into 1.5-ml
Eppendorf tube and stored in the freezer at -20 C.
Individual phenolics were quantified with a High Performance Liquid Chromatography- Diode Array
Detector (HPLC-DAD) system (Merck-Hitachi, Tokyo, Japan) that included a pump L-7100, a diode array
detector L-7455, a programmable autosampler L-7250, and an interface D-7000. The column used was
Superspher 100 RP-18 (75 x 4 mm i.d., 4 m, Merck, Germany). A 0.1 ml of extract was transferred into
vials for HPLC autosampler with 0.15 ml inserts and closed by caps with septa. Two solvents were used:
(A) 1% formic acid; (B) acetonitrile. The elution profile was: 0-3 min, 98% A (isocratic); 3-22 min, 2-23% B
in A (linear gradient); 22-45 min, 23-45% B in A (linear gradient); 45-50 min, 45% B in A (isocratic). Flow
rate was 1 ml/min; detection at 320 nm. The acquisition of UV spectra (230-355 nm) was done
automatically at the apex of each peak.
For quantification of individual phenolics, HPLC raw data of all samples were exported into
COWTool software (COWTool version 1.1., Department of Biotechnology, DTU, Denmark) in text format
and subjected to baseline correction to reduce the noise level. Afterwards, chromatograms derived from
each sample were aligned against the master chromatogram of polar phenolics to correct retention time
differences between different samples. Processed raw data were used for determination of retention time
and height of all peaks of individual phenolics. Relative content of individual phenolics was calculated as a
peak height per 1 g of dry mass of leaf sample.
Statistical analysis
Two-way repeated measures ANOVA was used, where the repeated (within-subject) factor was ramet age
and the fixed between-subject factors were genotype and defense-induction. Phenolic profiles were
analyzed using a canonical discriminant analysis (SAS procedure CANDISC) with leaf age and induction
state as the group variable. All analyses were conducted with SAS 9.1 (SAS Institute Inc., Cary, NC,
USA).

A risk-spreading strategy in clonal plants?
65
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Control Induced
Leaf
are
a da
mag
ed (c
m2 )
young
mature
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Control Induced
Leaf
are
a da
mag
ed (c
m2 )
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Control Induced
Leaf
are
a da
mag
ed (c
m2 )
young
mature
young
mature
RESULTS
Herbivore preference
Ramet age had a significant effect on the amount of leaf area consumed by the larvae when they were
offered young and mature ramets (Table 1). The results of the dual-choice tests on uninduced control
plants showed that S. exigua larvae consistently preferred young ramets over mature ramets of T. repens
(Figure 2). Young ramets incurred on average a three times higher damage than mature ramets.
Defense induction had a very strong effect on
the damage inflicted on the ramets (Table 1). The
dual-choice tests between young and mature ramets
from defense induced plants showed that larvae
preferred mature ramets over young ones, revealing
a significant change in preference after systemic
defense induction (Table 1). The average leaf area
consumed from the mature ramets was 50% lower
than the leaf area eaten from young ramets. There
were no significant differences in palatability
among genotypes, but there was a significant
genotype by defense induction effect.
Leaf total content of carbon and nitrogen
Genotypes differed significantly with respect to leaf carbon and nitrogen percentage as well as C/N ratio
(Table 2). Young ramets contained a significantly higher percentage of nitrogen, which resulted in a lower
C/N ratio. Percent carbon did not differ for young and mature ramets (Tables 2 and 3). Defense induction
did not affect carbon and nitrogen percentage, nor did it alter C/N ratios. However, there was a marginally
significant interaction effect of ramet age by induction on leaf carbon percentage.
TABLE 1. Herbivores preference. Repeated measures ANOVA.
FIGURE 2. Average leaf area (+1SE) consumed by 3 P
rdP-instar S. exigua larva in dual-choice tests.
In each choice test, the youngest fully expanded (young) and fifth-youngest (mature) leaf were confronted. Choice-tests were performed separately for control and defense-induced plants. The results of the four genotypes are pooled in this figure; n=40 for each bar. For more details see materials and methods.
df MS F
3 0.030 0.491 1.159 19.25***3 0.242 4.02*
72 0.060
1 0.073 4.75*3 0.152 0.991 3.252 21.05***3 0.259 1.68*
72 0.154Error
Source
Gen x IndError
Age x IndAge x Gen x Ind
Between subjects effectsGenotype
Within subjects effects
Age x Gen
Induction
Age
P
P < 0.0001***, 0.01< P < 0.05 *

Chapter 5
66
TABLE 2 . Nutritional leaf traits. Repeated measures ANOVA
Leaf morphology, biomechanics and anatomy
The punch-and-die revealed a significant effect of ramet age on specific punch strength and specific
punch toughness (Table 4). After defense induction, both young and mature leaves became significantly
stronger but not tougher per unit thickness. Genotypes differed significantly in all the morphological traits
except for specific punch strength. Ramet age had a significant effect on dry and fresh mass, LMA and a
marginally significant effect on leaf area. Mature ramets were heavier and had a higher LMA than young
ramets. Defense induction significantly increased thickness and dry mass in both young and mature
ramets (Table 4). The thickness of the upper and lower epidermis was genotype and age dependent. The
thickness of the individual layers measured in the anatomical study was not significantly affected by
defense induction and the same was true for the number of cells per cross section (Table 3).
Leaf specific punch strength differed for the different age classes in control and defense-induced
plants. Young ramets from control plants had the lowest leaf specific punch strength, which was coupled
with the highest leaf damage scored in the choice tests (Figure 3). The ramets in the other three
categories (defense-induced young, defense-induced mature and control mature) received approximately
similar amounts of damage, while their leaf specific punch strength slightly varied.
df MS F MS F MS F3 9.048 8.46** 9.14 5.66** 89.6 6.03**1 0.389 0.360 0.08 0.05 17.0 1.143 0.987 0.920 0.63 0.39 12.3 0.831 2.908 2.720 3.11 1.92 28.8 1.94
21 1.069 1.61 14.9
1 2.489 8.85** 0.08 0.07 27.97 6.05*3 2.872 10.21*** 0.98 0.91 26.72 5.76**1 0.008 0.030 3.32 3.08$ 1.89 0.413 0.304 1.080 0.46 0.42 2.58 0.561 0.019 0.070 0.49 0.45 2.09 0.45
21 0.281 1.08 4.63Error
Age x GenAge x Ind
Age x Gen x IndAge x Block
Between subject effects
Error Within subject effects
Age
Genotype (Gen) Induction (Ind)
Gen x IndBlock
Source N% C% C/N
P < 0.0001***, 0.001< P <0.01 **, 0.01< P < 0.05 *, 0.05 <P<0.1 $
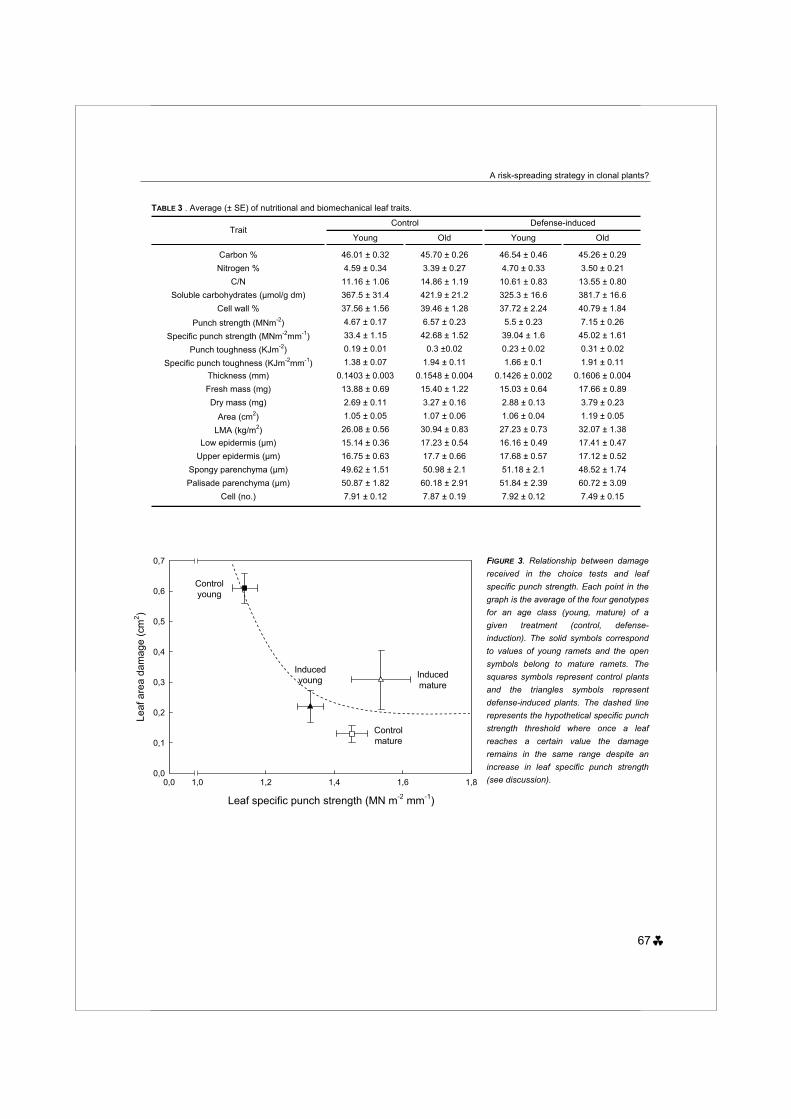
A risk-spreading strategy in clonal plants?
67
Leaf shear strength (MPa)
0,0 1,0 1,2 1,4 1,6 1,8
Leaf
are
a da
mag
e (c
m2 )
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Control young
Inducedyoung
Control mature
Inducedmature
Leaf shear strength (MPa)
0,0 1,0 1,2 1,4 1,6 1,8
Leaf
are
a da
mag
e (c
m2 )
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Control young
Inducedyoung
Control mature
Inducedmature
Leaf shear strength (MPa)
0,0 1,0 1,2 1,4 1,6 1,8
Leaf
are
a da
mag
e (c
m2 )
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Control young
Inducedyoung
Control mature
Inducedmature
Young Old Young Old
Carbon % 46.01 ± 0.32 45.70 ± 0.26 46.54 ± 0.46 45.26 ± 0.29Nitrogen % 4.59 ± 0.34 3.39 ± 0.27 4.70 ± 0.33 3.50 ± 0.21
C/N 11.16 ± 1.06 14.86 ± 1.19 10.61 ± 0.83 13.55 ± 0.80Soluble carbohydrates ( mol/g dm) 367.5 ± 31.4 421.9 ± 21.2 325.3 ± 16.6 381.7 ± 16.6
Cell wall % 37.56 ± 1.56 39.46 ± 1.28 37.72 ± 2.24 40.79 ± 1.84
Punch strength (MNm-2) 4.67 ± 0.17 6.57 ± 0.23 5.5 ± 0.23 7.15 ± 0.26
Specific punch strength (MNm-2mm-1) 33.4 ± 1.15 42.68 ± 1.52 39.04 ± 1.6 45.02 ± 1.61
Punch toughness (KJm-2) 0.19 ± 0.01 0.3 ±0.02 0.23 ± 0.02 0.31 ± 0.02
Specific punch toughness (KJm-2mm-1) 1.38 ± 0.07 1.94 ± 0.11 1.66 ± 0.1 1.91 ± 0.11Thickness (mm) 0.1403 ± 0.003 0.1548 ± 0.004 0.1426 ± 0.002 0.1606 ± 0.004Fresh mass (mg) 13.88 ± 0.69 15.40 ± 1.22 15.03 ± 0.64 17.66 ± 0.89Dry mass (mg) 2.69 ± 0.11 3.27 ± 0.16 2.88 ± 0.13 3.79 ± 0.23
Area (cm2) 1.05 ± 0.05 1.07 ± 0.06 1.06 ± 0.04 1.19 ± 0.05
LMA (kg/m2) 26.08 ± 0.56 30.94 ± 0.83 27.23 ± 0.73 32.07 ± 1.38Low epidermis ( m) 15.14 ± 0.36 17.23 ± 0.54 16.16 ± 0.49 17.41 ± 0.47
Upper epidermis ( m) 16.75 ± 0.63 17.7 ± 0.66 17.68 ± 0.57 17.12 ± 0.52Spongy parenchyma ( m) 49.62 ± 1.51 50.98 ± 2.1 51.18 ± 2.1 48.52 ± 1.74Palisade parenchyma ( m) 50.87 ± 1.82 60.18 ± 2.91 51.84 ± 2.39 60.72 ± 3.09
Cell (no.) 7.91 ± 0.12 7.87 ± 0.19 7.92 ± 0.12 7.49 ± 0.15
TraitControl Defense-induced
TABLE 3 . Average (± SE) of nutritional and biomechanical leaf traits.
Leaf specific punch strength (MN mP
-2P mm P
-1P)
FIGURE 3. Relationship between damage received in the choice tests and leaf specific punch strength. Each point in the graph is the average of the four genotypes for an age class (young, mature) of a given treatment (control, defense-induction). The solid symbols correspond to values of young ramets and the open symbols belong to mature ramets. The squares symbols represent control plants and the triangles symbols represent defense-induced plants. The dashed line represents the hypothetical specific punch strength threshold where once a leaf reaches a certain value the damage remains in the same range despite an increase in leaf specific punch strength (see discussion).

Chapter 5
68
dfM
SF
MS
FM
SF
MS
FM
SF
MS
F3
7.24
11.0
7***
0.47
4.46
**21
6.7
9.64
***
0.00
4640
.74*
**0.
101.
390.
464.
68**
14.
246.
47*
0.13
1.24
47.3
2.1
0.00
065.
4*0.
476.
32*
0.14
1.44
31.
281.
960.
100.
9859
.02.
62$
0.00
000.
290.
040.
480.
101
11.
962.
99$
0.00
0.00
186.
18.
28**
0.00
075.
98*
0.35
4.65
*0.
090.
955
0.65
0.11
22.5
0.00
01(5
3)0.
07(5
2)0.
10(5
2)
13.
014.
43*
0.09
1.59
58.2
93.
53$
0.00
010.
60.
284.
22*
0.63
5.04
*3
0.11
0.16
0.12
2.03
80.1
74.
85**
0.00
2114
.82*
**0.
162.
48$
0.25
1.97
10.
941.
380.
091.
50.
000.
000.
0001
0.53
0.03
0.42
0.11
0.85
30.
600.
880.
040.
732.
020.
120.
0002
1.31
0.01
0.19
0.02
0.14
10.
190.
280.
030.
560.
950.
060.
0004
2.63
0.00
40.
070.
090.
7555
0.68
0.06
16.5
30.
0001
(53)
0.07
(52)
0.13
(52)
Erro
r
Age
x G
enA
ge x
Ind
Age
x G
en x
Ind
Age
x B
lock
Bloc
kEr
ror
With
in s
ubje
ct e
ffect
sA
ge
Betw
een
subj
ect e
ffect
sG
enot
ype
(Gen
) In
duct
ion
(Ind)
Gen
x In
d
Spec
ific
punc
h to
ughn
ess
Sour
ce
Dry
mas
sLe
af a
rea
LMA
Thic
knes
sSp
ecifi
c pu
nch
stre
ngth
PVal
ues
inbr
acke
tsin
dica
te a
ccom
pany
ing
degr
ees
offre
edom
. P
< 0
.000
1***
,0.0
01<
P <
0.01
**,0
.01<
P <
0.05
*,0.
05 <
P<0
.1$
TAB
LE4.
Bio
mec
hani
cal l
eaf t
raits
. Rep
eate
d m
easu
res
AN
OV
A.
A risk-spreading strategy in clonal plants?
69
TABLE 5. Carbohydrate analysis. Repeated measures ANOVA.
Leaf carbohydrate content
The cell wall percentage of leaves
was strongly genotype dependent
(Table 5). Ramet age also had a
significant effect on the
percentage of cell walls per unit
dry mass. Mature ramets had a
higher relative content of cell walls
than younger ramets (Table 3).
After defense induction, there was
significant genotypic variation with
respect to leaf cell wall contents.
In genotype B11, for instance, cell
wall content per unit leaf dry mass
decreased after defense induction from 39 to 31%, while genotype D28 showed an increase from 40 to
49%. The amount of total soluble carbohydrates in white clover leaves was genotype dependent (Table
5). Age also had a significant effect on carbohydrates content. On average, mature ramets had a larger
content of soluble carbohydrates than young ramets (Table 3). Defense induction also had a significant
effect on soluble carbohydrates content. On average, the total amount in the leaf per unit dry mass was
lower in defense-induced plants (395 and 353 mol glucose per unit dry mass, respectively).
Leaf phenolics
98 phenolic compounds were recorded
and quantified in T. repens leaves.
Combinations of these compounds
proved to be specific among particular
combinations of leaf ages and defense-
induction treatments. 98% of the
between-groups variation was explained
by the first two canonical axes (Table 6).
There was a significant difference in the
phenolic profile between different age
groups, and also between young control
and young induced leaves. In contrast, phenolic compounds did not significantly discriminate between
mature control and mature induced leaves (Figure 4; Table 7).
TABLE 6. Canonical correlations.
TABLE 7. Squared Mahalanobis Distance.
P
Each value represents the distance between two groups based on the content of individual phenolic
compounds. Asterisks represent statistically significant values. *** P<0.0001 P
Axis Canonicalcorrelation Eigenvalue Proportion Cumulative Pr > F
1 0.9925 66.28 0.7732 0.7732 <.00012 0.9731 17.81 0.2078 0.981 0.0083 0.7874 1.63 0.019 1 0.995
Source
Between subject effects df MS F df MS FGenotype (Gen) 3 86191 8.51*** 3 1783 34.63***Induction (Ind) 1 47301 4.67* 1 20.23 0.39
Gen x Ind 3 18361 1.81 3 383.93 7.46***Block 1 21625 2.14 1 95.45 1.85Error 54 10126 55 51.49
Within subject effectsAge 1 91275 5.93* 1 208.75 4.99*
Age x Gen 3 59789 3.88* 3 173.04 4.14*Age x Ind 1 9.298 0 1 13.38 0.32
Age x Gen x Ind 3 6494 0.42 3 17.49 0.42Age x Block 1 26647 1.73 1 138.43 3.31$
Error 54 15395 55 41.82
Solublecarbohydrates Cell wall %
P
P < 0.0001***, 0.01< P < 0.05 *, 0.05 <P<0.1 $
Group Control mature Induced young Induced mature
Control young 297.4 *** 137.5 *** 328.6 ***Control mature 277.3 *** 15.3Induced young 272.4 ***

Chapter 5
70
DISCUSSION
Intra-clonal variation and defense-induction
Our study provides clear evidence for age-dependent intra-clonal variation in leaf tissue palatability. Dual-
choice tests with larvae of the generalist caterpillar Spodoptera exigua revealed strong and consistent
herbivore preference for young rather than for mature ramets of the stoloniferous herb Trifolium repens.
After systemic defense induction, however, herbivore preference changed dramatically. Young ramets
were no longer preferred over mature ones. On the contrary, larvae consumed larger amounts of leaf
tissue from mature than from young ramets after defense induction. This striking interaction between
feeding preference and systemic defense activation through localized herbivore attacks was largely
explained by a strong reduction in herbivore acceptance of young, induced ramets and to a lesser degree
by an increase in leaf tissue consumed from mature ramets (Figure 2)
Systemic defense induction clearly decreased intra-clonal variation in tissue quality for feeding
herbivores in our system by annihilating strong variation in tissue attractiveness between ramets of
different ontogenetic stages. Equalizing within-host plant palatability may be part of a defense strategy
acting to spread the risk of herbivore damage among tissues of different importance for plant functioning
and fitness. In our system, defense induction clearly reduced damage to vulnerable and valuable young
ramets. In more natural conditions, such protection may reduce herbivory pressures from selectively
FIGURE 4. Canonical discriminant analysis for phenolic compounds in T. repens. . The axes can 1 and can 2 explain 98% of the observed between-groups variation. There was a significant difference in the content of individual phenolics between different age groups, and also between young control (solid squares) and young induced (solid triangles) ramets. Phenolic compounds did not significantly discriminate between mature control (open squares) and mature induced (open triangles) ramets.
-15
-10
-5
0
5
10
15
-15 -10 -5 0 5 10 15
Can 1
Can
2
Control mature Induced mature Control young Induced young

A risk-spreading strategy in clonal plants?
71
feeding insect larvae. A balanced distribution of within-host plant damage is likely to be less detrimental
than the same amount of damage specifically affecting certain tissues or ramet cohorts (Edwards et al.
1992; Mauricio et al. 1993). The phenotypically plastic nature (i.e., inducibility) of this defense syndrome
suggests potentially high costs associated with an effective protection of young ramets (DeWitt et al.
1998; Miner et al. 2005; Sultan 2000)
Our previous studies on T. repens have shown that the systemic expression of induced defense
after local herbivore attacks is dictated by source-sink relationships within clonal plant networks (Gómez
and Stuefer 2006). As phloem flows are mainly directed towards growing stolon tips (Marshall 1990),
systemic induced resistance is unilaterally expressed in acropetal direction. In the absence of small-scale
heterogeneity in light conditions, attacked older ramets can induce developmentally younger ramets, but
younger ramets cannot warn older ramets of impending herbivore threats due to directional constraints on
the transport of the induction signal. Consequently, natural clones of T. repens may often consist of
induced and uninduced ramets, resembling a patchy within-plant defense induction shown for non-clonal
systems (Stout et al. 1996; Orians et al. 2000). The exact pattern of induction depends on complex spatio-
temporal interactions between the points of herbivore attack, the feeding dynamics of the herbivore and
plant-internal source-sink dynamics. The latter strongly depend on small-scale environmental
heterogeneity (Hutchings and Wijesinghe 1997; Stuefer 1996).
We have recently shown that feeding larvae of S. exigua preferentially attack young ramets of T.
repens in mesocosm-like systems (Gómez et al. 2007). In combination with a strictly unilateral defense
expression (Gómez and Stuefer 2006), this preference could potentially inactivate or circumvent the
defense expression pattern described in the current study. However, herbivore preference for young
leaves was strong, but did not exclude other ramets (Gómez et al. 2007), suggesting that major parts of
the ramet network may become defense induced after some time. Even more importantly, defense
induction through a localized consumption of mature and young ramets is carried on to newly developing
leaves on the same stolon, thereby protecting tissues which have not yet been formed at the time of
herbivore attacks (Gómez, van Dijk and Stuefer, submitted).
Leaf nutritional values, ontogeny and defense-induction
The observed herbivore preference for young ramets of uninduced T. repens plants is in agreement with
results of previous studies on clonal (Bråthen et al. 2004; Gómez et al. 2007) and non-clonal plants (Coley
1983; Fenner et al. 1999). This preference is usually linked to higher nitrogen contents and less
developed physical properties of young leaves (McKey 1974; Coley and Barone 1996; Coley et al. 2006).
In our study young ramets of T. repens contained significantly more nitrogen per leaf dry mass. After
defense induction we observed an overall reduction in the amount of total soluble carbohydrates in the
leaves, while the total amount of foliar carbon, nitrogen and C/N ratio were not affected. Similar effects
were reported for western wheatgrass (Pascopyrum smithii), in which foliage quality in terms of
carbohydrate and foliar nitrogen content decreased after grazing by grasshoppers (Redak and Capinera
1994). Inducible decreases in leaf quality can act as defense response, forcing herbivores with low

Chapter 5
72
mobility to feed for longer periods of time and thereby exposing them to increased predation risks (Feeny
1976; Augner 1995). The observed decrease in foliar soluble carbohydrates after herbivory might be the
result of an increase in the carbon export towards other organs. Reallocating resources towards
herbivore-free places may enable plants to better cope with localized herbivory (Babst et al. 2005).
Inducible biomechanical defense
Biomechanical tissue properties are usually associated with constitutive plant defense. Leaf
biomechanical traits, such as punch strength, are widely recognized as anti-herbivore defense traits
(Coley 1983; Choong 1996; Lucas et al. 2000). In the framework of inducible plant defenses, however,
biomechanical tissue properties have not received nearly as much attention as chemical defense traits.
Some well-studied, inducible changes in plant morphology include thorns (Milewski et al. 1991) and
trichomes (Agrawal 2000; Traw and Dawson 2002), while leaf traits such as size (Massei and Hartley
2000) and mechanical strength (Lowell et al. 1991; Xiang and Chen 2004) are not well-studied, but have
occasionally been reported to change after herbivory.
Biomechanical leaf properties changed significantly as a result of herbivory on connected ramets.
These changes took place in less than four days, and they were observed in both young developing and
in mature ramets. As to be expected, leaf specific punch strength was lower in young than in mature
ramets. This trend was coupled by increased levels of leaf damage on young as compared to mature
ramets, suggesting that biomechanical properties may be playing an important role as constitutive
defense in T. repens. Regardless of significant differences in leaf specific punch strength, young and
mature ramets of defense-induced plants were equally damaged by feeding herbivores. This result might
be explained by two, mutually not exclusive, defense responses consisting of (1) inducible changes in
biomechanical properties and (2) plastic changes in leaf chemistry. After defense induction, leaf specific
punch strength, leaf biomass and leaf thickness increased in both young and mature ramets. Young
ramets may have reached a biomechanical threshold (see Figure 3), which made young and mature
ramets equally unpalatable for 3P
rdP-instar larvae of S. exigua. Aide and Londoño (1989) have shown that
small changes in leaf strength associated with leaf development can play an important role in reducing
survival rates of hatching insect larvae. Plastic changes in biomechanical leaf properties after defense-
induction, such as observed in our study, may effectively hinder or prevent tissue damage from early-
instar larvae of chewing herbivores or small sucking insects (Alvarez 2007). The mechanistic basis for
plastic changes in biomechanical leaf properties remains unknown for this system.
Leaf phenolics
Phenolic compounds are frequently associated with plant defense (Bi et al. 1997; Riipi et al. 2002;
Treutter 2006). Some studies have shown their negative effect on herbivore growth (Kause et al. 1999),
while others report associations with reduced levels of herbivory (Dudt and Shure 1994). The phenolic
composition of leaves and the total amount of phenolic compounds varies considerably with leaf age and
after herbivore damage (Arnold et al. 2004; Van Dam et al. 2005). Our results clearly demonstrate that

A risk-spreading strategy in clonal plants?
73
young ramets of Trifolium repens exhibit a markedly different phenolic profile than mature ramets.
Defense induction through controlled herbivore attacks led to an obvious differentiation in phenolic profile
in young, but not in mature ramets. This supports the idea that the observed reversal in herbivore
preference after defense-induction may be caused by plastic changes in the leaf chemistry and/or
inducible changes in biomechanical properties of young ramets. More detailed analyses of phenolic
compounds and their inducible variation in relation to leaf age and induction status are necessary to
identify specific mechanisms of chemical defense, and to link chemical defense induction to
biomechanical changes in leaf properties.
Optimal defense and risk-spreading
The optimal defense theory predicts that plant tissues with a high contribution to fitness and a high risk of
attack should be better protected than other plant tissues. Young ramets of clonal plants are among the
most valuable tissues for growth and fitness and should therefore be especially well-protected (McKey
1974). Our results show that young ramets of uninduced plants get more damaged than mature ramets.
This increased herbivory risk of young ramets may at least partly be compensated for by rapid leaf
development (ca. 7 days; Carlson 1966), which limits the period of time during which ramets are
vulnerable for selectively feeding herbivores (Herms and Mattson 1992; Coley and Barone 1996). In
addition, young ramets strongly benefit from systemic defense induction, as shown in this study. After
localized herbivore damage, young ramets become strikingly unappealing for feeding caterpillars. This
disproportional increase in the protection of young as compared to mature ramets after defense induction
resulted in a dramatic change in the feeding preferences of foraging larvae, and effectively eliminated
conspicuous differences in the attractiveness of young and mature ramets, such as seen in uninduced
plants. The results of this study hence fit predictions from optimal defense theory applied to inducible
changes in traits that confer protection from herbivory.
In conclusion, systemic induced resistance in clonal plant networks limits the impact of herbivory by
reducing herbivore damage, especially on young ramets, and by equalizing within-plant variation in
attractiveness of different plant tissues. This can be seen as a modular risk-spreading strategy conferring
advantages for the protection of valuable parts of clonal plant networks.
Acknowledgements. We are grateful to I. Hendriks and A. Smit-Tiekstra for the carbohydrate
measurements, B. Verduyn and H. Noordman for assistance in the leaf anatomical analysis and J.
Eygensteyn for the CN analysis. The authors also thank V. Latzel and W. van Dijk for practical assistance
and H. de Kroon and C. Orians for useful comments on a previous version of the manuscript.

Chapter 5
74

S Gómez, V Latzel, Y Verhulst & JF Stuefer. Oecologia (2007)
CHA
PTER6
Costs & benefits of induced resistancein a clonal plant network

Chapter 6
76
ABSTRACT
Plant defense theory suggests that inducible resistance has evolved to reduce the costs of constitutive
defense expression. To assess the functional and potentially adaptive value of induced resistance it is
necessary to quantify the costs and benefits associated with this plastic response. The ecological and
evolutionary viability of induced defenses ultimately depends on the long-term balance between advan-
tageous and disadvantageous consequences of defense induction. Stoloniferous plants can use their
inter-ramet connections to share resources and signals and to systemically activate defense expression
after local herbivory. This network-specific early-warning system may confer clonal plants with potentially
high benefits. However, systemic defense induction can also be costly if local herbivory is not followed by
a subsequent attack on connected ramets. We found significant costs and benefits of systemic induced
resistance by comparing growth and performance of induced and control plants of the stoloniferous herb
Trifolium repens in the presence and absence of herbivores.

Costs & benefits of induced resistance in a clonal plant network
77
INTRODUCTION
Plants can allocate a limited pool of resources to the three main functions of growth, reproduction and
defense, suggesting that increased investments in one function may compromise the others. Empirical
studies have shown that constitutive defense can be costly (e.g., tannins, Sagers and Coley 1995;
glandular trichomes, Elle et al. 1999). Plant defense theory postulates that inducible defense mechanisms
have evolved to reduce these costs by optimizing the temporal match between resource investment into
defense and herbivory threats (Herms and Mattson 1992, Agrawal et al. 1999c). In the last decades,
numerous studies have attempted to find costs of induced defense (reviewed in Bergelson and Purlington
1996 and Strauss et al. 2002) but evidence for costs of inducible plant defense remains scarce and
inconclusive. More recently, empirical evidence has emerged supporting the allocation cost theory with the
help of improved experimental designs, genetic engineering, and biochemical methodology (Baldwin
1998; Van Dam and Baldwin 1998, 2001; Heil and Baldwin 2002).
Inducible resistance is a form of phenotypic plasticity as it allows plants to express an adequate
phenotype in response to temporally and spatially variable herbivore damage. Herbivore-induced changes
in the phenotypes of plants often relate to trait alterations which reduce the palatability and digestibility of
consumed tissue by producing toxic metabolites and/or by up-regulating the production of a variety of
defensive compounds. The ecological viability of induced resistance as an efficient defense strategy
depends on the balance of costs and benefits associated with plastic defense induction. Assessing the
benefits of induced defense in conjunction with possible costs is a prerequisite for estimating the
advantages and disadvantages of plastic defense induction as a potentially adaptive form of phenotypic
plasticity (Dudley and Schmitt 1996; Schmitt et al. 1999) and hence for understanding potential selection
pressures leading to the evolution of induced plant defenses (Agrawal 2000).
Costs of defense have been traditionally measured in terms of decreased plant fitness. Allocation
costs refer to a direct fitness decrease as a consequence of resource-mediated trade-offs between
defense investment and other plant functions. Recent empirical and conceptual work has provided
convincing arguments for the notion that defense induction can also affect fitness in an indirect manner,
via a multitude of potentially complex ecological interactions (Van Dam and Baldwin 1998; 2001; Heil and
Baldwin 2002; Strauss et al. 2002). These costs are commonly referred to as ecological costs.
Allocation theory suggests that costs of plastic defense induction should be more apparent in low-
resource environments than under optimal growth conditions (Herms and Mattson 1992, Bergelson 1994,
Purrington and Bergelson 1996, but see van Dam and Baldwin 2001) as the diversion of resources to
defense cannot easily be compensated for by enhanced resource acquisition. In addition, experiments to
detect costs of defense conducted under quasi-optimal conditions are unlikely to reflect realistic situations,
and therefore, tend to underestimate plasticity costs. To overcome this problem, several studies have
used competitive and/or low-resource environments to quantify costs of induced defense (Siemens et al.
2003 and studies quoted therein). Additionally, previous studies have shown that controlling the genetic
background of plants can substantially enhance the chances to detect costs, by removing confounding

Chapter 6
78
effects due to genetic variation in the induced response (Bergelson and Purlington 1996, Strauss et al.
2002).
Stoloniferous plants consist of multiple, genetically identical individuals (ramets) that are
interconnected by above-ground horizontal stems (stolons). Resource transport within clonal plant
networks has been extensively described in the literature (Pitelka and Ashmun 1985, Marshall 1990,
Alpert 1996, Alpert and Stuefer 1997). Nevertheless, the importance of stolon connections for the
transport of defense agents is a novel aspect (Stuefer et al. 2004) that has only recently been
demonstrated (Gómez and Stuefer 2006). Ramets of the stoloniferous herb Trifolium repens are able to
systemically induce other ramets after local herbivore damage. On one hand, this form of physiological
integration may confer clonal plant networks with considerable benefits by allowing for a fast, specific and
efficient early-warning system among interconnected ramets. On the other hand, the potentially large
spatial scale of clonal plant networks may also lead to substantial costs if network members become
induced without being threatened by herbivores (Gómez and Stuefer 2006). These costs are due to a
potential mismatch in the spatio-temporal scale of plastic defense expression and the dynamics and
patterns of herbivore attacks.
To assess the potentially adaptive nature of plastic responses, “it is necessary to demonstrate that
the phenotype induced in each relevant environment confers higher fitness in that environment relative to
alternative phenotypes” (Schmitt et al. 1999). This is analogous to stating that the induced phenotype
should incur costs in herbivore-free environments, while defense induction should lead to benefits in
herbivore-exposed environments. To quantify costs and benefits we measured traits related to plant
fitness and performance of induced and uninduced T. repens plants in the absence and presence of
herbivores. Growing induced and uninduced plants in the absence of herbivores allows for a quantification
of possible costs of induced resistance, simulating localized damage (e.g., by small herbivores with a low
mobility) and the activation of defense in ramets beyond the feeding range of the herbivore. Benefits of
induction, however, can only be assessed in the presence of herbivores after an initial attack, thereby
simulating a scenario with mobile herbivores showing active foraging behavior beyond the first place of
attack.
In this study we tested the following specific hypotheses: 1) in the absence of herbivores,
systemically induced ramets of clonal plants perform worse than uninduced ramets of the same genotype.
This is due to costs of defense induction when defense is not needed. 2) In the presence of herbivores,
induced ramets of clonal plants perform better than uninduced plants, due to an enhanced protection
through induced defense.
To test these hypotheses we grew induced and uninduced (control) plants of the stoloniferous herb
Trifolium repens together to expose them to mutual competitive interactions, resembling sub-optimal
growing conditions in a sward. To quantify costs and benefits of induced resistance we grew plants in
herbivore-free and herbivore-exposed environments, respectively.

Costs & benefits of induced resistance in a clonal plant network
79
MATERIALS AND METHODS Study organisms
Five genotypes of the stoloniferous herb Trifolium repens L. were vegetatively propagated in a
greenhouse at a mean temperature of 21ºC/19ºC (day/night), and at a 16h/8h (light/dark) photoperiod.
The genotypes originated from natural riverine grassland populations situated along the river Waal, The
Netherlands. They had been collected four years prior to the start of this experiment and were grown
under common garden conditions, eliminating possible maternal and environmental carry-over effects.
The beet armyworm (Spodoptera exigua Hübner) used in this study is a generalist caterpillar with a
broad host range. The caterpillar colony was maintained at a constant temperature of 24 ºC and 16h/8h
(light/dark) photoperiod. The larvae were reared on an artificial diet described in Biere et al. (2004).
Pre-growth of plant material
We started the experiment with 64 cuttings of each of the five genotypes. The cuttings were planted in
pairs into plastic trays (16cm x 12cm x 5cm) using sterilized clay grains as a substrate (Seramis,
Masterfoods GmbH, Germany). Each tray was fertilized weekly with 50ml full-strength Hoagland solution
before the start of the experiment. At the beginning of the experiment, all cuttings consisted of a main
stolon with at least eight fully developed ramets. If present, side branches were removed immediately
before starting the experiment.
Experimental design
The experimental set-up (Figure 1) to measure costs and benefits of systemic induced resistance
consisted of four peripheral trays placed around a central tray, which we will refer to as the competition
tray. All trays were of similar dimensions (16cm x 12cm x 5cm). Each of the peripheral trays contained two
cuttings with at least eight ramets each. The cuttings in two of those trays received a treatment to induce
defense during the entire duration of the experiment (for details see below), while the cuttings in the other
two trays remained uninduced (control). Trays receiving the same treatment were placed diagonally
opposite of each other. The competition tray was placed inside a metal frame (20cm x 15cm x 20cm)
covered by mosquito netting (mesh gauge 0.2 cm2) with four small openings on both longitudinal sides.
The two youngest ramets of each cutting were inserted through the mesh openings and allowed to grow
(proliferate and root) in the competition tray for 19 days. We used five T. repens genotypes, each of which
was replicated four times to measure costs and four times to measure benefits of defense induction. All
induced and control plants grown together in the same experimental set-up (as described above)
belonged to the same genotype. The experimental systems were randomly distributed on greenhouse
benches.

Chapter 6
80
Systemic induction of resistance
Systemic induction of resistance was achieved through a controlled herbivore attack. One Spodoptera
exigua larva was confined with two leaves in one petri dish mounted on the plants (Gómez and Stuefer
2006). The corresponding ramets of uninduced control plants were similarly enclosed in modified petri
dishes but without adding any larvae. The controlled herbivore attack was maintained throughout the
course of the experiment, starting on the ramet on the 8th position (counting from the tip of the stolon) from
each cutting. When the two ramets inside the petri dish had lost at least 50% of leaf tissue, the petri dish
was moved forward on the stolon and the adjacent, younger ramet was inserted into the petri dish.
Whenever the induction treatment was moved forward on the induced cuttings, a comparable leaf area
was removed with scissors from one ramet of each cutting in the control trays. This was done to
compensate for the leaf area loss due to caterpillar feeding in the induced plants. Cutting the leaves with
scissors does not induce resistance against Spodoptera exigua in T. repens (Gómez S and Stuefer JF,
Costs
Competition tray
Benefits
CostsCosts
Competition tray
Benefits
Competition tray
Benefits
FIGURE 1. Schematic representation of the experimental set-up to measure costs and benefits of Systemic Induced Resistance (SIR) in a clonal plant network. Control (white) and defense-induced (gray) plants grew from four peripheral trays into a common, central competition tray. The circles represent petri dishes used for a continued controlled herbivore attack (defense induction treatment). To measure costs of SIR, plants grew together in the absence of herbivores in the competition tray (upper drawing). To measure benefits, 10 caterpillars (wavy black lines) were added to the competition tray (lower drawing). See material and methods for more details.

Costs & benefits of induced resistance in a clonal plant network
81
unpublished data). The induction treatment started one day after the cuttings were placed into the
competition tray. If the caterpillar inside the petri dish died, it was replaced by a new one to maintain
defense induction.
In order to enhance plant interactions, induced and control plants were grown together in the
competition tray. Since all plants growing together belonged to the same genotype, induction effects
cannot be confounded with genetic differences in plant traits, including competitive ability, between
induced and control plants. All measurements described below were performed on ramets growing in the
competition trays.
Costs of Systemic Induced Resistance
Costs of defense induction were measured as a reduction in plant performance. Costs can be measured
after initial herbivore damage (and consequent defense induction) in the absence of subsequent herbivore
attacks. To quantify costs of defense induction we measured the following traits which are known to be
closely related to plant performance and fitness: total biomass production, relative biomass allocation to
leaves, petioles, stolons, and roots, number and length of the main and side stolons and number of ramets
on the main and side stolons. We also measured the petiole length, petiole dry mass, leaf area, leaf dry
mass of the 4th and 5th youngest ramets of each cutting.
Benefits of Systemic Induced Resistance
To quantify benefits of systemic induced resistance we exposed the plants in the competition tray to a
second, controlled herbivory attack (referred to as “herbivory treatment”). We released five 4th instar
caterpillars on day 16 in the competition tray and then added two and three more on day 17 and 18,
respectively, to achieve substantial levels of herbivore damage. The plants were harvested 19 days after
the start of the experiment. We quantified benefits of induced resistance by scoring herbivory damage in
the induced and in the control plants. At the time of harvesting, each ramet on the main stolon was
classified according to the leaf area consumed. We visually estimated the damage and assigned each
ramet a damage category ranging from 0 to 3. The values corresponded to the following amounts of
damage: 0= no damage, 1= 1-33%, 2= 33-66% and 3= 66-100% of leaf area consumed. We also
recorded the position of the damaged ramet on the stolon to investigate possible intra-clonal variation in
the damage pattern according to ramet age. In addition to the degree of damage, we measured the dry
mass of leaves, petioles, stolons and roots in induced and control plants.
Herbivore preference test
One day before releasing the caterpillars (herbivory treatment), we performed two dual choice tests per
competition tray to check whether plants assigned to the defense-induction treatment were systemically
induced. In each competition tray, we cut off two control and two induced ramets of a similar
developmental stage (third-youngest fully expanded leaf). Each control ramet was paired with an induced
one and placed together on a moist filter paper in a petri dish to perform a dual choice test. A fourth instar

Chapter 6
82
S. exigua caterpillar was placed in the middle and allowed to feed until more than 30% of one of the
leaves was consumed or for 48 hours. By means of visual estimates the leaf with the largest area
consumed was recorded for each choice test. In 78 % of the cases the control leaf was more consumed
(Sign test M = 23, P < 0.0001; n = 77) than the induced one, confirming that plants in the competition trays
that had received local herbivore damage (defense induction treatment) were induced before the herbivory
treatment started.
Statistical Analysis
Central competition trays were considered the units of replication in all statistical analyses. To avoid
pseudo-replication and a consequent inflation of degrees of freedoms (Hurlbert 1984), all traits measured
on plants (cuttings) in the competition trays were pooled per treatment (by averaging the four control
cuttings and the four defense-induced cuttings, respectively) prior to data analysis. Consequently, our
experiment had 20 replicates for measuring costs and 20 replicates for assessing benefits. Competing
plants cannot be considered independent from each other as, by definition, they change each other’s
environment, growth and development. To take this dependence into account we used a repeated
measures design to analyze differences between competing plants that belonged to different treatment
groups. Repeated measures analysis explicitly considers intrinsic relationships between treatment groups
(Potvin et al. 1990).
Costs of Systemic Induced Resistance
Repeated measures ANOVA was performed to test for costs of defense induction in number and length of
the main and side stolons and number of ramets on the main and side stolons, relative biomass allocation
to roots, stolons, petioles and leaves and petiole length, petiole dry mass, leaf area, leaf dry mass of the
4th and 5th youngest ramets. Defense induction (induced vs. control) was considered a within-subjects
effect, and genotype was treated as between-subjects effect. Absolute dry masses of roots, stolons,
petioles and leaves were analyzed using two-way repeated measures ANOVA (within-subjects effect:
defense induction; between-subjects effects: plant genotype and herbivory).
Benefits of Systemic Induced Resistance
The amount of damage in the herbivory treatment was assessed with doubly repeated measures ANOVA
using ramet age and defense induction as repeated factors and genotype as main effect. The analysis
included a profile analysis (SAS procedure GLM; profile statement) to test for differences in the degree of
damage between adjacent ramets on the stolons. To correct for differences in the developmental stage of
different cuttings we used only the six youngest ramets of each cutting in the damage analysis. All
analyses were conducted with SAS 9.1 (SAS Institute Inc., Cary, NC, USA).
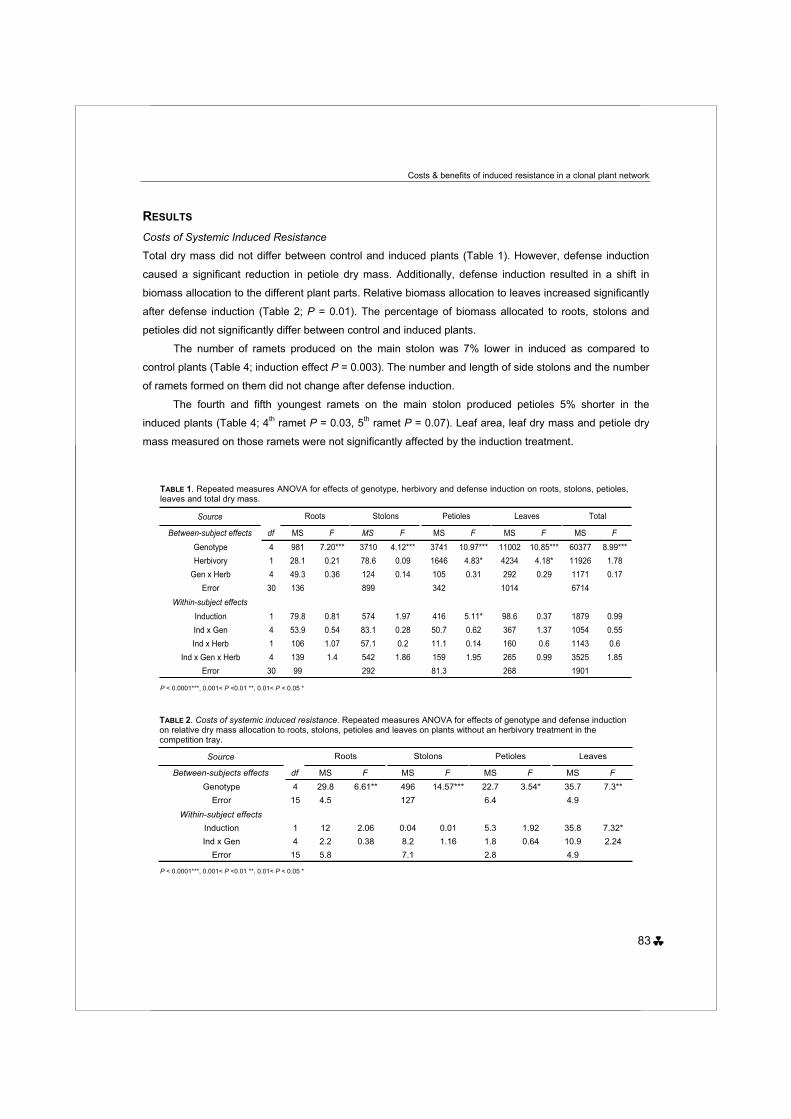
Costs & benefits of induced resistance in a clonal plant network
83
RESULTS
Costs of Systemic Induced Resistance
Total dry mass did not differ between control and induced plants (Table 1). However, defense induction
caused a significant reduction in petiole dry mass. Additionally, defense induction resulted in a shift in
biomass allocation to the different plant parts. Relative biomass allocation to leaves increased significantly
after defense induction (Table 2; P = 0.01). The percentage of biomass allocated to roots, stolons and
petioles did not significantly differ between control and induced plants.
The number of ramets produced on the main stolon was 7% lower in induced as compared to
control plants (Table 4; induction effect P = 0.003). The number and length of side stolons and the number
of ramets formed on them did not change after defense induction.
The fourth and fifth youngest ramets on the main stolon produced petioles 5% shorter in the
induced plants (Table 4; 4th ramet P = 0.03, 5th ramet P = 0.07). Leaf area, leaf dry mass and petiole dry
mass measured on those ramets were not significantly affected by the induction treatment.
P < 0.0001***, 0.001< P <0.01 **, 0.01< P < 0.05 *
Source
Between-subject effects df MS F MS F MS F MS F MS FGenotype 4 981 7.20*** 3710 4.12*** 3741 10.97*** 11002 10.85*** 60377 8.99***Herbivory 1 28.1 0.21 78.6 0.09 1646 4.83* 4234 4.18* 11926 1.78
Gen x Herb 4 49.3 0.36 124 0.14 105 0.31 292 0.29 1171 0.17Error 30 136 899 342 1014 6714
Within-subject effectsInduction 1 79.8 0.81 574 1.97 416 5.11* 98.6 0.37 1879 0.99Ind x Gen 4 53.9 0.54 83.1 0.28 50.7 0.62 367 1.37 1054 0.55Ind x Herb 1 106 1.07 57.1 0.2 11.1 0.14 160 0.6 1143 0.6
Ind x Gen x Herb 4 139 1.4 542 1.86 159 1.95 265 0.99 3525 1.85Error 30 99 292 81.3 268 1901
TotalRoots Stolons Petioles Leaves
TABLE 1. Repeated measures ANOVA for effects of genotype, herbivory and defense induction on roots, stolons, petioles, leaves and total dry mass.
Source
Between-subjects effects df MS F MS F MS F MS FGenotype 4 29.8 6.61** 496 14.57*** 22.7 3.54* 35.7 7.3**
Error 15 4.5 127 6.4 4.9
Within-subject effectsInduction 1 12 2.06 0.04 0.01 5.3 1.92 35.8 7.32*Ind x Gen 4 2.2 0.38 8.2 1.16 1.8 0.64 10.9 2.24
Error 15 5.8 7.1 2.8 4.9
Roots Stolons Petioles Leaves
P < 0.0001***, 0.001< P <0.01 **, 0.01< P < 0.05 *
TABLE 2. Costs of systemic induced resistance. Repeated measures ANOVA for effects of genotype and defense induction on relative dry mass allocation to roots, stolons, petioles and leaves on plants without an herbivory treatment in the competition tray.
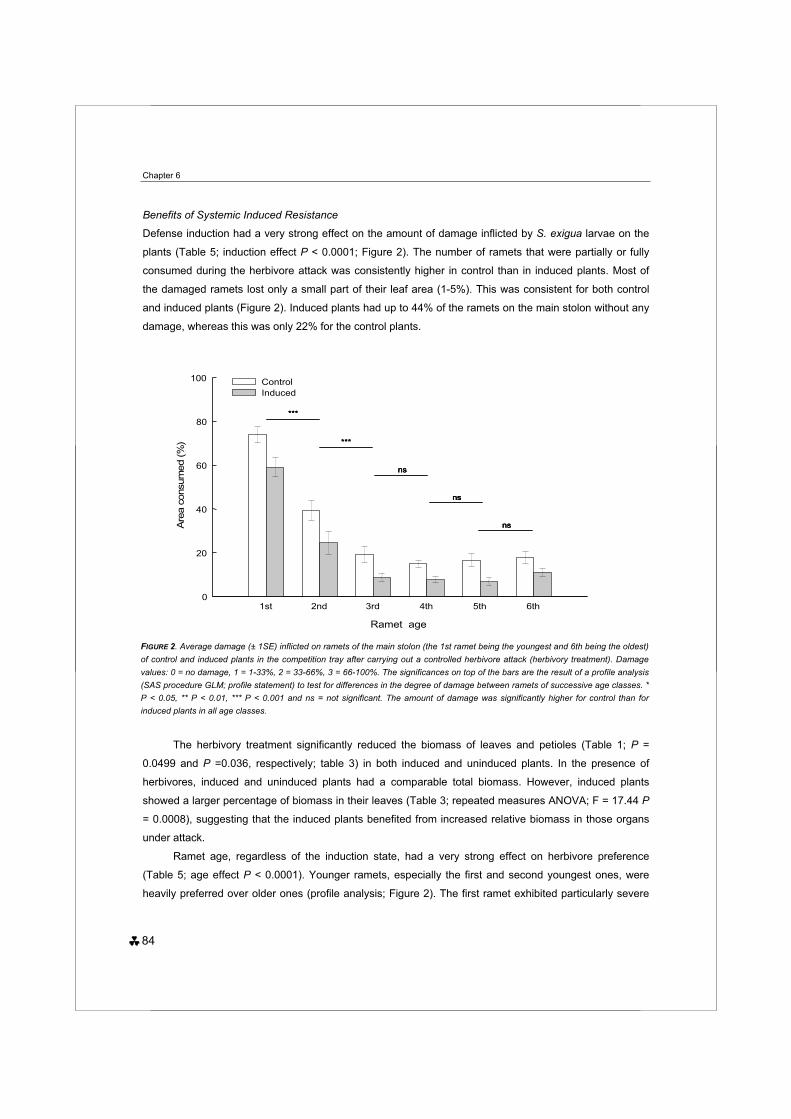
Chapter 6
84
Benefits of Systemic Induced Resistance
Defense induction had a very strong effect on the amount of damage inflicted by S. exigua larvae on the
plants (Table 5; induction effect P < 0.0001; Figure 2). The number of ramets that were partially or fully
consumed during the herbivore attack was consistently higher in control than in induced plants. Most of
the damaged ramets lost only a small part of their leaf area (1-5%). This was consistent for both control
and induced plants (Figure 2). Induced plants had up to 44% of the ramets on the main stolon without any
damage, whereas this was only 22% for the control plants.
The herbivory treatment significantly reduced the biomass of leaves and petioles (Table 1; P =
0.0499 and P =0.036, respectively; table 3) in both induced and uninduced plants. In the presence of
herbivores, induced and uninduced plants had a comparable total biomass. However, induced plants
showed a larger percentage of biomass in their leaves (Table 3; repeated measures ANOVA; F = 17.44 P
= 0.0008), suggesting that the induced plants benefited from increased relative biomass in those organs
under attack.
Ramet age, regardless of the induction state, had a very strong effect on herbivore preference
(Table 5; age effect P < 0.0001). Younger ramets, especially the first and second youngest ones, were
heavily preferred over older ones (profile analysis; Figure 2). The first ramet exhibited particularly severe
FIGURE 2. Average damage (± 1SE) inflicted on ramets of the main stolon (the 1st ramet being the youngest and 6th being the oldest) of control and induced plants in the competition tray after carrying out a controlled herbivore attack (herbivory treatment). Damage values: 0 = no damage, 1 = 1-33%, 2 = 33-66%, 3 = 66-100%. The significances on top of the bars are the result of a profile analysis (SAS procedure GLM; profile statement) to test for differences in the degree of damage between ramets of successive age classes. * P < 0.05, ** P < 0.01, *** P < 0.001 and ns = not significant. The amount of damage was significantly higher for control than for induced plants in all age classes.
Ramet age
1st 2nd 3rd 4th 5th 6th
Are
a co
nsum
ed (%
)
0
20
40
60
80
100 ControlInduced
***
ns
ns
ns
***
Ramet age
1st 2nd 3rd 4th 5th 6th
Are
a co
nsum
ed (%
)
0
20
40
60
80
100 ControlInduced
Ramet age
1st 2nd 3rd 4th 5th 6th
Are
a co
nsum
ed (%
)
0
20
40
60
80
100 ControlInduced
Ramet age
1st 2nd 3rd 4th 5th 6th
Are
a co
nsum
ed (%
)
0
20
40
60
80
100 ControlInduced
***
ns
ns
ns
***
******
nsns
nsns
nsns
******

Costs & benefits of induced resistance in a clonal plant network
85
damage in both induced and control plants (average leaf area consumed > 65%; Figure 2). Defense
induction had a significant effect on leaf area loss due to herbivory in all ramet age classes (Figure 2). The
degree to which systemic defense induction reduced herbivory damage was similar for ramets of all age
classes (Table 5; no age x induction effect). There was a marginally significant genotype effect on the
feeding of the caterpillars (Table 5; genotype effect P = 0.09).
DISCUSSION
Our study provides empirical evidence of significant costs and benefits of systemic induced resistance in a
clonal plant network. In agreement with our hypotheses, induced and control plants showed clear
differences in performance and fitness related traits when grown in the absence and presence of
herbivores. In environments without herbivores, induced plants produced fewer ramets, shorter petioles
and exhibited a shift in biomass allocation patterns. In environments with herbivores, control plants
suffered consistently higher degrees of leaf damage than induced plants. Even though defense induction
resulted in changes in plant growth, and significantly affected the amount of damage caused by the
herbivores, total plant biomass did not respond as expected under the adaptive plasticity hypothesis
(Dudley and Schmitt 1996; Schmitt et al. 1999), as we could not find a significant induction x herbivory
interaction effect. However, we propose that the differences observed in our study (e.g., reduced ramet
production rates and shorter petioles in the cost experiment, decreased amount of leaf damage in the
benefits experiment) are likely to translate into substantial differences in plant productivity, and hence
biomass, in the longer term.
Costs of Systemic Induced Resistance
Biomass production and allocation. Total plant biomass production did not change as a consequence of
defense induction, implying that defense induction did not incur direct and immediate productivity costs.
After induction, however, biomass allocation shifted significantly towards the leaves. We suggest that this
Roots (mg) Stolons (mg) Petioles (mg) Leaves (mg) Total (mg)
15.6 ± 3.0 103.2 ± 6.7 72.6 ± 4.8 107.5 ± 8.3
(4.5 ± 0.7%) (34.9 ± 1.0%) (24.9 ± 0.6%) (35.7 ± 0.7%)
11.3 ± 2.1 96.1 ± 6.0 67.3 ± 4.5 106.8 ± 8.3
(3.4 ± 0.6%) (34.8 ± 1.0%) (24.2 ± 0.5%) (37.6 ± 0.6%)
14.5 ± 2.2 99.5 ± 4.8 62.8 ± 3.7 90.1 ± 5.6
(5.1 ± 0.8%) (37.6 ± 0.8%) (23.8 ± 0.8%) (33.5 ± 0.8%)
14.8 ± 3.7 95.8 ± 6.1 59.0 ± 4.5 95.1 ± 7.6
(4.8 ± 0.9%) (36.8 ± 1.1%) (22.6 ± 0.6%) (35.8 ± 0.6%)
Cos
ts
Uninduced 298.9 ± 21.1
Induced 281.6 ± 19.1
Ben
efits
Uninduced 266.9 ± 13.2
Induced 264.7 ± 19.7
TABLE 3. Average ( SE) absolute and relative dry mass allocated to roots, stolons, petioles and leaves of uninduced and induced plants in absence of a subsequent herbivory treatment (costs) and in presence of a subsequent herbivory treatment (benefits) in the competition tray.

Chapter 6
86
TAB
LE4.
Cos
ts o
f sys
tem
ic in
duce
d re
sist
ance
. Rep
eate
d m
easu
res
AN
OV
A fo
r effe
cts
of g
enot
ype
and
defe
nse
indu
ctio
n on
pla
nt
fitne
ss a
nd p
erfo
rman
ce re
late
d tra
its in
abs
ence
of h
erbi
vore
s
P <
0.0
001*
**,0
.001
<P
<0.
01**
,0.0
1<P
< 0
.05
*
Bet
wee
n-su
bjec
ts e
ffect
sdf
MS
FM
SF
MS
FM
SF
MS
FM
SF
MS
FG
enot
ype
42.
383.
41*
23.5
5.4*
**71
.96.
72**
30.8
1.63
1612
.97*
**18
14.9
3***
8.57
15.3
6***
Erro
r15
0.69
4.31
10.7
18.9
1.23
1.2
0.55
With
in-s
ubje
ct e
ffect
sIn
duct
ion
11.
812
.13*
*7.
572.
451.
250.
250.
780.
230.
150.
314.
385.
22*
0.1
0.23
Ind
x G
en4
0.17
1.19
3.92
1.27
1.83
0.37
4.1
1.22
0.12
0.24
0.35
0.42
0.09
0.21
Erro
r15
0.14
3.08
4.95
3.37
0.51
0.84
0.46
Sid
e st
olon
s le
ngth
R
amet
num
ber
side
sto
lons
S
ourc
eM
ain
stol
on
le
ngth
Ram
et n
umbe
r m
ain
stol
on4t
h ra
met
le
af a
rea
4th
ram
et
pe
tiole
leng
thS
ide
stol
ons
num
ber

Costs & benefits of induced resistance in a clonal plant network
87
allocation shift may enable plants to better cope with current and future herbivory by reducing resource
allocation to those organs that are not currently impacted by herbivore damage. While potentially
beneficial in the short term, this response might result in indirect costs in the longer term due to reduced
performance under certain environmental conditions, such as drought, root herbivory and severe root
competition. A similar shift in the biomass allocation pattern was observed in Lepidium virginicum plants
after defense induction. Induced plants grown in high density showed a reduction in root biomass and an
increase in aboveground biomass (Agrawal 2005). In agreement with our findings, total biomass
production was not significantly altered by defense induction in that study. A reduction in below-ground
biomass was also reported for induced wild parsnip plants. In this case, however, the above-ground
biomass did not change significantly after defense induction (Zangerl et al. 1997). Further studies are
necessary to assess the generality, functional significance (including costs and benefits) and mechanistic
basis of changes in root-shoot allocation in response to induced resistance to herbivory.
Reduction in developmental growth rate. Defense induction negatively affected plant fitness by reducing
the number of ramets produced. This delayed developmental growth rate was expressed as a reduction in
the number of ramets on the main stolon produced during the experiment (7.4 ramets on the control and
7.0 on the induced plants). In the short run (i.e. time span of this experiment) this effect is unlikely to
translate into biomass differences. In time, however, subtle changes in the developmental growth rate are
known to result in very major divergences in performance, structure and clonal fitness of stoloniferous
plants (Birch and Hutchings 1992a; Birch and Hutchings 1992b; Huber and Stuefer 1997; Stuefer and
Huber 1998).
Reduction in petiole length. Defense induction had significant negative effects on petiole length. This
effect can have severe performance and fitness consequences for stoloniferous plants like T. repens,
which often grows in dense herbaceous canopies, and which relies on petiole elongation for shade-
avoidance (Huber 1997). Petiole length largely determines the ability of stoloniferous plants to place their
leaves higher up in the canopy (Huber and Wiggerman, 1997; Wijschedé et al. 2006). Even a small
reduction in petiole length could have serious performance costs since differences in the relative position
of leaves in herbaceous canopies are likely to be amplified by asymmetric competition for light (Weiner
1990, Pierik et al. 2003). Defense induction may also cause physiological trade-offs which impede the
simultaneous expression of plasticity to herbivores and to shading by competitors (Cipollini 2004). A
decrease in petiole length as a result of defense induction can hence, compromise the competitive ability
of plants and result in an enhanced risk of induced plants being over-shaded by neighbors. A recent study
by Kurashige and Agrawal (2005) supports this notion by showing that Chenopodium album plants, which
had previously been damaged by herbivores, were able to elongate stems to a similar proportional degree
as undamaged plants when grown in competition for light. However, the damaged plants were smaller due
to the expression of induced resistance, thereby incurring potential opportunity costs due to asymmetric
competition.

Chapter 6
88
Benefits of Systemic Induced Resistance
Reduced damage. Our results provide direct evidence
for short term benefits of having an early-warning
system in clonal plant networks. In the presence of
herbivores, induced plants suffered considerably less
damage than control plants. As many as 50% fewer
ramets were attacked in induced plants as compared
to controls. Localized damage (defense-induction
treatment) resulted in a greater degree of protection
against herbivores for ramets further along that main
stolon and its side branches. The reduced damage did
not translate into a significant effect of defense
induction on biomass production, due to the fact that
the youngest, usually not fully developed leaves were
heavily preferred by the herbivores. The biomass loss
due to young leaf consumption is very likely to strongly underestimate the negative effects of herbivory
and defense induction on future plant growth and performance. Coleman and Leonard (1995)
demonstrated how leaf area consumption, and its consequences for plant performance, can be severely
underestimated if the developmental stage of leaves is not taken into account. They showed that a certain
amount of damage inflicted on young expanding Nicotiana tabaccum leaves is more detrimental than the
same amount received by mature, fully developed leaves. As leaf tissue expanded, the area of the holes
increased almost 4 times and the final area of the leaf decreased by approximately 40%. In addition, they
observed a 35% decrease in the number and mass of the fruits on the plants that received the damage on
expanding young tissues. Therefore, an initially small amount of damage inflicted on young developing
leaves may have dramatic consequences for plant performance and fitness over time. Similarly, the
differences found in our experiment can be expected to result in considerable performance differences
between induced and uninduced plants as an increased damage and loss of young leaves in uninduced
plants will compromise plant productivity by reducing the number of future source ramets.
Our results show that ramet age largely determines herbivore damage. The first and second ramets
were heavily attacked as compared to the rest. This damage, although still large, was significantly reduced
in induced plants. The reduction in leaf area loss in induced young ramets likely increases their chance of
survival and establishment. Young ramets in clonal plants constitute the most valuable tissue since they
represent the future reproductive potential of the plant (Huber and During 2000) and their protection is
critical since they are responsible for a high proportion of the future biomass production (Beinhart 1963).
We present evidence supporting the hypothesis that an early-warning system after herbivory in a clonal
plant network grants vulnerable young offspring ramets with parental support (Stuefer et al. 2004) that
non-clonal plants are unable to confer their offspring at the moment of the attack (but see Agrawal et al.
1999b).
TABLE 5. Benefits of systemic induced resistance. Doubly repeated measures ANOVA for effects of genotype, defense induction and ramet age on leaf area loss due to herbivory.
P < 0.0001***, 0.001< P <0.01 **, 0.01< P < 0.05 *, 0.1 > P > 0.05 $
Source df MS F
Between subjects effectsGenotype (Gen) 4 1102 2.39 $
Error 15 461Within subjects effects
Induction (Ind) 1 6847 63.92 ***Ind x Gen 4 244 2.28
Error 15 107Age 5 18988 133.0 ***
Age x Gen 20 630 4.41 ***Error 75 142
Ind x Age 5 125 1.22Ind x Age x Gen 20 172 1.67$
Error 75 103

Costs & benefits of induced resistance in a clonal plant network
89
Our study provides evidence for significant costs and benefits of systemic defense induction in T.
repens. The experimental approach used in this study, however, does not allow for balancing costs and
benefits in terms of plant fitness and overall plant performance, because both positive and negative effects
of induction reported here, although likely to have significant longer-term effects on productivity and
ultimately on fitness, did not have an effect on biomass at the short time scale during which the
experiment took place. While our results indicate clear advantages and disadvantages of network
induction in the subsequent presence and absence of herbivores, respectively, an accurate and reliable
quantification of the cost-benefit ratio should make use of long-term experiments and different ecological
settings.
In conclusion, the present study shows that in the short term, the activation of early-warning
responses in clonal plant networks has both costs and benefits. In the absence of herbivores, the
performance of the induced phenotype was compromised as compared to the uninduced phenotype in
terms of potential competitive ability. In the presence of herbivores, the induced phenotype was favored by
suffering considerably less herbivore damage suggesting potential advantages for the phenotype correctly
matching its environment. Whether this represents an adaptive value of the induced responses remains to
be demonstrated in longer term studies where the initial small changes observed in our study can be
measured directly in terms of fitness. The long term balance of costs and benefits of induced resistance in
clonal plant networks is likely to be strongly context-dependent and a function of the match between
spatio-temporal aspects of systemic defense expression and the feeding behavior of herbivores.
Acknowledgements: We thank P. Walker, C. Jansen, T. van Mölken, H. de Caluwe, X. Chen, J.
Weijschedé, A. Smit-Tiekstra, P. Rodersman and T. Koubek for practical assistance, and H. de Kroon, T.
van Mölken, and N. van Dam for useful comments on a previous version of this manuscript. This study
was funded by the Netherlands Organization for Scientific Research (NWO, VIDI-fellowship to JFS). VL
was funded by the Czech Science Foundation, grant nr. 206/03/H034.

Chapter 6
90

Sara Gómez
CHA
PTER7
Summarizing Discussion

Chapter 7
92
SUMMARIZING DISCUSSION
Plants can activate their defense system in response to damage. This can take place locally at the site of
attack, but it can also be activated systemically in distant, undamaged parts of the plant. In this thesis I
have investigated the systemic expression of induced defense not within plant individuals but between
connected individuals of the clonal plant Trifolium repens. The research presented in this thesis sheds
light on the mechanisms and ecological implications of sharing defense agents in clonal plant networks.
Evidence is provided showing that members of the stoloniferous herb Trifolium repens are able to share
information about herbivore attacks. The defense response in undamaged members consists of changes
in nutrition, biomechanics, morphology and chemistry of leaves, which results in a reduced preference for
induced ramets by herbivores. I propose that an early warning system confers advantages for clonal
plants owing to the use of private channels for specific information exchange among network members
only. This internal warning system minimizes the risk of information abuse by neighboring competitors and
ensures parental support to young ramets which develop during periods of herbivore presence.
In this thesis I also provide experimental evidence for constraints and costs associated with the
early warning system in T.repens. Because benefits, costs and constraints are strongly dependent on
species-specific physiological, vascular and architectural traits, different clonal plant species are predicted
to exhibit different induced defense expression. In the following sections I will discuss the main findings of
this research project with regard to processes, patterns and implications of ramet-to-ramet communication
in networks of the stoloniferous herb Trifolium repens.
Early warning systems in clonal plant networks Whilst there is a wealth of published information regarding the principles, mechanisms and constraints of
resource transport between ramets of clonal plants (reviewed in Pitelka and Ashmun 1985; Marshall 1990;
Alpert 1996; Alpert and Stuefer 1997), the transport of non-resource substances such as defense signals
has been largely overlooked. As described in chapter 2, the main aim of this thesis was to investigate
whether stolon connections can provide means for internal transport of defense agents between
connected ramets. The experiments in chapter 3 provide conclusive evidence that members of a clonal
plant network can activate an early warning system in response to local herbivory. I have shown that the
transport of the unknown induction signal takes place internally following phloem source-sink dynamics
(Figure 1). This provides clonal plant networks with a fast, reliable and flexible defense system.
Clonal growth allows for fast local propagation, circumvents the vulnerable seedling stage, spreads
the risk of genet mortality over numerous ramets and allows for the sharing of resources in heterogeneous
habitats (Buss et al. 1985; van Groenendael and de Kroon 1990; de Kroon and van Groenendael 1997).
Furthermore, Kolar and Lodge (2001) identified asexual reproduction as a trait that increases the
probability of spread and invasion success of exotic plant species. Early warning in response to local
herbivory, as shown in this thesis, might also play an important role for the ecological (and invasive)
success of stoloniferous and rhizomatous species.

Summarizing discussion
93
Early warning among ramets of clonal plant networks has its limitations. Phloem transport of the
induction signal through physiological integration implies that defense expression is constrained by factors
controlling phloem flows. In Trifolium repens, phloem flows move mainly acropetally (Marshall 1990;
Kemball and Marshall 1995). This unidirectional vascular flow entails that after local damage inflicted on a
ramet, defense expression is restricted to ramets which are located further towards the stolon tip (chapter 3; experiments 1 and 2a). This can affect many or only a few other ramets, depending on the location of
the initial attack. Under undisturbed conditions, the internal transport of the defense agents results in a
highly predictable, acropetal expression of inducible defense. Herbivores may therefore develop an
adaptive behavior to avoid the negative effects of defense induction by feeding exclusively on the
youngest tissues, or by severing stolons before consuming the ramet. Phloem flows are quite flexible in T.
repens and can hence be modified by local conditions that influence source-sink dynamics such as light
availability (chapter 3; experiment 2b). Small-scale patterns in resource availability can therefore be
expected to have profound implications for the expression pattern of the systemic defense response.
Highly sectorial T. repens clones show restricted vascular contacts between groups of ramets
belonging to the same network (Lötscher and Hay, 1996; Marshall and Price 1997). Consequently,
resources or other substances that rely on the vascular system for their transport cannot be shared
among ramets belonging to different orthostichies. The limitations imposed by plant vasculature in the
expression of induced defenses have been shown in several non-clonal systems such as poplar (Jones et
al. 1993), tomato (Orians et al. 2000), tobacco (Schittko and Baldwin 2003) and bittersweet nightshade
(Viswanathan and Thaler 2004). Sectoriality may act as a source of variation in the expression pattern of
defense and thus, reduce the predictability of the spatio-temporal defense expression.
Despite of the advantages of being connected, there are also some associated risks. Stolons
provide internal transport means for defense signaling, but they can also provide physical bridges for
foraging herbivores to find new hosts of the same genetic identity. This potentially makes local damage a
very reliable cue for indicating future damage. Pathogens such as viruses might also take advantage of
the connections between ramets. They can disperse throughout the network making use of the vascular
system, posing a serious risk for other network members (T. van Mölken and J.F. Stuefer, unpublished
data).
Time is precious: temporal dynamics of induced resistance in clonal plants Inducible defense systems may optimize the allocation of resources to various key functions by adjusting
the plant’s phenotype to current needs. This plastic response allows plants to withstand unpredictable
herbivore attacks without investing in costly constitutive defense structures. However, the activation and
decay of induced defenses is not an immediate process (Schultz 1988; Underwood 1998). Plants must
fine-tune the timing of induced defenses in order to maximize the efficiency of protection against current
and near-future enemies and minimize the costs of the temporary investment in defense. The length and
strength of the response will ultimately depend upon the balance between the costs of producing
defensive compounds and the benefits yielded by accurate predictability of future damage.

Chapter 7
94
Due to the extensive areas clonal plants can cover (Cook 1985), it is important to understand the
spatio-temporal dynamics of induced responses to assess the ecological significance of early warning
systems. In chapter 4 of this thesis, I studied the temporal dynamics of induced defense in clonal
fragments. The experiments showed that it takes about 1.5 to 2 days to activate systemic defense in a
ramet that is positioned adjacent to the damaged ramet. This activation time comprises the initial phloem
loading time (Babst et al. 2005), long-distance transport time, and the time necessary to build up defense
in the undamaged leaf (Baldwin et al. 1994). Phloem sap moves at a rate of ca. 1-2 cm/min in herbaceous
C3 plants (P. Minchin, pers. communication). This suggests that the defense signal likely reaches distant
members rather fast, with the defense activation time being mainly controlled by the time of phloem
loading and defense expression in the target ramet. Therefore, defense induction in distant ramets is
predicted to take place relatively faster than observed in the ramet adjacent to the attacked ramet.
Ultimately, the efficiency of the response can only be assessed in relation to the temporal dynamics of the
herbivore. In other words, the defense expression must be faster than the feeding speed of the herbivore
within the network.
Chapter 3 showed that systemic defense is only expressed in an acropetal direction. The choice
tests of chapter 5 indicated that herbivores have a strong preference for young ramets. This means that
young ramets are very likely to be attacked first. Due to physiological constraints associated with source-
sink dynamics (see previous section) no other ramet will be warned (Figure 1). But, is systemic induction
of defense then a beneficial strategy? The answer to this question is implicit in the results of chapter 4,
which provide evidence that newly formed ramets (not present at the time of attack) also become defense-
induced. This is a great advantage for clonal plants since the young ramets are in need of protection due
to their vulnerability and fitness value. Non-clonal plants lack the ability of conferring such parental support
to their offspring (but see Agrawal et al. 1999b). The temporary protection of the newly produced ramets
may last long enough for the developing ramet to escape the vulnerable time window. Early warning
systems can hence be considered a functional strategy in clonal plants regardless of the initial point of
attack.
Induced-defense: killing me strongly Plants have a wide array of defenses to protect themselves against their enemies. These range from
passive defense, such as low nutritional value, to more active defenses such as the production of toxic
metabolites or antimetabolic proteins and physical structures such as thorns and trichomes.
Biomechanical properties have often been studied from the point of view of constitutive defenses (Feeny
1976, Coley 1983, Choong 1996, Lucas et al. 2000), while their role as an inducible response is usually
neglected. Most studies examining induced defenses focus on chemical defense, possibly due to the
notion that mechanical changes are slow responses. This, however, is only partially true (e.g., inducible
formation of thorns or trichomes; Karban and Baldwin 1997). Other biochemical traits, such as leaf
strength, toughness and thickness may change on a much shorter time scale. In chapter 5, I showed that

Summarizing discussion
95
significant changes in biochemical leaf properties can occur as fast as within 4 days, and that those
changes are not restricted to developing leaves but also take place in mature ramets.
The mechanisms allowing for such rapid changes in the material properties of the leaves remain
unknown (chapter 5). Cell wall content did not explain the plastic changes in biomechanical leaf trait after
defense induction. Changes in cell wall chemistry rather than total cell wall content might be responsible
for an increase in leaf strength. This view is indirectly supported by the observed change in the phenolic
composition of leaf tissue. A major group of the identified phenolic compounds discriminating between
induced and control leaves belong to hydroxycinnamic acid derivatives, which may include compounds
related to cell wall structural properties. Factors such as changes in leaf turgor pressure after herbivory
may also be involved in the observed increase in leaf strength.
Inducible changes of leaf biomechanical traits may play an important defense role in particular
against young instar larvae and other small herbivores that are sensitive to leaf surface resistance, such
as aphids or mites (Jiang and Ridsdill-Smith 1996; Alvarez 2007; Figure 2). It is unknown whether
changes in leaf biomechanics are reversible and whether they have negative consequences for the
development of young leaves, for example by constraining leaf expansion, which may result in a reduced
final photosynthesizing area.
HERBIVORE ENEMIESATTRACTION
INTERNAL INTRANET COMMUNICATION
EXTERNAL INTRANET COMMUNICATION
(PRIMING)
HERBIVORE ENEMIESATTRACTION
INTERNAL INTRANET COMMUNICATION
EXTERNAL INTRANET COMMUNICATION
(PRIMING)
FIGURE 1. Early warning systems in a clonal plant network. This is a schematic model of (hypothetical) defense strategies in a clonal plant network. The black arrows indicate the internal transport of defense agents (following phloem source-sink dynamics) as shown in chapter 3. This model includes two possible roles of volatile organic compounds (VOCs) in clonal plant networks: a) attraction ofnatural enemies (wavy solid lines) of the herbivores by systemically releasing VOCs from multiple ramets coupled with a decreasingconcentration gradient and/or volume with increasing distance from the point of attack (block arrows in gray scale); and b) external intra-clonal communication to overcome physical constraints imposed by the clone vascular flow dynamics or architecture (wavy dashed lines). Defense-induced ramets are represented in black and uninduced ramets in white.
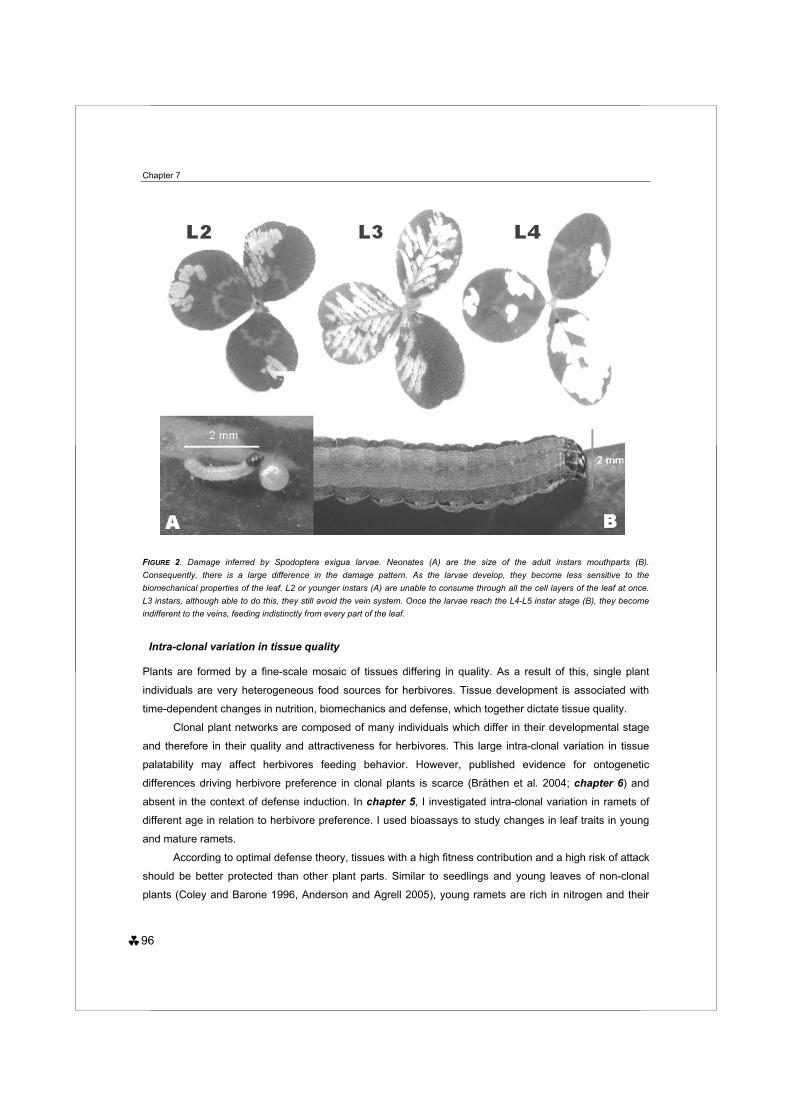
Chapter 7
96
Intra-clonal variation in tissue quality
Plants are formed by a fine-scale mosaic of tissues differing in quality. As a result of this, single plant
individuals are very heterogeneous food sources for herbivores. Tissue development is associated with
time-dependent changes in nutrition, biomechanics and defense, which together dictate tissue quality.
Clonal plant networks are composed of many individuals which differ in their developmental stage
and therefore in their quality and attractiveness for herbivores. This large intra-clonal variation in tissue
palatability may affect herbivores feeding behavior. However, published evidence for ontogenetic
differences driving herbivore preference in clonal plants is scarce (Bråthen et al. 2004; chapter 6) and
absent in the context of defense induction. In chapter 5, I investigated intra-clonal variation in ramets of
different age in relation to herbivore preference. I used bioassays to study changes in leaf traits in young
and mature ramets.
According to optimal defense theory, tissues with a high fitness contribution and a high risk of attack
should be better protected than other plant parts. Similar to seedlings and young leaves of non-clonal
plants (Coley and Barone 1996, Anderson and Agrell 2005), young ramets are rich in nitrogen and their
© M
. Dor
enbo
sch
FIGURE 2. Damage inferred by Spodoptera exigua larvae. Neonates (A) are the size of the adult instars mouthparts (B). Consequently, there is a large difference in the damage pattern. As the larvae develop, they become less sensitive to the biomechanical properties of the leaf. L2 or younger instars (A) are unable to consume through all the cell layers of the leaf at once. L3 instars, although able to do this, they still avoid the vein system. Once the larvae reach the L4-L5 instar stage (B), they become indifferent to the veins, feeding indistinctly from every part of the leaf.

Summarizing discussion
97
biomechanical properties are poorly developed (chapter 5). This makes young tissues both very
appealing and vulnerable to herbivores (Coley 1983, Fenner et al. 1999 but see de Boer 1999 and van
Dam et al. 1995 for exceptions). Young ramets have a high photosynthetic potential and are responsible
for the production of a large part of the future offspring (Beinhart 1963). This makes them important
contributors of the clone’s future performance and fitness. Therefore, they should be highly protected.
In contrast to those expectations, the experiments in chapter 5 revealed that young ramets are
highly preferred over mature ramets. Strong herbivory pressure on certain plant parts (e.g. fruits and
young tissues) or plants in certain developmental stages (e.g. seedlings) can compromise plant perfor-
mance, and may ultimately affect population and community dynamics (Halpern and Underwood 2006).
However, in defense-induced clonal fragments, the damage inferred to young ramets was significantly
reduced, resulting in a higher level of damage incurred by the mature ramets. These results suggest that
the defense of young ramets is strongly up-regulated in the presence of herbivores, potentially allowing
the investment of resources into their fast growth when defense is not needed. A fast development of
young ramets (T. repens: approximately 7 days at 24°C, Carlson 1966) might be an alternative strategy
for ramets to reduce the risk of herbivory because swift growth reduces the vulnerability period (Ernest
1989; Aide and Londoño 1989; Coley and Barone 1996). Once ramets reach maturity, herbivore damage
is dramatically reduced, often remaining below 5% of the total leaf area (chapter 6).
Induced defense as a risk-spreading strategy Herbivore attack leads to alterations in tissue quality both locally and systemically in undamaged parts of
the plant (Karban and Baldwin 1997). Induced responses therefore become an additional source of within-
plant variation in tissue quality (Stout et al. 1996a, Shelton 2005). In chapter 5 I provide evidence
indicating that systemic induced resistance simultaneously increases and decreases intra-clonal variation.
After local damage, variation in tissue quality may increase due to constraints imposed by source-sink
dynamics, resulting in groups of defense-induced and uninduced ramets in the same clone (Figure 1).
Within-plant variation in terms of tissue quality has been shown to be a hindering factor for the
performance and survival of some herbivores (Berenbaum and Zangerl 1993, Stockhoff 1993, Anderson
and Agrell 2005, Rodriguez-saona and Thaler 2005). However, systemic induction of resistance can also
decrease intra-clonal variation by making both mature and young ramets equally unpalatable, thereby
acting as a risk-spreading strategy among groups of ramets and enhancing the protection of the otherwise
more vulnerable young members of the network (chapter 5).
Protection has its price: costs & benefits of induced resistance The induction of defense has clear benefits, such as reduced damage, increased survival and improved
competitive ability in the presence of herbivores (Karban and Baldwin 1997, Tollrian and Harvell 1999).
However, defense also involves costs. These can become visible directly as a decrease in resource
investment to growth or reproduction, and indirectly via complex biotic and abiotic interactions such as

Chapter 7
98
reduced competitive ability or lower number of visits by pollinators (Bergelson and Purlington 1996;
Strauss et al. 2002; Heil and Baldwin 2002).
Clonal plant networks can be formed by hundreds of members, and a single clone can cover
hundreds of square meters or even several hectares (Cook 1985). Large scale networks could experience
considerable costs if many interconnected members of the network become defense-induced. In chapter 6, I investigated the costs and benefits of defense-induction by growing uninduced and defense-induced
clonal fragments together in the presence and absence of herbivores, respectively. The results showed
that the activation of early warning systems in clonal networks have costs when the plants grow without
herbivores. This resembles a scenario in natural conditions when there is a mismatch in the defense
expression and subsequent herbivory. The costs were expressed as fewer ramets being produced on the
main stolon, reduced petiole length and a shift in dry mass allocation towards the leaves in defense-
induced plants. The benefits consisted of markedly reduced herbivory damage in the presence of
herbivores. Twice as many uninduced than induced ramets were damaged and the damage was up to
50% larger in uninduced as compared to defense-induced ramets.
Perspectives for future research: role of volatiles in intranet communication As described in the previous sections, members of clonal plant networks can use private internal channels
to share information about herbivore attacks. This is a beneficial feature that may allow members of the
network to counterattack the herbivore’s offense as a group rather than individually. In contrast, non-clonal
plants are restricted to using external channels for the exchange of information when they are under
herbivore attack. After damage, plants release info-chemicals into their environment such as volatile
organic compounds (VOCs; Dicke and van Loon 2000; Pichersky and Gershenzon 2002), which mediate
complex interactions between plants, herbivores and higher trophic levels (Soler et al. 2005, Snoeren et
al. 2007). This type of plant-plant communication may allow non-clonal neighbors profit from a joint
warning system. However, the release of info-chemicals by the damaged plant may have negative
consequences if it is used as a cue by direct competitor plants (i.e., eavesdropping). The eavesdroppers
may activate their defense system before they become attacked (Karban et al. 2003; 2004), potentially
gaining competitive ability through an increased performance in the presence of herbivores. Clonal plants
may minimize eavesdropping by using internal intranet communication.
Nevertheless, the release of VOCs by damaged plants may confer additional advantages in the
struggle against herbivores. VOCs emitted by damaged plants are known to attract natural enemies of
herbivores above (Kessler and Baldwin 2001) and below-ground (Rasmann et al. 2005). In clonal plants,
the systemic release of volatiles by multiple members of the network might result in an increased volume
of the warning volatile blend as compared to the volume emitted by one individual non-clonal plant (Figure
1; chapter 2). This may help predators or parasitoids finding a prey patch (Kobayashi and Yamamura
2007). A decreasing concentration gradient with increasing distance from the point of attack, as shown
within individual plants (Dicke 1994/1995), may ease the location of the prey.

Summarizing discussion
99
In addition, as recently suggested (Karban et al. 2006; Heil and Silva Bueno 2007; Frost et al.
2007), VOCs may play a role in intra-plant communication. In analogy with non-clonal plants, VOCs
emission by damaged ramets may allow the alerting of ramets which cannot be reached by internal
defense signaling due to vascular constraints (Figure 1). This would act as a complementing defense
mechanism in addition to the internal, more reliable transport of defense signals. However, further studies
are needed to confirm these hypotheses, opening new avenues for plant defense research in clonal plant
networks.
Concluding remarks This thesis is the first study exploring inducible defense strategies in the context of clonal plant networks. I
have shown that stolon connections play a crucial and novel role in defense signaling between connected
individuals. This advantageous feature allows as of yet undamaged members of the network to undergo
plastic changes that reduce future herbivore damage. Furthermore, it confers young developing ramets
with parental support, increasing their chances of survival.
The experimental approaches developed in this thesis allow the studying of the underlying
principles and constraints of induced defense in clonal plants. As a next step, the generality of early
warning systems in network-forming species needs to be assessed. Additional experiments under natural
conditions will provide deeper insight into the ecological significance of intranet communication.
In conclusion, the study of plant-insect interactions in clonal plant networks adds a new dimension
to the fields of inducible defenses, plant-plant and plant-animal communication. This thesis is a keystone
in the field of defense in clonal plants, providing the basis (in terms of quantitative data and experimental
approaches) for further research into the mechanisms as well as ecological and evolutionary implications
of this interesting phenomenon.

Chapter 7
100

REFEREN
CES

References
102
REFERENCES
Agrawal AA, Gorski PM, Tallamy DW (1999a) Polymorphism in plant defense against herbivory:
Constitutive and induced resistance in Cucumis sativus. Journal of Chemical Ecology 25: 2285-
2304
Agrawal AA, Laforsch C, Tollrian R (1999b) Transgenerational induction of defences in animals and
plants. Nature 401: 60-63
Agrawal AA, Strauss SY, Stout MJ (1999c) Costs of induced responses and tolerance to herbivory in male
and female fitness components of wild radish. Evolution 53: 1093-1104
Agrawal AA Tuzun S, Bent E (1999d) Induced plant defenses against pathogens and herbivores:
biochemistry, ecology and agriculture. APS Press
Agrawal AA (2000) Benefits and costs of induced plant defense for Lepidium virginicum (Brassicaceae).
Ecology 81: 1804-1813
Agrawal AA (2005) Future directions in the study of induced plant responses to herbivory. Ent Exp App
115: 97-105
Agrell J, Oleszek W, Stochmal A, Olsen M, Anderson P (2003) Herbivore-induced responses in alfalfa
(Medicago sativa). J Chem Ecol 29: 303-320
Aide TM, Londono EC (1989) The effects of rapid leaf expansion on the growth and survivorship of a
lepidopteran herbivore. Oikos 55: 66-70
Akhtar Y, Isman MB (2004) Comparative growth inhibitory and antifeedant effects of plant extracts and
pure allelochemicals on four phytophagous insect species. J App Ent 128: 32-38
Alborn HT, Rose USR, McAuslane HJ (1996) Systemic induction of feeding deterrents in cotton plants by
feeding of Spodoptera spp larvae. J Chem Ecol 22: 919-932
Alpert P (1991) Nitrogen sharing among ramets increases clonal growth in Fragaria chiloensis. Ecology
72: 69-80
Alpert P (1996) Nutrient sharing in natural clonal fragments of Fragaria chiloensis. J Ecol 84: 395-406
Alpert P, Stuefer JF (1997) Division of labour in clonal plants. In: de Kroon H and van Groenendael J
(eds.) The ecology and evolution of clonal plants. Backhuys Publishers, The Netherlands, pp. 137-
154

References
103
Alpert P, Holzapfel C, Slominski C (2003) Differences in performance between genotypes of Fragaria
chiloensis with different degrees of resource sharing. J Ecol 91: 27-35
Alvarez AE (2007) Resistance mechanisms of Solanum species to Myzus persicae. PhD thesis.
Wageningen University, The Netherlands
Alves MN, Sartoratto A, Trigo JR (2007) Scopolamine in Brugmansia suaveolens (Solanaceae): defense,
allocation, costs, and induced response. J Chem Ecol 33: 297-309
Anderson P, Jonsson M, Morte U (2001) Variation in damage to cotton affecting larval feeding preference
of Spodoptera littoralis. Ent Exp App 101: 191-198
Anderson P, Agrell J (2005) Within-plant variation in induced defence in developing leaves of cotton
plants. Oecologia 144: 427-434
Aranwela N, Sanson G, Read J (1999) Methods of assessing leaf-fracture properties. New Phytol 144:
369-383
Arnold T, Appel H, Patel V, Stocum E, Kavalier A, Schultz J (2004) Carbohydrate translocation determines
the phenolic content of Populus foliage: a test of the sink-source model of plant defense. New
Phytol 164: 157-164
Augner M (1995) Low nutritive quality as a plant defense - Effects of herbivore-mediated interactions. Evol
Ecol 9: 605-616
Babst BA, Ferrieri RA, Gray DW, Lerdau M, Schlyer DJ, Schueller M, Thorpe MR, Orians CM (2005)
Jasmonic acid induces rapid changes in carbon transport and partitioning in Populus. New Phytol
167: 63-72
Baldwin IT, Schmelz EA, Ohnmeiss TE (1994) Wound-induced changes in root and shoot jasmonic acid
pools correlate with induced nicotine synthesis in Nicotiana sylvestris spegazzini and comes. J
Chem Ecol 20: 2139-2157
Baldwin IT (1998) Jasmonate-induced responses are costly but benefit plants under attack in native
populations. PNAS 95: 8113-8118
Baur R, Binder S, Benz G (1991) Nonglandular leaf trichomes as short-term inducible defense of the gray
alder, Alnus incana L, against the chrysomelid beetle, Agelastica alni L. Oecologia 87: 219-226
Beinhart G (1963) Effects of environment on meristematic development, leaf area, and growth of white
clover. Crop Sci 3: 209-213

References
104
Berenbaum MR, Zangerl AR (1993) Furanocoumarin metabolism in Papilio polyxenes - Biochemistry,
genetic variability, and ecological significance. Oecologia 95: 370-375
Bergelson J (1994) The effects of genotype and the environment on costs of resistance in lettuce. Am Nat
143: 349-359
Bergelson J, Purrington CB (1996) Surveying patterns in the cost of resistance in plants. Am Nat 148: 536-
558
Bernays EA (1997) Feeding by lepidopteran larvae is dangerous. Ecological Entomology 22: 121-123
Bernays EA (1998) Evolution of feeding behavior in insect herbivores. Bioscience 48: 35-44
Bi JL, Felton GW, Murphy JB, Howles PA, Dixon RA, Lamb CJ (1997) Do plant phenolics confer
resistance to specialist and generalist insect herbivores? J Agric Food Chem 45: 4500-4504
Biere A, Marak HB, van Damme JMM (2004) Plant chemical defense against herbivores and pathogens:
generalized defense or trade-offs? Oecologia 140: 430-441
Birch CPD, Hutchings MJ (1992a) Stolon growth and branching in Glechoma hederacea L - An application
of a plastochron index. New Phytol 122: 545-551
Birch CPD, Hutchings MJ (1992b) Analysis of ramet development in the stoloniferous herb Glechoma
heredacea using a plastochron index. Oikos 63: 387-394
Bluthgen N, Metzner A (2007) Contrasting leaf age preferences of specialist and generalist stick insects
(Phasmida) Oikos 116: 1853-1862
Brathen KA, Agrell J, Berteaux D, Jonsdottir IS (2004) Intraclonal variation in defence substances and
palatability: a study on Carex and lemmings. Oikos 105: 461-470
Bruin J, Sabelis MW, Dicke M (1991). Mite herbivory induces airborne information transfer between
plants. In: Dusbabek F, Bukva V (eds.) Modern acarology. Academia, Prague and SPB Academic
Publishing, The Hague, pp. 599-605
Bruin J, Dicke M, Sabelis MW (1992) Plants are better protected against spider-mites after exposure to
volatiles from infested conspecifics. Experientia 48: 525-529
Bruin J, Dicke M (2001) Chemical information transfer between wounded and unwounded plants: backing
up the future. Biochem Syst Ecol 29: 1103-1113

References
105
Buss LW, Cook RE, Jackson JBC (1985) Population biology and evolution of clonal organisms. Yale
University Press
Cain ML (1994) Consequences of foraging in clonal plant species. Ecology 75: 933-944
Caraco T, Kelly CK (1991) On the adaptive value of physiological integration in clonal plants. Ecology 72:
81-93
Carlson GE (1966) Growth of clover leaves - Developmental morphology and parameters at then stages.
Crop sci 6: 293-294
Chamberlain K, Guerrieri E, Pennacchio F, Pettersson J, Pickett JA, Poppy GM, Powell W, Wadhams LJ,
Woodcock CM (2001) Can aphid-induced plant signals be transmitted aerially and through the
rhizosphere? Biochem Syst Ecol 29: 1063-1074
Chapman DF, Robson MJ, Snaydon RW (1990a) Short-term effects of manipulating the source - sink ratio
of white clover (Trifolium repens) plants on export of carbon from, and morphology of, developing
leaves. Physiol Plant 80: 262-266
Chapman DF, Robson MJ, Snaydon RW (1990b) The carbon economy of developing leaves of white
clover (Trifolium repens L). Ann Bot 66: 623-628
Chapman DF, Robson MJ, Snaydon RW (1991) The influence of leaf position and defoliation on the
assimilation and translocation of carbon in white clover (Trifolium repens L.). 2. Quantitative carbon
movement. Ann Bot 67: 303-308
Chapman DF, Robson MJ, Snaydon RW (1992a) Physiological integration in the clonal perennial herb
Trifolium repens L. Oecologia 89: 338-347
Chapman DF, Robson MJ, Snaydon RW (1992b) The carbon economy of clonal plants of Trifolium repens
L. J Exp Bot 43: 427-434
Chen SX, Petersen BL, Olsen CE, Schulz A, Halkier BA (2001) Long-distance phloem transport of
glucosinolates in Arabidopsis. Plant Physiol 127: 194-201
Cheng N-H, Su C-L, Carter SA, Nelson RS (2000) Vascular invasion routes and systemic accumulation
patterns of tobacco mosaic virus in Nicotiana tabacum. Plant J 23: 349-362
Chia T- F, He J (1999) Photosynthetic capacity in Oncidium (Orchidaceae) plants after virus eradication.
Environ Exp Bot 42: 11-16

References
106
Choong MF (1996) What makes a leaf tough and how this affects the pattern of Castanopsis fissa leaf
consumption by caterpillars. Funct Ecol 10: 668-674
Cipollini D, Purrington CB, Bergelson J (2003) Costs of induced responses in plants. Basic Appl Ecol 4:
79-89
Cipollini D (2004) Stretching the limits of plasticity: Can a plant defend against both competitors and
herbivores?. Ecology 85: 28–37
Clay K (1990) Fungal endophytes of grasses. Annu Rev Ecol Syst 21: 275-297
Coleman JS, Leonard AS (1995) Why it matters where on a leaf a folivore feeds. Oecologia 101: 324-328
Coley PD (1983) Herbivory and defensive characteristics of tree species in a lowland tropical forest. Ecol
Monogr 53: 209-233
Coley PD, Barone JA (1996) Herbivory and plant defenses in tropical forests. Ann Rev Ecol Syst 27: 305-
335
Coley PD, Bateman ML, Kursar TA (2006) The effects of plant quality on caterpillar growth and defense
against natural enemies. Oikos 115: 219-228
Conrath U, Pieterse CMJ, Mauch-Mani B (2002) Priming in plant-pathogen interactions. Trends Pl Sci 7:
210-216
Cook RE (1978) Asexual reproduction: a further consideration. Am Nat 113: 769-772
Cook RE (1985) Growth and development in clonal plant populations In: Jackson JBC, Buss LW, Cook
RE (eds.) Population biology and evolution of clonal organisms Yale University Press, New Haven,
pp. 259-296
Cronin G, Hay ME (1996) Within plant variation in seaweed palatability and chemical defenses: optimal
defense theory versus the growth differentiation balance hypothesis. Oecologia 105: 361-368
Davis JM, Gordon MP, Smit BA (1991) Assimilate movement dictates remote sites of wound-induced
gene-expression in Poplar leaves. PNAS 88: 2393-2396
Day T (2003) Virulence evolution and the timing of disease life-history events. Trend Ecol Evo 18: 113-
118
de Boer NJ (1999) Pyrrolizidine alkaloid distribution in Senecio jacobaea rosettes minimises losses to
generalist feeding. Ent Exp App 91: 169-173

References
107
de Kroon H, Hutchings MJ (1995) Morphological plasticity in clonal plants - The foraging concept
reconsidered. J Ecol 83: 143-152
de Kroon H, van Groenendael J (1997) The ecology and evolution of clonal plants. Backhuys publishers,
Leiden, The Netherlands
de Kroon H, Huber H, Stuefer JF, van Groenendael JM (2005) A modular concept of phenotypic plasticity
in plants. New Phytol 166: 73-82
Dewitt TJ, Sih A, Wilson DS (1998) Costs and limits of phenotypic plasticity. Trend Ecol Evo 13: 77-81.
Dicke M, Bruin J, Sabelis MW (1993a). Herbivore-induced plant volatiles mediate plant-carnivore, plant-
herbivore and plant-plant interactions: talking plants revisited. In Schultz JC, Raskin I (eds.) Plant
signals in interactions with other organisms. Current Topics in Plant Physiology, American Society
of Plant Physiologists Series, pp. 182-196
Dicke M, Vanbaarlen P, Wessels R, Dijkman H (1993b) Herbivory induces systemic production of plant
volatiles that attract predators of the herbivore - Extraction of endogenous elicitor. J Chem Ecol 19:
581-599
Dicke M. (1994/1995). Why do plants talk?. Chemoecology 5/6: 159-165.
Dicke M, van Loon JJA (2000) Multitrophic effects of herbivore-induced plant volatiles in an evolutionary
context. Ent Exp App 97: 237-249
Dicke M, Bruin J (2001) Chemical information transfer between plants: back to the future. Biochem Syst
Ecol 29: 981-994
Dicke M, Dijkman H (2001) Within-plant circulation of systemic elicitor of induced defence and release
from roots of elicitor that affects neighbouring plants. Biochem Syst Ecol 29: 1075-1087
Dicke M, Hilker M (2003) Induced plant defenses: from molecular biology to evolutionary ecology. Basic
App Ecol 4: 3-14
Dirzo R, Harper JL (1982a) Experimental studies on slug-plant interactions - 3. Differences in the
acceptability of individual plants of Trifolium repens to slugs and snails. J Ecol 70: 101-117
Dirzo R, Harper JL (1982b) Experimental studies on slug-plant interactions - 4. The performance of
cyanogenic and acyanogenic morphs of Trifolium repens in the field. J Ecol 70: 119-138

References
108
Dolch R, Tscharntke T (2000) Defoliation of alders (Alnus glutinosa) affects herbivory by leaf beetles on
undamaged neighbours. Oecologia 125: 504-511
Dudley SA, Schmitt J (1996) Testing the adaptive plasticity hypothesis: Density-dependent selection on
manipulated stem length in Impatiens capensis. Am Nat 147: 445-465
Dudt JF, Shure DJ (1994) The influence of light and nutrients on foliar phenolics and insect herbivory.
Ecology 75: 86-98
Edwards PJ, Wratten SD, Parker EA (1992) The ecological significance of rapid wound-induced changes
in plants - Insect grazing and plant competition. Oecologia 91: 266-272
Elle E, Van Dam NM, Hare JD (1999) Cost of glandular trichomes, a "resistance" character in Datura
wrightii Regel (Solanaceae). Evolution 53: 22-35
Eriksson O, Jerling L (1990) Hierarchical selection and risk spreading in clonal plants. In van Groenendael
J, de Kroon H (eds.) Clonal growth in plants: regulation and function. SPB Academic publishing, The
Hague, pp. 79-94
Ernest KA (1989) Insect herbivory on a tropical understory tree - Effects of leaf age and habitat. Biotropica
21: 194-199
Evans JP (1991) The effect of resource integration on fitness related traits in a clonal dune perennial,
Hydrocotyle bonariensis. Oecologia 86: 268-275
Evans JP (1992) The effect of local resource availability and clonal integration on ramet functional-
Morphology in Hydrocotyle bonariensis. Oecologia 89: 265-276
Feeny P (1976) Plant apparency and chemical defense. In Wallace JW, MansellRL (eds.) Biochemical
interaction between plants and insects. Plenum Press, New York and London, pp. 1-36
Fenner M, Hanley ME, Lawrence R (1999) Comparison of seedling and adult palatability in annual and
perennial plants. Funct Ecol 13: 546-551
Frantzen J (1994) The role of clonal growth in the pathosystem Cirsium arvense - Puccinia punctiforis.
Can J Bot 72: 832-836
Frost CJ, Appel M, Carlson JE, De Moraes CM, Mescher MC, Schultz JC (2007) Within-plant signalling via
volatiles overcomes vascular constraints on systemic signalling and primes responses against
herbivores. Ecol Lett 10: 490-498

References
109
Funayama S and Terashima I. (1999) Effects of geminivirus infection and growth irradiance on the
vegetative growth and photosynthetic production of Eupatorium makinoi. New Phytol 142: 483-494
Garcia-Guzman G, Burdon JJ (1997) Impact of the flower smut Ustilago cynodontis (Ustilaginaceae) on
the performance of the clonal grass Cynodon dactylon (Gramineae). Am J Bot 84: 1565-1571
Gershenzon J (1994) Metabolic costs of terpenoid accumulation in higher-plants. J Chem Ecol 20: 1281-
1328
Gibbs A, Harrison B (1976) Effects of viruses on plants. In Plant virology. Edward Arnold, London, pp. 19-
32
Gomez S, Stuefer JF (2006) Members only: induced systemic resistance to herbivory in a clonal plant
network. Oecologia 147: 461-468
Gomez S, Latzel V, Verhulst YM, Stuerfer JF (2007) Costs and benefits of induced resistance in a clonal
plant network. Oecologia 153: 921-930
Gozzo F (2003) Systemic acquired resistance in crop protection: from nature to chemical approach. Agric
Food Chem 51: 4487-4503
Groppe K, Steinger T, Sanders I, Schmid B, Wiemken A, Boller T (1999) Interaction between the
endophytic fungus Epichloë bromicola and the grass Bromus erectus: effects of endophyte infection,
fungal concentration and environment on grass growth and flowering. Mol Ecol 8: 1827-1835
Gruntman M, Novoplansky A (2004) Physiologically mediated self/non-self discrimination in roots. PNAS
101: 3863-3867
Guerrieri E, Poppy GM, Powell W, Rao R, Pennacchio F (2002) Plant-to-plant communication mediating
in-flight orientation of Aphidius ervi. J Chem Ecol 28: 1703-1715
Halpern SL, Underwood N (2006) Approaches for testing herbivore effects on plant population dynamics.
J Appl Ecol 43: 922-929
Hamilton NRS, Hay MJM (1998) Vascular architecture of a large-leafed genotype of Trifolium repens. Ann
Bot 81: 441-448
Haukioja E, Ruohomaki K, Senn J, Suomela J, Walls M (1990) Consequences of herbivory in the
mountain birch (Betula pubescens Ssp Tortuosa) - Importance of the functional organization of the
tree. Oecologia 82: 238-247

References
110
Haukioja E (1991) The influence of grazing on the evolution, morphology and physiology of plants as
modular organisms. Philos Trans R Soc Lond B Biol Sci 333: 241-247
Hay MJM, Sackville Hamilton R (1996) Influence of xylem vascular architecture on the translocation of
phosphorus from nodal roots in a genotype of Trifolium repens during undisturbed growth. New
Phytol 132: 575-582
Hayden KJ, Parker IM (2002) Plasticity in cyanogenesis of Trifolium repens L.: inducibility, fitness costs
and variable expression. Evol Ecol Res 4: 155-168
Heil M (2001) The ecological concept of costs of induced systemic resistance (ISR). Eur J Plant Pathol
107: 137-146
Heil M (2002) Ecological costs of induced resistance. Curr Opin Plant Biol 5: 345-350
Heil M, Baldwin IT (2002) Fitness costs of induced resistance: emerging experimental support for a
slippery concept. Trends Pl Sci 7: 61-67
Heil M, Silva Bueno JC (2007) Within-plant signaling by volatiles leads to induction and priming of an
indirect plant defense in nature. PNAS 104: 5467-5472
Herms DA, Mattson WJ (1992) The dilemma of plants - to grow or defend. Quart Rev Biol 67: 283-335
Holzapfel C, Alpert P (2003) Root cooperation in a clonal plant: connected strawberries segregate roots.
Oecologia 134: 72-77
Honkanen T, Haukioja E (1998) Intra-plant regulation of growth and plant-herbivore interactions.
Ecoscience 5: 470-479
Honkanen T, Haukioja E, Kitunen V (1999) Responses of Pinus sylvestris branches to simulated herbivory
are modified by tree sink/source dynamics and by external resources. Funct Ecol 13: 126-140
Huber H (1997) Architectural plasticity of stoloniferous and erect herbs in response to light climate. PhD
thesis. Utrecht University, The Netherlands
Huber H, Stuefer JF (1997) Shade-induced changes in the branching pattern of a stoloniferous herb:
functional response or allometric effect? Oecologia 110: 478-486
Huber H, Wiggerman L (1997) Shade avoidance in the clonal herb Trifolium fragiferum: a field study with
experimentally manipulated vegetation height. Plant Ecol 130: 53-62

References
111
Huber H, During HJ (2000) No long-term costs of meristem allocation to flowering in stoloniferous
Trifolium species. Evol Ecol 14: 731-748
Hurlbert SH (1984) Pseudoreplication and the design of ecological field experiments. Ecol Monogr 54:
187-211
Hutchings MJ, Wijesinghe DK (1997) Patchy habitats, division of labour and growth dividends in clonal
plants. Trend Ecol Evo 12: 390-394
Jaremo J, Tuomi J and Nilsson P (1999) Adaptive status of localized and systemic defense responses in
plants. In: Tollrian R, Harvell D (eds) The ecology and evolution of inducible defenses. Princeton
University Press, Princeton, New Jersey, USA, pp. 33-44
Jerling L (1988) Clone dynamics, population dynamics and vegetation pattern of Glaux maritima on a
Baltic sea shore meadow. Vegetatio 74: 171-185
Jiang Y, RidsdillSmith TJ (1996) Antixenotic resistance of subterranean clover cotyledons to redlegged
earth mite, Halotydeus destructor. Ent Exp Appl 79: 161-169
Jones CG, Hopper RF, Coleman JS, Krischik VA (1993) Control of systemically induced herbivore
resistance by plant vascular architecture. Oecologia 93: 452-456
Jones RAC (1992) Further studies on losses in productivity caused by infection of annual pasture legumes
with three viruses. Aust J Agric Res 43: 1229-1241
Karban R, Adler FR (1996) Induced resistance to herbivores and the information content of early season
attack. Oecologia 107: 379-385
Karban R, Baldwin IT (1997). Induced responses to herbivory. The University of Chicago Press, Chicago
Karban R, Agrawal AA, Thaler JS, Adler LS (1999) Induced plant responses and information content
about risk of herbivory. Trend Ecol Evo 14: 443-447
Karban R, Baldwin IT, Baxter KJ, Laue G, Felton GW (2000) Communication between plants: induced
resistance in wild tobacco plants following clipping of neighboring sagebrush. Oecologia 125: 66-71
Karban R, Maron J, Felton GW, Ervin G, Eichenseer H (2003) Herbivore damage to sagebrush induces
resistance in wild tobacco: evidence for eavesdropping between plants. Oikos 100: 325-332
Karban R, Huntzinger M, Mccall AC (2004) The specificity of eavesdropping on sagebrush by other plants.
Ecology 85: 1846-1852

References
112
Karban R, Shiojiri K, Huntzinger M, Mccall AC (2006) Damage-induced resistance in sagebrush: volatiles
are key to intra- and interplant communication. Ecology 87: 922-930
Kause A, Ossipov V, Haukioja E, Lempa K, Hanhimaki S, Ossipova S (1999) Multiplicity of biochemical
factors determining quality of growing birch leaves. Oecologia 120: 102-112
Kelly CK (1995) Thoughts on clonal integration - Facing the evolutionary context. Evol Ecol 9: 575-585
Kemball WD, Palmer MJ, Marshall C (1992) The effect of local shading and darkening on branch growth,
development and survival in Trifolium repens and Galium aparine. Oikos 63: 366-375
Kemball WD, Marshall C (1995) Clonal integration between parent and branch stolons in white clover - A
developmental study. New Phytol 129: 513-521
Kemperman JA, Barnes BV (1976) Clone size in American aspens. Can J Bot 54: 2603-2607
Kempster VN, Scott ES, Davies KA (2002) Evidence for systemic, cross-resistance in white clover
(Trifolium repens) and annual medic (Medicago truncatula var truncatula) induced by biological and
chemical agents. Bio Sci Tec 12: 615-623
Kessler A, Baldwin IT (2001) Defensive function of herbivore-induced plant volatile emissions in nature.
Science 291: 2141-2144
Kobayashi Y, Yamamura N (2007) Evolution of signal emission by uninfested plants to help nearby
infested relatives. Evol Ecol 21: 281-294
Kolar CS, Lodge DM (2001) Progress in invasion biology: predicting invaders. Trend Ecol Evo 16: 199-
204
Korves TM, Bergelson J (2003) A developmental response to pathogen infection in Arabidopsis. Plant
Physiol 133: 339-347
Kurashige NS, Agrawal AA (2005) Phenotypic plasticity to light competition and herbivory in Chenopodium
album (Chenopodiaceae). Am J Botany 92: 21-26
Leisner SM, Turgeon R (1993) Movement of virus and photoassimilate in the phloem: a comparative
analysis. Bio Essays 15: 741-748
Levin DA (1973) Role of trichomes in plant defense. Quart Rev Biol 48: 3-15
Lipsitch M, Moxon ER (1997) Virulence and transmissibility of pathogens: what is the relationship?.
Trends Micr 5: 31-37

References
113
Lively CM (2001) Propagule interactions and the evolution of virulence. J Evol Biol 14: 317-324
Lockwood JR (1998) On the statistical analysis of multiple-choice feeding preference experiments.
Oecologia 116: 475-481
Lotscher M, Hay MJM (1996) Distribution of mineral nutrient from nodal roots of Trifolium repens:
genotypic variation in intra-plant allocation of P-32 and Ca-45. Physiol Plant 97: 269-276
Lotscher M, Hay MJM (1997) Genotypic differences in physiological integration, morphological plasticity
and utilization of phosphorus induced by variation in phosphate supply in Trifolium repens. J Ecol
85: 341-350
Lowell RB, Markham JH, Mann KH (1991) Herbivore-like damage induces increased strength and
toughness in a seaweed. Proc Royal Soc B-Biological Sciences 243: 31-38
Lucas PW, Turner IM, Dominy NJ, Yamashita N (2000) Mechanical defences to herbivory. Ann Bot 86:
913-920
Magori K, Oborny B, Dieckmann U, Meszena G (2003) Cooperation and competition in heterogeneous
environments: the evolution of resource sharing in clonal plants. Evol Ecol Res 5: 787-817
Magyar G, Kun A, Oborny B, Stuefer JF (2007) Importance of plasticity and decision-making strategies for
plant resource acquisition in spatio-temporally variable environments. New Phytol 174: 182-193
Maleck K, Dietrich RA (2003) Defense on multiple fronts: how do plants cope with diverse enemies?.
Trends Pl Sci 4: 215-219
Marshall C (1990) Source-sink relations of interconnected ramets. In: van Groenendael J, de Kroon H
(eds.) Clonal growth in plants: regulation and function. SPB Academic Publishing, The Hague, pp.
23-41
Marshall C, Price EAC (1997) Sectoriality and its implications for physiological integration. In: de Kroon H,
van Groenendael J (eds.) The ecology and evolution of clonal plants. Backhuys publishers, Leiden,
pp. 79-107
Massei G, Hartley SE (2000) Disarmed by domestication? Induced responses to browsing in wild and
cultivated olive. Oecologia 122: 225-231
Massey FP, Hartley SE (2006) Experimental demonstration of the antiherbivore effects of silica in
grasses: impacts on foliage digestibility and vole growth rates. Proc Royal Soc B-Biological
Sciences 273: 2299-2304

References
114
Massey FP, Ennos AR, Hartley SE (2007) Herbivore specific induction of silica-based plant defences.
Oecologia 152: 677-683
Mattner SW, Parbery DG (2001) Rust enhanced allelopathy of perennial ryegrass against white clover.
Agron J 93: 54-59
Mauricio R, Bowers MD, Bazzaz FA (1993) Pattern of leaf damage affects fitness of the annual plant
Raphanus sativus (Brassicaceae). Ecology 74: 2066-2071
McAuslane HJ, Alborn HT, Toth JP (1997) Systemic induction of terpenoid aldehydes in cotton pigment
glands by feeding of larval Spodoptera exigua. J Chem Ecol 23: 2861-2879
McCrea KD, Abrahamson WG (1985) Evolutionary impacts of the goldenrod ball gallmaker on Solidago
altisima clones. Oecologia 68: 20-22
Mckey D (1974) Adaptive patterns in alkaloid physiology. Am Nat 108: 305-320
Milewski AV, Young TP, Madden D (1991) Thorns as induced defenses - Experimental evidence.
Oecologia 86: 70-75
Miner BG, Sultan SE, Morgan SG, Padilla DK, Relyea RA (2005) Ecological consequences of phenotypic
plasticity. Trend Ecol Evo 20: 685-692
Oborny B, Kun A, Czaran T, Bokros S (2000) The effect of clonal integration on plant competition for
mosaic habitat space. Ecology 81: 3291-3304
Oborny B, Czaran T, Kun A (2001) Exploration and exploitation of resource patches by clonal growth: a
spatial model on the effect of transport between modules. Ecol Model 141: 151-169
Oborny B (2003) External and internal control in plant development. Adaptive plasticity in plants. Journal
of Systems Science and Complexity 16:22-28.
Oinonen E (1967) The correlation between the size of Finish bracken (Pteridium aquiliunum (L.) Kuhn.)
clones and certain periods of history. Acto Forestal Fennica 83: 1-51
Orians CM, Pomerleau J, Ricco R (2000) Vascular architecture generates fine scale variation in systemic
induction of proteinase inhibitors in tomato. J Chem Ecol 26: 471-485
Pan JJ, Clay K (2002) Infection by the systemic fungus Epichloë glyceriae and clonal growth of its host
grass Glyceria striata. Oikos 98: 37-46

References
115
Pederson GA, Brink GE (1998) Cyanogenesis effect on insect damage to seedling white clover in a
bermudagrass sod. Agro J 90: 208-210
Pichersky E, Gershenzon J (2002) The formation and function of plant volatiles: perfumes for pollinator
attraction and defense. Curr Opin Plant Biol 5: 237-243
Pierik R, Visser EJW, de Kroon H, Voesenek LACJ (2003) Ethylene is required in tobacco to successfully
compete with proximate neighbours. Pl Cell Env 26: 1229-1234
Piqueras J, Klimes L (1998) Demography and modelling of clonal fragments in the pseudoannual plant
Trientalis europaea L. Plant Ecol 136: 213-227
Piqueras J. (1999) Infection of Trientalis europaea by the systemic smut fungus Urocystis trientalis:
disease incidence, transmission and effects on performance of host ramets. J Ecol 87: 995-1004
Pitelka LF, Ashmun JW (1985) Physiology and integration of ramets in clonal plants. In: Jackson JBC
(ed.) Population Biology and Evolution of Clonal Organisms. Yale University Press, New Haven, pp.
399-435
Potter LR (1993) The effects of white clover mosaic virus on vegetative growth and yield of clones of
S.100 white clover. Plant Pathol 42: 797-805
Potvin CM, Lechowicz J, Tardif S (1990) The statistical analysis of ecophysiological response curves
obtained from experiments involving repeated measures. Ecology 71: 1389-1400
Price EAC, Hutchings MJ (1992a) Studies of growth in the clonal herb Glechoma hederacea - 2. The
effects of selective defoliation. J Ecol 80: 39-47
Price EAC, Marshall C, Hutchings MJ (1992b) Studies of growth in the clonal herb Glechoma hederacea -
1. Patterns of physiological integration. J Ecol 80: 25-38
Price EA, Hutchings MJ and Marshall C (1996) Causes and consequences of sectoriality in the clonal
herb Glechoma hederacea. Vegetatio 127: 41-54
Prince JS, LeBlanc WG, Macia S (2004) Design and analysis of multiple choice feeding preference data.
Oecologia 138: 1-4
Puustinen S, Mutikainen P (2001) Host-parasite-herbivore interactions: implications of host cyanogenesis.
Ecology 82: 2059-2071

References
116
Qureshi FA, Spanner DC (1971) Unidirectional movement of tracers along the stolon of Saxifraga
sarmentosa. Planta 101: 133-146
Qureshi FA, Spanner DC (1973) Movement of 14C sucrose along the stolon of Saxifraga sarmentosa.
Planta 110: 145-152
Rasmann S, Kollner TG, Degenhardt J, Hiltpold I, Toepfer S, Kuhlmann U, Gershenzon J, Turlings TCJ
(2005) Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 434:
732-737
Rautio P, Markkola A, Martel J, Tuomi J, Harma E, Kuikka K, Siitonen A, Riesco IL, Roitto M (2002)
Developmental plasticity in birch leaves: defoliation causes a shift from glandular to nonglandular
trichomes. Oikos 98: 437-446
Redak RA, Capinera JL (1994) Changes in western wheatgrass foliage quality following defoliation -
Consequences for a graminivorous grasshopper. Oecologia 100: 80-88
Read J, Sanson GD (2003) Characterizing sclerophylly: the mechanical properties of a diverse range of
leaf. New Phytol 160: 81-99
Riipi M, Ossipov V, Lempa K, Haukioja E, Koricheva J, Ossipova S, Pihlaja K (2002) Seasonal changes in
birch leaf chemistry: are there trade-offs between leaf growth, and accumulation of phenolics?
Oecologia 130: 380-390
Roa R (1992) Design and analysis of multiple-choice feeding-preference experiments. Oecologia 89: 509-
515
Roda AL, Baldwin IT (2003) Molecular technology reveals how the induced direct defenses of plants work.
Basic Appl Ecol 4: 15-26
Rodriguez-Saona C, Thaler JS (2005) Herbivore-induced responses and patch heterogeneity affect
abundance of arthropods on plants. Ecol Entomol 30: 156-163
Sagers CL, Coley PD (1995) Benefits and costs of defense in a neotropical shrub. Ecology 76: 1835-1843
Sampol B, Bota J, Riera D, Medrano H, Flexas J (2003) Analysis of the virus-induced inhibition of
photosynthesis in malmsey grapevines. New Phytol 160: 403-412
Schittko U, Baldwin IT (2003) Constraints to herbivore-induced systemic responses: bidirectional signaling
along orthostichies in Nicotiana attenuata. J Chem Ecol 29: 763-770

References
117
Schmitt J, Dudley SA, Pigliucci M (1999) Manipulative approaches to testing adaptive plasticity:
phytochrome-mediated shade-avoidance. Am Nat 154: 43-54
Schultz JC (1988) Plant responses induced by herbivores. Trend Ecol Evo 3: 45-49
Shelton AL (2004) Variation in chemical defences of plants may improve the effectiveness of defence.
Evol Ecol Res 6: 709-726
Shelton AL (2005) Within-plant variation in glucosinolate concentrations of Raphanus sativus across
multiple scales. J Chem Ecol 31: 1711-1732
Shumway SW (1995) Physiological integration among clonal ramets during invasion of disturbance
patches in a New England salt marsh. Ann Bot 76: 225-233
Siemens DH, Lischke H, Maggiulli N, Schurch S, Roy BA (2003) Cost of resistance and tolerance under
competition: the defense-stress benefit hypothesis. Evol Ecol 17: 247-263
Slade AJ, Hutchings MJ (1987a) The effects of light-intensity on foraging in the clonal herb Glechoma
hederacea. J Ecol 75: 639-650
Slade AJ, Hutchings MJ (1987b) The effects of nutrient availability on foraging in the clonal herb
Glechoma hederacea. J Ecol 75: 95-112
Snoeren TAL, De Jong PW, Dicke M (2007) Ecogenomic approach to the role of herbivore-induced plant
volatiles in community ecology. J Ecol 95: 17-26
Solangaarachchi SM, Harper JL (1989) The growth and asymmetry of neighboring plants of white clover
(Trifolium repens L). Oecologia 78: 208-213
Soler R, Bezemer TM, Van der Putten WH, Vet LEM, Harvey JA (2005) Root herbivore effects on above-
ground herbivore, parasitoid and hyperparasitoid performance via changes in plant quality. J Animal
Ecol 74: 1121-1130
Steinger T, Körner C, Schmid B (1996) Long-term persistence in a changing climate: DNA analysis
suggests very old ages of clones of alpine Carex curvula. Oecologia 105: 94-99
Stockhoff BA (1993) Diet Heterogeneity - Implications for growth of a generalist herbivore, the gypsy
moth. Ecology 74: 1939-1949
Stout MJ, Workman KV, Duffey SS (1996a) Identity, spatial distribution, and variability of induced
chemical responses in tomato plants. Ent Exp App 79: 255-271

References
118
Stout MJ, Workman KV, Workman JS, Duffey SS (1996b) Temporal and ontogenetic aspects of protein
induction in foliage of the tomato, Lycopersicon esculentum. Biochem Syst Ecol 24: 611-625
Stratmann JW (2003) Long distance run in the wound response - Jasmonic acid is pulling ahead. Trends
Pl Sci 8: 247-250
Strauss SY, Rudgers JA, Lau JA, Irwin RE (2002) Direct and ecological costs of resistance to herbivory.
Trend Ecol Evo 17: 278-285
Stuefer JF, During HJ, de Kroon H (1994) High benefits of clonal integration in two stoloniferous species
in response to heterogeneous light environments. J Ecol 82: 511-518
Stuefer JF, Hutchings MJ (1994) Environmental heterogeneity and clonal growth: a study of the capacity
for reciprocal translocation in Glechoma hederacea L. Oecologia 100: 302-308
Stuefer JF (1996) Potential and limitations of current concepts regarding the response of clonal plants to
environmental heterogeneity. Vegetatio 127: 55-70
Stuefer JF, de Kroon H, During HJ (1996) Exploitation of environmental heterogeneity by spatial division
of labour in a clonal plant. Funct Ecol 10: 328-334
Stuefer JF (1997) Division of labor in clonal plants? On the response of stoloniferous herbs to
environmental heterogeneity. PhD thesis. Utrecht University, The Netherlands
Stuefer JF, During HJ, Schieving F (1998) A model on optimal root-shoot allocation and water transport in
clonal plants. Ecol Model 111: 171-186
Stuefer JF, Huber H (1998) Differential effects of light quantity and spectral light quality on growth,
morphology and development of two stoloniferous Potentilla species. Oecologia 117: 1-8
Stuefer JF, Gómez S, Van Mölken T (2004) Clonal integration beyond resource sharing: implications for
defence signalling and disease transmission in clonal plant networks. Evol Ecol 18: 647-667
Sultan SE (2000) Phenotypic plasticity for plant development, function and life history. Trend Pl Sci 5:
537-542
Sutherland WJ (1987) Plant Ecology - Growth and foraging behavior. Nature 330: 18-19
Takabayashi J, Dicke M (1996) Plant-carnivore mutualism through herbivore-induced carnivore
attractants. Trends Pl Sci 1: 109-113

References
119
Takabayashi J, Dicke M (1997). Herbivore-induced plant volatiles with multifunctional effects in
ecosystems: a complex pattern of biotic interactions. In Abe T, Levin SA, Higashi M (eds.)
Biodiversity, an ecological perspective. Springer, pp. 131-145
Thaler JS, Farag MA, Pare PW, Dicke M (2002a) Jasmonate-deficient plants have reduced direct and
indirect defenses against herbivores. Ecol Lett 5: 764-774
Thaler JS, Karban R, Ullman DE, Boege K, Bostock RM (2002b) Cross-talk between jasmonate and
salicylate plant defense pathways: effects on several plant parasites. Oecologia 131: 227-235
Thomas H (2002) Ageing in plants. Mech Ageing Dev 123: 747-753
Thompson GA Schulz A (1999) Macromolecular trafficking in the phloem. Trends Pl Sci 4: 354-360
Till I (1987) Variability of expression of cyanogenesis in white clover (Trifolium repens L). Heredity 59:
265-271
Tollrian R, Harvell D (1999) The ecology and evolution of inducible defenses. Princeton University Press,
Princeton, New Jersey
Traw MB, Dawson TE (2002) Differential induction of trichomes by three herbivores of black mustard.
Oecologia 131: 526-532
Treutter D (2006) Significance of flavonoids in plant resistance: a review. Environ Chem Lett 4: 147-157
Turkington R, Hamilton RS, Gliddon C (1991) Within-population variation in localized and integrated
responses of Trifolium repens to biotically patchy environments. Oecologia 86: 183-192
Underwood NC (1998) The timing of induced resistance and induced susceptibility in the soybean
Mexican bean beetle system. Oecologia 114: 376-381
Underwood N, Morris W, Gross K, Lockwood JR (2000) Induced resistance to Mexican bean beetles in
soybean: variation among genotypes and lack of correlation with constitutive resistance. Oecologia
122: 83-89
van Dam NM, Verpoorte R, van der Meijden E (1994) Extreme differences in pyrrolizidine alkaloid levels
between leaves of Cynoglossum officinale. Phytochemistry 37: 1013-1016
van Dam NM, Vuister LWM, Bergshoeff C, Devos H, Vandermeijden E (1995) The raison-detre of
pyrrolizidine alkaloids in Cynoglossum officinale - Deterrent effects against generalist herbivores. J
Chem Ecol 21: 507-523

References
120
van Dam NM, Baldwin IT (1998) Costs of jasmonate-induced responses in plants competing for limited
resources. Ecol Lett 1: 30-33
van Dam NM, Baldwin IT (2001) Competition mediates costs of jasmonate-induced defences, nitrogen
acquisition and transgenerational plasticity in Nicotiana attenuata. Funct Ecol 15: 406-415
van Dam NM, Raaijmakers CE, van der Putten WH (2005) Root herbivory reduces growth and survival of
the shoot feeding specialist Pieris rapae on Brassica nigra. Ent Exp App 115: 161-170
van Groenendael J, de Kroon H (1990) Clonal growth in plants: regulation and function. SPB Academic
Publishing, The Hague
van Kleunen M, Stuefer JF (1999) Quantifying the effects of reciprocal assimilate and water translocation
in a clonal plant by the use of steam-girdling. Oikos 85: 135-145
Verburg RW, During HJ (1998) Vegetative propagation and sexual reproduction in the woodland
understorey pseudo-annual Circaea lutetiana L. Plant Ecol 134: 211-224
Viswanathan DV, Thaler JS (2004) Plant vascular architecture and within-plant spatial patterns in
resource quality following herbivory. J Chem Ecol 30: 531-543
Vuorisalo T, Hutchings MJ (1996) On plant sectoriality, or how to combine the benefits of autonomy and
integration. Vegetatio 127: 3-8
Waterhouse PM, Wang M-B, Lough T (2001) Gene silencing as an adaptive defense against viruses.
Nature 411: 834-842
Watson MA, Casper BB (1984) Morphogenetic constraints on patterns of carbon distribution in plants. Ann
Rev Ecol Syst 15: 233-258
Weijschede J, Martinkova J, De Kroon H, Huber H (2006) Shade avoidance in Trifolium repens: costs and
benefits of plasticity in petiole length and leaf size. New Phytol 172: 655–666
Weijschede J, Berentsen R, de Kroon H, Huber H (2008) Consequences of petiole elongation and
internode lengths and their plasticity for plant performance of Trifolium repens under competition
and disturbance. Evol Ecol in press.
Weiner J (1990) Asymmetric competition in plant-populations. Trend Ecol Evo 5: 360-364
Wennstrom A ,Ericson L (1992) Environmental heterogeneity and disease transmission within clones of
Lactuca sibirica. J Ecol 80: 71-77

References
121
Wennstrom A (1999) The effect of systemic rusts and smuts on clonal plants in natural systems. Plant
Ecol 141: 93-97
Wilhalm T (1995) A comparative study of clonal fragmentation in tussock-forming grasses. Abstracta
Botanica 19: 51-60
Wittstock U, Gershenzon J (2002) Constitutive plant toxins and their role in defense against herbivores
and pathogens. Curr Opin Plant Biol 5: 300-307
Xiang H, Chen J (2004) Interspecific variation of plant traits associated with resistance to herbivory among
four species of Ficus (Moraceae). Ann Bot 94: 377-384
Yemm EW, Willis AJ (1954) The estimation of carbohydrates in plant extracts by anthrone. Biochem J 57:
508-51
Zangerl AR, Arntz AM, Berenbaum MR (1997) Physiological price of an induced chemical defense:
photosynthesis, respiration, biosynthesis, and growth. Oecologia 109: 433-441
Zvereva EL, Kozlov MV, Niemela P, Haukioja E (1997) Delayed induced resistance and increase in leaf
fluctuating asymmetry as responses of Salix borealis to insect herbivory. Oecologia 109: 368-373

References
122

Nederlandse samenvatting
123
NEDERLA
NDSE SA
MEN
VATTIN
G
NETWERKEN VAN KLONALE PLANTEN
In de natuur is klonaliteit een algemene vorm van plantengroei die bij een groot aantal soorten en in een
uiteenlopend scala aan vormen voorkomt. Klonale groei kan bijvoorbeeld tot stand komen door de
vorming van bovengrondse, horizontale uitlopers die meerdere genetisch identieke individuen (ramets)
met elkaar verbinden en daardoor hele netwerken van klonale planten vormen. De omvang van deze
netwerken kan variëren van enkele tot grote aantallen verbonden ramets die deel uitmaken van hetzelfde
netwerk. In sommige gevallen kunnen individuele klonen dan ook grote gebieden bedekken. Klonale
netwerken worden niet uitsluitend door kruidachtige planten, zoals aardbei en klaver, gevormd. De
grootste netwerken van klonale planten worden in de regel door houtachtige planten, zoals populieren en
bamboe, gevormd en kunnen zelfs enkele hectaren beslaan.
Klonale soorten zijn in staat om grondstoffen, signalen en ziekteverwekkers zowel vertikaal, dat wil
zeggen binnen het ramet, als ook horizontaal tussen ramets van dezelfde kloon te transporteren
(hoofdstuk 1; Figuur 1). Dit in tegenstelling tot rechtopstaande planten waarbij alleen vertikaal transport
mogelijk is. In heterogene en/of stressvolle omstandigheden kan de mogelijkheid van horizontaal transport
bijzonder voordelig zijn voor klonale planten. Onderling verbonden individuen kunnen door middel van
horizontaal transport informatie uitwisselen, waardoor het effect van competitie geminimaliseerd en de
opname van grondstoffen geoptimaliseerd wordt. Naast de uitwisseling van grondstoffen tussen ramets,
biedt horizontaal transport ook de mogelijkheid tot uitwisseling van andere signalen. Zoals in dit
proefschrift aangetoond wordt (hoofdstuk 2), kunnen ramets verdedigingssignalen uitwisselen in reactie
op schade die ze van herbivoren ondervinden.
Elke ramet is functioneel onafhankelijk doordat een ramet zijn eigen bladeren, wortels en uitlopers
(zijtakken) kan vormen, en daardoor in staat is om zijn eigen grondstoffen op te nemen en te produceren
(bijvoorbeeld water, mineralen en fotoassimilaten). De ramets kunnen grondstoffen blijven uitwisselen
zolang ze met elkaar verbonden en dus fysiologisch geïntegreerd zijn. Zoals gezegd is in heterogene
milieus fysiologische integratie tussen leden van dezelfde kloon met name bijzonder voordelig voor wat
betreft het succes van de hele kloon.
Het intranet-transport van grondstoffen tussen ramets wordt bepaald door de vasculaire “source-
sink” dynamiek. Volwassen, volgroeide ramets (sources) ondersteunen de jonge, zich ontwikkelende
ramets (sinks), door transport van grondstoffen dat hoofdzakelijk in de richting van de jonge ramets
verloopt. De dynamiek van deze vasculaire grondstoffenstroom is echter flexibel, waardoor tijdelijke
veranderingen in de richting mogelijk zijn. De richting kan bijvoorbeeld veranderen doordat “source-sink”
patronen lokaal worden beïnvloed door omgevingsfactoren (zoals schaduw). Individuele ramets van een
klonale plant zijn in staat om plastisch op hun veranderende omgeving te reageren. Daarnaast kunnen de
ramets hun individuele respons ook coördineren met andere leden van het netwerk, waardoor klonale
planten extra mogelijkheden hebben om beperkende milieuomstandigheden te overleven en deze beter te
benutten. Het concept van plasticiteit wordt hierdoor dus naar een hoger niveau gebracht.

Nederlandse samenvatting
124
VERDEDIGING VAN PLANTEN TEGEN HERBIVOREN
Omdat planten vastzitten aan de bodem en dus niet voor herbivoren weg kunnen lopen, moeten ze
specifieke verdedigingsmechanismen ontwikkelen. De verdediging kan continu tot expressie komen
(constitutieve verdediging) door middel van morfologische kenmerken of chemische stoffen die het succes
of de kans op overleving van de aanvallende herbivoor verminderen. Verdediging is echter een kostbaar
proces en de investering van grondstoffen in dit proces kan ten koste gaan van de investering in andere
processen zoals groei en reproductie. Om de verdeling van grondstoffen tussen verschillende processen
te optimaliseren en de kosten van verdediging zoveel mogelijk te beperken, wordt er in de literatuur over
plantenverdediging vaak aangenomen dat planten een vorm van verdediging hebben ontwikkeld die
alleen geactiveerd wordt als dat nodig is (geïnduceerde verdediging). Zo kunnen planten in reactie op een
aanval van herbivoren chemische stoffen produceren of fysieke structuren (zoals haren of doornen)
ontwikkelen waardoor ze hun staat van verdediging verhogen en daarmee de negatieve gevolgen door
herbivorie beperken.
Nadat de verdediging lokaal geactiveerd is, kan deze ook in andere onbeschadigde delen van de
plant tot expressie komen (systemische inductie van afweer) waardoor de toekomstige schade
gereduceerd kan worden. De “source-sink” dynamiek en de vasculaire stroming bepalen waar de
verdedigingsstoffen in de plant terecht kunnen komen. Systemische inductie als gevolg van lokale
beschadiging kan alleen tot stand komen in die delen van de plant die rechtstreeks, door middel van
vaatbundels, verbonden zijn. Planten kunnen de beperkingen van de interne verspreiding van
verdedigingssignalen (zoals sectorialiteit) omzeilen door gebruik te maken van andere, externe
communicatiekanalen, zoals het uitstoten van vluchtige stoffen.
Als reactie op herbivorie kunnen planten signalen afgeven die naburige planten als sein kunnen
gebruiken. Deze signalen kunnen door de beschadigde plant in de vorm van vluchtige stoffen en door de
wortels uitgescheiden stoffen worden afgegeven, waardoor er in andere onbeschadigde delen van
dezelfde plant een verdedigingsreactie kan optreden. Het afgeven van informatie via externe kanalen kan
echter ook risico’s met zich mee brengen voor de plant in kwestie. Naburige, concurrenten zouden de
informatie over het gevaar van herbivorie kunnen gebruiken (als een soort afluisteren) om hun eigen
verdedigingssysteem te activeren voordat de eigenlijke aanval van herbivoren plaats vindt. De
afluisterende plant kan de toegebrachte schade beperken wanneer deze vervolgens door herbivoren
wordt aangevallen. Dit levert de afluisterende plant een concurrentievoordeel op ten opzichte van de plant
die de signalen afgeeft.
In netwerken van klonale planten bieden de horizontale uitlopers (stolonen) echter een mogelijkheid
die ontbreekt in niet-klonale planten. Via de stolonen kan informatie over een mogelijk risico op herbivorie
op een veilige manier worden doorgegeven, waardoor het gevaar van afluisteren, en andere problemen
van externe transport routes (zoals luchtstromingen, afstand tussen de zender en ontvanger, enz.),
worden ontweken. Het uitwisselen van informatie over het risico op herbivorie binnen netwerken van
klonale planten kan gezien worden als een bijzonder geval van plant-plant communicatie, waarbij de
zender en ontvanger met elkaar verbonden zijn. Wanneer een ramet beschadigd wordt door een

Nederlandse samenvatting
125
herbivoor kan deze informatie doorgeven aan de aangrenzende ramets. De resulteert in de inductie van
het verdedigingsmechanisme in de onbeschadigde ramets, voordat de eigenlijke aanval door de herbivoor
heeft plaatsgevonden. Schade die door een individuele ramet wordt ondervonden, is waarschijnlijk een
betrouwbare indicatie voor toekomstige schade, in het bijzonder omdat stolonen als fysieke bruggen
tussen ramets kunnen dienen. Sommige kleine herbivoren (bijv. mijten) kunnen namelijk de stolonen
volgen tijdens hun zoektocht naar voedsel.
DOEL EN UITEENZETTING VAN DIT PROEFSCHRIFT
Het belangrijkste doel van dit proefschrift was het testen van het algemene idee dat ramets van uitloper-
vormende planten de verbindingen tussen hun ramets voor meer gebruiken dan het transport van
grondstoffen alleen. De nadruk ligt hierbij op het uitwisselen van verdedigingssignalen en de ecologische
implicaties daarvan. Ik heb voor alle experimenten de uitloper-vormende plant Trifolium repens gebruikt.
T. repens is in het onderzoek aan klonale planten een klassiek modelsysteem. De morfologische eenvoud
en de grote hoeveelheid beschikbare literatuur over intra-klonaal transport van grondstoffen maken T.
repens een ideale kandidaat om vragen over de dynamiek (in tijd en ruimte) van systemisch geïnduceerde
afweer in netwerken van klonale planten te beantwoorden. Daarbij heb ik de larven van de generalistische
herbivoren Spodoptera exigua en Mamestra brassicae gebruikt om de verdediging in de planten te
induceren en te testen.
Dit proefschrift begint met het formuleren van voorspellingen over de ecologische en evolutionaire
implicaties van fysiologische integratie tussen ramets (hoofdstuk 2). Ik voorspel dat er na een lokale
aanval door herbivoren een early warning systeem zal worden geactiveerd. Ik verwacht dat dit systeem
bescherming biedt aan andere leden van het netwerk, door het activeren van directe (effecten op de
prestatie en/of voorkeur van de herbivoor) en indirecte (aantrekken van natuurlijke vijanden van de
herbivoor) verdedigingskenmerken. Daarnaast doe ik in dit hoofdstuk ook voorspellingen over de risico’s
van fysiologische integratie, waarbij de nadruk ligt op virus-transmissie in netwerken van klonale planten.
In de volgende hoofdstukken richt ik mij uitsluitend op het transport van verdedigingsstoffen als gevolg
van herbivorie.
In hoofdstuk 3 heb ik de hypothese over het early warning systeem, die in hoofdstuk 2 aan bod is
gekomen, getest. Ik heb met name getest of ramets van T. repens, als reactie op lokale herbivorie, in
staat zijn om hun verdedigingsmachinerie in andere delen van de kloon aan te zetten. Daarnaast hebben
we door het experimenteel manipuleren van de floëemstroom onderzocht welke route en welke richting de
verdedigingssignalen in het netwerk volgen.
In hoofdstuk 4 heb ik de timing van de geïnduceerde reactie in klonale fragmenten onderzocht. Ik
heb bestudeerd hoeveel tijd er nodig is voordat het verdedigingssignaal van ramet naar ramet
getransporteerd wordt en daar tot expressie komt, en hoelang het duurt voordat de geïnduceerde
weerstand na activering weer afneemt.
Uit hoofdstuk 3 kan geconcludeerd worden dat de inductie van de verdediging voornamelijk in de
richting van de jongere ramets tot expressie komt. Dit éénrichtingstransport suggereert dat het initiële punt

Nederlandse samenvatting
126
van aanval cruciaal is voor de veiligheid van de andere leden van het netwerk. Jonge ramets fungeren als
sterke “sinks” en als deze als eerste aangevallen worden, zal het signaal niet naar de andere leden van
het netwerk getransporteerd worden. Om vast te kunnen stellen wat de ecologische waarde van
systemische weerstand in netwerken van klonale planten is, heb ik in hoofdstuk 5 onderzocht of er een
associatie is tussen de voorkeur van de herbivoor, en de leeftijd van de ramet en de inductie van de
verdediging. Het tweede doel van dit hoofdstuk was het in kaart brengen van de kenmerken en
mechanismen die de waargenomen voorkeur zouden kunnen verklaren. Om de kwaliteit van het weefsel
te kunnen vaststellen, zijn er drie groepen van kenmerken geselecteerd: biomechanica, voedingswaarde
en chemische verdediging. Deze kenmerken zijn gemeten in jonge en volwassen ramets van zowel
controle planten als planten waarin de verdediging geïnduceerd was. Vervolgens zijn de kenmerken door
middel van tweezijdige keuzetesten gecorreleerd aan de voorkeur van de herbivoor
Bij een ecologisch relevante verdedigingsstrategie, moeten de voordelen van de verdediging even
groot als, of groter dan de kosten zijn. Na lokale herbivorie kunnen de kosten van systemische expressie
van verdediging uitzonderlijk groot zijn in klonale planten, wanneer alle leden van het netwerk
geïnduceerd worden en er geen aansluitende aanval in andere ramets plaatsvindt. Aan de andere kant
kan een tijdige waarschuwing van andere ramets binnen het netwerk enorme voordelen met zich
meebrengen, doordat de schade door herbivoren beperkt wordt wanneer er na initiële vraat een verdere
aanval op andere ramets plaatsvindt. Om de kosten en baten van geïnduceerde weerstand in een klonaal
netwerk vast te stellen, heb ik geïnduceerde en controleplanten samen laten groeien in de aan- en
afwezigheid van herbivoren en heb ik een aantal fitness-gerelateerde kenmerken en het succes van de
planten gemeten (hoofdstuk 6).
Ten slotte eindigt dit proefschrift met een samenvattende discussie (hoofdstuk 7) van de
belangrijkste resultaten en de mogelijke implicaties van early warning systemen in netwerken van klonale
planten.
CONCLUDERENDE OPMERKINGEN
Dit proefschrift is de eerste studie die induceerbare verdedigingsstrategieën in de context van netwerken
van klonale planten onderzoekt. Ik heb laten zien dat de verbindingen tussen ramets een cruciale rol
spelen als het gaat om de verspreiding van verdedigingssignalen tussen verbonden individuen. Door deze
voordelige eigenschap zijn onbeschadigde leden van het netwerk in staat om plastische veranderingen in
afweerkenmerken te ondergaan die de toekomstige schade door herbivoren kunnen beperken. Daarnaast
voorzien deze verbindingen de jonge, ontwikkelende ramets van ouderlijke ondersteuning, waardoor hun
kansen op overleving toenemen.
De onderliggende principes en beperkingen van geïnduceerde verdediging in klonale planten zijn in
dit proefschrift beschreven. Als volgende stap zou onderzocht moeten worden hoe algemeen het gebruik
van early warning systemen is bij netwerkvormende soorten. Aanvullende experimenten onder natuurlijke
omstandigheden zouden vervolgens een beter inzicht kunnen geven in de ecologische relevantie van
intranet-communicatie.

Nederlandse samenvatting
127
We kunnen concluderen dat de studie naar plant- insect interacties een nieuwe dimensie toevoegt
aan de onderzoeksvelden van geïnduceerde verdediging, plant-plant communicatie en plant-dier
communicatie. Dit proefschrift legt een fundament wat betreft het onderzoek naar verdediging in klonale
planten, doordat het de basis biedt voor toekomstig onderzoek naar zowel de mechanismen als de
ecologische en evolutionaire implicaties van dit interessante fenomeen.

Nederlandse samenvatting
128

Acknowledgements
129
AC
KNO
WLEDG
EMEN
TS
The outcome of this book is by no means a one person job. This thesis has been possible through the
direct (e.g. technical assistance, discussions, etc) and indirect help (e.g. casual chitchats, borrels, etc)
from a number of people, providing clear evidence of the importance of being connected.
Undertaking my PhD in the Netherlands has given me the opportunity to meet many people both
within and outside the science community. I have come across very peculiar individuals and it has given
me the chance to learn a great deal about people and the way to deal with different personalities and
cultures. Moving from Spain to the Netherlands was a major step in my life and I am very grateful to have
found an exceptional working and social atmosphere that made my time in Nijmegen an unforgettable
experience.
First of all, I’d like to thank the person who gave me the opportunity to accomplish this. Josef, where
to start? We go back a long way!. During my ERASMUS period in Utrecht in 1999/2000 I already
concluded you were one-of-a-kind supervisor and person. As a supervisor, I’ve learned a lot from you
(definitely not only random facts). You always let me have my freedom and independence from the very
beginning and yet, you were always there for me anytime that I needed to discuss something. On the
personal level, I enjoyed your permanent cheerful and positive attitude, our “core uitjes” and how you
never say “no” to trying something. Thank you so much for making my PhD a great experience. And by
the way, T2YgYWxsIHJhbmRvbSB0aGluZ3MsIHRoaXMgd2FzIG15IGZhdiA6KQ==.
Hans, thank you for your general guidance and discussions whenever I had “luxury problems” as
you use to call the multiple side ramifications of my project. I greatly appreciated your feed-back and
careful reading of my papers and your encouragement on my networking activities.
The people in the Experimental Plant Ecology department have all (without exception) helped me at
one point or another and therefore my thanks go to you all. Eric tio, your honesty and critical opinions are
a trademark (together with your pocketknife, of course). They were very helpful to me on many occasions
(the knife included). Thank you for taking me to Binn, you can always count on me for that one! Heidi,
thank you for keeping me up till past 2 am every night in the conference in London. Jokes aside, I enjoyed
our chitchats and your contributions in the lab meetings. José B., you made my life so much easier. You
always made sure that those things that needed to get done, got done and done fast! That must be why
you are the “Major”, right? Gerard, I appreciate the time and dedication you spent whenever I asked for
your help in developing an instrument. Annemiek, thank you for providing assistance in my experiments
and making sure my caterpillars always made it to Nijmegen early in the morning. Harry, your kindness
and willingness to help is unique. Hannie, you add a colorful note to the department, always successfully
organizing events. José A., thank you for patiently listening to me babbling in Dutch whenever I went to
your room to get something. Peter Charpentier, my days were fueled with the indispensable coffee you
make. Thanks to everyone in the Aquatic Ecology department, especially Dries, your help with occasional
computer problems saved the day more than once, especially the time my computer crashed (mental
note: make back-up). Martin, you always provided me with an immediate solution to any needs I ever had.
I am truly thankful to the greenhouse staff. Walter, I really appreciate your interest not only in my
project but also in my personal life. My dear Yvette, you always made me laugh. Harry, you always

Acknowledgements
130
stopped everything you were doing to help me find something. Gerard, you were
always a great source of information, from diseases on clover to the biology of
Solanum stoloniferum. I am glad I could make use of that valuable database you
dedicate so much effort to. I miss all of you, our casual chats and your kindness.
Special thanks go to my fellow PhDs, who came and went through the years. In
the beginning, I enjoyed the company of Liesje & Ronald, my first office roommates.
Linda, Julia, Werner and Eelke, we only shared a very brief time together in the
department but I am glad we could at least share a couple of “PhD dinners” together.
When we moved to the new building, I got new office roommates: Corien, you were
my inspiration to be orderly and the most disciplined person when it comes to
speaking in Dutch to me, your patience is amazing. I really enjoyed having you
around. Jelmer, I indeed understood your humor (almost all the time). We started
almost at the same time and since then you have got a house, got married, had a
baby… jeeez slow down, I can’t catch up! Xin, I enjoyed your curiosity. You always
had a random question for the rest of us. Tamara, you introduced me to the ALT
function which has saved me countless hours and headaches (especially while laying
out my thesis!). Besides that, you and I have shared a lot of happy, less happy, fun
and crazy moments. You started as a colleague and became one of my closest
friends. You were/are always there for me no matter for what (including the translation
of my Dutch summary!), thank you paranymfie humana. I miss you a lot. As for my
roommates at the other side of the wall: my dear Iain Russon and Peter Walker. Iain,
the very first time our paths crossed you almost kicked my face (accidentally) while
parking your bike. Obviously, after that we could only become very good friends or
enemies for life. Luckily, it was the first. We did so much stuff together: swimming,
cooking, going out…I especially enjoyed our trip to England…good old times, eh?
Don’t become a stranger. Pete, 3.14, the connection I have with you is something I
don’t have with many people (the freaky stone says it all), you know why? “because
you are like that”. You are one those loyal friends one wants to keep for life. I miss
your advice, our long talks and the way you always make me laugh. Thanks for
helping with the English bloopers in this thesis!
Throughout the last few years I had the pleasure of collaborating with other
PhDs from different countries who came to Nijmegen for short stays. Iva Špa ková
(University of South Bohemia), shortly after you arrived you found a four-leaved clover
among our stock plants (which I hadn’t yet found despite a whole year of looking!). I
should have believed myself that it was a lucky sign, because during that time it was
the turning point of my project. Vítek Latzel (University of South Bohemia), writing
paper bags with you was an unforgettable experience and our effort was rewarded by

Acknowledgements
131
a nice paper! Naomi Hosaka (Tokyo Metropolitan University), the science world keeps
bringing us together in different parts of the planet. Michelle Sampaio (Universidade
Federal do Rio de Janeiro) and Omer Falik (Ben-Gurion University of the Negev)
during the time we shared in my first year you became almost like family to me. I hope
we will meet again soon. Dulce Mantuano (Universidade Federal do Rio de Janeiro)
also kindly known as “el caballo de Atila”, it was fun having your craziness around in
the lab. Thank you for taking such good care of me in Brazil. My sincere gratitude
goes to the students that have helped during my thesis for they have contributed with
valuable data for my project: Yolanda Verhulst, William van Dijk, Sabrina van
Nieuwkerk, Fabiën Bruine and Nils van Rooijen.
I also want to thank a number of people for helping me doing measurements
and side pilot experiments. Thanks to Luc-Jan Laarhoven, Marco Steeghs and Simona
Cristescu who invited me to make use of their equipment and for putting in so much
time helping me with the collection of data.Yusuke Onoda (UU) for his insight into the
world of plant biomechanics. Vladimir Ossipov (University of Turku) for phenolic
profiling. Heinjo During (UU) for help with leaf reflectance measurements. Rieta Gols
(WUR) and Marcel Dicke (WUR) for help with the olfactometer experiments. Jelle
Eygensteyn for CN analysis. Ine Hendriks for help with sugar measurements. Merijn
Kant (UvA) for help with explorative proteinase inhibitors measurements. Huub op den
Camp, for his time, resources and enthusiasm to explore my samples. Marijn Nijssen,
thanks for identifying any bug I brought to you and for the gezellige chats.
I would also like to thank my fellow PhDs from the NIOO and Wageningen
University (Roxina, Hanneke, Maaike, Colette, Adriana and many others) whom I had
great times with at every workshop or congress we attended together. Roxina: 34, 34,
34, 34!!! In particular, I want to mention Tjeerd Snoeren and Erik Poelman. I enjoyed
very much organizing the “1st annual symposium on ecogenomics of plant defense”
with you guys and I was very glad to see there was a second one! I am also grateful to
M. Carmen López de Bustamante (Instituto de Agricultura Sostenible; CSIC) for her
kind provision of articles otherwise impossible to get. Martijn Frieling, you make
“burocrazy” a lot less painful. Dennis Whigham (Smithsonian Institution), thank you for
supporting my “scientific endeavors”. I would also like to thank Colin Orians (Tufts
University) and Richard Ferrieri (Brookhaven National Laboratory), my current
collaborators, for having opened a new scientific path for me.
The years I have spent in Nijmegen haven’t been all “work, work, work”. I have
been lucky enough to meet many people who made my social life very enjoyable, but I
will restrict myself to mention a few who had a significant impact in my life (and have
not been mentioned yet).

Acknowledgements
132
Andrew, you were my very first buddy, I will never forget that long and cold
night we spent in Utrecht Station after the Cranberries concert, it was worth it though!.
Then I met J9. Janine, you relieved my burden of being the “new person” and
introduced me to the social life in Nijmegen, which remarkably sped up my Dutch
learning process and my education on the most terrible 80’s music. Then Tom came
along, and we introduced the frietje tradition at sunrise. Marc, you are responsible for
broadening my music horizons. I really enjoyed all those “Holgersson moments” with
you, Mascha, Stefan, Johan and Michael. The weekend in Texel was unforgettable
and so was the Haldern pop festival. Stefan R. & Filip, I enjoyed having drinks with
you. Stefan, I loved listening to your funny and complicated life stories. I will definitely
miss the Merleyn nights and chilling in the Bison baai with my Dutch-international
crew: Wout & Floor, Gideon & Mari, Aniko & Bart, Pete & Henne, Iain and associates.
Martijn, I loved your amazing stories about remote places and people. Thanks for
taking me to the most beautiful places around Nijmegen. Although overall we are very
different, it’s funny how similar we can be sometimes. Job, I enjoyed our many
conversations in Spanish about life and science.
Thanks to the Smith family, in particular Becky, thank you for treating me like a
daughter from the first time you met me. I would like to thank the few people that keep
my ties with Spain: Azahara, personajillo genuino, no importa cuanto tiempo pase
desde nuestra última conversación, siempre parece que fué ayer. Marila, hemos
pasado por mucho pero seguimos como el primer día, del que ya hacen 15 años!.
María, tu has estado ahí siempre que lo he necesitado y mucho más, siempre
recordandome que la vida no está llena de “nada con tomate”. Gustavorrrr riau!
Gracias por ayudarme con todas mis preguntas y encargos tecnológicos (nk nk).
Elena te agradezco que me presentases a Venas. Venas, lo pasé genial en Nueva
York. James, thanks for always being so supportive. Ia , gracias por tus hermani-
conversaciones sobre el mundillo de la ciencia, tu me entiendes mejor que nadie.
Mamá, las hay morenas, pelirrojas, incluso rubias. Pero como tú, ninguna! Papá, que
haria yo sin el cañonero? Sin vosotros dos nada de esto hubiera sido posible y por
eso esta tesis os la dedico a vosotros.
And last, but definitely not least, Tyrell. Words are not enough to show you my
appreciation for your unconditional support in every aspect of my life and for the great
sacrifices you make so we can be together. Thank you for being you and for being
there. We make a great team!.

133
LISTOF PUBLIC
ATIO
NS
Gómez S, Latzel V, Verhulst Y, Stuefer JF (2007) Costs and benefits of induced resistance in clonal plant
networks. Oecologia 153: 921-930.
Perea J, García A, Gómez G, Acero R, Peña F, Gómez S (2007) Effect of light and substrate structural
complexity on microhabitat selection of the snail Helix aspersa Müller”. Journal of Molluscan Studies
73: 39-43.
Gómez S, Stuefer JF (2006) Members only: induced systemic resistance to herbivory in a clonal plant
network. Oecologia 147: 461-468.
Stuefer JF, Gómez S, van Mölken T * (2004) Clonal integration beyond resource sharing: implications for
defence signalling and disease transmission in clonal plant networks. Evolutionary Ecology 18: 647–
667. * All authors contributed equally to the ideas and preparation of this manuscript.
Hosaka N, Gómez S, Kachi N, Stuefer JF, Whigham DF (2005) The ecological significance of clonal
growth in the understorey tree, Pawpaw (Asimina triloba). Northeastern Naturalist 12: 11-22.
Gómez S, Onoda Y, Ossipov V, Stuefer JF. Systemic induced resistance: a risk-spreading strategy in
clonal plant networks?. Submitted.
Gómez S, van Dijk W, Stuefer JF. Time is precious: dynamics of induced resistance in a clonal plant
network. Submitted.

134
CUR
RIC
ULUM
VITA
E
SARA GÓMEZ
Sara “Coco” Gómez was born on August 22nd, 1978 in Córdoba (Spain). In 1996 she started her
undergraduate studies in Environmental Sciences at the University of Córdoba. In 1999 she received an
ERASMUS fellowship from the European Union to conduct research and follow undergraduate courses in
an European university. She established contact with the Plant Ecology department in Utrecht University
(The Netherlands), becoming one of the first exchange students of what has since proved to be a long-
lasting collaboration between both universities.
During her stay in Utrecht University, she became acquainted with the field of plant ecology and in
particular with clonal plants. This new interest resulted into two internships from the
Andrew W. Mellon Foundation in 2000 and 2001 respectively, which allowed her
to conduct further research at the Smithsonian Institution (Maryland, USA). In
October 2002, shortly after graduating from the University of Córdoba, she
started her PhD in the department of Experimental Plant Ecology at the Radboud
University Nijmegen.
In December 2007, she was appointed as a post-doctoral research
fellow at Tufts University (Massachusetts, USA) where she now conducts
research on plant resources partition in response to herbivory.


















