
STUDIE VAN DE BACTERIËLE MICROBIOTA IN HONING...Deze stoffen en de mate waarin ze voorkomen bepalen...
38
STUDIE VAN DE BACTERIËLE MICROBIOTA IN HONING Aantal woorden: 9593 Daan Quadvlieg Studentennummer: 01200218 Promotor: Prof. dr. Kurt Houf Promotor: Dr. Kris Baert Onderdeel van de Masterproef voorgelegd voor het behalen van de graad master in de diergeneeskunde Academiejaar: 2017 – 2018
Transcript of STUDIE VAN DE BACTERIËLE MICROBIOTA IN HONING...Deze stoffen en de mate waarin ze voorkomen bepalen...

STUDIE VAN DE BACTERIËLE
MICROBIOTA IN HONING
Aantal woorden: 9593
Daan Quadvlieg Studentennummer: 01200218
Promotor: Prof. dr. Kurt Houf
Promotor: Dr. Kris Baert
Onderdeel van de Masterproef voorgelegd voor het behalen van de graad master in de
diergeneeskunde
Academiejaar: 2017 – 2018

Universiteit Gent, haar werknemers of studenten bieden geen enkele garantie met betrekking tot de
juistheid of volledigheid van de gegevens vervat in deze masterproef, noch dat de inhoud van deze
masterproef geen inbreuk uitmaakt op of aanleiding kan geven tot inbreuken op de rechten van derden.
Universiteit Gent, haar werknemers of studenten aanvaarden geen aansprakelijkheid of
verantwoordelijkheid voor enig gebruik dat door iemand anders wordt gemaakt van de inhoud van de
masterproef, noch voor enig vertrouwen dat wordt gesteld in een advies of informatie vervat in de
masterproef

3
Voorwoord
Hierbij wil ik graag de personen bedanken die een bijdrage hebben geleverd aan de
totstandkoming van deze masterproef:
Als eerste wil ik mijn promotor, Prof. Dr. Kurt Houf, van harte bedanken voor zijn
niet-aflatende betrokkenheid gedurende het hele proces. Prof. Dr. Kurt Houf heeft
mij waardevolle adviezen gegeven bij elk deelproces van de masterproef. Vooral zijn
toegankelijke en persoonlijke manier van feedback, zoals de gesprekken via Skype,
weet ik ontzettend te waarderen.
Als tweede wil ik een woord van dank richten tot Dr. Kris Baert, enerzijds voor zijn
verhelderende feedback bij masterproef I anderzijds voor het feit dat hij mijn
co-promotor is gebleven toen door externe factoren het onderwerp aangepast moest
worden.
Tenslotte wil ik Drs. Pieter-Jan Kerkhof bedanken voor zijn ondersteuning bij het
verwerken en interpreteren van de onderzoeksresultaten en Sara Carton voor de
goede uitleg over de werking van het laboratorium en de persoonlijke rondleiding
aldaar.

4
Inhoudsopgave
1. Samenvatting…………………………………………………………………………………. p.5
2. Inleiding…………………………………………………………………………………………. p.6 2.1 Nut van de honingbij………………………………………………………………………………….. p.6
2.2 Wat is honing precies?.................................................................................... p.7
2.3 Toepassingen van honing ……………………………..…………………………………………… p.7
2.4 Micro-organismen en honing…………………………………………………………………….. p.8
2.4.1. Antibacteriële eigenschappen van honing…………………………………………p.8
2.4.2. Intrinsieke en extrinsieke invloeden op microbiële groei…....…………… p.9
2.4.3. Microbiële contaminatie………………………………………………………………….. p.10
2.4.4. Associatie tussen primaire bronnen en micro-organismen………………. p.11
2.4.5. Micro-organismen aangetroffen in honing………………………………………. p.12
2.5 Probleemstelling……………………………………………………………………………………….. p.14
2.6 Doelstelling……………………………………………………………………………………………….. p.15
3. Onderzoek………………………………………………………………………………….... p.15
3.1. Materiaal ………………………………………………………………………………………………… p.15
3.1.1. Geanalyseerde soorten honing …...………………………………………………… p.15
3.1.2. Mediums ……………………………….……………………………………………………… p.15
3.1.3. Overige materialen ………………………………………………………..……….…… p.16
3.2. Methodiek .……………………………………………………………………………………………. p.18
3.2.1. Staalname ……………………………………………………………………………………… p.18
3.2.2. Uitplating ……………………………………………………………………………………… p.18
3.2.3. MALDI-TOF MS staal voorbereiding en data-analyse ……………………. p.18
3.3. Uitleg van de weergaven van de resultaten.……………..……………………………. p.19
3.3.1. Betekenis van de betrouwbaarheidsscore……………………………………… p.19
3.3.2. Betekenis van de consistentie score…………………………………………….. p.20
3.4. Resultaten per honingsoort …………………………………………………………………….p.22
3.4.1. Klimophoning ………………………………………………………………………………..p.22
3.4.2. Koolzaadhoning …………………………………………………………………………….p.23
3.4.3. Paardenbloemhoning ………………………………………………………………….. p.25
3.5. Totaal resultaat ……………………………………………………………………………………… p.26
4. Discussie….………………………………………………….………………………………. p.28
Literatuurlijst ……….…………………………………………………………………….. p.31
Addenda ……………………………………………………………………………………… p.34

5
1. Samenvatting
Honing is een suikerrijk mengsel, geproduceerd door honingbijen. Het kent veel toepassingen in de
geneeskunde maar is ook een veelgebruikt voedingsmiddel. Door innovatie worden er steeds meer
toepassingsmogelijkheden voor honing in de voedselproductie bedacht. Bacteriën in honing kunnen
de kwaliteit en veiligheid van het product beïnvloeden. Een goede kennis van bacteriële microbiota in
honing kan helpen om potentiële biologische gevaren in beeld te brengen. Zodoende kan men
passende maatregelen bedenken. Met als doel, een veilig gebruik van honing en honing bevattende
producten. Tot nu toe was de kennis over het voorkomen van bacteriën in honing zeer beperkt en
dateert vaak reeds van een aantal decennia geleden. Wanneer de bacteriële microbiota bij een
beoordeling van honing werd geanalyseerd, beperkte men zich meestal tot een beperkt aantal
parameters. Staalnames maar ook de bacteriële analyses van honing zijn dan ook, in tegenstelling tot
veel andere levensmiddelen, niet gestandaardiseerd. Men gaat er immers vanuit dat door het hoge
suikergehalte, en bijgevolg een lage Aw, samen met de vorming van waterstofperoxide, de meeste
vegetatieve bacteriën worden afgedood of geremd. Met uitzondering van de detectie van Clostridium
species, wordt er weinig verder onderzoek uitgevoerd.
De algemene doelstelling van dit onderzoek was het isoleren en identificeren van bacteriën van een
aantal soorten koud geslingerde honing, namelijk van klimophoning, koolzaadhoning en
paardenbloemhoning. Dit zijn honing variëteiten die geen hitte behandeling bij de productie hebben
ondergaan. Hiervoor werd gebruik gemaakt van klassieke bacteriële analyses, met identificatie door
middel van de Matrix Assisted Laser Desorption Ionization Time of Flight Mass Spectrometry (MALDI-
TOF MS) methode. Zowel het kiemgetal als het aantal geïdentificeerde genera was daarbij eerder
verassend hoog, aangezien honing een groot aantal antibacteriële eigenschappen bezit. Enkel de
paardenbloemhoning bleek meerdere genera te bevatten die voor de mens pathogeen kunnen zijn. In
de meeste gevallen werden echter opportunische bacteriën geïsoleerd, afkomstig van de honing
productie. Desondanks is het wellicht verstandig om consumptie van honing te ontraden aan mensen
met een grotere kans op infectie door opportunistische species, zoals ouderen, jonge kinderen,
zwangere vrouwen en immunodeficiënte mensen.

6
2. Inleiding
2.1 Nut van de honingbij
Het belang van de honingbij (Apis mellifera) voor de mens kan nauwelijks worden overschat. De
honingbij zorgt voor de bevruchting van zeer veel gewassen die, als voedsel onontbeerlijk zijn voor de
mens (Olofsson and Vasquez, 2008). In de akkerbouw wordt men steeds afhankelijker van bestuiving
door, door imkers gehouden, honingbijen (Allsopp et al., 2008). Hieruit kan men afleiden dat het
belang van het bestaan van de honingbij, voor onze manier van voedselproductie, toeneemt.
Naast de belangrijke rol als bestuiver kan men honingbijen ook zien als een bijzondere groep
landbouwdieren. Honingbijen produceren immers diverse substanties die door de imker geoogst
kunnen worden en allen op een of andere manier nuttig zijn voor de mens1.
Zo produceren honingbijen propolis, enerzijds om de bijenkast af te dichten en zodoende het
binnendringen van micro-organismen te voorkomen, anderzijds ter bevordering van de gezondheid
van de honingbijen zelf. Propolis bevat naast stoffen die nadelig zijn voor bacteriën, virussen en
schimmels, substanties die onder meer anti-oxidatief en ontstekingsremmend werken. De precieze
samenstelling van propolis is afhankelijk van het gebied waar de honingbijen gehuisvest zijn. De
plaats van herkomst is dus van invloed op de eigenschappen van propolis. Het is gebleken dat
propolis uit een bepaald gebied in Brazilië een cytotoxische activiteit had tegen diverse lijnen van
maligne tumoren, vergelijkbaar met courant gebruikte chemotherapeutica (de Mendonca et al.,
2015).
Een ander weinig gekend product is bijengif. Het gif van de honingbij werd reeds onder meer bij de
behandeling van bepaalde vormen van artritis toegepast alsook bij sommige patiënten met multiple
sclerose. Bij de laatst genoemde aandoening werd symptomatische verbetering gezien bij patiënten,
alsook een vertraging van deze progressieve ziekte (Fratellone and Fratellone, 2016).
Voedsterbijen, dit zijn jonge vrouwelijke bijen die instaan voor het voeden van het bijenvolk,
produceren koninginnengelei2. Koninginnengelei is de voeding voor de (toekomstige) koningin van
een bijenvolk. Dit exclusievere product wordt door de mens gebruikt in cosmetica alsook in
voedingssupplementen. Diverse gezondheid bevorderende eigenschappen worden toegeschreven
aan koninginnengelei, onder meer het vermogen om verouderingsprocessen te vertragen. Bij
proefdieren werd vastgesteld dat koninginnengelei de eigenschap had om een verhoogde bloeddruk
tegen te gaan, alsook een verhoogde cholesterolspiegel in het bloed (Ramadan and Al-Ghamdi,
2012).
Het bekendste product dat door honingbijen wordt geproduceerd is zonder enige twijfel honing. Enkel indien het product afkomstig is van de honingbij mag het volgens de Raad van de Europese Unie ook daadwerkelijk honing heten3. Dit ten spijt van de mensen die hommels in hun tuin houden, immers mocht het hen al lukken iets te oogsten, dan nog mogen ze het geen honing noemen.
1 anoniem, 2011a. Bijenproducten. http://www.imkerpedia.nl/wiki/index.php?title=Bijenproducten (accessed 03-04-2017
2017). 2 Anoniem, 2011b. Voedsterbij.
http://www.imkerpedia.nl/wiki/index.php?title=Voedsterbij (accessed 04-04-2011 2017).
3 Richtlijn 2001/110/EG

7
2.2 Wat is honing precies? Honing is een suikerrijke oplossing gecreëerd door honingbijen. Het wordt voornamelijk gevormd uit nectar, maar kan in principe gevormd worden uit elke zoete excretie van plantaardige oorsprong. Het vochtgehalte bedraagt gemiddeld 17,2 procent. De kwantitatief belangrijkste suikers aanwezig in honing zijn glucose en fructose (Snowdon and Cliver, 1995). Dit zijn echter slechts twee van de 24 verschillende mogelijk aanwezige suikers in honing. De belangrijkste karakteristieken van honing zijn een gevolg van de aanwezigheid van de eerder genoemde suikers. Honing bevat naast suikers en water nog andere substanties, zoals pigmenten, zuren en mineralen. Deze stoffen en de mate waarin ze voorkomen bepalen in grote mate de verschillen tussen de diverse honingsoorten (Amna Jabbar Siddiqui, 2016).
2.3 Toepassingen van Honing Honing wordt door veel mensen geapprecieerd als zoetstof of broodbeleg, maar het kan op veel meer manieren worden gebruikt. In oude beschavingen werd honing reeds door artsen gebruikt bij diverse kwalen, maar ook vandaag de dag wordt honing nog in de geneeskunde gebruikt. Een scala aan wondsoorten wordt tegenwoordig succesvol behandeld met honing, zo verloopt de reiniging van geïnfecteerde wonden beter, verdwijnen onaangename geuren vlugger en wordt de heling gestimuleerd, waardoor er een betere regeneratie plaatsvindt en er minder littekens ontstaan. In de pediatrie heeft men vastgesteld dat diarree bij een bacteriële gastro-enteritis sneller stopt wanneer men honing gebruikt in plaats van glucose bij de rehydratietherapie. Ook in de oftalmologie wordt honing succesvol toegepast als therapeutisch middel, onder andere bij letsels aan de cornea (Molan, 1999). Honing kent, zoals hierboven vermeld werd, diverse toepassingen in de geneeskunde. Het wordt
echter ook vaak gebruikt als ingrediënt. Door innovatie worden er steeds meer
toepassingsmogelijkheden voor honing bedacht. Micro-organismen in honing kunnen de kwaliteit en
veiligheid van het product beïnvloeden. Een goede kennis van de aanwezige microbiota in honing is
van belang om een veilig gebruik van honing en honing bevattende producten te kunnen garanderen.
De honingindustrie heeft dan ook, in toenemende mate, interesse in de microbiële samenstelling van
honing (Snowdon and Cliver, 1995).
De huidige eisen die aan in de handel te brengen honing worden gesteld zijn, voor wat betreft micro-
organismen, veelal geëxtrapoleerd uit eisen gesteld aan andere levensmiddelen. Honing is echter een
uniek product, niet in het minst door zijn antibacteriële eigenschappen. Hierdoor zijn sommige eisen,
voorschriften en adviezen mogelijk niet relevant of geschikt voor honing (Snowdon and Cliver, 1995).
Zoals het advies van het wetenschappelijk comité van het federaal agentschap voor de veiligheid van
de voedselketen om honing te aanzien als ongeschikt voor kinderen jonger dan één jaar wegens het
sporadisch voorkomen van Clostridium botulinum in honing. Aangezien een infectie met Clostridium
botulinum bij kinderen jonger dan één jaar aanleiding kan geven tot infantiel botulisme4.
4 Advies 34-2006, het wetenschappelijk comité van het federaal agentschap voor de veiligheid van de voedselketen

8
Een goede kennis van de in honing aanwezige micro-organismen kan helpen om potentiële biologische
gevaren in beeld te brengen. Zodoende kan men passende maatregelen bedenken, ten gunste van de
volksgezondheid en voedselveiligheid.
2.4 Micro-organismen en honing
2.4.1. Antimicrobiële eigenschappen van honing
Micro-organismen zijn, in relatie tot honing, enkel interessant als ze niet afgedood worden in honing.
In honing is de concentratie suiker relatief hoog en heerst er een zuur milieu. Tevens heeft honing
specifieke antimicrobiële en met name antibacteriële eigenschappen. Deze laatste verdienen
aandacht aangezien zij een alternatief zouden kunnen vormen voor het gebruik van antibiotica bij
infecties met multiresistente bacteriën (Sereia et al., 2017).
Een kleine eeuw geleden werden deze antimicrobiële eigenschappen al beschreven (Sacket, 1919).
Het zou erna nog meer dan veertig jaar duren voordat het enzym ontdekt zou worden, dat
verantwoordelijk is voor één van de belangrijkste antimicrobiële eigenschappen in honing. Dit
enzym, genaamd glucose oxidase, produceert waterstofperoxide en gluconzuur uit kleine
hoeveelheden glucose. Met name het geproduceerde waterstofperoxide heeft een bactericide
werking. Niet-bacteriële micro-organismen zijn echter weinig tot niet gevoelig hieraan. Glucose
oxidase is overigens hitte-labiel en wordt dus vernietigd bij pasteuriseren van honing (White et al.,
1963).
Smith et al. (1969) zagen bij onderzoek van acht soorten honing dat niet alle honingsoorten een even
sterk antibacterieel effect hebben tegenover alle onderzochte Gram negatieve bacteriën. Allen et al.
(1991) concludeerden dat de bloemsoort waarvan de nectar afkomstig was, een invloed had op de
antibacteriële activiteit van de honing. Willix et al. (1992) onderzochten monoflorale honingsoorten
en stelde eveneens vast dat de soort antibacteriële activiteit verschillend was tussen de diverse
soorten honing.
Tysset and de la Roy (1974) somden een lijst karakteristieken van honing op, die aan de grondslag
zouden kunnen liggen van het antibacterieel karakter van honing: “een lage eiwitconcentratie, een
hoge koolstof-stikstof ratio en een zuur milieu zoals in honing zijn niet voordelig voor bacteriële
proliferatie. De reducerende suikers die in hoge gehaltes aanwezig zijn in honing zorgen voor een
laag redox-potentiaal, hetgeen de groei van aerobe bacteriën en gisten remt. De hoge viscositeit gaat
spreiding van kiemen in de honing tegen en beperkt de toestroom van zuurstof, noodzakelijk voor de
groei van aerobe micro-organismen, in honing. Door de hoge osmotisch druk, dehydrateren micro-
organismen en sterven deze sneller af.”
Molan (1992) noemde, net als Tysset en de la Roy (1974), de lage pH, de hoge osmolariteit (Tabel 1)
resulterend in een lage wateractiviteit (Aw) en het door glucose oxidase gevormde
waterstofperoxide als factoren die de antibacteriële activiteit van honing tot stand brengen. Molan
(1992) besprak echter nog andere factoren zoals de aanwezigheid van een antioxidant genaamd
pinocembrine, een lysozyme, zuren (waaronder fenolcarboxylzuren) en daarnaast ook de mogelijke
aanwezigheid van terpenen, benzylalcohol en gasvormige substanties. Het geheel van deze factoren
zou een antimicrobieel effect hebben, waar niet alleen bacterie species gevoelig aan zouden zijn,

9
maar ook schimmels zoals Aspergillus spp. en Penicillum spp., en gisten (Candida spp. en
Saccharomyces spp.).
Tabel 1: Samenstelling en belangrijkste eigenschappen van honing
(Snowdon and Cliver, 1995; White et al., 1962b)
Samenstelling Procentueel
Vochtgehalte (gemiddeld 17.2%)
fructose 38.4%
glucose 30.3%
sucrose Gemiddeld 1.3%
overige koolhydraten
12%
Mineralen 0.169%
Eiwitten 0.169%
pH 3.4-6.1 (gemiddeld 3.9)
Wateractiviteit (Aw)
0.5-0.6
Werksterbijen zijn jonge vrouwelijke honingbijen, verantwoordelijk voor het verzamelen van nectar.
Gilliam (1978) rapporteerde dat en er een lage prevalentie is van infectie van B. cereus in
werksterbijen en merkte tevens op dat honing een antibacteriële activiteit vertoonde tegen B.
cereus. Gilliam (1978) concludeerde hieruit dat de lage prevalentie van B. cereus bij werksterbijen het
gevolg zou kunnen zijn van consumptie van honing door deze werksterbijen.
2.4.2. Intrinsieke en extrinsieke invloeden op microbiële groei
De groei van micro-organismen wordt in honing, net zoals in alle voedingsmiddelen, beïnvloed door
intrinsieke en extrinsieke factoren. Intrinsieke factoren omvatten karakteristieken van de honing,
zoals de pH, de oxidatie-reductie-potentiaal en de wateractiviteit. De intrinsieke factoren van honing
hebben een bacteriostatisch tot zelfs bacteriocide effect en werden hierboven reeds uitvoeriger
besproken (Snowdon and Cliver, 1995).
Wat betreft extrinsieke factoren kunnen, ten aanzien van microbiële groei in honing, onder meer
omgevingstemperatuur, relatieve luchtvochtigheid, de aanwezigheid van gassen zoals stikstof,
zuurstof en koolstofdioxide én de partiële druk van elk aanwezig gas, tijdens bewaring als belangrijk
worden aanzien (Jay, 1992).
De natuur als voorbeeld
Tijdens de omvorming van nectar naar honing wordt onder meer het vochtgehalte van het product
verlaagd. Wanneer de volledige omzetting tot honing is gebeurd, wordt deze door de bijen

10
opgeslagen in raten. Een raat is opgebouwd uit honderden cellen, opgebouwd uit was. De cellen in
die raten, worden na het vullen met honing, luchtdicht afgesloten met een laagje bijenwas. Op deze
manier wordt microbiële groei tijdens het bewaren voorkomen.
Snowdon and Cliver (1995) adviseerden om de natuur als voorbeeld te nemen bij het voorkomen van
bederf na het oogsten van de honing. Enerzijds is het volgens hen van belang om zo min mogelijk
lucht (en zodoende zuurstof) in het recipiënt te hebben, anderzijds moet men de relatieve
luchtvochtigheid en de temperatuur in de omgeving van de honing beperken, om condensatie te
vermijden. De achterliggende gedachte is dat de groei van aërobe micro-organismen direct in de
hand gewerkt wordt door de aanwezigheid van zuurstof en dat een toename aan vocht in de honing
ook nog een indirect effect heeft op de groei van micro-organismen, namelijk door middel van dilutie
van de suikers in de honing. Wanneer men honing als ingrediënt in voedingsmiddelen gebruikt, vindt
er ook eenzelfde verdunning plaats, waardoor het remmend effect op de groei van micro-
organismen deels verloren gaat. Honing kan in elk geval als vector dienen voor micro-organismen.
Waarbij het risico bestaat dat ze in het voedingsmiddel, in tegenstelling tot in de honing, wel kunnen
vermeerderen en zelfs voor bederf kunnen zorgen (Snowdon, 1999).
Micro-organismen die voor de mens pathogeen zijn, zouden niet van nature in honing kunnen
voorkomen en dus enkel via contaminatie in honing terecht kunnen komen. De overlevingstijd van
deze organismen in honing tijdens bewaring is sterk afhankelijk van de temperatuur. Bij
kamertemperatuur is de overlevingstijd sterk beperkt (Snowdon and Cliver, 1995).
Bewaring bij minder dan 10°C kan de overlevingstijd van bepaalde pathogene bacteriën echter sterk
verlengen, zo kan Salmonella dublin bij deze temperatuur meer dan twee jaar overleven in honing
(Tysset and Durand, 1976).
Fermentatie wordt bij maximaal 10°C én een gelijktijdig relatieve luchtvochtigheid lager dan 50%
juist voorkomen. Suikertolerante gisten kunnen bij deze omstandigheden namelijk niet prolifereren
(Tysset and Rousseau, 1981). Wanneer honing een vochtgehalte van 17% of meer heeft is het
vatbaar voor fermentatie. Vanaf 19% is de kans op fermentatie zelfs zeer groot, hetgeen een sterke
verkorting van de houdbaarheidstermijn impliceert (Graham, 1992).
2.4.3. Microbiële contaminatie
Zoals hierboven vermeld produceren honingbijen honing uit nectar. Gilliam et al. (1983) konden
nauwelijks tot geen micro-organismen uit nectar isoleren. Aangezien het substraat, waaruit bijen
honing maken, dus vrijwel steriel is, kunnen we concluderen dat contaminatie met micro-organismen
tijdens het ontstaan en gedurende de verwerking van honing, een zeer belangrijke bijdrage levert in
het tot stand komen van de specifieke samenstelling van micro-organismen in honing.
Microbiële contaminatie van honing is onder te verdelen in primaire en secundaire contaminatie.
Primaire contaminatie vindt plaats voor de oogst van de honing. Voorbeelden zijn contaminatie vanuit
aarde, stuifmeel, lucht en stof en het verteringsstelsel van de honingbij, aangezien nectar en honing
vervoerd worden in de honingmaag.
Secundaire contaminatie vindt plaats tijdens en na de oogst van de honing. Voorbeelden van bronnen
verantwoordelijk voor microbiële contaminatie van honing in deze fase zijn, net als ook voor veel

11
andere levensmiddelen, mensen die betrokken zijn bij het productieproces, lucht, niet-steriele
materialen en gebouwen alsook kruiscontaminatie met andere producten.
De mate van secundaire contaminatie is sterk afhankelijk van de mate van hygiëne toegepast door
imkers en verwerkers. Wanneer men van een micro-organisme duizenden kolonievormende eenheden
detecteert in een honingstaal, terwijl men eigenlijk niet verwacht dit micro-organisme in honing aan
te treffen, en deze ook niet in honing kan vermeerderen, kan men ervanuit gaan dat het een secundaire
vorm van contaminatie betreft (Snowdon and Cliver, 1995). Uit een onderzoek van Tysset et al. (1970)
is gebleken dat er grotere kans is op secundaire contaminatie van honing dan op primaire
contaminatie, hierbij dient wel opgemerkt te worden dat deze conclusie in dit onderzoek gebaseerd
is op analyses van slechts twaalf stalen.
2.4.4. Associatie tussen primaire bronnen en micro-organismen
Vaak zijn bepaalde micro-organismen eigen aan bepaalde, specifieke onderdelen van een milieu (Jay,
1992). Zo is de kans groter om micro-organismen die in en op honingbijen, bijenkasten, bloemen en
aarde leven, aan te treffen in honing dan om een willekeurig ander micro-organisme aan te treffen.
Het is vanuit dat oogpunt interessant om de voornaamste primaire contaminatiebronnen en de ermee
geassocieerde micro-organismen te benoemen:
Aarde
In aarde worden vooral bacteriën aangetroffen zoals: Actinetobacter, Bacillus, Clostridium,
Corynebacterium, Pseudomonas, Psychrobacter en Vagococcus (Snowdon and Cliver, 1995).
Lucht en stof
Lucht en stof zijn eveneens bronnen voor contaminatie met Bacillus en Clostridium maar ook met
Micrococcus species (Snowdon and Cliver, 1995).
Het verteringsstelsel van de honingbij
Aangezien nectar in een deel van het verteringsstelsel van de honingbij vervoerd en bewerkt wordt, zijn micro-organismen die eigen hieraan zijn ook van belang. Onderstaande micro-organismen werden bij onderzoek van stalen van het verteringsstelsel van honingbijen reeds waargenomen: niet-geïdentificeerde pleomorfe bacteriën, Bacillus spp, Enterobacteriaceae, Penicillium spp., Aspergillus spp., Bacteridium (sic), Streptococcus species, Clostridium species, Achromobacter, Citrobacter, Enterobacter, Escherichia coli, Flavobacterium, Klebsiella, Proteus, Pseudomonas en gisten (veelal Torulopsis spp) (Snowdon and Cliver, 1995; Gilliam et al., 1988). Volgens Gilliam et al. (1983) zouden de micro-organismen, die het microbioom van het
verteringsstelsel van de honingbij vormen, afkomstig zijn van opgenomen stuifmeel en uitgewisseld
worden binnen het bijenvolk middels het doorgeven van voedsel. Dit gesuggereerde causale verband
tussen de samenstelling van het microbioom van het verteringsstelsel van de honingbij en stuifmeel
wordt ondersteund door conclusies uit eerdere onderzoeken. Gilliam (1971) stelde namelijk vast dat
het verteringsstelsel van de pas ontpopte honingbij vaak nog geen micro-organismen bevat, terwijl er
in het verteringsstelsel van honingbijen die enkele dagen ouder zijn en dus stuifmeel hebben kunnen
opnemen, met regelmaat bacteriën en schimmels aangetoond kunnen worden. Gilliam (1978)
suggereerde dat stuifmeel de belangrijkste toevoer zou zijn wat betreft de microbiota in het

12
verteringsstelsel van werksterbijen, aangezien nectar, zoals eerder vermeld, veelal vrij van micro-
organismen zou zijn. Hierbij moet dan wel opgemerkt worden dat nectar reeds gecontamineerd zou
kunnen worden, in ieder geval met gisten, vanuit de bloem en door aarde, alvorens deze door de
honingbij verzameld wordt (Graham, 1992). De grootste bron van contaminatie van nectar is echter
de honingbij zelf. Contaminatie van nectar door de honingbij zou in de meeste gevallen reeds bij het
verzamelen ervan plaatsvinden (Troller, 1979).
Bijenvoeder Aangezien het merendeel van de in de bijenkast opgeslagen honing wordt verwijderd door de imker, is substitutie nodig om hongersnood in het bijenvolk in het najaar en de winter te voorkomen. Imkers verstrekken daarom suikerrijke mengsels aan bijenvolken. Nakano et al. (1992) suggereren dat deze suikerrijke mengsels een contaminatiebron van sporen van C. botulinum zouden kunnen zijn. Mengsels die sporen bevatten, moeten tijdens de productie of verwerking gecontamineerd zijn, aangezien suiker op zichzelf een steriel product is. De bron van contaminatie van de suikers is echter niet gekend.
2.4.5. Micro-organismen in honing
Micro-organismen aangetroffen in honing, behoren steeds tot het reik van de bacteriën of tot het
reik van de fungi. Tot dit laatste reik behoren zowel schimmels als gisten. Hieronder zullen eerst de
schimmels en de gisten besproken worden en vervolgens de bacteriën.
Schimmels
Snowdon and Cliver (1995) vermelden een lage incidentie in het voorkomen van schimmels in honing.
Wanneer er al schimmel in honingstalen aanwezig is, gaat het volgens hen nagenoeg altijd om lage
concentraties. Dit werd weliswaar geconcludeerd op basis van niet gepubliceerde data uit een verslag
dat door de honingindustrie werd opgesteld en niet op basis van wetenschappelijke publicaties.
Als voorbeeld van schimmels die in honing aangetroffen kunnen worden, kunnen Aspergillus spp. en
Penicillum spp. worden genoemd. Van Aspergillus spp. is bekend dat ze enkel in de vorm van sporen
in honing kunnen voorkomen. Dit betekent dat Aspergillus spp. niet in honing kunnen vermeerderen
(Graham, 1992).
Gisten
Gisten onderscheiden zich van de meeste andere micro-organismen doordat ze in staat zijn om te
prolifereren in honing. Dit kunnen ze door hun vermogen om te overleven in een zuur milieu en
doordat ze niet geremd worden door de aanwezige sucrose.
De luchtvochtigheid in het gebied waar de bloemen zich bevinden, waaruit de honingbijen de nectar
onttrekken, is van invloed op de hoeveelheid gist in de daaruit geproduceerde honing. Indien de
luchtvochtigheid in een bepaald gebied erg hoog is, kan de gistvorming in de honing dusdanig
uitgesproken zijn, dat de zelfs de raat erdoor kan bederven.

13
Saccharomyces spp. vormen nagenoeg altijd de kwantitatief belangrijkste gisten in honing. De
concentratie aan gisten in honing verschilt zeer sterk tussen verschillende soorten. Dit verschil kan
oplopen tot een factor van maar liefst één miljoen tussen sommige soorten honing.
Bij industriële verwerking van honing worden maatregelen genomen om de groei van gisten tegen te
gaan. Hierdoor bedraagt de hoeveelheid gisten in gecommercialiseerde honing, in de meeste
gevallen, niet meer dan enkele honderden kolonievormende eenheden per gram (kve/g) (Snowdon
and Cliver, 1995).
Bacteriën
De bacteriën die men uit honing kan isoleren, kunnen in vier categorieën worden ingedeeld:
1. Indicatoren voor microbiologische kwaliteit
Deze groep wordt gevormd door de totale en thermotolerante bacteriën en Escherichia coli. Het zijn
Gram-negatieve bacillen. Ze behoren tot de familie van de Enterobacteriaceae en vormen in het
algemeen geen groot gevaar voor de voedselveiligheid of volksgezondheid. Ze kunnen enkel in honing
terechtkomen door secundaire contaminatie. Daarom kunnen ze worden gebruikt als indicator
species. Men kan ze gebruiken als maatstaaf voor de toegepaste hygiëne. Een te hoog gehalte kan
wijzen op een product met een slechte microbiële kwaliteit, dat potentieel micro-organismen bevat
die wél schadelijk kunnen zijn voor de gezondheid van de consument (Sereia et al., 2017).
2. Ziekteverwekkers voor de mens
De belangrijkste ziekteverwekker voor de volwassen mens, die in honing kan worden aangetroffen, is
Salmonella. Consumptie van een met Salmonella, gecontamineerd levensmiddel kan leiden tot infectie
bij zowel mens als dier. Men ziet een hoge morbiditeit en mortaliteit bij de mens na opname van
gecontamineerd voedsel (Sereia et al., 2017).
Clostridium botulinum is een andere ziekteverwekker bij de mens die in honing aanwezig kan zijn en is
vooral gevaarlijk voor zuigelingen. Het is een sporenvormende bacterie. De sporen van deze bacterie
zijn resistent in de omgeving en kunnen zeer lang overleven in honing. Honing is het enige
voedingsmiddel waarvan zuigelingen, na opname, botulisme kunnen krijgen. Het toedienen van
honing aan kinderen jonger dan één jaar oud wordt dan ook ten strengste ontraden. Hoewel deze
waarschuwing veelal op het etiket van commercieel verkrijgbare honing vermeld staat, zijn er al veel
gevallen van botulisme bij baby’s gemeld. Bacillus cereus en Clostridium perfringes zijn net als
Clostridium botulinum pathogene, sporenvormende bacteriën, die echter ook bij volwassenen
aanleiding tot ziekte kunnen geven (Estevinho et al., 2012; Sereia et al., 2017).
3. Ziekteverwekkers voor bijen
De enige bacteriële ziekteverwekker waar honing regelmatig op onderzocht wordt is Paenibacillus
larvae larvae. Deze bacterie kan aanleiding geven tot Amerikaans Vuilbroed, een wettelijke bestreden
ziekte met verstrekkende gevolgen. Bij een infectie met Paenibacillus larvae larvae dient het bijenvolk
zelfs te worden vernietigd (Iurlina, 2005).
4. Commensale micro-organismen
Dit zijn micro-organismen die gewoonlijk aangetroffen worden in honing, zoals bepaalde gisten.

14
2.5 Probleemstelling
Tot nu toe is de kennis over het voorkomen van bacteriën in honing zeer beperkt, en dateert vaak
reeds van een aantal decennia geleden. Wanneer de bacteriële microbiota bij een beoordeling van
honing worden geanalyseerd dan beperkt men zich meestal tot een relatief klein aantal parameters
zoals het totaal aëroob kiemgetal en de aanwezigheid van Clostridia species, zonder daarbij verder
taxonomisch te differentiëren. Of als men bijvoorbeeld de geschiktheid voor humane consumptie wil
nagaan, zal men meestal het totaal aantal fecale coliformen vaststellen, om de hygiëne van de imker
en verwerkers te kunnen beoordelen. Eerder zelden zullen andere (ziekte) kiemen, zoals Clostridia en
Salmonella worden onderzocht (Estevinho et al., 2012).
Bacteriële analyses worden eveneens uitgevoerd wanneer een bijenvolk wordt verdacht van een
aangifteplichtige ziekte zoals Amerikaans vuilbroed. Het verantwoordelijk agens voor deze ziekte is,
zoals hierboven vermeld, een bacterie genaamd Paenibacillus larvae larvae. De bijen kunnen op
verschillende manieren besmet raken5, maar de ziekte wordt voornamelijk overgebracht via besmette
honing (Alippi et al., 2004).
Zoals hierboven geïllustreerd worden bacteriële analyses van honing wel toegepast om specifieke
groepen van bacteriën of eventuele pathogenen aan te tonen, maar er is dan geen aandacht voor
andere bacteriën die in honing voorkomen. Er zijn zodoende weinig tot geen gegevens over het
bacterieel microboom van de diverse, courant geconsumeerde soorten honing, alsook over de
veranderingen in de bacteriële samenstelling van honing gedurende de bewaring.
Staalnames maar ook de bacteriële analyses van honing zijn dan ook, in tegenstelling tot veel andere
levensmiddelen, niet gestandaardiseerd (Snowdon and Cliver, 1995). De manier van oogsten heeft ook
invloed op de resultaten van bacteriële analyses van honing. Bij het verwijderen van de honing uit de
raten kan men immers kiezen tussen ‘koud slingeren’ en ‘warm slingeren’. Bij beide methoden wordt
honing uit de raten gehaald door middel van een centrifuge, die men een honingslinger noemt.
Wanneer men ‘warm slingert’ verhoogt men de temperatuur van de raten, hierdoor loopt de honing
er sneller uit en verloopt het slingeren makkelijker. De temperatuur dient echter minder dan 62.8
graden Celsius te bedragen, aangezien de bijenwas waaruit de raten bestaan bij deze temperatuur
smelt6. Deze verhoogde temperatuur kan invloed hebben op de bacteriën, die aanwezig zijn in honing
en zodoende de bacteriële samenstelling wijzigen. Bij ‘koud slingeren’ verhoogt men de temperatuur
niet en zal er dus geen invloed zijn op de bacteriële samenstelling door het slingeren.
5 FAVV, 2017. Aangifteplichtige ziekten en schadelijke organismen (KB 3 februari 2014)
http://www.afsca.be/bijenteelt/dierengezondheid/#aangifteplichtige (accessed 03-04-2017 2017).
6 FISHER, SCIENTIFIC, 1997. Material Safety Data Sheet Beeswax. https://fscimage.fishersci.com/msds/02556.htm (accessed 14-05-2018 2018).

15
2.6 Doelstelling
De algemene doelstelling van dit onderzoek is het isoleren en identificeren van bacteriën van een
aantal soorten koud geslingerde honing: een gecommercialiseerde paardenbloemhoning,
klimophoning en koolzaadhoning.
Het karakteriseren van het bacteriële microbioom van honing heeft diverse potentiële toepassingen
met betrekking tot de uitvoer van kwaliteitscontroles en de detectie van fraude. Zo is het denkbaar
dat men rauwe honing zou kunnen onderscheiden van verhitte honing op basis van de kwantiteit en
de soorten aanwezige bacteriën. Men kan wellicht een database creëren van de bacteriële microbiota
van diverse soorten honing uit diverse gebieden, om aangeboden honing op het vlak van hun
bacteriële microbiota te vergelijken met referentiestalen, om de claim van oorsprong op het etiket al
dan niet te bevestigen.
3. Onderzoek
3.1. Materialen
3.1.1. Geanalyseerde soorten honing
Voor dit onderzoek werd gebruik gemaakt van een gecommercialiseerde paardenbloemhoning,
klimophoning en koolzaadhoning (aangekocht bij Van Weyn’s Honingbedrijf). Alle stalen zijn afkomstig
van ‘koud geslingerde’ honing.
3.1.2. Isolatiemethoden
Voor de bacteriële analyse van de honingstalen werd steeds gebruik gemaakt van genormaliseerde
methoden voor de analyse van voedingsmiddelen. Voor de meeste parameters zijn immers geen
specifieke genormaliseerde methoden beschikbaar. De volgende parameters werden daarbij bepaald:
totaal aëroob kiemgetal, op Plate Count Agar (PCA); totaal anaëroob kiemgetal; telling van
vermoedelijke Pseudomonas spp.; telling van Bacillus cereus; telling van Staphylococcus aureus; telling
van schimmels en gisten; telling van totaal aantal melkzuurbacteriën; telling van totaal aantal
coliforme bacteriën.
Van elk honingstaal werd 100 gram onderzocht en uitgeplaat op diverse media (Tabel 2 en 3) en
geïncubeerd bij een specifieke temperatuur en atmosfeer. Er werd zowel gebruik gemaakt van
universele alsook van selectieve media. Informatie over de samenstelling en pH van de agars, alsook
de verantwoordelijke producent is te vinden in addendum A.1.
Tabel 2: De in dit onderzoek toegepaste universele media met de toegepaste atmosfeer, de
incubatietemperatuur en de duur van incubatie.
Naam (afkorting)
van universeel
medium
Atmosfeer Incubatie-
temperat
uur
Incubatietijd
Plate Count Agar
(PCA)
Aëroob 30 °C 2 dagen
16
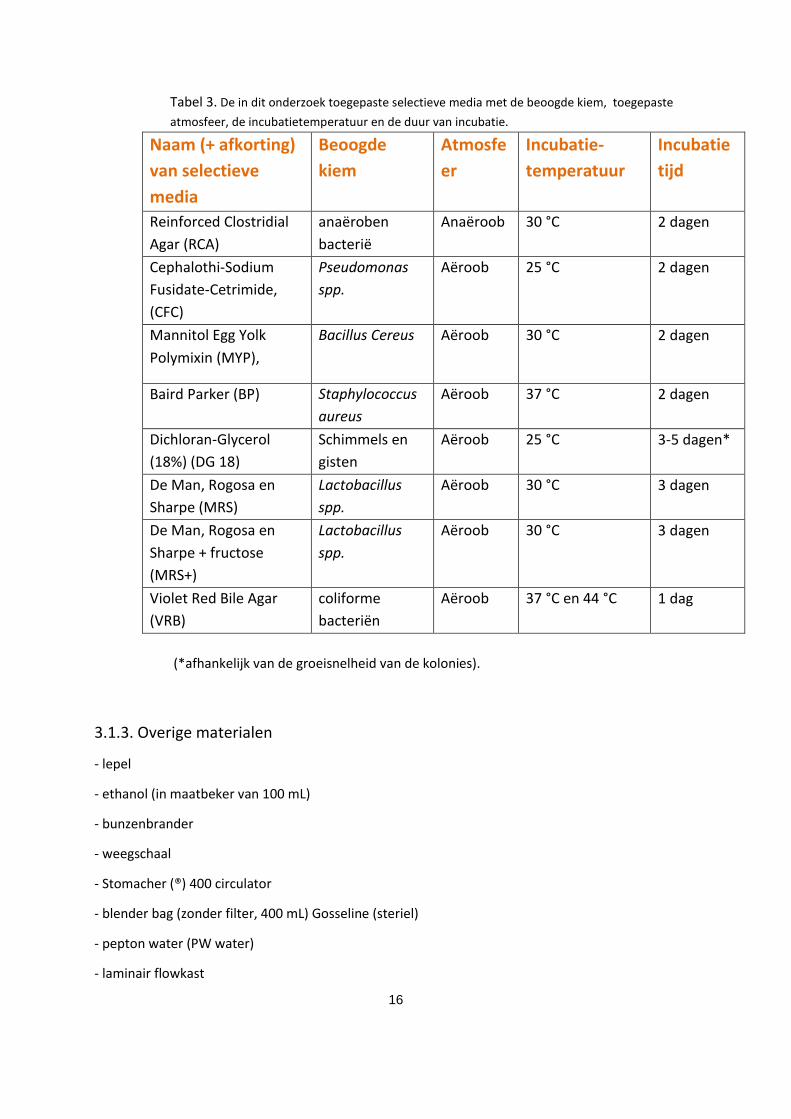
Tabel 3. De in dit onderzoek toegepaste selectieve media met de beoogde kiem, toegepaste
atmosfeer, de incubatietemperatuur en de duur van incubatie.
Naam (+ afkorting)
van selectieve
media
Beoogde
kiem
Atmosfe
er
Incubatie-
temperatuur
Incubatie
tijd
Reinforced Clostridial
Agar (RCA)
anaëroben
bacterië
Anaëroob 30 °C 2 dagen
Cephalothi-Sodium
Fusidate-Cetrimide,
(CFC)
Pseudomonas
spp.
Aëroob 25 °C 2 dagen
Mannitol Egg Yolk
Polymixin (MYP),
Bacillus Cereus Aëroob 30 °C 2 dagen
Baird Parker (BP) Staphylococcus
aureus
Aëroob 37 °C 2 dagen
Dichloran-Glycerol
(18%) (DG 18)
Schimmels en
gisten
Aëroob 25 °C 3-5 dagen*
De Man, Rogosa en
Sharpe (MRS)
Lactobacillus
spp.
Aëroob 30 °C 3 dagen
De Man, Rogosa en
Sharpe + fructose
(MRS+)
Lactobacillus
spp.
Aëroob 30 °C 3 dagen
Violet Red Bile Agar
(VRB)
coliforme
bacteriën
Aëroob 37 °C en 44 °C 1 dag
(*afhankelijk van de groeisnelheid van de kolonies).
3.1.3. Overige materialen
- lepel
- ethanol (in maatbeker van 100 mL)
- bunzenbrander
- weegschaal
- Stomacher (®) 400 circulator
- blender bag (zonder filter, 400 mL) Gosseline (steriel)
- pepton water (PW water)
- laminair flowkast

17
- warmwaterbad (35 °C voor PW water en 50 °C voor mediums MYP en BP)
- rekje
- afvalbakje
- mediums als voedingsbodems (zie vorige paragraaf voor details)
- Spiral plater (Easy spiral®)
- Cupjes (steriel)
- 1 Liter fles ethanol
- 1 Liter fles steriel water
- 1 pipetboy
-10 mL pipet (steriel)
- L-spreader (Drigalski)
- incubatoren (voor 25°C, 28 °C, 30°C, 37°C, 44°C)
- luchtdichte pot
- Gaspak ™ (BD)
- Anaerobic indicator (Firma: Oxoid)
- plastic dozen voor opslag
- alcoholstiften
- 100 PCA platen (voor elke transfer van een bacteriële kolonie)
- Microloops (10 microliter)
- handschoenen
- afvalbak voor biologisch afbreekbaar afval
- koelkast (ingesteld op 4 °C)
- diepvries (ingesteld op minus 20°C)
- glycerol (15%) en TSB (Tryptone Soya Broth)
- Cryotube boxen
- +- 400 PCA platen (voor uitplaten vanuit glycerol)
- plate setups voor MALDI-TOF MS en glycerol stocks

18
3.2 Methodiek
3.2.1. Staalname
De staalname van elke honing werd onder steriele omstandigheden uitgevoerd, daarom werd er
gebruik gemaakt van een bunzenbrander, de lepel waarmee een staal werd genomen, werd
voorafgaande aan de staalname in alcohol ondergedompeld en de tafel werd ontsmet. Er werd steeds
100 gram honing van één staal gemengd op 1:1 (w/w) met 100 gram steriel peptonwater (Pw).
3.2.2. Uitplating
Na homogenisatie in een stomacher, 2 minuten bij normale snelheid, werd een tienvoudige
verdunningsreeks in Pw aangemaakt, waarna 100 µL van elke verdunning in duplo werd uitgeplaat op
de diverse media. De media werden geïncubeerd in de atmosfeer en temperatuur zoals hierboven
beschreven. Na incubatie werden de verschillende kolonies op de platen geteld (niet-selectieve
platen: platen met een bacteriële groei tussen 30 en 300 kolonies, selectieve platen: tussen 15 en
150 kolonies). Na telling werden de kolonies individueel geoogst en overgezet op een vaste agar
(PCA/ MRS). Na een tweede incubatie werd de bacteriële groei overgebracht in 5 mL Trypticase soy
broth (TSB) en glycerol en ingevroren tot verdere analyse.
3.2.3. MALDI-TOF MS staalvoorbereiding en data-analyse
Voor de identificatie door middel van MALDI-TOF MS werden de bewaarde isolaten overnacht
opgekweekt op PCA volgens de incubatietijd zoals weergegeven in tabel 2.
De isolaten werden vervolgens voorbereid voor identificatie door middel van de directe
smeermethode. Hierbij werd een kleine hoeveelheid bacterieel cel materiaal van de groeiplaten direct
op de roestvrij stalen ‘Target 96 sample spots’ gesmeerd met een houten tandenstoker. Vervolgens
werd het organisch materiaal bedekt met een fixatiematrix 1 µl α-Cyano-4-hydroxycinnamisch zuur
(HCCA) matrix en werden de platen gedurende vijftien minuten aan de lucht gedroogd, alvorens de
analyses op te starten
Matrix voorbereiding
HCCA (α-Cyano-4-hydroxycinnamisch zuur) matrix werd toegevoegd aan een standaardoplossing
bestaande uit acetonitrile 50%, water 47.5%, trifluoro-azijnzuur 2.5%, om een oplossing te verkrijgen
met een concentratie van 10 mg HCCA/ml. Om de HCCA matrix volledig opgelost te krijgen werd de
oplossing grondig gemixt. Aangezien een dergelijke matrix oplossing gevoelig is voor UV-straling
werd het donker bewaard, namelijk verpakt in aluminiumfolie. Het werd op een warmteplaat (40°C)
bewaard.
Het genereren van een massa spectrum en het analyseren van de resultaten
Massa spectra werden gegenereerd in de lineaire positieve mode met een bereik van 2-20 kDa (kilo Dalton) met een Microflex BiotyperTM van de Duitse firma Brüker Daltonics. De standaardinstellingen van de fabrikant werden gebruikt. Elke serie metingen werd voorafgegaan door een kalibratie met een bacteriële standaardtest van de firma Brüker Daltonics (test: BTS 155

19
255343). Identificaties werden verkregen door massa spectra te vergelijken met de Brüker MSP databases (versie DB 7311), waarbij gebruik werd gemaakt van de Brüker Compass software (Brüker Daltonics), met gebruik van de standaardinstellingen. In figuur 1 worden de procedurele stappen van een MALDI-TOF analyse schematisch weergegeven.
Figuur 1: Schematische weergave van de procedurele stappen van een MALDI-TOF analyse. A: een ‘MALDI target
plate’ (met een staal gemixt met matrix) en een ioniserende laser (rode straal). B: Desorptie (Het verschijnsel
waarbij een stof vrijkomt uit of via een oppervlak) en versnelling van eenwaardige ionen doorheen
hoogspanning. C: een geladen veld; scheiding van ionen op basis van hun massa/lading ratio (m/z ratio). D: TOF-
detector met spiegel; meting van de reistijd in de richting van de detector. E: een peptide massa vingerafdruk
(PMV) toont de intensiteit (procentueel weergegeven op de y-as) tegenover de m/z ratios (weergegeven op de
x-as).
Identificatie van micro-organismen door middel van MALDI-TOF wordt bewerkstelligd door het
vergelijken van de peptide massa vingerafdrukken (MS spectra) van niet-gekende micro-organismen
met peptide massa vingerafdrukken die zich in de database bevinden. De identiteit van micro-
organismen kan accuraat worden vastgesteld tot op genus niveau of zelfs tot op species niveau. Het
spectrum van de uitgevoerde analyse wordt vergeleken met referentie spectra (MSPs) uit een
commerciële database (Brüker Daltonics, Duitsland). Deze vergelijking of overeenkomst krijgt een
logaritmische score toegekend die de accuraatheid van de overeenkomst beschrijft.
3.3. Uitleg van de weergaven van de resultaten
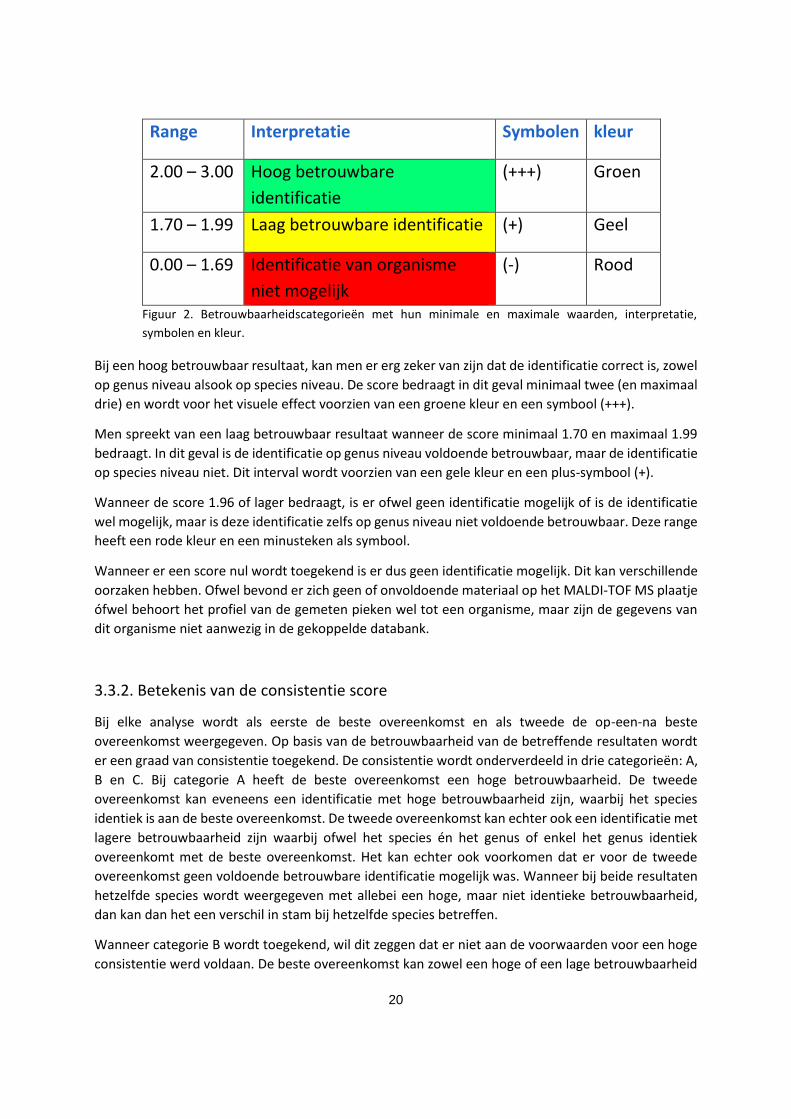
3.3.1. Betekenis van de betrouwbaarheidsscore
Aan elk resultaat dat het MALDI-TOF MS toestel genereert, verbindt het een mate van
betrouwbaarheid in de vorm van een score. Deze score heeft een bereik van nul tot drie. Er zijn drie
categorieën van betrouwbaarheid, die voor het visuele effect voorzien zijn van een kleur en een
symbool (voor mensen die kleurenblind zijn) zoals weergegeven in figuur 2. De hoogte van de score is
afhankelijk van het aantal overeenkomende pieken, gemeten op één plaatje en van de intensiteit van
de pieken en wordt via een logaritme berekend.
20
Range Interpretatie Symbolen kleur
2.00 – 3.00 Hoog betrouwbare
identificatie
(+++) Groen
1.70 – 1.99 Laag betrouwbare identificatie (+) Geel
0.00 – 1.69 Identificatie van organisme
niet mogelijk
(-) Rood
Figuur 2. Betrouwbaarheidscategorieën met hun minimale en maximale waarden, interpretatie,
symbolen en kleur.
Bij een hoog betrouwbaar resultaat, kan men er erg zeker van zijn dat de identificatie correct is, zowel
op genus niveau alsook op species niveau. De score bedraagt in dit geval minimaal twee (en maximaal
drie) en wordt voor het visuele effect voorzien van een groene kleur en een symbool (+++).
Men spreekt van een laag betrouwbaar resultaat wanneer de score minimaal 1.70 en maximaal 1.99
bedraagt. In dit geval is de identificatie op genus niveau voldoende betrouwbaar, maar de identificatie
op species niveau niet. Dit interval wordt voorzien van een gele kleur en een plus-symbool (+).
Wanneer de score 1.96 of lager bedraagt, is er ofwel geen identificatie mogelijk of is de identificatie
wel mogelijk, maar is deze identificatie zelfs op genus niveau niet voldoende betrouwbaar. Deze range
heeft een rode kleur en een minusteken als symbool.
Wanneer er een score nul wordt toegekend is er dus geen identificatie mogelijk. Dit kan verschillende
oorzaken hebben. Ofwel bevond er zich geen of onvoldoende materiaal op het MALDI-TOF MS plaatje
ófwel behoort het profiel van de gemeten pieken wel tot een organisme, maar zijn de gegevens van
dit organisme niet aanwezig in de gekoppelde databank.
3.3.2. Betekenis van de consistentie score
Bij elke analyse wordt als eerste de beste overeenkomst en als tweede de op-een-na beste
overeenkomst weergegeven. Op basis van de betrouwbaarheid van de betreffende resultaten wordt
er een graad van consistentie toegekend. De consistentie wordt onderverdeeld in drie categorieën: A,
B en C. Bij categorie A heeft de beste overeenkomst een hoge betrouwbaarheid. De tweede
overeenkomst kan eveneens een identificatie met hoge betrouwbaarheid zijn, waarbij het species
identiek is aan de beste overeenkomst. De tweede overeenkomst kan echter ook een identificatie met
lagere betrouwbaarheid zijn waarbij ofwel het species én het genus of enkel het genus identiek
overeenkomt met de beste overeenkomst. Het kan echter ook voorkomen dat er voor de tweede
overeenkomst geen voldoende betrouwbare identificatie mogelijk was. Wanneer bij beide resultaten
hetzelfde species wordt weergegeven met allebei een hoge, maar niet identieke betrouwbaarheid,
dan kan dan het een verschil in stam bij hetzelfde species betreffen.
Wanneer categorie B wordt toegekend, wil dit zeggen dat er niet aan de voorwaarden voor een hoge
consistentie werd voldaan. De beste overeenkomst kan zowel een hoge of een lage betrouwbaarheid
21
hebben. Het op-een-na beste resultaat kan net als bij categorie A een hoge of lage betrouwbaarheid
hebben of niet tot een voldoende betrouwbare identificatie hebben geleid.
Wanneer er geen sprake is van consistentie en er dus niet voldaan werd aan de voorwaarden voor
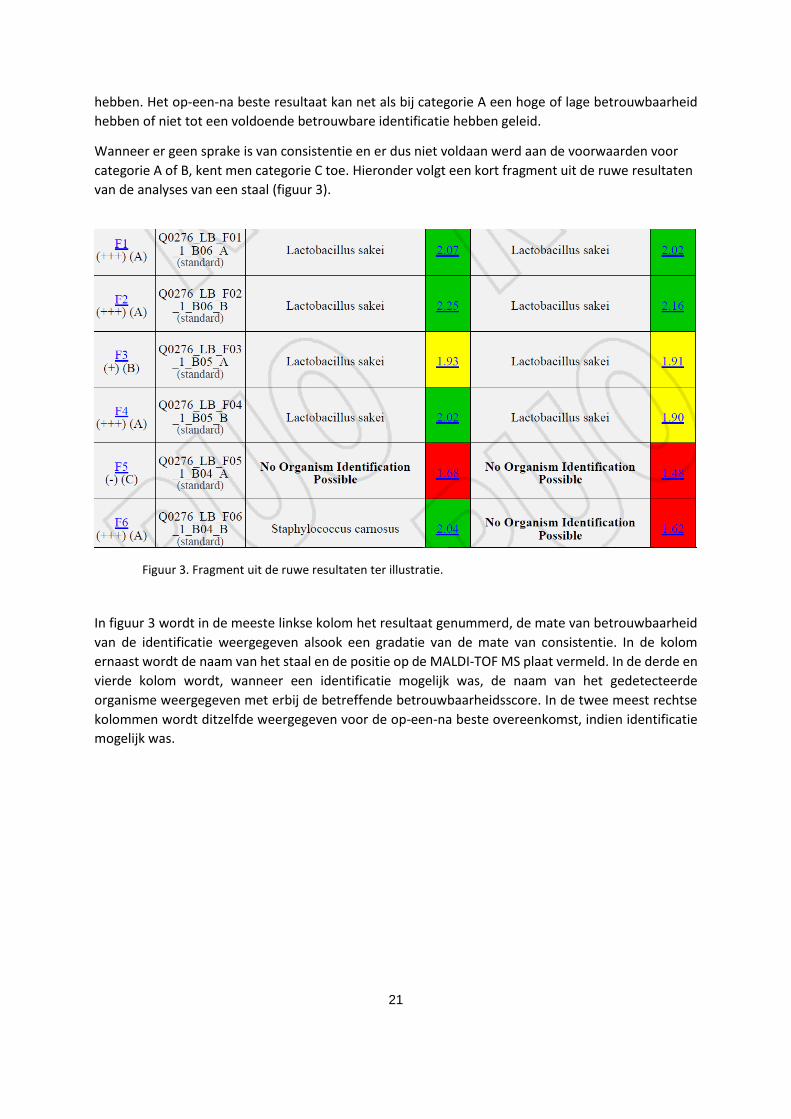
categorie A of B, kent men categorie C toe. Hieronder volgt een kort fragment uit de ruwe resultaten
van de analyses van een staal (figuur 3).
Figuur 3. Fragment uit de ruwe resultaten ter illustratie.
In figuur 3 wordt in de meeste linkse kolom het resultaat genummerd, de mate van betrouwbaarheid
van de identificatie weergegeven alsook een gradatie van de mate van consistentie. In de kolom
ernaast wordt de naam van het staal en de positie op de MALDI-TOF MS plaat vermeld. In de derde en
vierde kolom wordt, wanneer een identificatie mogelijk was, de naam van het gedetecteerde
organisme weergegeven met erbij de betreffende betrouwbaarheidsscore. In de twee meest rechtse
kolommen wordt ditzelfde weergegeven voor de op-een-na beste overeenkomst, indien identificatie
mogelijk was.
22
3.4. resultaten per honingsoort
3.4.1. Klimophoning
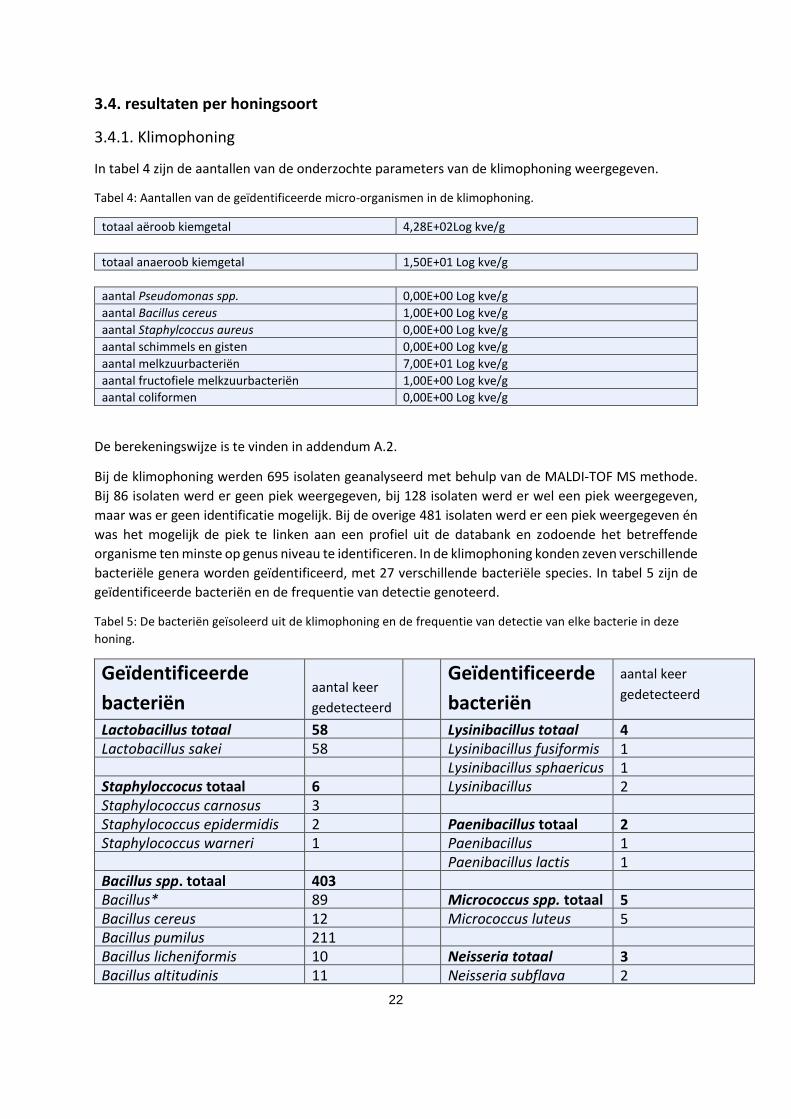
In tabel 4 zijn de aantallen van de onderzochte parameters van de klimophoning weergegeven.
Tabel 4: Aantallen van de geïdentificeerde micro-organismen in de klimophoning.
totaal aëroob kiemgetal 4,28E+02Log kve/g totaal anaeroob kiemgetal 1,50E+01 Log kve/g
aantal Pseudomonas spp. 0,00E+00 Log kve/g aantal Bacillus cereus 1,00E+00 Log kve/g aantal Staphylcoccus aureus 0,00E+00 Log kve/g aantal schimmels en gisten 0,00E+00 Log kve/g aantal melkzuurbacteriën 7,00E+01 Log kve/g aantal fructofiele melkzuurbacteriën 1,00E+00 Log kve/g aantal coliformen 0,00E+00 Log kve/g
De berekeningswijze is te vinden in addendum A.2.
Bij de klimophoning werden 695 isolaten geanalyseerd met behulp van de MALDI-TOF MS methode.
Bij 86 isolaten werd er geen piek weergegeven, bij 128 isolaten werd er wel een piek weergegeven,
maar was er geen identificatie mogelijk. Bij de overige 481 isolaten werd er een piek weergegeven én
was het mogelijk de piek te linken aan een profiel uit de databank en zodoende het betreffende
organisme ten minste op genus niveau te identificeren. In de klimophoning konden zeven verschillende
bacteriële genera worden geïdentificeerd, met 27 verschillende bacteriële species. In tabel 5 zijn de
geïdentificeerde bacteriën en de frequentie van detectie genoteerd.
Tabel 5: De bacteriën geïsoleerd uit de klimophoning en de frequentie van detectie van elke bacterie in deze
honing.
Geïdentificeerde
bacteriën aantal keer
gedetecteerd
Geïdentificeerde
bacteriën
aantal keer
gedetecteerd
Lactobacillus totaal 58 Lysinibacillus totaal 4 Lactobacillus sakei 58 Lysinibacillus fusiformis 1 Lysinibacillus sphaericus 1 Staphyloccocus totaal 6 Lysinibacillus
massiliensis
2 Staphylococcus carnosus 3 Staphylococcus epidermidis 2 Paenibacillus totaal 2 Staphylococcus warneri 1 Paenibacillus
illinoisensis
1 Paenibacillus lactis 1 Bacillus spp. totaal 403 Bacillus* 89 Micrococcus spp. totaal 5 Bacillus cereus 12 Micrococcus luteus 5 Bacillus pumilus 211 Bacillus licheniformis 10 Neisseria totaal 3 Bacillus altitudinis 11 Neisseria subflava 2
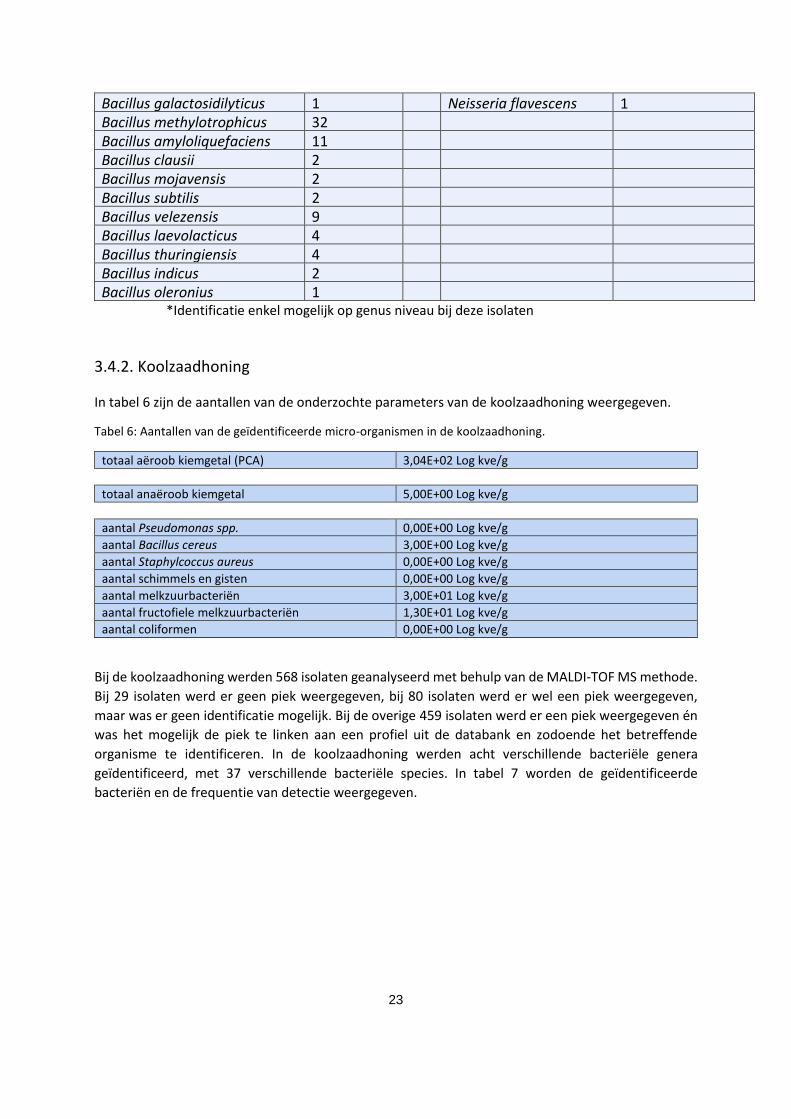
23
Bacillus galactosidilyticus 1 Neisseria flavescens 1 Bacillus methylotrophicus 32 Bacillus amyloliquefaciens 11 Bacillus clausii 2 Bacillus mojavensis 2 Bacillus subtilis 2 Bacillus velezensis 9 Bacillus laevolacticus 4 Bacillus thuringiensis 4 Bacillus indicus 2 Bacillus oleronius 1
*Identificatie enkel mogelijk op genus niveau bij deze isolaten
3.4.2. Koolzaadhoning
In tabel 6 zijn de aantallen van de onderzochte parameters van de koolzaadhoning weergegeven.
Tabel 6: Aantallen van de geïdentificeerde micro-organismen in de koolzaadhoning.
totaal aëroob kiemgetal (PCA) 3,04E+02 Log kve/g
totaal anaëroob kiemgetal 5,00E+00 Log kve/g
aantal Pseudomonas spp. 0,00E+00 Log kve/g
aantal Bacillus cereus 3,00E+00 Log kve/g
aantal Staphylcoccus aureus 0,00E+00 Log kve/g
aantal schimmels en gisten 0,00E+00 Log kve/g
aantal melkzuurbacteriën 3,00E+01 Log kve/g
aantal fructofiele melkzuurbacteriën 1,30E+01 Log kve/g
aantal coliformen 0,00E+00 Log kve/g
Bij de koolzaadhoning werden 568 isolaten geanalyseerd met behulp van de MALDI-TOF MS methode.
Bij 29 isolaten werd er geen piek weergegeven, bij 80 isolaten werd er wel een piek weergegeven,
maar was er geen identificatie mogelijk. Bij de overige 459 isolaten werd er een piek weergegeven én
was het mogelijk de piek te linken aan een profiel uit de databank en zodoende het betreffende
organisme te identificeren. In de koolzaadhoning werden acht verschillende bacteriële genera
geïdentificeerd, met 37 verschillende bacteriële species. In tabel 7 worden de geïdentificeerde
bacteriën en de frequentie van detectie weergegeven.
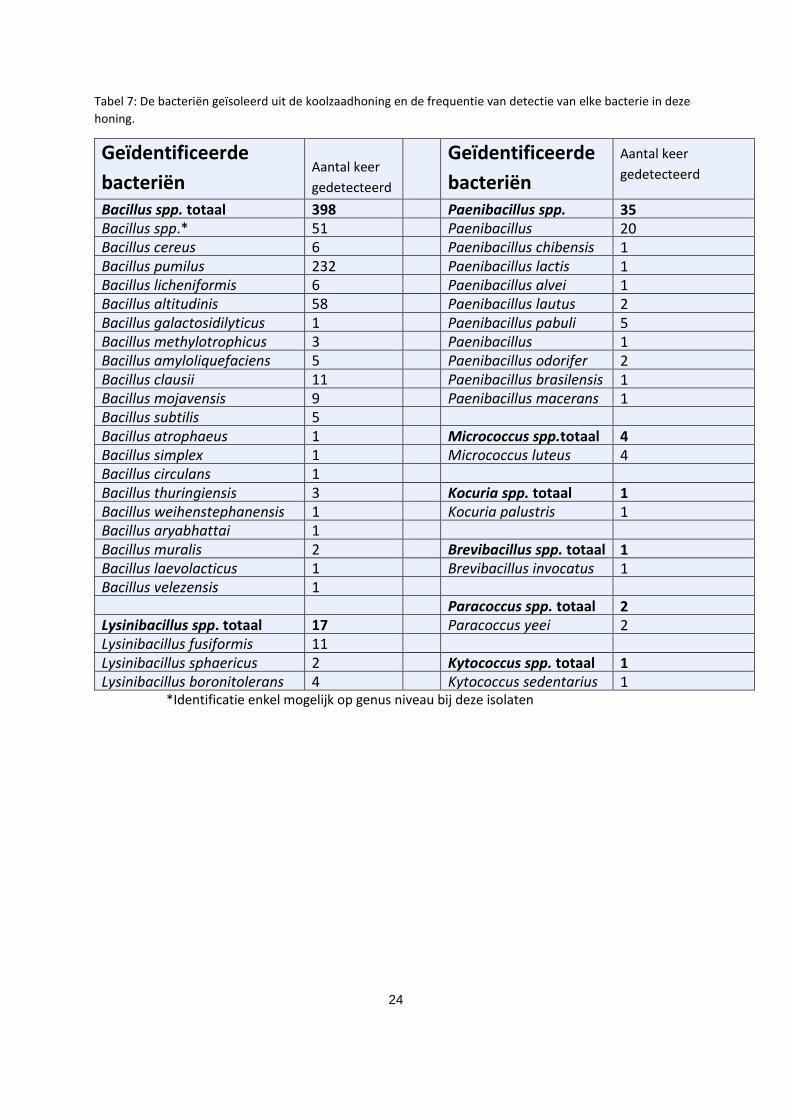
24
Tabel 7: De bacteriën geïsoleerd uit de koolzaadhoning en de frequentie van detectie van elke bacterie in deze
honing.
Geïdentificeerde
bacteriën Aantal keer
gedetecteerd
Geïdentificeerde
bacteriën
Aantal keer
gedetecteerd
Bacillus spp. totaal 398 Paenibacillus spp.
totaal
35 Bacillus spp.* 51 Paenibacillus
amylolyticus
20 Bacillus cereus 6 Paenibacillus chibensis 1 Bacillus pumilus 232 Paenibacillus lactis 1 Bacillus licheniformis 6 Paenibacillus alvei 1 Bacillus altitudinis 58 Paenibacillus lautus 2 Bacillus galactosidilyticus 1 Paenibacillus pabuli 5 Bacillus methylotrophicus 3 Paenibacillus
illinoisensis
1 Bacillus amyloliquefaciens 5 Paenibacillus odorifer 2 Bacillus clausii 11 Paenibacillus brasilensis 1 Bacillus mojavensis 9 Paenibacillus macerans 1 Bacillus subtilis 5 Bacillus atrophaeus 1 Micrococcus spp.totaal 4 Bacillus simplex 1 Micrococcus luteus 4 Bacillus circulans 1 Bacillus thuringiensis 3 Kocuria spp. totaal 1 Bacillus weihenstephanensis 1 Kocuria palustris 1 Bacillus aryabhattai 1 Bacillus muralis 2 Brevibacillus spp. totaal 1 Bacillus laevolacticus 1 Brevibacillus invocatus 1 Bacillus velezensis 1 Paracoccus spp. totaal 2 Lysinibacillus spp. totaal 17 Paracoccus yeei 2 Lysinibacillus fusiformis 11 Lysinibacillus sphaericus 2 Kytococcus spp. totaal 1 Lysinibacillus boronitolerans 4 Kytococcus sedentarius 1
*Identificatie enkel mogelijk op genus niveau bij deze isolaten
25
3.4.3. Paardenbloemhoning
In tabel 8 zijn de aantallen van de onderzochte parameters van de paardenbloemhoning weergegeven.
Tabel 8: Aantallen van de geïdentificeerde micro-organismen in de paardenbloemhoning.
totaal aëroob kiemgetal (PCA) 2,04E+02 Log kve/g
totaal anaëroob kiemgetal 3,67E+03 Log kve/g
aantal Pseudomonas spp. 0,00E+00 Log kve/g
aantal Bacillus cereus 5,00E+00 Log kve/g
aantal Staphylcoccus aureus 0,00E+00 Log kve/g
aantal schimmels en gisten 0,00E+00 Log kve/g
aantal melkzuurbacteriën 2,56E+02 Log kve/g
aantal fructofiele melkzuurbacteriën 8,00E+00 Log kve/g
aantal coliformen 0,00E+00 Log kve/g
Bij de paardenbloemhoning werden 639 isolaten geanalyseerd met behulp van de MALDI-TOF MS
methode. De berekeningswijze is te vinden in addendum A.2.
Bij 144 isolaten werd er geen piek weergegeven, bij 176 isolaten werd er wel een piek weergegeven,
maar was er geen identificatie mogelijk. Bij de overige 319 isolaten werd er een piek weergegeven én
was het mogelijk de piek te linken aan een profiel uit de databank en zodoende het betreffende
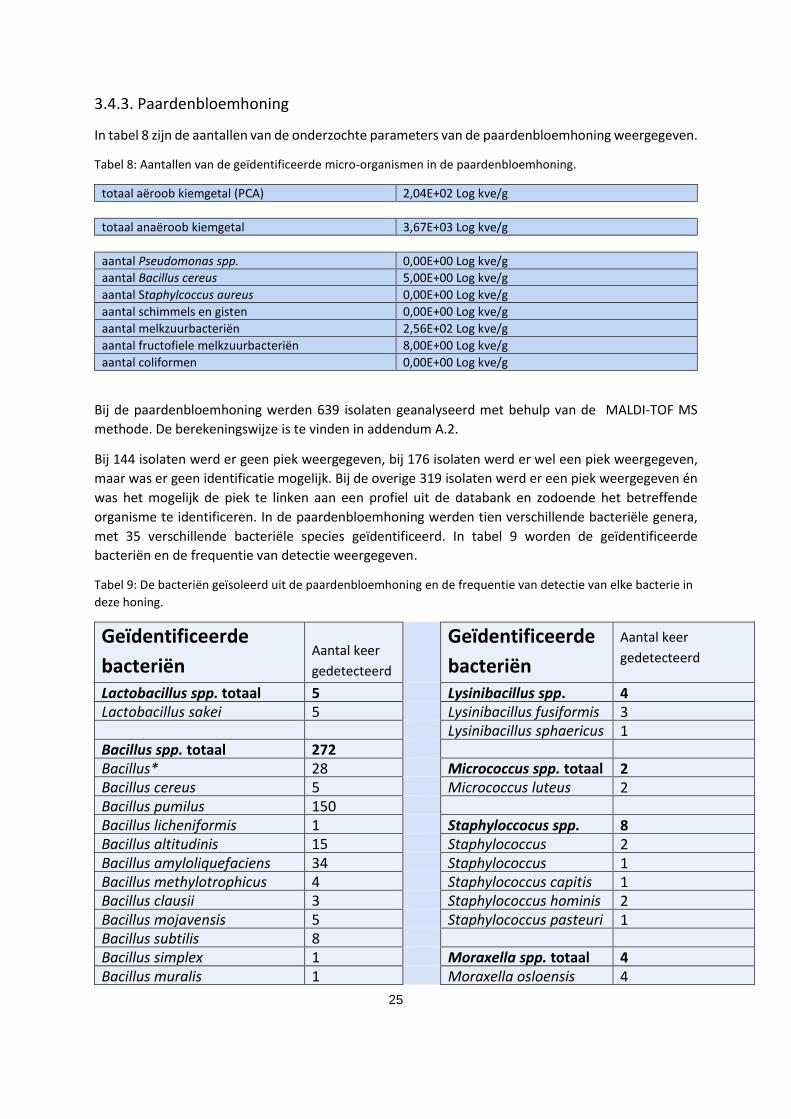
organisme te identificeren. In de paardenbloemhoning werden tien verschillende bacteriële genera,
met 35 verschillende bacteriële species geïdentificeerd. In tabel 9 worden de geïdentificeerde
bacteriën en de frequentie van detectie weergegeven.
Tabel 9: De bacteriën geïsoleerd uit de paardenbloemhoning en de frequentie van detectie van elke bacterie in
deze honing.
Geïdentificeerde
bacteriën Aantal keer
gedetecteerd
Geïdentificeerde
bacteriën
Aantal keer
gedetecteerd
Lactobacillus spp. totaal 5 Lysinibacillus spp.
totaal
4 Lactobacillus sakei 5 Lysinibacillus fusiformis 3 Lysinibacillus sphaericus 1 Bacillus spp. totaal 272 Bacillus* 28 Micrococcus spp. totaal 2 Bacillus cereus 5 Micrococcus luteus 2 Bacillus pumilus 150 Bacillus licheniformis 1 Staphyloccocus spp.
totaal
8 Bacillus altitudinis 15 Staphylococcus
epidermidis
2 Bacillus amyloliquefaciens 34 Staphylococcus
saprophyticus
1 Bacillus methylotrophicus 4 Staphylococcus capitis 1 Bacillus clausii 3 Staphylococcus hominis 2 Bacillus mojavensis 5 Staphylococcus pasteuri 1 Bacillus subtilis 8 Bacillus simplex 1 Moraxella spp. totaal 4 Bacillus muralis 1 Moraxella osloensis 4
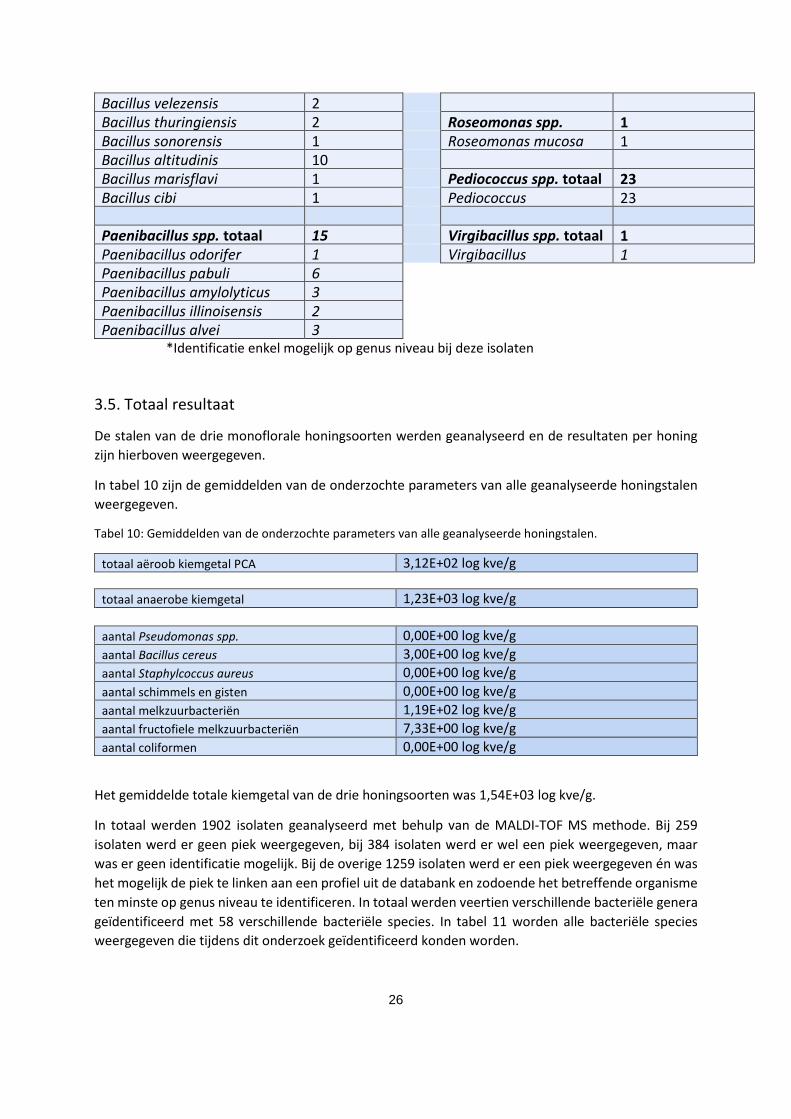
26
Bacillus velezensis 2 Bacillus thuringiensis 2 Roseomonas spp.
totaal
1 Bacillus sonorensis 1 Roseomonas mucosa 1 Bacillus altitudinis 10 Bacillus marisflavi 1 Pediococcus spp. totaal 23 Bacillus cibi 1 Pediococcus
pentosaceus
23 Paenibacillus spp. totaal 15 Virgibacillus spp. totaal 1 Paenibacillus odorifer 1 Virgibacillus
pantothenticus
1 Paenibacillus pabuli 6 Paenibacillus amylolyticus 3 Paenibacillus illinoisensis 2 Paenibacillus alvei 3
*Identificatie enkel mogelijk op genus niveau bij deze isolaten
3.5. Totaal resultaat
De stalen van de drie monoflorale honingsoorten werden geanalyseerd en de resultaten per honing
zijn hierboven weergegeven.
In tabel 10 zijn de gemiddelden van de onderzochte parameters van alle geanalyseerde honingstalen
weergegeven.
Tabel 10: Gemiddelden van de onderzochte parameters van alle geanalyseerde honingstalen.
totaal aëroob kiemgetal PCA 3,12E+02 log kve/g
totaal anaerobe kiemgetal 1,23E+03 log kve/g
aantal Pseudomonas spp. 0,00E+00 log kve/g
aantal Bacillus cereus 3,00E+00 log kve/g
aantal Staphylcoccus aureus 0,00E+00 log kve/g
aantal schimmels en gisten 0,00E+00 log kve/g
aantal melkzuurbacteriën 1,19E+02 log kve/g
aantal fructofiele melkzuurbacteriën 7,33E+00 log kve/g
aantal coliformen 0,00E+00 log kve/g
Het gemiddelde totale kiemgetal van de drie honingsoorten was 1,54E+03 log kve/g.
In totaal werden 1902 isolaten geanalyseerd met behulp van de MALDI-TOF MS methode. Bij 259
isolaten werd er geen piek weergegeven, bij 384 isolaten werd er wel een piek weergegeven, maar
was er geen identificatie mogelijk. Bij de overige 1259 isolaten werd er een piek weergegeven én was
het mogelijk de piek te linken aan een profiel uit de databank en zodoende het betreffende organisme
ten minste op genus niveau te identificeren. In totaal werden veertien verschillende bacteriële genera
geïdentificeerd met 58 verschillende bacteriële species. In tabel 11 worden alle bacteriële species
weergegeven die tijdens dit onderzoek geïdentificeerd konden worden.
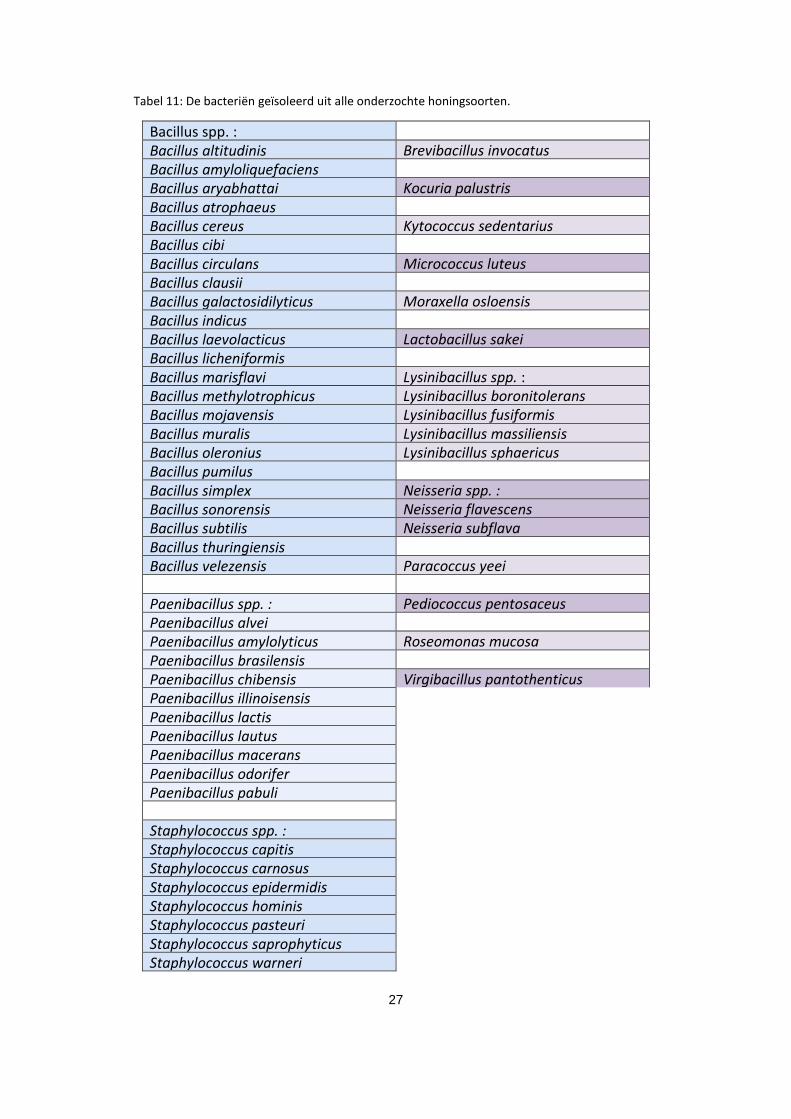
27
Tabel 11: De bacteriën geïsoleerd uit alle onderzochte honingsoorten.
Bacillus spp. : Bacillus altitudinis Brevibacillus invocatus Bacillus amyloliquefaciens Bacillus aryabhattai Kocuria palustris Bacillus atrophaeus Bacillus cereus Kytococcus sedentarius Bacillus cibi Bacillus circulans Micrococcus luteus Bacillus clausii Bacillus galactosidilyticus Moraxella osloensis Bacillus indicus Bacillus laevolacticus Lactobacillus sakei Bacillus licheniformis Bacillus marisflavi Lysinibacillus spp. : Bacillus methylotrophicus Lysinibacillus boronitolerans Bacillus mojavensis Lysinibacillus fusiformis Bacillus muralis Lysinibacillus massiliensis Bacillus oleronius Lysinibacillus sphaericus Bacillus pumilus Bacillus simplex Neisseria spp. : Bacillus sonorensis Neisseria flavescens Bacillus subtilis Neisseria subflava Bacillus thuringiensis Bacillus velezensis Paracoccus yeei Paenibacillus spp. : Pediococcus pentosaceus Paenibacillus alvei
Paenibacillus amylolyticus Roseomonas mucosa Paenibacillus brasilensis Paenibacillus chibensis Virgibacillus pantothenticus Paenibacillus illinoisensis Paenibacillus lactis Paenibacillus lautus
Paenibacillus macerans Paenibacillus odorifer Paenibacillus pabuli
Staphylococcus spp. : Staphylococcus capitis Staphylococcus carnosus Staphylococcus epidermidis Staphylococcus hominis Staphylococcus pasteuri Staphylococcus saprophyticus Staphylococcus warneri

28
4. Discussie
Het gemiddelde totale kiemgetal van de drie honingsoorten was 1,54E+03 log kve/g. Het laagste totale
kiemgetal had de koolzaadhoning (3,09E+02 log kve/g), het kiemgetal van de klimophoning was hoger
(4,43E+02 log kve/g) en de paardenbloemhoning had het grootste kiemgetal (3,88E+03 log kve/g). Het
feit dat er bacteriën geïsoleerd konden worden en dan zeker in deze hoeveelheden, is opvallend.
Honing heeft immers diverse antibacteriële eigenschappen. Zoals eerder in dit verslag vermeld heeft
honing onder andere een relatief lage pH, een lage wateractiviteit, een hoge osmolariteit en bevat het
antibacteriële substanties zoals pinocembrine, lysozyme, zuren en een enzym dat glucose oxideert tot
waterstofperoxide (Molan, 1992; Tysset and de la Roy, 1974).
Er werden in totaal veertien verschillende bacteriële genera geïdentificeerd met 58 verschillende
bacteriële species. Zoals hierboven vermeld heeft honing een niet gering aantal antibacteriële
eigenschappen, die door middel van verschillende mechanismen de groei maar ook de overleving van
bacteriën belemmeren en vaak zelfs onmogelijk maken. Daarom is niet alleen het kiemgetal verassend,
maar ook de diversiteit van het bacteriële microbioom van elk van deze honingsoorten. Het aantal
geïdentificeerde bacteriën per parameter komt overigens in relatief grote mate overeen tussen de
verschillende honingsoorten.
In dit onderzoek werd het bacterieel microbioom van klimophoning, koolzaadhoning en
paardenbloemhoning onderzocht. Er was tot nu toe niets beschreven in de wetenschappelijke
literatuur, over het bacteriële diversiteit van deze drie soorten monoflorale honingen. De identificatie
van de aanwezige bacteriële species vond plaats door middel van MALDI-TOF MS. Dit is een relatief
nieuwe methode waarmee de identificatie van bacteriën op grote schaal en in relatieve korte tijd kan
plaatsvinden. Hoewel kiemen van diverse voedingsmiddelen hiermee reeds succesvol werden
geïdentificeerd, werd dit tot op heden niet eerder gebruikt voor honing. In dit opzicht is het gebruik
van MALDI-TOF MS in dit onderzoek een primeur. De bacteriën die in de honingstalen werden
aangetroffen, kunnen in verschillende categorieën worden verdeeld:
- Commensale bacteriën: deze bacteriën komen voor in honing en leveren geen voordeel of nadeel
op voor de organismen die zich voeden met de honing, namelijk de mens en de honingbij zelf. In
tabel 12 zijn de commensale bacteriën opgelijst die in dit onderzoek geïdentificeerd werden
(Heyrman, 2015; Humphreys, 2012; Reddy et al., 2003; Snowdon and Cliver, 1995).
Tabel 12: Commensale bacteriën geïdentificeerd tijdens dit onderzoek.
Bacillus spp.: Staphylococcus spp. : Neisseria spp. :
Bacillus cereus Staphylococcus capitis Neisseria flavescens
Bacillus cibi Staphylococcus carnosus Neisseria subflava
Bacillus circulans Staphylococcus epidermidis
Bacillus clausii Staphylococcus hominis Micrococcus luteus
Bacillus galactosidilyticus Staphylococcus pasteuri Brevibacillus invocatus
Bacillus indicus Staphylococcus saprophyticus Kocuria palustris
Bacillus laevolacticus Staphylococcus warneri
Bacillus licheniformis
Bacillus marisflavi
Bacillus methylotrophicus
Bacillus mojavensis

29
Bacillus sonorensis
Bacillus subtilis
Bacillus thuringiensis
Bacillus velezensis
- Voor de mens pathogene bacteriën:
Moraxella osloensis is een Gram negatieve bacterie die uit de paardenbloemhoning werd geïsoleerd.
Deze bacterie valt enigszins tussen wal en schip bij het categoriseren. Deze kiem leeft in symbiose met
Phasmarhabditis hermaphrodita, een endoparasiet bij slakken. Infecties bij de mens zijn beschreven,
maar zijn zeldzaam. Dat dat deze bacterie wel in paardenbloemhoning werd gevonden en niet in de
klimophoning en koolzaadhoning, komt wellicht doordat de bloemen van de paardenbloem zich
dichter bij de grond bevinden dan deze van klimop en koolzaad. Hierdoor is de kans op contaminatie
door contact met slakken groter (Hleba, 2014; Tan and Grewal, 2001)
Paenibacillus spp. werden uit elke geanalyseerde honingsoort geïsoleerd. In tegenstelling tot
Paenibacillus larvae, de veroorzaker van Amerikaans Vuilbroed, zijn de hier geïdentificeerde species
niet pathogeen voor honingbijen. Ze kunnen echter als opportunisten wel pathogeen zijn bij de mens,
al is de kans op ziekte zeer klein (Grady et al., 2016)
Paracoccus yeei werd geïsoleerd uit de paardenbloemhoning. Het is een voor de mens pathogene
bacterie. Infecties met Paracoccus yeei worden zelden beschreven. De kiem heeft een zeer laag
pathologisch potentieel. Aarde wordt gezien als bron van contaminatie (Funke et al., 2004).
Roseomonas mucosa wordt aanzien als een voor de mens commensale bacterie, die bij mensen op de
huid leeft, maar steeds vaker als opportunist aanleiding geeft tot infecties. Deze bacterie werd enkel
aangetoond in de paardenbloemhoning. Het is aannemelijk dat tijdens de oogst of verwerking van de
honing, contaminatie via de huid van de mens naar de honing plaats heeft gevonden (Romano-
Bertrand et al., 2016)
- Voor de honingbij pathogene bacteriën:
Lysinibacillus spp.: Er werden Lysinibacillus spp. geïdentificeerd in alle onderzochte honingsoorten.
Recent werd ontdekt dat Lysinibacillus sphaericus pathogeen is voor bijen. De overige geïsoleerde
species zijn wellicht eveneens pathogeen voor bijen, meer onderzoek daarover is nodig (Fünfhaus et
al., 2018).
- Bacterie met activiteit tegen schimmels:
Pediococcus pentosaceus werd enkel aangetroffen in de paardenbloemhoning. Deze bacterie zet
suikers om in melkzuur en verlaagt zodoende de pH van de honing, hetgeen de groei van andere
microorganismen en in het bijzonder van schimmels in honing tegengaat (Bulgasem et al., 2016)

30
Conclusies ten aanzien van de voedselveiligheid
In tegenstelling tot de verwachting gebaseerd op internationale literatuur en microbiologische
veiligheidsordes als pH en Aw, bleken de drie stalen toch bacterieel besmet te zijn. Bijzonder is dat het
hier geen analyse onmiddellijk na het oogsten betrof, maar gecommercialiseerde honing die reeds
gestabiliseerd was in pH en Aw waarde. Het besmettingsniveau was weliswaar laag, maar een grote
diversiteit van genera en species werd geïdentificeerd. Er werd een beperkt aantal bacteriële species
aangetroffen, geassocieerd met ziekte bij de mens. Het betrof in alle gevallen bacteriën met een sterk
beperkt pathogeen vermogen die vooral als opportunist aanleiding kunnen geven tot infectie. Behalve
de Paenibacillus spp. werden de bacteriële species die pathogeen kunnen zijn voor de mens enkel
aangetroffen in de paardenbloemhoning. Hetgeen zou kunnen duiden op secundaire contaminatie.
Om de kans op ziekte bij de mens door consumptie van honing zoveel mogelijk te beperken, is het aan
te bevelen om de hygiënemaatregelen omtrent de oogst en verwerking van honing strikt na te leven.
Daarnaast is het wellicht verstandig om consumptie van honing te onraden aan mensen met een
vergrote kans op infectie door opportunistische bacteriën.

31
Literatuurlijst
Alippi, A.M., Reynaldi, F.J., Lopez, A.C., De Giusti, M.R., Aguilar, O.M., 2004. Molecular epidemiology of Paenibacillus larvae larvae and incidence of American foulbrood in Argentinean honeys from Buenos Aires province. J. Apic. Res. 43, 135-143. Allen, K.L., Molan, P.C., Reid, G.M., 1991. A survey of the antibacterial activity of some New Zealand honeys. The Journal of pharmacy and pharmacology 43, 817-822. Allsopp, M.H., de Lange, W.J., Veldtman, R., 2008. Valuing Insect Pollination Services with Cost of Replacement. Plos One 3, 8. Amna Jabbar Siddiqui, S.G.M., M. Iqbal Choudhary, Atta-ur- Rahman, 2016. Application of analytical methods in authentication and adulteration
of honey. Food Chem. 217. anoniem, 2011a. Bijenproducten. http://www.imkerpedia.nl/wiki/index.php?title=Bijenproducten (accessed 03-04-2017 2017). Anoniem, 2011b. Voedsterbij. http://www.imkerpedia.nl/wiki/index.php?title=Voedsterbij (accessed 04-04-2011 2017). Bulgasem, B.Y., Lani, M.N., Hassan, Z., Yusoff, W.M.W., Fnaish, S.G., 2016. Antifungal Activity of Lactic Acid Bacteria Strains Isolated from Natural Honey against Pathogenic Candida Species. Mycobiology 44, 302-309. de Mendonca, I.C.G., Porto, I., do Nascimento, T.G., de Souza, N.S., Oliveira, J.M.D., Arruda, R.E.D., Mousinho, K.C., dos Santos, A.F., Basilio, I.D., Parolia, A., Barreto, F.S., 2015. Brazilian red propolis: phytochemical screening, antioxidant activity and effect against cancer cells. Bmc Complementary and Alternative Medicine 15, 12. Estevinho, L.M., Feas, X., Seijas, J.A., Vazquez-Tato, M.P., 2012. Organic honey from Tras-Os-Montes region (Portugal): Chemical, palynological, microbiological and bioactive compounds characterization. Food Chem. Toxicol. 50, 258-264. FAVV, 2017. Aangifteplichtige ziekten en schadelijke organismen (KB 3 februari 2014) http://www.afsca.be/bijenteelt/dierengezondheid/#aangifteplichtige (accessed 03-04-2017 2017). FISHER, SCIENTIFIC, 1997. Material Safety Data Sheet Beeswax. https://fscimage.fishersci.com/msds/02556.htm (accessed 14-05-2018 2018). Fratellone, P.M., Fratellone, G., 2016. Apitherapy Products for Medicinal Use. Journal of Alternative and Complementary Medicine 22, 1020-1022. Fünfhaus, A., Ebeling, J., Genersch, E., 2018. Bacterial pathogens of bees. Current Opinion in Insect Science 26, 89-96. Funke, G., Frodl, R., Sommer, H., 2004. First Comprehensively Documented Case of Paracoccus yeei Infection in a Human. Journal of Clinical Microbiology 42, 3366-3368. Gilliam, M., 1971. MICROBIAL STERILITY OF INTESTINAL CONTENT OF IMMATURE HONEY BEE, APIS-MELLIFERA HYMENOPTERA-APIDAE. Ann. Entomol. Soc. Am. 64, 315-&. Gilliam, M., 1978. BACTERIA BELONGING TO GENUS BACILLUS ISOLATED FROM SELECTED ORGANS OF QUEEN HONEY BEES, APIS-MELLIFERA. J. Invertebr. Pathol. 31, 389-391. Gilliam, M., Lorenz, B.J., Richardson, G.V., 1988. DIGESTIVE ENZYMES AND MICROORGANISMS IN HONEY BEES, APIS-MELLIFERA - INFLUENCE OF STREPTOMYCIN, AGE, SEASON AND POLLEN. Microbios 55, 95-114. Gilliam, M., Moffett, J.O., Kauffeld, N.M., 1983. EXAMINATION OF FLORAL NECTAR OF CITRUS, COTTON, AND ARIZONA DESERT PLANTS FOR MICROBES. Apidologie 14, 299-302. Grady, E.N., MacDonald, J., Liu, L., Richman, A., Yuan, Z.-C., 2016. Current knowledge and perspectives of Paenibacillus: a review. Microbial cell factories 15, 203. Graham, J., 1992. THE 1992 EDITION OF THE HIVE AND THE HONEY-BEE .3. Am. Bee J. 132, 721-725. Heyrman, J.V., P. D.; Logan, N., 2015. Virgibacillus, In: Bergey's Manual of Systematics of Archaea and Bacteria.

32
Hleba, L., 2014. Antibiotic resistance of microbial contaminations isolated from husbandry animals and foodstuffs. Scientific Papers Animal Science and Biotechnologies 47, 104-108. Humphreys, H., 2012. 15 - Staphylococcus: Skin infections; osteomyelitis; bloodstream infection; food poisoning; foreign body infections; MRSA A2 - Greenwood, David, In: Medical Microbiology (Eighteenth Edition). Churchill Livingstone, Edinburgh, pp. 176-182. Iurlina, F., 2005. Characterization of microorganisms in Argentinean honeys from
different sources. International Journal of Food Microbiology, 297-304. Jay, J.M., 1992. Microbiological food safety. Critical Reviews in Food Science and Nutrition 31, 177-190. Molan, P.C., 1992. The antibacterial activity of honey: 1. The nature of the antibacterial activity. Bee world 73, 5-28. Molan, P.C., 1999. Why honey is effective as a medicine. I. Its use in modern medicine. Bee World 80, 80-92. Nakano, H., Yoshikuni, Y., Hashimoto, H., Sakaguchi, G., 1992. Detection of Clostridium botulinum in natural sweetening. International Journal of Food Microbiology 16, 117-121. Olofsson, T.C., Vasquez, A., 2008. Detection and identification of a novel lactic acid bacterial flora within the honey stomach of the honeybee Apis mellifera. Curr. Microbiol. 57, 356-363. Ramadan, M.F., Al-Ghamdi, A., 2012. Bioactive compounds and health-promoting properties of royal jelly: A review. Journal of Functional Foods 4, 39-52. Reddy, G.S.N., Prakash, J.S.S., Prabahar, V., Matsumoto, G.I., Stackebrandt, E., Shivaji, S., 2003. Kocuria polaris sp. nov., an orange-pigmented psychrophilic bacterium isolated from an Antarctic cyanobacterial mat sample. Int. J. Syst. Evol. Microbiol. 53, 183-187. Romano-Bertrand, S., Bourdier, A., Aujoulat, F., Michon, A.L., Masnou, A., Parer, S., Marchandin, H., Jumas-Bilak, E., 2016. Skin microbiota is the main reservoir of <em>Roseomonas mucosa</em>, an emerging opportunistic pathogen so far assumed to be environmental. Clinical Microbiology and Infection 22, 737.e731-737.e737. Sacket, W.G., 1919. HONEY AS A CARRIER OF INTESTINAL DISEASES. The Agricultural Experiment Station
OF THE
Colorado Agricultural College 252, 18. Sereia, M.R.F.G.P., Paulo Henrique Março, R.S.P., Erica Gomes de, Anjo, L.a.F.A., 2017. Techniques for the Evaluation of Microbiological Quality in Honey, In: Techniques for the Evaluation of Microbiological Quality in Honey. Smith, M.R., McCaughey, W.F., Kemmerer, A.R., 1969. Biological Effects of Honey. J. Apic. Res. 8, 99-110. Snowdon, Cliver, 1995. Microorganisms in honey. International Journal of
Food Microbiology, 1-26. Snowdon, J.A., 1999. The microbiology of honey - Meeting your buyers' specifications (Why they do what they do). Am. Bee J. 139, 51-60. Tan, L., Grewal, P.S., 2001. Pathogenicity of Moraxella osloensis, a bacterium associated with the nematode Phasmarhabditis hermaphrodita, to the slug Deroceras reticulatum. Applied and environmental microbiology 67, 5010-5016. Troller, J.A., 1979. FOOD SPOILAGE BY MICROORGANISMS TOLERATING LOW-AW ENVIRONMENTS. Food Technol. 33, 72-75. Tysset, C., de la Roy, Y.d.R., 1974. Assays on the study of ‘osmophilic’yeasts, organisms causing fermentations of honey collected in France. Association diplomacy microbiology faculty pharmacy university, Nancy Bull 134, 1-26. Tysset, C., Durand, C., 1976. On the survival of some enterobacteria in honey stored in a refrigerated place at 10 degree C. Bulletin de l'Academie Veterinaire de France (France).

33
Tysset, C., Durand, C., Taliergio, Y., 1970. Contribution to the study of the microbial contamination and the hygiene of commercial honey. Review Medical Veterinary 146, 1471-1492. Tysset, C., Rousseau, M., 1981. Problem of microbes and hygiene of commercial honey. Rev. Med. Vet 132, 591-600. White, J.j., Riethof, M., Subers, M., Kushnir, I., 1962b. Composition of American honeys. Tech. Bull. White, J.W., Schepartz, A.I., Subers, M.H., 1963. IDENTIFICATION OF INHIBINE, ANTIBACTERIAL FACTOR IN HONEY, AS HYDROGEN PEROXIDE AND ITS ORIGIN IN A HONEY GLUCOSE-OXIDASE SYSTEM. Biochimica Et Biophysica Acta 73, 57-&. Willix, D.J., Molan, P.C., Harfoot, C.G., 1992. A comparison of the sensitivity of wound-infecting species of bacteria to the antibacterial activity of manuka honey and other honey. The Journal of applied bacteriology 73, 388-394.

34
Addendum
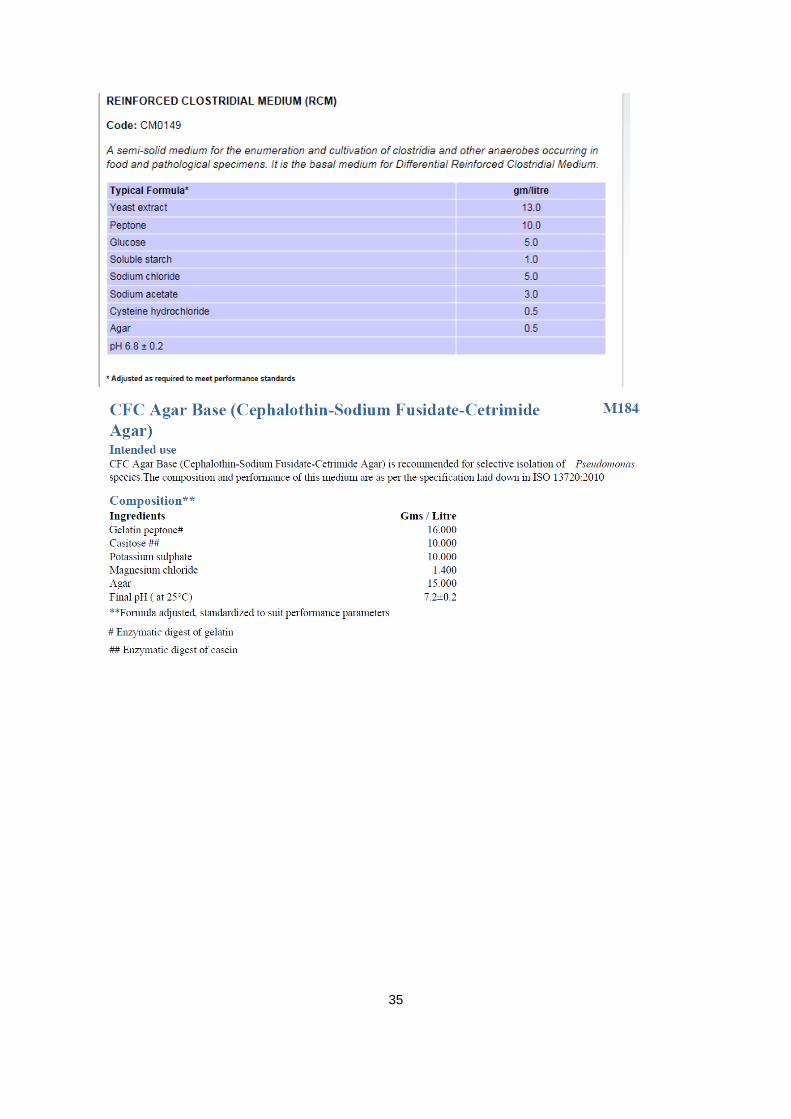
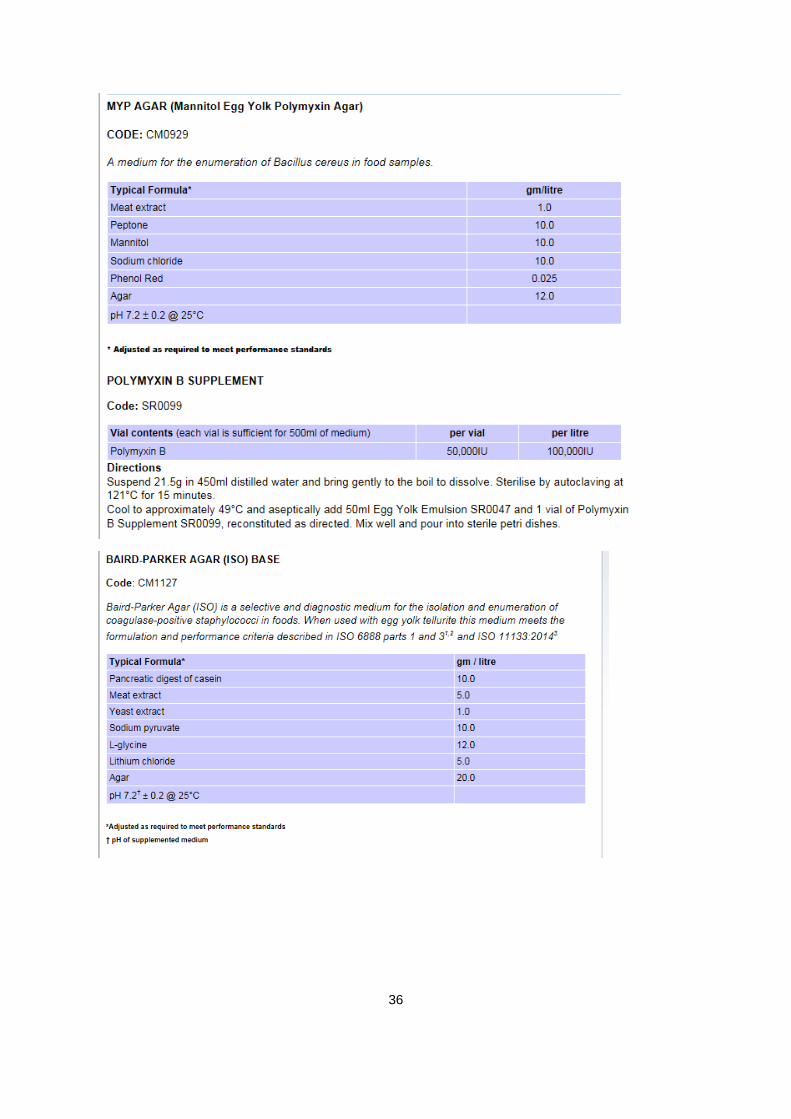
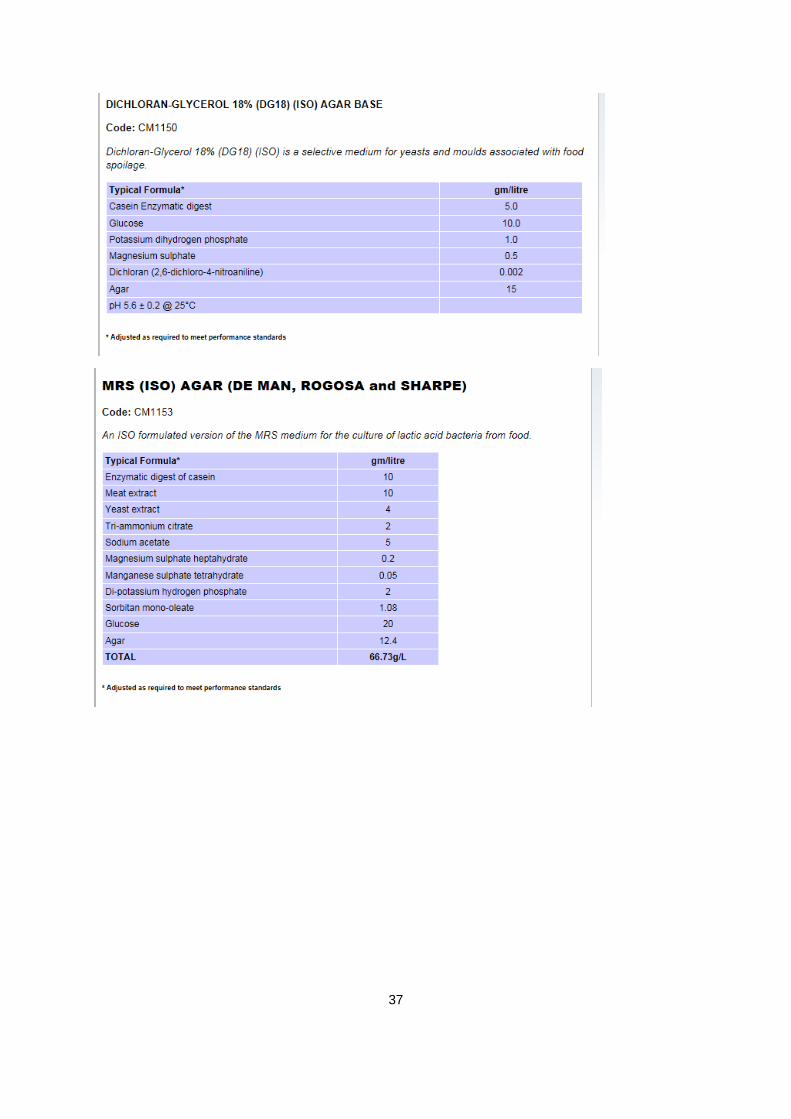
A.1. informatie over agars:
Verantwoordelijke producent van alle gebruikte agars: Oxoid limited
35
36
37

38
Addendum A.2. Berekeningswijze kve/g


















