In vitro vermeerdering van Ulva lactuca - lib.ugent.be · Abstract Voedsel en energie zullen...
77
Faculteit Bio-ingenieurswetenschappen Academiejaar 2013 – 2014 In vitro vermeerdering van Ulva lactuca. Klaartje Bunkens Promotor: Prof. dr. ir. Stefaan Werbrouck Tutor: dr. Julia Wald Masterproef voorgedragen tot het behalen van de graad van Master of Science in de biowetenschappen: land- en tuinbouwkunde
Transcript of In vitro vermeerdering van Ulva lactuca - lib.ugent.be · Abstract Voedsel en energie zullen...

Faculteit Bio-ingenieurswetenschappen
Academiejaar 2013 – 2014
In vitro vermeerdering van Ulva lactuca.
Klaartje Bunkens
Promotor: Prof. dr. ir. Stefaan Werbrouck
Tutor: dr. Julia Wald
Masterproef voorgedragen tot het behalen van de graad van
Master of Science in de biowetenschappen: land- en tuinbouwkunde

De auteur en de promotor geven de toelating deze scriptie voor consultatie beschikbaar te
stellen en delen ervan te kopiëren voor persoonlijk gebruik. Elk ander gebruik valt onder de
beperkingen van het auteursrecht, in het bijzonder met betrekking tot de verplichting uitdruk-
kelijk de bron te vermelden bij het aanhalen van resultaten uit deze scriptie.
Promotor:
Prof. dr. ir. Stefaan Werbrouck
De auteur:
Klaartje Bunkens

Voorwoord
Onze samenleving ziet zich geplaatst voor grote ecologische uitdagingen. Een groeiende we-
reldbevolking enerzijds en een zich doorzettende klimaatsverandering anderzijds dwingen ons
tot een dringende herziening van onze conventionele voedselproductie. Zeewierteelt kan een
bescheiden bijdrage leveren om ons huidige landbouwareaal uit te breiden. Bovendien kunnen
wieren ook ingezet worden als biobrandstof en als waterfilter. Wieren verbruiken immers uit-
gespoelde en/of overtollige nutriënten en kunnen zout water converteren naar zoet water. Al-
lemaal redenen om meer kennis te verzamelen over wieren en zeewier in het bijzonder. Dank-
zij deze inzichten kunnen we de fysiologie van de wieren beter begrijpen en kan het teeltsys-
teem verbeterd en duurzamer worden.
Omdat ik graag mijn steentje wil bijdragen aan het duurzamer maken van onze voedselpro-
ductie, heb ik voor dit onderwerp gekozen. Het is een klein begin, maar alle beetjes helpen.
Natuurlijk zou ik hier nooit alleen geraakt zijn. Daarom wil ik mijn promotor prof. Dr. Ir.
Werbrouck bedanken voor het ter beschikking stellen van zijn labo en zijn hulp bij de labora-
toriumproeven. Ook bedank ik graag Dr. Julia Wald en Dr. Willem Brandenburg van de Wa-
geningen Universiteit, om mij toe te laten tot dit unieke project. Ze hebben mij geïnspireerd
en ook steun geboden toen het onderzoek wat moeilijker ging. Ook dank aan alle medewer-
kers in het Laboratorium voor In Vitro Plantenbiotechnologie om een helpende hand te bieden
telkens het nodig was.
Het is in dit onderzoek gebleken dat wetenschappelijke resultaten niet altijd deze zijn die men
verwacht. Er waren heel wat tegenslagen en onverwachte resultaten. Hieruit valt te leren.
Als laatste wil ik ook heel graag mijn ouders en mijn vriend bedanken. Zij hebben mij gedu-
rende het proces gesteund en een luisterend oor geboden toen het nodig was. Dank jullie, voor
het nalezen en het advies.
Ondanks alle moeilijkheden is deze masterproef toch tot een goed einde gekomen en ik heb
veel bijgeleerd. Ik hoop dat dit eindwerk even leerzaam is voor anderen als voor mezelf en dat
het nuttig blijkt voor toekomstig onderzoek. Verder onderzoek is zeker nodig want er is geen
planeet B.

Abstract
Voedsel en energie zullen schaarser worden in de toekomst. Men zal de omvang van de voed-
selproductie moeten verdubbelen tegen 2050 en dit zal niet mogelijk zijn door conventionele
landbouw alleen. Er is voor conventionele landbouw te weinig plaats, zoet water en energie.
Een mogelijke bijdrage tot de oplossing is voedsel telen in de zee, waaronder het telen van
algen en wieren. Een duurzame optimalisering van deze teelt dringt zich op. Fundamenteel
onderzoek naar zeewier ondersteunt dit proces. Dit onderzoek beperkt zich tot één type wier,
namelijk Ulva lactuca. Het doel van dit eindwerk is de mogelijkheid te onderzoeken om Ulva
lactuca in vitro te vermeerderen.
Een in vitro cultuur van een weefsel vereist steriel weefsel. Er werden verschillende pogingen
ondernomen om een steriel stuk thallus te verkrijgen. De eerste methode was gebaseerd op de
‘landplantmethode’, namelijk via de rechtstreekse initiatie van uitwending gesteriliseerd thal-
lusweefsel op agarmedium. Verschillende klassieke sterilisatiemethoden werden uitgetest,
zoals spoelen met javel en ethanol, gassterilisatie of werken met bijzondere voedingsbodems
waarin proclin en antibiotica was verwerkt. Ulva lactuca thallus bleek zeer fragiel en niet be-
stand tegen de meeste sterilisatiebehandelingen. Ook de samenstelling van de voedingsbodem
moet verder onderzocht worden. Want hoewel zeesla een plant is, reageert het toch op een
aparte manier. De tweede methode om steriel weefsel te bekomen, bestond erin steriele sporen
of gameten te verkrijgen. Hiervoor moesten er eerst sporen of gameten gevormd worden. Dit
bleek een grotere uitdaging dan verwacht. De literatuur hieromtrent was immers niet eendui-
dig en de aangehaalde methodes niet reproduceerbaar. Een goed reproduceerbare methode om
sporen en gameten in het lab te induceren bleef uit. Het thallus reageert wel op stresscondities
door de vorming van reproductieorganen maar welke deze stresscondities precies zijn, bleef
nog onduidelijk. Het is wel zo dat het mogelijk is om deze sporen of gameten te isoleren en
steriel te krijgen. Maar voor de vorming van een effectief thallus zijn de resultaten uitgeble-
ven. De derde en laatste aanpak was om via een protoplastencultuur een steriel thallus te re-
genereren. Het was mogelijk om protoplasten te bekomen met 0,50mM MES, 1,5% cellulase,
1,5% abalone, 0,6M mannitol en 1%NaCl in miliQ water. De vorming van steriel thallus kon-
den we niet realiseren. Verder onderzoek rond de in vitro vermeerdering van Ulva lactuca is
dus nog nodig.
KERNWOORDEN: Ulva lactuca,, in vitro, protoplasten, sporen, gameten

Abstract
Food and energy will be scarce in the future. The food production must double by 2050 and
this will not be possible by conventional agriculture. There is little space, fresh water and en-
ergy for conventional agriculture. A possible contribution to the solution is to grow food in
the sea, like the cultivation of algae and seaweed. Sustainable optimization of this crop is im-
perative. Fundamental research of seaweed supports this process. This study is limited to one
type of seaweed, namely Ulva lactuca. The purpose of this thesis is to investigate how to mul-
tiply Ulva Lactuca in vitro.
An in vitro culture requires a sterile tissue. Several attempts were made to obtain a sterile
piece of thallus. The first method was based on the land plant method, namely through the
direct initiation of sterilized thallus tissue on agar medium. Several classic sterilization meth-
ods were tested , such ethanol rinse and bleach incubation, gas sterilization or administration
of ProClin 150 and antibiotics. Ulva lactuca thallus proved to be very fragile and not be re-
sistant to most sterilization treatments. The composition of the media needs to be further in-
vestigated. Even though sea lettuce looks like a land plant, it still responds differently. The
second method to obtain sterile tissue was by means of sterile spores or gametes. For this pur-
pose, the induction of spores or gametes was studied. This proved more challenging than ex-
pected. The literature was , after all, not so unambiguous and reproducible. A reproducible
method to induce spores and gametes in the lab was not developed. Normally, the thallus re-
sponds to stress conditions through the formation of reproductive organs, but the exact stress
conditions remained unclear. Although it was possible to induce and spores or gametes and to
isolate them under sterile conditions, they didn’t form a new thallus. The third and final ap-
proach was to regenerate a sterile thallus by means of a protoplast culture It was possible to
obtain protoplast with 0.50 mM MES, 1.5% cellulase, 1.5% abalone , 0.6 M mannitol, and 1%
NaCl in miliQ water solution. But we couldn’t realize the formation of sterile thallus. Further
research on the in vitro multiplication of Ulva lactuca is still needed .
KEYWORDS : Ulva lactuca, in vitro, protoplasts, spores, gametes

Inhoudstafel
Voorwoord ................................................................................................................................. 3
Abstract ...................................................................................................................................... 4
Abstract ...................................................................................................................................... 5
Inhoudstafel ................................................................................................................................ 6
Lijst met afkortingen .................................................................................................................. 9
Lijst met figuren ......................................................................................................................... 9
Lijst met tabellen ...................................................................................................................... 11
1 Inleiding .............................................................................................................................. 1
1.1 Probleemstelling .......................................................................................................... 1
1.2 Doelstelling .................................................................................................................. 2
1.3 Opbouw eindwerk ........................................................................................................ 2
2 Literatuur ............................................................................................................................. 3
2.1 Zeewier ........................................................................................................................ 3
2.1.1 Classificatie en naamgeving ................................................................................. 3
2.1.2 Morfologie van Ulva lactuca ............................................................................... 4
2.1.3 Verspreiding en nut van Ulva lactuca .................................................................. 5
2.1.4 Levenscyclus van Ulva lactuca ............................................................................ 5
2.1.5 De teelt van Ulva lactuca ..................................................................................... 6
2.2 In vitro teelttechniek .................................................................................................... 7
2.2.1 Ontstaan en werking in vitro teelt. ....................................................................... 7
2.2.2 In vitro teelt van landplanten ................................................................................ 7
2.2.3 Voor- en nadelen van in vitro teelt ....................................................................... 8
2.3 De in vitro teelt van zeewier ........................................................................................ 8
2.4 Vermeerdering door sporen en/of gameten ................................................................. 9
2.4.1 Vergelijkende literatuurstudie over sporen/gametenvorming .............................. 9
2.4.2 Sporen- en gametenvorming .............................................................................. 12
2.5 Protoplasten ............................................................................................................... 14
2.5.1 Definitie .............................................................................................................. 14
2.5.2 De celwand ......................................................................................................... 14
2.5.3 Enzymen ............................................................................................................. 15
2.5.4 Vergelijkende literatuurstudie ............................................................................ 17

3 Algemene materialen en methoden ................................................................................... 19
3.1 Herkomst Ulva lactuca .............................................................................................. 19
3.2 Teelt in aquaria .......................................................................................................... 19
3.3 Voedingsbodems ....................................................................................................... 20
3.3.1 Vast medium met zeewater (VMA) ................................................................... 20
3.3.2 Vast medium met zout water (VMB) ................................................................. 20
3.3.3 Vloeibaar medium (VlM) ................................................................................... 20
3.4 Microscopen .............................................................................................................. 21
3.5 Groei- of proefomstandigheden ................................................................................. 21
4 Experimenteel gedeelte ..................................................................................................... 22
4.1 Initiatie in vitro op agarmedium ................................................................................ 22
4.1.1 Doel .................................................................................................................... 22
4.1.2 Vaste voedingsbodems ....................................................................................... 22
4.1.3 Proeven in vloeibare voedingsbodems ............................................................... 30
4.2 Algemeen besluit in vitro initiatie met vast celweefsel ............................................. 36
4.3 Inductie sporulatie van Ulva lactuca ......................................................................... 36
4.3.1 Doel .................................................................................................................... 36
4.3.2 Inleiding ............................................................................................................. 36
4.3.3 Materiaal ............................................................................................................. 37
4.3.4 Methoden en resultaten ...................................................................................... 37
4.3.5 Bespreking sporulatie methoden ........................................................................ 47
4.3.6 Besluit sporulatie methoden ............................................................................... 47
4.4 Identificatie en isolatie sporen en gameten van U. lactuca ....................................... 48
4.4.1 Doel .................................................................................................................... 48
4.4.2 Identificatie van sporen en gameten ................................................................... 48
4.4.3 Isolatie van sporen .............................................................................................. 48
4.4.4 Bespreking identificatie en isolatie sporen en gameten ..................................... 51
4.4.5 Besluit identificatie en isolatie sporen en gameten ............................................ 51
4.5 Vorming van protoplasten ......................................................................................... 52
4.5.1 Doel .................................................................................................................... 52
4.5.2 Inleiding ............................................................................................................. 52
4.5.3 Vormen van protoplasten ................................................................................... 52
4.5.4 Bespreking vorming van protoplasten ................................................................ 57

4.5.5 Besluit: vorming van protoplasten ..................................................................... 57
5 Discussie ........................................................................................................................... 58
6 Algemeen besluit .............................................................................................................. 61
Referentie lijst .......................................................................................................................... 63
Bijlage ......................................................................................................................................... i

Lijst met afkortingen
AB Antibiotica.
AD Aqua destilata of gedestilleerd water.
BA 6-Benzyladenine.
FAO Food and Agricultural Organisation of the United Nations.
FDA Fluoresceïne diacetaat.
MES 2-(N-morpholino)ethane sulphonic acid.
MS Murashige en Skoog medium.
PAR Aantal μmol fotonen per vierkante meter per seconde = Photosynthetic Active
Radiation.
PM Protoplast medium.
VIB Vlaams instutuut voor biotechnologie.
VLM Vloeibaar medium.
VMA Vast medium gemaakt met zeewater.
VMB Vast medium met extra zout.
Lijst met figuren
Figuur 1: De verschillende structuren van gedifferentieerd weefsel van Ulva lactuca eigen
bewerking van (Tormo Molina, 2012) ....................................................................................... 4
Figuur 2: De reproductieve voortplantingscyclus van Ulva lactuca (Ponette & Bertin) ........... 5
Figuur 3: De vorming van gameten in Ulva mutabilis (Wichard & Oertel, 2010) .................. 12
Figuur 4: Bovenaan gesporuleerd weefsel (40X) Onderaan een duidelijke scheidingslijn
tussen vegetatief weefsel en gesporuleerd weefsel (20X) Microscopiefoto's genomen door
Norbert De Ruiter, het weefsel gevonden door Jetze Van Zwol .............................................. 13
Figuur 5: De celstructuur van Ulva sp. (Sperber, 2010) .......................................................... 14
Figuur 6: Turgor druk in plantencellen (wikipedia) ................................................................. 18
Figuur 7: Schematische weergave van de zeewier proefhoeve van Wageningen Universiteit 19
Figuur 8: Aquaria met Ulva lactuca ......................................................................................... 19
Figuur 9: Afgestorven, wit zeewier na één minuut in 10% javel oplossing ............................. 23
Figuur 10: Gespoelde Ulva lactuca met 1% NaClO (javel) ..................................................... 23
Figuur 11: Links onidentificeerbare doorzichtige vloeistof rond uitgesneden schijfjes, rechts
een duidelijke schimmel, bacterie en/of gist contaminatie....................................................... 24
Figuur 12: De grootte van het stukje thallus heeft geen invloed op de overleving op een
voedingsbodem ......................................................................................................................... 26
Figuur 13: Links bij de start van het experiment, midden en rechts dode en gecontamineerde
thallus schijfjes ......................................................................................................................... 27
Figuur 14: Resultaat antibioticaproef na 1 week: plaat 9 is gecontamineerd, het weefsel in
plaat 7 daarentegen leeft nog en is niet gecontamineerd .......................................................... 29
Figuur 15: Thallusschijfjes in vloeibaar zeewiermedium, links bij start rechts een week later
.................................................................................................................................................. 30

Figuur 16: Thallus gespoeld met steriel zeewater en Tween, links na dag 1 - rechts na 6 dagen
.................................................................................................................................................. 31
Figuur 17: Glazen stolp in trekkast waarin de gassterilisatie werd uitgevoerd ........................ 33
Figuur 18: 15 minuten natte gassterilisatie na tien dagen. ....................................................... 34
Figuur 19: Links de start van de antibiotica-proefopzet, rechts de proef negen dagen later,
gecontamineerd door micro organismen. ................................................................................. 35
Figuur 20: Links bij start van de proef en rechts een gesporuleerde versie, achtendertig dagen
later ........................................................................................................................................... 38
Figuur 21: Links het weefsel bij de start van het experiment en rechts zevenenveertig dagen
later. Het weefsel is dan zeer broos en het water rondom is licht groen gekleurd .................. 38
Figuur 22: Vermoedelijk gameten van Ulva lactuca. .............................................................. 39
Figuur 23: Herhaling van de tweede methode van Sluis (1.2). Links bij start van de
proefopzet rechts drieëndertig dagen later. Er is dan een groene wolk zichtbaar, dit zou
kunnen wijzen op het vrijkomen van sporen of gameten. ....................................................... 39
Figuur 24: Weefsel en omringend zeewater van de tweede keer methode 1.2 na 33 dagen, 40
maal vergroot. ........................................................................................................................... 40
Figuur 25: Derde uitvoering van de tweede methode van Sluis (1.2). Links met oud weefsel
dat al bijna een jaar in het opslagaquarium zat, rechts met nieuw, vers geoogst, weefsel ...... 41
Figuur 26: Beeld van de witte stukken weefsel bij 40X vergroting ......................................... 42
Figuur 27: Resultaat methode 4 ............................................................................................... 43
Figuur 28: Dood weefsel bij 40x vergroting ............................................................................ 44
Figuur 29: 11B na 17 dagen ..................................................................................................... 46
Figuur 30: 5A, het groene gedeelte is levend weefsel het zwarte gedeelte afgestorven weefsel
en het witte zou gesporuleerd weefsel kunnen zijn .................................................................. 46
Figuur 31: 11B: Links 20X vergroting van de rand. Midden een 40X vergroting van de
overgang van levend naar dood weefsel. Rechts lege cellen waarvan het niet duidelijk was of
ze gesporuleerd zijn .................................................................................................................. 46
Figuur 32: Links gecontamineerde voeding met weefselcellen. Midden vers overgeënte cellen
in VIM, Rechts groei van gameten na drie weken .................................................................. 49
Figuur 33: Sporen op VMA ..................................................................................................... 49
Figuur 34: Microscopisch beeld (40X vergroting) van de inhoud van de falcon met VLM. ... 49
Figuur 35: Gameten incubatieopstelling volgens (Lascaux, 2002) met links een aantal
opgepipetteerde losse gameten en rechts een stukje gevormd weefsel uit gameten. ............... 50
Figuur 36: Het vermenigvuldigen van de sporen in de duranfles met VLM ........................... 50
Figuur 37: Het VLM verdampte onder invloed van de droge perslucht die er in geblazen werd
.................................................................................................................................................. 50
Figuur 38: Zeewier weefsel gevormd na 70 dagen .................................................................. 50
Figuur 39: Na drie uur beginnen de cellen los te komen (20X vergroot) ................................ 55
Figuur 40: Links, één van de weinige protoplasten bij 0M mannitol na 16u incubatie. Rechts,
de herhaling, 24u incubatie ook 0M mannitol. (40X vergroting) ............................................ 55
Figuur 41: 0,8M mannitol na 24u incubatie (40X vergroting) ................................................. 55
Figuur 42: 0,6M mannitol na 24u incubatie (40X vergroting) ................................................. 55
Figuur 43: Celwand die aan het loskomen is, bekeken door de geïnverteerde microscoop..... 56

Lijst met tabellen
Tabel 1: Vergelijking van de verschillende in de literatuur beschreven methodes betreffende
de vorming van sporen en/of gameten bij gerelateerde zeewiersoorten Bronnen: (Azanza &
Aliza, 1999; Goudsmit, 2013; Han et al., 2008; Mantri et al., 2010; Niesenbaum, 1988; Ohno
& Dut Triet, 1997; Van Der Sluis, 2012; Wichard & Oertel, 2010). ....................................... 11
Tabel 2: Samenvatting van de verschillende manieren om protoplasten te vormen (Björk et al.
,1992; Deryckere et al.,2012; Reddy & Fujita, 1991; Reddy et al. ,2007; Uchida et al. , 1992;
Uppalopati & Fujita, 2002; Van Ginneken et al. , 2013) ......................................................... 17
Tabel 3: Tweede proefopstelling met antibiotica, de nummers in de tabel zijn de nummers van
de platen ................................................................................................................................... 28
Tabel 4: Abiotische parameters van de verschillende fytotrons, gebruikt in het experiment .. 37
Tabel 5: Proefopzet methode 7 ................................................................................................. 45
Tabel 6: Dag drie methode 7 .................................................................................................... 45
Tabel 7: Dag vier methode 7 .................................................................................................... 45
Tabel 8: Dag zeven methode 7 ................................................................................................. 45
Tabel 9: Dag 17 methode 7 ...................................................................................................... 46

1
1 Inleiding
1.1 Probleemstelling
De groeiende wereldbevolking stelt ons voor grote uitdagingen. Voorspeld wordt dat we in
2050 met negen miljard mensen zullen zijn (Fischer, 2009). Voedsel en energie zullen schaar-
se goederen worden. De huidige omvang van voedselproductie zal moeten verdubbelen om
aan de groeiende vraag te voldoen. Deze stijging kan niet door de conventionele landbouw
alleen opgevangen worden (UN, 2013; NCDO, 2011). De conventionele landbouw beschikt
over een steeds krimpend landbouwareaal en verbruikt op een haast verkwistende wijze kost-
baar water en eindige bronnen zoals fosfaten (Berkhout et al., 2010).
Naast het matigen van onze huidige energie- en voedselconsumptie hier in het Westen, is het
telen van voedsel in de zee een mogelijke oplossingsstrategie. Zo zou gecontroleerde en sys-
tematische productie van eetbaar zeewier een bescheiden bijdrage kunnen leveren. Bij deze
evolutie is duurzaamheid het codewoord. Er moet op toegekeken worden dat niet het Aziati-
sche voorbeeld gevolgd wordt, waar de teelttechniek de zeefauna en –flora aantast en het
zeewater vervuild wordt door overvloedig nutriëntengebruik (Bandenburg, 2013).
Het duurzame telen van zeewier staat nog in de kinderschoenen. Er is voorlopig nog een ge-
brek aan wetenschappelijk kennis en bijhorende vaardigheden om dit voluit te laten slagen. Er
zijn reeds duurzame teeltexperimenten zoals op de proefhoeve “de wierderij” van Wagenin-
gen Universiteit in de Oosterschelde en de commerciële boerderij Zeewaar, van Rebecca Wie-
ring en Jennifer Breaton. Er is echter nood aan een hogere teeltefficiëntie.
In dit werk hebben de Wageningen Universiteit, met haar reeds bestaande kennis voor wat
betreft het telen van zeewier, en de Universiteit Gent met haar ervaring met in vitro cultuur
van planten elkaar ontmoet in een gezamenlijke zoektocht naar een gecontroleerde en effici-
ënte vermeerderingsstrategie voor zeewier. Tot nu toe hanteert men een arbeidsintensieve
methode van vegetatieve vermeerdering waarbij stukken thallus tussen touwen worden ge-
bonden en zo in zee gehangen om te groeien. Misschien kan Ulva lactuca in vitro vermeer-
derd worden, net zoals andere planten. Op die manier zouden selecties snel en arbeidsbespa-
rend vermeerderd kunnen worden. Het zou ook in vitro veredelingstechnieken toelaten zoals
ploïdiemanipulatie of celfusie.

2
1.2 Doelstelling
Studieobject was Ulva lactuca. Ulva lactuca werd gekozen omdat dit een modelwier is waar-
aan meerdere studies gewijd zijn. De doelstelling van dit project was het in vitro initiëren van
Ulva lactuca. Er werden verschillende strategieën gevolgd: klassieke uitwendige sterilisatie,
inductie van sporen- en gameten en isolatie van protoplasten. Dit alles om te kunnen starten
met in vitro-teelt en vermeerdering.
1.3 Opbouw eindwerk
De literatuurstudie biedt informatie over wieren in het algemeen en Ulva lactuca in het bij-
zonder. Vermeerdering via sporen en/of gameten wordt vanuit deze optiek begrijpelijk. Ook
in vitro cultuur wordt kort besproken. Dit betreft voornamelijk in vitro cultuur van hogere
planten aangezien er over de in vitro teelt van algen en zeewieren maar sporadisch gepubli-
ceerd werd. We wilden begrijpen en vergelijken. Zoals eerder vermeld, bleek het steriel krij-
gen van de zeesla een hele uitdaging. Steriliteit is een noodzakelijke voorwaarde voor het in
vitro bestaan. Tijdens het onderzoek drongen zich andere methodes op. Vandaar een stuk lite-
ratuurstudie over sporen en gameten en een deel over protoplasten. Om protoplasten te isole-
ren was het nodig om de celwand van Ulva lactuca te beschrijven, net zoals de enzymen die
nodig zijn om deze celwand af te breken.
Na de literatuurstudie komt er een hoofdstuk handelend over algemene materialen en metho-
den. Dit beschrijft de materiële context van het onderzoek: het in leven houden van het wier in
aquaria, de groei- en proefomstandigheden, de gebruikte voedingsbodems e.d.
Hierna volgt het experimenteel gedeelte. Dit werd opgedeeld in drie grote delen: de in vitro
initiatie op agarmedium, de inductie en identificatie van sporen en gameten en de productie
van protoplasten. Van al deze onderwerpen werden materiaal en methode uitgewerkt en wer-
den de resultaten besproken. Dit eindwerk eindigt met een discussie, besluit en aanbevelingen
voor mogelijk toekomstig onderzoek.

3
2 Literatuur
2.1 Zeewier
2.1.1 Classificatie en naamgeving
Zeewier behoort tot het rijk van de algen of wieren. De afwezigheid van hoog gedifferentieer-
de cellen vormt het onderscheid tussen het rijk van de Plantae (gewone planten) en de Protis-
ta (algen). Wieren zijn autotrofe chlorofylhoudende organismen, zeer gevarieerd qua morfo-
logie en kleur. Ze behoren tot de Thallophyta, planten met een thallus, vroeger ook wel de
‘lagere planten’ genoemd (Coppejans, 1995).
Wieren omvatten alle chlorofylhoudende organismen die autotroof zijn t.o.v. koolstof maar
geen echte wortels, stengels noch bladeren bevatten. Er zijn landwieren, bijvoorbeeld lichenen
(korstmos, symbiose tussen wier en schimmel op rotsen of boomschors) en wieren die in zoet,
brak, en zout water leven (Coppejans, 1995).
Er zijn micro- en macro-algen. Microalgen zijn eencellig en al dan niet mobiel via flagellen of
zweepharen. Met macro-algen wordt een meercellige structuur bedoeld. Deze kan vasthangen
of vrij zwevend zijn (groenwieren, 2011). Zeewieren zijn zee-macroalgen. Daarom worden ze
beschouwd als planten ook al hebben ze geen vaatbundels (Coppejabs, 1995).
Er zijn verschillende soorten zeewieren: roodwieren (Rhodophyta), bruinwieren (Phaeophyta)
en groenwieren (Chlorophyta). Het onderzoek werd beperkt tot één type van Chlorophyta,
namelijk Ulva lactuca. Chlorophyta zijn groenwieren. Ze hebben deze naam verkregen door
hun groene kleur. Chlorophyta verkrijgen hun groene kleur door de chlorofyl a en b (groen
pigment), β-caroteen (geel pigment) en verschillende karakteristieke xanthophyllen (geel
bruin) in hun cellen (Guiry, 2000). Het zijn de evolutionaire voorlopers van de landplant
(Knee, 2008). Groenwieren kunnen in diverse biotopen voorkomen. Er zijn 5500 soorten en
meer van 90% van die soorten komt voor in zoet water. Er zijn ook groenwieren die voorko-
men in brak water, aan lucht blootgestelde substraten (bodem, rotsen bomen…) en uiteraard
ook in zeewater (Coppejans, 1995). Ze leven tussen 0 en 10 m diepte in zee om voldoende
licht te capteren voor fotosynthese (Brandenburg, 2013).
Rijk: Protoctista
Fylum: Chlorophyta
Genus: Ulva
Soort: Ulva lactuca L.
(BioInformatics, 2013)

4
Ulva lactuca, ook wel gekend onder de naam zeesla, is een opportunistische soort met een
variërende zouttolerantie en een extreem bereik van K+/Na
+ ratio. Dit wier is efficiënt in het
verwijderen van N en P (Van Ginneken et al., 2013). De soort Ulva lactuca is van het ge-
slacht Ulva. Dit is een meercellige stationaire “plant” met een eencellige mobiele generatieve
fase (BioInformatics, 2013; Knee, 2008).
2.1.2 Morfologie van Ulva lactuca
Ulva lactuca is een statisch multicellulair type wier met een mobiele eencellige reproductieve
fase. Dit wil zeggen dat de thallus bestaat uit meerdere cellen die een geordende structuur
vormen. Deze is statisch, dus niet bewegend. De seksuele en aseksuele voortplanting gebeurt
door middel van sporen of gameten. Deze hebben zweepharen en zijn mobiel. Zeesla bevat
een graad van cellulaire differentiatie en organisatie met verschillende structuren (Knee,
2008) (zie Figuur 1).
Figuur 1: De verschillende structuren van gedifferentieerd weefsel van Ulva lactuca
eigen bewerking van (Tormo Molina, 2012)
Ulva lactuca is groen tot donkergroen. De thallus kan tot één meter lang en 30 cm breed wor-
den en is zacht, gerimpeld en doorzichtig. Het is een membraan dat uit twee onregelmatig
geordende cellagen bestaat. Ulva kan zich vasthechten aan steen of een andere vaste substan-
tie met een klein schijfvormig voetje dat via een versterkt tussenstukje verbonden is met de
thallus. Deze hechtschijf is verstevigd door rizoïden (net zoals schimmeldraden) (Burrows
1991; Keizer, 2013; Coppejans, 1995). Ulva lactuca is een polymorfe soort die niet makkelijk
te differentiëren is (Van Ginneken et al., 2013). Dit wil zeggen dat er geen verschil waar te
nemen is tussen de diploïde en haploïde fase (zie 2.1.4).

5
2.1.3 Verspreiding en nut van Ulva lactuca
Ulva lactuca kan overal ter wereld voorkomen, van gematigde tot subtropische gebieden. Het
gedijt bij verschillende temperaturen en zoutgehaltes. Het komt zowel voor in het intergetij-
dengebied als ook dieper in zee. Omdat zeesla veel chlorofiel bevat (groenwier) heeft het wel
veel licht nodig om te kunnen groeien en komt niet dieper dan 10m diepte voor in zee (Keizer,
2013).
Zeewier kan gebruikt worden voor verschillende doeleinden. Als voedselcomponent voor
humane consumptie zoals bijvoorbeeld het gebruik als geleermiddel, farmaceutische toepas-
singen, maar ook voor dierenvoeding. Naast humane toepassingen kan ook de opvang en ver-
werking van de nutriëntenverliesstroom en de mogelijke bron als biobrandstof een extra
waarde geven aan de zeewierteelt (Goudsmit, 2013; Wolkers, 2011).
2.1.4 Levenscyclus van Ulva lactuca
De levenscyclus van Ulva lactuca start met vrouwelijke of mannelijke haploïde planten (ga-
metofyt) die gameten produceren. Uit de versmelting van mannelijke en vrouwelijke gameten
ontstaat de zygote waaruit een diploïde plant (sporofyt) ontspruit. Deze sporofyt levert na
reductiedeling haploïde sporen. De sporen vormen dan weer een volledig haploïde thallus
waardoor de cyclus gesloten is (figuur 2) (Guiry, 2000).
Figuur 2: De reproductieve voortplantingscyclus van Ulva lactuca (Ponette & Bertin)

6
Ulva lactuca is een haplodiplont, dit wil zeggen dat er zowel een haploïde als een diploïde
fase bestaat die beiden als zelfstandige plant kunnen voorkomen. Zeewieren kunnen zich op
drie manieren voortplanten: via de vegetatieve voortplanting, via de ongeslachtelijke voort-
planting en via de geslachtelijke voortplanting. De vegetatieve voortplanting is via scheuren
van stukjes thallus. De meest voorkomende manier van voortplanting is de ongeslachtelijke
voortplanting. Dit gebeurt door middel van zoösporen. Deze hebben vier flagellen en zijn iets
groter dan de gameten. Geslachtelijke voortplanting vindt plaats door middel van gametenco-
pulatie, zeesla heeft geflagelleerde gameten met twee flagellen. De mannelijke gameten zijn
iets kleiner dan de vrouwelijke gameten (Busink, et al., 2009; Eichhorn et al., 2005).
De haploïde en de diploïde fase wisselen af en zijn morfologisch nauwelijks te onderscheiden.
Er zijn maar enkele verschillen tussen de sporen en de twee types gameten, namelijk hun
grootte, de differentiatie van de cel voor de sporulatie en het aantal flagellen (Knee, 2008).
2.1.5 De teelt van Ulva lactuca
Volgens de Food and Agricultural Organization of the United Nations, de FAO, hebben enkel
China, Zuid-Korea en Japan een geregistreerde zeewierteelt voor voedselconsumptie
(www.fao.org). Naast de productie van Zeewaard, die in 2013 begonnen is, komt er in Neder-
land en België enkel experimentele teelt voor.
De teelt van zeesla kan op verschillende manieren gebeuren. Hieronder worden de twee voor-
naamste teeltvormen besproken, met name de intensieve en de extensieve teeltmethoden.
Bij intensieve teelt wordt op kleine schaal zeewier geteeld in tanks of in vijvers, dit meestal
als afvalbehandeling of als onderdeel van een geïntegreerde visboerderij. Deze teeltmethode
vraagt veel ruimte en aanzienlijke financiële middelen gezien de benodigde infrastructuur en
het onderhoud: het verversen van het water, de aangepaste belichting en verwarming indien
nodig (Van Ginneken et al., 2013; Busink et al., 2009).
Bij extensieve teelt wordt op grote schaal zeewier gecultiveerd met beperkte technologische
input. Men gebruikt gefixeerde bodems en drijvende touwen in de kustzones of verder in zee.
Dit kan gebeuren op drijvende vlotten of met netten tussen vast geankerde palen. De gefixeer-
de bodems zijn vaak lijnen van bamboe die op de zeebodem worden gefixeerd door betonnen
blokken (Van Ginneken et al., 2013; Busink et al., 2009; FAO, 2003).
De commerciële teelt gebeurt nu door vegetatieve vermeerdering. Kleine tukjes thallus wor-
den tussen de touwen geknoopt of in de tanks vrijgelaten. Deze groeien dan verder uit tot gro-
tere stukken thallus. Deze manier van werken is vergelijkbaar met het stekken bij gewone
landplanten (Brandenburg, 2013).

7
2.2 In vitro teelttechniek
De zeewierteelt op zee is nog niet voldoende efficiënt. Hier komen we bij het doel van dit
onderzoek. Door in vitro cultuur zouden we meer kennis, informatie en misschien wel een
efficiëntere manier van telen kunnen bekomen. Nu wordt zeewier vegetatief vermeerderd.
Stukjes zeewier worden manueel tussen touwen gestoken die aan pontons hangen. Dit is een
moeilijk en arbeidsintensief werk. Bovendien is de kwaliteit niet altijd even uniform en be-
staat er momenteel ook nog onduidelijkheid over bepaalde ziektesymptomen. Een cultuur in
vitro geeft mogelijkheid tot een snelle vermeerdering, kruising, selectie en verder onderzoek
naar zeewier.
2.2.1 Ontstaan en werking in vitro teelt.
In vitro betekent letterlijk “in glas”. In deze context wordt de term vooral gebruikt voor biolo-
gische technieken die buiten de normale omgeving, in gecontroleerde omstandigheden wor-
den toegepast. Dit hoeft uiteraard geen ‘glazen’ recipiënt te zijn. Onder in vitro cultuur ver-
staat men het kweken of telen van plantendelen op een kunstmatig samengestelde voedings-
bodem en dit onder gecontroleerde en steriele omstandigheden. Deze teeltvorm is mogelijk bij
verschillende plantendelen zoals geïsoleerde organen, weefsels, embryo’s, zaden, vruchtbe-
ginsels, meeldraden, cellen en zelfs protoplasten. In vitro teelt is mogelijk doordat plantencel-
len plastisch en totipotent zijn. De plasticiteit staat voor het grote aanpassingsvermogen van
plantencellen in zeer wisselende omstandigheden. Totipotentie stelt dat uit elke plantencel een
volledig nieuwe plant kan verkregen worden (Huys, 2002; INRA, 2013). De vraag is of dit
ook geldt voor zeewiercellen.
2.2.2 In vitro teelt van landplanten
De kennis over het in vitro telen van zeewier is bijna nihil, dus moesten we dit project baseren
op de kennis bij landplanten. Tot nu toe wordt de teelt van planten in vitro immers bijna uit-
sluitend op landplanten toegepast, en dan nog wel voornamelijk op zaadplanten.
De celwand van een landplant bevat veel lignine om de plant rechtop te houden en te be-
schermen tegen externe en interne factoren. Ook is er een waslaag omheen de celwand die
extra bescherming kan bieden (Verkerk et al., 2004). Dankzij deze beschermende lagen is het
makkelijker om deze planten steriel te krijgen. Deze steriliteit is noodzakelijk voor het over-
leven in vitro. De in vitro voedingsbodem met zijn vele nutriënten is als het ware een feest-
maal voor allerlei parasieten. Ze koloniseren het medium en tasten vaak ook de plant aan.
Dus, zonder een steriele plant, geen in vitro teelt. Het steriliseren van planten gebeurt met
ethanol en javel. Een groeipunt wordt steriel gemaakt, afgesneden en in de voedingsbodem
geplant. Door de rijke hoeveelheid aan nutriënten gaat deze extreem goed groeien en is een
snelle vermeerdering mogelijk (Werbrouck, 2011; INRA,2013).

8
2.2.3 Voor- en nadelen van in vitro teelt
In vitro teelt zorgt niet alleen voor een snelle vermenigvuldiging op basis van vegetatieve
vermeerdering, het maakt het ook mogelijk om op een makkelijke manier aan wetenschappe-
lijk onderzoek te doen en planten te veredelen. Onder deze gecontroleerde omstandigheden is
het niet moeilijk om de invloed van één specifieke parameter te controleren zonder dat er in-
terferentie is van bijvoorbeeld andere organismen of onbekende chemische stoffen
(Werbrouck, 2011; Huys, 2002). Ook fysiologisch onderzoek zou mogelijk kunnen zijn. Hoe
reageert deze primitieve plant op bepaalde plantenhormonen? Welke biochemische processen
zijn er al aanwezig? (Lascaux, 2002). Dit zou allemaal kunnen bijdragen tot een efficiëntere
manier van planten/zaaien in de commerciële zeewierteelt.
Een nadeel is wel dat in vitro telen een aanzienlijke financiële en infrastructurele inspanning
vraagt. Alle materiaal en installaties voor cultuurontwikkeling, zoals bijvoorbeeld een lami-
naire airflow of fytotrons, zijn duur. Het vereist bovendien een omkadering met een hoog tot-
voldoende wetenschappelijk geschoold personeel. (Werbrouck, 2011).
2.3 De in vitro teelt van zeewier
In de literatuur is er weinig te vinden over de in vitro teelt van zeewier en al helemaal niets
over het in vitro telen van Ulva lactuca. Azanza en Aliza (1999) vermelden het vrijkomen van
sporen in kunstmatig verrijkt zeewater. Zij voegen minieme hoeveelheden antibiotica toe aan
de oplossing om de groei van epifyten en ongewenste micro-organismen tegen te gaan. Ook
Lascaux (2002) vermeldt het in vitro telen van wieren. Hij benadrukt de hoge moeilijkheids-
graad en stelt dat deze specifieke teeltmethodes nog niet op punt staan. Zijn artikel is eerder
een uiteenzetting rond de bestaande werkpunten. De beste uitgewerkte methoden in dit artikel
zijn die voor kranswieren (Characeae). Dit zijn zoetwaterwieren en de rechtstreekse voorou-
ders van de landplanten. Bij Characeae is er al een duidelijke differentiatie tussen de stengel
en bladachtige structuren al wordt deze nog steeds “thalli” genoemd. Deze zijn iets steviger en
makkelijker steriel te krijgen (Lascaux, 2002).

9
2.4 Vermeerdering door sporen en/of gameten
Een alternatieve werkmethode om steriel wierweefsel te bekomen, is door sporen en/of game-
ten steriel te isoleren en deze te laten uitgroeien tot een steriel thallus. Deze werkwijze loopt
analoog met de in vitro cultuur van mossen, die taxonomisch dicht bij wieren staan
(Demeulenaere, Sterilizeren mossen, 2013).
2.4.1 Vergelijkende literatuurstudie over sporen/gametenvorming
Sporen en/of gameten zijn de voortplantingscellen van zeewieren. Deze kunnen gebruikt wor-
den voor een snelle manier van ‘zaaien’ van het zeewier (Azanza & Aliaza, 1999; Clifton &
Clifton, 1999). Gameten en/of sporen worden synchroon vrijgegeven op bepaalde tijdstippen.
De trigger tot dit gebeuren is nog niet helemaal achterhaald. Clifton (1999) maar ook Gouds-
mit (2013), Niesenbaum (1988), Ohno en Duy Triet (1997) en Han et al (2008) zeggen dat dit
bepaald wordt door omgevingsfactoren zoals temperatuur en licht. Deze veranderingen zijn
dus afhankelijk van het seizoen en de getijdenwerking in natuurlijke omstandigheden. Niese-
baun (1998) en Goudsmit (2013) beweren dat er een productie is van sporen na een koudebe-
handeling als een soort van overlevingsreactie. Voor Ohno & Duy Triet (1997) is het juist een
temperatuurverhoging die de productie van zoösporen vrijstelt. Bij Han et al. (2008) is het in
stukken snijden al genoeg om sporen te laten vrijkomen. Zij beweren dat indien er toxische
stoffen in het water zitten, deze de productie van sporen zullen minderen of helemaal tegen-
houden.
Wichard & Oertel (2010) kwamen met een andere theorie op de proppen. Zij beweren dat als
je de sporulatie inhibitoren -die door het zeewier zelf geproduceerd worden- weghaalt, er
spontaan gameetvorming ontstaat. Ze stellen ook dat er een donkere periode nodig is voor het
omzetten van de cellen in gameten. Martri et al. (2010) beweert dan weer dat de zoutconcen-
tratie de trigger is om zoösporen te induceren. Bij een lager dan normaal zoutgehalte, zouden
er spontaan sporen moeten ontstaan.
Veel van deze voorgaande artikels vonden het telkens weer de moeite waard om het artikel
van Smith (1947) en Lünig & Kadel (2008) over het maanafhankelijk zijn van sporulatie te
vermelden of tegen te spreken. Volgens hen zouden er vooral sporen en/of gameten gevormd
worden bij volle maan en nieuwe maan. Dit inzicht is plausibel omdat de getijdenwerking
nieuw, vers en koud water aanvoert.

10
Het is dus nodig al deze verschillende methoden naast elkaar te leggen (zie Tabel 1). Er werd
geen onderscheid gemaakt tussen de literatuur die specifiek bezig is met vorming van game-
ten of deze die over sporenvorming handelde. De standaard test-temperatuur lag tussen de 15
en de 25°C (meest voorkomend 20°C). Er was een grote variatie in de hoeveelheid licht die
nodig was voor het bekomen van sporen en/of gameten. Het zoutgehalte in het water lag tus-
sen de 15 en de 32 %. (meestal 30%). De pH was de standaard zeewater pH tussen 7 en 9. En
de meeste artikels creëerden sporen en/of gameten door het weefsel een shock te geven. Dit
door het zeewierweefsel bloot te stellen aan temperatuursverandering, het toebrengen van
schade door uitdroging of versnijding of het plaatsen in duisternis gedurende een bepaalde
periode.

11
Tabel 1: Vergelijking van de verschillende in de literatuur beschreven methodes betreffende de vorming van sporen en/of gameten bij gerelateerde zeewiersoorten Bronnen: (Azanza
& Aliza, 1999; Goudsmit, 2013; Han et al., 2008; Mantri et al., 2010; Niesenbaum, 1988; Ohno & Dut Triet, 1997; Van Der Sluis, 2012; Wichard & Oertel, 2010).
methode type wier temperatuur
(°C)
licht (µmol
fotonen m-2
s-
1)
licht
regime
zout
gehalte
(‰)
pH Hoeveelheid
wier
cultuurmedium
(ml)
extra
Azanza and
Aliaza
(1999)
Kappaphy-
cus alvarezii
27 200 12uL
12uD
30 twee takken? 50 één nacht drogen
Goudsmit
(2013)
Ulva lactuca 20 ? ? 30 - 35 8 -
8,5
40g (10 thal-
li)
150 000 bij start een koudebehande-
ling 24u bij 4°C in het don-
ker
Han et al.
(2008)
Ulva sp. 15 - 20 30 12uL
12uD
25 - 35 7,0-
9,0
één schijfje
met diameter
6mm
10
Mantri et al.
(2010)
Ulva fasciata 25 15 12uL
12uD
15 7,8 schijfje met
diameter
5mm
30
Niesen-
baum
(1988)
Ulva lactuca 21 44,87 9uL
15uD
? 8 -
8,5
12,5g 2 000 eerste 24u 2°C en maar 9u
licht per dag
Ohno and
Duy Triet
(1997)
Monostroma 21 verhoogd
in 0,5u tot 27
(+ vers warm
water)
100 ? 30 ? 5kg ? één nacht drogen
van der
Sluis (2012)
Ulva lactuca 15 ? 16uL
8uD
30- 35 ? een schijfje
met diameter
5mm
? start bij 11° C.
bij verkleuren van de schijf-
jes: 15°C
Wichard
and Oertel
(2010)
Ulva mutabi-
lis en Ulva
lactuca
20 60-120 16uL
8uD
? ? blade in stuk-
jes snijden
van ± 1 á 3
mm
10mg biomassa
per liter
bij start eerst 15 min. was-
sen met gedestilleerd water
vóór in cultuurmedia te
stoppen en derde dag medi-
um verversen.

12
2.4.2 Sporen- en gametenvorming
Aanvankelijk is er een gewone vegetatieve G1 cell-cyclus fase waarin gametogenesis kan
geïnduceerd worden, door het verwijderen van de populatie-inhibitor in het medium (Wichard
& Oertel, 2010). Dan gaat de cel over naar een vegetatieve S fase in dewelke het genoom ge-
repliceerd wordt. De daarop volgende fase is terug een G1 fase. Na deze tweede G1 fase komt
een S fase met inductie van gametogenese en een accumulatie van zetmeelgranulaten. Daarop
volgt een irreversibele gametangium differentiatie, een periode van progameet vorming, chlo-
roplastheroriëntatie en papilla-initiatie. Daarna worden de progameten gemultipliceerd (16
vrouwelijk, 8 mannelijk) en de papilla maturatie. Dit wordt gevolgd door de maturatie van de
gameten en de porie kap. Hierna opent de porie kap (Wichard & Oertel, 2010).
Figuur 3: De vorming van gameten in Ulva mutabilis (Wichard & Oertel, 2010)
De ongeslachtelijke voortplanting verloopt door de vorming van zoösporen. De sporophyte
produceert haploïde sporen door meiose (Eichorn et al., 2005). Eerst vormen de cellen die aan
de rand liggen zoösporen. Daarna worden steeds meer cellen door zoöspoorvorming geledigd.
Tenslotte blijft slechts een kleurloos vel met lege cellen van de plant over (Hardenberg,
2013).

13
Tijdens reproductie van het groen zeewier Ulva, is het afstaan van levende cellen geassoci-
eerd met een kleurverandering. Eerst wordt het weefsel geelachtig groen dan donker olijfkleu-
rig tot wit. De witte kleur reflecteert het feit dat de reproductieve delen vrijgelaten zijn (Han,
et al., 2008). Afhankelijk van het percentage kleurverandering kan er bepaald worden hoeveel
sporen en/of gameten er gevormd werden. Het is wel belangrijk om het verschil te zien tussen
dood weefsel (ook wit) en gesporuleerd weefsel. Weefsel dat sporuleerde, zou een scherpe
rand vertonen (Goudsmit, 2013). Op een stuk thallus zou het van groen weefsel plots over-
gaan naar kleurloos weefsel. Dood weefsel anderzijds is meer slijmerig en valt na aanraking
uit elkaar. Onder de microscoop is dit onderscheid duidelijk zichtbaar. Lege cellen met een
zwart puntje wijzen op gesporuleerd weefsel. Dit zwarte puntje is de opening waardoor de
sporen of gameten de cel verlaten hebben (Van Der Sluis, 2012; Ruiter, 2013; Wald, 2013).
Figuur 4: Bovenaan gesporuleerd weefsel (40X)
Onderaan een duidelijke scheidingslijn tussen vegetatief weefsel en gesporuleerd weefsel (20X)
Microscopiefoto's genomen door Norbert De Ruiter, het weefsel gevonden door Jetze Van Zwol

14
2.5 Protoplasten
Een tweede alternatieve weg om steriel weefsel te verkrijgen is te werken via een protoplas-
tencultuur. Isolatie en regeneratie van protoplasten zijn belangrijke stappen om genetische
variatie in te voeren. Deze genetische variatie kan bekomen worden via het gebruik van so-
maclonale variatie, door mutagenese, via somatische hybridisatie of door transformatie. Maar
bij de algen zijn er voor fusie en regeneratie nog veel problemen op te lossen (Lascaux, 2002).
2.5.1 Definitie
Protoplasten zijn “naakte cellen”. Het zijn levende cellen zonder celwand. Deze celwand kan
op mechanische, enzymatische, immunologische, chemische, spontane of elektrische wijze
verwijderd worden, zodat enkel de celinhoud en het celmembraan overblijven. Dit geeft een
uniek eencellig systeem (Lascaux, 2002; Reddy et al., 2007).
2.5.2 De celwand
Voor het starten van een protoplastencultuur is het nodig om te weten waaruit de celwand van
Ulva lactuca is opgebouwd (figuur 5). De celwand kan immers maar afgebroken worden in-
dien de juiste enzymen gebruikt worden voor de verschillende celwandcomponenten.
Figuur 5: De celstructuur van Ulva sp. (Sperber, 2010)
Bij A is er een schematisch doorzicht van de sectie te zien, bij
B is er een associatie voorgesteld tussen de verschillende celwand polysacchariden

15
Figuur 5 geeft de celstructuur weer van Ulva sp. Deze is verschillend van landplanten. De
celwand bestaat voornamelijk uit suikers. Vanwege de moeilijke omgevingsomstandigheden
waarin zeewieren in de natuur voorkomen bevat de zeewiercelwand veel beschermende com-
ponenten. Dit zijn fenolische componenten en grote hoeveelheden van polyanionische slijm-
laag. Ulva lactuca bevat ulvan, dit bevindt zich tussen de twee lagen waaruit het thallus be-
staat. De celwand bevat ook glucuronen. Deze bestanddelen zitten dicht bij het cytoplasma-
membraan, aan de buitenkant van de aanliggende zeewiercellen. De xylose en glucose bevat-
tende polysachariden vormen gestapelde lagen rondom de cellen (Van Ginneken et al., 2013).
2.5.2.1 Polysachariden
De polysachariden bestaan uit een water oplosbaar gedeelte (ulvan) en een in water onoplos-
baar gedeelte. Ulvan kan je karakteriseren als glucuronorhamnoxyloglycan. Het in water on-
oplosbaar gedeelte kan je indelen in een alkalisch oplosbaar gedeelte namelijk β-1,4-
glucoxylan en β-1,4-glucoranan, en een niet in basen oplosbaar gedeelte namelijk cellulose en
β-&,4 xyloglucans (Sperber, 2010).
2.5.2.2 Aminozuren en eiwitten
Ulva lactuca bevat veel hoogwaardige essentiële aminozuren en deze komen voor in dezelfde
concentraties als bijvoorbeeld in kikkererwten (Van Ginneken et al., 2013). Daarom kan zee-
wier ook gebruikt worden als vleesvervanger.
2.5.2.3 Lipiden
Zeesla bevat ook essentiële vetten waarvan de westerse mens denkt ze enkel in vis en noten te
vinden zijn. In veel literatuur wordt hiermee geen rekening gehouden als het aankomt op het
oplossen van de celwand. Waarschijnlijk omdat deze vetten zich vooral in de cellen bevinden
en niet zo zeer deel uitmaken van de celwand.
2.5.3 Enzymen
Om protoplasten te vormen is het nodig om de celwand af te breken. Zoals hierboven aange-
geven bestaat de celwand van Ulva lactuca voornamelijk uit polysachariden. Om polysacha-
riden af te breken heb je cellulase enzymen nodig (Reddy et al., 2007).
Uppalopati & Fujita (2002), Uchida, Yoshikawa, Ishida, & Saga (1992), Reddy C., Gupta,
Mantri, & Jha (2007) en Reddy & Fujita (1991) gebruiken Cellulase Onozuka R-10. Deze
enzymen zijn afkomstig van de schimmel Trichoderma viride. Deze cellulase is een multi-

16
enzymatisch systeem dat bestaat uit cellulase, α-amylase, hemicellulase, pectinase en protease
activiteit. Deze enzymen modificeren celluloses, lichines en graan β-D-glucans.
De modificatie gebeurt door de hydrolyze van 1,4-β-D glucosidische bindingen (amylase) en
het breken van 1,4-α-D-galastosiduronic bindingen (pectinase).
Cellulase onozuka R-10 bevat 1U/mg van cellulase activiteit, 1U/mg protease activiteit,
0,4U/mg pectinase activiteit en 0,01 U/mg protease activiteit. Die 1U/mg betekent dat één
eenheid cellulase 1µM van glucose van carboxymethyl cellulose zal vrijgeven (laboratories,
2011).
De meest algemene bron van niet-cellulitische zeewiercelwand-degraderende enzymen zijn
herbivore marine invertebraten (zeedieren die van algen leven) en marinebacteriën (Reddy et
al. ,2007). Deze hebben in de loop van evolutie efficiënte systemen gevonden om de celwand
van deze zeewieren af te breken. Abalone aceton poeder is gemaakt van abalone, een zeeslak.
De enzymen bevatten β-glucoronidase en sulfaten (Van Ginneken et al., 2013). Extracten van
digestieve enzymen van herbivore organismen bevatten echter potentieel schadelijke substan-
ties voor protoplasten. Het beste worden lage concentraties en korte incubatietijden gebruikt.
Ook het toevoegen van protease inhibitoren tijdens isolatie is mogelijk (Björk et al.,1992).
Tabel 2 op volgende pagina toont de vergelijking tussen de gegevens van een aantal artikels
over de vorming van protoplasten. De meeste artikels zijn gelijklopend qua enzymensamen-
stelling, temperatuur, pH en incubatieduur. Bij landplanten duurt het duidelijk langer om pro-
toplasten te vormen.

17
2.5.4 Vergelijkende literatuurstudie
Tabel 2: Samenvatting van de verschillende manieren om protoplasten te vormen (Björk et al. ,1992; Deryckere et al.,2012; Reddy & Fujita, 1991; Reddy et al. ,2007; Uchida et al. , 1992; Uppalopati &
Fujita, 2002; Van Ginneken et al. , 2013)
methode type
wier/plant
pH buf-
fer
osmotische
waarde
enzymen oplosmid-
del
incuba-
tie duur
zee-
wier
(mg)
oplosmid-
del (ml)
Tempera-
tuur (°C)
rotatie extra
Björk et
al (1992)
Ulva rigida 7 20m
M
He-
pes
0,4M manitol 1,5% abalone ace-
tone poeder, 1,5%
cellulysin
zeewater tot cel
digestie
100 10 20 60 rpm 30min met een
0,8M oplos-
sing behande-
len geeft extra
opbrengst.
Derycke-
re et al
(2012)
Cichorium 5,5 0,5M mannitol 1mg cellulase cay-
lase 345, 0,5mg
pectinase caylase
M2 (per ml)
miliQ wa-
ter
16u ? ? 23 25 rpm in het donker
Reddy
and Fuji-
ta (1991)
Entero-
morpha
6 50
mM
MES
0,6M Mannitol
0,5% kaliumdex-
transulfaat
3% cellulase RS,
2% top shell ace-
tone poeder
gefilterd
zeewater
3u 50 5 20 15-20
rpm
donker
Reddy et
al.
(2008)
all seaweeds 5 -
6
? 0,4M osmocicum 2% cellulase
onozuka R-10 2%
abalone acetone
poeder, 1% agarase
50U, 1% protease
1%NaCl in
H2O
? ? ? 20 - 25 schudden in
het donker
Uchida
et al.
(1991)
Ulva per-
tusa
5,5 50m
M
MES
1,2M D sorbitol 5% abalone ace-
tone poeder, 2%
cellulase onozuka
R10, 0,1%
macerozyme R-200
PES medi-
um
1u in-
ductie
5u en-
zymen
250 2,5 20 30 rpm
Uppala-
pati &
Fujita
(2002)
Ulva per-
tusa
6,5 50m
M
MES
kalium dextran
sulfaat 0,5%
0,6M mannitol
2% cellulase Ono-
zuka R-10, 2%
Macerozyme R-10
3% NaCl,
1mM CaCl
2- 2,5 ? 5 22 15-20
rpm
donker
Van
Ginne-
ken et al
(2013)
Ulva lactu-
ca
6 MES Mannitol, CaCl2 1,5% abalone,
1,5% cellysine
zeewater 30 min.-
1u
? ? 20 schud-
den
?

18
Vooral de osmotische waarde van de oplossing varieerde tussen de verschillende artikels
(zie Tabel 2: Samenvatting van de verschillende manieren om protoplasten te vormen). Het
aantal mol mannitol of sorbitol bepaalt de osmotische waarde. Door het verlies van de cel-
wand zijn de cellen niet meer bestand tegen hoge verschillen in omgevingsconcentratie aan
osmotisch actieve stoffen (molariteit)
Figuur 6: Turgor druk in plantencellen (wikipedia)
Indien de omgevingsconcentratie van deeltjes te laag is zal de protoplastcel veel water op-
nemen en uiteindelijk exploderen. Als er te veel deeltjes opgelost zijn zal de protoplast in-
wendig water verliezen en kan de cel imploderen.
Björk, Gómez-Pinchetti, Reina, & Pedersén (1992) en Reddy et al (2008) gebruikten 0,4M
mannitol als osmoticum. 0,5M mannitol was populair bij Deryckere et al (2012) en Uppala-
pati & Fujita (2002). Reddy and Fujita (1991) gebruikten 0,6M mannitol. Uchida et al.
(1991) was de enige die sorbitol gebruikte als osmoticum.
De pH was in alle artikels tussen de 5 en de 7 en bijna overal werd ook 50mM MES (2-(N-
morpholino) ethane sulphonic acid) gebruikt als buffer. De incubatieduur was in de meeste
publicaties over het algemeen vrij kort. Het is duidelijk dat het verwijderen van een land-
plantcelwand meer tijd vraagt (Dereyckere et al., 2012).
Ook de temperatuur (20°C) en het zachtjes schudden in het donker kwam algemeen voor.
Het zoutgehalte van het water kon variëren van 1% NaCl (Reddy et al., 2007) tot 3,2% (echt
zeewater) (Björk et al., 1992; Van Ginneken et al., 2011; Reddy & Fujita, 1991).

19
3 Algemene materialen en methoden
3.1 Herkomst Ulva lactuca
De Ulva lactuca was afkomstig uit de proefhoeve van Wageningen Universiteit in de
Schelphoek (Oosterschelde), coördinaten (40' 55'' N, 3° 49' 44'' E). Hier wordt de Ulva ge-
teeld aan touwen die aan pontons in het zeewater hangen. Er is ook materiaal gebruikt van
Nergena het zeewierlab van Wageningen Universiteit, waar Ulva gekweekt wordt in M3-
bakken.
Figuur 7: Schematische weergave van de zeewier proefhoeve van Wageningen Universiteit
3.2 Teelt in aquaria
De Ulva lactuca werd verdeeld in twee aquaria met afmetingen 20cm op 20cm op 35cm
(figuur 8). Elk plastic aquarium kan 14l bevatten. Elke maand werd het zeewater ververst en
de aquaria schoongemaakt. Het schoonmaken gebeurde met leidingwater en een sponsje. In
de tussentijdse periode werd ongeveer om de drie dagen het water aangevuld met gedestil-
leerd water om de verdampte hoeveelheid te compenseren. Om de twee weken werd er
vloeibare plantenvoeding toegevoegd (indien niet ververst). De vloeibare blad- en bodem-
bemesting waren van het merk Wulax met een NPK samenstelling van 8-8-6. Er werd 1ml
vloeibare voeding toegevoegd per 10l zeewater. Er werd continu lucht in de aquaria gebla-
zen met behulp van een klassiek aquariumpompje met bijhorend steentje (Crawfish airpump
3500 (2x144L/H)).
Figuur 8: Aquaria met Ulva lactuca

20
3.3 Voedingsbodems
Alle voedingsbodems in dit eindwerk zijn gemaakt op basis Murashige en Skoog medium
(MS). MS bestaat uit: NH4NO3 1,650 mg/l, CaCl2.2H2O 440 mg/l, MgSO4.7H2O 370 mg/l,
KH2PO4 170 mg/l, KNO3 1,900 mg/l, H3BO3 6.2 mg/l, CoCl2 6H2O 0.025 mg/l, Cu-
SO4.5H2O 0.025 mg/l, FeSO4.7H2O 27.8 mg/l, MnSO4.4H2O 22.3 mg/l, KI 0.83 mg/l,
Na2MoO4.2H2O 0.25 mg/l, ZnSO4.7H2O 8.6 mg/l, Na2EDTA.2H2O 37.2 mg/l, Inositol 100
mg/l, Niacin 0.5 mg/l, Pyridoxine HCl 0.5 mg/l, Thiamine HCl 0.1 mg/l, IAA 1–30 mg/l,
Kinetin 0.04–10 mg/l, Glycine 2.0 mg/l, Edamine S 1.0 g/l en agar-agar 10 g/l.
Er zijn twee verschillende vaste voedingsbodems gemaakt en één type vloeibare bodem.
3.3.1 Vast medium met zeewater (VMA)
Medium (voor 1l):
1025ml gefiltreerd Noordzeewater;
4,405g Murashige en Skoog medium (Duchelfa);
7g agar;
30g Sucrose;
pH 8 door eventueel correctie met KOH.
Het water, MS en de suiker werd in een fles gedaan. De pH werd gecontroleerd (Hanna in-
struments HI 2550) en aangepast. Daarna kon het agar toegevoegd worden. De volledige fles
werd gesteriliseerd in de autoclaaf (130°C bij 1atm gedurende 15 tot 30 min).
3.3.2 Vast medium met zout water (VMB)
Medium (voor 1l):
32g NaCl;
1025ml miliQ water;
4,405g Murashige en Skoog medium (Duchefa);
7g agar;
30g Sucrose;
pH 8 door eventueel correctie met KOH.
Het water, MS en de suiker werd in een duranfles gedaan. De pH werd gecontroleerd (Han-
na instruments HI 2550) en aangepast. Daarna kon het agar toegevoegd worden. De volledi-
ge fles werd gesteriliseerd in de autoclaaf (130°C bij 1atm overdruk gedurende 15 minuten).
3.3.3 Vloeibaar medium (VlM)
De samenstelling van het VIM is hetzelfde als het vast medium met zeewater (VMA) maar
dan zonder het toevoegen van de agar.

21
3.4 Microscopen
In dit onderzoek werd een gewone lichtmicroscoop van het merk Olympus UTVX (Japan)
gebruikt. Deze microscoop was voorzien van een camera gekoppeld aan een computer. Er
werd ook een omkeermicroscoop gebruikt, een Nikon Eclipse2000S. Er werden objectieven
van 10x, 20x en 40x vergroting gebruikt.
3.5 Groei- of proefomstandigheden
De aquaria, petriplanten en falconbuizen met zeewierweefsel werden allen onder gecontro-
leerde klimatologische omstandigheden gehouden door ze in een in vitro groeikamer of een
fytotron te plaatsen. Hierdoor was het mogelijk ze altijd onder ‘lente’ condities te houden.
De verschillende fytotrons gebruikt in dit experiment hadden temperaturen van 15°C, 20°C,
23°C of 26°C. Er was altijd een dag-nacht ritme van 16u licht en 8u donker. De lichtintensi-
teit was afhankelijk van de afstand tot de lampen en het aantal lampen. In alle fytotrons wer-
den lampen gebruikt van het type TL-D Philips 38W/840. De lichtintensiteit werd gemeten
met Photosynthetic Active Radiation (PAR). Dit is het aantal μM fotonen per vierkante me-
ter en per seconde. De PAR werd gemeten met een PAR meter (Jaz lichtmeter van Ocean
Optics).
Voor wat betreft de zuurtegraad kan men stellen dat de pH in de zee (natuurlijke groeiom-
standigheid van zeewier) vrij hoog is, rond pH 8. De pH van een medium beïnvloedt de op-
name van heel wat componenten en reguleert heel wat biochemische reacties. Tijdens de
cultuur veranderde de pH door de selectieve opname en uitwisseling van ionen door de
plant.
Ook het zoutgehalte is belangrijk bij het telen van zeewier. Door verdamping van water
stijgt de zoutconcentratie tijdens de cultuur. Bij vaste bodems echter, is deze verdamping
minimaal.

22
4 Experimenteel gedeelte
4.1 Initiatie in vitro op agarmedium
4.1.1 Doel
Het doel is om Ulva lactuca in steriele omstandigheden op een kunstmatig medium te telen.
Om dit te realiseren moet het zeewier steriel gemaakt worden. De bestaande literatuur over
het in vitro telen van zeewier is beperkt, daarom is het nuttig terug te grijpen naar informatie
over het in vitro telen van zaadplanten. Heel wat landplanten zijn in vitro te telen op Muras-
hige and Skoog medium (MS) daarom werd dit medium ook voor Ulva lactuca gebruikt.
4.1.2 Vaste voedingsbodems
4.1.2.1 Inleiding
Omdat condities gemakkelijk te bestuderen zijn op een vast medium en Ulva lactuca een
zeewier is dat tegen een periodische droogte kan, leek het testen op vaste voedingsbodems
een goed begin. Eerst werd de standaard in vitro procedure voor landplanten gevolgd. Alle
proeven werden steriel uitgevoerd in een laminaire airflow.
4.1.2.2 0,7cm diameter, 10% javeloplossing
4.1.2.2.1 Inleiding
Door gebrek aan kennis over het in vitro initiëren van zeewieren werd de standaard land-
plant methode getest (Werbrouck, 2011).
4.1.2.2.2 Specifieke materialen en methoden
Het wier moest gesteriliseerd worden voor het in vitro geïnitieerd werd. Deze sterilisatie
gebeurde door de gewenste plantendelen eerst even heel kort met 70% ethanol te spoelen
dan een kwartier in 10% javel oplossing. Na het spoelen met javel werd er drie keer met
steriel water gespoeld. (Werbrouck, 2011)
4.1.2.2.3 Resultaat en bespreking
Na 1 minuut in de 10% javel oplossing veranderde het zeewier al van kleur (Figuur 9). De
concentratie van 10% javel bleek dodelijk voor het wier. Na opname in de cellen stierf het
af.

23
Figuur 9: Afgestorven, wit zeewier na één minuut in 10% javel oplossing
4.1.2.3 0,7cm diameter, 1% javeloplossing
4.1.2.3.1 Inleiding
Omdat sterilisatie met javel moeilijk bleek, werd de concentratie van de vorige proef (zie
4.1.2.2) verlaagd tot een minimale concentratie om nog in staat te zijn contaminanten de
doden.
4.1.2.3.2 Specifieke materialen en methoden
Eerst werd het zeewier een minimaal aantal seconden in
ethanol gedompeld, dan 10 minuten in een 1% javelop-
lossing. Na 10 minuten spoelen van het zeewier in een
1% javeloplossing werd er met steriel zeewater gespoeld.
Er werd dan thallus in schijfjes gesneden met behulp van
een steriele kurkboor met diameter 0,7cm. De nog steeds
groen uitziende thallusschijfjes (Figuur 10) werden dan
op VMB gelegd en gecultiveerd in de klimaatkamer bij
15°C en 60µmol fotonen m-2s. Ook de platen, mesjes,
pipetten, pincetten, kurkboren e.d. waren steriel bij ge-
bruik. Deze waren steriel aangekocht of steriel gemaakt
via autoklaveren (130°C bij 1atm gedurende 15 tot 30
min.) of met behulp van een parelsterilisator.
Figuur 10: Gespoelde Ulva lac-
tuca met 1% NaClO (javel)
4.1.2.3.1 Resultaat en bespreking
Alle thallus-schijfjes waren dood binnen de vier dagen. Er was wel geen contaminatie zicht-
baar. Vermoedelijk tast de NaClO de cellen van het ultradunne thallus aan door de oxide-
rende werking van chloor en zuurstof. Pas na enkele dagen worden de gevolgen hiervan dui-
delijk.

24
4.1.2.4 0,7cm diameter, gespoeld met steriel zeewater met verschil-lende hormoonconcentraties
4.1.2.4.1 Inleiding
De invloed van het groeihormoon 6-Benzyladenine (BA) werd getest. Dit cytokinine stimu-
leert de groei en celdeling van landplanten en het kan levensduur van de plant verlengen
(Werbrouck, 2011; Van Labeke, 2013). Het was daarom nuttig om de invloed van BA bij
zeewieren te bestuderen. Het steriliseren met javel had tot nu toe nog geen succes gehad,
daarom leek het interessant om het zeewier te laten groeien zonder het op voorhand te steri-
liseren, maar gewoon te spoelen met steriel zeewater. Het zou immers kunnen dat de zout-
concentratie in de voedingsbodem hoog genoeg was om de meeste bacteriën en schimmels
te onderdrukken.
4.1.2.4.2 Specifieke materialen en methoden
Schijfjes van 0,7cm diameter zeewier werden uit het midden van het thallus gesneden met
een kurkboor. Deze werden grondig gespoeld met steriel zeewater en op VMB gelegd. De
concentraties 0µM BA, 0,1µm BA, 1µM BA en 10µM BA werden getest met elk nog een
controle plaat. Ook deze platen werden bij 15°C en 60µmol fotonen m-2
s gezet.
4.1.2.4.3 Resultaat en bespreking
De eerste drie dagen bleven de thallusschijfjes groen, maar vanaf dag vier kregen de meeste
contaminanten de overhand. Alle zeewierschijfjes waren na een week al gecontamineerd. De
verschillende concentraties aan BA leken hier helemaal geen invloed uit te oefenen. Op fi-
guur 11 is te zien dat er vloeistof rond het thallus aanwezig was voordat het wier stierf. En-
kele dagen later was deze vloeistof gecontamineerd.
Figuur 11: Links onidentificeerbare doorzichtige vloeistof rond uitgesneden schijfjes,
rechts een duidelijke schimmel, bacterie en/of gist contaminatie

25
4.1.2.5 0,7cm diameter, 1% javel oplossing overgoten met 5ml steriel zoutwater
4.1.2.5.1 Inleiding
Het uitdrogen bleek toch een groter probleem te zijn dan aanvankelijk gedacht. Daarom
werd er geopteerd om steriel zoutwater op de voedingsbodem te gieten en zo het weefsel
vochtig te houden. Bij alle voorgaande proeven leek het zeewier veel water te verliezen na
verloop van tijd. Misschien was dit wel de oorzaak van sterfte. In de VMB zouden mogelijks
essentiële elementen kunnen ontbreken daarom werd in deze voedingsbodem steriel zeewa-
ter gebruikt in plaats van miliQ water met zout.
4.1.2.5.2 Specifieke materialen en methoden
De opzet van de proef was gelijklopend als proef 4.1.2.3. Het zeewier werd met 1% javel
gespoeld en nadien in schijfjes van 0,7cm gesneden in een laminaire airflow. De schijfjes
werden op petriplaten met 20ml VMA gelegd. Er werd 5ml steriel zoutwater (16g NaCl in
500ml water) over de geënte plaat gegoten om uitdroging van het zeewierschijfje tegen te
gaan.
4.1.2.5.3 Resultaat en bespreking
De opname van NaClO in de cellen bleek toch de doodsoorzaak te zijn, want de schijfjes
zeewier stierven al na twee dagen. Als zeewier steriel gemaakt wordt met NaClO sterft het
af. De controle, die niet gesteriliseerd werd, was binnen de week gecontamineerd en stierf
uiteindelijk.
4.1.2.6 2cm diameter, 1% javeloplossing
4.1.2.6.1 Inleiding
Misschien was het inoculum bij de vorige proeven te klein. Het kon zijn dat het zeewier niet
groeide omdat er te weinig of zelfs geen meristeemcellen aanwezig waren in het thallus-
schijfje. Bij landplanten wordt er meestal ook meristematisch weefsel geënt zoals bijvoor-
beeld een stengeltje met okselknop. Uit voorgaande literatuur bleek dat Ulva lactuca het
meest groeikrachtig was in het midden van het thallus (Sluis, 2012). Om dit te testen werden
grotere schijfjes zeewier uit het midden van het thallus gesneden met een kurkboor.

26
4.1.2.6.2 Specifieke materialen en methoden
Het VMA werd steriel in platen gegoten in een laminaire airflow zoals bij alle vorige proe-
ven. Het Ulva lactuca thallus werd tien minuten voorgespoeld met 1% javel oplossing om
het steriel te maken. Maar nu had de kurkboor een diameter van 2cm. Er werden schijfjes uit
het midden van het thallus gesneden.
Figuur 12: De grootte van het stukje thallus heeft geen invloed op de overleving op
een voedingsbodem
4.1.2.6.3 Resultaat en bespreking
Het gebruik van grotere thallusschijfjes gaf geen verschil. Ook deze gingen ook al na twee
dagen dood (zie Figuur 12). De grootte van het inoculum maakt dus niet uit. Een mogelijke
verklaring is dat het groeipunt van Ulva lactuca toch niet in het midden van het thallus ligt.
4.1.2.7 2cm diameter, 5 keer spoelen met steriel zeewater
4.1.2.7.1 Inleiding
Om zeker te zijn dat het sterven van het zeewier in vorige proeven niet aan de opname van
NaClO in de cellen lag, werd deze proef herhaald maar zonder gebruik van javel. De veron-
derstelling was dat als er vaak genoeg gespoeld werd met steriel zeewater de concentratie
aan bacteriën, schimmels en gisten zou verkleind worden.
4.1.2.7.2 Specifieke materialen en methoden
Er werd dus een cirkel met 2cm diameter uit het midden van het thallusoppervlak gesneden.
Deze werden nadien vijf keer gespoeld met steriel zeewater in een laminaire airflow. Na de
spoeling werden de schijfjes op een mediumplaat gelegd met 20ml VMA.
4.1.2.7.3 Resultaat en bespreking
Deze hypothese bleek ook fout. Al na een viertal dagen was er contaminatie waar te nemen.
Spoelen met steriel zeewater spoelt de contaminatie niet weg.

27
4.1.2.8 Toevoegen van Proclin 150
4.1.2.8.1 Inleiding
Als de zeesla zelf niet steriel te krijgen was, was het misschien mogelijk om de voedingsbo-
dem zelf ongevoelig te maken voor contaminatie. Hiervoor werd gebruik gemaakt van Pro-
clin 150, een commercieel product om voedingsbodems steriel te maken en te houden. De
samenstelling van het product is 5-chloro-2-methyl-4-isothiazolin-3-one, 2-methyl-4-
isothiazolin-3-one, magnesiumchloride, magnesiumnitraat en water (Supelco, 2013).
4.1.2.8.2 Specifieke materialen en methoden
Aan de vaste voedingsbodem werd 0; 1,5; 1 en 10µM BA gecombineerd met 0; 5; 10 en
20µl Proclin 150. Beide werden in de plaat gepipetteerd met behulp van micropipet en ste-
riele filtertip. Daarna werd er 20ml medium in de petriplaat gegoten en moest er gewacht
worden tot de voedingsbodems gestold waren voor de thallus schijfjes, gemaakt met de ste-
riele kurkboor met diameter 0,7cm, op het medium gelegd werden.
4.1.2.8.3 Resultaat en bespreking
Figuur 13 toont enkele foto’s drie dagen na het maken van de platen. De platen zonder pro-
clin waren dood en gecontamineerd. Die met proclin leven nog nauwelijks. Zes dagen na het
maken van de platen zijn ze allemaal dood. De platen vanaf 5µl proclin zijn niet gecontami-
neerd. Enige invloed van BA is niet waarneembaar.
Figuur 13: Links bij de start van het experiment,
midden en rechts dode en gecontamineerde thallus schijfjes

28
4.1.2.9 Toevoegen van antibiotica
4.1.2.9.1 Inleiding
Een alternatief om een voedingsbodem steriel te houden is het toevoegen van antibiotica
(AB).
4.1.2.9.2 Specifieke materialen en methoden
Er werden platen gemaakt die verschillende soorten AB bevatten in verschillende concentra-
ties. De gebruikte concentraties zijn gebaseerd op toepassingen bij gewone zaadplanten
(Werbrouck, mondelinge mededeling, 2013). Om AB te besparen werden de tests in kleine
petriplaatjes uitgevoerd met maar 4ml medium. Er werd eerst steriel een druppel AB in de
plaat gebracht, respectievelijk 0, 2, 4, 6 en 8µl AB waarna het medium over de petriplaat
gegoten werd.
Er werden vijf soorten AB getest in de hierboven genoemde concentraties. De AB die getest
werden waren in volgende stockoplossingen beschikbaar: vancomycine 50mg/ml, stepromy-
cin 300mg/ml, cefotaxime125mg/ml, kananycin 50mg/ml, Ampiciline 100mg/ml. Op elk
plaatje werden twee cirkeltjes gelegd met diameter van 0,9cm. Het thallus werd op voorhand
gespoeld met steriel zeewater.
De platen met 2, 4 en 6µl cefotaxime 125mg/ml en de platen met vancomycine 50mg/ml
waren niet gecontamineerd en bleven zelfs nog een vijftal dagen groen.
In een tweede proefopstelling werd dan enkel met deze twee antibiotica gewerkt. Er werd
telkens een combinatie gemaakt van het vancomycine en het cetotaxime zoals beschreven in
Tabel 3. In deze opstelling werden ook twee thallus schijfjes per plaat gebruikt. De platen
werden in de fytotron bij 15°C geïncubeerd bij 60µmol fotonen m-2
s-1
.
Tabel 3: Tweede proefopstelling met antibiotica, de nummers in de tabel zijn de nummers van de platen
0 µl cef** 2 µl cef 4 µl cef 8 µl cef
0 µl van* 1 2 3 4
2 µl van 5 6 7 8
4 µl van 9 10 11 12
8 µl van 13 13 15 16
* van = vancomycine; ** cef = cefotaxime

29
4.1.2.9.3 Resultaat en bespreking
Bij de eerste antibiotica proef bleken cefotaxime en vancomycine een positieve invloed te
hebben op het overleven van de zeesla. Na een test met de combinatie van verschillende
concentratie cefotaxime 125mg/ml en cancomycine 50mg/ml bleek echter dat een te hoge
concentratie AB dodelijk was en zelfs niet in staat was om contaminatie tegen te gaan. Want
bij proef 2 waren na twee dagen de meeste petriplaten al beschimmeld. Met uitzondering
van plaatjes met nummers: 3, 4, 7, 8 en 15 die niet gecontamineerd waren (zie Tabel 3). Het
wier was zelfs nog een klein beetje groen. Na een week was het gecontamineerde weefsel
dood (wit). Pas na twee weken bleken ook de platen met nummers: 3, 4, 7 en 8 gecontami-
neerd (zie Figuur 14 ). Dus de antibiotica waren niet sterk genoeg om contaminatie tegen te
gaan. Een hoge concentratie van cefotaxime met een lage concentratie vancomycine bleek in
deze proef het beste resultaat te bekomen.
Figuur 14: Resultaat antibioticaproef na 1 week:
plaat 9 is gecontamineerd, het weefsel in plaat 7 daarentegen leeft nog en is niet gecontamineerd
4.1.2.10 Bespreking vaste voedingsbodem
Zeewier steriel krijgen is problematisch. Als het zeewier niet steriel is kan het ook niet in
vitro geteeld worden. Steriliseren met 10% javeloplossing betekende de dood voor het weef-
sel al na één minuut. Met 1% javel oplossing blijft het weefsel steriel maar is het ook binnen
de week na steriliseren dood. De grootte van het inoculum maakte geen verschil. Het toe-
voegen van het groeihormoon 6-Benzyladenine had eveneens geen invloed op de lengte van
de levensduur.
Vaste voedingsbodems bleken niet geschikt voor zeewier, vermoedelijk door een te lage
vochtigheidsheidsgraad van deze bodem. Maar zelfs na toevoegen van een laagje zout water
gaat het stukje weefsel toch dood. Door het toevoegen van proclin of antibiotica aan de voe-
dingsbodem blijven de thallusschijfjes wel iets langer leven, maar niet voldoende lang. Het
is niet mogelijk om via de standaardmethode voor landplanten zeewier in vitro te telen op
een vaste voedingsbodem. Sterfte kan te wijten zijn aan het gebrek aan symbiotische orga-
nismen, aan uitdroging of aan concurrentie met schadelijke organismen.

30
4.1.3 Proeven in vloeibare voedingsbodems
4.1.3.1 Inleiding
Ulva lactuca is een zeewier dat continu in de zee leeft. Uit voorgaande proeven leek vaste
bodemcultuur geen succes. Door de thallusschijfjes in een vloeibaar medium te steken zou
dit probleem eventueel opgelost kunnen worden. Vloeibare voedingsbodem is dezelfde MS
bodem als bij de vaste voedingsbodems maar dan zonder het toevoegen van de agar (zie
3.3.3). Om zuurstof toe te voegen, werden de buisjes met medium in een schudtoestel ge-
plaatst.
4.1.3.2 2cm diameter, 1% javeloplossing
4.1.3.2.1 Inleiding
De uitgevoerde procedure is dezelfde als bij de proef met vaste voedingsbodem en 1% javel
(zie 4.1.2.3) maar in dit geval met vloeibaar medium. Bij vaste voedingsbodems zou vocht-
verlies een mogelijke doodsoorzaak kunnen zijn. Dit zou bij vloeibare voedingsbodems geen
probleem mogen zijn.
4.1.3.2.2 Specifieke materialen en methoden
Het vloeibare medium werd in duranflessen van 500ml gemaakt. Nadien werd 20ml van
deze voedingsbodem in steriele falconbuisjes gegoten met behulp van steriele pipetten in
een laminaire airflow. Het thallus werd 10 minuten gespoeld met een 1% javeloplossing,
daarna op steriel papier gelegd en met een gesteriliseerde kurkboor met twee cm diameter
werden er schijfjes uit het thallus gesneden. Deze schijfjes werden met behulp van een ste-
riel pincet in de falconbuis met medium gelegd. Het schudtoestel stond in de fytotron bij
15°C
4.1.3.2.3 Resultaat en bespreking
De zeewierschijfjes stierven snel af door opname van NaClO in de cellen. De mediumoplos-
sing bleef wel steriel.
Figuur 15: Thallusschijfjes in vloeibaar zeewiermedium,
links bij start rechts een week later

31
4.1.3.3 2cm diameter, 5 keer spoelen met steriel zeewater
4.1.3.3.1 Inleiding
Bij voorgaande test bleek het gebruik van NaClO (javel) dodelijk. Er werd getest of de con-
centratie aan contaminanten door spoelen met steriel zeewater kon worden verminderd.
4.1.3.3.2 Specifieke materialen en methoden
Hier gebeurde dezelfde procedure als de vorige. Het enige verschil was dat er vijf keer ge-
spoeld werd met steriel water in plaats van het gebruik van de 1% javeloplossing. De fal-
conbuis werd in een schudder gezet in de klimaatkamer bij 15°C.
4.1.3.3.3 Resultaat en bespreking
Na een week trad er contaminatie op in de vloeibare voedingsbodem en al snel volgde de
dood van de cellen.
4.1.3.4 Afspoelen met Tween (detergent) in steriel zeewater
4.1.3.4.1 Inleiding
In het artikel van Azanza & Aliaza (1999) wordt er besproken dat het zeewier met detergent
werd gereinigd voor initiatie in het medium. Dus werd hier Tween als detergent uitgetest.
4.1.3.4.2 Specifieke materialen en methoden
Voordat de thalluscirkel van twee centimeter diameter in de steriele falconbuis met vloei-
baar medium werd ondergedompeld, werd het zeewier grondig gereinigd in steriel zeewater
waarin detergent was opgelost. Het idee was om het zeewier “proper” te maken. Er werd
ook een controle gemaakt die niet gereinigd was.
4.1.3.4.3 Resultaat en bespreking
Na zeven dagen is het weefsel dood. Of het gecontamineerd was, was moeilijk te zeggen
want het vloeibaar medium was al een beetje troebel van bij de start.
Figuur 16: Thallus gespoeld met steriel zeewater en Tween,
links na dag 1 - rechts na 6 dagen

32
4.1.3.5 Verschillende tijdspannes in 70% ethanoloplossing
4.1.3.5.1 Inleiding
Het artikels van Lascaux (2002) vermeldt het steriel krijgen van weefsel via ethanol. Daar-
om leek dit wel eens het proberen waard.
4.1.3.5.2 Specifieke materialen en methoden
Hier werden de thallusschijfjes respectievelijk 0, 1, 5, 10 en 15 minuten in 70% ethanol ge-
houden om ze daarna af te spoelen in steriel zeewater en dan in een falcon buis te brengen
met 20ml vloeibaar medium. Alles werd uitgevoerd in een laminaire airflow.
4.1.3.5.3 Resultaat en bespreking
Het weefsel dat één minuut in de ethanol gezeten had, heeft het drie dagen overleefd. De
andere thallusschijfjes waren allemaal na twee dagen dood. De ethanol werd door de cellen
opgenomen en dat kan mogelijkerwijze de cellen van binnenuit aangetast hebben. Opvallend
was dat de controle ook al na twee dagen dood was. Dus misschien was een tekort aan zuur-
stof de echte doodsoorzaak.
4.1.3.6 Pasteurisatie gewoon wier
4.1.3.6.1 Inleiding
Bij voedingswaren zoals zuivelproducten, wordt de techniek van de pasteurisatie gebruikt.
Pasteurisatie is een proces waarbij schadelijke organismen vernietigd worden door het weef-
sel gedurende een korte periode aan hoge temperatuur bloot te stellen. Dit zou ervoor zorgen
dat de bacteriën gedood worden maar het weefsel (eiwitten, suikers, …) gevrijwaard blijft.
4.1.3.6.2 Specifieke materialen en methoden
De Ulva lactuca werd 15s in zeewater van 72°C gelegd. Nadien in VLM gestoken in een
laminaire airflow met een steriel pincet. Er was ook een controle met gewoon zeewater ge-
maakt.
4.1.3.6.3 Resultaat en bespreking
Al na twee dagen lijkt het stukje thallus dat gepasteuriseerd was, bleek te worden. Na vier
dagen was het stukje thallus gestorven. De zeesla kan waarschijnlijk niet tegen hitte, vermits
de optimale groei temperaturen tussen de 10°C en de 25°C liggen. De controle was na een
week gecontamineerd en wit van kleur.

33
4.1.3.7 Gassterilisatie
4.1.3.7.1 Inleiding
Een andere methode om planten steriel te krijgen is met behulp van gassterilisatie. In het
Vlaams Instituut voor Biotechnologie (VIB) steriliseren ze op deze wijze zaadjes voor in
vitro teelt en ook sporen van mossen.
4.1.3.7.2 Specifieke materialen en methoden
De gassterilisatie vond plaats in een glazen stolp onder een trekkast. In een beker onder een
rekje werd 100ml bleekwater van 8% samengevoegd met 5ml van 38% HCl. Door de men-
ging van de vloeistoffen komt er een chloordamp vrij die alle schadelijke micro-organismen
zou doden. Er werd telkens een droog thallusschijfje en een thallusschijfje in zeewater op
het rekje in de glazen stolp gezet. Er werd ook een controle in gewone lucht gehouden. Ver-
schillende tijdspannes werden getest. 5 minuten, 15 minuten, en 1 uur.
Voor het weefsel in een steriele omgeving in het vloeibaar medium werd gebracht, werd het
eerst afgespoeld met steriel zeewater om alle schadelijke chloor af te wassen.
Figuur 17: Glazen stolp in trekkast waarin de gassterilisatie werd uitgevoerd
4.1.3.7.3 Resultaat en bespreking
Na 15 minuten in de gassterilisator was de droge methode al dood. De natte methode leefde
nog, waarschijnlijk omdat het gas meer tijd nodig had om door te dringen door het water tot
aan het weefsel. Na drie dagen waren alle droge methoden dood.
De controles en de natte methodes waren nog groen. Maar wel lichtgroen of groen met witte
vlekken. Na vijf dagen waren de controles ook bruin-wit geworden. Van de vijf minuten
natte methode is na vijf dagen nog maar 10% groen. Bij de vijftien minuten nat zijn de ran-
den wel wit geworden maar het midden gedeelte ziet nog donkergroen. Tien dagen later is
de vijftien minuten methode nog steeds groen (Figuur 18).

34
Figuur 18: 15 minuten natte gassterilisatie na tien dagen.
Alle andere methoden en controles waren na tien dagen al dood. Na zestien dagen begon de
oplossing er troebel uit te zien en het weefsel begon uit elkaar te vallen. Dit kan mogelijk
door een tekort aan nutriënten zijn. Als het medium ververst zou geweest zijn, had het stukje
thallus misschien nog langer overleefd. Het is wel zo dat enkel de controles gecontamineerd
waren (witte en bruine substanties in het medium). Al de gasmethodes bleven steriel.
4.1.3.8 Antibiotica
4.1.3.8.1 Inleiding
De methode met het toevoegen van antibiotica aan de voedingsbodem (zie 4.1.2.9) werd hier
herhaald, maar dan in vloeibaar medium. Misschien was de oorzaak van de sterfte toen uit-
droging en kan dit bij deze methode vermeden worden.
4.1.3.8.2 Specifieke materialen en methoden
Om uitdroging te voorkomen werd de gewenste concentratie antibiotica gemengd met de
juiste hoeveelheid VLM in een steriele proefbuis zoals weergegeven in Tabel 3. Voor de
thallusschijfjes (gesneden met een steriele kurkboor) in het medium gestoken werden, wer-
den ze eerst gespoeld met steriel zeewater. De steriele proefbuizen werden op een schudder
gezet om zuurstof in het medium te brengen. De incubatie vond plaats in de klimaatkamer
bij 15°C.

35
4.1.3.8.1 Resultaat en bespreking
De thallusschijfjes bleven wel langer leven in vloeibaar medium dan op vast medium, maar
na negen dagen waren alle proefbuisjes besmet. De hoeveelheid antibiotica was dus niet
voldoende.
Figuur 19: Links de start van de antibiotica-proefopzet,
rechts de proef negen dagen later, gecontamineerd door micro organismen.
4.1.3.9 Bespreking vloeibare voedingsbodem
Het groeien van Ulva lactuca in vloeibare voedingsbodems was geen succes. Mogelijke oor-
zaken van sterfte kunnen zijn: contaminatie, opname javel, te lage concentratie zuurstof,
opname ethanol of andere onbekende factoren. Een andere verklaringsgrond kan zijn dat
zeewieren niet overleven zonder mogelijkheid tot symbiose met bepaalde zeeorganismen.
Volgens Wichard & Oertel (2010) leeft zeewier in symbiose met bepaalde zeebacteriën (Ro-
seobacter MS2 en Cyphaga MS6) Ook de website ecomare zegt dat de bacterie YM2-23
nodig is voor de uitgroei van zeesla (Ecomare, 2013). Het feit dat deze bacteriën in de bo-
dem ontbreken zou mogelijk een aanleiding kunnen zijn dat de groei van het zeewier stopt.
(Bij orchideeën die ook in symbiose leven met bepaalde grondbacteriën is de groei in vitro
wel gelukt.) Ook de problematiek van de zuurstoftoevoer is een mogelijke factor. Er moet
een meer geschikte manier van zuurstoftoevoeging zijn dan schudden. Na verloop van tijd
kan ook een tekort aan essentiële nutriënten een rol spelen.

36
4.2 Algemeen besluit in vitro initiatie met vast celweefsel
Het is zeer moeilijk en misschien onmogelijk om met delen van zeesla een in vitro cultuur
op te starten. Ulva lactuca is moeilijk steriel te krijgen. Ulva lactuca heeft bepaalde compo-
nenten nodig uit het zeewater, componenten waar we nu nog geen zicht op hebben. Of we de
testen nu uitvoerden met VMA of VMB, het resultaat bleef hetzelfde: sterfte van het weef-
sel. Als besluit kan gesteld worden dat de precaire specificiteit van het zeeslaweefsel ver-
hindert dat het behandelingen met javel, ethanol, antibiotica, proclin overleeft. Het dunne
weefsel bleek niet bestand te zijn tegen deze producten. Het kon ook niet concurreren tegen
andere micro-organismen. Vanaf een te hoge temperatuur, een te laag vochtgehalte of te
weinig zuurstof stierf het weefsel dan ook af.
4.3 Inductie sporulatie van Ulva lactuca
4.3.1 Doel
Het doel is het vormen van sporen en/of gameten bij Ulva lactuca op een gecontroleerde
manier. Deze sporen kunnen nadien voor tal van toepassingen gebruikt worden voor zowel
rechtstreeks zaaibed of als start bij in vitro groei na sterilisatie. Misschien zijn sporen wel
makkelijker te steriliseren dan gewoon weefsel. Deze sporen werden geïnduceerd met be-
hulp van verschillende methoden beschreven in de literatuur.
4.3.2 Inleiding
Een collega van Marlies Demeulenaere van het VIB maakt sporen van mossen steriel door
gassterilisatie. Dit in tegenstelling Tijana et al. (2007) en Jen-Tung & Weickun (2003) die
hun mossen steriel kregen op de conventionele manier door ze enkele ogenblikken in een
lage concentratie NaClO te houden. Maar voor dit getest kon worden, moesten er eerst spo-
ren en/of gameten gevormd worden. Hier is heel wat literatuur rond te vinden. Elke auteur
gebruikt een andere techniek om sporen of gameten te verkrijgen. Het is wel zo dat in dit
onderzoek er geen duidelijke scheiding gemaakt is tussen het vormen van sporen en het
vormen van gameten. Eigenlijk zouden elk nog eens apart moeten onderzocht worden.

37
4.3.3 Materiaal
Er werd gebruikgemaakt van gefilterd Noordzeewater. De filtratie gebeurde met een ederol
JC Bunzer No 15 (menge 100) filter. Het zeewater bevat 32‰ NaCl en heeft een pH 8. Er
werden twee soorten petriplaten gebruikt met verschillende diameter: 8,4cm en 4cm, en een
hoogte van 1cm. De abiotische parameters staan beschreven in Tabel 4
Tabel 4: Abiotische parameters van de verschillende fytotrons, gebruikt in het experiment
Temperatuur Aantal µmol fotonen m-2
s-1
* Hoeveelheid TL-D lampen
15°C 60 2
20°C 100 4
23°C 60 2
26°C 60 2
*Het aantal fotonen m-2
s-1
werd gewijzigd door de platen dichter bij de lampen te zetten
Er werden koelkasten gebruikt met de temperaturen 4°C of 0°C. De pH en het zoutgehalte
werd gemeten door een NaCl- en pH-meter (Hanna instruments HI 2550)
4.3.4 Methoden en resultaten
4.3.4.1 Inleiding
Al deze methoden tot het vormen van sporen of gameten zijn afkomstig uit de literatuur en
werden elk minstens twee keer getest. Hieronder volgt een samenvatting van de verschillen-
de testen.
4.3.4.2 Methode 1 (Sluis, 2012), (Niesenbaum, 1988)
Methode 1.1 Er werd koud zeewater (2°C) gegoten in een petriplaat en er werd met 5,00g
gecentrifugeerd thallus ( 3min. bij 5000 rpm) in het zeewater gelegd. Dit werd
24u bij 2°C gehouden. Daarna werd het water ververst (terug 2°C) en de plaat
werd dan in een klimaatkamer van 20°C en 100 µmol fotonen m-2
s-1
geïncu-
beerd. Volgens het artikels zou het 3500lux = 44,87 µmol fotonen m-2
s-1
9u
donker 15u licht moeten zijn. (Bij TL 33 (Philips): 1000 lux = 12.82
μmol/s.m2.
Methode 1.2 Voor de tweede opzet werd er ± 5,00g thallus in kleine stukjes gesneden, ge-
centrifugeerd (3min. bij 5000 rpm) om overtollig vocht te verwijderen en.
Deze thallusstukjes werden dan in zeewater van 2°C gelegd. Om de twee da-
gen werd het zeewater ververst (2°C) gedurende twee weken. De incubatie
was bij 15°C en 60 µmol fotonen m-2
s-1
.

38
Methode 1.3 Een derde manier bestond erin om ±5,00g stukjes thallus van 3 á 4 mm te
nemen en deze grondig te spoelen met koud (0°C) zeewater. Centrifugeer na
spoelen (3min. bij 5000 rpm). De gespoelde stukken thallus werden nadien in
een petriplaat met 40 ml zeewater van 20°C gelegd. Er werd in een klimaat-
kamer van 20°C met 100 µmol fotonen m-2
s-1
geïncubeerd.
4.3.4.3 Resultaat methode 1
Bij de eerste en derde methode van Sluis (1.1 en 1.3) bleef het weefsel gewoon groen, hier
en daar wat lichter en wit, maar bij microscopisch onderzoek bleek dit afgestorven weefsel
te zijn.
Bij de tweede methode van Sluis (1.2) had het thallus na achtendertig dagen gesporuleerd.
Het omringende water zag groen (zie Figuur 20). Onder de microscoop waren er ovale ga-
meten waar te nemen met twee flagellen. Bij het openen van de petriplaat kwam er stank
vrij. Dit kon duiden op een anaerobe omgeving. Het thallus zag bleekgroen en bij aanraken
voor microscopisch onderzoek viel alles uit elkaar.
Figuur 20: Links bij start van de proef en rechts een gesporuleerde versie, achtendertig dagen later
Bij het experiment in Figuur 20 werd een groot stuk thallus gebruikt. Bij Figuur 21 werd het
stuk thallus verdeeld in kleinere deeltjes door het thallus te scheuren.
Figuur 21: Links het weefsel bij de start van het experiment en rechts zevenenveertig dagen later.
Het weefsel is dan zeer broos en het water rondom is licht groen gekleurd

39
Onderstaande foto (Figuur 22) toont vermoedelijk gameten bij een microscopische vergro-
ting van 40 maal. Er zijn twee flagellen zichtbaar. Bij sporen zouden er vier flagellen zicht-
baar zijn. Deze gameten werden verder gebruikt in proef 4.4.3: Isolatie van sporen.
Figuur 22: Vermoedelijk gameten van Ulva lactuca
Herhaling tweede proef van Sluis (1.2)
Deze methode werd nogmaals herhaald als controle. Bij herhaling kleurde het omliggende
zeewater na drieëndertig dagen groen. Bij elke verversingsbeurt werd ook de pH en het
zoutgehalte gemeten. Bij de start van de proef was de pH 7,99 en het percentage NaCl
3,21%. Na twee dagen was de pH al 8,14 en was het zoutgehalte gestegen tot 3,28%. Na vijf
dagen zonder verversing was de pH al tot 9,38 gestegen en het zoutgehalte gedaald tot
2,79%. Dit zou er op kunnen wijzen dat door verbruik van de nutriënten door de zeesla de
pH van het zeewater stijgt en het zoutgehalte daalt. Of dit sporulatie stimuleerde was ondui-
delijk.
Figuur 23: Herhaling van de tweede methode van Sluis (1.2).
Links bij start van de proefopzet rechts drieëndertig dagen later.
Er is dan een groene wolk zichtbaar, dit zou kunnen wijzen
op het vrijkomen van sporen of gameten
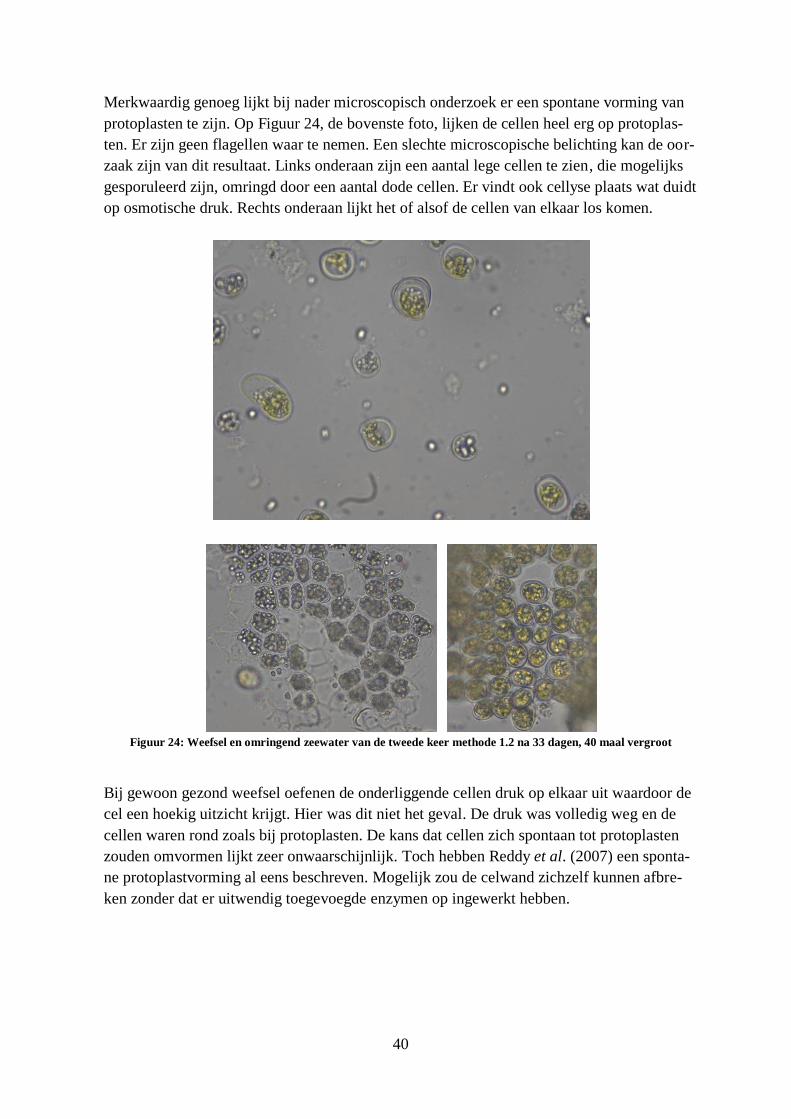
40
Merkwaardig genoeg lijkt bij nader microscopisch onderzoek er een spontane vorming van
protoplasten te zijn. Op Figuur 24, de bovenste foto, lijken de cellen heel erg op protoplas-
ten. Er zijn geen flagellen waar te nemen. Een slechte microscopische belichting kan de oor-
zaak zijn van dit resultaat. Links onderaan zijn een aantal lege cellen te zien, die mogelijks
gesporuleerd zijn, omringd door een aantal dode cellen. Er vindt ook cellyse plaats wat duidt
op osmotische druk. Rechts onderaan lijkt het of alsof de cellen van elkaar los komen.
Figuur 24: Weefsel en omringend zeewater van de tweede keer methode 1.2 na 33 dagen, 40 maal vergroot
Bij gewoon gezond weefsel oefenen de onderliggende cellen druk op elkaar uit waardoor de
cel een hoekig uitzicht krijgt. Hier was dit niet het geval. De druk was volledig weg en de
cellen waren rond zoals bij protoplasten. De kans dat cellen zich spontaan tot protoplasten
zouden omvormen lijkt zeer onwaarschijnlijk. Toch hebben Reddy et al. (2007) een sponta-
ne protoplastvorming al eens beschreven. Mogelijk zou de celwand zichzelf kunnen afbre-
ken zonder dat er uitwendig toegevoegde enzymen op ingewerkt hebben.

41
Tweede herhaling tweede proef van Sluis (1.2)
Na nogmaals een herhaling van de tweede methode van Sluis (1.2) is er merkwaardig ge-
noeg na vijftig dagen nog steeds geen groene wolk waar te nemen. Dit werd zowel getest
met oud weefsel als met vers geoogst weefsel. Beiden bleven hardnekkig groen zonder enige
tekenen van sporulatie, zelfs geen afstervende deeltjes (zie Figuur 25).
Figuur 25: Derde uitvoering van de tweede methode van Sluis (1.2).
Links met oud weefsel dat al bijna een jaar in het opslagaquarium zat,
rechts met nieuw, vers geoogst, weefsel

42
4.3.4.4 Methode 2 (Goudsmit, 2013)
Bij de methode volgens Goudsmit werd ± 5g Ulva lactuca thallus genomen en gecentrifu-
geerd (6min. tegen 8000rpm). Dit thallus werd in een petriplaat gelegd gevuld met 40ml
zeewater. Deze platen werden gedurende 24u in het donker bewaard en dit bij 4°C in een
koelkast. Het stuk thallus werd nadien in een petriplaat met vers zeewater van 20°C geïncu-
beerd bij 100µmol fotonen m-2
s-1
.
4.3.4.5 Resultaat methode 2
Het materiaal leek gesporuleerd. Er waren witte stukken zichtbaar in het groene thallus. Na
de koelkastperiode en het verversen van het water werd geen verandering waargenomen. Pas
vanaf dag tien kwamen er hier en daar witte vlekken aan de rand van het thallus. Maar bij
nader microscopisch onderzoek bleken deze witte randen gewoon afgestorven weefsel te
zijn (zie Figuur 26). Dit weefsel stierf waarschijnlijk af doordat de randcellen beschadigd
werden bij het in stukken snijden. Ook bij de herhaling waren er geen sporen of gameten
waargenomen.
Figuur 26: Beeld van de witte stukken weefsel bij 40X vergroting
4.3.4.6 Methode 3 (Han, et al., 2008) (oorspronkelijk voor Ulva pertusa)
Met een kurkboor werden schijfjes van 0,6cm diameter uit de rand van de thallus gesneden.
Het cirkeltje werd in een petriplaat met 10ml zeewater met 32‰ NaCl gelegd. Er werd geïn-
cubeerd in en fytotron van 15°C bij een licht - donker regime van 16u licht en 8u donker. De
platen waren zo gepositioneerd dat ze 80-100µmol fotonen m-2
s-1
opvingen. De pH was
tussen 7 en 9. Volgens het artikel zouden er binnen de 72 tot 96 uur al sporen zichtbaar moe-
ten zijn.
4.3.4.7 Resultaat methode 3
Er waren witte randjes waar te nemen maar geen groene wolk. Bij nader microscopisch on-
derzoek bleken de witten raden gewoon afgestorven weefsel. Deze proef had dus twee keer
geen succes.

43
4.3.4.8 Methode 4 (Azanza & Aliaza, 1999)
± 1,5 g Ulva lactuca thallus werd 1u gedroogd door dit op filterpapier (Whatman No 15) te
leggen in een petrischaaltje. Deze petriplaat lag in een kamer van 15°C onder licht van
60µmol fotonen m-2
s-1
. Na deze droge periode werd het thallus in een petriplaat gelegd met
20ml vers gefilterd zeewater van 15°C en 32‰NaCl.
4.3.4.9 Resultaat methode 4
Het materiaal leek gesporuleerd. Vanaf dag twee waren er kleine witte vlekjes waar te ne-
men (zie links onderaan Figuur 27). Maar het grootste deel van het thallus bleef wel zeker
twintig dagen donker groen. Deze bevindingen waren niet in overeenstemming met het arti-
kel. Volgens Azanza & Aliaza (1999) zouden er binnen de vijf dagen al sporen gevormd
moeten zijn. Dit was niet het geval. De kleine witte vlekjes waren ook hier weer afgestorven
weefsel.
Figuur 27: Resultaat methode 4

44
4.3.4.10 Methode 5 (Wichard & Oertel, 2010)
Er werd ± 5g Ulva lactuca in stukjes van 1 tot 3mm gesneden. Dit thallus werd 15min. ge-
spoeld met gedestilleerd water en gecentrifugeerd (6 min. 8000rpm). Daarna werd het thal-
lus in vers zeewater in een kamer van 20°C bij 100µmol fotonen m-2
s-1
gelegd. Na drie da-
gen mocht het thallus voor een tweede keer gespoeld worden. Na deze tweede spoeling zou
er direct sporulatie moeten optreden, althans volgens het artikel.
4.3.4.11 Resultaat methode 5
Er zijn geen sporen en geen verkleuring waar te nemen. Zelfs 87 dagen later is het zeewier
nog steeds groen, zelfs zonder extra toevoeging van zuurstof en nutriënten (zie Figuur 28).
4.3.4.12 Methode 6 (De Ruiter, mondelinge overdracht 2013)
Een stuk thallus dat al kleurverandering vertoonde werd in een vierkante petriplaat gelegd.
Volgens De Ruiter zou dit stuk thallus al met sporulatie bezig zijn of zouden de cellen reeds
gedifferentieerd zijn. Deze plaat werd dan met zeewater gevuld tot het thallus volledig be-
dekt was. Één kant van de plaat werd verduisterd en de andere kant werd verlicht (in dit ex-
periment zonlicht). Na enkele uren zouden de eventuele gevormde gameten naar het licht
zwemmen en dus in de bovenhoek een groen laagje vormen. De sporen zouden naar de bo-
dem zinken om zich vast te hechten onder het thallus in het donkere gedeelte.
4.3.4.13 Resultaat methode 6
Er zijn geen sporen waar te nemen. Bij verder microscopisch onderzoek bleken de witte
stukken gewoon dood weefsel (Figuur 28). Door lange tijd in volle zon te staan begonnen de
andere organismen aanwezig in het gefilterd zeewater zich te vermeerderen. Hierdoor werd
de zeesla aangetast en opgegeten. Deze methode is niet echt reproduceerbaar. De sporulatie
moet in de bewaaraquaria al begonnen zijn. Hoe de sporulatie geïnduceerd werd bleef on-
duidelijk.
Figuur 28: Dood weefsel bij 40x vergroting

45
4.3.4.14 Methode 7 (Mantri, Singh, Bijo, Kumari, Reddy, & Jha, 2010)
Volgens Mantri et al. (2010) zou het zoutgehalte in combinatie met de temperatuur invloed
kunnen hebben op het sporulatiegedrag van Ulva faciata. Daarom werd deze methode getest
op Ulva lactuca. Per petriplaatje werd er een cirkel thallus gelegd met diameter van 0,6cm.
Er werd 10ml gefilterd zeewater, gedestilleerd water (AD) of een mengsel van elk 50% toe-
gevoegd. Dit werd dan verdeeld over verschillende temperaturen 15, 20, 23 en 26°C (zie
Tabel 5). De plaatjes kregen 60µmol fotonen m-2
s-1
. Van alle plaatjes werd ook een controle
exemplaar gemaakt.
Tabel 5: Proefopzet methode 7
AD 50% AD 50% zeewater zeewater
15°C 1 A,B 2 A,B 3 A,B
20°C 4 A,B 5 A,B 6 A,B
23°C 7 A,B 8 A,B 9 A,B
26°C 10 A,B 11 A,B 12 A,B
4.3.4.15 Resultaat methode 7
Al vanaf dag twee begonnen platen 10 A en 10 B het moeilijk te krijgen. 50% van het blad-
oppervlak was bleek geworden. De thallusschijfjes in AD hadden het duidelijk moeilijker
dan de platen met zeewater. In onderstaande tabellen staat de opvolging van de resultaten na
3 dagen, vier dagen en zeven dagen.
Tabel 6: Dag drie methode 7
AD 50% AD 50% zeewater zeewater
15°C 1B 100% wit; 1A groen groen lichtgroen
20°C 4A 30% wit; 4B groen groen groen
23°C 7A 30% wit; 7B 10%wit groen groen
26°C 50% wit groen groen
Tabel 7: Dag vier methode 7
AD 50% AD 50% zeewater zeewater
15°C 1B 100% wit; 1A groen groen Licht groen
20°C 4A 30% wit; 4B groen groen groen
23°C 7A 40% wit; 7B 20%wit groen 0,5% wit
26°C 50% wit groen Licht groen
Tabel 8: Dag zeven methode 7
AD 50% AD 50% zeewater zeewater
15°C 1B 100% wit; 1A groen Licht groen 3B 10% wit
20°C 4A wit; 4B20% wit groen groen
23°C 7A 90% wit; 7B 30%wit groen 1% wit
26°C 100% wit groen Licht groen

46
Het is duidelijk dat water zonder zout niet zo goed was voor Ulva lactuca. Halvering van het
zoutgehalte maakte niet veel verschil in vergelijking met zuiver zeewater. De witte randen
waren scherp uitgelijnd en het leek of de stukken met lichtere kleur gingen sporuleren. Bij
microscopisch onderzoek bleek dit niet waar te zijn. Bij microscopisch onderzoek van 1A,
2A, 3B, 4A, 5A bleken de witte vlekken gewoon afgestorven weefsel. Hier en daar waren
ook wat lege cellen te zien, maar nergens sporen of gameten. Na zeventien dagen was het
resultaat als beschreven in tabel 9.
Tabel 9: Dag 17 methode 7
AD 50% AD 50% zeewater zeewater
15°C 1B 100% wit; 1A 50% wit 2A 20% wit; 2B groen 3B wit
20°C 100% wit 5A 100% wit; 5B 40%wit groen
23°C 7A 100% wit; 7B 60%wit groen 1% wit
26°C 100% wit 10% wit Licht groen
Onderstaande foto’s tonen het resultaat van plaat 11B en 5B.
Figuur 29: 11B na 17 dagen Figuur 30: 5A, het groene gedeelte is levend weefsel het
zwarte gedeelte afgestorven weefsel en het witte zou
gesporuleerd weefsel kunnen zijn
Figuur 31: 11B: Links 20X vergroting van de rand. Midden een 40X vergroting van de overgang van levend naar
dood weefsel. Rechts lege cellen waarvan het niet duidelijk was of ze gesporuleerd zijn

47
4.3.5 Bespreking sporulatie methoden
De meeste methoden bleken niet succesvol. Of dit te wijten was aan de anaerobe omgeving
of aan het feit dat het meeste zeewier al een tijd in een aquarium had gelegen was niet hele-
maal duidelijk. Dit heeft verder onderzoek nodig. Het is nodig om al deze proeven nog eens
te herhalen en alle parameters zoals licht, temperatuur maar vooral saliniteit en pH beter vast
te leggen. Momenteel zijn de resultaten niet representatief en niet betrouwbaar. De enige
methode die werkte was deze waarbij om de twee dagen het zeewater werd ververst met
koud zeewater, gedurende twee weken ( zie 4.3.4.2).Hier werden bij de eerste poging game-
ten waargenomen. De proef leek niet consistent want herhalingen mislukten. Alle toepassin-
gen, overgenomen van de literatuur, bleken niet te lukken. Of dit lag aan het gebruik van
Noordzeewater of aan de kunstmatige belichting, was niet helemaal duidelijk. De literatuur
liet soms belangrijke details weg waardoor de proeven niet exact konden herhaald worden.
4.3.6 Besluit sporulatie methoden
Hoe sporen en gameten gevormd worden blijft onduidelijk. Een specifieke combinatie van
temperatuur, licht, pH en zoutgehalte moet de vorming van sporen of gameten induceren.
Welke combinatie dit mogelijk maakt blijft onduidelijk. Het besluit was dat verder onder-
zoek nodig zal zijn om de juiste omstandigheden te vinden. Door regelmatig een koude-
shock te geven en lang te wachten hadden we nog het meeste kans op sporulatie.

48
4.4 Identificatie en isolatie sporen en gameten van U. lactuca
4.4.1 Doel
Het doel was sporen of gameten op te wekken, te identificeren om ze vervolgens steriel te
maken en ze dan in een voedingsbodem te initiëren. Na deze initiatie kon de spore of gameet
uitgroeien tot een steriel thallus.
4.4.2 Identificatie van sporen en gameten
Sporen of gameten zijn waar te nemen doordat er een groene schijn in het water verschijnt
en het thallus wit kleurt en scherpe randen krijgt. Onder de microscoop met een vergroting
van 40 zien de sporen en gameten er ovaal uit. Sporen hebben vier flagellen. Gameten heb-
ben twee flagellen en zijn kleiner dan sporen. Mannelijke gameten zouden kleiner en talrij-
ker zijn dan vrouwelijke gameten (mondeling, Wald, Van Zwol en de Ruiter) (Goudsmit,
2013; Sluis, 2012).
4.4.3 Isolatie van sporen
4.4.3.1 Spoelen met steriel zeewater
4.4.3.1.1 Specifieke materialen en methoden
Voor deze proef werden de gameten van de 4.3.4.2: methode 1 gebruikt. Het water rond het
gesporuleerde thallus zag wazig groen (zie Figuur 20). Er werd 90µl zeewater met groene
wolk gepipetteerd en in een eppendorfje gedaan. Het eppendorfje werd met een toerental
van 4000rpm gecentrifugeerd. Het supernatans werd verwijderd. 270µl steriel zeewater werd
toegevoegd in een steriele omgeving (laminaire airflow). De verschillende deeltjes werden
gehersuspendeerd en opnieuw gecentrifugeerd. Dit werd vijf keer herhaald. De pellet beko-
men na de vijfde spoeling werd opgelost in 90µl steriel zeewater en op/in VMA, VLM gego-
ten. De vloeibare oplossing zat in een falconbuis en werd in een schudtoestel geplaatst
(15°C) om de zuurstof toevoer te bevorderen. De vaste planten werden in de fytotron bij
15°C en 60µmol fotonen m-2
s-1
gezet.
Na groei werden een aantal cellen overgeënt in een andere falcon met 40ml medium. Na drie
weken groei werden de cellen microscopisch onderzocht en in en nieuwe opstelling met
perslucht ondergebracht. Alles werd bij 15°C bewaard.

49
4.4.3.1.2 Resultaat en bespreking
De sporen op VMA schimmelden na twee weken (zie Figuur 33). De sporen in VLM waren
ook gecontamineerd. Maar na een plukje gedifferentieerd weefsel over te enten in nieuw
VLM bleef het steriel en had het de kans om verder te groeien (zie Figuur 32). De inhoud
van de falcons onder de microscoop zag er uit als Figuur 34. Deze groene bolletjes waren
bewegelijk en draaiden rond.
Figuur 32: Links gecontamineerde voeding met weefselcellen. Midden vers overgeënte cellen in VIM,
Rechts groei van gameten na drie weken
Figuur 33: Sporen op VMA
Figuur 34: Microscopisch beeld (40X vergroting) van de inhoud van de
falcon met VLM

50
4.4.3.2 Groei in duranflessen met perslucht
4.4.3.2.1 Specifieke materialen en methoden
Wanneer het medium van Figuur 32 uitgeput begon te raken werden deze sporen na micro-
scopisch onderzoek overgezet in een opstelling met duranflessen waar perslucht door gebla-
zen werd. (Lascaux, 2002) (zie Figuur 35). Perslucht werd door luchtfilters in steriele rubbe-
ren buizen geblazen. Hierdoor kon er een zuurstofstroming in het steriele VLM gecreëerd
worden. Alle recipiënten werden samen geautoclaveerd (130°C bij 1atm gedurende 15 tot 30
min.).
Figuur 35: Gameten incubatieopstelling volgens (Lascaux, 2002) met links een aantal opgepipetteerde losse gameten
en rechts een stukje gevormd weefsel uit gameten
4.4.3.2.2 Resultaat en bespreking
De gameten begonnen zich al snel te vermenigvuldigen en vormden groene vlakken aan de
randen waar ze zich konden vasthechten (zie Figuur 36). Helaas verdampte het VLM vrij
snel door de droge perslucht die er in geblazen werd (zie Figuur 37). Hierdoor moest er om
de twee weken minstens 150ml steriel medium bijgegoten worden in de eerste duranfles.
Het afdekken van de dop met folie hielp een klein beetje tegen de verdamping maar niet
voldoende. Figuur 38 toont de groei na 70 dagen.
Figuur 36: Het vermenigvuldigen
van de sporen in de duranfles met
VLM
Figuur 37: Het VLM verdampte onder invloed
van de droge perslucht die er in geblazen werd
Figuur 38: Zeewier weefsel
gevormd na 70 dagen

51
4.4.4 Bespreking identificatie en isolatie sporen en gameten
De vaste bodems droogden uit en beschimmelden. In de vloeibare bodems waren er bruine
contaminaties waar te nemen. Het was pas na overenten dat er een zuivere cultuur kon be-
komen worden. Maar zelfs dan was het niet zeker dat het een Ulva lactuca cultuur betrof.
De groei in een vloeibaar medium gaat goed, zolang er voldoende nutriënten aanwezig blij-
ven (gedurende iets meer dan een maand). Er komen meer cellen en deze groeperen zich in
klontertjes. De groei in duranflessen met perslucht moet echter nog geoptimaliseerd worden.
Een groot probleem was de sterke verdamping van het medium. Bij het verversen was er
kans op contaminatie, zelfs al werd er in een laminaire airflow gewerkt. Na veertig dagen
was er een groene smurrie waar te nemen (zie Figuur 37 ) Maar er was nog geen sprake van
zeeslaweefsel klaar voor de teelt. De foto genomen na 70 dagen was een klompje cellen en
geen geordend thallusweefsel. De groei duurt langer dan aangegeven door Ohno & Duy Tret
(1997). Zij beweren dat er na 20 dagen al mooi weefsel kan verkregen worden.
4.4.5 Besluit identificatie en isolatie sporen en gameten
Spoelen met steriel zeewater om de gameten steriel te krijgen was niet geschikt. Er moet nog
een betere methode gezocht worden. Gameten groeien niet op VMA, ze overleven alleen in
VLM. Aan de opkweekmethode in duranflessen moet nog gewerkt worden. Deze methode
was arbeidsintensief en was gevoelig voor besmetting Ook de groei van weefsel duurde
lang.

52
4.5 Vorming van protoplasten
4.5.1 Doel
Ook via een protoplascultuur kan steriel weefsel verkregen worden dat dan verder in vitro
geteeld kan worden. Alle hinderlijke organismen kunnen samen met de celwand verwijderd
worden. Als de celwand nadien terug geregenereerd wordt, kan er thallusweefsel gevormd
worden. De eerste doelstelling was dus het creëren van protoplastische zeewieriecellen.
4.5.2 Inleiding
Protoplasten kunnen op verschillende manieren bekomen worden. Deze celwand kan op
mechanische, enzymatische, immunologische, chemische, spontane of elektrische wijze
verwijderd worden, zodat enkel de celinhoud en het celmembraan overblijven. In dit onder-
zoek werd geopteerd om protoplasten te bekomen via enzymatische weg omdat de enzymen
in het labo voorhanden waren vanwege een voorgaand onderzoek.
Op pagina 166 van ‘a simple method for mass isolation of protoplasts from species of Mo-
nostroma, Enteromorpha and Ulva (chlorophyta, Ulvales)’ van Uppalapati en Fujita (2002)
staat te lezen dat zij een steriele mutant uit India cadeau kregen van Dr C.R.K. Reddy. Het
was toen niet duidelijk of het steriel van ‘niet mogelijk om zich voort te planten’ of steriel
van ‘niet besmet’ bedoeld werd. Na het lezen van een aantal artikels van C.R.K. Reddy ont-
stond het idee om via plantregeneratie van protoplasten een steriel van ‘zonder ongewenste
micro-organismen’ algenweefsel te bekomen.
De in de literatuur gevonden methoden om protoplasten te vormen zijn niet altijd even uni-
form. Daarom worden eerst twee artikels uitgetest. Nadien worden de osmotische waarde
aangepast om tot het beste resultaat te komen.
4.5.3 Vormen van protoplasten
4.5.3.1 Poging 1 afgeleid van protoplastvorming bij zaadplanten + aba-lonepoeder
4.5.3.1.1 Inleiding
Eerst werd de methode voor landplanten uitgetest omdat de kennis hiervan het grootste is.
De geraadpleegde literatuur rond protoplastvorming bij zeewieren, gebruikt abalonepoeder
om de celwand af te breken, daar waar cellulase als enzym alleen niet voldoende was.

53
4.5.3.1.2 Specifieke materialen en methoden
Er werd een 50mM 2-(N-morpholino)ethane sulphonic acid (MES) buffer gemaakt met pH 6
in gefilterd zeewater. 0,2M mannitol, 2% cellulaze onozuka R-10 en 2% abalonepoeder
werd hierin opgelost en gefiltreerd. 50mg zeewier samen met 4ml van bovenstaande oplos-
sing werd in een petriplaatje gedaan. Deze petriplaat werd gedurende 16u al schuddend ge-
ïncubeerd op kamertemperatuur en met 25 toeren per minuut.
4.5.3.1.3 Resultaat en bespreking
Alle cellen waren kapot. Geen enkele protoplast te zien. Deze methode is maar één keer uit-
gevoerd.
4.5.3.2 Poging 2 afgeleid van (Reddy & Fujita, 1991; Reddy et al., 2007)
4.5.3.2.1 Inleiding
De methode voor standaard landplanten werkte niet. Daarom werden zeewiermethoden ge-
bruikt beschreven in de literatuur.
4.5.3.2.2 Specifieke materialen en methoden
Jong vegetatief thallus werd gewassen met gefilterd zeewater. De enzymen werden in de
gewenste concentratie opgelost in water met 1% NaCl en 50mM MES buffer met pH6 +
0,6M Mannitol. De enzymconcentraties zijn 2% Fulka cellulase van Aspergillus niger en 2%
abalonepoeder (halias Iris aroma New-Zealand). Gefiltreerd omwille van de grove stukjes in
het abalonepoeder. Er werd 50mg weefsel in kleine stukjes gesneden van ongeveer 1mm2 en
hieraan toegevoegd. De plaatjes incubeerden bij 20°C in het donker op een roterende schud-
der (15-20rpm) gedurende drie uur. De protoplasten werden na drie uur door filtratie en cen-
trifugatie geïsoleerd. De scheiding van grote resten werd gedaan met een nylonfilter en er
werd gecentrifugeerd gedurende 10min. bij 1000rpm. Na de isolatie werden de protoplasten
in 1ml van 0,2M mannitol 50mM MES en 1% NaCl gehersuspendeerd.
4.5.3.2.1 Resultaat en bespreking
Ook hier was geen enkele protoplast te zien, wel veel ongewenste contaminanten. Waar-
schijnlijk is drie uur incubatietijd niet genoeg. Hierdoor is besloten deze methode opnieuw
te testen met een langere incubatie duur (zie 0)

54
4.5.3.3 Poging 3 testen van verschillende concentraties mannitol
4.5.3.3.1 Inleiding
De literatuur betreffende deze methode was eensluidend, behalve op het vlak van osmoti-
sche waarden en incubatieduur. Daarom werden verschillende osmotische concentraties en
incubatietijden getest.
4.5.3.3.2 Specifieke materialen en methoden
Het proces was gelijklopend met poging 2, maar met dit verschil dat er 0,2; 0,3; 0,4; 0,5;
0,6; 0,8 en 0;8 M mannitol uitgeprobeerd werd. Ook de invloed van incubatieduur werd na-
gegaan door regelmatig met de geïnverteerde microscoop naar het weefsel te kijken.
Samenstelling oplossing Y:
1% NaCl
0,75g abalone acetone poeder (halias Iris aroma New-Zealand) / 50ml (1,5%)
0,75g Fulka cellulase van Aspergillus niger / 50ml (1,5%)
0,83 MES / 50ml
50ml miliQ water
oplossing Y werd gefiltreerd en in 2 maal 25 ml verdeeld.
Met de 1e 25ml werd oplossing X gemaakt.
Samenstelling oplossing X:
4,55g mannitol/ 25ml (om 1M oplossing te maken)
50mM MES
25ml miliQ water
1% NaCl
Nadien werden in kleine petriplaatjes de volgende oplossingen verdeeld:
Voor 0,3M: 1,2ml X + 2,8ml Y
Voor 0,4M: 1,6 ml X + 2,4 ml Y
Voor 0,5M: 2 ml X + 2 ml Y
Voor 0,6M: 2,4 ml X + 1,6 ml Y
Voor 2,8M: 1,6 ml X + 1,2 ml Y
Voor 3,2M: 1,6 ml X + 0,8 ml Y
De plaatjes werden gevuld en het zeewier werd in stukjes gesneden. Daarna werd alles in
aluminiumfolie op de schudder geplaatst en regelmatig met de microscoop gecontroleerd op
het loskomen van cellen.

55
Na het loskomen van de cellen werd de oplossing gefilterd met een nylonfilter en in eppen-
dorfjes verdeeld. Deze werden dan gecentrifugeerd gedurende 10min. bij 1000rpm. Het su-
pernatans werd verwijderd en de pellets geresuspendeerd met de juiste concentratie van op-
lossing X. Deze proef werd twee keer uitgevoerd.
4.5.3.3.1 Resultaat en bespreking
Bij 0,3M is niets te zien, hier en daar een geëxplodeerde cel. Bij 0,4M hangen de cellen nog
aan elkaar, maar geen protoplasten te zien. Een voorbeeld van cellen die loskomen is te zie
in Figuur 39. Bij 0,5M en 0,7 M mannitol was er niets te zien.
Op het eerste zicht waren er bij de tweede poging na 24u incuberen bij de 0,6M mannitol het
meeste protoplasten aanwezig (zie Figuur 42). Op één objectglaasje zijn er zeker 109 proto-
plasten zichtbaar. Bij de 0,4M mannitol slechts acht. Maar ook een osmotische concentratie
met 0M mannithol kan protoplasten vormen, of deze ook levensvatbaar zijn valt te betwijfe-
len (zie Figuur 40) Hier was duidelijk veel van het omgevingsvocht opgenomen. Voor een
exactere telling kan er best een Bürker chamber telkamer gebruikt worden.
Figuur 39: Na drie uur beginnen de
cellen los te komen (20X vergroot)
Figuur 40: Links, één van de weinige protoplasten bij 0M mannitol na 16u
incubatie. Rechts, de herhaling, 24u incubatie ook 0M mannitol. (40X ver-
groting)
Figuur 41: 0,8M mannitol na 24u
incubatie (40X vergroting)
Figuur 42: 0,6M mannitol na 24u incubatie (40X vergroting)

56
4.5.3.4 Poging 4: gelukte poging herhalen + testen zonder cellulase
4.5.3.4.1 Inleiding
Voorgaande tests waren niet eenduidig, daarom werden nog enkele herhalingen uitgevoerd.
4.5.3.4.2 Specifieke materialen en methoden
De oplossingen X en Y werden bewaard bij -80°C. Hiermee werden de proeven van 0,6M,
de 0,4M en de 0M mannitol herhaald met 24u incuberen.
Voor de proef met enkel abalonepoeder als enzym werd er een volledig nieuwe oplossing
gemaakt. Deze test werd maar één keer uitgevoerd.
Figuur 43: Celwand die aan het loskomen is, bekeken door de geïnverteerde microscoop
4.5.3.4.1 Resultaat en bespreking
Hier weer een identiek resultaat. Het meeste protoplasten bij 0,6M mannitol, het minste pro-
toplasten bij 0M mannitol. Als er enkel abalonepoeder als enzym gebruikt werd, werden er
geen protoplasten gevormd.

57
4.5.4 Bespreking vorming van protoplasten
De methode met 0,50mM MES, 1,5% cellulase, 1,5% abalone, 0,6M maninitol en 1%NaCl
in miliQ water was de beste methode. Er moet wel 24u in het donker geïncubeerd worden al
zacht schuddend. Deze incubatieduur was veel langer dan in de vergelijkende literatuur. Dit
kan te wijten zijn aan het gebruikte cellulasepoeder dat al vrij oud was. De temperatuur was
kamertemperatuur (±20°C), dit zou de reactiesnelheid vermoedelijk niet beïnvloeden.
Om het onderzoek af te ronden zouden er nog enkele proeven uitgevoerd kunnen worden:
levensvatbaarheid van de protoplasten via oplossing van fluoresceïne diacetaat
(FDA) (Thielemans, 2005).
degradatiegraad van de celwand testen met behulp van calcofluor-White
regeneratie van de protoplasten via lowmeltingpoint agar
telling van het aantal protoplasten via de Bürker chamber
4.5.5 Besluit: vorming van protoplasten
De methode met 0,50mM MES, 1,5% cellulase, 1,5% abalone, 0,6M maninitol 1%NaCl in
miliQ met 24u incuberen al zacht schuddend in het donker was de beste methode. Natuurlijk
is er nog onderzoek nodig naar onbekende factoren zoals het celaantal, de levensvatbaar-
heid, de aanwezigheid van de celwand, de verdere groei in een medium enz..

58
5 Discussie
Zeesla in vitro vermeerderen bleek heel moeilijk. Bij de eerste initiatieproeven van weefsel
op agarmedium bleek dat vaste voedingsbodems geen succes waren. Ook in de weinige lite-
ratuur over in vitro wierteelt, wordt er altijd gebruik gemaakt van vloeibare bodems (Las-
caux, 2002; Azanza & Aliaza, 1999). De stukjes zeesla die gebruikt werden, verloren ongeï-
dentificeerd doorzichtig vocht of beschimmelden. De standaardprocedure voor landplanten
met 10% javel zoals beschreven bij Werbrouck (2011) was dodelijk. Maar 1% javel werkte
wel gedeeltelijk. Waarom Ulva lactuca nadien afstierf, was niet duidelijk. Was de javel de
doodsoorzaak of was het de voedingsbodem? Alleszins bleken de leefomstandigheden niet
ideaal. De reactie van zeewier op het cytokinine BA is door de korte levensduur van het
stukje thallus ook niet duidelijk geworden. Beschikt het wier wel over een BA receptor?
Misschien lag de weefselsterfte wel aan de samenstelling van de bodem of aan het ontbreken
van essentiële componenten zoals symbiotische bacteriën (Ecomare, 2013; Wichard &
Oertel, 2010).
Ook andere componenten kunnen de oorzaak zijn. In de experimenten was er gevarieerd
gebruik gemaakt van medium met zout water en medium met zeewater. Alle weefsel ging
toch dood ongeacht het type voedingsbodem. Het is nodig om verder te experimenteren met
verschillende type voedingsbodems. Er zouden nog testen moeten gebeuren met andere me-
dia zoals IMR- of PES medium.
Waar meristematisch weefsel zit of hoe groot het innoculum moet zijn, is ook niet duidelijk.
Hier was de veronderstelling dat de schijfjes van eender welke grootte om het even waar in
het thallus genomen bruikbaar zouden zijn. Want het “midden” zo als vermeld bij Van Der
Sluis (2012) was een vage omschrijving. Dit is ook niet verder onderzocht vermits alle
weefsel stierf na enkele dagen. Met dit onderzoek is ook bewezen dat ProClin en antibiotica
toevoegen de zeesla vermoordt. Nochtans hadden Azanza & Aliaza (1999) een bruikbaar
vloeibaar medium met minieme hoeveelheden antibiotica. Maar zelf met hogere concentra-
ties AB dan in het onderzoek van Azana & Aliaza (1999), was in dit experiment na een tijd
toch alles gecontamineerd. De levensduur werd wel kortstondig verlengd dankzij het ge-
bruik van AB.
Ulva lactuca telen in vloeibare MS bodem met zeewater moet mogelijk zijn, kijk maar bij-
voorbeeld naar de sporenoplossing die maanden overleefde. Volgende problemen zijn nog te
overwinnen: het zeewier steriel maken, een goede zuurstoftoevoer vinden en op een makke-
lijke steriele manier de nodige nutriënten aan vullen. Dit zou eventueel kunnen opgelost
kunnen worden door de teelt in een bioreactor. Ook alternatieve sterilisatiemethoden werden
getest, maar zonder enig succes. Pasteurisatie, Tween, ethanol en gasserilisatie bleken ook
dodelijk. Hier was wel één uitzondering bij namelijk die met de vijf minuten natte gassterili-
satie. Hierbij heeft één van de twee controles het bijna twintig dagen overleefd. Wat hiervan
de oorzaak was, is nog onduidelijk.

59
Het uittesten van de gassterilisatie was eigenlijk bedoeld voor sporen want in het VIB wor-
den sporen van mossen gesteriliseerd met behulp van gassterilisatie. Maar door een tekort
aan sporen is dit niet verder uitgetest. Dit sprak andere literatuur tegen van Tijana et al
(2007) en Jen-Tung & Weickun (2003) die hun mossen steriel kregen op de conventionele
manier door ze enkele ogenblikken in een lage concentratie NaClO te houden. Tijdens het
onderzoek werd geen onderscheid gemaakt tussen de vorming van sporen en de vorming van
gameten. Er bestaat voorlopig geen eenduidige methode om sporen of gameten te vormen.
Na het uittesten van de verschillende methoden beschreven in de literatuur, bleek geen enke-
le methode te lukken. Bij toeval had er één keer gameetvorming plaatsgevonden bij de me-
thode met het regelmatig verversen met koud zeewater gedurende 2 weken. Wel veel later
dan vermeld in deze ongepubliceerde thesis. Het groen verkleurde zeewater bleek na micro-
scopisch onderzoek gameten te bevatten, er waren maar twee flagellen zichtbaar. Waarom
hier gameten gevormd werden is dus nog steeds een raadsel. Vermoedelijk heeft de tempera-
tuurverandering van 15°C naar 0°C en de anaerobe condities in de plaat hier iets mee te ma-
ken. Maar ook de verandering in pH en saliniteit zullen een rol gespeeld hebben.
In de toekomst kunnen er daarom best nog eens test gedaan worden met verschillende com-
binaties van pH en zoutgehalte omdat deze twee parameters haast niet beschreven staan in
de literatuur enkel Mantri et al., (2010) vermelden het zoutgehalte. In dit eindwerk bleek de
pH te stijgen en het zoutgehalte te dalen in de loop van de tijd. Maar er zijn te weinig metin-
gen gedaan om dit besluit te kunnen ondersteunen vermits in de meeste platen te weinig
vloeistof zat om te meten met de NaCl en pH meter Hanna instruments HI 2550.
Het is wel mogelijk, zodra er sporen gevormd zijn, deze steriel te krijgen. In dit onderzoek
werden sporen gesteriliseerd door ze vijf keer te spoelen met steriel zeewater en ze nadien
over te enten. Het is opletten geblazen om niet het foute weefsel op te zuiveren. Om de spo-
ren te laten opgroeien tot steriel gezond weefsel is veel voeding en plaats nodig. Perslucht in
duranflessen met medium blazen, bleek niet de ideale methode te zijn, waarschijnlijk kan dit
best eens in de toekomst uitgeprobeerd worden door een bioreactor op te starten. Volgens
(Ohno & Duy Triet, 1997) zou het maar twintig dagen duren om van vastgehechte zoöspore
om een bladachtige monostomatisch front te cultiveren. In deze proef was er na 50 dagen
nog geen tallus gevormd, vermoedelijk door het telkens terug loskomen van het opgroeiend
weefsel bij hervullen of door het af en toe gedeeltelijk uitdrogen. Na 70 dagen was een on-
geordende klonter weefsel gevormd van ongeveer vijf cm lang en drie cm breed.

60
Een derde methode om steriel weefsel te bekomen was via een protoplastencultuur. De wijze
waarom men van Ulva lactuca weefsel protoplasten kon maken stond mooi beschreven in
verschillende artikels uit de literatuur (Björk et al., 1992; Reddy & Fujita, 1991; Reddy et
al., 2007; Uchida et al., 1992; Uppalopati & Fujita, 2002). De regeneratie in een steriel me-
dium lukte niet door tijdsgebrek. Het moet perfect mogelijk zijn om via protoplasten een
steriel thallus te bekomen in een in vitro conditie. De methode met 0,50mM MES, 1,5%
cellulase, 1,5% abalone, 0,6M maninitol en 1%NaCl in miliQ water was de beste. Er moet
evenwel 24u in het donker zacht schuddend geïncubeerd worden. Deze incubatieduur was
veel langer dan in de vergelijkende literatuur. Dit is vermoedelijk omdat het cellulasepoeder
gebruikt bij deze tests al oud was. Het poeder was nog aanwezig in het labo van vorige ex-
perimenten. Hier kunnen we in tegenstelling tot het artikel van Björk et al. (1992) besluiten
dat ongeacht de tijd dat het zeewier in aquaria had doorgebracht, het toch protoplasten
vormden. Maar ook hier is nog bijkomend onderzoek nodig. De levensvatbaarheid van de
protoplasten kan gecontroleerd worden door toevoeging van een oplossing van FDA aan de
protoplast oplossing (Thielemans, 2005). De aanwezigheid van de celwand kan getest wor-
den door een kleuring met Calcofluor White M2R (Uchida et al., 1992). Ook een telling van
het aantal protoplasten is mogelijk met behulp van een Bürker chamber telling. De regenera-
tie met agarose beats besproken in het artikel van Deryckere, Eeckhaut, Van Huylenbroeck,
& Van Bockstaele (2012) kan ook uitgetest worden. Deze methode staat uitgewerkt in de
bijlage. Het was de bedoeling deze uit te testen indien er meer tijd zou geweest zijn.
Tot slot van de discussie kan gesteld worden dat het in vitro telen van zeesla niet onmogelijk
is, maar dat verder onderzoek nodig is om dit in de praktijk te brengen.
Hier volgen nog de aanbevelingen voor toekomstig onderzoek kort samengevat.
Onderzoek van het vocht na afsterven van het thallusweefsel op vaste voedingsbo-
dem om de mogelijke de doodsoorzaak vaststellen;
Uittesten van de invloed van verschillende plantenhormonen op zeewier;
Uittesten van IMR, PES en andere media;
Onderzoek naar waar het groeipunt of het meristeem van zeesla zich bevindt;
Bij sporulatie onderzoek moet de pH en het zoutgehalte nauwlettender in de gaten
gehouden worden. Dit zou mogelijks wel de oorzaak kunnen zijn van sporulatie;
Opstarten van een bioreactor met een sporenoplossing om hieruit weefsel te kunnen
oogsten;
De regeneratie van protoplasten met het medium beschreven in bijlage A kan nog
bekeken worden.

61
6 Algemeen besluit
Om Ulva lactuca in een in vitro cultuur te krijgen moet er eerst een steriele “plant” verkre-
gen worden.
In dit onderzoek zijn er drie manieren uitgetest om steriel weefsel te krijgen. De eerste ma-
nier was een stuk weefsel rechtstreeks steriliseren, de tweede manier was om sporen of ga-
meten te bekomen en deze steriel te krijgen en een derde manier was om via een protoplast-
cultuur aan een steriel thallus te geraken.
Ulva lactuca weefsel is op zich wel steriel te krijgen, maar overleeft daarna niet lang meer.
Ook de vorming van sporen of gameten ging niet zo vlot. Sporen en/of gameten vorming
gebeurde nu door louter toeval. De geraadpleegde literatuur klopte niet, was onvolledig of
werd hier niet juist uitgevoerd. Het vormen van protoplasten daarentegen vlotte wel, maar
door tijdsgebrek zijn deze proeven niet afgewerkt.
We kunnen besluiten dat Ulva lactuca weefsel zeer fragiel is. Het verdraagt in het algemeen
geen gassterilisatie, noch contaminatie, noch pro clin, noch antibiotica, noch droogte, noch
tween, noch ethanol, noch hoge temperatuur en noch 10% javeloplossing. Tien minuten in
1% javel geeft wel steriel levend weefsel maar dit overleeft geen drie dagen.
We besluiten ook dat bij het vormen van sporen of gameten nog onderzoek nodig is. Waar-
om er maar één keer toevallig sporen gevormd zijn, is onduidelijk. De meeste literatuur is
het er wel over eens dat er eerst stresscondities moeten zijn om sporulatie te induceren. In dit
onderzoek was het gelukt gameten te vormen door koude shock en anaerobe omstandighe-
den na ongeveer een maand incuberen bij 15°C met 8u donker en 16u licht. De toen ge-
vormde gameten zijn dan geïsoleerd door vijf keer te spoelen met steriel zeewater. Deze
steriele zeewater-gameten oplossing werd dan in VLM gegoten. Na groei werd een kleine
hoeveelheid groen weefsel overgeënt naar nieuw VLM waar een volledig steriele cultuur
verkregen werd. Omdat de schudtoestellen te weinig zuurstof boden en de cultuur niet goed
kon groeien werd een deel overgeënt in duranflessen met perslucht. Maar deze opstelling
moet duidelijk nog verbeterd worden. Of er dan effectief een steriel thallus ontstaat, is on-
duidelijk. In dit project leek dit niet te lukken wellicht door tijdsgebrek, of door het vele
overenten. Volgens Ohno & Duy Triet (1997) zou het maar twintig dagen duren om van
vastgehechte zoöspore naar een bladachtige thallus uit te groeien. In deze proef was er na 50
dagen nog geen echt tallus gevormd, dit kwam waarschijnlijk door het telkens terug los ko-
men bij hervullen en door het af en toe gedeeltelijk uitdrogen. Er was een ongedifferentieer-
de en ongestructureerde massa van cellen gevormd na 70 dagen.

62
Het onderzoek naar de vorming van protoplasten ging vlotter. Dankzij de literatuur was er
snel een ideale protoplastvormingsmethode gevonden. Een enzymen oplossing van 1,5%
Fulka cellulase van Aspergillus niger en 1,5% abalonepoeder in een bufferoplossing van
0,50mM MES, 0,6M mannithol en 1% NaCl in miliQ water was de ideale oplossing om de
Ulva lactuca cellen te ontdoen van hun celwand. Het loskomen van de protoplasten gebeur-
de onder zacht schudden (20rpm) in het donker. De protoplasten kwamen pas na 24u los.
Een mogelijke verklaring hiervoor is dat de cellulase enzymen al iets ouder waren. Hier
kunnen we in tegenstelling tot het artikel van Björk et al. (1992) besluiten dat ongeacht de
tijd dat het zeewier in aquaria had doorgebracht, het toch protoplasten vormde. De verder
verwerking van de protoplasten en de regeneratie van steriel weefsel heeft niet kunnen
plaatsvinden door tijdsgebrek. In bijlage staat wel een potentiële methode tot opvolging van
de groei van de protoplasten die in de toekomst kan uitgevoerd worden.
Dit onderzoek is dus lang niet afgerond. Vele parameters zijn ongekend zodat geen alomvat-
tende besluiten getrokken kunnen worden. Zeesla in een in vitro cultuur krijgen moet moge-
lijk zijn, maar er dient nog tijd in het onderzoek gestoken worden. Ook de kennis over de
precieze trigger die leidt tot voortplanting bij Ulva lactuca is beperkt. Het is pas mogelijk
om uit sporen en gameten een volgroeid thallus te bekomen indien we deze mechanismen
beter begrijpen. De methode om steriel weefsel te verkrijgen via protoplasten lijkt het meest
plausibel. Het was relatief makkelijk om protoplasten te vormen. De eerstvolgende opdracht
is nu om deze protoplasten te isoleren in een steriel medium en ze zo te laten regenereren.

63
Referentie lijst
Azanza, R., & Aliaza, T. (1999). In Vitro carpospore release and germination in
Kappaphycus alvaverzii (Doty) Doty from Tawi-Tawi, Philippines. Botanica
Marina. 42, pp. 281-284.
Berkhout, P., Bakker, E., & Bruchem, C. (2010). Vlees, waarom niet?, eds pp. 22-27. Den
Haag: Lei Wagenignen UR.
BioInformatics, E. (2013). soortengroep classificatie. Geraadpleegd op 09 30, 2013, via:
<http://www.soortenbank.nl/soorten.php?soortengroep=duikgids&selected=beschrijv
ing&menuentry=soorten&record=Ulva%20lactuca>
Björk, M., Gómez-Pinchetti, J. L., Reina, G. G., & Pedersén, M. (1992). Protoplast isolation
from Ulva rigida (Chlorophyta). European Journal of Phycology, pp.401-407.
Brandenburg, W. Seaweed, commodities for the future. pouwerpoint presentatie.op 15
februari 2013, Gent, Oostvlaanderen, België: Universiteit Wageningen.
Busink, S., van Dalen, J., & van Dalen, P. (2009). Inventarisatie en beoordeling van
soorten. Zeewieren uit de Zeeuwse Delta [eindwerk]. Hogeschool Zeeland,
Aquatische EcoTechnologie
Clifton, K., & Clifton, L. (1999). The phenelogy of sexual reproduction by green algae
(Bryopsidales) on caribbean coral reefs. J. Phycol, pp.24-34.
Coppejans, E. (1995). Flora van de Noord-Franse en Belgische zeewieren. Leuven:
Nationale plantentuin van België.
Demeulenaere, M. (2013, 17 03).(Phd student VIB). (Gassterilizatie mossporen)
persoonlijke communicatie [interview]. VIB, Gent.
Deryckere, D., Eeckhaut, T., Van Huylenbroeck, J., & Van Bockstaele, E. (2012). Low
melting point agarose beads as a standard method for plantlet regeneration from
protoplasts within the Cichorium genus. Pant cell Rep., pp.2261-2269.
Ecomare. (2013). Zeesla. Geraadpleegd op 11 28, 2013, via ecomare:
<http://www.ecomare.nl/nl/ecomare-encyclopedie/organismen/planten/algen-en-
wieren/groenwieren/zeesla/>
Eichhorn, R., Evert, S., & Raven, P. (2005). Biology of plants. US: W. H. Freeman.
FAO. (2003). Seaweeds used als human food. Geraadpleegd op 11 30, 2013, via FAO:
<http://www.fao.org/docrep/006/y4765e/y4765e0b.htm>
Fischer, G. "World Food and Agriculture to 2030/50: How do climate change and bioenergy
alter the long-term outlook for food, agriculture and resource availability?"
Geprezenteerd op: How to feed the World in 2050. Proceedings of a technical
meeting of experts, Rome, Italy, 24-26 juni 2009
Goudsmit, E. (2013). Use of a cold water treatment to induce sporulation in Ulva lactuca
[Bachelorproef]. Wageningen Universiteit, faculteit plant science.
groenwieren. (2011). Geraadpleegd op 09 30, 2013, via zeewierwijzer:
<http://www.zeewierwijzer.nl>
Guiry, M. D. (2000). Ulva: Sea lettuce. Geraadpleegd op 11 16, 2013, via Seaweed:
<http://www.seaweed.ie/algae/ulva.php>

64
Han, T., Han, Y.-S., Park, C. Y., Jun, Y. S., Kwon, M. J., Kang, S.-H., et al. (2008). Spore
release by green alge Ulva: A quantitative assay to evaluate aquatic toxicants.
Elsevier, pp.699-705.
Hardenberg. (2013, 03). Beschr. 01 afdeling Bacteriophyta t.m. fam. Delesseriaceae.
Geraadpleegd op 11 16, 2013, via Beschrijving afbeeldingen flora op postzegels:
<http://www.floraoppostzegels.nl/index.php/beschrijvingen/20-beschr-01-afdeling-
bacteriophyta-t-m-fam-delesseriaceae>
Huys, H. (2002). In vitro-cultuur van Asimina triloba[Masterproef]. Hogeschool Gent,
Departement toegepaste bio-ingenieurswetenschappen.
INRA sience & impact (2013) in vitro culture methods, geraadpleegd op 12 20, 2013 via
Green biotechnologies: <http://www.inra.fr/en/Scientists- Students
/Biotechnologies/All-reports/green-biotechnologies/In-vitro-culture-methods>
Keizer, G. (2013). zeesla (Ulva lactuca). Geraadpleegd op 11 14, 2013, via Soortenbank:
<http://www.soortenbank.nl/soorten.php?soortengroep=duikgids&id=339>
Knee, M. (2008, 9 12). Algae. Geraadpleegd op 10 6, 2013 via Hort & crop science:
<http://hcs.osu.edu/hcs300/algae.htm>
laboratories, P. t. (2011). Cellulase onozuka R-10. Geraadpleegd op 11 17, 2013, via Phyto
technology laboratories: <http://www.phytotechlab.com/detail.aspx?ID=193>
Lascaux, B. (2002). Établissement et entretien de cultures in vitro awéniques de plantes
"inférieures" aquatiques. Biotechnologies végétales. Parijs, Frankrijk: Université
Paris-Sud.
Mantri, V. A., Singh, R. P., Bijo, A. J., Kumari, P., Reddy, C., & Jha, B. (2010). Differential
response of varying salinity and temperature on zoospore induction, regeneration and
daily growth rate in Ulva fasciata (Chlorophyta, Uvales). Springer science +
Business Media, pp.243-250.
NCDO (2011), Bevolkingsgroei en de gevolgen ervan. Factsheet 2011. NCDO, Amsterdam,
Nederland
Niesenbaum, R. A. (1988, maart 10). The ecology of sporulation by the macroalga Ulva
lactuca L. (chlorophyceae). Aquatic Botany, pp. 155-166.
Oertel, T. W. (2010). Gametogenesis and gamete release of ulva mutabilis and ulva lactuca
(chlorophyta): Regulatory effects and chemical characterization of the "swarming
inhibitor". Phycological Society of America, 46, pp.248-259.
Ohno, M., & Duy Triet, V. (1997). Artifecial seeding of the green seaweed Monostroma for
cultivation. Journal of applied Phycology, 417-423.
Ponette, Q., & Bertin, P. (sd). Les plantes (1 de 2). Geraadpleegd op 10 6, 2013, via
Formation en biologie végétale: <http://www.afd-
ld.org/~fdp_bio/content.php?page=cara_plantes&skin=modi>
Reddy, C., & Fujita, Y. (1991). Regeneration of plantlets from Enteromorpha (Ulvales,
Chlorophyta) protoplasts in axenic culture. Journal of applied phycology, pp.265-
275.
Reddy, C., Gupta, M. K., Mantri, V. A., & Jha, B. (2007). Seaweed protoplasts: status,
biotechnological perspectives and needs. Springer Science + Business Media, 619-
632.

65
Ruiter, D. (2013, 14 augustus) (Manager Wageningen Light Microscopy Center (WLMC)
WUR, Lab. of Cell Biology). Visuele kenmerken sporulatie[interview]. Wageningen
Universiteit, Wageningen.
Sperber, B. (2010). Ulva biochemical composition. Wageningen: AFSG Wageningen
Universiteit.
Supelco. (2013). ProClin® 150 Preservative for Diagnostic Reagents. Geraadpleegd op 11
04, 2013, via ProClin® 150 Preservative for Diagnostic Reagents:
<https://www.sigmaaldrich.com/Graphics/Supelco/objects/4800/4758.pdf>
Thielemans, F. (2005). Asymetrische protoplastenfusie bij Araceae [Masterproef].
Hogeschool Gent, departement industrieel ingenieur tuinbouw.
Tormo Molina, R. (2012, 12 3). Uvales. Opgeroepen op 10 6, 2013, van Plantas y Hongos:
<http://www.plantasyhongos.es/algas/Ulvales.htm>
Uchida, A., Yoshikawa, T., Ishida, Y., & Saga, N. (1992). Stable protoplast isolation and its
regeneration into thallus of the marine graan alga Ulva persuta. Nippon Suisan
Gakkaishi, pp.153-157.
UN (2013) World Population Prospects: The 2012 Revision, Key Findings and Advance
Tables. Working Paper No. ESA/P/WP.227. Department of Economic and Social
Affairs, Population Division, New York, USA.
Uppalopati, S. R., & Fujita, y. (2002). A simple method for mass isolation of protoplasts
from species of Monostroma, Enteromorpha and Ulva (Chlorophyta,Ulvales).
Journal of Applied Phycology, pp.165-168.
Van Der Sluis, J. (2012). Growth analysis of Ulva lactuca[Bachelorproef]. Wageningen
Universiteit, Departement Organismal an Developmental biology.
Van Ginneken, V., Verdegem, M., Waller, U., Blisseling, T., Wald, J., Helsper, J., et al.
(2013). [nog niet gepubliceerde publicatie]. Ulva lactuca, applications of an
opportunistic macro-algae species for modern mariculture. Wageningen, Nederland:
Wageningen universiteit.
Van Labeke, M.C. (2013). Groei- en ontwikkelingsbeheersing bij planten.[syllabus]. Gent:
Universiteit Gent.
Verkerk, G., Broens, JB., Bouwens, R.E.A., de Groot, P.A.M., Kranendonk, W., Vogelzang.
M.J., Westra, J., Wevers-Prijs, I.M,(2004) Binas, Informatieboek voor natuurweten
schappen en wiskunde., vijfde druk, Groningen: Wolters- Noordhoff.
Wald, J.(research assistent seaweed cultivation) [Thesis bespreking]. op 10 oktober 2013.
UGent, Gent.
Werbrouck. (2012). Grondloze teelten en hydroteelt[syllabus]. Gent: Hogeschool Gent.
Werbrouck, S. (2013, 17 oktober) (Hoofd plantentechnologie labo). Persoonlijke
communicatie [interview] op 17 oktober 2013. Universiteit Gent, Gent.
Werbrouck, S. (2011). Plantenweefselteelt[syllabus]. Hogeschool Gent, Departement
toegepaste bio-ingenieruswetenschappen.
Wichard, T., & Oertel, W. (2010). Gametogenesis and gamete release of ulva mutabilis and
ulva lactuca (chlorophyta): Regulatory effects and chemical characterization of the
"swarming inhibitor". Phycological Society of America, 46, pp.248-259.
Wolkers, H. (20011). Boeren op zee. Wagenignenworld, pp.16-17.

i
Bijlage
De regeneratie van protoplasten:
Materiaal:
Protoplast medium (PM)
Protoplast medium A (voor 1l):
2,2g MS
0,4molair (osmotische waarde):
o 0,3M mannose = 36g
o 0,1M sucrose = 34g
1% NaCl = 10g
Protoplast medium B (voor 1l)
Zie protoplast medium A maar dan met 10g low melting point (LMP) agarose.
Steriel zeewater
Steriele petriplaten
Steriele pipetten
Laminaera airflow
Steriele eppendors
Seriele nylon filters
centrifuge
Methode:
De protoplastoplossing die al in steriel zeewater gemaakt is, na het regelmatig steriel afspoe-
len van het thallus gebruikt voor protoplast vorming wordt door de nylonfilter gefiltreerd en
nadien gecentrifugeerd. Het supernatans wordt dan verwijderd en de pellet wordt geresus-
pendeerd in medium A. Hierna worden de eppedorfjes terug gecentrifugeerd. Deze hersus-
pendatiestap worden een aantal keer herhaald. (bijvoorbeeld 5 keer). Maak best een controle
dubbel zo dat er op contaminatie kan gecontroleerd worden onder de microscoop voor het
mengen met het low-meltingpoint medium. Voeg bij de protopasten 1ml medium A en 1ml
medium B (op 36°C) Verdeer deze in druppels over de plaat en voeg de gewenste hoeveel-
heid medium A aan de plaat toe nadat de agardruppels gestold zijn. Plaats het geheel op een
schudtoestel voor zuurstoftoevoer.
Opstelling:
A
A+B


















