01 metabolisme inleiding
38
Metabolisme: Inleiding T.H. Groep T E. De Herdt p. 1 METABOLISMEN: INLEIDING 1Inleiding Intermediair metabolisme kan zeer onvolledig gedefinieerd worden als het geheel van gecoördineerde enzymatisch gecatalyseerde reacties die in de cel voorkomen. Deze reacties hebben een drietal hoofdfuncties. - Voldoende chemische energie bekomen uit de verbranding van substraten of uit de absorptie van zonlicht; - Exogene “voedingsstoffen” omzetten in celeigen kompone nten. Dit omvat: - afbraak van de voedingsstoffen tot eenvoudige bouwstenen = KATABOLISME - opbouwen uit deze eenvoudige bouwstenen van celeigen eiwitten, nucleïnezuren, lipiden, suikers = ANABOLISME; - Vorming en afbraak van biomoleculen met zeer gespecialiseerde functies in de cel of in het totaal van het organisme (mRNA, ribosomen, ...). Deze omschrijving slaat in eerste instantie op de individuele cel, maar geldt, bij uitbreiding, eveneens voor het weefsel, het orgaan en zelfs voor het totale organisme.
Transcript of 01 metabolisme inleiding

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 1/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 1
METABOLISMEN: INLEIDING
1Inleiding
Intermediair metabolisme kan zeer onvolledig gedefinieerd worden als het geheel vangecoördineerde enzymatisch gecatalyseerde reacties die in de cel voorkomen. Deze reactieshebben een drietal hoofdfuncties.
- Voldoende chemische energie bekomen uit de verbranding van substraten of uit deabsorptie van zonlicht;- Exogene “voedingsstoffen” omzetten in celeigen komponenten. Dit omvat:
- afbraak van de voedingsstoffen tot eenvoudige bouwstenen = KATABOLISME - opbouwen uit deze eenvoudige bouwstenen van celeigen eiwitten, nucleïnezuren,
lipiden, suikers = ANABOLISME;- Vorming en afbraak van biomoleculen met zeer gespecialiseerde functies in de cel of inhet totaal van het organisme (mRNA, ribosomen, ...).
Deze omschrijving slaat in eerste instantie op de individuele cel, maar geldt, bij uitbreiding,eveneens voor het weefsel, het orgaan en zelfs voor het totale organisme.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 2/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 2
1.1Klassificatie van de organismen op basis van hun C- enenergiebron.
Vanuit metabool oogpunt kunnen de organismen ingedeeld worden:
1.1.1Op basis van hun C-bron- autotrofen: uitgaande van CO2 kunnen alle biomoleculen gesynthetiseerd worden(fotosynthetische cellen en sommige bacteriën).- heterotrofen: vereisen min of meer komplexe koolstofverbindingen om hun biomoleculente kunnen synthetiseren.
1.1.2Op basis van hun energiebron en hun electrondonorDe energiebron kan ofwel zonlicht zijn (fototrofen) ofwel een oxidatie-reductie reactie(chemotrofen). De hierbij gebruikte electrondonors kunnen ofwel eenvoudige anorganische
komponenten zijn (litrotrofen) ofwel meer komplexe organische moleculen (organotrofen).
Deze indeling wordt samengevat in onderstaande tabel.
De heterotrofe organismen kunnen op basis van het feit dat de uiteindelijkeelectronencacceptor al dan niet moleculaire zuurstof is onderverdeeld worden in aëroob ofanaëroob. De meeste cellen of organismen kunnen zowel aëroob als anaëroob leven en zijndus facultatief aëroob of anaëroob.
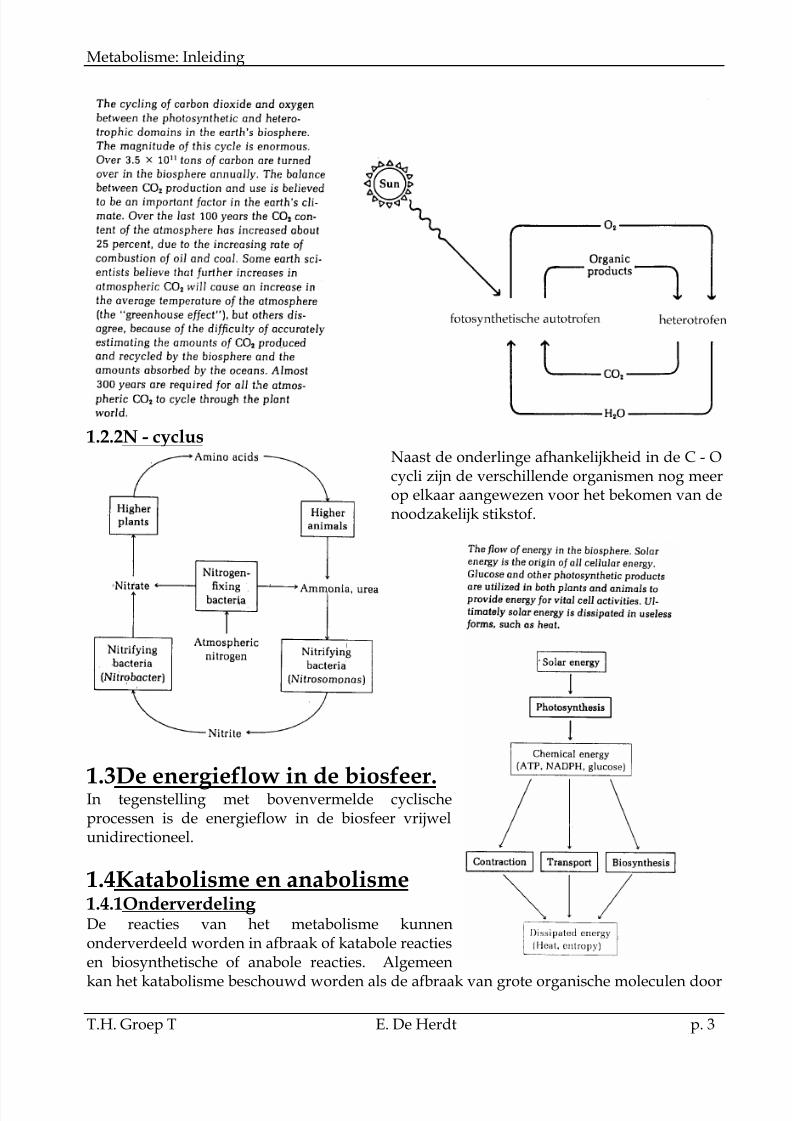
1.2Koolstof - zuurstof cyclusDe levende organismen in de biosfeer en bij uitbreiding de verschillende cellen vanéénzelfde systeem zijn onderling volkomen afhankelijk.
1.2.1C - O cyclusZo wordt zonneënergie, door de fotosynthetische organismen vastgelegd tijdens de fixatievan CO2 tot koolhydraten met vrijstelling van zuurstof. De heterotrofe organismen stellende vastgelegde energie vrij tijdens de ademhaling:
hυ CO2 + H2O (CH2O)n + O2
(CH2O)n + O2 CO2 + H2O + energie
5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 3/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 3
1.2.2N - cyclusNaast de onderlinge afhankelijkheid in de C - Ocycli zijn de verschillende organismen nog meerop elkaar aangewezen voor het bekomen van denoodzakelijk stikstof.
1.3De energieflow in de biosfeer.In tegenstelling met bovenvermelde cyclischeprocessen is de energieflow in de biosfeer vrijwelunidirectioneel.
1.4Katabolisme en anabolisme1.4.1Onderverdeling De reacties van het metabolisme kunnenonderverdeeld worden in afbraak of katabole reactiesen biosynthetische of anabole reacties. Algemeenkan het katabolisme beschouwd worden als de afbraak van grote organische moleculen door

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 4/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 4
oxidatieve reacties tot kleine organische moleculen met vrijstelling van energie. Devrijgekomen energie wordt hoofdzakelijk vastgelegd onder de vorm van energierijkefosfaatbindingen in ATP en in reductie equivalenten onder de vorm van NAD(P)H.
Het anabolisme zal de kleine geoxideerde organische moleculen terug opbouwen tot grote
organische moleculen met gebruik van de energie vastgelegd in ATP en van reductieequivalenten.
1.4.2StadiaKatabolisme en anabolisme verlopen in drie grote stadia.1.4.2.1Katabolisme
I: depolymerisatie van de belangrijkste biopolymeren, eiwitten, polysacchariden enlipiden tot hun respectievelijke monomeren, aminozuren, hexosen, pentosen en
vetzuren. Nucleïnezuren worden door de meeste cellen niet gebruikt als eenverbrandbaar substraat.II: afbraak van de monomeren tot een beperkt aantal eenvoudige intermediairen zoalsacetylCoA.III: een gemeenschappelijke oxidatiecyclus (Krebs of TCA = tricarboxylzuur cyclus)waarin de bekomen acetylresten verbrand worden tot CO2 en H2O met vrijstelling vanenergie.
1.4.2.2AnabolismeIII: α-ketozuren uit de Krebscyclus (α-ketoglutaarzuur, oxaalacetaat) kunnen in
II: geamineerde worden tot aminozuren (Glu, Asp) om vervolgens inI: ingebouwd te worden in polypeptideketens.Op eenzelfde manier worden acetyl-groepen ingebouwd in vetzuren en gaan deze ophun beurt verschillende lipiden vormen.
Merk op dat de afbraak van biopolymeren gepaard gaat met een verlies aan informatie(sequentie van nucleïnezuren en eiwitten) en aan specificiteit van de intermediairen(specifieke aminozuren en vetzuren worden afgebroken tot aspecifieke acetylresten) terwijlbiosynthetische reacties een winst aan specificiteit en informatie inhouden.
1.4.2.3Anabole en katabole wegen zijn niet elkaars tegengestelde

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 5/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 5
- In veel gevallen is dit energetisch onmogelijk omdat de afbraakwegen exergonischereacties bevatten, die niet noodzakelijk samenvallen met de endergonische reacties inde biosynthese.- Indien de anabolische en katabolischereactiewegen volkomen zouden samenvallen
zou een onafhankelijke regulatie van afbraaken biosynthese onmogelijk zijn.- De kompartementalisatie van de eucaryote celmaakt dit in een aantal gevallen fysischonmogelijk.Een gedeelte van de anabolische enkatabolische reactiewegen van een komponentkunnen wel samenvallen en gevormd wordendoor reversiebele reacties (glycolyse -gluconeogenese).
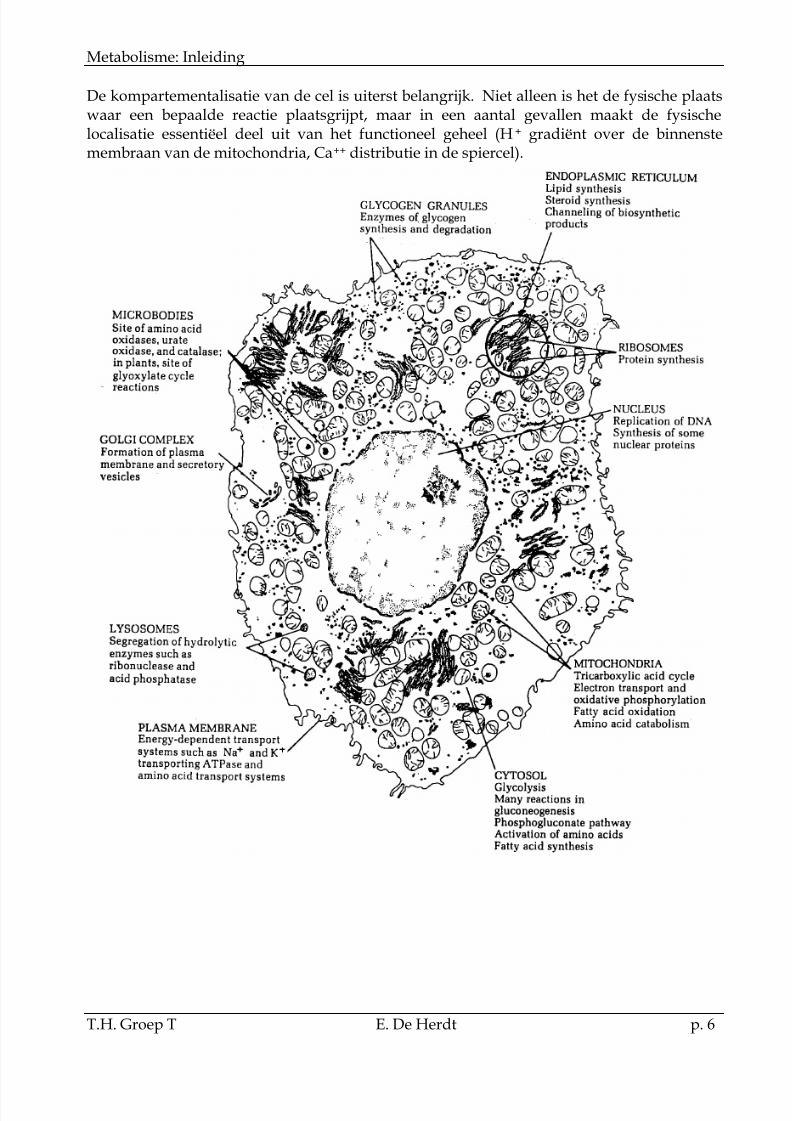
1.5Kompartementalisatie van deeucaryote celDe eukaryote cel bevat meerdere organellen waarinbepaalde delen van het celgebeuren zeer specifiekgelokaliseerd zijn.Van belang zijn onder andere:Plasmamembraan: Deze begrenst de cel als eenidentiteit ten opzichte van zijn omgeving. Hij neemteen belangrijke plaats in in de regulatie van het
transport van extra- naar intracellulair en in derelaties van de cel met zijn omgeving(membraanreceptoren).Cytoplasma: Deze vormt het oplosbare gedeeltevan de cel en bevat een groot aantal enzymen vanmeerdere metabolische wegen.Mitochondria: Dit is een eukaryoot celorganel meteen zeer gespecialiseerde structuur en functie en kanals de krachtcentrale van de cel omschreven worden. Volgens de symbiose theorie zijn demitochondria door evolutie ontstaan uit een aërobe heterotroof.
5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 6/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 6
De kompartementalisatie van de cel is uiterst belangrijk. Niet alleen is het de fysische plaatswaar een bepaalde reactie plaatsgrijpt, maar in een aantal gevallen maakt de fysischelocalisatie essentiëel deel uit van het functioneel geheel (H+ gradiënt over de binnenstemembraan van de mitochondria, Ca++ distributie in de spiercel).

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 7/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 7
2Bioënergetische principes enenergietransfer
2.1PrincipesDe reacties van het metabolisme zijn zuiver chemische reacties en kunnen geschrevenworden als:
aA + bB cC + dD
(C)c (D)d met: K’eq =
(A)a (B)b
Als zodanig voldoen ze aan alle wetten van de thermodynamica. De tweede wet van dethermodynamica stelt dat bij elk proces de entropie (∆S) van het systeem en zijn omgevingsteeds naar een maximum streeft. De veranderingen in entropie zijn echter niet geschikt omde richting en het evenwicht van de reactie te bepalen. Dit kan eenvoudiger gebeuren aan dehand van de veranderingen in vrije energie. Elke komponent bezit een karakteristiekeintrinsieke vrije energie als gevolg van zijn moleculaire structuur.
De standaard vrije energieverandering (∆G°) van een reactie is het verschil tussen de somvan de vrije energie van de produkten en die van de reactantia waarbij reactantia enproducten in hun standaardtoestand aanwezig zijn. Deze standaardtoestand van elk van de
komponenten is bij conventie een concentratie van 1M bij 25°C en bij 1 atm.
G° = Σ G°prod - ΣG°react
G° is dus de hoeveelheid vrije energie die opgenomen of vrijgesteld wordt wanneer A en Bomgezet worden tot C en D onder zulke voorwaarden dat de concentratie van A, B, C en Dop 1M blijft bij standaard temperatuur en druk.Voor een reactie die onder niet-standaardvoorwaarden doorgaat wordt de vrijeenergieverandering (G) gegeven door:
(C)c (D)d
G = G° + RT ln(A)a (B)b
ofG = G° + RT ln K’eq
Deze verandering in vrije energie, G, kan gedefinieerd worden als dat gedeelte van detotale energieverandering van een systeem dat naar een evenwicht gaat bij een gegeventemperatuur, druk en volume, dat beschikbaar is om arbeid te verrichten.Wanneer een reactie op zijn evenwichtstoestand is, dan is de vrije energie van het systeemminimaal en is er dan ook geen verandering in vrije energie (G=0). We krijgen dus:
0 = G° + RT ln K’eq of G°= - RT ln K’eq
of G°= - 2,303 RT log K’eq
5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 8/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 8
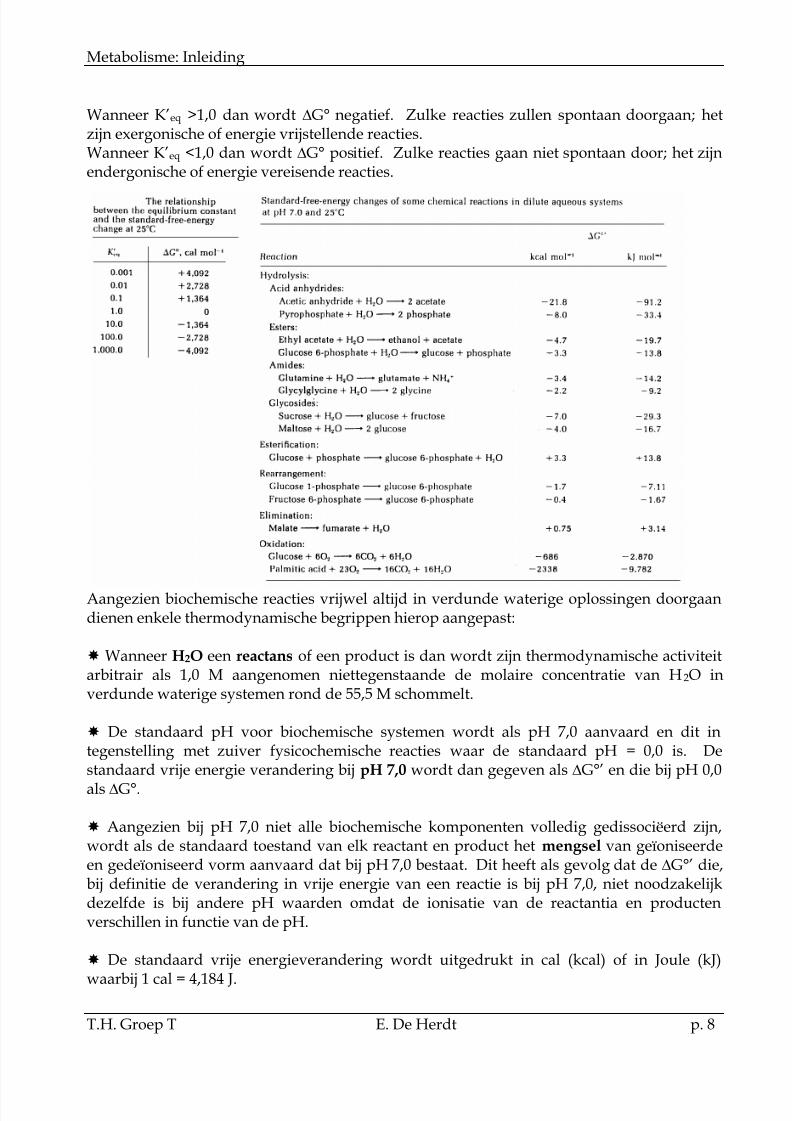
Wanneer K’eq >1,0 dan wordt G° negatief. Zulke reacties zullen spontaan doorgaan; hetzijn exergonische of energie vrijstellende reacties.Wanneer K’eq <1,0 dan wordt G° positief. Zulke reacties gaan niet spontaan door; het zijnendergonische of energie vereisende reacties.
Aangezien biochemische reacties vrijwel altijd in verdunde waterige oplossingen doorgaandienen enkele thermodynamische begrippen hierop aangepast:
Wanneer H2O een reactans of een product is dan wordt zijn thermodynamische activiteitarbitrair als 1,0 M aangenomen niettegenstaande de molaire concentratie van H2O inverdunde waterige systemen rond de 55,5 M schommelt.
De standaard pH voor biochemische systemen wordt als pH 7,0 aanvaard en dit integenstelling met zuiver fysicochemische reacties waar de standaard pH = 0,0 is. Destandaard vrije energie verandering bij pH 7,0 wordt dan gegeven als G°’ en die bij pH 0,0als G°.
Aangezien bij pH 7,0 niet alle biochemische komponenten volledig gedissociëerd zijn,wordt als de standaard toestand van elk reactant en product het mengsel van geïoniseerdeen gedeïoniseerd vorm aanvaard dat bij pH 7,0 bestaat. Dit heeft als gevolg dat de G°’ die,bij definitie de verandering in vrije energie van een reactie is bij pH 7,0, niet noodzakelijkdezelfde is bij andere pH waarden omdat de ionisatie van de reactantia en productenverschillen in functie van de pH.
De standaard vrije energieverandering wordt uitgedrukt in cal (kcal) of in Joule (kJ)waarbij 1 cal = 4,184 J.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 9/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 9
Het intermediair metabolisme wordt inhoofdzaak gevormd door sequenties vanconsecutieve reacties, ieder met zijn eigen G°’. DeG°’ van de totale sequentie is de algebraïsche som
van de G°’ waarden van de verschillendedeelreacties.
De G°’ van een reactie kan ook berekendworden aan de hand van de standaard vrijeenergie van vorming (G°’f) van zijn substraten enzijn producten. De G°’f wordt gedefiniëerd als dedaling in vrije energie wanneer één mol van eenkomponent gevormd wordt uit zijn elementen elkin zijn standaard toestand:
G°’ = Σ G°’fprod - ΣG°’freact
De G°’f van enkele belangrijke moleculen wordtgegeven in de tabel!
2.2Energietransfer
2.2.1Adenosinetrifosfaat
2.2.1.1De ATP cyclusATP functioneert op eencyclische manier als eencarrier van chemischeenergie van catabole
reacties die energie vrijstellen naar de verschillende processen in de cel die energie vereisen.ATP wordt gevormd uit ADP en Pi door de koppeling van de fosforylatiereactie aan eenenergieproducerende reactie (oxidatieve fosforylatie of fosforylatie op substraatniveau). Hetaldus gevormde ATP kan dan gebruikt worden om energie vereisende reacties te doendoorgaan waarbij de terminale fosfaatgroep afgesplitst wordt.
2.2.1.2ATP2.2.1.2.1Structuur
Adenosine trifosfaat (ATP) komt in de cel voor in concentraties tussen de 2 à 10 mM. De
molecule bevat 3 fosfaatgroepen waarvan 2 (β, γ ) in anhydride binding en één (α) inesterbinding.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 10/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 10
Bij pH 7 zijn ATP en ADP sterk geladen anionen. ATP heeft3 van de 4 protonen volledig geïoniseerd terwijl het vierdeproton voor 50 % gedissociëerd is. De totale lading bij pH 7is dus -3,5.
In de cel komt ATP en ADP vrijwel altijd voor onder devorm van hun Mg ++ chelaat. Het Mg ATP2- is het echtesubstraat voor de meeste enzymatische reacties waarin ATPdeelneemt.
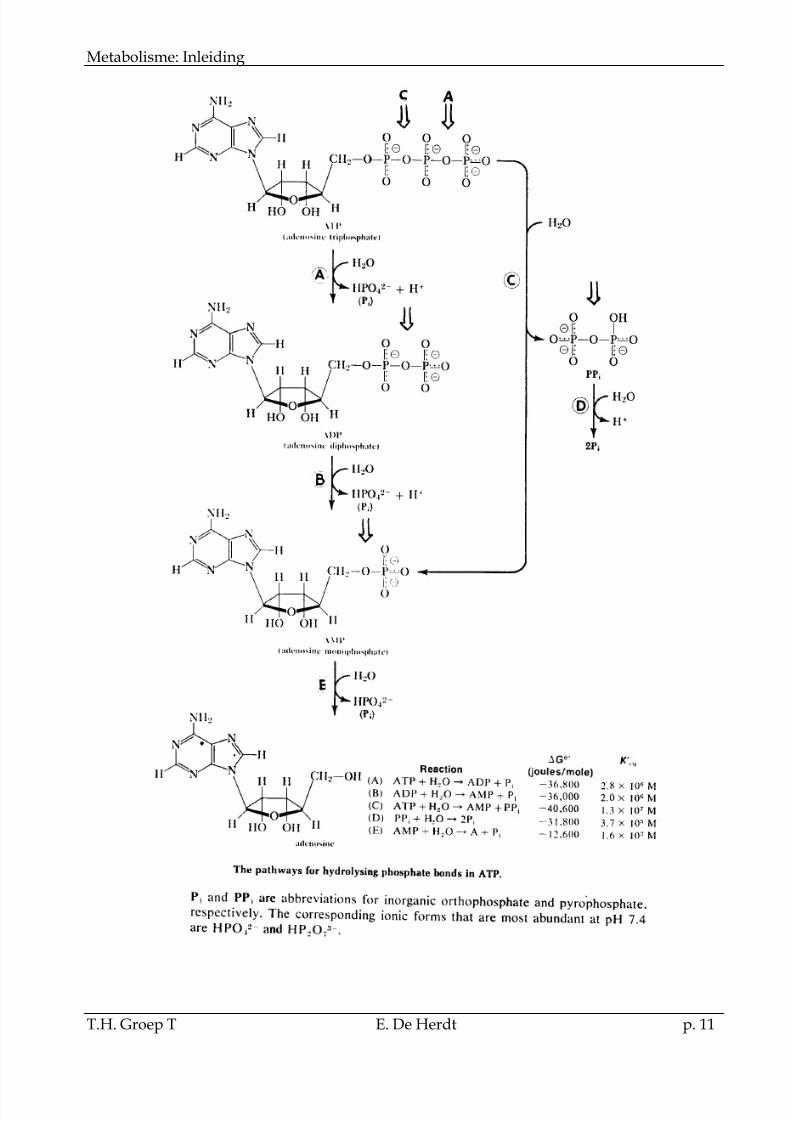
2.2.1.2.2ATP hydrolyse
De fosfaatgroepen van ATP kunnen op verschillende
wijze gehydrolyseerd worden zoals wordt aangegeven in de figuur.
De standaard vrije energie van hydrolyse van verschillende fosfaatverbindingen wordtmeestal uitgedrukt met H2O als arbitrair gekozen acceptor molecule volgens de reactie:
R-O-PO32- + H2O R-OH + HO- PO32-
De G°’ van hydrolyse van de terminale fosfaatgroep van ATP kan bekomen worden:- uit de evenwichtsconstante van de reactie:
ATP + H2O ADP + Pi G°’= -2,303RTln K’eq
Het reactieëvenwicht ligt echter zo ver naar rechts dat in de praktijk een nauwkeurige K’eq niet kan bepaald worden.- uit de additie van enkele ATP afhankelijke reacties:
hexokinase Gluc + ATP Gluc6P + ADP G°’= - 4 kcal/mol
fosfatase Gluc6P + H2O Gluc + Pi G°’= -3,3 kcal/mol
ATP + H2O ADP + Pi G°’= -7,3 kcal/mol
5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 11/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 11

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 12/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 12
Op dezelfde manier kan de G°’ van de verschillende fosfaatgroepen van ATP bepaaldworden
De G°’ voor hydrolyse van ATP in ADP en Pi variëert in de literatuur tussen -7 à -8
kcal/mol. De meest geaccepteerde waarde bij pH 7,0 en 25°C is echter - 7,3 kcal/mol.
De standaard vrije energie vanhydrolyse van enkelefosfaatverbindingen wordtgegeven in de volgende tabel:
De hoge G°’ waarde voorhydrolyse van ATP is in eersteinstantie het gevolg van het feit
dat de P-bindingen allenresonantiehybriden zijn, m.a.w.de dubbele bindingen hebben eenbelangrijk enkel bindingskarakteren vice versa. Deze resonantiehybrieden zijn stabieler dan deeenvoudige enkele of dubbelebinding en bevatten een zekere hoeveelheid resonantieënergie. Bij hydrolyse van eendergelijke binding komt er dan ook meer energie vrij.ATP draagt bij pH 7,0 ongeveer 3,5 negatieve ladingen. Deze zeer dicht bij elkaar gelegen
negatieve ladingen stoten elkaar electrostatisch sterk af. Dit betekent dat bij hydrolyse vanATP tot ADP + Pi een gedeelte van deze electrostatische afstoting opgeheven wordt en dezeenergie vrijkomt. De fundamentele reden van de hoge G°’ bij hydrolyse van ATP of eenandere “energierijke” komponent, is dus steeds te wijten aan de electronenconfiguratie in ofin de omgeving van de binding die gehydrolyseerd wordt. De energie van een“energierijke” fosfaatbinding is niet de energie van de P - O binding (bindingsenergie) maaris het verschil tussen de vrije energie van de reactantia en die van de producten wanneer eengefosforyleerde verbinding gehydrolyseerd wordt.
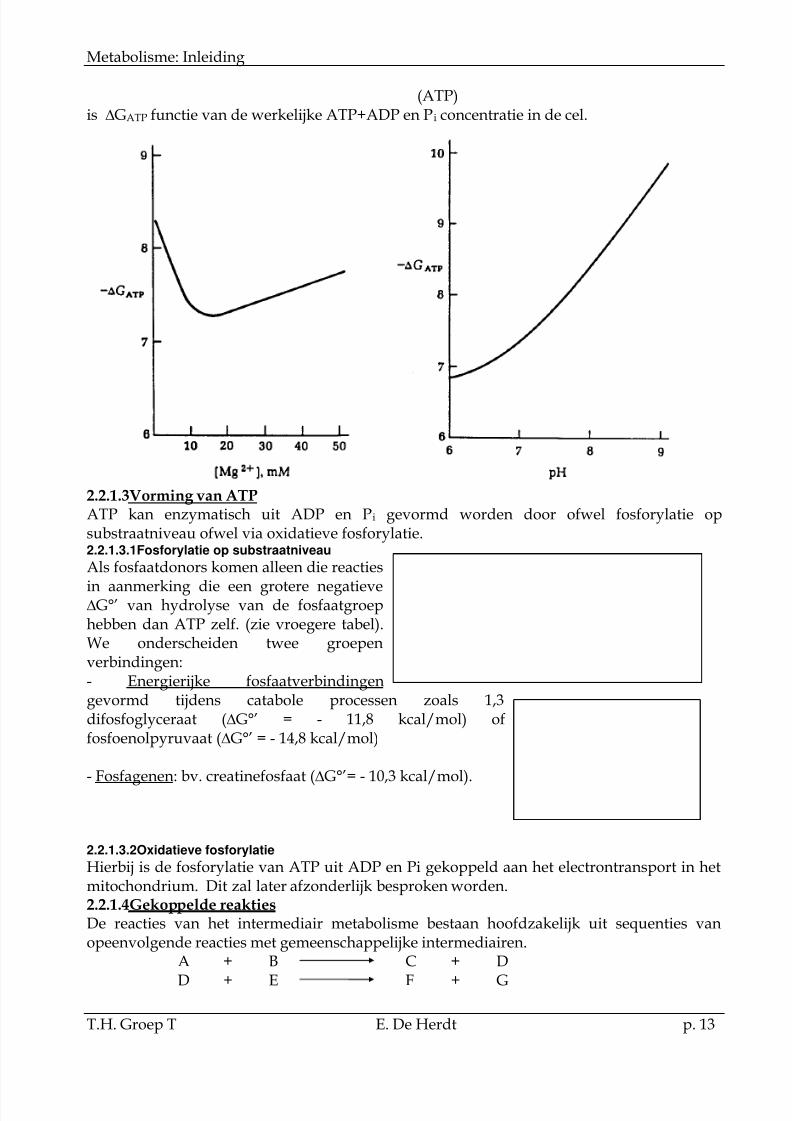
De G°ATP wordt beïnvloed door verschillende factoren:
- pH: bij stijgende pH stijgt de De G°ATP . Dit wordt veroorzaakt door de verhoogdeionisatie van ATP. (grotere electrostatisch afstoting!!)
HATP3- ATP4- + H+ pK’1 = 6,95
- Mg 2+ : G°ATP is ook functie van de Mg2+ concentratie. De affiniteit van ATP voor Mg2+ isafhanlijk van de pH. Dit betekent dat voor elke pH er een G°ATP /Mg2+ curve bestaat.
- De concentratie van ATP/ADP en Pi zelf: de G°ATP is gedefinieerd voor een concentratievan reactantia en producten van 1 M. De concentratie van ATP in de intacte cel variëert
echter tussen 2 à 10 mM. Aangezien:(ADP) (Pi)
GATP = G° + RT ln
5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 13/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 13
(ATP)is GATP functie van de werkelijke ATP+ADP en Pi concentratie in de cel.
2.2.1.3Vorming van ATPATP kan enzymatisch uit ADP en Pi gevormd worden door ofwel fosforylatie opsubstraatniveau ofwel via oxidatieve fosforylatie.2.2.1.3.1Fosforylatie op substraatniveau
Als fosfaatdonors komen alleen die reactiesin aanmerking die een grotere negatieveG°’ van hydrolyse van de fosfaatgroephebben dan ATP zelf. (zie vroegere tabel).We onderscheiden twee groepenverbindingen:- Energierijke fosfaatverbindingengevormd tijdens catabole processen zoals 1,3difosfoglyceraat (G°’ = - 11,8 kcal/mol) offosfoenolpyruvaat (G°’ = - 14,8 kcal/mol)
- Fosfagenen: bv. creatinefosfaat (G°’= - 10,3 kcal/mol).
2.2.1.3.2Oxidatieve fosforylatie
Hierbij is de fosforylatie van ATP uit ADP en Pi gekoppeld aan het electrontransport in hetmitochondrium. Dit zal later afzonderlijk besproken worden.2.2.1.4Gekoppelde reaktiesDe reacties van het intermediair metabolisme bestaan hoofdzakelijk uit sequenties vanopeenvolgende reacties met gemeenschappelijke intermediairen.
A + B C + DD + E F + G

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 14/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 14
G + H I + J J + .......
De transfer van chemische energie in de cel gebeurt volgens hetzelfde principe waarbij ATPhet gemeenschappelijk intermediair is tussen een exergonische en een endergonische reactie.Dit kan als volgt worden voorgesteld.
pyruvaat kinasePEP + ADP Pyr + ATP
hexokinase
Gluc + ATP Gluc6P + ADP
Dit betekent dat ATP op zichzelfnooit de energiebron is voor defosforylatie van een komponentmaar enkel de energiecarrier van een
exergonische naar een endergonischereactie volgens het principe van hetgemeenschappelijk intermediair.
2.2.1.5De fosfaat-pool
ATP is in het intermediairmetabolisme het meest frequentgebruikt gemeenschappelijkintermediair voor de energietransfer,
maar het is niet het enige. Deoverige drie nucleoside trifosfaten(GTP, UTP en CTP) worden in eenaantal gevallen eveneensingeschakeld in de energietransfer.Aangezien de configuratie van defosfaatgroepen in de vier NTP gelijkis, zijn de G°’ waarden van hun hydrolyse ook zeer gelijkend. De gehydrolyseerde NTPworden steeds ten koste van ATP geregenereerd met behulp van specifieke nucleoside di- enmono-fosfaatkinases. Dit betekent dat er constant een NTP pool betrokken is bij de
energietransfer.
De NTP ‘s vormen slechts een zeer kleine reserve voor de energievorming van de cel. Zekunnen niet als stockage dienst doen van energierijke fosfaatbindingen omdat ze maar inlage concentratie in de cel voorkomen en daarenboven een belangrijke regulatorische functievervullen in het nucleïnezuurmetabolisme. Als opslag van fosfaatbindingen worden vooralguanidoderivaten gebruikt (fosfagenen).Creatinefosfaat komt vooral voor in spierweefsel (skelet). Pinguïns en zeehonden bevattenzeer hoge concentraties ervan waardoor ze lang onder water kunnen blijven. Vogels hebbenminder nodig omdat tijdens het lange vliegen de ademhaling zorgt voor een blijven aëroobmetabolisme.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 15/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 15
creatineP kinase
creatineP + ADP creatine + ATPG°’ = - 3,0 kcal/mol
2.2.2Energietransfer tijdens een oxidatie - reductie reactie2.2.2.1Standaard oxidatie - reductie poltentiaalRedox reacties of oxidatie-reductie reacties zijn reacties waarbij er een transfer vanelectronen gebeurt van een electrondonor (reductans) naar een electron acceptor (oxidans).In sommige redoxreacties gebeurt de transfer van één of meerdere electronen via de transfervan H-atomen. Een “dehydrogenatie” is dus equivalent aan een oxidatie. Oxidans enreductans vormen steeds een redoxpaar:
A + e- Doxidans reductanse- acceptor e- donorwordt gereduceerd wordt geoxideerd
De neiging van een reductans om electronen te verliezen of van een oxidans om electronenop te nemen wordt uitgedrukt door de oxidatie - reductie potentiaal. Dit is deelectromotieve kracht (emf) uitgedrukt in V gegeven door een halfcel waarin het reductansen oxidans beiden aanwezig zijn in een concentratie van 1,0 M bij 25°C en pH 7,0 inevenwicht met een electrode die reversiebel electronen kan opnemen van het reductans. Dereferentiepotentiaal is die van de waterstofelectrode:
2 H+ + 2 e- H2
Bij H2 gas = 1 atmosfeer, [H+] = 1,0 M (pH=0,0) en 25°C. Na correctie van de
refentiepotentiaal naar pH 7 ([H+] = 10-7) wordt de standaard oxidatie reductiepotentiaal(Eo’) = - 0,42 V. De gemeten potentiaal (Eh) staat in verband met de standaard oxidatie

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 16/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 16
reductiepotentiaal Eo’ zoals de pK in verband staat met de pH en de proton donors enacceptors. Ze wordt gegeven door de Nernst vergelijking:
2,303 RT [electronacceptors]Eh = Eo’ + log
nF [electrondonors]waar:
Eo’ = standaard oxidatie - reductiepotentiaal bij pH 7 en 25°C en alle concentratiesgelijk aan 1 M.n = aantal getransfereerde electronenF = Faraday (23,062 Kcal/V)
De Nernst vergelijking drukt in feite de titratie uit van de electrondonor met een oxiderend
agens. De standaard oxidatie - reductie potentiaal wordt gedefinieerd als die potentiaal bij50 % oxidatie/reductie. Hiervoor geldt dus: Eh = Eo’.
De Eo’ maakt het mogelijk de richting van de electronoverdracht te bepalen in een reactie.
OxAc + 2 H+ + 2 e- Malaat Eo’ = - 0,18 V
NAD+ + 2 H+ + 2 e- NADH + H+ Eo’ = - 0,32 V
Het koppel NAD+/NADH heeft een sterker negatieve E en zal dus sneller electronen afstaan
aan OxAc dat een grotere affiniteit voor electronen heeft.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 17/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 17
2.2.2.2Standaard vrije energieverandering van een oxidatie - reductie reactieDe G°’ die optreedt wanneer twee geconjugeerde redoxparen met een gekende standaardoxidatie - reductiepotentiaal reageren wordt gegeven door:
G°’ = - nFEo’ waarbij:
n = aantal getransfereerde electronen;F = Faraday constante (23,062 kcal/v);Eo’ = Eo’ van de electronacceptor - Eo’ van de electrondonor.
Dit wordt geïllustreerd aan de hand van het volgende voorbeeld:
Malaat OxAc + 2H+ + 2 e- Eo’ = (-)- 0,18 V
NAD+ + 2 H+ + 2 e- NADH + H+ Eo’ = - 0,32 V
Malaat + NAD+ OxAc + NADH + H+ Eo’ = - 0,14 V
Hieruit volgt dat: G°’ = -2(23,062 kcal/V)(-0,14 V) = 6,5 kcal/molOnder standaardvoorwaarden is deze reactie dus niet spontaan!

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 18/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 18
De uiteindelijke electronacceptor in de ademhalingsketen is moleculaire zuurstof zodat:
1/2 O2 + 2H+ + 2 e- H2O Eo’ = + 0,82 V
NADH + H+ NAD+ + 2H+ + 2 e- Eo’ = (-)- 0,32 V
NADH + H+ + 1/2 O2 H2O + NAD+ Eo’ = + 1,14 V
Hieruit volgt dat: G°’ = - 52,6 kcal/molDit is een spontane, exergonische reactie onder standaardvoorwaarden.
Combineren we bovenstaande reacties, dan bekomen we:
Malaat + NAD+ OxAc + NADH + H+ G°’ = 6,5 kcal/mol
NADH + H+ + 1/2 O2 H2O + NAD+ G°’=-52,6kcal/mol
Malaat + 1/2 O2 OxAc + H2O G°’=-46,1kcal/mol
Bij de oxidatie van malaat naar oxaalazijnzuur kan dus potentiëel energie vrijgesteld wordenter grootte orde van 46 kcal/mol. Er bestaan uiteraard geen enzymesystemen die malaatrechtstreeks oxideren met moleculaire zuurstof (O) zodat het reductieëquivalent (dit zijn deelectronen en/of H-atomen die aan een oxidatie - reductie deelnemen) via NADH en de
electron carriers van de ademhalingsketen naar O2 kunnen getransfereerd worden.
Tijdens deze transfer kan een zeker percentage energie benut worden en vastgelegd wordenals energierijke fosfaatbindingen van ATP in het proces van oxidatieve fosforylatie(electronentransportketen in de mitochondria).
Reductieëquivalenten bekomen bij de oxidatie van substraten kunnen ook rechtstreeksgebruikt worden voor de reductie van precursoren. In dit geval is er dan eveneens eenenergietransfer onder de vorm van energierijke electronen. Bijvoorbeeld: NADPH bekomentijdens de pentose fosfaat cyclus (oxidatie van glucose) kunnen aangewend worden tijdens
de vetzuursynthese.
3Electronen transport en oxidatievefosforylatie
3.1Ademhaling en fermentatieIn het catabolisme worden verschillende substraten (suikers, vetten aminozuren, ...)
enzymatisch afgebroken door oxidatieve reacties. De oxidatie van het substraat gebeurtstapsgewijs door het onttrekken van telkens één electronenpaar aan het substraat.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 19/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 19
Voor deze zeer uiteenlopende oxidaties worden slechts een zeer beperkt aantal cofactoren(NAD, NADP, FAD)gebruikt waarvan de concentratie in de cel beperkt is.
Als gevolg van de oxidatie van het substraat wordt de cofactor zelf gereduceerd en dient opzijn beurt gereoxideerd te worden opdat de afbraak van het substraat zou kunnen blijven
doorgaan. De reoxidatie van de cofactor kan aëroob of anaëroob gebeuren.
3.1.1Anaërobe regeneratie van de cofactorEen anërobe regeneratie van de cofactor gebeurt via de tegenovergestelde reactie, namelijkde reductie van een substraat, waarbij de uiteindelijke electronacceptor een uitgebreide reekskomponenten kan zijn. De energie vrijgekomen door de substraatoxidatie wordt vastgelegdin ATP door substraatafhankelijke fosforylatie. Het type metabolisme wordt fermentatie genoemd.
ADP + Pi
Substr.red Coenzymeox Substr.red
Substr. ox Coenzymered Substr.ox
ATP
3.1.2Aërobe regeneratie van de cofactorBij een aërobe regeneratie van de cofactor is de uiteindelijke electronacceptor moleculairezuurstof. Het reductieëquivalent zal via een aantal electroncarriers zuurstof bereiken. De
aaneenschakeling van deze electroncarriers wordt de electronenetransportketen ofademhalingsketen genoemd en het type metabolisme is aërobe respiratie.
Sommige van deze electrontransferstappen zijn voldoende exergonisch om de vorming vaneen energierijke fosfaatbinding toe te laten tijdens de gekoppelde oxidatieve fosforylatie. Deprocessen van electrontransport en deze van de oxidatieve fosforylatie zijn volkomengelokaliseerd in de mitochondria en zijn sterk membraangebonden.
Substr.red Coenzymeox 1/2 O2 meerdere electron
carriersSubstr. ox Coenzymered H2O
ADP + Pi ATP
3.2Structuur van het mitochondrium
Het mitochondrium is van de grootte-orde van een E.coli cel maar variëert in grootte envorm sterk in functie van zijn metabole activiteit. Het is begrensd door een dubbelemembraan.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 20/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 20
3.2.1Buitenste membraan
De buitenste membraan is glad en elastisch en zeer vlot permeabel voor komponenten totcirca 10 000 dalton. Het wordt gekenmerkt door een hoge fosfolipide - eiwit verhouding
maar met een lage concentratie cardiolipine, terwijl de concentratie aan fosfatidylinositol encholesterol hoog is. Voor de structuur van deze moleculen verwijzen we naar het hoofdstukover vetten.
3.2.2Binnenste membraan
De binnenste membraan is gekenmerkt door een reeks instulpingen waarvan de vorm en hetaantal sterk afhankelijk is van de soort en de energietoestand van de cel. De membraan isimpermeabel voor de meeste moleculen tenzij neutrale moleculen met een molecuulgewicht<150 Dalton. De overige moleculen dienen dus via carriers getransporteerd te worden. Defosfolipide - eiwitverhouding is laag terwijl cardiolipine het belangrijkste fosfolipide is.
Alle electroncarriers komen voor in deze membraan. Op het oppervlak van de membraan -langs de matrixzijde - zijn kleine knobbeltjes zichtbaar met het electronenmicroscoop; dit dede koppelingsfactor F1-FoATPase of het ATP synthetase.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 21/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 21
Tussen de buitenste en binnenste membraan ligt de tussenmembraanruimte. Meerdereenzyme systemen en electroncarriers zijn in de membraan ingebed. Ze zijn hoofdzakelijkvectoriëel gericht - d.w.z. dat ze specifiek gericht zijn naar de cytoplasmazijde © of naar demitochondriale zijde (M). Deze oriëntatie wordt vooral bepaald door de richting vanwaar de
enzymes hun substraat zullen ontvangen (glycerolfosfaat dehydrogenase: C zijde; succinaatdehydrogenase: M zijde). Andere daarentegen overspannen volledig de membraan(Cytochroom C oxidase).
1.Het mitochondrion is niet alleen de plaatsvan de Krebscyclus; oxidatieve fosforylatie, boxidatie, enz.; maar het speelt ook een zeerbelangrijke rol in de distributie van ionen (o.a.Ca2+) en van intermediairen tussen hetcytoplasma en het mitochondrium. Deze
distributie is in vele gevallen vanregulatorische betekenis.
3.3Electrontransportketen
3.3.1ElectroncarriersDe oxidatie van het substraat en het daaraangekoppelde electronentransport kan globaal
gezien onderverdeeld worden in vierverschillende deelreacties:
1. Substraat dehydrogenasen2. NAD dehydrogenase komplex (I)3. Ubiquinon dehydrogenase komplex (II)4. Cytochroom C oxidase komplex (III)
3.3.1.1 Substraat dehydrogenasenDit zijn die enzymesystemen die de verschillendesubstraten rechtstreeks kunnen oxideren. Ze bezittenmeestal NAD of FAD als cofactor. We vinden ze zowel inde mitochondria als in het cytoplasma.
3.3.1.1.1 NAD afhankelijke substraatdehydrogenasen
De structuur van NAD of nicotinamide adenosinedinucleotide kan je terugvinden in het hoofdstuk overvitaminen. Tijdens de oxidatie wordt aan het substraateen hydride ion (H-) onttrokken. De reductie van de ringveroorzaakt een zeer specifieke verandering van hetabsorptiespectrum die de basis is van tal van
enzymatische bepalingsmethoden. Het NAD is nooitcovalent gebonden aan de polypeptideketen maar eerderlos geassociëerd via waterstofbruggen. Het kan
Distributie van enzymen in ratlever mitochondria

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 22/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 22
gemakkelijk dissociëren van het enzyme.
NAD en NADH komen zowel voor in het cytoplasma als in de mitochondriale matrix.Tussen beide celkompartimenten bestaat er echter geen uitwisseling aangezien NAD+ eensterk polair molecule is waarvoor
de binnenste mitochondriale membraan niet permeabel is. Dit betekent dat er specialetransportsystemen moeten bestaan om reductieëquivalenten van het cytoplasma naar hetmitochondrium over te brengen.
NAD afhankelijke substraatdehydrogenasen zullen aangewend worden daar waar hetsubstraat een sterk negatieve standaard oxidatie-reductiepotentiaal Eo’ heeft (< -200 mV).Voorbeelden van dehydrogenasen met hun specifieke localisatie treft u in de tabel.
3.3.1.1.2 Flavine afhankelijke substraatdehydrogenasen
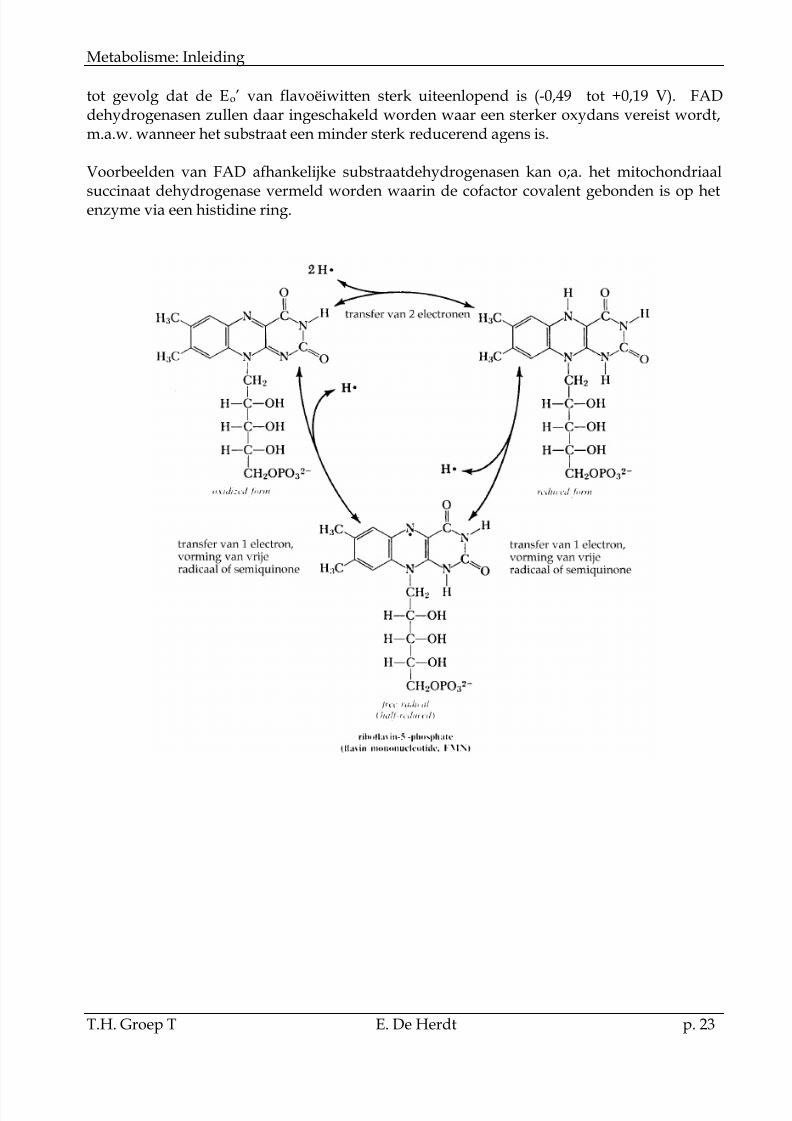
De structuur van de coënzymen FMN (flavine mononucleotide) en FAD (flavine adeninedinucleotide) wordt gegeven in het hoofdstuk over vitaminen. Bij reductie van het flavinecoënzyme kan zowel één als twee electronen getransfereerd worden. Wanneer één electrongetransfereerd wordt, ontstaat een vrij radicaal of semiquinone.
De flavine coënzymen zijn veel steviger gebonden dan NAD en komen tijdens de oxidatie -reductie cyclus niet los van het enzyme. De bindingssterkte van de cofactor aan hrschillendin de geoxideerde of gereduceerde vorm en karakteristiek voor alle enzymen. Dit heeft o.a.
5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 23/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 23
tot gevolg dat de Eo’ van flavoëiwitten sterk uiteenlopend is (-0,49 tot +0,19 V). FADdehydrogenasen zullen daar ingeschakeld worden waar een sterker oxydans vereist wordt,m.a.w. wanneer het substraat een minder sterk reducerend agens is.
Voorbeelden van FAD afhankelijke substraatdehydrogenasen kan o;a. het mitochondriaal
succinaat dehydrogenase vermeld worden waarin de cofactor covalent gebonden is op hetenzyme via een histidine ring.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 24/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 24
Het substraat dehydrogenase transfereert de electronen uiteindelijk naar ubiquinone (CoQ).Wegens het verschil in Eo’ tussen NAD en FAD afhankelijke dehydrogenasen is er ook eenbeduidend verschil in G van de reacties. Zo kan bij de transfer van een reductieëquivalentvan het substraat naar ubiquinone via een NAD afhankelijk substraat dehydrogenase éénATP gevormd worden, waar ditvia een FAD afhankelijksubstraatdehydrogenase nietmogelijk is.
3.3.1.2 NADH dehydrogenasekomplex (I)De oxidatie van NADH totNAD+ door ubiquinone wordtuitgevoerd door een NADHdehydrogenase. Dit is eencomplex enzymesysteem van de
mitochondriale membraanbestaande uit verschillendeeiwitten waarvan ieder één deelvan de totale reactie uitvoert.Het complex bevat:
3.3.1.2.1Flavo eiwit
Dit eiwit bevat FMN als coënzyme en wordt rechtstreeks door NADH gereduceerd langs deM zijde van de binnenste mitochondriale membraan.3.3.1.2.2IJzer - zwavel eiwitten
Ijzer - zwavel eiwitten bevatten Fe-atomen als chelaten gebonden aan S-atomen van cysteïne.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 25/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 25
Dank zij de Fe-atomen kunnen ze als electron carriers fungeren. Slechts één electron wordtgetransfereerd (Fe2+......Fe3+). De electron affiniteit van het Fe-S centrum wordt inhoofdzaak bepaald door de natuur van de eiwitketen.
Het
reductieëquivalentwordt van NADHdoor het NADdehydrogenasecomplexgetransfereerd naarubiquinone (CoQ).Ubiquinone kan ookrechtstreeksgereduceerd worden
door een FADafhankelijk substraatdehydrogenase.Ubiquinonenbevatten eenquinone ring en eensysteem bestaandeuit een variabel aantal isopreen eenheden.Ubiquinone kan gereduceerd worden door één electron waarbij het vrij radicaal(ubismiquinone) gevormd wordt, of door twee electronen tot ubiquinol. Ubiquinone komt
voor in het lipide gedeelte van de binnenste mitochondriale membraan en is hierin vrijbeweegbaar.
3.3.1.3. Ubiquinol dehydrogenase komplex (II)Het reductieëquivalent wordt van ubiquinol naar Cyt C getransfereerd door middel van hetubiquinol dehydrogenase. Dit is een sterk membraangebonden enzymecomplex dat tweetypes Cytb, minimum één Fe-S eiwit en cytochroom c1 als carrier bevat.
5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 26/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 26
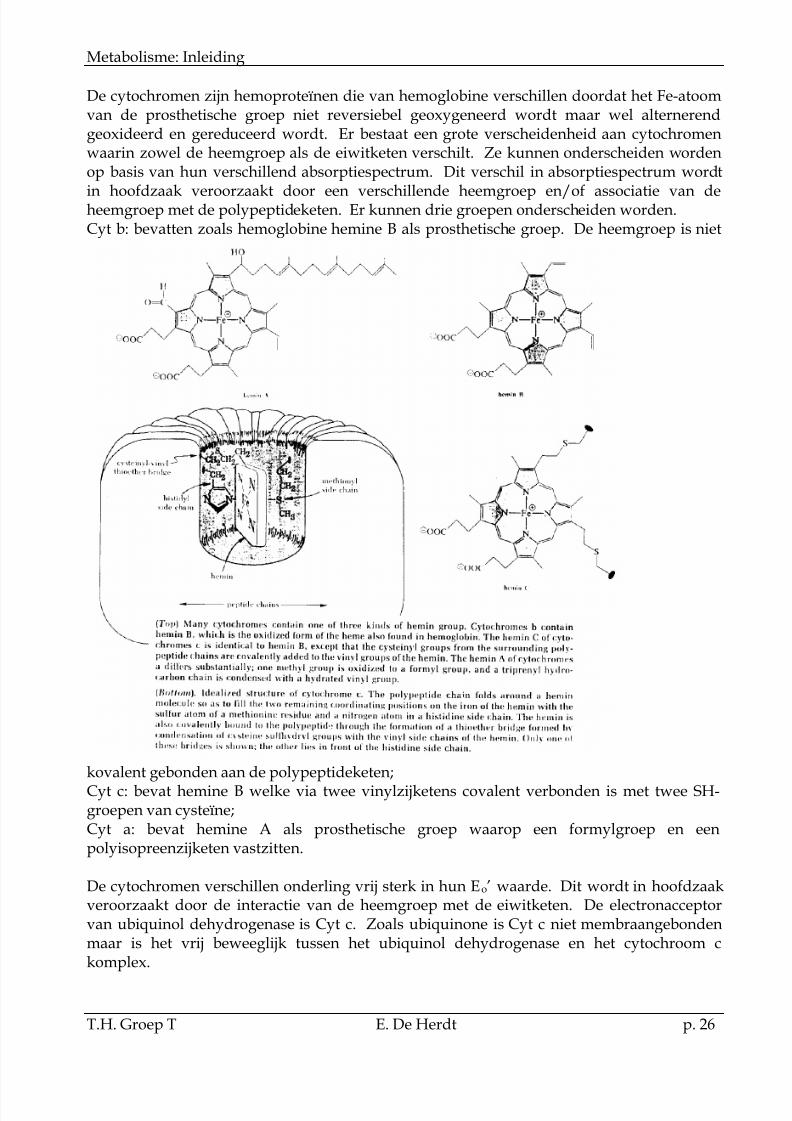
De cytochromen zijn hemoproteïnen die van hemoglobine verschillen doordat het Fe-atoomvan de prosthetische groep niet reversiebel geoxygeneerd wordt maar wel alternerendgeoxideerd en gereduceerd wordt. Er bestaat een grote verscheidenheid aan cytochromenwaarin zowel de heemgroep als de eiwitketen verschilt. Ze kunnen onderscheiden wordenop basis van hun verschillend absorptiespectrum. Dit verschil in absorptiespectrum wordt
in hoofdzaak veroorzaakt door een verschillende heemgroep en/of associatie van deheemgroep met de polypeptideketen. Er kunnen drie groepen onderscheiden worden.Cyt b: bevatten zoals hemoglobine hemine B als prosthetische groep. De heemgroep is niet
kovalent gebonden aan de polypeptideketen;Cyt c: bevat hemine B welke via twee vinylzijketens covalent verbonden is met twee SH-groepen van cysteïne;Cyt a: bevat hemine A als prosthetische groep waarop een formylgroep en eenpolyisopreenzijketen vastzitten.
De cytochromen verschillen onderling vrij sterk in hun Eo’ waarde. Dit wordt in hoofdzaakveroorzaakt door de interactie van de heemgroep met de eiwitketen. De electronacceptorvan ubiquinol dehydrogenase is Cyt c. Zoals ubiquinone is Cyt c niet membraangebondenmaar is het vrij beweeglijk tussen het ubiquinol dehydrogenase en het cytochroom ckomplex.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 27/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 27
3.3.1.4. Cytochroom C oxidase komplex (III)Cytochroom c oxidase bestaat uit een hexameer komplex gevormd door cyt a, een Cu2+ atoom en cyt a3. De electronen worden getransfereerd van cyt c naar moleculaire zuurstof.Cytochroom c oxidase vormt zoals hemoglobine irreversiebele complexen met CO en Cu+
waardoor de electronenoverdracht onmogelijk wordt.
3.3.2. Het flowschema van de electronenstroom
3.3.2.1. Relatie: oxidatie - reductiepotentiaal en de stromingsrichting De overdracht van electronen gebeurt van NADH naar moleculaire zuurstof:
NADH + H+ NAD+ + 2H+ + 2 e- Eo’ = (-)- 0,32 V
1/2 O2 + 2H+ + 2 e- H2O Eo’ = + 0,82 V
NADH + H+ + 1/2 O2 H2O + NAD+ Eo’ = + 1,14 V
Hieruit volgt dat: G°’ = - 52,6 kcal/mol
De electronen gaan dus van het meest electronegatieve naar het meest electropositievesysteem. De Eo’ van de verschillende electroncoarriers is tussen deze twee uitersten (-0,32 Ven +0,81 V) gelegen. De Eo’ van de verschillende carriers is uitgezet in de volgende figuur.
Hieruit blijkt duidellijk dat de drie complexen:
- NAD dehydrogenase komplex (I)- Ubiquinon dehydrogenase komplex (II)- Cytochroom C oxidase komplex (III)
drie overlappende groepen vormen. Dit betekent dat in elk complex steeds één carrieraanwezig is die een Eo’ heeft overeenkomstig met die van de electrondonor en één carriermet een Eo’ overeenkomstig met die van de electronacceptor.
De Eo’ in elk complex bedraagt respectievelijk 0,33; 0,31 en 0,32 V wat overeenkomt met eenG van respectievelijk 15,22; 14,3 en 14,76 kcal/mol. dit is ruim voldoende voor de vormingvan een energierijke fosfaatbinding (7,3 kcal/mol).

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 28/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 28
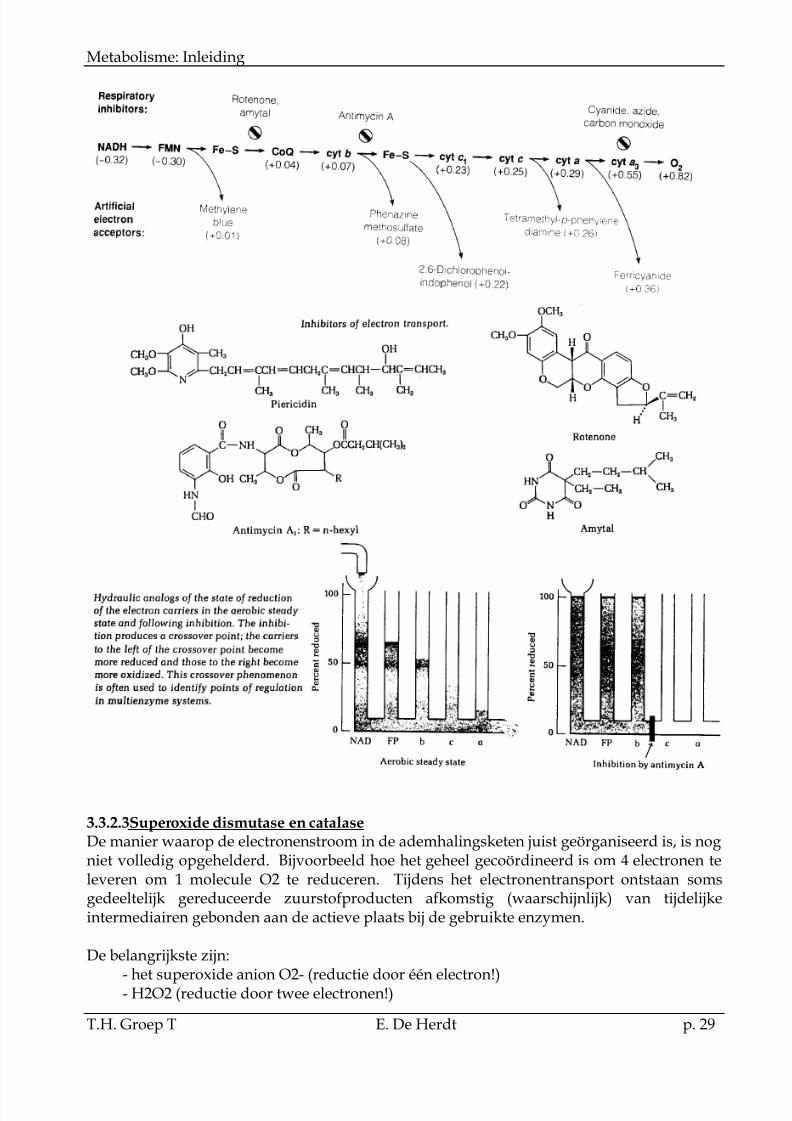
3.3.2.2Inhibitoren van het electronentransport
Er zijn verschillende inhibitoren van het electronentransport gekend. Deze hebbenbijgedragen in de opheldering van de volgorde van de gebruikte carriers. Dit wordtaangetoond in het hydraulische model.
5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 29/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 29
3.3.2.3Superoxide dismutase en catalaseDe manier waarop de electronenstroom in de ademhalingsketen juist geörganiseerd is, is nogniet volledig opgehelderd. Bijvoorbeeld hoe het geheel gecoördineerd is om 4 electronen televeren om 1 molecule O2 te reduceren. Tijdens het electronentransport ontstaan somsgedeeltelijk gereduceerde zuurstofproducten afkomstig (waarschijnlijk) van tijdelijkeintermediairen gebonden aan de actieve plaats bij de gebruikte enzymen.
De belangrijkste zijn:- het superoxide anion O2- (reductie door één electron!)- H2O2 (reductie door twee electronen!)

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 30/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 30
Ze zijn enorm reactief en schadelijk voor heel wat biomoleculen. Aërobe cellen bevattenechter het enzyme syperoxide dismutase dat de volgende reactie catalyseert:
2O2- + 2H+ H2O2 + O2
Het enzyme komt in vrij hoge concentratie voor en is buitengewoon actief wat doetvermoeden dat superoxide radicalen continu geproduceerd worden door verschillendeenzymen.
Het gevormde H2O2 wordt ontbonden door het enzyme catalase:
H2O2 H2O + 1/2 O2
Dit enzyme komt voornamelijk voor in de peroxysomen, kleine celorganellen.
3.4Oxidatieve fosforylatie
3.4.1Koppeling van de exergonische en endergonische reactie
De transfer van een electronenpaar naar moleculaire zuurstof is een sterk exergonischereactie zoals blijkt uit zijn sterk negatieve G°’. Een zeker percentage van deze energie kanbenut worden o.a. voor de vorming van energierijke fosfaatbindingen.
NADH + H+ + 1/2 O2 H2O + NAD+ G°’ = - 52,6 kcal/mol
ADP + Pi ATP + H2O G°’ = - 7,3 kcal/mol
De hoeveelheid anorganisch fosfaat vastgelegd in ATP per zuurstofatoom (P/O ratio)bedraagt 3. Dit betekent dat per electronenpaar 3 ATP kunnen gevormd worden:
3 x 7,3 kcal = 21,9 kcal of 42%
en dit respectievelijk bij de doorgang van de drie complexen:
- NAD dehydrogenase komplex (I)- Ubiquinon dehydrogenase komplex (II)- Cytochroom C oxidase komplex (III)
De volledige reactievergelijking van het fosforylerend electronentransport van NADH naarzuurstof is dus:
NADH + H+ + 1/2 O2 + 3 Pi + 3 ADP 4 H2O + 3 ATP + NAD+
Indien succinaat wordt gedehydrogeneerd door het flavine gebonden succinaatdehydrogenase worden er maar 2 ATP’s gevormd per electronenpaar aangezien deze

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 31/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 31
electronen maar ter hoogte van ubiquinone in de electrontransportketen terechtkomen! (zieFAD substraat dehydrogenasen!!!)
3.4.1.1Het ATP synthetiserende enzyme
De binnenste mitochondriale membraan bevat een paddestoelachtig enzymatisch complex,nml. het ATP synthetase of FoF1 ATPase. Het bestaat uit twee hoofdcomponenten Fo en F1.F1 lijkt op een deurknop welke via een staafje verbonden is met de matrix van de binnenstemembraan waarin Fo ligt.
De index o betekent dat het dat deel van het ATPase is dat bindt met het giftig antibioticumoligomycine, een sterke inhibitor van dit enzyme. F1 alleen kan geen ATP synthetiseren uitADP en Pi maar wel de hydrolyse van ATP tot ADP en Pi en wordt daarom wel een ATPase(F1) genoemd.
Deze ATPase activiteit wordt verondersteld het inverse te zijn van de normale functie van F1in intacte mitochondriën.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 32/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 32
3.4.1.2Chemiosmotische koppelingshypothese
3.4.1.2.1Hypothese electrochemische potentiaal
Deze is gebaseerd op een aantal experimentele gegevens:
Om gekoppelde oxidatieve fosforylatie mogelijk te maken dient de binnenstemitochondriale membraan intact te zijn.
Electrontransport veroorzaakt de verwijdering van H+ van de matrix naar deintermembraanruimte zodanig dat er een pH verschil ontstaat van ongeveer 1,5 pHeenheden.
Parallel met het electrontransport kunnen Ca2+ ionen actief vanuit deintermembraanruimte naar de matrix getransporteerd worden tegen de concentratiegradiënt
van Ca2+
in.
Beide ionenpompen worden “aangedreven” door het electrontransport en blijvendoorgaan zelfs wanneer de ATP productie experimenteel geblokkeerd wordt. Dit kanbijvoorbeeld gebeuren door reagentia zoals 2,4 dinitrofenol. Hierbij zal de vrije energiewelke door het electrontransport wordt gegenereerd vrijgesteld worden onder de vorm vanwarmte i.p.v. onder ATP productie. Deze reagentia verhogen in grote mate depermeabiliteit van de binnenste membraan voor H+ ionen.
Wanneer in het mitochondrion experimenteel een H+ gradiënt opgebouwd wordt in
afwezigheid van electrontransport, dan kan deze gradiënt de vorming van ATP aandrijven.
De hypothese samengevat in de volgende figuurstelt dat de binnenste mitochondriale membraaneen eiwit/ionen pomp bevat die aangedrevenwordt door het electrontransport en dat deprotonen vanuit de matrix naar deintermembraanruimte getransporteerd worden.
Dit veronderstelt dat de binnenste membraanimpermeabel is voor protonen. Carrier B wordtgereduceerd en verkrijgt 2 H+. Deze kunnen ofwelvan de gereduceerde carrier AH2 komen ofwel uitde matrix. Wanneer BH2 geoxideerd wordt doorC worden H+ vrijgesteld in deintermembraanruimte. Dit veronderstelt hetvectoriëel gericht zijn van de carriers in de wand.
Het transport van protonen doorheen demembraan gaat al dan niet gepaard met hettransport van tegenionen X-. In het geval van transport van tegenionen zal er een

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 33/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 33
concentratiegradiënt van H+ ontstaan met als gevolg een daling van de pH aan de C-zijde.Wanneer geen tegenionen getransporteerd worden dan zullen de H+ zich ophopen terhoogte van de membraan met als gevolg het ontstaan van een membraanpotentiaal. Deconcentratiegradiënt en/of de membraanpotentiaal vormt een hoog-energetische toestand.
De energie vastgelegd in dit electrochemisch potentiaalverschil wordt gegeven door:G = 5,708 pH + FΦ
waarin:pH = de concentratiegradiënt van H+F = constante van Faraday
Φ = het potentiaalverschil over de membraan.De aldus vastgelegde “energie” kan dan benut worden voor de vorming van energierijkefosfaatbindingen in ATP of voor membraantransport.
3.4.1.2.2Vorming van ATP
Per gevormde ATP zouden twee protonen teruggetransporteerd worden naar de matrix. Ditbetekent dat er drie protonenpompen dienen te zijn. Waar deze voorkomen en welke juistde opeenvolging van de carriers is, is niet duidelijk, maar een aanvaardbare voorstellingwordt gegeven in de volgende figuur.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 34/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 34
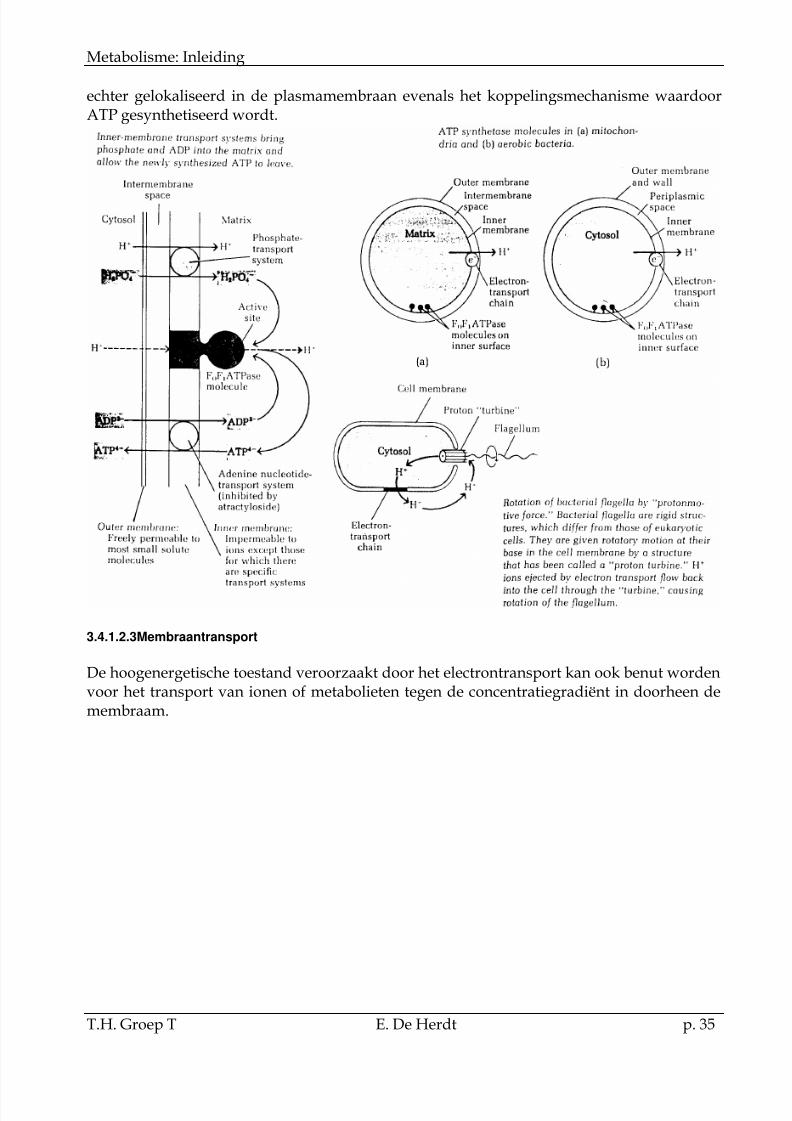
De productie van ATPgebeurt ter hoogte vanhet ATP synthetase. Zekan voorgesteld wordenals een vectoriëleafsplitsing van H2O vanPi en ADP.
Bacteriën enchloroplasten bevatten
ook protonentransfersystemen aangedreven door electronentransport. De dehydrogenasen zijngelokaliseerd in het bacteriëel cytosol, de electroncarriers van de ademhalingsketen zijn
5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 35/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 35
echter gelokaliseerd in de plasmamembraan evenals het koppelingsmechanisme waardoorATP gesynthetiseerd wordt.
3.4.1.2.3Membraantransport
De hoogenergetische toestand veroorzaakt door het electrontransport kan ook benut wordenvoor het transport van ionen of metabolieten tegen de concentratiegradiënt in doorheen demembraam.

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 36/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 36
Zo wordtATP gevormdin hetmitochondrionen
hoofdzakelijkverbruikt in hetcytoplasma. Ditvereist eenconstanteuitwisselingvan ADP en Pi tegen ATP. Ditgebeurt doortwee
afzonderlijkeantiportcarriers. DePi/OH- carrierwordtaangedrevendoor de protonengradiënt en behoudt electrische neutraliteit terwijl de ADP/ATP carrieraangedreven wordt door de membraanpotentiaal.
Ook andere metabolieten zoals dicarboxylzuren en aminozuren kunnen via carriers
doorheen de binnenste mitochondriale wand getransporteerd worden. Deze carriers kunnenook zonder gekoppeld te zijn aan het electronentransport functioneren maar werken dansteeds in de richting van de concentratiegradiënt (= passieve diffusie).
Het membraantransport is van zeer groot belang om reductieëquivalenten, ontstaan in hetcytoplasma door de oxidatie van substraten over te brengen naar het mitochondrion.Aangezien de binnenste membraan impermeabel is voor NAD+/NADH dient hetreductieëquivalent onrechtstreeks getransporteerd te worden via een reversiebel oxideerbaar- reduceerbaar metaboliet (= shuttle). De belangrijkste hiertoe zijn de glycerolfosfaat shuttle en de malaat-aspartaat shuttle.
In de glycerolfosfaat shuttle wordtdihydroxyacetonfosfaat gereduceerddoor het NADH afhankelijkeglycerol3P dehydrogenase totglycerol3P. Dit diffundeert doorheende buitenste mitochondriale membraanen wordt geoxideerd door het FADafhanelijk glycerol3P dehydrogenaseop het oppervlak van de binnenste
membraan. Het reductieëquivalentwordt dan verder doorgegeven aanubiquinone terwijl het

5/16/2018 01 metabolisme inleiding - slidepdf.com
http://slidepdf.com/reader/full/01-metabolisme-inleiding 37/37
Metabolisme: Inleiding
T.H. Groep T E. De Herdt p. 37
dihydroxyacetonfosfaat naar het cytoplasma terugkeert. Het reductieëquivalent afkomstigvan NADH komt dus in de electrontransportketen na complex I en kan dus slechts 2 ATP’sgeven.
De glycerolfosfaat shuttle is unidirectioneel, wat essentiëel is om een hoge NAD+/NADH
verhouding, nodig voor de oxidatie van het substraat in het cytoplasma, te behouden t.o.v.een lage NAD+/NADH verhouding in het mitochondrion.
In de malaat-aspartaat shuttle wordt cytoplasmatisch oxaalacetaat gereduceerd door eenspecifiek NADH afhankelijk malaat dehydrogenase tot malaat dat via een dicarboxylaatcarrier uitgewisseld wordt voor mitochondriaal alfa ketoglutaarzuur. In het mitochondrionwordt malaat geoxideerd door een mitochondriaal NAD+ afhankelijk malaat dehydrogenasedat het reductieëquivalent in de electrontransportketen brengt.
De mitochondriale membraan is vrijwel impermeabel voor oxaalacetaat zodat dit via een
transaminase omgezet wordt tot aspartaat dat via de Asp/Glu carrier naar het cytoplasmagetransporteerd wordt waar het opnieuw getransamineerd wordt tot oxaalacetaat.
De shuttle is in principe reversiebel, maar wordt irreversiebel door het koppelen van hettransport aan het electrontransport.