
T E S I S · 2016-10-27 · MARGARITA ELIZABETH RICO LEMUS DIRECTOR DE TESIS DR. BENJAMÍN...
128
CENTRO DE INVESTIGACIÓN Y ASISTENCIA EN TECNOLOGÍA Y DISEÑO DEL ESTADO DE JALISCO, A.C. Efecto del nitroprusiato de sodio en la regeneración in vitro de un híbrido del género Polianthes T E S I S PARA OBTENER EL GRADO ACADÉMICO DE MAESTRO EN CIENCIAS DE LA FLORICULTURA PRESENTA MARGARITA ELIZABETH RICO LEMUS DIRECTOR DE TESIS DR. BENJAMÍN RODRÍGUEZ GARAY Guadalajara, Jalisco, Febrero del 2016
Transcript of T E S I S · 2016-10-27 · MARGARITA ELIZABETH RICO LEMUS DIRECTOR DE TESIS DR. BENJAMÍN...

CENTRO DE INVESTIGACIÓN Y ASISTENCIA EN
TECNOLOGÍA Y DISEÑO DEL ESTADO DE JALISCO, A.C.
Efecto del nitroprusiato de sodio en la
regeneración in vitro de un híbrido del género
Polianthes
T E S I S
PARA OBTENER EL GRADO ACADÉMICO DE
MAESTRO EN CIENCIAS DE LA FLORICULTURA
PRESENTA
MARGARITA ELIZABETH RICO LEMUS
DIRECTOR DE TESIS
DR. BENJAMÍN RODRÍGUEZ GARAY
Guadalajara, Jalisco, Febrero del 2016

DECLARACIÓN DE PROPIEDAD
Declaro que la información contenida en la sección de materiales y métodos
experimentales, los resultados y discusión de este punto proviene de las
actividades de experimentación realizadas durante el periodo que se me asignó
para desarrollar mi trabajo de tesis, en las unidades y laboratorios del Centro de
Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco, A.C., y
que a razón de lo anterior y en contraprestación de los servicios educativos o de
apoyo que me fueron brindados, dicha información, en términos de la Ley Federal
del Derecho de Autor y la Ley de la Propiedad Industrial, le pertenecen
patrimonialmente a dicho Centro de Investigación. Por otra parte, en virtud de lo ya
manifestado, reconozco que de igual manera los productos intelectuales o
desarrollados pertenecen patrimonialmente al Centro de Investigación y Asistencia
en Tecnología y Diseño del Estado de Jalisco, A.C., y en el mismo tenor,
reconozco que si derivasen de este trabajo productos intelectuales o desarrollos
tecnológicos, en lo especial, estos se regirán, en todo caso por lo dispuesto por la
Ley Federal del Derecho de Autor y la Ley de la Propiedad Industrial, en el tenor
de lo expuesto en la presente declaración.
____________________________
MARGARITA ELIZABETH RICO LEMUS

ÍNDICE GENERAL
Pág.
I. INTRODUCCIÓN 1
II. ANTECEDENTES 2
2.1 Origen e Importancia del género Polianthes 2
2.2 Características morfológicas de P. tuberosa, especies silvestres y
sus híbridos
4
2.3 Reproducción de Polianthes tuberosa y especies silvestres 6
2.4 Propagación in vitro del género Polianthes L. 7
2.5 Embriogénesis somática 12
2.5.1 Picloram y 2,4-D en la inducción de embriogénesis 19
2.6 Óxido nítrico 21
2.6.1 Definición y propiedades fisicoquímicas del óxido nítrico 21
2.6.2 Función del óxido nítrico en las células vegetales 22
2.6.2.1 Rol del óxido nítrico en la síntesis de clorofila 23
2.6.3 Biosíntesis de óxido nítrico en las plantas 24
2.6.4 Agentes químicos donadores de óxido nítrico 27
2.6.5 Complejos de transición metal-óxido nítrico 27
2.6.5.1 Nitroprusiato o nitroprusida de sodio (NPS) 28
2.6.5.2 NPS en el cultivo in vitro 28
III. JUSTIFICACIÓN 31
IV. HIPÓTESIS 32

ÍNDICE GENERAL
Pág.
V. OBJETIVOS 33
5.1 Objetivo general 33
5.2 Objetivos específicos 33
VI. METODOLOGÍA 34
6.1 Embriogénesis somática 36
6.1.1 Germinación de semillas 36
6.1.2 Desdiferenciación de tejido somático de plántulas del híbrido 37
6.1.3 Doble tinción diferencial de células 39
6.1.4 Subcultivo de callo potencialmente embriogénico 39
6.1.4.1 Subcultivo utilizando medios de inducción 39
6.1.4.2 Subcultivo utilizando medio de inducción enriquecido
con aminoácidos
40
6.1.5 Subcultivo a medio de expresión 41
6.2 Micropropagación 41
6.2.1 Descripción inicial de plántulas 41
6.2.2 Estimulación de la producción de brotes en plántulas 44
6.2.3 Establecimiento de yemas axilares de la inflorescencia del
híbrido 1008157
46
6.2.4 Propagación de clones del híbrido 1008157 47
6.2.5 Estimulación de la producción de brotes en clones del
híbrido 1008157
47
6.2.6 Extracción y cuantificación de clorofila 50
6.3 Preparación de medios de cultivo con NPS 51

ÍNDICE GENERAL
Pág.
6.4 Análisis estadístico 52
6.5 Enraizado y aclimatación de explantes clones del híbrido
1008157 52
VII. RESULTADOS Y DISCUSIÓN 54
7.1 Embriogénesis somática 54
7.1.2 Desdiferenciación de tejido somático de plántulas del híbrido 54
7.1.3 Doble tinción diferencial de células 60
7.1.4 Subcultivo de callo potencialmente embriogénico 62
7.1.4.1 Subcultivo utilizando medios de inducción 62
7.1.4.2 Subcultivo utilizando medio de inducción enriquecido
con aminoácidos
68
7.1.5 Subcultivo a medio de expresión 68
7.2 Micropropagación 70
7.2.1 Descripción inicial de plántulas 70
7.2.2 Estimulación de la producción de brotes en plántulas 70
7.2.3 Establecimiento de yemas axilares de la inflorescencia del
híbrido 1008157
81
7.2.4 Propagación de clones del híbrido 1008157 82
7.2.5 Estimulación de la producción de brotes en clones del
híbrido 1008157
84
7.2.5.1 Clones del híbrido con al menos un brote antes del
cultivo en NPS
85

ÍNDICE GENERAL
Pág.
7.2.5.1 Clones del híbrido con al menos un brote antes del
cultivo en NPS
87
7.2.6 Extracción y cuantificación de clorofila 90
7.2.5 Enraizado y aclimatación de explantes clones del híbrido
1008157
91
IX. CONCLUSIONES 92
Xl. BIBLIOGRAFÍA 94

ÍNDICE DE FIGURAS
No. Descripción Pág.
1 Distribución del género Polianthes en México 2
2 Morfología de P. tuberosa var. doble 4
3 Morfología de la especie silvestre P. howardii Verh-Will 5
4 Morfología del híbrido 1008157 6
5 Principales técnicas para la propagación in vitro 9
6 Embriogénesis cigótica en angiospermas y gimnospermas 14
7 Eventos característicos de la embriogénesis somática 16
8 Rutas enzimáticas y no enzimáticas para la producción de óxido
nítrico en las plantas
25
9 Diagrama de los principales grupos de compuestos donadores de
óxido nítrico (NO)
27
10 Principales compuestos del grupo complejos de transición metal-
óxido nítrico
28
11 Diagrama general de la metodología experimental 35
12 Proceso de germinación de las semillas 37
13 Siembra de los segmentos de hoja en el medio de cultivo 38
14 Diseño factorial 24 para evaluar el efecto de picloram y el
nitroprusiato de sodio (NPS) sobre la desdiferenciación de tejido
vegetal de plántulas del híbrido 1008157
38
15 Plántulas resembradas en medio con tidiazuron después de 30 d de
cultivo
42
16 Proceso de establecimiento de las yemas axilares de la
inflorescencia del híbrido 1008157
46
17 Diseño experimental unifactorial utilizado en la evaluación de
multiplicación de brotes adicionando NPS (0, 10, 20, 30 y 40 M)
50
18 Proceso de aclimatación de los explantes clones del híbrido 1008157 54
19 Similitud con los embriones somáticos formados a partir de
secciones tallo en Agave fourcroydes
56

ÍNDICE DE FIGURAS
No. Descripción Pág. 20 Comparación de medias para la respuesta en los 16 tratamientos del
diseño factorial entre NPS y picloram
57
21 Desdiferenciación de tejido a partir de segmentos de hoja de
plántulas provenientes de semillas de un híbrido del género
Polianthes después de 30 d de cultivo
61
22 Comparación de medias para la presencia de estructuras globulares
en los 16 tratamientos del diseño factorial entre NPS y picloram
61
23 Tipos de respuesta en la inducción de la embriogénesis somática a
partir de segmentos de hoja de plántulas de un híbrido del género
Polianthes
64
24 Estructuras globulares sometidas a la doble tinción diferencial 66
25 Comparación de la respuesta de dos genotipos diferentes del
tratamiento con 2.07 y 10 M de picloram y NPS respectivamente,
en medios de cultivo para inducción MIA y MIP
69
26 Comparación de la respuesta de dos genotipos diferentes del
tratamiento con 2.07 y 20 M de picloram y NPS respectivamente,
en medios de cultivo para inducción MIA y MIP
70
27 Respuesta de tres genotipos diferentes del tratamiento con 2.07 y 30
M de picloram y NPS respectivamente, en medio de cultivo para
inducción MIA
71
28 Comparación de la respuesta de dos genotipos diferentes del
tratamiento con 3.1 y 10 M de picloram y NPS respectivamente, en
medios de cultivo para inducción MIA y MIP
71
29 Comparación de la respuesta de tres genotipos diferentes del
tratamiento con 3.1 y 30 M de picloram y NPS respectivamente, en
medios de cultivo para inducción MIA y MIP
72
30 Comparación de la respuesta de los 4 genotipos diferentes del
tratamiento con 4.54 y 0 M de picloram y NPS respectivamente, en
medios de cultivo para inducción MIA y MIP
73

ÍNDICE DE FIGURAS
No. Descripción Pág.
31 Comparación de la respuesta de los 4 genotipos diferentes del
tratamiento con 4.54 y 20 M de picloram y NPS respectivamente,
en medios de cultivo para inducción MIA y MIP
74
32 Respuesta de los genotipos a la resiembra en medios de inducción
enriquecido con aminoácidos
76
33 Plantas control con diferencias fenotípicas en su morfología después
de 60 d en medio de cultivo sin NPS
79
34 Efecto del NPS sobre el desarrollo de las plantas durante el primer
cultivo
80
35 Porcentaje de plantas estimuladas para la producción de nuevos
brotes durante el primer cultivo
81
36 Efecto del NPS sobre el desarrollo de las plantas durante el segundo
cultivo
82
37 Porcentaje de plantas que fueron estimuladas hasta el segundo
cultivo para la producción de nuevos brotes
83
38 Porcentaje de plantas que fueron estimuladas al final del
experimento para la producción de nuevos brotes
84
39 Plántulas con mayor número de brotes en cada tratamiento 84
40 Plantas cultivadas en medio con 10 M de NPS después de 60 d de
cultivo
85
41 Plantas cultivadas en medio con 20 M de NPS después de 60 d de
cultivo
86
42 Plantas cultivadas en medio con 30 M de NPS después de 60 d de
cultivo
87
43 Plantas cultivadas en medio con 40 M de NPS después de 60 d de
cultivo
88
44 Establecimiento de yemas axilares a partir de la inflorescencia del
hibrido 1008157
90

ÍNDICE DE FIGURAS
No. Descripción Pág.
45 Propagación de brotes vegetativos utilizando la multiplicación por
yemas axilares
91
46 Explantes clones del híbrido después de incubarlos en oscuridad 92
47 Marchitamiento y muerte del tejido del ápice de las hojas adultas
durante el cultivo con NPS
94
48 Efecto del NPS sobre las características de desarrollo de los
explantes clones del híbrido con al menos un brote
95
49 Efecto del NPS sobre las características morfológicas de los
explantes clones del híbrido sin brotes al inicio del cultivo
97
50 Prueba de rangos múltiples (LSD) para contenido de clorofila 99
51 Comparación del contenido de clorofila entre los tratamientos con
NPS para los clones del híbrido con o sin brotes durante el cultivo
100
52 Plantas del híbrido 1008157 aclimatadas 101

ÍNDICE DE TABLAS
No. Descripción Pág.
1 Producción de P. tuberosa L. (gruesa) en México 3
2 Propagación in vitro del género Polianthes 10
3 Embriogénesis somática en el género Agave y otros 18
4 El uso de NPS en combinación con reguladores de crecimiento en la
propagación in vitro
30
5 Descripción inicial de las 75 plántulas del híbrido 1008157 42
6 Diseño experimental unifactorial utilizado para evaluar el efecto de la
concentración de NPS sobre la producción de brotes de plántulas del
híbrido 1008157
45
7 Propagación de plantas del híbrido 1008157 47
8 Descripción inicial de los explantes del híbrido con presencia de al
menos un brote
48
9 Descripción inicial de los explantes del híbrido sin brotes 49
10 Ecuaciones para la determinación clorofila 51
11 Efecto del NPS y picloram en la inducción de embriogénesis
somática utilizando tejido somático de plántulas de un híbrido del
género Polianthes
58
12 Comparación de las respuestas de desarrollo de las plantas durante
el cultivo en medio sin NPS
75
13 Comparación de las respuestas de desarrollo de las plantas durante
el cultivo en medio con 10 M NPS
81
14 Comparación de las respuestas de desarrollo de las plantas durante
el cultivo en medio con 20 M NPS
82
15 Comparación de las respuestas de desarrollo de las plantas durante
el cultivo en medio con 30 M NPS
83
16 Comparación de las respuestas de desarrollo de las plantas durante
el cultivo en medio con 40 M NPS
84

ÍNDICE DE TABLAS
No. Descripción Pág.
17 Comparación entre las variables de respuesta para los clones del
híbrido con al menos un brote durante el cultivo en medio con NPS
89
18 Promedio y desviación estándar para las variables de respuesta en
clones del híbrido con presencia de brotes durante el cultivo en
medio con NPS
90
19 Comparación entre las variables de respuesta para los clones del
híbrido sin brotes durante el cultivo en medio con NPS
92
20 Promedio y desviación estándar para las variables de respuesta en
clones del híbrido con presencia de brotes durante el cultivo en
medio con NPS
93
21 Promedio para el contenido de clorofila a y b en explantes clones del
híbrido después del cultivo en medio con NPS
95

A G R A D EC I M I E N T O S
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca
otorgada para la posible realización de mis estudios de Maestría a través del
Centro de Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco
(CIATEJ) dentro de la Unidad de Biotecnología Vegetal.
A mi Director de tesis, el Dr. Benjamín Rodríguez Garay por todo el apoyo,
los consejos, las enseñanzas, el tiempo brindado y sobre todo, la paciencia que
mostró para el desarrollo del proyecto.
Al Maestro Manuel Rodríguez por el tiempo otorgado para el análisis de
resultados, la revisión de avances de tesis y seminarios, siempre mostrando una
actitud de aprecio y confianza para solucionar cualquier duda.
Al Dr. Ernesto Tapia por sus comentarios y correcciones en los seminarios y
el tiempo brindado para revisión de avances de tesis. Agradeciendo especialmente
su atenta disposición para la donación del material vegetal de trabajo.
Al Maestro Octavio García Depraect por su tiempo brindado para la
revisión y corrección del documento escrito, por su apoyo, sus consejos y el ánimo
de un buen amigo para culminar a tiempo la maestría.
A mis amigos y compañeros de laboratorio Alejandra Flores, Isabel Ibarra,
Daniela Rodríguez y Ángel Barranco por todo su apoyo en las etapas de trabajo
más pesadas, sin su apoyo hubiera sido más difícil lograrlo.
A mis amigos Magdalena, Samuel, Michelle, Felipe, Jaime Silva, Ismael,
Jesús, Diego, Luzmy, Bahena, Fredy, Anahi, Mary, Nelly por su apoyo y amistad
en los buenos momentos que pasamos juntos, momentos que hicieron mi estancia
en Guadalajara inolvidable y muy placentera.
A cada uno de los profesores de la Maestría en Floricultura, en especial a la
Doctora Antonia, directora de la unidad de Biotecnología Vegetal por su apoyo y
estimación.

D E D I C A T O R I A
A Dios
Por permitirme concluir satisfactoriamente una etapa más en mi vida, por
cuidarme y apoyarme en los momentos decisivos durante estos dos años.
A mi Padres
A mi Papá que me apoyó económicamente pero sobre todo dándome su
confianza y consejos para crecer como persona; a mi Mamá que fue la luz y fuerza
en los momentos más difíciles, que a pesar de la distancia siempre tenía una
palabra de aliento y/o consuelo para continuar. Por recibirme con tanto cariño
cuando estaba en casa y despedirme dándome ánimos y fortaleza a mi regreso.
A mis hermanos por su apoyo incondicional, por creer en mí y sentirse
orgullosos de mis logros.
Al Dr. José Luis Cabrera Ponce, por impulsarme en realizar una Maestría y
vincularme con el CIATEJ, sin su apoyo y consejos, esta gran meta no la hubiera
comenzado en cuanto salí de la universidad.

RESUMEN
El género Polianthes L. (Asparagaceae), es endémico de México y posee
gran variedad de especies silvestres distribuidas en gran parte del territorio
nacional. Este género es representado económica y culturalmente por la variedad
comercial P. tuberosa L. la cual en tiempos prehispánicos fue utilizada por los
aztecas como flor de corte y para usos medicinales. Actualmente, está siendo
utilizada en la industria farmacéutica y para uso ornamental. Se han realizado
cruzas entre P. tuberosa L. y especies silvestres para obtener híbridos con
características especiales por ejemplo el color de la flor, el aroma, el tamaño, etc.
que las hagan atractivas para un nicho de mercado. La propagación in vitro del
género Polianthes se ha realizado principalmente por la técnica de yemas axilares
utilizando reguladores de crecimiento como auxinas y citocininas. Escasamente la
regeneración in vitro de esta planta se ha estudiado y realizado por la técnica de
embriogénesis somática. En el presente estudio se realizó la regeneración de un
híbrido producto de la cruza de P. tuberosa L. y P. howardii mediante las técnicas
de yemas axilares y embriogénesis somática. Además de adicionar al medio de
cultivo reguladores de crecimiento, se utilizó diferentes concentraciones de
nitroprusiato de sodio (NPS), un agente químico donador de oxido nítrico, para
estimular la respuesta del tejido vegetal en cada una de las técnicas utilizadas.
Dentro de los resultados, únicamente se logró la regeneración del hibrido por la
técnica de yemas axilares. Sin embargo, la adición de NPS en las diferentes
concentraciones (10, 20, 30 Y 40 M) no aportó diferencia estadística para la
producción de brotes con respecto al control que contenía solo reguladores de
crecimiento (10 mg/L de BA y 0.025 mg/L de 2,4-D). Sin embargo, numéricamente,
la producción de nuevos brotes del híbrido sí fue mayor cuando el medio de cultivo
fue adicionado con 40 M de NPS. Debido a estos resultados se podría
incrementar la concentración de NPS en estudios posteriores del género
Polianthes.
Palabras clave: Polianthes, híbrido, regeneración, in vitro, nitroprusiato de sodio.

ABSTRACT
The genus Polianthes L. (Asparagaceae), is endemic to Mexico. It has a
high number of species distributed over a great part of Mexican territory. This
genus is represented economically and culturally by the commercial variety P.
tuberosa L. which in ancient times was used by the Aztecs as a cut flower and for
medicinal uses. Currently, it is being used in the pharmaceutical industry and for
ornamental uses. Crosses have been made between P. tuberosa L. and wild
species generating hybrids with special characteristics such as flower color, flavor,
size, etc. that make them attractive to a specific niche market. In vitro propagation
of genus Polianthes has been mainly carried out by axillary buds culture technique
using plant growth regulators such as auxins and cytokinins. Rarely in vitro
regeneration of this plant has been studied and applied by somatic embryogenesis
technique. In this study, the regeneration of a hybrid product obtained by cross of
P. tuberosa L. with P. howardii was performed using axillary buds culture and
somatic embryogenesis technique. Besides adding to the culture medium growth
regulators, different concentrations of sodium nitroprusside (SNP), a chemical
agent used as nitric oxide donor, to stimulate a response on plant tissue. Results
showed that only hybrid regeneration was achieved by using axillary buds
technique. On the other hand, the addition of NPS in different concentrations (10,
20, 30 and 40 uM) no provided significant statistical difference for shoot production
over the control, which contained only growth regulators (10 mg BA /L and 0.025
mg 2,4-D /L). However, numerically, the production of new shoots from hybrid was
higher when the culture medium was added with NPS (40 uM). Finally, these
results point out that the effect of NPS concentration on the genus Polianthes must
be studied deeply.
Keywords: Polianthes, hybrid, regeneration, in vitro, sodium nitroprusside.

I. INTRODUCCIÓN
En México los productos ornamentales han ganado terreno en cuanto a
exportaciones y al valor de la producción. En el año 2011 el valor de la floricultura
fue de 5,646 millones de pesos, además, la superficie destinada al cultivo de flores
y plantas en maceta fue de 18,629 ha. Los estados con mayor participación por el
valor de la producción de este cultivo destacaron Estado de México, Puebla,
Morelos, Distrito Federal, Baja California y Jalisco.
P. tuberosa L. se cultiva extensivamente dentro del territorio nacional debido
a sus flores intensamente aromáticas y su uso en la industria del perfume. La
forma más común y fácil de propagación es de manera asexual usando bulbos
tanto para la variedad comercial como para las especies silvestres sin embargo
esto tiene desventajas en tiempo, costo y enfermedades de las plantas. Por lo
tanto, es necesario implementar protocolos de cultivo in vitro debido a las ventajas
que esta técnica ofrece como son el número elevado de plantas en un espacio
reducido, la producción en cualquier época del año, el crecimiento más vigoroso y
rejuvenecimiento de la planta así, como plantas libres de enfermedades. De esta
manera, se podrían propagar especies silvestres o híbridos del género Polianthes
con características especiales para que resulten atractivas para un nuevo
mercado.
Recientemente se está implementado el uso de agentes químicos donadores
de óxido nítrico (NO) como potenciadores del crecimiento de las plantas. Tal es el
caso del nitroprusiato de sodio (NPS) un compuesto que libera NO al medio de
cultivo. El NPS ha sido evaluado en su mayoría en protocolos de propagación y en
estudios para la protección de las células contra daños por patógenos u otro tipo
de estrés. En el presente estudio se realizó una evaluación del efecto de NPS en
la regeneración in vitro de un híbrido del género Polianthes. Se evaluó la
respuesta del tejido somático de plántulas a la inducción de embriogénesis y por
otro lado se evaluó el efecto de NPS en la producción de brotes a partir de yemas
axilares.

2
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
II. ANTECEDENTES
2.1 Origen e Importancia del género Polianthes
El género Polianthes L. (Asparagaceae), es endémico de México y está
conformado por catorce especies, tres variedades y dos cultivares (Solano, 2000).
Desde tiempos prehispánicos, este género ha tenido importancia en aspectos
económicos, científicos y culturales, siendo utilizado comúnmente para propósitos
ornamentales y ceremoniales. Polianthes L. se distribuye principalmente desde el
sur de Chihuahua y Tamaulipas hasta el centro-sur de Oaxaca. La Figura 1
muestra las áreas con mayor variedad de especies que se localizan a lo largo de
la Sierra Madre Occidental y la Faja Volcánica Transmexicana (Solano y Feria,
2007). Por otra parte, en base a estudios citogenéticos del género Polianthes, este
comparte una cercana relación a la familia Agavaceae, presentando un número
cromosómico haploide x=30, de los cuales, 25 cromosomas son pequeños y 5 son
grandes con forma de L (Barba-Gonzalez y col., 2012).
Figura 1. Distribución del género Polianthes en México (tomado de Barba-González y col., 2012).
La principal especie conocida del género Polianthes es P. tuberosa L.,
conocida comúnmente como nardo. En México, los aztecas la cultivaban desde
antes de la conquista como planta ornamental y medicinal. Ellos nombraron a sus
flores "omixochitl" o flor de hueso por poseer un color blanco-crema y ser
intensamente fragantes. La planta ornamental P. tuberosa L. se cultiva
extensivamente dentro del territorio nacional principalmente en el estado de

3
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Jalisco (Trueblood, 1973). Además de México; en India, Nueva Zelanda y Japón el
nardo se cultiva comercialmente debido a sus flores intensamente aromáticas y es
usado en la industria del perfume en India y Francia (Barba-González y col.,
2012).
De acuerdo a la Secretaria de Agricultura, Ganadería, Desarrollo Rural,
Pesca y Alimentación (SAGARPA), México reportó en el año 2014 una producción
de P. tuberosa L. de aproximadamente 328 mil toneladas representando un valor
de producción cercano a 47 millones de pesos (Tabla 1).
Tabla 1. Producción de P. tuberosa L. (gruesa) en México
(SAGARPA, 2014)
En México, cinco especies de Polianthes (P. densiflora, P. howardii, P.
longiflora, P. palustris y P. platyphylla) se encuentran incluidas en la categoría de
protección especial en la Norma Oficial Mexicana (NOM-059-ECOL-2001)
(SEMARNAT, 2002), anteriormente estas especies estaban catalogadas como
raras ante la IUCN (por sus siglas inglés, International Union for Conservation of
Nature) (Oldfield, 1997). Actualmente, el término "raro" ya no es utilizado, sin
embargo, cabe resaltar que la rareza es una categoría natural de distribución o
abundancia del taxón y no necesariamente indica riesgo de extinción.
Parámetro Valor
Superficie sembrada (ha) 282.50
Superficie cosechada (ha) 282.50
Producción (t) 327,467.15
Rendimiento (t/ha) 1,159.18
Precio medio rural (pesos (MXN)/t) 143.74
Valor de producción (miles de pesos (MXN)) 47,069.96

4
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
No obstante, tal concepto, puede considerarse un factor importante en la
evaluación del estatus de conservación de las especies (Solano y Feria, 2007).
Siendo importante efectuar una revisión continua de dicho estatus sobre todo
cuando existe una transformación de los hábitats naturales (Oldfield, 1997).
2.2 Características morfológicas de P. tuberosa, especies silvestres y
sus híbridos
La morfología de P. tuberosa (Figura 2) se describe como una planta
herbácea, perenne, de hojas verdes lineares a lanceoladas (Figura 2c). Sus flores
intensamente fragantes son blancas, cerosas y se agrupan por parejas. Cada flor
posee un sépalo, seis estambres y un solo pistilo (Figura 2a). La planta completa
(desde la base del bulbo) suele medir entre 60 cm y 1 m de altura. El órgano de
reproducción asexual de P. tuberosa es el bulbo, este tiene una forma ovoide y, su
longitud y diámetro es de 3 a 3.5 cm y 2 cm respectivamente; presenta de 6 a 10
hojas por roseta y de la parte central del bulbo emerge el tallo floral o
inflorescencia, siendo esta, de tipo espiga con flores tubulares, alcanzando una
altura promedio de 40 cm (desde donde aparecen las primeras flores hasta el
ápice floral, Figura 2b) (Solano, 2000).
Figura 2. Morfología de P. tuberosa var. doble. a) Estructura de las flores b)Inflorescencia floral c) Morfología de las hojas

5
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Respecto a la morfología de las especies silvestres de Polianthes, la especie
P. howardii Verh-Will (distribuida principalmente en los estados de Colima y
Jalisco) (Figura 3) se caracteriza por presentar una longitud de planta completa
entre 62 a 108 cm; su bulbo, al igual que P. tuberosa, es de forma ovoide y
generalmente tanto su longitud como su diámetro mide 8 mm; este bulbo posee de
5 a 6 hojas por roseta (con manchas purpuras en la base) con una longitud y
ancho promedio de 22 a 27 cm y 1.5 a 2.5 cm respectivamente. Por otra parte, su
inflorescencia con forma de racimo tiene de 15 a 41 nudos fértiles y, la distancia
que existe entre dos nudos es mayor en el caso de los nudos estériles que en los
fértiles. Con relación a la sección donde se encuentran las flores en la
inflorescencia, esta sección presenta un color morado o rojizo; sus flores sin
fragancia aparecen solitarias con pedicelos rojizos con una longitud de 2 a 4.7 cm
y un tubo floral de 1.2 a 2.1 cm de largo y 3.2 a 5 mm de ancho (Solano, 2000).
Figura 3. Morfologia de la especie silvestre P. howardii Verh-Will. (a) tomada de Barba-González y col., 2012; (b) tomada de Feria-Arroyo y col., 2010).
Se han realizado cruzas entre Polianthes tuberosa L. y especies silvestres
para obtener híbridos que conserven o no características de ambos progenitores y
puedan ser de interés comercial y/o designadas para el registro de nuevas

6
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
variedades. En este sentido, en el presente trabajo se utilizó un híbrido (No. de
registro interno 1008157 proporcionado por el Centro de Investigación y Asistencia
en Tecnología y Diseño del Estado de Jalisco A.C., CIATEJ) producto de la cruza
realizada entre P. tuberosa L. (madre) y P. howardii Verh-Will. (padre). La Figura 4
muestra las principales carecteristicas morfológicas del híbrido.
Figura 4. Morfología del híbrido 1008157.
2.3 Reproducción de Polianthes tuberosa y especies silvestres.
La reproducción comercial de P. tuberosa L. se realiza comúnmente
utilizando el bulbo, que se siembra al comienzo de la estación templada, en un
terreno rico en materia orgánica y con un porcentaje elevado de arcilla y de arena.
Cuando es tiempo de floración, la inflorescencia suele durar más de 40 d en
disposición de ser cortada, ya que éstas deben mantenerse para completar el ciclo
de multiplicación de los nuevos bulbos, que por regla general se extraen de la
tierra a la llegada de la estación fría. La reproducción de Polianthes tiene un ciclo

7
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
de tres años. Durante el primer año los bulbillos nacen junto al bulbo madre, en el
segundo se engrosan y finalmente durante el tercer año, los bulbillos son
separados del bulbo madre, estando listos para la etapa de floración (SIAP, 2014).
Por otra parte, las especies silvestres de Polianthes pueden ser reproducidas
tanto vegetativamente mediante bulbos (al igual que P. tuberosa) como
sexualmente a partir de la semilla. La técnica de propagación por semilla es
utilizada en programas de mejoramiento, siendo importante en el mantenimiento
de la diversidad genética y la propagación de las especies de Polianthes en
peligro de extinción. Sin embargo, desde un punto de vista comercial, la
propagación por semilla presenta una gran desventaja; y es que para alcanzar la
madurez en la planta es necesario un tiempo demasiado prolongado. Aunado a
esta desventaja, muchas especies silvestres de Polianthes presentan una
producción deficiente en el número y la viabilidad de las semillas (Barba-González
y col., 2012).
2.4 Propagación in vitro del género Polianthes L.
Como se mencionó previamente, dentro de las características de
reproducción para el género Polianthes (tanto comercial como silvestre), la forma
más común y fácil de propagación es de manera asexual usando bulbos. Sin
embargo, debido a sus ventajas, la propagación in vitro también conocida como
micropropagación es la principal técnica utilizada para la generación de millones
de plántulas de diferentes especies cada año, ya que teóricamente, para
micropropagar un cultivo, cualquier órgano y tejido de las plantas pueden ser
potencialmente utilizados como punto de inicio (Mendoza, 2013).
Las ventajas de la propagación in vitro abarcan dos vertientes, la primera, es
la cantidad de material vegetal utilizado para poder realizar la técnica, es decir,
para iniciar la micropropagación se puede partir de una parte muy pequeña de la
planta y así generar nuevos brotes. La segunda, es el desarrollo del cultivo masivo
de una especie, esto permite producir un número elevado de plantas en un

8
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
espacio reducido, lo que conlleva a producir plantas no solo en una temporada
sino durante cualquier época del año y, disminuir los insumos y el personal
destinado para el cuidado de las plantas. Además, el crecimiento de las plantas
propagadas in vitro con frecuencia es más vigoroso que el de las plantas
propagadas in vivo, debido al rejuvenecimiento de la planta y a la obtención de
plantas libres de enfermedades (George, 1993; George y col., 2008).
Las técnicas para la micropropagación que son teóricamente disponibles
para la regeneración de plantas in vitro se ilustran en la Figura 5. Resumiendo, las
técnicas se pueden realizar a partir de la multiplicación de brotes partiendo de (1)
yemas axilares o apicales y (2) por la formación de brotes adventicios y/o
embriones somáticos adventicios (George, 1993).
Dentro de las técnicas para realizar la propagación, en el presente estudio se
utilizó la multiplicación por medio de yemas axilares y la embriogénesis somática
(mostrada en sección 2.5). Respecto a la primera, es importante mencionar que el
fundamento de esta técnica se basa en promover el desarrollo de puntos de
crecimiento ya existentes en las plantas conocidos como meristemos. Por otra
parte, teóricamente la multiplicación por medio de yemas axilares mantiene la
fidelidad genética de la descendencia con respecto a la planta madre, ya que la
variación somaclonal está prácticamente ausente cuando se cultivan estructuras
organizadas como lo son meristemos y yemas. Por lo que puede considerarse que
este es el sistema ideal para la propagación clonal de cultivares (Molphe y col.,
1999).
Debido a que no existe un método general para la micropropagación, es
necesario realizar estudios específicos con el fin de efectuar y mejorar esta técnica
consiguiendo de esta forma la regeneración y multiplicación de una especie o
variedad en particular. En la actualidad los estudios que conllevan a la
regeneración de plantas in vitro son enfocados principalmente a estimular una
respuesta positiva en el crecimiento de la planta. Esto se consigue básicamente a

9
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
través de la evaluación de (1) diferentes tipos de tejido como explante inicial y (2)
diferentes combinaciones de reguladores de crecimiento (RC) y agentes químicos,
por ejemplo, la adición de nitroprusiato de sodio como donador de óxido nítrico al
medio de cultivo para favorecer los eventos fisiológicos y bioquímicos de las
plantas (Rico-Lemus y Rodríguez-Garay, 2014).
Figura 5. Principales técnicas para la propagación in vitro. Adaptado de George y col., 2008.
Aunque la propagación in vitro de especies silvestres o híbridos de
Polianthes es nula, existe información escasa para la micropropagación de P.
tuberosa L. En la siguiente tabla se muestra en orden cronológico y de forma
resumida información de diversos trabajos realizados para la micropropagación de
este género. Cabe mencionar que hasta la fecha, no se reporta el desarrollo de
protocolos de micropropagación para el género Polianthes utilizando agentes
químicos donadores de óxido nítrico.
10
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
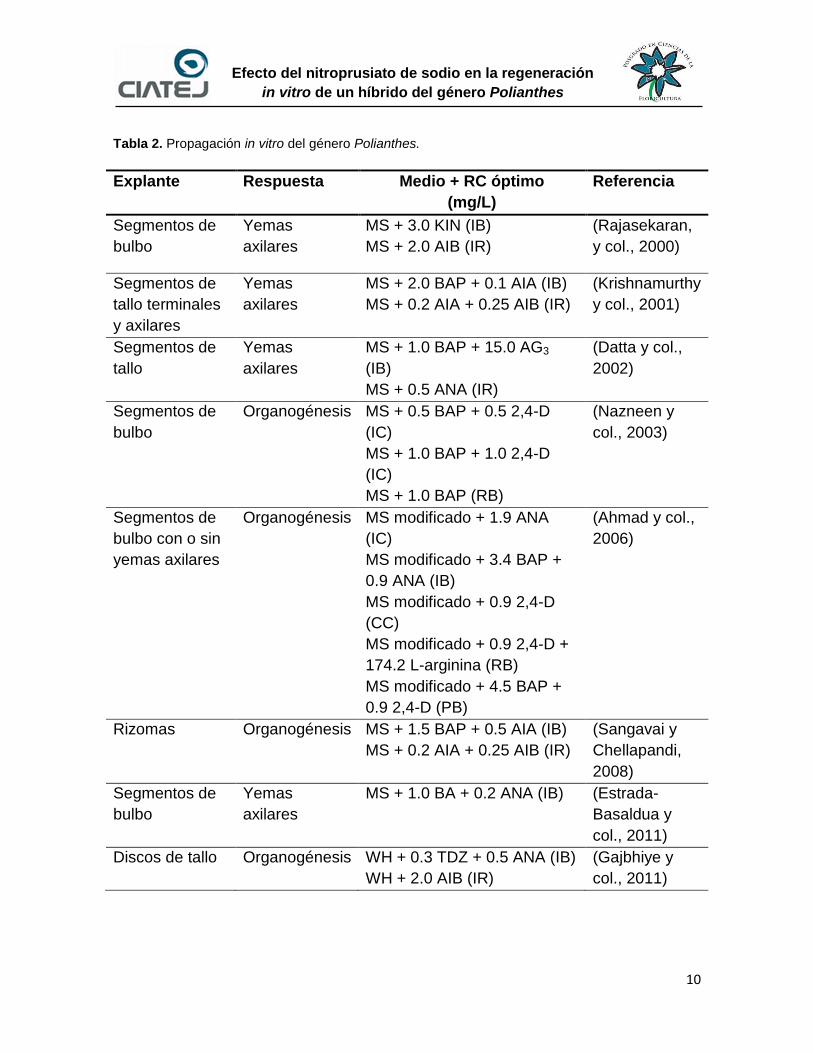
Tabla 2. Propagación in vitro del género Polianthes.
Explante Respuesta Medio + RC óptimo
(mg/L)
Referencia
Segmentos de
bulbo
Yemas
axilares
MS + 3.0 KIN (IB)
MS + 2.0 AIB (IR)
(Rajasekaran,
y col., 2000)
Segmentos de
tallo terminales
y axilares
Yemas
axilares
MS + 2.0 BAP + 0.1 AIA (IB)
MS + 0.2 AIA + 0.25 AIB (IR)
(Krishnamurthy
y col., 2001)
Segmentos de
tallo
Yemas
axilares
MS + 1.0 BAP + 15.0 AG3
(IB)
MS + 0.5 ANA (IR)
(Datta y col.,
2002)
Segmentos de
bulbo
Organogénesis MS + 0.5 BAP + 0.5 2,4-D
(IC)
MS + 1.0 BAP + 1.0 2,4-D
(IC)
MS + 1.0 BAP (RB)
(Nazneen y
col., 2003)
Segmentos de
bulbo con o sin
yemas axilares
Organogénesis MS modificado + 1.9 ANA
(IC)
MS modificado + 3.4 BAP +
0.9 ANA (IB)
MS modificado + 0.9 2,4-D
(CC)
MS modificado + 0.9 2,4-D +
174.2 L-arginina (RB)
MS modificado + 4.5 BAP +
0.9 2,4-D (PB)
(Ahmad y col.,
2006)
Rizomas Organogénesis MS + 1.5 BAP + 0.5 AIA (IB)
MS + 0.2 AIA + 0.25 AIB (IR)
(Sangavai y
Chellapandi,
2008)
Segmentos de
bulbo
Yemas
axilares
MS + 1.0 BA + 0.2 ANA (IB) (Estrada-
Basaldua y
col., 2011)
Discos de tallo Organogénesis WH + 0.3 TDZ + 0.5 ANA (IB)
WH + 2.0 AIB (IR)
(Gajbhiye y
col., 2011)
11
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
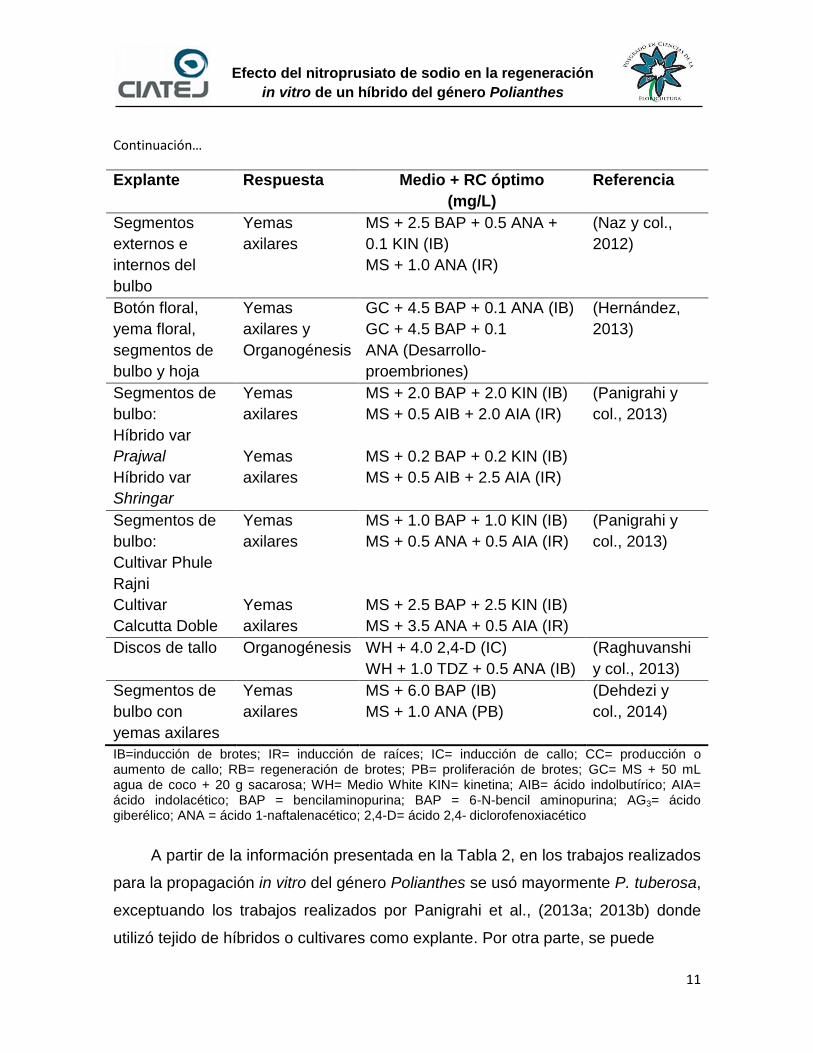
Continuación…
Explante Respuesta Medio + RC óptimo
(mg/L)
Referencia
Segmentos
externos e
internos del
bulbo
Yemas
axilares
MS + 2.5 BAP + 0.5 ANA +
0.1 KIN (IB)
MS + 1.0 ANA (IR)
(Naz y col.,
2012)
Botón floral,
yema floral,
segmentos de
bulbo y hoja
Yemas
axilares y
Organogénesis
GC + 4.5 BAP + 0.1 ANA (IB)
GC + 4.5 BAP + 0.1
ANA (Desarrollo-
proembriones)
(Hernández,
2013)
Segmentos de
bulbo:
Híbrido var
Prajwal
Híbrido var
Shringar
Yemas
axilares
Yemas
axilares
MS + 2.0 BAP + 2.0 KIN (IB)
MS + 0.5 AIB + 2.0 AIA (IR)
MS + 0.2 BAP + 0.2 KIN (IB)
MS + 0.5 AIB + 2.5 AIA (IR)
(Panigrahi y
col., 2013)
Segmentos de
bulbo:
Cultivar Phule
Rajni
Cultivar
Calcutta Doble
Yemas
axilares
Yemas
axilares
MS + 1.0 BAP + 1.0 KIN (IB)
MS + 0.5 ANA + 0.5 AIA (IR)
MS + 2.5 BAP + 2.5 KIN (IB)
MS + 3.5 ANA + 0.5 AIA (IR)
(Panigrahi y
col., 2013)
Discos de tallo Organogénesis WH + 4.0 2,4-D (IC)
WH + 1.0 TDZ + 0.5 ANA (IB)
(Raghuvanshi
y col., 2013)
Segmentos de
bulbo con
yemas axilares
Yemas
axilares
MS + 6.0 BAP (IB)
MS + 1.0 ANA (PB)
(Dehdezi y
col., 2014)
IB=inducción de brotes; IR= inducción de raíces; IC= inducción de callo; CC= producción o aumento de callo; RB= regeneración de brotes; PB= proliferación de brotes; GC= MS + 50 mL agua de coco + 20 g sacarosa; WH= Medio White KIN= kinetina; AIB= ácido indolbutírico; AIA= ácido indolacético; BAP = bencilaminopurina; BAP = 6-N-bencil aminopurina; AG3= ácido giberélico; ANA = ácido 1-naftalenacético; 2,4-D= ácido 2,4- diclorofenoxiacético
A partir de la información presentada en la Tabla 2, en los trabajos realizados
para la propagación in vitro del género Polianthes se usó mayormente P. tuberosa,
exceptuando los trabajos realizados por Panigrahi et al., (2013a; 2013b) donde
utilizó tejido de híbridos o cultivares como explante. Por otra parte, se puede

12
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
resaltar el éxito que se ha tenido en la regeneración de plantas utilizando la
técnica de propagación por yemas axilares.
Además, las concentraciones óptimas de RC para una mejor respuesta en la
micropropagación encontradas empíricamente durante los diferentes estudios,
representan un amplio espectro de opciones que sirven de guía para el desarrollo
de nuevos protocolos. Es importante subrayar que el efecto de los RC es diferente
para cada planta, y está en función de la especie, el explante que se utilice, el
estado de desarrollo en que se encuentre la planta de donde se toma el explante
inicial, la concentración e interacción hormonal, y los factores ambientales a la
cual sea expuesta.
2.5 Embriogénesis sómatica
Antes de la descripción de las etapas y eventos celulares que ocurren
durante el proceso de embriogénesis somática, es importante tomar en cuenta
algunos conceptos básicos del proceso de embriogénesis cigótica, el cual se lleva
a cabo durante la reproducción sexual de las plantas.
Hace aproximadamente 300 millones de años las plantas se dividieron en
angiospermas y gimnospermas. Actualmente, en ambos grupos las etapas básicas
del desarrollo del embrión se encuentran bien definidas (von Arnold y col., 2002).
En plantas angiospermas, se genera simultáneamente el embrión y el endospermo
durante el proceso llamado doble fertilización, este propicia el desarrollo de una
semilla viable (Dodeman y col., 1997). En contraste, en plantas gimnospermas,
generalmente no ocurre el proceso de la doble fertilización.
En ambos tipos de plantas, la formación del embrión requiere la secuencia
de etapas características, para las angiospermas comienza con la aparición del
cigoto (célula que resulta de la unión de las células sexuales masculina y
femenina), y continúa con su desarrollo a través de divisiones celulares y cambios

13
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
morfológicos hasta llegar a un estado cotiledonar. A todo este proceso, se le
conoce como embriogénesis cigótica (Figura 6a) (von Arnold y col., 2002).
De acuerdo con Dodeman y colaboradores (1997) el desarrollo embrionario
puede ser dividido en dos eventos principales:
1. Embriogénesis sensu stricto: Como se mencionó anteriormente, en el caso
de plantas angiospermas, el desarrollo comienza con la formación del cigoto, el
cual sufre una serie de divisiones de manera ordenada. Estas divisiones fueron
descritas y agrupadas en tres eventos por (Goldberg y col.,1994): (1) División
asimétrica del cigoto, la cual da origen a una célula pequeña apical y una célula
grande basal; (2) Formación de un patrón específico, la cual da lugar al estadio de
un embrión globular y (3) Transición a la etapa cotiledonar, en la cual se presenta
la aparición de cotiledones primarios.
Por otro lado, en plantas gimnospermas, la secuencia para el desarrollo
embrionario está dividida en tres eventos descritos por Singh (1978) y que se
esquematizan en la Figura 6b: (1) Proembriogénesis, la cual representa a todas
las etapas antes de la elongación del suspensor; (2) Embriogénesis temprana, la
cual engloba a todas las etapas después de la elongación del suspensor y antes
del establecimiento del meristemo radicular y (3) Embriogénesis tardía,
histogénesis intensiva que incluye la formación de la raíz y los meristemos
apicales.
2. Maduración del embrión seguido por su germinación: A partir de esta
etapa, tanto en angiospermas como en gimnospermas la actividad meristemática
se dispara dando lugar a cambios fisiológicos donde inician los procesos de
crecimiento, almacenamiento de sustancias de reserva y maduración. Los
cambios fisiológicos, tales como la desecación, y en la mayoría de los casos la
dormancia, completan el proceso de la formación de semillas (Dodeman y col.,
1997).

14
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 6. Embriogénesis cigótica. (a) en angiospermas, ejemplo Arabidopsis, dibujos basados en Goldberg y col., 1994. (b) en gimnospermas, ejemplo de la familia Pinacea, dibujos basados en Gifford y Foster, 1989). Abreviaturas traducidas del inglés: EP = parte embrionaria, pU = suspensor primario, pE = parte embrionaria primaria, E = sección de embrión, S = sección suspensor, U= tejido somático, EM = tejido embrionario, sS = suspensor secundario. La ilustración no está a escala.
Por otra parte y, retomando el enfoque del presente estudio, la
embriogénesis somática se define como un proceso de desarrollo en el cual
células somáticas son sometidas a un serie de reestructuraciones para generar
células embriogénicas. Las células somáticas experimentan cambios morfológicos
y bioquímicos que resultan en la formación de un embrión somático (también
llamado no-cigótico) con la capacidad de regenerarse y formar una nueva planta
(Yang y Zhang, 2010).
Los primeros reportes de embriogénesis somática fueron publicados en 1958
por Reinert; Stewards y colaboradores, utilizando un cultivo de células en
suspensión de Daucus carota L. Desde entonces, el potencial de la embriogénesis
somática ha sido probado exitosamente en una amplia gama de sistemas de

15
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
de tejidos tanto para plantas dicotiledóneas como monocotiledóneas (Mathieu y
col., 2006; Quiroz-Figueroa y col., 2006).
La embriogénesis somática es una técnica muy útil para la conservación de
germoplasma in vitro y para el mejoramiento genético. Como método para la
micropropagación, esta aumenta la eficiencia del sistema de multiplicación,
disminuye los costos de producción, y permite la automatización parcial del
proceso mediante el uso de biorreactores. Es decir, esta técnica ha permitido
desarrollar la micropropagación eficiente de varias especies económicamente
importantes. En el proceso se modulan factores biológicos, físicos y químicos los
cuales deben ser optimizados para cada especie, variedad, o incluso línea
genética con el fin de generar el éxito de un protocolo y/o hacerlo comercialmente
viable (Monja-Mio y Robert, 2013).
La embriogénesis somática puede derivar de una célula somática individual o
de un grupo de células. Además, es posible clasificarla en dos tipos de
embriogénesis: (1) Directa, cuando los embriones somáticos se diferencian desde
el explante inicial sin pasar por una fase de callo y (2) Indirecta, donde el explante
inicial forma un callo que posteriormente se diferencia para formar embriones
somáticos (Williams y Maheswaran, 1986).
En resumen, para realizar la embriogénesis somática se requiere de una
serie de etapas que siguen una única ruta de desarrollo que involucra un número
de eventos celulares complejos (Figura 7). Dichas etapas se explican en base a
von Arnold y col., (2002)
1. Iniciación de cultivos embriogénicos, para ello se cultiva el explante inicial
en un medio suplementado con reguladores de crecimiento, principalmente
auxinas, pero comúnmente dicho medio también es adicionado con citocininas.

16
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
2. Proliferación de cultivos embriogénicos, se realiza en medio sólido o
líquido suplementado con reguladores de crecimiento a concentraciones similares
o menores a las utilizadas en el medio de iniciación.
3. Premaduración de embriones somáticos, esto se lleva a cabo en medio sin
RC, para inhibir la proliferación de células no embriogénicas y, estimular la
formación y el desarrollo temprano de embriones.
4. Maduración de embriones somáticos, se realiza un cultivo utilizando un
medio suplementado con ABA y/o con un potencial osmótico reducido.
5. Desarrollo de plantas, para ello se utiliza un medio que carece de RC, lo
cual propicia la aparición y el fortalecimiento de raíces.
Figura 7. Eventos característicos de la embriogénesis somática.
Nomura y Komamine (1985) y Quiroz-Figueroa y colaboradores (2002)
coinciden en llamar a la primera fase de la embriogénesis somática, iniciación
embriogénica, donde células somáticas diferenciadas pasan por un proceso de
desdiferenciación para adquirir competencia embriogénica y proliferar como
células embriogénicas. Esta fase de iniciación de la vía embriogénica se limita a
ciertas células sensibles que tienen el potencial para activar los genes que
participan en la generación de células embriogénicas. Una vez que estos genes se
1. Desdiferenciación celular
2. Activación de la división celular
3. Reprogramación celular (fisiología, metabolismo, patrones de expresión de
genes)

17
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
activan, la expresión programada de genes embriogénicos reemplaza el patrón de
genes establecido en el tejido del explante (Quiroz-Figueroa y col., 2002).
Una vez que las células embriogénicas se han formado, estas continúan
proliferando y forman masas proembriogénicas (PEMs). Las auxinas son
requeridas para la etapa de proliferación pero son inhibidoras para el desarrollo
posterior de embriones somáticos (Filonova y col., 2000; Nomura y Komamine,
1995). El grado de diferenciación de embriones que tiene lugar en presencia de
concentraciones de auxina varía en diferentes especies. En la mayoría de los
cultivos, el potencial embriogénico disminuye con el cultivo prolongado en un
medio que contiene reguladores de crecimiento y eventualmente llega a perderse
esa capacidad embriogénica (von Arnold y col., 2002).
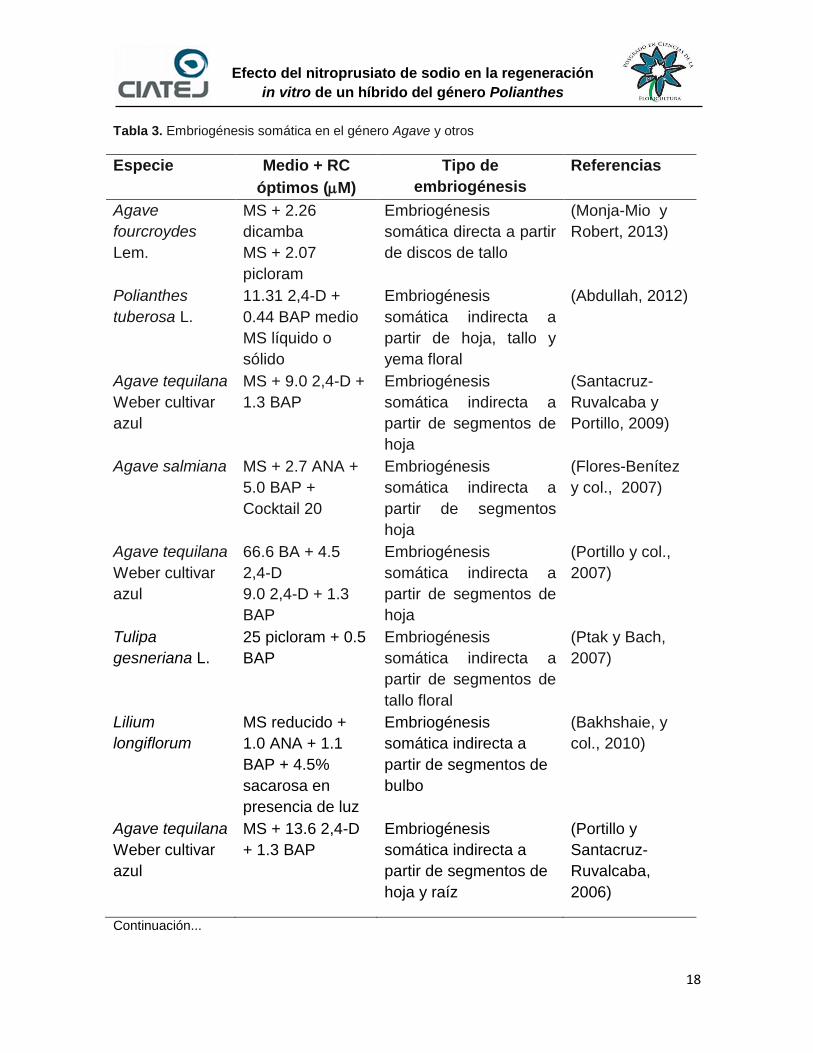
Dentro del acervo bibliográfico de la técnica de embriogénesis somática en el
género Polianthes la información es casi nula, sin embargo, existe información de
géneros cercanamente emparentados como es el caso del género Agave, donde
se ha logrado exitosamente la regeneración de embriones somáticos. También, la
embriogénesis somática se ha realizado satisfactoriamente en otras plantas
ornanmentales bulbosas como es el caso de Tulipa gesneriana L. y Lilium
longiflorum Thunb. Basado en esta revisión (Tabla 3), en el presente trabajo de
investigación fueron seleccionados aquellos reguladores de crecimiento que
pudieran resultar más efectivos para el desarrolo de esta técnica.
18
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Tabla 3. Embriogénesis somática en el género Agave y otros
Especie Medio + RC
óptimos (M)
Tipo de
embriogénesis
Referencias
Agave
fourcroydes
Lem.
MS + 2.26
dicamba
MS + 2.07
picloram
Embriogénesis
somática directa a partir
de discos de tallo
(Monja-Mio y
Robert, 2013)
Polianthes
tuberosa L.
11.31 2,4-D +
0.44 BAP medio
MS líquido o
sólido
Embriogénesis
somática indirecta a
partir de hoja, tallo y
yema floral
(Abdullah, 2012)
Agave tequilana
Weber cultivar
azul
MS + 9.0 2,4-D +
1.3 BAP
Embriogénesis
somática indirecta a
partir de segmentos de
hoja
(Santacruz-
Ruvalcaba y
Portillo, 2009)
Agave salmiana MS + 2.7 ANA +
5.0 BAP +
Cocktail 20
Embriogénesis
somática indirecta a
partir de segmentos
hoja
(Flores-Benítez
y col., 2007)
Agave tequilana
Weber cultivar
azul
66.6 BA + 4.5
2,4-D
9.0 2,4-D + 1.3
BAP
Embriogénesis
somática indirecta a
partir de segmentos de
hoja
(Portillo y col.,
2007)
Tulipa
gesneriana L.
25 picloram + 0.5
BAP
Embriogénesis
somática indirecta a
partir de segmentos de
tallo floral
(Ptak y Bach,
2007)
Lilium
longiflorum
MS reducido +
1.0 ANA + 1.1
BAP + 4.5%
sacarosa en
presencia de luz
Embriogénesis
somática indirecta a
partir de segmentos de
bulbo
(Bakhshaie, y
col., 2010)
Agave tequilana
Weber cultivar
azul
MS + 13.6 2,4-D
+ 1.3 BAP
Embriogénesis
somática indirecta a
partir de segmentos de
hoja y raíz
(Portillo y
Santacruz-
Ruvalcaba,
2006)
Continuación...

19
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Especie Medio + RC
óptimos (M)
Tipo de
embriogénesis
Referencias
Agave victoria
reginae
MS + 2.26 2,4-D Embriogénesis
somática indirecta
(Martínez-
Palacios y col.,
2003)
Agave victoria
reginae Moore
MS + 1.4 2,4-D Embriogénesis
somática directa a partir
de hoja
(Rodríguez-
Garay y col.,
1996)
2.5.1 Picloram y 2,4-D en la inducción de embriogénesis.
Se ha propuesto que los reguladores de crecimiento vegetal y el estrés
celular desempeñan un papel central en la mediación de la cascada de
transducción de señales. Esto conduce a la reprogramación de la expresión
génica, seguido por una serie de divisiones celulares que inducen el crecimiento
del callo ya sea de manera desorganizada o mostrando un crecimiento polarizado
que conduce a la embriogénesis somática (de Jong y col., 1993; Dudits y col.,
1991).
La iniciación o activación en función de una respuesta auxínica puede ser un
evento clave en la adaptación celular y la reprogramación genética, metabólica, y
fisiológica, conduciendo a que células vegetales somáticas sean potencialmente
embriogénicas (Yang y Zhang, 2010). Además de las auxinas como un inductor
principal para la embriogénesis somática, también hay estudios en respuesta
aotros RC como citocininas o el ácido abscísico (ABA). Recientemente la
aplicación de una nueva generación de RC como los oligosacáridos, jasmonatos,
poliaminas y brasinoesteriodes han demostrado ser útiles para la iniciación de la
embriogénesis en muchas especies vegetales (Jiménez, 2005).
Las auxinas comúnmente utilizadas en el cultivo de tejidos son el ácido 2,4-
diclorofenoxiacético (2,4-D) y el ácido 1-naftalenacético (ANA). El 2,4-D es usado
para la inducción de callo y en la técnica de suspensiones celulares. Otras auxinas
como dicamba (ácido 3,6-dicloro-O-anísico) y picloram (ácido 4-amino-3,5,6

20
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
trichloropyridine-2-carboxilico) son empleadas comercialmente como herbicidas
selectivos, en contraste, estos mismos son comúnmente efectivos en la inducción
de formación de tejido embriogénico y en el mantenimiento de la viabilidad de
suspensiones celulares (Gaspar y col., 1996).
La eficiencia del picloram en la inducción de callo es comparable a la
eficiencia del 2,4-D en terminos de la inducción de crecimiento, y es superior a
éste en cuanto a lograr la friabilidad de los callos (Gaspar y col., 1996). Sin
embargo, como se ha mencionando anteriormente, el nivel de respuesta de los
grupos de RC es muy variable. Especificamente la acción de las auxinas se
diferencia no sólo de planta a planta, sino también de órgano a órgano, tejido a
tejido, célula a célula y, por otra parte, también con la edad y el estado fisiológico
de la planta (Davies, 2010). Algunos ejemplos comparativos en el uso de picloram
y 2,4-D son mostrados en la Tabla 3. La auxina inductora de la embriogénesis
somática indirecta que ha presentado mayor éxito sobre las especies Agave
tequilana y Agave victoria reginae es 2,4-D. Sin embargo, dentro del mismo
genero pero diferente especie (Agave fourcroydes) resultó ser mejor el uso de
picloram para la inducción de embriogénesis somática y coincide con trabajos
realizados en otras especies de plantas hornamentales y bulbosas.
Respecto a otras especies, donde se ha empleado el picloram como el
principal regulador de crecimiento para el desarrollo de embriogénesis somatica,
se encuentra el estudio realizado en Fragaria ananassa L. que utilizó hojas como
explante inicial y el medio óptimo para la inducción de embriones somáticos
contenía 2 mg/L de picloram (Kordestani y Karami, 2008). Otros ejemplos de éxito
para la inducción de embriogénesis somática utilizando picloram son en: Triticum
aestivum L. donde el uso del picloram aumentó la producción de callo (Mendoza y
Kaeppler, 2002); cultivares de Hordeum vulgare (Castillo y col., 1998); Paspalum
scrobiculatum L. donde la formación directa de embriones somáticos tuvo lugar
utilizando un medio de cultivo que contenía una combinación de picloram y

21
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
cinetina (Kaur y Kothari, 2004) y Lilium longiflorum donde se adicionó picloram en
una concentración de 2 M para obtener un callo friable (Tribulato y col., 1997).
2.6 Óxido nítrico
2.6.1 Definición y propiedades fisicoquímicas del óxido nítrico
El óxido nítrico (NO) es un gas incoloro, inorgánico, lábil, de bajo peso
molecular (30.01 g/mol), que tiene un electrón no apareado en su capa exterior y
debido a ello posee las características de un radical que puede ganar o perder un
electrón, intercambiándose en tres especies diferentes: el radical (NO•), el catión
nitrosonio (NO+) y el anión nitroxilo (NO-) lo que explica su elevada reactividad y
su tendencia a unirse con hemoproteínas reducidas (Thomas y col., 2001). Estas
tres formas moleculares en las que se puede encontrar el NO, son intercambiables
dentro de la célula y dependen fuertemente del potencial redox de la misma. El
radical (NO•) se difunde libremente en soluciones acuosas y es capaz de atravesar
membranas lipídicas; moverse dentro de compartimentos celulares y de célula a
célula (Subczynski y col, 1996).
El NO es inestable y ligeramente soluble en medio acuoso, un poco más
soluble en solventes orgánicos y, en presencia de oxígeno se oxida a nitrito y
nitrato los cuales son moléculas más estables. Por otra parte, el NO tiene una vida
media de menos de seis segundos, siendo el nitrito el producto más abundante de
su catabolismo (Airaki y col., 2012).
El óxido nítrico fue descubierto en 1772 por Joseph Priestley como “aire
nitroso” y anteriormente se creía que era solo un gas venenoso que causaba la
lluvia ácida (Khan y col., 2013). La primera función biológica del NO fue reportada
por Ignarro y colaboradores en 1987 (Hou y col., 1999). La importancia biológica
del óxido nítrico ganó considerable atención en todo el mundo cuando en el año
1992 fue nominada como la “molécula del año” por las revistas de ciencia (Khan y
col., 2013). En 1998 tres científicos americanos Robert F. Furchgott, Ferid Murad y

22
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Louis J. Ignarro identificaron el NO como una molecula señalizadora en el sistema
cardiovascular ganando el premio nobel de Medicina y Fisiología en ese mismo
año. Debido a este aporte en el campo de la medicina humana, actualmente, el
NO ha sido blanco de estudios para evaluar su efecto en los procesos fisiológicos
y patológicos en plantas (Delledonne, 2005; Hou y col., 1999).
2.6.2 Función del óxido nítrico en las células vegetales
Inicialmente, los estudios para dilucidar la funcionalidad del NO se
desarrollaron utilizando células animales, donde el NO funciona como un factor de
relajación endotelial (EDRF, endothelium-derived relaxing factor) que se genera a
partir de la conversión de L-arginina, por la enzima óxido nítrico sintasa (NOS) y
sus 3 isoformas (nNOS, eNOS, iNOS), algunas de ellas dependientes de la
concentración de calcio libre intracelular (Lamattina y col., 2003; Wendehenne y
col., 2001).
En plantas, el NO juega el rol de mensajero químico, ya que este participa
tanto en procesos fisiológicos, patológicos y en el desarrollo de las células
vegetales. Muchos estímulos hormonales y medioambientales son transmitidos
tanto directa o indirectamente por cascadas de señalización donde participa el NO.
La capacidad del NO para actuar simultáneamente en diferentes rutas bioquímicas
sin relación y sus propiedades homeostáticas redox sugieren que podría ser una
molécula de sincronización en las plantas las cuales pueden generar NO por
sistemas tanto enzimáticos como no enzimáticos (Lamattina y col., 2003).
Los procesos donde participa el NO son diversos pudiendo enumerarlos de
la siguiente manera: (1) resistencia a enfermedades causadas por patógenos; (2)
respuesta al estrés abiótico; (3) dormancia y germinación de semillas; (4)
estimulación del crecimiento de plántulas; (5) etapa de floración; (6) cierre de los
estomas; (7) regulación del crecimiento y orientación del tubo polínico; (8)
maduración de frutos; (9) retardo de la senescencia de las hojas; (10) actividades
enzimáticas; (11) rutas de señalización MAP quinasas; (12) expresión de genes

23
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
del ciclo celular; (13) desarrollo de nódulos funcionales y (14) crecimiento de
raíces primarias y laterales (del Rio y col., 2014; Khan y col., 2013). Además, otros
estudios indicaron que el NO puede participar en la división celular y por lo tanto
afectar la regeneración y multiplicación de brotes adventicios (Han y col., 2009).
2.6.2.1 Rol del óxido nítrico en la síntesis de clorofila
Los cloroplastos son el principal sitio de la célula donde se produce NO, es
por ello que el funcionamiento correcto de estos es esencial para mantener los
niveles adecuados de NO en las hojas. El óxido nítrico estimula la biosíntesis de
clorofila, la diferenciación de los cloroplastos (Tewari y col., 2013) y preserva los
niveles de clorofila durante la senescencia de las hojas. Sin embargo, el
mecanismo por el cual el NO regula el proceso de degradación de clorofila
permanece aún desconocido (Liu y Guo, 2013).
Dependiendo de factores como la concentración, tipo de tejido, edad de la
planta y el tipo de estrés, el NO puede ser un agente inductor de estrés, ó bien, el
NO puede desempeñar un rol en la protección de las células ante el daño
oxidativo (Lipton y col., 1993). Lazalt y colaboradores (1997) han reportado que el
óxido nítrico fue capaz de prevenir parcialmente la degradación de clorofila
producida por Phytophtora infestans en hojas de papa, postulando la capacidad de
NO como una molécula protectora, tanto para preservar la membrana de los
cloroplastos en contra de los efectos tóxicos de las especies reactivas de oxígeno
en secciones de hoja infectadas o como una molécula directamente involucrada
en cualquiera de los pasos de la vía metabólica de la clorofila. Por otro lado,
(Beligni y Lamattina, 1999) y (Hung y col., 2002) han reportado que el NO es
capaz de contrarrestar la toxicidad de herbicidas como el paraquat y diquat en
hojas de papa y arroz, respectivamente.
En otros términos, respecto al conocimiento en el área de cuidados post-
cosecha de frutas y hortalizas existen estudios donde se ha evaluado la actividad
protectora de NO (Eum y col., 2009; Manjunatha y col., 2010). Por ejemplo, la

24
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
aplicación externa de NO en flores de brócoli retrasa el proceso de amarillamiento
y el inicio de la degradación de clorofila durante el almacenamiento a 20°C (Eum y
col., 2009). Otro estudio realizado en plantas de plátano sugiere que el NO puede
mejorar el sistema antioxidante de las plantas de manera enzimática y no
enzimática retardando la degradación de clorofila. Debido a su efecto inhibitorio
sobre la actividad de las enzimas que degradan la clorofila y la promoción del
sistema de defensa antioxidante, se sugiere que el NO tiene potencial para
preservar el contenido de clorofila y mantener la calidad de los frutos después de
almacenamiento en frío (Wang y col., 2015).
2.6.3 Biosíntesis de óxido nítrico en las plantas
Las plantas pueden generar NO por procesos tanto enzimáticos como no
enzimáticos. Hasta la fecha, se han reconocido y estudiado ocho diferentes rutas
(Figura 8) para la producción de NO en plantas. Los principales sitios en la célula
donde se realiza la biosíntesis de NO son los cloroplastos, mitocondrias y
peroxisomas (Rőszer, 2014). El citoplasma, la membrana celular, el retículo
endoplasmático y el apoplasto también pueden generar NO en plantas vasculares
(Fröhlich y Durner, 2011).
La síntesis enzimática de NO en plantas se divide básicamente en dos vías
una reductiva y otra oxidativa. En la primera, la generación de NO se realiza
utilizando la enzima nitrato reductasa (NR). Esta reacción ocurre en el citoplasma
de la célula donde NR cataliza la reacción de reducción de nitrato a nitrito usando
NADH como donador de electrones. Esta misma enzima NR también cataliza la
reacción de reducción de nitrito a NO (Airaki y col., 2012). La síntesis de NO
mediante la NR requiere concentraciones relativamente bajas de nitrato y altas de
nitrito porque la afinidad por el nitrito (KM = 100 µM) es mayor comparada con la
constante de inhibición del nitrato (Ki = 50 µM) (Rockel y col., 2002) y, por lo tanto,
la producción de NO depende de la acumulación de nitrito. Además, las
modificaciones post-traduccionales de la enzima NR afectan la producción de NO
in vitro e in vivo (Airaki y col., 2012).

25
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 8. Rutas enzimáticas y no enzimáticas para la producción de NO en las plantas: (1) Producción de NO por reducción de nitrito que puede ser catalizado por a= nitrato reductasa, b= cadena de trasporte de electrones en la mitocondria y c= hemoproteínas desoxigenadas; (2) bajo condiciones de acidez, el ácido nitroso es protonado y forma nitrito que puede producir NO en un proceso no enzimático; (3) síntesis oxidativa de NO a partir de L-arginina catalizada por la enzima óxido nítrico sintasa; (4, 5 y 6) poliaminas e hidroxilamina pueden incrementar la síntesis de NO aun por mecanismos desconocidos; (7) liberación no enzimática del NO cuando el mismo NO reacciona con el glutatión para formar S-nitrosoglutation (Adaptado de Rőszer 2014).
Por otro lado, dentro de la generación oxidativa de NO en células vegetales,
se encuentra la síntesis de NO mediante la actividad de la enzima putativa óxido
nítrico sintasa (NOS). Así como en células animales, en tejido vegetal fue
reportada la actividad de NOS dependiente del aminoácido L-arginina, donde
también se muestra que la actividad enzimática de NOS puede ser inhibida por la
presencia de derivados estructurales de dicho aminoácido (Khan y col., 2013). En
cuanto a la estructura proteica o al gen que participa en la expresión de esta, de
acuerdo a Fröhlich y Durner (2011), no se ha tenido éxito en la identificación de
dichos genes o enzimas en plantas superiores. Además, Lamattina y
colaboradores (2003) establecieron que no se han encontrado ni el gen o cDNA de
ninguna proteína con alta similitud de secuencia conocida a NOS de animales.
En la actualidad ha sido posible la detección de la actividad de NOS
dependiente del metabolismo de L-arginina en al menos 11 especies diferentes de
plantas, esto mediante métodos bioquímicos, fisicoquímicos y moleculares

26
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
(Corpas y col., 2009; Del Río, 2011). La actividad de NOS fue reportada en
peroxisomas, en células de tejido de raíces, tallos y hojas de plántulas de chícharo
(Barroso y col., 1999; Corpas y col., 2006; del Rio y col., 2002). Así mismo,
Valderrama y colaboradores (2007) reportaron que la salinidad en arboles de olivo
incrementa la actividad de NOS.
Por otra parte, Guo y colaboradores (2003) identificaron en Arabidopsis una
proteína (AtNOS1) que produce NO en respuesta a señales hormonales. Dicha
proteína exhibió en sus genes una secuencia homologa del 16% con respecto a la
posible NOS encontrada en una especie de caracol (Helix pomatia). Sin embargo,
ni la proteína de caracol ni la proteína de Arabidopsis fueron similares a ningún
tipo de NOS de animales, sin embargo, sí fue evidente su actividad enzimática
debido a que existió un incremento de NO mediante la síntesis dependiente de L-
arginina.
Profundizando en el estudio antes mencionado, se utilizó un mutante de
Arabidopsis (Atnos1) para la producción de NO, donde la proteína purificada
(AtNOS1) al igual que en células animales empleaba L-arginina y NADPH como
sustratos y necesitó Ca+2 y calmodulina para su activación. De tal forma que
ninguna secuencia mostró similitud con cualquier isoforma de NOS en mamíferos.
AtNOS1 se propuso como una enzima distinta y se sugirió cambiar el nombre a
AtNOA1 (óxido nítrico asociada 1). En conclusión, la actividad de la posible NOS
en plantas superiores es importante para la generación de NO en la célula, así
como en el crecimiento y la señalización hormonal de esta (Guo, Okamoto, &
Crawford, 2003). Sin embargo, en la actualidad siguen siendo desconocidas las
secuencias homólogas de los genes que codifican para la expresión de NOS en
mamíferos (Mur y col., 2013).
2.6.4 Agentes químicos donadores de óxido nítrico
Existen agentes químicos donadores de óxido nítrico (NO) los cuales han
sido estudiados principalmente en medicina cardiovascular desde hace más de

27
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
dos décadas. Por definición, un agente donador de NO se refiere a cualquier
compuesto que pueda generar NO en alguna de sus formas (radical, anión o
catión) a través de rutas enzimáticas, químicas, electroquímicas o fotoquímicas. La
mayoría de estos agentes son compuestos orgánicos, pero hay un pequeño grupo
de compuestos que forman un complejo de transición metal-NO al que pertenece
el nitroprusiato de sodio. Los donadores de NO pueden ser clasificados en 6
categorías (Figura 9) basados en el átomo al que está unido el resto de la
molécula liberadora de NO (Hou y col., 1999).
Figura 9. Diagrama de los principales grupos de compuestos donadores de óxido nítrico.
2.6.5 Complejos de transición metal-óxido nítrico
Representan una clase importante de donadores de óxido nítrico. En este
tipo de compuestos el NO actúa como un poderoso ligando, en el cual, el
nitrógeno, se une al metal. Los principales compuestos de este tipo usados en
investigación se mencionan en la figura 9.
DONADORES DE ÓXIDO NÍTRICO
C-NO N-NO O-NO S-NO DONADORES HETEROCÍCLICOS
COMPLEJOS DE TRANSICIÓN
METAL-ÓXIDO NÍTRICO

28
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 10. Principales compuestos del grupo complejos de transición metal-óxido nítrico, tomado de Hou y col., 1999.
2.6.5.1 Nitroprusiato o nitroprusida de sodio (NPS)
Es el compuesto más ampliamente estudiado de la familia de los nitrosilos de
hierro, su nombre sistemático según la IUPAC (Unión Internacional de Química
Pura y Aplicada, por sus siglas en inglés) es pentacianidonitrosilferrato de sodio II,
un complejo inorgánico donde el hierro está en estado ferroso (Fe2-) y el óxido
nítrico se une formalmente como NO+. A pesar de estudios intensivos de su
reactividad química, en particular, con tioles, el mecanismo de liberación de NO
sigue siendo claramente no entendido. Es claro, sin embargo, que el NPS requiere
tanto la irradiación con luz o la reducción de un electrón para liberar NO (Feelisch,
1998). El NPS en solución acuosa, se degrada cuando se expone a la luz blanca o
azul, pero no a la luz roja, aun cuando se degrada los subproductos siguen siendo
biológicamente activos (Arnold y col., 1984).
2.6.5.2 NPS en el cultivo in vitro
La adición de NPS en el cultivo in vitro es actualmente un tema de interés.
Ötvös y colaboradores (2005) utilizaron NPS como donador de óxido nítrico en
combinación con auxinas como promotor de la activación de la división celular y la
formación de células embriogénicas en Medicago sativa. Dentro de sus resultados

29
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
ellos postulan que en ausencia de una auxina endógena y con una concentración
de 10 M de NPS no hay influencia para la división celular. Sin embargo en
presencia tanto de 0.22 y 1 M de 2,4-D y la misma cantidad de NPS incrementa
significativamente la frecuencia de células en división y viables.
Un estudio más reciente sin relación con el género anterior pero
perteneciente al mismo orden que Polianthes fue realizado por Tan y
colaboradores en el 2013, donde evaluaron el efecto del NPS en la multiplicación
de brotes y regeneración de Vanilla planifolia Andrews. En ambos estudios se
utilizó una concentración de 10 µM de NPS adicionado al medio MS y las
concentraciones de auxinas y citocininas fueron muy diferentes. En el caso de la
producción de brotes en Vanilla ellos reportaron que el NO estimuló el desarrollo
de los brotes y fue un intermediario en la regeneracion de brotes adventicios y
raíces como se ha sugerido para otras especies como Cucumis sativus donde
adicionó la misma concentración y en Solanum lycopersicum donde la
concentración óptima fue de 200 µM (Correa-Aragunde y col., 2006; Pagnussat y
col., 2002).
En relación con las concetraciones de NPS usadas hay un rango amplio en
la bibliografía reportada pues en el caso de He y colaboradores (2004) los cuales
evaluaron la respuesta del NO en la trancisión a la etapa de floración donde sus
tratamientos de semillas germinadas en medio con NPS elevaron su crecimiento
vegetativo y retrasaron su floración. NPS incrementó el crecimiento de brotes en
concentraciones menores o iguales a 100 µM de NPS y lo inhibió en
concentraciones por encima de la mencionada. La concentración óptima de NPS
para promover el crecimiento de brotes fue aproximadamente de 100 µM.
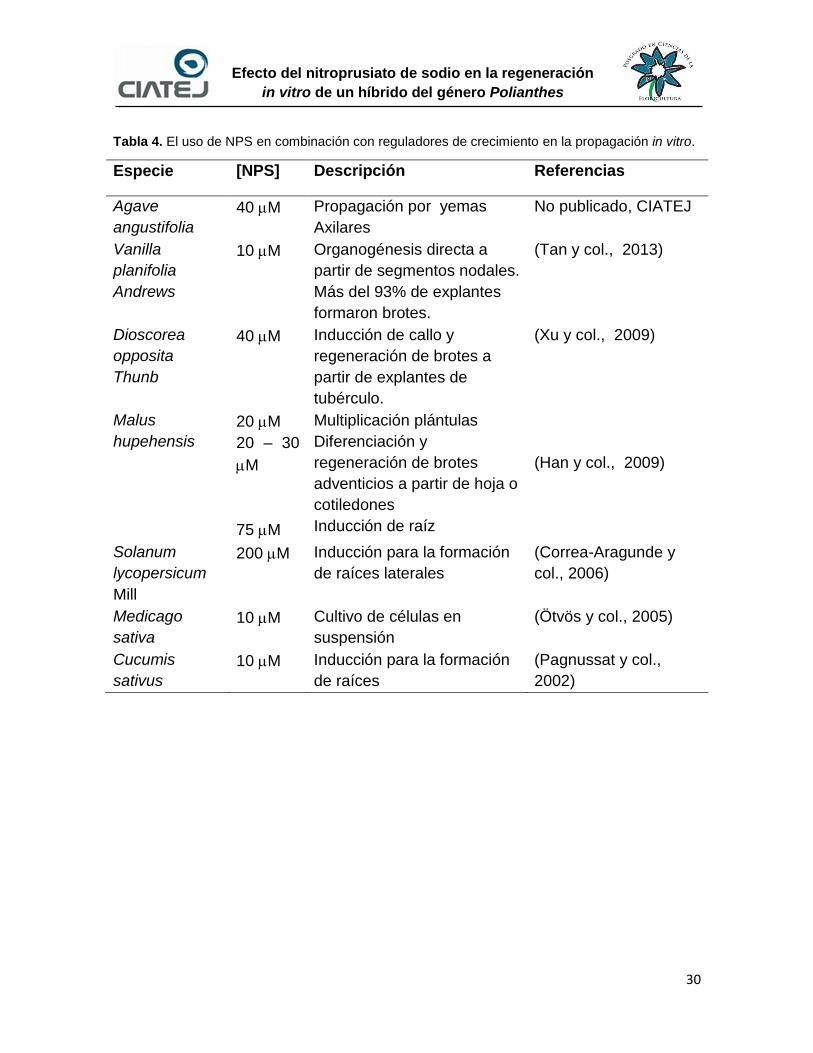
En la tabla 4 se agrupan otros trabajos realizados. En conclusión la
respuesta de la planta al efecto por el nitroprusiato de sodio no es característica
de un solo género en particular, lo que permite sugerir que el uso de NPS pudiera
tener un efecto sobre el cultivo in vitro de Polianthes.
30
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Tabla 4. El uso de NPS en combinación con reguladores de crecimiento en la propagación in vitro.
Especie [NPS] Descripción Referencias
Agave
angustifolia
40 M Propagación por yemas
Axilares
No publicado, CIATEJ
Vanilla
planifolia
Andrews
10 M Organogénesis directa a
partir de segmentos nodales.
Más del 93% de explantes
formaron brotes.
(Tan y col., 2013)
Dioscorea
opposita
Thunb
40 M Inducción de callo y
regeneración de brotes a
partir de explantes de
tubérculo.
(Xu y col., 2009)
Malus
hupehensis
20 M
20 – 30
M
75 M
Multiplicación plántulas
Diferenciación y
regeneración de brotes
adventicios a partir de hoja o
cotiledones
Inducción de raíz
(Han y col., 2009)
Solanum
lycopersicum
Mill
200 M Inducción para la formación
de raíces laterales
(Correa-Aragunde y
col., 2006)
Medicago
sativa
10 M Cultivo de células en
suspensión
(Ötvös y col., 2005)
Cucumis
sativus
10 M Inducción para la formación
de raíces
(Pagnussat y col.,
2002)

31
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
II. JUSTIFICACIÓN
El género Polianthes es endémico de México, su uso comercial principal es
como planta ornamental (como flor de corte), además se utiliza en la industria del
perfume y farmacéutica. En el país, el cultivo de Polianthes representa una
importante fuente de generación de ingresos, en el año 2014 se reportó un valor
de producción de 47 millones de pesos (SAGARPA, 2014). Sin embargo, para
producir la variedad comercial de Polianthes (P. tuberosa) se utiliza la
reproducción asexual mediante bulbos, disminuyendo la variabilidad genética y
produciendo plantas con inflorescencias de color blanco. Por lo tanto, es necesario
implementar protocolos de cultivo in vitro que permitan propagar nuevas especies
silvestres o híbridos del género Polianthes con características especiales (por ej.
modificación de color de flor, aroma, tamaño, etc.) que las hagan atractivas para
un nicho de mercado, además, de propagar especies en peligro de extinción y
conservar la diversidad genética nativa.
Actualmente, existe un número limitado de trabajos de investigación
utilizados para propagar Polianthes, en su mayoría protocolos de
micropropagación a partir de segmentos de bulbo y en menor medida protocolos
de embriogénesis somática. Estos protocolos han sido probados
satisfactoriamente utilizando medio MS y diferentes reguladores de crecimiento.
Por otro lado, en otras especies vegetales se ha utilizado exitosamente el NPS
para promover la iniciación de células embriogénicas y/o la generación de brotes.
Sin embargo, su uso no ha sido investigado en el cultivo in vitro del género
Polianthes. Debido a esto, en el presente trabajo se estableció un protocolo de
embriogénesis somática y micropropagación para reproducir un híbrido de
Polianthes evaluando el efecto de la adición de NPS. Esto permitirá obtener un
mayor número de plantas en un menor tiempo y contribuirá a la obtención de
registros de nuevas variedades.

32
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
IV. HIPÓTESIS
La adición de nitroprusiato de sodio al medio de cultivo in vitro, sólo o en
combinación con reguladores de crecimiento favorece la iniciación de células
embriogénicas y/o estimula la producción de brotes en el cultivo in vitro de un
híbrido del género Polianthes.

33
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
V. OBJETIVOS
5.1 Objetivo general
Evaluar el efecto de la adición de nitroprusiato de sodio sobre la
regeneración in vitro de un híbrido del género Polianthes mediante la
embriogénesis somática y proliferación de yemas axilares.
5.2 Objetivos específicos
1. Obtener tejido juvenil a través de la germinación de semillas del híbrido.
2. Evaluar el efecto de la adición de NPS sobre la desdiferenciación de tejido de
hoja para la inducción de embriogénesis somática.
3. Inducir el proceso de embriogénesis somática utilizando medios cultivo
específicos para la etapa de inducción y de expresión.
4. Establecer y propagar el híbrido mediante la técnica de yemas axilares
utilizando la inflorescencia.
5. Evaluar el efecto de la adición de NPS sobre la producción de brotes
utilizando plántulas del híbrido o explantes generados por la técnica de
yemas axilares de la inflorescencia del hibrido, así como su efecto sobre la
síntesis de clorofila.

34
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
VI. METODOLOGÍA EXPERIMENTAL
Para cumplir con los objetivos planteados en el presente proyecto de
investigación, la metodología experimental fue dividida en dos etapas (Figura 11).
Durante la primera etapa se realizaron estudios de embriogénesis para evaluar el
efecto de NPS sobre la inducción de embriones somáticos a partir del tejido de
hoja. Al inicio se efectuó la germinación de semillas del híbrido 1008157 P.
howardii x P. tuberosa (proporcionado el Dr. Ernesto Tapia Campos-CIATEJ) para
obtener tejido juvenil. Este tejido fue utilizado para evaluar el efecto de la
combinación entre NPS y un regulador de crecimiento perteneciente al grupo de
las auxinas (picloram) sobre la desdiferenciación del tejido vegetal. Debido a que
después del proceso de desdiferenciación se observó la formación de estructuras
relacionadas con la formación de embriones, como lo son las estructuras
globulares y callos friables se procedió a realizar la comprobación a través de una
técnica de doble tinción diferencial con el fin de detectar estructuras
embriogénicas y polarización celular. Una vez corroborada la presencia de células
potencialmente embriogénicas se realizaron resiembras a medios de inducción
para seguir estimulando el proceso de diferenciación. Primero, se resembró el
tejido utilizando medios de cultivo que contenían reguladores de crecimiento (2,4-
D y BA) en diferentes concentraciones. Después, el tejido que presentó viabilidad
embriogénica vinculada a la morfología estructural (estructuras globulares) fue
resembrado en un medio que contenía (ANA y BA) como reguladores de
crecimiento y enriquecido con aminoácidos para favorecer selectivamente la
formación de estructuras embriogénicas. Debido a que las estructuras que se
formaron una vez concluido el proceso de desdiferenciación de tejido e inducción
de embriogénesis sólo presentaron un estadio globular se procedió a realizar una
resiembra utilizando un medio de expresión sin reguladores de crecimiento y
reduciendo la concentración de nutrientes, esto con el fin de coadyuvar el avance
en los estadios que componen el proceso de desarrollo de un embrión somático a
través del estrés inducido por limitación de nutrientes.
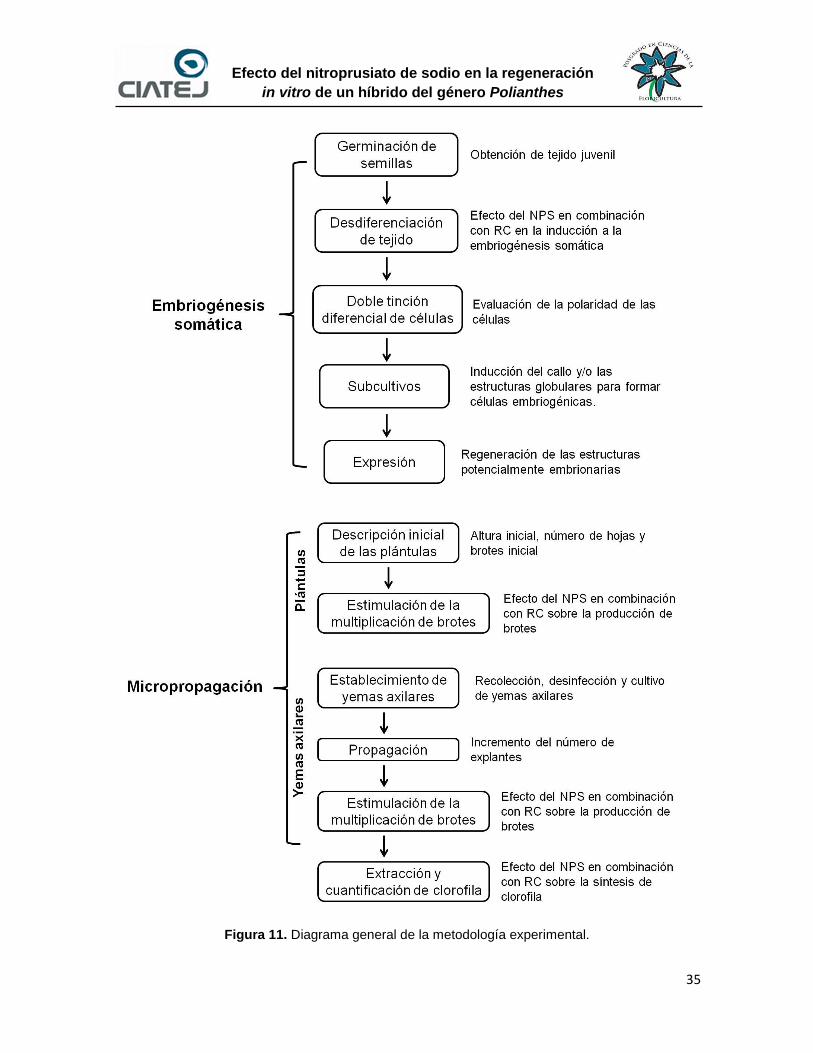
35
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 11. Diagrama general de la metodología experimental.

36
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Con el objetivo de incrementar el número de clones del híbrido 1008157 P.
howardii x P. tuberosa durante la segunda etapa, se realizaron estudios de
micropropagación. En estos estudios se evaluó el efecto de la concentración de
NPS sobre la estimulación de la producción de brotes utilizando plántulas y yemas
axilares florales. Respecto a las plántulas, éstas primeramente fueron
seleccionadas y caracterizadas morfológicamente. Por otra parte, se colectaron
yemas axilares de la inflorescencia del híbrido, para obtener brotes vegetativos a
partir de estas yemas, estas fueron desinfectadas y cultivadas utilizando un medio
de propagación con reguladores de crecimiento (BA y 2,4-D). Los brotes
vegetativos resultantes se propagaron con la finalidad de incrementar el número
de explantes, y así disponer de material suficiente para evaluar el efecto de la
adición de NPS sobre la multiplicación de brotes y la síntesis de clorofila.
Finalmente, la evaluación del efecto de NPS sobre la estimulación de la
producción de brotes se realizó de manera independiente tanto en plántulas como
en yemas axilares.
6.1 Embriogénesis somática
6.1.1 Germinación de semillas
200 semillas del híbrido 1008157 provenientes de polinización abierta fueron
utilizadas para la germinación (Figura 12). Las semillas se desinfectaron con
hipoclorito de sodio comercial (Cloralex) diluido al 3% v/v, para ello se agregó 10
mL de NaClO más dos gotas de detergente líquido a un tubo tipo Falcon (50 mL)
estéril, el cual fue agitado vigorosamente durante 15 min. Después, se aplicó
lavado por triplicado con agua destilada estéril en periodos de 5 min.
Posteriormente, para prevenir contaminación microbiana, las semillas se lavaron
con una solución de PPM (2 mL/L) (Plant Preservative Mixture, Plant Cell
Technology, USA) durante 10 min. Transcurrido el tiempo, se retiró la solución de
PPM y se adicionó una solución de cefotaxima (300 mg/L) durante 1 h.

37
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Finalmente, se retiró completamente el exceso de esta solución y las semillas
fueron vertidas sobre una caja Petri estéril.
Una vez desinfectadas las semillas, estas fueron cultivadas en frascos de
vidrio con medio GE sin reguladores de crecimiento. Cada frasco contenía 4
semillas y 25 mL de medio GE que contenía 10 mL/L de medio MS (Murashige y
Skoog, 1962) (concentrado 10 veces), 30 g/L de sacarosa, 8 g/L de agar. Se
ajustó el pH a 5.8 utilizando HCl o NaOH (ambos 1N). El medio fue esterilizado
utilizando un autoclave a 121 °C durante 20 min. Aunque la germinación de las
plántulas ocurrió durante los primeros 5 d, el tiempo de desarrollo del cultivo duró
60 d, tiempo necesario para observar una altura de las plántulas mayor a 7 cm.
Los frascos se incubaron a una temperatura de 27 °C y con un fotoperiodo de 16
h luz y 8 h oscuridad.
Figura 12. Proceso de germinación de las semillas: (a) desinfección; (b); vertido en placa estéril; (c) cultivo en medio GE y (d) plántulas germinadas con 30 d de cultivo.
6.1.2 Desdiferenciación de tejido somático de plántulas del híbrido
Después de la germinación, las plántulas fueron retiradas del medio GE. A
cada plántula se le cortaron las hojas en segmentos pequeños de
aproximadamente 0.5 cm. Es importante mencionar que únicamente se utilizó
tejido de hojas excluyéndose la raíz. Los segmentos se sembraron verticalmente
cuidando de no hundirlos completamente en el medio de cultivo (Figura 13). Este
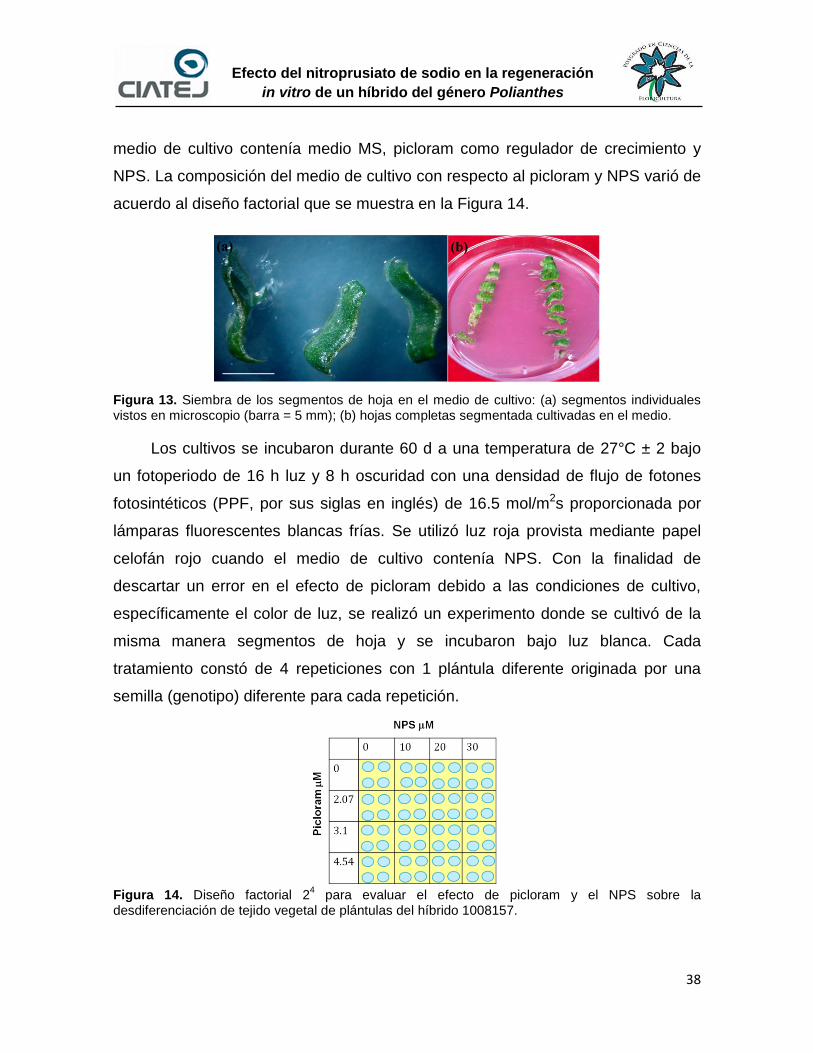
38
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
medio de cultivo contenía medio MS, picloram como regulador de crecimiento y
NPS. La composición del medio de cultivo con respecto al picloram y NPS varió de
acuerdo al diseño factorial que se muestra en la Figura 14.
Figura 13. Siembra de los segmentos de hoja en el medio de cultivo: (a) segmentos individuales vistos en microscopio (barra = 5 mm); (b) hojas completas segmentada cultivadas en el medio.
Los cultivos se incubaron durante 60 d a una temperatura de 27°C ± 2 bajo
un fotoperiodo de 16 h luz y 8 h oscuridad con una densidad de flujo de fotones
fotosintéticos (PPF, por sus siglas en inglés) de 16.5 mol/m2s proporcionada por
lámparas fluorescentes blancas frías. Se utilizó luz roja provista mediante papel
celofán rojo cuando el medio de cultivo contenía NPS. Con la finalidad de
descartar un error en el efecto de picloram debido a las condiciones de cultivo,
específicamente el color de luz, se realizó un experimento donde se cultivó de la
misma manera segmentos de hoja y se incubaron bajo luz blanca. Cada
tratamiento constó de 4 repeticiones con 1 plántula diferente originada por una
semilla (genotipo) diferente para cada repetición.
Figura 14. Diseño factorial 2
4 para evaluar el efecto de picloram y el NPS sobre la
desdiferenciación de tejido vegetal de plántulas del híbrido 1008157.

39
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
6.1.3 Doble tinción diferencial de células
Para conocer si el callo y/o las estructuras globulares formadas a partir de la
hoja en los tratamientos del diseño factorial poseen células potencialmente
embriogénicas se realizó la técnica de doble tinción diferencial de acuerdo a
Portillo y colaboradores (2007). Se colocó una muestra de callo de
aproximadamente 8 mm3 en un tubo Eppendorf (1.5 mL), cuando la concistencia
del callo era dura fue necesario disgregar el tejido cuidando no aplicar una fuerza
excesiva. La muestra se suspendió en 0.5 mL de agua destilada y se agregaron 3
gotas de acetocarmin al 0.5%, luego, se llevó a baño María a 50°C durante 30 s.
Posteriormente, las células fueron lavadas tres veces con agua destilada para
remover el exceso de colorante. Después, a la muestra ya teñida se le agregaron
dos gotas de azul de Evans al 0.5% y se dejó reposar durante 30 s.
Inmediatamente, se lavó tres veces con agua destilada para remover el exceso de
colorante. La muestra doblemente teñida se resuspendió en 0.5 mL de agua
destilada y fue observada en el microscopio Olympus BH-2 RFCA acoplado a una
cámara Leica® DFC 450C.
6.1.4 Subcultivos de callo y/o estructuras potencialmente
embriogénicas
6.1.4.1 Subcultivo utilizando medios de inducción
Después del tiempo de desdiferenciación del tejido (60 d) se resembró el
material vegetal que presentó formación de callo y/o estructuras globulares para
continuar con la inducción del proceso de embriogénesis. Las muestras fueron
cultivadas utilizando medios de inducción: medio MIA para Agave y medio MIP
para Polianthes. El tejido desdiferenciado se incubó durante 60 d utilizando luz
blanca bajo las mismas condiciones descritas en la sección 5.1.2. El medio MIA
contenía 100 mL/L de medio MS (Murashige y Skoog, 1962) (concentrado 10
veces), 30 g/L de sacarosa, 8 g/L de agar, 1.3 M de BA y 9.0 M de 2,4-D.

40
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Además, el pH fue ajustado a 5.8 utilizando HCl o NaOH (ambos 1N). El
medio fue esterilizado utilizando un autoclave a 121 °C durante 20 min. Por otra
parte, el medio MIP contenía 100 mL/L de medio MS (Murashige y Skoog, 1962)
(concentrado 10 veces), 30 g/L de sacarosa, 8 g/L de agar, 0.1 mg/L de BA y 2.5
mg/L de 2,4-D. Se ajustó el pH a 5.8 utilizando HCl o NaOH (ambos 1N). Este
medio también fue esterilizado en autoclave a 121 °C durante 20 min.
6.1.4.2 Subcultivo utilizando medio de inducción enriquecido con
aminoácidos
Una vez transcurrido el tiempo de incubación descrito en la sección anterior,
algunos tratamientos presentaron estructuras potencialmente embriogénicas,
particularmente estructuras asociadas a los estadios embrionarios. Estas
estructuras permanecieron en forma globular y no continuaron su proceso de
desarrollo, debido a esto, el tejido fue resembrado en un medio de cultivo de
inducción (MIc20) enriquecido con aminoácidos e incubado en oscuridad durante
30 d a una temperatura de 27°C ± 2. El medio MIc20 contenía medio MS sin
vitaminas, 30 g/L de sacarosa, 8 g/L de agar, en mg/L: L-asparagina 10, L-arginina
10, L-ác. aspártico 7.5, glicina 23, glutamina 60, ác. glutámico 7.5, biotina (B12) 1,
ác. fólico 1, ác. nicotínico 1.5, piridoxina 1.5, riboflavina 0.1, tiamina 3, myo-
inositol 145, urea 45, BA 1 y ANA 0.5. Se ajustó el pH a 5.8 utilizando HCl o NaOH
(ambos 1N). Para preparar medio Mic20, una parte del medio la cual contenía
medio MS sin vitaminas, sacarosa, agar y RC y fue esterilizado en autoclave (121
°C durante 20 min). Otra parte que contenía la solución de aminoácidos se
esterilizó en frio (microfiltración, 0.22 m), a continuación se mezclaron ambas
soluciones.
6.1.5 Subcultivo a medio de expresión
Para corroborar la viabilidad celular embrionaria, pseudoembriones en
estado globular y callo friable fueron transferidos a un medio de expresión (ME)
durante 30 d utilizando luz blanca y condiciones idénticas de temperatura y

41
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
fotoperiodo descritas en la sección de germinación. El medio ME únicamente
contenía 100 mL/L de medio MS (Murashige y Skoog, 1962) (concentrado 10
veces), 30 g/L de sacarosa, 6 g/L de Phytagel, 250 mg/L hidrolizado de caseína y
500 mg/L glutamina, posteriormente, el pH fue ajustado a 5.8 utilizando HCl o
NaOH (ambos 1N) y fue esterilizado utilizando un autoclave a 121 °C durante 20
min.
6.2 Micropropagación
6.2.1 Descripción inicial de plántulas
En la etapa de embriogénesis se germinaron 200 semillas del híbrido
1008157, las plántulas derivadas de la germinación se utilizarón para la realización
de estudios de micropropagación. En esta segunda etapa el objetivo fue evaluar el
efecto de la concentración de NPS sobre la inducción de nuevos brotes en
plántulas. Las plántulas sin hojas obtenidas después del proceso de diferenciación
de tejido fueron resembradas en medio TDZ durante 30 d (Figura 15). El medio
TDZ contenía 100 mL/L de medio MS concentrado, 30 g/L de sacarosa, 8 g/L de
agar y 0.25 mg/L de tidiazuron (TDZ). Se ajustó el pH a 5.8 utilizando HCl o NaOH
(ambos 1N) y, fue esterilizado utilizando un autoclave a 121 °C durante 20 min,
excepto el TDZ que fue esterilizado a través de microfiltración utilizando una
membrana de 0.22 m. La resiembra se realizó con el propósito de fortalecer y
acelerar el crecimiento de las plántulas. Cuando se observó la recuperación
(crecimiento) de las plántulas, 75 de ellas fueron seleccionadas para realizar la
evaluación del efecto de NPS sobre la producción de brotes. Estas plántulas
seleccionadas se describieron morfológicamente (Tabla 5) cuantificando el número
de hojas, número de brotes y altura de la planta (tomando como referencia la hoja
de mayor longitud del meristemo central).
42
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 15. Plántulas resembradas en medio TDZ después de 30 d de cultivo.
Tabla 5. Descripción inicial de las 75 plántulas del híbrido 1008157.
No. Código de identificación Altura (cm) No. Hojas No. Brotes
1 1 2.2 3 0
2 2 2.5 14 3
3 3 2.4 5 1
4 4 5.8 4 0
5 5 4.7 19 3
6 6 3.3 5 0
7 7 5.8 17 2
8 8 5.5 4 0
9 9 3.0 5 0
10 10 4.4 7 1
11 11 1.3 8 2
12 12 2.4 4 0
13 14 2.1 4 0
14 15 2.0 5 1
15 21 3.2 4 0
16 22 2.5 5 0
17 23 4.1 4 0
18 24 1.1 2 1
19 25 3.2 6 1
20 26 1.9 3 0
21 27 1.9 7 1
22 28 1.0 4 0
23 30 1.8 5 1
24 31 0.6 2 0
Continuación…
43
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
No. Código de identificación Altura (cm) No. Hojas No. Brotes
25 32 1.5 2 0
26 33 1.4 3 0
27 34 1.2 7 1
28 35 2.2 4 0
29 36 2.3 4 0
30 37 1.3 1 0
31 40 2.0 2 0
32 41 7.0 7 1
33 42 3.2 4 0
34 43 1.8 4 0
35 44 4.0 8 2
36 45 4.3 10 1
37 46 1.7 2 0
38 47 4.8 10 2
39 48 1.4 3 0
40 49 8.1 7 1
41 50 1.3 3 0
42 51 2.5 5 1
43 52 1.3 8 1
44 53 1.9 13 3
45 54 5.0 6 1
46 61 9.5 8 1
47 62 6.2 4 0
48 63 5.0 12 1
49 64 0.8 5 0
50 65 1.6 4 0
51 66 1.5 5 3
52 70 2.8 4 0
53 71 1.7 7 1
54 72 2.7 6 1
55 73 2.2 7 1
56 74 3.4 4 0
57 75 0.7 3 0
58 76 7.5 9 1
59 77 2.5 4 0
60 84 3.9 10 2
Continuación…
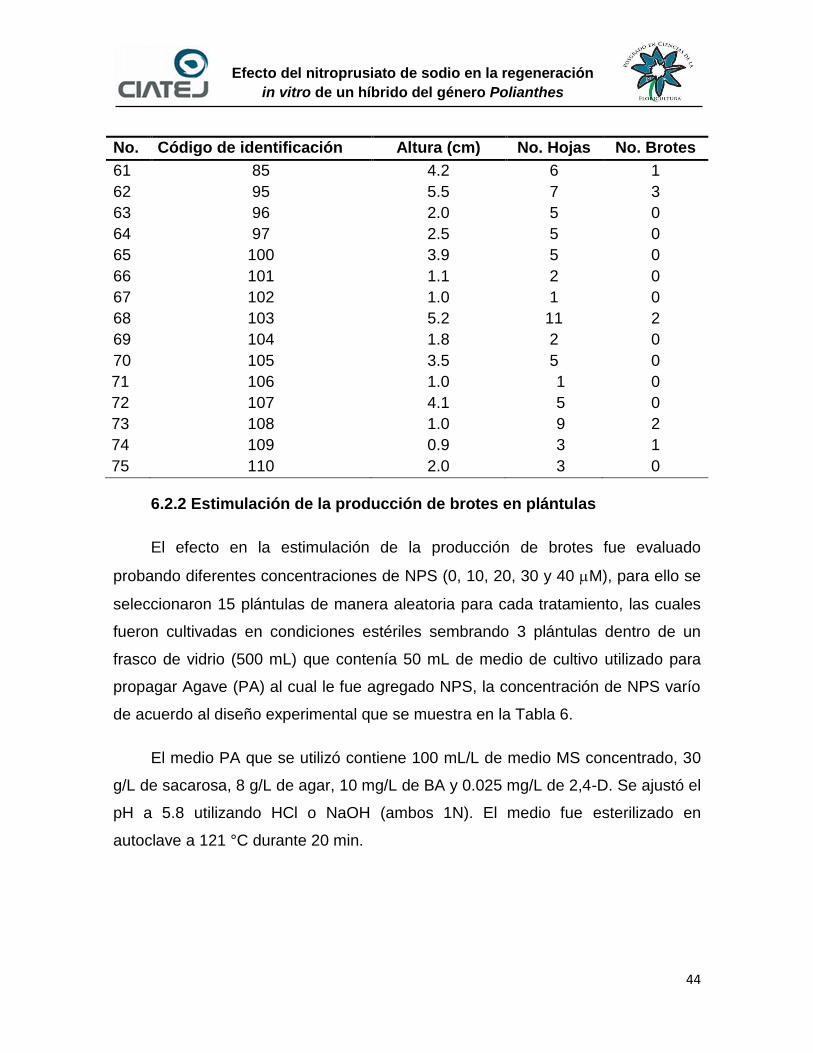
44
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
No. Código de identificación Altura (cm) No. Hojas No. Brotes
61 85 4.2 6 1
62 95 5.5 7 3
63 96 2.0 5 0
64 97 2.5 5 0
65 100 3.9 5 0
66 101 1.1 2 0
67 102 1.0 1 0
68 103 5.2 11 2
69 104 1.8 2 0
70 105 3.5 5 0
71 106 1.0 1 0
72 107 4.1 5 0
73 108 1.0 9 2
74 109 0.9 3 1
75 110 2.0 3 0
6.2.2 Estimulación de la producción de brotes en plántulas
El efecto en la estimulación de la producción de brotes fue evaluado
probando diferentes concentraciones de NPS (0, 10, 20, 30 y 40 M), para ello se
seleccionaron 15 plántulas de manera aleatoria para cada tratamiento, las cuales
fueron cultivadas en condiciones estériles sembrando 3 plántulas dentro de un
frasco de vidrio (500 mL) que contenía 50 mL de medio de cultivo utilizado para
propagar Agave (PA) al cual le fue agregado NPS, la concentración de NPS varío
de acuerdo al diseño experimental que se muestra en la Tabla 6.
El medio PA que se utilizó contiene 100 mL/L de medio MS concentrado, 30
g/L de sacarosa, 8 g/L de agar, 10 mg/L de BA y 0.025 mg/L de 2,4-D. Se ajustó el
pH a 5.8 utilizando HCl o NaOH (ambos 1N). El medio fue esterilizado en
autoclave a 121 °C durante 20 min.
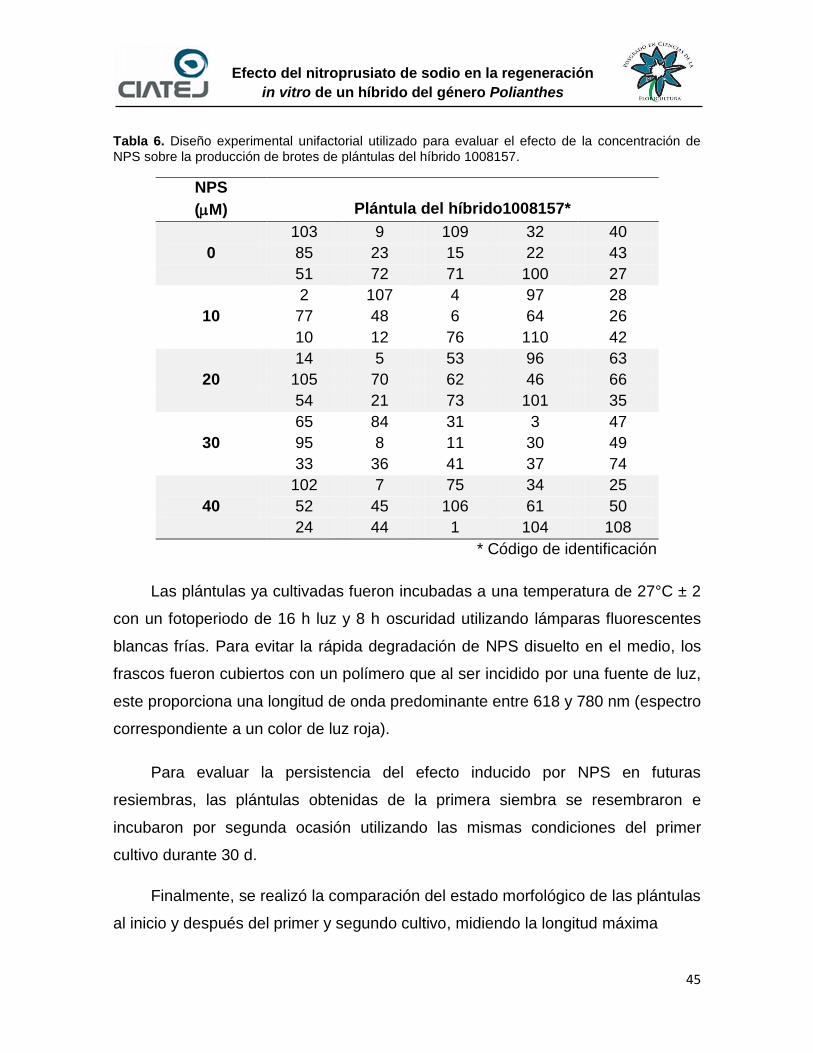
45
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Tabla 6. Diseño experimental unifactorial utilizado para evaluar el efecto de la concentración de
NPS sobre la producción de brotes de plántulas del híbrido 1008157.
NPS
(M)
Plántula del híbrido1008157*
0
103 9 109 32 40
85 23 15 22 43
51 72 71 100 27
10
2 107 4 97 28
77 48 6 64 26
10 12 76 110 42
20
14 5 53 96 63
105 70 62 46 66
54 21 73 101 35
30
65 84 31 3 47
95 8 11 30 49
33 36 41 37 74
40
102 7 75 34 25
52 45 106 61 50
24 44 1 104 108
* Código de identificación
Las plántulas ya cultivadas fueron incubadas a una temperatura de 27°C ± 2
con un fotoperiodo de 16 h luz y 8 h oscuridad utilizando lámparas fluorescentes
blancas frías. Para evitar la rápida degradación de NPS disuelto en el medio, los
frascos fueron cubiertos con un polímero que al ser incidido por una fuente de luz,
este proporciona una longitud de onda predominante entre 618 y 780 nm (espectro
correspondiente a un color de luz roja).
Para evaluar la persistencia del efecto inducido por NPS en futuras
resiembras, las plántulas obtenidas de la primera siembra se resembraron e
incubaron por segunda ocasión utilizando las mismas condiciones del primer
cultivo durante 30 d.
Finalmente, se realizó la comparación del estado morfológico de las plántulas
al inicio y después del primer y segundo cultivo, midiendo la longitud máxima

46
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
(tomando como referencia la hoja de mayor longitud del meristemo central) así
como el número de hojas y brotes.
6.2.3 Establecimiento de yemas axilares de la inflorescencia del híbrido
1008157
Para el establecimiento de yemas axilares (Figura 16) se realizó una colecta
de tallos de la inflorescencia del híbrido 1008157. Los tallos se cortaron en 50
segmentos internodales de 2 a 3 cm de longitud los cuales presentaban yemas
axilares florales. Los segmentos fueron desinfectados, para ello se lavaron con
hipoclorito de sodio comercial (10% v/v) durante 15 min, después se les retiró la
solución de hipoclorito de sodio, se enjuagaron con agua destilada y se colocaron
dentro de un frasco de vidrio (1 L) estéril con una solución de cefotaxima (600
mg/L) con PPM (2 mL/L) agitándose durante 24 h. Posteriormente, se enjuagaron
con agua destilada estéril tres veces, después, las yemas fueron cultivadas
durante 120 d en frascos de vidrio con medio estéril PA, manteniendo las
siguientes condiciones de incubación: temperatura 27°C ± 2, fotoperiodo de 16 h
luz y 8 h oscuridad y PPF de 16.5 mol/m2s. Posterior al último cultivo, los
segmentos se subcultivaron en medio TDZ y se incubaron bajo las condiciones
mencionadas por 90 d hasta cuando presentaron respuesta.
Figura 16. Proceso de establecimiento de las yemas axilares de la inflorescencia del híbrido 1008157: (a) colecta; (b) adición de cefotaxima y PPM; (c) yemas axilares desinfectadas.

47
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
6.2.4 Propagación de clones del híbrido 1008157
Para incrementar el número de clones del híbrido se utilizaron los brotes
vegetativos resultantes del establecimiento de yemas axilares florales. Estos
brotes fueron separados del tallo madre, luego, se dividieron en varios segmentos
y se cultivaron durante 202 d utilizando un medio diferente como se muestra en la
Tabla 7. Los explantes cultivados en medio MSC fueron incubados en oscuridad
durante 12 d.
Tabla 7. Propagación de plantas del híbrido.
Medio de cultivo Fecha
PA 16/Abril/15
PA 16/Mayo/15
TDZ 03/Julio/15
PA 07/Agosto/15
10 NPS 17/Septiembre/15
MSC 23/Octubre/15
*El medio 10 NPS contenía 100 mL/L de medio MS concentrado, 30 g/L de sacarosa, 8 g/L de
agar, 10 mg/L de BA y 0.025 mg/L de 2,4-D y 10M de NPS. Se ajustó el pH a 5.8 utilizando HCl o NaOH (ambos 1N). El medio fue esterilizado utilizando una autoclave a 121 °C durante 20 minutos excepto el NPS que fue esterilizado en frio. **El medio MSC contenía 100 mL/L de medio MS concentrado, 30 g/L de sacarosa y 8 g/L de agar. Se ajustó el pH a 5.8 utilizando HCl o NaOH (ambos 1N). El medio fue esterilizado utilizando una autoclave a 121 °C durante 20 min.
6.2.5 Estimulación de la producción de brotes en clones del híbrido
1008157
Una vez propagados los brotes se obtuvo un número suficiente de explantes,
los cuales fueron cultivados para evaluar el efecto de la combinación de NPS y
reguladores de crecimiento sobre la producción de brotes a partir de clones del
híbrido. Dicha evaluación se llevó a cabo en dos experimentos, en la primera se
utilizaron explantes que presentaban al menos un brote (Tabla 8) y en la segunda
se utilizaron explantes individuales, los cuales carecían de brotes (Tabla 9). Se
realizaron 4 y 6 repeticiones por concentración de NPS para el primer y segundo
experimento respectivamente (Figura 17). Los cultivos fueron incubados durante
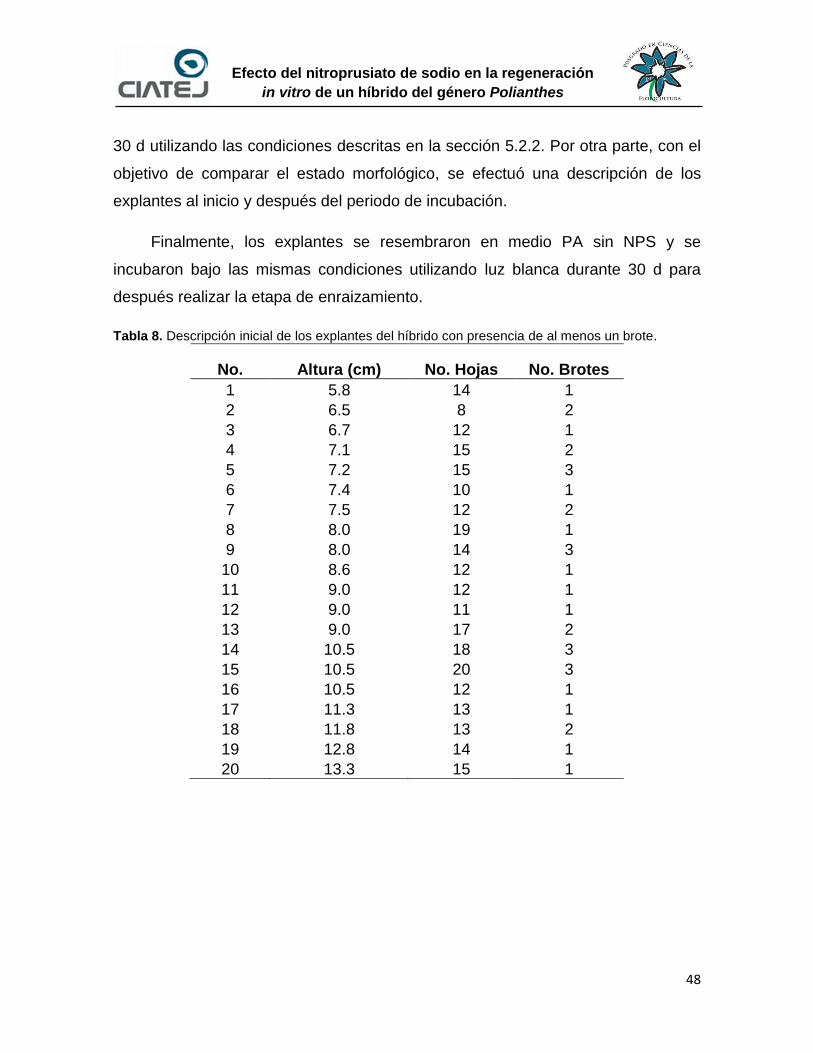
48
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
30 d utilizando las condiciones descritas en la sección 5.2.2. Por otra parte, con el
objetivo de comparar el estado morfológico, se efectuó una descripción de los
explantes al inicio y después del periodo de incubación.
Finalmente, los explantes se resembraron en medio PA sin NPS y se
incubaron bajo las mismas condiciones utilizando luz blanca durante 30 d para
después realizar la etapa de enraizamiento.
Tabla 8. Descripción inicial de los explantes del híbrido con presencia de al menos un brote.
No. Altura (cm) No. Hojas No. Brotes
1 5.8 14 1
2 6.5 8 2
3 6.7 12 1
4 7.1 15 2
5 7.2 15 3
6 7.4 10 1
7 7.5 12 2
8 8.0 19 1
9 8.0 14 3
10 8.6 12 1
11 9.0 12 1
12 9.0 11 1
13 9.0 17 2
14 10.5 18 3
15 10.5 20 3
16 10.5 12 1
17 11.3 13 1
18 11.8 13 2
19 12.8 14 1
20 13.3 15 1
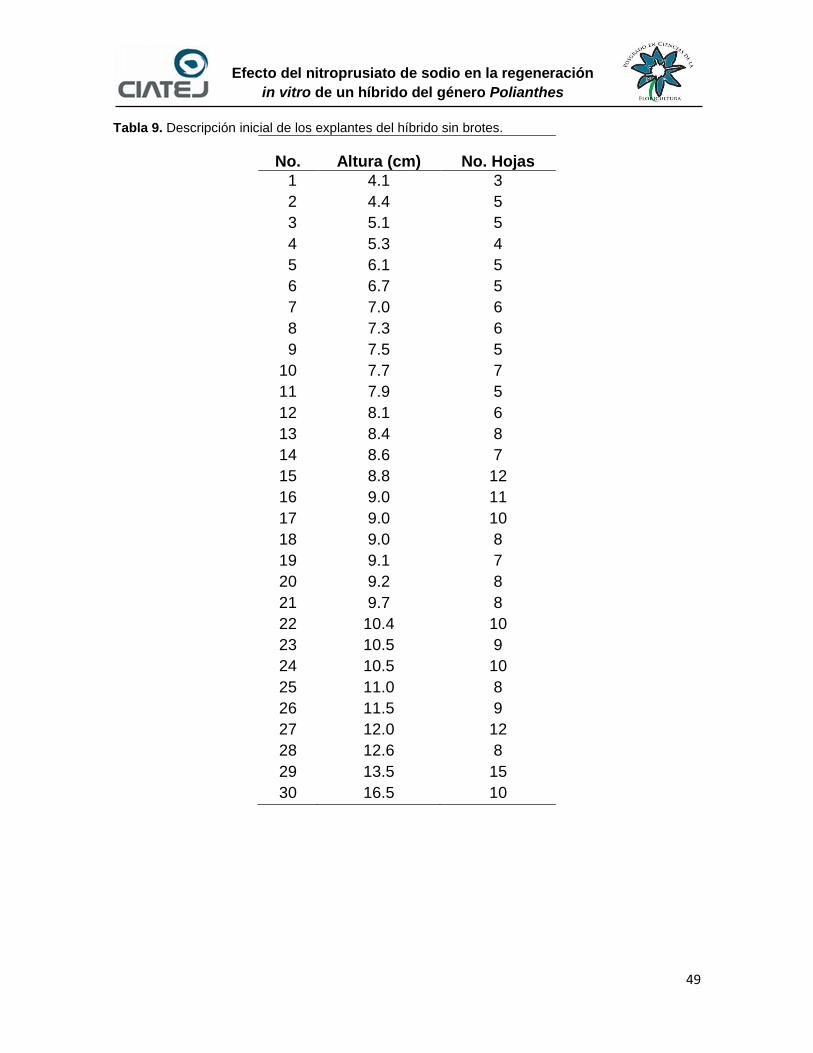
49
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Tabla 9. Descripción inicial de los explantes del híbrido sin brotes.
No. Altura (cm) No. Hojas
1 4.1 3
2 4.4 5
3 5.1 5
4 5.3 4
5 6.1 5
6 6.7 5
7 7.0 6
8 7.3 6
9 7.5 5
10 7.7 7
11 7.9 5
12 8.1 6
13 8.4 8
14 8.6 7
15 8.8 12
16 9.0 11
17 9.0 10
18 9.0 8
19 9.1 7
20 9.2 8
21 9.7 8
22 10.4 10
23 10.5 9
24 10.5 10
25 11.0 8
26 11.5 9
27 12.0 12
28 12.6 8
29 13.5 15
30 16.5 10

50
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 17. Diseño experimental unifactorial utilizado en la evaluación de multiplicación de brotes
adicionando NPS en diferentes concentraciones (0, 10, 20, 30 y 40 M); (a) explantes con al menos un brote; (b) explantes individuales.
6.2.6 Extracción y cuantificación de clorofila
Después de haber realizado la propagación de clones del híbrido 1008157,
se realizó una cuantificación de clorofila (a y b). La determinación de clorofila
permitió conocer el estado fisiológico de los explantes después de haber sido
sometidos a estrés ocasionado por la incubación en oscuridad durante 12 d. La
concentración de clorofila fue determinada siguiendo la metodología propuesta por
Bojovic y Stojanovic (2005). 500 mg de tejido fresco fueron cortados
(aproximadamente 0.5 cm) y molidos en un mortero, en seguida, se colocaron en
un tubo Falcon (50mL) y se agregó 10 mL de acetona al 80% (v/v).
Posteriormente, la mezcla se dejó macerar durante 30 min y fue centrifugada a
2500 rpm durante 5 min. Entonces, se tomó 1 mL del sobrenadante y se diluyó en
9 mL de acetona al 80% (v/v). La muestra diluida se analizó en un
espectrofotómetro UV-Vis (DR 5000, HACH) utilizando una longitud de onda de
664 y 647 para clorofila a y b respectivamente. Para calcular el contenido de
clorofila (a y b) se utilizó un conjunto de ecuaciones propuestas por Wellburn
(1994) (Tabla 10).

51
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Después de la estimulación de la producción de brotes en clones del híbrido,
se evaluó el efecto de la concentración de NPS sobre la síntesis clorofila. Para
extraer la clorofila se utilizó metanol, se tomaron tres discos (0.5 cm de diámetro)
de diferentes hojas del explante y se colocaron en tres tubos de ensayo de vidrio
de 10 mL. Se agregó 1 mL de metanol a cada tubo y se cubrió con Parafilm,
enseguida, los tubos fueron refrigerados a 4C durante 24 h. Pasado el tiempo, se
tomó una alícuota y se analizó por espectrofotometría utilizando un
espectrofotómetro para microplacas (Multiskan™ GO). De igual manera, el
contenido de clorofila (a y b) fue calculado utilizando el conjunto de ecuaciones
propuestas por Wellburn (1994).
Tabla 10. Ecuaciones para la determinación clorofila.
Solvente Ecuación (g/mL)
Acetona 80%
Metanol
6.3 Preparación de medios de cultivo con nitroprusiato de sodio.
Para la preparación de los medios de cultivo que contenían NPS (10, 20, 30,
40 M) se realizó el siguiente procedimiento. Primeramente, el medio de cultivo
MS con sacarosa, agar y RC fue esterilizado. Mientras el medio esterilizado
disminuía su temperatura, se preparó una solución stock de NPS (1x105M) la
cual se esterilizó en frio usando un filtro de membrana (0.22 m) y se mantuvo
protegida de la luz. En función de la concentración de NPS utilizada en cada
tratamiento, se tomó el volumen específico de la solución stock al medio de

52
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
cultivo. Dependiendo del objetivo, se vacío el medio sobre cajas Petri o en frascos
de vidrio estériles. Inmediatamente, para evitar la degradación del compuesto por
la luz (solar o blanca) se cubrieron con plástico de color rojo o se mantuvieron en
oscuridad hasta su uso. Los medios que contienen NPS se utilizaron el mismo día
o máximo un día después de su elaboración.
6.4 Análisis Estadístico
Para la etapa de embriogénesis somática se analizó estadísticamente el
efecto del NPS y el picloram sobre la desdiferenciación del tejido de plántulas
utilizando una estadística no paramétrica mediante la prueba para muestras
independientes Kruskal Wallis (α = 0.05), donde 1 representaba la presencia de
respuesta, y 0 representaba la ausencia de respuesta. Para la etapa de
micropropagación las variables de respuesta se analizaron mediante un análisis
de varianza a través de la comparación de muestras múltiples (α = 0.05). Para
ambos análisis se utilizó el software Statgraphics Centurion versión 16.1
(Statistical Graphics, Rockville, MD).
6.5 Enraizado y aclimatación de explantes clones del híbrido 1008157
Una vez finalizados los experimentos con el NPS, los explantes que
presentaron brotes se cultivaron en medio PA para continuar la propagación. Los
explantes que no presentaban brotes se colocaron en un medio MSC que se
preparó igual que la en la sección 6.2.4 pero con la diferencia que se incrementó
la cantidad de sacarosa a 45 g/L esto con el fin de estimular la producción de
raíces en el explante. Una vez que los explantes se cultivaron en el medio, se
incubaron por 40 d bajo las condiciones mencionadas en la sección 6.2.3.
Después de los 40 d, las plantas fueron retiradas del medio de cultivo para
colocarlas en el sustrato. El sustrato fue una mezcla de tres tipos diferentes de
suelo comúnmente utilizados en el cultivo del género Polianthes: turba, arena y
agrolita en un proporción 3:2:1 respectivamente. Esta mezcla de sustratos fue

53
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
humedecida y esterilizada en autoclave a 121 °C durante 20 min. Una vez que él
sustrato se encontraba a temperatura ambiente se colocó dentro de macetas de
plástico limpias (Figura 18a).
Los explantes una vez retirados del frasco (Figura 18b) fueron lavados con
agua corriente hasta eliminar completamente el agar (Figura 18c) y a continuación
se pasaron brevemente por una solución fungicida Captan 50 PH (ingrediente
activo: N-triclorometiltio-4-ciclohexeno-1,2-dicarboximida) en una dosis de 1 g/L
(Figura 18d). Inmediatamente se les colocó en la base enraizador en polvo Radix
1500 (ingrediente activo ácido indolbutírico 1500 ppm) (Figura 18e) y se plantaron
dentro de la maceta con sustrato (Figura 18f). En cada maceta se colocó uno o
dos explantes dependiendo el tamaño de los mismos. Después de plantar los
explantes se le agregó agua a cada maceta evitando que el sustrato quedara
demasiado húmedo. Finalmente fueron dispuestos en bolsas de plástico cubriendo
totalmente el (Figura 18g). Las macetas fueron incubadas durante 15 d con riegos
de una vez por semana. Transcurridos los 15 d, se realizaron dos perforaciones a
los extremos de la bolsa de plástico que cubría la maceta (Figura 18h) y el riego
se modificó una vez cada tercer día. Después de 15 d, se les retiró la cubierta de
plástico y el riego continúo siendo el mismo.

54
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 18. Proceso de aclimatación de los explantes clones del híbrido 1008157. (a) Sustrato estéril colocado en macetas limpias y previamente humedecido. (b) Planta retirada del frasco, aún presenta agar en la parte de la raíz. (c) Planta lavada para retirar el agar. (d) Plantas en solución de Captan. (e) Enraizador en polvo. (f) Planta cultivada en el sustrato. (g) Plantas con cubierta plástica para la aclimatación y colocadas en el incubador. (h) Planta a los 15 d de cultivada indicando las perforaciones que se le hicieron a la cubierta plástica para apoyar a la aclimatación.

55
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
VII. RESULTADOS Y DISCUSIÓN
7.1 Embriogénesis somática
7.1.1 Desdiferenciación de tejido somático de plántulas del híbrido
El tejido somático que se utilizó como explante inicial fue obtenido de la
germinación de semillas del híbrido 1008157. Las hojas de las plántulas fueron
segmentadas y distribuidas en 16 tratamientos (TR) variando las concentraciones
de NPS y picloram. En cada tratamiento, cuatro plántulas diferentes fueron
sometidas de manera independiente a desdiferenciación. Diferentes respuestas
fueron inducidas por efecto de la desdiferenciación resaltando la formación de
raíces, formación de callo (con o sin potencial embriogénico) y estructuras
globulares. El tipo de respuesta estuvo determinada en función de la
concentración de NPS, solo ó en combinación con picloram. Fehér y
colaboradores (2003) mencionan que el proceso de embriogénesis somática es
modulado por varios factores como el tipo de explante, el nivel endógeno y
exógeno de auxinas, las condiciones in vitro y el genotipo. El picloram fue
seleccionado como auxina inductora debido a que ha sido reportado por otros
autores (Gaspar y col., 1996; Ptak y Bach, 2007; Monja-Mio y Robert, 2013) en
relación a la inducción de embriogénesis en otras monocotiledóneas cercanas al
género Polianthes. Tal es el caso de Monja-Mio y Robert (2013) que realizaron
embriogénesis somática a partir de discos de tallo de Agave obteniendo 82.7
embriones por explante, las estructuras que se formaron en el proceso de
desdiferenciación del tejido son similares a las estructuras globulares identificadas
en TR13 y TR15, teniendo un diámetro y tiempo de expresión aproximado (Figura
19).
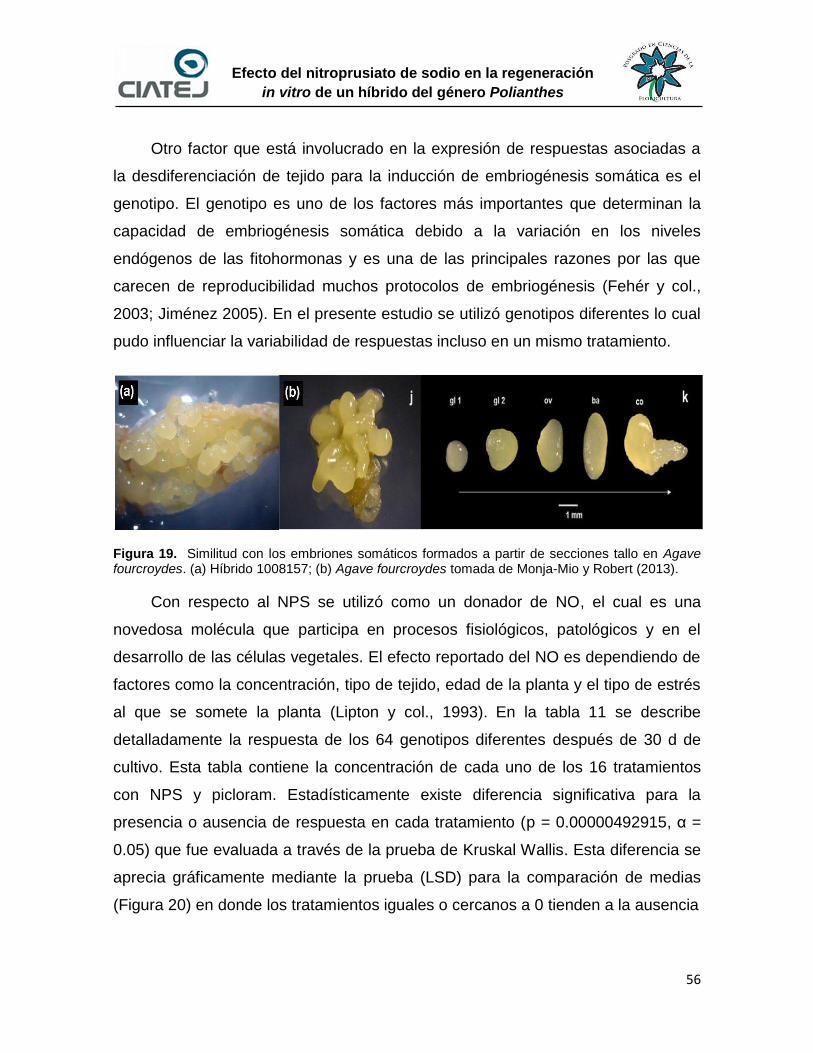
56
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Otro factor que está involucrado en la expresión de respuestas asociadas a
la desdiferenciación de tejido para la inducción de embriogénesis somática es el
genotipo. El genotipo es uno de los factores más importantes que determinan la
capacidad de embriogénesis somática debido a la variación en los niveles
endógenos de las fitohormonas y es una de las principales razones por las que
carecen de reproducibilidad muchos protocolos de embriogénesis (Fehér y col.,
2003; Jiménez 2005). En el presente estudio se utilizó genotipos diferentes lo cual
pudo influenciar la variabilidad de respuestas incluso en un mismo tratamiento.
Figura 19. Similitud con los embriones somáticos formados a partir de secciones tallo en Agave fourcroydes. (a) Híbrido 1008157; (b) Agave fourcroydes tomada de Monja-Mio y Robert (2013).
Con respecto al NPS se utilizó como un donador de NO, el cual es una
novedosa molécula que participa en procesos fisiológicos, patológicos y en el
desarrollo de las células vegetales. El efecto reportado del NO es dependiendo de
factores como la concentración, tipo de tejido, edad de la planta y el tipo de estrés
al que se somete la planta (Lipton y col., 1993). En la tabla 11 se describe
detalladamente la respuesta de los 64 genotipos diferentes después de 30 d de
cultivo. Esta tabla contiene la concentración de cada uno de los 16 tratamientos
con NPS y picloram. Estadísticamente existe diferencia significativa para la
presencia o ausencia de respuesta en cada tratamiento (p = 0.00000492915, α =
0.05) que fue evaluada a través de la prueba de Kruskal Wallis. Esta diferencia se
aprecia gráficamente mediante la prueba (LSD) para la comparación de medias
(Figura 20) en donde los tratamientos iguales o cercanos a 0 tienden a la ausencia
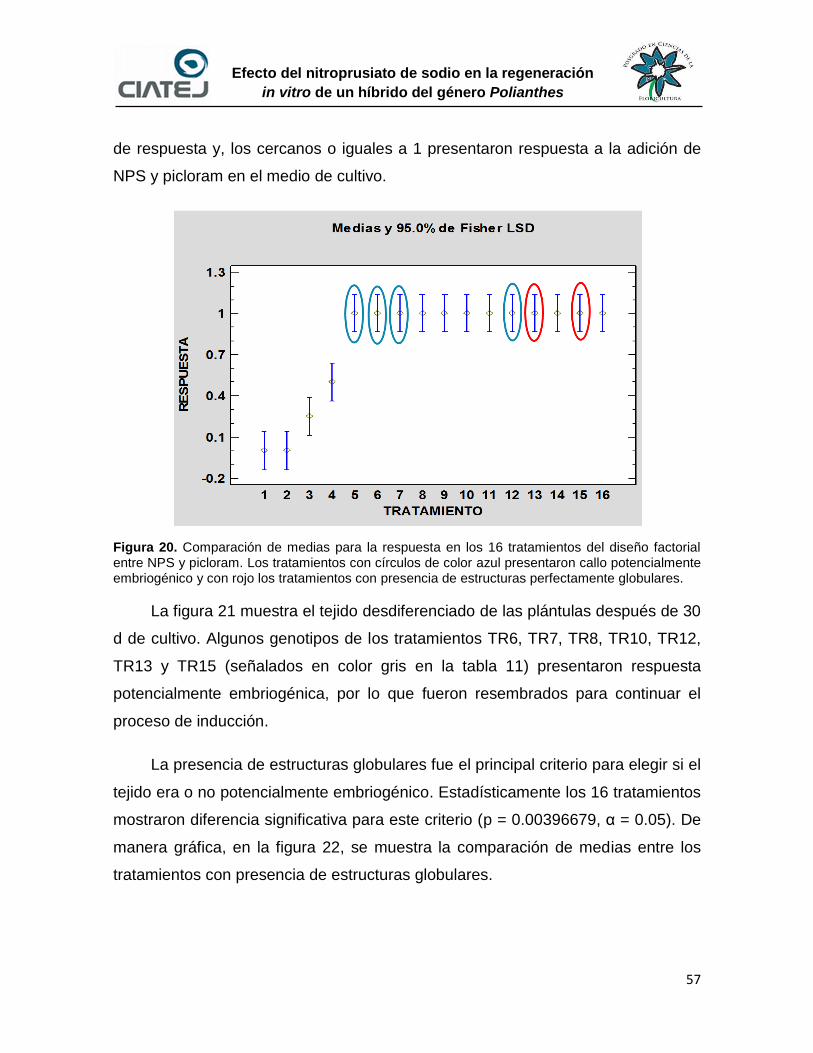
57
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
de respuesta y, los cercanos o iguales a 1 presentaron respuesta a la adición de
NPS y picloram en el medio de cultivo.
Figura 20. Comparación de medias para la respuesta en los 16 tratamientos del diseño factorial entre NPS y picloram. Los tratamientos con círculos de color azul presentaron callo potencialmente embriogénico y con rojo los tratamientos con presencia de estructuras perfectamente globulares.
La figura 21 muestra el tejido desdiferenciado de las plántulas después de 30
d de cultivo. Algunos genotipos de los tratamientos TR6, TR7, TR8, TR10, TR12,
TR13 y TR15 (señalados en color gris en la tabla 11) presentaron respuesta
potencialmente embriogénica, por lo que fueron resembrados para continuar el
proceso de inducción.
La presencia de estructuras globulares fue el principal criterio para elegir si el
tejido era o no potencialmente embriogénico. Estadísticamente los 16 tratamientos
mostraron diferencia significativa para este criterio (p = 0.00396679, α = 0.05). De
manera gráfica, en la figura 22, se muestra la comparación de medias entre los
tratamientos con presencia de estructuras globulares.
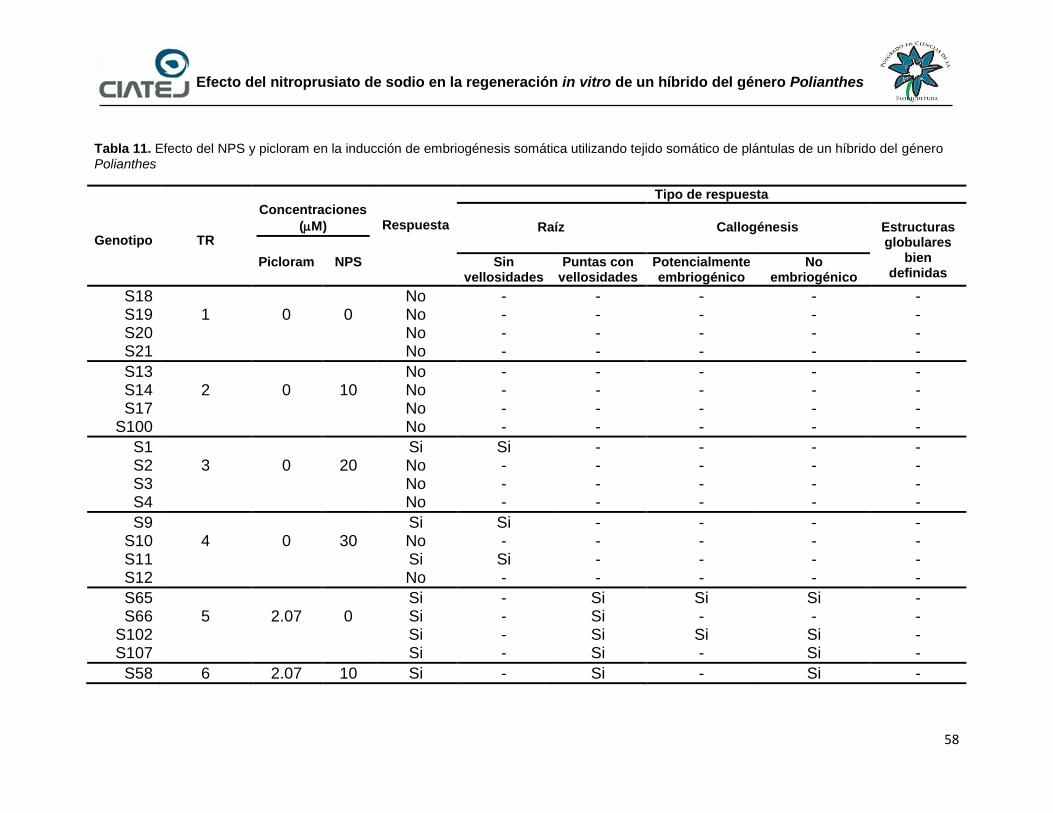
58
Efecto del nitroprusiato de sodio en la regeneración in vitro de un híbrido del género Polianthes
Tabla 11. Efecto del NPS y picloram en la inducción de embriogénesis somática utilizando tejido somático de plántulas de un híbrido del género Polianthes
Genotipo
TR
Concentraciones
(M)
Respuesta
Tipo de respuesta
Raíz
Callogénesis
Estructuras globulares
bien definidas
Picloram NPS Sin vellosidades
Puntas con vellosidades
Potencialmente embriogénico
No embriogénico
S18 1
0
0
No - - - - - S19 No - - - - - S20 No - - - - - S21 No - - - - -
S13 2
0
10
No - - - - - S14 No - - - - - S17 No - - - - -
S100 No - - - - -
S1 3
0
20
Si Si - - - - S2 No - - - - - S3 No - - - - - S4 No - - - - -
S9 4
0
30
Si Si - - - - S10 No - - - - - S11 Si Si - - - - S12 No - - - - -
S65 5
2.07
0
Si - Si Si Si - S66 Si - Si - - -
S102 Si - Si Si Si - S107 Si - Si - Si -
S58 6 2.07 10 Si - Si - Si -
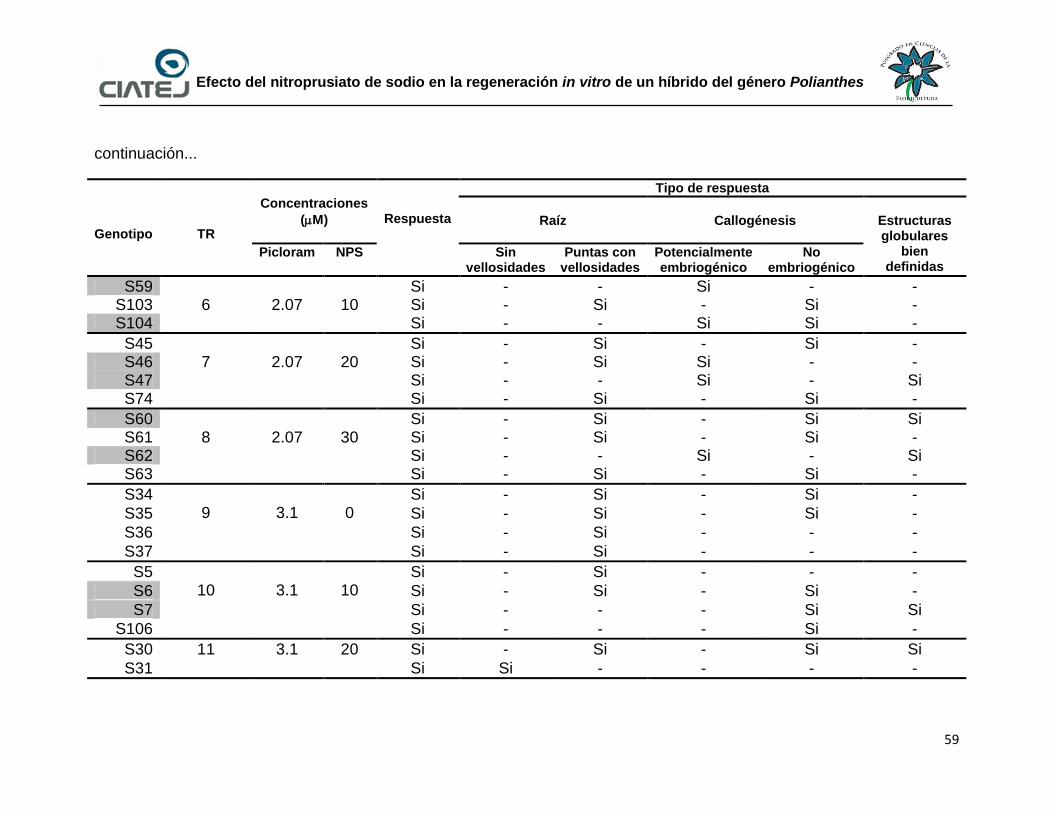
59
Efecto del nitroprusiato de sodio en la regeneración in vitro de un híbrido del género Polianthes
continuación...
Genotipo
TR
Concentraciones
(M)
Respuesta
Tipo de respuesta
Raíz
Callogénesis
Estructuras globulares
bien definidas
Picloram NPS Sin vellosidades
Puntas con vellosidades
Potencialmente embriogénico
No embriogénico
S59 6
2.07
10
Si - - Si - - S103 Si - Si - Si - S104 Si - - Si Si -
S45 7
2.07
20
Si - Si - Si - S46 Si - Si Si - - S47 Si - - Si - Si S74 Si - Si - Si -
S60 8
2.07
30
Si - Si - Si Si S61 Si - Si - Si - S62 Si - - Si - Si S63 Si - Si - Si -
S34 9
3.1
0
Si - Si - Si -
S35 Si - Si - Si -
S36 Si - Si - - -
S37 Si - Si - - -
S5 10
3.1
10
Si - Si - - -
S6 Si - Si - Si -
S7 Si - - - Si Si
S106 Si - - - Si -
S30 11 3.1 20 Si - Si - Si Si
S31 Si Si - - - -
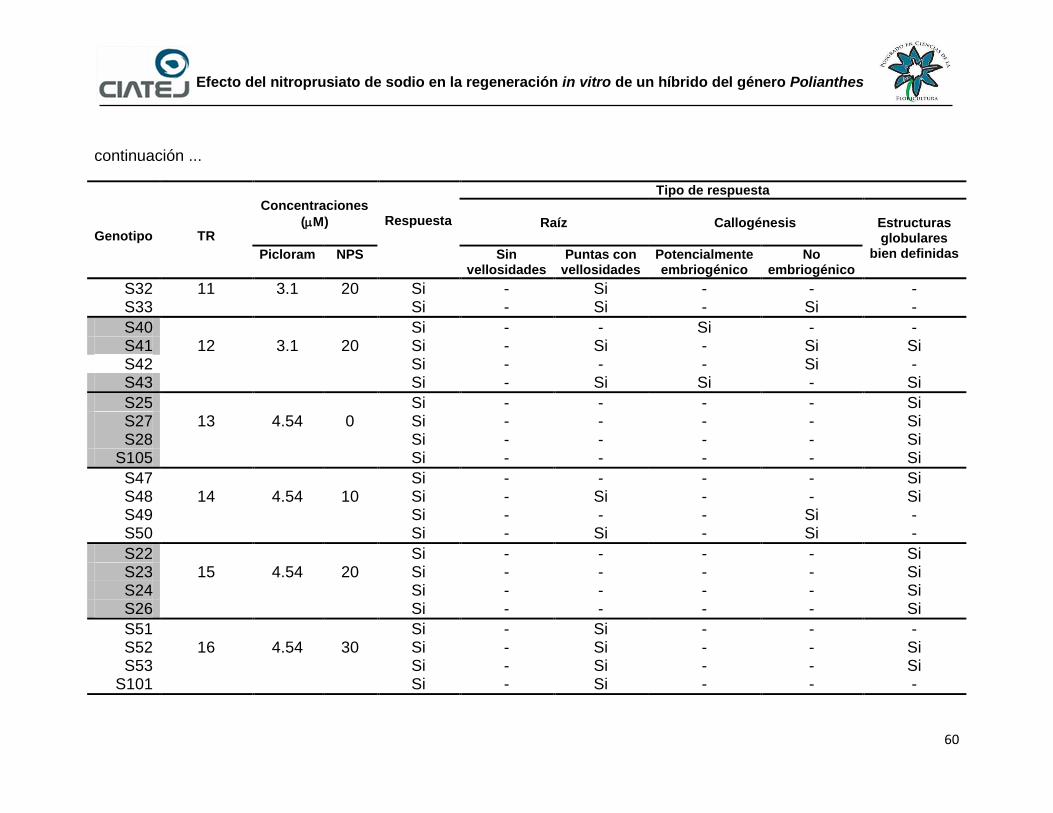
60
Efecto del nitroprusiato de sodio en la regeneración in vitro de un híbrido del género Polianthes
continuación ...
Genotipo
TR
Concentraciones
(M)
Respuesta
Tipo de respuesta
Raíz
Callogénesis
Estructuras globulares
bien definidas Picloram NPS Sin vellosidades
Puntas con vellosidades
Potencialmente embriogénico
No embriogénico
S32 11
3.1 20 Si - Si - - - S33 Si - Si - Si -
S40 12
3.1
20
Si - - Si - - S41 Si - Si - Si Si S42 Si - - - Si - S43 Si - Si Si - Si
S25 13
4.54
0
Si - - - - Si S27 Si - - - - Si S28 Si - - - - Si
S105 Si - - - - Si
S47 14
4.54
10
Si - - - - Si S48 Si - Si - - Si S49 Si - - - Si - S50 Si - Si - Si -
S22 15
4.54
20
Si - - - - Si S23 Si - - - - Si S24 Si - - - - Si S26 Si - - - - Si
S51 16
4.54
30
Si - Si - - - S52 Si - Si - - Si S53 Si - Si - - Si
S101 Si - Si - - -
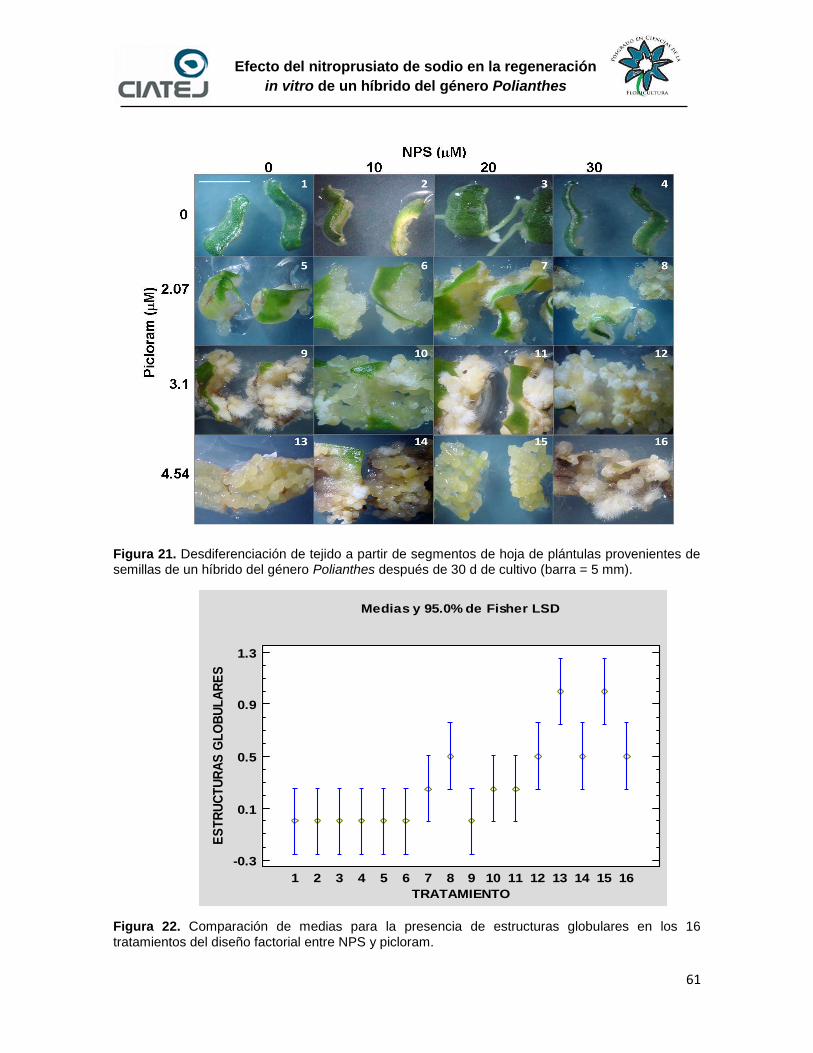
61
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 21. Desdiferenciación de tejido a partir de segmentos de hoja de plántulas provenientes de semillas de un híbrido del género Polianthes después de 30 d de cultivo (barra = 5 mm).
Figura 22. Comparación de medias para la presencia de estructuras globulares en los 16
tratamientos del diseño factorial entre NPS y picloram.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Medias y 95.0% de Fisher LSD
TRATAMIENTO
-0.3
0.1
0.5
0.9
1.3
ES
TR
UC
TU
RA
S G
LO
BU
LA
RE
S

62
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Las estructuras globulares fueron claramente identificadas en los cuatro
genotipos en los tratamientos 13 y 15 presentando algunas características
diferentes entre ellos. En el TR13, los genotipos con mayor formación de
estructuras globulares (tipo 1) fueron S27 y S28, siendo perfectamente globulares,
compactas, de una tenue coloración amarilla, fácil de separar y con un diámetro
menor a 1 mm (Figura 23e). En el TR15 fueron tres los genotipos con una mejor
respuesta para la formación de estructuras globulares (tipo 2). La diferencia con el
TR13 fue el tamaño y la coloración, siendo más traslúcidas y con un diámetro
menor (figura 23f).
En base al objetivo del presente estudio que es inducir embriogénesis
somática, todos aquellos tratamientos en los cuales el tipo de respuesta después
de 30 d de cultivo estuvo relacionada con la formación de raíces o estructuras tipo
raíces y callo no embriogénico fueron descartados (TR1, TR2, TR3, TR4, TR5,
TR9, TR11, TR14, TR16). En TR1 y TR2 el tejido somático no presentó respuesta,
y permaneció vivo hasta que los nutrientes en el medio de cultivo se agotaron
(Figura 23a). En TR3 y TR4, sólo algunos segmentos del tejido presentaron
respuesta para la formación de raíces gruesas sin vellosidades. Es decir, los
tratamientos con 0 M de picloram, donde el NPS no tuvo efecto de inducción y
requiere una concentración mayor al nivel endógeno de auxina para que el tejido
se desdiferencie. Ötvös y colaboradores (2005) utilizaron NPS en combinación con
2,4-D como promotor de la activación de la división celular y la formación de
células embriogénicas en Medicago sativa, postulando que en ausencia de una
auxina endógena y con una concentración de 10 M de NPS no hay influencia
para la división celular. Sin embargo en presencia de 2,4-D (0.22 y 1 M) y NPS
(10 M) se incrementa significativamente la frecuencia de células en división y
viables.

63
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Por otra parte, TR9 fue el primero en mostrar una respuesta de
desdiferenciación del tejido a los 10 d de cultivo (figura 23b), sin embargo, el
escaso callo que se formó en los bordes de algunos segmentos de hoja pronto se
convirtió en un callo no friable (figura 23h) y más tarde aumentó de tamaño
formando estructuras tipo raíces deformes (figura 23d). Además, en este mismo
tratamiento un genotipo presentó estructuras tipo raíces con abundantes
vellosidades. Para TR11, TR14 y TR16 la respuesta en dos genotipos fue la
formación de estructuras globulares bien definidas de color amarillo pálido. Al paso
de los días dichas estructuras globulares adquirieron coloraciones verdes. Para los
otros genotipos de estos mismos tratamientos, las estructuras globulares
aumentaron de tamaño y se deformaron en estructuras tipo raíces donde apareció
abundante vello (Figura 23g).
El efecto en la formación de raíces primarias y laterales fue reportado por
Khan y colaboradores (2013); Del Río y colaboradores (2014). Con relación a este
tipo de respuesta retomemos que en los tratamientos donde solo había NPS, se
presentó la formación de raíces, y cuando se adicionaba en conjunto con picloram
principalmente para los tratamientos con 2.07 y 3.1 M producía raíces con
vellosidades.

64
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 23. Tipos de respuesta en la inducción de la embriogénesis somática a partir de segmentos de hoja de plántulas de un híbrido del género Polianthes:(a) Raíz sin vellocidades, (b) Desdiferenciación del tejido somático después de 9 d, (c) Dos respuesta en un mismo genotipo, formación de estructuras globulares y tipo puntas, (d y g) Estructuras tipo raíz con abundantes vellocidades, (e) Estructuras globulares de color amarillo pálido, (f) Estructuras globulares translúcidas, (h) Callo no friable. Barra = 5.0 mm

65
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
7.1.2 Doble tinción diferencial de células
Posterior a los 30 d de cultivo y después de observar cuidadosamente cada
unos de los 7 tratamientos seleccionados como potencialmente embriogénicos se
tomó una muestra para realizar la prueba de la doble tinción diferencial de células
como se describe en la sección 6.1.3. Con respecto a la muestra, se tomaron
varias estructuras globulares con el diámetro menor en cada uno de los genotipos
de cada tratamiento. La manipulación del tejido fue meticulosa para evitar la
contaminación y debido a esto sólo se realizó la tinción una vez. Se tomó
suficiente muestra para poder realizar más de una observación en el microscopio.
Sin embargo, la forma globular y compacta de las estructuras impidió una visión
clara de las mismas en el microscopio. En varias ocasiones la muestra se presionó
con una fuerza excesiva y aún así no se logró observar claramente la polaridad en
las células. Por lo tanto estos resultados, no comprobaron la presencia de células
polarizadas, característica fundamental en la inducción de embriogénesis
somática. Sin embargo, estos resultados fueron comparados con análisis
histológicos de estructuras globulares embriogénicas (Monja-Mio y Robert, 2013;
Portillo y col., 2007; Sáenz y col., 2006). En estos estudios, las estructuras
embriogénicas que se analizaron histológicamente son similares a las encontradas
durante la doble tinción diferencial.
En la mayoría de las estructuras las células del contorno se tiñeron con azul
de Evans y por dentro con acetocarmin, aunque existió la presencia de células
azules, estas eran menores. De acuerdo al estudio de embriogénesis somática de
coco realizado por Sáenz y colaboradores (2006), el conjunto de células teñidas
de azul formaron algo similar a la cubierta de células que denominan protodermo,
y debajo están las posibles células meristemáticas que se activan para dar inicio al
desarrollo de estructuras embriogénicas.

66
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
En la figura 24 se puede observar una imagen representativa de la doble
tinción diferencial celular. Las estructuras Tipo 1 (Figura 24a) que se formaron
principalmente en el TR13 presenta una mayor organización en las células del
contorno de las estructuras teñidas (Figura 24 b y c). Para el caso de las
estructuras Tipo 2 (Figura 24d) que se formaron en el TR15 presentan también
células del contorno teñidas mayormente con azul de Evans y tienen una
apariencia menos compacta que las del tratamiento 13 (Figura 24e).
Como se menciono previamente otros tratamientos presentaron estructuras
globulares pero con otras características (Figura 24g) como delgadas
vellosidades, apariencia translucida, apariencia porosa y otras compactas pero de
color amarillo. Este tipo de estructuras también se tiñeron y lo que se observo en
el microscopio también presentaba células del contorno teñidas (Figura 24f) pero
desorganizadas. En el TR8 se encontraron otro tipo de estructuras semejante a lo
que los autores denominan embrión ovalado con el mismo patrón de tinción en las
células del contorno y del interior (Figura 24h-i).
Figura 24. Estructuras globulares sometidas a la doble tinción diferencial.

67
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
7.1.3 Subcultivos de callo y/o estructuras potencialmente embriogénicas
7.1.3.1 Subcultivo utilizando medios de inducción.
Con los resultados de la doble tinción diferencial no fue posible concluir la
inducción de embriogénesis somática en algunos de los 7 tratamientos
seleccionados. Transcurridos 60 d de cultivo bajo observaciones periódicas los
tratamientos no mostraban cambios aparentes, las estructuras globulares no
parecían continuar el desarrollo normalmente embrionario por lo que fue necesario
realizar una resiembra. Se utilizaron dos medios de cultivo que han sido utilizados
para inducción a embriogénesis somática. Se utilizó medio MIA reportado por
varios autores para inducir embriogénesis somática en Agave el cuál contiene 2
mg/L de 2,4-D y 0.3 mg/L de BA (Portillo y col., 2007). El otro medio de inducción
que se utilizó fue el medio MIP reportado recientemente (Abdullah, 2012) que
contiene 2.5 mg/L de 2,4-D y 0.1 mg/L de BA. La posibilidad de una respuesta
positiva para continuar el proceso de inducción a la embriogénesis somática
estuvo en base de que estos medios han sido utilizados exitosamente en el
género Agave (cercano al género Polianthes), y el mismo género para Polianthes
tuberosa. Además la activación de las celulas en respuesta a auxinas puede ser
un evento clave en la adaptación celular genética, metabólica y de reprogramación
fisiológica, dando lugar a la competencia embriogénica de las células vegetales
somáticas. los RC en ambos medios son los mismos y las concentraciones muy
cercanas. De esta manera el efecto de ambos RC pudo verse aun cuando se
trabajo con genotipos diferentes.
Portillo y col., 2007 mencionan que el efecto de aumenar la concentración de
citoquininas es benefico para la formación de embriones somaticos en Agave; el
medio MIA tenia mayor concentración de BA en comparación con el MIP, sin
embargo, esta diferencia de BA no es tan elevada como la reportada por esos
mismos autores (44.4 y 66.6 M BA) por lo tanto se pensó en una respuesta muy
parecida en ambos medios. Abdullah (2012) estudio la embiogénesis somática en

68
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Polianthes tuberosa y utilizó el medio MIP para promover efectivamente la
embriogénesis somática. Obtuvo apartir de callo, embriones somáticos en sus
diferentes estadios (globular, torpedo y cotiledonar) despúes de tres meses de
cultivo. Utilizando medio MIP líquido o sólido obtuvo un promedio de embriones de
26.67±0.42 and 20.53±0.50 respectivamente a partir de 0.5 cm de callo.
El proceso de resiembra fue seleccionar muestra en cada genotipo donde
hubiera posibilidad de inducción embriogénica y cultivarla tanto en medio MIA
como en medio MIP. Los resultados después de 30 d de cultivo se observan para
cada tratamiento (figura 25 - 31). No se puede generalizar una respuesta, debido
principalmente al origen del tejido genotipicamente diferente. Sin embargo si se
distingue un efecto más positivo en el medio MIA con respecto a la inducción pero
sin llegar a obtener embriones en ningun tratamiento.
El genotipo S59 del tratamiento 6 formó en ambos medios de cultivo cúmulos
celulares que sobresalían al tejido los cuales no estaban presentes antes de la
resiembra. Sin embargo la coloración era muy amarilla y no parecían ser
estructuras embrionarias sino más bien el inicio de masas pro embriogénicas. En
el genotipo S104 la respuesta en los dos medios fue diferente. En el medio MIA
fue similar a la antes descrita añadiendo que parte del tejido se necroso y murió.
Para el medio MIP se formaron las mismas estructuras ya identificadas como
puntas con abundantes vellosidades y también se presento tejido necrótico y seco.
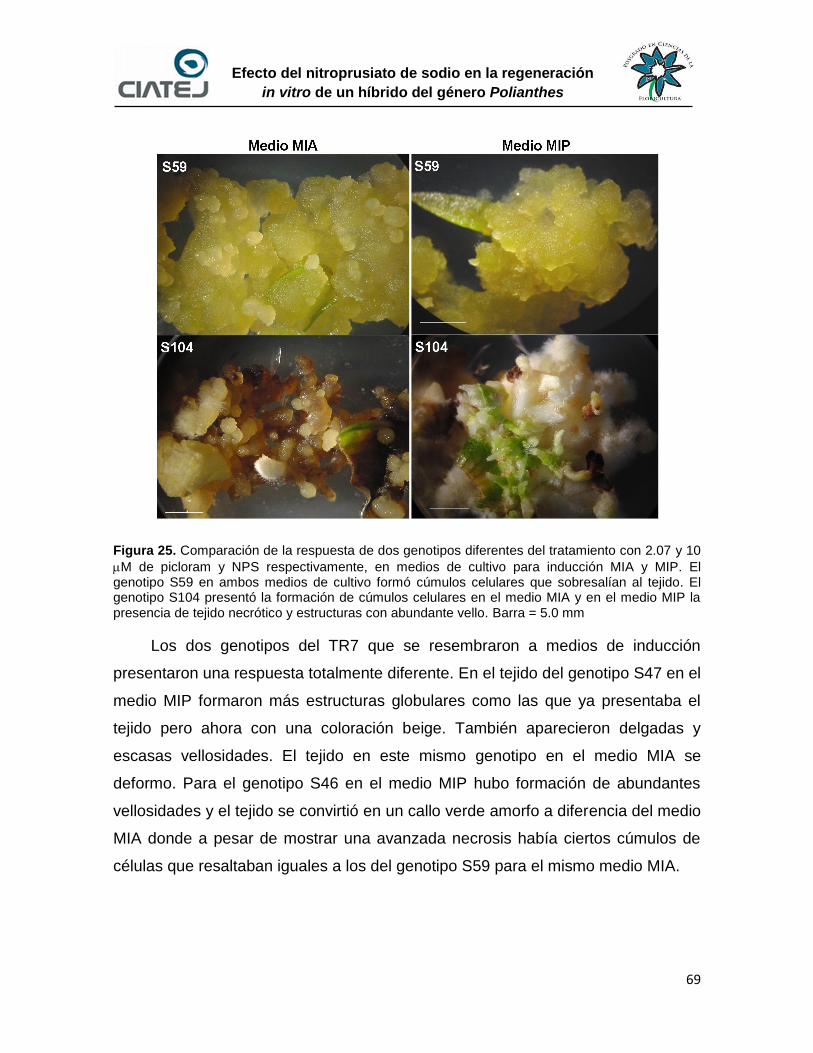
69
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 25. Comparación de la respuesta de dos genotipos diferentes del tratamiento con 2.07 y 10
M de picloram y NPS respectivamente, en medios de cultivo para inducción MIA y MIP. El genotipo S59 en ambos medios de cultivo formó cúmulos celulares que sobresalían al tejido. El genotipo S104 presentó la formación de cúmulos celulares en el medio MIA y en el medio MIP la presencia de tejido necrótico y estructuras con abundante vello. Barra = 5.0 mm
Los dos genotipos del TR7 que se resembraron a medios de inducción
presentaron una respuesta totalmente diferente. En el tejido del genotipo S47 en el
medio MIP formaron más estructuras globulares como las que ya presentaba el
tejido pero ahora con una coloración beige. También aparecieron delgadas y
escasas vellosidades. El tejido en este mismo genotipo en el medio MIA se
deformo. Para el genotipo S46 en el medio MIP hubo formación de abundantes
vellosidades y el tejido se convirtió en un callo verde amorfo a diferencia del medio
MIA donde a pesar de mostrar una avanzada necrosis había ciertos cúmulos de
células que resaltaban iguales a los del genotipo S59 para el mismo medio MIA.

70
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 26. Comparación de la respuesta de dos genotipos diferentes del tratamiento con 2.07 y 20
M de picloram y NPS respectivamente, en medios de cultivo para inducción MIA y MIP. El genotipo S47 en medio MIA presentó deformación y en medio MIP formación de estructuras globulares de coloración beige. El genotipo S46 en medio MIA presentó tejido necrótico y áreas donde hubo formación de estructuras globulares y en medio MIP deformación de callo con abundantes vellosidades. Barra = 5.0 mm
El efecto de la resiembra de tres genotipos para el TR8 fue muy heterogéneo
entre los mismos. El genotipo que tuvo una respuesta más relacionada con la
inducción fue el S60 en ambos medios ya que aparecieron algunas estructuras
globulares y en forma de arroz de apariencia translúcidas rodeadas de tejido
necrótico. Los otros dos genotipos no mostraron este mismo efecto para el medio
MIA. En el S61, las estructuras globulares existentes adquirieron una coloración
verde y abundantes vellosidades. En el S62 el tejido se necrosó o se formó un
callo blanco de aspecto poroso.
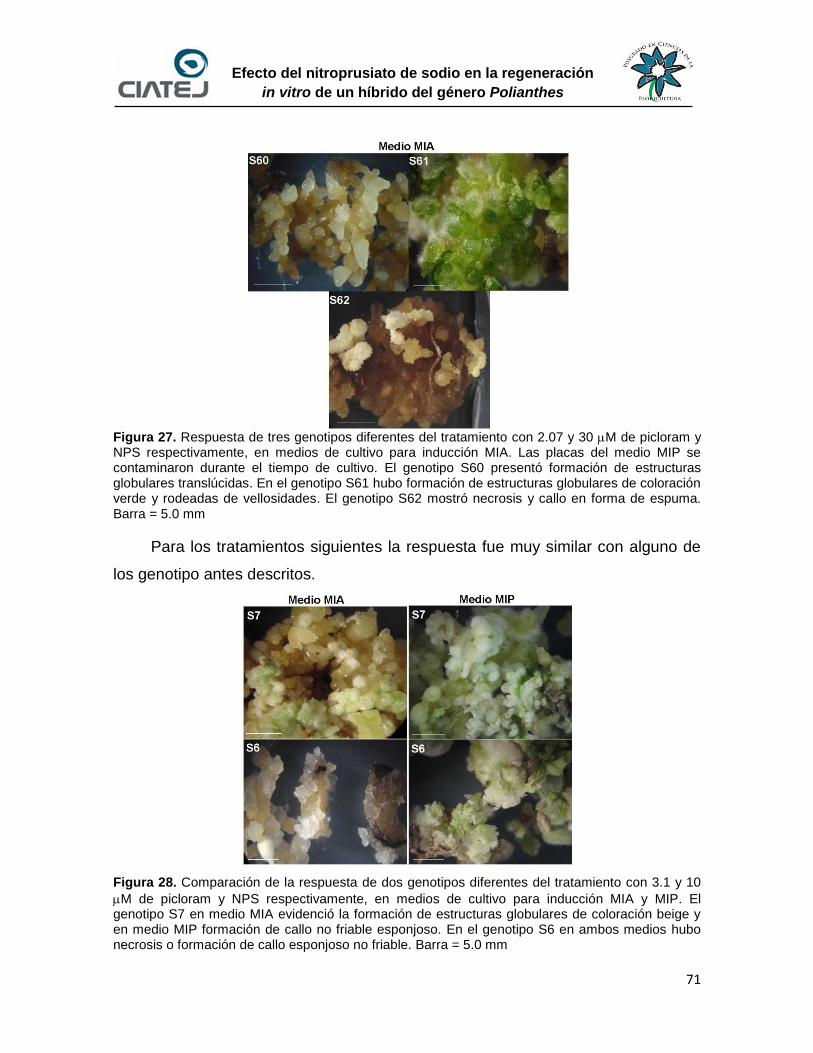
71
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 27. Respuesta de tres genotipos diferentes del tratamiento con 2.07 y 30 M de picloram y NPS respectivamente, en medios de cultivo para inducción MIA. Las placas del medio MIP se contaminaron durante el tiempo de cultivo. El genotipo S60 presentó formación de estructuras globulares translúcidas. En el genotipo S61 hubo formación de estructuras globulares de coloración verde y rodeadas de vellosidades. El genotipo S62 mostró necrosis y callo en forma de espuma. Barra = 5.0 mm
Para los tratamientos siguientes la respuesta fue muy similar con alguno de
los genotipo antes descritos.
Figura 28. Comparación de la respuesta de dos genotipos diferentes del tratamiento con 3.1 y 10
M de picloram y NPS respectivamente, en medios de cultivo para inducción MIA y MIP. El genotipo S7 en medio MIA evidenció la formación de estructuras globulares de coloración beige y en medio MIP formación de callo no friable esponjoso. En el genotipo S6 en ambos medios hubo necrosis o formación de callo esponjoso no friable. Barra = 5.0 mm
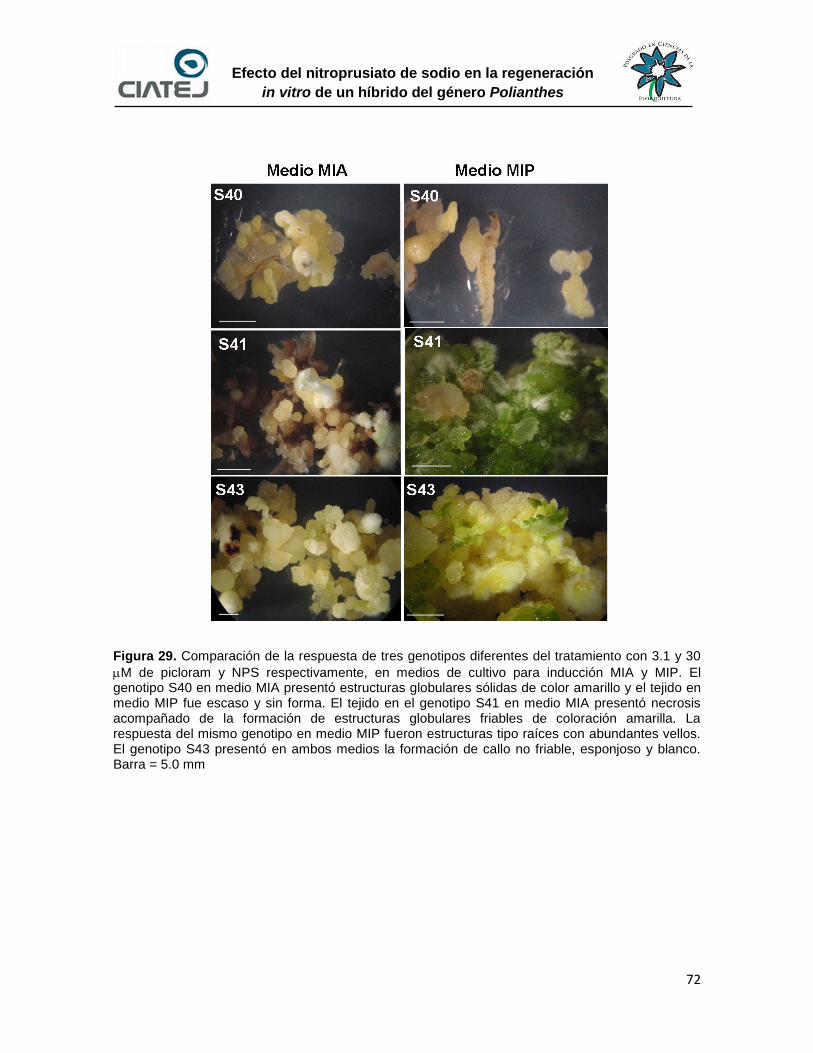
72
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 29. Comparación de la respuesta de tres genotipos diferentes del tratamiento con 3.1 y 30
M de picloram y NPS respectivamente, en medios de cultivo para inducción MIA y MIP. El genotipo S40 en medio MIA presentó estructuras globulares sólidas de color amarillo y el tejido en medio MIP fue escaso y sin forma. El tejido en el genotipo S41 en medio MIA presentó necrosis acompañado de la formación de estructuras globulares friables de coloración amarilla. La respuesta del mismo genotipo en medio MIP fueron estructuras tipo raíces con abundantes vellos. El genotipo S43 presentó en ambos medios la formación de callo no friable, esponjoso y blanco. Barra = 5.0 mm
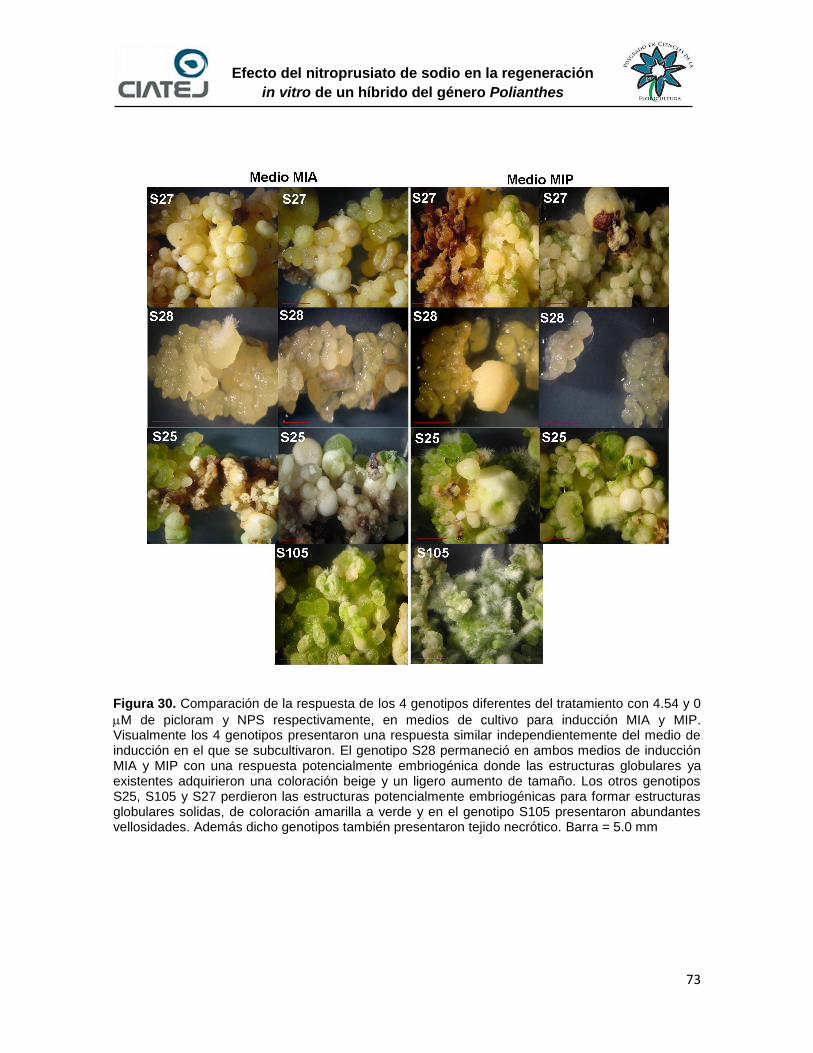
73
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 30. Comparación de la respuesta de los 4 genotipos diferentes del tratamiento con 4.54 y 0
M de picloram y NPS respectivamente, en medios de cultivo para inducción MIA y MIP. Visualmente los 4 genotipos presentaron una respuesta similar independientemente del medio de inducción en el que se subcultivaron. El genotipo S28 permaneció en ambos medios de inducción MIA y MIP con una respuesta potencialmente embriogénica donde las estructuras globulares ya existentes adquirieron una coloración beige y un ligero aumento de tamaño. Los otros genotipos S25, S105 y S27 perdieron las estructuras potencialmente embriogénicas para formar estructuras globulares solidas, de coloración amarilla a verde y en el genotipo S105 presentaron abundantes vellosidades. Además dicho genotipos también presentaron tejido necrótico. Barra = 5.0 mm
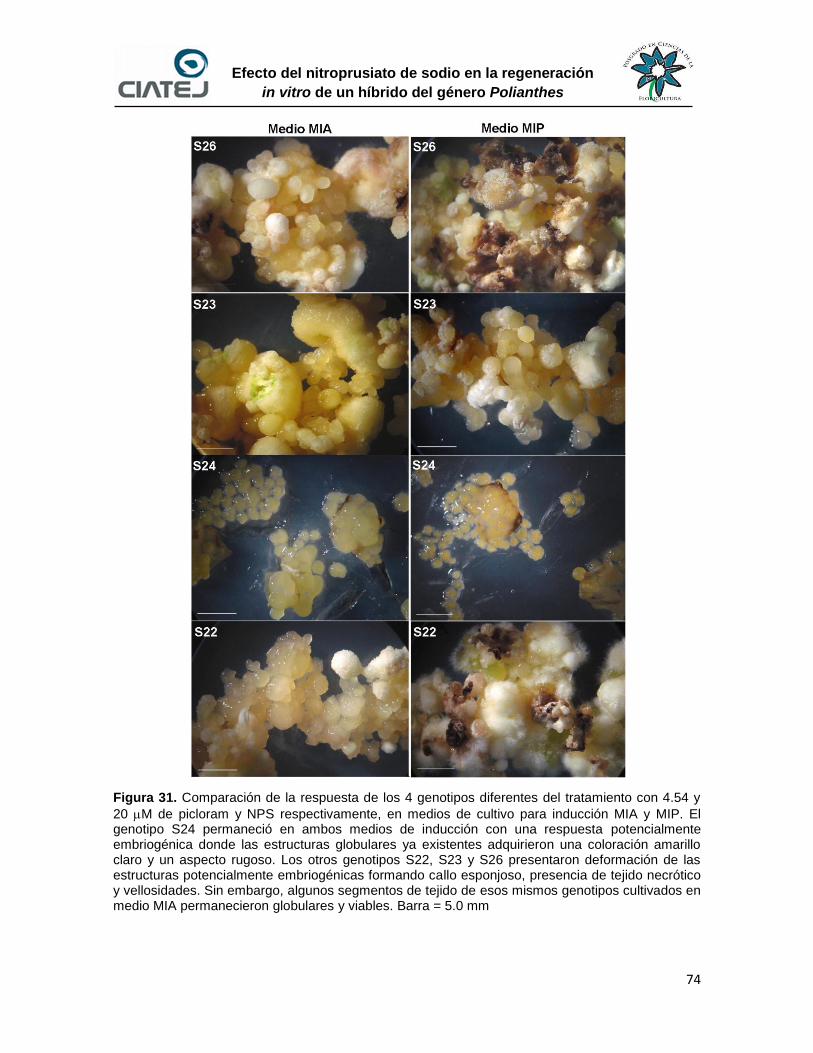
74
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 31. Comparación de la respuesta de los 4 genotipos diferentes del tratamiento con 4.54 y
20 M de picloram y NPS respectivamente, en medios de cultivo para inducción MIA y MIP. El genotipo S24 permaneció en ambos medios de inducción con una respuesta potencialmente embriogénica donde las estructuras globulares ya existentes adquirieron una coloración amarillo claro y un aspecto rugoso. Los otros genotipos S22, S23 y S26 presentaron deformación de las estructuras potencialmente embriogénicas formando callo esponjoso, presencia de tejido necrótico y vellosidades. Sin embargo, algunos segmentos de tejido de esos mismos genotipos cultivados en medio MIA permanecieron globulares y viables. Barra = 5.0 mm

75
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
7.1.3.2 Subcultivo utilizando medio de inducción enriquecido con
aminoácidos.
Al finalizar la resiembra utilizando los medios de inducción MIA y MIP solo se
logró mantener con viabilidad el tejido. Después de 90 d aún existían en algunos
tratamientos la presencia de estructuras globulares potencialmente viables. Mere y
Vázquez (2003) utilizaron una solución enriquecida con factores de crecimiento,
específicamente, vitaminas y aminoácidos, además de compuestos orgánicos
como la urea para producir callos embriogénicos en Daucus carota L. Flores-
Benítez y colaboradores (2007) indujeron embriones somáticos en Agave a partir
de callo mediante la adición de una mezcla de aminoácidos y vitaminas que
denominaron coctel 20, estos fueron inducidos después de seis a ocho semanas
resultando friables. Por lo tanto, en el presente trabajo se realizó un último
subcultivo adicionando al medio de cultivo MS la misma solución de aminoácidos y
vitaminas reportada por Benítez y colaboradores (2007) cuya compocición es rica
en nitrógeno orgánico.
Nitrógeno reducido fue identificado como otro factor importante en la
inducción y desarrollo del embrión. Trigiano y colaboradores (1992), confirmaron
que una fuente adicional de nitrógeno reducido indujo la desdiferenciación y
proliferación de células en la epidermis para iniciar embriones secundarios en
Dactylis glomerata. Además, Stuart y Strickland (1984), incrementaron el número
de embriones producidos por cultivos de alfalfa al adicionar nitrógeno reducido al
medio de cultivo. Sin embargo, en el presente estudio el efecto de la adición del
coctel 20 al medio de cultivo MIc20 (en conjunto con los RC) sobre el tejido de
todos los genotipos resembrados no influyó en la desdiferenciación y proliferación
de células en epidermis de las estructuras globulares resembradas. Al contrario,
exceptuando el genotipo S24 del TR15 en donde las estructuras globulares
persistieron sin mostrar ningún cambio aparente en la resiembra, en todos los
demás genotipos la adición del coctel 20 y RC causó una desorganización del
tejido, producción de callo no embriogénico, aparición de vellosidades y la
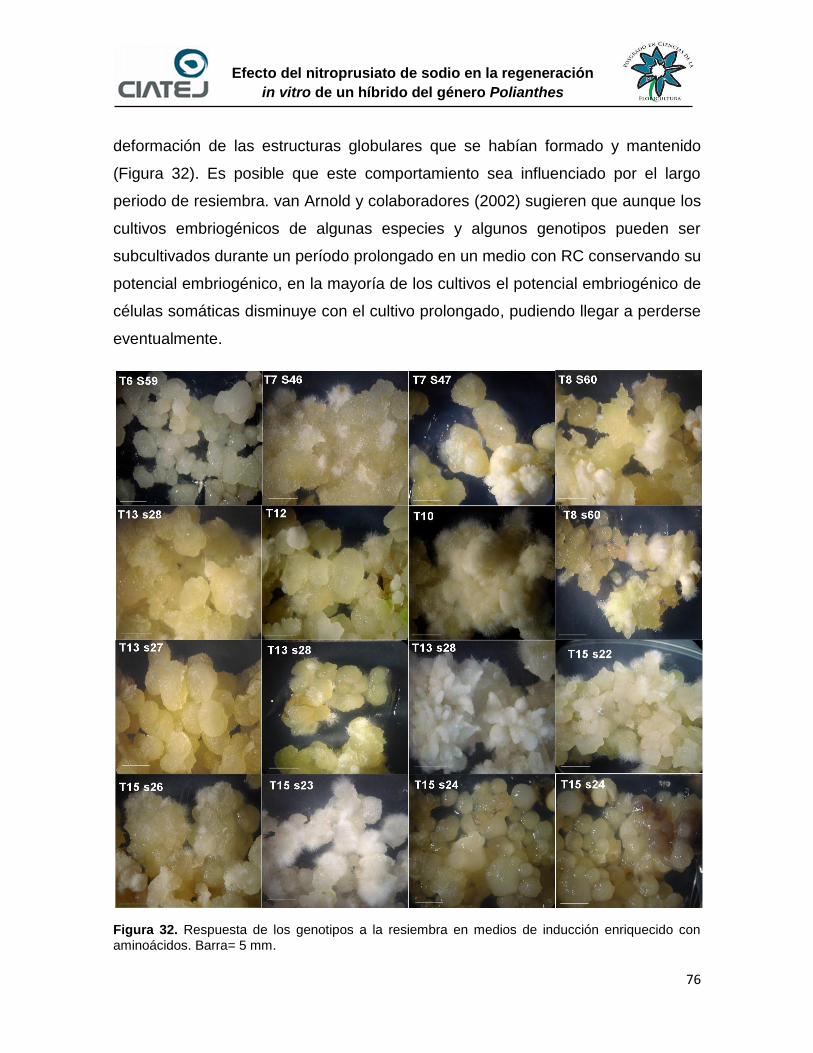
76
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
deformación de las estructuras globulares que se habían formado y mantenido
(Figura 32). Es posible que este comportamiento sea influenciado por el largo
periodo de resiembra. van Arnold y colaboradores (2002) sugieren que aunque los
cultivos embriogénicos de algunas especies y algunos genotipos pueden ser
subcultivados durante un período prolongado en un medio con RC conservando su
potencial embriogénico, en la mayoría de los cultivos el potencial embriogénico de
células somáticas disminuye con el cultivo prolongado, pudiendo llegar a perderse
eventualmente.
Figura 32. Respuesta de los genotipos a la resiembra en medios de inducción enriquecido con
aminoácidos. Barra= 5 mm.

77
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
7.2 Micropropagación
7.2.1 Descripción inicial de plántulas
Para evaluar el efecto del NPS sobre la micropropagación de plántulas del
híbrido 1008157 se utilizaron 75 plantas provenientes de la germinación de
semillas, debido a esto, las plantas fueron genotípicamente diferentes entre sí. La
Tabla 5 (mostrada en la sección 6.2.1) muestra la descripción inicial de las 75
plántulas obteniendo en promedio una longitud de la planta de 2.94 cm ± 1.89, un
número de hojas igual a 5.61 ± 3.44 y 0.68 ± 0.90 brotes. Además, respecto al
número de brotes, el 45.3 % del total de las plantas presentaron brotes, de los
cuales, el 64.7 % presentó un solo brote, el 20.6 % dos brotes y el 14.7 % tres
brotes.
7.2.2 Estimulación de la producción de brotes en plántulas
El efecto del NPS en la estimulación de la producción de brotes fue evaluado
en un primer cultivo, donde las 75 plantas fueron distribuidas aleatoriamente en
cinco tratamientos variando la concentración de NPS (0, 10, 20, 30 y 40 M).
Además, para evaluar la posible persistencia o disminución del efecto inducido por
el NPS en cultivos subsecuentes, las plantas obtenidas del primer cultivo se
resembraron por segunda vez utilizando la misma concentración de NPS para
cada tratamiento. Al finalizar cada cultivo, se realizó una descripción del desarrollo
de la planta donde las variables de respuesta fueron la altura, el número de hojas
y brotes.
La adición de NPS en el cultivo in vitro de células vegetales es actualmente
un tema de interés. Han y colaboradores (2009) indicaron que el NO puede
participar en la división celular y por lo tanto afectar la regeneración y
multiplicación de brotes adventicios. Tan y colaboradores (2013), evaluaron el
efecto de la NPS en la multiplicación de brotes y regeneración de Vanilla planifolia

78
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Andrews. Sus resultados indicaron que el NPS a un concentración 10 µM
estimulaba el desarrollo de organogénesis directa produciendo brotes a partir de
segmentos nodales con un éxito del 93% de los explantes producían brotes.
Pagnussat y colaboradores (2002) adicionaron la misma concentración de
NPS en Cucumis sativus y sugieren su efecto como un intermediario de
regeneración de brotes adventicios y raíces. Esto también se ha sugerido por
Correa-Aragunde y colaboradores (2006) en otras especies como Solanum
lycopersicum donde la concentración óptima fue menor (200 µM). Los resultados
en este estudio no pudieron demostrar el efecto potenciador del NPS en la
producción de brotes debido principalmente al número reducido de repeticiones
por tratamiento. Sin embargo si se observó una influencia numéricamente positiva
al producir brotes.
En el tratamiento 0 M NPS (Tabla 12), al finalizar el primer cultivo presentó
un promedio para la altura de las plantas igual a 1.52 cm ± 1.13, este mismo
tratamiento en el segundo cultivo tuvo una disminución en la altura de las plantas,
las cuales presentaron un promedio igual a 0.59 cm ± 1.62. Para las variables de
respuesta, estadísticamente no hay diferencia significativa entre el primer y
segundo cultivo p = 0.0846, 0.0595, 0.8150, α = 0.05 para altura, número de hojas
y número de brotes respectivamente. Respecto al número de hojas, al finalizar el
primer cultivo el promedio de las plantas fue igual a 5.53 hojas ± 3.34. Esta misma
variable de respuesta al finalizar el segundo cultivo mostró un incremento
obteniendo un promedio de 6.46 hojas ± 5.72. Respecto al número de brotes, el
promedio del primer y segundo cultivo fue igual a 0.93 ± 0.88 y 0.85 ± 1.03 brotes
respectivamente. Debido a los fines de la técnica de micropropagación, el número
de brotes es la variable de respuesta de mayor interés en este estudio (Figura 39)
entonces, es importante resaltar que al finalizar el segundo cultivo 11 plantas
diferentes (73.3%) mostraron una estimulación en la producción de nuevos brotes.
Sin embargo, el número de plantas estimuladas fue mayor durante el primer

79
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
cultivo (nueve), a diferencia del segundo cultivo en el que únicamente dos plantas
fueron estimuladas para la producción de nuevos brotes (Figura 33).
Tabla 12. Comparación de las respuestas de desarrollo de las plantas durante el cultivo en medio sin NPS.
(0 M)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 PROM
ALTURA
(cm)
1ER C 0.0 2.0 1.2 2.0 3.0 0.9 2.8 3.0 2.3 0.0 0.0 0.1 1.5 2.7 1.3 1.52
2DO C -2.2 -0.7 -0.5 1.5 -1.6 1.6 2.0 - - 0.2 3.4 -0.4 2.2 1.0 1.1 0.59
HOJAS
1ER C 7 4 3 3 2 6 8 13 10 3 5 2 9 6 2 5.53
2DO C 17 12 -2 15 5 10 2 - - 2 6 2 9 4 2 6.46
BROTES
1ER C 2 0 0 1 0 0 1 2 2 1 1 0 2 2 0 0.93
2DO C 3 1 0 2 0 2 0 - - 0 1 0 2 0 0 0.85
*Los espacios vacíos en la tabla corresponden a plantas contaminadas durante la manipulación en el subcultivo. Por lo tanto, estas plantas no fueron tomadas en cuenta para la determinación de los promedios, C: cultivo.
Al finalizar el primer cultivo, ninguno de los tratamientos presentó diferencia
significativa en la altura (0, 10, 20, 30 y 40 M) (p = 0.6971, α = 0.05). Además, la
altura promedio mayor fue la del control (1.52 cm ± 1.13), mientras que el
tratamiento con menor altura promedio fue 20 M (0.73 cm ± 1.77) (Tabla 14)
seguido de 30 M (0.86 cm ± 1.90) (Tabla 15), 40 M (1.37 cm ± 1.40) (Tabla 16)
y 10 M (1.44 cm ± 2.66) (Tabla 13).
Figura 33. Plantas control con diferencias fenotípicas en su morfología después de 60 d en medio de cultivo sin NPS.
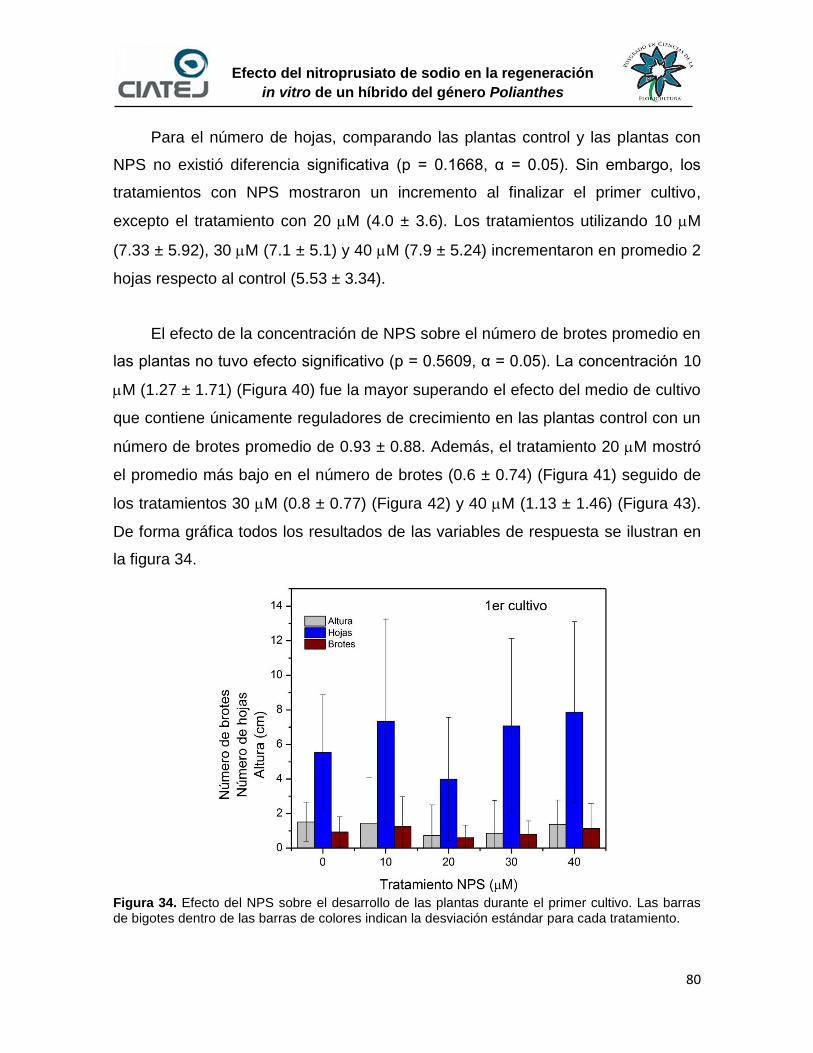
80
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Para el número de hojas, comparando las plantas control y las plantas con
NPS no existió diferencia significativa (p = 0.1668, α = 0.05). Sin embargo, los
tratamientos con NPS mostraron un incremento al finalizar el primer cultivo,
excepto el tratamiento con 20 M (4.0 ± 3.6). Los tratamientos utilizando 10 M
(7.33 ± 5.92), 30 M (7.1 ± 5.1) y 40 M (7.9 ± 5.24) incrementaron en promedio 2
hojas respecto al control (5.53 ± 3.34).
El efecto de la concentración de NPS sobre el número de brotes promedio en
las plantas no tuvo efecto significativo (p = 0.5609, α = 0.05). La concentración 10
M (1.27 ± 1.71) (Figura 40) fue la mayor superando el efecto del medio de cultivo
que contiene únicamente reguladores de crecimiento en las plantas control con un
número de brotes promedio de 0.93 ± 0.88. Además, el tratamiento 20 M mostró
el promedio más bajo en el número de brotes (0.6 ± 0.74) (Figura 41) seguido de
los tratamientos 30 M (0.8 ± 0.77) (Figura 42) y 40 M (1.13 ± 1.46) (Figura 43).
De forma gráfica todos los resultados de las variables de respuesta se ilustran en
la figura 34.
Figura 34. Efecto del NPS sobre el desarrollo de las plantas durante el primer cultivo. Las barras de bigotes dentro de las barras de colores indican la desviación estándar para cada tratamiento.
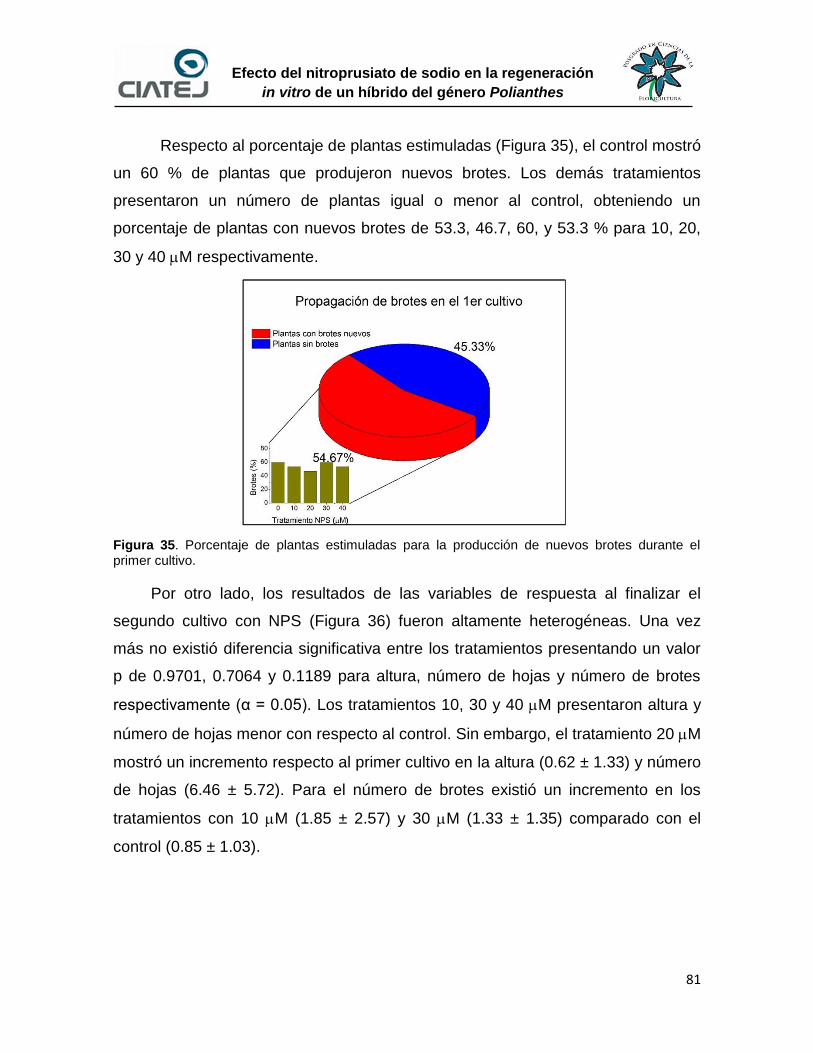
81
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Respecto al porcentaje de plantas estimuladas (Figura 35), el control mostró
un 60 % de plantas que produjeron nuevos brotes. Los demás tratamientos
presentaron un número de plantas igual o menor al control, obteniendo un
porcentaje de plantas con nuevos brotes de 53.3, 46.7, 60, y 53.3 % para 10, 20,
30 y 40 M respectivamente.
Figura 35. Porcentaje de plantas estimuladas para la producción de nuevos brotes durante el primer cultivo.
Por otro lado, los resultados de las variables de respuesta al finalizar el
segundo cultivo con NPS (Figura 36) fueron altamente heterogéneas. Una vez
más no existió diferencia significativa entre los tratamientos presentando un valor
p de 0.9701, 0.7064 y 0.1189 para altura, número de hojas y número de brotes
respectivamente (α = 0.05). Los tratamientos 10, 30 y 40 M presentaron altura y
número de hojas menor con respecto al control. Sin embargo, el tratamiento 20 M
mostró un incremento respecto al primer cultivo en la altura (0.62 ± 1.33) y número
de hojas (6.46 ± 5.72). Para el número de brotes existió un incremento en los
tratamientos con 10 M (1.85 ± 2.57) y 30 M (1.33 ± 1.35) comparado con el
control (0.85 ± 1.03).
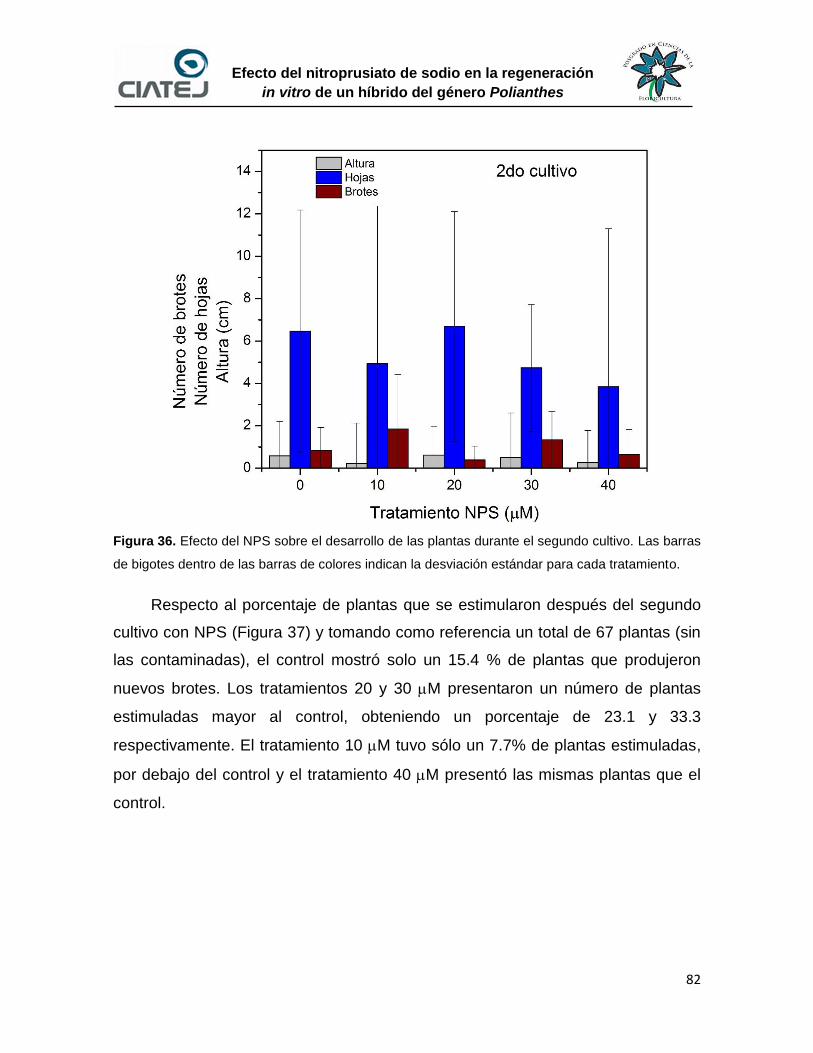
82
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 36. Efecto del NPS sobre el desarrollo de las plantas durante el segundo cultivo. Las barras
de bigotes dentro de las barras de colores indican la desviación estándar para cada tratamiento.
Respecto al porcentaje de plantas que se estimularon después del segundo
cultivo con NPS (Figura 37) y tomando como referencia un total de 67 plantas (sin
las contaminadas), el control mostró solo un 15.4 % de plantas que produjeron
nuevos brotes. Los tratamientos 20 y 30 M presentaron un número de plantas
estimuladas mayor al control, obteniendo un porcentaje de 23.1 y 33.3
respectivamente. El tratamiento 10 M tuvo sólo un 7.7% de plantas estimuladas,
por debajo del control y el tratamiento 40 M presentó las mismas plantas que el
control.
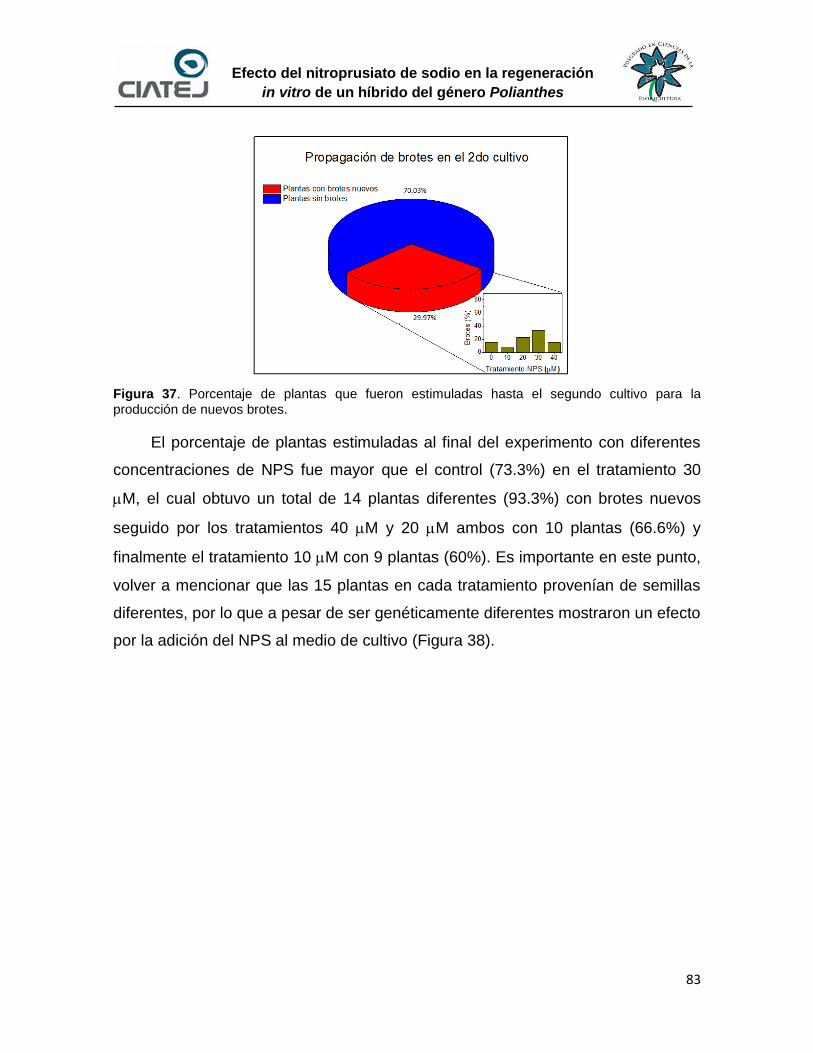
83
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 37. Porcentaje de plantas que fueron estimuladas hasta el segundo cultivo para la producción de nuevos brotes.
El porcentaje de plantas estimuladas al final del experimento con diferentes
concentraciones de NPS fue mayor que el control (73.3%) en el tratamiento 30
M, el cual obtuvo un total de 14 plantas diferentes (93.3%) con brotes nuevos
seguido por los tratamientos 40 M y 20 M ambos con 10 plantas (66.6%) y
finalmente el tratamiento 10 M con 9 plantas (60%). Es importante en este punto,
volver a mencionar que las 15 plantas en cada tratamiento provenían de semillas
diferentes, por lo que a pesar de ser genéticamente diferentes mostraron un efecto
por la adición del NPS al medio de cultivo (Figura 38).
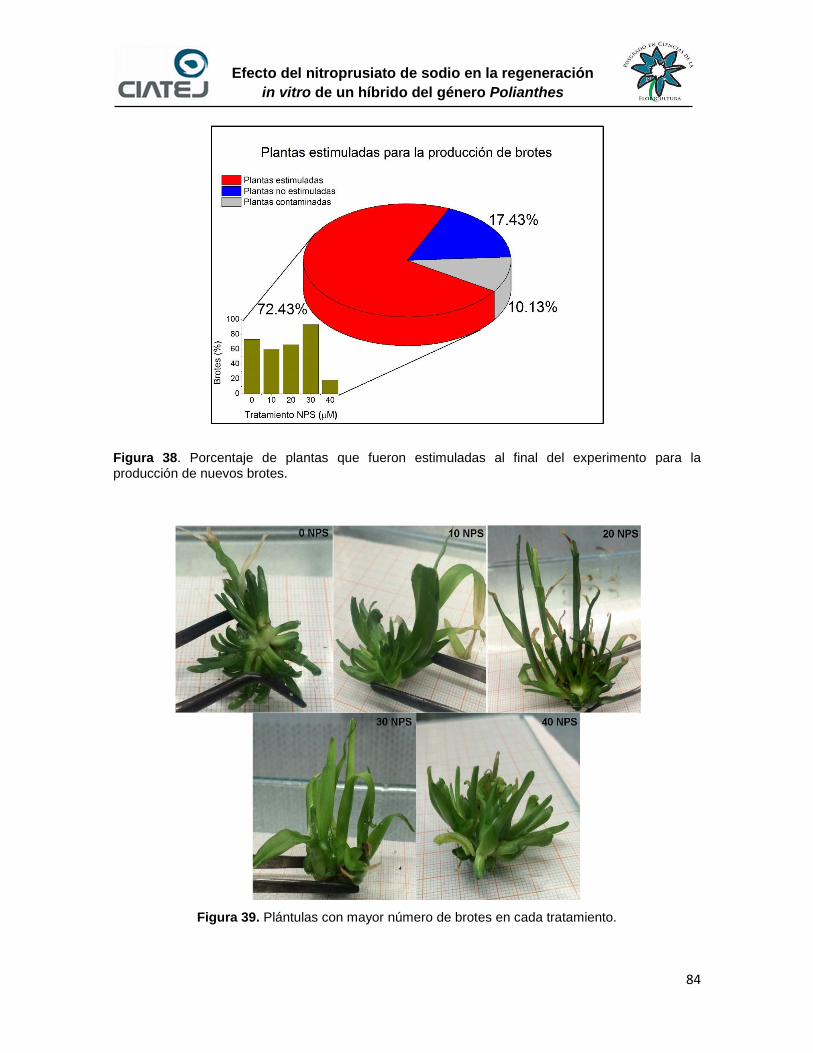
84
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 38. Porcentaje de plantas que fueron estimuladas al final del experimento para la
producción de nuevos brotes.
Figura 39. Plántulas con mayor número de brotes en cada tratamiento.
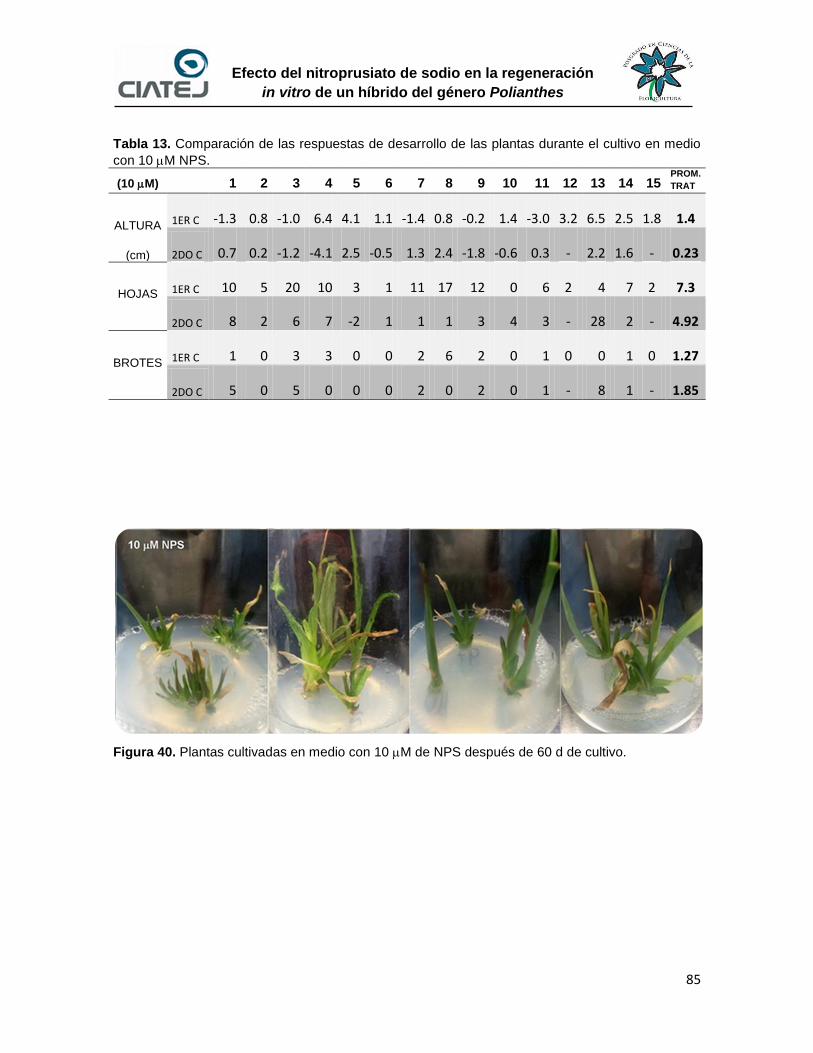
85
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Tabla 13. Comparación de las respuestas de desarrollo de las plantas durante el cultivo en medio
con 10 M NPS.
(10 M)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 PROM.
TRAT
ALTURA
(cm)
1ER C -1.3 0.8 -1.0 6.4 4.1 1.1 -1.4 0.8 -0.2 1.4 -3.0 3.2 6.5 2.5 1.8 1.4
2DO C 0.7 0.2 -1.2 -4.1 2.5 -0.5 1.3 2.4 -1.8 -0.6 0.3 - 2.2 1.6 - 0.23
HOJAS
1ER C 10 5 20 10 3 1 11 17 12 0 6 2 4 7 2 7.3
2DO C 8 2 6 7 -2 1 1 1 3 4 3 - 28 2 - 4.92
BROTES
1ER C 1 0 3 3 0 0 2 6 2 0 1 0 0 1 0 1.27
2DO C 5 0 5 0 0 0 2 0 2 0 1 - 8 1 - 1.85
Figura 40. Plantas cultivadas en medio con 10 M de NPS después de 60 d de cultivo.

86
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Tabla 14. Comparación de las respuestas de desarrollo de las plantas durante el cultivo en medio
con 20 M NPS.
(20 M)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 PROM.
TRAT
ALTURA
(cm)
1ER C 2.8 0.2 0.5 1.8 3.6 0.1 0.4 -1.1 0.6 1.0 -1.5 2.3 2.8 0.2 0.5 0.73
2DO C 0.3 1.5 -0.5 -0.9 1.6 0 0 -1.8 1.3 1.6 0 2.1 - 2.9 - 0.62
HOJAS
1ER C 1 4 6 8 3 4 2 6 1 3 1 4 -2 13 6 4.0
2DO C 1 7 7 19 1 4 7 6 5 5 5 3 - 17 - 6.69
BROTES
1ER C 0 0 0 2 0 1 1 2 0 1 0 0 1 0 1 0.6
2DO C 0 1 0 2 0 0 0 0 1 0 1 0 - 0 - 0.39
Figura 41. Plantas cultivadas en medio con 20 M de NPS después de 60 d de cultivo.

87
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Tabla 15. Comparación de las respuestas de desarrollo de las plantas durante el cultivo en medio
con 30 M NPS.
(30 M)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 PROM.
TRAT
ALTURA
(cm)
1ER C 4.7 -2.5 1.1 1.6 0.5 3.0 1.7 0.3 0.9 0.4 2.3 1.8 -1.3 -2.4 0.9 0.86
2DO C -3.5 -0.4 -0.4 0.8 1.5 0.3 3.0 2.2 5.6 0.8 -1.4 -0.7 -0.5 1.0 -0.8 0.5
HOJAS
1ER C 8 18 12 14 2 4 3 8 7 4 12 1 3 8 2 7.1
2DO C 7 9 7 3 2 7 1 -2 3 6 4 7 5 8 4 4.73
BROTES
1ER C 2 1 1 1 0 1 0 1 1 0 2 0 0 2 0 0.8
2DO C 1 5 3 0 0 2 0 1 1 2 1 2 1 0 1 1.3
Figura 42. Plantas cultivadas en medio con 30 M de NPS después de 60 d de cultivo.

88
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Tabla 16. Comparación de las respuestas de desarrollo de las plantas durante el cultivo en medio
con 40 M NPS.
(40 M)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 PROM.
TRAT
ALTURA
(cm)
1ER C 3.0 2.4 1.8 -3.1 2.4 2.0 2.0 0.8 2.4 1.3 1.5 1.3 0.8 1.1 0.9 1.37
2DO C 0.0 1.3 0.6 0.0 -3.1 -2.0 0.2 1.2 0.4 1.0 0.0 -0.3 2.2 - 2.4 0.26
HOJAS
1ER C 5 5 7 19 4 6 1 4 5 14 11 2 10 9 16 7.9
2DO C 2 8 2 0 -1 6 0 2 24 11 2 2 -2 - -6 3.85
BROTES
1ER C 0 1 0 5 0 0 0 0 1 3 2 0 1 2 2 1.13
2DO C 0 2 0 0 0 1 0 1 4 1 0 0 0 - 0 0.64
Figura 43. Plantas cultivadas en medio con 40 M de NPS después de 60 días de cultivo.

89
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
7.2.3 Establecimiento de yemas axilares de la inflorescencia del hibrido
Para la fase del establecimiento de las yemas axilares a partir de la
inflorescencia del híbrido 1008157 se colectaron dos tipos de tallo. El primero
pertenecía a una inflorescencia muy desarrollada (Figura 44a) de tejido rígido,
leñoso y con botones florales abiertos. El segundo tipo de tallo que se colectó era
de una inflorescencia juvenil (Figura 44c), su tejido presentaba mayor plasticidad,
botones florales cerrados y todos agrupados en el ápice formando espiga. Ambos
tallos fueron colectados en diferente fecha. Se cortaron, desinfectaron y
establecieron como se mencionó en la sección 6.2.3. Se realizaron más de dos
colectas sin tener éxito en la respuesta de los explantes. Los segmentos de tallo
se necrosaban y el tejido moría. Solo dos establecimientos de tallos realizados por
separado tuvieron respuesta.
Sólo tres explantes en el primer establecimiento tuvieron respuesta. Fueron
segmentos de tallo tipo leñoso. Estos segmentos contenían yemas axilares
florales las cuales no terminaron de diferenciarse para formar un botón floral y
formaron un botón vegetativo como respuesta al medio de cultivo u otro factor
(Figura 44b). El tejido somático que se obtuvo en este establecimiento se utilizó
para desdiferenciarlo en la estandarización de la sección 6.1.2 y no fue
propagado. En el segundo establecimiento solo dos segmentos de tallo tuvieron
respuesta (Figura 44d). Los segmentos presentaban yemas axilares destinadas a
formar un botón floral. Sin embargo, siete meses después de que se establecieron
los segmentos de tallo, estos formaron brotes vegetativos estimulados por el
medio de cultivo, la plasticidad de las células del explante, entre otros factores.
Debido al prolongado tiempo de respuesta de las yemas axilares de la
inflorescencia, se optó por propagar los dos brotes obtenidos en lugar de realizar
un tercer establecimiento.

90
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 44. Establecimiento de yemas axilares a partir de la inflorescencia del hibrido 1008157.
7.2.4 Propagación de clones del híbrido 1008157
La propagación de los brotes vegetativos obtenidos a partir de yemas
axilares de la inflorescencia se realizó con la finalidad de incrementar el material
vegetal disponible para realizar los experimentos posteriores es este estudio. La
propagación se realizó por más de ocho meses utilizando diferentes medios de
cultivo (sección 6.2.2). Debido a que el híbrido 1007157 es una planta con
crecimiento vigoroso y sano (Figura 45a), los dos brotes iniciales se multiplicaron
hasta formar más de 50 explantes (Figura 45c). Algunos de esos explantes
presentaban brotes axilares pequeños (Figura 45b) los cuales permanecieron
unidos al brote central para propiciar un crecimiento más rápido de ambos. En la

91
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
última fase de la propagación, los explantes se cultivaron en medio MSC sin
reguladores de crecimiento y se colocaron en oscuridad con la finalidad de inducir
un decremento en la síntesis de clorofila.
Figura 45. Propagación de brotes vegetativos utilizando la multiplicación por yemas axilares.
Los explantes después de 12 días en oscuridad presentaron una disminución
del color verde intenso que habían presentado durante toda la etapa de
propagación (Figura 46a). Además las hojas de mayor longitud perdieron la
dureza, el vigor, mostraron un aspecto de marchitez y un color amarillo en
contraste con las hojas del explante cultivado en el mismo medio MSC pero en
presencia de luz (Figura 46b) el cual no mostró ninguno de los síntomas
mencionados.

92
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 46. Explantes clones del híbrido después de incubarlos en oscuridad. a) Algunos de los explantes clones del híbrido después de incubarlos durante 12 días en oscuridad. b) Comparación de explantes incubados en presencia de luz (centro) y oscuridad (laterales).
7.2.5 Estimulación de la producción de brotes en clones del híbrido
1008157
Después de cultivar los explantes en medio MSC durante 12 días en
oscuridad, los explantes fueron divididos en dos experimentos diferentes para
evaluar el efecto del NPS en la producción de brotes. En el primer experimento se
utilizó explantes que presentaban brotes (al menos uno) y en el segundo
experimento se utilizó explantes que no presentaron brotes. En ambos
experimentos los explantes se cultivaron en un medio con diferentes
concentraciones de NPS (0, 10, 20, 30 y 40 M) y se incubaron durante 30 d.

93
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Transcurridos los 30 d, en cada uno de los experimentos se realizó una
descripción de características de desarrollo donde los valores de cada variable de
respuesta reflejan la diferencia entre el estado final e inicial. Las variables de
respuesta consideradas fueron la altura de la planta; el número de hojas y brotes.
7.2.5.1 Clones del híbrido con al menos un brote antes del cultivo en
NPS
Cuatro repeticiones por tratamiento fueron realizadas por experimento. Se
evaluó la respuesta de los explantes clones del hibrido que tenían brotes antes del
cultivo con diferentes concentraciones de NPS. De acuerdo a los valores
obtenidos (Tabla 17), para la altura, independientemente del tratamiento, el 50%
del total de las plantas, no mostraron crecimiento o en su defecto presentaron una
disminución. Esto se traduce, en que, la mayoría de las hojas que presentaron
síntomas de marchitez después del periodo de incubación en oscuridad, no
lograron recuperarse durante el cultivo con NPS. Dicho tejido se secó (Figura 47)
dando como resultado una disminución en la longitud de la hoja que se tomó como
referencia para medir la altura de la planta. Respecto al número de explantes que
fueron estimulados para producir brotes nuevos, en el control, el 100% de los
explantes produjo brotes, en contraste con los tratamientos con NPS donde solo el
tratamiento 40 M mostró un 100% de explantes con producción de brotes. Sin
embargo, un 75% del total de explantes presentó un estímulo para la producción
de brotes nuevos.
Tabla 17. Comparación entre las variables de respuesta para los clones del híbrido con al menos un brote durante el cultivo en medio con NPS.
NPS (M)
0 0 0 0 10 10 10 10 20 20 20 20 30 30 30 30 40 40 40 40
ALTURA
(cm) 0.0 2.1 2.1 -0.6 -0.6 1.4 4.6 1.1 -3.3 -2.8 5.4 6.0 -0.5 -4.2 1.0 -1.7 1.5 1.8 0.0 -1.8
HOJAS -1 14 5 11 7 2 14 8 12 12 21 2 5 0 4 11 8 33 10 6
BROTES 1 3 1 1 0 0 1 2 2 2 2 0 1 0 0 4 3 3 1 2

94
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 47. Marchitamiento y muerte del tejido del ápice de las hojas adultas durante el cultivo con NPS.
En términos del valor promedio, para la altura, los tratamientos 10 y 20 M
fueron numéricamente superiores que el control, mientras que los tratamientos con
mayor concentración de NPS (30 y 40 M) estuvieron por debajo del crecimiento
en el control (Tabla 18). Para la variable número de hojas, todos los tratamientos
con NPS superaron al control con excepción del tratamiento 30 M que fue menor.
Para la producción de nuevos brotes, y retomando que esta variable es la de
mayor interés, el control mostró un promedio mayor o igual que los tratamientos
10, 20 y 30 M. Únicamente el tratamiento con 40 M numéricamente superó el
número de brotes, sin embargo, no existió diferencia significativa de acuerdo al
ANOVA (p= 0.5996, 0.4880 y 0.3662, α= 0.05, para altura, número de hojas y
número de brotes respectivamente, Figura 48).
Tabla 18. Promedio y desviación estándar para las variables de respuesta en clones del híbrido con presencia de brotes durante el cultivo en medio con NPS.
NPS (M)
0 10 20 30 40
PROM DS PROM DS PROM DS PROM DS PROM DS
ALTURA
(cm) 0.9 1.41 1.63 2.8 1.33 5.1 -1.35 2.2 0.63 1.23
HOJAS 7.5 4.58 7.75 4.92 11.75 7.76 6.67 4.55 14.25 12.6
BROTES 1.5 1.0 0.75 0.96 1.5 1.0 1.25 1.89 2.25 0.96
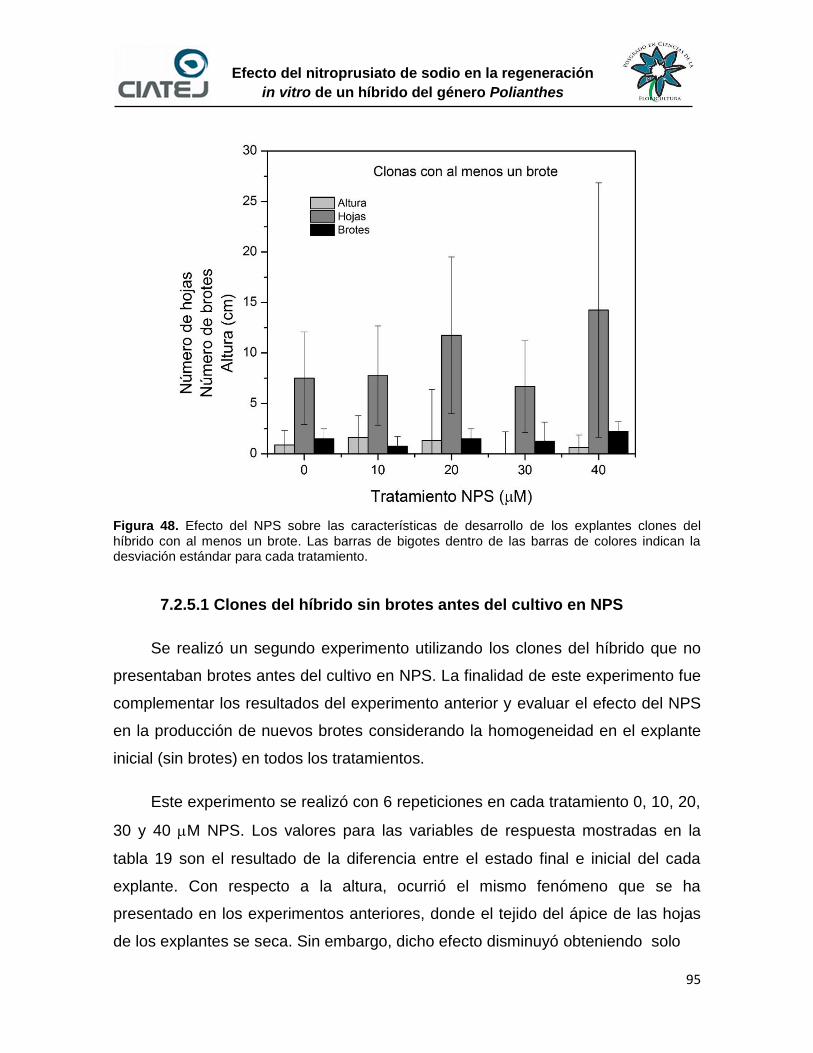
95
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 48. Efecto del NPS sobre las características de desarrollo de los explantes clones del híbrido con al menos un brote. Las barras de bigotes dentro de las barras de colores indican la desviación estándar para cada tratamiento.
7.2.5.1 Clones del híbrido sin brotes antes del cultivo en NPS
Se realizó un segundo experimento utilizando los clones del híbrido que no
presentaban brotes antes del cultivo en NPS. La finalidad de este experimento fue
complementar los resultados del experimento anterior y evaluar el efecto del NPS
en la producción de nuevos brotes considerando la homogeneidad en el explante
inicial (sin brotes) en todos los tratamientos.
Este experimento se realizó con 6 repeticiones en cada tratamiento 0, 10, 20,
30 y 40 M NPS. Los valores para las variables de respuesta mostradas en la
tabla 19 son el resultado de la diferencia entre el estado final e inicial del cada
explante. Con respecto a la altura, ocurrió el mismo fenómeno que se ha
presentado en los experimentos anteriores, donde el tejido del ápice de las hojas
de los explantes se seca. Sin embargo, dicho efecto disminuyó obteniendo solo
96
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
un 33.3% del total de explantes que disminuyeron su altura en comparación con el
50% en el experimento de clones con brotes. Es importante considerar que la
mitad del número de explantes del 33.3% pertenecen únicamente al tratamiento
40 M. Para el número de brotes (la variable de mayor interés), se observó una
disminución con respecto al experimento anterior (clones con al menos un brote)
ya que solo el 56.6% del total de explantes fueron estimuladas para la producción
de brotes. Con referencia a esto, el tratamiento 40 M tuvo un 83.3% de plantas
estimuladas superando al control (66.6%) y a los otros tratamientos con NPS.
Tabla 19. Comparación de las variables de respuesta (altura, hojas y brotes) para los clones del híbrido sin brotes durante el cultivo en medio con NPS. (6 repeticiones por tratamiento)
NPS (M)
0 10 20 30 40
ALTURA
(cm)
-2.9 0.5 1.8 3.7 1.7 0.7 4.5 1.5 -3.4 1.8 2.1 1.3 7.8 -2.7 -0.8
-0.3 1.3 1.2 -1.2 0.3 0.0 2.3 2.5 1.2 2.1 1.1 -0.2 -0.2 -1.7 -0.3
HOJAS
6 3 2 4 5 13 1 4 8 4 4 4 3 3 10
4 8 3 5 -2 4 5 2 3 4 2 7 6 2 1
BROTES
2 0 0 0 0 5 0 1 3 1 1 0 2 1 2
1 3 1 1 0 0 2 0 0 0 0 4 2 1 0
En términos de promedio (Tabla 20) para la altura, todos los tratamientos con
NPS fueron superiores que el control, sin embargo, esté valor no tiene el suficiente
respaldo debido a que sólo uno de los 6 explantes aporto este valor. Por lo tanto,
sería más razonable considerar el tratamiento 30 M como el mejor debido a que
5 de 6 plantas incrementaron su longitud. Para la variable número de hojas, el
tratamiento 10 M superó al control y los tratamientos 20, 30 y 40 M fueron
menores al control. El efecto en el número brotes, y retomando que esta variable
es la de mayor interés, los tratamientos 10, 20 y 30 M tuvieron el mismo
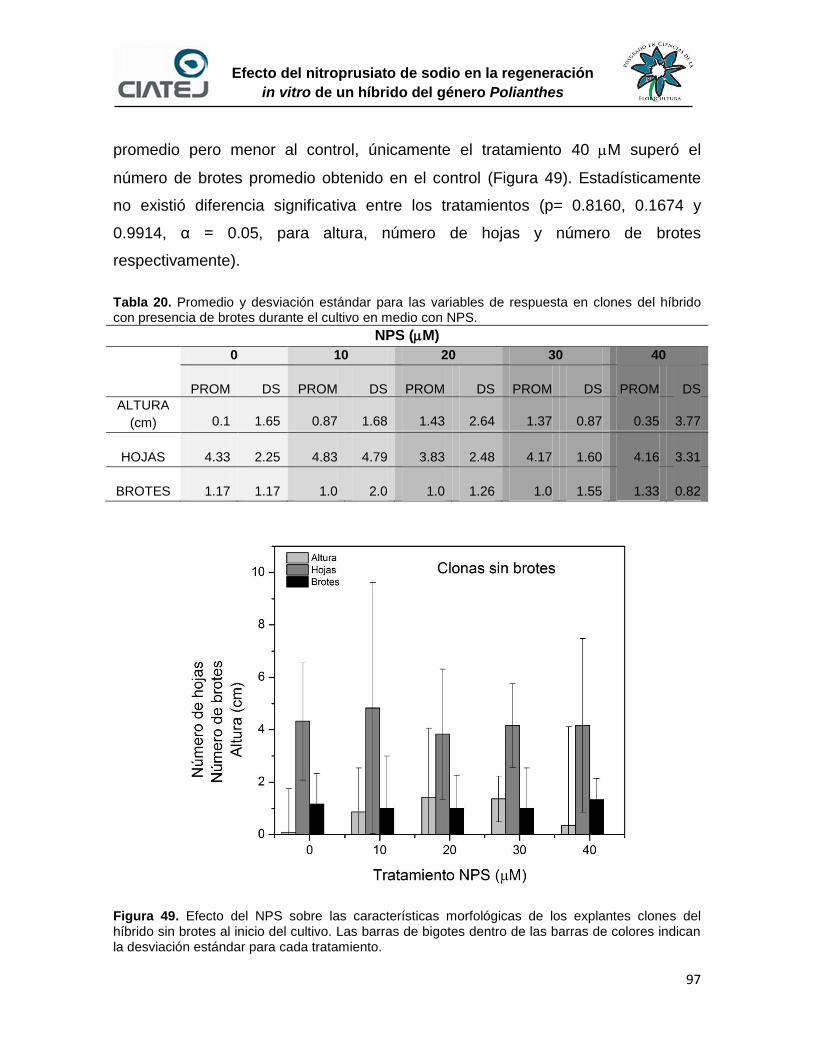
97
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
promedio pero menor al control, únicamente el tratamiento 40 M superó el
número de brotes promedio obtenido en el control (Figura 49). Estadísticamente
no existió diferencia significativa entre los tratamientos (p= 0.8160, 0.1674 y
0.9914, α = 0.05, para altura, número de hojas y número de brotes
respectivamente).
Tabla 20. Promedio y desviación estándar para las variables de respuesta en clones del híbrido con presencia de brotes durante el cultivo en medio con NPS.
NPS (M)
0 10 20 30 40
PROM DS PROM DS PROM DS PROM DS PROM DS
ALTURA
(cm) 0.1 1.65 0.87 1.68 1.43 2.64 1.37 0.87 0.35 3.77
HOJAS 4.33 2.25 4.83 4.79 3.83 2.48 4.17 1.60 4.16 3.31
BROTES 1.17 1.17 1.0 2.0 1.0 1.26 1.0 1.55 1.33 0.82
Figura 49. Efecto del NPS sobre las características morfológicas de los explantes clones del híbrido sin brotes al inicio del cultivo. Las barras de bigotes dentro de las barras de colores indican la desviación estándar para cada tratamiento.

98
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
7.2.6 Extracción y cuantificación de clorofila
Los explantes clones del híbrido (con y sin brotes) se cultivaron en un medio
con diferentes concentraciones de NPS. Después de hacer la evaluación para las
variables de respuesta se tomó una muestra de material vegetal de cada explante
para extraer la clorofila utilizando etanol por 24 hrs. Posteriormente se cuantificó
clorofila a y b por medio de espectrofotometría. Las absorbancias obtenidas para
los dos tipos de clorofila en cada explante fueron convertidas a g/mL mediante
las fórmulas de Wellburn (1994).
Se obtuvo el promedio de clorofila a y b para cada tratamiento con NPS (0,
10, 20, 30 y 40 M) de acuerdo a los dos experimentos con o sin brotes antes del
cultivo. Este promedio representa la cantidad de clorofila en g/mL en los
explantes de cada tratamiento. El promedio para la clorofila a y b en el tratamiento
0 M NPS fue menor en los explantes con al menos un brote respecto a los
explantes sin brotes. Sin embargo, estadísticamente no existió diferencia
significativa para el contenido de clorofila tanto a como b entre ambos
experimentos (clorofila a p = 0.2893; clorofila b: p = 0.2802, α = 0.05).
Para los explantes con brotes, el contenido de clorofila a fue menor en el
control, seguido de 20, 10, 40 y 30 M NPS. Estadísticamente dicha disminución
en el control no tuvo efecto significativo con respecto a todos los tratamientos (p =
0.2062, α = 0.05). Para la clorofila b, en este mismo experimento, los tratamientos
10, 30 y 40 M superaron al control, excepto el tratamiento 20 M siendo una vez
más numéricamente el tratamiento 30 M NPS el de mayor contenido y
presentando diferencia estadísticamente significativa entre los tratamientos (p =
0.0255, α = 0.05). La diferencia que existió fue para los tratamientos 10 y 30 M
con el tratamiento 20 M, lo que sólo confirmó que los niveles más bajos de
clorofila se dieron en el tratamiento 20 M (Figura 50).
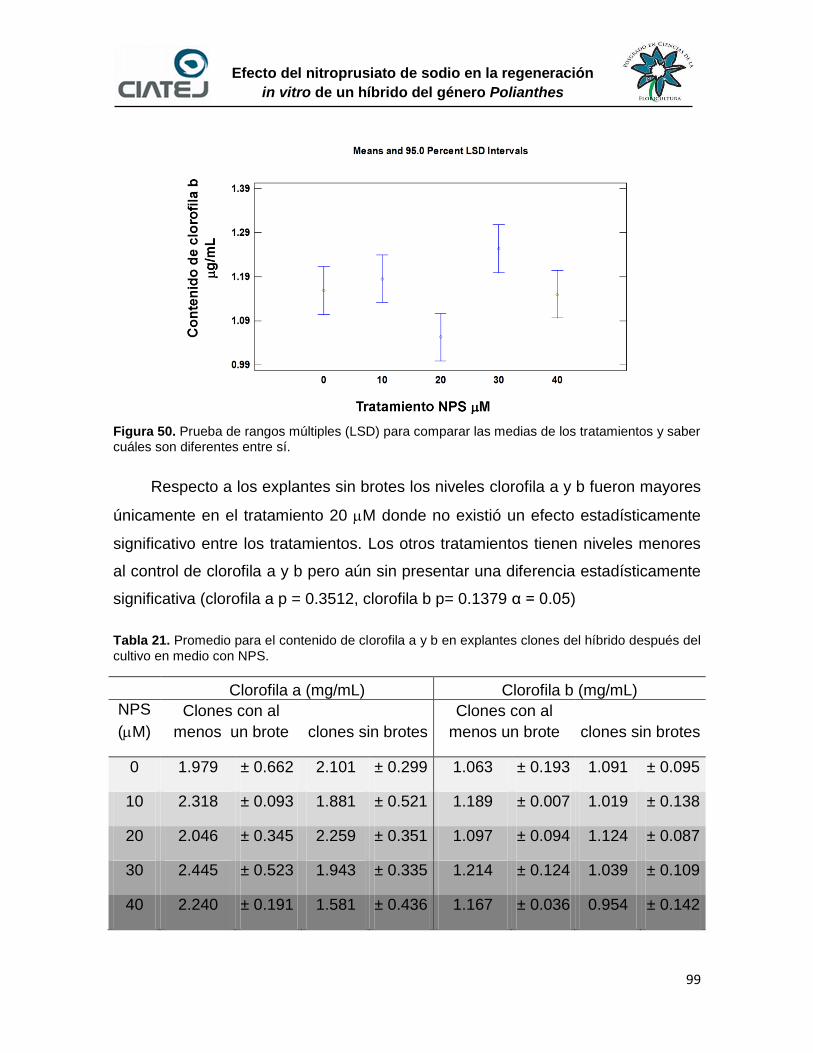
99
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 50. Prueba de rangos múltiples (LSD) para comparar las medias de los tratamientos y saber cuáles son diferentes entre sí.
Respecto a los explantes sin brotes los niveles clorofila a y b fueron mayores
únicamente en el tratamiento 20 M donde no existió un efecto estadísticamente
significativo entre los tratamientos. Los otros tratamientos tienen niveles menores
al control de clorofila a y b pero aún sin presentar una diferencia estadísticamente
significativa (clorofila a p = 0.3512, clorofila b p= 0.1379 α = 0.05)
Tabla 21. Promedio para el contenido de clorofila a y b en explantes clones del híbrido después del
cultivo en medio con NPS.
Clorofila a (mg/mL) Clorofila b (mg/mL)
NPS
(M)
Clones con al
menos un brote clones sin brotes
Clones con al
menos un brote clones sin brotes
0 1.979 ± 0.662 2.101 ± 0.299 1.063 ± 0.193 1.091 ± 0.095
10 2.318 ± 0.093 1.881 ± 0.521 1.189 ± 0.007 1.019 ± 0.138
20 2.046 ± 0.345 2.259 ± 0.351 1.097 ± 0.094 1.124 ± 0.087
30 2.445 ± 0.523 1.943 ± 0.335 1.214 ± 0.124 1.039 ± 0.109
40 2.240 ± 0.191 1.581 ± 0.436 1.167 ± 0.036 0.954 ± 0.142
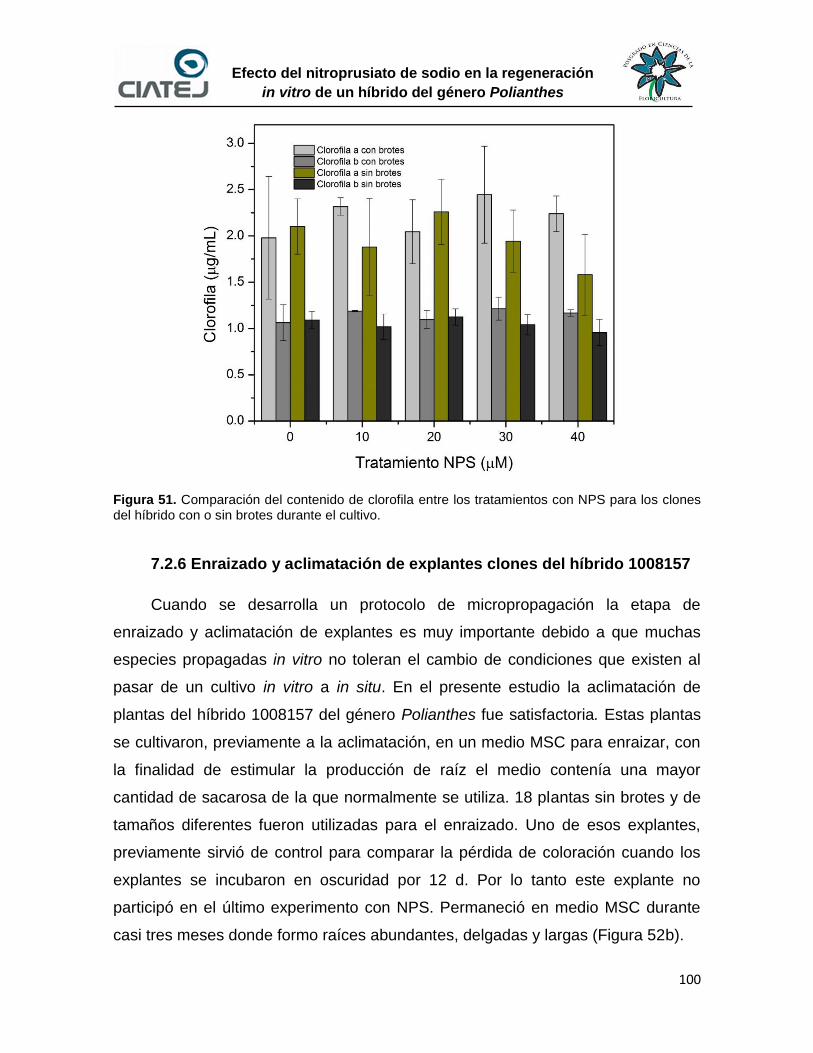
100
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Figura 51. Comparación del contenido de clorofila entre los tratamientos con NPS para los clones del híbrido con o sin brotes durante el cultivo.
7.2.6 Enraizado y aclimatación de explantes clones del híbrido 1008157
Cuando se desarrolla un protocolo de micropropagación la etapa de
enraizado y aclimatación de explantes es muy importante debido a que muchas
especies propagadas in vitro no toleran el cambio de condiciones que existen al
pasar de un cultivo in vitro a in situ. En el presente estudio la aclimatación de
plantas del híbrido 1008157 del género Polianthes fue satisfactoria. Estas plantas
se cultivaron, previamente a la aclimatación, en un medio MSC para enraizar, con
la finalidad de estimular la producción de raíz el medio contenía una mayor
cantidad de sacarosa de la que normalmente se utiliza. 18 plantas sin brotes y de
tamaños diferentes fueron utilizadas para el enraizado. Uno de esos explantes,
previamente sirvió de control para comparar la pérdida de coloración cuando los
explantes se incubaron en oscuridad por 12 d. Por lo tanto este explante no
participó en el último experimento con NPS. Permaneció en medio MSC durante
casi tres meses donde formo raíces abundantes, delgadas y largas (Figura 52b).

101
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Las otras 17 plantas que se pusieron en medio MSC para enraizar, después
de los 40 d al retirarlos del medio de cultivo presentaron apenas la punta de una
raíz en la base. Un mes después de que las 18 plantas se cultivaron en suelo, 16
explantes lograron aclimatarse (Figura 52c) y solo dos murieron por contaminación
por hongos. Las plantas aclimatadas presentaron un crecimiento vigoroso, poseen
hojas muy verdes, gruesas y sanas (Figura 52d). Además, las plantas fueron
retiradas del suelo para ver el desarrollo de sus raíces. Comparando las raíces
antes y después de la aclimatación en la Figura 52a, se logra observar que las
raíces son más gruesas independientemente del tamaño del explante. Se
comprobó la característica del vigor en el crecimiento que posee este híbrido tanto
radicular como apical.
Figura 52. Plantas del híbrido 1008157 aclimatadas a) Plantas con presencia de raíces después
de un mes de aclimatación barra= 5 cm; b) Planta recién retirada del medio de cultivo; c) 16 plantas aclimatadas; d) Las plantas aclimatadas presentaron un crecimiento vigoroso, poseen hojas muy verdes, gruesas y sanas.

102
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
IX. CONCLUSIONES
La regeneración del híbrido 1008157 del género Polianthes fue satisfactoria
únicamente por la técnica de propagación por yemas axilares de la inflorescencia.
No fue posible comprobar que la adición de nitroprusiato de sodio al medio
de cultivo favorece la inducción de embriogénesis somática o estimula la
producción de brotes en el cultivo in vitro del híbrido. Sin embargo, se reporta la
descripción detallada del proceso de desdiferenciación del tejido somático de
plántulas utilizando NPS y picloram en 16 concentraciones diferentes. Por otro
lado, en la producción de brotes, el NPS si tiene un efecto numéricamente mayor
en comparación con el medio sin NPS para una concentración de 40 M.
Debido al crecimiento vigoroso y capacidad de multiplicación del híbrido
1008157 fue posible establecer exitosamente un protocolo de micropropagación
desde el inicio hasta la etapa de aclimatación contribuyendo así en incrementar el
número de plantas existentes del híbrido 1008157.
Este estudio puede servir de referencia para trabajos posteriores en la
propagación de otros híbridos o especies cercanas al género.

103
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
XI. BIBLIOGRAFIA
Abdullah, S. (2012). Somatic embryogenesis and regeneration of Polianthes
tuberosa L. Universidad de Malaya. Tesis de Maestria. 191 pp
Ahmad, M. S., Ahmad, T., Zaidi, N., y Ahmad, N. I. (2006). High frequency in vitro
propagation of Polianthes tuberosa. Pakistan Journal of Scientific and
Industrial Research, 49(5), 344-348.
Airaki, M., Corpas Aguirre, F. J., Martínez, P., y Manuel, J. (2012). Función de las
especies de oxígeno y nitrógeno reactivo (ROS y RNS) en plantas de
pimiento (Capsicum annuum L.) durante el desarrollo y en estrés por baja
temperatura. Universidad de Granada. Tesis de Doctorado. 225 pp.
Arnold, W., Longnecker, D., y Epstein, R. (1984). Photodegradation of sodium
nitroprusside: biologic activity and cyanide release. Anesthesiology, 61(3),
254-260.
Bakhshaie, M., Babalar, M., Mirmasoumi, M., y Khalighi, A. (2010). Effects of light,
sucrose, and cytokinins on somatic embryogenesis in Lilium ledebourii
(Baker) Bioss. via transverse thin cell-layer cultures of bulblet microscales.
The Journal of Horticultural Science and Biotechnology, 85(6), 491-496.
Barba-Gonzalez, R., Rodríguez-Domínguez, J. M., Castañeda-Saucedo, C.,
Rodríguez, A., Van Tuyl, J. M., y Tapia-Campos, E. (2012). Mexican
Geophytes I. The Genus Polianthes. Floriculture and Ornamental
Biotechnology Global Science Books, 122-128.
Barroso, J. B., Corpas, F. J., Carreras, A., Sandalio, L. M., Valderrama, R., Palma,
J., del R o, L. A. (1999). Localization of nitric-oxide synthase in plant
peroxisomes. Journal of Biological Chemistry, 274(51), 36729-36733.
Beligni, M. V., y Lamattina, L. (1999). Nitric oxide counteracts cytotoxic processes
mediated by reactive oxygen species in plant tissues. Planta, 208(3), 337-
344.

104
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Castillo, A., Egana, B., Sanz, J., y Cistue, L. (1998). Somatic embryogenesis and
plant regeneration from barley cultivars grown in Spain. Plant Cell Reports,
17(11), 902-906.
Corpas, F. J., Barroso, J. B., Carreras, A., Valderrama, R., Palma, J. M., León, A.
M., y del Río, L. A. (2006). Constitutive arginine-dependent nitric oxide
synthase activity in different organs of pea seedlings during plant
development. Planta, 224(2), 246-254.
Corpas, F. J., Palma, J. M., del Río, L. A., y Barroso, J. B. (2009). Evidence
supporting the existence of l‐arginine‐dependent nitric oxide synthase
activity in plants. New Phytologist, 184(1), 9-14.
Correa-Aragunde, N., Graziano, M., Chevalier, C., y Lamattina, L. (2006). Nitric
oxide modulates the expression of cell cycle regulatory genes during lateral
root formation in tomato. Journal of Experimental Botany, 57(3), 581-588.
Datta, S. K., Misra, P., Mandal, A. K., y Chakrabarty, D. (2002). Direct shoot
organogenesis from different explants of chrysanthemum, marigold, and
tuberose. Israel Journal of Plant Sciences, 50(4).
Davies, P. J. (2010). Regulatory factors in hormone action: Level, location and
signal transduction in Plant Hormones (16-35): Ed. Springer
De Jong, A. J., Schmidt, E. D., & de Vries, S. C. (1993). Early events in higher-
plant embryogenesis. Plant molecular biology, 22(2), 367-377.
Dehdezi, A. A., sadat Mousavi, E., & Azadi, P. (2014). Evaluation of Different
Growth Regulators on Proliferation of Polianthes tuberosa. Bull. Env.
Pharmacol. Life Sci, 3, 172-174.
Del Río, L. A. (2011). Peroxisomes as a cellular source of reactive nitrogen species
signal molecules. Archives of biochemistry and biophysics, 506(1), 1.
Del Río, L. A., Corpas, F. J., Barroso, J. B., López-Huertas, E., & Palma, J. M.
(2014). Function of peroxisomes as a cellular source of nitric oxide and
other reactive nitrogen species Nitric Oxide in Plants: Metabolism and Role
in Stress Physiology (33-55): Ed. Springer.

105
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Del Río, L. A., Corpas, J. F., Sandalio, L. M., Palma, J. M., Gómez, M., y Barroso,
J. B. (2002). Reactive oxygen species, antioxidant systems and nitric oxide
in peroxisomes. Journal of Experimental Botany, 53(372), 1255-1272.
Delledonne, M. (2005). NO news is good news for plants. Current opinion in plant
biology, 8(4), 390-396.
Dodeman, V. L., Ducreux, G., y Kreis, M. (1997). Zygotic embryogenesis versus
somatic embryogenesis. Journal of Experimental Botany, 48(8), 1493-1509.
Dudits, D., Bogre, L., y Gyorgyey, J. (1991). Molecular and cellular approaches to
the analysis of plant embryo development from somatic cells in vitro. Journal
of Cell Science (United Kingdom).
Estrada-Basaldua, J. A., Pedraza-Santos, M. E., de la Cruz-Torres, E., Martínez-
Palacios, A., Sáenz-Romero, C., y Morales-García, J. L. (2011). Efecto de
rayos gamma 60Co en nardo (Polianthes tuberosa L.). Revista mexicana de
ciencias agrícolas (3), 445-458.
Eum, H. L., Hwang, D. K., y Lee, S. K. (2009). Nitric oxide reduced chlorophyll
degradation in broccoli (Brassica oleracea L. var. italica) florets during
senescence. Food Science and Technology International.
Feelisch, M. (1998). The use of nitric oxide donors in pharmacological studies.
Naunyn-Schmiedeberg's archives of pharmacology, 358(1), 113-122.
Fehér, A., Pasternak, T. P., y Dudits, D. (2003). Transition of somatic plant cells to
an embryogenic state. Plant Cell, Tissue and Organ Culture, 74(3), 201-228.
Feria-Arroyo, T. P., Solano, E., y García-Mendoza, A. (2010). Reevaluación del
riesgo de extinción de cinco especies del género Polianthes L.(Agavaceae).
Acta botánica mexicana 1(92), 11-28.
Filonova, L. H., Bozhkov, P. V., y von Arnold, S. (2000). Developmental pathway of
somatic embryogenesis in Picea abies as revealed by time‐lapse tracking.
Journal of Experimental Botany, 51(343), 249-264.

106
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Flores-Benítez, S., Jiménez-Bremont, J. F., Rosales-Mendoza, S., Argüello-
Astorga, G. R., Castillo-Collazo, R., y Alpuche-Solís, Á. G. (2007). Genetic
transformation of Agave salmiana by Agrobacterium tumefaciens and
particle bombardment. Plant Cell, Tissue and Organ Culture, 91(3), 215-
224.
Fröhlich, A., y Durner, J. (2011). The hunt for plant nitric oxide synthase (NOS): is
one really needed? Plant science, 181(4), 401-404.
Gajbhiye, S., Tripathi, M., Vidya, M., Singh, M., Baghel, B., y Tiwari, S. (2011).
Direct shoot organogenesis from cultured stem disc explants of tuberose
(Polianthes tuberosa Linn.). Journal of Agricultural Technology, 7(3), 695-
709.
Gaspar, T., Kevers, C., Penel, C., Greppin, H., Reid, D. M., y Thorpe, T. A. (1996).
Plant hormones and plant growth regulators in plant tissue culture. In Vitro
Cellular & Developmental Biology-Plant, 32(4), 272-289.
George, E. F. (1993). Plant propagation by tissue culture. Part 1: the technology:
Exegetics limited.
George, E. F., Hall, M. A., y De Klerk, G. J. (2008). Micropropagation: uses and
methods in Plant propagation by tissue culture (29-64): Ed. Springer.
Gifford, E. M., y Foster, A. S. (1987). Morphology and evolution of vascular plants
(626): Ed. W.H. Freeman and Company.
Goldberg, R. B., de Paiva, G., y Yadegari, R. (1994). Plant Embryogenesis: Zygote
to Seed. Science, 266, 605-614.
Guo, F.-Q., Okamoto, M., y Crawford, N. M. (2003). Identification of a plant nitric
oxide synthase gene involved in hormonal signaling. Science, 302(5642),
100-103.
Han, X., Yang, H., Duan, K., Zhang, X., Zhao, H., You, S., y Jiang, Q. (2009).
Sodium nitroprusside promotes multiplication and regeneration of Malus
hupehensis in vitro plantlets. Plant Cell, Tissue and Organ Culture, 96(1),
29-34.

107
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Hernández, F. M. (2013). Caracterización de las condiciones para la
micropropagación de Polianthes tuberosa L. a partir de cormo, hoja, flor y
yema. Colegio de Postgraduados. Tesis Maestria.
Hou, Y., Janczuk, A., y Wang, P. (1999). Current trends in the development of
nitric oxide donors. Current pharmaceutical design, 5(6), 417-442.
Jiménez, V. M. (2005). Involvement of plant hormones and plant growth regulators
on in vitro somatic embryogenesis. Plant Growth Regulation, 47(2-3), 91-
110.
Kaur, P., y Kothari, S. (2004). In vitro culture of kodo millet: influence of 2, 4-D and
picloram in combination with kinetin on callus initiation and regeneration.
Plant Cell, Tissue and Organ Culture, 77(1), 73-79.
Khan, M. N., Mobin, M., Mohammad, F., y Saquib, M. A. (2013). Nitric oxide
synthesis in plants: a recent update. Global Journal of Botanical Science, 1,
22-28.
Kordestani, G. K., y Karami, O. (2008). Picloram-induced somatic embryogenesis
in leaves of strawberry (Fragaria ananassa L.). Acta Biológica Cracoviensia
Series Botanica, 50(1), 69-72.
Krishnamurthy, K., Mythili, J., y Meenakshi, S. (2001). Micropropagation studies in
"single" vs."double" types of tuberose (Polianthes tuberosa L.). Journal of
Applied Horticulture, 3(2), 82-84.
Lamattina, L., García-Mata, C., Graziano, M., y Pagnussat, G. (2003). Nitric oxide:
the versatility of an extensive signal molecule. Annual Review of Plant
Biology, 54(1), 109-136.
Lazalt, A., Beligni, M. V., y Lamattina, L. (1997). Nitric oxide preserves the level of
chlorophyll in potato leaves infected by Phytophthora infestans. European
Journal of Plant Pathology, 103(7), 643-651.
Lipton, S. A., Choi, Y.-B., Pan, Z.-H., Lei, S. Z., Chen, H.-S. V., Sucher, N. J., y
Stamler, J. S. (1993). A redox-based mechanism for the neuroprotective
and neurodestructive effects of nitric oxide and related nitroso-compounds.
Nature, 364, 626-632.

108
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Liu, F., y Guo, F.-Q. (2013). Nitric oxide deficiency accelerates chlorophyll
breakdown and stability loss of thylakoid membranes during dark-induced
leaf senescence in Arabidopsis. PloS ONE, 8(2),1-12.
Manjunatha, G., Lokesh, V., y Neelwarne, B. (2010). Nitric oxide in fruit ripening:
trends and opportunities. Biotechnology advances, 28(4), 489-499.
Martínez-Palacios, A., Ortega-Larrocea, M. P., Chávez, V. M., y Bye, R. (2003).
Somatic embryogenesis and organogenesis of Agave victoriae-reginae:
Considerations for its conservation. Plant Cell, Tissue and Organ Culture,
74(2), 135-142.
Mathieu, M., Lelu-Walter, M., Blervacq, A., David, H., Hawkins, S., y Neutelings, G.
(2006). Germin-like genes are expressed during somatic embryogenesis
and early development of conifers. Plant molecular biology, 61(4-5), 615-
627.
Mendoza, M. G., y Kaeppler, H. F. (2002). Auxin and sugar effects on callus
induction and plant regeneration frequencies from mature embryos of wheat
(Triticum aestivum L.). In Vitro Cellular & Developmental Biology-Plant,
38(1), 39-45.
Molphe, E., Ramírez, R., Núñez, H., y Ochoa, N. (1999). Introducción al cultivo de
tejidos vegetales en México: Universidad Autónoma de Aguascalientes.
Monja-Mio, K. M., y Robert, M. L. (2013). Direct somatic embryogenesis of Agave
fourcroydes Lem. through thin cell layer culture. In Vitro Cellular &
Developmental Biology-Plant, 49(5), 541-549.
Mur, L. A., Mandon, J., Persijn, S., Cristescu, S. M., Moshkov, I. E., Novikova, G.
V., y Gupta, K. J. (2013). Nitric oxide in plants: an assessment of the current
state of knowledge. AoB Plants, 5. 1-17
Murashige, T., y Skoog, F. (1962). A revised medium for rapid growth and bio
assays with tobacco tissue cultures. Physiologia plantarum, 15(3), 473-497.
Naz, S., Aslam, F., Ilyas, S., Shahzadi, K., y Tariq, A. (2012). In vitro propagation
of tuberose (Polianthes tuberosa). Journal of Medicinal Plants Research,
6(24), 4107-4112.

109
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Nazneen, S., Jabeen, M., y Ilahi, I. (2003). Micropropagation of Polyanthus
tuberosa (Tuberose) through Callusformation. Pakistan Journal of Botany
(Pakistan).
Nomura, K., y Komamine, A. (1985). Identification and isolation of single cells that
produce somatic embryos at a high frequency in a carrot suspension
culture. Plant Physiology, 79(4), 988-991.
Nomura, K., y Komamine, A. (1995). Physiological and biochemical aspects of
somatic embryogenesis in In vitro embryogenesis in plants (249-265): Ed.
Springer.
Oldfield, S. (1997). Cactus and succulent plants: status survey and conservation
action plan: International Union for Conservation of Nature and Natural
Resources (IUCN).
Ötvös, K., Pasternak, T. P., Miskolczi, P., Domoki, M., Dorjgotov, D., Bottka, S., y
Fehér, A. (2005). Nitric oxide is required for, and promotes auxin‐mediated
activation of, cell division and embryogenic cell formation but does not
influence cell cycle progression in alfalfa cell cultures. The Plant Journal,
43(6), 849-860.
Pagnussat, G. C., Simontacchi, M., Puntarulo, S., y Lamattina, L. (2002). Nitric
oxide is required for root organogenesis. Plant Physiology, 129(3), 954-956.
Panigrahi, J., Mistry, N. M., y Patel, I. C. (2013). Direct shoot organogenesis from
bulbs explants of Polianthes tuberosa cultivars (Prajwal and Shringar).
Indian Journal of Applied Research, 3(7), 73-75.
Panigrahi, J., Rana, V. D., y Patel, I. C. (2013). Multiple Shoot Regeneration of
Polianthes tuberosa cultivars Phulerajni and Calcutta double. Indian Journal
of Applied Research, 3(7), 12-14.
Portillo, L., y Santacruz-Ruvalcaba, F. (2006). Obtención de embrioides de Agave
tequilaza Weber a partir de explantes de raíz. Zonas Áridas(10), 11-19.
Portillo, L., Santacruz-Ruvalcaba, F., Gutiérrez-Mora, A., y Rodríguez-Garay, B.
(2007). Somatic embryogenesis in Agave tequilana Weber cultivar azul. In
Vitro Cellular & Developmental Biology-Plant, 43(6), 569-575.

110
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Ptak, A., y Bach, A. (2007). Somatic embryogenesis in tulip (Tulipa gesneriana L.)
flower stem cultures. In Vitro Cellular & Developmental Biology-Plant, 43(1),
35-39.
Quiroz-Figueroa, F. R., Rojas-Herrera, R., Galaz-Avalos, R. M., y Loyola-Vargas,
V. M. (2006). Embryo production through somatic embryogenesis can be
used to study cell differentiation in plants. Plant Cell, Tissue and Organ
Culture, 86(3), 285-301.
Raghuvanshi, S., Tripathi, M. K., Vidhya-Sankar, M., y Singh, O. P. (2013).
Establishment of low-cost effective protocol for massive in vitro propagation
in Polianthes tuberosa Linn. Plant Cell Biotechnology and Molecular Biology,
14(1y2), 49-59
Rajasekaran, V., Haripriya, K., y Shakila, A. (2000). In vitro propagation of
tuberose (Polianthes tuberosa L.). Paper presented at the Spices and
aromatic plants: challenges and opportunities in the new century.
Contributory papers. Centennial conference on spices and aromatic plants,
Calicut, Kerala, India, 20-23 September, 2000.
Rico-Lemus, M., y Rodríguez-Garay, B. (2014). SNP as an Effective Donor of Nitric
Oxide for in vitro Plant Cell and Tissue Culture. Journal of Plant
Biochemistry & Physiology.
Rodríguez-Garay, B., Gutiérrez-Mora, A., y Acosta-Duefias, B. (1996). Somatic
embryogenesis of Agave victoria-reginae Moore. Plant Cell, Tissue and
Organ Culture, 46(1), 85-87.
Rodríguez-Sahagún, A., Acevedo-Hernández, G., Rodríguez-Domínguez, J. M.,
Rodríguez-Garay, B., Cervantes-Martínez, J., y Castellanos-Hernández, O.
A. (2011). Effect of light quality and culture medium on somatic
embryogenesis of Agave tequilana Weber var. Azul. Plant Cell, Tissue and
Organ Culture (PCTOC), 104(2), 271-275.
Rőszer, T. (2014). Biosynthesis of Nitric Oxide in Plants in Nitric Oxide in Plants:
Metabolism and Role in Stress Physiology (17-32): Ed. Springer.

111
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
SAGARPA. (2014). Secretaria de Agricultura, Ganaderia, Desarrollo Rural, Pesca
y Alimentación. Obtenido de Cierre de la producción agrícola por cultivo:
http://www.siap.gob.mx/cierre-de-la-produccion-agricola-por-cultivo
Sangavai, C., y Chellapandi, P. (2008). In vitro Propagation of a Tuberose Plant
(Polianthes tuberosa L.). Electronic Journal of Biology, 4(3), 98-101.
Santacruz-Ruvalcaba, F., y Portillo, L. (2009). Thin cell suspension layer as a new
methodology for somatic embryogenesis in Agave tequilana Weber cultivar
azul. Industrial crops and products, 29(2), 609-614.
SEMARNAT. (2002). Norma Oficial Mexicana NOM-059-ECOL-2001, protección
ambiental-especies nativas de México de flora y fauna silvestres-categorías
de riesgo y especificaciones para su inclusión, exclusión o cambio-lista de
especies en riesgo. Diario Oficial, 6 de marzo del, 2002, 1-56.
SIAP. (2014). Servicio de Información Agroalimentaria y Pesquera. Obtenido de
Polianthes tuberosa: http://www.siap.gob.mx/nardo/
Singh, H. (1978). Embryology of gymnosperms in Zimmerman W., Carlquist Z.,
Ozenda P. y Wulff H.D. 187-241: Ed. Handbuch der Pflanzenanatomie.
Solano, C. (2000). Sistemática del género Polianthes L.(Agavaceae). Facultad de
Ciencias, Universidad Nacional Autónoma de México. Tesis Doctorado.
Solano, E., y Feria, T. P. (2007). Ecological niche modeling and geographic
distribution of the genus Polianthes L.(Agavaceae) in Mexico: using niche
modeling to improve assessments of risk status. Plant Conservation and
Biodiversity, 311-326.
Subczynski, W. K., Lomnicka, M., y Hyde, J. S. (1996). Permeability of nitric oxide
through lipid bilayer membranes. Free radical research, 24(5), 343-349.
Tan, B. C., Chin, C. F., y Alderson, P. (2013). Effects of sodium nitroprusside on
shoot multiplication and regeneration of Vanilla planifolia Andrews. In Vitro
Cellular & Developmental Biology-Plant, 49(5), 626-630.
Tewari, R. K., Prommer, J., y Watanabe, M. (2013). Endogenous nitric oxide
generation in protoplast chloroplasts. Plant Cell Reports, 32(1), 31-44.

112
Efecto del nitroprusiato de sodio en la regeneración
in vitro de un híbrido del género Polianthes
Thomas, D. D., Liu, X., Kantrow, S. P., y Lancaster, J. R. (2001). The biological
lifetime of nitric oxide: implications for the perivascular dynamics of NO and
O2. Proceedings of the National Academy of Sciences, 98(1), 355-360.
Tribulato, A., Remotti, P., Löffler, H., y Van Tuyl, J. (1997). Somatic embryogenesis
and plant regeneration in Lilium longiflorum Thunb. Plant Cell Reports,
17(2), 113-118.
Trueblood, E. W. E. (1973). “Omixochitl”—the tuberose (Polianthes tuberosa).
Economic Botany, 27(2), 157-174.
Valderrama, R., Corpas, F. J., Carreras, A., Fernández-Ocaña, A., Chaki, M.,
Luque, F., y Barroso, J. B. (2007). Nitrosative stress in plants. FEBS Letters,
581(3), 453-461.
von Arnold, S., Sabala, I., Bozhkov, P., Dyachok, J., y Filonova, L. (2002).
Developmental pathways of somatic embryogenesis. Plant Cell, Tissue and
Organ Culture, 69(3), 233-249.
Wang, Y., Luo, Z., y Du, R. (2015). Nitric oxide delays chlorophyll degradation and
enhances antioxidant activity in banana fruits after cold storage. Acta
Physiologiae Plantarum, 37(4), 1-10.
Wendehenne, D., Pugin, A., Klessig, D. F., y Durner, J. (2001). Nitric oxide:
comparative synthesis and signaling in animal and plant cells. Trends in
plant science, 6(4), 177-183.
Williams, E., y Maheswaran, G. (1986). Somatic embryogenesis: factors
influencing coordinated behaviour of cells as an embryogenic group. Annals
of Botany, 57(4), 443-462.
Xu, J., Yin, H., Wang, W., Mi, Q., y Liu, X. (2009). Effects of sodium nitroprusside
on callus induction and shoot regeneration in micropropagated Dioscorea
opposita. Plant Growth Regulation, 59(3), 279-285.
Yang, X., y Zhang, X. (2010). Regulation of somatic embryogenesis in higher
plants. Critical Reviews in Plant Science, 29(1), 36-57.


















