
Encapsulatie van cellen in tailor-made poreuze ......biomateriaal interacties worden geëvalueerd...
65
Encapsulatie van cellen in tailor-made poreuze polymeerscaffolds via bioplotten Evelien KIECKENS Verhandeling ingediend tot het verkrijgen van de graad van Master in de Biomedische Wetenschappen Promotor: Dr. DECLERCQ Heidi Begeleidster: BERNEEL Elke Vakgroep: Medische Basiswetenschappen Academiejaar 2010-2011
Transcript of Encapsulatie van cellen in tailor-made poreuze ......biomateriaal interacties worden geëvalueerd...

Encapsulatie van cellen in tailor-made poreuze
polymeerscaffolds via bioplotten
Evelien KIECKENS
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Dr. DECLERCQ Heidi
Begeleidster: BERNEEL Elke
Vakgroep: Medische Basiswetenschappen
Academiejaar 2010-2011


Encapsulatie van cellen in tailor-made poreuze
polymeerscaffolds via bioplotten
Evelien KIECKENS
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Dr. DECLERCQ Heidi
Begeleidster: BERNEEL Elke
Vakgroep: Medische Basiswetenschappen
Academiejaar 2010-2011

“De auteur en de promotor geven de toelating deze masterproef voor consultatie beschikbaar
te stellen en delen ervan te kopiëren voor persoonlijk gebruik. Elk ander gebruik valt onder
de beperkingen van het auteursrecht, in het bijzonder met betrekking tot de verplichting
uitdrukkelijk de bron te vermelden bij het aanhalen van resultaten uit deze masterproef.”
Datum
Evelien Kieckens Dr. Heidi Declercq

Voorwoord
Bij het verwezenlijken van deze masterproef werd ik bijgestaan door een aantal mensen die ik
in het bijzonder wil bedanken.
Ik zou graag als eerste Prof. Dr. Maria Cornelissen bedanken voor het ter beschikking stellen
van het labo en voor de kennis die ze me heeft bijgebracht. Zij heeft me geleerd dat je in de
wetenschap moet blijven proberen tot het lukt (ook al lukt het niet altijd).
Dr. Heidi Declercq, mijn promotor, wil ik graag bedanken om me in te wijden in de wondere
wereld van tissue engineering met scaffolds en om talloze keren mijn teksten na te lezen en te
verbeteren.
Een bijzonder woordje van dank gaat uit naar Elke Berneel, mijn begeleidster, voor het
beestig leuke onderwerp en voor de fijne tijd die we samen onder de flow hebben
doorgebracht. Ze heeft me de ‘know-how’ van het celcultuur labo bijgebracht en me ondanks
een druk en bewogen jaar zo goed mogelijk geholpen.
Johanna Aernoudt zou ik graag willen bedanken voor haar logistieke ondersteuning, het
uitoefenen van haar slachterscapaciteiten en haar luisterend oor.
Leen Pieters, onze levende encyclopedie van histologische kleuringen, bedank ik voor de
hulp, tips en anekdotes die de wachttijd tijdens de uren durende immuunkleuringen konden
verzachten.
Verder wil ik graag Elien, Karolien , Pamela, Veerle, Evi, Veronique, Julie, Toke, Sylvia en
Greet bedanken voor de gezellige tijd en de toffe samenwerking in het labo.
Ook Annelot en Wai Long, mijn teaching lab-maatjes en partners in crime , wil ik bedanken
voor de fijne tijd, pijnlijke voeten veroorzaakt door de flair schoenen en de liters warme
chocomelk met slagroom (in ikea tas) die we samen hebben mogen nuttigen.
Elisabeth Brabants, ‘carpoolmate’ maar vooral vriendin, bedankt voor de fijne autoritten en
middagpauzes die we steeds druk taterend doorbrachten.
Verder wil ik ook de PBM-groep bedanken, voor het leveren van het materiaal, alle
medewerkers van het animalarium om me de mogelijkheid te geven zelf menisci te leren
verwijderen en hechtingen te plaatsen.
Tenslotte gaat mijn allergrootste dank uit naar mijn ouders en mijn vriend Kevin, die op elk
moment voor me klaarstonden. Bedankt voor jullie onvoorwaardelijke steun en geduld in dit
drukke jaar.
Evelien

Inhoud
Lijst met gebruikte afkortingen .................................................................................................................
Samenvatting ........................................................................................................................................... 1
1 Inleiding .............................................................................................................................................. 2
1.1 Anatomie ................................................................................................................................. 2
1.2 Histologie ................................................................................................................................ 3
1.2.1 Extracellulaire matrix ........................................................................................................... 3
1.2.2 De cellen ............................................................................................................................... 4
1.2.2.1 Fibrochondrocyten ....................................................................................................... 4
1.2.2.2 Fibroblastachtigen ....................................................................................................... 5
1.2.2.3 Superficiële zone cellen ............................................................................................... 5
1.3 Functies van de meniscus ........................................................................................................ 5
1.4 Meniscusscheuren ................................................................................................................... 6
1.5 Huidige therapieën .................................................................................................................. 6
1.5.1 Meniscectomie ...................................................................................................................... 6
1.5.2 Meniscustransplantatie ......................................................................................................... 7
1.5.2.1 Bewaring van donormenisci ........................................................................................ 7
1.5.2.2 Vasthechting van de donormeniscus ........................................................................... 8
1.5.2.3 ‘Size-matching’ ........................................................................................................... 9
1.5.2.4 Complicaties ................................................................................................................ 9
1.5.3 Meniscus reconstructie met substituten ................................................................................ 9
1.5.3.1 Actifit TM
Orteq Bioengineering .................................................................................... 10
1.5.3.2 Menaflex TM
Collagen Meniscus Implant ...................................................................... 10
1.6 ‘Tissue engineering’ voor menisci ........................................................................................ 10
1.6.1 Plotten versus printen ......................................................................................................... 12
1.6.2 Mogelijke materialen .......................................................................................................... 12
1.6.3 ‘Cell free’ of ‘cell seeded’ scaffolds? ................................................................................. 14
1.6.4 Mogelijke celtypes voor meniscus ‘tissue engineering’ ..................................................... 14
1.6.4.1 Fibrochondrocyten ................................................................................................. 14
1.6.4.2 Mesenchymale stamcellen ..................................................................................... 14
1.6.4.3 Synoviale cellen........................................................................................................ 15
1.6.5 Theoretisch model voor bioplotten van menisci ................................................................. 15
1.6.6 Groeifactoren die meniscusherstel kunnen bevorderen ...................................................... 16
1.6.7 Gebruik van pelletculturen ................................................................................................. 16
1.7 Doel van de masterproef ........................................................................................................ 17

2 Materialen en methoden .................................................................................................................... 17
2.1 Isolatie van primaire fibrochondrocyten .................................................................................. 17
2.1.1 Isoleren van de menisci ...................................................................................................... 17
2.1.2 Vrijstellen van de cellen uit de ECM.................................................................................. 17
2.2 Pelletculturen van primaire fibrochondrocyten ........................................................................ 18
2.3 Encapsulatie van primaire fibrochondrocyten in hydrogels ..................................................... 18
2.3.1 Encapsulatie in gelatine-methacrylamide hydrogel (gel-MOD)......................................... 18
2.3.2 Encapsulatie in alginaat hydrogel ....................................................................................... 19
2.3.3 Encapsulatie in agarose ...................................................................................................... 20
2.3.4 Viabiliteit en proliferatie in hydrogels ............................................................................... 20
2.3.4.1 Viabiliteit en proliferatie in gel-MOD ........................................................................... 20
2.3.4.1.1 MTT analyse ........................................................................................................... 20
2.3.4.1.2 „Live/dead assay‟ .................................................................................................... 22
2.3.4.1.3 Histologie ............................................................................................................... 22
2.3.4.2 Viabiliteit en proliferatie in alginaat .............................................................................. 22
2.3.4.2.1 MTT analyse ........................................................................................................... 22
2.3.4.2.2 „Live/dead assay‟ .................................................................................................... 22
2.3.4.3 Viabiliteit in agarose...................................................................................................... 23
2.4 Celadhesie van HFF’s op 2D-coatings ..................................................................................... 23
2.5 Cultuur van primaire fibrochondrocyten op 3D-scaffolds ....................................................... 23
2.5.1 Uitzaaien op commerciële scaffolds ................................................................................... 23
2.5.1.1 Becton DickinsonTM
Three Dimensional Collagen Composite Scaffold ....................... 23
2.5.1.2 Sigma-Aldrich® 3D Biotek 3D Inserts PCL scaffoldsTM
.............................................. 24
2.5.2 Uitzaaien op geplotte scaffolds .......................................................................................... 24
2.5.3 Viabiliteit en proliferatie op scaffolds ................................................................................ 24
2.6 Histochemische karakterisatie .................................................................................................. 24
2.6.1 Haematoxyline eosine kleuring .......................................................................................... 24
2.6.2 Alciaanblauw kleuring........................................................................................................ 25
2.7 Statistische verwerking ............................................................................................................. 25
3 Resultaten .......................................................................................................................................... 25
3.1 Encapsulatie van primaire fibrochondrocyten in hydrogels ..................................................... 25
3.1.1 Encapsulatie in gel-MOD ................................................................................................... 25
3.1.2 Encapsulatie in alginaat ...................................................................................................... 26
3.1.3 Encapsulatie in agarose ...................................................................................................... 28
3.1.4 Statistische vergelijking tussen Gel-MOD en commerciële hydrogel (alginaat). .............. 29
3.2 Celadhesie van HFF’s op coatings ........................................................................................... 30

3.3 Primaire fibrochondrocyten op scaffolds ................................................................................. 33
3.3.1 Viabiliteit ............................................................................................................................ 33
3.3.2 Histologie ........................................................................................................................... 38
3.4 Pelletculturen van primaire fibrochondrocyten ........................................................................ 40
4 Discussie ............................................................................................................................................ 40
5 Conclusies ......................................................................................................................................... 48
6 Referenties ......................................................................................................................................... 48
Bijlagen ................................................................................................................................................... 1
Bijlage I: Samenstelling Ringeroplossing ........................................................................................... 1
Bijlage II: Samenstelling basis medium .............................................................................................. 1
Bijlage III: Samenstelling pronase (protease) medium ....................................................................... 1
Bijlage IV: Samenstelling collagenase medium .................................................................................. 1
Bijlage V: Samenstelling fibrochondrocyten medium ........................................................................ 1
Bijlage VI: Samenstelling basis medium en chondrogeen medium voor differentiatie ...................... 1
Bijlage VII: Samenstelling PBS .......................................................................................................... 2
Bijlage VIII: Samenstelling Dulbecco’s PBS ...................................................................................... 2
Bijlage IX: Samenstelling HFF medium ............................................................................................. 2
Bijlage X: Protocol Actine-cytoskelet focal adhesion kleuring .......................................................... 3
Bijlage XI: Samenstelling neutraal gebufferde formol........................................................................ 4
Bijlage XII: Haematoxyline eosine kleuring ....................................................................................... 4
Bijlage XIII: Protocol Alciaanblauw kleuring .................................................................................... 4

Lijst met gebruikte afkortingen
AA : ascorbinezuur
AEMA : 2-aminoethyl methacrylaat
AL : antilichaam
AD : gedestilleerd water
ATP : adenosinetrifosfaat
BD : Becton Dickinson
bFGF : ‘basic Fibroblast Growth Factor’
BMP : ‘bone morphogenic protein’
BSA : ‘bovine serum albumine’
Ca/PI : Calceïne AM/propidium iodide
CAD : ‘computer aided design’
CAM : ‘cell associated matrix’
CD34 : ‘cluster of differentiation 34’
CMI : ‘Collagen Menicus Implant’
COMP : ‘cartilage oligomeric matrix protein’
DAB : 3,3’-diaminobenzidine tetrahydrochloride
DAPI : ‘4',6-diamidino-2-phenylindole’
DMEM : ‘Dulbecco's Modified Eagle Medium’
DNA : deoxyribonucleïnezuur
DS : ‘degree of saturation’
ECM : extracellulaire matrix
eq : equivalent
F12 : ‘nutrient mixture F-12’
FC : fibrochondrocyten
FC medium : fibrochondrocyten medium
FCS : ‘Fetal Calf Serum’
FDA : ‘Food and drug administration’
FGF-2 : ‘fibroblast growth factor 2’
Fn : fibronectine
GAG's : glycosaminoglycanen
GelB : gelatine type B
gel-MOD : gelatine-methacrylamide
Gem. : gemiddelde

GFP : ‘Green Fluorescent Protein’
H0 : nulhypothese
HA : alternatieve hypothese
HBSS : ‘Hanks Buffered Salt Solution’
HE : haematoxyline eosine
HFF's : ‘Human Foreskin Fibroblasts’
HLA : humaan leukocyt antigeen
Ig : Irgacure 2959
IGF-1 : ‘insulin-like growth factor 1’
ITS : ‘insulin, transferrin, and sodium selenite’
MSC : Mesenchymale stamcellen
MTS : 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyfenyl)-2-(4-sulfofenyl)-
2H-tetrazolium
MTT : 3-(4,5-Dimethylthiazol-2-yl)-2,5-difenyltetrazolium bromide
NGS : ‘normal goat serum’
NRS : ‘normal rabbit serum’
NSS : ‘normal swine serum’
OA : osteoartritis
P : passage
P/S : Peniciline/Streptomycine
PBS : ‘phosphate buffered saline’
PCL : poly-ε-caprolacton
PDGFE : ‘platelet derived growth factor E’
PKC : proteïne kinase C
qRT-PCR : ‘quantitative Real Time-Polymerase Chain Reaction’
RGD : arginine-glycine-asparagine
rpm : ‘rounds per minute’
SD : standaarddeviatie
TB : trisbuffer
TGF-β1 : ‘transforming growth factor β1’
w/v : ‘weight/volume’
α-SMA : ‘α-smooth muscle actine’

1
Samenvatting
De menisci in de knie zijn 2 halvemaanvormige structuren die het gewicht dragen, de
schokken absorberen, het gewricht stabiliseren en vochtig houden. Scheuren in de menisci
vormen een belangrijk probleem omdat menisci een zo goed als onbestaande
regeneratiecapaciteit hebben. Vroeger was de meest gekozen behandeling een meniscectomie.
Men is echter tot het inzicht gekomen dat dit kan leiden tot osteoartritis en
kraakbeendegeneratie. Daarom zoekt men nu vooral naar meniscus-sparende alternatieven.
Hiertoe kan ‘tissue engineering’ met een belangrijke bijdrage leveren.
In deze masterproef wordt de cel-biomateriaal interactie bestudeerd van mogelijke nieuwe
materialen voor meniscus ‘tissue engineering’. Hier worden zowel hydrogels als scaffolds
gebruikt om fibrochondrocyten op uit te zaaien. De cytotoxiciteit, viabiliteit en cel-
biomateriaal interacties worden geëvalueerd aan de hand van MTT-analyses, actine cytoskelet
kleuringen, ‘live-dead assays’ en histologie.
Er wordt een vergelijkende studie gemaakt tussen inkapseling van primaire fibrochondrocyten
in gelatine-methacrylamide (gel-MOD) en commerciële hydrogels (alginaat en agarose) als
controle. De ingekapselde cellen worden opgevolgd in de tijd en verschillende analyses
gebeuren op dag 0, dag 1 en dag 3. Het viabiliteitspercentage voor cellen in alginaat (78,62 ±
1,67) ligt significant hoger (p<0,05) dan voor cellen in gel-MOD (8,54 ± 0,65).
Daarnaast wordt de proliferatie en differentiatie van fibrochondrocyten op verschillende
commercieel beschikbare scaffolds en niet-commerciële scaffolds, die geplot worden met de
bioplotter, bestudeerd. De celbezaaide scaffolds worden in verschillende media (basis,
chondrogeen of fibrochondrocyten (FC) medium) gekweekt. De viabiliteit, proliferatie en
differentiatie wordt bestudeerd op dag 7, 14 en 21. De scaffolds worden gekoloniseerd door
fibrochondrocyten in FC medium. De PCL-GelB Fn scaffolds geven betere resultaten dan de
PCL-O en de commerciële PCL scaffolds. In chondrogeen medium is het effect van de Fn
coating duidelijk zichtbaar. De differentiatie en extracellulaire matrix (ECM) productie op
scaffolds wordt vergeleken met pelletculturen en hydrogels en geëvalueerd op verschillende
tijdstippen. De pelletculturen in passage 0 vertonen een betere ECM vorming dan
pelletculturen in passage 2. Er is een duidelijke ECM productie op scaffolds in FC medium,
maar minder in chondrogeen medium. In hydrogels is weinig ECM productie te zien in
vergelijking met de pelletculturen in passage 0.

2
1 Inleiding
Er is een grote problematiek rond beschadiging en scheuren in menisci omdat niet alleen
ouderen hiermee geconfronteerd worden, maar ook jonge topsporters al te vaak getroffen
worden door dergelijk letsel. Jaarlijks worden er in Europa meer dan 400.000 operaties
uitgevoerd ten gevolge van een meniscusscheur. [1] Deze letsels hebben een grote socio-
economische impact. Bij jongere patiënten komen meniscusletsels vooral voor door
sportkwetsuren, bijvoorbeeld door het draaien van het bovenlichaam bij een gebogen knie. Bij
oudere patiënten zijn meniscusletsels vooral het gevolg van de vermindering van elasticiteit
van het kraakbeen. [2]
Menisci spelen een belangrijke rol in het goed functioneren van het kniegewricht en hebben
een dragende, krachtoverbrengende, schokabsorberende, gewrichtstabiliserende en smerende
functie. [3,4]
Het grootste probleem bij een meniscusletsel is het feit dat het fibrocartilago, waaruit de
meniscus is opgebouwd, niet of nauwelijks herstelt. [3] Dit is vooral te wijten aan de
afwezigheid van bloedvaten in het binnenste deel van de meniscus. Vroeger bestond de
behandeling uit een volledige of gedeeltelijke meniscectomie afhankelijk van de grootte van
het defect. Een meniscectomie heeft echter als groot nadeel dat er kraakbeendegeneratie en
osteoartritis (OA) kan ontstaan in het gewricht van de patiënt. Dit leidt tot ernstige pijn en
immobiliteit. [1] Omdat meniscustransplantatie haar beperkingen heeft, wordt in deze
masterproef gekeken naar mogelijkheden voor ‘tissue engineering’ van de meniscus.
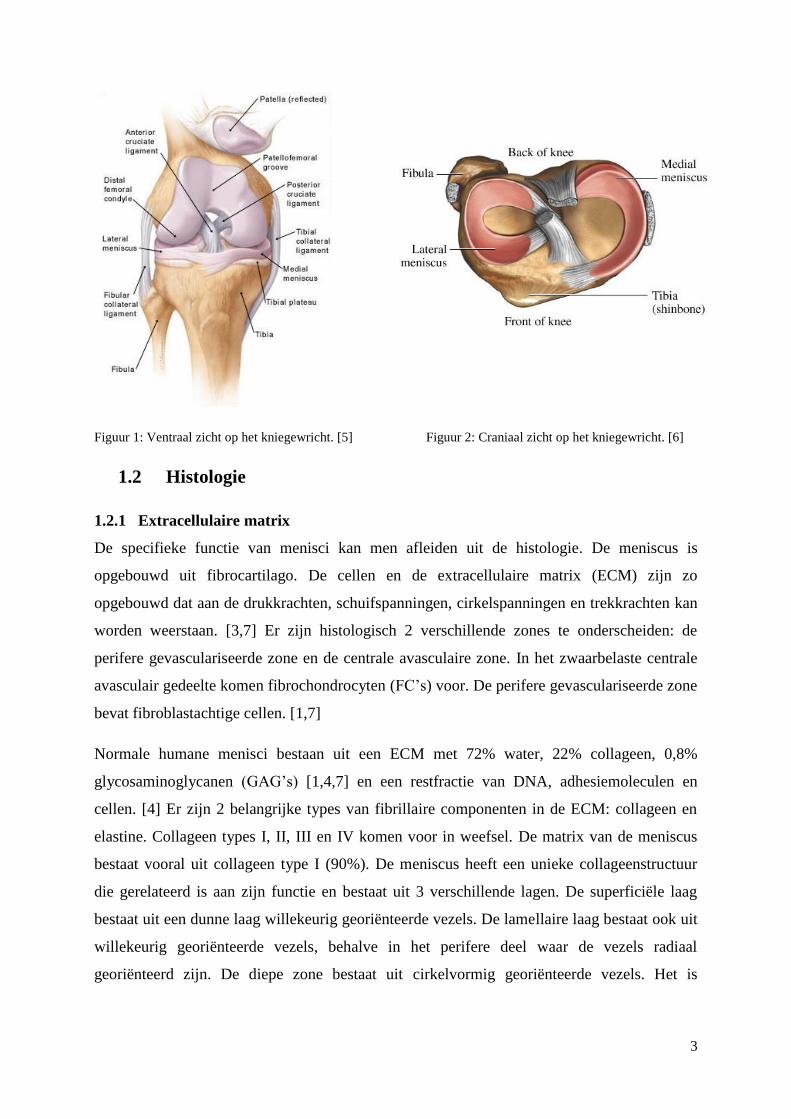
1.1 Anatomie
Elke knie bevat 2 halvemaanvormige schijven die menisci heten. Ze zijn gelegen tussen de
tibia en de femur. In elke knie zit een laterale en een mediale meniscus. De menisci zijn wit
en glanzend en de buitenzijde is bedekt door de synoviale membraan. De mate waarin de
meniscus gevasculariseerd is, verschilt naargelang de leeftijd van het individu. Een
kindermeniscus is volledig gevasculariseerd, terwijl een meniscus van een volwassene enkel
vascularisatie vertoont in het buitenste derde van het weefsel. De hoorns zijn de plaatsen die
het sterkste geïnnerveerd zijn binnen de meniscus. Het buitenste derde van de meniscus is ook
geïnnerveerd, daar kan men grote zenuwvezels die circumferentieel en kleinere vezels die
radiaal geschikt zijn observeren. Er komt geen innervatie voor in het meest centraal gelegen
deel. [3,4]
3
Figuur 1: Ventraal zicht op het kniegewricht. [5] Figuur 2: Craniaal zicht op het kniegewricht. [6]
1.2 Histologie
1.2.1 Extracellulaire matrix
De specifieke functie van menisci kan men afleiden uit de histologie. De meniscus is
opgebouwd uit fibrocartilago. De cellen en de extracellulaire matrix (ECM) zijn zo
opgebouwd dat aan de drukkrachten, schuifspanningen, cirkelspanningen en trekkrachten kan
worden weerstaan. [3,7] Er zijn histologisch 2 verschillende zones te onderscheiden: de
perifere gevasculariseerde zone en de centrale avasculaire zone. In het zwaarbelaste centrale
avasculair gedeelte komen fibrochondrocyten (FC’s) voor. De perifere gevasculariseerde zone
bevat fibroblastachtige cellen. [1,7]
Normale humane menisci bestaan uit een ECM met 72% water, 22% collageen, 0,8%
glycosaminoglycanen (GAG’s) [1,4,7] en een restfractie van DNA, adhesiemoleculen en
cellen. [4] Er zijn 2 belangrijke types van fibrillaire componenten in de ECM: collageen en
elastine. Collageen types I, II, III en IV komen voor in weefsel. De matrix van de meniscus
bestaat vooral uit collageen type I (90%). De meniscus heeft een unieke collageenstructuur
die gerelateerd is aan zijn functie en bestaat uit 3 verschillende lagen. De superficiële laag
bestaat uit een dunne laag willekeurig georiënteerde vezels. De lamellaire laag bestaat ook uit
willekeurig georiënteerde vezels, behalve in het perifere deel waar de vezels radiaal
georiënteerd zijn. De diepe zone bestaat uit cirkelvormig georiënteerde vezels. Het is

4
aangetoond dat de collageen hoeveelheid gesynthetiseerd door de fibrochondrocyten afneemt
met de veroudering van het individu. [4]
Figuur 3: Schikking van de collageenvezels in de meniscus. [4]
Proteoglycanen (GAG’s en aggrecan) zijn verantwoordelijk voor het behoud van een optimaal
visco-elastisch gedrag, de compressiestijfheid en de hydratatie van het weefsel. Door hun
negatieve lading trekken GAG’s water aan en zorgen zo voor een sterke hydratatie. Daarnaast
zorgen GAG’s en andere proteïnen aan het oppervlak er ook voor dat er een zachte,
wrijvingsloze beweging van de menisci over de gewrichtsoppervlakken van de tibia en de
femur mogelijk zijn. Aggrecan is het meest voorkomende proteoglycaan. [1,3]
De adhesiemoleculen zijn gedeeltelijk verantwoordelijk voor de binding van andere
matrixmoleculen en cellen. Er zijn momenteel 3 molecules van dit type geïdentificeerd in de
meniscus: collageen type IV, fibronectine en thrombospondine. [4]
1.2.2 De cellen
De cellen aanwezig in de meniscus kan men opdelen in 3 grote groepen: fibrochondrocyten,
fibroblastachtigen en superficiële zone cellen.
1.2.2.1 Fibrochondrocyten
Fibrochondrocyten komen voor in het binnenste deel van de meniscus. Ze zijn rond tot ovaal
van vorm en hebben een duidelijke cel geassocieerde matrix (CAM) met een intense
pericellulaire proteoglycaan kleuring. Ze synthetiseren een grote hoeveelheid fibreus
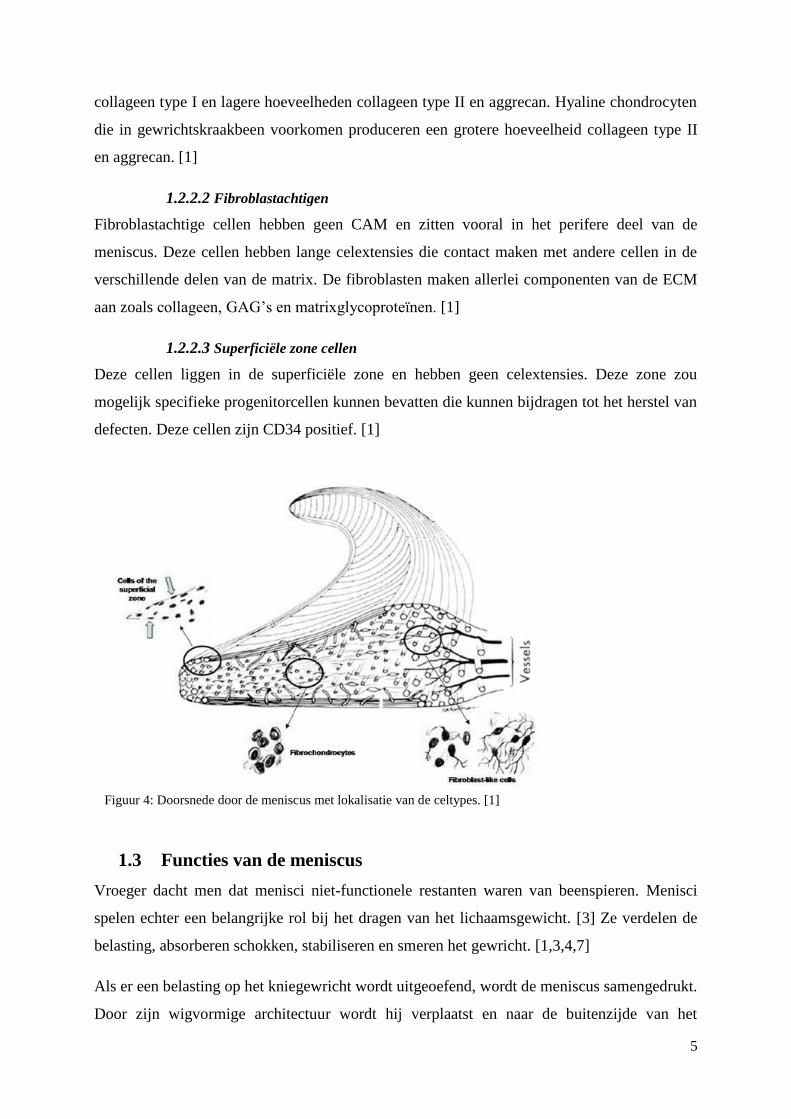
5
collageen type I en lagere hoeveelheden collageen type II en aggrecan. Hyaline chondrocyten
die in gewrichtskraakbeen voorkomen produceren een grotere hoeveelheid collageen type II
en aggrecan. [1]
1.2.2.2 Fibroblastachtigen
Fibroblastachtige cellen hebben geen CAM en zitten vooral in het perifere deel van de
meniscus. Deze cellen hebben lange celextensies die contact maken met andere cellen in de
verschillende delen van de matrix. De fibroblasten maken allerlei componenten van de ECM
aan zoals collageen, GAG’s en matrixglycoproteïnen. [1]
1.2.2.3 Superficiële zone cellen
Deze cellen liggen in de superficiële zone en hebben geen celextensies. Deze zone zou
mogelijk specifieke progenitorcellen kunnen bevatten die kunnen bijdragen tot het herstel van
defecten. Deze cellen zijn CD34 positief. [1]
1.3 Functies van de meniscus
Vroeger dacht men dat menisci niet-functionele restanten waren van beenspieren. Menisci
spelen echter een belangrijke rol bij het dragen van het lichaamsgewicht. [3] Ze verdelen de
belasting, absorberen schokken, stabiliseren en smeren het gewricht. [1,3,4,7]
Als er een belasting op het kniegewricht wordt uitgeoefend, wordt de meniscus samengedrukt.
Door zijn wigvormige architectuur wordt hij verplaatst en naar de buitenzijde van het
Figuur 4: Doorsnede door de meniscus met lokalisatie van de celtypes. [1]

6
gewricht gedrukt. Dit zorgt voor trekkrachten op de pezen waarmee de meniscus vastgehecht
zit aan het tibiaal plateau. In het algemeen wordt, tijdens compressie, de laterale meniscus
meer verplaatst dan de mediale. Het contactoppervlak is het grootst als de knie niet geplooid
is. Als de knie geplooid wordt, neemt het contactoppervlak af. Als extra kracht wordt
uitgeoefend zal de laterale meniscus de meeste belasting dragen, daar waar de mediale
meniscus de belasting splitst in 2 delen: 50% wordt door de mediale meniscus gedragen en
50% door het gewrichtskraakbeen. Als er een beschadiging van de meniscus is of de meniscus
verwijderd wordt, stijgt de druk op het tibiaal plateau. [4]
1.4 Meniscusscheuren
Meniscusscheuren komen zeer frequent voor, met een incidentie van ongeveer 61 per 100.000
personen per jaar. Bij jongere patiënten ziet men vooral scheuren als gevolg van
sportongevallen, door te draaien met een gebogen knie, vaak bij voetbal, rugby en skiën. Bij
oudere patiënten zijn meniscusscheuren vooral te wijten aan de degeneratie van fibreus
kraakbeen waardoor het minder elastisch wordt. [2]
Scheuren komen vooral voor in het avasculaire centrale gedeelte dat geen spontaan herstel
toelaat. In de jaren 60 van vorige eeuw dacht men dat de beste behandeling voor
meniscusscheuren het volledig verwijderen van de meniscus was. Bij een volledige
meniscectomie wordt het kniegewricht geopend met behulp van een lange incisie waarlangs
het beschadigde meniscuskraakbeen volledig wordt verwijderd. Op korte termijn gaf dit zeer
goede resultaten, maar op lange termijn begon men ontwikkeling van OA in het kniegewricht
te zien. Dit komt doordat na verwijdering van de meniscus het contactoppervlak tussen de
verschillende delen van het kniegewricht afneemt waardoor hogere spanningen en druk
ontstaan. Deze overmatige stress op de gewrichtsoppervlakken van de femur en de tibia leiden
tot schade aan het articulair kraakbeen, wat op zijn beurt leidt tot OA. Aangezien OA traag
ontwikkelt heeft het enkele tientallen jaren geduurd vooraleer men een correlatie zag tussen
meniscectomie en OA. [3]
1.5 Huidige therapieën
1.5.1 Meniscectomie
Vroeger ging men bij beschadiging van de meniscus vaak over tot een volledige
meniscectomie. Pas in de jaren 60 en 70 kwamen de nadelige gevolgen van deze behandeling
aan het licht. De patiënten kregen last van kraakbeendegeneratie en OA. [4]

7
Daarom is men overgeschakeld op een partiële meniscectomie, waarbij enkel het beschadigde
deel van de meniscus wordt verwijderd. Hiertoe heeft men met nieuwere toestellen een
techniek ontwikkeld die veel minder invasief is: de arthroscopische operatie waarbij men
gebruik maakt van een veel kleinere inscisie. Deze behandeling is enkel mogelijk indien het
defect niet te groot is. De kortetermijneffecten van deze behandeling zijn goed, maar over
langere periodes moet men er rekening mee houden dat het resterende deel van de meniscus
niet alle lasten zal kunnen blijven dragen en dus onvoldoende zijn functie zal kunnen
uitoefenen. Dit zal uiteindelijk ook leiden tot degradatie van het gewrichtskraakbeen, ernstige
pijn en verlies van functie van het gewricht. [1,4]
De beslissing om een meniscus te verwijderen dan wel de defecten te herstellen is afhankelijk
van verschillende klinische parameters, zoals de vorm en geometrie van de scheur, de plaats,
vascularisatie, grootte, stabiliteit, weefselbeschikbaarheid en kwaliteit. Leeftijd is niet
noodzakelijk een bepalende parameter, al dient men er wel rekening mee te houden dat met
toenemende leeftijd de kans op onherstelbare scheuren en een gebrek aan weefselviabiliteit
toeneemt. Dit leidt bij ouderen tot een groter aantal verwijderingen en minder herstellingen.
Daarnaast moet men ook rekening houden met de voorkeur van de patiënt. In het algemeen
gaat een verwijdering gepaard met een kortere herstelperiode, maar op lange termijn is
herstellen van de meniscus toch een meer aangewezen mogelijkheid. Men schakelt meer en
meer over naar herstel waar mogelijk. [2]
1.5.2 Meniscustransplantatie
In geval van sterk beschadigde menisci is een volledige meniscectomie een veel gebruikte
methode. Men heeft dan een plaatsvervangende meniscus nodig. Dit kan een allograft zijn
afkomstig van donoren, een meniscus substituut of in de toekomst een ‘tissue engineered’
weefselconstruct. Allograften van donoren zijn echter slechts beperkt beschikbaar. Een
belangrijk punt dat moet in acht genomen worden als men gebruik maakt van donorweefsel is
het verwerken en het bewaren van het materiaal. Men kan gebruik maken van verse ‘viabele’
allograften, diepgevroren allograften of gecryopreserveerde allograften die men op een
steriele manier isoleert. [1]
1.5.2.1 Bewaring van donormenisci
De verse allograften worden in een cultuurmedium bewaard tot aan de transplantatie. Een
nadeel van deze methode is dat het donormateriaal maximum 2 weken kan bewaard worden.
Het gebruik van deze allograften maakt dat men moet screenen naar microbiologische ziekten

8
die kunnen worden overgebracht naar de acceptor. Deze testen kunnen soms langer duren dan
de bewaartijd van een meniscus. Een oplossing voor dit probleem is het gebruik van
weefselbanken waarin het materiaal bevroren wordt bewaard. Hierdoor kan het
donormateriaal veel langer bewaard worden. In weefselbanken is het ook mogelijk om
menisci van verschillende afmetingen te stockeren waardoor er kan gezocht worden naar de
best passende meniscus voor de patiënt. [2]
Er zijn 2 mogelijke technieken om materiaal in te vriezen: cryopreservatie en versvriezen. Bij
cryopreservatie maakt men gebruik van een gecontroleerde invriessnelheid bij temperaturen
tot -196°C. Het materiaal wordt bewaard in een bad van cryoprotectans dat de cellen
beschermt tegen celdood door waterkristalvorming. Ondanks alles blijft slechts 10% van de
cellen metabolisch actief. De cellen die viabel blijven zitten meestal het dichtst bij het
cryoprotectans. Dit perifere deel van de meniscus wordt ook het snelste weer geherpopuleerd
na transplantatie in de acceptor. De diepere delen van het materiaal bevatten dus de minst
viabele cellen en deze delen worden dan ook nog eens traag geherpopuleerd. [2]
Bij versvriezen wordt het donormateriaal gewassen, verpakt en diepgevroren bij
-80°C. Dit is geen celsparende methode. Alvorens in te vriezen bestaan er verschillende
methodes voor sterilisatie zoals chemische decontaminatie of sterilisatie door bestraling met
γ-stralen. [2]
Over het effect van cryopreservatie op de microarchitectuur van collageenvezels bestaan
tegenstrijdige publicaties. De onderzoeksgroep van Gelber heeft in 2009 aangetoond dat
invriezen de microstructuur van het collageennetwerk niet behoudt, waar cryopreservatie dat
wel doet. [8,9] Het artikel van McDermott [2] gaat ervan uit dat cryopreservatie de
mechanische integriteit van de meniscus niet beter in stand houdt dan de versvries methode.
1.5.2.2 Vasthechting van de donormeniscus
De mechanische functie van de meniscus is volledig afhankelijk van de ultrastructurele
organisatie van het weefsel. De implantatie van een meniscus is dan ook een veeleisende
techniek. Bij de naaitechniek heeft men een stevige perifere rand nodig waaraan de allograft
kan worden vastgehecht. De huidige operatietechnieken blijven nog steeds verbeteren. Er zijn
nu ook specifieke instrumenten op de markt die toelaten om nog preciezer te werken. [2]

9
1.5.2.3 ‘Size-matching’
Een ander belangrijk punt is de ‘size-matching’ van een donormeniscus aan de acceptorknie.
Onderzoek heeft aangetoond dat een slechte positionering van een meniscus, als gevolg van
de verkeerde afmetingen, de functie van de graft aanzienlijk vermindert. Het is dus belangrijk
dat men kan beschikken over een ruime weefselbank met menisci van verschillende
afmetingen om zo de best passende meniscus uit te zoeken. Hiervoor kan ‘tissue engineering’
een oplossing bieden want de volledige structuur kan gemaakt worden op maat van de patiënt.
[2]
1.5.2.4 Complicaties
Complicaties na transplantatie zijn zeldzaam. De grootste zorg van de patiënten is het risico
van ziektetransmissie van het donorweefsel. De incidentie van contaminatie van het weefsel
met daaropvolgende infectie is zeldzaam omdat de weefselbanken een groot aantal
veiligheidsregels in acht moeten nemen. Het meniscusweefsel zelf is geïmmunoprivileerd
omdat de cellen in een dense mucopolysaccharide matrix ingebed zitten. Zo worden ze
beschermd tegen een mogelijke immuunrespons. Er zijn wel enkele zeldzame cases bekend
waarin patiënten antilichamen (AL) ontwikkelden tegen het humaan leukocyt antigeen (HLA)
van de donor. Maar er is slechts 1 geval van gefaalde transplantatie als gevolg van afstoting
bekend. [2]
1.5.3 Meniscus reconstructie met substituten
Men maakt reeds gebruik van implantaten om beschadigde menisci te herstellen. Twee
frequent gebruikte materialen zijn ActifitTM
van Orteq Bioengineering en MenaflexTM
‘Collagen Meniscus Implant’. [10,11] Het gebruik van deze substituten geeft goede resultaten
[12], maar toch zijn er belangrijke beperkingen aan verbonden. De scaffolds hebben vaste
afmetingen en moeten door de chirurg op maat gesneden worden, om zo goed mogelijk te
kunnen passen in het defect. Dit zal echter nooit zo accuraat kunnen gebeuren als met
beeldvorming en bioplotten. Daarnaast worden de scaffolds ook acellulair geïmplanteerd en
kan men de celgroei en celkolonisatie in vivo niet controleren.
De methode die gebruikt wordt bij patiënten bestaat uit verschillende stappen. Eerst wordt het
defect verwijderd en gemeten zodat men de scaffold zo goed mogelijk kan aanpassen naar
vorm en grootte. Vervolgens wordt de scaffold in de knie gebracht en vastgehecht aan de
resterende meniscusrand. [11,13]

10
1.5.3.1 Actifit TM
Orteq Bioengineering
De ActifitTM
scaffold van Orteq Bioengineering is een biodegradeerbare, poreuze,
synthetische scaffold, opgebouwd uit polycaprolacton (zachte component) en polyurethaan
(stijve component) die weefselingroei toelaten. Polycaprolacton is een degradeerbaar
polyester dat ook wordt gebruikt in implanteerbare biodegradeerbare ‘medical devices’ zoals
hechtingsdraden. Het polyurethaan zal langzaam degraderen waardoor putrescine vrijkomt.
Deze stof wordt ook geproduceerd door zoogdiercellen en bevordert celdeling. [10,11]
1.5.3.2 Menaflex TM
Collagen Meniscus Implant
De MenaflexTM
‘Collagen Meniscus Implant’ (CMI) is een resorbeerbare poreuze collageen-
glycosaminoglycaan matrix met een gedefinieerde geometrie, densiteit, thermische stabiliteit
en mechanische sterkte. De CMI is samengesteld uit 97% gezuiverd collageen type I,
afkomstig van de achillespezen van runderen en glycosaminoglycanen die chondroïtinesulfaat
en hyaluronzuur bevatten. Het collageen-GAG complex is chemisch gecrosslinkt om de in-
vivostabiliteit en de handelbaarheid te verbeteren. [12]
In vivo tests met proefdieren hebben aangetoond dat de scaffold, eens ingeplant, zich goed
vormt naar het gewricht. Er werden geen negatieve effecten gezien en het nieuw gegenereerde
weefsel had een histologisch uitzicht dat sterk op natief meniscusweefsel leek. Verder is
gebleken dat deze materialen biocompatibel zijn en weefselregeneratie ondersteunen. [12]
1.6 ‘Tissue engineering’ voor menisci
‘Tissue engineering’ is een domein dat verschillende disciplines uit de biologie, chemie en
ingenieurswetenschappen combineert. Het uiteindelijke doel is het verkrijgen van een
bioimplantaat om uitgevallen, beschadigde of slecht werkende organen of weefsels te
vervangen of te ondersteunen. [14]
Een schematisch overzicht van het ‘tissue engineering’ proces is weergegeven in figuur 5.
Een eerste luik is de medische beeldvorming: men kan het defect in de patiënt met behulp van
x-stralentomografie in beeld brengen en dan met software omzetten tot een 3D-
computermodel. Daarna bestaan er 2 verschillende benaderingen. Men kan een poreuze
scaffold plotten waarop men in de volgende stap (al dan niet patiënteigen) cellen zal uitzaaien
of men kan de cellen rechtsteeks opmengen in het biomateriaal en de cellen en het materiaal
samen plotten. Deze scaffolds brengt men dan in cultuur waarna men de 3D-structuur zal
implanteren. Daar zal het implantaat geïntegreerd worden in het betreffende weefsel en zijn

11
functie kunnen uitoefenen. Afhankelijk van de biostabiliteit en de degradatiesnelheid van het
gebruikte materiaal zal de scaffold sneller of minder snel worden geresorbeerd en vervangen
door lichaamseigen materiaal. Door te variëren in de samenstelling van materialen kan men
naast de degradatiesnelheid ook het cellulair fenotype beïnvloeden.
Figuur 5: Overzicht van het ‘tissue engineering’ proces.
De eerste stappen voor ‘tissue engineering’ in de orthopedie werden reeds gezet in 1988, toen
de onderzoeksgroep van Arnoczky probeerde om een fibrineklonter te gebruiken om de
meniscus hersteltechnieken te verbeteren. Dit idee was gebaseerd op de toen geldende kennis
van de aanwezigheid van groeifactoren en stimulerende cytokines in bloedklonters. Deze
factoren, aanwezig in de klonter, zouden het helingsproces kunnen versnellen net zoals dat
met wondheling gebeurt. Men dacht ook dat deze klonter, fungerend als scaffold, een brug
zou vormen tussen de gescheurde fragmenten. Hierop zouden cellen van het synovium en
andere cellen in de omgeving zich hechten om zo de beschadiging te herstellen. Hierna
volgden andere onderzoeken met permanente en biodegradeerbare synthetische materialen die
gebruikt werden als scaffolds. [15]
In 1992 rapporteerden Stone et al. voor het eerst het gebruik van een collageen scaffold om
beschadigde menisci te herstellen. Deze collageenscaffold liet ingroei van meniscuscellen toe

12
die konden deelnemen aan het helingsproces. Deze studies hebben geleid tot een ‘research’
explosie in het ‘tissue engineering’ domein van kraakbeen. [15]
‘Tissue engineering’ heeft een aantal belangrijke voordelen tegenover het gebruik van
allograften. Zo kan men, door het variëren met verschillende synthetische polymeren en hun
modificaties, polymeerconstructen maken die variëren in sterkte en degradatiesnelheid. Men
kan deze materialen ook naar wens vormgeven, eventueel met behulp van de bioplotter.
Indien nodig kan men complexe polymeren synthetiseren die geleidelijk aan groeifactoren
vrijzetten om zo de adhesie aan de scaffold en differentiatie te bevorderen. Daarnaast heeft
men geen probleem met donor-acceptor compatibiliteit of tekort aan donoren omdat men
patiënteigen cellen kan uitzaaien op de scaffolds. [15] Verder biedt ‘tissue engineering’ ook
de mogelijkheid om een meniscus te maken die bestaat uit gelatineuze kern die wordt
verstevigd met een stevige poreuze scaffold. Zo kan men tot een construct komen dat een
maximale gelijkenis vertoont met een natuurlijke meniscus.
‘Tissue engineering’ kan een onderdeel worden van de nieuwe behandeling van patiënten met
defecte menisci. Het belangrijkste voordeel van ‘tissue engineering’ is de mogelijkheid om
een volledig functionerend weefsel te maken op maat van de patiënt. Daar waar
voorgevormde commercieel beschikbare scaffolds steeds op maat moeten aangepast worden
door de behandelende arts, kan men met ‘tissue engineering’ een meer ingewikkeld construct
met verschillende materialen en verschillende celtypes op maat maken.
1.6.1 Plotten versus printen
In de literatuur worden vaak de begrippen plotten en printen door elkaar gebruikt. Toch is er
een belangrijk onderscheid. Bij het 3D-bioprinting proces maakt men gebruik van
poederdeeltjes die met een bindmiddel of solvent aan elkaar gelinkt worden. Het niet
gebonden poeder wordt weggeblazen door een luchtstroom. De 3D-bioplotter procedure
maakt gebruik van een visceus plotmateriaal dat geplot wordt in een medium of in lucht. [16]
1.6.2 Mogelijke materialen
Een ideale scaffold moet biocompatibel en biodegradeerbaar zijn op lange termijn. Hij moet
een degradatieprofiel hebben dat ingroei van nieuw weefsel en hermodellering van deze
weefsels onder invloed van belasting toelaat. De degradatie mag niet gebeuren alvorens de
scaffold volledig is opgevuld met nieuw weefsel om ‘instorten’ van de structuur te vermijden.
Daarnaast moet de scaffold onbeperkte cellulaire ingroei en vrije diffusie van nutriënten

13
toelaten. Voor cel ingroei zijn intergeconnecteerde macroporiën met een diameter van
155-355 µm nodig. De scaffold kan ook gebruikt worden als drager voor stimulerende of
inhiberende groeifactoren en moet sterk genoeg zijn om te weerstaan aan de mechanische
belasting in het gewricht. Ook moet de scaffold een optimale treksterkte hebben, om te
voorkomen dat hij zal doorscheuren bij het vastnaaien aan het weefsel. [3]
Veel scaffoldmaterialen zijn al onderzocht voor de applicatie bij ‘tissue engineering’ van
fibrocartilago. Scaffolds kan men opdelen in natuurlijke en synthetische scaffolds. Men heeft
4 types natuurlijke scaffolds uitgeprobeerd voor de ‘tissue engineering’ van menisci. Deze
bestaan uit collageen, dunne darm submucosa, periost weefsel of perichondraal weefsel. Van
deze 4 toonde onderzoek met periost weefsel de minst goede resultaten. De meest
veelbelovende resultaten werden verkregen uit experimenten met collageen scaffolds. Een
belangrijk nadeel van deze natuurlijke weefsels is dat de initiële mechanische eigenschappen
en de poriegrootte niet kunnen gevarieerd worden. [4]
Geïsoleerde weefselcomponenten zoals collageen, proteoglycanen of elastinemoleculen
kunnen gebruikt worden om een op maat gemaakte scaffold met optimale 3D-structuur op te
bouwen. Populaire scaffolds voor het onderzoek naar kraakbeen ‘tissue engineering’ zijn o.a.
fibrine, polyglycolzuur, alginaat en collageen type I en II scaffolds. Een groot nadeel van de
collageen en alginaat scaffolds zijn de initieel zwakke mechanische eigenschappen. Dit kan
leiden tot een ongecontroleerd krimpen van de scaffolds in cultuur. [3]
Een andere mogelijkheid is het gebruik van een volledig synthetische polymeergebaseerde
scaffold. Vaak zijn dit polyesters omdat deze degradeerbaar zijn door graduele hydrolyse.
Ook copolymeren op basis van polyglycolzuur, polymelkzuur en polyurethaan worden
gebruikt. Een groot voordeel hierbij is dat de porositeit, de degradatiesnelheid en de
mechanische eigenschappen kunnen aangepast worden aan de noden. De mechanische
eigenschappen kunnen gecontroleerd worden door de graad van poreusheid. Zo zal een
materiaal met een grotere poreusheid - theoretisch - sneller degraderen door hydrolyse omdat
de totale vrije oppervlakte onderhevig aan oppervlakte-erosie groter is. [3]
Er bestaat een grotere variëteit aan synthetische scaffolds, maar nog geen enkele is al zover
gevorderd in klinische trials als de collageen scaffold. Er werd al onderzoek gedaan met
synthetische scaffolds bestaande uit teflon, koolstofvezels, samenstellingen van
koolstofvezels en polyurethanen, zuivere polyurethanen, koolstofvezels en polyesters,
polyurethaan-polylactide en caprolacton samenstellingen. [4]

14
1.6.3 ‘Cell free’ of ‘cell seeded’ scaffolds?
Men kan voor implantatie zowel een celvrije als een celbezaaide scaffold gebruiken.
Onderzoek bij diermodellen heeft aangetoond dat men zelfs bij het implanteren van
acellulaire scaffolds nog voldoende kolonisatie kan verkrijgen om de functie van de meniscus
te onderhouden. [8] Men heeft aangetoond dat repopulatie van acellulaire implantaten gebeurt
door cellen afkomstig van synoviaal weefsel en cellen uit het parameniscaal bindweefsel. [17]
Studies op humane diepgevroren allograften tonen echter een beperkte en superficiële
herpopulatie van het implantaat door gastheercellen. De kern van de graft blijft acellulair. [1]
Op basis van deze bevindingen zou men meer geneigd zijn om te kiezen voor de celvrije
scaffolds, omdat deze eenvoudig kunnen geïmplanteerd worden zonder dat er tijd nodig is om
cellen uit te zaaien. Toch hebben de celbezaaide scaffolds belangrijke voordelen. Zo kan men
met behulp van de bioplotter verschillende celtypes op verschillende plaatsen plotten. Men is
ook zeker van een goede celinfiltratie voor de implantatie.
1.6.4 Mogelijke celtypes voor meniscus ‘tissue engineering’
1.6.4.1 Fibrochondrocyten
De cellen uit het binnenste avasculaire deel zijn fibrochondrocyten. Ze produceren meer
GAG’s als ze in cultuur gebracht worden dan de cellen uit het perifere deel. Dit heeft
implicaties voor het gebruik van autologe cellen voor ‘tissue engineering’. Men kan
verwachten dat alleen de cellen vanuit het binnenste gescheurde deel beschikbaar zijn. Dit
celaantal is echter zeer laag en kan nog verminderd zijn door het trauma. Daarom kan men er
sterk aan twijfelen of deze cellen wel zullen prolifereren en differentiëren in het juiste
fenotype. [3]
Fibrochondrocyten kunnen prolifereren in een driedimensionele matrix in vitro. In deze
omstandigheden exprimeren ze hetzelfde fenotype als in hun originele matrix. In klinische
settings worden deze autologe fibrochondrocyten geoogst door biopsie en geïsoleerd.
Omwille van hun geringe beschikbaarheid kunnen we stellen dat deze bron van cellen niet de
meest optimale is voor ‘tissue engineering’. [1]
1.6.4.2 Mesenchymale stamcellen
Waarschijnlijk zullen mesenchymale stamcellen (MSC) een betere bron zijn voor
meniscusherstel. Deze MSC kunnen, afhankelijk van het cultuurmedium en de
cultuuromstandigheden, in vitro differentiëren tot verschillende mesenchymale weefsels zoals

15
bot, kraakbeen, vet, zenuwweefsel, spier en beenmerg. Hierbij wordt gebruik gemaakt van
heel dense celculturen die men micropellets noemt. Er is aangetoond dat MSC, als ze in
micropellets gekweekt worden in medium met dexamethasone en ‘Transforming Growth
Factor β’ (TGF-β), zullen differentiëren in (fibro)chondrogene lineage. [1] Ook andere
groeifactoren die de synthese van de ECM stimuleren en de afbraak inhiberen, kunnen heel
nuttig zijn. Zo kan TGF-β1, de groeifactor die betrokken is bij de ontwikkeling van bot en
kraakbeen, chondrogenese induceren. Onderzoek heeft aangetoond dat ‘Platelet-Derived
Growth Factor E’ (PDGFE) betrokken is bij de shift van een chondrogeen naar een
meniscusachtig celtype. ‘Insulin-like growth factor I’ (IGF-I) is de belangrijkste anabole
groeifactor van gewrichtskraakbeen onder normale omstandigheden, ook deze factor zou een
rol kunnen spelen. ‘Fibroblast Growth Factor 2’ (FGF2) lijkt een sterke modulator voor
stamcel proliferatie. Hij houdt de stromale beenmergfractie immatuur. ‘Bone Morphogenic
Protein 6’ (BMP-6) lijkt chondrogenese te stimuleren in een subpopulatie van
beenmergcellen. [1,3]
1.6.4.3 Synoviale cellen
Een andere potentiële celbron is het synovium van gewrichten. Men heeft gezien dat
fibroblasten die op acellulaire scaffolds groeien in een gewricht, onder invloed van belasting
differentiëren tot fibrocartilagoachtig weefsel. [3]
1.6.5 Theoretisch model voor bioplotten van menisci
Figuur 6: Schematisch overzicht van een doorsnede door de geplotte meniscus.

16
Voor een meniscus kan men best inkapseling en plotten combineren omdat men een
gelatineuze kern nodig heeft, de hydrogel, die verstevigd wordt met een poreuze scaffold. In
deze masterproef worden zowel rigidere scaffolds als gelachtige hydrogels onderzocht. In
bovenstaand model voor ‘tissue engineering’ worden centraal fibrochondrocyten en perifeer
fibroblastachtige cellen gebruikt.
1.6.6 Groeifactoren die meniscusherstel kunnen bevorderen
Groeifactoren werken in op targetcellen door binding op een specifieke receptor. Dit signaal
triggert de activatie van genen die celproliferatie, celdifferentiatie en celdood reguleren.
Insuline groeifactoren zoals IGF-1 stimuleren de synthese van proteoglycanen, collageen type
II en integrines. Ze inhiberen ook de destructie van de ECM en stimuleren de
fibrochondrocyten om contact te maken met fibronectine en collageen type II. TGF-β1
induceert ook de synthese van specifieke proteoglycanen en collageen type II door
mesenchymale cellen. FGF-2 is een krachtig mitogeen dat in combinatie met TGF-β1 een
efficiënte expansie toelaat van de gewrichtskraakbeencellen, terwijl hun mogelijkheid tot
differentiatie bewaard blijft. BMP-2 werkt stimulerend op de anabole activiteit in normale en
osteoartritische cellen. [18]
1.6.7 Gebruik van pelletculturen
Pelletculturen worden gebruikt als controle om het behoud van het fenotype aan te tonen.
Gebaseerd op embryonale chondrogenese hebben onderzoekers vooropgesteld dat het
mogelijk is om kraakbeen aan te maken door chondrocyten in pelletculturen te kweken. Het
chondrocytfenotype kan gekarakteriseerd worden door het aantonen van de synthese van
collageen type II en aggrecan. [19]
De fenotypische instabiliteit van chondrocyten in een conventionele monolayercultuur is een
grote uitdaging geweest voor kraakbeen ‘engineering’. In een monolayercultuur ontwikkelen
chondrocyten een fibroblastachtige morfologie en secreteren collageen type I maar verliezen
de expressie van collageen type II en aggrecan. Het is cruciaal om deze dedifferentiatie te
vermijden tijdens het proces van kraakbeen aanmaak. De celdensiteit is een kritische
parameter die het chondrocytfenotype kan stabiliseren. Bij hogere densiteiten (4x105
cellen/cm2) verandert het fenotype niet. [19] Ook fibrochondrocyten vertonen dedifferentiatie
in een monolayercultuur. Ze vertonen een gedaalde expressie van collageen type II, ‘cartilage
oligomeric matrix protein’ (COMP) en aggrecan. [20]

17
1.7 Doel van de masterproef
In deze masterproef wordt een antwoord gezocht op de volgende vragen:
- Zijn primaire fibrochondrocyten na inkapseling in hydrogels nog voldoende viabel?
- Kunnen mature fibrochondrocyten een 3D-scaffold voldoende koloniseren?
- Kunnen fibrochondrocyten in pelletculturen, hydrogels en scaffolds differentiëren tot
fibrocartilago?
2 Materialen en methoden
2.1 Isolatie van primaire fibrochondrocyten
2.1.1 Isoleren van de menisci
Voor de isolatie van primaire fibrochondrocyten worden menisci afkomstig van achterpoten
van varkens (Sus scrofa), die maximaal 24 uur voor het experiment geslacht waren, gebruikt.
De huid en overtollig spierweefsel worden verwijderd zonder het gewrichtskapsel open te
snijden. Daarna wordt alles ontsmet met 70% alcohol en wordt het gewrichtskapsel onder
steriele omstandigheden geopend. De menisci worden verwijderd en enkele seconden
ondergedompeld in 70% alcohol, om ze vervolgens over te brengen in een Ringeroplossing
(bijlage I) met dubbelgeconcentreerd peniciline/streptomycine (P/S). Hierin blijven ze
gedurende 30 minuten staan om ze uiteindelijk over te brengen naar basis medium (bijlage II).
De volledige menisci worden overnacht bewaard in het medium bij 37°C.
2.1.2 Vrijstellen van de cellen uit de ECM
Het binnenste 2/3 van de meniscus wordt losgesneden. Dit centrale deel wordt verder
gebruikt. Eerst worden de menisci overgebracht in petrischalen met een kleine hoeveelheid
medium. In die petrischalen worden ze mechanisch verkleind met behulp van een scalpel tot
stukjes van ongeveer 2 mm x 2 mm x 2 mm. De volgende stap is de enzymatische
behandeling met protease (60 mg Protease type XIV van Streptomyces griseus, Sigma-
Aldrich® in 20 ml protease medium (bijlage III) per poot) gedurende 2 uur bij 37°C. Hierna
wordt het medium verwijderd en volgt een wasstap met DMEM/F12 (Gibco®) gedurende 10
minuten bij 37°C. Het DMEM/F12 medium wordt verwijderd en dan volgt een tweede
enzymatische behandeling met collagenase (80 mg Collagenase type II van Clostridium

18
histolyticum, Sigma-Aldrich® in 40 ml collagenase medium (bijlage IV) per poot) gedurende
3 uur bij 37°C. Daarna wordt 5 ml FCS (Gibco®) toegevoegd.
De overgebleven celsuspensie wordt door een Becton Dickinson (BD)TM
Falcon cellstrainer
gebracht om zoveel mogelijk ‘single cells’ te bekomen. Deze cellen worden gecentrifugeerd
(10 min, 1000 ‘rounds per minute’(rpm)). Dan wordt het supernatans verwijderd en de pellet
wordt geresuspendeerd in fibrochondrocyten medium (FC medium) (bijlage V). De bekomen
fibrochondrocyten worden geteld met een Bürker telkamer.
2.2 Pelletculturen van primaire fibrochondrocyten
Van primaire fibrochondrocyten (passage 0 (P0) en passage 2 (P2)) worden pelletculturen
opgestart om de differentiatie na te gaan in basis medium (controle) en chondrogeen medium.
Hiervoor worden pellets gemaakt van 250.000 fibrochondrocyten. De benodigde hoeveelheid
cellen wordt geresuspendeerd in 1 ml basis medium. Dan worden de cellen gecentrifugeerd (5
min, 1000 rpm). Hierna wordt het supernatans verwijderd. De pellets die zullen dienen als
negatieve controle worden geresuspendeerd in 0,5 ml basis medium (bijlage VI). De pellets
die als positieve controle zullen dienen, worden in chondrogeen medium (bijlage VI)
geresuspendeerd. Hierna worden de cellen nog eens gecentrifugeerd (5 min, 1000 rpm). De
pellets worden opgevolgd gedurende 28 dagen en het medium wordt 3 keer per week ververst.
Op de gekozen tijdstippen worden de pellets gefixeerd en gekleurd. Fixatie gebeurt overnacht
met 4% paraformaldehyde (Merck) in PBS (bijlage VII).
2.3 Encapsulatie van primaire fibrochondrocyten in hydrogels
Een eerste deel van het praktisch werk bestaat uit het inkapselen van primaire
fibrochondrocyten in gel-MOD, alginaat en agarose hydrogels. Alginaat en agarose zijn gels
die vaak als referentieculturen gebruikt worden.
2.3.1 Encapsulatie in gelatine-methacrylamide hydrogel (gel-MOD)
Gelatine is een biopolymeer dat afgeleid is van collageen door hydrolytische degradatie. De
gebruikte gelatine wordt gemodificeerd met methacrylamide om crosslinking via een foto-
initiator en uv-licht mogelijk te maken. [21] Voor de inkapseling van fibrochondrocyten in
gel-MOD (1eq 10% weight/volume (w/v)) (‘Polymer Chemistry and Biomaterials Group’-
Universiteit Gent) wordt telkens een concentratie van 300.000 cellen/well (24-well plaat)
gebruikt.

19
De Irgacure 2959 (Ig) foto-initiator oplossing wordt buiten de flow aangemaakt en daarna
gefilterd m.b.v. een 0.22 µm Millex filter (MilliporeTM
). Hiervoor wordt eerst 40 mg Ig
afgewogen en aan 5 ml steriel water toegevoegd. De oplossing wordt overgebracht in een
kolfje dat wordt ingepakt in aluminiumfolie om de foto-initiator te beschermen tegen licht.
Dan wordt het kolfje in een warmwaterbad van 60°C geplaatst en geroerd met een magneet.
Vervolgens wordt de gel-MOD opgelost. Hiervoor wordt 1 g steriele gel-MOD onder steriele
omstandigheden opgelost in 5 ml PBS in een kolfje met stop. Het kolfje wordt verwarmd in
een warmwaterbad bij 40°C en geroerd. Bij 1 g gel-MOD met 67% vernetbare zijgroepen
(DS) moet 0,143 ml Ig toegevoegd worden. De oplossing wordt dan ontgast m.b.v. een
ontgaspompje (KNF laboport) gedurende 5 minuten en telkens als de oplossing begint te
borrelen wordt het vacuüm verwijderd.
De bekomen gel-MOD oplossing (1eq) is een 20% (w/v) oplossing. Om te komen tot een 10%
(w/v) gel-MOD oplossing (1eq) moet er een gelijke hoeveelheid celsuspensie aan een
bepaalde hoeveelheid gel-MOD met Ig worden toegevoegd. Aan de celsuspensie moet
eenzelfde hoeveelheid gel-MOD worden toegevoegd. De celsuspensie wordt afgedraaid en de
pellet wordt geresuspendeerd in een gelijke hoeveelheid PBS. Dit mengsel wordt goed
geroerd in een warmwaterbad bij 37°C onder de flow. Hiervan wordt telkens 200 µl in een
48-well suspensie plaat gepipetteerd. Als controle worden fibrochondrocyten in suspensie
gebruikt (300.000 cellen/well). De well platen met gels worden eerst gedurende 30 minuten in
de koelkast geplaatst en vervolgens 30 minuten onder een uv-lamp (intensiteit 8 mW/cm2)
gezet op een afstand van 10 cm. De plaat die de gels zonder uv bevat wordt niet onder de uv-
lamp geplaatst. Daarna wordt aan elke well 600 µl FC medium toegevoegd.
2.3.2 Encapsulatie in alginaat hydrogel
De inkapseling van cellen in alginaat (Low viscosity alginate van Macrocystis pyrifera,
Sigma-Aldrich®) gebeurt met een eindconcentratie van 1% alginaat en 5x106 cellen/ml.
Hiervoor wordt 0,5 g alginaat in 25 ml HBSS (Gibco®) opgelost bij 37°C (2 g/100 ml). De
alginaatoplossing wordt dan doorgespoten door een filter. Een gelijk volume van de
celsuspensie (3.000.000 cellen/ml HBSS) wordt toegevoegd aan eenzelfde volume van de
alginaatoplossing.
De alginaat beads worden gevormd door de alginaatoplossing met behulp van 10 µl
Gilsontips in een 102 mM CaCl2 (in gedestilleerd water (AD)) oplossing te druppelen. De
beads laat men dan gedurende 10 minuten bij kamertemperatuur polymeriseren. Hierna

20
worden de beads 2 keer gewassen in 0,9% steriele NaCl en in AD overgebracht in een 6-well
plaat aan een concentratie van 20 beads per well. Dit komt overeen met 300.000 cellen per
well.
Er worden wells gemaakt voor MTT op alginaatbeads, wells voor calceïne/propidium iodide
(Ca/PI) en wells voor coupes, wells met cellen in suspensie en wells met cellen die met Na-
citraat behandeld worden. Dit wordt verder beschreven op pagina 23.
2.3.3 Encapsulatie in agarose
Cellen worden ingekapseld aan een celdensiteit van 1,5 miljoen cellen/ml. Eerst wordt 0,8 g
agarose (type IX, Sigma-Aldrich®, A5030-10G) opgelost in 20 ml AD. Deze oplossing wordt
vervolgens opgekookt in de microgolf. De gewenste hoeveelheid cellen wordt opgelost in
dubbelgeconcentreerd DMEM medium. Hiervoor wordt 1,338 g DMEM poeder (Gibco®, art°
32500-035) en 0,37 g NaHCO3 poeder (Merck, Pro Analysi, art° 6329) opgelost in 50 ml AD.
Deze oplossing wordt doorgespoten. Vervolgens wordt aan 40 ml van de oplossing 10 ml
FCS en 100 µg/ml ascorbinezuur (L-Ascorbic acid 2-phosphate sesquimagnesium salt,
Sigma-Aldrich®, A8960-5G) toegevoegd. Dan wordt een gelijk volume van de opgeloste
agarose gemengd met een gelijk volume van de celsuspensie in dubbelgeconcentreerd DMEM
medium. Er wordt 0,5 ml van deze oplossing per well in een 24-well plaat gevoegd. Dan laat
men de oplossing gelifiëren bij 4°C gedurende 30 minuten. Hierna wordt basis, chondrogeen
of FC medium toegevoegd, dat 3 keer per week wordt ververst.
2.3.4 Viabiliteit en proliferatie in hydrogels
2.3.4.1 Viabiliteit en proliferatie in gel-MOD
2.3.4.1.1 MTT analyse
Op dag 0 voert men een MTT-test uit om een idee te krijgen van de viabiliteit van niet-
ingekapselde cellen. Hiervoor worden 300.000 cellen gebruikt. De benodigde celsuspensie
wordt in steriele eppendorfs gebracht en gecentrifugeerd (5 min, 1000 rpm). Het supernatans
wordt verwijderd en 500 µl MTT mengsel (5 mg MTT poeder (Calbiochem, N°475989)
opgelost in 1 ml steriel water en 9 ml FC medium (bijlage V)) wordt toegevoegd. De cellen
worden dan gedurende 4 uur bij 37°C op een schudplaat in het donker geplaatst. Hierna wordt
gecentrifugeerd en wordt het MTT mengsel verwijderd. Er wordt 500 µl lysebuffer
toegevoegd (100 µl Triton X100 (Fluka Analytical, reduced form) in 900 µl steriel water en
9 ml isopropanol, met 0,04 M HCl) en gedurende 30 minuten bij 37°C op de schudplaat in het

21
donker gezet. Daarna wordt 200 µl in een 96-well plaat overgebracht en wordt de absorbantie
bepaald met behulp van een ‘plate absorbance reader’ (EL 800, Biotec Instruments).
Op dag 1 en dag 3 worden eerst de uv-bestraalde gels en de niet-uv bestraalde gels
enzymatisch behandeld. Deze behandeling is nodig om de cellen vrij te stellen uit de gels. Ter
controle worden ook cellen in suspensie enzymatisch behandeld.
De uv-bestraalde gels worden uit de 48-well plaat gehaald en in fragmentjes gesneden. Deze
fragmenten worden dan in een 24-well plaat overgebracht in 0,5 ml collagenase oplossing
(Collagenase type IA van Clostridum histolyticum, Sigma-Aldrich) (16 mg collagenase in
10 ml warm basis medium (bijlage II)). Het achtergebleven medium wordt in eppendorfs
gebracht en afgedraaid (1000 rpm, 5 min). Dan wordt het supernatans verwijderd en wordt de
pellet geresuspendeerd in 0,5 ml collagenase oplossing. Deze celsuspensie wordt dan
toegevoegd aan de corresponderende gelfragmenten in de 24-well plaat.
De niet-uv bestraalde gels worden gebruikt als controle voor het effect van uv-belichting op
de cellen. Deze gels zijn visceus maar minder stevig en dienen daarom niet gesneden te
worden. Ze kunnen rechtstreeks in de 24-well plaat gebracht worden. Het bovenstaand
medium wordt in eppendorfs gebracht en afgedraaid (1000 rpm, 5 min). De pellet wordt
geresuspendeerd in 1 ml collagenase oplossing en toegevoegd aan de niet-uv bestraalde gels
in een 24-well plaat.
De cellen in suspensie dienen als controle voor het effect van collagenase op de
fibrochondrocyten. Deze celsuspensie wordt in eppendorfs gebracht en afgedraaid (1000 rpm,
5 min). Het supernatans wordt verwijderd en de cellen worden geresuspendeerd in 1 ml
collagenase oplossing. Dit wordt dan overgebracht in een 24-well plaat.
De cellen in suspensie die niet met enzym worden behandeld, worden gebruikt als
controlewaarde in de grafieken. Ze vormen een controle voor de celviabiliteit van cellen die
niet-ingekapseld of enzymbehandeld zijn.
De well plaat met cellen in suspensie, uv-bestraalde cellen in gel en niet-uv bestraalde cellen
in gel wordt dan gedurende 90 minuten in de incubator geplaatst bij 37°C en elk half uur
wordt er geschud.
De cellen in suspensie en cellen vrijgesteld uit gels worden uit de 24-well plaat gehaald en
overgebracht in steriele eppendorfs. De cellen worden afgedraaid (5 min, 1000 rpm). Het

22
supernatans wordt afgehaald en er wordt 500 µl MTT mengsel (zoals bij MTT-test dag 0)
toegevoegd. De cellen worden gedurende 4 uur bij 37°C in het donker op de schudplaat gezet.
Daarna worden de cellen afgedraaid gedurende 5 min (1000 rpm) en het MTT mengsel wordt
verwijderd. Vervolgens wordt 500 µl lysebuffer toegevoegd. De cellen worden gedurende
30 minuten opnieuw op de schudplaat in het donker bij 37°C geplaatst. Hierna wordt er goed
geresuspendeerd en er wordt 200 µl in een 96-well plaat gebracht. Vervolgens wordt de
absorbantie bepaald.
2.3.4.1.2 „Live/dead assay‟
Na de inkapseling in gel-MOD wordt een Calceïne AM/propidium iodide (Ca/PI) kleuring
(‘live/dead’ assay) uitgevoerd om de viabele cellen (groen) van de dode cellen (rood) te
onderscheiden. Hiervoor maakt men gebruik van een oplossing met 1 ml steriele PBS, 2 µl
calceïne AM (Anaspec, N°89201) en 2 µl propidium iodide (Sigma-Aldrich®, P4170). Deze
oplossing wordt goed gemengd en in het donker bewaard. Het bovenstaande medium wordt
van de gel verwijderd en de gel wordt 2 keer gespoeld met PBS, telkens gedurende 1 minuut.
Vervolgens wordt 500 µl van het Ca/PI mengsel aan de well toegevoegd. Deze oplossing laat
men 10 minuten bij kamertemperatuur in het donker staan. Vervolgens wordt er nog 2 keer
gespoeld met PBS, telkens gedurende 1 minuut. Dan worden er met behulp van de
fluorescentiemicroscoop (Olympus IX81) foto’s gemaakt met filters voor ‘Green Fluorescent
Protein’ (GFP) en Texas Red.
2.3.4.1.3 Histologie
De gels die gebruikt worden voor coupes worden gespoeld met PBS en dan gefixeerd in een
4% paraformaldehyde in PBS. De gels worden tot aan de inbedding bewaard in de koelkast.
Na verwerking wordt aan de hand van een haematoxyline eosine (HE) kleuring (bijlage XII)
bepaald welke coupes verder gekleurd worden met de methodes beschreven in 2.6.
2.3.4.2 Viabiliteit en proliferatie in alginaat
2.3.4.2.1 MTT analyse
Alle testen op dag 0 zijn volledig analoog met deze beschreven bij inkapseling in gel-MOD
(2.3.4.1.1). Het isoleren van cellen uit alginaat voor de MTT test op dag 1 en dag 3 gebeurt
met Natrium-citraat (55 mM) in AD gedurende 30 minuten.
2.3.4.2.2 „Live/dead assay‟
Voor de Ca/PI kleuring wordt PBS vervangen door Dulbecco’s PBS (bijlage VIII). De
‘live/dead’ kleuring wordt uitgevoerd op dag 1, 3, 7, 14, 21 en 28.

23
Er worden per tijdstip 20 alginaatbeads gefixeerd in natriumcacodylaat (0,1 M) met 1%
CaCl2.2H2O, 4% formaldehyde en 0,5% glutaraldehyde. Deze beads worden overnacht in de
koelkast geplaatst tot verdere doorwerking gebeurt.
2.3.4.3 Viabiliteit in agarose
Voor cellen in agarose wordt een ‘live-dead assay’ uitgevoerd op dag 21 en worden
histologische coupes gemaakt.
2.4 Celadhesie van HFF’s op 2D-coatings
Voordat cellen uitgezaaid worden op scaffolds bekijkt men het celadhesief effect van
chemische modificaties op biomaterialen. Hiervoor wordt gebruik gemaakt van glazen
schijfjes die gecoat worden met poly-ε-caprolacton (PCL) waarop verschillende chemische
modificaties zijn aangebracht. Een PCL film wordt na een plasmabehandeling gegraft met 2-
aminoethyl methacrylaat (AEMA). De AEMA molecule bevat een positief geladen primaire
aminegroep die biologisch actieve molecules toelaat om te binden op deze groep. Een
voorbeeld hiervan is gelatine (GelB) dat een niet-toxisch en biodegradeerbaar materiaal is.
Van gelatine is geweten dat het de celadhesie bevordert. Na binding van gelatine op PCL
coatings is het mogelijk om het oppervlak verder te modificeren met fibronectine (Fn) via een
dip-coating. Fibronectine is een belangrijk bestanddeel van de extracellulaire matrix en
reguleert celadhesie, differentiatie, proliferatie en migratie. [22]
HFF’s worden uitgezaaid op coatings (PCL, PCL-AEMA, PCL-GelB en PCL-GelB Fn) in
een 24-well plaat met een densiteit van 30.000 cellen per well in HFF medium (bijlage IX).
Men laat de cellen vasthechten gedurende 4 uur of 24 uur. De coatings worden gespoeld met
PBS. Vervolgens worden de cellen gefixeerd met 4% paraformaldehyde (in PBS) en
gepermeabiliseerd met Triton. Daarna voert men een actine cytoskelet en focale
adhesiepunten kleuring uit volgens het protocol in bijlage X. Alle gebruikte coatings zijn
afkomstig van de ‘Polymer Chemistry and Biomaterials Group’ (Universiteit Gent).
2.5 Cultuur van primaire fibrochondrocyten op 3D-scaffolds
2.5.1 Uitzaaien op commerciële scaffolds
2.5.1.1 Becton DickinsonTM
Three Dimensional Collagen Composite Scaffold
Deze scaffold is opgebouwd uit een mengsel van collageen type I en collageen type II
afkomstig van runderhuid. Per experiment worden de scaffolds (diameter 4,2-5,2 mm; hoogte
3,9-4,5 mm; gemiddelde poriegrootte 100-200 µm) doormidden gesneden. Ze worden 1 dag

24
voor uitzaaien in medium ondergedompeld. De halve scaffolds worden in eppendorfs met
1 ml DMEM/F12 gebracht en daarna met behulp van een naald en spuit vacuüm gezogen
zodat ze volledig ondergedompeld blijven en naar de bodem zakken. Hierna worden de
eppendorfs overnacht bij 37°C geplaatst.
De volgende dag worden de scaffolds overgebracht in een 96-well plaat en per scaffold
worden 1.000.000 cellen in 20 µl medium (FC medium, chondrogeen of basis medium
(bijlage V, VI)) voorzichtig op de scaffold gedruppeld. Dan laat men de cellen 4 uur
vasthechten en daarna vult men aan tot 200 µl met het corresponderende medium. De
scaffolds worden dan bij 37°C geplaatst. De volgende dag worden de scaffolds overgebracht
in een 12-well suspensieplaat. Aan elke well wordt dan 3 ml medium toegevoegd en elke 2
dagen wordt het medium ververst. De scaffolds worden geëvalueerd op dag 7, 14 en 21.
2.5.1.2 Sigma-Aldrich® 3D Biotek 3D Inserts PCL scaffoldsTM
Men zaait primaire fibrochondrocyten uit op 3D Biotek Inserts van Sigma-Aldrich (Z724513)
(diameter 5,2 mm; hoogte 1,6 mm; gemiddelde poriegrootte 300 µm) volgens bovenstaand
protocol, maar zonder ze doormidden te snijden. Deze scaffolds zijn opgebouwd uit PCL.
2.5.2 Uitzaaien op geplotte scaffolds
Primaire fibrochondrocyten zaait men uit op PCL, PCL-O en PCL-gelB Fn scaffolds
gesynthetiseerd door de ‘Polymer Chemistry and Biomaterials Group’ (Universiteit Gent)
(hoogte 3 mm; diameter 4,5 mm; gemiddelde poriegrootte 518 µm). De werkwijze voor
modificeren van scaffolds staat beschreven in een artikel van Desmet et al. [22] Het gebruikte
protocol voor uitzaaien is analoog aan dit beschreven in 2.5.1.1. De scaffolds worden echter
niet doormidden gesneden.
2.5.3 Viabiliteit en proliferatie op scaffolds
Op dag 7, 14 en 21 worden de scaffolds gekleurd met Ca/PI kleuring en worden foto’s
gemaakt zoals beschreven in 2.3.4.1.2. Daarna worden de scaffolds overnacht gefixeerd in
neutraal gebufferde formol (4%) (bijlage XI) en verwerkt tot coupes.
2.6 Histochemische karakterisatie
2.6.1 Haematoxyline eosine kleuring
De routine haematoxyline eosine (HE) kleuring wordt uitgevoerd om een algemeen overzicht
te krijgen van de morfologie en de celdistributie op de coupes. HE kleurt celkernen blauw-
paars en de kraakbeenmatrix roze. (bijlage XII) [23]

25
2.6.2 Alciaanblauw kleuring
Alciaanblauw is een basische kleurstof met Cu-kern die bij lage pH reageert met zure negatief
geladen groepen uit zure mucopolysacchariden, zoals chondroïtinesulfaat en hyaluronzuur uit
kraakbeen. Hierdoor ontstaat een permanente, onoplosbare blauwe kleur.(bijlage XIII) [24]
2.7 Statistische verwerking
De statistische verwerking van de gegevens van de MTT-testen voor fibrochondrocyten in
gel-MOD en in alginaat gebeurt aan de hand van een ‘Mann-Withney U’-test m.b.v. SPSS
Statistics 19. [25]
Deze test laat toe na te gaan of 2 onafhankelijke (ongepaarde) steekproeven afkomstig zijn uit
dezelfde populatie. Gezien de kleine steekproefaantallen en gezien het feit dat de waarden niet
beantwoorden aan de normale verdeling wordt voor de ‘Mann-Withney U’-test gekozen. [25]
De nulhypothese (H0) kan als volgt geformuleerd worden:
H0: beide steekproeven zijn afkomstig uit dezelfde populatie. In dit geval zou er geen verschil
mogen gezien worden tussen de MTT waarden van de cellen ingekapseld in alginaat en deze
ingekapseld in gel-MOD. Als er wel een verschil gezien wordt, geldt de alternatieve
hypothese HA: beide steekproeven zijn niet afkomstig uit dezelfde populatie.
De nulhypothese wordt verworpen ten voordele van de alternatieve hypothese als p < 0,05.
3 Resultaten
3.1 Encapsulatie van primaire fibrochondrocyten in hydrogels
3.1.1 Encapsulatie in gel-MOD
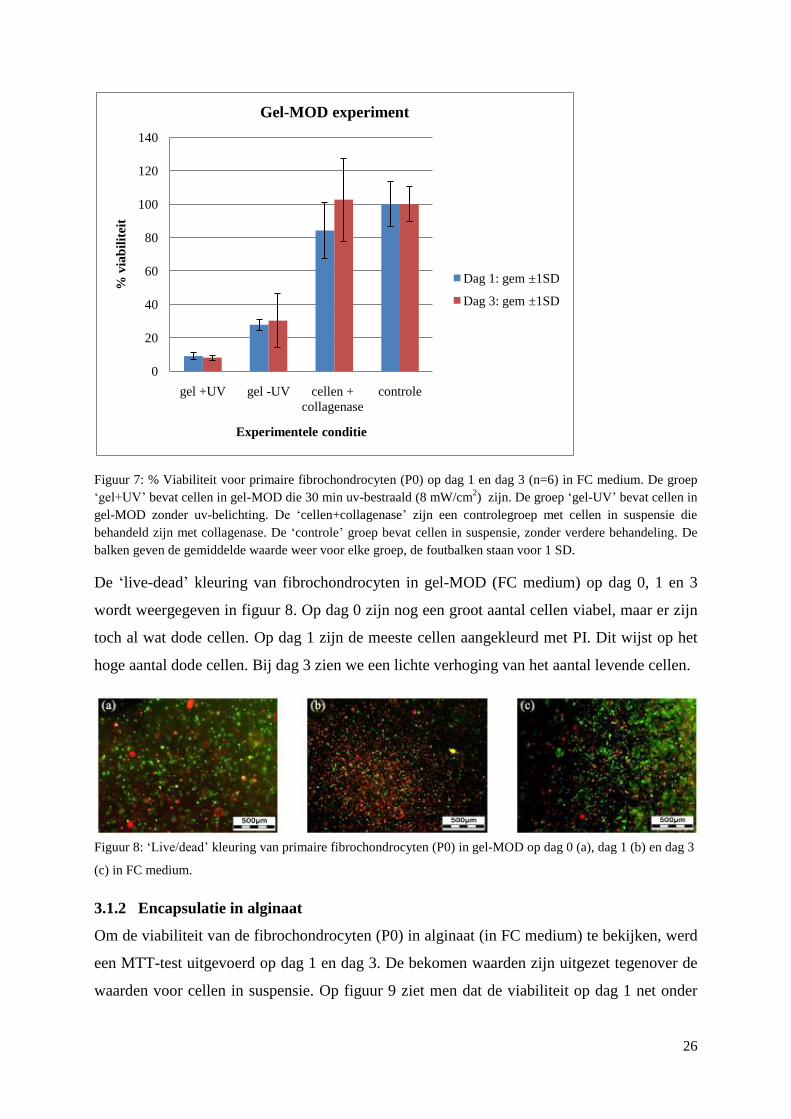
Primaire fibrochondrocyten (P0) werden ingekapseld in gel-MOD (1eq 10% w/v, DS 67%).
In figuur 7 worden de waarden voor dag 1 en dag 3 weergegeven, uitgezet t.o.v. cellen in
suspensie (controle). Opmerkelijk is hier de zeer lage viabiliteit voor de cellen in gel-MOD
die uv-bestraald zijn. De cellen die ingekapseld zijn in gel-MOD zonder uv-bestraling hebben
een iets hogere viabiliteit. Cellen die enkel collagenase behandeld zijn, hebben een viabiliteit
die ongeveer 8 keer hoger ligt in vergelijking met de cellen in gel-MOD die uv-bestraald zijn.
26
Figuur 7: % Viabiliteit voor primaire fibrochondrocyten (P0) op dag 1 en dag 3 (n=6) in FC medium. De groep
‘gel+UV’ bevat cellen in gel-MOD die 30 min uv-bestraald (8 mW/cm2) zijn. De groep ‘gel-UV’ bevat cellen in
gel-MOD zonder uv-belichting. De ‘cellen+collagenase’ zijn een controlegroep met cellen in suspensie die
behandeld zijn met collagenase. De ‘controle’ groep bevat cellen in suspensie, zonder verdere behandeling. De
balken geven de gemiddelde waarde weer voor elke groep, de foutbalken staan voor 1 SD.
De ‘live-dead’ kleuring van fibrochondrocyten in gel-MOD (FC medium) op dag 0, 1 en 3
wordt weergegeven in figuur 8. Op dag 0 zijn nog een groot aantal cellen viabel, maar er zijn
toch al wat dode cellen. Op dag 1 zijn de meeste cellen aangekleurd met PI. Dit wijst op het
hoge aantal dode cellen. Bij dag 3 zien we een lichte verhoging van het aantal levende cellen.
Figuur 8: ‘Live/dead’ kleuring van primaire fibrochondrocyten (P0) in gel-MOD op dag 0 (a), dag 1 (b) en dag 3
(c) in FC medium.
3.1.2 Encapsulatie in alginaat
Om de viabiliteit van de fibrochondrocyten (P0) in alginaat (in FC medium) te bekijken, werd
een MTT-test uitgevoerd op dag 1 en dag 3. De bekomen waarden zijn uitgezet tegenover de
waarden voor cellen in suspensie. Op figuur 9 ziet men dat de viabiliteit op dag 1 net onder
0
20
40
60
80
100
120
140
gel +UV gel -UV cellen +
collagenase
controle
% v
iab
ilit
eit
Experimentele conditie
Gel-MOD experiment
Dag 1: gem ±1SD
Dag 3: gem ±1SD
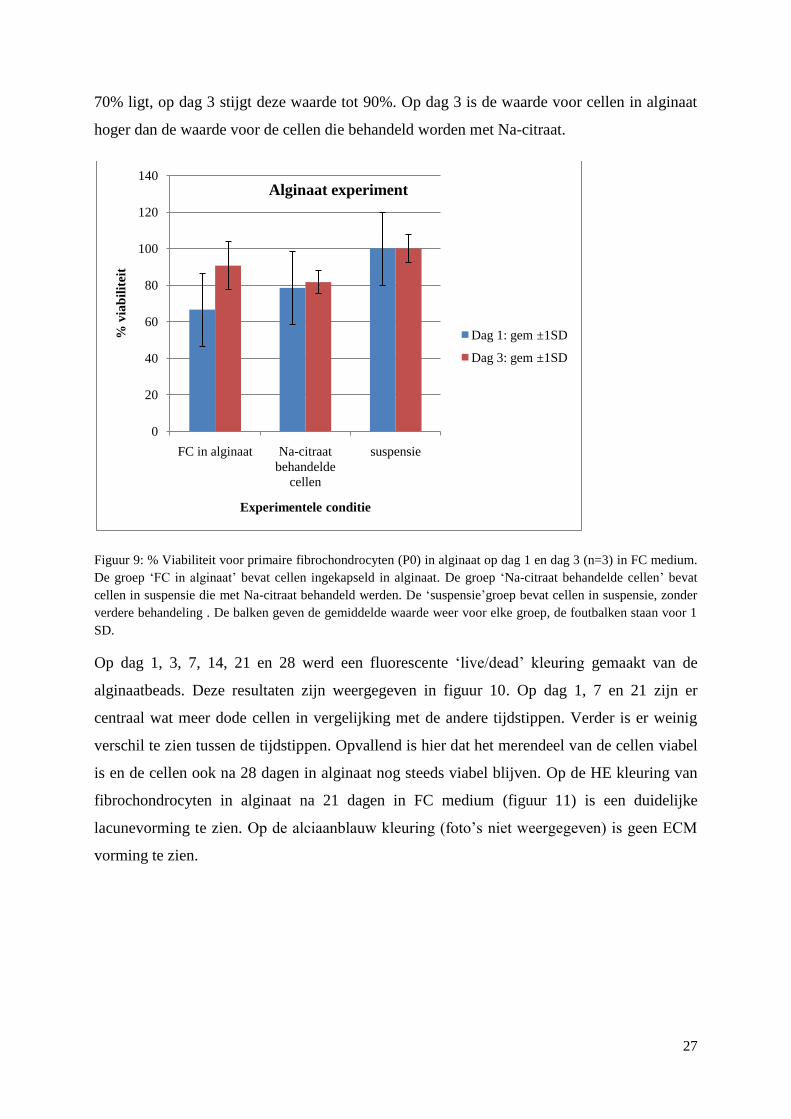
27
70% ligt, op dag 3 stijgt deze waarde tot 90%. Op dag 3 is de waarde voor cellen in alginaat
hoger dan de waarde voor de cellen die behandeld worden met Na-citraat.
Figuur 9: % Viabiliteit voor primaire fibrochondrocyten (P0) in alginaat op dag 1 en dag 3 (n=3) in FC medium.
De groep ‘FC in alginaat’ bevat cellen ingekapseld in alginaat. De groep ‘Na-citraat behandelde cellen’ bevat
cellen in suspensie die met Na-citraat behandeld werden. De ‘suspensie’groep bevat cellen in suspensie, zonder
verdere behandeling . De balken geven de gemiddelde waarde weer voor elke groep, de foutbalken staan voor 1
SD.
Op dag 1, 3, 7, 14, 21 en 28 werd een fluorescente ‘live/dead’ kleuring gemaakt van de
alginaatbeads. Deze resultaten zijn weergegeven in figuur 10. Op dag 1, 7 en 21 zijn er
centraal wat meer dode cellen in vergelijking met de andere tijdstippen. Verder is er weinig
verschil te zien tussen de tijdstippen. Opvallend is hier dat het merendeel van de cellen viabel
is en de cellen ook na 28 dagen in alginaat nog steeds viabel blijven. Op de HE kleuring van
fibrochondrocyten in alginaat na 21 dagen in FC medium (figuur 11) is een duidelijke
lacunevorming te zien. Op de alciaanblauw kleuring (foto’s niet weergegeven) is geen ECM
vorming te zien.
0
20
40
60
80
100
120
140
FC in alginaat Na-citraat
behandelde
cellen
suspensie
% v
iab
ilit
eit
Experimentele conditie
Alginaat experiment
Dag 1: gem ±1SD
Dag 3: gem ±1SD
(a) (b) (c)
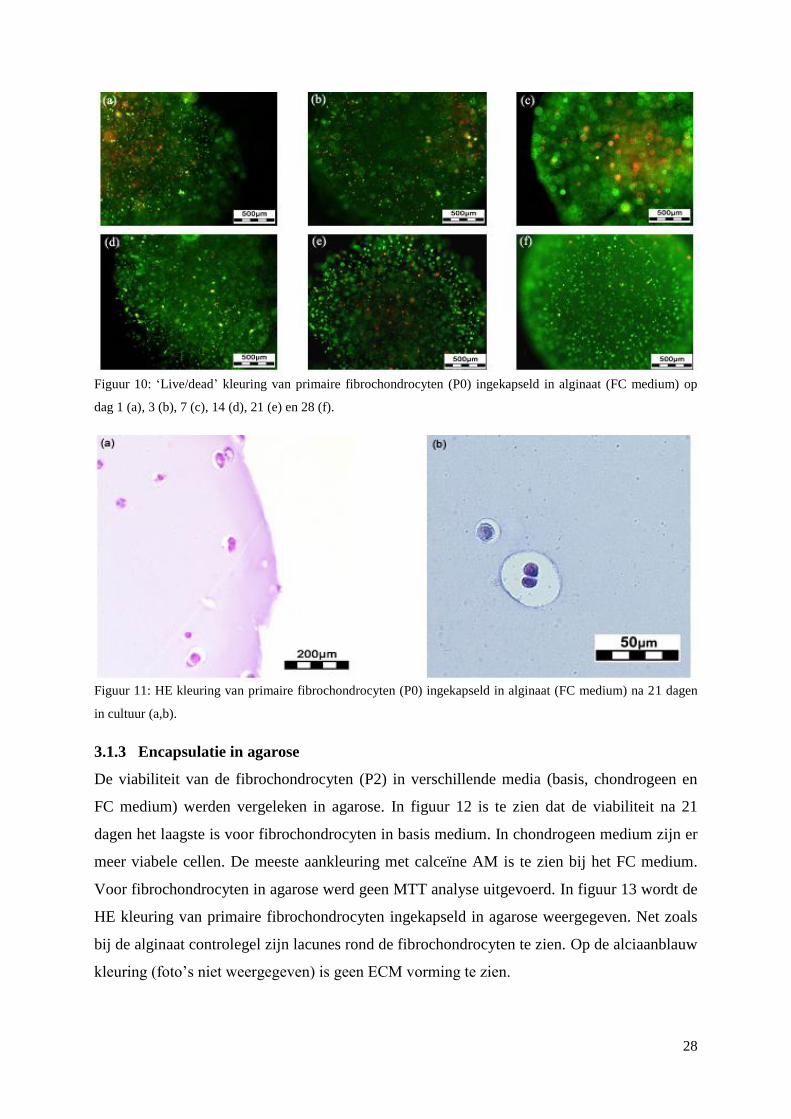
28
Figuur 10: ‘Live/dead’ kleuring van primaire fibrochondrocyten (P0) ingekapseld in alginaat (FC medium) op
dag 1 (a), 3 (b), 7 (c), 14 (d), 21 (e) en 28 (f).
Figuur 11: HE kleuring van primaire fibrochondrocyten (P0) ingekapseld in alginaat (FC medium) na 21 dagen
in cultuur (a,b).
3.1.3 Encapsulatie in agarose
De viabiliteit van de fibrochondrocyten (P2) in verschillende media (basis, chondrogeen en
FC medium) werden vergeleken in agarose. In figuur 12 is te zien dat de viabiliteit na 21
dagen het laagste is voor fibrochondrocyten in basis medium. In chondrogeen medium zijn er
meer viabele cellen. De meeste aankleuring met calceïne AM is te zien bij het FC medium.
Voor fibrochondrocyten in agarose werd geen MTT analyse uitgevoerd. In figuur 13 wordt de
HE kleuring van primaire fibrochondrocyten ingekapseld in agarose weergegeven. Net zoals
bij de alginaat controlegel zijn lacunes rond de fibrochondrocyten te zien. Op de alciaanblauw
kleuring (foto’s niet weergegeven) is geen ECM vorming te zien.
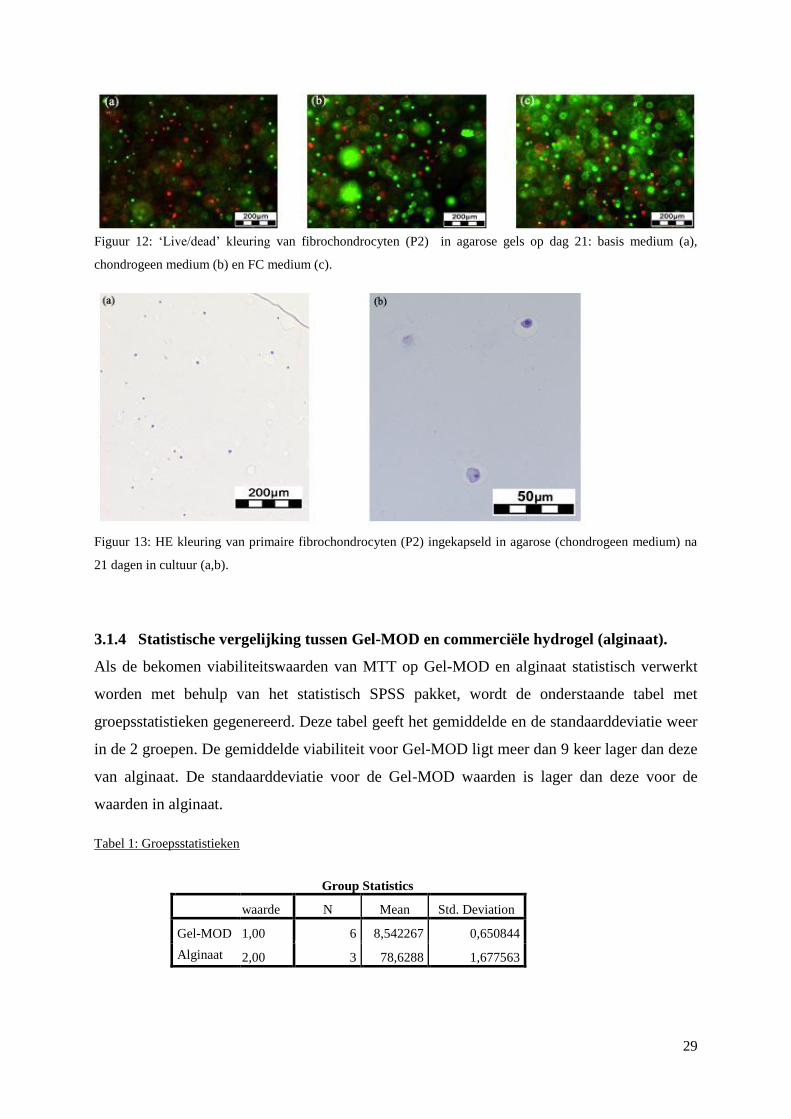
29
Figuur 12: ‘Live/dead’ kleuring van fibrochondrocyten (P2) in agarose gels op dag 21: basis medium (a),
chondrogeen medium (b) en FC medium (c).
Figuur 13: HE kleuring van primaire fibrochondrocyten (P2) ingekapseld in agarose (chondrogeen medium) na
21 dagen in cultuur (a,b).
3.1.4 Statistische vergelijking tussen Gel-MOD en commerciële hydrogel (alginaat).
Als de bekomen viabiliteitswaarden van MTT op Gel-MOD en alginaat statistisch verwerkt
worden met behulp van het statistisch SPSS pakket, wordt de onderstaande tabel met
groepsstatistieken gegenereerd. Deze tabel geeft het gemiddelde en de standaarddeviatie weer
in de 2 groepen. De gemiddelde viabiliteit voor Gel-MOD ligt meer dan 9 keer lager dan deze
van alginaat. De standaarddeviatie voor de Gel-MOD waarden is lager dan deze voor de
waarden in alginaat.
Tabel 1: Groepsstatistieken
Group Statistics
waarde N Mean Std. Deviation
Gel-MOD
Alginaat
1,00 6 8,542267 0,650844
2,00 3 78,6288 1,677563
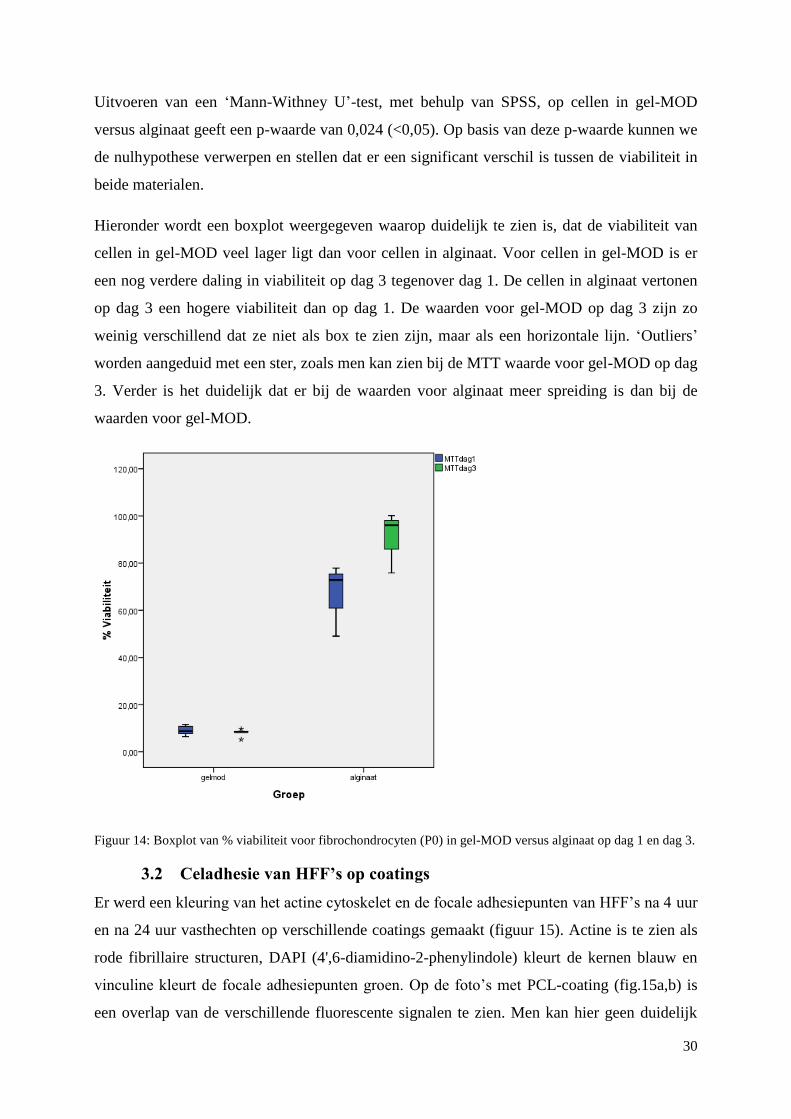
30
Uitvoeren van een ‘Mann-Withney U’-test, met behulp van SPSS, op cellen in gel-MOD
versus alginaat geeft een p-waarde van 0,024 (<0,05). Op basis van deze p-waarde kunnen we
de nulhypothese verwerpen en stellen dat er een significant verschil is tussen de viabiliteit in
beide materialen.
Hieronder wordt een boxplot weergegeven waarop duidelijk te zien is, dat de viabiliteit van
cellen in gel-MOD veel lager ligt dan voor cellen in alginaat. Voor cellen in gel-MOD is er
een nog verdere daling in viabiliteit op dag 3 tegenover dag 1. De cellen in alginaat vertonen
op dag 3 een hogere viabiliteit dan op dag 1. De waarden voor gel-MOD op dag 3 zijn zo
weinig verschillend dat ze niet als box te zien zijn, maar als een horizontale lijn. ‘Outliers’
worden aangeduid met een ster, zoals men kan zien bij de MTT waarde voor gel-MOD op dag
3. Verder is het duidelijk dat er bij de waarden voor alginaat meer spreiding is dan bij de
waarden voor gel-MOD.
Figuur 14: Boxplot van % viabiliteit voor fibrochondrocyten (P0) in gel-MOD versus alginaat op dag 1 en dag 3.
3.2 Celadhesie van HFF’s op coatings
Er werd een kleuring van het actine cytoskelet en de focale adhesiepunten van HFF’s na 4 uur
en na 24 uur vasthechten op verschillende coatings gemaakt (figuur 15). Actine is te zien als
rode fibrillaire structuren, DAPI (4',6-diamidino-2-phenylindole) kleurt de kernen blauw en
vinculine kleurt de focale adhesiepunten groen. Op de foto’s met PCL-coating (fig.15a,b) is
een overlap van de verschillende fluorescente signalen te zien. Men kan hier geen duidelijk

31
actine cytoskelet waarnemen en ook de focale adhesiepunten zijn niet te zien. De cellen
vertonen op beide foto’s een eerder afgeronde morfologie. Op de PCL-AEMA-coatings
(fig.15c,d) is het actine cytoskelet aangekleurd en zijn duidelijk actinevezels te zien. Na 4 uur
vasthechten zijn al lichte focale adhesiepunten zichtbaar (fig.15c). Na 24 uur vertonen de
cellen een sterk uitgestrekte morfologie (fig.15d). Op de PCL-gelB coating zijn de cellen na 4
uur al sterk uitgestrekt (fig.15e), de focale adhesiepunten zijn ook zichtbaar. De HFF’s op
PCL-gelB Fn coatings (fig.15g,h) vertonen een duidelijk actine cytoskelet en focale
adhesiepunten, zowel na 4 uur (fig.15g) als na 24 uur (fig.15h). De cellen zijn minder lang,
maar breder in vergelijking met de PCL-GelB coating.
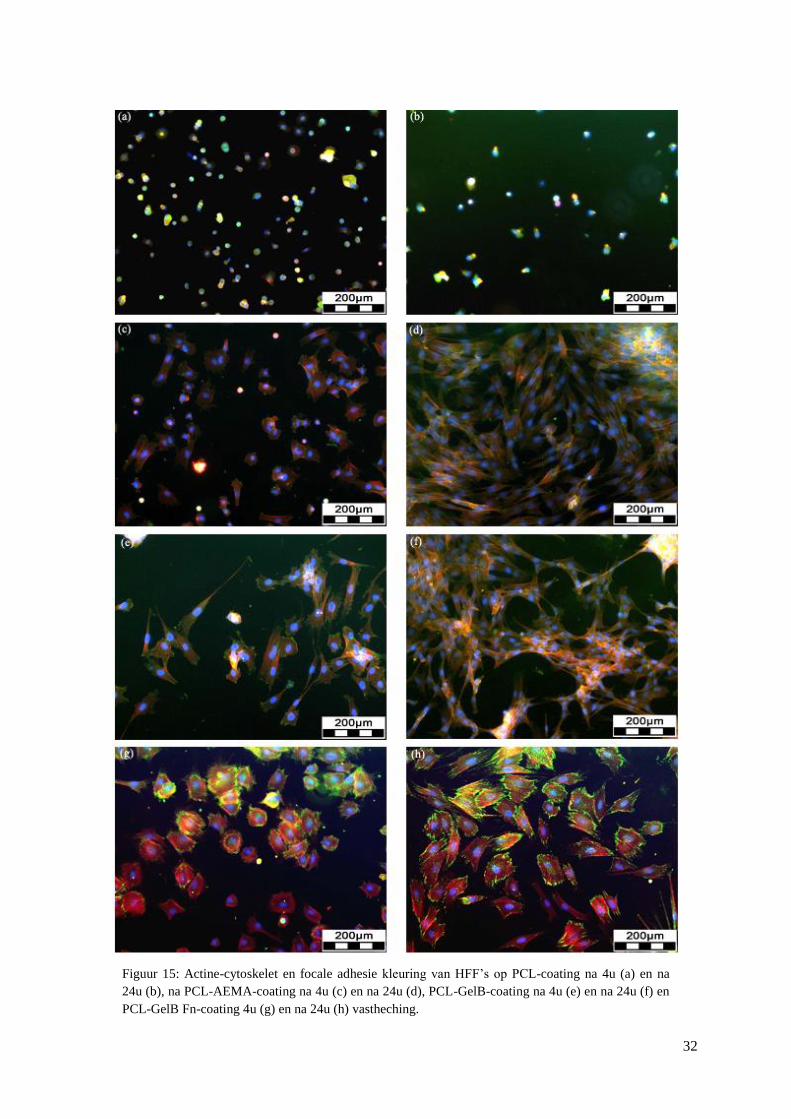
32
Figuur 15: Actine-cytoskelet en focale adhesie kleuring van HFF’s op PCL-coating na 4u (a) en na
24u (b), na PCL-AEMA-coating na 4u (c) en na 24u (d), PCL-GelB-coating na 4u (e) en na 24u (f) en
PCL-GelB Fn-coating 4u (g) en na 24u (h) vastheching.

33
3.3 Primaire fibrochondrocyten op scaffolds
Om voldoende cellen te bekomen voor het uitzaaien op scaffolds, werden de
fibrochondrocyten eerst geëxpandeerd als monolayer in falcons. De fibrochondrocyten waren
na 4 dagen confluent en hadden zoveel ECM aangemaakt dat het niet mogelijk was om de
cellen te isoleren met trypsine. Het was noodzakelijk om de ECM af te breken met
collagenase.
In een screeningstest werd gezien dat collageenscaffolds (BDTM
) die bezaaid werden met pas
geïsoleerde fibrochondrocyten (P0), oplosten na 24 uur. Dit werd ook gezien bij
fibrochondrocyten (P2) die behandeld waren met collagenase na expansie. Hieruit werd
besloten dat de collagenase, ondanks 3 keer spoelen en centrifugeren, nog steeds niet was
verwijderd. Daarom werden de cellen na de collagenasebehandeling opnieuw in cultuur
gebracht gedurende 24 uur. Als de cellen vervolgens uitgezaaid werden op de
collageenscaffolds, gebeurden er geen zichtbare veranderingen.
In een tweede experiment werden fibrochondrocyten (P2) op scaffolds uitgezaaid in FC
medium en na 1 dag overgebracht naar basis medium, chondrogeen medium en FC medium.
De scaffolds werden gedurende 21 dagen opgevolgd. Er werden geen cellen waargenomen op
de scaffolds in basis medium en chondrogeen medium. De scaffolds in FC medium
vertoonden wel goede resultaten.
Voor het derde experiment werden de cellen (P2) eerst 7 dagen vastgehecht in FC medium en
vervolgens overgebracht naar chondrogeen medium of FC medium.
3.3.1 Viabiliteit
Om de viabiliteit op verschillende scaffolds te onderzoeken, werd gebruik gemaakt van een
‘live/dead’ kleuring op dag 7, 14 en 21.
Figuur 16 geeft de fluorescentiebeelden weer van fibrochondrocyten op PCL. Op dag 7 (a,b)
is de scaffold in FC medium al relatief goed begroeid met fibrochondrocyten. In figuur b zijn
‘celbruggen’ te zien tussen de verschillende struts. Op dag 14 (c,d) is de scaffold in FC
medium mooi begroeid met cellen, er zijn ook celbruggen te zien. Er zijn echter wat meer
dode cellen. Opvallend is ook dat de scaffold minder regelmatig gevormd is waardoor de
celbruggen niet overal even breed zijn. Na 21 dagen in FC medium (e,f) zijn er veel minder
cellen te zien. In chondrogeen medium ziet men op dag 14 (g,h) en dag 21 (i,j) een duidelijk
lager celaantal in vergelijking met de corresponderende tijdstippen in FC medium.
(a) (b) (c)
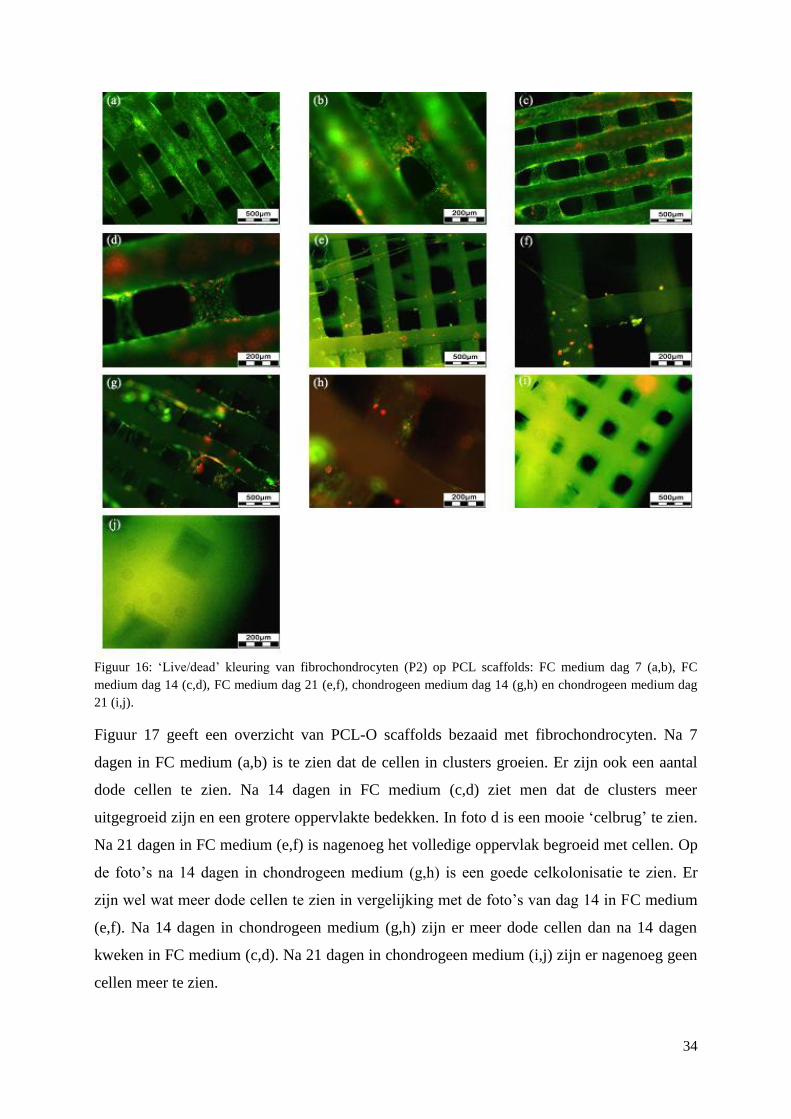
34
Figuur 16: ‘Live/dead’ kleuring van fibrochondrocyten (P2) op PCL scaffolds: FC medium dag 7 (a,b), FC
medium dag 14 (c,d), FC medium dag 21 (e,f), chondrogeen medium dag 14 (g,h) en chondrogeen medium dag
21 (i,j).
Figuur 17 geeft een overzicht van PCL-O scaffolds bezaaid met fibrochondrocyten. Na 7
dagen in FC medium (a,b) is te zien dat de cellen in clusters groeien. Er zijn ook een aantal
dode cellen te zien. Na 14 dagen in FC medium (c,d) ziet men dat de clusters meer
uitgegroeid zijn en een grotere oppervlakte bedekken. In foto d is een mooie ‘celbrug’ te zien.
Na 21 dagen in FC medium (e,f) is nagenoeg het volledige oppervlak begroeid met cellen. Op
de foto’s na 14 dagen in chondrogeen medium (g,h) is een goede celkolonisatie te zien. Er
zijn wel wat meer dode cellen te zien in vergelijking met de foto’s van dag 14 in FC medium
(e,f). Na 14 dagen in chondrogeen medium (g,h) zijn er meer dode cellen dan na 14 dagen
kweken in FC medium (c,d). Na 21 dagen in chondrogeen medium (i,j) zijn er nagenoeg geen
cellen meer te zien. (b) (c)
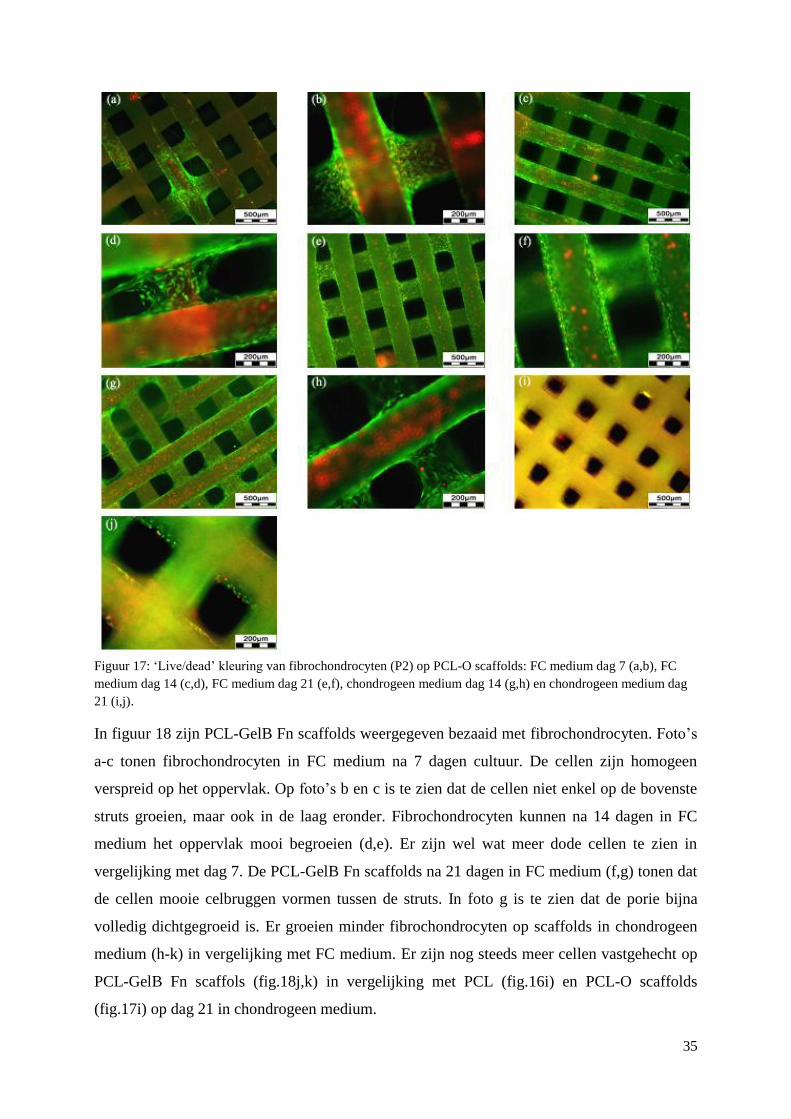
35
Figuur 17: ‘Live/dead’ kleuring van fibrochondrocyten (P2) op PCL-O scaffolds: FC medium dag 7 (a,b), FC
medium dag 14 (c,d), FC medium dag 21 (e,f), chondrogeen medium dag 14 (g,h) en chondrogeen medium dag
21 (i,j).
In figuur 18 zijn PCL-GelB Fn scaffolds weergegeven bezaaid met fibrochondrocyten. Foto’s
a-c tonen fibrochondrocyten in FC medium na 7 dagen cultuur. De cellen zijn homogeen
verspreid op het oppervlak. Op foto’s b en c is te zien dat de cellen niet enkel op de bovenste
struts groeien, maar ook in de laag eronder. Fibrochondrocyten kunnen na 14 dagen in FC
medium het oppervlak mooi begroeien (d,e). Er zijn wel wat meer dode cellen te zien in
vergelijking met dag 7. De PCL-GelB Fn scaffolds na 21 dagen in FC medium (f,g) tonen dat
de cellen mooie celbruggen vormen tussen de struts. In foto g is te zien dat de porie bijna
volledig dichtgegroeid is. Er groeien minder fibrochondrocyten op scaffolds in chondrogeen
medium (h-k) in vergelijking met FC medium. Er zijn nog steeds meer cellen vastgehecht op
PCL-GelB Fn scaffols (fig.18j,k) in vergelijking met PCL (fig.16i) en PCL-O scaffolds
(fig.17i) op dag 21 in chondrogeen medium.
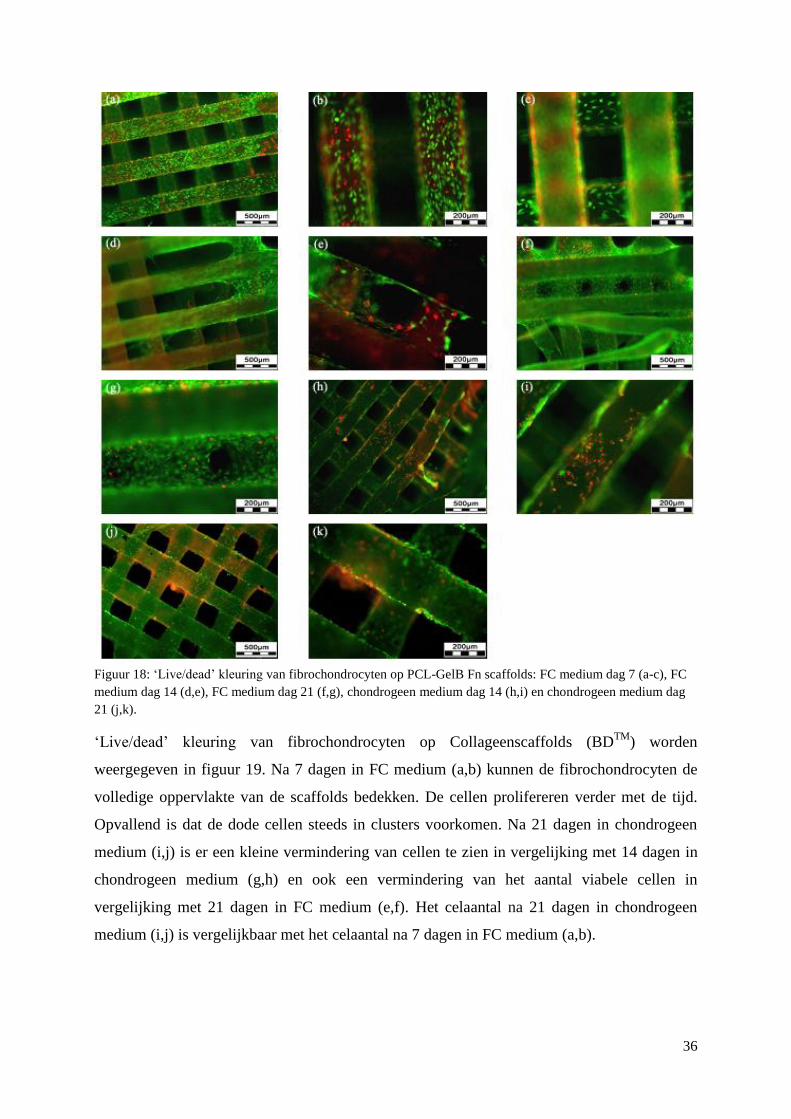
36
Figuur 18: ‘Live/dead’ kleuring van fibrochondrocyten op PCL-GelB Fn scaffolds: FC medium dag 7 (a-c), FC
medium dag 14 (d,e), FC medium dag 21 (f,g), chondrogeen medium dag 14 (h,i) en chondrogeen medium dag
21 (j,k).
‘Live/dead’ kleuring van fibrochondrocyten op Collageenscaffolds (BDTM
) worden
weergegeven in figuur 19. Na 7 dagen in FC medium (a,b) kunnen de fibrochondrocyten de
volledige oppervlakte van de scaffolds bedekken. De cellen prolifereren verder met de tijd.
Opvallend is dat de dode cellen steeds in clusters voorkomen. Na 21 dagen in chondrogeen
medium (i,j) is er een kleine vermindering van cellen te zien in vergelijking met 14 dagen in
chondrogeen medium (g,h) en ook een vermindering van het aantal viabele cellen in
vergelijking met 21 dagen in FC medium (e,f). Het celaantal na 21 dagen in chondrogeen
medium (i,j) is vergelijkbaar met het celaantal na 7 dagen in FC medium (a,b). (c)
(a)
(b)
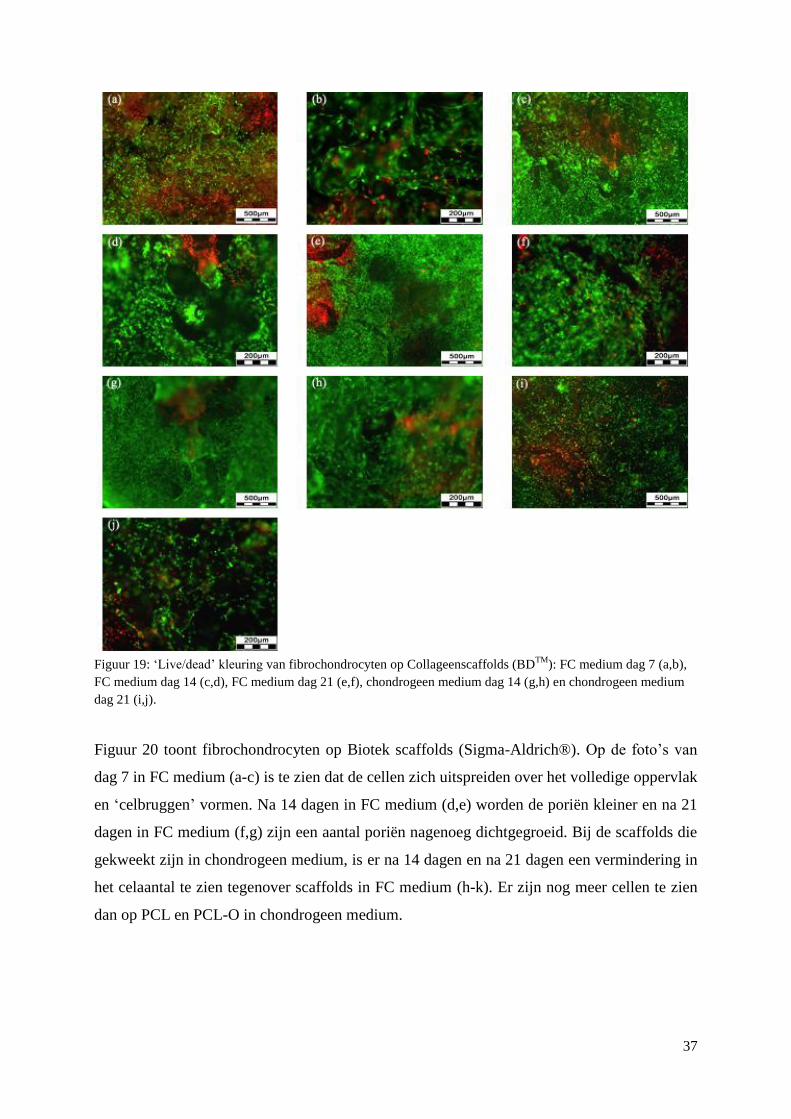
37
Figuur 19: ‘Live/dead’ kleuring van fibrochondrocyten op Collageenscaffolds (BDTM
): FC medium dag 7 (a,b),
FC medium dag 14 (c,d), FC medium dag 21 (e,f), chondrogeen medium dag 14 (g,h) en chondrogeen medium
dag 21 (i,j).
Figuur 20 toont fibrochondrocyten op Biotek scaffolds (Sigma-Aldrich®). Op de foto’s van
dag 7 in FC medium (a-c) is te zien dat de cellen zich uitspreiden over het volledige oppervlak
en ‘celbruggen’ vormen. Na 14 dagen in FC medium (d,e) worden de poriën kleiner en na 21
dagen in FC medium (f,g) zijn een aantal poriën nagenoeg dichtgegroeid. Bij de scaffolds die
gekweekt zijn in chondrogeen medium, is er na 14 dagen en na 21 dagen een vermindering in
het celaantal te zien tegenover scaffolds in FC medium (h-k). Er zijn nog meer cellen te zien
dan op PCL en PCL-O in chondrogeen medium. (c)
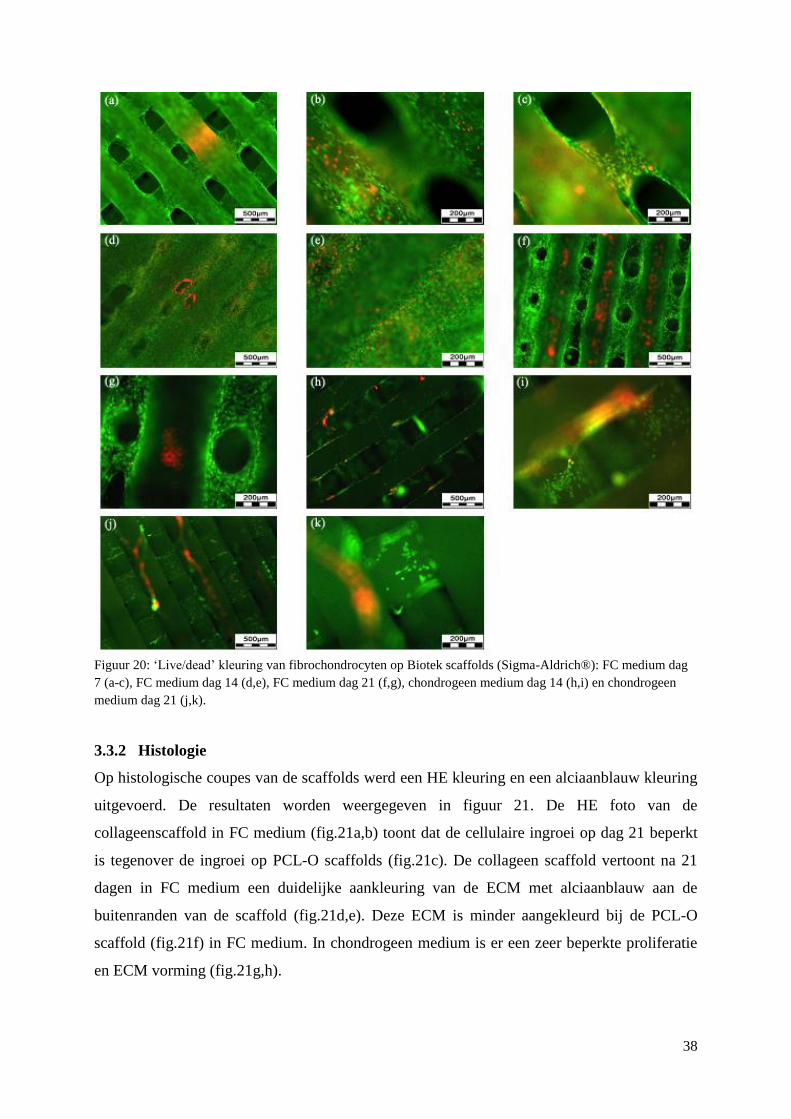
38
Figuur 20: ‘Live/dead’ kleuring van fibrochondrocyten op Biotek scaffolds (Sigma-Aldrich®): FC medium dag
7 (a-c), FC medium dag 14 (d,e), FC medium dag 21 (f,g), chondrogeen medium dag 14 (h,i) en chondrogeen
medium dag 21 (j,k).
3.3.2 Histologie
Op histologische coupes van de scaffolds werd een HE kleuring en een alciaanblauw kleuring
uitgevoerd. De resultaten worden weergegeven in figuur 21. De HE foto van de
collageenscaffold in FC medium (fig.21a,b) toont dat de cellulaire ingroei op dag 21 beperkt
is tegenover de ingroei op PCL-O scaffolds (fig.21c). De collageen scaffold vertoont na 21
dagen in FC medium een duidelijke aankleuring van de ECM met alciaanblauw aan de
buitenranden van de scaffold (fig.21d,e). Deze ECM is minder aangekleurd bij de PCL-O
scaffold (fig.21f) in FC medium. In chondrogeen medium is er een zeer beperkte proliferatie
en ECM vorming (fig.21g,h).
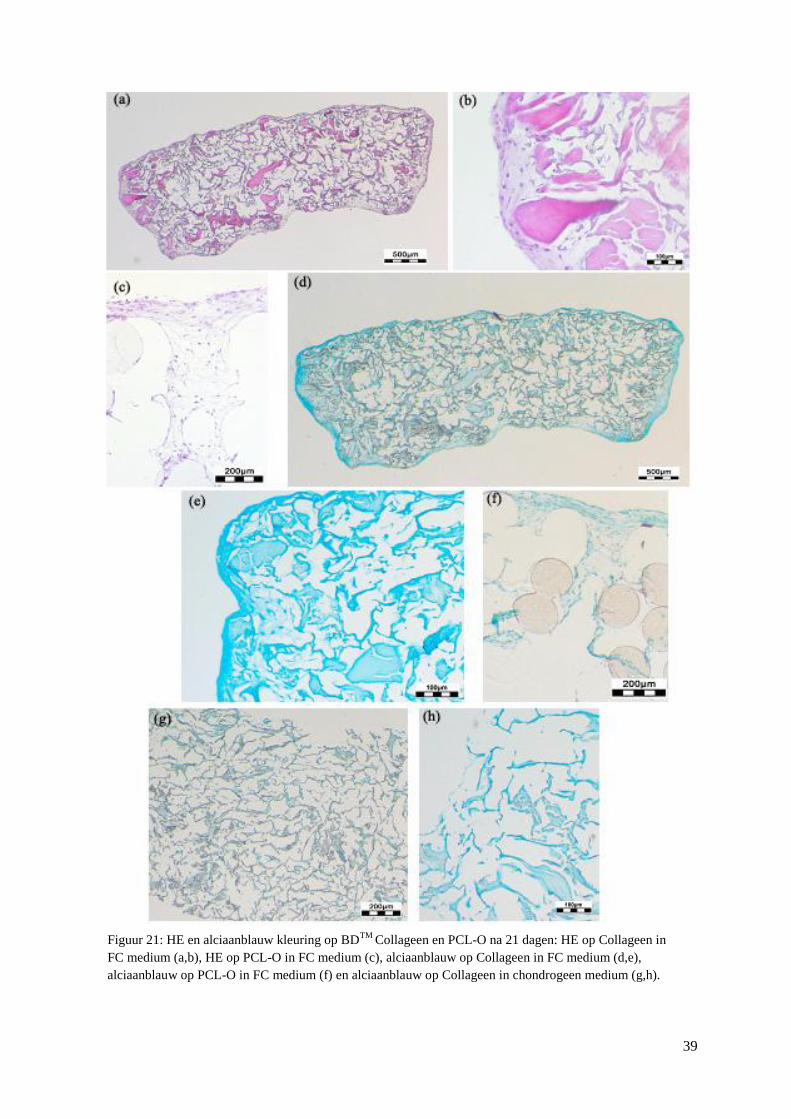
39
Figuur 21: HE en alciaanblauw kleuring op BDTM
Collageen en PCL-O na 21 dagen: HE op Collageen in
FC medium (a,b), HE op PCL-O in FC medium (c), alciaanblauw op Collageen in FC medium (d,e),
alciaanblauw op PCL-O in FC medium (f) en alciaanblauw op Collageen in chondrogeen medium (g,h).
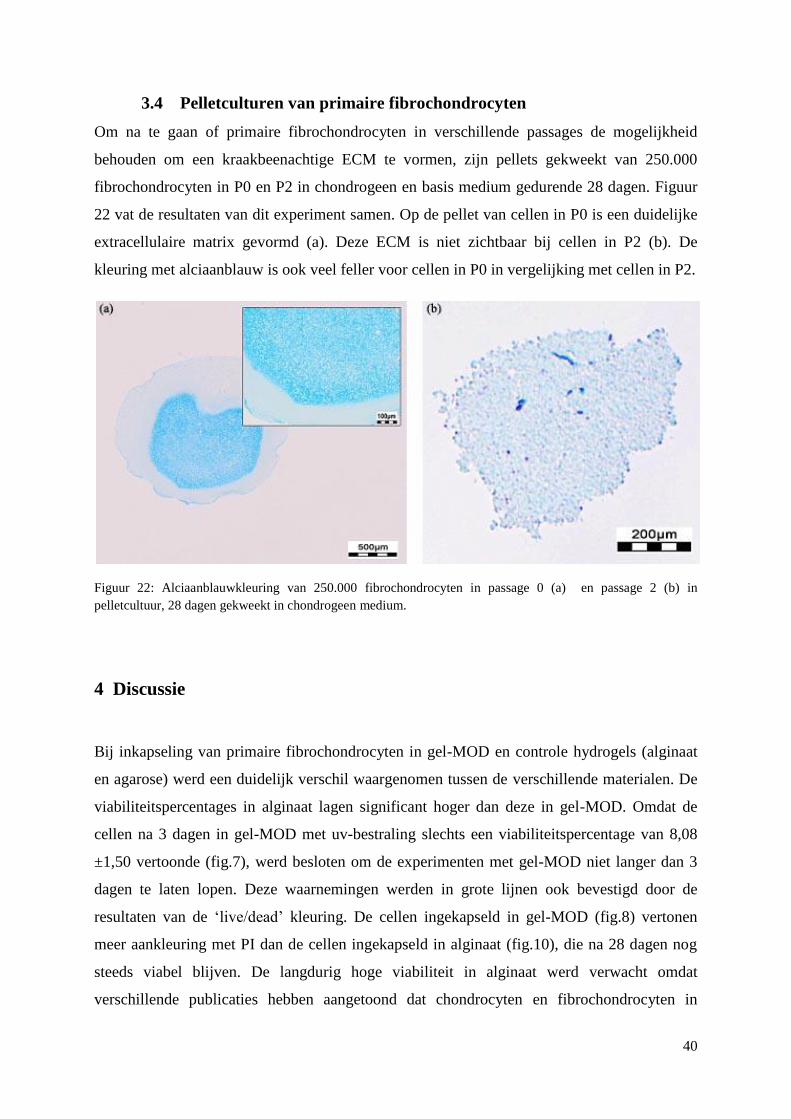
40
3.4 Pelletculturen van primaire fibrochondrocyten
Om na te gaan of primaire fibrochondrocyten in verschillende passages de mogelijkheid
behouden om een kraakbeenachtige ECM te vormen, zijn pellets gekweekt van 250.000
fibrochondrocyten in P0 en P2 in chondrogeen en basis medium gedurende 28 dagen. Figuur
22 vat de resultaten van dit experiment samen. Op de pellet van cellen in P0 is een duidelijke
extracellulaire matrix gevormd (a). Deze ECM is niet zichtbaar bij cellen in P2 (b). De
kleuring met alciaanblauw is ook veel feller voor cellen in P0 in vergelijking met cellen in P2.
Figuur 22: Alciaanblauwkleuring van 250.000 fibrochondrocyten in passage 0 (a) en passage 2 (b) in
pelletcultuur, 28 dagen gekweekt in chondrogeen medium.
4 Discussie
Bij inkapseling van primaire fibrochondrocyten in gel-MOD en controle hydrogels (alginaat
en agarose) werd een duidelijk verschil waargenomen tussen de verschillende materialen. De
viabiliteitspercentages in alginaat lagen significant hoger dan deze in gel-MOD. Omdat de
cellen na 3 dagen in gel-MOD met uv-bestraling slechts een viabiliteitspercentage van 8,08
±1,50 vertoonde (fig.7), werd besloten om de experimenten met gel-MOD niet langer dan 3
dagen te laten lopen. Deze waarnemingen werden in grote lijnen ook bevestigd door de
resultaten van de ‘live/dead’ kleuring. De cellen ingekapseld in gel-MOD (fig.8) vertonen
meer aankleuring met PI dan de cellen ingekapseld in alginaat (fig.10), die na 28 dagen nog
steeds viabel blijven. De langdurig hoge viabiliteit in alginaat werd verwacht omdat
verschillende publicaties hebben aangetoond dat chondrocyten en fibrochondrocyten in

41
alginaat tot zelfs 10 weken viabel kunnen blijven. [7,26] Alginaat wordt dan ook vaak
gebruikt als referentiecultuur.
Het gebruik van P0 fibrochondrocyten zou mogelijk een rol kunnen spelen in de lage MTT
waarden na inkapseling in gel-MOD. De cellen moeten zich immers zowel aan de
cultuuromstandigheden als aan het materiaal aanpassen. Mogelijk kunnen hogere resultaten
voor de viabiliteit bekomen worden als de cellen ingekapseld worden na 1 of 2 passages. Een
aanwijzing voor het feit dat primaire fibrochondrocyten een langere aanpassingsperiode
hebben, is de trage vasthechting op de bodems van de falcons na enzymatische isolatie. Na 1
dag is er slechts een aanhechting van 20% van de cellen. Deze vastgehechte cellen vertonen
een fibroblastachtige morfologie, de niet-vastgehechte cellen vertonen een ronde morfologie.
Na 4-5 dagen zijn bijna alle cellen vastgehecht. In de controlegels (alginaat en agarose)
vertonen de cellen in P0 echter een hogere viabiliteit. Om deze reden is het meer
waarschijnlijk dat de lage viabiliteit veroorzaakt wordt door de eigenschappen van het
materiaal. Het gel-MOD materiaal is gederiveerd van collageen, wat een belangrijke
component is van de ECM, maar om een stevige hydrogel te vormen is modificatie met
methacrylamidegroepen en belichting met uv-licht noodzakelijk. [21] Onderzoek heeft
aangetoond dat het gebruik van een foto-initiator in combinatie met uv-belichting kan leiden
tot significant hogere niveaus van celdood door het ontstaan van vrije radicalen. [27] Er is dan
ook een duidelijk verschil in viabiliteitspercentage te zien op dag 1 tussen de uv-bestraalde
(9,00 ± 1,97) en de niet-uv bestraalde (27,72 ± 3,12) fibrochondrocyten. Een mogelijke
oplossing hiervoor zou zijn om de uv-belichtingstijd drastisch te reduceren (vb. 10 min).
Verder onderzoek is echter nog nodig om na te gaan wat de minimale belichtingstijd is om
een goede crosslinking van de hydrogel toe te laten. Daarnaast zou men ook kunnen zoeken
naar geschikte ‘radicaalscavangers’, zoals ascorbinezuur, om het negatieve effect van vrije
radicalen te minimaliseren. [27]
De viabiliteit in gel-MOD kan ook afhankelijk zijn van het gebruikte celtype. Binnen de
vakgroep Medische Basiswetenschappen (Universiteit Gent) werden al verschillende cellijnen
(MC3T3’s, UMRGFP’s en HepG2’s) ingekapseld in gel-MOD. Enkel de HepG2-cellijn gaf
aanleiding tot een relatief hoge viabiliteit. In deze masterproef werd gebruik gemaakt van
primaire cellen, die mogelijk nog minder robuust zijn dan cellijnen.
Ondanks de lage viabiliteit is, gezien de lage kostprijs en relatief eenvoudige
productiemethode, verder onderzoek met gelatine hydrogels aangewezen. [28] Naast het

42
reduceren van de uv-belichtingstijd en het zoeken naar ‘radicaalscavangers’ kan men ook
proberen de 30 minuten incubatie in de koelkast te verminderen. Door deze wachttijd te
reduceren, vermindert de contacttijd tussen de niet-gecrosslinkte gel-MOD met Ig en de
cellen. Als men de viabiliteit op dag 1 van niet-uv bestraalde cellen in gel-MOD (27,72 ±
3,12) vergelijkt met de cellen in suspensie die collagenase behandeling ondergaan (84,19 ±
16,73), ziet men een opmerkelijk verschil. Deze resultaten suggereren dat de niet-
gecrosslinkte gel-MOD met Ig een mogelijk cytotoxisch effect heeft.
Er werden experimenten met een HFF-cellijn op coatings uitgevoerd om een eerste indicatie
te krijgen van de celadhesie op verschillende materialen. Deze adhesie werd bestudeerd met
behulp van PCL coatings met verschillende modificaties.
De coatingexperimenten toonden dat de cellen niet vasthechten op de PCL coating, noch na 4
uur noch na 24 uur. De cellen uitgezaaid op een PCL-AEMA coating begonnen na 4 uur vast
te hechten. Dit is te zien aan de vorming van focale adhesiepunten en het cytoskelet. Na 24
uur op PCL-AEMA coating waren de cellen volledig vastgehecht, dit ziet men aan de
uitgestrekte morfologie. Op een PCL-GelB coating waren de cellen na 4 uur al vastgehecht.
Op de PCL-GelB Fn coating waren het actine cytoskelet en focale adhesiepunten al duidelijk
te zien na 4 uur, wat op een goede aanhechting van de cellen wijst. Deze goede vasthechting
is te wijten aan de fibronectine-eiwitten die celadhesie bevorderen. [22] Op basis van de
resultaten met coatings werd ook een selectie gemaakt voor de gebruikte scaffolds.
Op de ‘live/dead’ kleuring van PCL scaffolds (fig.16) en PCL-O scaffolds (fig.17) is te zien
dat cellen na 7 dagen in FC medium in clusters vasthechten en daarna verder uitgroeien. Deze
clusters worden gevormd omdat het materiaal hydrofoob is en weinig celadhesie toelaat. Bij
de PCL-GelB Fn scaffold (fig.18) is op dag 7 al een meer homogene spreiding van de cellen
te zien. De oorzaak voor dit verschil in aanhechting ligt waarschijnlijk aan het feit dat
fibronectine een eiwit is dat celadhesie stimuleert. [22] Op de ‘live/dead’ kleuring van de
PCL-O, PCL-GelB Fn en Sigma® Biotek scaffolds (fig.17,18,20) is er een toename van het
aantal cellen te zien in functie van de tijd bij kweken in FC medium. Dit komt omdat de cellen
in een geschikt medium zullen prolifereren. Op PCL scaffolds (fig.16) in FC medium is echter
een duidelijke vermindering van het celaantal na 21 dagen cultuur te zien. Dit zou kunnen
veroorzaakt worden door enerzijds de gebrekkige ECM productie van de fibrochondrocyten in
passage 2 en anderzijds het materiaaloppervlak dat geen goede celadhesie toelaat. Hierdoor
komen de cellen toch los van de hydrofobe scaffold. Op de PCL-GelB Fn scaffolds ziet men,

43
i.t.t. PCL en PCL-O scaffolds, een relatief hoog aantal cellen dat vastgehecht blijft op dag 21
in chondrogeen medium (fig.18j,k). Een oorzaak hiervoor is te vinden in de
oppervlaktemodificatie van de scaffold. De PCL-GelB Fn is dankzij de fibronectine-eiwitten
in staat om celadhesie te stimuleren, zelfs al zijn er geen celadhesie bevorderende
componenten aanwezig in het medium. Op de collageen scaffolds van BDTM
(fig.19) is er
geen zichtbare verhoging van het celaantal waar te nemen. Dit komt waarschijnlijk omdat het
oppervlak na 7 dagen cultuur al volledig bedekt is met cellen. De ‘live/dead’ kleuring geeft
enkel een beeld weer van de oppervlakkige laag en op deze manier kan dus geen verdere
celgroei worden aangetoond. Dit zou echter wel kunnen met een kwantitatieve methode zoals
MTT of met histologische coupes (fig.21). Op alle scaffolds, behalve BDTM
Collageen, is een
duidelijke afname van het celaantal na kweken in chondrogeen medium te zien.
Als men de fluorescentiefoto’s van commerciële PCL (Sigma Biotek insert) scaffolds (fig.20)
vergelijkt met de geplotte PCL scaffolds (fig.16) ziet men een duidelijk verschil. De geplotte
PCL scaffold laat minder celadhesie en celgroei toe dan de commerciële variant. Dit kan
verschillende oorzaken hebben. Om te beginnen zijn de poriën bij geplotte scaffolds groter
(±518 µm) dan deze van de commercieel beschikbare scaffold (±300 µm). Daarnaast lopen bij
de geplotte scaffolds de evenwijdige strengen recht onder mekaar. Hierdoor vallen de cellen
bij uitzaaiing makkelijker door de scaffold en kan men verwachten dat de uitzaai efficiëntie
lager is. Dit is niet het geval bij een commerciële PCL scaffold (Sigma®) waar de
evenwijdige strengen ‘in shift’ tegenover elkaar verplaatst worden in de verschillende lagen.
Dit effect zou kunnen bestudeerd worden door de oppervlaktemodificaties uit te voeren op de
PCL scaffolds van Sigma-Aldrich®. Een andere mogelijke oorzaak die dit verschil kan
verklaren, is het feit dat de commerciële PCL en geplotte PCL na synthese een verschillende
sterilisatiemethode ondergaan. De niet-commerciële scaffolds worden gesteriliseerd m.b.v.
een etyleenoxidebehandeling. Sigma-Aldrich® gebruikt γ-irradiatie als sterilisatiemethode.
Onderzoek naar het effect van γ-irradiatie op de eigenschappen van PCL scaffolds heeft
aangetoond dat er door deze methode mogelijk significante veranderingen optreden voor
celadhesie en celgroei op de scaffolds. [29]
De histologische coupes van de commerciële collageen scaffolds tonen een beperkte cellulaire
infiltratie na 21 dagen in FC medium (fig.21a,b). Er is een goede cellulaire ingroei te zien op
de niet-commerciële PCL-O scaffolds (fig.21c). Alciaanblauw kleuring op deze scaffolds
toont dat er meer ECM gevormd wordt op de collageenscaffold (fig.21d,e) dan op de PCL-O
scaffold (fig.21f). Een mogelijke verklaring hiervoor is dat fibrochondrocyten op de

44
collageenscaffold minder prolifereren en meer ECM aanmaken, terwijl ze op de PCL-O
scaffold meer zullen prolifereren om de scaffold dieper te kunnen koloniseren langs de grote
poriën.
In een eerste experiment werden fibrochondrocyten op scaffolds gezaaid in FC medium en na
1 dag overgebracht in basis, chondrogeen of FC medium. Het basis medium is een serumvrij
medium dat geen groeifactoren bevat. Het chondrogeen medium bevat TGF-β1, dat wordt
gebruikt om dedifferentiatie van fibrochondrocyten op scaffolds te voorkomen. Daarnaast is
er in het chondrogeen medium ook dexamethasone aanwezig dat de differentiatie bevordert.
In de literatuur wordt beschreven dat dexamethasone chondrogenese en initiële ECM
productie van MSC kan bevorderen. [30] De afwezigheid van adhesiefactoren in het basis en
chondrogeen medium zou het verschil in celadhesie tussen de verschillende media kunnen
verklaren. Het basis medium en chondrogeen medium bevat ITS. Dit is een
mediumsupplement dat insuline, transferrine en sodium seleniet bevat. ITS wordt vaak
gebruikt in serumvrije of serumarme media ter vervanging van de componenten die normaal
in serum aanwezig zijn. Het FC medium is een medium dat wordt gebruikt om
fibrochondrocyten te kweken. Dit medium bevat 10% FCS en een hoge concentratie AA
(172 µM), wat nodig is om de ECM vorming te stimuleren. Aan de hand van de gemaakte
‘live/dead’ kleuring kan men zien dat de cellen goed vasthechten in FC medium maar niet in
de andere media. De initiële vasthechting op deze scaffolds wordt waarschijnlijk bevorderd
door de aanwezigheid van FCS in het FC medium. Deze stelling wordt ondersteund door
bevindingen van Gunja et al. [31]. Dit effect werd ook in deze masterproef waargenomen,
wanneer de fibrochondrocyten rechtstreeks werden uitgezaaid in basis, chondrogeen of FC
medium. Bij gebruik van basis en chondrogeen medium werd geen kleuring waargenomen
met de ‘live/dead assay’ na 7 dagen. Om deze reden liet men in het volgende experiment de
cellen eerst 7 dagen vasthechten in FC medium. Daarna werden de scaffolds overgebracht in
chondrogeen of FC medium.
In veel celculturen wordt gebruik gemaakt van een hoge concentratie (10%) FCS om
celadhesie en proliferatie te stimuleren. De mogelijkheid om een cultuurmedium te gebruiken
met gedefinieerde componenten, verhoogt de veiligheid van ‘tissue engineered’ constructen
voor klinische toepassingen. Een gedefinieerd cultuurmedium zoals ‘knock-out serum
replacement’ is al met succes gebruikt bij articulair kraakbeen ‘engineering’. Ook van
serumvrij medium met ‘basic fibroblast growth factor’ (bFGF) is aangetoond dat het kan
worden gebruikt voor meniscuscellen. [30] Voor culturen op scaffolds is in de literatuur

45
aangetoond dat eliminatie van serum tijdens de fibrochondrocytenkweek de mogelijkheid van
cellen om te prolifereren en ECM te vormen significant verlaagd. Er werd een vermindering
van 60% in het celaantal per scaffold waargenomen. Dezelfde onderzoeksgroep heeft ook
aangetoond dat dit effect kan worden vermeden door TGF-β1 toe te voegen in een
concentratie van 10 ng/ml in plaats van 10% FCS. [31] In deze masterproef werd dezelfde
concentratie gebruikt. Echter, de onderzoeksgroep van Gunja et al. [31] maakte gebruik van
een andere mediumsamenstelling voor het (chondrogeen) differentiatiemedium. Er werd in de
publicatie Gunja et al. van gebruik gemaakt van chondrogeen medium met TGF-β1, maar
zonder dexamethasone. Mogelijk heeft dexamethasone een negatief effect op de proliferatie in
chondrogeen medium.
Een grote moeilijkheid aan deze reeks van experimenten met primaire fibrochondrocyten is de
beschikbaarheid van cellen. Per knie konden 10.000.000 tot 15.000.000 cellen geïsoleerd
worden. Om voldoende cellen te bekomen, moesten voor de scaffold experimenten de cellen
eerst een tweetal passages geëxpandeerd worden. Deze expansie zorgt echter voor een aantal
moeilijkheden zoals bijvoorbeeld de veranderde genexpressie [20] en het vormen van een
ECM in monolayerculturen.
Een eerste probleem na monolayerexpansie is het feit dat fibrochondrocyten door de hoge
concentratie ascorbinezuur, eens ze vastgehecht zijn, veel ECM aanmaken waardoor ze, bij
trypsiniseren, loskomen als een monolayer. Hierdoor diende men na de trypsinisatie ook een
collagenase behandeling uit te voeren. Bij P0 en P2 werd gezien dat de BDTM
Collageen
scaffolds oplosten door deze enzymbehandeling. Daarnaast bevatten de modificaties van
PCL-GelB en PCL-GelB Fn scaffolds gelatine, dat opgebouwd is uit gedenatureerd collageen.
Hierdoor zouden de modificaties verloren gaan bij het uitzaaien van collageen behandelde
cellen. Om het negatieve effect van deze collagenase behandeling op de gebruikte scaffolds in
de volgende experimenten te vermijden, werden de fibrochondrocyten na collagenase
behandeling nog 3 keer gespoeld met DMEM/F12 en gedurende 24u opnieuw in cultuur
gebracht.
Een tweede probleem is het feit dat men heeft aangetoond dat de genexpressie kan veranderen
door kweken in monolayers. Onderzoek naar het effect van monolayerexpansie op de
genexpressie van fibrochondrocyten uit de meniscus is gebeurd door de onderzoeksgroep van
Gunja et al. Zij onderzochten verschillen in genexpressie tussen primaire fibrochondrocyten
in passage 0 t.e.m. passage 4 aan de hand van qRT-PCR. Ze zagen dat de collageen type I

46
expressie steeg met het aantal passages, terwijl de collageen type II en de COMP expressie
daalden. De expressie van aggrecan veranderde niet significant. Ze probeerden ook de
veranderingen in genexpressie terug te normaliseren door gebruik te maken van coatings met
collageen type I en aggrecan. Hieruit bleek dat zowel de collageen type I coatings als de
aggrecan coatings de verhoogde genexpressie van collageen type I met meer dan 50% konden
reduceren. De coatings hadden echter, tegen alle verwachtingen in, geen effect op de
collageen type II genexpressie. De expressie van COMP werd door de collageen type I
coating opgereguleerd en kon na 3 passages opnieuw het expressieniveau van passage 0
bereiken. Voor de aggrecan coating kon niet aangetoond worden dat er significante
veranderingen waren opgetreden. Geen van beide coatings toonde een effect op de
aggrecanexpressie. Deze coatings zijn dus nog geen ideale methode om dedifferentiatie van
fibrochondrocyten tegen te gaan. [20]
Een mogelijkheid om de dedifferentiatie van fibrochondrocyten tegen te gaan is het gebruik
van staurosporine. Dit is een actine-modificerende stof die de genexpressie en synthese van de
fibrochondrocytmatrix verhoogt. Staurosporine is een proteïne kinase C (PKC) inhibitor die
verhindert dat adenosine trifosfaat (ATP) bindt op het kinase. [32] De expressie van collageen
type II kan verhoogd worden door het toedienen van TGF-β1 en IGF-1. Daarnaast heeft men
ook aangetoond dat de combinatie van TGF-β1 en FGF-2 een efficiënte expansie van
gewrichtskraakbeencellen toelaat, terwijl hun mogelijkheid tot differentiatie bewaard blijft.
[18] Deze groeifactoren bieden zeker nog mogelijkheden voor verder onderzoek.
Door tijdsgebrek werd de genexpressie in deze masterproef echter niet onderzocht. Wel kan
aan de hand van de vergelijking van de coupes van pellets in P0 en P2 gezegd worden dat de
fibrochondrocyten in P0 in staat zijn om een duidelijke ECM te vormen, waar dit bij
fibrochondrocyten in P2 niet het geval is. Hieruit zou men dus besluiten dat primaire
fibrochondrocyten in pellets in staat zijn om een kraakbeenmatrix te vormen in P0 maar niet
meer in P2.
Om een vergelijking te maken tussen de viabiliteit op de verschillende scaffoldmaterialen
werd geen gebruik gemaakt van een kwantitatieve methode, maar werd enkel een vergelijking
gemaakt op basis van een ‘live/dead’ kleuring en histologie. De ‘live/dead’ kleuring heeft als
groot nadeel dat er enkel naar de oppervlakkige lagen gekeken wordt en men dus geen beeld
heeft van de ingroei in de lager gelegen zones van de scaffolds. Met coupes kan men wel
kijken naar lager gelegen gebieden in de scaffold. Bij verschillende materialen deed zich

47
echter het probleem voor dat de coupes loskwamen van de draagglaasjes bij het dehydrateren
na de kleuring.
Men zou een MTT- of MTS-test kunnen uitvoeren om de viabiliteit en proliferatie op de
scaffolds na te gaan. Dit werd in deze masterproef niet gedaan, omdat elke test dan ‘in
triplicate’ zou moeten uitgevoerd worden en er dan meer fibrochondrocyten nodig zouden zijn
dan er voorhanden waren.
Ondanks de nadelen van PCL zijn er ook een aantal voordelen. PCL heeft een degradatietijd
van meer dan 24 maand, dit laat de cellen toe om de scaffold voldoende te begroeien en ECM
te synthetiseren, alvorens de scaffold degradeert. Daarnaast is het productieproces voor PCL
ook relatief eenvoudig en niet duur en heeft het materiaal een FDA goedkeuring. [33] Om
deze redenen is het belangrijk dat verdere optimalisatie op dit type scaffolds wordt
bestudeerd.
Het uiteindelijke doel is patiënteigen cellen te gebruiken voor het herstellen van
kraakbeendefecten. Een belangrijk toekomstperspectief in dit onderzoek is het gebruik van
(humane) MSC om uit te zaaien op of in te kapselen in verschillende materialen. Men weet
welke groeifactoren belangrijk kunnen zijn voor de differentiatie van MSC naar een
fibrocartilagineuse lineage, maar het is nog onduidelijk welke combinaties van groeifactoren
optimaal zijn. [34] Naast groeifactoren kan ook mechanische belasting van MSC een
belangrijke rol spelen. De onderzoeksgroep van Connelly heeft recent aangetoond dat de
combinatie van groeifactoren en belasting door treksterkte een fibrochondrocytachtige
differentiatie van beenmerg MSC bevorderen. [35]
Een belangrijk aspect dat moet worden onderzocht, eens een volledige bioactieve meniscus
geplot is, is de mechanische sterkte van het construct. Gezien het kniegewricht onderhevig is
aan enorme belastingen, moet men er zeker van zijn dat het gekozen materiaal stevig genoeg
is om aan deze krachten te weerstaan.

48
5 Conclusies
Een eerste luik van dit onderzoek ging na of primaire cellen na inkapseling in hydrogels nog
voldoende viabel bleven. Aan de hand van de viabiliteitspercentages kan gesteld worden dat
de viabiliteit in de onderzochte gel-MOD onvoldoende hoog is voor tissue engineering
applicaties. Er moet worden gezocht naar factoren die deze viabiliteit kunnen verbeteren,
zoals kortere uv-belichtingstijd, kortere contacttijd met niet-vernette hydrogel,
‘radicaalscavangers’…
Een tweede deel van het onderzoek focuste op de viabiliteit en kolonisatie van mature
fibrochondrocyten op een 3D-scaffold. In deze masterproef is aangetoond dat mature
fibrochondrocyten zowel commerciële als niet-commerciële scaffolds kunnen koloniseren. In
de PCL-O scaffold werd gezien dat de cellen dieper ingroeiden dan in de BDTM
Collageen
scaffold.
In een laatste deel werd nagegaan of fibrochondrocyten in pelletculturen, op scaffolds en in
hydrogels kunnen differentiëren tot fibrocartilago. Hier is aangetoond dat bij pelletculturen in
P0 een duidelijke ECM gevormd wordt. Bij P2 cellen is dit echter niet meer het geval. Coupes
van fibrochondrocyten na 21 dagen in alginaat en agarose toonden de vorming van lacunes,
maar geen ECM vorming. Op de BDTM
Collageen scaffold werd een duidelijke ECM
vorming aangetoond bij cellen in P2 in FC medium maar niet in chondrogeen medium.
6 Referenties
[1] Van der Bracht H, Verdonk R, Verbruggen G, Elewaut D, Verdonk P (2007). Topics in tissue engineering 3
In: Cell-based meniscus tissue engineering.
[2] McDermott I (2010). What tissue bankers should know about the use of allograft meniscus in orthopaedics.
Cell tissue bank 11: 75-85.
[3] P Buma, NN Ramrattan, TG van Tienen, RPH Veth (2004). Tissue engineering of the meniscus.
Biomaterials 25: 1523-1532.
[4] Sweigart MA, Athanasiou KA (2001). Towards tissue engineering of the knee meniscus. Tissue engineering
7 (2): 111-129.
[5] http://medchrome.com/basic-science/the-knee-joint/
[6] http://www.emedicinehealth.com/script/main/
[7] Verdonk PCM, Forsyth RG, Wang J, Almqvist KF, Verdonk R, Veys EM, Verbruggen G (2005).
Characterisation of human knee meniscus cell phenotype. Osteoarthritis and Cartilage 13: 548-560.
[8] Gelber PE, Gonzalez G, Torres R, Giralt NG, Caceres E, Monllau JC (2009). Cryopreservation does not alter
the ultrastructure of the meniscus. Knee surgery, sports traumatology, arthroscopy 17: 639-644.

49
[9] Gelber PE, Gonzalez G, Lloreta JL, Reina F, Caceres E, Monllau JC (2008). Freezing causes changes in the
meniscus collagen net: a new ultrastructural meniscus disarray scale. Knee surgery, sports traumatology,
arthroscopy 16: 353-359.
[10] Verdonk P, Huysse W, Forsyth R, Verdonk R on behalf of the actifit study group. Implantation of a novel
synthetic scaffold for meniscus tissue regeneration. Bone-tec, Oct 8-11, 2009, Hannover.
[11] http:// www.orteq.com
[12] Rodkey WG (2010). MenaflexTM
Collagen Meniscus Implant: Basic Science. In: The meniscus. Beaufils P,
Verdonk R, editors. Berlin Heidelberg: Springer-Verslag, pp. 367-379.
[13] http://www.menaflex.com/en-us/en/procedure.php
[14] Cornelissen M, Beele H (2010). Cursus: weefselengineering. In: Culture of human embryonic stem cells for
tissue engineering.
[15] Ibarra C, Koski JA, Warren RF (2000). Tissue engineering meniscus: cells and matrix. Orthopedic Clinics
of North America 31(3).
[16] Pfister A, Launders R, Laib A, Hübner U, Schmelzeisen R, Mülhaupt R (2004). Biofunctional rapid
prototyping for tissue engineering applications: 3D bioplotting versus 3D printing. Journal of polymer science.
Part A 42: 624-638.
[17] Rijk P (2004) Meniscal allograft transplantation – part I: background, results, graft selection and
preservation, and surgical considerations. Arthroscopy 20:728–743.
[18] Forriol F (2009). Growth factors in cartilage and meniscus repair. Injury 40 (3).
[19] Zhang Z, McCaffery M, Spencer RGS, Francomano CA (2004). Hyaline cartilage engineered by
chondrocytes in pellet culture: histological, immunohistochemical and ultrastructural analysis in comparison
with cartilage explants. Journal of anatomy 204: 229-237.
[20] Gunja NJ, Athanasiou KA (2007). Passage and reversal effects on gene expression of bovine meniscal
fibrochondrocytes. Arthritis research and therapy: 9 (5).
[21] Van Vlierberghe S, Dubruel P, Schacht E (2011). Biopolymer-based hydrogels as scaffolds for tissue
engineering applications: a review. Biomacromolecules 12 (5): 1387-1408.
[22] Desmet T, Billiet T, Berneel E, Cornelissen R, Schaubroeck D, Schacht E, Dubruel P (2010). Post-plasma
grafting of AEMA as a versatile tool to biofunctionalise polyesters for tissue engineering. Macromolecular
Journals 10.
[23] Schmitz N, Laverty S, Kraus VB, Aigner T (2010). Basic methods in histopathology of joint tissues.
Osteoarthritis and cartilage 18: S113-S116.
[24] De Rijk H (1999). Histotechniek deel 5. In Kleurtechnieken histologie 7: 28-29.
[25] De Moor G, Van Maele G (2008). Continue Variabele: niet-parametrische Mann-Withney U test. In:
Inleiding tot de biomedische statistiek. Leuven: Acco, pp. 236-242.
[26] Mc Conell K, Jarman-Smith M, Stewart K, Chaudhuri JB (2004). Culture of meniscal chondrocytes on
alginate hydrogel matrices. Food and bioproducts processings 82 (C2): 126-132.
[27] Sabnis A, Rahimi M, Chapman C, Nguyen K (2009). Cytocompatibility studies of an in situ
photopolymerized thermoresponsive hydrogel nanoparticle system using human aortic smooth muscle cells.
Journal of biomedical materials research 91 (1): 52-59.

50
[28] Nichol JW, Koshy ST, Bae H, Hwang CM, Yamanlar S, Khademhosseini A (2010). Cell-laden
microengineered gelatin metacrylate hydrogels. Biomaterials 31: 5536-5544.
[29] Cottam E, Hukins DWL, Lee K, Hewitt C, Jenkins MJ (2009). Effect of sterilisation by gamma irradiation
of the ability of polycaprolactone (PCL) to act as a scaffold material. Medical engineering and physics 31: 221-
226.
[30] Heng BC, Cao T, Lee EH (2004). Directing stem cell differentiation into the chondrogenic lineage in vitro.
Stem cells 22: 1152-1167.
[31] Gunja NJ, Uthamanthil RK, Athanasiou KA (2009). Effects of TGFbeta1 and hydrostatic pressure o
meniscus cell-seeded scaffolds. Biomaterials 30: 565-573.
[32] Hoben GM, Athanasiou KA (2008). Use of staurosporine, an actin-modifying agent, to enhance
fibrochondrocyte matrix gene expression and synthesis. Cell tissue research 334: 469-476.
[34] Woodruff MA, Hutmacher DW (2010). The return of a forgotten polymer-Polycaprolactone in the 21st
century. Progress in Polymer Science 35 (10): 1217-1256.
[35] Pabbruwe MB, Kafienah W, Tarlton JF, Mistry S, Fox DJ, Hollander AP (2010). Repair of meniscal
cartilage white zone tears using a stem cell/collagen-scaffold implant. Biomaterials 31: 2583-2591.
[36] Connely JT, Vanderploeg EJ, Mouw JK, Wilson CG, Levenston ME (2010). Tensile loading modulates
bone marrow stromal cell differentiation and the development of engineered fibrocartilage constructs. Tissue
engineering part A 16 (6): 1913-1923.

1
Bijlagen
Bijlage I: Samenstelling Ringeroplossing
Voor 1 liter Ringeroplossing wordt volgende samenstelling gebruikt:
- 9 g NaCl
- 0,42 g KCl
- 0,24 g CaCl2 in 1 liter gedestilleerd water
Bijlage II: Samenstelling basis medium
- 90 ml DMEM/F12 (Gibco®, N°31330)
- 10 ml FCS (Gibco®, N°10270)
- 0,5 ml P/S (Gibco®, N° 15140, + 10.000 u/ml penicilline + 10.000 u/ml streptomycine)
- 1 ml Fungizone (Gibco®, N°15290-018, 250 µg/ml)
- 1 ml Na-pyruvaat (Gibco®, N°11360, 100 mM)
Bijlage III: Samenstelling pronase (protease) medium
- 100 ml DMEM/F12
- 0,5 ml P/S
- 0,5 ml Fungizone
- 1 ml Na-pyruvaat
Bijlage IV: Samenstelling collagenase medium
- 90 ml DMEM/F12
- 10 ml FCS
- 0,5 ml P/S
- 0,5 ml Fungizone
- 1 ml Na-pyruvaat
Bijlage V: Samenstelling fibrochondrocyten medium
- 90 ml DMEM/F12
- 10 ml FCS
- 0,5 ml Ascorbinezuur (Sigma-Aldrich®, A8960-5G) (172 µg/ml)
- 0,5 ml P/S
- 1 ml Fungizone
- 1 ml Na-pyruvaat
Bijlage VI: Samenstelling basis medium en chondrogeen medium voor differentiatie
Basis medium (8 ml)
- 8 ml DMEM/F12

2
- 8 µl ITS (BDTM
Universal Culture Supplement premix, N°354351)
- 8 µl P/S
- 80 µl Na-pyruvaat
Chondrogeen medium (4 ml)
- 4 ml Basis medium
- 80 µl Ascorbinezuur (200 µM)
- 4 µl Dexamethasone (Sigma-Aldrich®, D2915)
- 2 µl TGF-β 1 (R&D systems®, 240B002 ,20 µg/ml)
Bijlage VII: Samenstelling PBS
10 mM, pH 7,2
- 1,78 g Na2HPO4
- 0,42 g KH2PO4
- 7,2 g NaCl
- 1000 ml AD
Bijlage VIII: Samenstelling Dulbecco’s PBS
Voor 1 liter Dulbecco’s PBS wordt volgende samenstelling gebruikt:
- 8 g NaCl
- 0,20 g KCl
- 0,2 g KH2PO4
- 1,15 g Na2HPO4
- 0,133 g CaCl2.2H2O
- 0,1 g MgCl2.6H2O in 1000 ml AD bij pH 7,5
Bijlage IX: Samenstelling HFF medium
- 83 ml DMEM+GlutamaxTM
(Gibco®,N°61965 )
- 15 ml FCS
- 100 µl Na-pyruvaat
- 2 ml L-glutamine (Sigma-Aldrich®, G7513)
- 100 µl P/S

3
Bijlage X: Protocol Actine-cytoskelet focal adhesion kleuring
1. PRINCIPE
- TRITC-conjugated Phalloidin (actinekleuring)
- Anti-Vinculin (focale punten)
- DAPI (kernkleuring)
(MilliporeTM
, Actin cytoskeleton and focal adhesion staining kit, FAK100)
2. MATERIALEN
- fixatief : 4% paraformaldehyde
- permeabiliserende stof: 0,1% Triton X-100:
5 µl Triton + 4995 µl PBS
Triton tijdig uit frigo halen want visceus.
- blokkingsoplossing: 1% BSA in 1 x PBS:
5 ml PBS + 0,05 g BSA + 250 µl NGS1 + 100 µl Tween 20
- secundair AL: fluorescent gelabeld anti-muis AL
- wasbuffer: 1 x PBS
- mounting oplossing
- draag- en dekglaasjes
3. PROTOCOL
Cellijn die voor maximaal 50-60% confluent is.
Stap Duur en omstandigheden2
Fixeer in 4% paraformaldehyde in 1x PBS 20 min (0,5 ml/well)
2 x wassen met PBS 2 x 5 min
Cellen permeabiliseren 0,1% Triton X-100 in
1x PBS
5 min
2 x wassen met PBS 2 x 5 min
Blokkingsserum toevoegen 30 min
1e AL (anti-Vinculin) (1/800) in
verdunningsbuffer 3
Afdekken met aluminium folie
1 uur
3 x wassen met PBS 5 min
2e AL: TRITC-phalloïdine (1/100) + GA-
FITC (1/200) in verdunningsbuffer 4
1 uur
3 x wassen met PBS 5 min
DAPI kleuring 1/1000 5 5 min
3 x wassen met PBS 5-10 min
Foto’s
1 Normal goat serum (DakoCytomation, X0907)
2 Alle stappen worden uitgevoerd bij kamertermperatuur.
3 Vinculine: 300 µl/well: 1/800 = 1 µl vinculine + 800 µl verdunningsbuffer.
4 FITC/ TRITC: 1/200 = 3/600 = 3 µl FITC + 600 µl verdunningsbuffer + 6 µl TRITC (TRITC : 1/100)
5 DAPI: 1/1000 => 2/2000

4
Bijlage XI: Samenstelling neutraal gebufferde formol
Voor 1 liter: (4%, pH 6,9)
- 1,78 g Na2HPO4
- 0,42 g KH2PO4
- 100 ml formaldehyde (40%)
- Aanlengen met AD tot 1 liter.
Bijlage XII: Haematoxyline eosine kleuring
Deze routinekleuring wordt uitgevoerd m.b.v. het Micron ® kleurtoestel.
Stap Duur
3 x tolueen 5 min
2 x isopropanol 2 min
2 x alcohol 96% 2 min
Leidingswater 2 min
AD 1 min
Haematoxyline 15 sec
Leidingswater 2 min
Clarifier I 1 min
Leidingswater 1 min
Bluing reagent 1 min
Leidingwater 1 min
AD 1 min
Eosine+phloxine 2 min
Leidingwater 2 min
2 x alcohol 96% 2 min
2 x isopropanol 2 min
3 x tolueen 2 min
Bijlage XIII: Protocol Alciaanblauw kleuring
Alciaanblauw 1% bij pH 2,5
1) Deparaffineren
2) 30 min in Alciaanblauw6 1% in 3% ijsazijn
3) Spoelen in AD
4) Dehydrateren en monteren
5) Drogen aan de lucht
6 Sigma-Aldrich®, A5268



















