
Web view, National Center for Science ... in such changes.2 Titel van Part 3, ... its descendants to...
45
Macro evolutie http://tsjok45.multiply.com/photos/album/2028/Macro_evolutie _ inleiding tot de grondbeginsellen van de macro-evolutie- biologie Deze uitgave van het op schrift gesteld getuigenis van Dr. kevin Padian tijdens het kitzmiller proces ___ waaraan de oorspronkelijke gebruikte "power point "presentaties zijn toegevoegd ___verleent het een totaal nieuwe kracht .... Het hier voorliggend geheel is in feite een erg goede inleiding tot de grondbeginsellen van de macro-evolutie-biologie Wanneer één of andere creationist in de toekomst beweert dat hij micro-evolutie aanvaard , maar bij gebrek aan bewijs macro-evolutie voor hem niet te accepteren valt ...dan kan men hem op deze webpagina wijzen ... De uiteenzetting was oorspronkelijk aan niet- wetenschappers gericht , maar aan de rechters en de advocaten tijdens het Dover proces ; zodat hetgetuigenis bj uitmuntendheid begrijpelijk is voor om het even welke intelligente leek... en het verplettert de valse verwerping en afwijzingen van de macro-evolutie , die de creationisten zo dierbaar zijn http://www.sciohost.org/ncse/kvd/Padian/Padian_transcript.html#s006 The testimony of Kevin Padian in Kitzmiller v. Dover Edited by Nick Matzke , National Center for Science Education Version 1.0 This web page contains the testimony given by U.C. Berkeley paleontologist Kevin Padian during his appearance as an expert witness in the 2005 Kitzmiller v. Dover case. Padian is also the president of the board of directors of the National Center for Science Education . Padian's exhibit slides are included in the transcript, and are also available separately . Sections: Background (by Nick Matzke) Copyright information Expert Qualifications Direct Examination Introduction Classification, Ancestors, And Relationships Creationism and the Fossil Record I. "Irreducible Complexity" and the evolution of major adaptations II. The "Cambrian Explosion" III. How vertebrates gained land (the "fish-amphibian" transition) IV. The Origin of Birds V. Fossil Mammals The Evolution of the Ear in Mammals The Origin of Whales Creationist misrepresentations of Homology and Analogy Pandas on homology: the real wolf and Tasmanian "wolf" IDCers prefer the explanation of special creation over descent Cross Examination Cross Examination (continued) Redirect Acknowledgements Gastblog: Macroevolutie - deel 1 door renefransen op apr.28, 2011, onder Extra stof Vandaag kreeg ik van Gerdien de Jong een gastbijdrage over macroevolutie - het eerste deel in een vierluik. In veel discussies hoor ik ‘Ja, microevolutie bestaat, maar macroevolutie niet’. Ik heb Gerdien gevraagd dit thema eens uit te diepen. Macroevolutie: omschrijving.
-
Upload
nguyendien -
Category
Documents
-
view
214 -
download
0
Transcript of Web view, National Center for Science ... in such changes.2 Titel van Part 3, ... its descendants to...

Macro evolutie
http://tsjok45.multiply.com/photos/album/2028/Macro_evolutie _
inleiding tot de grondbeginsellen van de macro-evolutie-biologie Deze uitgave van het op schrift gesteld
getuigenis van Dr. kevin Padian tijdens het kitzmiller proces ___ waaraan de oorspronkelijke gebruikte "power point "presentaties zijn toegevoegd ___verleent het een totaal nieuwe kracht .... Het hier voorliggend geheel is in feite een erg goede inleiding tot de grondbeginsellen van de macro-evolutie- biologie Wanneer één of andere creationist in de toekomst beweert dat hij micro-evolutie aanvaard , maar bij gebrek aan bewijs macro-evolutie voor hem niet te accepteren valt ...dan kan men hem op deze webpagina wijzen ... De uiteenzetting was oorspronkelijk aan niet- wetenschappers gericht , maar aan de rechters en de advocaten tijdens het Dover proces ; zodat hetgetuigenis bj uitmuntendheid begrijpelijk is voor om het even welke intelligente leek... en het verplettert de valse verwerping en afwijzingen van de macro-evolutie , die de creationisten zo dierbaar zijn
http://www.sciohost.org/ncse/kvd/Padian/Padian_transcript.html#s006
The testimony of Kevin Padian in Kitzmiller v. Dover
Edited by Nick Matzke, National Center for Science EducationVersion 1.0
This web page contains the testimony given by U.C. Berkeley paleontologist Kevin Padian during his appearance as an expert witness in the 2005 Kitzmiller v. Dover case. Padian is also the president of the board of directors of the National Center for Science Education. Padian's exhibit slides are included in the transcript, and are also available separately.
Sections:Background (by Nick Matzke)Copyright informationExpert QualificationsDirect Examination Introduction Classification, Ancestors, And Relationships Creationism and the Fossil Record I. "Irreducible Complexity" and the evolution of major adaptations II. The "Cambrian Explosion" III. How vertebrates gained land (the "fish-amphibian" transition) IV. The Origin of Birds V. Fossil Mammals The Evolution of the Ear in Mammals The Origin of Whales Creationist misrepresentations of Homology and Analogy Pandas on homology: the real wolf and Tasmanian "wolf" IDCers prefer the explanation of special creation over descentCross Examination Cross Examination (continued)RedirectAcknowledgements
Gastblog: Macroevolutie - deel 1
door renefransen op apr.28, 2011, onder Extra stof
Vandaag kreeg ik van Gerdien de Jong een gastbijdrage over macroevolutie - het eerste deel in een vierluik.
In veel discussies hoor ik ‘Ja, microevolutie bestaat, maar macroevolutie niet’. Ik heb Gerdien gevraagd dit thema eens uit te diepen.
Macroevolutie: omschrijving.
Een constante bewering van creationisme is dat microevolutie bewezen is maar macroevolutie niet. Microevolutie kan direct waargenomen worden, dus dat is moeilijk te ontkennen. Macroevolutie?Eerst een post over wat het woord macroevolutie betekent (hier), dan als tweede over de grote patronen in de evolutie, als derde over de grote processen in de evolutie; en daarna wordt een samenvatting onvermijdelijk.
Wat wordt onder ‘macroevolutie’ verstaan?
I De verschillende studieboeken geven verschillende definities van macroevolutie, en gebruiken het word verschillend.
Minkoff, E.C., 1984. Evolutionary Biology. Addison-Wesley Publishing Company.1 Definitie in Glossary: Macroevolution: Evolution above the species level2 Titel van Unit 3, met onder andere de volgende hoofdstukonderwerpen: Geologic record, classification, biogeography, extinction, rates of evolution.
Skelton, P. (editor) 1993. Evolution: a biological and palaeontological approach. Addison-Wesley Publishing Company.1 Definitie in Glossary: Macroevolution: Evolution above the species level; that is, the patterns of change in the forms of organisms and in their diversity arising from the appearance and disappearance of species, and the processes involved in such changes.2 Titel van Part 3, met onder andere de volgende hoofdstukonderwerpen: Evolutionary history, fossilization, geography, evolution of form, phylogenetic patterns.

Stearns, S.C. & Hoekstra, R.F., 2000. Evolution: an introduction. (2nd edition) Oxford Unversity Press.1 Definitie in Glossary: Macroevolution: The pattern of evolution at and above the species level, including most of fossil history and much of systematics.2 Part 3 heeft als titel Principles of Macroevolution, met de volgende hoofdstukonderwerpen: Speciation, phylogeny and systematics, compararative methods.
Futuyma, D.J., 1993. Evolutionary Biology (3rd edition). Sinauer.1 Definitie in Glossary: Macroevolution: a vague term for the evolution of great phenotypic changes, usually great enough to allocate the changed lineage and its descendants to a distinct genus or higher taxon.2 Part 5 heeft als title: Macroevolution: evolution above the species level, met de volgende hoofdstukonderwerpen: Development and evolution, Pattern and process in macroevolution, Evolution of biological diversity.
Futuyma, D.J., 2005. Evolution. Sinauer.1 Definitie in Glossary: Macroevolution: a vague term, usually meaning the evolution of substantial phenotypic changes, usually great enough to place the changed lineage and its descendants to a distinct genus or higher taxon.2 Chapter 21: Macroevolution: evolution above the species level.
Freeman, S. & Herron, J.C., 2007. Evolutionary Analysis. (4th edition), Pearson.1 Definitie in Glossary: Macroevolution: large evolutionary change, usually in morphology; typically refers to the evolution of differences among populations that would warrant their placement in different genera of higher level taxa.2 Geen Unit of hoofdstuk, alleen een paragraaftitel: 18.3: Macroevolutonary patterns.
Ridley, M. 2004. Evolution (3rd edition). Blackwell Publishing.1 Definitie in Glossary: Macroevolution: Evolution on the grand scale: the term refers to events above the species level. The origin of a new higher group, such as the vertebrates, would be an example of a macroevolutionary event.2 Part 5 heeft als title: Macroevolution met de volgende hoofdstukonderwerpen: The history of life, evolutionary genomics, evolutionary developmental biology, rates of evolution, coevolution, extinction, radiation.
Hall, B.K. & Hallgrimsson, B, 2008. Strickberger’s Evolution. (4th edition). Jones & Bartlett.1 Definitie in Glossary: Macroevolution: The pattern of evolution at and above the level of the species. Fossils are our chief evidence for macroevolution.2 Niet als hoofdstuk titel oid..
Barton, N.H., Briggs, D.E.G., Eisen, J.A., Goldstein, D.B. & Patel, N.H., 2007. Evolution. CSHL Press.1 Definitie in Glossary: Macroevolution: evolutionary change at or above the of the species level.2 Niet als hoofdstuk titel oid. In de index wordt onder macroevolution verwezen naar de fossil record.
Al met al blijkt dat in de studieboeken evolutiebiologie macroevolutie betekent: evolutie boven het niveau van de soort. Het woord macroevolutie wordt vaak gebruikt voor evolutiepatronen, vooral de patronen die de fossielen laten zien. Het woord is weinig specifiek, handig voor titel van grote afdelingen in een boek, maar verder niet. Macroevolutie is geen technische term in de biologie, maar een van die losse woorden die handig zijn om te gebruiken en waarvan iedereen wel ongeveer weet wat het zou betekenen. Iets met ‘groot’ en ‘evolutie’, waarbij ‘groot’ inhoudt: op grote schaal.
II Voor de creationistische positie gebruik ik de vertaling van het boek van Junker & Scherer:Junker, R. & Scherer, S., 2010. Evolutie – het nieuwe studieboek. Stichting De Oude Wereld.
1 In het glossarium staat: macro-evolutie: het ontstaan van nieuwe organen, structuren of typen bouwplan, ontstaan van kwalitatief nieuwe genen. (schuin in origineel).
2 De titel van onderdeel 4.3 is: Micro- en macro-evolutie. Daar wordt macroevolutie omschreven als:“Macro-evolutie: ontstaan van nieuwe, niet eerder aanwezige organen, structuren of bouwplantypen; daaraan gerelateerd ook het ontstaan van kwalitatief nieuw genetisch materiaal. Het begrip “opwaartse ontwikkeling” (anagenese) heeft in dit boek ook deze betekenis (afbeelding 4.8).” (schuin in origineel).
Afbeelding 4.8 (gedownload van de Duitse site met toestemming dr. R. Junker)
De titel van hoofdstuk 6 is: Macro-evolutie. Dit gaat over wat de schrijvers zien als eventuele mechanismen voor macro-evolutie; microevolutie + tijd wordt niet voldoende geacht om grote verschillen op te leveren.
III Het zal duidelijk zijn dat de omschrijving van macroevolutie in Junker & Scherer wel iets maar niet veel met de omschrijving van macroevolutie in evolutiebiologie van doen heeft.
Wat punten bij de omschrijving van macroevolutie als in onderdeel 4.3 van Junker & Scherer.
1 ‘Anagenese’ betekent niet ‘opwaartse ontwikkeling’. Anagenese (dat niet in hun glossarium voorkomt) betekent geleidelijke morfologische verandering binnen een (fossiele) soort, zonder soortsplitsing. Het hele idee ‘opwaarts’ bestaat niet binnen de evolutiebiologie.2 In figuur 4.8 links is geen evolutie weergegeven, maar een ouder en lang verlaten idee, de scala naturae. Of zouden Junker en Scherer werkelijk denken dat trilobieten (trilobieten zijn geleedpotigen, de club waarbij ook kreeften en insecten behoren) van kwallen afstammen, en gewervelde dieren van trilobieten?
3 Bouwplan staat wel in de definitie van macroevolutie die Junker & Scherer geeft, maar komt niet in glossarium of index voor. Daarmee is niet bekend wat onder ‘bouwplan’ verstaan wordt. Hebben honden en katten een ander bouwplan? Nee? Er is een technische term ‘Bauplan’: die wijst op het verschil in bouw tussen bijvoorbeeld het fylum slakken, het fylum geleedpotigen, of het fylum gewervelden. Een Bauplan hoort bij een fylum, een hoofdafdeling. Het woord ‘fylum’ komt niet in Junker & Scherer voor.

4 Honden en katten zijn volgens Junker & Scherer verschillende basistypen: en basistypen mogen van J&S geen gemeenschappelijke voorouder hebben. Het verschil tussen honden en katten is desondanks geen macroevolutie: tenminste, niets in de definitie van macroevolutie als in Junker & Scherer is toepasbaar op honden en katten: zelfde bouwplan, zelfde organen, zelfde genen. Volgens de Junker & Scherer definitie van macroevolutie zouden honden en katten een gemeenschappelijke voorouder mogen hebben.
Nu komt de joker te voorschijn: macroevolutie gaat over gemeenschappelijke afstamming van alle levende organismen. Dat is waar creationisten bezwaar tegen hebben.
Gastblog macroevolutie - 2
mei.17, 2011, door Gerdien de Jong.
Vergeet evolutie. Kijk naar de organismen .
Het eerste wat de biologie doet is indelen, tenminste, van die organismen die bekend zijn. Hoe? In hoeveel groepen worden schelvis, koolvis, witvis, walvis, bruinvis ingedeeld? Waterbeesten, één groep? Zoet- of zoutwaterbeesten, twee groepen? Nee, drie vissensoorten, twee zoogdiersoorten. De uiterlijke min of meer gestroomlijnde vorm van walvis en bruinvis houdt verband met hun levenswijze. Op levenswijze wordt niet ingedeeld, wel op diepe overeenkomst in anatomie. Daarom worden walvissen, bruinvissen samen met de vleermuizen ingedeeld.
DEEL I
De diepste overeenkomst in anatomie wordt gegeven door de basenvolgorde in het DNA. Die basenvolgorde heeft niet of heel erg weinig met de levenswijze van een soort te maken, en kan daarom goed gebruikt worden voor het maken van indelingen. Mutaties van basen zijn grotendeels selectief neutraal, en van de afwezigheid van selectie wordt gebruik gemaakt. De indeling is op overeenkomst van basen in het DNA.. Er zijn een aantal methoden om indelingen te maken op DNA basenvolgorden; bijvoorbeeld de zuinigste manier van indelen. De methoden leveren een indeling die soorten die het meest op elkaar lijken op volgorde groepeert. Indelingen worden weergegeven in een boomstructuur.
Zo’n indelingsboom ziet er bijvoorbeeld zo uit:
Figuur 1:Boven: Soort A wordt samen met soort B in een groep ingedeeld. De soorten A en B heten zustersoorten. Soorten A en B staan even ver van soort C.
Onder: Soort C is de zustergroep van de groep met de soorten A en B. Soort C wordt met (A, B) in een groep ingedeeld. Soort D staat even ver van de soorten A, B, en C – soort D is de zustergroep van de groep (C, (A, B)).
In zo’n boomstructuur mag je alleen langs de lijntjes lopen: verwissel niet de lay-out met de indeling!
Figuur 2: hier staat precies hetzelfde als in figuur 1, het is alleen een andere lay-out. De lay-out is niet belangrijk.

Figuur 3. Links: De soorten A, B en C behoren tot de groep die ingedeeld gaat worden. Soort D behoort tot een andere groep, dat is bekend, en wordt als buitengroep gebruikt. Dit zet een ongeordende indeling (links boven) om in een geordende indeling met een leesbare layout (linksonder). Rechts. De indeling is hierarchisch: de groep gevormd door soorten Aen B valt binnen de groep gevormd door de soorten C en (A,B).
In figuur 3 staat een hiërarchische indeling. De groep gevormd door soorten A en B behoort binnen de groep gevormd door de soorten (A,B) en C. Een hiërarchische indeling geeft aan wat de relatie van alle soorten tot elkaar is. A lijkt meer op B dan op C, C lijkt evenveel op A als op B.
Er komt nu een moleculaire indeling van het leven, op volgorde met steeds wijdere groepen.
Eerst komt de moleculaire indeling van de katten.:
Figuur 4. Moleculaire indeling van de kattensoorten binnen de familie katachtigen, niet-kat roofdieren als buitengroep (Johnson et al 2006)
Dit is de figuur zoals in de publicatie, en daarom staan er ook de splitsingstijden in. Vergeet die splitsingstijden nu maar, het gaat alleen om de indeling. Het is een indeling op volgorde: huiskatgroep als zustergroep van de luipaardkatgroep, die twee samen hebben de puma-cheetahgroep als zustergroep, en zo verder. De katten worden ingedeeld in acht groepen; tenslotte de Grote Katten met een groep van zeven groepen, de Kleine Katten, als zustergroep.
Dat waren dan de katten. Daarna komt er een indeling van alle roofdieren. Daarbij is het niet nodig alle katten weer in de studie op te nemen, vijf soorten katten is genoeg, beetje verspreid over de groepen. Vergeet weer die splitsingstijden, het gaat alleen om de indeling. Eerst het origineel, dan in schema met Nederlandse namen.

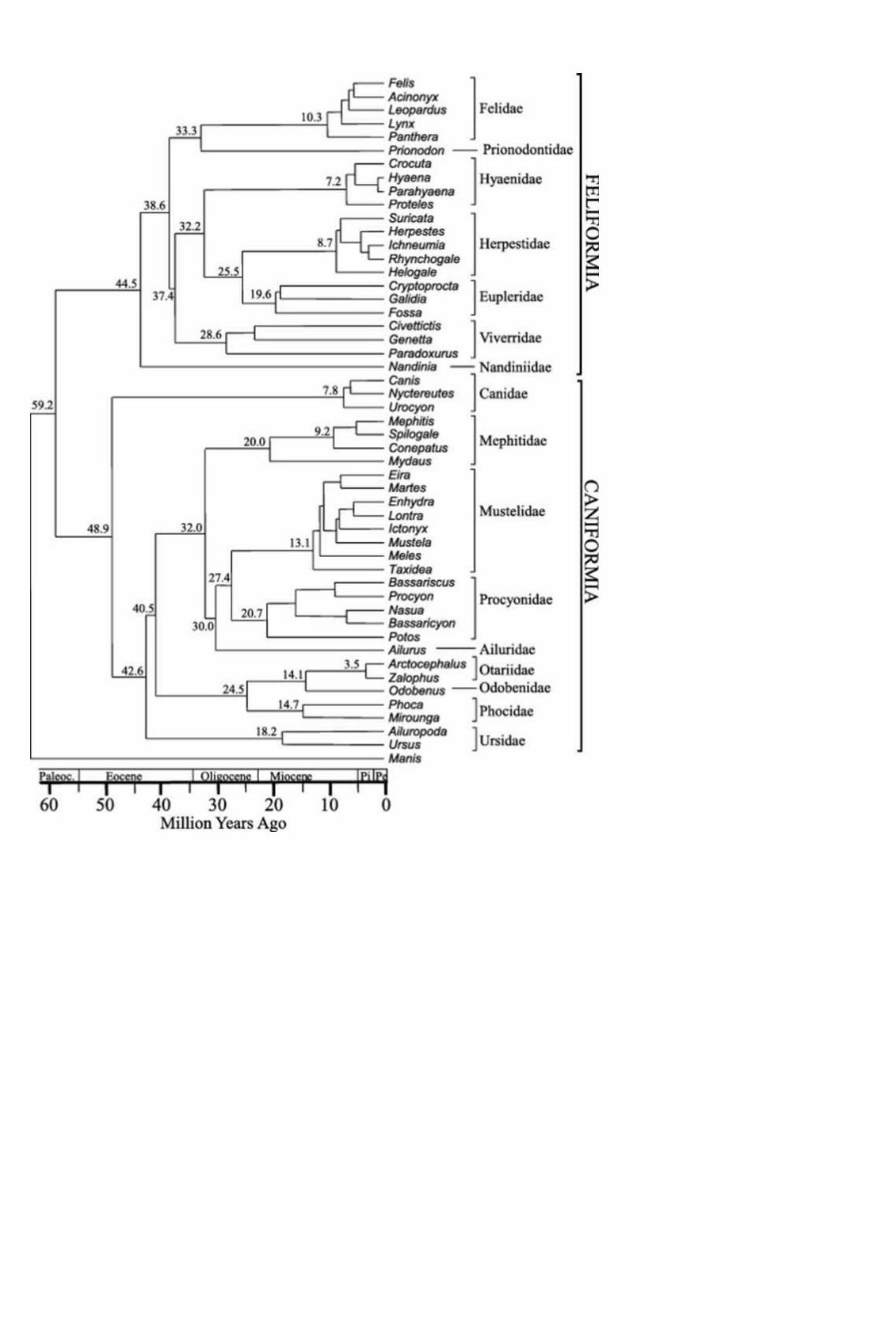
Figuur 5. Moleculaire indeling van de families binnen de orde roofdieren, schubdier als buitengroep (Eizirik et al 2010). Beneden is in schema maar hetzelfde als boven, alleen zijn de gebruikte soorten vervangen door de Nederlandse namen per familie.
De Hondvormigen en Katvormigen zijn de twee groepen binnen de roofdieren. De schubdieren zijn de zustergroep van alle roofdieren.
Wat betekent zo’n boom? Moleculair hebben de (stinkdieren, (rode panda, (wasberen, marters))) iets in hun DNA basenvolgorde gemeen, maar waarin zij allen samen verschillen van de zeeroofdieren. De (beren, (zeeroofdieren, (stinkdieren, (rode panda, (wasberen, marters))))) hebben iets in hun DNA basenvolgorde gemeen waarin zij verschillen van de honden. De Hondvormigen en de Katvormigen verschillen in hun DNA basenvolgorden – maar lijken meer op elkaar dan een van deze groepen op de schubdieren. Onderling komen de Katvormigen meer overeen dan elk van deze met een Hondvormige, en andersom. Elke Katvormige lijkt evenveel op elke Hondvormige.
De roofdieren zijn een van de ordes van de placentale zoogdieren. Over alle placentale zoogdieren lijkt de beste moleculaire indeling:
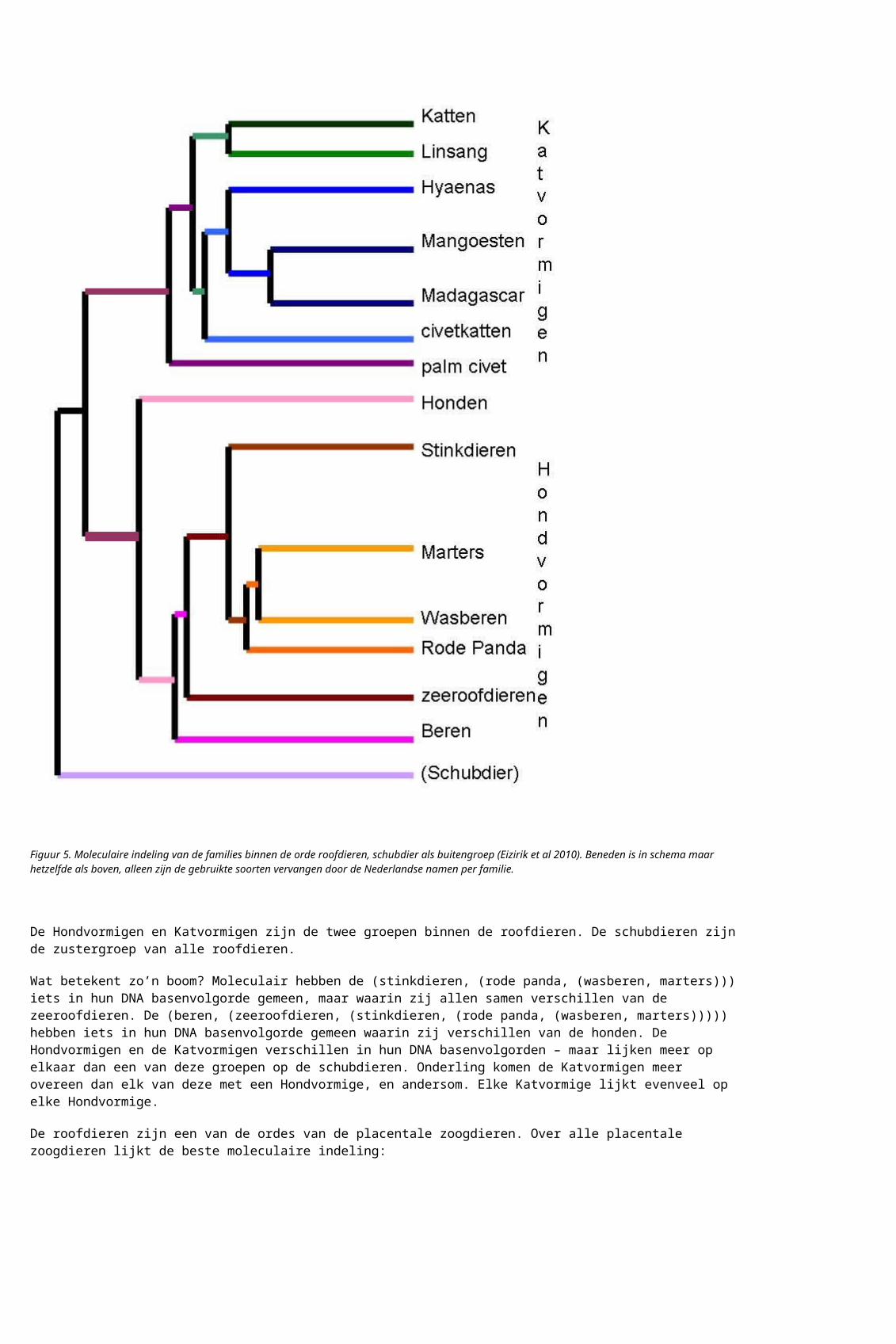
Figuur 6. Moleculaire indeling van de ordes binnen placentale zoogdieren, buideldier als buitengroep. De figuurtjes zijn uit Nishihara et al PNAS 2006; ik heb de indeling iets gewijzigd naar andere, eerdere en latere studies. Niet elke studie heeft evenveel resolutie.
De hierarchische moleculaire indeling wordt voortgezet. De placentale zoogdieren en de buideldieren zijn zustergroepen, en staan moleculair even ver van de eierleggende zoogdieren.
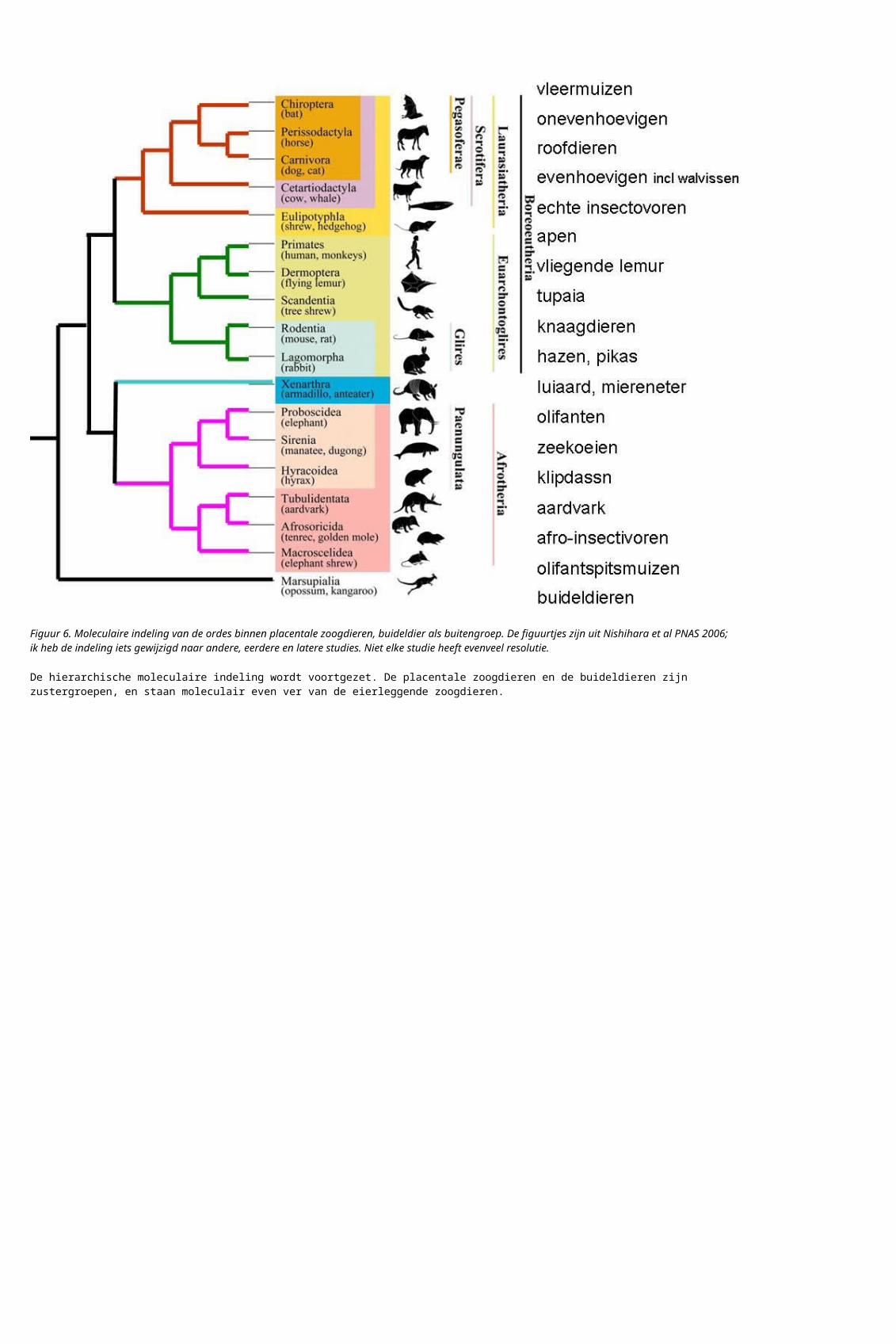
Figuur 7 Moleculaire indeling van de verschillen onderklassen zoogdieren, met andere landvertebraten als buitengroep. Figuur uit Van Rheede et al 2006, boven origineel, beneden schema.
Nu alle zoogdieren langsgeweest zijn komen de vierpotigen, de landvertebraten + amfibieën, aan de beurt. De indeling van de landvertebraten (zonder schilpad) is een van de eerste die met moleculaire methoden gedaan is – in feite, de moleculaire indeling van de vertebraten gaf omstreeks 1975 het idee dat dit wel eens zou kunnen gaan werken.
Figuur 8 Moleculaire indeling van de vierpotigen (twee studies gegeven in Zardoya & Meyer 2001), met ‘vissen’ als buitengroep. Rechts schema. Klassieke indelingen op morfologie zijn duidelijk herkenbaar.
De drie longvissen en de twee soorten coelacanth staan binnen de huidige gewervelde dieren dichter bij de landvertebraten dan alle echte vissen. De vissen zijn de grootste groep gewervelde beesten, met zelf een duidelijke indeling.
De gewervelde dieren behoren met lancetvisje (Cephalochordaten) en de tunicaten (Urochordaten) tot de Chordaten, de beesten met een chorda – waar de wervelkolom om aanlegd wordt bij gewervelde dieren. De Chordaten en de Ambulacraria vormen samen de Deuterostomia. De stekelhuidigen (zeesterren en zo, Echinodermata) samen met de Hemichordata (eikelwormen en zo) vormen de Ambulacraria.
Hier geef ik twee moleculaire indelingen van de Deuterostomia, uit een studie van Blair & Hedges 2005 en uit het artikel met het genoom van het lancetvisje (Putnam et al 2008)
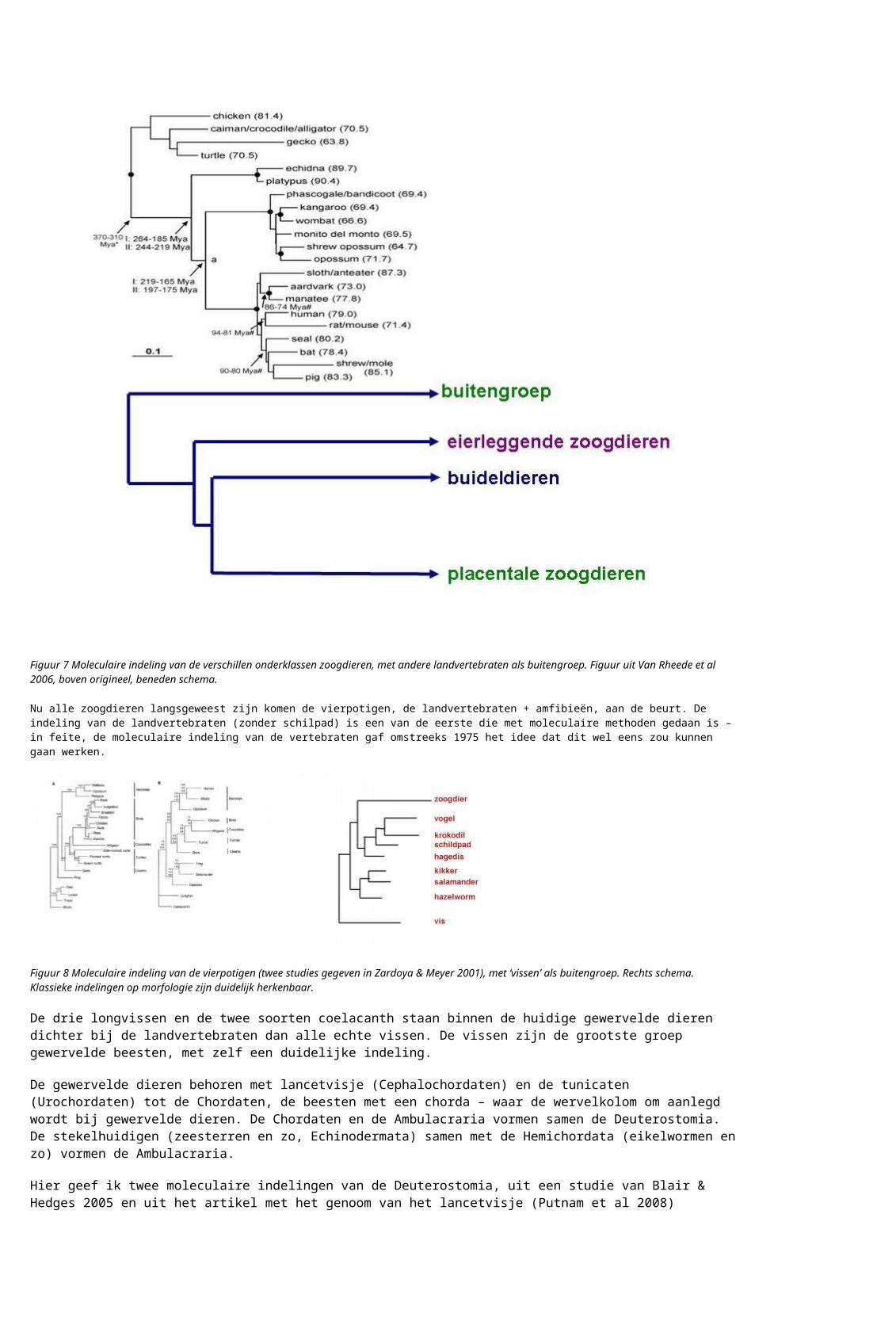
Figuur 9 Moleculaire indeling van de Deuterostomia. Chordaten bij knoop i, Ambulacraria bij knoop l. De Urochordaten (zakpijpen , tunicaten) staan dichter bij de gewervelden (knoop g) dan het lancetvisje (de Cephalochordaat). Hier is geen buitengroep in beeld.
Figuur 10 Moleculaire indeling van de Deuterostomia, met zeeanemoon en fruitvlieg als buitengroep. Ambulacraria zijn de eikelworm en zeeegel. De Urochordaten (larvaceae, sea squirt)) staan dichter bij de gewervelden (benedenste groep) dan het lancetvisje (Amphioxus), ondanks de grote veranderingen in de Urochordaten. Putnam et al 2008.
Nu is het de beurt aan alle beesten:
Figuur 11. Schema van de moleculaire indeling van het dierenrijk, uit Raible et al 2005. Rechts zoals uit Junker & Scherer, 2010, figuur 9.5. Acoela zijn platte zeebeestjes zonder lichaamsholte.
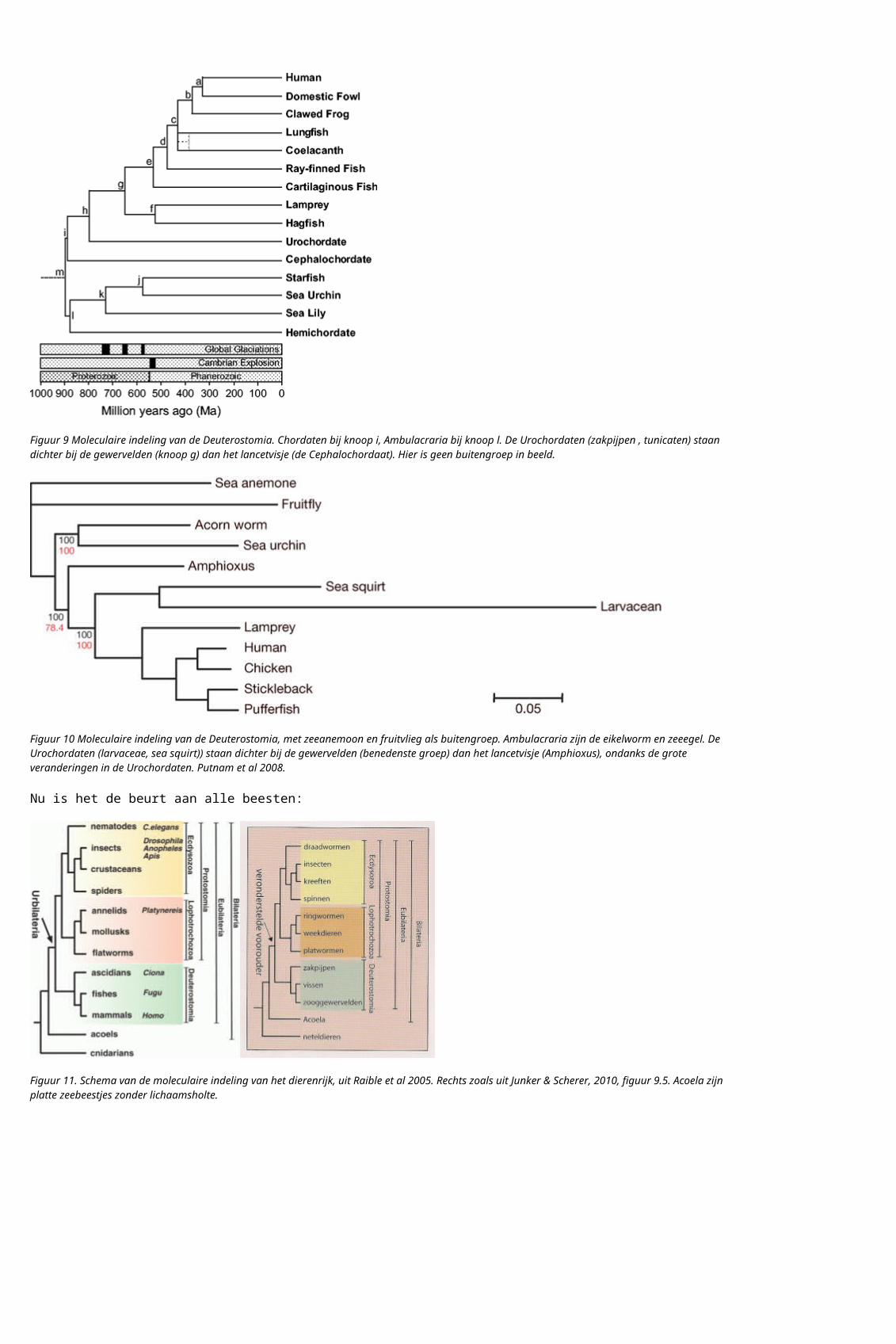
Figuur 12. Schema van de moleculaire indeling van het dierenrijk, met schimmels en planten als buitengroep, Adoutte et al 2000.
De beesten, de planten en de schimmels zijn meercellige organismen met een celkern. Meercellig doet er niet toe volgens de moleculaire indeling, de celkern wel. Er zijn veel eencelligen met een celkern, met een veel groter diversiteit dan de multicellulaire organismen met een celkern. De moleculaire indeling van alles met een celkern is als volgt:
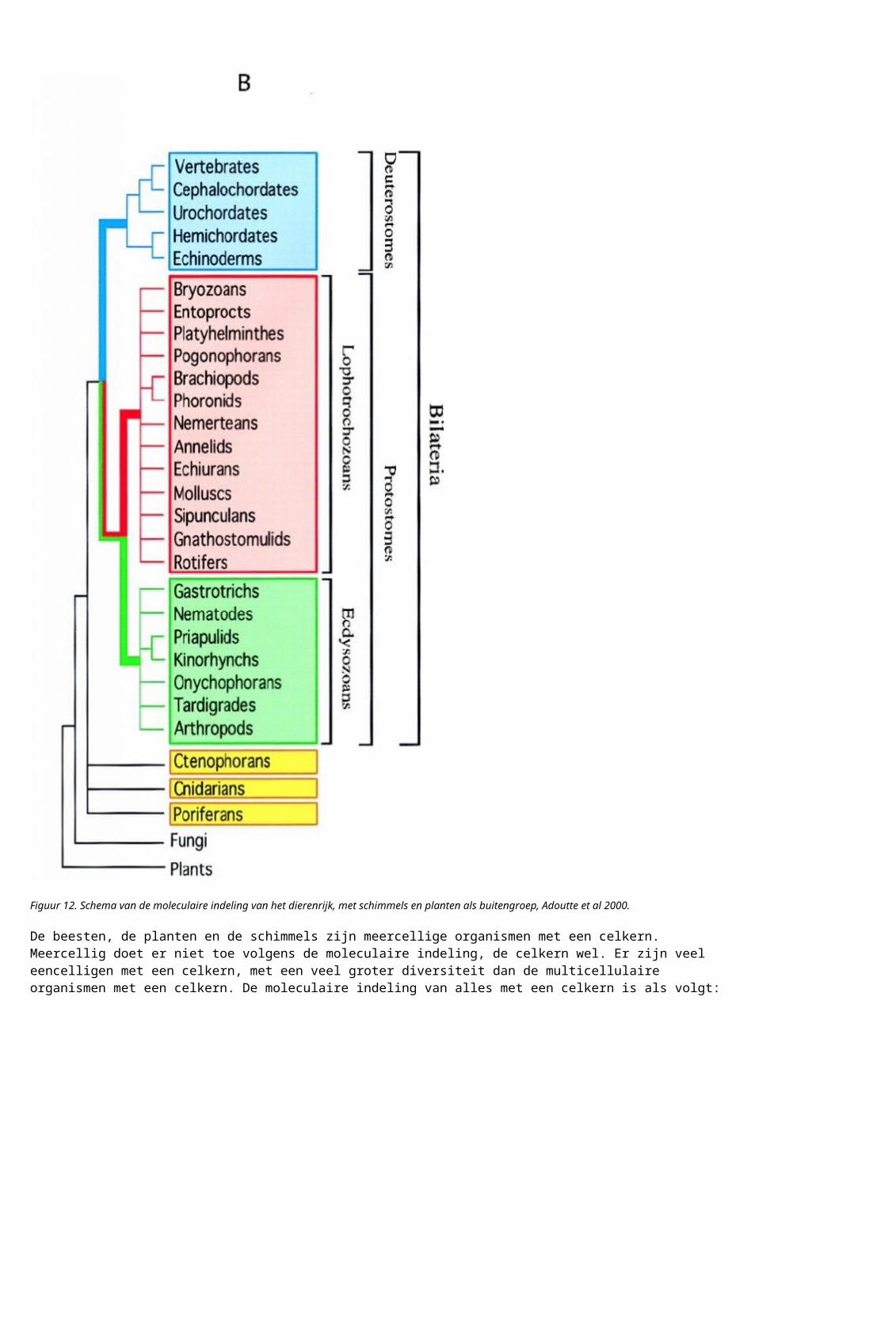
Figuur 13. Schema van de moleculaire indeling van de organismen met een celkern, uit Desmond et al 2011. De beesten staan erin als Metazoa, links. LECA staat voor Last Eukaryote Common Ancestor, hoewel het over een indeling gaat. De buitengroep zijn is hier de organismen zonder echte celkern – en die staan op de LECA plaats.
Naast organismen met een celkern zijn er twee groepen organismen zonder echte celkern, de Bacterian en de Archaea.
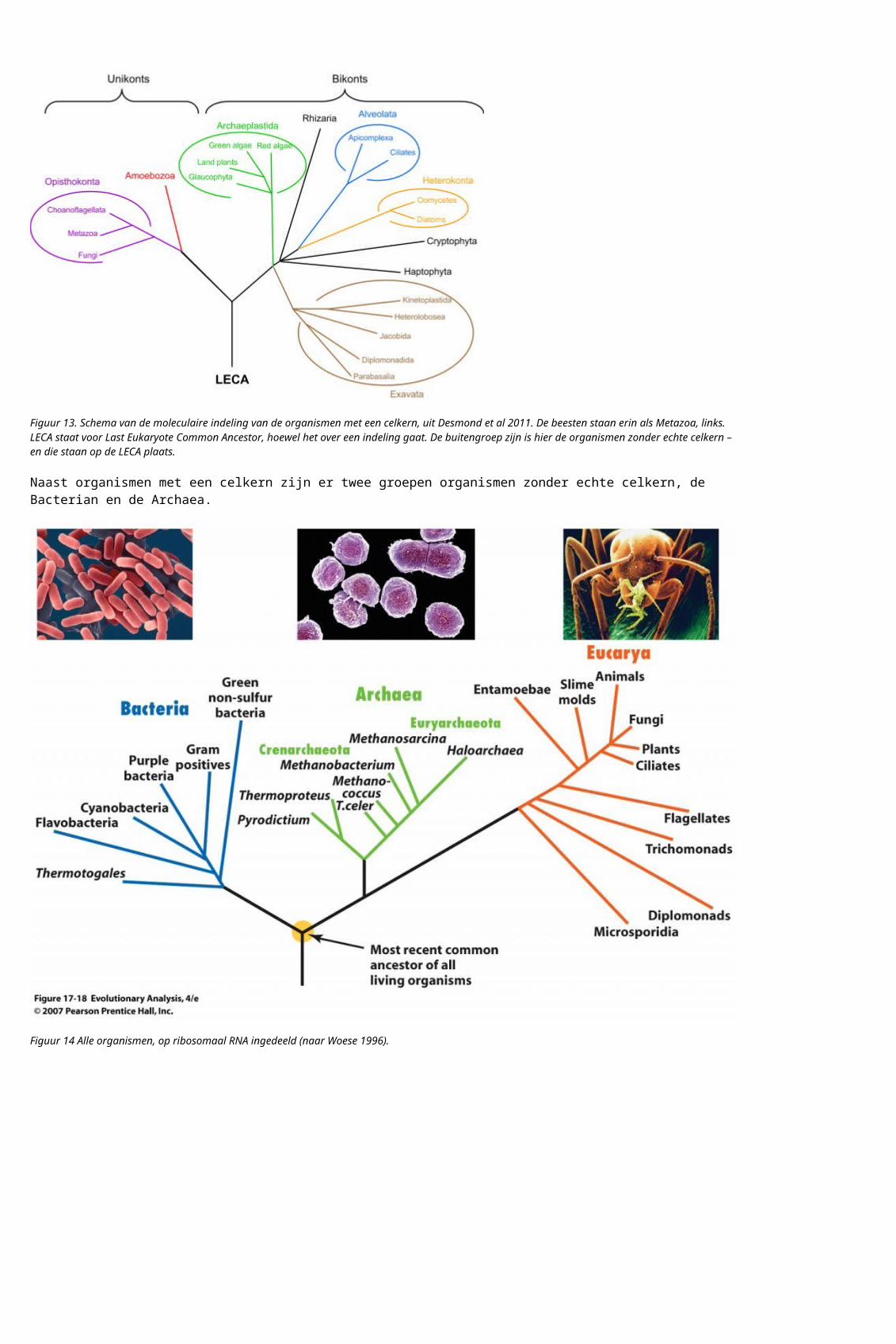
Figuur 14 Alle organismen, op ribosomaal RNA ingedeeld (naar Woese 1996).
Figuur 15. Alle organismen ingedeeld op 31 universele genen (naar Ciccarelli et al 2006).
Vanaf de katten in figuur 4 tot en met alle organismen in figuur 15 bestaat er een hiërarchische moleculaire indeling.
Werkt dit nu altijd zo goed? Waar zitten er moeilijkheden? Na 35 jaar gecumuleerde ervaring van velen zijn veel van de bekende voetangels en klemmen in de methoden wel te ontwijken. De verbetering zit in betere statistische methoden voor indelen, grotere computers en snellere methoden voor sequencen van DNA.
Veel soorten met weinig DNA lengte geeft fouten. Ongebalanceerd aantal soorten over wat wel groepen zullen zijn geeft geen resolutie voor een groep met heel laag aantal gebruikte soorten.
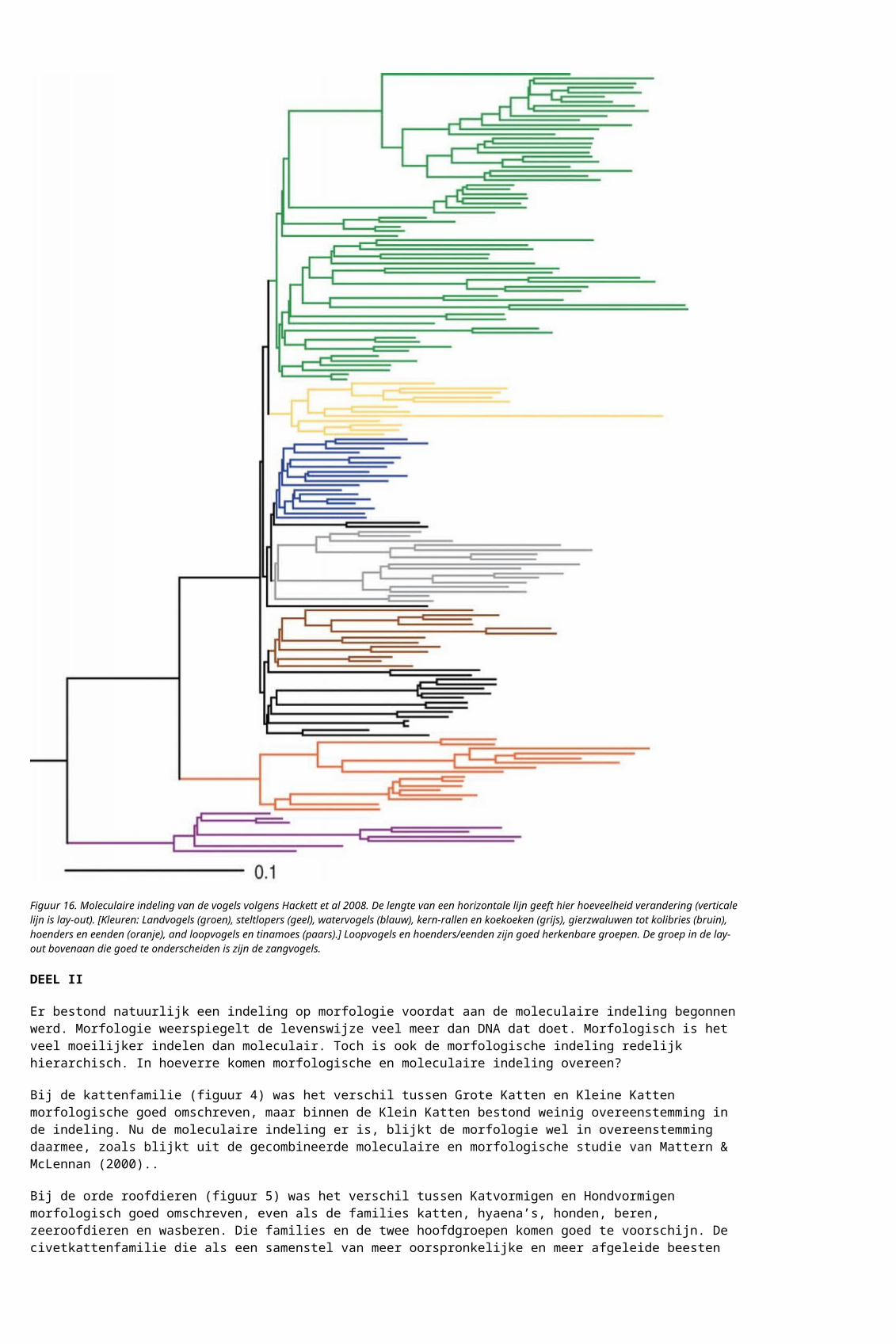
Sommige problemen zien er uit als onoplosbaar. Een diepe vertakking met heel weinig soorten aan de ene kant en veel soorten aan de andere kant is lastig, heeft weinig oplossend vermogen aan de kant met weinig soorten. Een tweede probleem is als er veel moleculaire divergentie heel dicht op elkaar zit. De vogels hebben daar last van. Er is overeenstemming over een deel van de moleculaire indeling, maar sommige families veranderen van plaats tussen studies.

Figuur 16. Moleculaire indeling van de vogels volgens Hackett et al 2008. De lengte van een horizontale lijn geeft hier hoeveelheid verandering (verticale lijn is lay-out). [Kleuren: Landvogels (groen), steltlopers (geel), watervogels (blauw), kern-rallen en koekoeken (grijs), gierzwaluwen tot kolibries (bruin), hoenders en eenden (oranje), and loopvogels en tinamoes (paars).] Loopvogels en hoenders/eenden zijn goed herkenbare groepen. De groep in de lay-out bovenaan die goed te onderscheiden is zijn de zangvogels.
DEEL II
Er bestond natuurlijk een indeling op morfologie voordat aan de moleculaire indeling begonnen werd. Morfologie weerspiegelt de levenswijze veel meer dan DNA dat doet. Morfologisch is het veel moeilijker indelen dan moleculair. Toch is ook de morfologische indeling redelijk hierarchisch. In hoeverre komen morfologische en moleculaire indeling overeen?
Bij de kattenfamilie (figuur 4) was het verschil tussen Grote Katten en Kleine Katten morfologische goed omschreven, maar binnen de Klein Katten bestond weinig overeenstemming in de indeling. Nu de moleculaire indeling er is, blijkt de morfologie wel in overeenstemming daarmee, zoals blijkt uit de gecombineerde moleculaire en morfologische studie van Mattern & McLennan (2000)..
Bij de orde roofdieren (figuur 5) was het verschil tussen Katvormigen en Hondvormigen morfologisch goed omschreven, even als de families katten, hyaena’s, honden, beren, zeeroofdieren en wasberen. Die families en de twee hoofdgroepen komen goed te voorschijn. De civetkattenfamilie die als een samenstel van meer oorspronkelijke en meer afgeleide beesten werd gezien is opgesplitst. De stinkdieren zijn van de marters afgesplitst. De rode panda bleef moleculair alleen, in zijn eigen familie, na langdurige morfologische discussie over zijn plaats .
De ordes van de placentale zoogdiern waren morfologisch niet allemaal te groeperen. De indeling van de ordes van de placentale zoogdieren (figuur 6) bracht behoorlijke verrassingen, vooral omdat drie van de vier hoofdgroepen morfologisch nooit voorgesteld waren, en moeilijk morfologisch te karakteriseren zijn. Moleculair is het signaal voor de hoofdgroepen sterk. Sommige groeperingen van ordes tot zusterordes die voorgesteld waren komen ook moleculair terug, maar andere morfologische voorstellen bleken geen moleculaire tegenhanger te hebben. De vleermuizen waren bijvoorbeeld ongeveer naast de apen gezet, omdat ze naar voren gerichte ogen hebben. Die
naar voren gerichte ogen moet toch met vliegen, met leefwijze, te maken hebben: de vleermuizen staan, met de insecteneters, in een andere hoofdgroep.
Bij de klasse zoogdieren (figuur 7) komt de indeling in subklassen moleculair en morfologisch overeen. tussen de placentale zoogdieren, buideldieren en eierleggende zoogdieren.
De indeling van de vertebraten in klassen en onderklassen (figuur wordt moleculair teruggevonden, met dezelfde hierarchische indeling van de landvertebraten morfologisch als moleculair. Alleen kan de schildpad minder precies geplaatst worden morfologisch dan moleculair.
De vertebraten behoorden altijd al tot het fylum Chordaten, en de Chordaten tot de Deuterostomia (figuur 9, 10). Er is een zekere verandering in de indeling binnen de Deuterostomia opgetreden: morfologisch werd altijd gedacht dat de Cephalochordaten (dat is, het lancetvisje) de zustergroep van de Chordaten zou zijn, en de Urochordaten (tunicaten, zakpijpen, sea squirt, larvaceae in figuur) de zustergroep van Chordaten + Cephalochordaten. Moleculair is dat anders: de Cephalochordaten zijn moleculair de zustergroep van Chordaten + Urochordaten, ondanks de afwijkende morfologie en levenswijze van de tunicaten.
De indeling van het dierenrijk ging op de schop. De traditionele indeling van de tweezijdig symmetrische beesten werkte met de aan/afwezigheid en bouw van de lichaamsholte, segmentatie enwat er van de eerste instulping van de embryonale ontwikkeling wordt (grofweg: wordt de blastopore mond of anus). De Deuterostomia, beesten met een tweede instulping als mond, waren morfologisch onderscheiden en kwamen moleculair terug, met dezelfde fyla; zoals we al gezien hebben. Lichaamsholte en segmentatie bleken geen enkele weerspiegeling te hebben in de moleculaire indeling, en de hele overige club van de fyla, de Protostomia in figuur 11, 12, wordt nu anders ingedeeld.
Heeft de moleculaire indeling dan geen morfologische tegenhanger? Jawel, maar het waren deels kenmerken waar niet aan gedacht was. Vroeger zaten de gelede wormen (als de regenworm) naast de geleedpotigen (als de duizendpoten, insecten en zo). Moleculair zitten de gelede wormen naast de weekdieren, met een reeks minder bekende beesten. Op grond van de embryologie is dat veel beter. Deze groep, Lophotrochozoa in figuur 11 en 12, hebben spiraalklieving in hun embryo, bijvoorbeeld. Het was ook allang bekend dat gelede wormen die in zee leven en weekdieren in zee hetzelfde type larve hadden. De indeling op lichaamsholte en segmentatie gooide de larven uit elkaar; bij de moleculaire indeling zitten de larven bij elkaar.
De Ecdysozoa vormen de tweede hoofdgroep in de Protostomia, en zijn de beestenrijkste groep van het dierenrijk. Het zijn de geleedpotigen, de fluweelwormen, de beerdiertjes, de rondwormen, en nog een paar fyla. Ecdysozoa betekent beesten die een dikke cuticula van chitine hebben, vervellen om te groeien, en het hormoon ecdyson daarbij gebruiken. Vervellen als morfologisch indelingskenmerk van een grote groep fyla was nooit voorgesteld. Het komt wel overeen met de moleculaire indeling. (Over morfologie en moleculen in het dierenrijk: Minelli 2009).
De eencelligen met een celkern zijn morfologisch lastig (en geen erg favoriete groep voor studie). De onderscheiding tussen Eukaryoten aan de ene kant en Bacterien, Archaea aan de andere kant is een echte celkern (en nog het een en ander in celstructuur). Bacterien verschillen van Archaea in celwand, celmembraan en nog een lijstje kenmerken.
Dat wil zeggen, al met al: de moleculaire indeling werkt beter dan de morfologische. Veel klassieke morfologische indeling komt terug, of de alternatieve moleculaire indeling blijkt toch ook een morfologische component te hebben. De moleculaire indeling geeft lost ook problemen op, als de indeling van de kleine katten, de plaats van de rode panda, de indeling van de ordes van de placentale zoogdieren. En een oplossing van het grootste indelingsprobleem: hoe de geleedpotigen in te delen.
De geleedpotigen zijn de insecten, cheliceraten, duizendpoten en kreeftachtigen. De cheliceraten zijn de spinnen, molukkenkreeft en zeespinnen.De fluweelwormen zijn morfologisch meestal beschouwd als het fylum naast het fylum geleedpotigen, of soms binnen de geleedpotigen gezet.
Morfologisch werden de insecten altijd als zustergroep van de duizendpoten gezien: landbeesten met tracheeën. Er waren morfologisch drie rivaliserende opvattingen over hoe het verder moest. De eerste opvatting is: (fluweelworm, (duizendpoot, insect)) tegen (cheliceraten, kreeften). Dan heb je landbeesten met onvertakte poten tegen beesten met vertakte poten (in de molukkenkreeft voor de cheliceraten). Deze opvatting heeft niet erg school gemaakt. In elke andere opvatting is de fluweelworm een fylum naast de geleedpotigen.
In de tweede opvatting werd de indeling: (insecten, duizenpoten) tegen (kreeftachtigen, cheliceraten). In de derde opvatting is de indeling ((insecten, duizendpoten), kreeftachtigen) tegen cheliceraten. Dan zijn de kreeften de zustergroep van de insecten + duizendpoten. Daar is morfologisch wat voor te zeggen op grond van de bouw van de kop; tenminste, als het aantal kopsegmenten hetzelfde is, en de kopaanhangsels vergelijkbaar zijn. Kreeften hebben zes kopsegmenten. Als het bij elke groep om zes segmenten gaat, ook bij de insecten en duizendpoten, draagt kopsegment 4 bij elke groep een aanhangsel geheten mandibel – het meest kaakachtige van de aanhangsels. De groep insecten + kreeften + duizendpoten heet daarom Mandibulata, mandibeldieren. De meeste morfologische discussie ging over de vraag hoe de kop van de insecten en duizendpoten in elkaar zat, met hetzelfde aantal segmenten of niet. De morfologische consensus is nu dat het bij alle drie groepen om zes kopsegmenten gaat, waarbij segment 2 geen aanhangsel draagt in insecten en duizendpoten.
Dat was morfologie, voor 1995 grotendeels. Moleculair kwam de grootste verrassing van alle moleculaire studies: de insecten bleken binnen de kreeften terecht te komen. In 1995 kwam de eerste en overtuigende moleculaire studie dat insecten en kreeften zustergroepen waren, en 1997 of zo kwamen de insecten binnen de kreeften terecht. Formeel zijn daarmee de insecten een landvorm van de kreeftachtigen. Waar precies in de kreeften was nog de vraag, en waar de duizendpoten dan bleven ook. De laatste studie is van Regier en medewerkers, van 2010; deze studie onderscheidt zich omdat er een behoorlijk aantal duizendpoten in is opgenomen, en een diversiteit aan kreeftengroepen, ook de wat minder bekende kreeftachtigen.
De moleculaire indeling ziet er dan als volgt uit (Regier 2010):

Figuur 17. Moleculaire indeling van de geleedpotigen, met fluweelwormen en beerdiertjes als buitengroep (Regier et al 2010). Horizontale lijnlengte hoeveelheid verandering.

Figuur 18 Schema van de moleculaire indeling van de geleedpotigen, naar Regier et al (2010). Zelfde kleuren als figuur 17.
De insecten komen binnen de kreeftachtigen terecht; en de hele groep heet daarmee Pancrustaceae. Dit valt morfologisch ook wel te verdedigen: uit hersenstructuur en de structuur van de oogfacetten.
Ook vindt Regier dat de duizendpoten naast de Pancrustaceae terecht komen – en dat de Mandibeldieren, die op morfologie en genexpressie verdedigd waren, moleculair terugkomen.
En de morfologische overeenkomst tussen insecten en duizendpoten? Kopsegment 2 heeft bij deze beesten geen antennes: dat moet dan onafhankelijk verlies van een segmentaanhangsel zijn. Tracheeën? Insecten en duizendpoten hebben tracheeën, maar er zijn genoeg verschillen in structuur en positie om die tracheeën niet als overeenkomstig te beschouwen (Minelli 2009 geeft de referenties daarvoor). Tracheeën horen bij de levenswijze op het land.
DEEL III
Dit ging allemaal over de huidige beesten. Wat gebeurt er met de patronen als we de fossielen erbij nemen? Niets – dat is te zeggen, er gebeurt niets met de patronen, omdat dezelfde patronen verschijnen. De indeling van de hoofdgroepen van de landvertebraten komt terug in de fossielen, op dezelfde manier als bij de huidige beesten. Alleen kun je bij de fossielen het voorkomen in de tijd volgen, en alle beesten zien die er ook nog geweest zijn. Zoals dinosauriers. De vogels delen in binnen de dinosauriers, maar dat is intussen wel algemeen bekend. Formeel zijn de dinosauriers dus niet uitgestorven: ze zitten in de bomen en zingen. De niet-vogel dino’s zijn uitgestorven.
De walvissen worden moleculaire binnen de evenhoevigen ingedeeld, met de nijlpaarden als zustergroep: en dat betekent natuurlijk dat walvissen evenhoevigen zijn.
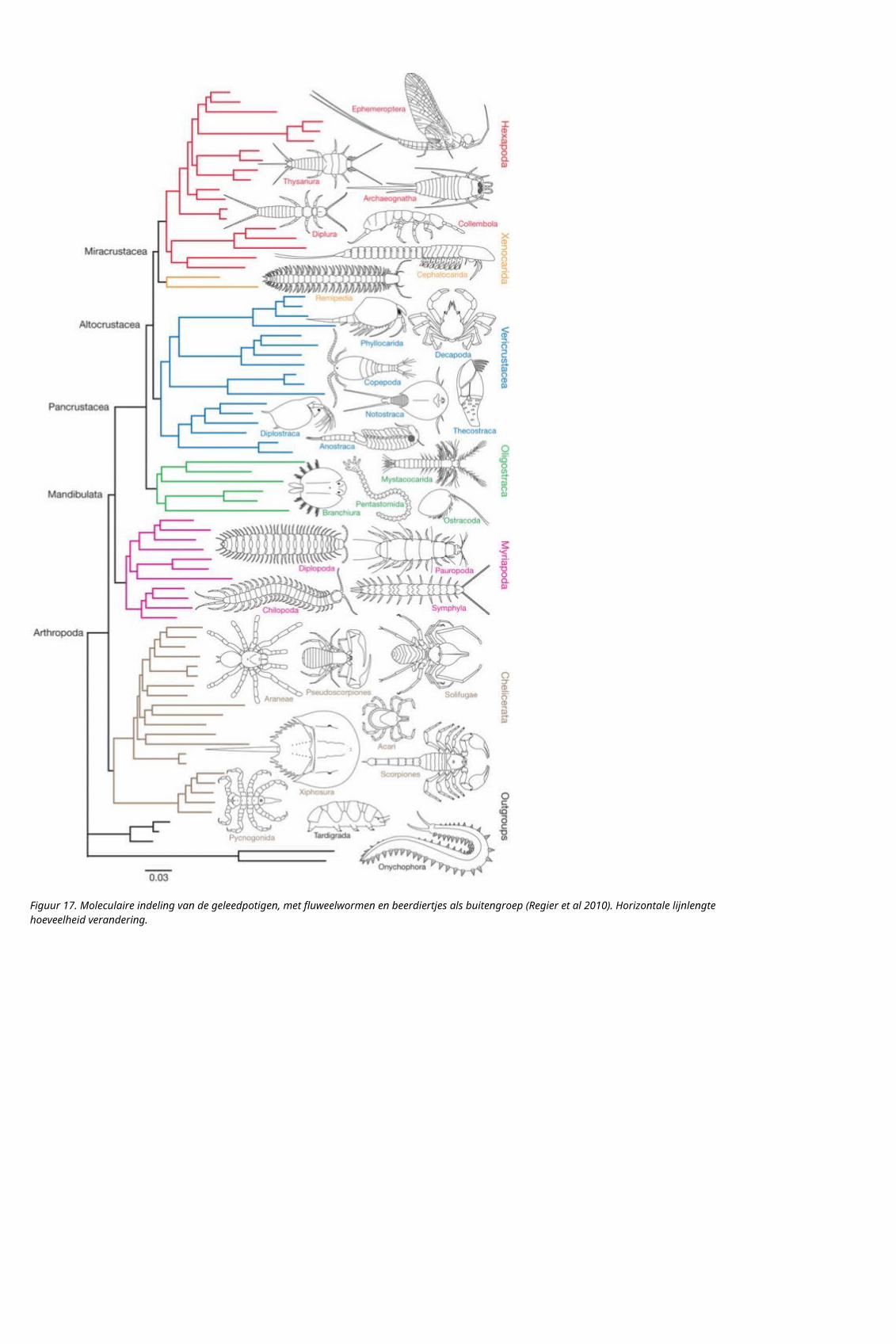
Figuur 19. Walvissen worden moleculair ingedeel binnen de evenhoevigen: links tabel met genetische afstand, rechts indelingsboom (Freeman & Herron 2007). Indeling op betacaseine.
figuur 20. Moleculaire indeling op mtDNA: deel van de indeling van de placentale zoogdieren door Arnason et al (2008). De walvissen (rode stip) worden ingedeeld binnen de evenhoevigen (groene stip).
De indeling van de fossielen is dezelfde:
Figuur 21. Fossiel zijn de Anthracoteren de zustergroep van de nijlpaarden, en de walvissen de zustergroep van de nijlpaarden + Anthracotheren; fossiel worden al deze beesten ingedeeld binnen de evenhoevigen. Lichte lijn: lay-out van de indelingsboom. Donkere lijn: tijd van voorkomen van in de indeling voorkomende soort. Grijs: geeft groep aan. ( Boisserie et al 2005).
Er is een morfologisch kenmerk dat alleen bij de walvissen voorkomt en niet bij de andere huidige placentale zoogdieren. Het heet involucrum, en is een soort verdikking van het kapsel om het middenoor. Dit kenmerk komt ook voor in het fossiel Pakicetus uit het Eoceen: en daarom werd geconcludeerd dat Pakicetus ingedeeld moest worden bij de walvissen. Verder graven leverde de poten van Pakicetus op: een landbeest en een hoefdier. Dat Pakicetus een hoefdier bleek te zijn was nogal spectaculair (Thewissen et al 2001: er is ook een Noorderlicht programma hierover).
De vormverandering van de walvissen loopt (heel erg schematisch! er zijn veel meer beesten bekend):

Figuur 23 Fossiele walvissen.
In figuur 7 staat de moleculaire indeling van de eierleggende zoogdieren, de buideldieren en de placentale zoogdieren. Alle huidige zoogdieren worden samen met een redelijk aantal fossiele soorten, uit het mesozoicum voornamelijk, ingedeeld binnen de zoogdiervormigen. De zoogdiervormigen worden ingedeeld binnen de therapsida, die met een volledig verouderde naam zoogdierachtige reptielen hebben geheten.
Er zijn nu 310 geslachten zoogdiervormigen uit het mesozoicum bekend. De indeling van de fossiele zoogdiervormigen is volgens Luo (2007):
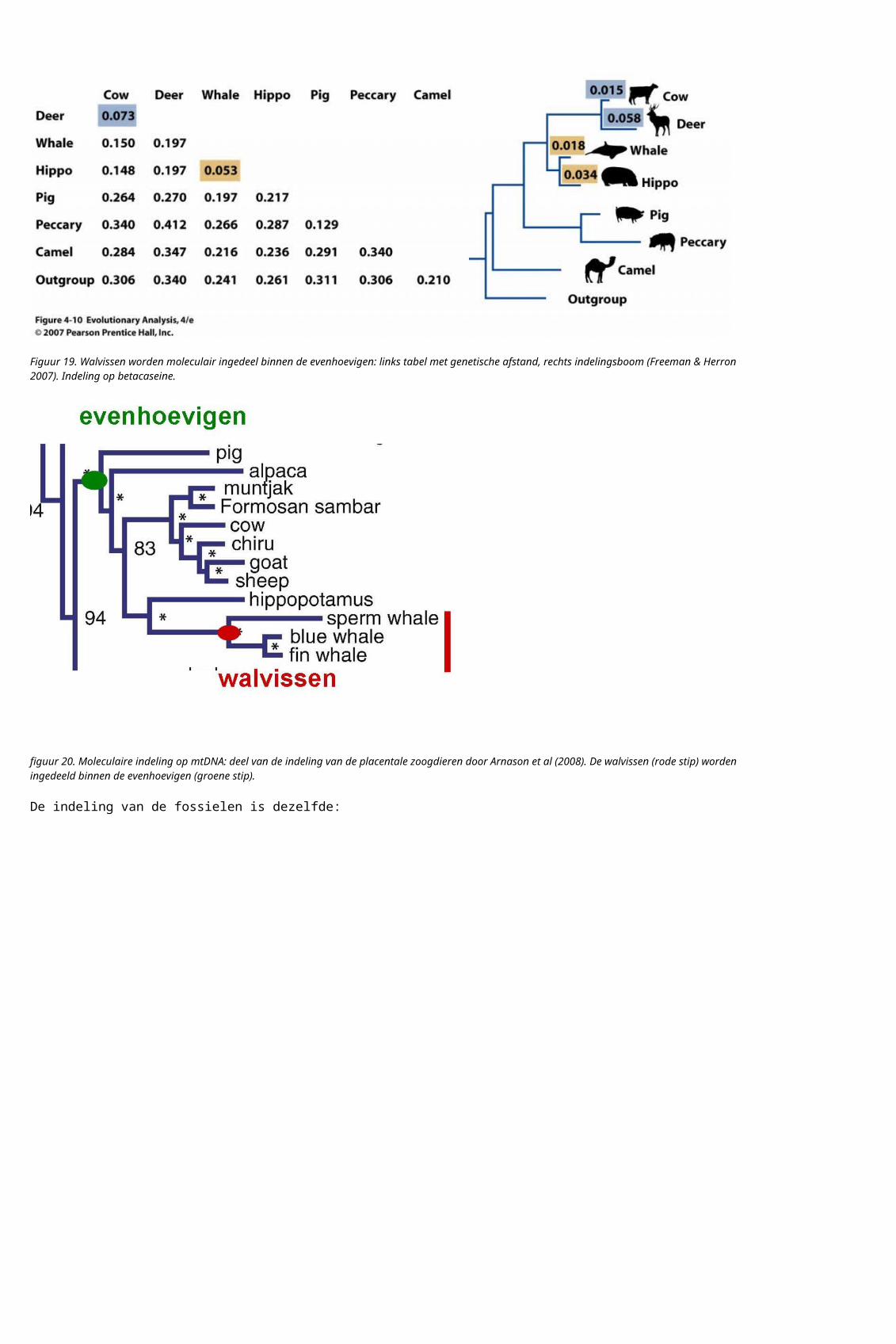
Figuur 24. Indeling van de fossiele zoogdiervormigen. a: soorten en groepen. b: indeling waarbij schematisch de diversiteit binnen de groep is weergegeven, voor zo ver bekend. Rood: placentale zoogdieren en fossiele verwanten. Blauw: buideldieren en fossiele verwanten. Groen: eierleggende zoogdieren en fossiele verwanten. Geel uitgestorven groepen die binnen de omschrijving zoogdier vallen, als zoogdier op grond van de huidige eierleggende zoogdieren, buideldieren en placentale zoogdieren wordt gedefinieerd. Donkerblauw: stam zoogdieren.
De indeling van de fossiele zoogdieren komt overeen met de indeling van de huidige zoogdieren. Veranderingen in morfologie, als structuur van het middenoor en kenmerken van het gebit, zijn te volgen over deze indeling.
DEEL IV
Dat zijn de patronen bij de indeling van soorten in de biologie. Er zijn meer patronen: in overeenkomst van genen tussen groepen soorten, in overeenkomst in genexpressie tussen groepen soorten, in genfamilies, in vergelijkende anatomie, in ontwikkeling, in ecologie, …, maar ik laat het hierbij.
Wat heb ik laten zien?
1 het is mogelijk alle levende organismen in te delen op moleculaire overeenkomst
2 dat geeft een hiërarchische indeling over de levende wezens, met groepen binnen groepen, steeds herhaald.
3 moleculair, morfologisch en fossiel geeft dezelfde indeling
De hiërarchische indeling laat zien welk organisme het meest overeenkomt met welk ander organisme, welke groep overeenkomt met welke andere groep, welke groep een onderdeel is van een wijdere groep, welke groep een nauwere groep omvat. Al die indeling werkt met overeenkomst.

Waar komt overeenkomst vandaan? Er zijn twee waarneembare bronnen van overeenkomst, levenswijze en verwantschap. Een moleculaire indeling vermijdt levenswijze. Verwantschap geeft een duidelijke verklaring van de indeling. Verwantschap is het gevolg van gedeelde afstamming van een gezamenlijke voorouder. De katten hebben een gezamenlijke voorouder, de roofdieren hebben een gezamenlijke voorouder, de placentale zoogdieren hebben een gezamenlijke voorouder, (etc). De indeling wijst op gedeelde gezamenlijke afstamming van alle levende wezens.
DEEL V
Iedereen, inclusief creationisten, aanvaardt de indeling van de katten (figuur 4) als een weergave van verwantschap. De kattensoorten binnen de huiskatgroep zijn verwant met elkaar, de huiskatgroep is verwant met de zustergroep van de luipaardkatten, die twee samen hebben de puma-cheetahgroep als meest verwante zustergroep, en zo verder, tot de Grote Katten elk even verwant zijn met elke Kleine Kat. Binnen de katten wordt verwantschap als gezamenlijke afstamming geinterpreteerd, door iedereen.
De daaropvolgende moleculaire indelingen werken precies hetzelfde als de moleculaire indeling van de katten. Daarmee is het niet mogelijk de moleculaire indeling van de katten wel als verwantschap maar de moleculaire indeling van de roofdieren niet als verwantschap te interpreteren. En ook is het daarmee niet mogelijk de moleculaire indeling van de katten wel als verwantschap maar de moleculaire indeling van de ordes van de zoogdieren niet als verwantschap te interpreteren. Weigeren de moleculaire indeling van de roofdieren, de zoogdieren als verwantschap te interpreteren zou een andere interpretatie leggen op zelfde materiaal. De indeling laat duidelijk zien dat het idee van gescheiden basistypen geen grond heeft.
Het creationistische boek van Junker & Scherer (2006) zegt op blz 305:
“Als er een algemene evolutie van organismen zou bestaan, zouden de vele soorten minstens macro-taxonomisch in een boomvormige structuur en daarmee in een hiërarchisch, compact systeem ingepast moeten kunnen worden. Dit is tot op zekere hoogte mogelijk, maar er zijn over de gehele linie zwaarwegende uitzonderingen waarbij dit niet lukt.”.
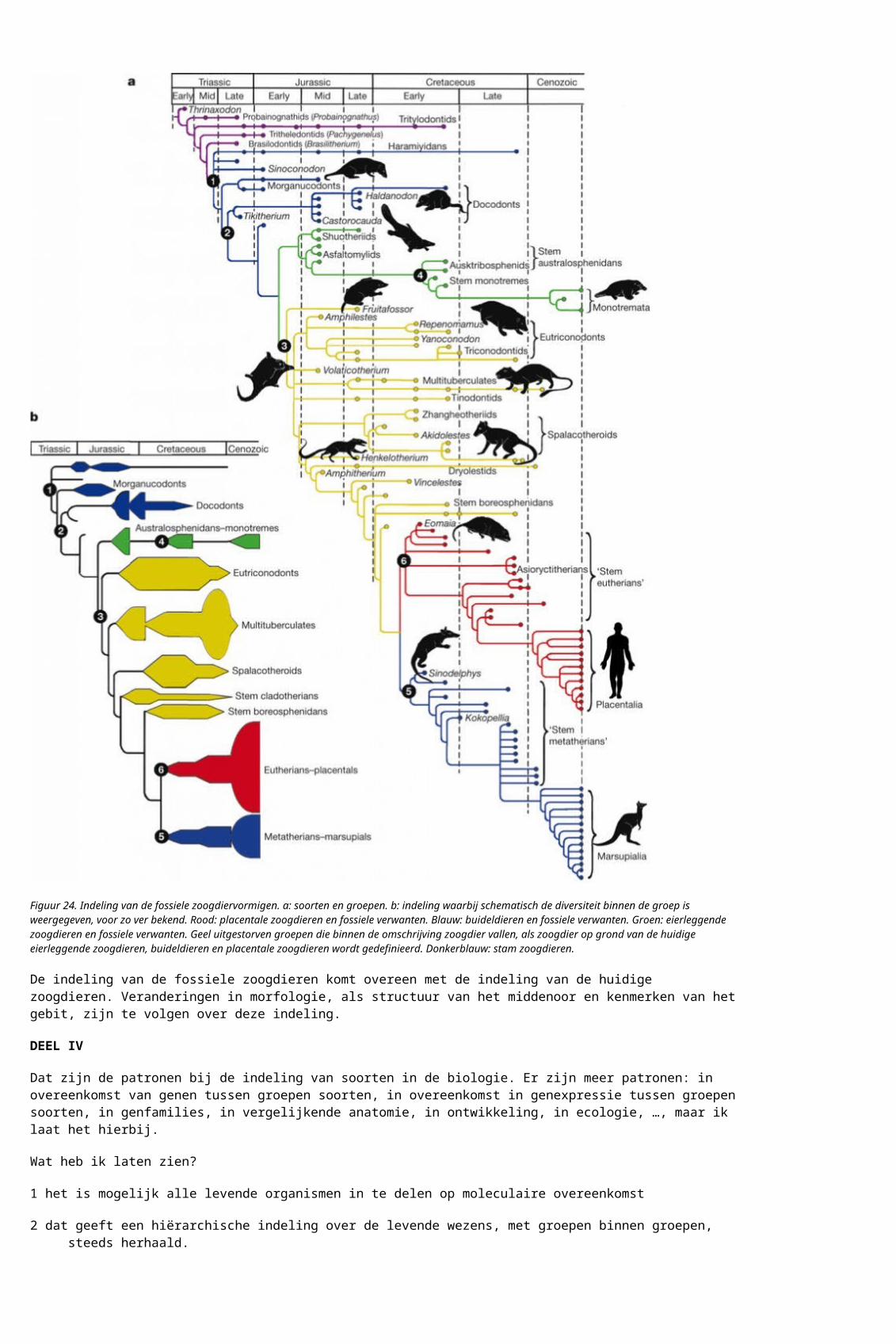
Inpassing in een hiërarchisch, compact is in tegenstelling tot de bewering van J&S over de hele linie mogelijk. J&S noemt niet welke de ‘zwaarwegende uitzonderingen’ zijn, of waar het ‘over de gehele linie’ op slaat, maar even later verwijst J&S naar figuur 16.35 op blz 315. Dat is een figuur die de relaties tussen de groepen van de geleedpotigen onderling (de insecten, duizendpoten, kreeften, cheliceraten) en de fluweelwormen beoogt aan te geven. De figuur komt uit een wetenschappelijk boek uit 2001 over de techniek van het indelen.
Figuur 25 = figuur 16.35 J&S. Bijschrijft in J&S: “Relaties tussen organismen op basis van overeenkomsten kunnen als netwerk voorgesteld worden. Als voorbeeld zijn de verwantschapsrelaties tussen arthropoden (geleedpotigen) weergegeven. Afhankelijk van het gehanteerde kenmerk ontstaan verschillende groepen. De bijbehorende onderzoekingen stammen uit de 1990-er jaren.”
Als er “afhankelijk van het gehanteerde kenmerk .. verschillende groepen (ontstaan)” is er geen netwerk, maar een serie alternatieve hypothesen.
Hierboven, in figuur 17 en 18, staat de indeling van de geleedpotigen. Een dergelijke indeling, maar minder mooi weergegeven, was ook in 2006 bekend.
Ook benadrukt Junker & Scherer convergentie. Convergentie betekent dat de levenswijze weerspiegeld wordt in de morfologie. Walvissen, ichthyosaurs, haaien en tuna’s zijn convergent in lichaamsvorm: stroomlijn voor snelle zwemmer, maar verder verschillend. De mierenegel legt eieren, de numbat is een buideldier, de miereneter hoort bij de Xenarthra, het schubdier bij de Laurasiatheria, het aardvark bij de
Afrotheria (zie figuur 6 voor de hoofdgroepen van de placentale zoogdieren); ze eten mieren of termieten, hebben een spitse snuit met gereduceerde tanden en een lange tong. Hier is het duidelijk dat de levenswijze de morfologie bepaalt. Er is niets bekend over de genetische achtergrond van deze overeenkomst in morfologie. Convergentie is overal: dezelfde ecologische niche op een andere plaats geeft een min of meer hetzelfde beest uit een andere groep een opening.
Junker & Scherer doen alsof convergentie voor de reconstructie van de fylogenie een groot struikelblok is. Dat is niet correct: morfologische convergentie wordt ondervangen door moleculaire fylogenie. Moleculaire convergentie in sommige eiwitten (die ook bestaat) wordt ondervangen door moleculaire fylogenie op het geheel van het DNA. Convergentie valt te herkennen. Morfologische convergentie is niet bestand tegen studie van morfologisch detail: de tracheeën van insecten en duizendpoten verschillen in embryologie en bouw, het vierkamerige hart van vogels en zoogdieren verschilt in embryologische ontwikkeling, een walvis golft op/neer en een vis links/rechts – de walvis galoppeert, en de kiezen van een Tasmaanse tijger verschillen van die van de placentale roofdieren.
*********
Voor namen van beesten zie wikipedia
-Adoutte, A., G. Balavoine, N. Lartillot, O. Lespinet, B. Prud’homme, & R. de Rosa. 2000. The new animal phylogeny: Reliability and implication. PNAS 97: 4453-4456.
-Arnason, U., J.A. Adegoke, A. Gullberg, E.H. Harley, A. Janke, M. Kullberg, 2008. Mitogenomic relationships of placental mammals and molecular estimates of their divergences. Gene 421:37–51
-Blair, J.E., & S.B. Hedges, 2005. Molecular phylogeny and divergence times of deuterostome animals. Molecular biology and evolution 22: 2275-2284.
-Boisserie, J-R, F Lihoreau & M Brunet, 2005. The position of Hippopotamidae within Cetartiodactyla. PNAS 102: 1537-1541
-de Muizon, 2001. Walking with whales. Nature 413: 259-260.
-Desmond E, Brochier-Armanet C, Forterre P, et al. 2011. On the last common ancestor and early evolution of eukaryotes: reconstructing the history of mitochondrial ribosomes. Researches in microbiology: 162: 53-70
-Eizirik, E., W.J. Murphy, K.P. Koepfli, W.E. Johnson, J.W. Dragoo, R.K.Wayne, en S.J. O’Brien, 2010. Pattern and timing of the diversification of the mammalian order Carnivora inferred from multiple nuclear gene sequences. Molecular Phylogenetics and Evolution 56: 49-63.
Freeman, S., & J.C. Herron, 2007. <i>Evolutionary Analysis</i> 4th edition. Pearson Education Inc. ISBN 978-0-13-239789-6.
-Hackett, S.J., et al., 2008. A phylogenomic study of birds reveals their evolutionary history. Science 320: 1763-1768.
-Johnson, W.E., E. Eizirik, J. Pecon-Slattery,W.J. Murphy, A. Antunes, E. Teeling, S.J. O’Brien, 2006. The Late Miocene Radiation of Modern Felidae: A Genetic Assessment. Science 311; 73-77.
-Luo, Z-X., 2007. Transformation and diversification in early mammal evolution. Nature 450:1011-1019.
-Mattern, M.Y., & D.A.. McLennan, 2000. Phylogeny and Speciation of Felids. Cladistics 16: 232–25
-Minelli, A. 2009. <i>Perspectives in Animal Phylogeny and Evolution.</i> Oxford University Press. ISBN 978-0-19-856621-2
-Nishihara, H., M.Hasegawa, & N. Okada, 2006. Pegasoferae, an unexpected mammalian revealed by tracking ancient retroposon. PNAS 103:9929-9934.
-Putnam, N.H., Butts T, Ferrier DEK, et al., 2008. The amphioxus genome and the evolution of the chordate karyotype. Nature 453: 1064-1075
-Raible F, Tessmar-Raible K, Osoegawa K, et al., 2005. Vertebrate-type intron-rich genes in the marine annelid <i>Platynereis dumerilii</i>. Science 310: 1325-1326
-Regier J.C., J.W. Shultz, A. Zwick, A. Hussey, B.Ball, R. Wetzer, J.W. Martin & C.W. Cunningham, 2010. Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences. Nature 463:1079-1084.
-van Rheede T, Bastiaans T, Boone DN, Hedges SB, de Jong WW, & Madsen O. 2006. The platypus is in its place: Nuclear genes and indels confirm the sister group relation of monotremes and therians. Molecular Biology and Evolution 23: 587-597
-Zardoya R, & A. Meyer, 2001. The evolutionary position of turtles revised. Naturwissenschaften 88: 193-200.
Gastblog macroevolutie - 3
Bij deze het derde deel van de serie gastblogs die Gerdien de Jong schrijft over macroevolutie. Een stevige kluif, maar het is dan ook geen onbelangrijk onderwerp.
Verandering in de levende natuur
Een meteoorinslag, uitbarstingen van vulkanen, verschuiven van continenten: alles heeft zijn weerslag op de levende natuur, welke organismen er zijn en of ze blijven bestaan. dat is weer een ander verhaal. Hier wil ik het hebben over de beesten (soms planten) zelf: hoe die veranderen.
Er zijn vier onderwerpen bij verandering: toeval, ontwikkeling, erfelijkheid en selectie. Bij erfelijkheid kan ik genetische verandering ook mutatie noemen, maar sommige mensen denken bij mutatie alleen aan puntmutatie, een mutatie van één base in het DNA. Mutatie gelijk te stellen aan puntmutatie is een beetje kortzichtig. Er bestaat heel veel genetische verandering, inclusief verdubbeling van het hele genoom, en het mag allemaal mutatie heten.

Eerst komen komt er een overzicht met per onderwerp een lijstje van mogelijke verandering, steeds met een klein voorbeeld. Die lijstjes pretenderen geen volledigheid. Er komt een beetje commentaar, maar interessante gevallen moeten wachten tot de volgende aflevering.
I Toeval
Toeval: alle zaken waarvoor geen eenduidige oorzaak die betrekking heeft op het bestudeerde valt aan te geven.Toeval op alle manieren: een groep sterft uit of niet na meteoor inslag, een groep komt op een eilandcontinent terecht. Toeval bestaat ook in morfologie. De Indische neushoorn heeft één hoorn, de Afrikaanse neushoorns hebben er twee. Bij de herkauwers hebben de runderen en de schapen / geiten hoorns en de herten een gewei. Ook zulk verschil valt onder toeval. Toeval in het genoom is een bekender idee. Puntmutatie treedt op volgens toeval (en ik bedoel niet uniform verdeeld, en zelfs niet Poisson verdeeld), chromosoomreorganisatie en genduplicatie zijn algemeen. Polyploidie is bij planten veelvuldig. Dat zijn allemaal zaken die geen aanwijsbare oorzaak hebben, en daarmee vallen onder toeval. Ook van toevallige zaken kun je nagaan wat de uitwerking is.
II Ontwikkeling
II-1 Epigenese.
Het DNA bevat geen blauwdruk voor het organisme. Blauwdruk is een misleidend idee, want het laat denken dat in het DNA alles precies vastligt. Dat is niet zo. Het DNA geeft een recept, min of meer, en al naar gelang de omstandigheden komt daar een wat ander organisme uit. De ontwikkeling van bevrucht ei tot beest (eh, plant) verloopt volgens een min of meer gegeven procedure, op volgorde, met speling. Genen geven instructies voor de procedure, en worden geïnstrueerd, om een proces uit te voeren, onder inspraak van de omgeving. Net zoiets als een ballet, alleen minder strak geregisseerd. Embryologie heeft dat laten zien, met alle regulatienetwerken van genen, organizers en zelforganisatie.
Verandering tijdens ontwikkeling is het gevolg van verandering van bestaande structuren op een manier die epigenetisch heet. De cellen en weefsels zijn in contact met elkaar, en wisselen boodschappen uit. Bloedvaten en bot ontstaan waar nodig is, en bloedvaten en zenuwen concurreren met elkaar om klandizie. Bot heeft vele voorgangers, en de vorm van een bot geeft aan waar de meeste kracht op een bot werd uitgeoefend tijdens het leven. Verbening van bindweefsel als daar veel kracht op werkt is standaard; bijvoorbeeld, er zijn zogenaamde sesambotjes in de pees van de buigspier van de grote teen, bij mensen. De knieschijf is een groot uitgevallen sesambotje. Sesambotjes verschillen tussen soorten.
Alle verandering binnen een soort in een kenmerk als snavelgrootte (de Darwinvinken gaan weer eens langs komen) is dan ook een epigenetische verandering, waarbij de communicatie over en weer tussen weefsels iets anders verloopt bij de verschillende soorten..
Grote punten bij epigenese zijn hoe cellen communiceren, en hoe het netwerk van genregulatie verloopt over cellen. Daar zijn vele eiwitten voor, die meestal als functie hebben genen aan of uit te schakelen (soms direct soms via boodschappers van celmembraan naar kern).
II-2 Patroonvorming
Bij tweezijdig symmetrische beesten wordt eerst de voor-achter as aangelegd in het embryo (de A-P as). De initiatie van de A-P as kan op allerlei manieren gebeuren; voor de as zelf zijn signaalsystemen van genetische regulatoren nodig. Een gradient in genexpressie langs de A-P as specificeert afstand vanaf de voorkant. Die genexpressiegradient schakelt een volgende reeks genen aan, die de identiteit van een plek of segment bepalen en daarmee de verdere ontwikkeling. Dit zijn de zogenaamde Hox-genen. De Hoxgenen worden genummerd of aangeduid met namen die afkomstig zijn van de namen van de fruitvlieg.
De expressie van Hoxgenen langs de A-P as laat zien hoe de verschillende groepen hun specifieke vorm regelen. De Hoxgenen komen op volgorde van voor tot achter tot expressie. Bij de geleedpotigen zijn er specifieke Hoxgenen voor kop en voor lijf.
Figuur 1. Expressie van de Hox genen van een duizendpoot. De namen van de Hoxgenen zijn die van Drosophila melanogaster, en de Hoxgen nummers zijn aan de hand van hun volgorde in het gencomplex in D. melanogaster. Janssen & Damen 2006.
Bij de duizendpoot komt het gen Hox1 = labial , afgekort lab, tot expressie vanaf het derde segment; het gen Hox4=Deformed, afgekort Dfd, komt tot expressie vanaf het vierde segment. Hox1 tot en met Hox 4 zijn specifiek voor de kop. Hox10 geeft het einde van het beest aan. Hox7 tot en met Hox9 zijn specifiek voor de thoraxsegmenten. Het is duidelijk te zien dat de thoraxsegmenten identiek zijn in Hoxexpressie, na de overgang vanaf de kop.
Figuur 2. Expressie van de Hox genen van de fruitvlieg en een spin. Ga uit van de segment indeling die als NEW aangegeven staat. Grijs balkje sterke expressie, wit balkje zwakke expressie. Damen 2009.
Bij de spin komt het gen Hox1 = labial tot expressie vanaf het derde segment, bij de fruitvlieg is het derde segment het enige segment waarin labial tot expressie komt.
Het gen Hox4=Deformed, komt tot expressie vanaf het vierde segment, bij de spin over het hele kopborststuk, bij de vlieg over de eerste twee segmenten van de achterkop. Hox7 tot en met Hox9 (Antp, Ubx en abd-A) geven bij de spin het achterlijf. Bij de vlieg geven Hox7 en Hox8 (met de niet aangegeven Hox5) het borststuk, terwijl Hox9 het voornaamste gen voor de identiteit van het achterlijf is.
Voor de kreeften heb ik geen mooi plaatje (wel een slecht plaatje uit 1999). Ook bij de kreeften is Hox1 specifiek voor het derde segment, terwijl Hox4 vanaf het vierde segment begint. Hox1 komt tot expressie in het laatste segment van de ‘oude’ geleedpotigenkop, de segmenten 1-3. Bij de Mandibulata (zie vorige blog) geeft Hox4 de ‘nieuwe’ onderdelen van de kop, segment 4-6; bij de spin geeft Hox4 het verdere kopborststuk. Hox 7-9 zijn voor het achterlijf.
II-3 Heterochronie
Heterochronie betekent dat een gen eerder of later tot expressie komt, of dat bij vergelijken tussen twee soorten een onderdeel zich op een ander tijdstip ontwikkeld.
De vorm van de bloem in Delphinium nudicaule kan als een geval van heterochronie worden opgevat. Delphinium nudicaule heeft rode bloemen van een kleur die verder onder riddersporen schaars is, en komt uit Californie en schijnt door kolibries bestoven te worden. De vorm van de bloem is nogal knopachtige, vergeleken met de een blauwe ridderspoor Delphinium decorum uit hetzelfde milieu, terwijl de spoor van gelijke grootte is.
Figuur 3 Delphinium nudicaule en Delphinium decorum. Zie ook hier.
II-4 Allometrie

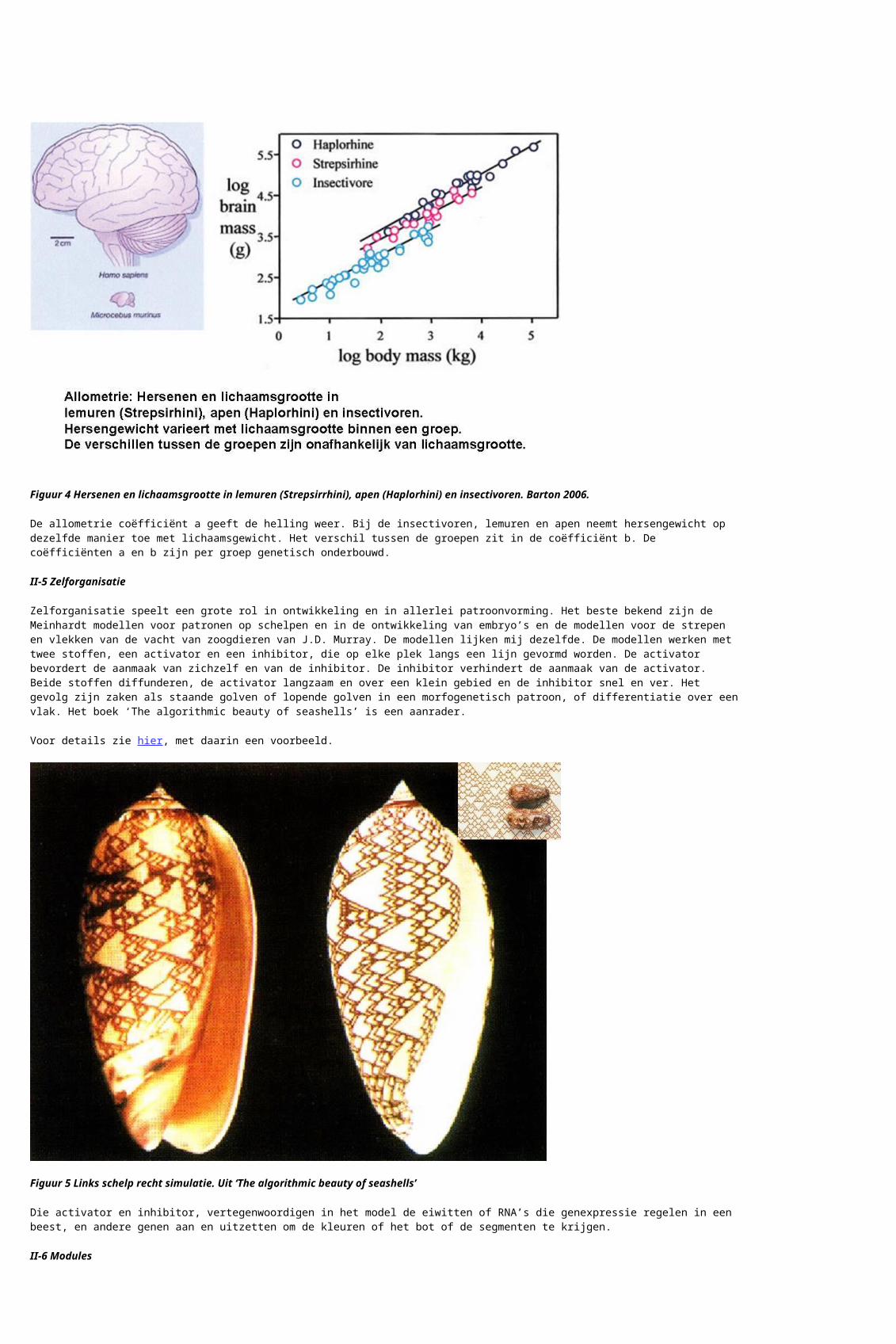
Allometrie betekent dat nagegaan wordt hoe de vorm van een beest verandert met zijn grootte. De vraag is bijvoorbeeld of de doorsnee en daarmee de sterkte van de poten gelijk kan blijven als een huiskat opgeschaald wordt tot een leeuw, of een big tot een olifant. Er blijkt dat delen op een andere manier veranderen dan het geheel, de grootte van het beest. Meestal blijkt het te gaan om een exponentiële verandering. Voor bijvoorbeeld hersengrootte x en lichaamsgrootte y bestaat het volgende verband: :
y = bxa
of, als logarithme geschreven en geplot:
log y =log b + a log x
Figuur 4 Hersenen en lichaamsgrootte in lemuren (Strepsirrhini), apen (Haplorhini) en insectivoren. Barton 2006.
De allometrie coëfficiënt a geeft de helling weer. Bij de insectivoren, lemuren en apen neemt hersengewicht op dezelfde manier toe met lichaamsgewicht. Het verschil tussen de groepen zit in de coëfficiënt b. De coëfficiënten a en b zijn per groep genetisch onderbouwd.
II-5 Zelforganisatie
Zelforganisatie speelt een grote rol in ontwikkeling en in allerlei patroonvorming. Het beste bekend zijn de Meinhardt modellen voor patronen op schelpen en in de ontwikkeling van embryo’s en de modellen voor de strepen en vlekken van de vacht van zoogdieren van J.D. Murray. De modellen lijken mij dezelfde. De modellen werken met twee stoffen, een activator en een inhibitor, die op elke plek langs een lijn gevormd worden. De activator bevordert de aanmaak van zichzelf en van de inhibitor. De inhibitor verhindert de aanmaak van de activator. Beide stoffen diffunderen, de activator langzaam en over een klein gebied en de inhibitor snel en ver. Het gevolg zijn zaken als staande golven of lopende golven in een morfogenetisch patroon, of differentiatie over een vlak. Het boek ‘The algorithmic beauty of seashells’ is een aanrader.
Voor details zie hier, met daarin een voorbeeld.
Figuur 5 Links schelp recht simulatie. Uit ‘The algorithmic beauty of seashells’

Die activator en inhibitor, vertegenwoordigen in het model de eiwitten of RNA’s die genexpressie regelen in een beest, en andere genen aan en uitzetten om de kleuren of het bot of de segmenten te krijgen.
II-6 Modules
Bij ontwikkeling kan onderscheid gemaakt worden in min of meer onafhankelijke modules. De schedel van vertebraten bijvoorbeeld bestaat uit een deel dat ook voorkomt bij kraakbeenvissen als de haai, en een deel dat afkomstig is benige buitendekplaten in de huid. Het eerste heet het chondrocranium, het tweede het dermatocranium. Bij de ontwikkeling van de snuit blijken dit onafhankelijke modules, gecontroleerd door verschillende genen. Waar in de ontwikkeling zulke onafhankelijke modules zitten is niet altijd voor alle beesten hetzelfde.
II-7 Fenotypische plasticiteit
Beesten met hetzelfde genotype kunnen in ander milieu een ander uiterlijk, fenotype, krijgen. Dat kan gebeuren tijdens het leven van één beest, waarbij het beest sterk verandert: zoals de kanoetstrandloper zijn spiermaag omzet in vet voor de trek. Het kan ook tijdens de ontwikkeling van het beest. Insecten die bij lage temperatuur gekweekt worden worden groter dan insecten die bij hoge temperatuur gekweekt worden. De rups van de vlinder Nemoria arizonaria http://whyevolutionistrue.wordpress.com/2011/03/07/mimicry-polymorphism-for-camouflage-in-a-caterpillar/ eet eikenblad en lijkt op een eikenkatje vroeg in het voorjaar en op een eikentakje wat later, onder invloed van de hoeveelheid tannine in het eikenblad. Fenotypische plasticiteit houdt in dat de mate van reageren op het milieu een erfelijke eigenschap van het beest is.
III Erfelijkheid
Erfelijkheid is een heel ruim thema: genetische variatie is er in soorten en maten, en voorlopig heb ik 13 rubrieken met genetische variatie. Het zou me niet verbazen als er meer zijn, maar deze waslijst verschijnselen is al lang genoeg.
Een punt moet even uit de weg: schadelijke mutaties. Schadelijke mutaties kunnen het gevolg zijn van een puntmutatie in het gen, of in het DNA dat het gen reguleert, of in een van de vele regulerende genen. Schadelijke mutaties kunnen ook het gevolg zijn van rondzwervende transposons of retrovirussen. De bron maakt eigenlijk niet uit. Mutaties die functie verstoren zijn niet interessant: er komt een mutatie-selectie evenwicht, en bij allerlei mutaties een stabiele verdeling van mutaties met hun frequenties in de populatie. Dat is de bagger die rond blijft drijven maar waar populaties geen last van hebben. Zo’n vijftig jaar geleden, voordat de moleculaire biologie bestond, werd gekeken naar hoeveel homozygoot lethale mutaties er in Drosophila melanogaster waren: zo’n 30% van de chromosomen heeft zeker één homozygoot lethale mutatie. Daar heeft de soort geen last van om voort te bestaan als leuke vlieg.
III-1 Puntmutatie
Een bekend voorbeeld van een interessante puntmutatie bestaat in het alcohol afbrekende enzym alcoholdehydrogenase in Drosophila. Het enzym heeft 255 aminozuren. In Drosophila simulans is het aminozuur op positie 192 lysine (code AAG). In D. melanogaster komen twee vormen voor, een met lysine op positie 192 en een met threonine op positie 192 (code ACG). In vrijwel alle D. melanogaster populatie komen beide vormen voor; de threonine variant werkt beter bij lagere temperatuur. Naast deze verandering die een gevolg heeft voor de functie van het enzym zijn nog 13 gevallen van alternatieve basen voor die geen effect hebben op de aminozuursamenstelling.
Drosophila melanogaster is van oorsprong een tropische soort, maar komt nu ook in gematigde streken voor. Overwinteren doet D. melanogaster daar door diapauze, een ruststadium. Diapauze komt in ?30% van de vliegen voor in zuid Florida tot in 90% in New England. Bij genetische analyse bleek dat het gen couch potato (cpo) belangrijk was. In het Couch Potato eiwit zijn er twee aminozuren vervangen ten opzichte van de oorspronkelijke tropische vorm: op plaats 356 valine in plaats van alanine, en op plaats 462 lysine in plaats van isoleucine. De frequentie van cpo3565Val en cpo462Lys in natuurlijke populaties van Florida tot Maine verandert van 15% tot 61%. (Schmidt et al 2008)
III-2 Genexpressie
Genetische variatie in hoe vaak of hoe snel een gen wordt afgeschreven is genetische variatie in gen expressie. Het kan binnen een soort optreden of tussen soorten. Verschil in gevormd eiwit en verschil in genexpressie kunnen samengaan, als bij alcolholdehydrogenase en couch potato in D. melanogaster. Genexpressie van cpo was lager in de vliegen met diapauze.
Een bekend voorbeeld is de verhoging van de expressie van het gen bone morphogenetic protein 2 (Bmp2) in embryonale vingers de vleermuis Carollia perspicillata vergeleken met de embryonale vingers huismuis. De vingers beginnen hetzelfde in beide soorten. Het lijkt erop dat verhoging van de activeit van het Bmp2 pathway bijdraagt tot de vorm van de vleermuisvleugel.
III-3 Genduplicatie
De verdubbeling van een gen, eventueel gevolgd door een nieuwe functie van dat gen, is een wijdverbreid verschijnsel. Het is soms te volgen binnen populaties, als bij het gen amylase in Drosophila melanogaster. Per chromosoom kan er één gen amylase of twee genen amylase aanwezig zijn.
Bekende voorbeelden van genduplicatie zijn hemoglobine en de Hoxgenen. Voor hemoglobine, zie HIER; op DEZE staan ook nog twee voorbeelden.
De Hoxgenen zijn lid van een veel grotere genfamilie, en het grote aantal Hoxgenen is het gevolg van uitgebreide genduplicatie. Die genduplicatie verloopt wat anders in de verschillende groepen beesten. Voor de indeling van het dierenrijk, zie vorige blogpost.
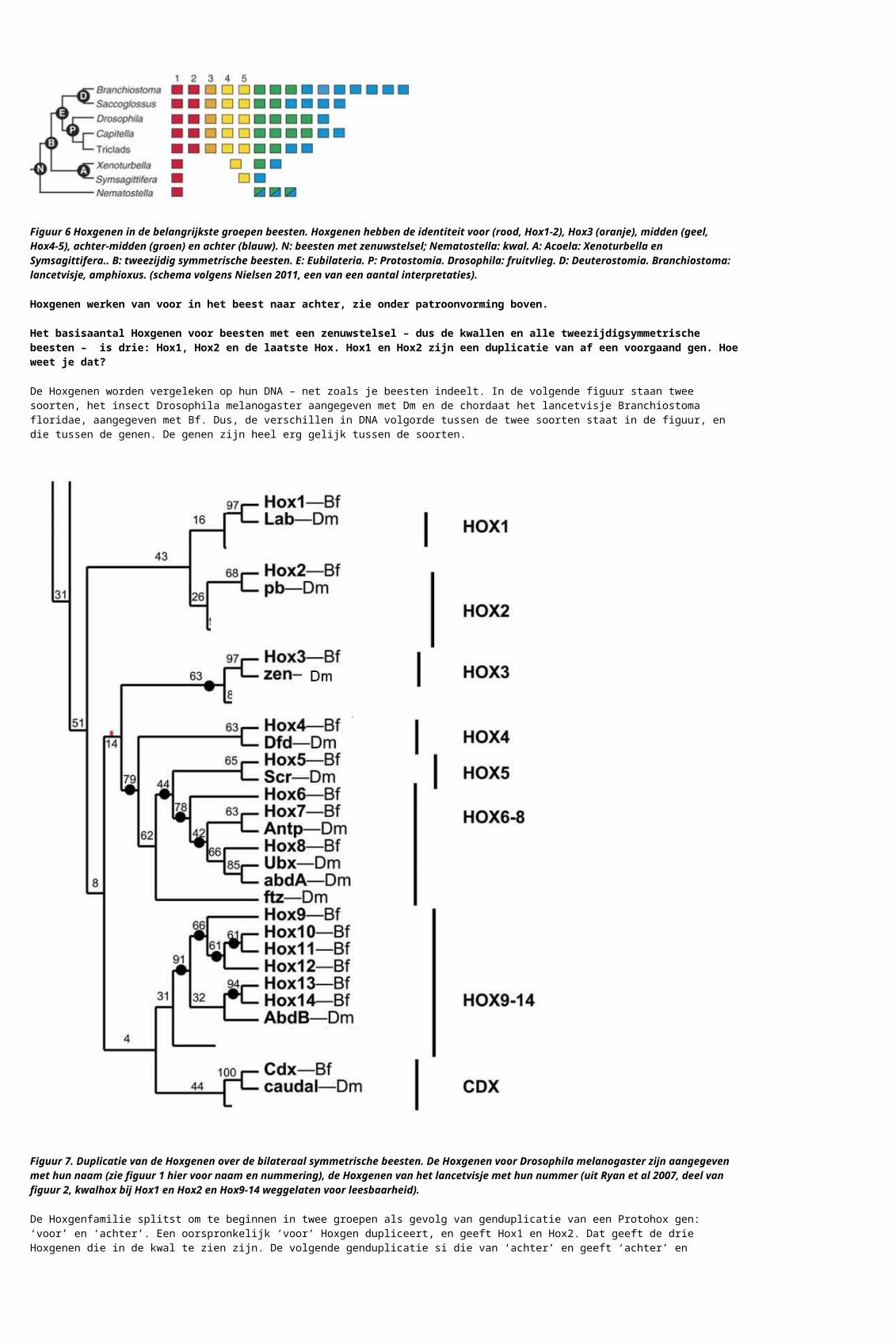
Figuur 6 Hoxgenen in de belangrijkste groepen beesten. Hoxgenen hebben de identiteit voor (rood, Hox1-2), Hox3 (oranje), midden (geel, Hox4-5), achter-midden (groen) en achter (blauw). N: beesten met zenuwstelsel; Nematostella: kwal. A: Acoela: Xenoturbella en Symsagittifera.. B: tweezijdig symmetrische beesten. E: Eubilateria. P: Protostomia. Drosophila: fruitvlieg. D: Deuterostomia. Branchiostoma: lancetvisje, amphioxus. (schema volgens Nielsen 2011, een van een aantal interpretaties).
Hoxgenen werken van voor in het beest naar achter, zie onder patroonvorming boven.
Het basisaantal Hoxgenen voor beesten met een zenuwstelsel – dus de kwallen en alle tweezijdigsymmetrische beesten – is drie: Hox1, Hox2 en de laatste Hox. Hox1 en Hox2 zijn een duplicatie van af een voorgaand gen. Hoe weet je dat?
De Hoxgenen worden vergeleken op hun DNA – net zoals je beesten indeelt. In de volgende figuur staan twee soorten, het insect Drosophila melanogaster aangegeven met Dm en de chordaat het lancetvisje Branchiostoma floridae, aangegeven met Bf. Dus, de verschillen in DNA volgorde tussen de twee soorten staat in de figuur, en die tussen de genen. De genen zijn heel erg gelijk tussen de soorten.

Figuur 7. Duplicatie van de Hoxgenen over de bilateraal symmetrische beesten. De Hoxgenen voor Drosophila melanogaster zijn aangegeven met hun naam (zie figuur 1 hier voor naam en nummering), de Hoxgenen van het lancetvisje met hun nummer (uit Ryan et al 2007, deel van figuur 2, kwalhox bij Hox1 en Hox2 en Hox9-14 weggelaten voor leesbaarheid).
De Hoxgenfamilie splitst om te beginnen in twee groepen als gevolg van genduplicatie van een Protohox gen: ‘voor’ en ‘achter’. Een oorspronkelijk ‘voor’ Hoxgen dupliceert, en geeft Hox1 en Hox2. Dat geeft de drie Hoxgenen die in de kwal te zien zijn. De volgende genduplicatie si die van ‘achter’ en geeft ‘achter’ en ‘midden’. ‘Achter’ is bij Drosophila vertegenwoordigd door gedupliceerde genen, het Hoxgen Abdominal-B en het embryonale gen Caudal. Bij het lancetvisje en alle vertebraten is ‘achter’ Hox sterk uitgebreid, tot Hox9-14. ‘Midden’ Hox geeft via een volgende genduplicatie Hox3, en door verdere genduplicaties Hox4-8. Hox1-5 laten dezelfde genduplicaties zien in insect en chordaat – wat betekent dat het heel vroege genduplicaties zijn. Hox6 gaat een eigen weg in het insect, en Hox8 van het lancetvisje laat een eigen duplicatie zien in Drosophila (Ryan et al 2007).
Trichoplax adhaerens is het enige beestje van de Placozoa, en wordt ingedeeld tussen de sponzen en alle andere beesten (die met een zenuwstelsel). Trichoplax is anatomisch het envoudigste beestje. Trichoplax heeft één Hoxgen, Trox-2, dat overeenkomt met Hox2. Sponzen hebben geen Hox, wel Parahox uit dezelfde genfamilie.
De genfamilie Hox maakt deel uit van een grotere genfamilie, allemaal genen die voor eiwitten coderen die andere genen reguleren. De functie van de genen is vaak min of meer overeenkomstig: signalering bij Hox en verwante families, hormoon bij de steroïden, zuurstofoverbrenging bij de globines.
Er zijn ook genduplicaties waarbij het gedupliceerde gen een totaal andere functie krijgt. Voorbeelden zijn te vinden bij de lenseiwitten van het vertebratenoog.

Figuur 8. De lenseiwitten zijn afkomstig van eiwitten met een andere functie. De ?-crystallines zijn afkomstig van hitteschokeiwitten, bijvoorbeeld. (Figuur 3.25 uit Freeman & Herron 2007. )
Er zijn vele genfamilies. David & Alm (2011) keken naar 3983 genfamilies, over alle levende organismen, en probeerden na te gaan wanneer de genfamilie ontstaan was. Belangrijke genetische innovatie komt uit een korte periode omstreeks 3.25 miljard jaar geleden, in het Archaean. Toen ontstond 27% van de huidige genfamilies. Welke genfamilies? Genfamilies die te maken hebben met elektronentransport en ademhaling. Genfamilies die met RNA of DNA te doen hebben komen als nog ouder uit de studie.
III-4 Exon shuffling
Genen bij beesten / planten / eukaryoten bestaan uit exonen die voor eiwit coderen, gescheiden door niet vertaalde delen DNA. Een exon komt vaak overeen met een eiwitdomein. Zo’n eiwitdomein blijkt vaak voor te komen in een aantal eiwitten, en het overeenkomstige exon dan in een aantal genen. Exon shuffling is een specialiteit van beesten, hun manier om het aantal eiwitten te vermenigvuldigen.
Figuur 9 De eiwitten die zorgen voor bloedstolling zijn opgebouwd door exon shuffling. (Graur en Li 2000 fig 6.13)
III-5 Nieuwe genen
Nieuwe genen ontstaan ook zonder een voorganger die er erg op leek. Een stuk niet-coderend DNA kan ingeschakeld worden, of een stukje DNA kan volledig omgebouwd wordne. De anti-vries bij zuidpool vissen is het bekendste. Er was in december 2010 een voorbeeld van een nieuw anti-vries gen dat samengeclubd was uit stukjes van andere genen. Zie ook hier.
III-6 Insertie, deletie, translocatie
Verschijnen, verdwijnen, van plaats veranderen van stukjes DNA.
III-7 Chromosoomverandering
Chromosomen kunnen in de delingsfase gekleurd worden en dan vertonen ze een aantal bandjes. De chromosomen van soorten kunnen vergeleken worden aan de hand van die bandjes. Er is precies te zien welk stukken chromosoom met elkaar overeenkomen. Zo is te zien dat het basisaantal chromosomen bij de roofdieren 2n=38 is, een aantal dat bij de wasbeerachtigen, oa de ringtail voorkomt. De kat heeft 2n=44 chromosomen. De eerste drie, de langste, chromosomen van de ringtail hebben het centromeer in het midden. Die drie zijn gesplits, elk in twee chromosomen met het centromeer aan het eind. Zo kom je van n=19 naar n=22.
Ook kan een hypothese over het basisaantal chromosomen voor de gewervelde dieren opgesteld worden: n=12.
Fig 10 Vertebrate chromosome set (Jaillon et al 2004)
III-8 Polyploidie
Dieren doen aan exon shuffling, planten aan polyploïdie. Muizenoren zijn berucht: 18, 27, 36, 45, 54 chromosomen. Paardenbloem is diploied of triploied – ten noorden van de Rijn. Lagere aantallen chromosomen planten zich seksueel voort, de hogere aantallen parthenogenetisc, aseksueel. Bramen zin berucht: een complex van polyploiden die onderling hybridiseren..
III-9 Whole genome duplication
Duplicatie van het volledige chromosoom (WGD) komt zo af ten toe voor. Bij de planten aan de basis van de zaadplanten, aan de basis van de bedektzadigen, nog een verdrievoudiging aan de basis van de ‘echte’ dicotylen, twee keer WGD in de lijn naar de grassen, twee keer bij de kruisbloemigen. Dat zijn de gevallen die in de figuur staan, de lijst zal nog wel langer worden.
Figuur 11. Verdubbeling van het volledige genoom in de landplanten. (Jiao et al 2011).
Aan de basis van de gewervelde dieren zitten er twee verdubbelingen van het genoom, vermoedelijk voor de splitsing tussen de prikken en de gewervelde dieren met kaken. Binnen de vissen is er een derde verdubbeling, aan de basis van Teleosten, de grootste groep van de straalvinnige vissen. (zie hier voor de indeling van de vissen).

Figuur 12 Verdubbeling van het volledige genoom bij de vissen. (Mulley & Holland 2004).
III-10 Epigenesis
Er zijn een aantal zaken die genexpressie beïnvloeden, maar zelf niet direct uit het DNA volgen.
Het milieu waarin de moeder leeft is bij de placentale zoogdieren van invloed op de standaard waarop het metabolisme van het kind gezet wordt.
Genen kunnen gemethyleerd worden. Dat beïnvloedt hun activiteit.
III-12 Niet-coderend DNA
Het meeste DNA in organismen met een celkern codeert niet voor eiwitten; welk percentage dat is verschilt tussen soorten. Een deel van het niet-coderende DNA bestaat uit promoters en na transcriptie regulerend RNA. Hoewel het overige niet-coderende DNA deels ook wordt overgeschreven naar RNA, is er geen functie voor aan te geven. Bij mensen bestaat veel niet-coderend DNA uit geïnactiveerde transposable elementen.
III-13 Transposable elements
Uitleg op Wikipedia: zie hier en hier.
Transposable elements zijn ‘zelfzuchtige genetische elementen’, stukjes DNA die voor zichzelf begonnen zijn. Actieve, rondspringende TE’s zijn schadelijk, omdat ze willekeurig genen uitschakelen. Het gastheer genoom deactiveert TE’s na verloop van tijd. Sommige DNA sequenties afkomstig van TE’s zijn ingeschakeld in het gasteergenoom als nieuwe promoters van gastheergenen. De expressie van het gastheergen wijkt weinig af van de oorspronkelijke expressie met de oorspronkelijke promoter. (Cohen et al 2009, Huda et al 2011).
IV Selectie
Selectie is er altijd en overal. Formeel is selectie een wiskundige relatie tussen een aantal grootheden, die altijd tussen twee generaties bestaat. Een wiskundige relatie? Ja, met statistische grootheden. Geen biologie? Ja, als achtergrond bij een van de statistische relaties.
Welke relatie tussen welke grootheden? In een populatie kan voor elk individu een kenmerk gemeten worden: lengte, of genexpressie tijdens de eerste week na de geboorte, of aantal zenuwcellen in een vierkante centimeter huid. Ook kan voor elk individu het aantal kinderen geteld worden, over het hele leven: dat getal noemen we fitness. Dan kunnen twee zaken berekend worden. De eerste is de variantie in de kenmerkwaarde, en omdat het de variantie in een waargenomen, dus fenotypisch kenmerk is, heet dat de fenotypische variantie. De tweede is de covariantie tussen kenmerkwaarde en fitnesswaarde. Het gemiddelde voor het kenmerk aan het begin van de generatie is bekend. Het kenmerkgemiddelde aan het einde van dezelfde generatie is te berekenen, en een beetje wiskunde vertelt dat:
de verandering in het gemiddelde – (dus het gemiddelde op het einde van de generatie min het gemiddelde aan het begin van de generatie) – gelijk is aan de covariantie van fenotypisch kenmerk en fitness, gedeeld door de gemiddelde fitness.
Ik wil niet steeds: gedeeld door gemiddelde fitness schrijven. Als ik de fitness van elk individu deel door de gemiddelde fitness, krijg ik relatieve fitness. De verandering in gemiddelde is gelijk aan de covariantie tussen fenotypische kenmerkwaarde en relatieve fitness: OK?.
Sommige individuen hebben kinderen (anders was alle fitness gelijk aan 0 en hield de zaak op). Maak een grafiek met de fenotypische waarde van de ouders op de x-as en de kinderen op de y-as. Trek een regressielijn. Dan kun je de gemiddelde kenmerkwaarde aan het begin van de volgende generatie aflezen. Die gemiddelde kenmerkwaarde kan anders uitvallen dan de gemiddelde kenmerkwaarde aan het begin van de oudergeneratie:
de verandering in het gemiddelde – (dus het gemiddelde aan het begin van de kindgeneratie min het gemiddelde aan het begin van de oudergeneratie) – gelijk is aan de covariantie tussen fenotype en relatieve fitness in de ouders maal de regressiehelling van kinderen op ouders.
Dit gaat altijd op. De regressielijn tussen ouders en kinderen geeft erfelijkheid in de ruimste zin. Het verband tussen ouders en kinderen mag ook liggen in het erven van een territorium; maar meestal gaat het om genetica, en is de regressielijn het gevolg van genetische verschillen in de populatie. Hoe het met genetica en DNA te maken heeft doet er niets toe. Het is een statistisch verband, en wat de mechanistische achtergrond daarvan is, is bijzaak. Dus alles van puntmutatie via epistasis of epigenetics tot whole genome duplication, van genexpressie tot gedrag: alles in dezelfde soep, geeft een genetische regressie van kenmerkwaarde in de kinderen op kenmerkwaarde in de ouders. Het is belangrijk dit te beseffen: de covariante waar het om draait is het eindresultaat van alle genetische, ontwikkelings, oecologische, gedrags-processen, en de manieren waarop dat eindresultaat bereikt wordt doen er niet toe. Dit betekent dat selectie over alles heen gaat, hoe ontwikkelingsprocessen of genetische rommel, of epigenetische warrigheden ook werken. Onontkoombaar.
Een beetje omzetten van de algebra levert het volgende op (ik wil ooit wel de algebra leveren, als iemand interesse heeft):
de verandering in het gemiddelde tussen twee generaties gelijk is aan de genetische covariantie tussen fenotype van de ouders en relatieve fitness van de ouders.
Er treedt selectie naar een hogere kenmerkwaarde op als die covariantie groter dan nul is. Er treedt selectie naar een lagere kenmerkwaarde op als die covariantie kleiner dan nul is. Er treedt geen selectie op als die covariantie gelijk aan nul is.
Biologisch is het interessant waar die covariantie tussen fenotype en fitness vandaan komt, welke ontwikkelingsbiologie, oecologie van de soort en gedrag tot een bepaalde covariantie leiden. Biologisch is het interessant of de langetermijnwaarde van die covariantie positief of negatief is. Dan gaat het over het verband tussen hoe het beest eruit ziet, zijn fenotype, en hoe het functioneert in zijn omgeving. Dus over hoe je aan zo’n covariantie komt. Dat is waar de studie van natuurlijke selectie zich mee bezig houdt.
Vier punten.
Ten eerste het bekende lesmateriaal. Er bestaat een één-locus model met drie genotypen en hun drie bijbehorende fitnessen dat altijd als lesmateriaal voor selectie gebruikt wordt. Uitschrijven laat zien dat dat een speciaal geval van de covariantieformulering is. Dit lesmateriaal moet niet als de standaardomschrijving van selectie gezien worden.
Ten tweede gaat het hier nu even om één kenmerk. Hetzelfde model kan ook voor een aantal kenmerken tegelijk – krijg je matrixalgebra. Het kan ook voor een aantal kenmerken die zelf een functie van het milieu zijn – krijg je een matrixfunctie. Met andere woorden, al die ontwikkelingszaken als allometrie of zelfsorganisatie of fenotypische plasticiteit of modules waarbij integratie tussen kenmerken optreedt valt ook onder dezelfde statistische omschrijving van selectie.
Ten derde, selectie is een statistische verband, en niet een zelfstandig iets, niet een soort ‘kracht’. Elke formulering van het soort: “als iets ergens goed voor is, zorgt selectie er wel voor dat we het tegenkomen”, “if there was selective value in trait X, then natural selection would have ensured it arose” is dwaasheid.
Ten vierde, selectie leidt niet per definitie tot een ‘leuk’ beest. Het ligt er maar aan wat die covariantie tussen fenotype en fitness is. Bij de gerenuk die veel moeite voor zijn blaadjes moet doen zal er een positieve covariantie zijn tussen een speciale constructie in de achterpoten om rechtop te kunnen staan en fitness, en bij Sacculina die in zijn voer zwemt zal de covariantie tussen het hebben van een zeepok uiterlijk en het aantal eieren negatief zijn.
V Veranderingen en patronen
De vraag is of het ontstaan van de grote patronen (die we HIER gezien hebben) te volgen is uit wat we weten over de huidige waargenomen mogelijkheden voor biologische veranderingen. Daarbij is nodig te onthouden dat we geen volledig inzicht hebben in de interactie tussen ontwikkeling en erfelijkheid: verre van dat. Terwijl juist begrip van de interactie tussen ontwikkeling en erfelijkheid nodig is.
Veranderingen komen bij beesten door ontwikkeling, erfelijkheid en toeval. In elk boek is terug te vinden dat verandering binnen en tussen de patronen neerkomt op verandering in de ontwikkeling. Wat doet selectie dan? Selectie – die covariantie tussen fenotype en fitness – zorgt voor het in elkaar passen van structuur en functie, voor ontwerp. Ontwerp? Ja: ontwerp, design. Niet intelligent design, dat is iets volslagen anders. In de biologie is design, ontwerp, het samenvallen van structuur en functie.
Veranderingen in erfelijkheid en in ontwikkeling veranderen de beesten. Daar begint het mee.
VI Hebben grote veranderingen te maken met grote patronen?
Nee, en ja.
De grootste verandering die boven genoemd staat is de verdubbeling van het gehele genoom (WGD zie III.9). Die twee verdubbelingen van het gehele genoom aan de basis van de gewervelde dieren, is dat de oorzaak van het ontstaan van het bouwplan van de gewervelde dieren? Ik vermoed van niet: binnen de vissen is er een derde verdubbeling, aan de basis van Teleosten, de grootste groep van de straalvinnige vissen. Dan is het bouwplan voor vissen er al.
Hetzelfde voor de planten: twee keer volledige genoomduplicaties (WGD’s) aan de basis van de orde kruisbloemigen, en binnen de kruisbloemigen trouwens nog een derde, bij een familie. Dit volledige genoom duplicatie aan de aan de basis van de zaadplanten, aan de basis van de bedektzadigen, nog een verdrievoudiging aan de basis van de ‘echte’ dicotylen. Het bouwplan bestaat.
Eerder lijkt het me dat volledige genoom duplicatie (WGD) staat aan de basis van de diversiteit, niet aan de basis van het bouwplan. WGD geeft veel overschot aan genetisch materiaal, zodat er veel diversiteit mogelijk is. WGD veroorzaakt niet een bouwplan of een verandering in een bouwplan: WGD maakt het succes van het bouwplan mogelijk, door alle genetische variatie die mogelijk is.
De grote belangrijke groepen die we zien zijn natuurlijk de groepen die succes hebben gehad: veel diversiteit, veel niches. Veel genetische variatie is een van de routes naar succes. De route naar ontstaan van een bouwplan en naar succes van een bouwplan in niet per definitie dezelfde.
Volledige genoom duplicatie is niet nodig voor succes van een groep: bij de kreeften of insecten heb ik er nooit over gehoord.
*****************************Edward O. Guerrant, Jr, 1982, Neotenic Evolution of Delphinium nudicaule (Ranunculaceae): A Hummingbird-Pollinated Larkspur. Evolution 36: 699-712 ROBERT A. BARTON, 2006. Primate Brain Evolution: Integrating Comparative, Neurophysiological, and Ethological Data. Evolutionary Anthropology 15:224–236 Hans Meinhardt, Przemyslaw Prusinkiewicz , Deborah R. Fowler , The Algorithmic Beauty of Sea Shells. Springer; 3 edition (February 12, 2003) ISBN-13: 978-3540440109 J. D. Murray, How the leopard gets its spots, Scientific American, March 1988, p. 80En boek van dezelfde naam 1988.http://www.authorstream.com/Presentation/Denise-37681-Turing-Patterns-Animal-Coat-Coats-Alan-1912-1954-James-Murray-author-in-as-Entertainment-ppt-powerpoint/http://www.eb.tuebingen.mpg.de/departments/former-departments/h-meinhardt/shells.htmlhttp://www.eb.tuebingen.mpg.de/departments/former-departments/h-meinhardt/shell-book.html Sears KE, Behringer RR, Rasweiler JJ, et al.2006. Development of bat flight: Morphologic and molecular evolution of bat wing digitsPROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA 103: 6581-6586 Nielsen C, 2010. all: Xenoturbella is an acoelomorph! EVOLUTION & DEVELOPMENT 12 : 241-243 Graur en Li 2000 Fundamentals of Molecular Evolution 2de drukblz 283-286`exon shuffling, fig 6.13, blood coagulatie Deng C, Cheng CHC, Ye H, et al. 2010. Evolution of an antifreeze protein by neofunctionalization under escape from adaptive conflict . PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA 107 :21593-21598 .

Jaillon et al 2004. Genome duplication in the teleost fish Tetraodon nigroviridis reveals the early vertebrate proto-karyotype. Nature 431: 946-957 Schmidt PS, Zhu CT, Das J, et al. 2008. An amino acid polymorphism in the couch potato gene forms the basis for climatic adaptation in Drosophila melanogaster. PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES OF THE UNITED STATES OF AMERICA : 105 : 16207-16211 RYAN JF, 2007. Pre-Bilaterian Origins of the Hox Cluster and the Hox Code: Evidence from the Sea Anemone, Nematostella vectensis . PLOS ONE 2 : ARTN e153 2007 David LA, Alm EJ (2011) Rapid evolutionary innovation during an Archaean genetic expansion. Nature 469(7328):93-6. Cohen CJ, Lock WM, Mager DL ,2009. Endogenous retroviral LTRs as promoters for human genes: A critical assessment . GENE 448: 105-114 Huda A, Bowen NJ, Conley AB, et al. 2011. Epigenetic regulation of transposable element derived human gene promoters GENE 475 : 39-48
Cursus van Raoul Van Damme UIA http://webh01.ua.ac.be/funmorph/raoul/ebc/h3.doc
Hoofdstuk 3.Macro-evolutie De evolutionaire processen die zich uitstrekken over een geologische tijdsschaal, en die leiden tot het ontstaan van opmerkelijk verschillende genera en hogere taxa, vat men samen onder de noemer “macro-evolutie”. Macro-evolutie is voornamelijk het domein van paleontologen, die fossiel materiaal gebruiken om langdurige trends en belangrijke gebeurtenissen (vb. adaptieve radiaties, massa-extincties, onregelmatigheden in de snelheid van evolutie) te bestuderen.Volgens Gould (2002) kan evolutie bestudeerd worden over drie grote tijdsvensters.
Het eerste tijdsvenster betreft perioden van tientallen tot duizenden jaren; dit is het domein van de populatie-genetica en aanverwante richtingen.
Het tweede venster omvat perioden van miljoenen jaren, perioden waarover snelheden van speciatie en extinctie kunnen geschat worden en vergeleken tussen verschillende groepen van organismen.
Het derde venster toont perioden van tien miljoen tot honderd miljoen jaren, en wordt gemarkeerd door een reeks massa-extincties. Macro-evolutionaire studies bekijken de geschiedenis van het leven door de laatste twee vensters.3.1. Snelheid van evolutie
3.1.1. Schatten van de snelheid van evolutieDe snelheid waarmee een kenmerk verandert in de loop van de evolutie wordt uitgedrukt in Darwin (d). Een snelheid van één Darwin betekent een verandering met een factor e (2.718) in 1 miljoen jaar. De evolutiesnelheid r wordt dus gedefinieerd alswaarbij x1 en x2 de gemiddelde afmetingen van het kenmerk zijn op tijdstip 1 en 2, en t de tijdspanne verstreken tussen beide punten in de tijd (in miljoenen jaren). Bijvoorbeeld, in de laatste 1 000 000 jaren steeg de herseninhoud in het genus Homo van 900 cm³ tot 1400 cm³ en dus bedroeg de evolutiesnelheid Er wordt op een logaritmische schaal gewerkt omdat anders de snelheid van evolutie zou lijken toe te nemen naargelang het bestudeerde organisme groter wordt. Een soort op 1 miljoen jaar tijd 1% aan massa wint, groeit zo steeds aan 0.00995 darwin. Zonder logaritmen zou de evolutiesnelheid van die soort 0.01 bedragen wanneer de soort van 1 g naar 1.01 g evolueert, maar 1 wanneer de soort groeit van 100g naar 101g. De logaritmen zorgen er dus voor dat eenzelfde proportioneel increment eenzelfde snelheid oplevert.Informatie over de snelheid van evolutie komt van drie fronten. De meest voor de handliggende bron is het fossiele materiaal. Hier worden metingen aan morfologische karakteristieken gerelateerd aan de ouderdom van de fossielen, meestal bepaald a.d.h.v. radioactieve dateringsmethoden. Een tweede bron van informatie vormen de veranderingen in kenmerken van soorten die in een recent verleden een nieuw gebied hebben gekoloniseerd. De grootte van verschillen tussen de nakomelingen van de kolonisators en die van de ouderlijke populatie, gedeeld door de tijd verstreken sinds kolonisatie, geeft eveneens een idee van de evolutiesnelheid. Een derde bron wordt gevormd door selectie-experimenten.Schattingen van evolutiesnelheden variëren tussen taxa, tussen kenmerken en tussen perioden. Op het eerste gezicht lijken mammalia bijvoorbeeld sneller te evolueren dan bivalven. De snelheid van verandering bij de mammalia lijkt het hoogst in het Holoceen, het laagst in het Tertiair en intermediair in het Pleistoceen. Dergelijke verschillen werden vaak geïnterpreteerd als het gevolg van verschillen in selectiedruk. Kurtén (1960), bijvoorbeeld, weet de hoge evolutiesnelheden in het Pleistoceen en het Holoceen aan de periodieke glaciaties, en aan de radicale milieuveranderingen die optraden na de recentste terugtrekking van de ijskappen. De vergelijking van evolutiesnelheden berekend over verschillende tijdspannen is echter een hachelijke onderneming. Dit bleek uit de compilatie die Philip D. Gingerich (1983) maakte van gegevens over de snelheid van evolutie in de literatuur. De compilatie omvat data over meer dan 500 kenmerken. Eén van de opvallendste resultaten van de literatuurstudie was namelijk dat er een omgekeerd evenredig verband bestaat tussen de berekende evolutiesnelheid en de tijdspanne waarover de snelheid berekend was.Dit verschijnsel is waarschijnlijk een gevolg van fluctuaties in de richting waarin evolutie verloopt.

Overzicht van evolutiesnelheden uit verschillende typen studies (naar Gingerich 1983).tijdsinterval evolutiesnelheid (d)
n range x range x
Selectie-experimenten
8 1.5-10 j 3.7 j 12000-200000 58700
Kolonisaties 104 70-300 j 170 j 0-79700 370
Post-pleistocene mammalia
46 1000-10000j 8200 j 0.11-32.0 3.7
Fossiele invertebraten
135 0.3-350 Mj 7.9 Mj 0-3.7 0.07
Fossiele vertebraten
228 8000j-98Mj 1.6 Mj 0-26.2 0.08
Totaal 521 1.5 j-350Mj 0.2 Mj 0-200000 0.73
Wanneer de richting van evolutie fluctueert, zal de schatting van de evolutiesnelheid dalen met toenemende lengte van het tijdsinterval waarover de snelheid berekend wordt. Vergelijkingen tussen evolutiesnelheden zijn dus slechts zinvol wanneer ze over vergelijkbare tijdsintervallen gebeuren. De snelheden in bovenstaande tabel suggereren bijvoorbeeld dat vertebraten 1.4 keer sneller zouden geëvolueerd zijn dan invertebraten, maar de intervallen waarover de veranderingen voor vertebraten gemeten werden zijn gemiddeld kleiner dan die voor invertebraten. Na correctie voor dit verschil in tijdspanne lijken invertebraten zelfs sneller te evolueren dan vertebraten.Een tweede opvallend resultaat was het verschil in schattingen bekomen uit de verschillende typen van studies. De snelheden berekend uit artificiële selectie-experimenten of uit de analyse van natuurlijke ecologische kolonisaties zijn aanzienlijk hoger dan die berekend uit het fossiele materiaal. Hiervoor zijn twee verklaringen mogelijk. Enerzijds kunnen de hoge snelheden uit de eerste twee typen datasets een gevolg zijn van extreem hoge selectie-intensiteiten. Anderzijds kunnen zij een artefact zijn van de korte tijdspanne waarover gemeten werd. Misschien volgde evolutie in die korte tijdspanne één richting, en niet het zig-zag pad dat kenmerkend is voor langere perioden.
3.2. De theorie van de onderbroken evenwichten (punctuated equilibria)In paleontologische reeksen ziet men vaak dat plots een nieuwe soort verschijnt, die vervolgens gedurende een tijdje vrijwel onveranderd blijft voortbestaan, om dan even plots opnieuw te verdwijnen. Dikwijls duikt dan een nieuwe, verwante soort op, maar zonder veel overgang tussen de voorouder en nakomeling. Darwin, en vele paleontologen na hem, weten dit ogenschijnlijk springerige evolutiepatroon aan de onvolledigheid van het fossiele materiaal. Zij zagen evolutie als een gradueel gebeuren, waarbij de veranderingen aan een vrij constant tempo geschieden, en waarbij nieuwe soorten geleidelijk uit voorouderlijke soorten ontstaan. Tegenwoordig noemt men deze zienswijze ‘fyletisch gradualisme’.
Twee modellen omtrent het tempo van evolutie : het model van het onderbroken evenwicht (rechts ) en dat van het fyletisch gradualisme.(linnks )
In 1972 formuleerden Niles Eldredge en Stephen Jay Gould een alternatieve verklaring voor het springerige karakter van paleontologische reeksen. Volgens hun theorie van de onderbroken evenwichten (Eng: punctuated equilibria) verloopt evolutie
De evolutie van een kenmerk over de tijd. Fluctuaties in de richting van evolutie maken dat de berekende snelheid lager wordt naarmate men langere tijdsintervallen beschouwt.In dit hypothetische voorbeeld bedraagt de snelheid waarmee het kenmerk toeneemt +0.27 darwin indien berekend tussen tijdstippen 1 en 3, -0.60 tussen 3 en 6, +2.20 tussen 6 en 7, en +0.25 tussen 1 en 7.
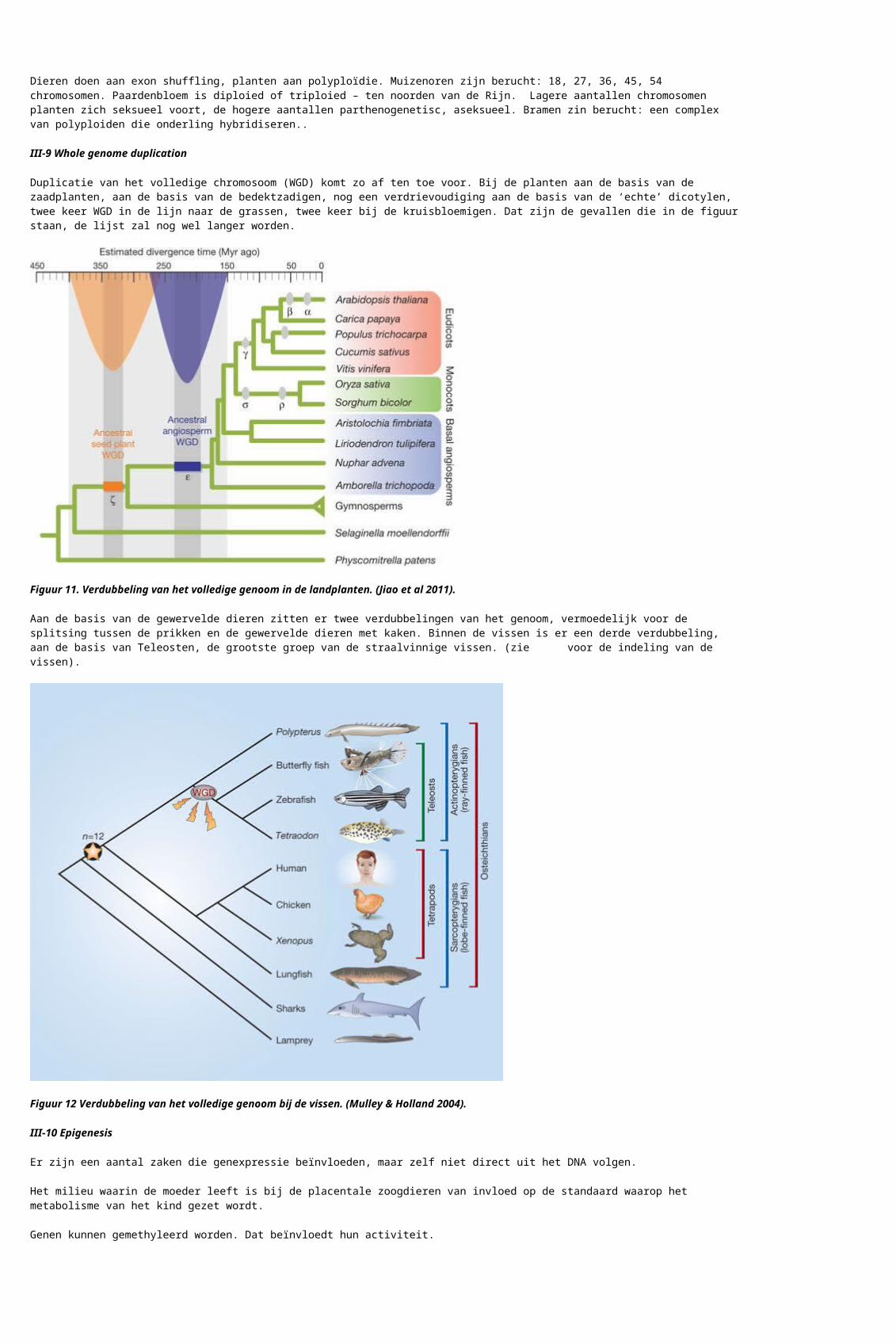
uitzonderlijk snel tijdens de speciatie, maar vervolgens ondergaat de populatie vrijwel geen veranderingen meer en verkeert ze in een vorm van ‘stasis’.

Speciatiemechanismen volgens het onderbroken evenwichtsmodelHet favoriete speciatiemodel van Eldredge en Gould is dat van perifere isolatie, waarbij een nieuwe soort allopatrisch en in kleine, geïsoleerde populaties wegevolueert van de ouderpopulatie. Oorspronkelijk zagen zij er geen graten in om de snelle evolutie tijdens de speciatie toe te schrijven aan klassieke processen zoals selectie en drift, maar in nieuwere versies van de theorie wordt het plotse uiteenwijken van de populaties toegeschreven aan bijzondere condities.Eén zo’n conditie is de desintegratie van de homeostase. In normale omstandigheden bevoordeelt stabiliserende selectie het ontstaan van homeostatische mechanismen. In extreme condities breken deze mechanismen op, en kan een ongewoon brede diversiteit aan fenotypen ontstaan. Misschien kan op dat moment genetische variatie, die voorheen door de homeostatische mechanismen gemaskeerd werd, tot uiting komen, waardoor het pad van evolutie verlegd wordt. Bovendien is gebleken dat onder stress merkwaardige genotypische effecten kunnen ontstaan. De mate van recombinatie, bijvoorbeeld, kan onder invloed van extreme, abnormale temperaturen verhogen.Een ander onorthodox idee van Gould om de plotse sprintjes van evolutie tijdens de speciatie te verklaren, is een moderne versie van de ‘hopeful monster’ hypothese van Goldschmidt. De evolutieve doorbraken zouden in dit geval komen van macro-mutaties, mutaties die optreden in de genen die de ontogenetische ontwikkeling sturen of andere belangrijke processen reguleren. Hierdoor zouden plots sterk aberrante fenotypen ontstaan. Macro-evolutie zou dan grotendeels een niet-adaptef gebeuren zijn, en adaptieve evolutie zou zich beperken tot de eerder geringe correcties binnen populaties. Stasis : stabiliserende selectie of evolutionaire remmen ?Het is momenteel onduidelijk waarom fossiele reeksen lange tijd vrijwel onveranderde fenotypen opleveren. Stabiliserende selectie lijkt de meest plausibele verklaring. Het is een welbeschreven en goed gedocumenteerd fenomeen. Zolang een populatie binnen bepaalde omgevingsextremen kan blijven, zal hetzelfde genotype bevoordeeld blijven. Worden de grenzen overschreden, komt de mogelijkheid voor plotse evolutie en speciatie vrij.De alternatieve verklaring voor het optreden van stasis, genetische remming, is minder onderbouwd. Volgens deze hypothese zou een soort tijdens de perioden van stasis niet evolueren omdat er geen genetische variatie voor handen is. In kleine, perifere populaties zou latente, of nieuwe, genetische variatie plots tot uiting komen en aanleiding geven tot snelle evolutie en speciatie. Deze hypothese voorspelt dat het kenmerk in kwestie een erfbaarheid nul heeft. Het is uiteraard onbegonnen werk om de erfbaarheid van fossiele kenmerken te schatten, maar vergelijking met analoge eigenschappen in moderne soorten leert dat dit zelden het geval is.Geografische variatie in het kenmerk is een bijkomend argument tegen de hypothese van stasis door evolutionaire remming.Gould versus DarwinIn een ongemeen strijdlustig artikel heeft Gould (1980) de theorie van de onderbroken evenwichten de ‘doodsteek’ van het neo-Darwinisme genoemd. We bespreken twee twistpunten.In de eerste plaats claimt Gould dat zijn model met tempowisselingen regelrecht indruist tegen het gradualisme van Darwin en de neo-Darwinisten. Dit berust wellicht op een misverstand. Darwin beklemtoonde inderdaad herhaaldelijk het trage, graduele karakter van evolutie, maar hij bedoelde daar waarschijnlijk iets anders mee dan het extreme fyletische gradualisme waartegen Gould zich afzet. Darwin sprak over geleidelijkheid wanneer het ging om het ontstaan van adaptaties, in het bijzonder complexe adaptaties. Hij moest er de mensen van overtuigen dat adaptaties konden worden bereikt via kleine evolutionaire tussenstadia. Wanneer hij het expliciet over het tempo van evolutie had, opperde Darwin zelfs de mogelijkheid van snelle veranderingen tijdens de speciatie en perioden van tragere evolutie daartussen.Een tweede beschuldiging van Gould aan het adres van de neo-Darwinisten was dat hun moderne synthese reductionistisch was, in de zin dat ze slechts één mechanisme zag voor evolutie : natuurlijke selectie onder allelen met differentiële fitness. Dit mechanisme wordt verantwoordelijk geacht voor de microevolutie binnen populaties, voor speciatie, en voor het ontstaan van de hogere taxa. De moderne synthese zou macro-evolutie reduceren tot geaccumuleerde micro-evolutionaire gebeurtenissen.De feitenOf de theorie van de onderbroken evenwichten nu in contradictie is met de neo-darwinistische synthese of niet, het blijft een interessante vraag of evolutie geleidelijk gebeurt, of getypeerd wordt door tempoveranderingen met perioden van abrupte veranderingen en perioden van stasis. Ondertussen gaan de meeste paleobiologen akkoord dat beide theorieën, zoals hierboven beschreven, slechts uitersten zijn in een continuüm van mogelijkheden. De vraag is dus verschoven naar de frequentiedistributie van evolutiesnelheden.
De beste manier om de vraag op te lossen is natuurlijk door naar de feiten te zoeken. Dit is echter niet zo eenvoudig : een goede test van beide theorieën vergt het opmeten van grote reeksen fossielen in een zo compleet mogelijke stratigrafische reeks * Caraibisch mosdiertje .
http://porites.geology.uiowa.edu/database/bryozoa/systemat/metrar.htm
Een continuüm van evolutiemodellen verbindt fyletisch gradualisme met het patroon van onderbroken evenwichten.
http://porites.geology.uiowa.edu/database/bryozoa/systemat/metaurgp.gif
M. auriculatumCanu and Bassler, 1923

Zo’n reeks werd gevonden in de mariene afzettingen van het boven-Mioceen tot het onder-Plioceen (8-3.5 miljoen jaar geleden) van de Dominicaanse republiek. Deze lagen bevatten verschillende opeenvolgende populaties van het geslacht Metrarabdotos, een groep van kleine, kolonievormende bentische invertebraten, behorende tot de Bryozoa. Alan Cheetham (1986) mat van elk van 1000 specimens, behorende tot ongeveer 100 populaties, 46 skeletkenmerken op, en vergeleek de snelheid van verandering binnen soorten met die tussen soorten. Zijn resultaten waren sterk in overeenkomst met de theorie van de onderbroken evenwichten. De meeste soorten van het genus bleven nagenoeg onveranderd gedurende perioden van verschillende miljoenen jaren, en nieuwe soorten ontstonden plots, zonder intermediaire overgangspopulaties. Als er toch overgangsvormen gevonden werden, dan verdwenen die snel; ze overleefden gemiddeld slechts 160 000 jaar. Bovendien bleven in een aantal gevallen de ancestrale soorten voortbestaan, samen met de afgesplitste soorten.
http://www.zoology.ubc.ca/~bio336/Bio336/Lectures/Lecture23/Overheads.htmlPatronen van evolutionaire verandering bij Metrarabdotus. De horizontale as stelt de morfologische differentiatie voor, de verticale as is de tijdschaal (naar Cheetham 1986). De Cenozoïsche molluscenfauna van het Turkana-bassinOok Peter G. Williamson (1981) vond aanwijzingen voor het bestaan van evolutionaire tempowisselingen. Hij voerde een morfometrische analyse uit op 3300 molluscen, behorende tot 13 soortlijnen, afkomstig uit de laat-Cenozoïsche afzettingen van het Turkana basin (Noord-Kenia), een ongemeen rijke vindplaats voor goed-bewaarde prosobranchen, pulmonaten en bivalven. Tot de bestudeerde kenmerken behoorden de coördinaten van bepaalde punten op de schelp, de vorm van de schelp, de positie van de indrukken van de sluitspieren en van het ligament. Ondanks grote variatie in ecologie, reproductieve strategieën en grootte van de bestudeerde lijnen, volgden zij allen een opvallend gelijkend evolutionair patroon. Het is opnieuw een lange periode van stasis (3-5 miljoen jaar), die onderbroken wordtTiming van evolutionaire veranderingen bij molluscen uit het Turkana bassin (naar Williamson 1981).
door drie perioden (van elk 5000-50 000 jaar) waarin plots grote morfologische veranderingen optreden, en speciatie plaats vindt. Volgens Cheetham ondersteunt de fylogenetische geometrie van deze molluscen het ‘founder-effect’-model voor speciatie (Mayr 1954). Volgens dit model zou soortvorming gebeuren door geografische isolatie, gepaard gaand met belangrijke omgevingsstress. De perioden van speciatie vallen in dit geval samen met belangrijke lacustrine regressies, waardoor de fauna’s van het bassin onderling geïsoleerd raakten én in moeilijkere milieuomstandigheden terecht kwamen.

Een pleidooi voor fyletisch gradualisme : Welse trilobietenMaar niet alle studies ondersteunen het model van de onderbroken evenwichten. Peter Sheldon (1987) onderzocht een uitzonderlijk rijke fossiele reeks trilobieten in zeven opeenvolgende afzettingen uit het Ordovicium in Wales. Hij controleerde meer dan 15000 individuele fossielen, telde het aantal pygidiale ribben van 3458 specimen behorende tot acht verschillende genera, en kwam tot het besluit dat in elk van de genera het aantal ribben gradueel toenam, en dit over een periode van ongeveer drie miljoen jaar. De overgang gebeurde zo geleidelijk, dat een vroegere indeling van opeenvolgende soorten, die gebaseerd was op het aantal ribben, onhoudbaar bleek. Er waren teveel intermediaire specimens.
http://www.zoology.ubc.ca/~bio336/Bio336/Lectures/
Lecture23/Overheads.html Dat in elk van de acht onderzochte genera trilobieten het aantal pygidiale ribben netto toenam (hier en daar gebeurden wel enkele tijdelijke omkeringen), lijkt erop te wijzen dat natuurlijke selectie, eerder dan drift, verantwoordelijk is voor de verandering. De functionele betekenis van de ribben is onbekend. Misschien verborg elke rib een aanhangsel met een kieuwboog. Misschien zorgden extra ribben gewoon voor meer stevigheid. Het is ook mogelijk dat de ontwikkeling van extra-ribben pleiotropisch verbonden was met een ander kenmerk waarop geselecteerd werd, en dat het aantal ribben zélf adaptief neutraal was, of zelfs ietwat ongunstig.
Sheldon zag in zijn resultaten een argument voor het fyletisch gradualisme, en John Maynard Smith (1987) besloot in een begeleidende commentaar dat er geen reden was om het “nieuwe” paradigma van de onderbroken evenwichten te aanvaarden, laat staan om het neo-darwinisme af te zweren. Maar Eldredge en Gould (1988) gingen niet akkoord. In een antwoord op Sheldons artikel en de commentaar geleverd door Maynard Smith, argumenteren zij dat de veranderingen, genoteerd door Sheldon, té miniem zijn om ons iets te leren over de patronen van de diversificatie van trilobieten. Voor Eldredge en Gould zijn de veranderingen niet meer dan wat knutselwerk, onvergelijkbaar met de grote morfologische veranderingen die de trilobieten ondergingen op andere momenten. Zij zien de periode beschreven door Sheldon als een periode van stasis. De woordenwisseling tussen de betrokken wetenschappers illustreert verschillende aspecten van het dispuut dat in dit hoofdstuk behandeld werd.
3.3. Speciatie- en extinctiesnelhedenTriops cancriformis (Notostraca) bestaat als soort al 200 miljoen jaar.
De soortenrijkdom van verschillende taxa verschilt aanzienlijk tussen taxa en doorheen de geologische tijd. Volgens Stanley (1998) houden de Notostraca, een orde Crustacea, het record van de traagste speciatie. De twee moderne genera zijn minstens 300 miljoen jaar oud, en er zijn geen andere, fossiele genera bekend. De orde heeft altijd maar uit enkele soorten bestaan. Nu zijn het er negen. De ginkgo’s vormen een gelijkaardig voorbeeld. De enige huidige soort, Ginkgo biloba, behoort tot de familie Ginkgoaceae, die voor het eerst opdook in het vroege Jura. Planten bijna identiek aan Ginkgo biloba leefden 120 miljoen jaar geleden. Gedurende gans haar geschiedenis bevatte de familie minder dan 20 soorten, en nooit leefden er meer dan 11 soorten tegelijkertijd (Zhou & Zeng 2003). Aan het andere extreem liggen de cichliden van het Nabugabo-meer, waar in de laatste 4000 jaar wellicht 5 nieuwe soorten ontstaan zijn (Owen et al. 1990).
De diversiteit aan soorten kan gezien worden als een balans tussen speciatie- en extinctie-snelheden. Welke kenmerken bepalen deze balans ?
Ginkgo biloba (Ginkgoaceae)
Kenmerken van organismen die speciatie bevorderen.Organismale kenmerken die reproductieve isolatie bevorderen, kunnen leiden tot een verhoogde speciatiesnelheid. Kandidaten zijn allerlei kenmerken die seksuele selectie bevorderen, of de tussenkomst van andere soorten bij de voortplanting (vb. pollinatie). De beweeglijkheid van organismen kan ook een effect hebben op de speciatiesnelheid. Kenmerken van organismen die extinctie voorkomen.Bijvoorbeeld kenmerken die zorgen voor grote populaties, een grote geografische verspreiding, een groot vermogen tot dispersie, enzovoort. Kenmerken van organismen die nieuwe ‘adaptieve zones’ openen.Sommige kenmerken zijn geassocieerd met hoge speciatiesnelheden, omdat ze een groep toelaten een ganse reeks nieuwe habitaten in te nemen. Voorbeelden zijn de vleugels van vogels en insekten, de eieren van reptielen. Soort-kenmerken.Sommige macro-evolutionairen denken dat speciatie en extinctie kan beïnvloed worden door emergente eigenschappen van een soort in zijn geheel. Voorbeelden zijn het verspreidingsareaal en de graad van versnippering. Als deze kenmerken “erfbaar” zijn over soorten (d.w.z., dochtersoorten lijken op de ancestrale soort), dan zal de proportie van soorten met deze emergente eigenschappen toenemen in de tijd. Dit proces wordt “species-selectie” genoemd (maar moet niet verward worden met groepselectie).
3.4. Massa-extinctiesDe geschiedenis van het leven op aarde wordt gekenmerkt door een aantal perioden waarin opmerkelijk veel soorten uitgestorven zijn. De bekendste van deze catastofale perioden zijn de zogenaamde Perm-extinctie (225 miljoen jaar geleden, op het eind van het Perm en dus op de overgang tussen Paleozoïcum en Mesozoïcum) en de Krijt-extinctie (65 miljoen jaar geleden, op het eind van het Krijt, en dus op de overgang tussen het Mesozoïcum en het Kainozoïcum). Deze twee golven van uitsterven waren bijzonder omvangrijk : tijdens de Perm-extinctie stierven zowat 96% van alle mariene soorten uit, tijdens de Krijt-extinctie zowat 65%.Over de oorzaak van deze cataclysmen is druk gespeculeerd. Tot de meer serieuze mogelijkheden behoren klimaatsveranderingen (vooral afkoeling en verdroging), stijgingen of dalingen van het zeeniveau, predatie, epidemieën en
Evolutie van het aantal ribben bij trilobieten uit het Ordovicium (naar Sheldon, 1987).
Trilobiet

competitie. In 1980 formuleerden Walter Alvarez en co-auteurs een ophefmakende hypothese om de Krijt-extinctie te verklaren. In rotsen uit deze periode troffen ze een aantal zeldzame elementen aan, waaronder iridium. Deze elementen komen ook in hoge concentraties voor in extraterrestrische objecten, en Alvarez wijtte de zogenaamde iridium-anamolie aan de botsing tussen een asteroïd en de aarde. Andere geologen hebben ondertussen geargumenteerd dat het iridium ook een terrestrische oorsprong zou kunnen hebben (vulkanische activiteit), maar de asteroïden blijven de meest aanvaarde verklaring. Volgens Alvarez et al. veroorzaakte de inslag (wiens kinetische energie wordt geschat op het equivalent van 108 megaton) een stofwolk die voor lange tijd alle zonlicht op aarde zou geblokkeerd hebben. Dat alleen al zou het massale uitsterven kunnen verklaren, maar de impact had daarnaast nog andere neven-effecten, waaronder een globale opwarming, zure regen, extreem vulkanisme, en misschien een aarde-omvattende brand. Cruciaal voor Alvarez’ theorie is (1) de tijdsspanne waarover de extincties plaatsvonden (Alvarez voorspelt een zeer plots en synchroon uitsterven) en (2) de aanwezigheid van fysische geologische veranderingen. Op dit moment zijn de meningen verdeeld over hoe goed empirische gegevens hierrond aan de verwachtingen beantwoorden.David Raup en John Sepkoski bekeken de extinctiesnelheden voor alle families van mariene organismen doorheen de geologische tijd en kwamen tot de verrassende vaststelling dat de hierboven geschetste cataclysmen deel uitmaakten van een patroon van cyclische veranderingen, met periodes van massaal uitsterven die terugkeerden om de 26 miljoen jaar. Geologische, statistische en taxonomische problemen ondergraven enigszins de conclusies van Raup en Sepkoski, maar het staat buiten kijf dat massa-extincties een weerkerend fenomeen zijn. De verklaring hiervoor moet wellicht in fysische processen gezocht worden – eerder dan in biologische.
Er is weinig bewijs dat de impact van asteroïden een rol heeft gespeeld bij andere extinctiegolven dan die op het einde van het Krijt. Voor de andere extinctie-perioden werd geen iridium-anomalie gevonden. De idee van een periodische collitie tussen de aarde en grote asteroiden lijkt daarom onwaarschijnlijk.
Sommige massa-extincties vallen samen met tijden van klimaatsverandering en de daarmee gepaard gaande veranderingen in zeeniveau, maar de correlatie is verre van perfect.
Massa-extincties kunnen verband houden met veranderingen in de vorm van de continenten. Om ecologische redenen bestaat er een verband tussen de oppervlakte van een geografisch gebied, en het aantal soorten dat er leeft. Bewegingen van continenten veroorzaken grote veranderingen in de oppervlakte en perimeter van beschikbaar land. Dit idee lijkt een goede verklaring voor de Perm-extinctie (op dat moment kwamen de continenten samen om het supercontinent Pangaea te vormen), maar lijkt minder van toepassing op andere perioden van uitsterven.
De geologische tijdsschaal
Hoofdtijdperk Periode Tijdvak Aanvang miljoen jaar geleden
Cenozoïcum Kwartair Holoceen 0,01
Pleistoceen 1,6
Tertiair Plioceen 5
Mioceen 23
Oligoceen 35
Eoceen 56
Paleoceen 65
Mesozoïcum (Secundair)
Krijt 145
Jura 208
Trias 245
Paleozoïcum (Primair)
Perm 290
Carboon Bovencarboon
Ondercarboon 362
Devoon Bovendevoon
Middendevoon
Onderdevoon 408
Siluur 439
Ordivicium 510
Cambrium 570
Precambrium 4500
http://www.geo.arizona.edu/geo3xx/geo308/FoldersOnServer/2005%20Lecture%20Notes/MicroEvoLectureNotes2005.pdf
Attachment: MicroEvoLectureNotes2005.pdf


















