
Verkennende studie van de regenworm als bodemingenieur in...
93
Faculteit Bio-ingenieurswetenschappen Academiejaar 2014 – 2015 Verkennende studie van de regenworm als bodemingenieur in private moestuinen Jolien Bracke Promotor: Prof. dr. ir. Jan Mertens Co-promotor: Dr. ir. Dieter Anseeuw Co-promotor: ir. Stephanie Schelfhout Masterproef voorgedragen tot het behalen van de graad van Master of Science in de biowetenschappen: land- en tuinbouwkunde
Transcript of Verkennende studie van de regenworm als bodemingenieur in...

Faculteit Bio-ingenieurswetenschappen
Academiejaar 2014 – 2015
Verkennende studie van de regenworm als bodemingenieur
in private moestuinen
Jolien Bracke
Promotor: Prof. dr. ir. Jan Mertens
Co-promotor: Dr. ir. Dieter Anseeuw
Co-promotor: ir. Stephanie Schelfhout
Masterproef voorgedragen tot het behalen van de graad van
Master of Science in de biowetenschappen: land- en tuinbouwkunde


Faculteit Bio-ingenieurswetenschappen
Academiejaar 2014 – 2015
Verkennende studie van de regenworm als bodemingenieur
in private moestuinen
Jolien Bracke
Promotor: Prof. dr. ir. Jan Mertens
Co-promotor: Dr. ir. Dieter Anseeuw
Co-promotor: ir. Stephanie Schelfhout
Masterproef voorgedragen tot het behalen van de graad van
Master of Science in de biowetenschappen: land- en tuinbouwkunde


I
AUTEURSRECHTELIJKE BESCHERMING
De auteur en de promotoren geven de toelating deze masterproef voor consultatie beschikbaar te
stellen en delen ervan te kopiëren voor persoonlijk gebruik. Elk ander gebruik valt onder de beperkingen
van het auteursrecht, in het bijzonder met betrekking tot de verplichting de bron uitdrukkelijk te
vermelden bij het aanhalen van resultaten uit deze masterproef.
Gent, juni 2015
Auteur:
Jolien BRACKE
Promotoren:
Prof. dr. ir. Jan MERTENS
Dr. ir. Dieter ANSEEUW
Ir. Stephanie SCHELFHOUT

II
WOORD VOORAF
Niet zonder enige trots stel ik deze masterproef voor. De totstandkoming ervan was mogelijk dankzij de
deskundige hulp van verschillende personen.
In de eerste plaats wil ik Stephanie Schelfhout bedanken: haar ervaring en kennis waarop ik steeds
beroep kon doen zijn een enorme hulp geweest. Naast assistentie bij veldwerk kon ik rekenen op
zorgvuldig verbeterde proefversies. Zonder haar toegewijde begeleiding was de totstandkoming van dit
werk niet mogelijk en een stuk minder aangenaam geweest.
Mijn dank gaat ook uit naar Jan Mertens wiens kritische insteek zeer op prijs gesteld werd. Dieter
Anseeuw ben ik dankbaar voor het ter beschikking stellen van dit boeiende onderwerp, maar ook voor
het aanreiken van respondenten en interessante literatuur.
Daarnaast wil ik ook Filip Debersaques bedanken voor de financiering van de bodemanalyses. Deze
schenking gaf me de mogelijkheid om dieper te graven naar het hoe en waarom. In deze context bedank
ik graag ook An De Schrijver voor het coördineren van de analyses, alsook Luc Willems van het Labo voor
Bos & Natuur (ForNaLab) en de laboranten van het Laboratorium Toegepaste Fysicochemie (ISOFYS)
voor de uitvoering ervan.
Ook Geert Baert, die me hielp met GIS en tevens de textuurbepaling van de bodemstalen uitvoerde,
verdient een woord van dank.
Vervolgens wil ik de respondenten bedanken die hun moestuin ter beschikking stelden voor mijn
onderzoek. Hopelijk is jullie regenwormpopulatie inmiddels weer op peil!
Tot slot mag ook mijn vader niet op deze pagina ontbreken, aangezien hij me door weer en wind bijstond
tijdens zo goed als al het veldwerk dat in het eerste semester verricht werd. Dit werd ongelooflijk
geapprecieerd.
Jolien Bracke
Gent, juni 2015

III
ABSTRACT
Regenwormen beïnvloeden door nutritionele en graafactiviteiten hun habitat en dat van andere
bodemdieren zodanig dat gesproken kan worden van bodemingenieurs. Door hun bijdrage aan een
vruchtbare bodem zijn ze graag geziene gasten in landbouwsystemen. Ook daarbuiten leveren ze diverse
ecosysteemdiensten zoals erosievermindering en overstromingsregulatie en dragen ze bij aan
biodiversiteit. Hun rol in moestuinen, die in het sterk verstedelijkte Vlaanderen een aanzienlijk deel van
het landschap vertegenwoordigen, werd in deze verkennende studie onderzocht. Daarbij werd rekening
gehouden met zowel beheermaatregelen als bodemparameters. In dit kader werden 37 moestuinen
onderzocht, waarvan 10 permacultuur (bedekte bodem en minimale bodembewerking), 11 ecologisch
en 16 conventioneel (gebruik van kunstmest) beheerd. Tijdens dit onderzoek werden via
mosterdextractie en handsortering 2664 regenwormen die tot 19 verschillende soorten behoren
gecollecteerd. Het gemiddelde totale aantal bedroeg 267 regenwormen m-2 en de gemiddelde totale
massa 78,7 g m-2. Permacultuurmoestuinen bleken (i) de grootste aantallen epigeïsche regenwormen te
bevatten; (ii) het grootst effectief aantal soorten te herbergen; en (iii) een groter totaal aantal
regenwormen te bevatten dan conventioneel beheerde moestuinen. Bijkomend kon geconcludeerd
worden dat overbemesting in Vlaamse moestuinen een actueel probleem is waardoor niet alleen de
plantengroei negatief beïnvloed kan worden maar ook de milieudruk te hoog is. Een regenworm-
vriendelijk beheer in combinatie bemesting op basis van bodemanalyses biedt perspectieven voor zowel
tuinier als milieu. Een samenwerking tussen organisaties die in nauw contact staan met particulieren,
zoals VELT en Tuinhier, en de overheid voor een sensibilisatieactie rond de bemestingsproblematiek in
moestuinen lijkt aangewezen.
Kernwoorden: regenwormen, Lumbricidae, moestuinen, ecosysteemdiensten, overbemesting,
permacultuur
Earthworms influence their own habitat and that of other soil animals by their digging and nutritional
activities. Within this scope they are called soil engineers. Because of their enhancing effects on soil
fertility, they are very welcome in agricultural systems. Also, they deliver important ecosystem services:
they decrease soil erosion, regulate flooding and stimulate the soil food web. In this pilot study we
investigated their impact in vegetable gardens, which represent an important part of the strongly
urbanized Flemish landscape. Management practices as well as soil characteristics were considered. We
examined 37 vegetable gardens: 10 permaculture (covered soil and reduced tillage), 11 ecological and
16 conventional (amendment of inorganic fertilizers) managed vegetable gardens. Earthworm sampling
resulted in the capture of 2664 individuals. Mean total abundance was 267 individuals m-2 and mean
total biomass was 78,7 g m-2. Permaculture vegetable gardens turned out to (i) contain the largest
amount of epigeics; (ii) have the largest effective number of species; and (iii) their earthworm
populations outnumber those from conventional managed vegetable gardens. Furthermore, we
conclude that overfertilization is an actual problem in Flemish vegetable gardens which not only can
have a negative effect on plant growth but also puts a pressure on the environment. Earthworm-friendly
gardening combined with fertilization based on soil analysis opens up new possibilities, both for

IV
gardener and environment. A sensitization concerning the overfertilization issue managed by the
government in cooperation with organisations like VELT en Tuinhier, which are in close contact with
gardeners, seems appropriate.
Keywords: earthworms, Lumbricidae, vegetable gardens, ecosystem services, overfertilization,
permaculture

V
INHOUDSOPGAVE
Auteursrechtelijke bescherming ........................................................................................................................ I
Woord vooraf .................................................................................................................................................... II
Abstract ............................................................................................................................................................ III
Inhoudsopgave .................................................................................................................................................. V
Lijst met figuren ............................................................................................................................................... VII
Lijst met tabellen .............................................................................................................................................. IX
Inleiding ............................................................................................................................................................. 1
1 Literatuur ................................................................................................................................................... 3
1.1 Tuinen .............................................................................................................................................. 3
1.1.1 Inleiding & belang van tuinen in Vlaanderen .............................................................................. 3
1.1.2 Moestuinen in Vlaanderen .......................................................................................................... 4
1.1.3 Ecosysteemdiensten van moestuinen ......................................................................................... 8
1.2 Regenwormen ................................................................................................................................ 11
1.2.1 Inleiding ..................................................................................................................................... 11
1.2.2 Morfologie en determinatiekenmerken .................................................................................... 11
1.2.3 Ecologie ..................................................................................................................................... 13
1.2.4 Sturende factoren ...................................................................................................................... 16
1.2.5 Ecosysteemdiensten van regenwormen.................................................................................... 19
2 Materiaal en methoden ........................................................................................................................... 23
2.1 Onderzoeksveld .............................................................................................................................. 23
2.2 Proefopzet ...................................................................................................................................... 25
2.3 Staalname ...................................................................................................................................... 26
2.3.1 Bodemstaalname en -analyse .................................................................................................... 26
2.3.2 Regenwormstaalname en determinatie .................................................................................... 28
2.4 Statistische analyses ...................................................................................................................... 30
2.4.1 Algemene eigenschappen.......................................................................................................... 30
2.4.2 Nutriënten en regenwormen ..................................................................................................... 30
2.4.3 Diversiteit .................................................................................................................................. 31
3 Resultaten en bespreking ........................................................................................................................ 32
3.1 Algemene moestuineigenschappen ............................................................................................... 32
3.2 Bodem ............................................................................................................................................ 34

VI
3.2.1 Zuurtegraad (pH) ........................................................................................................................ 35
3.2.2 Calciumgehalte ........................................................................................................................... 36
3.2.3 Vochtgehalte .............................................................................................................................. 37
3.2.4 Nitraatresidu .............................................................................................................................. 37
3.2.5 Fosforgehalte ............................................................................................................................. 39
3.2.6 Kaliumgehalte ............................................................................................................................ 40
3.2.7 Magnesiumgehalte ..................................................................................................................... 40
3.2.8 Koolstofgehalte .......................................................................................................................... 41
3.3 Regenwormen ................................................................................................................................. 44
3.3.1 Algemeen ................................................................................................................................... 44
3.3.2 Sturende factoren ...................................................................................................................... 46
Algemene conclusie ......................................................................................................................................... 55
Referentielijst ................................................................................................................................................... 58
Appendix .......................................................................................................................................................... 67

VII
LIJST MET FIGUREN
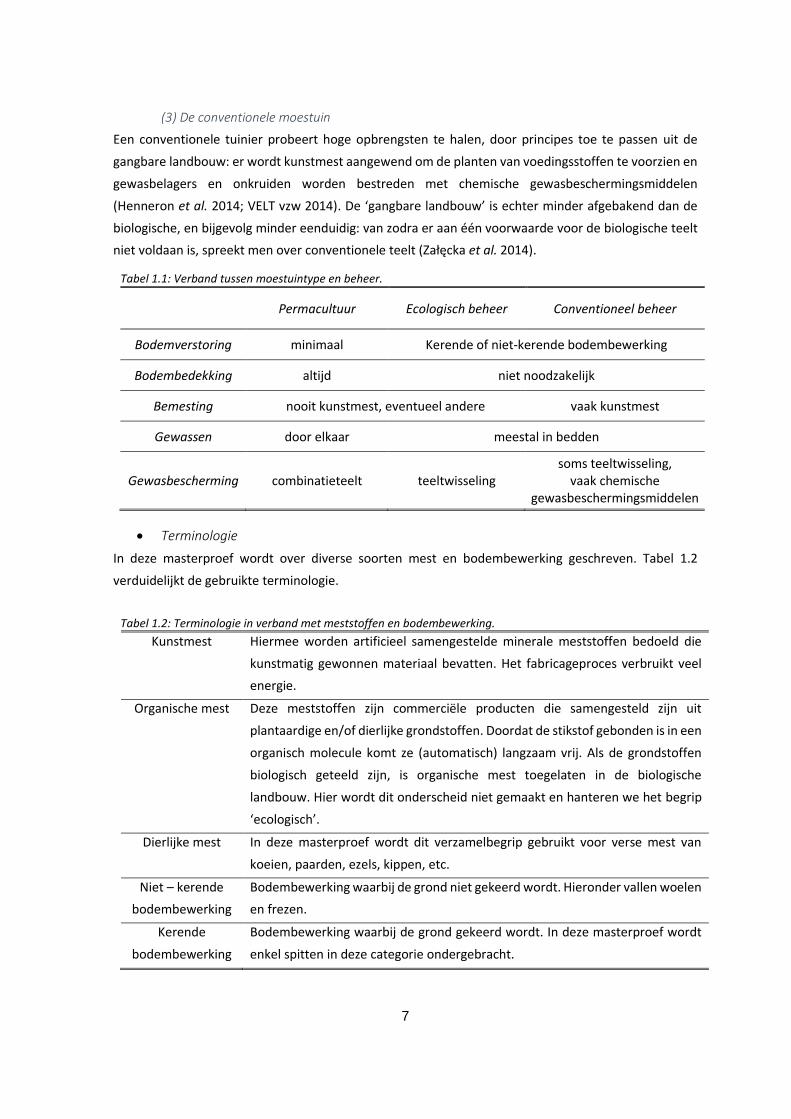
Figuur 1.1: Permacultuurtuinen kunnen natuurlijk (a) of gestileerd (b) ogen, maar de bodem zal nooit
onbedekt zijn zoals in een klassieke moestuin (c). Foto’s werden genomen in respectievelijk tuin 1
(24/10), tuin 3 (5/11) en tuin 7 (26/10). ................................................................................................... 6
Figuur 1.2: Dorsaal zicht van de (a) epilobe en (b) tanylobe kop. Het prostomium is op een verschillende
manier bevestigd (Herwerkt uit Sims & Gerard (1999)). ........................................................................ 12
Figuur 1.3: Determinatiekenmerken van Lumbricidae met van boven naar onder: prostomium (met mond),
borstels, mannelijke porus, clitellum en tubercula pubertatis (Herwerkt uit Edwards en Lofty (1977)).12
Figuur 1.4: De drie ecologische groepen, v.l.n.r. epigeïsch, endogeïsch en anekisch (illustratie: Klaas Van de
Moortel). ................................................................................................................................................. 13
Figuur 1.5: Doorsnede van gangen van anekische regenwormen op 0,30 meter diepte, gefotografeerd op
2/11 in tuin 20. ........................................................................................................................................ 14
Figuur 1.6: pHH2O-bereik waarbij 15 soorten regenwormenvoorkomen. Dendrodrillus rubidus, D. pygmea, A.
icterica en Microscolex phosphoreus zijn niet opgenomen in deze figuur. De pH-waarden zijn
overgenomen van Sims & Gerard (1999). ............................................................................................... 17
Figuur 2.1: Geografische ligging van de 37 moestuinen. Zwarte bolletjes geven permacultuurmoestuinen
weer, groene bolletjes ecologisch beheer en rode conventioneel beheer. De landbouwstreken werden
ter illustratie als achtergrond ingevoegd. ............................................................................................... 24
Figuur 2.2: Proefopzet met per tuin 3 plots, gefotografeerd op 29/10 in tuin 11. ......................................... 25
Figuur 2.3: Gutsboor voor bodemstaalname van 0,3 m diep, gefotografeerd op 1/11 in tuin 18. ................. 26
Figuur 2.4: Voorbeeld van een plot met houten frame te midden van een groenbemester (gele mosterd),
gefotografeerd op 1/11 in tuin 18. ......................................................................................................... 28
Figuur 2.5: Handsortering als derde en finale bemonsteringsstap, gefotografeerd op 1/11 in tuin 18. ........ 29
Figuur 2.6: De vangst van tuin 34, plot III (22/11). De pigmentatie van de regenwormen is een duidelijk
eerste kenmerk bij de determinatie. ...................................................................................................... 29
Figuur 3.1: Boxplot van de pHKCl voor de drie moestuintypes. Groene stippellijnen geven de streefzone aan.
De mediaan wordt weergeven door de horizontale lijn in de box. Vijf van de tien
permacultuurmoestuinen hebben een te lage pH. ................................................................................. 35
Figuur 3.2: Boxplot van het calciumgehalte (AL-extractie) voor de drie moestuintypes. Groene stippellijnen
geven de streefzone aan. De mediaan wordt weergeven door de horizontale lijn in de box. In tuin 8
wordt basaltmeel aangewend, wat het hoge Ca-gehalte daar kan verklaren. ....................................... 36
Figuur 3.3: Boxplot van het vochtgehalte voor de moestuintypes. ................................................................ 37
Figuur 3.4: Nitraatresidu (kg/ha) voor de drie moestuintypes. De rode lijn duidt de maximumgrens aan: drie
kwart van de permacultuur- en conventionele moestuinen overschrijdt de limiet. Bij ecologische
moestuinen zijn 50% van de waarnemingen te hoog. ............................................................................ 38
Figuur 3.5: Cirkeldiagram met het relatief voorkomen van de ingevulde beoordelingsklassen voor het
nitraatresidu. ........................................................................................................................................... 38
Figuur 3.6: Boxplot van het P-gehalte (AL-extractie) voor de drie tuintypes. Groene stippellijnen geven de
streefzone aan Moestuin 8 is al 40 jaar oud (sinds 2 jaar permacultuur en ervoor ecologisch beheer),
wat een mogelijke verklaring is voor de hoge concentratie P in deze moestuin. ................................... 39
Figuur 3.7: Boxplot van het K-gehalte (AL-extractie) voor de drie moestuintypes. Groene stippellijnen geven
de streefzone aan.................................................................................................................................... 40

VIII
Figuur 3.8: Boxplot van het Mg-gehalte (AL-extractie) voor de drie moestuintypes. Groene stippellijnen
geven de streefzone aan. In tuin 8 wordt basaltmeel aangewend, een bodemverbeteraar met een
hoog Mg-gehalte. In tuin 14 en 36 wordt jaarlijks Mg-kalk gebruikt....................................................... 41
Figuur 3.9: Boxplot van het totale koolstofgehalte voor de drie tuintypes.Tuin 3 en 8 bevatten grote
hoeveheelheden koolstof. In die tuinen lag een dikke laag mulch op de bodem. In tuin 10 ligt de
applicatie van zowel compost, turf, organische mestkorrels en kippen- en paardenmest aan de basis
van de hoge concentratie C. .................................................................................................................... 41
Figuur 3.10: Cirkeldiagram met het relatief voorkomen van de ingevulde beoordelingsklassen voor de pH,
calcium, fosfor, kalium en magnesium. ................................................................................................... 43
Figuur. 3.11: Procentueel voorkomen van de drie ecologische groepen (massa en aantal). Endogeïsche
regenwormen komen in aantal gemiddeld het vaakst voor: 174 ± 25 regenwormen m-2, gevolgd door
epigeïsche (47 ± 10 regenwormen m-2) en anekische (46 ± 7 regenwormen m-2). Qua massa staan de
forse anekische soorten op de eerste plaats (gemiddeld 40,1 ± 6,6 g m-2), gevolgd door de endogeïsche
(33,6 ± 3,6 g m-2) en de epigeïsche (4,93 ± 1,1 g m-2). ............................................................................. 44
Figuur 3.12: Top vijf van meest voorkomende soorten volgens aantal (a) en massa (b). De forse soort A.
longa die in aantal niet dominant is, is toch goed voor bijna 15% van de totale biomassa. Ook L.
terrestris stijgt van 5,38% naar 25,32% als de massa in plaats van het aantal in acht genomen wordt. 44
Figuur 3.13: Procentueel voorkomen op vlak van aantal en massa van de 19 aangetroffen
regenwormsoorten. ................................................................................................................................. 45
Figuur 3.14: Totale mediane massa (a) en totaal aantal (b) per m² met relatief voorkomen van de drie
ecologische groepen per moestuintype. Balken die dezelfde letter delen zijn niet significant
verschillend op vlak van totale massa en aantal (a<b<c; Kruskal-Wallis & Mann-Whitney U). Aangezien
met mediane waarden gewerkt wordt, is de som van de mediaan van de drie categorieën niet gelijk
aan de mediaan van de som. ................................................................................................................... 47
Figuur 3.15: Soortensamenstelling op vlak van aantal (a) en biomassa (b). Significante verschillen worden
aangeduid met kleine letters (a<b en sterretjes (*: p < 0.05; **: p < 0.01). Indien geen letter
weergegeven wordt werden geen verschillen genoteerd. ...................................................................... 51
Figuur 3.16: Boxplots van aantal regenwormen (a) en massa regenwormen (b) voor de drie textuurklassen.
Zowel aantal als massa zijn gelijkaardig bij moestuinen op zandleem en lichte zandleembodems. Bij
lemig zand werden slechts twee moestuinen onderzocht. ..................................................................... 53
Figuur A.1:Boxplot met de afmetingen van de drie moestuintypes. Er werden geen significante verschillen
gedetecteerd……………………………………………………………………………….……………………………………………………. 75
Figuur A.2: Boxplots voor totale ouderdom van de moestuinen (a) en van het aantal jaar dat huidig beheer
toegepast wordt. De laatste variabele was significant kleiner voor permacultuur dan voor de andere
moestuintypes (p < 0.05)……………………… ……………………………………………………………………….……………………76

IX
LIJST MET TABELLEN Tabel 1.1: Verband tussen moestuintype en beheer. ....................................................................................... 7
Tabel 1.2: Terminologie in verband met meststoffen en bodembewerking. .................................................... 7
Tabel 1.3: Aangetroffen soorten regenwormen met bijhorende ecologische groep, volgens Sims & Gerard
(1999) en * volgens Eisenhauer et al. (2008). Elke soort behoort tot de Lumbricidae, behalve Microscolex
phosphoreus die een exoot is en tot de familie van de Acanthodrilidae behoort.................................. 15
Tabel 2.1: Genoteerde moestuinvariabelen bij bezoek aan moestuin. ........................................................... 23
Tabel 2.2: Eigenschappen van de drie moestuincategorieën. ......................................................................... 24
Tabel 3.1: Mediane grootte, ouderdom (aantal jaar als moestuin) en aantal jaar dat de huidige methode
toegepast wordt voor de verschillende moestuintypes. Enkel deze laatste parameter bleek significant
kleiner (p < 0.05) bij permacultuurmoestuinen. ..................................................................................... 32
Tabel 3.2: Textuurklassen weergeven per moestuintype. .............................................................................. 34
Tabel 3.3: Overzichtstabel met gemiddelde waarden(µ) en standaardafwijkingen(SE) van de geanalyseerde
variabelen................................................................................................................................................ 34
Tabel 3.4: Overzichtstabel met het eerste kwartiel (Q1), de mediaan en het derde kwartiel (Q3) van de
geanalyseerde waarden die met een niet-parametrische test onderzocht werden. ............................. 35
Tabel 3.5: Spearmans rank correlatietest tussen biomassa en aantal regenwormen per m², p < 0,01. ......... 46
Tabel 3.6: Mediane aantal en biomassa voor de drie moestuintypes. Q1 = eerste kwartiel, Q3 = derde kwartiel.
Significante verschillen tussen moestuintypes worden aangeduid met kleine letters (a<b<c; Kruskal-
Wallis & Mann-Whitney U). Indien geen letter weergegeven wordt zijn er geen verschillen genoteerd.46
Tabel 3.7: Mediaan aantal en biomassa epigeïsche regenwormen voor de drie moestuintypes. Q1 = eerste
kwartiel, Q3 = derde kwartiel. Significante verschillen tussen moestuintypes worden aangeduid met
kleine letters (a<b<c; Kruskal-Wallis & Mann-Whitney U). Indien geen letter weergegeven wordt zijn er
geen verschillen genoteerd. .................................................................................................................... 47
Tabel 3.8: Mediaan aantal en biomassa epigeïsche regenwormen voor de drie moestuintypes. Q1 = eerste
kwartiel, Q3 = derde kwartiel. Significante verschillen tussen moestuintypes worden aangeduid met
kleine letters (a<b<c; Kruskal-Wallis & Mann-Whitney U). Indien geen letter weergegeven wordt zijn er
geen verschillen genoteerd. .................................................................................................................... 48
Tabel 3.9: Mediaan aantal en biomassa anekische regenwormen voor de drie moestuintypes. Significante
verschillen tussen moestuintypes worden aangeduid met kleine letters (a<b<c; Kruskal-Wallis & Mann-
Whitney U). Indien geen letter weergegeven wordt zijn er geen verschillen genoteerd. ...................... 49
Tabel 3.10: Overzichtstabel met diversiteitsvariabelen voor de drie moestuintypes. Weergegeven waarden
gelden voor een oppervlakte van 0,27 m². Q1 = eerste kwartiel, Q3 = derde kwartiel. Significante
verschillen worden aangeduid met kleine letters (a<b). Indien geen letter weergegeven wordt zijn er
geen verschillen genoteerd. .................................................................................................................... 52
Tabel A.1: Potentieel voorkomende gevaarlijke stoffen in de moestuin met bijhorende gezondheidsrisico's
www.gezonduiteigengrond.be). ............................................................................................................. 67
Tabel A.2: Coördinaten van de 37 moestuinen met bijhorend textuurtype. .................................................. 68
Tabel A.3: Verschillende gewasgroepen en er in voorkomende soorten . ...................................................... 69
Tabel A.4: Tuinen met correcte en overschatte OC-gehaltes, met bijhorende beoordelingsklasse volgens de
Bodemkundige dienst van België (Maes et al. 2012). ............................................................................. 70
Tabel A.5: Verdeelsleutel van juveniele soorten volgens drie determinatiekenmerken. ............................... 71

X
Tabel A.6: Transformatie van variabelen om normaliteit en homoscedasticiteit te bekomen. Variabelen met
een lege cel in de tweede kolom konden niet getransformeerd worden. Geen enkele van de variabelen
was initieel normaal verdeeld. ................................................................................................................ 72
Tabel A.7: Overzichtstabel met voor alle variabelen. Voor elk moestuintype is de het eerste kwartiel, de
mediaan en het tweede kwartiel weergegeven. De letters indiceren significante verschillen tussen de
drie tuintypes (p < 0,05) waarbij geldt: a < b, b < c; a = ab en a < bc. Indien geen letter weergegeven
wordt zijn er geen verschillen genoteerd. ............................................................................................... 73
Tabel A.8: Overzichtstabel met niet normaal verdeelde of heteroscedastische variabelen. Voor elk
moestuintype is de het eerste kwartiel, de mediaan en het tweede kwartiel weergegeven. De letters
indiceren significante verschillen tussen de drie tuintypes (p < 0,05) waarbij geldt: a < b, b < c; a = ab en
a < bc. Indien geen letter weergegeven wordt zijn er geen verschillen genoteerd. ............................... 74
Tabel A.9: Absoluut en procentueel voorkomen van de verschillende teeltgroepen per moestuintype. ....... 77
Tabel A.10: Beoordelingsklassen, eigen bewerking van de Bodemkundige Dienst van België en de Vlaamse
Landmaatschappij (Maes et al. 2012; VLM 2014). .................................................................................. 78
Tabel A.11: Bodemtypes waarvoor de beoordelingsklassen uit Tabel A.1 van toepassing zijn, eigen bewerking
van de Bodemkundige Dienst van België en de Vlaamse Landmaatschappij (Maes et al. 2012; VLM 2014).
................................................................................................................................................................. 79

1
INLEIDING
In het sterk verstedelijkte Vlaanderen bezitten tuinen – en hun bewoners – een vaak ongekende
natuurwaarde (Dewaelheyns et al. 2011). Regenwormen, door Darwin verheven tot de belangrijkste
diersoort op aarde, zijn een belangrijke groep organismen die in onze contreien een onmiskenbare
invloed uitoefenen op de bodem (Sims & Gerard 1999). Doordat ze hun eigen habitat en daarbij dat van
vele andere soorten zodanig beïnvloeden, worden ze bestempeld als bodemingenieurs.
De rol van regenwormen in de bodem is veelzijdig en positief van aard: naast het maken van de
kruimelstructuur en het versnellen van de nutriëntencirculatie, verbeteren ze met hun graafactiviteiten
de verluchting van de bodem waardoor ook een betere waterhuishouding ontstaat (Edwards & Lofty
1977; Sims & Gerard 1999; Bertrand et al. 2015). Als gevolg van deze grote impact op de
bodemvruchtbaarheid, zijn ze een belangrijke speler inzake plantengroei en de daarmee verbonden
gewasopbrengst in landbouwsystemen.
In Vlaanderen komen 25 inheemse soorten regenwormen voor, die op basis van hun gedrag en activiteit
onderverdeeld worden in drie verschillende groepen (Valckx et al. 2009). Voor een goede werking en
maximaal ‘rendement’ is het belangrijk dat deze alle drie aanwezig zijn in de bodem. In akkerland en
bossen werd hun voorkomen inmiddels veelvuldig onderzocht, en aangezien moestuinen een niet
onbelangrijke fractie van het Vlaamse landschap vertegenwoordigen (± 3,3 %), wordt in deze
masterproef de regenwormabundantie in deze systemen voor het eerst onder de loep genomen.
Er wordt vertrokken van drie moestuincategorieën: (i) de permacultuurmoestuin waarbij natuurlijke
situaties nagebootst worden (bedekte bodem en minimale bodembewerking), (ii) de ecologische
moestuin en (iii) de conventionele moestuin waar kunstmest aangewend wordt. De invloed van
pesticiden op regenwormen wordt in dit onderzoek niet onderzocht, omdat deze reeds gekend is en
nefast werd bevonden (Pelosi et al. 2014): vandaar de beslissing om te focussen op andere
beheerfactoren.
Dankzij de bereidwilligheid van 37 moestuineigenaars konden in totaal 2664 regenwormen gevangen
en gedetermineerd worden. In elke moestuin werd ook een bodemstaal genomen, waardoor
regenwormendata gekoppeld kon worden aan diverse bodemvariabelen zoals de zuurtegraad, het
organische stofgehalte en de concentratie van enkele belangrijke nutriënten.
Deze masterproef poogt een globaal beeld te schetsen van de diversiteit en kwantiteit van de
regenwormen in Vlaamse moestuinen. Waar mogelijk worden verbanden gelegd met zowel de
beheerfactoren, dikwijls gelinkt aan het moestuintype, als de bodemvariabelen. De hypothese dat
abundantie en diversiteit hiermee gecorreleerd zijn, wordt onderzocht. Er wordt verwacht dat
permacultuurmoestuinen meer en andere soorten zullen herbergen dan de twee andere
moestuintypes; enerzijds omdat er een permanente strooisellaag aanwezig is waarin sommige
regenwormen vertoeven en anderzijds omdat er bewust een minimale bodembewerking wordt

2
toegepast. Intensieve bodembewerking vormt in conventionele landbouw immers grote hindernissen
voor regenwormpopulaties (Curry 1998).
Naast dit hoofddoel wordt tevens nagegaan of moestuinen hun veronderstelde natuurwaarde bereiken.
Dragen ze bij aan de biodiversiteit, of net niet?
Deze masterproef begint met een literatuurstudie over de rol van tuinen met de focus op moestuinen,
om vervolgens in deel twee over te gaan op de regenwormen. Hierna volgt een beschrijving van de
onderzoeksmethoden en het praktische luik. Nadien worden de resultaten weergegeven en besproken
in eenzelfde hoofdstuk. Tot slot wordt nog een algemeen besluit vooropgesteld, waarin de voornaamste
bevindingen en eventuele pijnpunten geschetst worden.

3
1 LITERATUUR
1.1 TUINEN
1.1.1 Inleiding & belang van tuinen in Vlaanderen
Steeds meer aandacht wordt besteed aan de rol van private tuinen in het kader van biodiversiteit
(Gaston et al. 2005). Sinds de conferentie hieromtrent in Rio de Janeiro in 1992, is wereldwijd besloten
om halt te roepen aan het biodiversiteitverlies. Op deze toonaangevende conferentie werd
biodiversiteit gedefinieerd als: “de variabiliteit onder levende organismen van allerlei herkomst, met
inbegrip van, onder andere, terrestrische, mariene en andere aquatische ecosystemen en de
ecologische complexen waarvan zij deel uitmaken. Dit omvat mede de diversiteit binnen soorten, tussen
soorten en van ecosystemen”. Het verlies is bij uitstek te wijten aan de introductie van invasieve soorten,
overexploitatie van natuurlijke hulpbronnen en habitatverlies en –degradatie (IUCN; Bakkes et al. 2008).
De wereldwijd toenemende urbanisatie speelt hier een rol in: reductie van habitatoppervlakte enerzijds
en fragmentatie van deze kleinere habitats anderzijds (Saunders et al. 1992). Met urbanisatie neemt
tevens de hoeveelheid bodemafdichting toe, wat ervoor zorgt dat regenwater de bodem niet meer kan
binnensijpelen. Ook in Europa is dit het geval: Malta, met 13,3% ondoorlaatbare oppervlakte, voert de
top drie aan, gevolgd door België (7,4%) en Nederland (7,3%). Als we Vlaanderen apart beschouwen is
zelfs 12,9% van de bodem afgedicht. Een hallucinant cijfer, zeker in vergelijking met een gemiddelde
waarde van 1,8% voor Europa (De Meyer et al. 2011). Meer dan ooit moet resterende ruimte optimaal
benut worden om verder verlies van biodiversiteit te beperken. Bakkes et al. (2008) berekenden dat de
kosten van biodiversiteitverlies tegen 2050 wereldwijd kunnen oplopen tot 14 000 miljard euro – 7%
van het bruto wereldproduct – als geen stappen ondernomen worden om dit verlies te voorkomen. De
doelstellingen voor 2010 werden niet behaald (Europese Commissie 2011). Om de nieuwe
doelstellingen tegen 2020 wel te halen, wordt aanbevolen om private tuinen hierbij te betrekken (Loram
et al. 2007; Beumer & Martens 2014).
Bomans et al. concludeerden dat het totale tuinareaal in Vlaanderen 8,2% van de totale oppervlakte
vertegenwoordigt (2011). Ondanks deze significante oppervlakte – 12,9% is afgedicht, 10% is bos – is er
weinig informatie over deze categorie beschikbaar (Zmyslony & Gagnon 1998; Bomans et al. 2011;
Dewaelheyns et al. 2014). Thompson et al. wijzen dit hiaat aan kennis toe aan het private aspect: tuinen
liggen buiten de bevoegdheid van overheden en lokale besturen (2003). Informatie verkrijgen over de
toestand en het beheer ervan is bijgevolg ook moeilijk, mede doordat deze oppervlakte verdeeld is
onder veel individuele eigenaars en dus zeer heterogeen is (Zmyslony & Gagnon 1998; Thompson et al.
2003; Van Delm & Gulinck 2011).
De planning van een eigen tuin is deze dagen paradoxaal (Cameron et al. 2012; Dewaelheyns et al. 2014).
Het Structuurplan Ruimtelijk Beleid in Vlaanderen uit 1996 zegt dat de Vlaming kleiner zou moeten gaan

4
wonen, er meer in hoogte moet gebouwd worden en dat het landschap moet ont- in plaats van
versnipperen. Een recente evaluatie toont aan dat niet aan het gebeuren is (OVAM 2010). Het bezit van
een eigen huis met tuin wordt in Vlaanderen beschouwd als een verworven recht: een gedachte die
voortkomt uit de 19de en 20ste eeuw toen dit concept door de overheid sterk gepromoot werd (De Decker
2011). Niet iedereen hecht even veel belang aan een tuin: 15% van de respondenten van een rondvraag
aan 1614 Vlamingen geeft aan niet zonder een tuin te kunnen leven, terwijl 18% geen of weinig belang
hecht aan een tuin. Als gevraagd werd te kiezen tussen (1) een bos; (2) een nabijgelegen natuurgebied;
of (3) een privétuin, koos 70% voor de laatste optie (Beleidsdomein Leefmilieu Natuur en Energie 2014).
Aangezien tuinen niet weg te denken zijn in het huidige Vlaanderen, zou het potentieel van deze ruimte optimaal benut moeten worden. Bijvoorbeeld door tuinen niet te zien en beheren als individuele lapjes grond, maar als één groot tuincomplex (Dewaelheyns, Bomans, & Gulinck, 2011).
1.1.2 Moestuinen in Vlaanderen
Kenmerken
Steeds meer onderzoek gebeurt naar volkstuinen, terwijl private moestuinen wereldwijd nog vaak
genegeerd worden (Kortright & Wakefield 2011; Guitart et al. 2012; Taylor & Lovell 2014; Church et al.
2015). Diverse motivaties bestaan voor het houden van een moestuin: als educatie voor de kinderen,
als hobby, om de ecologische voetafdruk te verkleinen, het ‘weet-wat-je-eet’-idee, lekkerder voedsel,
etc. Schaduw, het ontbreken van kennis en vaardigheden en twijfels over voedselveiligheid zijn dan weer
redenen om geen groenten te verbouwen (Kortright & Wakefield 2011).
Ook over moestuinen in Vlaanderen is weinig geweten. Volgens de openbare afvalmaatschappij (OVAM)
bezit 30% van de 452 respondenten met een tuin naast gras (93%), verharding (84%), houtachtigen
(64%) en bloemperken (58%), een moestuin (2012). Een grootschaligere ondervraging (1138
respondenten) van Dewaelheyns en Gulinck komen uit bij een percentage van 41,6% (2008). De
gemiddelde moestuin zou volgens beide onderzoeken respectievelijk 233 m² en 178 m² groot zijn. Als
met deze cijfers de totale oppervlakte aan moestuin berekend wordt komt dit neer op 43 701 ha (3,23%
van Vlaanderen) en 46 295 ha (3,42% van Vlaanderen).
Cijfers over het pesticiden- en meststoffengebruik in Vlaamse moestuinen zijn niet voorhanden. Wel zijn
cijfers gekend die particulieren lieten uitvoeren bij de Bodemkundige Dienst van België (BDB). Deze
onderzoeksinstelling analyseert sinds 1946 bodemstalen en geeft bemestings-, belkalkings- en milieu-
advies aan particulieren (www.bdb.be). Tussen augustus 2007 en juli 2009 werden 393 grondstalen uit
Vlaamse moestuinen onderzocht (Elsen et al. 2011). Deze stalen zijn echter niet representatief voor de
gemiddelde Vlaamse moestuin, aangezien dikwijls pas stalen ingeleverd worden indien men problemen
ondervindt. Van elke staal werd de bodemtextuur, de zuurtegraad (pHKCl) en het koolstof-, fosfor -,
kalium-, magnesium -, calcium- en natriumgehalte bepaald:
Zuurtegraad: 67% heeft een te hoge pH, wat kan zorgen voor onevenwichtige opname van
nutriënten die nochtans in voldoende mate aanwezig zijn in de bodem. In bodems met een

5
te lage pH zullen daarbovenop ook basische kationen zoals calcium, kalium en magnesium
uitlogen bij een neerslagoverschot.
Organisch koolstofgehalte (C): 59% heeft een organisch C-gehalte hoger dan het optimum.
Dit indiceert een te hoge input van compost, stalmest of andere organische producten. Een
te hoog percentage kan tot verliezen leiden (Reubens et al. 2010).
Fosfor (P): 89% heeft een P-gehalte boven het optimum, wat duidt op excessieve
bemesting. Dit zorgt ervoor dat P accumuleert in de bovenste bodemlaag. Naast de
plantengroei kan ook het oppervlaktewater hierdoor negatief beïnvloed worden.
Kalium (K): 74% heeft een te hoog K-gehalte.
Magnesium (Mg): 76,3% heeft een te hoog Mg-gehalte.
Calcium (Ca): naar analogie met de pH is ook het Ca-gehalte meestal (±56%) te hoog.
Natrium (Na): zo een 53% heeft een te laag Na-gehalte.
Elsen et al. (2011) maakten ook de vergelijking tussen 1998 en 2009, maar voor moestuinen werd geen
evolutie waargenomen in de resultaten.

6
Moestuintypes in Vlaanderen
Grofweg wordt in Vlaanderen afhankelijk van het beheer onderscheid gemaakt tussen drie
moestuintypes (zie ook Tabel 1.1) :
(1) De ecologische moestuin
In deze tuin worden geen kunstmest, chemische pesticiden of genetisch gemodificeerde organismen
gebruikt. Verder probeert de ecologische tuinier met zo weinig mogelijk externe inputs de bodem zo
vruchtbaar mogelijk te houden, door bijvoorbeeld het organisch stofgehalte van de bodem op peil te
houden door eigen compost te maken van oogstresten en ander organisch materiaal. Om plagen te
vermijden wordt een teeltwisselingssysteem toegepast (VELT vzw 2014). De voordelen van ecologisch
tuinieren zijn duidelijk aangetoond voor biodiversiteit alsook voor water- en bodemkwaliteit (Dorais
2007; Henneron et al. 2014). Bovendien worden significant minder residu’s van pesticiden terug
gevonden op biologisch geteelde producten (Baker et al. 2002).
(2) De permacultuur moestuin
Permacultuur is een internationale beweging en een ecologisch ontwerpsysteem voor tuinen (Ferguson
& Lovell 2014). De uit Australië afkomstige term is afgeleid van ‘permanent agriculture’ en is ontsproten
uit de ideeën van David Holmgren en Bill Mollison die hun eerste boek erover publiceerden in 1978.
Concreet wordt gedoeld op een beheer dat aanleunt bij de permacultuurprincipes: minimale
bodembewerking, heterogene vegetatie (combinatieteelt), permanente bodembedekking (Figuur 1.1),
lage impact op milieu (geen chemicaliën en kunstmest), en dit alles in een landschap dat een natuurlijke
situatie nabootst om zo in voedsel, vezels en energie te voorzien (Mollison & Holmgren 1978; Ferguson
& Lovell 2014; LeVasseur 2014; VELT vzw 2014).
(a) (b) (c)
Figuur 1.1: Permacultuurtuinen kunnen natuurlijk (a) of gestileerd (b) ogen, maar de bodem zal nooit onbedekt zijn zoals in een klassieke moestuin (c). Foto’s werden genomen in respectievelijk tuin 1 (24/10), tuin 3 (5/11) en tuin 7 (26/10).
7
(3) De conventionele moestuin
Een conventionele tuinier probeert hoge opbrengsten te halen, door principes toe te passen uit de
gangbare landbouw: er wordt kunstmest aangewend om de planten van voedingsstoffen te voorzien en
gewasbelagers en onkruiden worden bestreden met chemische gewasbeschermingsmiddelen
(Henneron et al. 2014; VELT vzw 2014). De ‘gangbare landbouw’ is echter minder afgebakend dan de
biologische, en bijgevolg minder eenduidig: van zodra er aan één voorwaarde voor de biologische teelt
niet voldaan is, spreekt men over conventionele teelt (Załęcka et al. 2014).
Terminologie
In deze masterproef wordt over diverse soorten mest en bodembewerking geschreven. Tabel 1.2
verduidelijkt de gebruikte terminologie.
Tabel 1.1: Verband tussen moestuintype en beheer.
Permacultuur Ecologisch beheer Conventioneel beheer
Bodemverstoring minimaal Kerende of niet-kerende bodembewerking
Bodembedekking altijd niet noodzakelijk
Bemesting nooit kunstmest, eventueel andere vaak kunstmest
Gewassen door elkaar meestal in bedden
Gewasbescherming combinatieteelt teeltwisseling soms teeltwisseling,
vaak chemische gewasbeschermingsmiddelen
Tabel 1.2: Terminologie in verband met meststoffen en bodembewerking.
Kunstmest Hiermee worden artificieel samengestelde minerale meststoffen bedoeld die
kunstmatig gewonnen materiaal bevatten. Het fabricageproces verbruikt veel
energie.
Organische mest Deze meststoffen zijn commerciële producten die samengesteld zijn uit
plantaardige en/of dierlijke grondstoffen. Doordat de stikstof gebonden is in een
organisch molecule komt ze (automatisch) langzaam vrij. Als de grondstoffen
biologisch geteeld zijn, is organische mest toegelaten in de biologische
landbouw. Hier wordt dit onderscheid niet gemaakt en hanteren we het begrip
‘ecologisch’.
Dierlijke mest In deze masterproef wordt dit verzamelbegrip gebruikt voor verse mest van
koeien, paarden, ezels, kippen, etc.
Niet – kerende
bodembewerking
Bodembewerking waarbij de grond niet gekeerd wordt. Hieronder vallen woelen
en frezen.
Kerende
bodembewerking
Bodembewerking waarbij de grond gekeerd wordt. In deze masterproef wordt
enkel spitten in deze categorie ondergebracht.

8
1.1.3 Ecosysteemdiensten van moestuinen
Ecosysteemdiensten zorgen voor het welzijn van de mens en andere soorten. Het zijn producten
(voedsel, water, …), regulerende diensten (bestuiving, waterzuivering, …), culturele diensten
(esthetisch, educatief, …) door een ecosysteem geleverd of diensten die voorgaande ondersteunen
(Millennium Ecosystem Assessment 2004). Volgende paragrafen brengen enkele ecoysteemdiensten
naar voor waaraan tuinen en meer specifiek moestuinen een bijdrage kunnen leveren.
Voedselproductie
In de huidige maatschappij heeft de mens nog nooit zo ver gestaan van de productie van het voedsel
dat op het menu staat. Canadees onderzoek (Xuereb 2005) wees uit dat 58 vaak gegeten producten,
die bovendien lokaal geproduceerd zouden kunnen worden, gemiddeld 4497 km aflegden tot de
consument. Jones (2006) bracht aan het licht dat bonen geproduceerd in Kenia tot dertien keer
meer energie vergden dan bonen uit het Verenigd Koninkrijk. Het concept voedselkilometers is
echter een te eng begrip om de milieu-impact van de voedselindustrie te schetsen. Producten die
veel zonlicht vergen voor productie, kunnen een lagere impact hebben indien geïmporteerd uit
klimatologisch gunstigere regio’s (Avetisyan et al. 2014). Lokaal, seizoensgebonden voedsel lijkt een
uitweg voor dit probleem.
Hoewel moestuinen wijdverspreid zijn, ontbreekt vaak informatie over hun betekenis voor
voedselvoorziening (Taylor & Lovell 2014). Wel kunnen ze weerwerk bieden op de intensieve
landbouw die grotendeels afhangt van mechanisatie, brandstof, meststoffen en pesticiden
(Satterthwaite et al. 2010; Chen 2012).
Kleinschalig onderzoek in de gemeente Herent in Vlaanderen (Dewaelheyns & Gulinck, 2008)
berekende de gemiddelde groentenopbrengst en kwam uit op 0,287 kg per m². Deze schatting zegt
echter weinig: dit cijfer hangt namelijk sterk af van de geteelde groente die hierbij niet vermeld
staan, waardoor deze waarde niet te vergelijken valt met referentiewaarden. CoDyre et al. (2015)
brachten interessantere conclusies: (1) de meest productieve tuinen wijzen op het enorme
opbrengstpotentieel; (2) momenteel wordt maar een fractie van de potentiële opbrengst behaald;
(3) om zelfvoorzienend te zijn, is meer kennis/kunde nodig: slechts 10% van de tuinders bleek - in
het seizoen - zelfvoorzienend; en (4) tuiniers met meer dan zeven jaar ervaring hadden significant
hogere opbrengsten. Ook bleken de geoogste producten gemiddeld goedkoper te zijn in de winkel,
wanneer rekening gehouden werd met de prijs van materiaal, en zeker als arbeid werd
meegerekend. Dit was echter niet zo bij de ‘beste’ tuiniers. Bij enkelen waren de opbrengsten zelfs
vergelijkbaar met deze van professionele kwekers in de buurt (CoDyre et al. 2015).
Bijdrage aan biodiversiteit
In een stedelijke omgeving vertegenwoordigen private tuinen een aanzienlijk deel van de groene
ruimte, waardoor ze de mogelijkheid hebben om biodiversiteit te behouden en ecosysteemdiensten
te leveren (Gaston et al. 2005; Loram et al. 2007; Dewaelheyns et al. 2011). Het BUGS-project

9
(Biodiversity of Urban Gardens in Sheffield) van de Universiteit van Sheffield (Thompson et al. 2003)
voert al tien jaar onderzoek binnen dit thema. Conclusies van dit en andere onderzoeken zijn dat
privétuinen een aanzienlijke hoeveelheid wilde planten- en diersoorten kunnen bevatten en dat
allerlei landschapselementen zoals poelen, hagen, dood hout, etc. een habitat en voedselbron zijn
voor tal van soorten (Thompson et al. 2003; Gaston et al. 2005). Zelfs als individuele tuinen te klein
zijn om levensvatbare populaties te herbergen, kunnen ze dienen als matrix of corridors voor
populaties (Rudd et al. 2002; Goddard et al. 2010). Tuinen zouden volgens Hermy en Claessens
(2011) kunnen dienen als een soort ‘Ark van Noah’. Ze worden tevens gekenmerkt door planten met
een hoge intra- en interspecifieke genetische variabiliteit, vooral inzage gewasvariëteiten en
landrassen, en kunnen op deze manier bijdragen aan het behoud van de agro-biodiversiteit (Aguilar-
Støen et al. 2009; Galluzzi et al. 2010).
Tuinieren kan echter ook negatieve impact hebben op (lokale) ecosystemen en biodiversiteit: door
excessief watergebruik, onoordeelkundig pesticidengebruik, vermesting en introductie van
invasieve exoten (Steinberg 2006; Clayton 2007).
Specifiek voor Vlaanderen zijn in enkele studies gegevens terug te vinden over het gebruik van
chemische producten in de tuin. Een verkennende studie in opdracht van de Vlaamse
Milieumaatschappij naar de inputs en outputs in privétuinen in Vlaanderen (Dewaelheyns & Gulinck
2008) stelt dat de milieu-impact in vergelijking met landbouw weliswaar kleiner is, maar toch niet
verwaarloosbaar. Uit een rondvraag bij 1138 tuineigenaars in 2007 bleek dat 40% van de
respondenten chemische bestrijdingsmiddelen gebruikt. Een enquête van de Openbare
Afvalmaatschappij (OVAM) uit 2006 bracht aan het licht dat 49% van de ondervraagde tuineigenaars
chemische bestrijdingsmiddelen en/of kunstmeststoffen gebruikt (OVAM 2006). In 2012 werd dit
onderzoek herhaald en werd een vergelijkend rapport uitgegeven (OVAM 2012). Daaruit blijkt dat
in 2012 dit percentage gedaald is tot 36% . Verwacht wordt dat deze dalende trend zich verder zal
zetten gezien de definitieve ingang van de fytolicentie in september 2015. Dit certificaat bestaat
enkel voor professionele gebruikers en verbiedt op die manier particulieren professionele
chemische gewasbeschermingsmiddelen aan te schaffen.
Bij beide enquêtes van OVAM werden 500 Vlaamse gezinnen uit 38 verschillende gemeenten
bereikt. De drie voorgaande resultaten zijn niet representatief voor Vlaanderen, gezien de manier
van ondervragen (een internet-enquête) en tevens omdat er geen referentie waarden zijn op
Vlaams, gewestelijk, provinciaal of gemeentelijk niveau (Dewaelheyns & Gulinck, 2008). Ze kunnen
echter wel een indicatie geven. De Vlaamse overheid probeert de Vlaming bewust te maken van de
mogelijk heden van ecologisch tuinieren via campagnes als “Zonder is gezonder” en “Gezond uit
eigen grond”. Of deze campagnes vruchten afwerpen is nog niet geweten.
Of de balans van een tuin positief dan negatief is voor de biodiversiteit, hangt af van het beheer
(Guitart et al. 2012). Worden nutriënten toegevoegd door middel van artificiële meststoffen of via
compost? Worden plagen verhinderd door gebruik van pesticiden of eerder door gewasrotatie of

10
door bepaalde gewassen met elkaar te combineren? Wordt leidingwater gebruikt of wordt
regenwater opgevangen voor irrigatie? Lindemann-Matthies en Marty (2013) onderzochten de
verschillende invloeden van conventioneel en ecologisch beheer (frequentie van grasmaaien en
wieden en intensiteit van gebruik van pesticiden en kunstmeststoffen) op de soortenrijkdom in
tuinen en concludeerden dat deze in ecologisch beheerde tuinen significant hoger was.
Daarbovenop bleken deze tuinen esthetisch ook beter te scoren.
Welzijn van de mens
Uit onderzoek van onder andere Kaplan en Kaplan (1990) blijkt dat mensen die in contact staan met
de natuur zich beter voelen. Ook wijst onderzoek uit dat tuinieren een positieve invloed kan hebben
op de mentale gezondheid (Gaston et al., 2005; Ropars, 2013; Stuart, 2005; Währborg, Petersson &
Grahn, 2014). De tuin kan tevens stress reduceren en burn-outs verminderen (Ulrich et al. 1991;
Adevi & Mårtensson 2013). Naast positieve verbanden tussen mentale gezondheid en de nabijheid
van groen, bestaat er ook een correlatie met fysiologische gezondheid: bepaalde ziekten kwamen
beduidend minder voor bij proefpersonen die een eigen tuin of een andere vorm van groen in de
directe leefomgeving hadden (de Vries et al. 2003; Maas et al. 2009). De gezondheidslink met
openbaar groen is vaak te wijten aan de mogelijkheid om - al dan niet samen met anderen - aan
recreatie en sport te doen (Maas et al. 2009; Jennings & Gaither 2015). Of in de tuin werken een
invloed heeft op de gezondheid, is onvoldoende onderzocht (Roe & Thompson 2011).
Andere positieve aspecten voor de gezondheid zijn de verhoogde consumptie van groenten en fruit
(Alaimo et al. 2008; Morton et al. 2008) . Nochtans zijn er soms ook risico’s verbonden aan de
consumptie van groenten uit eigen tuin. Sterk verstedelijkte gebieden zoals Vlaanderen worden
blootgesteld aan een grote hoeveelheid metalen en organische contaminanten (Shinn et al. 2000;
Clark et al. 2006). Tabel A.1 (in de appendix) geeft een overzicht van potentieel in moestuinen
voorkomende gevaarlijke stoffen volgens de Vlaamse Overheid. Een analyse van gewassen uit
Berlijnse moestuinen bracht aan het licht dat de kritische concentraties voor metalen frequent
overschreden werden. Barrières zoals gebouwen, hoge beplanting of zelfs een afstand van meer
dan 10 meter tot zwaar verkeer, reduceerde het percentage overschrijdingen sterk (Säumel et al.
2012). Een kleinschalig onderzoek omtrent atmosferische contaminatie in de Gentse stadskern
toont enkel frequente overschrijdingen voor het element lood (De Wispelaere 2014). Ook Xiong et
al. (2014) wijzen op gezondheidsrisico’s geassocieerd met groenten die geteeld zijn in regio’s met
hoge gehalten aan fijn stof.
Andere ecosysteemdiensten die aangehaald kunnen worden zijn temperatuurbuffering, het vermogen
om water vast te houden (overstromingsregulatie) en te zuiveren en het sluiten van kringlopen door de
verwerking van groenten-, fruit- en tuinafval onder vorm van compost (Guitart et al. 2011; Cameron et
al. 2012). Dit laatste wordt in Vlaanderen al frequent gedaan: 58% van de Vlamingen composteert
(OVAM 2012). Bloeiende gewassen zijn bovendien gunstig voor bestuivers.

11
1.2 REGENWORMEN
1.2.1 Inleiding
In gematigde regio’s zoals West-Europa zijn regenwormen qua massa de dominante groep macro-
invertebraten in veel bodems (Edwards & Lofty 1977). Nog voor onze jaartelling begon, trok hun
vermogen om de grond om te woelen Aristoteles’ aandacht. 2000 jaar later focuste Darwins laatste boek
‘The Formation of Vegetable Mould through the Action of Worms’ op hun potentieel om plantaardig en
dierlijk materiaal om te zetten en hun onmiskenbare invloed op de bodem. Zijn werk was gebaseerd op
40 jaar observeren en was wetenschappelijk niet geheel waterdicht, maar wierp wel licht op deze
ongelooflijk belangrijke dieren (Edwards & Lofty 1977).
De regenwormen behoren net als zoetwaterwormen en enkele andere terrestrische wormen tot de
klasse van de Oligochaeta. Samen met de bloedzuigers worden ze ondergebracht in de groep van de
Clitellata welke behoort tot de stam Annelida ofwel de stam van de rondwormen. Deze herbergt het
grootste aantal wormachtige species die zowel terrestrisch als aquatisch kunnen zijn. Het belangrijkste
gemeenschappelijk kenmerk van deze stam is het gesegmenteerde karakter: het lichaam is
onderverdeeld in afzonderlijke units (Barnes, 1980). De Oligochaeta kunnen verder opgesplitst worden
in drie ordes. De regenworm behoort tot de Haplotaxida, meer bepaald tot de onderorde Lumbricina.
Deze wordt gekarakteriseerd door een clitellum (zie paragraaf 1.2.2) dat bestaat uit meerdere cellagen
en een oöcyt met kleine dooier, waaruit later de eicel zal ontstaan. De verdere onderverdeling van de
Lumbricina in superfamilies en families gebeurt op basis van verschillen in het vrouwelijke
voortplantingsstelsel (Edwards & Lofty, 1977; Sims & Gerard, 1999). In Europa worden hoofdzakelijk
regenwormen teruggevonden die behoren tot de familie van de Lumbricidae. Andere soorten zijn
exotisch (Sims & Gerard 1999).
Regenwormen zijn hermafrodiet en hebben zowel vrouwelijke als mannelijke geslachtsporiën. Enkele
soorten kunnen zich voortplanten via parthenogenese, maar in de meeste gevallen is paring nodig. Een
of meerdere embryo’s groeien op in cocons die afgezet worden na de paring. In de vrije natuur worden
regenwormen meestal niet ouder dan een paar jaar (Edwards & Lofty 1977).
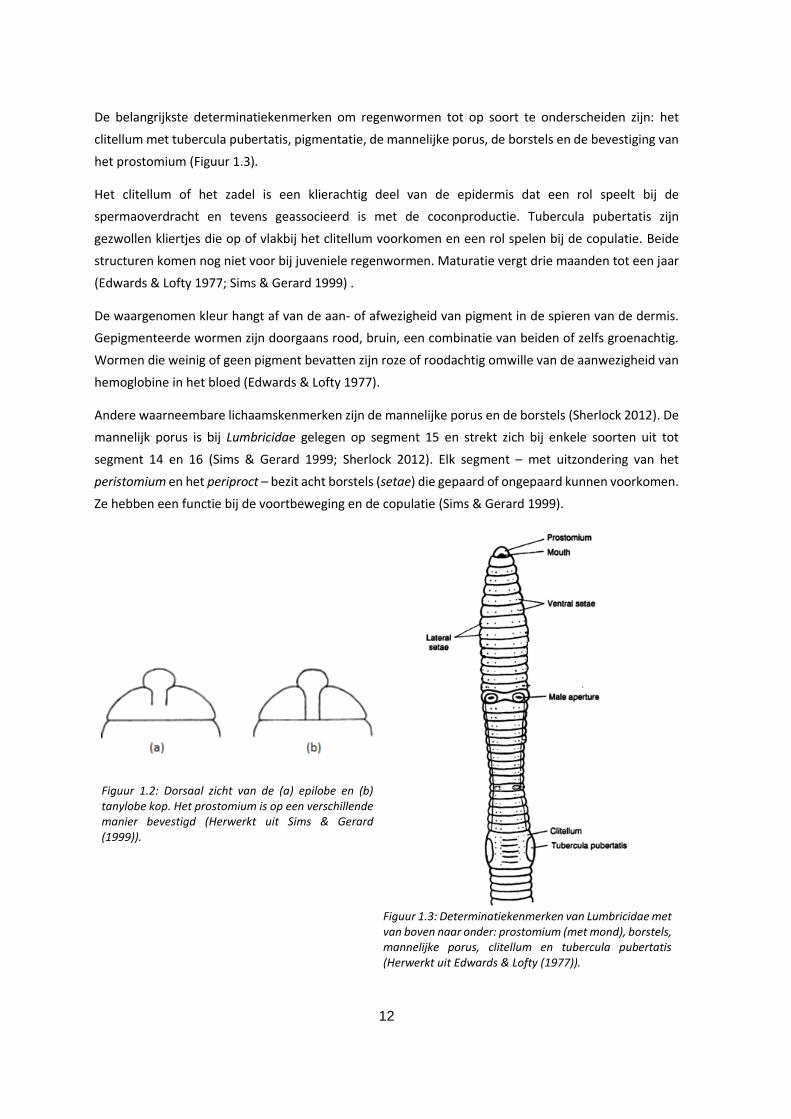
1.2.2 Morfologie en determinatiekenmerken
In West-Europa varieert de lichaamslengte van adulte species van 15 mm tot 300 mm waarbij de
diameter schommelt tussen 1 en 10 mm. De uitwendig zichtbare groeven corresponderen met de
inwendige septa die het langgerekte lichaam in segmenten onderverdelen. Deze zijn afgezien van het
eerste en het laatste segment - respectievelijk het peristomium en het periproct - gelijkwaardig: men
spreekt van homonome segmentatie. Het peristomium draagt een soort flap die de mond bedekt en
een functie heeft bij voedselopname, het prostomium (Figuur 1.2). Op het periproct is de anus zichtbaar
als een klein gleufje.
12
De belangrijkste determinatiekenmerken om regenwormen tot op soort te onderscheiden zijn: het
clitellum met tubercula pubertatis, pigmentatie, de mannelijke porus, de borstels en de bevestiging van
het prostomium (Figuur 1.3).
Het clitellum of het zadel is een klierachtig deel van de epidermis dat een rol speelt bij de
spermaoverdracht en tevens geassocieerd is met de coconproductie. Tubercula pubertatis zijn
gezwollen kliertjes die op of vlakbij het clitellum voorkomen en een rol spelen bij de copulatie. Beide
structuren komen nog niet voor bij juveniele regenwormen. Maturatie vergt drie maanden tot een jaar
(Edwards & Lofty 1977; Sims & Gerard 1999) .
De waargenomen kleur hangt af van de aan- of afwezigheid van pigment in de spieren van de dermis.
Gepigmenteerde wormen zijn doorgaans rood, bruin, een combinatie van beiden of zelfs groenachtig.
Wormen die weinig of geen pigment bevatten zijn roze of roodachtig omwille van de aanwezigheid van
hemoglobine in het bloed (Edwards & Lofty 1977).
Andere waarneembare lichaamskenmerken zijn de mannelijke porus en de borstels (Sherlock 2012). De
mannelijk porus is bij Lumbricidae gelegen op segment 15 en strekt zich bij enkele soorten uit tot
segment 14 en 16 (Sims & Gerard 1999; Sherlock 2012). Elk segment – met uitzondering van het
peristomium en het periproct – bezit acht borstels (setae) die gepaard of ongepaard kunnen voorkomen.
Ze hebben een functie bij de voortbeweging en de copulatie (Sims & Gerard 1999).
Figuur 1.2: Dorsaal zicht van de (a) epilobe en (b) tanylobe kop. Het prostomium is op een verschillende manier bevestigd (Herwerkt uit Sims & Gerard (1999)).
Figuur 1.3: Determinatiekenmerken van Lumbricidae met van boven naar onder: prostomium (met mond), borstels, mannelijke porus, clitellum en tubercula pubertatis (Herwerkt uit Edwards & Lofty (1977)).

13
1.2.3 Ecologie
Figuur 1.4: De drie ecologische groepen, v.l.n.r. epigeïsch, endogeïsch en anekisch (illustratie: Klaas Van de
Moortel).
Meer dan 40 jaar geleden verdeelde Bouché (1972) regenwormen aan de hand van morfologische en
gedragskarakteristieken in drie ecologische categorieën (Figuur 1.4): epigeïsche, endogeïsche en
anekische regenwormen (Sims & Gerard 1999). Epigeïsche regenwormen leven aan de oppervlakte in
een laag vers en verterend organisch materiaal, een strooisellaag, waar ze zich tevens mee voeden. Deze
kleine gepigmenteerde regenwormen graven geen gangen. Endogeïsche regenwormen leven
gedurende het grootste deel van het jaar in de toplaag van de minerale bodem (0-50 cm), behalve
wanneer het te koud of te droog is. Aangezien ze doorgaans niet aan de oppervlakte komen zijn deze
soorten niet gepigmenteerd. Ze graven horizontale gangen die ze niet hergebruiken, waardoor grote
hoeveelheden bodem hun gestel passeert: zij zijn de zogenaamde geofagen. Samen met bodem nemen
ze verterend organisch materiaal (organische stof) op waaruit ze hun energie halen. Anekische
regenwormen vormen een intermediaire categorie: ze foerageren ook op vers organisch materiaal aan
de oppervlakte, maar brengen de meeste tijd door in de minerale bodem. Ze graven een permanente,
verticale gang die tot 2,40 meter diep gaat (Edwards & Lofty 1977; Sims & Gerard 1999). De doorsnede
van zo een gang is zichtbaar op Figuur 1.5. Doordat ze zich steeds verplaatsen van de oppervlakte naar
diepere lagen en vice versa worden ze soms pendelaars genoemd.
Lowe en Butt (2002a) onderzochten interacties tussen enkele soorten waarbij ze tot volgende
vaststellingen kwamen: (1) juveniele anekische soorten (Lumbricus terrestris en Aporrectodea longa)
leven in het vaarwater van endogeïsche soorten, wanneer ze volwassen zijn verdwijnt deze overlap;
(2) Lumbricus rubellus beïnvloedt andere soorten negatief, wat waarschijnlijk te wijten is aan snelle
reproductie en competitie voor voedsel met soorten als L. terrestris; en (3) dat de intensiteit van
interacties te wijten is aan niche overlap, maar dat de resultaten niet altijd te verklaren zijn aan de hand
van de ecologische groep waartoe de soorten behoren en dat het ontwikkelingsstadium van belang is.

14
Uit onderzoek blijkt dat sommige soorten intermediair gedrag vertonen en dus moeilijk te klasseren zijn
in een van de drie categorieën (Eisenhauer et al. 2008; Emmerling & Felten 2009). In Tabel 1.3 is de
ecologische groep weergegeven van de 19 verschillende soorten regenwormen die tijdens dit onderzoek
gevonden werden. Op een uitzondering na werd steeds de classificatie van Sims & Gerard (1999)
gebruikt. L. rubellus die vrij goed lijkt op L. terrestris werd hier ook onderverdeeld bij de anekische
categorie gezien de bevindingen van Eisenhauer et al. (2008). In datzelfde (laboratorium-)onderzoek
wordt gesuggereerd dat A. longa mogelijks eerder bij de endogeïsche soorten behoort gezien de
kleinere affiniteit voor vers organisch materiaal en afwijkende graafactiviteiten. Aan L. rubellus wordt
ook soms een deels endogeïsch karakter toegewezen, aangezien de soort zich ook voedt in de bovenste
bodemlaag (Suárez et al. 2004; Emmerling & Felten 2009). Intra- en interspecifiek onderzoek is echter
zeer complex gezien vele factoren meespelen en onderzoek in veldomstandigeheden moeilijk is (Uvarov
2009).
Figuur 1.5: Doorsnede van gangen van anekische regenwormen op 0,30 meter diepte, gefotografeerd op 2/11 in tuin
20.
15
Tabel 1.3: Aangetroffen soorten regenwormen met bijhorende ecologische groep, volgens Sims & Gerard (1999) en * volgens Eisenhauer et al. (2008). Elke soort behoort tot de Lumbricidae, behalve Microscolex phosphoreus die een exoot is en tot de familie van de Acanthodrilidae behoort.
Soort regenworm Ecologische groep
Lumbricus terrestris anekisch
* Lumbricus rubellus anekisch
Lumbricus castanea epigeïsch
Lumbricus festivus epigeïsch
Eisenia fetida epigeïsch
Eiseniella tetraedra epigeïsch
Dendrodrilus rubidus epigeïsch
Dendrobaena attemsi epigeïsch
Dendrobaena octaedra epigeïsch
Dendrobaena pygmaea epigeïsch
Aporrectodea longa anekisch
Aporrectodea caliginosa endogeïsch
Aporrectodea limicola endogeïsch
Aporrectodea rosea endogeïsch
Aporrectodea icterica endogeïsch
Allolobophora chlorotica endogeïsch
Octalasion cyaneum endogeïsch
Octalasion lacteum endogeïsch
Microscolex phosphoreus endogeïsch

16
1.2.4 Sturende factoren
Klimaat: temperatuur en vochtigheid
Klimatologische omstandigheden beïnvloeden regenwormen zowel direct door sturing van biologische
processen als indirect door de invloed op het habitat en de voedselvoorziening. De meeste
regenwormen in gematigde regio’s kennen een temperatuuroptimum tussen de 10 en de 20 °C en zullen
sterven bij temperaturen rond 25 à 30 °C en bij temperaturen onder het vriespunt. Vochtigheid is een
andere sturende parameter die gerelateerd is aan de temperatuur en zeker in gematigde regio’s van
groter belang zal zijn. Regenwormen zijn door hun onvermogen vocht goed vast te houden erg gevoelig
aan droge omstandigheden.
Veel soorten hebben zich geadapteerd aan zulke ongunstige omstandigheden waardoor ze
seizoensschommelingen doorkomen. Gedragsveranderingen zijn in droge perioden cruciaal: bepaalde
soorten zoals Allolobophora sp., Aporrectodea sp. en Octalasion sp. zullen naarmate de toplaag van de
bodem droger wordt dieper kruipen in hun gangen en zichzelf oprollen. De ecologisch gestuurde
rustfase (facultatieve diapauze) die regenwormen doorgaan wordt estivatie genoemd (zomerslaap).
Bepaalde soorten zoals A. longa zullen onafhankelijk van klimatologische omstandigheden in rust gaan
(obligatoire diapauze). Ook te veel neerslag kan nefast zijn: overstroomde gangenstelsels zullen
regenwormen naar de oppervlakte drijven, waar ze een eenvoudige prooi zijn voor predatoren zoals
vogels (Edwards & Lofty 1977; Curry 1998; Sims & Gerard 1999).
Bij hoge temperaturen zal organisch materiaal sneller verteren waardoor de voor epigeïsche en
anekische soorten belangrijke strooisellaag sneller zal verdwijnen. Endogeïsche soorten die door een
efficiëntere vertering minder afhankelijk zijn van hoog kwalitatief organisch materiaal hebben hier
minder last van (Curry 1998).
Bodemtextuur
Regenwormen zouden een middelmatige korrelgrootte prefereren boven een zandige of zeer kleiige
bodem. Vooral de schurende werking van zand is problematisch. Verder heeft de bodemtextuur een
invloed op het vochtgehalte in de bodem en zo een indirecte invloed op regenwormen: in dichte
kleibodems zullen bij grote neerslaghoeveelheden snel anaerobe condities optreden en zandbodems
zijn dan weer gevoelig aan verdroging (Edwards & Lofty 1977; Curry 1998).
pH-H20
In sterk zure bodems (pH < 4,5) zullen weinig regenwormen voorkomen. Onder pH 3,5 zijn ze zelfs
afwezig. Hoewel er sterke verschillen zijn tussen soorten onderling komen de meeste gematigde soorten
voor in een range tussen pH 5 en pH 7,4 (Edwards & Lofty 1977). Figuur 1.6 geeft de pH range waarin
15 van de 19 gevonden soorten voorkomen.
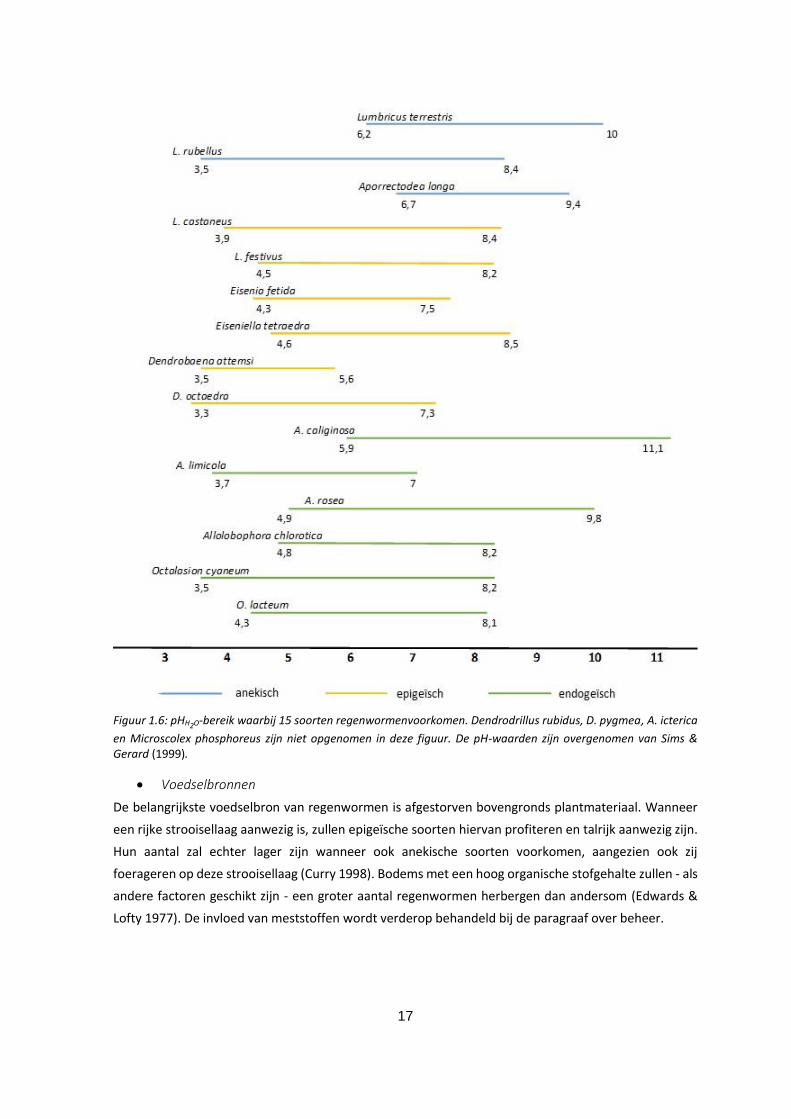
17
Figuur 1.6: pHH2O-bereik waarbij 15 soorten regenwormenvoorkomen. Dendrodrillus rubidus, D. pygmea, A. icterica
en Microscolex phosphoreus zijn niet opgenomen in deze figuur. De pH-waarden zijn overgenomen van Sims & Gerard (1999).
Voedselbronnen
De belangrijkste voedselbron van regenwormen is afgestorven bovengronds plantmateriaal. Wanneer
een rijke strooisellaag aanwezig is, zullen epigeïsche soorten hiervan profiteren en talrijk aanwezig zijn.
Hun aantal zal echter lager zijn wanneer ook anekische soorten voorkomen, aangezien ook zij
foerageren op deze strooisellaag (Curry 1998). Bodems met een hoog organische stofgehalte zullen - als
andere factoren geschikt zijn - een groter aantal regenwormen herbergen dan andersom (Edwards &
Lofty 1977). De invloed van meststoffen wordt verderop behandeld bij de paragraaf over beheer.

18
Voedsel afkomstig van gras, kruidachtigen en bepaalde loofbomen die groeien op basische gronden
(bv. linde, beuk) heeft een C/N-verhouding die 20/1 benadert en is doorgaans van hoge kwaliteit voor
regenwormen. C/N-verhoudingen hoger dan 60/1 zijn ongunstig. Stikstofconcentratie kan een nuttige
indicator zijn om strooiselkwaliteit tussen sterk verschillende habitats te vergelijken, maar zal in
landbouwsystemen met goed verteerbare oogstresten geen goede indicator noch limiterende factor
voor regenwormpopulaties zijn (Curry 1998).
Competitie
Intraspecifieke competitie komt voor met soorten die het habitat en de voedselvoorziening voor
regenwormen beïnvloeden, zoals grazers. Interspecifieke competitie tussen regenwormen onderling is
ondanks de relatief kleine hoeveelheid aanwezige soorten per habitat (1 – 16) een belangrijke factor die
de structuur van een levensgemeenschap kan vormen. Meestal is deze competitie om voedsel.
Interspecifieke interacties zijn niet altijd negatief: gangen worden soms hergebruikt en door anekische
regenwormen verzameld materiaal aan oppervlakte of in gangen kan verorberd worden door
respectievelijk epigeïsche en endogeïsche soorten (Curry 1998).
Beheer
Menselijke activiteit kan regenwormenpopulaties beïnvloeden door zowel directe fysieke storing
(bv. bodembewerking, betreding) als indirect door het habitat en de voedselvoorziening te wijzigen
(bv. meststoffen- en pesticidengebruik, bekalking, irrigatie). Enkele beheerfactoren uit de literatuur
relevant in het kader van de moestuin worden besproken. Opgemerkt dient te worden dat de gebruikte
bronnen over onderzoek in landbouwsystemen gaan en dat er geen wetenschappelijke informatie
voorhanden is specifiek voor moestuinen.
Mortaliteit geassocieerd met bodemverstoring staat in verband met de intensiteit en frequentie ervan.
Grote anekische regenwormen lijden het sterkst onder intensieve bodembewerking als ploegen, door
fysieke verwonding enerzijds en door de vernietiging van hun gangen anderzijds. Doordat de anekische
L. terrestris een lange levenscyclus heeft en zich traag voortplant is deze soort extra gevoelig. Ook het
verdwijnen van de voedselvoorziening aan de oppervlakte bij kerende bodembewerking beïnvloedt de
anekische regenwormen negatief. Dit is ook het geval bij epigeïsche (Curry 1998; Chan 2000; Curry et al.
2002; Rothwell et al. 2011). Het gebrek aan voedsel heeft een grotere impact dan de bodembewerking,
vooral het achterlaten van oogstresten met een lage C/N-verhouding heeft een positieve invloed
(Rothwell et al. 2011).
Endogeïsche soorten zoals A. caliginosa zouden wel kunnen varen bij het inploegen van organisch
materiaal (Ernst & Emmerling 2009). Regenwormen uit de familie Aporrectodea zijn vaak dominant in
akkerland waar de diversiteit doorgaans lager is (Valckx et al. 2009; Ernst & Emmerling 2009; Rothwell
et al. 2011).

19
Andere belangrijke indirecte effecten van bodembewerking die een invloed hebben op regenwormen
zijn veranderde bodemtemperaturen en vochtigheid bij een naakte bodem en versnelde afbraak van
organisch materiaal (Edwards & Lofty 1977; Curry 1998).
Het effect van minerale meststoffen (kunstmest) op regenwormen is variabel. Indien geen buitensporige
hoeveelheden worden aangebracht zijn de resultaten dikwijls positief door een betere strooiselkwaliteit
en hogere strooiselkwantiteit als reactie op de toegediende meststoffen, indien oogstresten op het veld
achtergelaten worden. Organische meststoffen (dierlijke mest) leveren daarbovenop nutriëntenrijk
voedsel voor de regenwormen (Whalen et al. 1998). Te grote hoeveelheden kunnen echter toxisch zijn
(Curry 1998). Een studie aan Universiteit Gent (Leroy et al. 2008) vergeleek het effect van eenzelfde
hoeveelheid toegevoegde organische koolstof doormiddel van stalmest, drijfmest, drie soorten compost
en kunstmest. Twee onbemeste controleplots hadden de kleinste hoeveelheid regenwormen
(± 150 individuen per m²) en de plots bemest met stal- en drijfmest de grootste (800 - 900 individuen
per m²). De compostbehandeling leverde intermediaire aantallen (400 - 500 individuen per m²).
Regenwormen kunnen ook een negatief beïnvloed worden door de pH-daling die gepaard gaat met het
gebruik van stikstofmeststoffen zoals ammoniumsulfaat (Ma et al. 1990).
Kotcon (2011) geeft een overzicht van verschillende studies omtrent regenwormen in alternatieve
(minimale grondbewerking, vanggewassen, etc.), biologische en conventionele landbouwsystemen. De
conclusie is op enkele uitzondering na steeds dat regenwormen talrijker zijn (twee tot negen keer, Chan
2000) in de alternatieve en biologische systemen. Bernard et al. (2014) tonen aan dat gereduceerde
grondbewerking en het inzaaien van vanggewassen op lange termijn voordeliger zijn voor h dan
conventionele systemen met pesticiden en kunstmeststoffen.
In een Europese review-studie van Pelosi et al. (2014) wordt de invloed van pesticiden op regenwormen
onderzocht. Deze studie houdt rekening met de op dat moment toegestane producten op verschillende
niveaus: intra-individueel (genexpressie en fysiologie), individueel en populatieniveau (mortaliteit,
populatiedichtheid en gedrag) en bracht aan het licht dat pesticiden een negatieve impact hebben op
elk niveau. Zo werd bijvoorbeeld de enzymatische activiteit verstoord, steeg de mortaliteit en daalde de
vruchtbaarheid. Insecticiden en fungiciden hadden de sterkste invloed. De grootte van de impact is
soort- en gedragsafhankelijk (Bertrand et al. 2015).
1.2.5 Ecosysteemdiensten van regenwormen
De aanhoudende interesse in regenwormen is te danken aan hun functie als ecosysteem ingenieurs.
Deze term werd geïntroduceerd door Jones et al. (1994) en houdt in dat dit organismen zijn die
rechtstreeks of onrechtstreeks het milieu van andere organismen beïnvloeden (Jones et al., 1994).
Gezien regenwormen extreem belangrijk zijn voor bodemvorming door consumptie, fragmentatie en
intense menging van bodempartikels en organisch materiaal (Edwards & Lofty 1977), werd in deze
masterthesis het woord bodemingenieur gekozen. Hoewel alle soorten de bodem beïnvloeden, zijn de
gevolgen sterk verschillend per ecologische groep. Volgende paragrafen illustreren hun impact.

20
Voedselproductie
Regenwormen kunnen de plantengroei en dus gewasopbrengst verhogen door hun graaf- en
nutritionele activiteiten die een grote invloed hebben op bodemvruchtbaarheid (Edwards & Lofty 1977).
Scheu (2003) vatte de resultaten van 67 experimenten samen waarin de plantrespons op de
aanwezigheid van regenwormen onderzocht werd. In 79% van de studies verhoogde de bovengrondse
biomassa significant. Ook van Groeningen et al. (2014) concludeerden na een meta-analyse dat de
bovengrondse biomassa gemiddeld met 25% steeg als regenwormen aanwezig waren, terwijl de
kwaliteit gelijk bleef. De grootteorde van de effecten was positief gecorreleerd met de aanwezigheid
van oogstresten en met de regenwormendensiteit, en negatief gecorreleerd met de hoeveelheid
toegevoegde anorganische stikstofbemesting (kunstmest). Dit laatste wil niet zeggen dat de
regenwormen negatief beïnvloed werden door de bemesting, maar dat de positieve effecten van
regenwormen op plantengroei marginaal zijn in dit geval.
De twee volgende paragrafen gaan dieper in op aspecten die verantwoordelijk zijn voor verhoogde
gewasopbrengsten.
Fragmentatie en afbraak van organisch materiaal
Regenwormen spelen een essentiële rol bij de eerste verwerking en verkleining van organisch materiaal
(Edwards & Lofty 1977). Dit materiaal is voornamelijk, maar zeker niet uitsluitend, van plantaardige aard.
Zowel levend (e.g. kiemplantjes, zaden) als dood (e.g. afgevallen bladeren, oogstresten) plantaardig
materiaal staan op de menu, net als micro-organismen die in de bodem leven en feces van diverse dieren
(Sims & Gerard 1999). Ook voor het inwerken van organisch materiaal in de bodem, zijn regenwormen
van groot belang. Forse Lumbricus soorten kunnen met hun monddelen zelfs volledige bladeren in hun
gangen trekken. De meeste soorten verorberen in meer of mindere mate ook minerale bodem of
prefereren een mix van beide (Doube et al. 1997). Organisch materiaal wordt verder afgebroken door
micro-organismen en ander bodemleven zoals mijten, springstaarten en andere geleedpotigen. Zachte
planten en dierlijk materiaal kan rechtstreeks of na verwering verteerd worden door micro-organismen,
maar divers plantaardig materiaal zal slechts vatbaar zijn voor vertering na fragmentatie door en
enzymwerking in het spijsverteringsstelsel van bodemdieren (Edwards & Lofty 1977; Lee 1985).
De aanwezigheid van de drie ecologische groepen is van belang voor een goede werking van dit proces.
De epigeïsche regenwormen die in de strooisellaag leven zullen een grote rol spelen bij deze
fragmentatie. Endogeïsche regenwormen, die vooral minerale bodem maar daarbij ook een zekere
hoeveelheid organisch materiaal opnemen, vermengen en verspreiden op die manier het organisch
materiaal in de toplaag van de bodem. Dit wordt uitgescheiden als ‘casts’. Casts of wormenhoopjes
vormen stabiele aggregaten en worden ook geproduceerd door anekische regenwormen. Zij eten grote
hoeveelheden organisch materiaal dat ze met hun monddelen tot diep in de bodem kunnen trekken en
zo verspreiden. Ook wordt door hun graafactiviteiten de bodem grondig vermengd waardoor nutriënten
terug naar de toplaag van de bodem verplaatst kunnen worden (Edwards & Lofty 1977; Sims & Gerard
1999).

21
Nutriëntencyclus
Regenwormen verorberen per dag een grote hoeveelheid organisch materiaal - ongeveer 30% van hun
eigen lichaamsgewicht - dat een aanzienlijk gehalte stikstof bevat. Een deel daarvan wordt
uitgescheiden als plantopneembare stikstof via urine, feces en slijm. Een ander deel wordt opgeslagen
in het lichaam van de worm en komt na de dood terug in systeem terecht. Stikstofmineralisatie wordt
door de aanwezigheid van regenwormen versneld (Edwards & Lofty 1977; Lee 1985; van Groeningen et
al. 2014). Een review-studie door Blouin et al. (2013) wijst op het feit dat aan een groot percentage van
de stikstofbehoefte voldaan kan worden door de aanwezigheid van regenwormen in gras- en akkerland.
Ondanks de doorgaans positieve berichtgeving omtrent regenwormen, staat hun activiteit ook soms in
negatief daglicht. Dit is omwille van hun invloed op de emissie van broeikasgassen (CO2 en N20) en
uitspoeling van nitraat in agro-ecosystemen (Lubbers et al. 2011; Lubbers et al. 2013; Marhan et al.
2015), hoewel verder onderzoek nodig is om de netto-balans op te stellen die rekening houdt met alle
factoren (Soane et al. 2012).
In systemen waar epi- en endogeïsche soorten dominant zijn kan de hoeveelheid uitwisselbaar fosfor
stijgen en eventueel uitspoelen indien er een overschot is (Suárez et al. 2004). Naast stikstof en fosfor
zou eveneens de omzetting van kalium, calcium en magnesium versneld doorgaan bij de aanwezigheid
van regenwormen (Adejuyigbe et al. 2006; Bhadauria & Saxena 2009).
In het darmkanaal van de regenwormen wordt voedsel geïnoculeerd met micro-organismen die er talrijk
voorkomen. Deze zorgen ook na uitscheiding voor een versnelde verdere afbraak van het organisch
materiaal, waardoor nutriënten sneller terug ter beschikking van planten komen. Op die manier oefenen
regenwormen ook indirect een grote invloed uit op de snelheid van de nutriëntencyclus. Ook wordt de
C/N-verhouding van opgegeten strooisel gradueel verlaagd tot ca. 20, waardoor stikstof opneembaar
wordt voor planten (Edwards & Lofty 1977).
Bijdrage aan biodiversiteit
Om het continue proces van opbouw en afbraak van organische stof in de bodem te reguleren is de
aanwezigheid van een goed functionerend voedselweb cruciaal. Het voedselweb bestaat uit alle levende
organismen in de bodem die elk hun eigen functie hebben (Reubens et al. 2010). Interacties tussen
micro-organismen en bodeminvertebraten bestaan niet enkel uit predator-prooi reacties maar ook uit
indirecte effecten zoals competitie voor voedsel en habitatvorming. Regenwormen spelen als
bodemingenieur hierin een belangrijke rol. Zoals eerder aangegeven, zijn uitscheidingen van
regenwormen hotspots voor bodemorganismen. Anekische regenwormen die organisch materiaal
vanop de oppervlakte in de bodem brengen, kunnen zo extra voedsel verschaffen aan endogeïsche
soorten of aan ander bodemleven (Lowe & Butt 2002a). Regenwormen zijn zelf een belangrijke
voedselbron voor mollen, vossen en vogels. Ook loop- en roofkevers zijn predatoren van regenwormen
(Sims & Gerard 1999).

22
Overstromings- en erosieregulatie
In het semi-permanente gangenstelsel van anekische wormen kan heel wat water ondergebracht
worden dat op die manier niet afvloeit én bovendien niet bijdraagt aan bodemerosie (Edwards &
Shipitalo 1998; Bertrand et al. 2015). In Vlaanderen is bodemerosie momenteel een van grootste
bedreigingen voor landbouwbodems (Valckx et al. 2009). Na textuur is aggregaatstabiliteit (samen met
organische stofgehalte) een van de belangrijkste factoren bepalend voor de erosiegevoeligheid van de
toplaag van de bodem. Een goede kruimelstructuur is van belang voor een goede bewerkbaarheid,
doorwortelbaarheid en toegankelijkheid voor het bodemleven. Door hun graafactiviteiten, vermenging
van organisch materiaal met minerale bodem en het produceren van biogene structuren met hoge
stabiliteit (aggregaten) verminderen regenwormen erosie en dragen ze bovendien bij aan een goede
bodemstructuur (Edwards & Lofty 1977; Reubens et al. 2010). Onder biogene structuren vallen onder
meer de onder- en bovengrondse wormenhoopjes en de gangstelsels van endogeïsche en anekische
regenwormen (Valckx et al. 2009). Doordat regenwormen door hun voedingsgewoonten de
bodembedekkingsgraad verkleinen zouden ze ook potentieel kunnen bijdrage aan een erosietoename,
doch is het netto-erosie effect negatief (Edwards & Shipitalo 1998; Valckx et al. 2009).

23
2 MATERIAAL EN METHODEN
2.1 ONDERZOEKSVELD
Dit onderzoek handelt over de private moestuin. Van Dale omschrijft een moestuin als “een tuin
bestemd voor de verbouw van groenten”. Met privaat wordt hier bedoeld dat de verbouwer van deze
groenten dit doet voor eigen gebruik. De moestuin zelf kan eventueel wel onderdeel zijn van een plaats
met een publiek karakter, zoals dit het geval is bij een volkstuin.
Door middel van een oproep in de media (Radio2, Het Nieuwsblad, De Weekbode, …) en een gerichte e-
mailing (TuinHier, VELT, PermacultuurNetwerk, …) werden vrijwilligers uitgenodigd om hun pesticide-
vrije moestuin ter beschikking te stellen voor deze studie. Om zoveel mogelijk dezelfde bodemtypes te
bekomen werden vrijwilligers gezocht die in dezelfde streek woonden. In dit onderzoek was dit de
omgeving rond Roeselare en Kortrijk. Om voldoende permacultuurmoestuinen te bekomen werd deze
randvoorwaarde opgeheven voor dit tuintype. Vervolgens werden de respondenten bevraagd omtrent
de bodembewerkingen en het mestgebruik die zij toepassen. Na selectie werd een afspraak gemaakt
voor de bemonstering van hun moestuin. De elektronisch verzamelde gegevens werden bij het bezoek
gecontroleerd en verder nauwgezet aangevuld op een informatieblad. Tabel 2.1 toont de genoteerde
eigenschappen. Uiteindelijk werden 37 moestuinen geselecteerd, waarvan 11 conventioneel beheerd,
16 ecologisch beheerd en 10 permacultuurtuinen. Staalname gebeurde in de herfst van 2015 tussen
24 oktober en 30 november en enkel in de delen van de tuin waar effectief groenten werden geteeld.
De geografische ligging van de verschillende moestuinen is weergegeven in Figuur 2.1. en coördinaten
zijn terug te vinden in de appendix (Tabel A.2). Eén moestuin (tuin 8, permacultuur) ligt onder de
taalgrens. Tabel 2.2 toont aan welke voorwaarden deze moestuinen minstens voldoen.
Tabel 2.1: Genoteerde moestuinvariabelen bij bezoek aan moestuin.
methode (permacultuur/ecologisch/conventioneel)
oppervlakte (m²)
laatste teelt op bemonsteringsplaats (8 groepen, zie Tabel A.3)
ouderdom moestuin (jaar) en beheersevolutie
weersomstandigheden (temperatuur, zon/bewolkt/regen)
bodembewerking (geen/woelen/spitten/frezen)
teeltwisseling (ja/nee)
bekalking (ja/nee)
compostgebruik (ja/nee)
gebruik van ongecomposteerd organisch materiaal (ja/nee)
mestgebruik (geen/kunstmest/organische mest/dierlijke mest)

24
Figuur 2.1: Geografische ligging van de 37 moestuinen. Zwarte bolletjes geven permacultuurmoestuinen weer, groene bolletjes ecologisch beheer en rode conventioneel beheer. De landbouwstreken werden ter illustratie als achtergrond ingevoegd.
Tabel 2.2: Eigenschappen van de drie moestuincategorieën.
Permacultuur
- Geen bodembewerking
- Steeds bedekte bodem
- Geen kunstmest, noch
pesticiden
Ecologisch - Geen kunstmest, noch
pesticiden
Conventioneel - Steeds kunstmest, nooit
pesticiden
Omwille van de bewezen negatieve invloed van pesticiden op regenwormen (Pelosi et al. 2014) werd
beslist om moestuinen waar pesticiden gebruikt werden buiten beschouwing te laten. Door deze
beslissing kon de beschikbare tijd geïnvesteerd worden in onderzoek in bovengenoemde
moestuintypes.

25
2.2 PROEFOPZET
In elk van 37 moestuinen werden drie plots geselecteerd binnen een gebied van 3 m op 3 m (Figuur 2.2).
Door de drie plots telkens op een vergelijkbare afstand van elkaar te plaatsen, krijgen we een beeld van
de regenworm- en bodemsamenstelling in de bemonsterde moestuinen en het verschil tussen de
tuinen. De variabiliteit binnen elke moestuin wordt buiten beschouwing van dit onderzoek gelaten. Om
rand- en betredingseffecten te vermijden werden geen stalen genomen op een afstand minder dan
0,2 m van randen van omheiningen, perceelgrenzen en paden.
Figuur 2.2: Proefopzet met per tuin 3 plots, gefotografeerd op 29/10 in tuin 11.

26
2.3 STAALNAME
2.3.1 Bodemstaalname en -analyse
Op 0,2 m naast elk plot werd een bodemstaal genomen van de 0 - 0,3 m bodemlaag met een gutsboor
(weergegeven op Figuur 2.3). Per locatie werden de bodemstalen samengevoegd tot een mengstaal. De
stalen werden bewaard bij -18°C.
Figuur 2.3: Gutsboor voor bodemstaalname van 0,3 m diep, gefotografeerd op 1/11 in tuin 18.
Voor analyse werden de bodemstalen ontdooid en werd onverteerd organisch materiaal verwijderd.
Een deel van het vers staal (afkomstig van diepvriesbewaring) werd gezeefd (1 mm) voor analyse van
biobeschikbaar ammonium (NH4+) en nitraat (NO3
-). Het reststaal werd gedroogd (48h bij 40°C in een
droogstoof) voor de bepaling van het vochtgehalte, de textuur, pH, koolstof (C), totale stikstof (N),
fosfor (P), kalium (K), magnesium (Mg) en calcium (Ca).
Alle chemische analyses werden uitgevoerd in het Labo voor Bos & Natuur (ForNaLab), behalve de
metingen van het biobeschikbaar NH4+ en NO3
-, welke uitgevoerd werden in het Laboratorium
Toegepaste Fysicochemie (ISOFYS). Beide labo's werken volgens een strikt kwaliteitssysteem. Volgende
methodes werden aangewend:
Biobeschikbaar NO3- werd colorimetrisch bepaald met een auto-analyzer (AA3 Bran+Luebbe
Continuous) na extractie met 1M kaliumchloride (KCl, 1:5 verdunning).
De pHKCl (potentiële zuurtegraad) werd gemeten met een glaselektrode (Orion, model 920A)
met 1:5 verhouding KCl (1M).
C en totale N werden bepaald via een element-analyse na droge verbranding bij 850°C (Vario
MAX CNS, Elementar).

27
De biobeschikbare concentraties aan P, K, Mg en Ca werden geëxtraheerd met
ammoniumlactaat (AL, 9.01 g melkzuur (88%), 18.75 g azijnzuur (99%), 7.75 g
ammoniumacetaat in 1 L water) in een 1:5 verhouding (massa bodem:volume AL). Vervolgens
werden zowel P (Varian, Cary 50) als de kationen (Varian, AA240 FS) colorimetrisch bepaald.
Bij een pH lager dan 6,5 komen geen vrije carbonaten voor en is het organische C-gehalte nagenoeg
gelijk aan het totale C-gehalte. Om het organische C-gehalte te bepalen bij bodems met een pHKCl hoger
dan 6,5 dient het totale C-gehalte verminderd te worden met het gehalte vrije carbonaten (De Neve et
al. 2006). Het vrij carbonaatgehalte werd echter niet bepaald. Daardoor is het organische C-gehalte in
24 van de 37 tuinen een overschatting (Tabel A.4), waardoor een geen conlusies uit getrokken konden
worden.
Uit het NO3--gehalte werd het nitraatresidu berekend om te kunnen vergelijken met de streefwaarden
voor landbouw (VLM 2014). Het nitraatresidu (N03--residu) is de hoeveelheid nitraat die tussen
1 oktober en 15 november in de bodem achterblijft (tot 0,9 m diep). Deze moet onder een bepaalde
grens liggen (90 kg N03- ha-1) om te vermijden dat nitraat uitspoelt naar het grond- en oppervlaktewater.
Nitraten in het grondwater beïnvloeden de drinkwaterproductie negatief en hoge nitraatconcentraties
in het oppervlaktewater zorgen ervoor dat dit te voedselrijk wordt (eutrofiëring) wat sterke algenbloei
veroorzaakt (Reubens et al. 2010; VLM 2015). Het nitraatresidu werd met onderstaande formule (EMIS
2010a) berekend:
𝐶90 = 3 ∗ 𝐶𝑁,30 × 𝜌30 × 𝐷30
100
C90 = nitraatresidu [kg N03--N ha-1];
CN,30 = gemeten N03--N in de bovenste 0,3 m bodem [mg N03
--N kg-1];
ρ30 = de dichtheid van de bovenste 0,3 m bodem. Hiervoor werd de gemiddelde waarde voor
leem- en zandgronden 1450 kg m-3 gebruikt (EMIS 2010b);
D30 = diepte van de bodemlaag in m (= 0,3 m).
Het vochtgehalte werd als volgt berekend:
𝑉𝑜𝑐ℎ𝑡𝑔𝑒ℎ𝑎𝑙𝑡𝑒 (%) =𝑔𝑒𝑤𝑖𝑐ℎ𝑡 𝑛𝑎𝑡 𝑠𝑡𝑎𝑎𝑙 (𝑔) − 𝑔𝑒𝑤𝑖𝑐ℎ𝑡 𝑑𝑟𝑜𝑜𝑔 𝑠𝑡𝑎𝑎𝑙 (𝑔)
𝑔𝑒𝑤𝑖𝑐ℎ𝑡 𝑛𝑎𝑡 𝑠𝑡𝑎𝑎𝑙 (𝑔) × 100
Van ieder grondstaal werd de textuur bepaald door Geert Baert aan de hand van palpatie, een techniek
waarbij de gezeefde en gedroogde grond aangevoeld wordt. Deze methode wordt ook gehanteerd door
de Bodemkundige Dienst van België (Maes 2012).

28
2.3.2 Regenwormstaalname en determinatie
De kwantitatieve bemonstering van regenwormen omvat drie stappen: handsortering van het strooisel,
mosterdextractie en handsortering van de bodem.
Met behulp van een houten frame van 0,3 m x 0,3 m werd elk plot afgebakend (Figuur 2.4). Indien een
strooisellaag aanwezig was binnen dit frame, werd deze in een afsluitbare plastic zak bewaard en de dag
zelf doorzocht op regenwormen. In het strooisel worden voornamelijk epigeïsche regenwormen
gevonden. Deze leven namelijk aan de oppervlakte en voeden zich met vers organisch materiaal (Sims
& Gerard 1999). Ook werd de bovengrondse vegetatie manueel verwijderd zodat bovenkomende
wormen in de volgende stap makkelijk te detecteren waren.
Figuur 2.4: Voorbeeld van een plot met houten frame te midden van een groenbemester (gele mosterd), gefotografeerd op 1/11 in tuin 18.
Overige regenwormen werden telkens verzameld door het toepassen van een extractie-methode
gevolgd door handsortering. Deze gecombineerde methode levert de beste resultaten (Bouché &
Gardner 1984). Om anekische soorten te verzamelen wordt eerst een uitdrijvingsmiddel gebruikt, dat in
de permanente gangen stroomt welke uitmonden aan de oppervlakte. Bij de meeste endogeïsche
species zal het middel het horizontale gangensysteem moeilijker kunnen bereiken (Zaborski 2003;
Bartlett et al. 2006; Čoja et al. 2008). Juveniele specimen zullen met hun lagere oppervlakte/volume-
verhouding minder adequaat reageren (Bartlett et al. 2006). Handsortering is nodig om een negatieve
bias te vermijden naar voorgenoemde soorten.
Hoewel de in 2011 gepubliceerde ISO-norm 23611 nog steeds formaldehyde aanwijst als
uitdrijvingsmiddel (NBN 2011), is deze chemische stof uiterst schadelijk. Zowel bodemdieren, micro-
organismen en planten ondervinden negatieve effecten (Eichinger et al. 2007). Formaldehyde is
kankerverwekkend (Zaborski 2003). Onderzoek wees uit dat mosterdpoeder opgelost in water een veilig
en effectief alternatief is (Gunn 1992; Iannone et al. 2012). Net als allylisothiocyanaat (AITC), een ander

29
doch minder aangewezen middel (Iannone et al. 2012), zal de mosterdpoederoplossing irriterend
werken op de huid van de wormen (Sims & Gerard 1999). Om voorgenoemde redenen werd bij deze
proefopzet mosterdpoeder (Indasia, art. 6761015) gebruikt als extractant. Op een vrijgemaakte bodem
werd met behulp van een gieter met sproeikop een mosterdpoederoplossing van 6 g L-1 gelijkmatig over
het oppervlak verdeeld. Om deze oplossing te bekomen werd op de avond voorafgaand aan de
bemonstering voor elk plot een stockoplossing van 1 L bereid met een concentratie van 30 g L-1. Op het
veld werd deze stockoplossing in bidons van 5 L gegoten, aangevuld met regenwater en grondig
geschud. Hoeveelheden en concentraties aanbevolen door Claperton et al. (2007) en het ecoworm-
project (Valckx et al. 2009) werden aangepast aan het te bemonsteren bodemvolume. Voorgaande
proeven bij gelijkaardige condities toonden aan dat een volume groter dan 5 L slecht in de grond drong.
De waterige modderpoel die hieruit voorspruit is hinderlijk voor het collecteren van de wormen. Daarom
werd een kleinere hoeveelheid mosterdpoederoplossing toegediend zodra bleek dat de bodem
voortijdig gesatureerd was. Wanneer gedurende 10 minuten geen wormen meer aan de oppervlakte
kwamen, werd de grond binnen het kader met een spade tot 0,30 m diep uitgegraven waarna werd
overgegaan op handsortering (Figuur 2.5).
Hierbij werd de uitgegraven grond op een plastic zeil handmatig verkruimeld en minutieus doorzocht.
Handsortering van uitgegraven bodem is arbeidsintensief, maar effectief voor endogeïsche soorten.
Anekische soorten daarentegen kunnen in de diepere lagen zitten en door deze methode slechts
gedeeltelijk bemonsterd worden (Čoja et al. 2008). Daarom werd eerst een extractie uitgevoerd.
Figuur 2.5: Handsortering als derde en finale bemonsteringsstap, gefotografeerd op 1/11 in tuin 18.
Alle gevangen wormen die binnen het frame naar boven kwamen werden onmiddellijk afgedood en
bewaard in 70% ethanol. In het labo werden de wormen gespoeld (Figuur 2.6) en overgebracht in een
5% formaldehyde oplossing om zo fixatie en goede bewaring te bekomen (Valckx et al. 2009). Na een
Figuur 2.6: De vangst van tuin 34, plot III (22/11). De pigmentatie van de regenwormen is een duidelijk eerste kenmerk bij de determinatie.

30
maand werden de wormen terug overgebracht in 70% ethanol. Determinatie gebeurde in een labo met
behulp van een microscoop en volgens de sleutel van Sims & Gerard (1999) en Sherlock (2012). Indien
mogelijk werd gedetermineerd tot op soort.
Overige (juveniele of onvolledige) wormen werden op basis van 3 kenmerken (epi- of tanylobe kop, al
dan niet gepaardheid van borstels en aan- of afwezigheid van pigment) onderverdeeld in 8 groepen (zie
Tabel A.5). Naargelang de procentuele aanwezigheid van adulte exemplaren (per tuin) corresponderend
met kenmerken van de groep in kwestie werden deze op naam gebracht en opgeteld bij de adulten van
desbetreffende plot.
Vervolgens werden de wormen per soort gewogen nadat de alcohol ca. een minuut verdampte.
2.4 STATISTISCHE ANALYSES
2.4.1 Algemene eigenschappen
Gemiddelden, medianen en standaardafwijkingen werden met behulp van SPSS Statistics 21 berekend
voor enkele eigenschappen van de onderzochte moestuinen. Zo wordt de ouderdom, oppervlakte,
bodembewerking, etc. besproken. Uitgevoerde testen zijn niet-parametrische Kruskal-Wallis
(> 2 groepen) of Mann-Whitney U testen (2 groepen). Om na te gaan welke variabelen (bij > 2 groepen)
precies significant verschilden werden Mann-Whitney U testen als post-hoc uitgevoerd.
2.4.2 Nutriënten en regenwormen
Het aantal en de massa regenwormen uit de drie plots werd per tuin omgerekend naar aantal en massa
per m². Om significante verschillen op te sporen werden verschillende statistische testen uitgevoerd in
SPSS Statistics 21.
Bij voorkeur werd een parametrische variantieanalyse (one-way ANOVA) uitgevoerd gevolgd door een
Tuckey HSD post-hoc test. Een one-way ANOVA test de nulhypothese dat de groepsgemiddelden uit de
verschillende populaties gelijk zijn. Voor deze test zijn twee voorwaarden van belang: normaal verdeelde
data en gelijkheid van de varianties. Deze werden respectievelijk getest door een Shapiro-Wilk test voor
kleine datasets (n < 2000) en een Levene’s test uit te voeren.
Wanneer de assumpties voor deze test niet voldaan waren (normaliteit en homoscedasticiteit van de
varianties), werd een logaritmische transformatie uitgevoerd op de data (Tabel A.6). Gemiddelde en
mediane waarden in figuren en tabellen werden steeds berekend op ruwe ongetransformeerde data.
Indien data-transformatie niet baatte werd een niet-parametrische Kruskal-Wallis (> 2 groepen) of
Mann-Whitney U test (2 groepen) uitgevoerd. Bij deze test worden de gemiddelde rangordes van de
waarnemingen per groep vergeleken in plaats van de groepsgemiddelden. Voor deze variabelen werden
de onderlinge verschillen besproken aan de hand van de mediaan die minder afhankelijk is van
uitschieters. Om na te gaan welke variabelen (bij > 2 groepen) precies significant verschilden werden
Mann-Whitney U testen als post-hoc uitgevoerd.

31
Voor correlatietesten werd gebruikt gemaakt van een bivariate niet-parametrische Spearmans rank
correlatietest.
Weergegeven boxplots zijn steeds Tuckey box plots: extreme waarden die verder dan de whiskers liggen
(1,5 x de interkwartielafstand) worden apart weergegeven en zijn gelabeld volgens het tuinnummer. De
lijn in de box stelt de mediaan voor en de boven- en ondergrens van de box wordt gevormd door het
eerste en derde kwartielpunt.
Grafieken werden gemaakt in MS Excel 2013 en in SPSS Statistics 21.
2.4.3 Diversiteit
Per plot werd het aantal soorten berekend met MS Excel 2013. Omdat niet alleen het aantal soorten,
maar vooral de verdeling ervan een maat is voor diversiteit, werd dit getal teruggerekend tot het
effectief aantal soorten (Jost 2006). In dit getal wordt rekening gehouden met het relatief voorkomen
van elke soort: een plot met bijvoorbeeld drie soorten regenwormen waarbij elke soort even veel
voorkomt zal een hoger effectief aantal soorten hebben dan een plot waarbij een van deze drie soorten
veel dominanter is.
Om het effectief aantal soorten (D) per plot te berekenen werd eerst per plot de Shannon-Weaver index
(H’) bepaald. Dit is een maat die iets zegt over de evenwichtigheid van de populatie en wordt als volgt
berekend:
𝐻′ = − ∑[𝑝𝑖 × ln(𝑝𝑖)]
𝑆
𝑖=1
𝑀𝑒𝑡 𝑆 = totaal aantal soorten in dat plot; 𝑝𝑖 = 𝑛𝑖
𝑁⁄ ; 𝑛𝑖 = aantal exemplaren van soort 𝑖; 𝑁 =
totaal aantal exemplaren
Uit H’ werd vervolgens D berekend met volgende formule:
𝐷 = exp (𝐻′)
Ook H’ en D werden berekend in MS Excel 2013.
Om deze parameters voor de drie moestuintypes onderling te vergelijken werd een one-way ANOVA
uitgevoerd gevolgd door een Tuckey HSD post-hoc test. Het aantal soorten werd vergeleken met het
niet-parametrische alternatief, een Kruskal-Wallis H test met als post-hoc de Mann-Whitney U test.
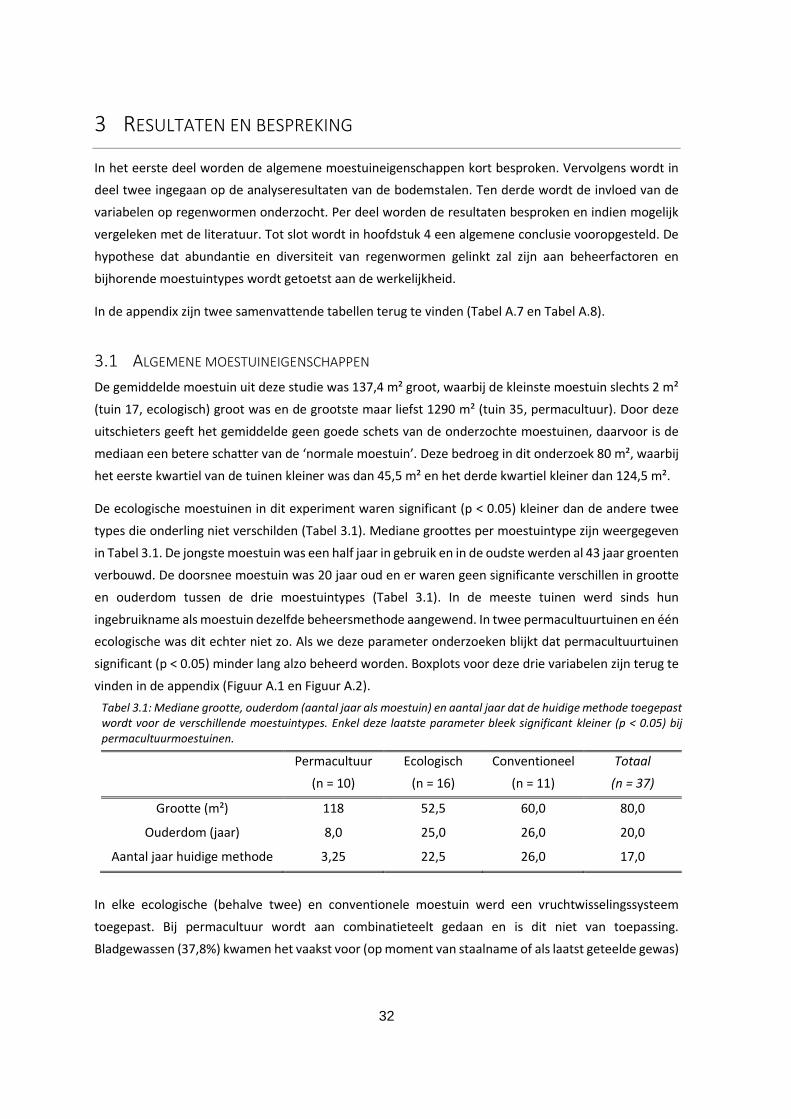
32
3 RESULTATEN EN BESPREKING
In het eerste deel worden de algemene moestuineigenschappen kort besproken. Vervolgens wordt in
deel twee ingegaan op de analyseresultaten van de bodemstalen. Ten derde wordt de invloed van de
variabelen op regenwormen onderzocht. Per deel worden de resultaten besproken en indien mogelijk
vergeleken met de literatuur. Tot slot wordt in hoofdstuk 4 een algemene conclusie vooropgesteld. De
hypothese dat abundantie en diversiteit van regenwormen gelinkt zal zijn aan beheerfactoren en
bijhorende moestuintypes wordt getoetst aan de werkelijkheid.
In de appendix zijn twee samenvattende tabellen terug te vinden (Tabel A.7 en Tabel A.8).
3.1 ALGEMENE MOESTUINEIGENSCHAPPEN
De gemiddelde moestuin uit deze studie was 137,4 m² groot, waarbij de kleinste moestuin slechts 2 m²
(tuin 17, ecologisch) groot was en de grootste maar liefst 1290 m² (tuin 35, permacultuur). Door deze
uitschieters geeft het gemiddelde geen goede schets van de onderzochte moestuinen, daarvoor is de
mediaan een betere schatter van de ‘normale moestuin’. Deze bedroeg in dit onderzoek 80 m², waarbij
het eerste kwartiel van de tuinen kleiner was dan 45,5 m² en het derde kwartiel kleiner dan 124,5 m².
De ecologische moestuinen in dit experiment waren significant (p < 0.05) kleiner dan de andere twee
types die onderling niet verschilden (Tabel 3.1). Mediane groottes per moestuintype zijn weergegeven
in Tabel 3.1. De jongste moestuin was een half jaar in gebruik en in de oudste werden al 43 jaar groenten
verbouwd. De doorsnee moestuin was 20 jaar oud en er waren geen significante verschillen in grootte
en ouderdom tussen de drie moestuintypes (Tabel 3.1). In de meeste tuinen werd sinds hun
ingebruikname als moestuin dezelfde beheersmethode aangewend. In twee permacultuurtuinen en één
ecologische was dit echter niet zo. Als we deze parameter onderzoeken blijkt dat permacultuurtuinen
significant (p < 0.05) minder lang alzo beheerd worden. Boxplots voor deze drie variabelen zijn terug te
vinden in de appendix (Figuur A.1 en Figuur A.2).
In elke ecologische (behalve twee) en conventionele moestuin werd een vruchtwisselingssysteem
toegepast. Bij permacultuur wordt aan combinatieteelt gedaan en is dit niet van toepassing.
Bladgewassen (37,8%) kwamen het vaakst voor (op moment van staalname of als laatst geteelde gewas)
Tabel 3.1: Mediane grootte, ouderdom (aantal jaar als moestuin) en aantal jaar dat de huidige methode toegepast wordt voor de verschillende moestuintypes. Enkel deze laatste parameter bleek significant kleiner (p < 0.05) bij permacultuurmoestuinen.
Permacultuur
(n = 10)
Ecologisch
(n = 16)
Conventioneel
(n = 11)
Totaal
(n = 37)
Grootte (m²) 118 52,5 60,0 80,0
Ouderdom (jaar) 8,0 25,0 26,0 20,0
Aantal jaar huidige methode 3,25 22,5 26,0 17,0

33
op de onderzochte plots (n = 111), gevolgd door wortel-, kool-, vrucht- en peulgewassen (27; 11,7; 9,9
en 7,2%). Beduidend minder vaak werden aardappelen (2,7%), aardbeien (1,8%) en doorlevende
gewassen (1,8%) genoteerd. Dit was deels te wijten aan de periode waarin bemonsterd werd. In Tabel
A.9 wordt het voorkomen van elke gewasgroep per moestuintype weergegeven. De geteelde
gewasgroepen waren niet significant verschillend voor de drie moestuintypes.
15 van de 37 respondenten (40,5%) bekalkte de moestuin in het afgelopen teeltseizoen. 31 personen
(83,3%) gebruikten compost en 17 (45,9%) ongecomposteerd organisch materiaal. 17 tuiniers (45,9%)
maakten gebruik van dierlijke mest, 21 (56,8%) van organische mest en 11 (29,7%) van kunstmest.
In 22 tuinen werd een niet-kerende bodembewerking uitgevoerd:
Permacultuur: acht maal geen bodembewerking, twee maal sporadisch woelen
Ecologisch: vier maal woelen, vier maal frezen
Conventioneel: twee maal woelen, twee maal frezen
In de overige vijftien tuinen werd wel een kerende bodembewerking uitgevoerd (spitten): acht keer in
ecologische en zeven keer in conventionele moestuinen.
Volgens de literatuur was de gemiddelde Vlaamse moestuin, afhankelijk van de bron, 178 m² of 233 m²
groot (Dewaelheyns & Gulinck 2008; OVAM 2012). De moestuinen uit dit onderzoek waren kleiner
(137,4 m² en 80 m² als mediaan) dan deze waarden, wat mogelijks te wijten was aan het relatief groot
aandeel stads- en stadsrandtuinen.
Sommige tuinen waren nog niet lang in gebruik. Omdat regenwormen jaarlijks aanzienlijke afstanden
kunnen afleggen (ca. 10 meter) werd ervan uit gegaan dat dit geen probleem betreft (Edwards & Lofty
1977). Elke onderzochte moestuin grensde aan minstens één ander tuinelementen (bv. grasveld,
bloemperk, struiken, …) waaruit regenwormen konden migreren.
83,1% van de respondenten van dit onderzoek doet aan thuiscompostering. Ook onderzoek van OVAM
wees ook op een hoge composteringsgraad bij Vlamingen (2012). Aangezien in ons onderzoek alle
respondenten een moestuin hebben, ligt dit percentage nog hoger (83,3% t.o.v. 59%), er was immers
een afzetplaats voor de compost: de moestuin. De niet-composteerders zijn in hoofdzaak (4 van de 6)
permacultuurtuiniers die hun tuin- en keukenafval rechtstreeks in de moestuin aanbrachten.
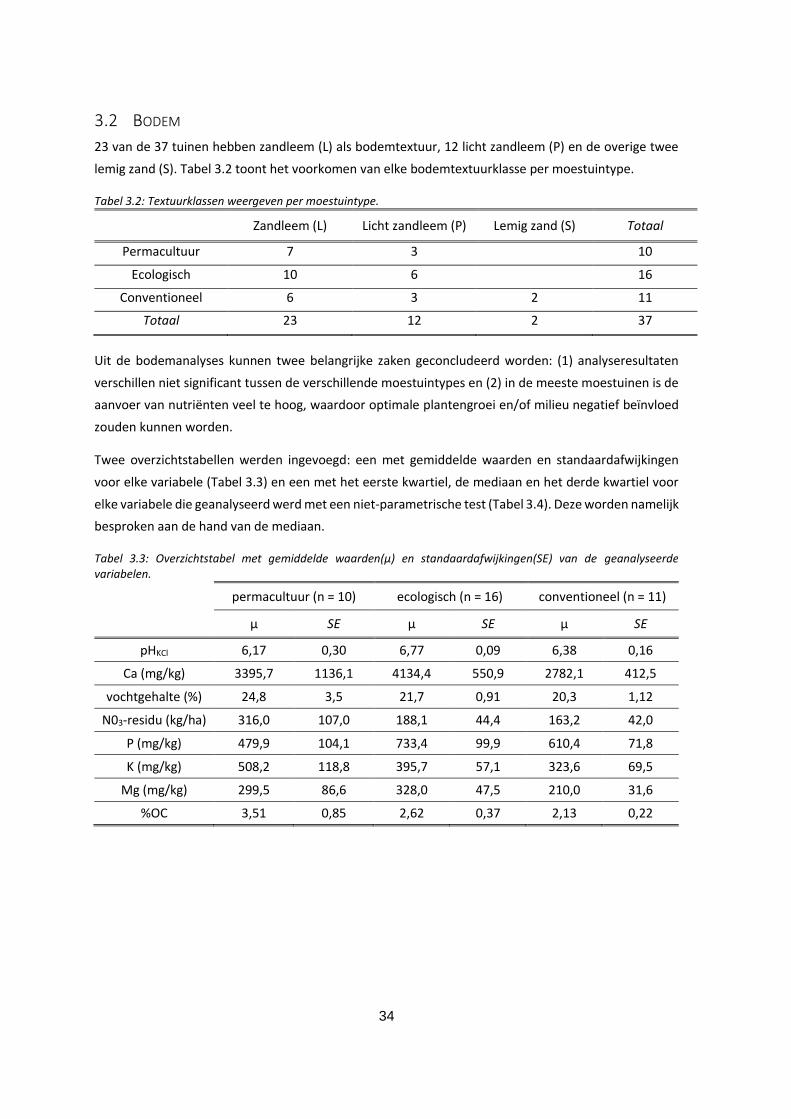
34
3.2 BODEM
23 van de 37 tuinen hebben zandleem (L) als bodemtextuur, 12 licht zandleem (P) en de overige twee
lemig zand (S). Tabel 3.2 toont het voorkomen van elke bodemtextuurklasse per moestuintype.
Tabel 3.2: Textuurklassen weergeven per moestuintype.
Zandleem (L) Licht zandleem (P) Lemig zand (S) Totaal
Permacultuur 7 3 10
Ecologisch 10 6 16
Conventioneel 6 3 2 11
Totaal 23 12 2 37
Uit de bodemanalyses kunnen twee belangrijke zaken geconcludeerd worden: (1) analyseresultaten
verschillen niet significant tussen de verschillende moestuintypes en (2) in de meeste moestuinen is de
aanvoer van nutriënten veel te hoog, waardoor optimale plantengroei en/of milieu negatief beïnvloed
zouden kunnen worden.
Twee overzichtstabellen werden ingevoegd: een met gemiddelde waarden en standaardafwijkingen
voor elke variabele (Tabel 3.3) en een met het eerste kwartiel, de mediaan en het derde kwartiel voor
elke variabele die geanalyseerd werd met een niet-parametrische test (Tabel 3.4). Deze worden namelijk
besproken aan de hand van de mediaan.
Tabel 3.3: Overzichtstabel met gemiddelde waarden(µ) en standaardafwijkingen(SE) van de geanalyseerde variabelen.
permacultuur (n = 10) ecologisch (n = 16) conventioneel (n = 11)
µ SE µ SE µ SE
pHKCl 6,17 0,30 6,77 0,09 6,38 0,16
Ca (mg/kg) 3395,7 1136,1 4134,4 550,9 2782,1 412,5
vochtgehalte (%) 24,8 3,5 21,7 0,91 20,3 1,12
N03-residu (kg/ha) 316,0 107,0 188,1 44,4 163,2 42,0
P (mg/kg) 479,9 104,1 733,4 99,9 610,4 71,8
K (mg/kg) 508,2 118,8 395,7 57,1 323,6 69,5
Mg (mg/kg) 299,5 86,6 328,0 47,5 210,0 31,6
%OC 3,51 0,85 2,62 0,37 2,13 0,22

35
Tabel 3.4: Overzichtstabel met het eerste kwartiel (Q1), de mediaan en het derde kwartiel (Q3) van de geanalyseerde waarden die met een niet-parametrische test onderzocht werden.
permacultuur (n = 10) ecologisch (n = 16) conventioneel (n = 11)
Q1 mediaan Q3 Q1 mediaan Q3 Q1 mediaan Q3
pHKCl 5,54 6,32 7,07 6,56 6,74 7,09 6,32 6,52 6,76
vochtgehalte (%) 16,0 22,3 29,9 19,3 21,6 25,1 18,3 19,9 24,4
N03-residu (kg/ha) 83,2 109,1 726,0 63,2 88,4 274,4 97,2 135,7 183,3
P (mg/kg) 258,6 367,5 667,0 408,0 518,9 1143 409,7 576,4 860,1
Indien referentiewaarden genoemd worden zijn deze afkomstig van de Bodemkundige Dienst van België
(Maes et al. 2012), met uitzondering van deze omtrent het nitraatresidu. Deze werden overgenomen
van de Vlaamse Landmaatschappij (VLM 2014). Een samenvattende tabel is te vinden in de appendix
(Tabel A.10). In Tabel A.11 wordt weergegeven voor welk bodemtype de referentiewaarden gelden.
3.2.1 Zuurtegraad (pH)
De zuurtegraad van de drie verschillende moestuintypes verschilde niet significant. Kwartielen, de
mediaan en extreme waarden voor elk moestuintype zijn zichtbaar op Figuur 3.1.
De pH beïnvloedt de opneembaarheid van voedingselementen voor planten en is bijgevolg een
belangrijke parameter in de moestuin (VELT vzw 2014). Omdat 31 tuiniers (83,1%) compost gebruikten
(pH 7 – 8, (Zmora-Nahum et al. 2007)) en 15 (40,5%) de moestuin in het afgelopen teeltseizoen
bekalkten, werd verwacht dat de meerderheid van de moestuinen geen suboptimale pH zou hebben. 14
moestuinen (37,8%) hadden een optimale pH (6,2 – 6,6), 15 (40,5%) een supra-optimale en 8 (21,6%)
een suboptimale zuurtegraad (Figuur 3.10). Compost werkt enkel contra-verzurend in combinatie met
Figuur 3.1: Boxplot van de pHKCl voor de drie moestuintypes. Groene stippellijnen geven de streefzone aan. De mediaan wordt weergeven door de horizontale lijn in de box. Vijf van de tien permacultuurmoestuinen hebben een te lage pH.
36
calcium (Sæbø & Ferrini 2006), een element dat onze bodems in voldoende mate aanwezig was (zie
paragraaf 3.2.2).
Ondanks de pH-waarden van de moestuintypes niet significant verschillenden, was opmerkelijk dat in
vijf van de acht tuinen met een suboptimale pH permacultuur het beheertype was. In slechts één van
de tien permacultuurmoestuinen werd bekalkt, wat hiervoor aan de basis kan liggen. Bekalken bleek
dikwijls overbodig of minstens overmatig te gebeuren: in tien van de vijftien bekalkte moestuinen lag
de pH boven het optimum. Tuin 25 (permacultuur) heeft een opmerkelijk lage pH (4,27). Deze moestuin
ligt op een voormalige bosbodem die inmiddels acht jaar gecultiveerd is maar nog steeds omringd wordt
door Alnus glutinosa (zwarte els) en Corylus avellana (hazelaar). Vooral zwarte els heeft een verzurend
effect op de bodem (Mohr & Topp 2005; De Schrijver et al. 2012).
Elsen et al. (2011) concludeerden dat op 393 moestuinbodemstalen (geanalyseerd door de
Bodemkundige Dienst van België) 18% een optimale pH had, 67% een te hoge pH en 15% een te lage.
Deze studie is echter niet representatief voor de gemiddelde Vlaamse moestuin, aangezien stalen
dikwijls pas ingeleverd worden indien problemen ondervonden worden. De stalen uit dit experiment
zijn er dan ook iets beter aan toe (40,5% binnen de streefzone t.o.v. 18%).
3.2.2 Calciumgehalte
Op zure bodems (lage pH, veel vrije protonen (H+)), zal calcium verdrongen worden door een grote
hoeveelheid protonen en kunnen tekorten ontstaan. Er was zoals verwacht een positieve correlatie
tussen het calciumgehalte en de pH waar te nemen (p < 0.001, ρ = 0.683). Minimale en maximale
waarden zijn terug te vinden in Figuur 3.2, net als de kwartielen, de mediaan en extreme waarden. Er
waren geen significante verschillen tussen de drie moestuintypes op vlak van het Ca-gehalte.
Figuur 3.2: Boxplot van het calciumgehalte (AL-extractie) voor de drie moestuintypes. Groene stippellijnen geven de streefzone aan. De mediaan wordt weergeven door de horizontale lijn in de box. In tuin 8 wordt basaltmeel aangewend, wat het hoge Ca-gehalte daar kan verklaren.
37
35,1% van de moestuinen had een optimaal Ca-gehalte (2000 – 2400 mg kg-1), 10,1% een suboptimaal
en 53,7% een supra-optimaal (Figuur 3.10). Calcium is belangrijk voor de stevigheid van de gewassen en
draagt ook bij aan een goede bodemstructuur. Een overmaat ervan zal echter de opname van andere
elementen, zoals fosfor en magnesium, bemoeilijken (Reubens et al. 2010; VELT vzw 2014).
3.2.3 Vochtgehalte
Het mediane vochtgehalte verschilde niet significant per moestype. Figuur 3.3 geeft de mediane
waarden weer in een boxplot. Tuin 3 heeft naast een hoog vochgehalte tevens een hoog koolstofgehalte
(zie later), wat wijst op veel organische stof in de bodem. Organische stof heeft namelijk een groot
waterbergend vermogen (Reubens et al. 2010), wat het hoge vochtgehalte in deze moestuin verklaart.
3.2.4 Nitraatresidu
Het gemiddelde nitraatresidu voor permacultuurmoestuinen bedroeg 316,0 ± 107 kg NO3- ha-1
tegenover 188,1 ± 44 kg NO3- ha-1 voor ecologische moestuinen en 163,2 ± 42 kg NO3
- ha-1 voor
conventionele moestuinen.
Figuur 3.4 illustreert de overschrijdingen per moestuintype. 62,1% van alle waarnemingen overschreed
de maximumgrens van 90 kg NO3- ha-1 (Figuur 3.5). Daarvan lag bovendien 32,4% in de categorie ‘zeer
veel te hoog’ ( > 200 kg NO3- ha-1). De meerderheid van de moestuinen uit deze studie zal daardoor een
negatieve impact hebben op de waterkwaliteit. Hoewel de conventionele moestuinen de norm
procentueel het vaakst overschreden, lagen de overschijdingen bij permacultuur zeer hoog. Dit is
opmerkelijk, daar permacultuurtuiniers net proberen om zulke zaken te vermijden. Met een mix van
diep- en oppervlakkig wortelende gewassen wordt er getracht aangebrachte nutriënten zoveel mogelijk
op te vangen.
Figuur 3.3: Boxplot van het vochtgehalte voor de moestuintypes.
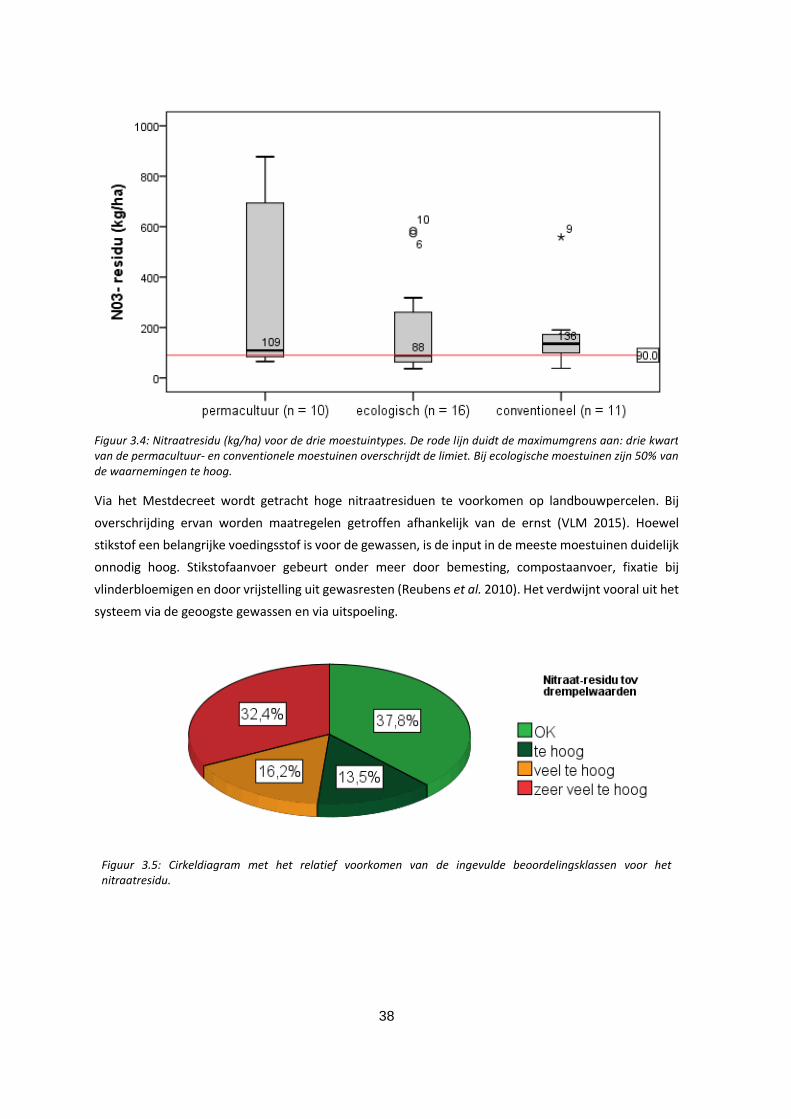
38
Figuur 3.4: Nitraatresidu (kg/ha) voor de drie moestuintypes. De rode lijn duidt de maximumgrens aan: drie kwart van de permacultuur- en conventionele moestuinen overschrijdt de limiet. Bij ecologische moestuinen zijn 50% van de waarnemingen te hoog.
Via het Mestdecreet wordt getracht hoge nitraatresiduen te voorkomen op landbouwpercelen. Bij
overschrijding ervan worden maatregelen getroffen afhankelijk van de ernst (VLM 2015). Hoewel
stikstof een belangrijke voedingsstof is voor de gewassen, is de input in de meeste moestuinen duidelijk
onnodig hoog. Stikstofaanvoer gebeurt onder meer door bemesting, compostaanvoer, fixatie bij
vlinderbloemigen en door vrijstelling uit gewasresten (Reubens et al. 2010). Het verdwijnt vooral uit het
systeem via de geoogste gewassen en via uitspoeling.
Figuur 3.5: Cirkeldiagram met het relatief voorkomen van de ingevulde beoordelingsklassen voor het nitraatresidu.
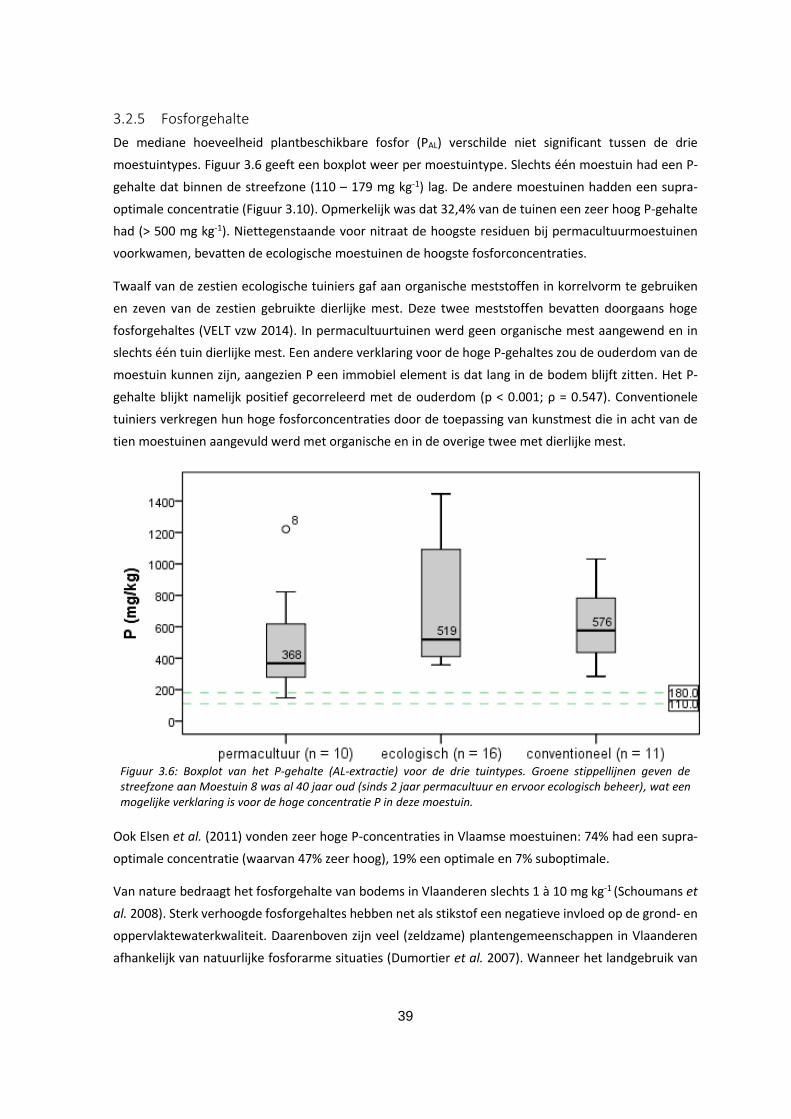
39
3.2.5 Fosforgehalte
De mediane hoeveelheid plantbeschikbare fosfor (PAL) verschilde niet significant tussen de drie
moestuintypes. Figuur 3.6 geeft een boxplot weer per moestuintype. Slechts één moestuin had een P-
gehalte dat binnen de streefzone (110 – 179 mg kg-1) lag. De andere moestuinen hadden een supra-
optimale concentratie (Figuur 3.10). Opmerkelijk was dat 32,4% van de tuinen een zeer hoog P-gehalte
had (> 500 mg kg-1). Niettegenstaande voor nitraat de hoogste residuen bij permacultuurmoestuinen
voorkwamen, bevatten de ecologische moestuinen de hoogste fosforconcentraties.
Twaalf van de zestien ecologische tuiniers gaf aan organische meststoffen in korrelvorm te gebruiken
en zeven van de zestien gebruikte dierlijke mest. Deze twee meststoffen bevatten doorgaans hoge
fosforgehaltes (VELT vzw 2014). In permacultuurtuinen werd geen organische mest aangewend en in
slechts één tuin dierlijke mest. Een andere verklaring voor de hoge P-gehaltes zou de ouderdom van de
moestuin kunnen zijn, aangezien P een immobiel element is dat lang in de bodem blijft zitten. Het P-
gehalte blijkt namelijk positief gecorreleerd met de ouderdom (p < 0.001; ρ = 0.547). Conventionele
tuiniers verkregen hun hoge fosforconcentraties door de toepassing van kunstmest die in acht van de
tien moestuinen aangevuld werd met organische en in de overige twee met dierlijke mest.
Ook Elsen et al. (2011) vonden zeer hoge P-concentraties in Vlaamse moestuinen: 74% had een supra-
optimale concentratie (waarvan 47% zeer hoog), 19% een optimale en 7% suboptimale.
Van nature bedraagt het fosforgehalte van bodems in Vlaanderen slechts 1 à 10 mg kg-1 (Schoumans et
al. 2008). Sterk verhoogde fosforgehaltes hebben net als stikstof een negatieve invloed op de grond- en
oppervlaktewaterkwaliteit. Daarenboven zijn veel (zeldzame) plantengemeenschappen in Vlaanderen
afhankelijk van natuurlijke fosforarme situaties (Dumortier et al. 2007). Wanneer het landgebruik van
Figuur 3.6: Boxplot van het P-gehalte (AL-extractie) voor de drie tuintypes. Groene stippellijnen geven de streefzone aan Moestuin 8 was al 40 jaar oud (sinds 2 jaar permacultuur en ervoor ecologisch beheer), wat een mogelijke verklaring is voor de hoge concentratie P in deze moestuin.
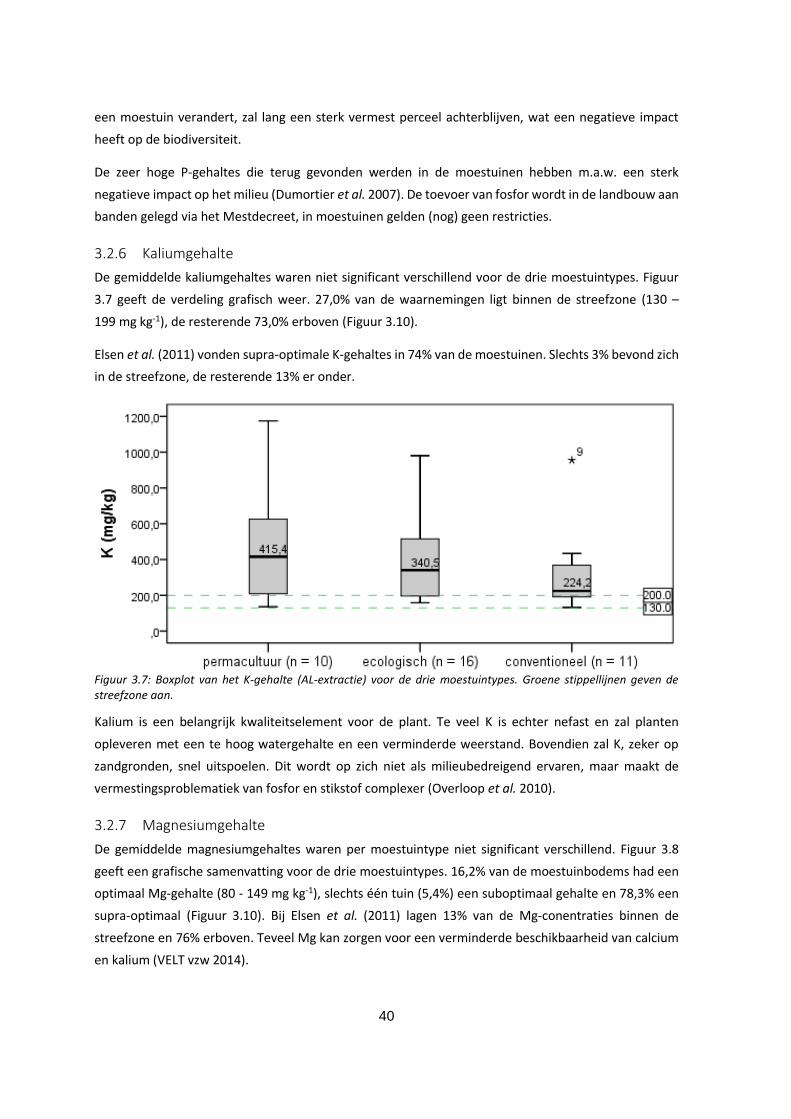
40
een moestuin verandert, zal lang een sterk vermest perceel achterblijven, wat een negatieve impact
heeft op de biodiversiteit.
De zeer hoge P-gehaltes die terug gevonden werden in de moestuinen hebben m.a.w. een sterk
negatieve impact op het milieu (Dumortier et al. 2007). De toevoer van fosfor wordt in de landbouw aan
banden gelegd via het Mestdecreet, in moestuinen gelden (nog) geen restricties.
3.2.6 Kaliumgehalte
De gemiddelde kaliumgehaltes waren niet significant verschillend voor de drie moestuintypes. Figuur
3.7 geeft de verdeling grafisch weer. 27,0% van de waarnemingen ligt binnen de streefzone (130 –
199 mg kg-1), de resterende 73,0% erboven (Figuur 3.10).
Elsen et al. (2011) vonden supra-optimale K-gehaltes in 74% van de moestuinen. Slechts 3% bevond zich
in de streefzone, de resterende 13% er onder.
Kalium is een belangrijk kwaliteitselement voor de plant. Te veel K is echter nefast en zal planten
opleveren met een te hoog watergehalte en een verminderde weerstand. Bovendien zal K, zeker op
zandgronden, snel uitspoelen. Dit wordt op zich niet als milieubedreigend ervaren, maar maakt de
vermestingsproblematiek van fosfor en stikstof complexer (Overloop et al. 2010).
3.2.7 Magnesiumgehalte
De gemiddelde magnesiumgehaltes waren per moestuintype niet significant verschillend. Figuur 3.8
geeft een grafische samenvatting voor de drie moestuintypes. 16,2% van de moestuinbodems had een
optimaal Mg-gehalte (80 - 149 mg kg-1), slechts één tuin (5,4%) een suboptimaal gehalte en 78,3% een
supra-optimaal (Figuur 3.10). Bij Elsen et al. (2011) lagen 13% van de Mg-conentraties binnen de
streefzone en 76% erboven. Teveel Mg kan zorgen voor een verminderde beschikbaarheid van calcium
en kalium (VELT vzw 2014).
Figuur 3.7: Boxplot van het K-gehalte (AL-extractie) voor de drie moestuintypes. Groene stippellijnen geven de streefzone aan.
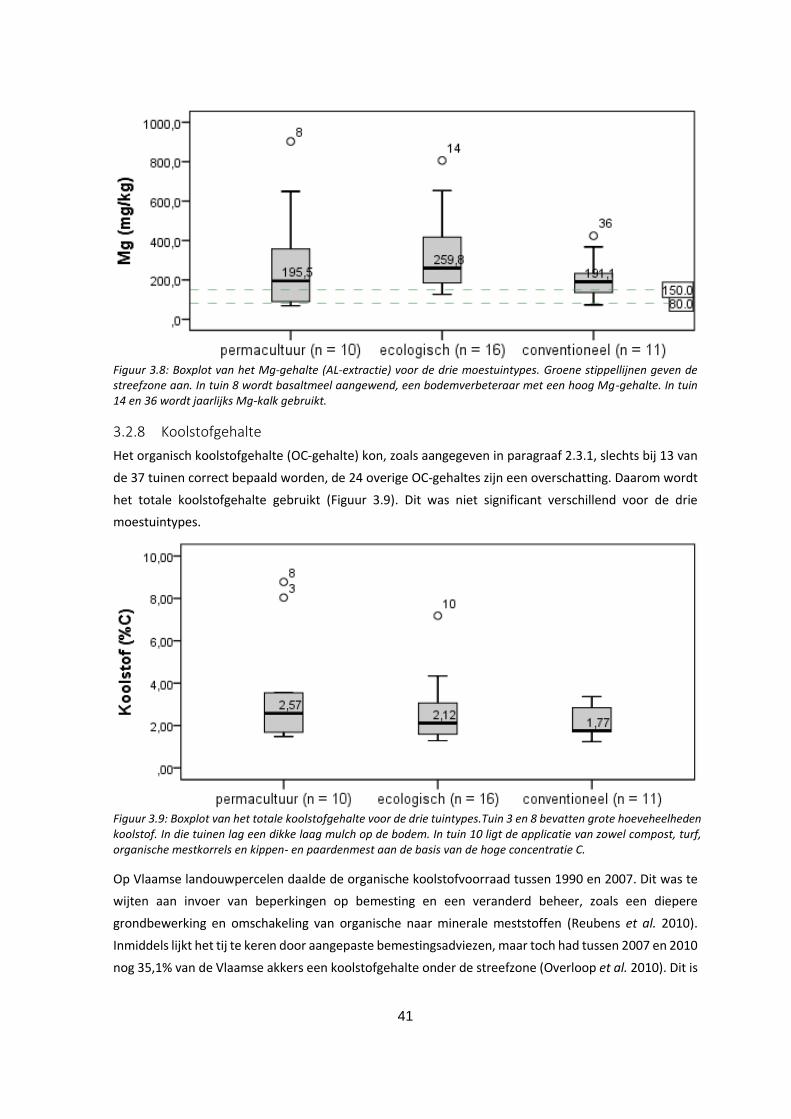
41
Figuur 3.8: Boxplot van het Mg-gehalte (AL-extractie) voor de drie moestuintypes. Groene stippellijnen geven de streefzone aan. In tuin 8 wordt basaltmeel aangewend, een bodemverbeteraar met een hoog Mg-gehalte. In tuin 14 en 36 wordt jaarlijks Mg-kalk gebruikt.
3.2.8 Koolstofgehalte
Het organisch koolstofgehalte (OC-gehalte) kon, zoals aangegeven in paragraaf 2.3.1, slechts bij 13 van
de 37 tuinen correct bepaald worden, de 24 overige OC-gehaltes zijn een overschatting. Daarom wordt
het totale koolstofgehalte gebruikt (Figuur 3.9). Dit was niet significant verschillend voor de drie
moestuintypes.
Op Vlaamse landouwpercelen daalde de organische koolstofvoorraad tussen 1990 en 2007. Dit was te
wijten aan invoer van beperkingen op bemesting en een veranderd beheer, zoals een diepere
grondbewerking en omschakeling van organische naar minerale meststoffen (Reubens et al. 2010).
Inmiddels lijkt het tij te keren door aangepaste bemestingsadviezen, maar toch had tussen 2007 en 2010
nog 35,1% van de Vlaamse akkers een koolstofgehalte onder de streefzone (Overloop et al. 2010). Dit is
Figuur 3.9: Boxplot van het totale koolstofgehalte voor de drie tuintypes.Tuin 3 en 8 bevatten grote hoeveheelheden koolstof. In die tuinen lag een dikke laag mulch op de bodem. In tuin 10 ligt de applicatie van zowel compost, turf, organische mestkorrels en kippen- en paardenmest aan de basis van de hoge concentratie C.

42
nefast voor de bodemkwaliteit: organische stof (bestaande uit ongeveer 58% OC) heeft namelijk een
positieve invloed op onder andere de bodemstructuur, het vochtvasthoudend vermogen en het
bodemleven en is bovendien een belangrijke aanvoerpost voor nutriënten gedurende het groeiseizoen
(Reubens et al. 2010).
Van de dertien moestuinen waarvoor het OC-gehalte correct is, liggen vier waarden binnen de optimale
range en de andere erboven (Tabel A.4). Hieruit kunnen we evenwel niets conluderen voor de overige
moestuinen, gezien dit niet om random gekozen moestuinen gaat, maar over deze met de laagste pH.
In de moestuinen onderzocht door de Bodemkundige Dienst van België (Elsen et al. 2011) heeft de
meerderheid van de tuinen (59%) een supra-optimaal OC-gehalte en 31% een optimaal. Deze hoge
concentraties zijn te wijten aan de vaak jaarlijkse toediening van compost en dierlijke en organische
meststoffen. Ook in de moestuinen uit deze studie worden jaarlijks veel koolstofrijke materialen
aangebracht: 83,1% gebruikt compost, 45,9% dierlijke meststoffen en 56,8% organische meststoffen.
Bovendien werd in elke permacultuurmoestuin (en in zeven andere) vers organisch materiaal
aangebracht. Om deze redenen wordt het OC-gehalte in de meerderheid van de moestuinen niet sub-
optimaal geacht.
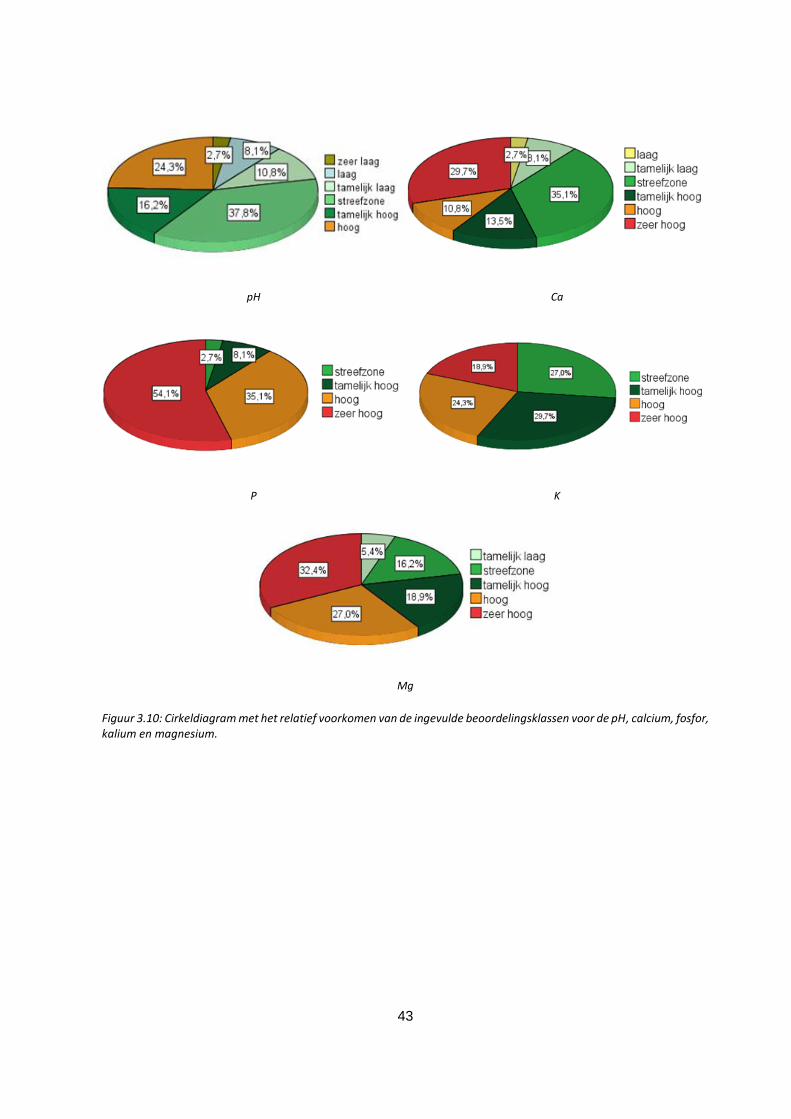
43
pH Ca
P K
Mg
Figuur 3.10: Cirkeldiagram met het relatief voorkomen van de ingevulde beoordelingsklassen voor de pH, calcium, fosfor, kalium en magnesium.
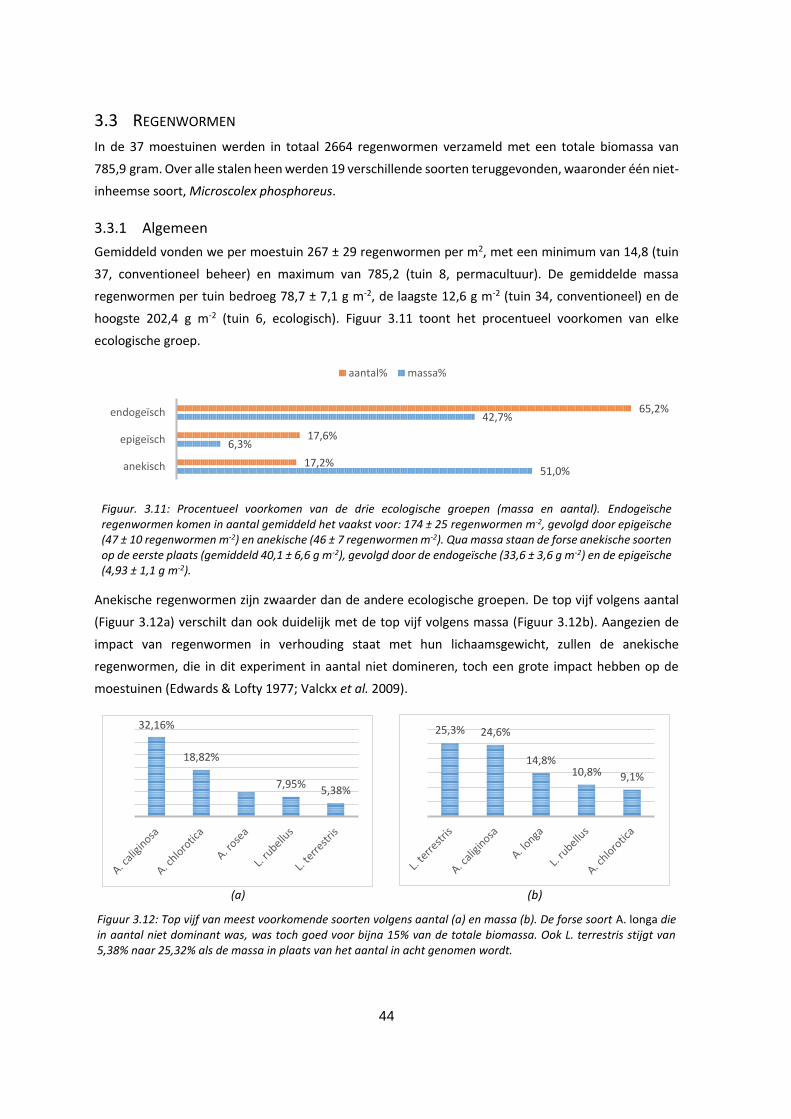
44
3.3 REGENWORMEN
In de 37 moestuinen werden in totaal 2664 regenwormen verzameld met een totale biomassa van
785,9 gram. Over alle stalen heen werden 19 verschillende soorten teruggevonden, waaronder één niet-
inheemse soort, Microscolex phosphoreus.
3.3.1 Algemeen
Gemiddeld vonden we per moestuin 267 ± 29 regenwormen per m2, met een minimum van 14,8 (tuin
37, conventioneel beheer) en maximum van 785,2 (tuin 8, permacultuur). De gemiddelde massa
regenwormen per tuin bedroeg 78,7 ± 7,1 g m-2, de laagste 12,6 g m-2 (tuin 34, conventioneel) en de
hoogste 202,4 g m-2 (tuin 6, ecologisch). Figuur 3.11 toont het procentueel voorkomen van elke
ecologische groep.
Figuur. 3.11: Procentueel voorkomen van de drie ecologische groepen (massa en aantal). Endogeïsche regenwormen komen in aantal gemiddeld het vaakst voor: 174 ± 25 regenwormen m-2, gevolgd door epigeïsche (47 ± 10 regenwormen m-2) en anekische (46 ± 7 regenwormen m-2). Qua massa staan de forse anekische soorten op de eerste plaats (gemiddeld 40,1 ± 6,6 g m-2), gevolgd door de endogeïsche (33,6 ± 3,6 g m-2) en de epigeïsche (4,93 ± 1,1 g m-2).
Anekische regenwormen zijn zwaarder dan de andere ecologische groepen. De top vijf volgens aantal
(Figuur 3.12a) verschilt dan ook duidelijk met de top vijf volgens massa (Figuur 3.12b). Aangezien de
impact van regenwormen in verhouding staat met hun lichaamsgewicht, zullen de anekische
regenwormen, die in dit experiment in aantal niet domineren, toch een grote impact hebben op de
moestuinen (Edwards & Lofty 1977; Valckx et al. 2009).
(a)
(b)
Figuur 3.12: Top vijf van meest voorkomende soorten volgens aantal (a) en massa (b). De forse soort A. longa die in aantal niet dominant was, was toch goed voor bijna 15% van de totale biomassa. Ook L. terrestris stijgt van 5,38% naar 25,32% als de massa in plaats van het aantal in acht genomen wordt.
51,0%
6,3%
42,7%
17,2%
17,6%
65,2%
anekisch
epigeïsch
endogeïsch
aantal% massa%
32,16%
18,82%
7,95%5,38%
25,3% 24,6%
14,8%10,8% 9,1%
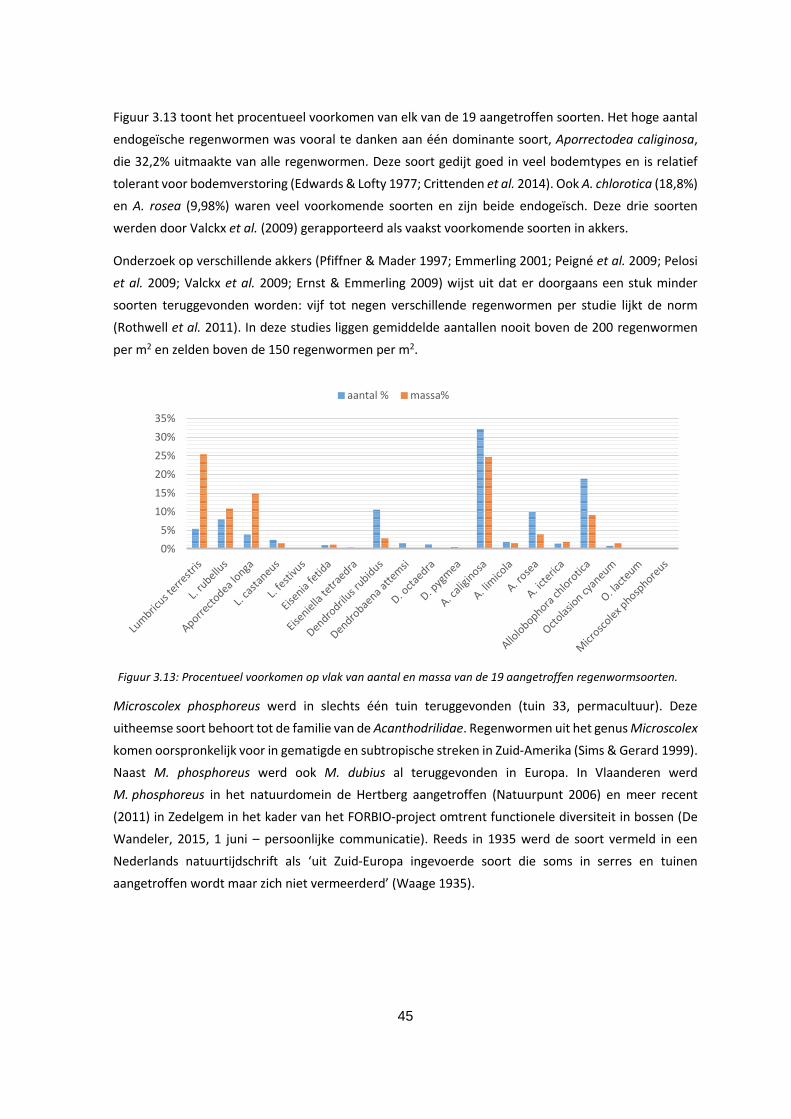
45
Figuur 3.13 toont het procentueel voorkomen van elk van de 19 aangetroffen soorten. Het hoge aantal
endogeïsche regenwormen was vooral te danken aan één dominante soort, Aporrectodea caliginosa,
die 32,2% uitmaakte van alle regenwormen. Deze soort gedijt goed in veel bodemtypes en is relatief
tolerant voor bodemverstoring (Edwards & Lofty 1977; Crittenden et al. 2014). Ook A. chlorotica (18,8%)
en A. rosea (9,98%) waren veel voorkomende soorten en zijn beide endogeïsch. Deze drie soorten
werden door Valckx et al. (2009) gerapporteerd als vaakst voorkomende soorten in akkers.
Onderzoek op verschillende akkers (Pfiffner & Mader 1997; Emmerling 2001; Peigné et al. 2009; Pelosi
et al. 2009; Valckx et al. 2009; Ernst & Emmerling 2009) wijst uit dat er doorgaans een stuk minder
soorten teruggevonden worden: vijf tot negen verschillende regenwormen per studie lijkt de norm
(Rothwell et al. 2011). In deze studies liggen gemiddelde aantallen nooit boven de 200 regenwormen
per m2 en zelden boven de 150 regenwormen per m2.
Figuur 3.13: Procentueel voorkomen op vlak van aantal en massa van de 19 aangetroffen regenwormsoorten.
Microscolex phosphoreus werd in slechts één tuin teruggevonden (tuin 33, permacultuur). Deze
uitheemse soort behoort tot de familie van de Acanthodrilidae. Regenwormen uit het genus Microscolex
komen oorspronkelijk voor in gematigde en subtropische streken in Zuid-Amerika (Sims & Gerard 1999).
Naast M. phosphoreus werd ook M. dubius al teruggevonden in Europa. In Vlaanderen werd
M. phosphoreus in het natuurdomein de Hertberg aangetroffen (Natuurpunt 2006) en meer recent
(2011) in Zedelgem in het kader van het FORBIO-project omtrent functionele diversiteit in bossen (De
Wandeler, 2015, 1 juni – persoonlijke communicatie). Reeds in 1935 werd de soort vermeld in een
Nederlands natuurtijdschrift als ‘uit Zuid-Europa ingevoerde soort die soms in serres en tuinen
aangetroffen wordt maar zich niet vermeerderd’ (Waage 1935).
0%
5%
10%
15%
20%
25%
30%
35%
aantal % massa%
46
Uitheemse regenwormen worden meestal geïntroduceerd door menselijk toedoen. Regenwormen of
cocons kunen tijdens transport van planten meereizen in de aardkluit, maar ook getransporteerd
worden via vermicompost (compost geproduceerd door regenwormen), als aas voor visvangst of
klevend aan gemotoriseerd verkeer (Hendrix et al. 2008).
Uit de Spearmans rank correlatietest blijkt dat de biomassa en het aantal regenwormen voor de drie
ecologische groepen zeer sterk gecorreleerd waren (Tabel 3.5). De totale massa was slechts matig
gecorreleerd met het totale aantal, wat te wijten is aan grote massaverschillen tussen de drie
ecologische groepen. Massa is een minstens even interessante parameter als aantal aangezien deze
gecorreleerd is met het werk dat ondergronds verricht wordt (Valckx et al. 2009). Omdat de totale massa
per m2 matig gecorreleerd was met het aantal én omdat massa niet in elke bron waarmee vergeleken
werd op dezelfde manier bepaald wordt, vermelden we in de bespreking steeds beide parameters.
Tabel 3.5: Spearmans rank correlatietest tussen biomassa en aantal regenwormen per m², p < 0,01.
Aantal (# m-2) en biomassa (g m-2) Spearmans correlatiecoëfficiënt ρ
epigeïsch 0,91 endogïsch 0,83 anekisch 0,78 totaal 0,40
3.3.2 Sturende factoren
(a) Beheer
Verklaringen voor verschillen tussen regenwormabundantie en -diversiteit tussen de moestuintypes
worden verder gezocht aan de hand van beheerfactoren. De bodemanalyses brachten immers aan het
licht dat de nutriëntenstatus van de verschillende moestuintypes niet significant verschilde.
Er was geen significant verschil tussen de drie beheertypes voor de totale biomassa (Tabel 3.6). Op vlak
van aantal was dit wel zo: in permacultuurmoestuinen was het aantal regenwormen per m² significant
hoger dan in conventioneel beheerde moestuinen (369 regenwormen m-2 t.o.v. 189 regenwormen m-2,
p < 0.01), terwijl ecologisch tuinen (202 regenwormen m-2) ten opzichte van beide types niet significant
verschilden.
Uit Figuur 3.14 blijkt dat het moestuintype en het daaraan gelinkte beheer de ecologische groepen
verschillend beïnvloedde. Daarom worden deze groepen apart besproken.
Tabel 3.6: Mediane aantal en biomassa voor de drie moestuintypes. Q1 = eerste kwartiel, Q3 = derde kwartiel. Significante verschillen tussen moestuintypes worden aangeduid met kleine letters (a<b<c; Kruskal-Wallis & Mann-Whitney U). Indien geen letter weergegeven wordt zijn er geen verschillen genoteerd.
Permacultuur (n = 10) Ecologisch (n = 16) Conventioneel (n = 11)
Q1 mediaan Q3 ≠ Q1 mediaan Q3 ≠ Q1 mediaan Q3 ≠
Totaal aantal regenwormen (#/m²)
262,0 368,5 399,1 b 114,8 201,9 457,4 ab 137,0 188,9 229,6 a
Totale biomassa regenwormen (g/m²)
47,0 83,8 102,7 51,7 87,3 125,7 15,9 60,4 87,7
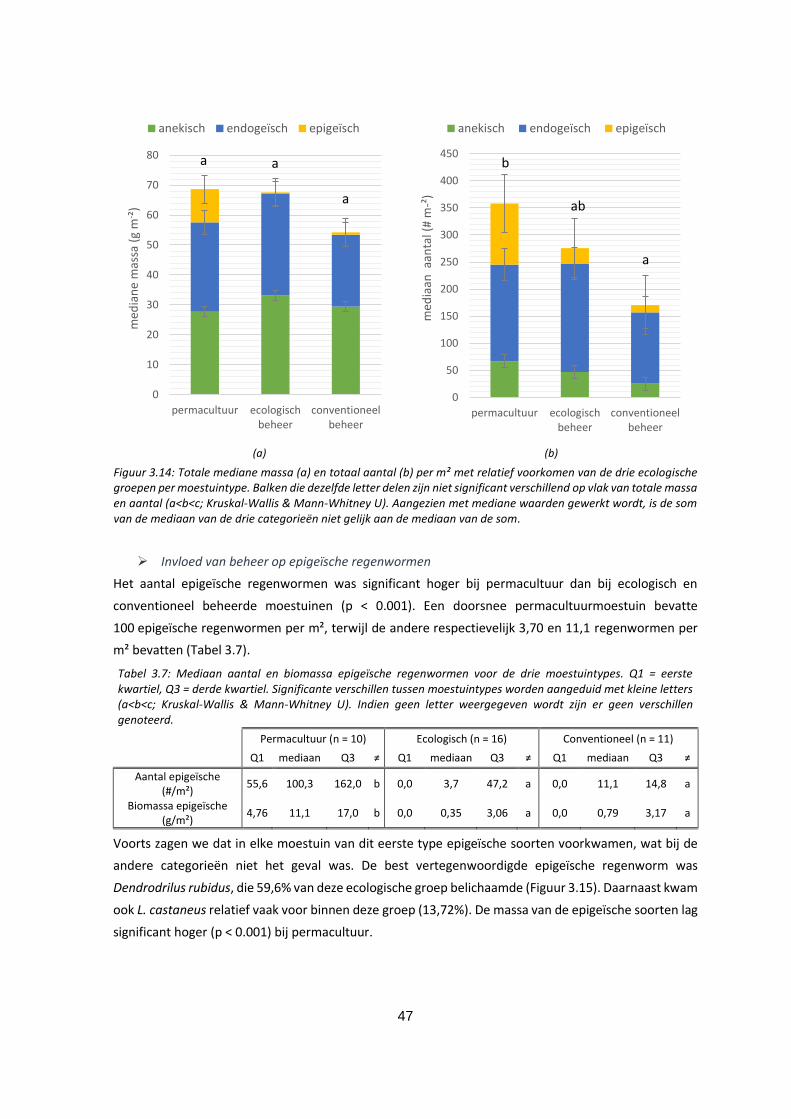
47
(a) (b)
Figuur 3.14: Totale mediane massa (a) en totaal aantal (b) per m² met relatief voorkomen van de drie ecologische groepen per moestuintype. Balken die dezelfde letter delen zijn niet significant verschillend op vlak van totale massa en aantal (a<b<c; Kruskal-Wallis & Mann-Whitney U). Aangezien met mediane waarden gewerkt wordt, is de som van de mediaan van de drie categorieën niet gelijk aan de mediaan van de som.
Invloed van beheer op epigeïsche regenwormen
Het aantal epigeïsche regenwormen was significant hoger bij permacultuur dan bij ecologisch en
conventioneel beheerde moestuinen (p < 0.001). Een doorsnee permacultuurmoestuin bevatte
100 epigeïsche regenwormen per m², terwijl de andere respectievelijk 3,70 en 11,1 regenwormen per
m² bevatten (Tabel 3.7).
Voorts zagen we dat in elke moestuin van dit eerste type epigeïsche soorten voorkwamen, wat bij de
andere categorieën niet het geval was. De best vertegenwoordigde epigeïsche regenworm was
Dendrodrilus rubidus, die 59,6% van deze ecologische groep belichaamde (Figuur 3.15). Daarnaast kwam
ook L. castaneus relatief vaak voor binnen deze groep (13,72%). De massa van de epigeïsche soorten lag
significant hoger (p < 0.001) bij permacultuur.
0
10
20
30
40
50
60
70
80
permacultuur ecologischbeheer
conventioneelbeheer
med
ian
e m
assa
(g
m- ²
)anekisch endogeïsch epigeïsch
a a
a
0
50
100
150
200
250
300
350
400
450
permacultuur ecologischbeheer
conventioneelbeheer
med
iaan
aan
tal (
# m
-²)
anekisch endogeïsch epigeïsch
b
ab
a
Tabel 3.7: Mediaan aantal en biomassa epigeïsche regenwormen voor de drie moestuintypes. Q1 = eerste kwartiel, Q3 = derde kwartiel. Significante verschillen tussen moestuintypes worden aangeduid met kleine letters (a<b<c; Kruskal-Wallis & Mann-Whitney U). Indien geen letter weergegeven wordt zijn er geen verschillen genoteerd.
Permacultuur (n = 10) Ecologisch (n = 16) Conventioneel (n = 11)
Q1 mediaan Q3 ≠ Q1 mediaan Q3 ≠ Q1 mediaan Q3 ≠
Aantal epigeïsche (#/m²)
55,6 100,3 162,0 b 0,0 3,7 47,2 a 0,0 11,1 14,8 a
Biomassa epigeïsche (g/m²)
4,76 11,1 17,0 b 0,0 0,35 3,06 a 0,0 0,79 3,17 a
48
De grote abundantie aan epigeïsche regenwormen kon hier verwacht worden. Deze soorten leven
immers in een laag organisch materiaal aan de oppervlakte, welke in elke permacultuurtuin te vinden
is.
Epigeïsche regenwormen komen in de regel weinig voor op akkers, hoewel hun aantal stijgt indien
overgegaan wordt op niet-kerende grondbewerking waarbij gewasresten aan de oppervlakte blijven
liggen in plaats van ondergeploegd worden (Valckx et al. 2009; Rothwell et al. 2011; Pelosi et al. 2015).
D. rubidus en L. castaneus werden in een studie op Vlaamse akkers (Valckx et al. 2009) ook
teruggevonden, zij het in zeer kleine aantallen. In moestuinen kunnen de epigeïsche regenwormen een
rol spelen bij de fragmentatie en primaire afbraak van oogstresten en ander organisch materiaal dat aan
de oppervlakte ligt, waardoor nutriënten sneller terug beschikbaar komen voor de gewassen. Samen
met de anekische regenwormen zullen ze het organisch materiaal verorberen. Een goede werking van
dit systeem en een evenwichtige aanvoer maakt manueel inwerken overbodig.
Zowel het aantal als de massa aan epigeïsche regenwormen was negatief gecorreleerd (paantal < 0.001,
ρaantal = -0.528 en pmassa < 0.05, ρmassa = -0.378) met het aanbrengen van organische meststoffen (in
korrelvorm). Deze meststof werd veel gebruikt bij de niet-permacultuurtuinen: 21 van de 27 tuiniers
maakten er gebruik van. Omdat geen enkele permacultuurtuinier deze meststof gebruikte én het aantal
epigeïsche regenwormen in deze moestuinen hoger ligt dan bij de andere, vermoeden we dat deze
correlatie niet wijst op een causaal verband. De toepassing van dierlijke meststoffen (vers) verklaarde
de abundantie of massa van epigeïsche regenwormen niet.
Dat de totale biomassa per m² tussen de moestuintypes niet significant verschilde en het totale aantal
per m² wel (Figuur 3.14), wordt verklaard door de lage biomassa van het grote aantal epigeïsche
regenwormen bij de permacultuurtuinen.
Invloed van beheer op endogeïsche regenwormen
Deze ecologische groep was in de drie moestuintypes gelijkwaardig vertegenwoordigd, zowel op vlak
van aantal als biomassa (Tabel 3.8). De drie vaakst voorkomende soorten waren steeds A. caliginosa,
A. rosea en A. chlorotica (Figuur 3.15a). Gezien deze soorten het best bodemverstoring verdragen
(Valckx et al. 2009; Rothwell et al. 2011) en bodemchemische eigenschappen van de drie moestuintypes
niet verschillen, lag deze uitkomst in de lijn van de verwachtingen.
Tabel 3.8: Mediaan aantal en biomassa epigeïsche regenwormen voor de drie moestuintypes. Q1 = eerste kwartiel, Q3 = derde kwartiel. Significante verschillen tussen moestuintypes worden aangeduid met kleine letters (a<b<c; Kruskal-Wallis & Mann-Whitney U). Indien geen letter weergegeven wordt zijn er geen verschillen genoteerd.
Permacultuur (n = 10) Ecologisch (n = 16) Conventioneel (n = 11)
Q1 mediaan Q3 ≠ Q1 mediaan Q3 ≠ Q1 mediaan Q3 ≠
Aantal endogeïsche (#/m²)
50,0 146,3 217,6 52,8 155,6 325,0 96,3 133,3 196,3
Biomassa endogeïsche (g/m²)
21,1 29,8 47,1 20,6 34,0 52,6 11,3 24,0 39,2
49
De endogeïsche regenwormen zullen in moestuinen de minerale bodem grondig vermengen met het
aanwezige organisch materiaal en zo bijdragen aan de vertering ervan. Doordat ze grote hoeveelheden
bodem opnemen en weer uitscheiden als stabiele aggregaten verbeteren ze bovendien de
bodemstructuur sterk. Hun activiteit is bevorderlijk voor de gewenste kruimelstructuur die
bewerkbaarheid, doorwortelbaarheid en toegankelijkheid voor ander bodemleven zal verhogen
(Edwards & Lofty 1977; Reubens et al. 2010).
Invloed van beheer op anekische regenwormen
Anekische regenwormen zullen net zoals de epigeïsche een belangrijke rol spelen bij de primaire afbraak
van organisch materiaal dat aan de oppervlakte ligt. Doordat ze dit materiaal meesleuren in hun
permanente gangen brengen ze voedingstoffen terug in de bodem. Hun gangstelsel is bovendien nuttig
voor waterberging bij hevige regenval en zal de luchthuishouding verbeteren (Edwards & Lofty 1977;
Bertrand et al. 2015).
Ook deze groep was gelijk vertegenwoordigd in de drie moestuintypes. Aangezien bodemverstoringen
bij permacultuur minimaal waren én er veel voedsel te vinden is aan de oppervlakte verwachtten we in
dit moestuintype het grootste aantal anekische regenwormen. Dit was echter niet zo: ondanks het feit
dat de mediane aantallen hoger waren bij permacultuur en ecologische tuinen ten opzichte van
conventionele tuinen (Tabel 3.9), was dit verschil niet statistisch onderbouwd. De bijhorende massa’s
waren ook niet significant verschillend bij de drie moestuintypes.
Omdat binnen het ecologische (4 x woelen, 8 x spitten en 4 x frezen) en conventionele moestuintypes
(2 x woelen, 7 x spitten en 2 x frezen) verschillende bodembewerkingen uitgevoerd werden, werd
nagegaan of het type bodembewerking aan de basis lag voor het gebrek aan verschil met permacultuur.
Verondersteld wordt dat woelen het minst verstorend is voor de permanente gangen. Ook hier konden
geen significante verschillen gedetecteerd worden. Wat niet strookte met de literatuur was dat A. longa,
die verondersteld wordt zeer slecht verstoring te verdragen (Chan 2000; Valckx et al. 2009), significant
meer voorkwam bij spitten dan bij de drie andere bodembewerkingstypes (paantal < 0.05, pmassa< 0.05).
In Figuur 3.15 valt op dat de meest voorkomende anekische worm L. terrestris was bij de ecologische
moestuinen terwijl dit bij permacultuur duidelijk L. rubellus was. De aanwezigheid van beide soorten
bleek negatief gecorreleerd (paantal < 0.05; ρaantal= -0.349; pmassa < 0.05; ρmassa= 0.381). Dit was
waarschijnlijk een geval van intraspecifieke competitie. Bij permacultuur werd wel een grote abundantie
van L. terrestris verwacht omdat de omstandigheden optimaal zijn (weinig bodemverstoring en veel
Tabel 3.9: Mediaan aantal en biomassa anekische regenwormen voor de drie moestuintypes. Significante verschillen tussen moestuintypes worden aangeduid met kleine letters (a<b<c; Kruskal-Wallis & Mann-Whitney U). Indien geen letter weergegeven wordt zijn er geen verschillen genoteerd.
Permacultuur (n = 10) Ecologisch (n = 16) Conventioneel (n = 11)
Q1 mediaan Q3 Q1 mediaan Q3 Q1 mediaan Q3
Aantal anekische (#/m²)
2,8 45,2 126,5 20,4 40,7 67,6 11,1 25,9 34,1
Biomassa anekische (g/m²)
0,50 27,7 57,7 14,5 33,3 108,6 8,07 29,4 40,4

50
voedsel). In labo-experimenten werd deze negatieve interactie al aangetoond (Lowe & Butt 2002a; Lowe
& Butt 2002b; Uvarov 2009). Ze werkte slechts in een richting (L. rubellus > L. terrestris) en kwam enkel
voor als er naast organisch materiaal aan de oppervlakte ook organisch materiaal in de bodem was
ondergemengd omdat L. rubellus zich zowel aan de oppervlakte als ondergronds voedt. Dit is niet het
geval bij L. terrestris, wat L. rubellus een competitief voordeel biedt. Het zou dus kunnen dat L. rubellus
in permacultuurtuinen - waar beide voedselbronnen aanwezig zijn - te competitief was voor L. terrestris
en deze wegconcurreerde op nutritioneel vlak. Uvarov (2009) beschrijft een gelijkaardige negatieve
invloed van L. rubellus op A. longa. Ook in ons experiment bleken beide soorten negatief gecorreleerd
(paantal < 0.05; ρaantal= -0.363).
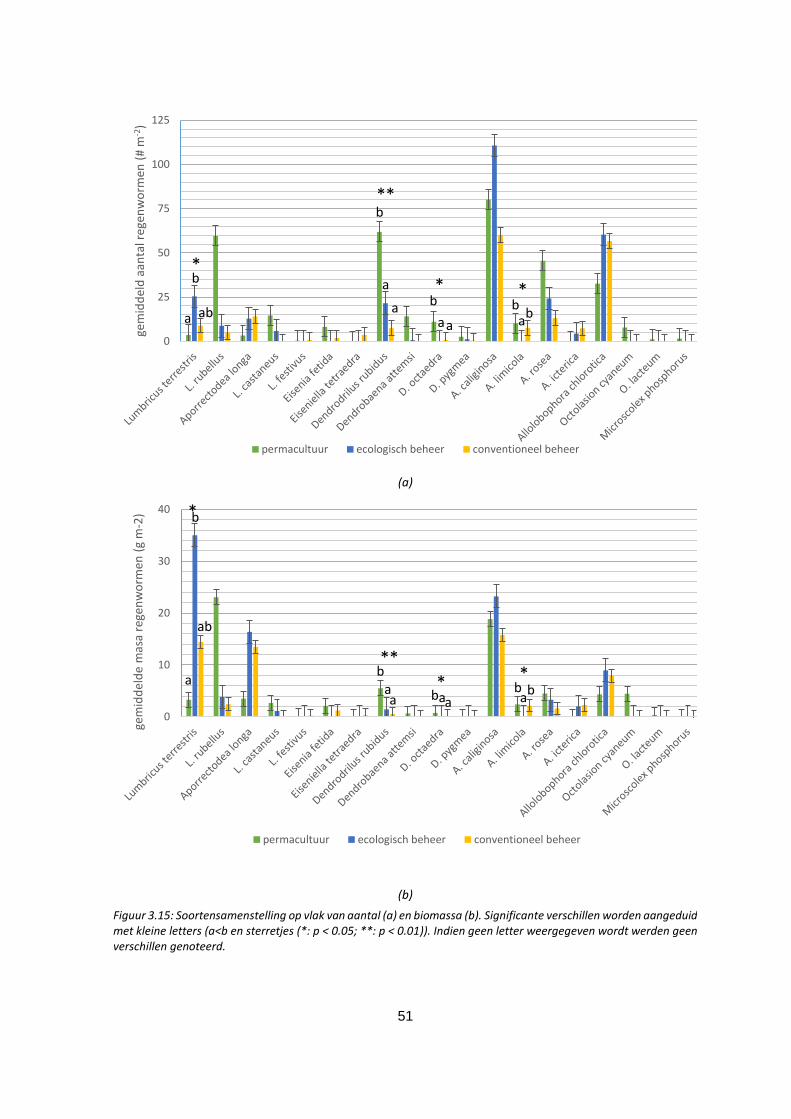
51
(a)
(b)
Figuur 3.15: Soortensamenstelling op vlak van aantal (a) en biomassa (b). Significante verschillen worden aangeduid met kleine letters (a<b en sterretjes (*: p < 0.05; **: p < 0.01)). Indien geen letter weergegeven wordt werden geen verschillen genoteerd.
0
25
50
75
100
125
gem
idd
eld
aan
tal r
egen
wo
rmen
(#
m-2
)
permacultuur ecologisch beheer conventioneel beheer
a ab
b
b
a
a
*
**
b
aa
*
ab
b
*
0
10
20
30
40
gem
idd
eld
e m
asa
rege
nw
orm
en (
g m
-2)
permacultuur ecologisch beheer conventioneel beheer
b
ab
ab
aa baa
b a
b
*
**
* *
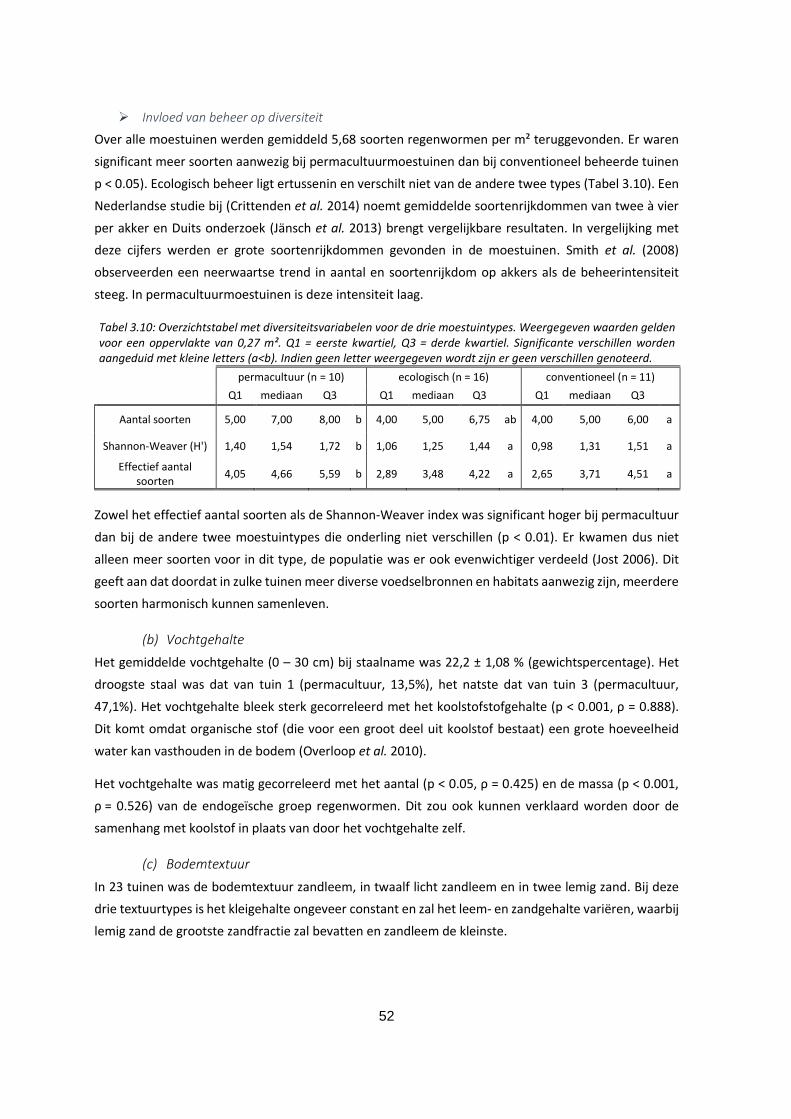
52
Invloed van beheer op diversiteit
Over alle moestuinen werden gemiddeld 5,68 soorten regenwormen per m² teruggevonden. Er waren
significant meer soorten aanwezig bij permacultuurmoestuinen dan bij conventioneel beheerde tuinen
p < 0.05). Ecologisch beheer ligt ertussenin en verschilt niet van de andere twee types (Tabel 3.10). Een
Nederlandse studie bij (Crittenden et al. 2014) noemt gemiddelde soortenrijkdommen van twee à vier
per akker en Duits onderzoek (Jänsch et al. 2013) brengt vergelijkbare resultaten. In vergelijking met
deze cijfers werden er grote soortenrijkdommen gevonden in de moestuinen. Smith et al. (2008)
observeerden een neerwaartse trend in aantal en soortenrijkdom op akkers als de beheerintensiteit
steeg. In permacultuurmoestuinen is deze intensiteit laag.
Zowel het effectief aantal soorten als de Shannon-Weaver index was significant hoger bij permacultuur
dan bij de andere twee moestuintypes die onderling niet verschillen (p < 0.01). Er kwamen dus niet
alleen meer soorten voor in dit type, de populatie was er ook evenwichtiger verdeeld (Jost 2006). Dit
geeft aan dat doordat in zulke tuinen meer diverse voedselbronnen en habitats aanwezig zijn, meerdere
soorten harmonisch kunnen samenleven.
(b) Vochtgehalte
Het gemiddelde vochtgehalte (0 – 30 cm) bij staalname was 22,2 ± 1,08 % (gewichtspercentage). Het
droogste staal was dat van tuin 1 (permacultuur, 13,5%), het natste dat van tuin 3 (permacultuur,
47,1%). Het vochtgehalte bleek sterk gecorreleerd met het koolstofstofgehalte (p < 0.001, ρ = 0.888).
Dit komt omdat organische stof (die voor een groot deel uit koolstof bestaat) een grote hoeveelheid
water kan vasthouden in de bodem (Overloop et al. 2010).
Het vochtgehalte was matig gecorreleerd met het aantal (p < 0.05, ρ = 0.425) en de massa (p < 0.001,
ρ = 0.526) van de endogeïsche groep regenwormen. Dit zou ook kunnen verklaard worden door de
samenhang met koolstof in plaats van door het vochtgehalte zelf.
(c) Bodemtextuur
In 23 tuinen was de bodemtextuur zandleem, in twaalf licht zandleem en in twee lemig zand. Bij deze
drie textuurtypes is het kleigehalte ongeveer constant en zal het leem- en zandgehalte variëren, waarbij
lemig zand de grootste zandfractie zal bevatten en zandleem de kleinste.
Tabel 3.10: Overzichtstabel met diversiteitsvariabelen voor de drie moestuintypes. Weergegeven waarden gelden voor een oppervlakte van 0,27 m². Q1 = eerste kwartiel, Q3 = derde kwartiel. Significante verschillen worden aangeduid met kleine letters (a<b). Indien geen letter weergegeven wordt zijn er geen verschillen genoteerd.
permacultuur (n = 10) ecologisch (n = 16) conventioneel (n = 11)
Q1 mediaan Q3 Q1 mediaan Q3 Q1 mediaan Q3
Aantal soorten 5,00 7,00 8,00 b 4,00 5,00 6,75 ab 4,00 5,00 6,00 a
Shannon-Weaver (H') 1,40 1,54 1,72 b 1,06 1,25 1,44 a 0,98 1,31 1,51 a
Effectief aantal soorten
4,05 4,66 5,59 b 2,89 3,48 4,22 a 2,65 3,71 4,51 a
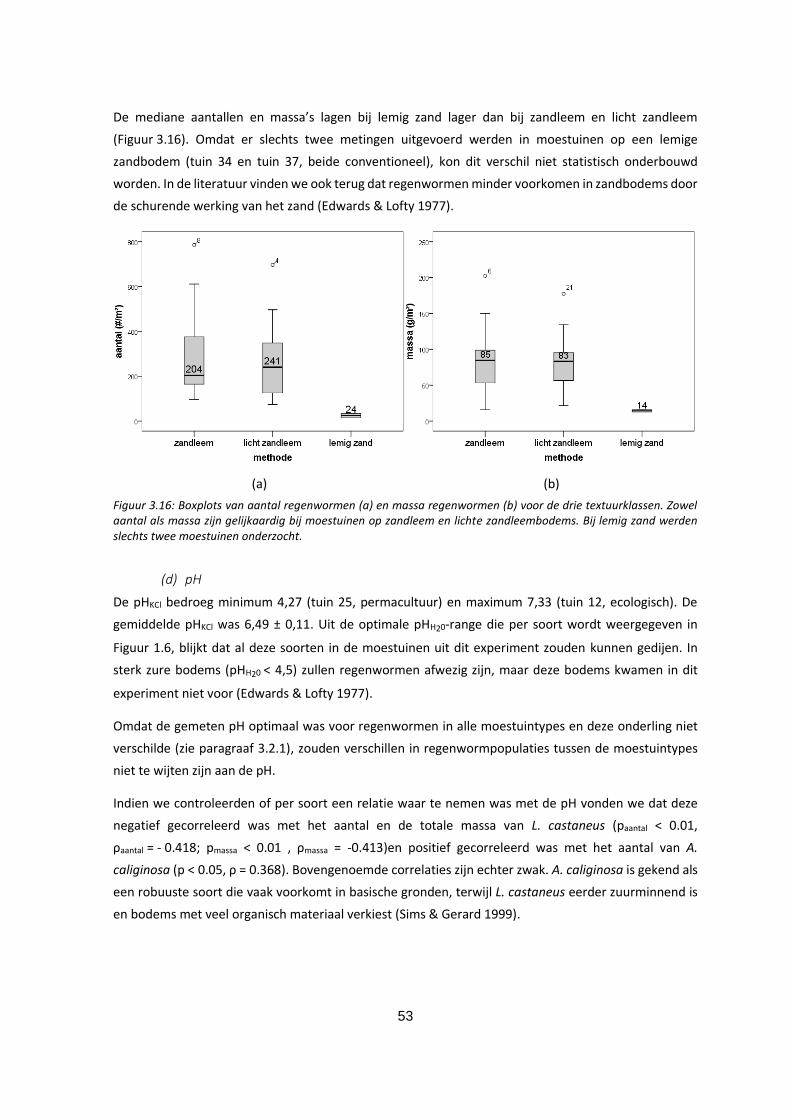
53
De mediane aantallen en massa’s lagen bij lemig zand lager dan bij zandleem en licht zandleem
(Figuur 3.16). Omdat er slechts twee metingen uitgevoerd werden in moestuinen op een lemige
zandbodem (tuin 34 en tuin 37, beide conventioneel), kon dit verschil niet statistisch onderbouwd
worden. In de literatuur vinden we ook terug dat regenwormen minder voorkomen in zandbodems door
de schurende werking van het zand (Edwards & Lofty 1977).
(a)
(b)
Figuur 3.16: Boxplots van aantal regenwormen (a) en massa regenwormen (b) voor de drie textuurklassen. Zowel aantal als massa zijn gelijkaardig bij moestuinen op zandleem en lichte zandleembodems. Bij lemig zand werden slechts twee moestuinen onderzocht.
(d) pH
De pHKCl bedroeg minimum 4,27 (tuin 25, permacultuur) en maximum 7,33 (tuin 12, ecologisch). De
gemiddelde pHKCl was 6,49 ± 0,11. Uit de optimale pHH20-range die per soort wordt weergegeven in
Figuur 1.6, blijkt dat al deze soorten in de moestuinen uit dit experiment zouden kunnen gedijen. In
sterk zure bodems (pHH20 < 4,5) zullen regenwormen afwezig zijn, maar deze bodems kwamen in dit
experiment niet voor (Edwards & Lofty 1977).
Omdat de gemeten pH optimaal was voor regenwormen in alle moestuintypes en deze onderling niet
verschilde (zie paragraaf 3.2.1), zouden verschillen in regenwormpopulaties tussen de moestuintypes
niet te wijten zijn aan de pH.
Indien we controleerden of per soort een relatie waar te nemen was met de pH vonden we dat deze
negatief gecorreleerd was met het aantal en de totale massa van L. castaneus (paantal < 0.01,
ρaantal = - 0.418; pmassa < 0.01 , ρmassa = -0.413)en positief gecorreleerd was met het aantal van A.
caliginosa (p < 0.05, ρ = 0.368). Bovengenoemde correlaties zijn echter zwak. A. caliginosa is gekend als
een robuuste soort die vaak voorkomt in basische gronden, terwijl L. castaneus eerder zuurminnend is
en bodems met veel organisch materiaal verkiest (Sims & Gerard 1999).

54
(e) Voedselbronnen
Het koolstofgehalte is zwak gecorreleerd met het aantal en de gemiddelde totale massa van de
endogeïsche regenwormen (paantal < 0.05, ρaantal = 0.363 en pmassa < 0.001, ρmassa = 0.328). Deze
ecologische groep voedt zich met bodem vermengd met organische stof (verterend organisch materiaal)
die voor een groot deel uit koolstof bestaat. Daarbij is ook een zwakke correlatie met het totaal aantal
regenwormen (p < 0.05, ρ = 0.351), waarschijnlijk te wijten aan de hoge abundantie endogeïsche
regenwormen (65,2%). De matige correlaties zijn in dit onderzoek waarschijnlijk te wijten aan de
combinatie van een nauwe koolstofrange (Figuur 3.9) en het feit dat koolstof waarschijnlijk overal
voldoende aanwezig was. Bovendien zullen populaties vanaf een bepaald voedselaanbod niet verder in
aantal toenemen omdat ze gelimiteerd zijn door andere factoren zoals ruimte.

55
ALGEMENE CONCLUSIE
Naast positieve eigenschappen van moestuinen op vlak van voedselproductie, welzijn en
overstromingsregulatie die terug te vinden zijn in de literatuur, bracht dit onderzoek een eerder negatief
aspect naar voren.
Alle onderzochte nutriënten waren in de meerderheid van de tuinen overmatig aanwezig. Dit is niet
alleen nefast voor de evenwichtige opname ervan, maar kan ook schadelijk zijn voor het milieu zoals het
geval is bij stikstof en fosfor. Beide (NO3--N en PAL) blijken in te hoge concentraties voor te komen in de
Vlaamse moestuinbodems: in respectievelijk 62,1 en 97,3 procent van de moestuinen waren de
concentraties veel hoger dan de streefwaarde. Op die manier kunnen de meeste moestuinen een
negatieve impact uitoefenen op het oppervlakte- en grondwater. Daarnaast blijkt bekalken dikwijls
overmatig te gebeuren: in tien van de vijftien bekalkte moestuinen ligt de pH boven het optimum.
Het Mestdecreet beperkt in land- en tuinbouw overmatige mestgebruik, maar moestuinen, waar (nog)
geen restricties gelden, glippen door de mazen van het net. Het lijkt verantwoord dat in de nabije
toekomst ingezet wordt op sensibilisering over de nadelen van onevenwichtige bemesting of
overbemesting. Een samenwerking tussen organisaties die nauw in contact staan met particulieren,
zoals VELT en Tuinhier, lijkt daarbij aangewezen.
Drie verschillende beheertypes werden onder de loep genomen. Tegen de verwachtingen in, bleek hun
impact op de nutriëntenstatus niet zo verschillend. Dikwijls wordt permacultuur- of ecologisch
getuinierd uit milieuoverweging, maar qua impact op de directe omgeving blijken deze moestuinen in
dit onderzoek niet beter te scoren dan conventionele moestuinen, integendeel. Bij de
permacultuurmoestuinen uit dit onderzoek kwamen opvallend hoge nitraatresiduen naar voren en een
omgekeerde trend deed zich voor op vlak van fosfor. Hoge fosforgehaltes zijn (over alle tuintypes heen)
gecorreleerd met de ouderdom van de moestuin, wat te verklaren is door het immobiele karakter van
dit element. Hoewel het aannemelijk is dat in ecologische en conventionele moestuinen de fosfor-input
via bemesting hoger was, zou ook hun ouderdom (respectievelijk 25 en 26 jaar als mediaan t.o.v. 8 jaar
bij permacultuur) verantwoordelijk kunnen zijn voor de hogere waarden.
Ondanks dat voorzichtigheid geboden is voor conclusies omtrent het organische stofgehalte in de
moestuinbodems van dit onderzoek, indiceren de koolstofgehaltes een voldoende hoge input. De
jaarlijkse aanbreng van koolstofhoudend materiaal zoals compost, ongecomposteerd plantaardig
materiaal, organische meststoffen en dierlijke mest ligt hiervoor aan de bron. Ook in conventionele
moestuinen, waar koolstofarme kunstmest aangewend wordt, wordt het C-gehalte op peil gehouden
door de aanvulling met eerder genoemde materialen.
Per moestuin kwamen gemiddeld 267 regenwormen per m² voor met een biomassa van 78,7 g per m².
Over alle stalen heen werden negentien verschillende soorten regenwormen gecollecteerd, waarvan
één uitheemse soort, Microscolex phosphoreus. Gemiddeld kwamen per moestuin 5,68 soorten

56
voor per m². Regenwormen zijn daarmee in moestuinen opmerkelijk abundant in vergelijking met
akkers, waar zelden meer dan 150 regenwormen per m² en doorgaans lagere soortenrijkdommen
teruggevonden worden.
Doordat regenwormen een positieve invloed uitoefenen op de bodemvruchtbaarheid en de daarmee
samenhangende gewasopbrengsten, en ze tevens zorgen voor een betere water- en luchthuishouding,
is hun talrijke aanwezigheid als positief te beschouwen voor de moestuinen. De fragmentering en
decompositie van organisch materiaal verloopt slechts optimaal als de drie ecologische groepen goed
vertegenwoordigd zijn binnen de regenwormgemeenschap. Dit komt omdat deze elk hun functie
hebben binnen het bodemecosysteem: epigeïsche regenwormen spelen een grote rol bij de
fragmentatie van organisch materiaal, anekische incorporeren het in de bodem en endogeïsche
vermengen het met en verspreiden het in de minerale bodem.
Endogeïsche regenwormen kwamen in aantal procentueel vaker voor (65,2%) dan epigeïsche (17,6%)
en de anekische (17,2%). Dit was in hoofdzaak toe te schrijven aan Aporrectodea caliginosa, een
robuuste soort die toleranter is voor verstoring en om die reden ook in akkers domineert. Als de massa
in acht genomen wordt staan de anekische regenwormen op de eerste plaats (51,0% t.o.v. 42,7%
endogeïsche en 6,3% epigeïsche). De forse anekische soorten domineren in moestuinen niet in aantal
maar zullen er een belangrijke rol spelen.
Tussen de drie moestuinbeheertypes (conventioneel, ecologisch of permacultuur) werden geen
significante verschillen in totale regenwormenbiomassa gedetecteerd, niettegenstaande het totale
aantal regenwormen wel significant hoger was bij permacultuurmoestuinen (369 regenwormen per m2)
in vergelijking met conventionele tuinen (189 regenwormen per m2). Dit kwam omdat vooral de lichte
epigeïsche wormen massaal voorkwamen in de permacultuurmoestuinen. Alhoewel deze hoge
abundantie aan epigeïsche regenwormen er verwacht kon worden, toont de huidige studie voor het
eerst aan dat een permanente laag aan organisch materiaal bij permacultuurbeheer kan leiden tot een
(minstens) vertienvoudiging van het aantal epigeïsche regenwormen (100 regenwormen per m2 t.o.v.
3,7 regenwormen per m2 bij ecologisch en 11,1 regenwormen per m2 bij conventioneel beheer).
Omdat ook anekische regenwormen organisch materiaal aan de oppervlakte halen, en ze door hun
permanent gangenstelsels geacht worden bodemverstoring slecht te verdragen, hadden we deze groep
ook abundanter verwacht in permacultuurmoestuinen. Dit bleek echter niet het geval. De endogeïsche
soorten waren zowel qua aantal als biomassa gelijkwaardig vertegenwoordigd in de drie moestuintypes.
Aangezien ondergronds voedsel overal rijkelijk aanwezig was en de bodemchemische eigenschappen
tussen de types niet verschillen, is dit een verklaarbaar resultaat.
De hypothese dat permacultuurmoestuinen een hoger aantal soorten bevatten dan de andere
moestuintypes blijkt te kloppen. Evenwel dient opgemerkt te worden dat dit in dit onderzoek meer te
wijten lijkt aan de permanente organische bodembedekking dan aan de verminderde bodemverstoring.
Naast de grotere soortenrijkdom blijken de populaties ook evenwichtiger verdeeld, wat een indicatie

57
kan zijn dat in permacultuurtuinen meerdere soorten harmonisch kunnen samenleven dankzij meer
diverse habitats en voedselvoorraden.
Alles bij elkaar beschouwd kunnen we stellen dat (1) overbemesting een actueel probleem is in elk type
moestuin en (2) regenwormen over het algemeen abundant zijn in moestuinen maar wel degelijk
beïnvloed worden door beheer.
Deze conclusie biedt perspectieven. Enerzijds wordt te veel bemest en anderzijds zijn er veel
regenwormen die, door hun positieve invloed op de bodem en de nutriëntencyclus, meststoffen deels
zouden kunnen vervangen. Dit zou de zowel de directe kosten (voor de tuinier) en de indirecte kosten
(op het milieu) kunnen verlagen. Met de groter wordende aandacht voor extensievere
productiesystemen, stadslandbouw en duurzame voedselproductie in een korte keten, is de vaststelling
dat een regenworm-vriendelijk beheer bestaat een waarneming die toegejuicht kan worden.

58
REFERENTIELIJST
Adejuyigbe CO, Tian G, Adeoye GO (2006) Microcosmic study of soil microarthropod and earthworm interaction in litter decomposition and nutrient turnover. Nutr Cycl Agroecosystems 75:47–55. doi: 10.1007/s10705-006-9010-5
Adevi AA, Mårtensson F (2013) Stress rehabilitation through garden therapy: The garden as a place in the recovery from stress. Urban For Urban Green 12:230–237. doi: 10.1016/j.ufug.2013.01.007
Aguilar-Støen M, Moe SR, Camargo-Ricalde SL (2009) Home gardens sustain crop diversity and improve farm resilience in Candelaria Loxicha, Oaxaca, Mexico. Hum Ecol 37:55–77. doi: 10.1007/s10745-008-9197-y
Alaimo K, Packnett E, Miles RA, Kruger DJ (2008) Fruit and Vegetable Intake among Urban Community Gardeners. J Nutr Educ Behav 40:94–101. doi: 10.1016/j.jneb.2006.12.003
Avetisyan M, Hertel T, Sampson G (2014) Is Local Food More Environmentally Friendly? The GHG Emissions Impacts of Consuming Imported versus Domestically Produced Food. Environ Resour Econ 58:415–462. doi: http://dx.doi.org/10.1108/17506200710779521
Baker BP, Benbrook CM, Groth E, Benbrook KL (2002) Pesticide residues in conventional , integrated pest management ( IPM ) -grown and organic foods : insights from three US data sets Pesticide residues in conventional , integrated pest management ( IPM ) -grown and organic foods : insights. Food Addit Contam 19:427–446. doi: 10.1080/0265203011011379
Bakkes J, Bolt K, Braeuer I, et al (2008) The Cost of Policy Inaction. The case of not meeting the 2010 biodiversity target. Environment 44:1–16.
Bartlett MD, Harris J a., James IT, Ritz K (2006) Inefficiency of mustard extraction technique for assessing size and structure of earthworm communities in UK pasture. Soil Biol Biochem 38:2990–2992. doi: 10.1016/j.soilbio.2006.04.036
Beleidsdomein Leefmilieu Natuur en Energie (2014) Tevredenheid over milieugerelateerde leefkwaliteit.
Bertrand M, Barot S, Blouin M, et al (2015) Earthworm services for cropping systems. A review. Agron Sustain Dev 553–567. doi: 10.1007/s13593-014-0269-7
Beumer C, Martens P (2014) Biodiversity in my (back)yard: towards a framework for citizen engagement in exploring biodiversity and ecosystem services in residential gardens. Sustain Sci 10:87–100. doi: 10.1007/s11625-014-0270-8
Bhadauria T, Saxena KG (2009) Role of Earthworms in Soil Fertility Maintenance through the Production of Biogenic Structures. In: Karmegam N (ed) Status, Trends, and Advances in Earthworm Research and Vermitechnology. Hindawi Publishing Corporation,
Blouin M, Hodson ME, Aranda Delgado E, et al (2013) A review of earthworm impact on soil function and ecosystem services. Eur J Soil Sci 64:161–182.

59
Bomans K, Dewaelheyns V, Heylen K, Gulinck H (2011) The spatial importance of gardens: a regional perspective. In: Dewaelheyns V, Bomans K, Gul (eds) The Powerful Garden - Emerging views on the garden complex. Garant, Antwerpen - Apeldoorn, pp 69–84
Bouché MB (1972) Lombriciens de France : écologie et systématique. Institut national de la recherche agronomique, Paris
Bouché MB, Gardner RH (1984) Earthworm functions VIII. - Population estimation techniques. Pedobiologia Int J soil Biol 21:37–63.
Cameron RWF, Blanuša T, Taylor JE, et al (2012) The domestic garden - Its contribution to urban green infrastructure. Urban For Urban Green 11:129–137. doi: 10.1016/j.ufug.2012.01.002
Chan KY (2000) An overview of some tillage impacts on earthworm population abundance and diversity - Implications for functioning in soils. Soil Tillage Res 57:179–191. doi: 10.1016/S0167-1987(00)00173-2
Chen S (2012) Civic Agriculture: Towards a Local Food Web for Sustainable Urban Development. APCBEE Procedia 1:169–176. doi: 10.1016/j.apcbee.2012.03.027
Church A, Mitchell R, Ravenscroft N, Stapleton LM (2015) “Growing your own”: A multi-level modelling approach to understanding personal food growing trends and motivations in Europe. Ecol Econ 110:71–80. doi: 10.1016/j.ecolecon.2014.12.002
Clapperton MJ, Baker GH, Fox CA (2007) Earthworms. Soil Sampling and Methods of Analysis, Second Edition. CRC Press, p 1264
Clark HF, Brabander DJ, Erdil RM (2006) Sources, sinks, and exposure pathways of lead in urban garden soil. J Environ Qual 35:2066–2074. doi: 10.2134/jeq2005.0464
Clayton S (2007) Domesticated nature: Motivations for gardening and perceptions of environmental impact. J Environ Psychol 27:215–224. doi: 10.1016/j.jenvp.2007.06.001
CoDyre M, Fraser EDG, Landman K (2015) How does your garden grow? An empirical evaluation of the costs and potential of urban gardening. Urban For Urban Green 14:72–79. doi: 10.1016/j.ufug.2014.11.001
Čoja T, Zehetner K, Bruckner A, et al (2008) Efficacy and side effects of five sampling methods for soil earthworms (Annelida, Lumbricidae). Ecotoxicol Environ Saf 71:552–565. doi: 10.1016/j.ecoenv.2007.08.002
Crittenden SJ, Eswaramurthy T, de Goede RGM, et al (2014) Effect of tillage on earthworms over short- and medium-term in conventional and organic farming. Appl Soil Ecol 1–9. doi: 10.1016/j.apsoil.2014.03.001
Curry JP (1998) Factors Affecting Earthworm Abundance in soils. In: Edwards CA (ed) Earthworm Ecology. CRC Press, Florida, pp 37 – 64
Curry JP, Byrne D, Schmidt O (2002) Intensive cultivation can drastically reduce earthworm populations in arable land. Eur J Soil Biol 38:127–130. doi: 10.1016/S1164-5563(02)01132-9

60
De Decker P (2011) A Garden of Eden? - The promotion of a single-family house with a garden in Belgium before the Second World War. In: Dewaelheyns V, Bomans K, Gulinck H (eds) The Powerful Garden - Emerging views on the garden complex. Garant, Antwerpen - Apeldoorn, pp 28–50
De Meyer A, Tirry D, Gulinck H, Van Orshoven J (2011) Ondersteunend Onderzoek. Actualisatie MIRA Achtergronddocument Bodem. Thema Bodemafdichting. Eindrapport.
De Neve S, Van Den Bossche A, Sleutel S, Hofman G (2006) Soil Nutrient Status of Organic Farms in Flanders: An Overview and a Comparison with the Conventional Situation. Biol Agric Hortic 24:217–235. doi: 10.1080/01448765.2006.9755023
De Schrijver A, De Frenne P, Staelens J, et al (2012) Tree species traits cause divergence in soil acidification during four decades of postagricultural forest development. Glob Chang Biol 18:1127–1140. doi: 10.1111/j.1365-2486.2011.02572.x
De Vries S, Verheij RA, Groenewegen PP, Spreeuwenberg P (2003) Natural environments...healthy environments? An exploratory analysis of the relationship between greenspace and health. Environ Plan A 35:1717–1731. doi: 10.1068/a35111
De Wispelaere A (2014) Hoe gezond is eigen kweek?
Dewaelheyns V, Bomans K, Gulinck H (eds) (2011) The Powerful Garden - Emerging views on the garden complex.
Dewaelheyns V, Gulinck H (2008) Inputs en outputs in privétuinen. Studie uitgevoerd in opdracht van de Vlaamse Milieumaatschappij, MIRA, MIRA/2008/02 , Afdeling Bos, Natuur en Landschap, K.U.Leuven.
Dewaelheyns V, Rogge E, Gulinck H (2014) Putting domestic gardens on the agenda using empirical spatial data: The case of Flanders. Appl Geogr 50:132–143. doi: 10.1016/j.apgeog.2014.02.011
Dorais M (2007) Organic production of vegetables: State of the art and challenges. Can J Plant Sci 87:1055–1066. doi: 10.4141/CJPS07160
Doube BM, Schmidt O, Killham K, Correll R (1997) Influence of mineral soil on the palatability of organic matter for lumbricid earthworms: A simple food preference study. Soil Biol Biochem 29:569–575. doi: 10.1016/S0038-0717(96)00032-6
Dumortier M, De Bruyn L, Hens M, et al (2007) Natuurrapport 2007. Toestand van de natuur in Vlaanderen: cijfers voor het beleid. Mededelingen van het Instituut voor Natuur- en Bosonderzoek nr. 4, Brussel.
Edwards CA, Lofty JR (1977) Biology of earthworms, 2nd edn. Chapman and Hall Ltd, London
Edwards WM, Shipitalo MJ (1998) Consequences of Earthworms in Agricultural Soils: Aggregation and Porosity. In: Edwards CA (ed) Earthworm Ecology. CRC Press, Florida, pp 147–162
Eichinger E, Bruckner A, Stemmer M (2007) Earthworm expulsion by formalin has severe and lasting side effects on soil biota and plants. Ecotoxicol Environ Saf 67:260–266. doi: 10.1016/j.ecoenv.2006.03.015
Eisenhauer N, Marhan S, Scheu S (2008) Assessment of anecic behavior in selected earthworm species: Effects on wheat seed burial, seedling establishment, wheat growth and litter incorporation. Appl Soil Ecol 38:79–82. doi: 10.1016/j.apsoil.2007.07.002

61
Elsen A, Deckers S, Vandendriessche H (2011) Soil fertility in domestic and public gardens in Flanders: the state of the art. In: Gulinck H, Dewaelheyns V, Bomans K (eds) The Powerful Garden - Emerging views on the garden complex. Garant, Antwerpen - Apeldoorn, pp 165–178
EMIS (2010a) Bodem – Bepaling van nitraatstikstof.
EMIS (2010b) Bodem – Schijnbaar soortelijk gewicht (dichtheid).
Emmerling C (2001) Response of earthworm communities to different types of soil tillage. Appl Soil Ecol 17:91–96. doi: 10.1016/S0929-1393(00)00132-3
Emmerling C, Felten D (2009) Earthworm burrowing behaviour in 2D terraria with single- and multi-species assemblages. Biol Fertil Soils 45:789–797.
Ernst G, Emmerling C (2009) Impact of five different tillage systems on soil organic carbon content and the density, biomass, and community composition of earthworms after a ten year period. Eur J Soil Biol 45:247–251. doi: 10.1016/j.ejsobi.2009.02.002
Europese Commissie (2011) Plan voor de bescherming van soorten en habitats. http://ec.europa.eu/news/environment/110503_nl.htm. Accessed 15 Apr 2015
Ferguson RS, Lovell ST (2014) Permaculture for agroecology: Design, movement, practice, and worldview. A review. Agron Sustain Dev 34:251–274. doi: 10.1007/s13593-013-0181-6
Galluzzi G, Eyzaguirre P, Negri V (2010) Home gardens: neglected hotspots of agro-biodiversity and cultural diversity. Biodivers Conserv 19:3635–3654. doi: 10.1007/s10531-010-9919-5
Gaston KJ, Smith RM, Thompson K, Warren PH (2005) Urban domestic gardens (II): Experimental tests of methods for increasing biodiversity. Biodivers Conserv 14:395–413. doi: 10.1007/s10531-004-6066-x
Goddard M a., Dougill AJ, Benton TG (2010) Scaling up from gardens: biodiversity conservation in urban environments. Trends Ecol Evol 25:90–98. doi: 10.1016/j.tree.2009.07.016
Guitart D, Pickering C, Byrne J (2012) Past results and future directions in urban community gardens research. Urban For Urban Green 11:364–373. doi: 10.1016/j.ufug.2012.06.007
Guitart DA, Byrne J, Pickering CM (2011) Greener growing: assessing the influence of gardening practices on the ecological viability of community gardens in South East Queensland, Australia. Gueensland
Gunn A (1992) the Use of Mustard To Estimate Earthworm Populations. Pedobiologia (Jena) 36:65–67.
Hendrix PF, Callaham M a., Drake JM, et al (2008) Pandora’s Box Contained Bait: The Global Problem of Introduced Earthworms *. Annu Rev Ecol Evol Syst 39:593–613. doi: 10.1146/annurev.ecolsys.39.110707.173426
Henneron L, Bernard L, Hedde M, et al (2014) Fourteen years of evidence for positive effects of conservation agriculture and organic farming on soil life. Agron Sustain Dev 169–181. doi: 10.1007/s13593-014-0215-8

62
Hermy M, Claessens B (2011) Gardens and plant biodiversity. In: Dewaelheyns V, Bomans K, Gulinck H (eds) The Powerful Garden - Emerging views on the garden complex. Garant, Antwerpen - Apeldoorn, pp 153–164
Iannone B V., Umek LG, Wise DH, Heneghan L (2012) A Simple, Safe, and Effective Sampling Technique for Investigating Earthworm Communities in Woodland Soils: Implications for Citizen Science. Nat Areas J 32:283–292. doi: 10.3375/043.032.0305
IUCN Why is biodiversity in crisis? http://www.iucn.org/what/biodiversity/about/biodiversity_crisis/. Accessed 15 May 2015
Jänsch S, Steffens L, Höfer H, et al (2013) State of knowledge of earthworm communities in German soils as a basis for biological soil quality assessment. Soil Org 85:215–233.
Jennings V, Gaither C (2015) Approaching Environmental Health Disparities and Green Spaces: An Ecosystem Services Perspective. Int J Environ Res Public Health 12:1952–1968. doi: 10.3390/ijerph120201952
Jones A (2006) A life cycle analysis of UK supermarket imported green beans from Kenya. Fresh insights 4. International Institute for Environment and Development, London
Jones CG, Lawton JH, Shachak M (1994) Organisms as Ecosystem Engineers. Oikos 69:373–386.
Jost L (2006) Entropy and diversity. Oikos 113:363–375. doi: 10.1111/j.2006.0030-1299.14714.x
Kaplan R, Kaplan S (1990) Restorative experience: the healing power of nearby nature. In: Francis M, Hester RT (eds) The Meaning of Gardens: Idea, Place and Action. The MIT Press, Cambridge, pp 238–243
Kortright R, Wakefield S (2011) Edible backyards: A qualitative study of household food growing and its contributions to food security. Agric Human Values 28:39–53. doi: 10.1007/s10460-009-9254-1
Kotcon JB (2011) Population Dynamics of Earthworms in Organic Farming Systems. In: Ayten K (ed) The Impact of Cultivation Techniques on Earthworm Populations. Springer Science & Business Media, Heidelberg, pp 299 – 310
Lee KE (1985) Earthworms: their ecology and relationships with soils and land use. Academic Press, Sydney
Leroy BLM, Schmidt O, Van den Bossche A, et al (2008) Earthworm population dynamics as influenced by the quality of exogenous organic matter. Pedobiologia (Jena) 52:139–150. doi: 10.1016/j.pedobi.2008.07.001
LeVasseur TJ (2014) Permaculture. In: Kaplan DM, Thompson PB (eds) Encyclopedia of Food and Agricultural Ethics. Springer Netherlands, Dordrecht, pp 1471–1473
Lindemann-Matthies P, Marty T (2013) Does ecological gardening increase species richness and aesthetic quality of a garden? Biol Conserv 159:37–44. doi: 10.1016/j.biocon.2012.12.011
Loram A, Tratalos J, Warren PH, Gaston KJ (2007) Urban domestic gardens (X): The extent & structure of the resource in five major cities. Landsc Ecol 22:601–615. doi: 10.1007/s10980-006-9051-9
Lowe CN, Butt KR (2002a) Growth of hatchling earthworms in the presence of adults: Interactions in laboratory culture. Biol Fertil Soils 35:204–209. doi: 10.1007/s00374-002-0471-7

63
Lowe CN, Butt KR (2002b) Influence of organic matter on earthworm production and behaviour: A laboratory-based approach with applications for soil restoration. Eur J Soil Biol 38:173–176. doi: 10.1016/S1164-5563(02)01141-X
Lubbers IM, Brussaard L, Otten W, Van Groenigen JW (2011) Earthworm-induced N mineralization in fertilized grassland increases both N2O emission and crop-N uptake. Eur J Soil Sci 62:152–161. doi: 10.1111/j.1365-2389.2010.01313.x
Lubbers IM, van Groenigen KJ, Fonte SJ, et al (2013) Greenhouse-gas emissions from soils increased by earthworms. Nat Clim Chang 3:187–194. doi: 10.1038/nclimate1692
Ma W-C, Brussaard L, de Ridder JA (1990) Long-term effects of nitrogenous fertilizers on grassland earthworms (Oligochaeta: Lumbricidae): Their relation to soil acidification. Agric Ecosyst Environ 30:71–80. doi: 10.1016/0167-8809(90)90184-F
Maas J, Verheij RA, de Vries S, et al (2009) Morbidity is related to a green living environment. J Epidemiol Community Health 63:967–973. doi: 10.1136/jech.2008.079038
Maes S (2012) Bodemkundige Dienst van België - Studienamiddag “Meten om te sturen.”
Maes S, Elsen A, Tits M, et al (2012) Wegwijs in de bodemvruchtbaarheid van de Belgische akkerbouw- en weilandpercelen (2008-2011). Bodemkundige Dienst van België
Marhan S, Auber J, Poll C (2015) Additive effects of earthworms, nitrogen-rich litter and elevated soil temperature on N2O emission and nitrate leaching from an arable soil. Appl Soil Ecol 86:55–61. doi: 10.1016/j.apsoil.2014.10.006
Millennium Ecosystem Assessment (2004) Living Beyond Our Means: Natural Assets and Human Well-being.
Mohr D, Topp W (2005) Hazel improves soil quality of sloping oak stands in a German low mountain range. Ann For Sci 62:23–29.
Mollison B, Holmgren D (1978) Permaculture one: a perennial agricultural system for human settlements. Tagari, Tyalgum
Morton LW, Bitto EA, Oakland MJ, Sand M (2008) Accessing food resources: Rural and urban patterns of giving and getting food. Agric Human Values 25:107–119. doi: 10.1007/s10460-007-9095-8
Natuurpunt (2006) Het spook van Hertberg is ontmaskerd.
NBN (2011) EN ISO 23611-1. http://shop.nbn.be/Search/SearchResults.aspx?a=NBN+EN+ISO+2361&UIc=nl#details. Accessed 28 Feb 2015
OVAM (2010) Evaluerend onderzoek naar effectiviteit van de uitvoering van het ruimtelijk beleid in Vlaanderen.
OVAM (2012) Evaluatieonderzoek materialenkringloop gft- en groenafval.
OVAM (2006) Preventie-evaluatieonderzoek voor GFT- en groenafval, KGA en AEEA. Mechelen

64
Overloop S, Tits M, Bries J, et al (2010) Milieurapport Vlaanderen MIRA, Achtergronddocument 2010 Bodem.
Peigné J, Cannavaciuolo M, Gautronneau Y, et al (2009) Earthworm populations under different tillage systems in organic farming. Soil Tillage Res 104:207–214. doi: 10.1016/j.still.2009.02.011
Pelosi C, Barot S, Capowiez Y, et al (2014) Pesticides and earthworms. A review. Agron Sustain Dev 34:199–228. doi: 10.1007/s13593-013-0151-z
Pelosi C, Bertrand M, Roger-Estrade J (2009) Earthworm community in conventional, organic and direct seeding with living mulch cropping systems. Agron Sustain Dev 29:287–295. doi: 10.1051/agro/2008069
Pelosi C, Bertrand M, Thénard J, Mougin C (2015) Earthworms in a 15 years agricultural trial. Appl Soil Ecol 88:1–8. doi: 10.1016/j.apsoil.2014.12.004
Pfiffner L, Mader P (1997) Effects of biodynamic, organic and conventional production systems on earthworm populations. 15:3–10.
Reubens B, D’Haene K, D’Hose T, et al (2010) Bodemkwaliteit en landbouw: een literatuurstudie. Act 1 van het Interregproject BodemBreed 203.
Roe J, Thompson CW (2011) The “green edge” in urban streets: impact of urban gardens and street trees on health and well-being. In: Dewaelheyns V, Bomans K, Gulinck H (eds) The Powerful Garden - Emerging views on the garden complex. Garant, Antwerpen - Apeldoorn, pp 198–220
Ropars K (2013) The Effects of Horticulture Therapy on the Positive Affect of Older Adults with Dementia. Temple University
Rothwell A, Chaney K, Haydock P (2011) The Impact of Cultivation Techniques on Earthworm Populations. In: Ayten K (ed) Biology of Earthworms. Springer Science & Business Media, Heidelberg, pp 159 – 172
Rudd H, Vala J, Schaefer V (2002) Importance of backyard habitat in a comprehensive biodiversity conservation strategy: A connectivity analysis of urban green spaces. Restor Ecol 10:368–375. doi: 10.1046/j.1526-100X.2002.02041.x
Sæbø A, Ferrini F (2006) The use of compost in urban green areas - A review for practical application. Urban For Urban Green 4:159–169. doi: 10.1016/j.ufug.2006.01.003
Satterthwaite D, McGranahan G, Tacoli C (2010) Urbanization and its implications for food and farming. Philos Trans R Soc Lond B Biol Sci 365:2809–2820. doi: 10.1098/rstb.2010.0136
Säumel I, Kotsyuk I, Hölscher M, et al (2012) How healthy is urban horticulture in high traffic areas? Trace metal concentrations in vegetable crops from plantings within inner city neighbourhoods in Berlin, Germany. Environ Pollut 165:124–132. doi: 10.1016/j.envpol.2012.02.019
Saunders DA, Hobbs RJ, Margules CR, et al (1992) Biological consequences of ecosystem fragmentation: A review. Biol Conserv 59:77. doi: 10.1016/0006-3207(92)90725-3
Scheu S (2003) Effects of earthworms on plant growth: patterns and perspectivesThe 7th international symposium on earthworm ecology · Cardiff · Wales · 2002. Pedobiologia (Jena) 47:846–856. doi: 10.1016/S0031-4056(04)70279-6

65
Schoumans OF, Willems J, van Duinhoven G (2008) 30 vragen en antwoorden over fosfaat in relatie tot landbouw en milieu. Wageningen, Alterra, 53 blz.
Sherlock E (2012) Key to the earthworms of the UK and Ireland. FSC & Natural History Museum, London
Shinn NJ, Bing-Canar J, Cailas M, et al (2000) Determination of spatial continuity of soil lead levels in an urban residential neighborhood. Environ Res 82:46–52. doi: 10.1006/enrs.1999.4004
Sims RW, Gerard BM (1999) Earthworms: keys and notes for the identification and study of the species. Field Studies Council, Shrewsbury
Smith RG, McSwiney CP, Grandy a. S, et al (2008) Diversity and abundance of earthworms across an agricultural land-use intensity gradient. Soil Tillage Res 100:83–88. doi: 10.1016/j.still.2008.04.009
Soane BD, Ball BC, Arvidsson J, et al (2012) No-till in northern, western and south-western Europe: A review of problems and opportunities for crop production and the environment. Soil Tillage Res 118:66–87. doi: 10.1016/j.still.2011.10.015
Steinberg T (2006) American Green. Norton, New York
Stuart SM (2005) Lifting spirits: Creating gardens in California domestic violence shelters. In: Bartlett PF (ed) Urban place: Reconnecting with the natural world. MIT press, Cambridge, pp 61–88
Suárez ER, Pelletier DM, Fahey TJ, et al (2004) Effects of exotic earthworms on soil phosphorus cycling in two broadleaf temperate forests. Ecosystems 7:28–44. doi: 10.1007/s10021-003-0128-x
Taylor JR, Lovell ST (2014) Urban home food gardens in the Global North: Research traditions and future directions. Agric Human Values 31:285–305. doi: 10.1007/s10460-013-9475-1
Thompson K, Austin KC, Smith RM, et al (2003) Urban domestic gardens (I): Putting small-scale plant diversity in context. J Veg Sci 14:71–78. doi: 10.1111/j.1654-1103.2003.tb02129.x
Ulrich RS, Simons RF, Losito BD, et al (1991) Stress recovery during exposure to natural and urban environments. J Environ Psychol 11:201–230. doi: 10.1016/S0272-4944(05)80184-7
Uvarov A V. (2009) Inter- and intraspecific interactions in lumbricid earthworms: Their role for earthworm performance and ecosystem functioning. Pedobiologia (Jena) 53:1–27. doi: 10.1016/j.pedobi.2009.05.001
Valckx J, Govers G, Hermy M, Muys B (2009) Dieper graven naar het belang van regenwormen in duurzaam akkerbeheer - een toolkit voor ecologische erosiecontrole. Departement Aard- en omgevingswetenschappen, K.U.Leuven, 34 p.
Van Delm A, Gulinck H (2011) Classification and quantification of green in the expanding urban and semi-urban complex: Application of detailed field data and ikonos-imagery. Ecol Indic 11:52–60. doi: 10.1016/j.ecolind.2009.06.004
Van Groeningen JW, Lubbers IM, Vos HMJ, et al (2014) Earthworms increase plant production: a meta-analysis. http://www.nature.com/srep/2014/140915/srep06365/pdf/srep06365.pdf. Accessed 28 Feb 2015

66
VELT vzw (2014) Handboek Ecologisch Tuinieren. Druk in de Weer (Graphius), Gent
VLM (2014) Drempelwaarden vanaf 2014. http://www.vlm.be/SiteCollectionDocuments/Mestbank/Algemeen/Drempelwaardeninfocusgebiedenvanaf 2014.pdf. Accessed 3 May 2015
VLM (2015) Begeleidende maatregelen 2015 bij een te hoog nitraatresidu.
Waage GH (1935) No Title. Natuurhistorisch Maandbl 24:48.
Währborg P, Petersson IF, Grahn P (2014) Nature-assisted rehabilitation for reactions to severe stress and/or depression in a rehabilitation garden: long-term follow-up including comparisons with a matched population-based reference cohort. J Rehabil Med 46:271–6. doi: 10.2340/16501977-1259
Whalen JK, Parmelee RW, Edwards CA (1998) Population dynamics of earthworm communities in corn agroecosystems receiving organic or inorganic fertilizer amendments. Biol Fertil Soils 27:400–407. doi: 10.1007/s003740050450
Xiong TT, Leveque T, Austruy A, et al (2014) Foliar uptake and metal(loid) bioaccessibility in vegetables exposed to particulate matter. Environ Geochem Health 897–909. doi: 10.1007/s10653-014-9607-6
Xuereb M (2005) Food miles: Environmental implications of food imports to Waterloo Region. Waterloo, Reg Waterloo Public Heal 42:3508–13.
Zaborski ER (2003) Allyl isothiocyanate: An alternative chemical expellant for sampling earthworms. Appl Soil Ecol 22:87–95. doi: 10.1016/S0929-1393(02)00106-3
Załęcka A, Bügel S, Paoletti F, et al (2014) The influence of organic production on food quality - research findings, gaps and future challenges. J Sci Food Agric 1–5. doi: 10.1002/jsfa.6578
Zmora-Nahum S, Hadar Y, Chen Y (2007) Physico-chemical properties of commercial composts varying in their source materials and country of origin. Soil Biol Biochem 39:1263–1276. doi: 10.1016/j.soilbio.2006.12.017
Zmyslony J, Gagnon D (1998) Residential management of urban front-yard landscape: A random process? Landsc Urban Plan 40:295–307. doi: 10.1016/S0169-2046(97)00090-X

67
APPENDIX
Tabel A.1: Potentieel voorkomende gevaarlijke stoffen in de moestuin met bijhorende gezondheidsrisico's www.gezonduiteigengrond.be).
Polyaromatische koolwaterstoffen (PAK’s) komen in de lucht terecht via uitlaatgassen en de verwarming van gebouwen. In regio’s met veel huizen en verkeer zitten er meer PAK’s in de lucht. Die komen ook op je groenten terecht. Door ze goed te wassen, kun je de meeste PAK’s verwijderen. Uit de bodem nemen groenten niet zoveel PAK’s op.
Dioxines en pcb’s komen in de lucht terecht tijdens verbrandingsprocessen: tuinvuurtjes, hout- en steenkoolkachels … Door je groenten goed te wassen en te schillen, kun je deze stoffen grotendeels verwijderen. Uit de bodem nemen groenten niet zoveel dioxines en pcb’s op. Een uitzondering zijn groenten uit de komkommerfamilie: komkommer, courgette, meloen, pompoen …
Persistente pesticiden zoals DDT mogen vandaag niet meer gebruikt worden. Hier en daar zitten ze wel nog in de grond, maar groenten nemen die stoffen maar beperkt op. Een uitzondering zijn groenten uit de komkommerfamilie: komkommer, courgette, meloen, pompoen … Zij nemen wel makkelijk pesticiden uit de grond op.
Minerale oliën kun je in de bodem aantreffen in de buurt van een tankstation (lekkende brandstoftank), een garage, een industrie- of landbouwbedrijf, een parkeerterrein voor vrachtwagens of bij particulieren met een lekkende stookolietank. Groenten nemen olie maar beperkt uit de bodem op. De vervuiling moet echt vlakbij zijn om een effect te hebben.
Zware metalen zoals cadmium worden vlot door planten opgenomen en kunnen een gevaar zijn voor de gezondheid. Ze komen vooral in de bodem terecht tijdens de productie en het gebruik van metaalhoudende producten. Na een vervuiling blijven ze lange tijd in de bodem zitten. Ook een vervuiling uit het verleden blijft dus een risico.
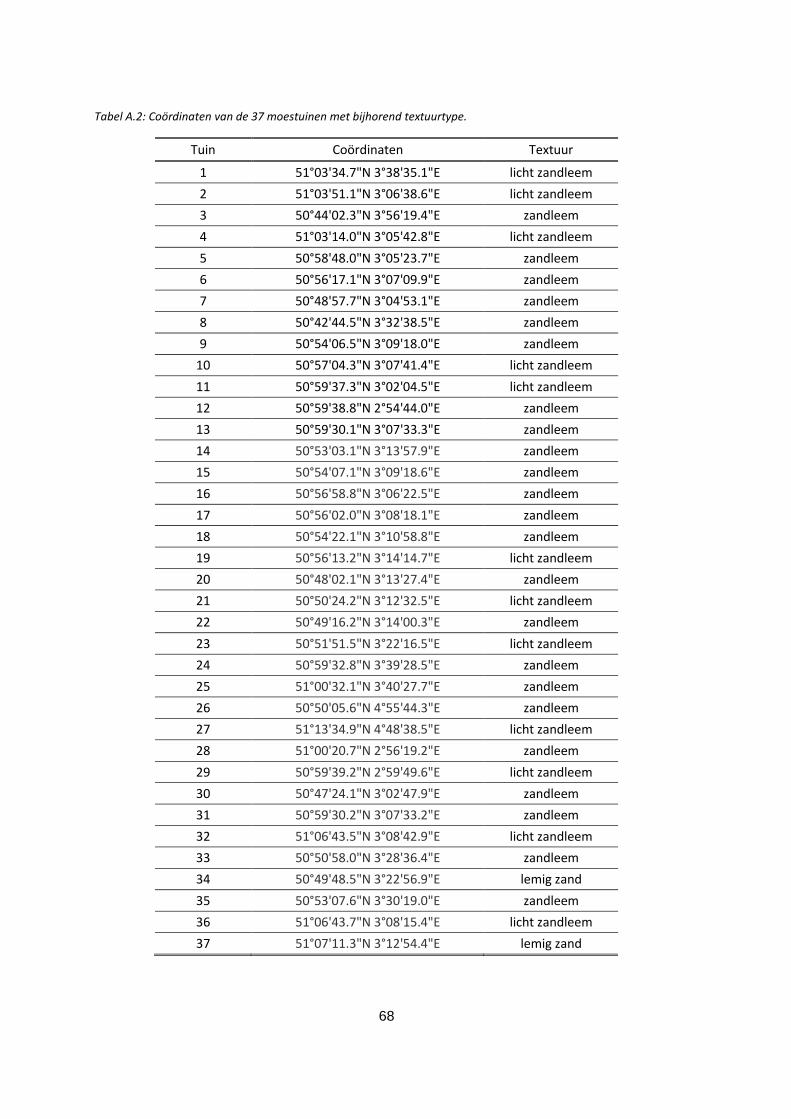
68
Tabel A.2: Coördinaten van de 37 moestuinen met bijhorend textuurtype.
Tuin Coördinaten Textuur
1 51°03'34.7"N 3°38'35.1"E licht zandleem
2 51°03'51.1"N 3°06'38.6"E licht zandleem
3 50°44'02.3"N 3°56'19.4"E zandleem
4 51°03'14.0"N 3°05'42.8"E licht zandleem
5 50°58'48.0"N 3°05'23.7"E zandleem
6 50°56'17.1"N 3°07'09.9"E zandleem
7 50°48'57.7"N 3°04'53.1"E zandleem
8 50°42'44.5"N 3°32'38.5"E zandleem
9 50°54'06.5"N 3°09'18.0"E zandleem
10 50°57'04.3"N 3°07'41.4"E licht zandleem
11 50°59'37.3"N 3°02'04.5"E licht zandleem
12 50°59'38.8"N 2°54'44.0"E zandleem
13 50°59'30.1"N 3°07'33.3"E zandleem
14 50°53'03.1"N 3°13'57.9"E zandleem
15 50°54'07.1"N 3°09'18.6"E zandleem
16 50°56'58.8"N 3°06'22.5"E zandleem
17 50°56'02.0"N 3°08'18.1"E zandleem
18 50°54'22.1"N 3°10'58.8"E zandleem
19 50°56'13.2"N 3°14'14.7"E licht zandleem
20 50°48'02.1"N 3°13'27.4"E zandleem
21 50°50'24.2"N 3°12'32.5"E licht zandleem
22 50°49'16.2"N 3°14'00.3"E zandleem
23 50°51'51.5"N 3°22'16.5"E licht zandleem
24 50°59'32.8"N 3°39'28.5"E zandleem
25 51°00'32.1"N 3°40'27.7"E zandleem
26 50°50'05.6"N 4°55'44.3"E zandleem
27 51°13'34.9"N 4°48'38.5"E licht zandleem
28 51°00'20.7"N 2°56'19.2"E zandleem
29 50°59'39.2"N 2°59'49.6"E licht zandleem
30 50°47'24.1"N 3°02'47.9"E zandleem
31 50°59'30.2"N 3°07'33.2"E zandleem
32 51°06'43.5"N 3°08'42.9"E licht zandleem
33 50°50'58.0"N 3°28'36.4"E zandleem
34 50°49'48.5"N 3°22'56.9"E lemig zand
35 50°53'07.6"N 3°30'19.0"E zandleem
36 51°06'43.7"N 3°08'15.4"E licht zandleem
37 51°07'11.3"N 3°12'54.4"E lemig zand

69
Tabel A.3: Verschillende gewasgroepen en er in voorkomende soorten.
1 bladgewassen slasoorten, warmoes, spinazie, prei, selder, kervel, peterselie,
knolselder, waterkers, snijbiet, postelein, mosterdzaad
2 koolgewassen koolsoorten, broccoli, raap, radijs, spruiten
3 vruchtgewassen pompoen, courgette, komkommer, tomaat, peper, aubergine, paprika,
suikermais
4 wortelgewassen wortel, pastinaak, witloof, rode biet, ui, sjalot, look, knoflook,
stengelui, venkel
5 aardappel aardappel
6 peulgewassen erwt, boon, sojaboon, facelia
7 aardbei aardbei
8 doorlevende
gewassen asperge, rabarber, aardpeer, bonenkruid
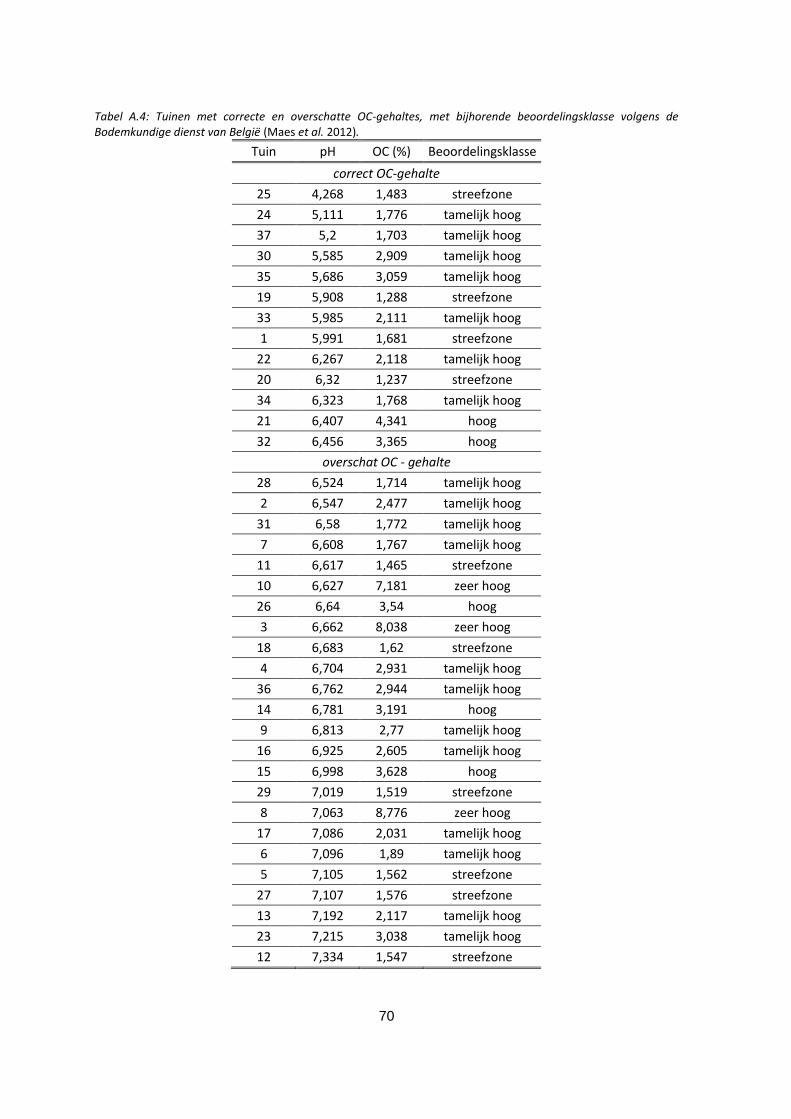
70
Tabel A.4: Tuinen met correcte en overschatte OC-gehaltes, met bijhorende beoordelingsklasse volgens de Bodemkundige dienst van België (Maes et al. 2012).
Tuin pH OC (%) Beoordelingsklasse
correct OC-gehalte
25 4,268 1,483 streefzone
24 5,111 1,776 tamelijk hoog
37 5,2 1,703 tamelijk hoog
30 5,585 2,909 tamelijk hoog
35 5,686 3,059 tamelijk hoog
19 5,908 1,288 streefzone
33 5,985 2,111 tamelijk hoog
1 5,991 1,681 streefzone
22 6,267 2,118 tamelijk hoog
20 6,32 1,237 streefzone
34 6,323 1,768 tamelijk hoog
21 6,407 4,341 hoog
32 6,456 3,365 hoog
overschat OC - gehalte
28 6,524 1,714 tamelijk hoog
2 6,547 2,477 tamelijk hoog
31 6,58 1,772 tamelijk hoog
7 6,608 1,767 tamelijk hoog
11 6,617 1,465 streefzone
10 6,627 7,181 zeer hoog
26 6,64 3,54 hoog
3 6,662 8,038 zeer hoog
18 6,683 1,62 streefzone
4 6,704 2,931 tamelijk hoog
36 6,762 2,944 tamelijk hoog
14 6,781 3,191 hoog
9 6,813 2,77 tamelijk hoog
16 6,925 2,605 tamelijk hoog
15 6,998 3,628 hoog
29 7,019 1,519 streefzone
8 7,063 8,776 zeer hoog
17 7,086 2,031 tamelijk hoog
6 7,096 1,89 tamelijk hoog
5 7,105 1,562 streefzone
27 7,107 1,576 streefzone
13 7,192 2,117 tamelijk hoog
23 7,215 3,038 tamelijk hoog
12 7,334 1,547 streefzone

71
Tabel A.5: Verdeelsleutel van juveniele soorten volgens drie determinatiekenmerken.
groep
Kenmerken
soorten
Gep
aard
(0 =
nee
; 1 =
ja)
Pig
men
t
(0 =
nee
; 1 =
ja)
Pro
sto
miu
m
(ep
iloo
b =
0;
tan
ylo
ob
= 1
) 1 1 1 1 Eisenia fetida, Aporrectodea longa
2 1 1 0 Lumbricus terrestris, L. rubellus, L. castaneus, L. festivus
3 1 0 0
4 1 0 1 Dendrodrilus rubidus, Dendrobaena attemsi, D. octaedra
5 0 1 1 Eisenia fetida, A. caliginosa, A. limicola, A. rosea, A. icterica, Allolobophora chlorotica
6 0 1 0
7 0 0 1 D. pygmea, Octolasion cyaneum, O. lacteum, Microscolex phosphorus
8 0 0 0

72
Tabel A.6: Transformatie van variabelen om normaliteit en homoscedasticiteit te bekomen. Variabelen met een lege cel in de tweede kolom konden niet getransformeerd worden. Geen enkele van de variabelen was initieel normaal verdeeld.
Variabele transformatie
pHKCl
vochtgehalte (%)
N03-residu (kg/ha)
P (mg/kg)
K (mg/kg) f(x) = log(x)
Mg (mg/kg) f(x) = log(x)
Ca (mg/kg) f(x) = log(x)
%OC f(x) = log(x)
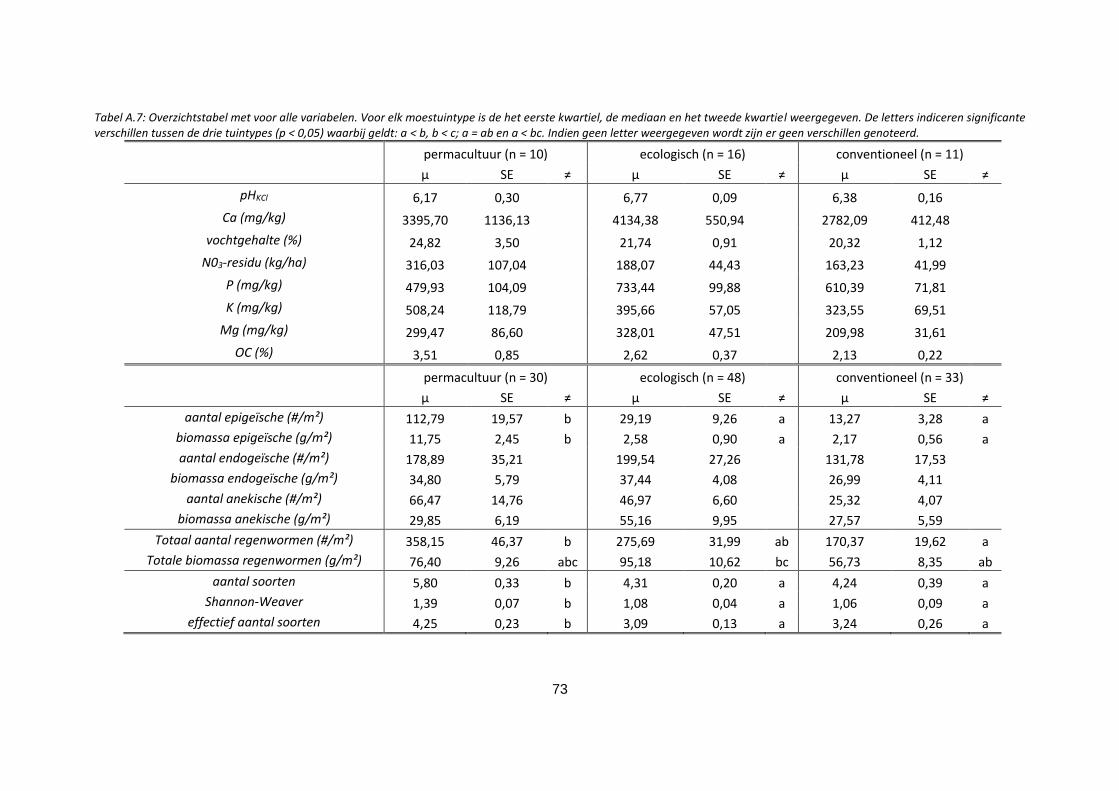
73
Tabel A.7: Overzichtstabel met voor alle variabelen. Voor elk moestuintype is de het eerste kwartiel, de mediaan en het tweede kwartiel weergegeven. De letters indiceren significante verschillen tussen de drie tuintypes (p < 0,05) waarbij geldt: a < b, b < c; a = ab en a < bc. Indien geen letter weergegeven wordt zijn er geen verschillen genoteerd.
permacultuur (n = 10) ecologisch (n = 16) conventioneel (n = 11)
µ SE ≠ µ SE ≠ µ SE ≠
pHKCl 6,17 0,30 6,77 0,09 6,38 0,16
Ca (mg/kg) 3395,70 1136,13 4134,38 550,94 2782,09 412,48
vochtgehalte (%) 24,82 3,50 21,74 0,91 20,32 1,12
N03-residu (kg/ha) 316,03 107,04 188,07 44,43 163,23 41,99
P (mg/kg) 479,93 104,09 733,44 99,88 610,39 71,81
K (mg/kg) 508,24 118,79 395,66 57,05 323,55 69,51
Mg (mg/kg) 299,47 86,60 328,01 47,51 209,98 31,61
OC (%) 3,51 0,85 2,62 0,37 2,13 0,22
permacultuur (n = 30) ecologisch (n = 48) conventioneel (n = 33)
µ SE ≠ µ SE ≠ µ SE ≠
aantal epigeïsche (#/m²) 112,79 19,57 b 29,19 9,26 a 13,27 3,28 a
biomassa epigeïsche (g/m²) 11,75 2,45 b 2,58 0,90 a 2,17 0,56 a
aantal endogeïsche (#/m²) 178,89 35,21 199,54 27,26 131,78 17,53
biomassa endogeïsche (g/m²) 34,80 5,79 37,44 4,08 26,99 4,11
aantal anekische (#/m²) 66,47 14,76 46,97 6,60 25,32 4,07
biomassa anekische (g/m²) 29,85 6,19 55,16 9,95 27,57 5,59
Totaal aantal regenwormen (#/m²) 358,15 46,37 b 275,69 31,99 ab 170,37 19,62 a
Totale biomassa regenwormen (g/m²) 76,40 9,26 abc 95,18 10,62 bc 56,73 8,35 ab
aantal soorten 5,80 0,33 b 4,31 0,20 a 4,24 0,39 a
Shannon-Weaver 1,39 0,07 b 1,08 0,04 a 1,06 0,09 a
effectief aantal soorten 4,25 0,23 b 3,09 0,13 a 3,24 0,26 a
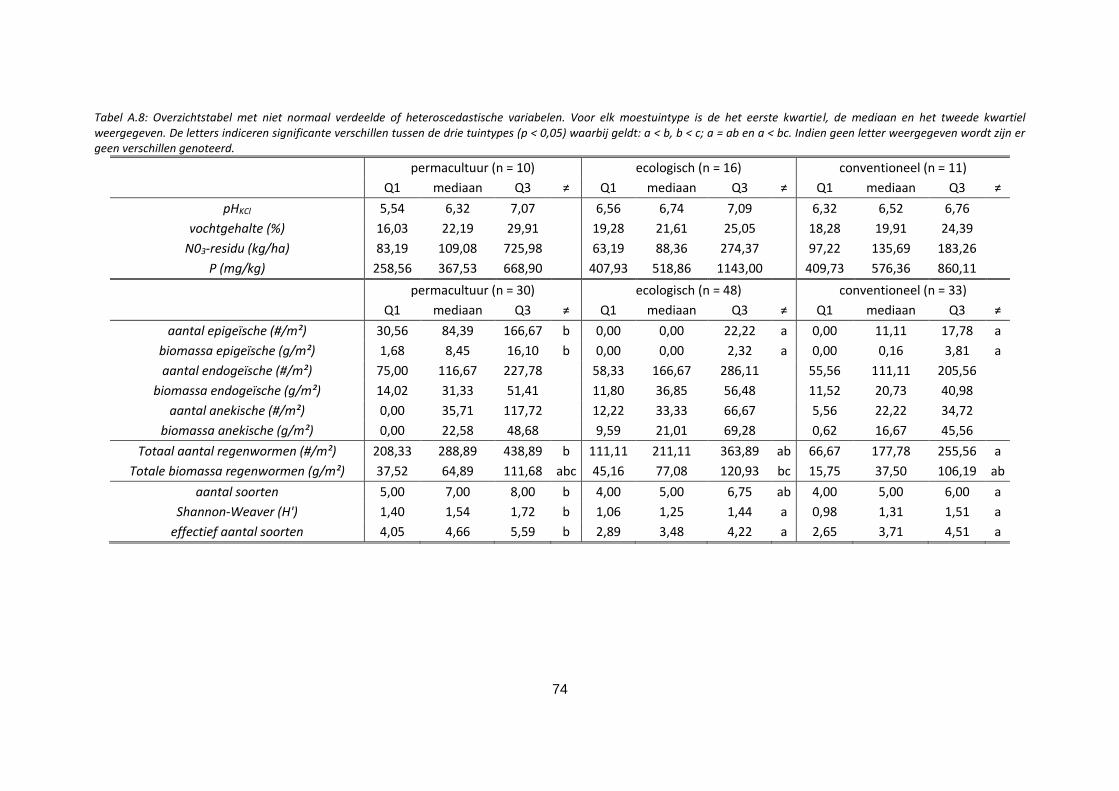
74
Tabel A.8: Overzichtstabel met niet normaal verdeelde of heteroscedastische variabelen. Voor elk moestuintype is de het eerste kwartiel, de mediaan en het tweede kwartiel weergegeven. De letters indiceren significante verschillen tussen de drie tuintypes (p < 0,05) waarbij geldt: a < b, b < c; a = ab en a < bc. Indien geen letter weergegeven wordt zijn er geen verschillen genoteerd.
permacultuur (n = 10) ecologisch (n = 16) conventioneel (n = 11)
Q1 mediaan Q3 ≠ Q1 mediaan Q3 ≠ Q1 mediaan Q3 ≠
pHKCl 5,54 6,32 7,07 6,56 6,74 7,09 6,32 6,52 6,76
vochtgehalte (%) 16,03 22,19 29,91 19,28 21,61 25,05 18,28 19,91 24,39
N03-residu (kg/ha) 83,19 109,08 725,98 63,19 88,36 274,37 97,22 135,69 183,26
P (mg/kg) 258,56 367,53 668,90 407,93 518,86 1143,00 409,73 576,36 860,11
permacultuur (n = 30) ecologisch (n = 48) conventioneel (n = 33)
Q1 mediaan Q3 ≠ Q1 mediaan Q3 ≠ Q1 mediaan Q3 ≠
aantal epigeïsche (#/m²) 30,56 84,39 166,67 b 0,00 0,00 22,22 a 0,00 11,11 17,78 a
biomassa epigeïsche (g/m²) 1,68 8,45 16,10 b 0,00 0,00 2,32 a 0,00 0,16 3,81 a
aantal endogeïsche (#/m²) 75,00 116,67 227,78 58,33 166,67 286,11 55,56 111,11 205,56
biomassa endogeïsche (g/m²) 14,02 31,33 51,41 11,80 36,85 56,48 11,52 20,73 40,98
aantal anekische (#/m²) 0,00 35,71 117,72 12,22 33,33 66,67 5,56 22,22 34,72
biomassa anekische (g/m²) 0,00 22,58 48,68 9,59 21,01 69,28 0,62 16,67 45,56
Totaal aantal regenwormen (#/m²) 208,33 288,89 438,89 b 111,11 211,11 363,89 ab 66,67 177,78 255,56 a
Totale biomassa regenwormen (g/m²) 37,52 64,89 111,68 abc 45,16 77,08 120,93 bc 15,75 37,50 106,19 ab
aantal soorten 5,00 7,00 8,00 b 4,00 5,00 6,75 ab 4,00 5,00 6,00 a
Shannon-Weaver (H') 1,40 1,54 1,72 b 1,06 1,25 1,44 a 0,98 1,31 1,51 a
effectief aantal soorten 4,05 4,66 5,59 b 2,89 3,48 4,22 a 2,65 3,71 4,51 a
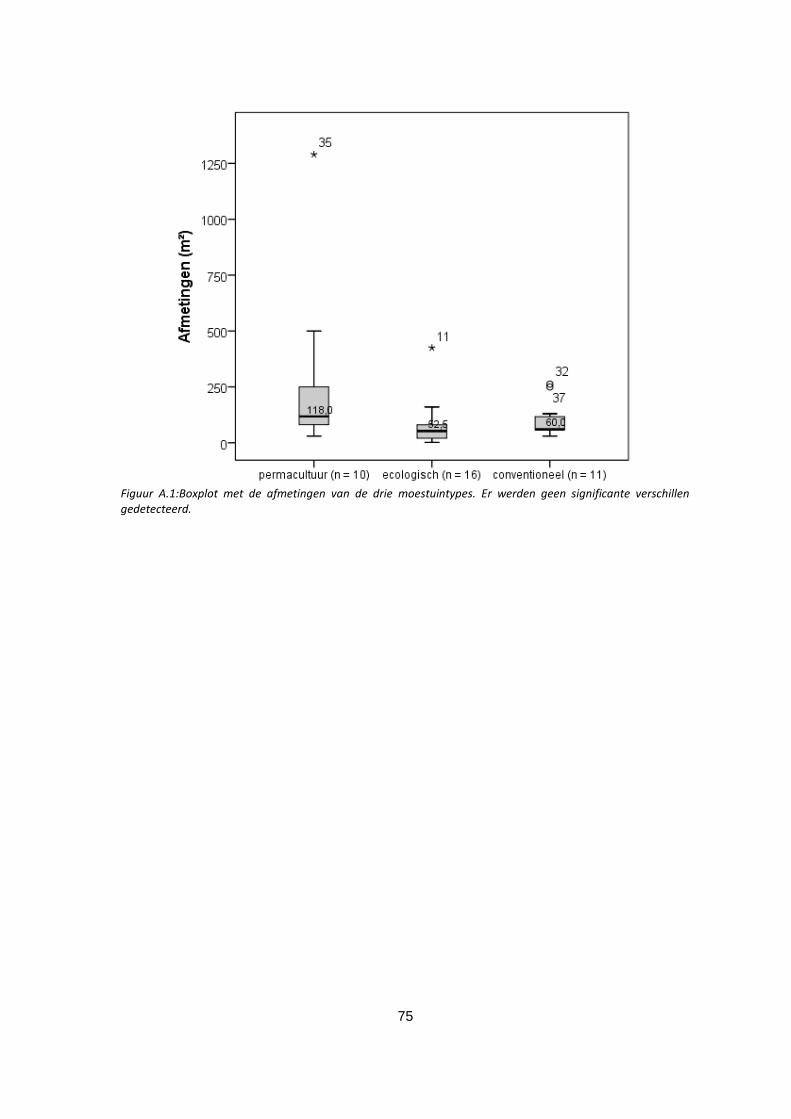
75
Figuur A.1:Boxplot met de afmetingen van de drie moestuintypes. Er werden geen significante verschillen gedetecteerd.
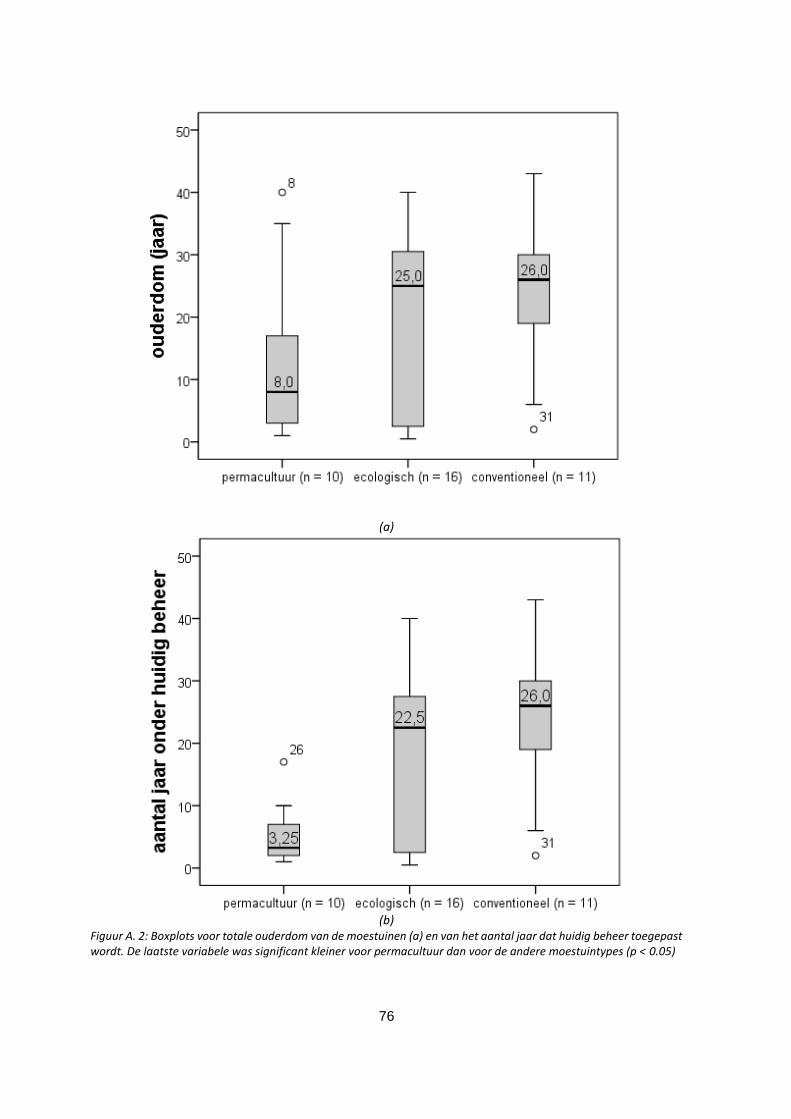
76
(a)
(b) Figuur A. 2: Boxplots voor totale ouderdom van de moestuinen (a) en van het aantal jaar dat huidig beheer toegepast wordt. De laatste variabele was significant kleiner voor permacultuur dan voor de andere moestuintypes (p < 0.05)
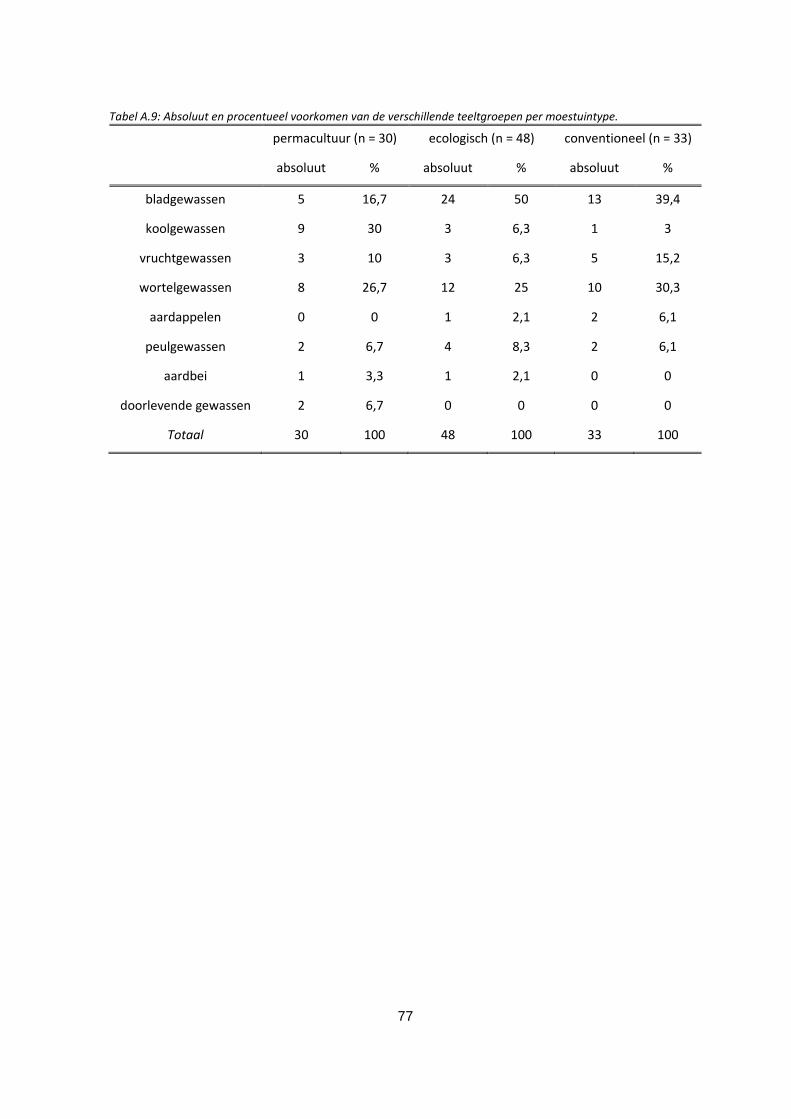
77
Tabel A.9: Absoluut en procentueel voorkomen van de verschillende teeltgroepen per moestuintype.
permacultuur (n = 30) ecologisch (n = 48) conventioneel (n = 33)
absoluut % absoluut % absoluut %
bladgewassen 5 16,7 24 50 13 39,4
koolgewassen 9 30 3 6,3 1 3
vruchtgewassen 3 10 3 6,3 5 15,2
wortelgewassen 8 26,7 12 25 10 30,3
aardappelen 0 0 1 2,1 2 6,1
peulgewassen 2 6,7 4 8,3 2 6,1
aardbei 1 3,3 1 2,1 0 0
doorlevende gewassen 2 6,7 0 0 0 0
Totaal 30 100 48 100 33 100
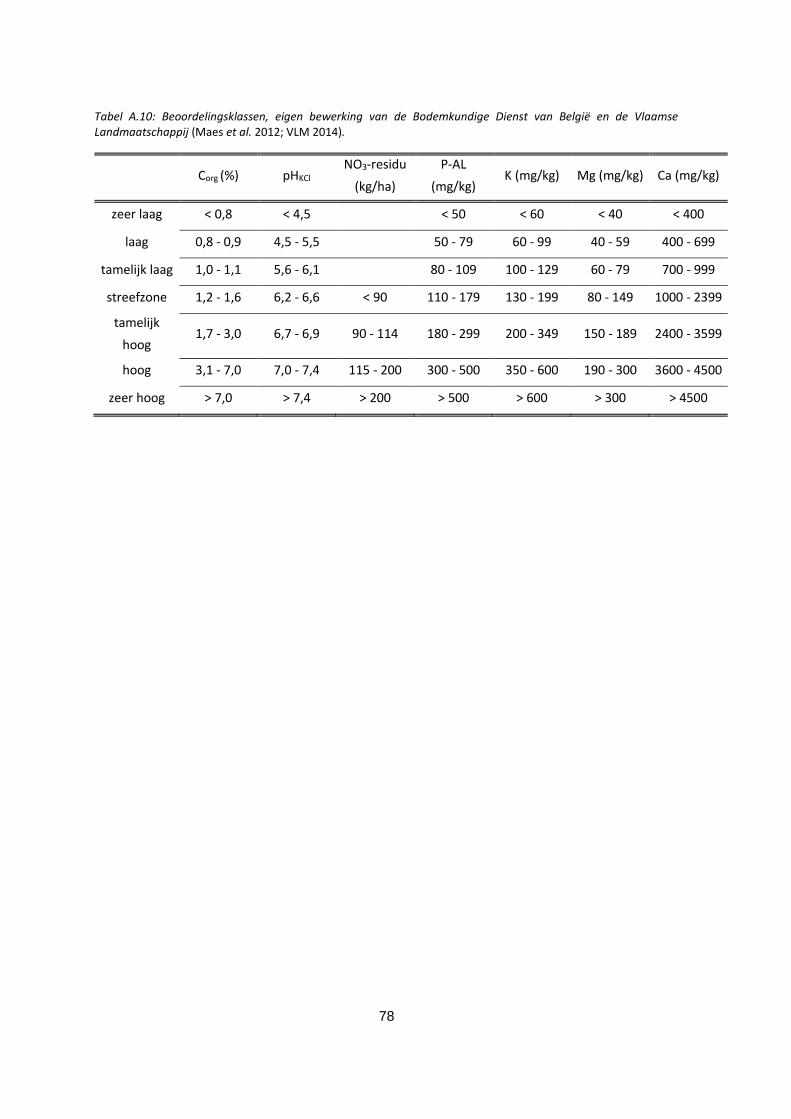
78
Tabel A.10: Beoordelingsklassen, eigen bewerking van de Bodemkundige Dienst van België en de Vlaamse Landmaatschappij (Maes et al. 2012; VLM 2014).
Corg (%) pHKCl NO3-residu
(kg/ha)
P-AL
(mg/kg) K (mg/kg) Mg (mg/kg) Ca (mg/kg)
zeer laag < 0,8 < 4,5 < 50 < 60 < 40 < 400
laag 0,8 - 0,9 4,5 - 5,5 50 - 79 60 - 99 40 - 59 400 - 699
tamelijk laag 1,0 - 1,1 5,6 - 6,1 80 - 109 100 - 129 60 - 79 700 - 999
streefzone 1,2 - 1,6 6,2 - 6,6 < 90 110 - 179 130 - 199 80 - 149 1000 - 2399
tamelijk
hoog 1,7 - 3,0 6,7 - 6,9 90 - 114 180 - 299 200 - 349 150 - 189 2400 - 3599
hoog 3,1 - 7,0 7,0 - 7,4 115 - 200 300 - 500 350 - 600 190 - 300 3600 - 4500
zeer hoog > 7,0 > 7,4 > 200 > 500 > 600 > 300 > 4500

79
Tabel A.11: Bodemtypes waarvoor de beoordelingsklassen uit Tabel A.1 van toepassing zijn, eigen bewerking van de Bodemkundige Dienst van België en de Vlaamse Landmaatschappij (Maes et al. 2012; VLM 2014).
bodemtype
Corg (%) zandleem - leem
pHKCl zandleem
NO3-residu (kg/ha) elk type bodem behalve zand- en zware kleigronden
P (mg/kg) elk type bodem
K (mg/kg) alle bodems behalve polders
Mg (mg/kg) zandleem - leem
Ca (mg/kg) Zandleem


















