
UNIVERSITEIT GENT FACULTEIT DIERGENEESKUNDE … · Subphylum Apicomplexa (coccidia) Genera Isospora...
34
UNIVERSITEIT GENT FACULTEIT DIERGENEESKUNDE Academiejaar 2011-2012 GASTRO-ENTERITIS BIJ HAGEDISSEN Door Tess ENGELEN Promotor : Dr. B. Leyman Copromotor: Prof. Dr. F. Pasmans Literatuurstudie in het kader van de masterproef
Transcript of UNIVERSITEIT GENT FACULTEIT DIERGENEESKUNDE … · Subphylum Apicomplexa (coccidia) Genera Isospora...

UNIVERSITEIT GENT
FACULTEIT DIERGENEESKUNDE
Academiejaar 2011-2012
GASTRO-ENTERITIS BIJ HAGEDISSEN
Door
Tess ENGELEN
Promotor : Dr. B. Leyman
Copromotor: Prof. Dr. F. Pasmans
Literatuurstudie in het kader van de
masterproef

De auteur en de promotor(en) geven de toelating deze studie als geheel voor consultatie beschikbaar
te stellen voor persoonlijk gebruik. Elk ander gebruik valt onder de beperkingen van het auteursrecht,
in het bijzonder met betrekking tot de verplichting de bron uitdrukkelijk te vermelden bij het aanhalen
van gegevens uit deze studie.
Het auteursrecht betreffende de gegevens vermeld in deze studie berust bij de promotor(en). Het
auteursrecht beperkt zich tot de wijze waarop de auteur de problematiek van het onderwerp heeft
benaderd en neergeschreven. De auteur respecteert daarbij het oorspronkelijke auteursrecht van de
individueel geciteerde studies en eventueel bijhorende documentatie, zoals tabellen en figuren.
De auteur en de promotor(en) zijn niet verantwoordelijk voor de behandelingen en eventuele
doseringen die in deze studie geciteerd en beschreven zijn.

VOORWOORD
Graag zou ik hierbij enkele personen willen bedanken die hebben bijgedragen tot de realisatie van
mijn literatuurstudie.
Vooreerst wil ik mijn promotor en medepromotor bedanken voor hun begeleiding, behulpzaamheid en
tijd. Daarnaast wil ik ook dierenarts L. Lambrechts bedanken voor het ter beschikking stellen van zijn
beeldmateriaal.
Ook mijn oprechte dankbetuiging aan mijn vriend voor zijn hulp, raad en mentale steun, en aan mijn
vrienden en familie.
Mag ik dan als laatste ook mijn honden en mijn kat niet vergeten te bedanken, om mijn voeten te
verwarmen tijdens het opzoekwerk en mij op tijd en stond op te beuren.

INHOUDSOPGAVE
VOORWOORD
INHOUDSOPGAVE
SAMENVATTING p. 1
INLEIDING p. 2
LITERATUURSTUDIE p. 3
1. Chromista en Protozoa p. 3
1.1. Inleiding p. 3
1.2. Etiologie p. 3
1.3. Epidemiologie p. 3
1.4. Pathogenese p. 5
1.5. Symptomen p. 5
1.6. Letsels p. 6
1.7. Diagnose p. 8
1.8. Behandeling p. 9
1.9. Preventie p. 10
2. Nematoden p. 11
2.1. Etiologie p. 11
2.2. Epidemiologie p. 11
2.3. Pathogenese p. 12
2.4. Symptomen p. 13
2.5. Letsels p. 14
2.6. Diagnose p. 15
2.7. Behandeling p. 16
2.8. Preventie p. 17
3. Trematoden en Cestoden p. 17
3.1. Etiologie p. 17
3.2. Epidemiologie p. 17
3.3. Pathogenese p. 18
3.4. Symptomen p. 19
3.5. Letsels p. 19
3.6. Diagnose p. 19
3.7. Behandeling p. 20
3.8. Preventie p. 20
4. Adenovirus p. 20
4.1. Etiologie p. 20
4.2. Epidemiologie p. 20
4.3. Pathogenese p. 21
4.4. Symptomen p. 21
4.5. Letsels p. 21
4.6. Diagnose p. 21
4.7. Behandeling p. 21
4.8. Preventie p. 22
5. Bacteriën en Fungi p. 22
5.1. Etiologie p. 22
5.2. Pathogenese p. 22
5.3. Diagnose p. 22
5.4. Behandeling p. 23
6. Niet infectieuze oorzaken p. 23
6.1. Etiologie, Epidemiologie en Pathogenese p. 23
6.2. Symptomen p. 24
6.3. Diagnose p. 24
6.4. Behandeling en Preventie p. 25
BESPREKING p. 26
REFERENTIELIJST p. 27

1
SAMENVATTING
Hagedissen worden meer en meer als gezelschapsdieren in huis gehouden. Aangezien gastro-
intestinale stoornissen hier zeer frequent de oorzaak zijn van ziekte, wordt er in deze literatuurstudie
verder ingegaan op de oorzakelijke agentia van gastro-enteritis bij deze dieren.
Er zullen 6 hoofdstukken aan bod komen. In deze hoofdstukken wordt telkens een oorzakelijk agens
besproken. Achtereenvolgens worden per agens de etiologie, epidemiologie, pathogenese,
symptomen, letsels, diagnose, behandeling en preventie overlopen.
In het eerste hoofdstuk ‘Protozoa en Chromista’ worden de eencellige parasieten besproken die van
pathologisch belang kunnen zijn bij gastro-intestinale stoornissen bij hagedissen, met nadruk op de
coccidieën, waaronder ook Cryptosporidium, de ciliaten, de amoeben en de flagellaten.
In het tweede hoofdstuk worden vervolgens de nematoden of rondwormen besproken. Hierbij wordt er
verder ingegaan op de Oxyurida (pinwormen), de Ascaridida (spoelwormen), de Rhabditoidea met als
voornaamste vertegenwoordiger de Strongyloides (haakwormen), de Strongylida, de Spirurida
(draadworm), de Enoplida met als belangrijkste genera Capillaria (haarwormen) en Trichinella en tot
slot de Gnathostomatoidea.
Het volgende hoofdstuk handelt over de Trematoden (platwormen) en Cestoden (lintwormen). Ook de
Acanthocephala zullen hier kort aan bod komen wegens de oppervlakkige gelijkenissen tussen deze
laatste en de Cestoden.
Vervolgens zal in het vierde hoofdstuk het enige pathogene gastro-intestinale virus dat beschreven is
voor hagedissen, namelijk Adenovirus, besproken worden.
Om de infectieuze oorzaken af te sluiten, worden ter volledigheid de bacteriën en de fungi (schimmels)
vermeld. Hoewel hun primair belang bij gastro-enteritis bij hagedissen nog niet duidelijk is, treden ze
toch vaak op als secundaire pathogenen.
Om af te ronden wordt er in het zesde hoofdstuk ingegaan op de niet infectieuze oorzaken met als
voornaamste toxines, impactie, obstructie, intussusceptie, cloacaprolaps en gastro-intestinale
neoplasieën.

2
INLEIDING
De populariteit van reptielen als huisdier neemt toe. Het is dan ook belangrijk dat er meer en meer
dierenartsen het belang ervan gaan inzien om ook deze dieren op een optimale manier te kunnen
benaderen. Vaak gaan er echter nog vele reptielen verloren doordat er nog veel onderzoek dient te
gebeuren in de herpetologie, of simpelweg ook omdat er nog onwetendheid heerst bij de
behandelende dierenarts.
Daarom handelt deze literatuurstudie over een zeer frequent voorkomende aandoening bij frequent
gehouden reptielen, namelijk gastro-enteritis bij hagedissen.
Enteritis is een van de belangrijkste oorzaken waarvoor reptielen worden aangeboden in de
dierenartspraktijk. Door meer te weten over de etiologie, epidemiologie, pathogenese en symptomen,
kan men de strategie plannen om een juiste diagnose te stellen, en vervolgens een aangepaste
behandeling inzetten. Men zal er in deze literatuurstudie achter komen, dat door meer te weten over
de etiologie en epidemiologie van de verschillende agentia, er vele gastro-intestinale klachten ook
vermeden kunnen worden.
In dit werkstuk zal vooral de aandacht gevestigd worden op de infectieuze oorzaken van gastro-
enteritis bij hagedissen. Hieronder zijn vooral de parasitosen erg belangrijk. Eén enkele virale
aandoening zal verder onder de loep genomen worden, namelijk Adenovirus. Ook bacteriën en
schimmels worden kort aangehaald. Ten slotte zal er nog ingegaan worden op de niet infectieuze
oorzaken.
Vaak zijn er nog geen uitgebreide onderzoeken gebeurd naar dit alles. Bijgevolg steunt deze
literatuurstudie vooral op case reports en informatie beschikbaar in handboeken.
Het is de bedoeling dat, na het lezen van deze literatuurstudie, een goed beeld verworven kan worden
van de gekende mogelijke aandoeningen in het gastro-intestinale stelsel van hagedissen, wat hopelijk
bijdraagt tot een betere diagnostiek en behandeling. Hopelijk kunnen er dan in de toekomst meer
reptielen geholpen of gered worden in de dierenartsenpraktijk.

3
LITERATUURSTUDIE
1. CHROMISTA EN PROTOZOA
1.1. INLEIDING
Infecties met Chromista en Protozoa worden beïnvloed door individuele verschillen tussen de
hagedissen, zoals het verschil in zuurtegraad, de microbiologie en de passage van voedingsstoffen in
de darmen. Er bestaan antagonismen tussen de verschillende soorten protozoa en ook predatie
tussen de soorten komt voor. De aanwezige protozoa hebben ook een invloed op de bacteriële
microflora van de darm, door competitie voor voedingsstoffen en predatie (1).
1.2. ETIOLOGIE
In volgende tabel worden de Chromista en Protozoa opgesomd die kunnen voorkomen in het gastro-
intestinale stelsel van hagedissen.
Tabel 1: Etiologie van Chromista en Protozoa bij hagedissen (2,3,4,5)
Koninkrijk Chromista
Subphylum Apicomplexa
(coccidia)
Genera Isospora (I. amphiboluri), Eimeria, Cyclospora, Caryospora,
Sarcocystis, Besnoita en Cryptosporidium (C. serpentis, C. saurofilus)
Phylum ciliophora
(ciliaten) Genera Balantidium, Nyctotherus
Koninkrijk Protozoa
Phylum Amoebozoa
(amoeben) Genus Entamoeba (E. invadens)
Phylum Flagellates
(flagellaten)
Genera Hexamita, Trichomonas, Leptomonas, Monocercomonoides,
Proteromonas, Monocercomonas, Giardia
1.3. EPIDEMIOLOGIE
1.3.1. Coccidia
Coccidia zijn gastheer specifieke organismen. Ze bevinden zich hoofdzakelijk in de enterocyten (1).
De meeste soorten bevinden zich intracytoplasmatisch, er zijn echter ook soorten gekend onder
Isospora, Eimeria en Cyclospora die zich intranucleair bevinden (4,6,7). Sommige genera, zoals
Isospora en Eimeria, hebben een directe levenscyclus, daartegenover staat bijvoorbeeld Sarcocystis,
die behoefte heeft aan een tussengastheer (2). Bij deze laatste kan de hagedis zowel tussengastheer
als eindgastheer zijn (8). Coccidia vormen oöcysten, na sporulatie zijn dezen de infectieuze stadia.
Sarcocystis is direct infectieus bij het verlaten van de gastheer. Andere soorten moeten eerst nog
sporuleren in de omgeving. Na opname verlaten de sporozoieten de oöcyst en penetreren de
enterocyten. Hier vindt er eerst aseksuele vermeerdering plaats, en daarna seksueel met vorming van
gameten en zygoten. Rond de zygote wordt er een wand gevormd, en deze vormt zich dan om tot
oöcyste. De oöcyste wordt uitgescheiden in de feces (2). Bij hagedissen behoort coccidiose tot de
meest voorkomende en meest bedreigende infectieziekten (9). Zware infecties met Coccidia worden

4
voornamelijk gezien bij hagedissen die in gevangenschap leven, waar er overbevolking heerst, en
slechte omgevingsfactoren aanwezig zijn (4).
De morbiditeit van Isospora amphiboluri bij jonge baardagamen (Pogona vitticeps) is zeer hoog (1).
Een slechte hygiëne en een beperkte oppervlakte kunnen de mortaliteit sterk doen toenemen (9).
Isospora wordt daarnaast vaak gezien bij kraaghagedissen (Chlamydosaurus kingii), jemenkameleons
(Chameleo calyptratus) en luipaardgekko’s (Eublepharis macularius) (10,11). Zelden word Isospora
waargenomen bij leguanen (Iguana spp.). Vooral jonge dieren zijn zeer gevoelig, oudere dieren treden
meestal op als dragers (9).
1.3.1.1. Cryptosporidium
Cryptosporidium werd al gediagnosticeerd bij 15 soorten hagedissen (12). De ontwikkeling vindt bij
hagedissen over het algemeen plaats in de dunne darm, vastgehecht aan de rand van de microvilli
door middel van een parasitofore vacuole (4,14,15). Cryptosporidium is echter ook al gediagnosticeerd
ter hoogte van de maag (13,14), de dikke darm (15) en de cloaca (16,17,18). Oöcysten met een
dunne wand zijn niet zeer resistent, en veroorzaken auto-infectie. De oöcysten met een dikke wand
worden uitgescheiden voor transmissie, en zijn zeer resistent in de omgeving. De oöcyst is direct
infectieus bij het verlaten van de gastheer (19). Transmissie kan dus zowel via direct ofwel via indirect
contact tot stand komen, en gebeurt zeer snel en efficiënt (9). De uiteindelijke ernst van infectie is
afhankelijk van de graad van reïnfectie, de graad van blootstelling in de omgeving en de immuniteit
van de gastheer. Vaak zijn erge infecties met Cryptosporidium geassocieerd met simultane ziekte of
suboptimale omgevingsfactoren. Bij massale infecties kunnen tot 80% van de cellen geïnfecteerd zijn
door Cryptosporidium (20).
1.3.2. Ciliaten
Adulte ciliaten bevinden zich in de dikke darm. Ze komen vooral voor bij herbivoren. Transmissie
gebeurt via feco-oraal contact (2,21).
1.3.3. Amoeben
Amoeben hebben 2 levensstadia, namelijk het trofozoiet stadium, waarin ze pseudopodia bevatten om
zich te voeden en rond te bewegen, en het ruststadium, waarin ze geëncysteerd zitten. Deze cysten
zijn infectieus bij uitscheiding. Na opname ontwikkelen ze zich tot trofozoiet, en planten ze zich voort
in de dikke darm. De cyclus is dus direct en een hoge infectiestatus kan snel bereikt worden bij de
hele populatie (2). Herbivore soorten lijken minder gevoelig aan infectie, waardoor ze als
asymptomatische dragers kunnen fungeren voor de carnivore hagedissen (22). Ook schildpadden
kunnen als reservoir optreden voor Entamoeba (9).
1.3.4. Flagellaten
Flagellaten hebben een directe levenscyclus. Ze komen verspreid voor in het gastro-intestinale stelsel
(2). Het zijn opportunistische organismen (9). Omgevingsfactoren zoals slechte temperaturen, sociale
stress, aanwezigheid van een andere ziekte, kunnen leiden tot een opflakkering van flagellaten (4).

5
1.4. PATHOGENESE
1.4.1. Coccidia
Coccidia zijn obligaat pathogene parasieten (1). De ziekteontwikkeling bij infecties met coccidia hangt
van een aantal factoren af, namelijk het aantal opgenomen oöcysten, snelheid van
epitheelregeneratie, de genetica van de parasiet en de immuniteitsstatus van de gastheer. De schade
die aangericht wordt, is evenredig met het aantal fasen van vermenigvuldiging dat heeft
plaatsgevonden in het darmepitheel (2). De schade veroorzaakt aan de darmmucosae, kan aanleiding
geven tot resorptiestoornissen (9). Wanneer geen ziekte ontstaat, spreekt men van coccidiase.
Wanneer wel ziekte ontstaan spreekt men van coccidiose (2). Cryptosporidium veroorzaakt een
overtollige mucussecretie en een gestoorde enzymsecretie in het gastro-intestinale kanaal, waardoor
een maldigestie ontstaat (13).
1.4.2. Ciliaten
Ciliaten zijn meestal niet pathogeen (2). Wanneer ze voorkomen in kleine aantallen treden ze op als
commensalen, wanneer er grote hoeveelheden ciliaten aanwezig zijn, kunnen ze echter wel ziekte
veroorzaken (10). Balantidium kan facultatief pathogeen zijn wanneer de darmwand op voorhand is
aangetast. Deze laatste kan ook pathologie veroorzaken door hyaluronidase te produceren. Daardoor
wordt de darmbarrière aangetast (9).
1.4.3. Amoeben
De meeste amoeben zijn apathogeen voor reptielen (4). Trofozoieten kunnen zich, afhankelijk van de
afweer van de gastheer, ingraven in de mucosa. Bij een immunocompetente hagedis zal dit niet
gebeuren (2).
Entamoeba invadens kan voor pathologie zorgen bij carnivore hagedissen, wanneer geen zetmeel
aanwezig is in het gastro-intestinale stelsel. De amoebe heeft behoefte aan deze energiebron, en bij
gebrek zal hij de cel binnendringen om glycogeen als alternatief te gebruiken (21).
1.4.4. Flagellaten
Flagellaten zijn, wanneer ze in kleine aantallen voorkomen, meestal
niet pathogeen (9). Bij zware infecties kunnen ze wel letsels
veroorzaken (10).
1.5 SYMPTOMEN
1.5.1. Coccidia
De meeste coccidia infecties verlopen symptoomloos (2). De
symptomen van Isospora amphiboluri bij baardagamen (Pogona
vitticeps) zijn anorexie, gewichtsverlies, diarree en groeivertraging. Bij
jonge baardagamen ligt de mortaliteit soms hoog. Sommige
baardagamen zijn echter klinisch gezond terwijl grote aantallen van
Fig.1 Baardagaam (Pogona vitticeps) met diarree en bevuilde cloaca (F. Pasmans)

6
oöcysten in de mest kunnen gevonden worden (2,23). Eimeria infecties kunnen symptomen zoals
waterige feces, anorexie, regurgitatie, dehydratatie en cachexie veroorzaken (7,21).
1.5.1.1. Cryptosporidium
Bij Cryptosporidium infecties kan chronisch vermageren
optreden (14,15,24). Symptomen die verder nog kunnen
voorkomen zijn onder andere anorexie, lethargie, diarree,
regurgitatie en een opgezwollen abdomen
(15,17,20,24,25,26). Er kunnen echter ook subklinische
infecties optreden, waarbij er soms een massale uitscheiding
aanwezig is. Deze dieren treden dan op als
asymptomatische dragers (14,20).
1.5.2. Ciliaten
Ciliaten veroorzaken meestal geen symptomen (2). Schneller en Pantchev beschrijven dat
Balantidium anorexie, diarree en gewichtsverlies kan veroorzaken bij productie van hyaluronidase
(2008).
1.5.3. Amoeben
De symptomen die kunnen optreden bij een Entamoeba infectie zijn anorexie, dehydratatie, een
slechte conditie en dysenterie met mucus en bloed (2). Ook een verharding van de darmwand kan
soms gevoeld worden, en dit mag niet verward worden met verharde mest. De aanwezigheid van
deze symptomen duidt op een ernstige infectie, en de kans is groot dat enkele dagen tot weken na het
verschijnen van de symptomen, de dieren zullen bezwijken (9).
1.5.4. Flagellaten
Flagellaten veroorzaken meestal geen symptomen. Wanneer een opflakkering aanwezig is, kunnen
anorexie en gewichtsverlies optreden (21). Bij kameleons (Chameleo sp.) werd al een uitbraak
besproken van Leptomonas waar de symptomen vooral anorexie en dehydratatie waren (27). Bij
infecties met Monocercomonas kan er anorexie optreden, gewichtsverlies, en gedragsveranderingen
zoals agressie (2). Lethargie en diarree zijn ook beschreven (28).
1.6. LETSELS
De vroege gastheerreactie bij een protozoaire infectie bestaat uit een infiltraat van heterofielen en
eosinofielen. Protozoa kunnen necrotiserende letsels veroorzaken in het gastro-intestinale stelsel, met
macrofaag en histiocyt granulomen tot gevolg (29).
1.6.1. Coccidia
Bij coccidia infecties worden er niet altijd letsels gevonden (30). Soms kan er verlies van intestinaal
epitheel optreden en hyperplasie van het epitheel. In de mucosa is er dan een ontstekingcellen
Fig.2 Cachexie luipaardgekko (Eublepharis macularius) door Cryptosporidiose (F. Pasmans)

7
infiltraat aanwezig (31). Coccidiose kan aanleiding geven tot hemorrhagische enteritis, intestinale
ulcers, fibrose en secundaire septicemie (21). Bij kameleons (Chameleo sp.) is er intussusceptie
beschreven door coccidiose (32).
1.6.1.1. Cryptosporidium
Cryptosporidium kan de oorzaak zijn van een chronische voortschrijdende enteritis, waarbij de schade
geleidelijk toeneemt (9). Er is dan losse inhoud aanwezig verspreid over de volledige darm. Er zijn
veel heterofielen, lymfocyten en macrofagen aanwezig in de mucosa (15,20). Er is een hypertrofische
enteritis en hyperplasie van de darmwand aanwezig (15,21,25). In een case report van Oros et al.
(1998) werd een kraaghagedis (Chlamydosaurus kingii) beschreven met een Cryptosporidium infectie
van de maag, die tevens secundair bacterieel geïnfecteerd was. De maag was hard en de
maagmucosa was verdikt, rood en bedekt met een dunne laag donkerrode mucus. Aan de pylorus
werd een geel-witte materie op het oppervlak gezien. De mucosa was hier necrotisch en er was een
infiltraat van mononucleaire ontstekingscellen aanwezig. Er werd een milde diffuse gastritis gevonden
met een infiltratie van ontstekingscellen, een diffuus verlies van granulaire cellen en grote aantallen
ronde kleine amfofiele organismen op het oppervlak. De bloedvaten vertoonden congestie. Er zijn
echter niet altijd lesies te zien bij Cryptosporidium infecties (3).
1.6.2. Ciliaten
Balantidium kan geassocieerd zijn met necrose en bloedingen in de dikke darm, wanneer deze
hyaluronidase produceert (9).
1.6.3. Amoeben
Amoeben kunnen ulceraties veroorzaken in de mucosa van de darm. Ulceratieve gastritis en colitis
kunnen gezien worden (2). Necrotiserende letsels zijn vaak aanwezig (33). Ook lever- en nierletsels
kunnen gezien worden met necrose en abcesvorming (2), doordat ze kunnen binnendringen in de
portale circulatie na ulceratie van de mucosa (4). Entamoeba invadens kan bij carnivore hagedissen
hemorrhagische enteritis veroorzaken (10). Gray et al., beschreven in 1966 bij necropsie van een
komodovaraan (Varanus komodoensis) hemorrhagische enteritis en colitis. Via lichtmicroscopie
werden amoeben gevonden nabij de ulceratieve letsels in het colon en de lever, en diffuse hepatische
necrose.
1.6.4.Flagellaten
Bij kameleons (Chameleo sp.) werd bij een uitbraak van Leptomonas vooral een bloederige colitis
gezien (27). Lesies die optreden bij Monocercomonas zijn onder andere ondermijning van het gastro-
intestinale epitheel, en het loskomen van het epitheel ten opzichte van de lamina propria (2). Bij
desquamatie van het epitheel kunnen er ontstekingscellen in de fibrinelaag op het mucosale oppervlak
gezien worden (4).
8
1.7 DIAGNOSE
Voor de diagnose van een protozoa infectie, kan
men gebruik maken van mestonderzoek. Voor
detectie van bewegende trofozoieten maakt men
gebruik van verse fecesuitstrijkjes, zo kan men
ciliaten, flagellaten en amoeben detecteren.
Flottatietechnieken met suiker kunnen gebruikt
worden voor coccidia-infecties. Men kan de feces
ook fixeren in polyvenylalcohol, en daarna kleuren
door middel van trichroom of ijzer-hematoxyline
(2). De PAS-kleuring kan ook aangewend worden
(34).
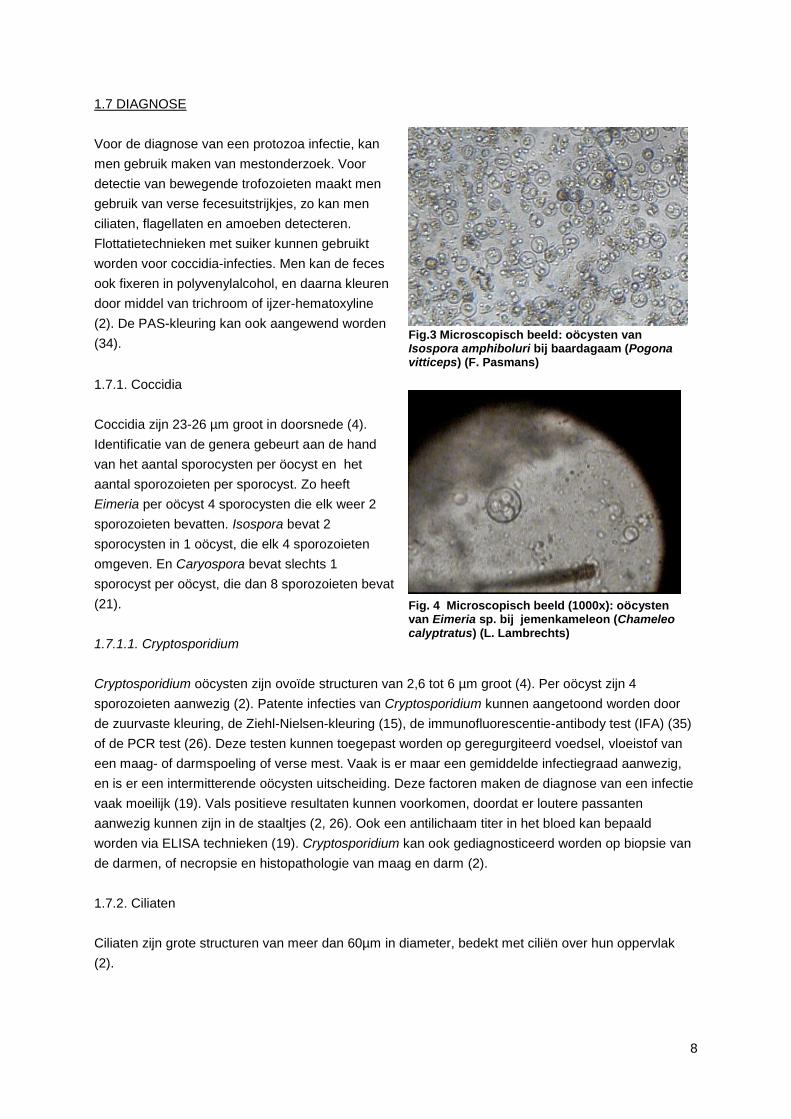
1.7.1. Coccidia
Coccidia zijn 23-26 µm groot in doorsnede (4).
Identificatie van de genera gebeurt aan de hand
van het aantal sporocysten per öocyst en het
aantal sporozoieten per sporocyst. Zo heeft
Eimeria per oöcyst 4 sporocysten die elk weer 2
sporozoieten bevatten. Isospora bevat 2
sporocysten in 1 oöcyst, die elk 4 sporozoieten
omgeven. En Caryospora bevat slechts 1
sporocyst per oöcyst, die dan 8 sporozoieten bevat
(21).
1.7.1.1. Cryptosporidium
Cryptosporidium oöcysten zijn ovoïde structuren van 2,6 tot 6 µm groot (4). Per oöcyst zijn 4
sporozoieten aanwezig (2). Patente infecties van Cryptosporidium kunnen aangetoond worden door
de zuurvaste kleuring, de Ziehl-Nielsen-kleuring (15), de immunofluorescentie-antibody test (IFA) (35)
of de PCR test (26). Deze testen kunnen toegepast worden op geregurgiteerd voedsel, vloeistof van
een maag- of darmspoeling of verse mest. Vaak is er maar een gemiddelde infectiegraad aanwezig,
en is er een intermitterende oöcysten uitscheiding. Deze factoren maken de diagnose van een infectie
vaak moeilijk (19). Vals positieve resultaten kunnen voorkomen, doordat er loutere passanten
aanwezig kunnen zijn in de staaltjes (2, 26). Ook een antilichaam titer in het bloed kan bepaald
worden via ELISA technieken (19). Cryptosporidium kan ook gediagnosticeerd worden op biopsie van
de darmen, of necropsie en histopathologie van maag en darm (2).
1.7.2. Ciliaten
Ciliaten zijn grote structuren van meer dan 60µm in diameter, bedekt met ciliën over hun oppervlak
(2).
Fig.3 Microscopisch beeld: oöcysten van Isospora amphiboluri bij baardagaam (Pogona vitticeps) (F. Pasmans)
Fig. 4 Microscopisch beeld (1000x): oöcysten van Eimeria sp. bij jemenkameleon (Chameleo calyptratus) (L. Lambrechts)

9
1.7.3. Amoeben
Ook onderzoek van histologische coupes van darm of lever kan aangewend worden voor het
opsporen van amoeben na necropsie op een pas gestorven hagedis. Deze coupes worden gekleurd
door middel van trichroom of ijzer-hematoxyline. De trofozoieten zijn dan aantoonbaar extracellulair
naast de letsels (2). PCR technieken kunnen gebruikt worden om verschillende soorten van amoeben
te differentiëren (4).
1.7.4. Flagellaten
Hexamita stadia zijn kleiner dan 8 µm en zwemmen altijd zeer snel uit het microscopische beeld.
Giardia daarentegen zwemt gracieus en maakt een rollende beweging. Dezen bevatten ook een
zuigschijf. Monocercomonas bevat een rigide axostyle in de lengte, 3 anterieure flagellen en 1
posterieure. Scanning elektronen microscopie bij flagellaten kan ook gebruikt worden om de soorten
onderling te kunnen onderscheiden en differentiëren (2).
1.8. BEHANDELING
Aangezien de meeste soorten niet pathogeen zijn, is behandeling enkel nodig in de gevallen waar er
massale infectie heerst en waar ziektesymptomen aanwezig zijn (1). Naast de specifieke behandeling
van de parasitose, dient soms ook een ondersteunende behandeling ingezet te worden die secundaire
pathologieën zoals dehydratatie, cachexie en secundaire bacteriële infecties te lijf gaat. Eveneens de
omgeving van de patiënt moet dan gesaneerd worden. Bij groepshuisvesting wordt quarantaine van
zieke dieren aanbevolen (9).
1.8.1. Coccidia
Coccidia zijn moeilijk te eradiceren. Men kan behandelen door middel van sulfa-dimethoxine (50
mg/kg peroraal sid gedurende 3 tot 4 weken). Ook trimetroprim-sulfadiazine kan zorgen voor een
daling van de uitscheiding (eenmalig 30 mg/kg peroraal, gevolgd door 15 mg/kg sid peroraal
gedurende 3 tot 4 weken) (1). In 50% van de gevallen wordt door middel van deze behandeling
volledige eliminatie bereikt (10). Men moet wel aandacht schenken aan de hydratatietoestand van de
hagedis tijdens zulke behandelingen, want na enkele toedieningen van sulfonamiden kan er anorexie
optreden. Het mag ook niet aan nierpatiënten gegeven worden (10). Ook behandeling met toltrazuril
(36) of ponazuril (37) wordt voorgesteld. Oöcysten van coccidia zijn zeer resistent in de omgeving. Ze
worden niet vernietigd door uitdroging of osmotische veranderingen (10).
1.8.1.1. Cryptosporidium
Een behandeling tegen Cryptosporidium bij hagedissen is genoodzaakt, omdat een infectie hier een
chronische vorm kan aannemen, en niet zelflimiterend is (20). De behandeling van Cryptosporidium is
echter zeer moeilijk en moet langdurig aangehouden worden (8). Aprinocid of halofuginone kunnen
niet toegediend worden, want dezen zijn toxisch voor reptielen (19).
Boviene hyperimmuun colostrum is effectief gebleken tegen C. saurofilus bij de savannevaraan
(Varanus exanthematicus) (1% van het volume van het lichaamsgewicht 1 maal per week gedurende
6 weken) (17,38,39,40). Dezelfde therapie bij luipaardgekko’s (Eublepharius macularius) haalde

10
minder positieve resultaten. Het serum bestrijdt Cryptosporidium door de bevattende antistoffen.
Daarnaast is het ook een zeer goede bron van makkelijk verteerbare eiwitten, die een
ondersteunende invloed hebben (40).
Andere behandelingspistes zoals paromomycine (100 mg/kg gedurende 1 week gevolgd door 2 maal
per week gedurende 3 maanden), spiromycine (160mg/kg gedurende 1 week gevolgd door 2 maal per
week gedurende 3 maanden) of trimetroprim-sulfa (eenmaal daags gedurende 2 weken, en daarna 1
tot 3 maal per week gedurende verschillende maanden) zorgden voor een gedaalde uitscheiding van
oöcysten bij slangen (20,35). Men moet bij aanwending van paromomycine genoeg drinkwater ter
beschikking stellen, aangezien dit een nefrotoxische stof is (9).
Cryptosporidium is tevens moeilijk te bestrijden in de omgeving, omdat het zo resistent is tegen vele
desinfecteermiddelen. Bestrijding kan door middel van vochtige hitte, invriezing of uitdroging.
Neutralisatie van de infectiviteit wordt bekomen na een vochtige hitte tussen de 45 tot 60°C
gedurende 5 tot 9 minuten (20). Ammonium 5% of formal saline 10% (dit is een mengsel van
formaldehyde en methanol) stoppen de infectiviteit van Cryptosporidium na 18 uren contact aan 4°C
(20,41). Na het reinigen van de terrarium met een ammoniumoplossing moet deze 3 dagen leeg
blijven staan om volledig op te drogen (20).
Dieren zijn pas officieel vrij verklaard van Cryptosporidium als ze gedurende 1 jaar geen symptomen
meer vertonen en geen oöcysten meer uitscheiden (9).
1.8.2. Ciliaten
Ciliaten kunnen behandeld worden door middel van metronidazole (21). Wanneer ziekte veroorzaakt
wordt door Balantidium, dient de predisponerende oorzaak eerst behandeld te worden (9).
1.8.3. Amoeben
De behandeling van een infectie met Entamoeba invadens kan door middel van metronidazole of
nitro-imidazole. Paromomycine en jodoquinol in combinatie met metronidazole wordt vaak
aangewend, omdat de eerste 2 hun werking uiten tegenover cysten, en metronidazole tegenover de
trofozoieten. Alle dieren uit dezelfde terrarium dienen gelijktijdig behandeld te worden. Verder moet
men er op letten dat de omgevingstemperatuur hoog genoeg is, zo’n 35°C (9).
1.8.4. Flagellaten
Flagellaten kunnen bestreden worden door middel van metronidazole (21). Als er echter meer dan 2
behandelingen nodig zijn om een daling in uitscheiding te komen, is het aan te raden naar een andere
oorzaak van de ziekte te zoeken (10). De behandeling van een Monocercomonas infectie kan door
toediening van ronidazole peroraal (10mg/kg gedurende 8 tot 10 dagen of 150mg/l in drinkwater) (2).
1.9 PREVENTIE
In de preventie van parasitose is het belangrijk een goede hygiëne aan te wenden in de terraria, zoals
het regelmatig verwijderen van organisch materiaal. Dit om de infectiedruk zo laag mogelijk te houden
(9). Nieuw aangekochte dieren blijven best gedurende minstens 30 dagen in quarantaine, om
eventuele infectie op te sporen. Ook een jaarlijks parasitologisch onderzoek is aan te raden om
infecties vroegtijdig op te sporen (2).

11
Preventie van een infectie met Cryptosporidium is van primordiaal belang, omdat het zo moeilijk te
bestrijden is (19). Hygiëne en een goed management is uiterst belangrijk (20). Ook dienen alle
bijkomende ziekmakende factoren, voedingsfouten en omgevingsfouten bestreden te worden (20,38).
Preventie van Entamoeba infecties is ook zeer belangrijk. Het apart houden van drager dieren en
gevoelige dieren is hier aan te raden (21).
2. NEMATODEN
2.1. ETIOLOGIE
Hier volgt een opsomming van de rondwormen die al frequent beschreven zijn bij hagedissen:
- Oxyuris spp. van de orde Oxyurida (1)
- de families Ascarididae, Anisakidae en Heterakidae van de orde Ascaridida (1,9)
- het genus Strongyloides van de orde Rhabditoidea (2)
- 3 genera van de orde Strongylida, namelijk Diaphanocephaloidea spp., Oswaldocruzia spp.,
en Kalicephalus spp. (1,42)
Zeldzamer (8) kunnen ook nog volgende nematoden voorkomen:
- Physaloptera en de Dracunculoidea spp. van de orde Spirurida (21)
- de families Trichuridae (oa. Capillaria spp.) en Trichinellidae van de orde Enoplida (2,21)
- de orde Gnathostomatoidea (43)
2.2. EPIDEMIOLOGIE
Nematoden kunnen zowel een directe als een indirecte levenscyclus hebben. Eieren of larven worden
mee uitgescheiden in de feces (2). Bij nematoden met een directe levenscyclus, kan de infectiegraad
snel hoog oplopen in een terrarium, waardoor auto-infectie kan ontstaan (9,44).
2.2.1. Oxyurida
De orde Oxyurida komt vooral voor bij insectivore of herbivore hagedissen (9). Soorten die hiertoe
behoren hebben een hoge gastheerspecificiteit. De meeste hagedissen zijn geïnfecteerd met
minstens 1 soort, sommigen bevatten zelfs meerdere soorten (2). Deze nematoden hebben een
directe levenscyclus. Na uitscheiding van de eieren in de mest, ontwikkelen dezen zich tot infectieuze
larven. Deze laatsten worden peroraal opgenomen. De adulten leven in de dikke darm (9,21).
Wanneer vele dieren in een kleine ruimte gehuisvest zijn en aan permanente auto-infectie onderhevig
zijn, kunnen zelfs aantallen van 60000 wormen in 1 dier voorkomen (27). In de natuur echter, ontstaan
er zelden zulke massale infecties, en stelt er zich een evenwicht tussen de gastheer en de parasiet
(11).
2.2.2. Ascaridida
De levenscyclus van de orde Ascaridida verloopt indirect bij hagedissen (9). Bij uitscheiding van
wormeieren ontwikkelt er zich in 7 dagen tijd een L3 larve, die na opname door de tussengastheer
gedurende 4 weken verder ontwikkelt tot het infectieuze stadium. Bij opname van een tussengastheer
die infectieuze stadia bevat, ontwikkelt de larve zich verder tot L4 in de darmwand, en wordt

12
vervolgens adult in het darmlumen (27). Indien de larve echter nog niet infectieus is wanneer de
tussengastheer ten prooi valt, boort de larve zich in de darmwand, waarna hij zich hier inkapselt (9).
2.2.3. Rhabditoidea
Strongyloides soorten hebben een directe levenscyclus. De L3 larven zijn de infectieuze stadia, en
kunnen ofwel peroraal opgenomen worden, ofwel de huid van de gastheer penetreren (2). Daarna
ondergaan ze een migratie langs hart, longen en slokdarm (11). De adulten komen vooral voor in de
dunne darm van de gastheer (2). Larven kunnen tot meerdere maanden infectieus blijven in de
omgeving (11).
2.2.4. Strongylida
Nematoden van de orde Strongylida hebben een directe levenscyclus. De eieren ontwikkelen in een
vochtige omgeving tot L3 larven (9). De infectieuze stadia kunnen door de huid penetreren of peroraal
opgenomen worden. De adulten leven in het gastro-intestinale kanaal van de oesofagus tot de dikke
darm (21). Ze hebben een lage gastheerspecificiteit (10).
2.2.5. Spirurida
De nematoden van de orde Spirurida bezitten een indirecte levenscyclus. Hierbij is de tussengastheer
meestal een arthropode. Physaloptera spp. gebruiken een mier als tussengastheer (4). De adulten
komen vooral voor in het bovenste gedeelte van het gastro-intestinale kanaal, meer bepaald de maag.
Hagedissen kunnen hier echter ook optreden als paratenische gastheer of als tussengastheer (4,21).
2.2.6. Enoplida
Capillaria spp. leven in de dunne darm (2). Ze bedden zich gedeeltelijk of volledig in in de mucosae.
De wormeieren van deze nematoden zijn zeer resistent in de omgeving (9). Larven van de familie
Trichinellidae kunnen geëncysteerd zitten in de mucosa van de maag (21).
2.3 PATHOGENESE
Meestal zijn de effecten van nematoden op hun gastheer onbekend, en vaak kunnen ze neutraal
aanwezig zijn (2). Pathologie bij nematoden kan zowel veroorzaakt worden door migrerende larven,
als door adulte wormen. Congregaties van grote aantallen nematoden kunnen onder andere leiden tot
orgaan- en weefseldestructie, ontsteking, verlies van voedingsstoffen en secundaire bacteriële
infecties (10).
2.3.1. Oxyurida
Oxyuris spp. zijn meestal niet pathogeen, tenzij bij massale infecties (1, 29). Er zijn zelfs positieve
eigenschappen toegekend aan oxyuriden infecties. In de dikke darm voeden ze zich met darminhoud
(9). Hier kunnen ze bijdragen tot een optimale vertering bij herbivore hagedissen, zoals de groene
leguaan (Iguana iguana). Ze helpen in de mechanische afbraak van ingesta en de preventie van
cellulose impactie (45). Toch is er al een fatale impactie gemeld door infectie met oxyuriden (46). Bij

13
hoge infectiegraad kan de hagedis lijden aan overmatige gasopstapeling. Door deze ruimte-inname
kunnen ademhalingsstoornissen ontstaan (9). In winterslaap kan deze parasiet voor problemen
zorgen, wanneer bij massale wormaantallen, de stofwisselingsproducten van de wormen niet meer
uitgescheiden kunnen worden. Dit kan dan aanleiding geven tot intoxicaties (27).
2.3.2.Ascaridida
Ascariden zijn de meest pathogene nematoden bij hagedissen. De larven boren zich door de
darmwand, hetzij voor ontwikkeling tot L4, hetzij voor inkapseling. Dit kan leiden tot ernstige
weefseldestructie bij massale invasie. Daarnaast kunnen ingekapselde larven bepaalde
orgaanfuncties verstoren. Migrerende larven kunnen voor verbloeding zorgen bij arteriële schade.
Adulten hechten zich vast aan de maag- of darmwand, en bij grote aantallen kan de darmwand
geperforeerd worden (9). Daarnaast kunnen de adulten soms aanleiding geven tot impactie van het
gastro-intestinale kanaal (10). Bij zware infecties kunnen ze tot 50 procent van de voedselopname van
de hagedis absorberen. Secundaire malnutritie kan dan optreden (10). Heterakiden infecties zijn
meestal apathogeen (9).
2.3.3.Rhabditoidea en Strongylida
Adulten van de orde Strongylida en Rhabditoidea hechten zich met hun getande mondplaat vast aan
de mucosae van het gastro-intestinale kanaal, waardoor schade ontstaat aan de mucosae. Hierdoor
kunnen resorptiestoornissen ontstaan. Ze voeden zich met bloed en weefselvocht (9,10,21).
2.3.4. Spirurida
Adulten van deze orde bevinden zich voornamelijk in
de mucosae van de maag. Bij massale aanwezigheid
van wormen kunnen onder andere resorptiestoornissen
en obstructies ontstaan (9,21).
2.3.5. Enoplida
Massale infecties met Capillaria spp. kunnen leiden tot
een functiestoornis van de darm (9).
2.4. SYMPTOMEN
Zwaar geparasiteerde hagedissen vertonen over het algemeen lage groeicijfers, lagere
reproductiecijfers, en een lagere resistentie tegenover andere ziektes (2).
2.4.1. Oxyuriden
Oxyuriden kunnen zelfs bij hoge infectiegraad asymptomatisch blijven (44). Soms kan men bij zwaar
geïnfecteerde hagedissen echter anorexie, apathie en diarree waarnemen, en treedt er langzaam
gewichtsverlies op. Andere symptomen zoals ademhalingsstoornissen, intoxicatie- en
deficiëntieverschijnselen kunnen aanwezig zijn. Ook cloacaprolaps wordt beschreven (9).
Fig. 5 Microscopisch beeld (40x) adulte Oxyuris sp. bij leguaan (Iguana sp.) (L. Lambrechts)

14
2.4.2. Ascaridida
De symptomen van een zware Ascariden infectie kunnen bestaan uit anorexie, diarree,
gewichtsverlies en obstipatie. Regurgitatie van halfverteerd voedsel wordt tevens beschreven (4,9,10).
Bloedverlies via de darm kan leiden tot symptomen van anemie (9). Bij schade aan grote bloedvaten
kan plotse dood voorvallen. Dit kan ook gebeuren door perforatie van het gastro-intestinale stelsel (9).
2.4.3. Rhabditoidea
De symptomen van een Strongyloiden infectie zijn lethargie, anorexie, regurgitatie, diarree en
steathoree. Ademhalingsstoornissen kunnen ontstaan door larvaire migraties (10,44). Symptomen van
anemie kunnen ook hier aanwezig zijn (9).
2.4.4. Strongylida
Bij een hoge infectiedruk kunnen Strongyliden infecties ernstige ziekte veroorzaken. In zulke situaties
kunnen symptomen voorkomen zoals anorexie, regurgitatie van wormen en ernstige zwakte en
apathie (21). Door de resorptiestoornissen treedt snelle vermagering op. Symptomen van anemie
kunnen ook aanwezig zijn (9).
2.4.5. Gnathostomidae
In het artikel van Heringbeck en Boomker (2000) waar Gnathostomatoidea werd gevonden in de maag
van varanen (Varanus spp.), werd er bij de hagedissen anorexie waargenomen.
2.5. LETSELS
2.5.1. Oxyurida
Oxyuriden veroorzaken zelden letsels (9). Bij
massale infecties kunnen er lesies ontstaan zoals
impactie (1,46).
2.5.2. Ascaridida
Hoge aantallen Ascariden kunnen gastro-intestinale
perforatie en purulente, ulceratieve of sclerotische
inflammatoire lesies veroorzaken (1,9,10,44). Necrotiserende letsels kunnen aanwezig zijn door
migratie of door secundaire bacteriële infecties (34). Perforatie van darmen kan leiden tot septicemie
en bacteriële coelomitis (44). Intestinale obstructie kan aanwezig zijn (47).
2.5.3. Rhabditoidea en Spirurida
Strongyloides spp. en Spirurida larven kunnen tot granulomateuze reacties leiden (4,44).
Fig. 6 Microscopisch beeld (400x) Oxyuris sp. eieren met lensvormige plug (a) bij baardagaam (Pogona vitticeps) (L. Lambrechts)
a

15
2.5.4. Strongylida
Strongyliden kunnen bij massale infectie bij hagedissen ernstige hemorrhagische ulceratie,
inflammatie, perforatie, coelomitis, intussusceptie en obstructie veroorzaken (1,10,42).
2.6. DIAGNOSE
De diagnose van nematoden kan gesteld worden via macroscopische waarneming tijdens necropsie,
natief mestonderzoek of flottatietechnieken voor het opsporen van zowel larven als eieren (2). Larven
kunnen ook aangetoond worden door middel van de Baermann techniek (2). Bij het vinden van eieren
in mestonderzoek moet men er altijd rekening mee houden dat dezen ook afkomstig kunnen zijn van
prooidieren, zonder dat de hagedis zelf geïnfecteerd is (10).
2.6.1. Oxyurida
Eieren van oxyuriden zijn groot (130 bij 40 µm). Ze hebben een dikke gladde wand, zijn langwerpig of
driehoekig en bevatten een lensvormige plug op het uiteinde (2,4,9,10). De adulten bevatten allemaal
een oesofagale bulbus. Ze zijn ongeveer een centimeter lang (9), en de wijfjes hebben een lange
dunne staart (2).
2.6.2. Ascaridida
Ascaridida zijn grote wormen met 3 prominente lippen (2). Ze hebben een gladde en lichte oppervlakte
en kunnen wel tot 17cm lang worden (9). Adulte wormen kunnen aanwezig zijn in geregurgiteerd
voedsel of in mest (10). Hun eieren hebben een dikke dubbele wand, zijn ongeëmbryoneerd en zijn
zo’n 80-100 µm bij 60-80 µm groot. Ascariden infecties kunnen ook gediagnosticeerd worden door
identificatie van geëncysteerde larven in een histologisch preparaat van de mucosa, of door aantonen
van eieren in een maagspoeling (4). Ook is het belangrijk bij Ascariden secundaire orgaanschade te
diagnosticeren, dit kan met behulp van bloedonderzoek, radiografie of echografie (9).
2.6.3. Rhabditoidea
Adulte wormen van Strongyloides zijn zeer klein (2). De
eieren zijn dunwandig en geëmbryoneerd (10).
2.6.4. Strongylida
Strongylida zijn kleine tot middelmatig grote wormen.
Adulten kunnen opgespoord worden in regurgitatie of
mest (21). De eieren zijn ongeëmbryoneerd, dunwandig
en ovaal (10).
Fig. 7 Microscopisch beeld (400x) Capillaria sp. eieren met bipolaire plugs (a) bij baardagaam (Pogona vitticeps) (L. Lambrechts)
a

16
2.6.5. Enoplida
Bij een infectie met Trichinella kunnen de migrerende larven histologisch aangetoond worden in de
maagmucosa (21). Capillaria eieren zijn dikschalig en bevatten bipolaire plugs (4). De wormen zelf zijn
vrij klein en dun (‘haarwormen’) (2,9).
2.7. BEHANDELING
Hoewel lang niet alle nematoden infecties pathologie veroorzaken, is het toch best om dezen te
behandelen. Dit doet de stress dalen in het terrarium en verbetert de levenskwaliteit van de hagedis
(10).
Een bestrijding van een parasiet met een directe levenscyclus is veel moeilijker dan de behandeling
van een parasiet met een indirecte levenscyclus. Bij deze laatste kan men immers de levenscyclus
doorbreken door de tussengastheer weg te nemen bij de eindgastheer. De cyclus van een parasiet
met een directe levenscyclus kan doorbroken worden door te verhinderen dat hagedissen direct
contact kunnen maken met geïnfecteerde soortgenoten of met gecontamineerde mest (2).
De controle en behandeling van nematodeninfectie vindt het beste plaats door een strikte hygiëne toe
te passen in het terrarium en alle dieren gelijktijdig te behandelen, dit doet de reïnfectie het efficiëntst
dalen (44). Geïnfecteerde dieren moeten apart gehuisvest worden gedurende 3 à 4 weken, of tot het
mestonderzoek heeft uitgewezen dat er geen uitscheiding van eieren meer is (9).
Spoelworminfecties kunnen behandeld worden door middel van fenbendazole (25-50 mg/kg per oraal
of per rectam eenmaal per dag gedurende 4 dagen met een herhaling na 10 dagen (1,2) of 100 mg/kg
peroraal eenmalig met een herhaling om de twee weken tot er geen uitscheiding van eieren meer
aanwezig is in de feces (44)).
Ook ivermectine kan toegediend worden (0,2 mg/kg intramusculair, minstens tweemaal met 2 weken
tussentijd) (1). Het is niet het geneesmiddel van eerste keuze omdat het de parasiet veel sneller
afdood, waardoor vele antigenen vrijkomen die voor immuunreacties kunnen zorgen. Dit kan
bijvoorbeeld gegeven worden wanneer de parasiet resistent is aan fenbendazole (10). Ook kan het
toxisch zijn bij kleine hagedissen, kameleons (Chameleo spp.) en skinken (Scincidae spp.) (9).
Daarnaast kan men ook albendazole toedienen (eenmalig 50 mg/kg) (1) of oxfendazole (66 mg/kg
peroraal om de twee weken tot er geen uitscheiding van eieren meer wordt gezien in de mest) (44).
De bestrijding van oxyuriden is niet gemakkelijk, soms berokkent men zelfs meer schade door
toediening van het medicament dan dat de worm zelf berokkend heeft (2). Een behandeling kan de
infectiegraad doen dalen en zware infecties vermijden bij dieren die in gecontamineerde terraria
gehuisvest zitten (44).
Naast de behandeling van de parasitaire infectie, moet men de hagedis ook altijd algemeen
ondersteunen. Ondersteunende voeding kan gegeven worden wanneer deficiëntieverschijnselen
aanwezig zijn. Dit zorgt onder andere ook voor een ondersteuning van de immunologische afweer.
Secundaire verschijnselen, zoals anemie en secundaire bacteriële infecties behoeven ook een
behandeling (9).

17
2.8. PREVENTIE
Preventie van een nematodeninfectie is zeer belangrijk (2). Hygiëne in het terrarium is van primordiaal
belang, ook bij nematoden met een indirecte cyclus zoals Ascariden, omdat zelfs onvolledig
ontwikkelde larven hier massale schade kunnen veroorzaken door migraties (9). Eenmaal per jaar
dienen hagedissen gecontroleerd te worden op infectie (2). Parasitaire controle en behandeling vlak
voor de winterslaap is aan te raden, vooral bij infecties met Oxyuriden, omdat dezen tijdens de
winterslaap intoxicaties kunnen veroorzaken (9). Er wordt aangeraden nieuw aangekochte dieren
gedurende minstens 30 dagen in quarantaine te zetten, alvorens te introduceren aan soortgenoten
(2,9). Bij parasieten met een indirecte levenscyclus is het van belang de hagedissen niet in contact te
laten komen met geïnfecteerde tussengastheren (de prooidieren). Men kan dezen gedurende 30
dagen invriezen alvorens ze te voederen aan de hagedis (2).
3. TREMATODEN EN CESTODEN
3.1. ETIOLOGIE
In het gastro-intestinale stelsel komt slechts één orde van trematoden (platwormen) voor: de Digenea.
De volgende genera komen voor bij hagedissen: Haplometroides, Ochetosoma, Pneumatophilus,
Stomatrema, Zeugorchis (1).
Van cestoden (lintwormen) zijn er meer orden die voorkomen bij hagedissen, namelijk de orden
Proteocephalidea, Pseudophyllidea en Cyclophyllidea (4).
Van de orde Proteocephalidea komen onder andere de genera Proteocephalus, Acanthotaeniae,
Crepidobothrium en Ophiotaenia bij hagedissen voor (2,4).
Ook zijn al enkele genera van de orde Pseudophyllidea beschreven, waaronder Duthiersia,
Scyphocephalus, Bothridium, Bothriocephalus en Spirometra (2,4).
Van de orde Cyclophyllidea zijn volgende families al bij hagedissen beschreven: de Nematotaenidae,
de Anoplocephalidae, de Dilepididae en de Mesocestoididae (2,4).
Een ander phylum, de Acanthocephala, komt ook voor bij reptielen (2).
3.2. EPIDEMIOLOGIE
3.2.1. Trematoden
De trematoden hebben een indirecte levenscyclus. Meestal is de cyclus gebonden aan een waterig
gebied (48). De tussengastheer is meestal een weekdier en de eindgastheer een carnivore vertebraat
(9). In de tussengastheer bevinden zich de geëncysteerde metacercaria (1). Reptielen kunnen ook
tussengastheer zijn van trematoden (9).
3.2.2. Cestoden
Cestoden hebben ook een indirecte levenscyclus (1,49). Cestoden die slechts 1 tussengastheer nodig
hebben, kunnen hun levenscyclus ook in gevangenschap volmaken. Cestoden die echter meerdere

18
tussengastheren nodig hebben in hun cyclus, kunnen dit niet. Ze komen daarom enkel voor bij
hagedissen in de vrije natuur, of bij wildvang hagedissen (9).
3.2.2.1. Proteocephalidea
De Proteocephalidea worden eerst opgenomen door aquatische Crustacea, waarin ze evolueren tot
procercoid en verder tot plerocercoid. Deze infectieuze stadia worden opgenomen door de
eindgastheer waarbij ze adult worden in de dunne darm na vasthechting aan de mucosae (2,4).
3.2.2.2. Pseudophyllidea
De Pseudophyllidea hebben ook een Crustacea als tussengastheer, waarin ze ontwikkelen tot de
procercoïden, hierna worden ze opgenomen door een tweede tussengastheer (een vertebraat) waarin
ze ontwikkelen tot plerocercoid, ook wel het spargana genoemd. Dit spargana wordt opgenomen door
een carnivore eindgastheer (2,4). Het genus Spirometra maakt soms ook gebruik van de hagedis als
tussengastheer (9).
3.2.2.3. Cyclophyllidea
Bij Cyclophyllidea zoals de Dilepididae en de Mesocestoididae is het reptiel tussengastheer.
Mesocestoididae gebruiken artropoden als eerste tussengastheer, waarin ze ontwikkelen van
oncosfeer tot cysticercoid. Daarna worden ze opgenomen door een tweede tussengastheer, dit kan
een hagedis zijn, hierbij ontwikkelen ze verder tot tetrathyridium. Deze tussengastheer wordt dan
opgenomen door de eindgastheer, een vogel of een zoogdier, waarin de parasiet adult wordt (4).
3.2.3. Acanthocephala
De Acanthocephala komen als volwassen worm voor in de dunne darm. Ze hebben een arthropode
als tussengastheer, en de hagedis kan eventueel optreden als paratenische gastheer. In dit laatste
geval zitten de larven geëncysteerd in de abdominale holte (2).
3.3. PATHOGENESE
De meeste trematoden zijn weinig pathogeen (1).
Cestoden worden meestal per toeval aangetroffen bij necropsie. In aanwezigheid van een groot aantal
wormen in de darm kan er echter pathologie ontstaan (4). De orden Proteocephalidea en
Pseudophyllidea kunnen pathogeen zijn wanneer er een hoge infectiestatus bereikt is, doordat ze zich
met hun scolex vasthechten aan de mucosa van de darm (2,9). Ze kunnen ook obstructies
veroorzaken of in competitie gaan voor voedingsstoffen (4). Wanneer de hagedis optreedt als
tussengastheer voor de cestode, kan deze laatste zich in verschillende organen inkapselen, zoals de
lever, darmen, coeloomholte of mesenteria. Door onder andere mechanische schade, kan dan op
deze plaatsen orgaandysfunctie ontstaan (4,50).
De groep van de Acanthocephala bevat een proboscis met tanden die in de mucosae kunnen
vasthechten (2).
19
3.4. SYMPTOMEN
Bij zware infecties met trematoden kan er anorexie
ontstaan, een daling in de vitaliteit en groeistoornissen
(9).
De cestoden die in de darm vertoeven, veroorzaken
zelden klinische ziekte, behalve bij hoge infectiegraad.
Wanneer dieren in een slechte conditie verkeren,
kunnen er symptomen ontstaan van
voedingsstoftekorten (2). Vitamine B tekorten worden
vermeld bij massale infecties, waardoor
coördinatiestoornissen zich zouden kunnen ontwikkelen. Een hogere voedselopname kan opvallen,
hoewel de dieren in gewicht verliezen. Daarnaast kunnen de dieren last hebben van diarree en
braken. Hagedissen die als tussengastheer fungeren kunnen symptomen vertonen zoals anorexie en
gewichtsverlies (9).
3.5. LETSELS
Trematoden kunnen darmschade veroorzaken bij
massale infectie (9).
Wanneer volwassen cestoden in grote aantallen in de
darm aanwezig zijn, kan secundaire malnutritie
ontstaan, enteritis door mechanische irritatie, obstructie
en impactie (10).
De orde Proteocephalidea kan bij hagedissen
intestinale necrose, epitheliaal verlies en een
celinfiltraat in de tunica muscularis veroorzaken (1).
De orde Pseudophyllidea kan ulceratie van de
intestinale mucosae, hemorrhagieën en oedemen
veroorzaken (1).
Als de hagedis tussengastheer is, kan ontsteking
aanwezig zijn op de plaats van de cyste (2). Zo
beschreef Goldberg (1985) witte nodules in onder
andere lever, darmen, coeloomholte en mesenteria van
de eilandnachthagedis (Xantusia riversiana) na infectie
door Mesocestoides. De parasiet veroorzaakte lichte
ontstekingsreacties, en zat geëncysteerd in een dun
fibreus kapsel.
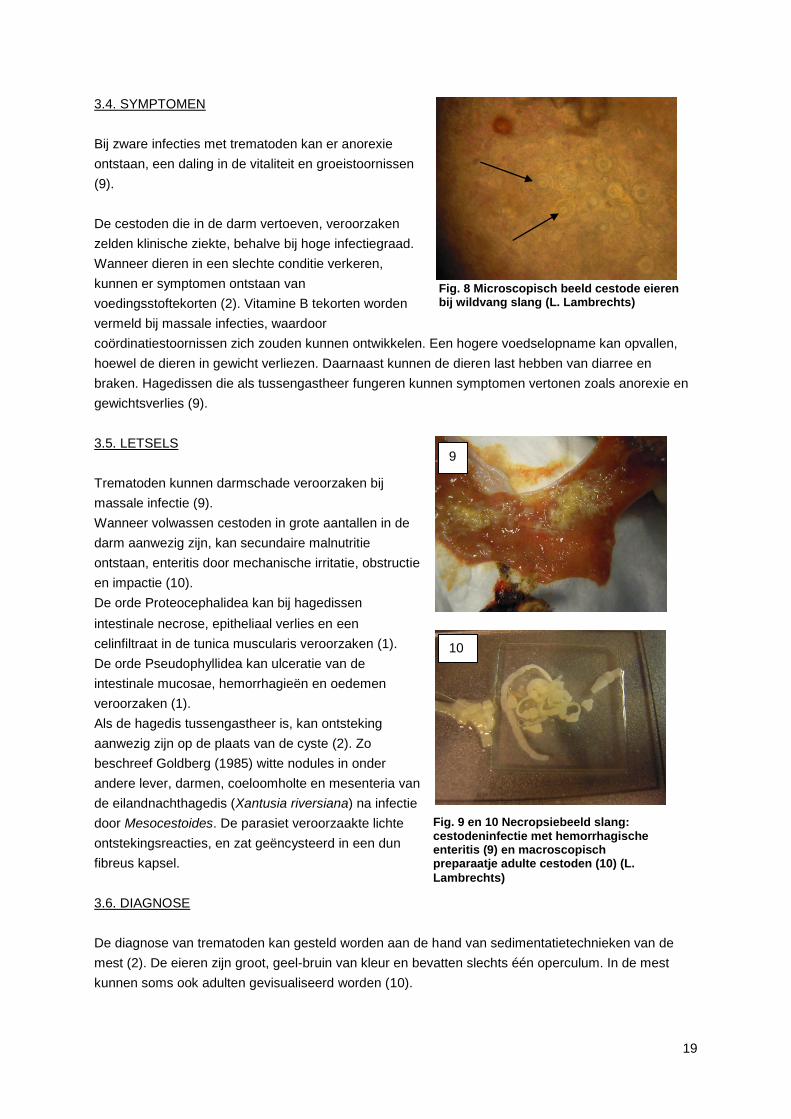
3.6. DIAGNOSE
De diagnose van trematoden kan gesteld worden aan de hand van sedimentatietechnieken van de
mest (2). De eieren zijn groot, geel-bruin van kleur en bevatten slechts één operculum. In de mest
kunnen soms ook adulten gevisualiseerd worden (10).
Fig. 9 en 10 Necropsiebeeld slang: cestodeninfectie met hemorrhagische enteritis (9) en macroscopisch preparaatje adulte cestoden (10) (L.
Lambrechts)
9
10
Fig. 8 Microscopisch beeld cestode eieren bij wildvang slang (L. Lambrechts)

20
Cestoden worden ook gediagnosticeerd aan de hand van mestonderzoek. Soms kunnen de adulte
lintwormen direct geïdentificeerd worden. Ook de proglottiden en de eieren kunnen teruggevonden
worden in de mest. De meest gebruikte technieken zijn sedimentatie en flottatie (2,4). De eieren van
Proteocephalidea bevatten volledig gevormde oncosferen met 6 haken, en zijn tussen 50 bij 40 µm
groot (2,4). De adulte wormen van Pseudophyllidae zijn tussen de 10 en de 80 cm groot. De cestoden
van de orde Cyclophyllidea kunnen gediagnosticeerd worden aan de hand van identificatie van de
cysten (2).
3.7. BEHANDELING
3.7.1. Trematoden
Trematodeninfecties kunnen behandeld worden door praziquantel toe te dienen 8 mg/kg peroraal of
subcutaan 2 of 3 maal met telkens een tussenperiode van 2 weken (1). Ronidazole kan ook
toegediend worden (9). Daarnaast is ook albendazole effectief (1). Een controle van effectiviteit is
gewenst na 3 à 4 weken (9).
3.7.2. Cestoden
Ook cestoden infecties kunnen behandeld worden door middel van praziquantel, volgens hetzelfde
schema als bij de trematoden. De mest dient na 4 tot 6 weken gecontroleerd te worden, aangezien
eieren resistent zijn aan deze behandeling. Geëncysteerde larven kunnen chirurgisch worden
verwijderd (2,9).
3.8. PREVENTIE
Bij infectie van één dier, moeten de andere dieren uit dezelfde omgeving ook onderzocht worden op
infectie (9). Omdat cestoden en nematoden parasieten zijn met een indirecte cyclus, is het van belang
de hagedissen niet in contact te laten komen met geïnfecteerde tussengastheren. Verder zijn een
jaarlijkse parasitaire controle en quarantaine van nieuw aangekochte dieren aangewezen (zie hoger).
4. ADENOVIRUS
4.1. ETIOLOGIE
Adenovirus is een virus zonder enveloppe, van 70-90 nm in diameter. Replicatie vindt plaats in de
celkern. Geaccumuleerde viruspartikels vormen zo inclusielichamen (51). Bij hagedissen komt
voornamelijk het genus Atadenovirus voor (52,53).
4.2. EPIDEMIOLOGIE
Adenovirus kan bij reptielen infecties veroorzaken met een hoge morbiditeit en mortaliteit. Er zijn al
verschillende uitbraken geobserveerd van Adenovirus bij baardagamen (Pogona vitticeps). Sinds
1990 is het een opkomend probleem in de Verenigde Staten van Amerika, en nu is het er endemisch
aanwezig onder de baardagamen. Er wordt gesuggereerd dat infectie plaatsvindt door oro-fecaal
contact omdat de meeste lesies gevonden worden in het gastro-intestinale stelsel en de lever.

21
Aangezien adulten persisterend subklinisch geïnfecteerd kunnen zijn, zouden ze als reservoir kunnen
optreden voor jonge hagedissen (51). Ook een verticale transmissie wordt niet uitgesloten (54,55).
Daarnaast wordt een natuurlijk verworven infectie ook mogelijk geacht (56).
4.3. PATHOGENESE
Meestal is het virus geadapteerd aan de gastheer, en ontstaat er weinig pathologie. Pathologie kan
echter wel ontstaan bij jonge hagedissen, of bij een verminderde afweer, bijvoorbeeld door
samenlopende parasitaire, bacteriële, fungale, of virale ziekte of stress (57). Zo is Adenovirus al
samen gediagnosticeerd met onder andere parasitaire infecties zoals Coccidia, Nematoden en
Microsporidia (58,59,60) en het Dependovirus (55,59).
4.4. SYMPTOMEN
Symptomen komen vaak acuut voor bij jonge hagedissen, en worden vaak gevolgd door sterfte (51).
Symptomen die al beschreven zijn, zijn onder andere groeistop/groeivertraging (54,60), anorexie
(54,58,56,61), gewichtsverlies (52), apathie (58), distentie coeloomholte (58), parese (54), head tilt
(59), cirkelgang (59), opisthotonus (59,61), sterfte zonder voorafgaande symptomen (55,60) en
regurgitatie (52). Subklinische infectie is echter ook al beschreven (56).
4.5. LETSELS
Macroscopische letsels die al gevonden zijn bij infecties met adenovirus zijn onder andere oesofagitis
(61,62), gastritis (52), enteritis (52) en hepatitis (54,60). Jacobson et al. (1996) beschreven
hemorrhagieën in de coeloomholte, de darmen en perirenaal en een vergrote lever die diffuse
geelrode vlekken bevatte. Hyndman en Shilton beschreven in 2011 gelijkaardige letsels op de lever.
Microscopisch werden eosinofiele of basofiele intranucleaire inclusielichamen gezien in epitheel en
mucosa van slokdarm, maag, darm en lever in de respectievelijke artikels. Een proliferatie van het
epitheel werd gevonden bij de case reports die oesofagitis en gastritis beschreven.
4.6. DIAGNOSE
Antemortem diagnose kan gesteld worden met behulp van orale of cloacale swabs of nasale
spoelingen (63). Ook in meststalen kan men trachten het virus terug te vinden (51). De diagnose kan
ook gesteld worden op biopsies van lever, maag, slokdarm en nier. Biopsies moeten genomen
worden van weefsels die er morfologisch ongezond uitzien. Op deze weefselstalen kan men
histopathologie doen, waar bij infectie de inclusielichamen kunnen worden gezien (63). Postmortem
kan men deze inclusies onder andere zoeken in galgangen, lever, pancreas, nier, gastro-intestinale
epitheel, kupfercellen, miltmacrofagen en endotheel (63). De diagnose kan bevestigd worden door
elektronenmicroscopie, in situ hybridisatie of PCR (52,53,60,63). Een positieve isolatie wijst op een
infectie, een negatief resultaat sluit geen infectie uit (54).
4.7. BEHANDELING
Voor deze virale infectie bestaat er geen specifieke behandeling. Men kan enkel de patiënt proberen
te ondersteunen, bijvoorbeeld door vochttoediening, antibioticum tegen secundaire bacteriële

22
infecties, vermindering van stress, een optimale temperatuur en vochtigheid en geassisteerde voeding
(57,58). Een eventuele onderliggende predisponerende oorzaak moet ook behandeld worden (57).
De omgeving moet ook behandeld worden. Adenovirus is een resistent virus. Het is resistent
tegenover hitte, organische solventen en pH extremen. Ook vele desinfectantia zijn niet werkzaam.
Inactivatie kan gebeuren door middel van meer dan 1 uur contact met formaline, aldehydes of
jodoforen (51).
4.8. PREVENTIE
Aangezien er nog weinig gekend is over de epidemiologie van dit virus, is preventie van groot belang.
Opruiming of strikte quarantaine van geïnfecteerde dieren moet toegepast worden. Nieuw
aangekochte dieren dienen ook minstens 6 maanden in quarantaine te verblijven, om subklinisch
geïnfecteerde dieren te onderkennen. Dezen kunnen opgespoord worden door middel van de PCR
techniek. Ook een goede hygiëne moet toegepast worden omdat het virus lang infectieus kan blijven
in de omgeving (51,57).
5. BACTERIËN EN FUNGI
5.1. ETIOLOGIE
De bacteriën die uit lesies van enteritis bij reptielen geïsoleerd kunnen worden, hebben meestal een
gram-negatieve spectrum. Zo zijn er onder andere al kiemen uit letsels geïsoleerd zoals E. coli,
Klebsiella, Salmonella, Enterococcus, Pseudomonas, Serratia, Proteus, Citrobacter, Alcaligines en
Pasteurella. Gram-positieve bacteriën worden soms ook geïsoleerd, zoals Clostridium,
Corynebacterium en Staphylococcus. Ook zuurvaste kiemen zoals Mycobacteria zijn beschreven (62).
Volgende fungi zijn al beschreven in het gastro-intestinale stelsel van reptielen: Chrysosporium
keratinophilum (64), Penicillium (44,65), Basidiobolus spp. (44,65), Paecilomyces (44), Candida,
Aspergillus, Fusarium en Mucor spp. (65).
5.2. PATHOGENESE
Er is weinig bekend over het primaire belang van bacteriën en fungi in het gastro intestinale kanaal bij
reptielen. Ze treden wel frequent op als secundaire pathogenen (1,9). Door bepaalde factoren, zoals
een onaangepast dieet, antibioticamisbruik, stress of andere infecties, kan er een onevenwicht
ontstaan in de intestinale flora van hagedissen. Hierdoor kan maldigestie ontstaan, ileus, diarree of
anorexie. Op zulke momenten kunnen opportunistische bacteriën het gastro-intestinale stelsel
koloniseren (62). Een hoge vochtigheid, malnutritie, overbevolking, onhygiënische omstandigheden,
overmatig antibiotica gebruik zijn tevens allen predisponerende factoren voor een fungale infectie (44).
5.3. DIAGNOSE
De diagnose van een fungale of bacteriële superinfectie steunt op cultuuronderzoek of histopathologie
van de waargenomen letsels. Histopathologie kan uitgevoerd worden op afdrukpreparaatjes of
biopsies (1).

23
5.4. BEHANDELING
Bij reptielen die lijden aan gastro-enteritis, kan een antibioticum toegediend worden, tegen eventuele
secundaire bacteriële infecties (9). Nystatine is een antimycoticum dat kan aangewend worden bij
intestinale infectie. Het wordt niet systemisch opgenomen (44). Indien er ook systemische infectie
aanwezig is kan men gebruik maken van onder andere ketoconazole, itraconazole, fluconazole,
fluorocytosine of amfotericine (66). Chirurgische interventie is soms aangeraden bij fungale infecties
om recidieven te voorkomen (1).
Om de intestinale flora terug te herstellen kan men sondevoeding geven, transfaunatie uitvoeren, of
probiotica toedienen, eventueel lactobacillus bevattend (44).
6. NIET INFECTIEUZE OORZAKEN
6.1. ETIOLOGIE, EPIDEMIOLOGIE EN PATHOGENESE
6.1.1. Toxines
Toxines kunnen bij hagedissen gastritis of enteritis veroorzaken (1). Toxines kunnen opgenomen
worden bij een onaangepast dieet. Ze kunnen bijvoorbeeld aanwezig zijn in insecten. Zo kunnen
vuurvliegjes (Photinus spp.) bijvoorbeeld toxines produceren, steroïdale pyronen, die mogelijk zeer
toxisch kunnen zijn voor hagedissen. Soms kan zelfs ingestie van 1 vuurvliegje fataal zijn (67). Ook
pesticiden en herbiciden kunnen potentieel voor toxiciteit zorgen bij hagedissen (1).
6.1.2. Impactie en obstructie
Vreemde voorwerpen zijn zeer vaak oorzaak van gastro-intestinale ziekte bij hagedissen. Dezen
kunnen leiden tot impactie en obstipatie (44,62). Zo kunnen bijvoorbeeld opgenomen substraten
leiden tot obstructie. Dit komt geregeld voor bij kleine insectivore hagedissen die op een zand of kiezel
substraat gehuisvest zitten (1). Ook een slechte calcium supplementatie, een onaangepaste
vochtigheidsgraad of onvoldoende toegang tot drinkwater kan tot deze pathologie bijdragen (44).
6.1.3. Intussusceptie
Ook intussusceptie van de darmen kan leiden tot enteritis. Het is al beschreven bij groene leguanen
(Iguana iguana), kameleons (Chameleo spp.) en een blauwtongskink (Tiliqua scincoides) (62).
Fig. 11 en 12 Impactie bij luipaardgekko (Eublepharis macularius), veroorzaakt door zand als bodembedekking (L. Lambrechts)
11 12

24
6.1.4. Cloacaprolaps
Bij prolaps van de cloaca kan trauma en contaminatie optreden. De prolaps kan ontstaan door een
constipatie of tenesmus. Dit komt onder andere voor bij dystocia, overvoeding, trichobezoaren,
neoplasieën, parasitisme, onaangepaste temperaturen of urolithen. Het kan ook ontstaan door een
drukstijging in de coeloomholte, door bijvoorbeeld ruimte innemende processen, nefromegalie,
obesitas of een pneumocoeloom. Ook een cloacitis of een hypocalcemie kan leiden tot een prolaps
(44). Omdat de veneuze retour van het geprolabeerde deel vaak geïnhibeerd wordt, zet het weefsel
snel uit, en wordt het zeer oedemateus en fragiel (1).
6.1.5. Neoplasie
Gastro-intestinale neoplasie is ook in belang aan het
toenemen bij hagedissen (68,69). Er is een mogelijkheid
dat een C-type oncorna virus een aandeel heeft in het
ontstaan van neoplasieën bij reptielen. Andere mogelijke
carcinogenen zijn toxines, genetica, parasitisme, fysische
agentia, trauma en radiatie, maar voor geen van dezen
zijn er bewijzen (68).
6.1.6. Andere
Een onaangepast dieet, of verrotting van voedsel in de maag na het verblijf in een omgeving met een
suboptimale postprandiale temperatuur, kan aanleiding geven tot gastritis (44). Daarnaast kan acute
gastritis ook ontstaan door thermale wonden, chirurgie, ontstekingsremmers en corticosteroïden.
Enteritis kan daarnaast veroorzaakt worden door antibioticamisbruik of stress (1).
6.2. SYMPTOMEN
Bij opname van vreemde voorwerpen kunnen acute symptomen ontstaan zoals lethargie, anorexie en
dehydratatie. Regurgitatie kan optreden als het vreemde voorwerp proximaal van de dunne darm
gelokaliseerd zit. Ook een chronisch verloop is mogelijk. Hierbij zal de hagedis langzaam aan
vermageren, lethargisch zijn, anorexie vertonen en geconstipeerd zijn. Het dier zal na een tijd
cachectisch worden en dehydratatie vertonen. Melena, hematoschezie en pijn tijdens het eten,
kunnen opgemerkt worden (1).
De symptomen van een gastro-intestinale tumor hangen af van de locatie en van het type van de
tumor. Zo kan onder andere emesis, hematemesis, vertraagde groei, anorexie, opzwellen van de
coeloomholte, melena, een verminderde mogelijkheid tot flexie van de achterhand, asymmetrische
cloacavergroting en constipatie aanwezig zijn. De structuren kunnen ook palpeerbaar zijn in de
coeloomholte (1,62).
6.3. DIAGNOSE
De diagnose van een vreemd voorwerp kan gesteld worden door middel van medische beeldvorming
technieken. Radiografie voor minerale materialen, contrast radiografie, echografie, CT-scan, MRI zijn
de verschillende mogelijkheden (1). Een maagspoeling kan ook tot de diagnose leiden (44).
Fig.13 Cloacaprolaps groene leguaan (Iguana iguana) (F. Pasmans)

25
Bloedonderzoek kan ook een indicatie geven. Anemie kan gevonden worden bij gastro-intestinale
hemorrhagieën, evenals leukocytose, een verhoging in lever- en spierenzymes en
elektrolytstoornissen (1). Tenslotte kan eventueel een coeliotomie uitgevoerd worden (44).
De diagnose van een gastro-intestinale tumor kan ook gesteld worden aan de hand van medische
beeldvorming. Een bevestiging van de diagnose kan behaald worden door histopathologie uit te
voeren op een dunnenaaldaspiraat of een biopsie (1).
6.4. BEHANDELING EN PREVENTIE
Bij een obstipatie is preventie van groot belang. Hier moet men een optimale huisvesting en
management nastreven (1,44). Bij de behandeling kan men eerst trachten een niet-invasieve methode
toe te passen, zoals het geven van lauwe baden, massage van het distale colon, toediening van
lactulose via een maagsonde (0,5 ml/kg per dag) of toediening van cisapride of metoclopramide.
Indien er een volledige obstructie aanwezig is, een cloacolith of een intussusceptie, dient er
chirurgisch ingegrepen te worden (44). Een vreemd voorwerp kan via chirurgie of onder
endoscopische begeleiding verwijderd worden (1). Daarnaast moet een mogelijke onderliggende
oorzaak aangepakt worden, bijvoorbeeld door optimalisatie van voeding en omgeving en eventueel
een antiparasitaire behandeling (44).
Bij de behandeling van een prolaps is het corrigeren van de onderliggende oorzaak zeer belangrijk.
Daarnaast dient men een repositie uit te voeren met behulp van een wateroplosbaar glijmiddel.
Eventueel kan men eerst trachten het volume van de prolaps te verkleinen door middel van glycerine
of een geconcentreerde suikeroplossing. Om recidieven te voorkomen, kan men een tijdelijke
beursnaad of een transversale cloacale hechting plaatsen. Soms, als repositie niet meer mogelijk is, is
amputatie geïndiceerd (1,44).
De behandeling van een tumor is afhankelijk van de locatie, de grootte en het type van de tumor.
Indien mogelijk, wordt de tumor geëxciseerd met een grote marge. Ook cryotherapie of
elektrochirurgie behoren tot de mogelijkheden (44). Men heeft al studies ondernomen in verband met
radiatietherapie en chemotherapie bij reptielen. Verder onderzoek is hierbij nodig. Cisplatine is
effectief tegen fibrosarcoma’s (1).
Fig. 14 en 15 RX opname van Zwart-wit Tegu (Tupinambis merianae) : impactie (14) en dezelfde Tegu na verwijdering veroorzakend agens (houtstukjes en hennepvezels) (15) (L. Lambrechts)
14 15

26
BESPREKING
Als er in de praktijk een hagedis aangeboden wordt met een verminderde algemene conditie en
symptomen zoals anorexie of polyfagie, diarree, regurgitatie, constipatie, prolaps of gewichtsverlies
moet men steeds de mogelijkheid van gastro-enteritis in acht nemen. Er moet echter ook op gewezen
worden dat niet elke hagedis die een symptoom vertoont van gastro-intestinale ziekte, ook effectief
gastro-enteritis heeft. Een hagedis kan bijvoorbeeld anorexie vertonen doordat hij gehuisvest is in een
suboptimale omgeving (62).
Om van de differentiaaldiagnose tot de diagnose te komen, is van primordiaal belang dat eerst het
signalement en de anamnese goed en uitgebreid aan bod komen. Het signalement is vooral belangrijk
om de leeftijd en herkomst van het dier te achterhalen. Zo zijn bijvoorbeeld protozoa zoals Isospora
spp. een zeer belangrijke differentiaaldiagnose bij juveniele baardagamen. De herkomst is belangrijk
omdat bijvoorbeeld trematoden en cestoden meestal enkel voorkomen bij wildvang dieren. In de
anamnese moeten zeker de huisvestingsfactoren uitgebreid aan bod komen. Ook al is een infectieuze
oorzaak soms de doorslaggevende factor in de pathologie, toch speelt de huisvesting hier vaak als
predisponerende factor een niet te onderschatten rol. Daarnaast zijn niet-infectieuze agentia ook
frequent de primaire oorzaak van gastro-enteritis. Naast parasitose is obstructie door een vreemd
voorwerp immers de meest voorkomende oorzaak van gastro-enteritis bij reptielen, en vaak is deze
veroorzaakt door fouten in het management (62).
In het algemeen klinisch onderzoek mag men zeker niet vergeten een natief mestpreparaatje
microscopisch te bekijken, want van de infectieuze agentia zijn parasieten meestal de oorzaak van de
gastro-enteritis. In natuurlijke omstandigheden zijn alle reptielen geparasiteerd. De meeste parasieten
zijn niet schadelijk, en dienen dus in principe ook niet behandeld te worden. In gevangenschap
kunnen echter lage graden van infectie leiden tot ziekte. Door overbevolking, stress en malnutritie, kan
er een onevenwicht ontstaan tussen parasiet en gastheer, met opflakkering van de parasitaire infectie
tot gevolg. Ook ziekte leidt tot een verminderde weerstand en stress, waardoor een lichte parasitaire
infectie significant kan worden, en bijdragen tot een meer uitgesproken ziektebeeld. Daarom is het
toch aangeraden parasitaire infecties bij hagedissen die in gevangenschap leven, te behandelen, en is
preventie ervan zeer belangrijk. Een jaarlijkse check-up is voor deze reden ook aan te raden (2).
Als zowel de niet-infectieuze als de parasitaire oorzaken uit de differentiaaldiagnose geschrapt zijn,
kan men nog steeds denken aan een virale oorsprong van de gastro-enteritis. Hierbij is adenovirose
als enige significante aandoening bekend. Daarnaast mag men ook fungi en bacteriën niet vergeten,
maar of dezen als primaire pathogenen gastro-intestinaal lijden veroorzaken, is nog niet gekend.
Qua behandeling dient men naast de etiologische bestrijding ook voldoende aandacht te besteden
aan een ondersteunende therapie. Hagedissen die in de praktijk als ziek aangeboden worden,
kampen vaak met dehydratatie en vermagering. Een antiparasitaire behandeling is zoals reeds
vermeld ook aan te raden als deel van een ondersteunende behandeling. Antibiotica kunnen
toegediend worden bij gastro-enteritis tegen secundaire bacteriële infecties.
Na een uitgebreid onderzoek, een adequate diagnose en een aangepaste behandeling, dient de
eigenaar dus zeker met vele tips naar huis gestuurd te worden omtrent de verzorging van zijn hagedis.
Het besluit is immers dat vele oorzaken van gastro-enteritis bij hagedissen vermeden kunnen worden
door een voldoende kennis van het management van de hagedis.

27
REFERENTIELIJST
1. Diaz-Figueroa O., Mitchell M.A. (2006). Gastrointestinal anatomy and physiology. In: Mader D.R.
(Editor) Reptile medicine and surgery, second edition. Elsevier, California, p.145-162.
2. Greiner E.C., Mader D.R. (2006). Parasitology. In: Mader D.R. (Editor) Reptile medicine and
surgery, second edition. Elsevier, California, p. 343-364.
3. Koudela B., Modry D. (1998). New species of Cryptosporidium (Apicomplexa: Cryptosporidiidae)
from lizards. Folia Parasitol 45, p. 93-100.
4. Jacobson E.R. (2007). Parasites and Parasitic Diseases of Reptiles. In: Jacobson E.R. (Editor)
Infectious diseases and pathology of reptiles. CRC Press, Florida, p. 571-666.
5. Appeltans W., Bouchet P., Boxshall G.A., De Broyer C., de Voogd N.J., Gordon D.P., Hoeksema
B.W., Horton T., Kennedy M., Mees J., Poore G.C.B., Read G., Stöhr S., Walter T.C., Costello M.J.
(2012). World register of Marine Species. Internetreferentie: http://www.marinespecies.org
(geconsulteerd op 22 maart 2012).
6. Atkinson C.Y., Ayala S.C. (1987). Isospora manchacensis n. sp., an intranuclear coccidian from the
Louisiana ground skink, Scincella lateralis (Say, 1823) (Lecertilia: Scincidae). J Parasitol 73, p. 817-
823.
7. Modry D., Daszak P., Volf J., Vessely M., Ball S.J., Koudela B. (2001a). Five new species of
coccidian (Apicomplexa: Eimeriidae) from Madagascan chameleons (Sauria: Chamaelonidae). Syst
Parasitol 48, p. 117-123.
8. Matuschka F.R. (1987). Reptiles as intermediate and/or final hosts of Sarcosporidia. Parasitol Res
73, p. 22-332.
9. Schneller P., Pantchev N. (2008). Parasitologie bei Schlangen, Echsen und Schildkröten, Ein
Handbuch für die Reptielenhaltung. Edition Chimaira, Frankfurt Am Main, p. 32-33, 35, 74-113,121-
179.
10. Klingenberg R.J. (2004). Parasitology. In: Girling S.J., Raiti P. (Editors) Manual of reptiles, Second
edition. British Small Animal Veterinary Association, Waterwells, p. 319-329.
11. Beck W., Pantchev N. (2006). Praktische Parasitologie bei Heimtieren. Schlütersche
Verslagsgesellschaft mbH & Co. KG, Hannover, p. 229-293.
12. O’Donoghue P.J. (1995). Cryptosporidium and cryptosporidiosis in man and animals. Int J
Parasitol 25, p. 139-195.
13. Oros J., Rodriguez J.L., Patterson-Kane J. (1998). Gastric cryptosporidiosis in a Wild Frilled Lizard
from Australia. Journal of Wildlife diseases 34(4), p.807-810.
14. Klingenberg R.J. (1996). Enteric cryptosporidiosis in a colony of indigo snakes, Drymarchon corais
spp., a panther chameleon, Chameleo pardalis, and a Savannah monitor, Varanus
exanthematicus. Bull Assoc Reptil Amphib Vet 6, p. 5-9.
15. Taylor M.A., Geach M.R., Cooley W.A. (1999). Clinical and pathological observations on natural
infections of cryptosporidiosis and flagellate protozoa in leopard geckos (Eublepharis macularius).
Vet Rec 145, p. 695-699.
16. Upton S.J., McAllister C.T., Freed P.S., Barnard S.M. (1989). Cryptosporidium spp. in wild and
captive reptiles. J Wildl Dis 25, p. 20-30.
17. Graczyk T.K., Cranfield M.R., Bostwick E.F. (1999a). Hyperimmune bovine colostrums treatment
of moribund Leopard geckos (Eublepharis macularius) infected with Cryptosporidium sp. Vet Res
30(4), p. 377.
18. Cranfield M.R., Graczyk T.K. (2000). Cryptosporidia in reptiles. In: Bonagura J.D. (Editor) Kirk’s
current veterinary therapy XIII small animal practice, WB Saunders, London, p. 387-394.

28
19. Cranfield M.R., Graczyk T.K. (2006). Cryptosporidiosis. In: Mader D.R. (Editor) Reptile medicine
and surgery, second edition. Elsevier, California, p. 756-762.
20. Cranfield M.R., Graczyk T.K., Wright K., Frye F.L., Raphael B. (1999a). Cryptosporidiosis. Bull
Assoc Reptl Amphib Vet 9(3), p. 15.
21. Hernandez-Divers S.J. (2006) Reptile Parasites- Summary Table. In: Mader D.R.
(Editor) Reptile medicine and surgery, second edition. Elsevier, California, p. 1159-1170.
22. Frank W. (1984). Non-hemoparasitic protozoans. In: Hoff G., Frye F., and Jacobson E. (Editors),
Diseases of Amphibians and Reptiles, Plenum Press, New York, p. 259-384.
23. McAllister C.T., Upton S.J., Jacobson E.R., Kopit W. (1995). A description of Isospora amphiboluri
(Apicomplexa: Eimeriidae) from the inland bearded dragon, Pogona vitticeps (Sauria: Agamidae).
J Parasitol 81, p. 281-284.
24. Coke R.L., Tristan T.E. (1998). Cryptosporidium infection in a colony of leopard geckos,
Eublepharis macularius. Proceedings of the Association of Reptilian and Amphibian Veterinarians
5th Annual Meeting, Kansas City, MO, p. 157-165.
25. Terrell S.P., Uhl E.W., Funk R.S. (2003). Proliferative enteritis in leopard geckos (Eublepharis
macularius) associated with Cryptosporidium sp. infection. J Zoo Wildl Med 34, p. 69-75.
26. Richter B., Nedorost N., Maderner A., Weisenböch H. (2011). Detection of Cryptosporidium
species in feces or gastric contents from snakes and lizards as determined by polymerase chain
reaction analysis and partial sequencing of the 185 ribosomal RNA gene. Journal of Veterinary
Diagnostic Investigation 23(3), p. 430-435.
27. Frank W., (1985). Amphibien und Reptilien. In: Isenbügel E., Frank W. (Editors)
Heimtierkrankheiten, Ulmer, Stuttgart, p. 161-402.
28. Zwart P., Tennis S.F.M., Cornelissen J.M.M. (1983). Monocercomoniasis in reptiles. In: Vago
C., Matz G. (Editors) Proceedings of the First International Colloquium on Pathology of Reptiles
and Amphibians, Les Presses de l’Université d’Angers, Angers, p. 73-76.
29. Terrell S.P., Stacy B.A. (2007). Reptile necropsy techniques. In: Jacobson E.R. (Editor) Infectious
diseases and pathology in reptiles, Color atlas and text. CRC Press, Boca Raton, p. 219-256.
30. Modry D., Slapeta J.R., Knotek Z., Koudela B. (2001b). Caryospora varaniornati sp. n.
(Apicomplexa: Eimeriidae) in the Nile monitor, Varanus (Polydaedalus) niloticus species complex.
Folia Parasitol, Praha 48, p. 7-10.
31. Greiner E.C. (2003). Coccidiosis in reptiles. Semin Avian Exotic Pet Med 12, p. 49-56.
32. Zwart P. (1973). Ziekten van reptielen V: infectieziekten. Lacerta 31, p. 116-120.
33. Kojimoto A., Uchida K., Horii Y., Okumura S., Yamaguchi R., Tateyama S. (2001). Amebiasis
in four ball pyhons (Python regius). J Vet Med Sci 63, p. 1365-1368.
34. Stacy B.A., Pessier A.P. (2007). Host Response to Infectious Agents and Identification of
Pathogens in Tissue Section. In: Jacobson E.R. (Editor). Infectious diseases and pathology in
reptiles, Color atlas and text. CRC Press, Boca Raton, p. 257-298.
35. Graczyk T.K., Cranfield M.R., Fayer R. (1996). Evaluation of commercial enzyme immunoassay
(EIA) and immunofluorescent antibody (IFA) test kits for detection of Cryptosporidium oocysts
other than Cryptosporidium parvum. AM J Trop Med Hyg 54(3), p. 274.
36. Modry D., Sloboda M. (2004). Control of Coccidiosis in Chameleons Using Toltrazuril – Results
of an Experimental Trial. Proceedings of the 7th International Symposium on Pathology and
Medicine in Reptiles and Amphibians, Berlin, Edition Chimaira, p.100-101.
37. Bogoslavsky B.A. (2007). The use of ponazuril to treat coccidiosis in eight inland bearded dragons
(Pogona vitticeps). Proceedings Association of Reptilian and Amphibian Veterinarians, ARAV
Fourteenth Annual Conference, p.8.

29
38. Cranfield M.R., Graczyk T.K., Bostwick E.F. (1999b). A comparative assessment of the therapeutic
efficacy of hyperimmune bovine colostrums treatment against Cryptosporidium infections in
Leopard geckos (Eublepharius macularius) and the Savanna monitors (Varanus exanthematicus).
In: Willette M.M. (Editor) Proceedings of the Association of Reptilian and Amphibian Veterinarians,
Colombus, Ohio, p. 119-121.
39. Graczyk T.K., Cranfield M.R., Bostwick E.F. (1999b). Therapeutical efficacy of hyperimmune
bovine colostrum therapy against cryptosporidium infections in reptiles. In: Baer C.K. (Editor)
Proceedings of the American Association of Zoo Veterinarians, Colombus, Ohio, p. 6-10.
40. Graczyk T.K., Cranfield M.R., Bostwick E.F. (2000). Successful hyperimmune bovine colostrums
treatment of Savanna monitors (Varanus exanthematicus) infected with Cryptosporidium sp. J
Parasitol 86(3), p. 631.
41. Campbell I., Tzipori S. (1982). Effects of disinfectants on survival of cryptosporidium oocysts. Vet
Rec 111, p. 414.
42. Bodri M.S. (1994). Common parasitic diseases of reptiles and amphibians. Proceedings of the
American Association of Reptilian and Exotic Veterinarian Annual Conference, p.11-17.
43. Hering-Hagenbeck S.F.B.N., Boomker J. (2000). A check-list of the nematode parasites of
South African serpents (snakes) and sauria (lizards). Onderstepoort J Vet Res 67, p. 1-13.
44. McArthur S., McLellan L., Brown S. (2004). Gastrointestinal system. In: Girling S.J., Raiti P.
(Editors) Manual of reptiles, Second edition. British Small Animal Veterinary Association,
Waterwells, p. 210-229.
45. Iverson J. (1982). Adaptations to herbivory in iguanine lizards. In: Burghardt G. and Rand A.
(Editors) Iguanas of the World: Their Behaviour, Ecology, and Conservation, Noyes Publications,
Park Ridge, NJ, p. 60-76.
46. Kane K.K., Corwin R.M., Boever W.J. (1976). Impaction due to oxyurid infection in a Fiji Island
Iguana. Vet Med Small Anim Clin 71, p.183-184.
47. Bone R.D. (1992). Gastrointestinal system. In: Beynon P.H., Lawton M.P.C., Cooper J.E. (Editors)
Manual of reptiles, Gloucestershire, p. 101-116.
48. Reichenbach-Klinke H.H. (1977). Krankheiten der Reptilien. Gustav Fischer Verlag, Stuttgart, p.
228.
49. Kearn G.C. (1998). Parasitism and the Platyhelminths. Chapman & Hall, London, p. 544.
50. Goldberg S.R. (1985). Larval cestodes (Mesocestoides sp.) in the liver of the island night lizard,
Xantusia riversiana. J Wildl Dis 21, p.310-312.
51. Ritchie B. (2006). Virology. In: Mader D.R. (Editor) Reptile medicine and surgery, second edition.
Elsevier, California, p. 391-417.
52. Wellehan J.F.X., Johnson A.J., Harrach B., Benkö M., Pessier A.P., Johnson C.M., Garner M.M.,
Childress A., Jacobson E.R. (2004). Detection and analysis of six lizards adenoviruses by
consensus primer PCR provides further evidence of a reptilian origin for the atadenoviruses.
Journal of Virology 78 (23), p. 13366-13369.
53. Papp T., Fledelius B., Schmidt V., Kajan G.L., Marschang R.E. (2009). PCR sequence
characterization of new adenoviruses found in reptiles and the first successful isolation of a lizard
adenovirus. Veterinary Microbiology 134, p 233-240.
54. Julian A.F., Durham J.K. (1985). Adenoviral hepatitis in a bearded dragon (Amphibolurus
barbatus). N Z Vet J 30, p. 59.
55. Jacobson E.R., Kopit W., Kennedy F.A., Funk R.S. (1996). Co-infection of a bearded dragon,
Pogona vitticeps, with adeno- and dependo-like viruses. Vet Path 33, p. 343-346.
56. Kinsel M.J., Barbiers R.B., Manharth A., Murnane R.D. (1997). Small intestinal adeno-like

30
virus in a mountain chameleon (Chameleo montium). J Zoo Wildl Med 28, p. 498-500.
57. Hernandez-Divers S.J. (2006). Reptile Viral Diseases- Summary Table. In: Mader D.R. (Editor)
Reptile medicine and surgery, second edition. Elsevier, California, p. 1147-1158.
58. Stahl S.J. (2001). Update on diseases of bearded dragons (Pogona vitticeps). Proceedings on 15th
Annual North American Veterinary Conference, Orlando, Fl, p. 822-825.
59. Kim D.Y., Mitchell M.A., Bauer R.W., Poston R., Cho D.Y.(2002). An outbreak of adenoviral
infection in inland bearded dragons (Pogona vitticeps) coinfected with dependovirus and coccidial
protozoa (Isospora sp.). J Vet Diagn Invest 14, p. 332-334.
60. Hyndman T., Shilton C.M. (2011). Molecular detection of two adenoviruses associated with
disease in Australian lizards. Australian Veterinary Journal 89 (6), p. 232-235.
61. Jacobson E.R., Gardiner C.H. (1990). Adeno-like virus in esophageal and tracheal mucosa of a
jacksons chameleon (Chameleo jacksonii). Vet Pathol 27, p. 210.
62. Benson K.B. (1999). Reptilian gastrointestinal diseases. Semin Avian Exotic Pet Med 8, p. 90-97.
63. Marschang R.E., Chitty J. (2004). Infectious diseases. In: Girling S.J., Raiti P. (Editors)
Manual of reptiles, Second edition. British Small Animal Veterinary Association, Waterwells, p.
330-345.
64. Zwart P., Poelma F.G., Strik W.J., Peters J.C., Polder J.J.W. (1968). Report on births and deaths
occurring in gardens of the Royal Rotterdam Zoo “Blijdorp” during the years 1961 and 1962.
Tijdschr Diergeneeskd 93, p. 348.
65. Enweani I.B., Uwajeh J.C., Bello C.S.S., Ndip R.N. (1997). Fungal carriage in lizards. Mycoses 40,
p. 115-117.
66. Paré J.A. (2003). Fungi and fungal diseases of reptiles. Proceedings of the American Association
of Reptilian and Amphibian Veterinarians Annual Conference, p.128-131.
67. Knight M., Glor R., Smedley S.R., Gonzales A., Adler K., Eisner T. (1999). Firefly toxicosis in
lizards. J Chem Ecol 25, p.1981-1986.
68. Jacobson E.R. (1981). Neoplastic diseases. In: Cooper J.E., Jackson O.F. (Editors) Diseases of
Reptilia, Vol. 2, Academic Press, San Diego, p. 429-468.
69. Frye F.L. (1991b). Common pathologic lesions and disease processes: neoplasia. Frye F.L.
(Editor) Reptile care: an atlas of diseases and treatments, Vol. 2, TFH Publishing, Neptune City,
NJ, p. 576-609.


















