
Role Antibody Leukocytosis-Promoting Hemagglutinin ... · Aerosol infection of mice. Aerosol...
9
INFECTION AND IMMUNITY, Mar. 1981, p. 1223-1231 Vol. 31, No. 3 0019-9567/81/031223-09$02.00/0 Role of Antibody to Leukocytosis-Promoting Factor Hemagglutinin and to Filamentous Hemagglutinin in Immunity to Pertussis YUJI SATO,' KAZUMI IZUMIYA,1 HIROKO SATO,2 JAMES L. COWELL,3* AND CHARLES R. MANCLARK3 First' and Second2 Departments of Bacteriology, National Institute of Health, Tokyo 141 Japan and Division of Bacterial Products, Bureau of Biologics, Food and Drug Administration, Bethesda, Maryland 202053 Gamma globulins were obtained from rabbits hyperimmunized with the fila- mentous hemagglutinin (FHA) or the leukocytosis-promoting factor hemaggluti- nin (LPF. HA) of Bordetella pertussis. Double immunodiffusion analyses showed the antibodies to be highly specific for the homologous antigen. Intraperitoneal injection of anti-FHA or anti-LPF HA into 10-day-old ICR mice 30 min before aerosol challenge with B. pertussis protected the mice from disease as evidenced by survival and a normal rate of gain in body weight. Both anti-LPF HA and anti-FHA enhanced the clearance of B. pertussis from the lung and prevented leukocytosis which normally occurs after aerosol infection of mice with B. pertus- sis. Anti-LPF. HA, but not anti-FHA, neutralized the leukocytosis-promoting and histamine-sensitizing activities of LPF HA when incubated with the purified protein in vitro. Anti-FHA, but not anti-LPF- HA, prevented the attachment of B. pertussis to mammalian cells (HeLa and Vero) in culture. Since Sauer (26) and Kendrick and Eldering (9) established that effective vaccines containing whole cells of Bordetella pertussis could be made, there have been many attempts to iden- tify the protective antigen(s) of B. pertussis. After Keogh et al. (11) discovered that cultures of B. pertussis produced a hemagglutinin (HA) some studies (5, 10) suggested the HA played an important role in immunity to pertussis, whereas other data (13, 20, 29) suggested that HA was not involved in immunity to pertussis. Recently Sato et al. (23) reported that immunization with a purified preparation containing HA and leu- kocytosis-promoting factor (LPF) protected mice from intracerebral infection. Arai and Sato (3) subsequently separated this preparation into two distinct HAs. Once HA (leukocytosis-promoting factor he- magglutinin or LPF.-HA) had low hemaggluti- nating activity and produced leukocytosis and an increased sensitivity to the lethal effects of histamine when injected into mice. This protein showed a single band on polyacrylamide gel electrophoresis and had a molecular weight of about 105,000. Electron micrographs of LPF. HA showed spherical molecules about 6 nm in diameter. Similar preparations of LPF- HA have been purified by Morse and Morse (15) and Irons and MacLennan (7, 8). LPF.-HA, when injected into mice, also stimulates the secretion of insulin upon glucose or epinephrine challenge (Y. Sato, J. L. Cowell, H. Sato, and C. R. Manclark, un- published data) and appears identical to the islet-activating protein of B. pertussis (32). The other HA, filamentous HA (FHA), had high hemagglutinating activity, was free of leu- kocytosis-promoting and histamine-sensitizing activities, and was antigenically distinct from the LPF .HA (3). Electron micrographs of FHA showed filamentous structures about 2 x 40 nm in size. Gel electrophoresis in sodium dodecyl sulfate showed one major band (molecular weight, 126,000) and several other bands with molecular weights between 120,000 and 52,000. Similar preparations of FHA have been obtained by Morse and Morse (15) and Irons and Mac- Lennan (7). As currently purified, FHA is a protein preparation of multiple components, free of LPF- HA, and enriched in an HA distinct from the LPF HA. Sato et al. (24) have pro- posed that the filamentous structures in the FHA are derived from fimbriae which they sug- gest are HA and adherence factors for B. pertus- sis. Mouse protective activity was re-examined with these two distinct HAs. Sato et al. (24), Morse and Morse (15) and Irons and Mac- Lennan (7) reported that active immunization with FHA protected mice from intracerebral challenge with B. pertussis. LPF HA made a toxoid by Formalin was a poor protective anti- gen for mice when tested by Sato et al. (24), and 1223 on April 12, 2020 by guest http://iai.asm.org/ Downloaded from
Transcript of Role Antibody Leukocytosis-Promoting Hemagglutinin ... · Aerosol infection of mice. Aerosol...

INFECTION AND IMMUNITY, Mar. 1981, p. 1223-1231 Vol. 31, No. 30019-9567/81/031223-09$02.00/0
Role of Antibody to Leukocytosis-Promoting FactorHemagglutinin and to Filamentous Hemagglutinin in
Immunity to PertussisYUJI SATO,' KAZUMI IZUMIYA,1 HIROKO SATO,2 JAMES L. COWELL,3* AND CHARLES R.
MANCLARK3First' and Second2 Departments ofBacteriology, National Institute of Health, Tokyo 141 Japan and
Division ofBacterial Products, Bureau of Biologics, Food and Drug Administration, Bethesda,Maryland 202053
Gamma globulins were obtained from rabbits hyperimmunized with the fila-mentous hemagglutinin (FHA) or the leukocytosis-promoting factor hemaggluti-nin (LPF. HA) of Bordetella pertussis. Double immunodiffusion analyses showedthe antibodies to be highly specific for the homologous antigen. Intraperitonealinjection of anti-FHA or anti-LPF HA into 10-day-old ICR mice 30 min beforeaerosol challenge with B. pertussis protected the mice from disease as evidencedby survival and a normal rate of gain in body weight. Both anti-LPF HA andanti-FHA enhanced the clearance of B. pertussis from the lung and preventedleukocytosis which normally occurs after aerosol infection of mice with B. pertus-sis. Anti-LPF. HA, but not anti-FHA, neutralized the leukocytosis-promoting andhistamine-sensitizing activities of LPF HA when incubated with the purifiedprotein in vitro. Anti-FHA, but not anti-LPF- HA, prevented the attachment ofB. pertussis to mammalian cells (HeLa and Vero) in culture.
Since Sauer (26) and Kendrick and Eldering(9) established that effective vaccines containingwhole cells of Bordetella pertussis could bemade, there have been many attempts to iden-tify the protective antigen(s) of B. pertussis.After Keogh et al. (11) discovered that culturesof B. pertussis produced a hemagglutinin (HA)some studies (5, 10) suggested the HA played animportant role in immunity to pertussis, whereasother data (13, 20, 29) suggested that HA wasnot involved in immunity to pertussis. RecentlySato et al. (23) reported that immunization witha purified preparation containing HA and leu-kocytosis-promoting factor (LPF) protectedmice from intracerebral infection. Arai and Sato(3) subsequently separated this preparation intotwo distinct HAs.Once HA (leukocytosis-promoting factor he-
magglutinin or LPF.-HA) had low hemaggluti-nating activity and produced leukocytosis andan increased sensitivity to the lethal effects ofhistamine when injected into mice. This proteinshowed a single band on polyacrylamide gelelectrophoresis and had a molecular weight ofabout 105,000. Electron micrographs of LPF.HA showed spherical molecules about 6 nm indiameter. Similar preparations ofLPF-HA havebeen purified by Morse and Morse (15) and Ironsand MacLennan (7, 8). LPF.-HA, when injectedinto mice, also stimulates the secretion of insulinupon glucose or epinephrine challenge (Y. Sato,
J. L. Cowell, H. Sato, and C. R. Manclark, un-published data) and appears identical to theislet-activating protein of B. pertussis (32).The other HA, filamentous HA (FHA), had
high hemagglutinating activity, was free of leu-kocytosis-promoting and histamine-sensitizingactivities, and was antigenically distinct fromthe LPF.HA (3). Electron micrographs of FHAshowed filamentous structures about 2 x 40 nmin size. Gel electrophoresis in sodium dodecylsulfate showed one major band (molecularweight, 126,000) and several other bands withmolecular weights between 120,000 and 52,000.Similar preparations ofFHA have been obtainedby Morse and Morse (15) and Irons and Mac-Lennan (7). As currently purified, FHA is aprotein preparation of multiple components, freeof LPF- HA, and enriched in an HA distinctfrom the LPF HA. Sato et al. (24) have pro-posed that the filamentous structures in theFHA are derived from fimbriae which they sug-gest are HA and adherence factors for B. pertus-sis.Mouse protective activity was re-examined
with these two distinct HAs. Sato et al. (24),Morse and Morse (15) and Irons and Mac-Lennan (7) reported that active immunizationwith FHA protected mice from intracerebralchallenge with B. pertussis. LPF HA made atoxoid by Formalin was a poor protective anti-gen for mice when tested by Sato et al. (24), and
1223
on April 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

1224 SATO ET AL.
active LPF-HA was not protective at the dosestested by Irons and MacLennan (7). However,Munoz and Bergman (16, 17) found that a prep-aration of LPF.HA, referred to as pertussigenand reported to be free of demonstrable FHA(2), protected mice from intracerebral B. pertus-sis infection.
In this paper we present passive immunizationdata to further evaluate the mouse protectiveactivity of FHA and LPF. HA. Using an aerosol-induced respiratory infection of mice as a labo-ratory model for pertussis (25), we show thatintraperitoneal injection of antibody to FHA orto LPF HA protects mice from disease. Datarelevant to possible mechanisms by which anti-FHA and anti-LPF. HA protect are also pre-sented.
MATERIALS AND METHODSBacterial strains and growth. B. pertussis strain
Tohama, phase I or phase III, was grown at 35°C onBordet-Gengou agar containing 20% defibrinated bo-vine blood. The cells from freeze-dried cultures weretransferred a maximum of three times before use. Onthe final passage, cells were allowed to grow for 20 h.
Viable counts were determined by diluting samplesin sterile 0.01 M sodium phosphate-buffered saline(pH 7.2) (PBS) and spreading 0.1 ml of the appropriatedilutions on Bordet-Gengou agar plates. Plates wereincubated at 35°C for 3 to 4 days. Viable counts wereexpressed in colony-forming units.Aerosol infection of mice. Aerosol infection of
mice was done as previously described (25). Specific-pathogen-free 10-day-old ICR mice were exposed(without mother) for 30 min to an aerosol generatedfrom 2 x 109 cells of B. pertussis strain Tohama, phaseI per ml suspended in PBS containing 1% CasaminoAcids (Difco Laboratories, Detroit, Mich.). Animalswere exposed to the aerosol by spraying 0.4 ml of thebacterial suspension per min at a nebulizer pressure of1.5 kg/cm2 with filter sterilized air. A specially builtnebulizer was used, but similar aerosols of B. pertussiscan be obtained by using the standard nebulizer dis-tributed by Fisons Corporation, Bedford, Mass. Anoutlet tube at the bottom of the chamber removed airto a sterilizing system at a negative pressure of about1.5 kg/cm2.About 20 min after stopping the aerosol generation,
the mice were removed from the chamber and placedwith mothers in cages with air filter covers. Thisrepresented zero time after infection. At this time andappropriate times thereafter, the following procedureswere performed. (i) Mice were weighed individually.(ii) Blood was removed from the tail vein of eachmouse for estimation of the leukocyte (WBC) countper cubic millimeter of blood (23). (iii) Mice wereanesthetized with ether and exsanguinated by heartpuncture, and the lungs were removed aseptically. (iv)Lungs were homogenized in sterile PBS and colony-forming units of B. pertussis in the lungs were deter-mined by plate count on Bordet-Gengou agar.
Preparation of gamma globulins. New ZealandWhite rabbits were bled before immunization as a
source of normal y globulins. For the preparationof anti-FHA, the FHA preparation in 50 mMtris(hydroxymethyl)aminomethane- hydrochloride(pH 8.0) containing 1 M NaCl was emulsified with anequal volume of incomplete Freund adjuvant (DifcoLaboratories), and 100 ,ug of protein was injected intothe footpads of each rabbit. After 6 weeks, a boosterinjection of 100 ,ug of soluble protein in the abovetris(hydroxymethyl)aminomethane-sodium chloridebuffer (pH 8.0) was injected subcutaneously, and theanimals were bled 2 weeks later. Anti-LPF.HA wasprepared as above except that the protein was treatedwith Formalin (23). Antibody to the crude antigenicmixture of B. pertussis was prepared as described foranti-FHA except that each rabbit received 500 ,ug ofprotein in the primary injection and 1 mg of proteininjected into the ear vein in the secondary injection.The y globulin fractions of the normal serum and
the antisera were prepared by the methods of Harboeand Ingild (6) except that the diethylaminoethyl-Sephadex chromatography step was omitted. The yglobulin preparations were stored at -20°C at proteinconcentrations of 38, 34, and 26 mg/ml for anti-LPF.HA, anti-FHA, and normal globulins, respectively.The agglutination titers of anti-LPF.HA and anti-FHA for B. pertussis strain Tohama, phase I, testedas described by Sato et al. (23), were 1:40 and 1:2560,respectively.
Effect of passive immunization with anti-FHAor anti-LPF.HA. Anti-FHA, anti-LPF.HA, or nor-mal y globulin were diluted 1:10 with PBS and 0.2 mlwas injected intraperitoneally into mice 30 min beforeaerosol challenge with B. pertussis. At zero time afterinfection and at appropriate times thereafter theweight of the mice, their peripheral WBC counts, andthe viable B. pertussis cells in the lungs were deter-mined as stated above.Attachment of B. pertussis to mammalian
cells. HeLa and Vero cell cultures were grown on glasscover slips (10.5 by 22 mm) placed in sterile plasticdishes (35 by 10 mm; Falcon Plastics, Oxnard, Califor-nia) containing 3 ml of the HeLa or Vero cell suspen-sion (about 105 cells per ml) in Eagle minimal essentialmedium containing 10% heat-inactivated fetal bovineserum and 100 U of penicillin per ml. The cell cultureswere incubated at 36°C in 5% CO2 in air for 24 h andthen washed with Eagle minimal essential mediumlacking penicillin before the addition of B. pertussis.Suspensions of B. pertussis strain Tohama, phase I orphase III, at 1 x 109 cells per ml in Eagle minimalessential medium were incubated at 23°C for 15 minwith or without a 1:40 dilution of anti-FHA, anti-LPF-HA, or normal y globulin. Portions (2 ml) of thesesuspensions were then added to the 24-h monolayersof HeLa or Vero cells and incubated for 2 h at 36°C in5% CO2 in air. The monolayer cells were then washedseveral times with Eagle minimal essential mediumand either fixed with 3 ml of 95% ethanol and stainedwith 1% crystal violet or fixed and stained by a deoxy-ribonucleic acid-staining fluorochrome procedure (4).
Purification ofFHA and LPF-HA. The proteinswere purified and assayed as described by Arai andSato (3). The preparations used for these studies wereobtained from the second agarose gel filtration step.
Neutralization of the leukocytosis-promoting
INFECT. IMMUN.
on April 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
PROTECTIVE ANTIBODIES TO B. PERTUSSIS 1225
and histamine-sensitizing activities of LPF.HA.Purified LPF.HA (10 Ag/ml) was mixed with an equalvolume of PBS or various dilutions of normal y glob-ulin, anti-FHA or anti-LPF. HA. After 30 min at 23°C,0.2 ml portions of the mixtures were injected into thetail veins of 14- to 16-g DDY mice. The WBC countper cubic millimeter of blood was assayed 3 days afterinjection. On day 4, histamine sensitivity was deter-mined by the intraperitoneal injection of 1 mg ofhistamine base per mouse. Mortality was used as ameasure of histamine-sensitizing activity.Crude antigenic preparation. A preparation con-
taining many antigens of B. pertussis was prepared byammonium sulfate precipitation of the culture super-natant from 4-day stationary cultures of B. pertussisTohama, phase I (23). Solid ammonium sulfate wasadded to 1 liter of culture supernatant to 80% satura-tion (561 g/liter) and stirred at 4°C for 20 min. After72 h at 40C, the precipitate was collected by centrifu-gation (27,000 x g for 15 min, 40C). The precipitatewas extracted with 5 ml of 0.01 M sodium phosphatebuffer (pH 8.0), centrifuged as above, and the pelletwas extracted with 5 ml of 0.1 M tris(hydroxy-methyl)aminomethane (pH 8.0) containing 0.5 MNaCl. The supernatants of these two extractions werepooled to give the crude antigen at 4.7 mg of proteinper ml. This crude antigen preparation showed about25 bands after sodium dodecyl sulfate gel electropho-resis with 7.5% gels as described by Weber et al. (31).
Immunodiffusion. Immunodiffusion was done in1% agarose buffered with 25 mM tris(hydroxy-methyl)aminomethane (pH 8.0) containing 0.5 MNaCl. Diffusion took place in a moist chamber at 23°Cfor about 24 h.
Protein determination. Protein concentrationswere estimated with Folin reagent (12) with bovineplasma albumin (Armour Pharmaceutical Co., Scotts-dale, Ariz.) as the standard.
RESULTSPassive protection of mice by anti-LPF.
HA and anti-FHA. To evaluate the protectivecapability of anti-LPF-HA and anti-FHA, eachwas injected intraperitoneally into mice 30 minbefore the mice were challenged with B. pertus-sis by aerosol inhalation. Mice receiving anti-LPF -HA or anti-FHA had a rate of gain in bodyweight equivalent to mice not exposed to B.pertussis aerosols (Fig. 1). No deaths occurredin the passively immunized or uninfected groupsof mice. In contrast, mice that were injectedintraperitoneally with normal y globulin 30 minbefore aerosol challenge had a significantly re-duced rate of body weight gain (Fig. 1), and 80%of the animals died with 14 days of aerosolinhalation.
Specificity of anti-LPF.HA and anti-FHA. Since anti-LPF.HA and anti-FHA pas-sively protected mice from disease, it was im-portant to evaluate the specificity of these anti-body preparations. This was done, in part, bythe immunodiffusion of anti-LPF- HA and anti-
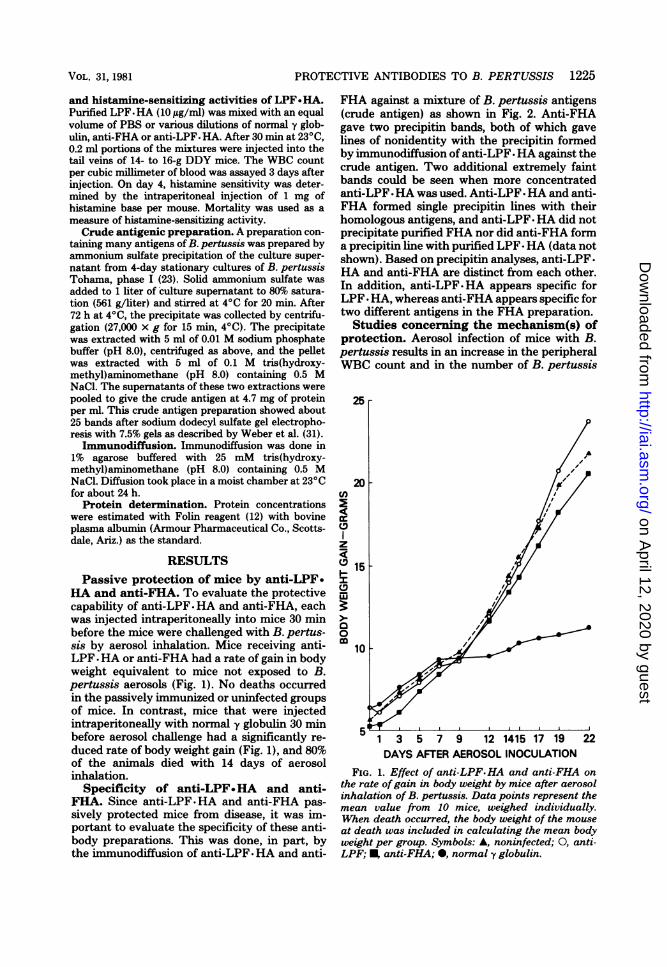
FHA against a mixture of B. pertussis antigens(crude antigen) as shown in Fig. 2. Anti-FHAgave two precipitin bands, both of which gavelines of nonidentity with the precipitin formedby immunodiffusion of anti-LPF. HA against thecrude antigen. Two additional extremely faintbands could be seen when more concentratedanti-LPF *HA was used. Anti-LPF * HA and anti-FHA formed single precipitin lines with theirhomologous antigens, and anti-LPF. HA did notprecipitate purified FHA nor did anti-FHA forma precipitin line with purified LPF-HA (data notshown). Based on precipitin analyses, anti-LPF-HA and anti-FHA are distinct from each other.In addition, anti-LPF-HA appears specific forLPF * HA, whereas anti-FHA appears specific fortwo different antigens in the FHA preparation.Studies concerning the mechanism(s) of
protection. Aerosol infection of mice with B.pertussis results in an increase in the peripheralWBC count and in the number of B. pertussis
20cn
CD)
z
a 15
x
010
5w!T1 3 5 7 9 12 1415 17 19 22DAYS AFTER AEROSOL INOCULATION
FIG. 1. Effect of anti-LPF.HA and anti-FHA onthe rate ofgain in body weight by mice after aerosolinhalation of B. pertussis. Data points represent themean value from 10 mice, weighed individually.When death occurred, the body weight of the mouseat death was included in calculating the mean bodyweight per group. Symbols: A, noninfected; 0, anti-LPF; I anti-FHA; 0, normal y globulin.
VOL. 31, 1981
on April 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
anti-FHA prevented leukocytosis in aerosol-in-fected mice, it was appropriate to determine ifanti-LPF.HA and anti-FHA could neutralizethe leukocytosis-promoting and histamine-sen-sitizing activities of the LPF * HA. The neutrali-zation data are shown in Table 1. At a finaldilution of 1:36 for anti-LPF * HA, neutralizationof LPF HA activity was complete and signifi-cant neutralization was obtained at 1:108. Nor-mal y globulin or anti-FHA at dilutions at 1:4did not affect the activities of LPF * HA.
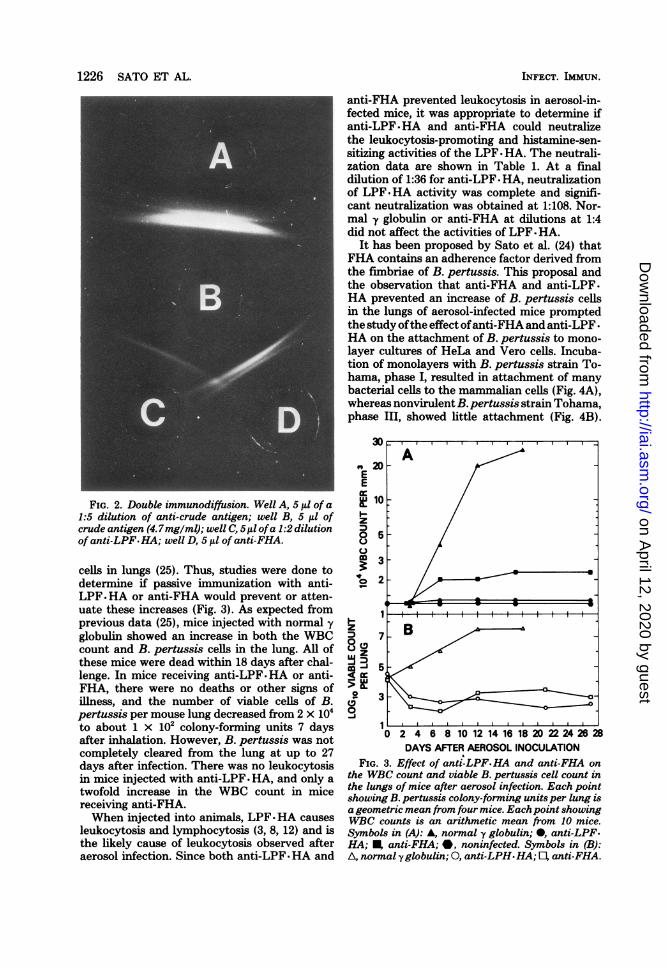
It has been proposed by Sato et al. (24) thatFHA contains an adherence factor derived fromthe fimbriae of B. pertussis. This proposal andthe observation that anti-FHA and anti-LPF.HA prevented an increase of B. pertussis cellsin the lungs of aerosol-infected mice promptedthe study ofthe effect ofanti-FHA and anti-LPF -
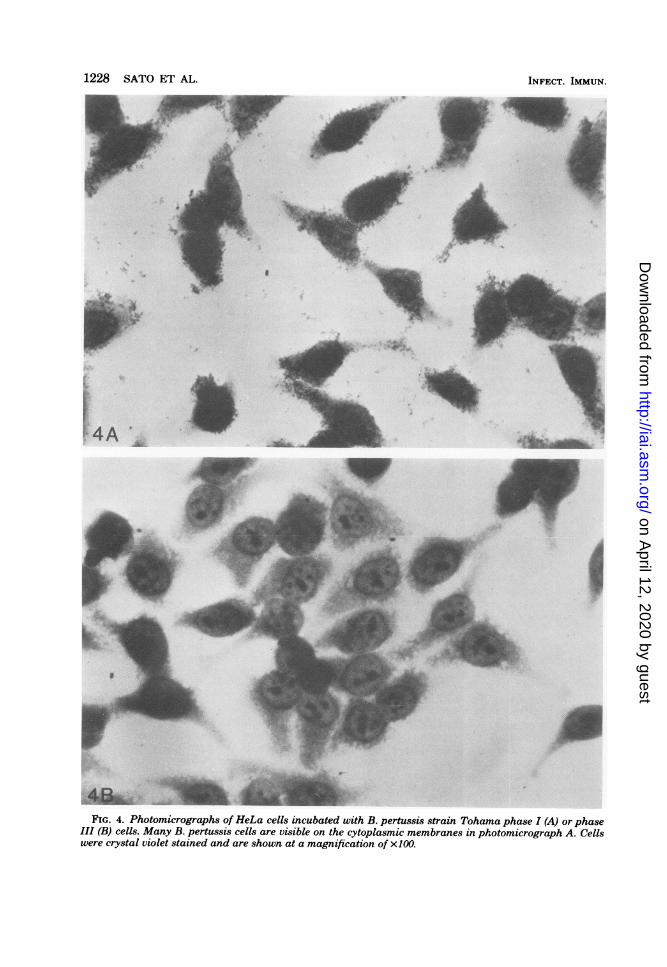
HA on the attachment of B. pertussis to mono-layer cultures of HeLa and Vero cells. Incuba-tion of monolayers with B. pertussis strain To-hama, phase I, resulted in attachment of manybacterial cells to the mammalian cells (Fig. 4A),whereas nonvirulent B. pertussis strain Tohama,phase III, showed little attachment (Fig. 4B).
_______________________ ~~~~~30 IIIIIIIIIII
Aco 20-~~~~~~~~~~~~~EE
FIG. 2. Double immunodiffusion. Well A, 5 1l of a t 101:5 dilution of anti-crude antigen; well B, 5 ,il of :crude antigen (4.7mg/ml); well C, 5,ul ofa 1:2 dilution D /of anti-LPF. HA; well D, 5 ,ll of anti-FHA.
m 3cells in lungs (25). Thus, studies were done to 3/determine if passive immunization with anti- 2-LPF.HA or anti-FHA would prevent or atten-uate these increases (Fig. 3). As expected from 1 , lprevious data (25), mice injected with normal 'yglobulin showed an increase in both the WBC D 7count and B. pertussis cells in the lung. All of 8these mice were dead within 18 days after chal- / 3lenge. In mice receiving anti-LPF-HA or anti-FHA, there were no deaths or other signs of > W.illness, and the number of viable cells of B.pertussis per mouse lung decreased from 2 x 104 °to about 1 x 102 colony-forming units 7 days 1 6 8 101214161820 2224262after inhalation. However, B. pertussis was notcompletely cleared from the lung at up to 27 DAYS AFTER AEROSOL INOCULATIONdays after infection. There was no leukocytosis FIG. 3. Effect of anti-LPF.HA and anti-FHA onin mice injected with anti-LPF. HA, and only a the WBC count and viable B. pertussis cell count intwofold increase in the WBC count in mice the lungs of mice after aerosol infection. Each pointtwodinc eanti-FHAn showing B. pertussis colony-forming unitsper lung isreceiving anti-FHA . a geometric mean from four mice. Eachpoint showingWhen injected into animals, LPF-HA causes WBC counts is an arithmetic mean from 10 mice.leukocytosis and lymphocytosis (3, 8, 12) and is Symbols in (A): A, normal y globulin; 0, anti-LPF.the likely cause of leukocytosis observed after HA; E, anti-FHA; *, noninfected. Symbols in (B):aerosol infection. Since both anti-LPF HA and A, normal yglobulin; 0, anti-LPH.HA; 0, anti-FHA.
1226 SATO ET AL. INFECT. IMMUN.
on April 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

PROTECTIVE ANTIBODIES TO B. PERTUSSIS 1227
TABLE 1. Effect of anti-LPF.HA and anti-FHA onthe leukocytosis-promoting and histamine-
sensitizing activities ofLPFGNormal Anti-FHA Anti-LPF
Dilu- y globulintions
WBC HS WBC HS WBC HS
1:4 78 x i03 5/5 84 x i03 5/5 27 x i03 0/51:12 71 x 103 5/5 97 x 103 5/5 28 x 103 0/51:36 86 x 103 5/5 92 x 103 5/5 27 x 103 0/51:108 99 X 103 5/5 40 x 103 1/51:324 104 x 103 5/5 58 x 103 3/51:972 73 x 103 4/51:2916 98 x 103 5/5
a Final dilutions of the y globulin preparations afterthe addition of an equal volume of LPF-HA (10 ug)are described in the text. WBC (White blood cellcount) values are expressed per cubic millimeter ofblood and represent arithmetic means from five mice;mice injected with PBS averaged 22 x 103 WBC permm3, and mice injected with LPF mixed with PBSgave 78 x 103 WBC per mm3. HS (histamine sensitiv-ity) values show the number of deaths per total num-ber of animals injected; mice injected with PBS hadan HS of 0/5, whereas mice injected with LPF HAmixed with PBS had an HS of 5/5.
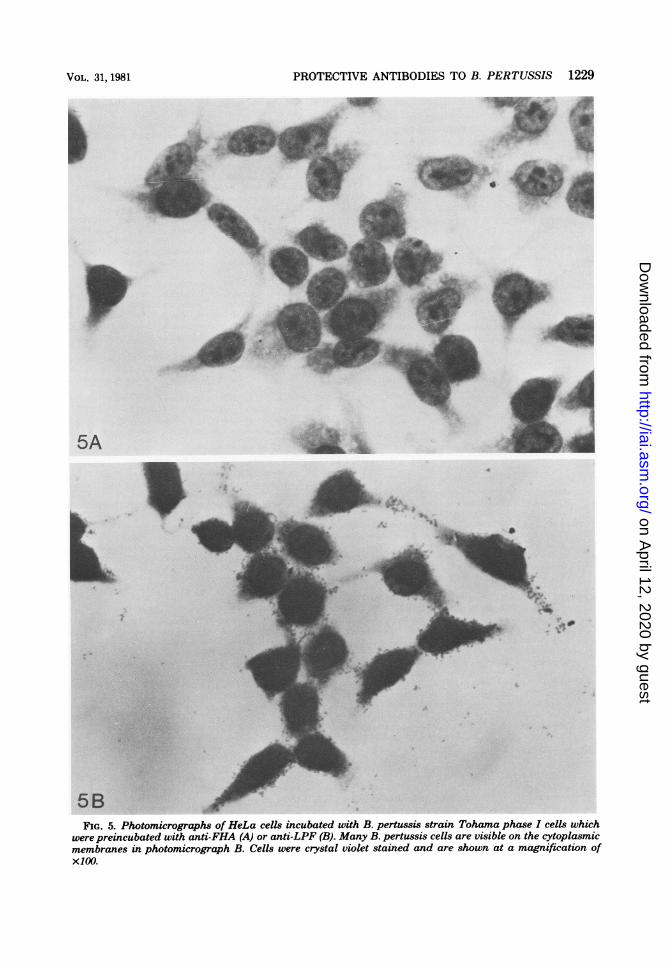
Preincubation of B. pertussis strain Tohamaphase I cells with anti-FHA appeared to preventattachment of these cells to the monolayer (Fig.5A), but preincubation of Tohama phase I cellswith anti-LPF-HA did not prevent attachment(Fig. 5B). Photomicrographic fields of HeLa orVero cells incubated with B. pertussis strainTohama phase I cells which were preincubatedwith normal y globulin were similar to Fig. 5B inthat many bacteria were attached to the mam-malian cells. Figures 4 and 5 show representativephotomicrographic fields of results obtainedwith HeLa cells stained with crystal violet. Sim-ilar results were obtained with Vero cells.
DISCUSSIONThe data presented show that mice infected
by aerosol inhalation of B. pertussis were pas-sively protected from disease by two distinctantibody preparations, anti-LPF-HA and anti-FHA. Although more information is required onthe mechanism(s) by which anti-LPF HA andanti-FHA protect, there are several possible ex-planations. We found that anti-LPF HA neu-tralized the leukocytosis-promoting and hista-mine-sensitizing activities of purified LPF- HA,and it is possible that anti-LPF. HA protectsfrom disease by neutralizing the potentiallyharmful activities of LPF-HA released duringthe infectious process. LPF-HA is released fromthe bacterial cell during growth in vitro (23).Also, since anti-LPF. HA and anti-FHA en-
hanced the clearance of B. pertussis from mouselungs, it is possible that either or both act toincrease phagocytosis of B. pertussis by pulmo-nary phagocytes. In vitro data show that alveolarmacrophages (18) and polymorphonuclear leu-kocytes (30) are activated by antisera to B. per-tussis to phagocytize virulent B. pertussis orga-nisms. In addition, anti-FHA may protect bypreventing attachment of B. pertussis to respi-ratory epithelial cells. We did find that anti-FHA prevented the attachment of B. pertussisto mammalian cells in vitro, and there are datasuggesting that attachment is a virulence factorfor B. pertussis (14, 24). Matsuyama (14), instudying excised rabbit trachea, showed thatcells of virulent B. pertussis strain Tohama,phase I, were not moved by ciliary activity fromthe point of inoculation, whereas nonvirulent B.pertussis Tohama phase III cells inoculated atthe same area of the tracheal segment werecleared toward the laryngeal end of the trachea.The fact that anti-FHA and anti-LPF. HA
passively protected mice from disease after res-piratory infection corrQborates active immuni-zation data identifying FHA as a protective an-tigenic preparation (7, 24), and suggests thatimmunization with LPF-HA should protectmice from infection with B. pertussis. PurifiedLPF. HA is lethal at relatively low concentra-tions, and Irons and MacLennan (7, 8), by usingthe standard mouse potency assay, found thelethal dose of LPF-HA to be smaller than theprotective dose. Sato et al. (24) made LPF-HAa toxoid with Formalin, but the toxoid poorlyprotected mice from intracerebral challengewith B. pertussis; it is possible that this toxoidwas a poor immunogen. The LPF preparationcalled pertussigen by Munoz and Bergman (16,17) has been reported to actively protect micefrom intracerebral challenge with B. pertussis;however, the purity of this preparation has notbeen adequately described, and it is not certainwhether the LPF *HA in this preparation is act-ing as a protective antigen or is enhancing theeffect of other protective antigens in the sample.Nakase and Doi (19), by using the mouse intra-cerebral challenge potency test, have shown thatsmall amounts of LPF-HA could enhance theprotective effect of FHA. Additional active im-munization studies with purified, well-character-ized, and appropriately inactivated LPF-HA areneeded.
In this discussion of protective antigens andantibody preparations, two different challengeroutes, intracerebral injection and aerosol inha-lation, have been used. Our passive protectiondata were obtained from an aerosol inducedrespiratory infection, whereas the active immu-
VOL. 31, 1981
on April 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
1228 SATO ET AL.
_-.1
4v-
A
I.
F;FIG. 4. Photomicrographs of HeLa cells incubated with B. pertussis strain Tohama phase I (A) or phase
III (B) cells. Many B. pertussis cells are visible on the cytoplasmic membranes in photomicrograph A. Cellswere crystal violet stained and are shown at a magnification of x100.
INFECT. IMMUN.
. IL-1i
A
on April 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
PROTECTIVE ANTIBODIES TO B. PERTUSSIS 1229
5A 1
||_ o; ,t~~~~
5B-FIG. 5. Photomicrographs of HeLa cells incubated with B. pertussis strain Tohama phase I cells which
were preincubated with anti-FHA (A) or anti-LPF (B). Many B. pertussis cells are visible on the cytoplasmicmembranes in photomicrograph B. Cells were crystal violet stained and are shown at a magnification ofX100.
VOL. 31, 1981
on April 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

1230 SATO ET AL.
nization data were obtained from an intracere-bral infection. The respiratory infection, as amodel for pertussis, is considered more satisfac-tory since intracerebral infection is not naturaland fresh disease isolates are usually of very lowvirulence when injected intracerebrally intomice (1, 21, 27). In addition, it has been sug-gested (22, 27, 28) that the host-parasite inter-action occurring after intracerebral infection isdifferent from that in respiratory infection.
Intracerebral challenge of vaccinated mice isthe accepted challenge route in the standardassay of the protective potency of pertussis vac-cines. Intracerebral infection is widely used andwill continue to be employed to evaluate poten-tial protective antigens of B. pertussis; however,respiratory infection should also be used in fur-ther active immunization studies necessary tounderstand the role of LPF -HA and FHA inimmunity to pertussis. In addition, biochemicalcharacterization of FHA is needed to evaluatethe nature of the protective antigen(s) in thispreparation.
ACKNOWLEDGMENTH. Yoshida gave greatly appreciated assistance in the ex-
periments with Vero cell cultures, and M. Oda provided expertassistance with the aerosol infection experiments.
LITERATURE CITED1. Andersen, E. K. 1952. Some observations made during
experiments on mice inoculated with H. pertussis. ActaPathol. Microbiol. Scand. 31:546-560.
2. Arai, H., and J. J. Munoz. 1979. Fimbrial hemagglutininin stationary and shake cultures of Bordetellapertussis.Infect. Immun. 25:764-767.
3. Arai, H., and Y. Sato. 1976. Separation and characteri-zation of two distinct hemagglutinins contained in pur-ified leukocytosis-promoting factor from Bordetellapertussis. Biochim. Biophys. Acta. 444:765-782.
4. Barile, M. F. 1977. Mycoplasma contamination of cellcultures: a status report, p. 291-334. In R. Actin (ed.),Cell culture: its applications. Academic Press, Inc., N.Y.
5. Fisher, S., M. F. Warburton, H. N. B. Wettenhall, andS. W. Williams. 1951. The haemagglutinin of Haemo-philus pertussis. IV. Reactions and antibody responsein babies and children following the administration ofhaemagglutinin adsorbed on aluminum phosphate.Aust. J. Exp. Biol. Med. Sci. 29:425-432.
6. Harboe, N., and A. Ingild. 1973. Immunization, isolationof imniunoglobulins, and estimation of antibody titer,p. 161-164. In N. H. Axelsen, J. Kroll, and B. Weeke(ed.), A manual of quantitative immunoelectrophoresis.Methods and applications. Scand. J. Immunol. 2 (Suppl.1):15-169.
7. Irons, L. I., and A. P. MacLennan. 1979. Substratespecificity and the purification by affinity combinationmethods of the two Bordetella pertussis hemaggluti-nins, p. 338-349. In C. R. Manclark and J. C. Hill (ed.),International symposium on pertussis. Department ofHealth, Education and Welfare Publication No. 79-1830. U.S. Government Printing Office, Washington,D.C.
8. Irons, L. I., and A. P. MacLennan. 1979. Isolation ofthe lymphocytosis promoting factor-haemagglutinin of
Bordetella pertussis by affinity chromatography.Biochim. Biophys. Acta. 580:175-185.
9. Kendrick, P., and G. Eldering. 1939. A study in activeimmunization against pertussis. Am. J. Hyg. 29:133-153.
10. Keogh, E. V., and E. A. North. 1948. The haemagglu-tinin as a protective antigen in experimental murinepertussis. Aust. J. Exp. Biol. Med. Sci. 26:315-322.
11. Keogh, E. V., E. A. North, and M. F. Warburton. 1947.Haemagglutinins of Haemophilus pertussis group. Na-ture (London). 160:63.
12. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193:265-275.
13. Masry, F. L. G. 1952. Production, extraction and purifi-cation of haemagglutinins of Haemophilus pertussis. J.Gen. Microbiol. 7:201-210.
14. Matsuyama, T. 1977. Resistance of Bordetella pertussisphase I to mucociliary clearance by rabbit trachaelmucous membrane. J. Infect. Dis. 136:609-616.
15. Morse, S. E., and J. H. Morse. 1976. Isolation andproperties of the leukocytosis- and lymphocytosis-pro-moting factor of Bordetellapertussis. J. Exp. Med. 143:1483-1502.
16. Munoz, J. J., and R. K. Bergman. 1977. Bordetellapertussis. Immunological and other biological activities.In N. Rose (ed.), Immunological series, vol. 4. MarcelDekker, Inc., N.Y.
17. Munoz, J. J., and R. K. Bergman. 1979. Mechanism ofaction of pertussigen, a substance from Bordetella per-tussis, p. 193-197. In D. Schlessinger (ed.) Microbiol-ogy-1979. American Society for Microbiology, Wash-ington, D.C.
18. Muse, K. E., D. Findley, L. Allen, and A. M. Collier.1979. In vitro model of Bordetella pertussis infection:pathogenic and microbicidal interactions, p. 41-50. InC. R. Manclark and J. C. Hill (ed.), International sym-posium on pertussis. Department of Health, Educationand Welfare Publication No. 79-1830. U.S. GovernmentPrinting Office, Washington, D.C.
19. Nakase, Y., and M. Doi. 1979. Toxicity and potency ofa purified pertussis vaccine, p. 350-356. In C. R. Man-clark and J. C. Hill (ed.), Intemational symposium onpertussis. Department of Health, Education and Wel-fare Publication No. 79-1830. U.S. Government PrintingOffice, Washington, D.C.
20. Pillemer, L. 1950. Absorption of protective antigen ofHemophilus pertussis on human red cell stromata.Proc. Soc. Exp. Biol. Med. 75:704-705.
21. Preston, N. W., and P. Evans. 1963. Type-specific im-munity against intracerebral pertussis infection in mice.Nature (London). 197:508-509.
22. Preston, N. W., and T. N. Stanbridge. 1976. Mouse orman? Which are pertussis vaccines to protect? J. Hyg.76:249-256.
23. Sato, Y., H. Arai, and K. Suzuki. 1974. Leukocytosis-promoting factor of Bordetella pertussis. III. Its iden-tity with protective antigens. Infect. Immun. 9:801-810.
24. Sato, Y., K. Izumiya, M. A. Oda, and H. Sato. 1979.Biological significance of Bordetella pertussis fimbriaeor hemagglutinin: A possible role of the fimbriae orhemagglutinin for pathogenesis and antibacterial im-munity, p. 51-57. In C. R. Manclark and J. C. Hill (ed.),International symposium on pertussis, Department ofHealth, Education and Welfare Publication No. 79-1830. U.S. Government Printing Office, Washington,D.C.
25. Sato, Y., K. Izumiya, H. Sato, J. L. Cowell, and C. R.Manclark. 1980. Aerosol infection of mice with Bor-detella pertussis. Infect. Immun. 29:261-266.
26. Sauer, L., 1933. Whooping cough. A study in immuniza-tion. J. Amer. Med. Assoc. 100:239-241.
INFECT. IMMUN.
on April 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

PROTECTIVE ANTIBODIES TO B. PERTUSSIS 1231
27. Standfast, A. F. B. 1958. Some factors influencing thevirulence for mice of Bordetella pertussis by the intra-cerebral route. Immunology. 1:123-134.
28. Standfast, A. F. B., and J. M. Dolby. 1961. A compari-son between the intranasal and intracerebral infectionof mice with BordeteUa pertussis. J. Hyg. 59:217-229.
29. Thiele, E. 1950. Studies on the hemagglutinin of He-mophilus pertussis. J. Immunol. 65:627-632.
30. Utsumi, S., S. Sonoda, T. Imagawa, and M. Kanoh.1978. Polymorphonuclear leukocyte-inhibitory factor of
Bordetella pertussis. I. Extraction and partial purifi-cation of phagocytosis- and chemotaxis-inhibitory ac-tivities. Biken J. 21:121-135.
31. Weber, K., J. R. Pringle, and M. Osborn. 1972. Mea-surement of molecular weights by electrophoresis onSDS-acrylamide gel. Methods Enzymol. 26:3-27.
32. Yajima, M., K. Hosoda, Y. Kanbayashi, T. Naka-mura, L. Takahashi, and M. Ui. 1978. Biological prop-erties of islets-activating protein (IAP) purified fromthe culture medium ofBordetellapertussis. J. Biochem.(Tokyo) 83:305-312.
VOL. 31, 1981
on April 12, 2020 by guest
http://iai.asm.org/
Dow
nloaded from


















