
PWS Op pad met Fermat
54
PWS Op pad met Fermat Een onderzoek naar het verband tussen het loopgedrag van mieren en Fermats principe van de kortste tijd. E.M. van Sark F.H.B. Somhorst dhr. Booltink 2 januari 2015 Het Assink Lyceum
description
Eindverslag profielwerkstuk 'Op pad met Fermat' Een onderzoek naar het verband tussen het loopgedrag van mieren en Fermats principe van de kortste tijd.
Transcript of PWS Op pad met Fermat

PWS
Op pad met Fermat
Een onderzoek naar het verband tussen het
loopgedrag van mieren en Fermats principe
van de kortste tijd.
E.M. van Sark
F.H.B. Somhorst
dhr. Booltink
2 januari 2015
Het Assink Lyceum

2
Op pad met Fermat
Een onderzoek naar het verband tussen het loopgedrag van mieren en
Fermats principe van de kortste tijd.
Auteur: E.M. van Sark
Kandidaatnummer: 117924
E-mailadres: [email protected]
Profielwerkstuk
Auteur: F.H.B. Somhorst
Kandidaatnummer: 117803
E-mailadres: [email protected]
Profielwerkstuk
Opleiding: Atheneum
Organisatie: Het Assink Lyceum
Begeleider: Dhr. B. Booltink
Externe begeleider: E. Dondergoor
Periode: juni 2014 tot januari 2015
Haaksbergen, 2 januari 2015

3
Voorwoord
Na een paar maanden hard gewerkt te hebben aan ons profielwerkstuk: Op pad
met Fermat ligt voor u het eindresultaat. Wij zijn zeer tevreden over het proces
en eindproduct, maar konden dit nooit behalen zonder hulp van een aantal
begeleiders.
Bij deze willen we graag onze begeleider dhr. Booltink bedanken. Hij heeft ons geholpen met het bedenken van de onderzoeksvraag en het vormen van ons
verslag. Daarnaast heeft hij ons ondersteuning geboden bij alle vragen die wij
hadden omtrent het werkstuk.
Ook willen wij dhr. Dondergoor bedanken voor het uitlenen van zijn mierenkolonie en het geven van advies over het houden van de mieren en het
uitvoeren van de experimenten. Bovendien willen wij hem bedanken voor het ter
beschikking stellen van materiaal om het experiment mee uit te voeren.
Verder gaat onze dank uit naar dhr. Rönitz voor het helpen met het houden van
de mieren en het uitvoeren van de experimenten. Tevens willen wij mevr. Keizer
bedanken voor haar hulp bij het bedenken van een onderwerp.
Eveneens willen wij de gebruikers van antforum.nl bedanken voor hun kennis en
kunde.
Wij wensen u veel leesplezier toe.
Frank Somhorst en Elles van Sark
Haaksbergen, 2 januari 2015.

4
Samenvatting
Uit het onderzoek: Fermat's Principle of Least Time Predicts Refraction of Ant
Trails at Substrate Borders is gebleken dat het looppad van de miersoort
Wasmannia auropunctata voorspeld kan worden met behulp van Fermats
principe van de kortste tijd. Wij willen graag weten of dit ook geldt voor de
Lasius niger. Onze onderzoeksvraag is:
Om een goed antwoord te geven op onze onderzoeksvraag beantwoorden we
eerst een paar deelvragen:
1. Wat houdt Fermats principe van de kortste tijd in?
Lichtstralen leggen de weg af die de minste tijd kost. Bij een overgang van twee
in optisch dichtheid verschillende stoffen is er sprake van breking. We kunnen
constante brekingsindex (n) op twee verschillende manieren berekenen.
en
2. Welke rol spelen feromonen bij de communicatie tussen de mieren van de
soort Lasius niger?
Een feromoon roept een reactie op in een ontvangend organisme. Als mieren op
zoek gaan naar voedsel leggen ze een onderbroken pad van feromonen neer. Als
ze voedsel gevonden hebben, leggen ze op de terugweg een doorlopend pad
neer. Zo roepen ze de andere mieren op om ook te gaan lopen. Uiteindelijk zal er
dus op het pad dat de minste tijd kost het dikste feromonenspoor liggen.
3. Bij welke omstandigheden is de soort Lasius niger gemotiveerd om deel te
nemen aan ons experiment?
Voor ons onderzoek is het belangrijk dat de mieren ondanks het jaargetijde
actief zijn. Hiervoor hebben we een warmtematje op het nest gelegd.
Wij stellen in onze hypothese het volgende: als = , dan is het
mogelijk om met Fermats principe van de kortste tijd de refractie van het
looppad van mierensoort Lasius niger te voorspellen.
Om n op de twee verschillende manieren te berekenen, hebben we mieren over twee verschillende oppervlaktes naar voedsel laten lopen en dit gefilmd. Hierdoor
is het mogelijk om ∠i, ∠r, en te berekenen.
Uit de meetresultaten volgt dat voor beide pogingen die we gedaan hebben
geldt: = 0,6 en = 0,4.
Bij de evaluatie hebben wij berekend dat statistisch gezien onze hypothese
aangenomen mag worden. Onze conclusie luidt dus: Het is wel mogelijk om met
Fermats principe van de kortste tijd de refractie van het looppad van miersoort
Lasius niger te voorspellen.
Het belangrijkste discussiepunt is dat de snelheid niet constant is. Uit de
standaarddeviatie blijkt dat er veel verschil zit tussen de gevonden snelheden.
Daarnaast maakt de miersoort die gebruikt is in ons onderzoek in mindere mate
gebruik van feromonen, waardoor het spoor wellicht niet volstaat.
Is het mogelijk om met Fermats principe van de kortste tijd de refractie
van het looppad van miersoort Lasius niger te voorspellen?

5
Inhoudsopgave
Inleiding 6
Theorie 7
- Wat houdt Fermats principe van de kortste tijd in? 8
- Welke rol spelen feromonen bij de communicatie tussen mieren
van de soort Lasius niger? 12
- Bij welke omstandigheden is de soort Lasius niger gemotiveerd
om deel te nemen aan ons experiment? 16
Onderzoeksvraag 17
Hypothese 18
Werkplan 19
- Meetmethode beeldopnames 19
- Meetmethode 21
- Meetmethode 24
Resultaten 25
- Poging 1 25
- Poging 2 26
Verwerking 27
- Poging 1 27
- Poging 2 28
Evaluatie 29
- Hypothesetoets 29
- Correlatie en regressie 33
Conclusie 34
Discussie 35
Literatuurlijst 37
Bijlagen 39

6
Inleiding
Na wekenlang brainstormen - zonder resultaat - bracht mevr. Keizer ons op het geweldige idee om het gedrag van mieren te bestuderen. Niet alleen wij, maar
ook de wetenschap is gefascineerd door het functioneren van de mierenkolonie
als één geheel waarbij het individu ondergeschikt is aan de groep.
De mens kijkt vaak technieken af van de natuur. Zo heeft men dat ook bij
mieren gedaan. Er zijn verscheidende optimalisatiealgoritmes gemaakt
gebaseerd op het gedrag van mierenkolonies. In het begin ging hier onze interesse naar uit. Gaandeweg bleek dit echter te hoog gegrepen.
Vervolgens stuitten wij op het onderzoek: Fermat’s Principle of Least Time Predicts Refraction of Ant Trails at Substrate Borders. Hierin werd onderzocht of
het looppad van mierensoort Wasmannia auropunctata voorspeld kan worden
met behulp van Fermats principe van de kortste tijd. De wetenschappers concludeerden dat dit het geval is. Hierdoor werden wij benieuwd of dit ook geldt
voor de Lasius niger, de Nederlandse huis-tuin-en-keukenmier.
Onze onderzoeksvraag luidt daarom:
Natuurlijk beschikken wij niet over de meest geavanceerde meetapparatuur, waardoor wij genoodzaakt zijn om het onderzoek op een versimpelde wijze uit te
voeren. Hiervoor is bepaalde theoretische kennis vereist. Voor het theoretische
deel van ons onderzoek proberen wij daarom te achterhalen wat het principe van Fermat eigenlijk inhoudt, hoe mieren met elkaar communiceren en hoe wij de
mieren kunnen motiveren om deel te nemen aan ons onderzoek.
Is het mogelijk om met Fermats principe van de kortste tijd de refractie
van het looppad van miersoort Lasius niger te voorspellen?

7
Theorie Om de kennis die we nodig hebben voor ons experiment te vergaren, hebben wij
de volgende deelvragen geformuleerd:
Wat houdt Fermats principe van de kortste tijd in?
Het is natuurlijk van belang om te weten wat het principe van Fermat inhoudt, voordat we er überhaupt iets mee kunnen proberen te voorspellen.
Welke rol spelen feromonen bij de communicatie tussen de mieren van
de soort Lasius niger?
Het is bekend dat de soort Lasius niger met elkaar communiceert door middel
van feromonen. Het is belangrijk om te weten hoe dit in zijn werk gaat. Op deze
manier weten we welke rol dit speelt tijdens het experiment.
Bij welke omstandigheden is de soort Lasius niger gemotiveerd om deel
te nemen aan ons experiment?
Het onderzoek is uitgevoerd in een periode waarin mieren in een winterslaap
zijn. Daarom moeten we zorgen dat deze winterslaap onderbroken wordt en de optimale activiteit tijdens het experiment behaald wordt.

8
Wat houdt Fermats principe van de kortste tijd in?
Het is de Nederlandse wis- en natuurkundige Willebrordus Snellius van Royen gelukt om dit principe in formulevorm op te schrijven. Deze formule is bekend als
de Wet van Snellius, waarmee voorspeld kan worden hoe een lichtstraal breekt
bij de overgang tussen twee in optische dichtheid verschillende stoffen:
Het principe van Fermat geldt niet alleen voor lichtstralen, maar ook bij andere
verschijnselen. Een tot de verbeelding sprekend voorbeeld hiervan is de strandwacht die intuïtief de snelste route naar een drenkeling neemt. [2] De
strandwacht kan sneller rennen over het strand dan zwemmen in de zee. Hij gaat
dus niet in een rechte lijn naar de drenkeling toe. Hij rent het grootste gedeelte
over het strand en pas op het laatste moment duikt hij de zee in om de drenkeling te redden.
Met behulp van Fermats principe van de kortste weg, de stelling van Pythagoras, goniometrie, differentiaalrekening en de snelheid van de strandwacht op het
strand en in het water is het mogelijk om de theoretisch snelste weg te
berekenen en te construeren.
∠
∠
De wet van Snellius
Waarbij geldt: i = hoek van inval
r = hoek van refractie
t = hoek van terugkaatsing n = een constante, specifiek voor de
overgang van stof I naar stof II
NB: n in de schematische tekening
staat voor de normaal, een lijn die
loodrecht op de grenslijn staat. Schematische tekening lichtstralengang [9]
Schematische tekening snelste weg strandwacht
De weg die een lichtstraal tussen twee punten aflegt, is die welke in de
kortste tijd afgelegd wordt. (Pierre de Fermat, 1657) [12]
9
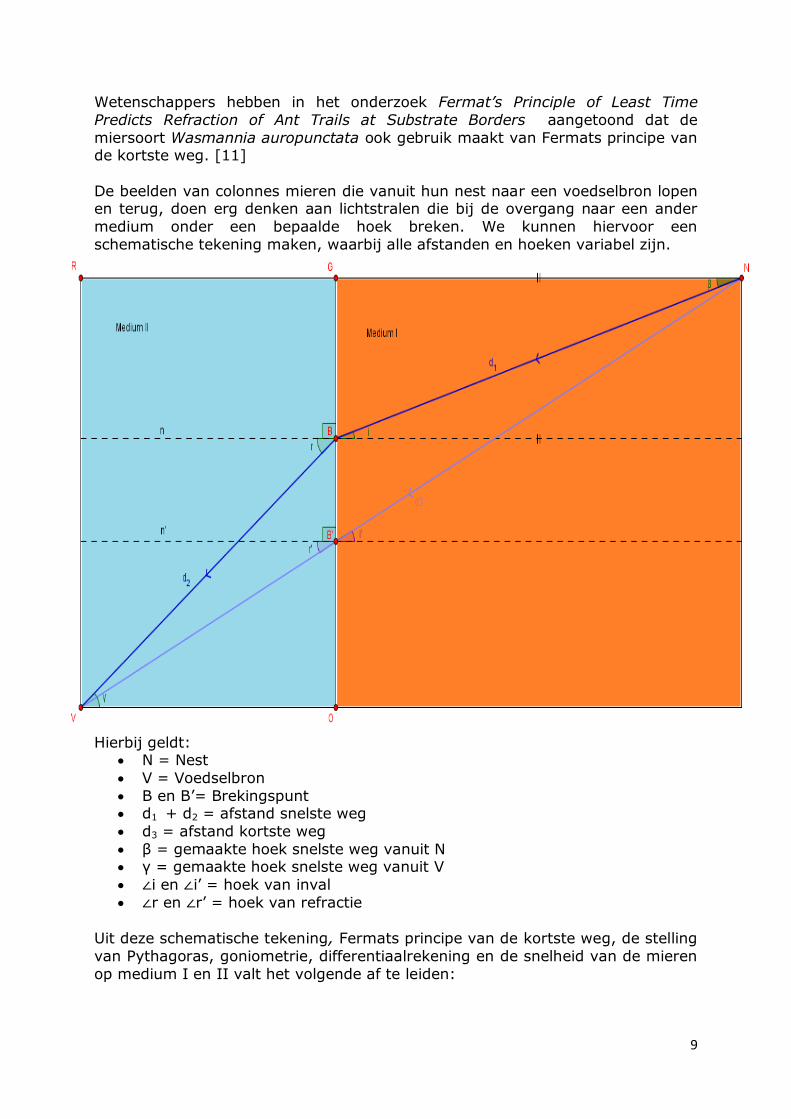
Wetenschappers hebben in het onderzoek Fermat’s Principle of Least Time
Predicts Refraction of Ant Trails at Substrate Borders aangetoond dat de
miersoort Wasmannia auropunctata ook gebruik maakt van Fermats principe van de kortste weg. [11]
De beelden van colonnes mieren die vanuit hun nest naar een voedselbron lopen en terug, doen erg denken aan lichtstralen die bij de overgang naar een ander
medium onder een bepaalde hoek breken. We kunnen hiervoor een
schematische tekening maken, waarbij alle afstanden en hoeken variabel zijn.
Hierbij geldt: N = Nest
V = Voedselbron
B en B’= Brekingspunt d1 + d2 = afstand snelste weg
d3 = afstand kortste weg
β = gemaakte hoek snelste weg vanuit N γ = gemaakte hoek snelste weg vanuit V
∠i en ∠i’ = hoek van inval
∠r en ∠r’ = hoek van refractie
Uit deze schematische tekening, Fermats principe van de kortste weg, de stelling
van Pythagoras, goniometrie, differentiaalrekening en de snelheid van de mieren
op medium I en II valt het volgende af te leiden:

10
Kortste weg
Voor de hoek van inval die gemaakt wordt voor de kortste weg geldt dat deze
gelijk is aan de hoek van refractie voor de kortste weg (overstaande hoeken):
∠ i’ = ∠ r’
Wanneer dit ingevuld wordt in de wet van Snellius, volgt hieruit dat n = 1. Snelste weg
Voor de afstand van de snelste weg geldt:
Voor de afstanden d1 en d2 geldt:
Voor de afstand van de snelste weg geldt:
Ervan uitgaande dat de snelheid op elk medium constant is, is het verband tussen snelheid, afstand en tijd als volgt uit te drukken:
Uit {3} en {4} samen volgt dan:
Hierbij geldt:
v1 = snelheid mier op medium 1 v2 = snelheid mier op medium 2
Om makkelijker te kunnen rekenen stellen we BG = x, dus OB = OG – x Dat betekent:
De afgeleide van t(x) kunnen we als volgt berekenen:
Hieruit volgt:
De kortste tijd is te vinden door de afgeleide van t(x) gelijk te stellen aan 0.
{5} invullen in
geeft:

11
Voor x schrijven we weer BG:
Beide kanten vermenigvuldigen we nu met v1:
Van hoek β weten we het volgende:
Als we hier {1} invullen dan volgt hieruit:
Van hoek γ weten we het volgende:
Als we hier {2} invullen dan volgt hieruit:
Substitueren we {7} en {8} in {6}, dan volgt hieruit:
Uit de schematische tekening kunnen we afleiden:
∠ β = ∠ i (Z-hoeken)
∠ γ = ∠ r (Z-hoeken)
Verder nemen we aan dat
constant is, we stellen daarom dat
Combineren we bovenstaande gegevens met {9}, dan krijgen we:
Deze afleiding is interessant voor ons onderzoek, omdat we in de afleiding gezien
hebben dat we de waarde van n zowel met de snelheden als met de hoeken
kunnen berekenen.
en
Theoretisch zijn deze waardes aan elkaar gelijk.
Op deze manier hebben we het principe van Fermat opgeschreven in
formulevorm. Dit is 'toevallig' de wet van Snellius!

12
Welke rol spelen feromonen bij de communicatie tussen de mieren van
de soort Lasius niger?
Feromonen zijn een subgroep van de semiochemicaliën. Semiochemicaliën zijn
stoffen die zorgen voor communicatie tussen verschillende organismen 15.
Feromonen zijn moleculen die individuen gebruiken om te communiceren met
andere individuen van dezelfde soort . Een feromoon is gedefinieerd als een stof
die wordt vrijgelaten door een organisme naar de buitenwereld, waardoor een
specifieke gedrags- of fysiologische reactie ontstaat bij een ontvangend
organisme van dezelfde soort. 10 Een feromoon is een molecuul met als basis
koolstof en waterstof.
De onderzoekers Peter Karlson en Martin Lüscher hebben in 1959 voor het eerste
de term pheromone gebruikt. Zij hebben dit gebaseerd op de Griekse woorden
pherein (overbrengen) en hormone (stimuleren). 6
Alle organismen maken in enige mate gebruik van feromonen. Het is ook het
oudste systeem dat gedeeld wordt door alle organismen. 5 Er zijn wel limieten
aan het nuttig gebruik van feromonen. De maximale feromonenproductie van
enkele organismen is kleiner dan de mate waarin de feromonen zich verspreiden.
4
Feromonen worden ingedeeld in primerferomonen en releaserferomonen.
Primerferomonen hebben invloed op stofwisseling of ontwikkeling van een
individu en zijn niet reversibel. Releaserferomonen werken rechtstreeks op het
gedragspatroon van een individu en zijn reversibel. Een subgroep van de laatst
genoemde groep, de spoorvolgferomonen, zijn voor ons onderzoek het meest
van belang. 7
Feromonen maken biologische communicatie mogelijk. Aangezien er geen
verwarring mag ontstaan over wat biologische communicatie is, gebruiken wij de
definitie van John Maynard Smith en David Harper. 13
Benzaldehyde (spoorvolgferomoon) 1
13
Biologische communicatie gaat volgens hen niet over het overdragen van
informatie. Een signaal wordt doorgegeven en daarop volgt automatisch een
respons.
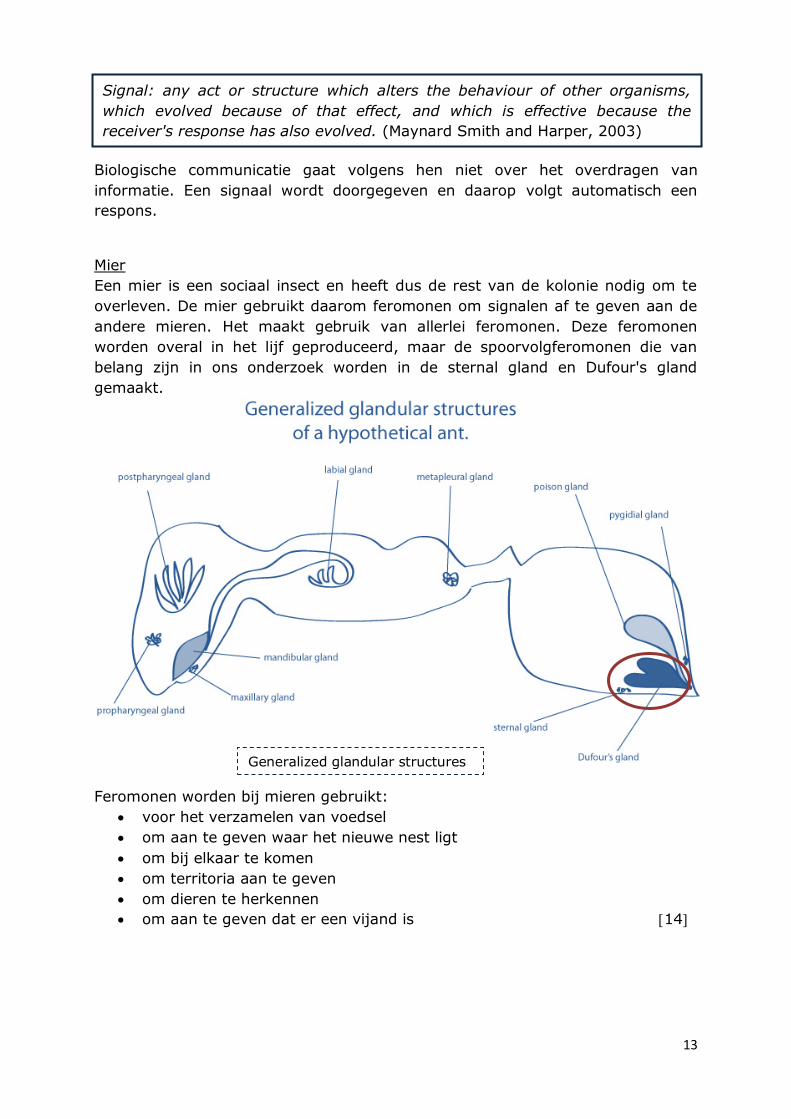
Mier
Een mier is een sociaal insect en heeft dus de rest van de kolonie nodig om te
overleven. De mier gebruikt daarom feromonen om signalen af te geven aan de
andere mieren. Het maakt gebruik van allerlei feromonen. Deze feromonen
worden overal in het lijf geproduceerd, maar de spoorvolgferomonen die van
belang zijn in ons onderzoek worden in de sternal gland en Dufour's gland
gemaakt.
Feromonen worden bij mieren gebruikt:
voor het verzamelen van voedsel
om aan te geven waar het nieuwe nest ligt
om bij elkaar te komen
om territoria aan te geven
om dieren te herkennen
om aan te geven dat er een vijand is 14
Signal: any act or structure which alters the behaviour of other organisms,
which evolved because of that effect, and which is effective because the
receiver's response has also evolved. (Maynard Smith and Harper, 2003)
Generalized glandular structures

14
Feromonen zijn dus voor de vorming van de ingewikkelde sociale samenleving
van mieren van uiterst belang. Mieren hebben daarom een uitzonderlijk goed
reukvermogen. Ze hebben vier tot vijf keer meer geurreceptoren dan andere
insecten. De mier heeft 400 receptoren die verschillende geuren kunnen
opvangen. 16
Homogene oppervlaktes en spoorvolgferomonen
Voor ons onderzoek is het belangrijk om te begrijpen hoe mieren met behulp van
feromonen tot padvorming komen. Het antwoord is eigenlijk erg logisch:
spoorvolgferomonen. Hieronder staat het gedetailleerd beschreven.
(1) Als mieren op zoek gaan naar voedsel leggen ze een onderbroken pad van
feromonen neer.
(2) Als ze terugkeren naar het nest met voedsel maken ze een doorlopend spoor
van feromonen.
(3) De andere individuen van de kolonie reageren hierop. Het volledige spoor
leidt hen naar de bron. In ons onderzoek zal dit voedsel zijn. De mieren leggen
ook allemaal een spoor met feromonen aan. Zolang de voedselbron er is, zullen
mieren het pad volgen en versterken. 8

15
Het pad dat mieren lopen, blijkt het snelst te zijn. Het kost mieren minder tijd
om heen en weer te lopen. De mieren lopen dus vaker en leggen steeds een
feromonenspoor neer. Mieren volgen het pad met de meeste feromonen.
Feromonen verdampen snel, waardoor nutteloze paden niet snel gevolgd worden.
Op dezelfde manier lossen mieren effectief een blokkade op:
Heterogene oppervlaktes en spoorvolgferomonen
Als we de mieren laten lopen op twee oppervlaktes die de loopsnelheid van de
mier verschillend beïnvloeden, is het lastiger om het snelste pad te vinden.
Mieren kunnen hun tijd optimaliseren door Fermats principe van de kortste tijd te
volgen.
Obstakel 3

16
Bij welke omstandigheden is de soort Lasius niger gemotiveerd om deel
te nemen aan ons experiment?
Om tot een antwoord te komen op deze vragen hebben we ze gesteld aan
Evertjan Dondergoor aangezien we zijn mieren hebben gebruikt voor het
uitvoeren van ons experiment.
Bij welke temperatuur zijn mieren actief?
Ik houd ze in een gipsnest op kamertemperatuur (15-19 °C). Mieren kunnen
zonder problemen tegen lagere temperaturen, maar zullen dan minder actief
worden. Om die reden heb ik jullie een warmtematje toegestuurd. Dit kan je
gebruiken om een gedeelte van het nest te verwarmen. Zo heb je meer kans van
slagen met je experiment.
Bij welke lichtintensiteit zijn de mieren actief?
Het is het best om het nest af te dekken en dus zo weinig mogelijk licht toe te
laten in het nest. Daarvoor gebruik ik een kleine handdoek die ik ook heb
meegestuurd.
Hoe vaak en welk voedsel hebben de mieren nodig?
Voedsel heb ik meegedaan in de vorm van suiker-honingwater. Eigenlijk hebben
ze dat niet meer nodig, maar je kunt het gebruiken voor je experiment. Ze zullen
er zeker op af komen. Water hebben ze wel nodig. Bij mij hebben ze altijd
toegang tot een reageerbuis met water. Voor het experiment kan je dit wel even
afsluiten zodat ze sneller gaan lopen voor het water.
Hoe kunnen we een mier verplaatsen?
Je kan een mier verplaatsen met het pincet dat ik heb meegestuurd. Het is een
speciaal ontwerp zodat de mieren niet geplet worden. Dit vergt enige oefening,
maar zal lukken. Zelf gebruik ik ook vaak een reageerbuis, die zet ik dan over de
mier heen die uiteindelijk omhoog zal lopen en dan leg ik de reageerbuis waar ik
hem wil hebben.
Hoe voorkomen we ontsnappende mieren?
Ze zullen niet echt proberen te ontsnappen, maar individuele mieren zullen op
verkenning gaan. En als er ergens een gaatje zit, dan vinden ze dat. Uiteindelijk
gaan ze wel weer gewoon terug naar het nest. In de natuur vallen mieren ook
ten prooi aan van alles, mocht dat dus gebeuren is dat niet echt een probleem.
Mocht er iets met de koningin misgaan, dan is dat het einde van de kolonie. Die
zou ik met rust laten. Gebeurt dat toch, dan is dat weliswaar jammer, maar ik zal
er niet van wakker liggen. Mieren houden niet van zeep of afwasmiddel dus pas
daar mee op.

17
Onderzoeksvraag
Beperkingen
Aangezien wij niet bij machte zijn om een experiment van toereikende omvang
uit te voeren, vereenvoudigen wij de werkelijkheid. We nemen daarom bepaalde
stellingen voor waar aan, zonder deze te bewijzen. Deze stellingen zijn niet perse juist, maar eventuele hierdoor veroorzaakte verschillen worden verwaarloosd in
ons experiment.
We stellen de volgende van deze zogenaamde axioma’s vast:
Onze mierenkolonie van 300 mieren gedraagt zich net zoals de
mierenkolonie in het onderzoek: Fermat’s Principle of Least Time Predicts Refraction of Ant Trails at Substrate.
Een kleinere kolonie kan anders reageren op invloeden van buitenaf. Er zijn
minder mieren die uit het nest treden. Hierdoor kan de uitkomst van het experiment veranderen.
Het jaargetijde heeft geen invloed op de activiteit van mieren.
Mieren hebben normaal in de winter een diapauze (soort winterslaap). Wij
voeren het experiment echter in de winter uit. De mieren zijn over het algemeen minder actief tijdens de diapauze. Hierdoor kan de uitkomst van
het experiment veranderen.
De loopsnelheid van de mieren is constant.
De loopsnelheid van mieren hebben we nodig om de waarde van nsnelheid te
bepalen. Als de loopsnelheid niet constant is, kunnen we deze niet berekenen.
Er wordt een volstaand feromonenspoor gevormd dat alle mieren zullen
volgen.
Feromonen verdampen vrij snel. Als er niet genoeg mieren lopen, is er geen
volstaand feromonenspoor. Hierdoor zoeken de mieren zelf een weg naar het
voedsel. Als er geen feromonenspoor is, ontstaat er dus geen looppad.
Is het mogelijk om met Fermats principe van de kortste tijd de refractie
van het looppad van mierensoort Lasius niger te voorspellen?
Is het mogelijk om met Fermats principe van de kortste tijd de refractie van
het looppad van miersoort Lasius niger te voorspellen?

18
Hypothese
Informatici hebben optimaliseringalgoritmes gemaakt gebaseerd op het gedrag van mierenkolonies. Wij verwachten daarom dat onze mierenkolonie zelf ook de
optimale route naar het voedsel zoekt.
Optimaal zal dan de snelste route zijn, zoals afgeleid kan worden uit het antwoord op de deelvraag: Welke rol spelen feromonen bij de communicatie
tussen de mieren van de soort Lasius niger?. De optimale route kan berekend
worden met behulp van het principe van Fermat, zoals afgeleid kan worden uit het antwoord op de deelvraag: Wat houdt Fermats principe van de kortste tijd
in?.
Indirect kan onze onderzoeksvraag dus beantwoord worden door te onderzoeken of = of ≠ . Hierbij geldt dan dat:
en
Daarbij hebben deze variabelen dezelfde betekenis als in de deelvraag: Wat
houdt Fermats principe van de kortste tijd in?.
Indirect stellen wij in onze hypothese dus het volgende:
Wij verwachten - met inachtneming van eerder geformuleerde axioma’s - dat het mogelijk is om met Fermats principe van de kortste tijd de refractie van
het looppad van miersoort Lasius niger te voorspellen.
Als = , dan is het mogelijk om met Fermats principe van de kortste
tijd de refractie van het looppad van miersoort Lasius niger te voorspellen.
19
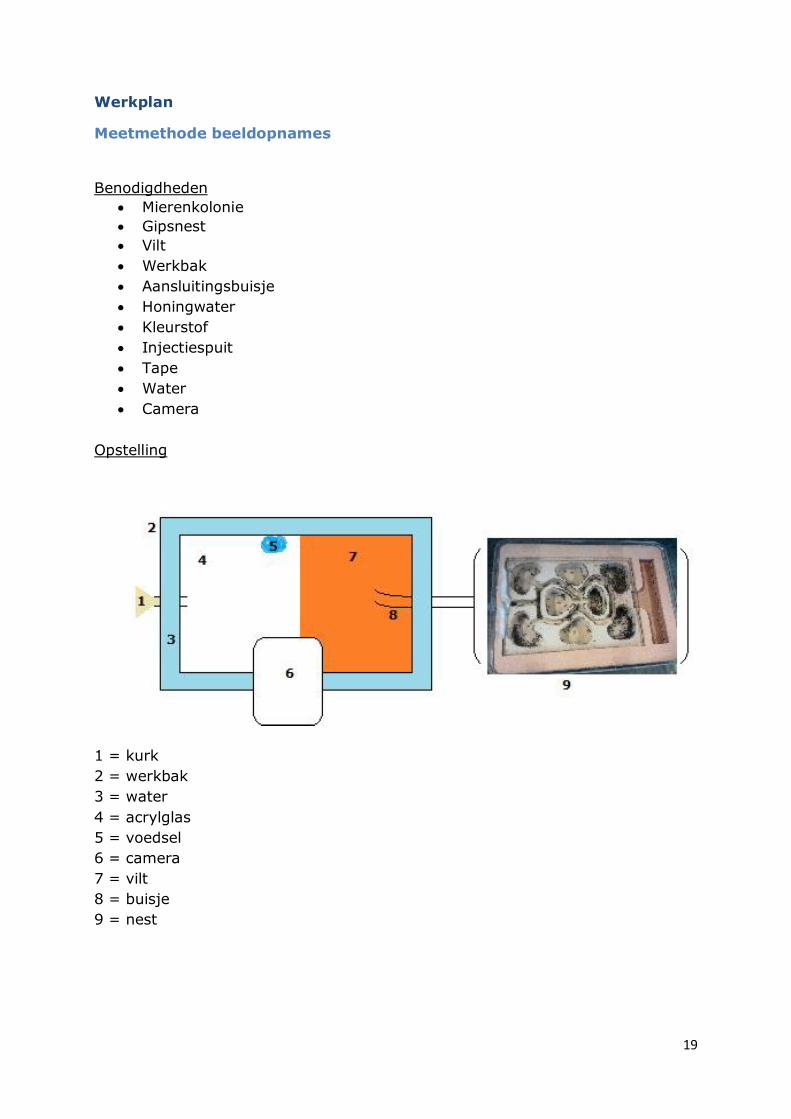
Werkplan
Meetmethode beeldopnames
Benodigdheden
Mierenkolonie
Gipsnest
Vilt
Werkbak
Aansluitingsbuisje
Honingwater
Kleurstof
Injectiespuit
Tape
Water
Camera
Opstelling
1 = kurk
2 = werkbak
3 = water
4 = acrylglas
5 = voedsel
6 = camera
7 = vilt
8 = buisje
9 = nest

20
Werkwijze
1. Zorg ervoor dat de mieren tijdelijk geen toegang hebben tot water, zodat ze
gemotiveerd zijn om naar het honingwater te lopen.
2. Giet water in de buitenste rand van de werkbak, zodat de mieren niet kunnen
ontsnappen.
3. Breng een stukje vilt in een helft van de werkbak en plak dit vast met tape.
4. Mix honingwater met kleurstof en breng dit aan met een injectiespuit bij de
zijkant van het acrylglasgedeelte.
5. Bevestig het buisje op het vilt met een hoek naar links of naar rechts.
6. Bevestig de camera boven de werkbak zodat deze goed in beeld is.
7. Zet de camera aan en zet hem op record.
8. Sluit met het aansluitingsbuisje de mierenkolonie aan.
9. Voer het experiment nogmaals uit.
Noot
Het is belangrijk dat de ondergrond, het voedsel, de temperatuur en de mieren
constant zijn. De hoek van het buisje op het vilt hoeft niet gelijk te zijn. N wordt
namelijk berekend door een verhouding van hoeken en deze zou bij elke hoek
van het buisje gelijk moeten zijn.

21
Meetmethode
Benodigdheden
GeoGebra 5.0
Beeldbewerkingsoftware (Paint) Media Player (Windows Media Player)
Beeldopnames experiment
Werkwijze
1. Start GeoGebra op en maak een nieuw bestand aan.
2. Maak een kader door het punt Lo te fixeren op (0,0), Ro op (24,0) en Lb op
(0,15). Deze punten zijn belangrijk om de verhoudingen te laten kloppen, want
in werkelijkheid waren de maten van het experimentoppervlak 15 x 24 cm:
3. Open de beeldopnames met de Media Player.
4. Vergroot deze zodat het hele beeldscherm opgevuld is met de opname. 5. Zoek een mier uit en maak gedurende de afgelegde weg van nest naar
voedsel een aantal printscreens.
6. Plak deze in beeldbewerkingsoftware. 7. Selecteer alleen het binnenste vlak:
8. Voeg deze afbeelding in in het GeoGebra bestand.
22
9. Zorg ervoor dat het hoekpunt linksonder vast zit aan het punt Lo, rechtsonder
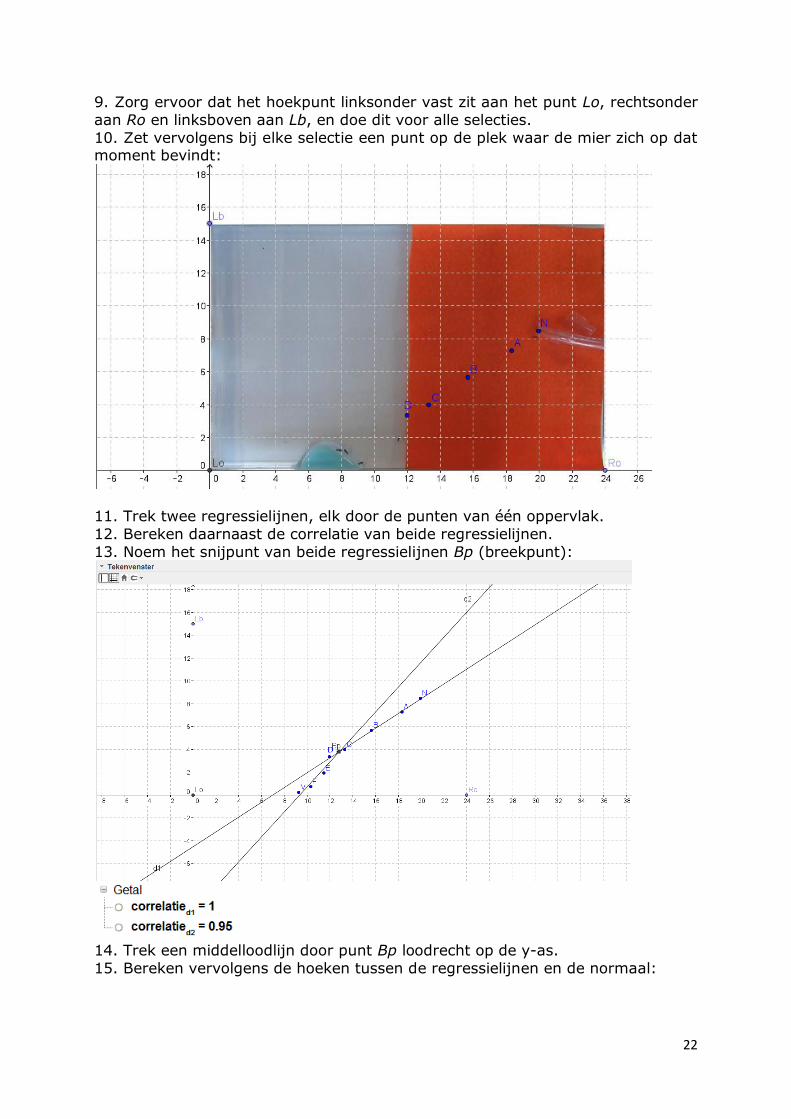
aan Ro en linksboven aan Lb, en doe dit voor alle selecties.
10. Zet vervolgens bij elke selectie een punt op de plek waar de mier zich op dat moment bevindt:
11. Trek twee regressielijnen, elk door de punten van één oppervlak.
12. Bereken daarnaast de correlatie van beide regressielijnen.
13. Noem het snijpunt van beide regressielijnen Bp (breekpunt):
14. Trek een middelloodlijn door punt Bp loodrecht op de y-as.
15. Bereken vervolgens de hoeken tussen de regressielijnen en de normaal:
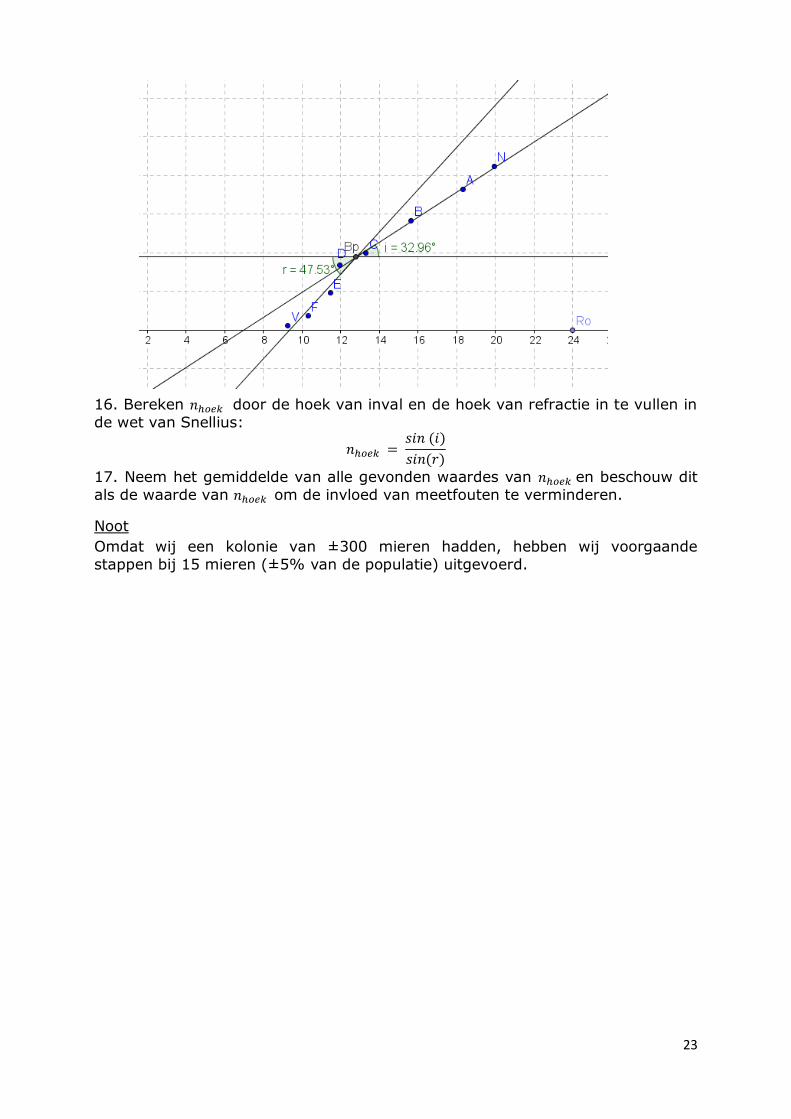
23
16. Bereken door de hoek van inval en de hoek van refractie in te vullen in
de wet van Snellius:
17. Neem het gemiddelde van alle gevonden waardes van en beschouw dit
als de waarde van om de invloed van meetfouten te verminderen.
Noot
Omdat wij een kolonie van ±300 mieren hadden, hebben wij voorgaande
stappen bij 15 mieren (±5% van de populatie) uitgevoerd.

24
Meetmethode
Benodigdheden
Media Player (Windows Media Player) Meetinstrument voor afstand tot op mm nauwkeurig (Geodriehoek)
Kladpapier + schrijfspullen
Stopwatch Beeldopnames
Werkwijze
1. Open de beeldopnames met de Media Player.
2. Meet met behulp van het meetinstrument de lengte op van het binnenste vlak.
3. Berekenen hiermee de verhouding tussen de gemeten waardes en de echte
waardes. Deze waardes zijn belangrijk om de snelheid van de mier in het echt te kunnen berekenen.
4. Kies nu willekeurig een aantal mieren uit op beide oppervlakten. Probeer een interval uit te zoeken waarin de mier een rechte baan beschrijft.
5. Meet de afgelegde afstand van dit interval op en reken deze met behulp van
eerder berekende verhoudingen om tot de afstand die de mier in het echt heeft
afgelegd. 6. Meet met de stopwatch het bijbehorende tijdsinterval op.
7. Bereken nu de snelheid bij elk interval door de afgelegde afstand te delen door het bijbehorende tijdsinterval. Bereken deze snelheid in mm s-1.
8. Bereken v1 door het gemiddelde te nemen van alle gemeten snelheden op
medium I. 9. Bereken v2 door het gemiddelde te nemen van alle gemeten snelheden op
medium II.
Door de gemiddelde snelheid te nemen, neemt de invloed van meetfouten af.
10. Deel de gevonden waarde van v1 door de gevonden waarde van v2 om de
waarde van te berekenen:
Noot
Omdat wij een kolonie van ±300 mieren hadden, hebben wij voorgaande stappen bij 15 mieren (±5% van de populatie) uitgevoerd. We hebben dus de
snelheid van 15 mieren op medium I berekend en de snelheid van 15 mieren op
medium II.
25
Resultaten
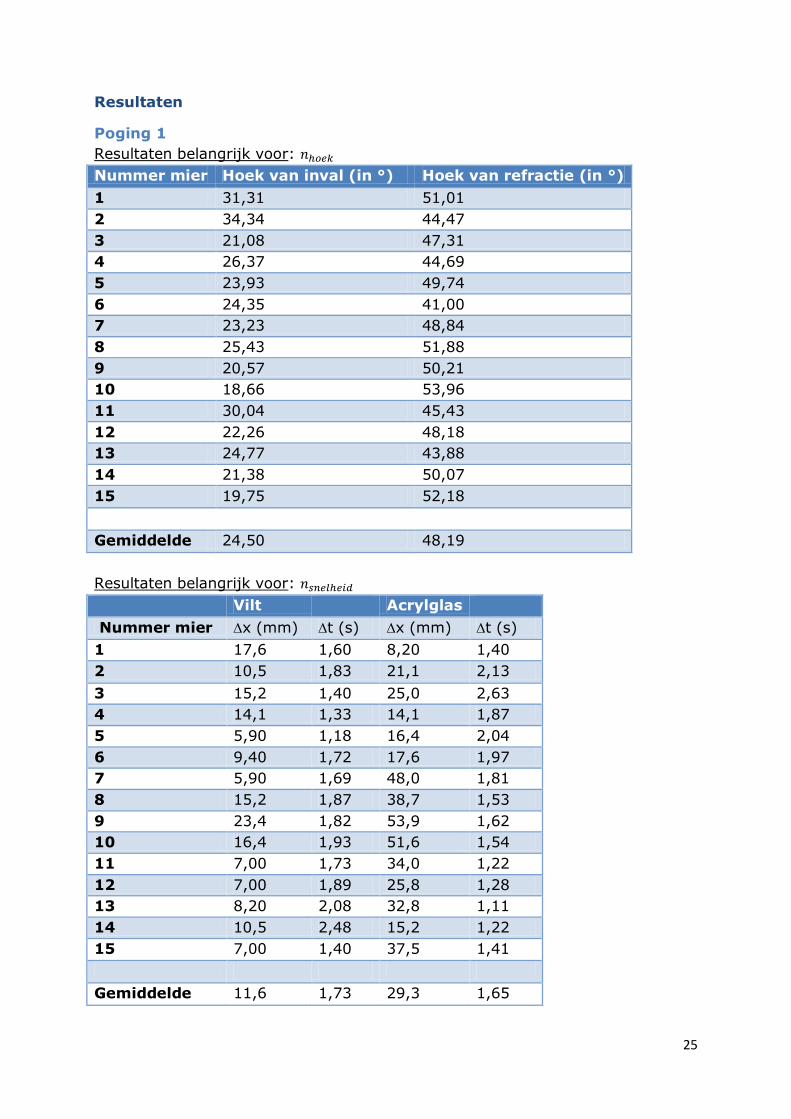
Poging 1
Resultaten belangrijk voor:
Nummer mier Hoek van inval (in °) Hoek van refractie (in °)
1 31,31 51,01
2 34,34 44,47
3 21,08 47,31
4 26,37 44,69
5 23,93 49,74
6 24,35 41,00
7 23,23 48,84
8 25,43 51,88
9 20,57 50,21
10 18,66 53,96
11 30,04 45,43
12 22,26 48,18
13 24,77 43,88
14 21,38 50,07
15 19,75 52,18
Gemiddelde 24,50 48,19
Resultaten belangrijk voor:
Vilt Acrylglas
Nummer mier x (mm) t (s) x (mm) t (s)
1 17,6 1,60 8,20 1,40
2 10,5 1,83 21,1 2,13
3 15,2 1,40 25,0 2,63
4 14,1 1,33 14,1 1,87
5 5,90 1,18 16,4 2,04
6 9,40 1,72 17,6 1,97
7 5,90 1,69 48,0 1,81
8 15,2 1,87 38,7 1,53
9 23,4 1,82 53,9 1,62
10 16,4 1,93 51,6 1,54
11 7,00 1,73 34,0 1,22
12 7,00 1,89 25,8 1,28
13 8,20 2,08 32,8 1,11
14 10,5 2,48 15,2 1,22
15 7,00 1,40 37,5 1,41
Gemiddelde 11,6 1,73 29,3 1,65
26
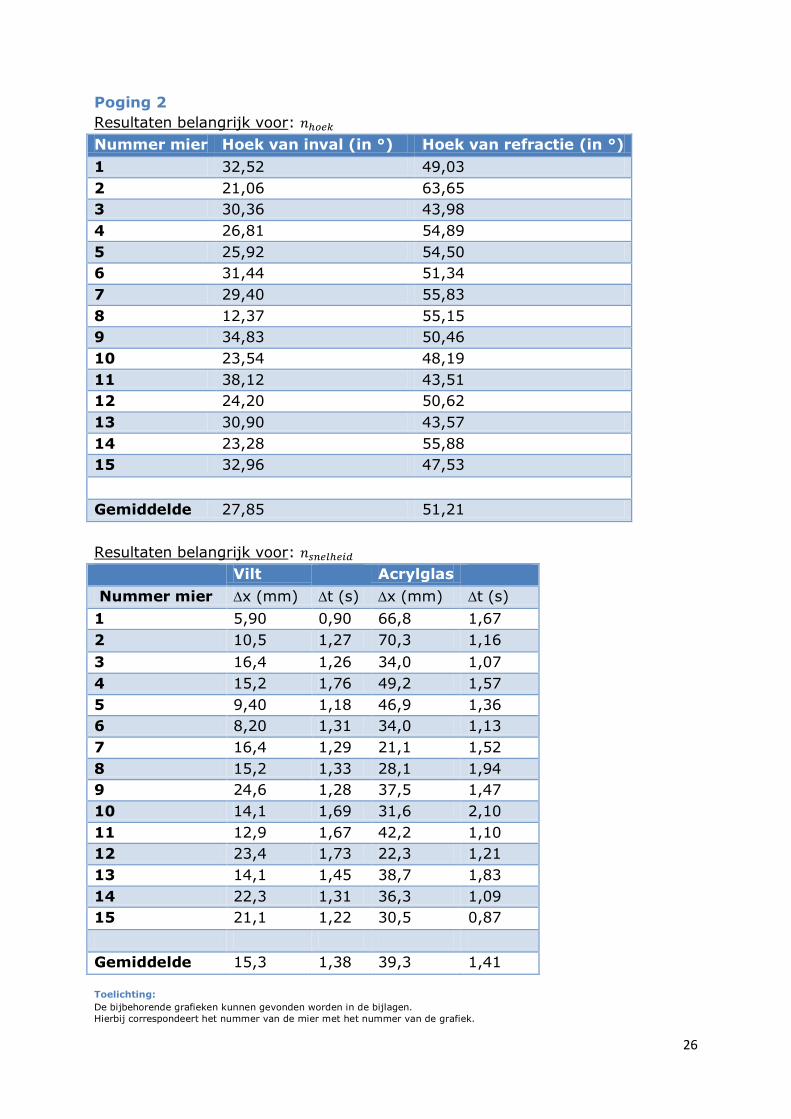
Poging 2
Resultaten belangrijk voor:
Nummer mier Hoek van inval (in °) Hoek van refractie (in °)
1 32,52 49,03
2 21,06 63,65
3 30,36 43,98
4 26,81 54,89
5 25,92 54,50
6 31,44 51,34
7 29,40 55,83
8 12,37 55,15
9 34,83 50,46
10 23,54 48,19
11 38,12 43,51
12 24,20 50,62
13 30,90 43,57
14 23,28 55,88
15 32,96 47,53
Gemiddelde 27,85 51,21
Resultaten belangrijk voor:
Vilt Acrylglas
Nummer mier x (mm) t (s) x (mm) t (s)
1 5,90 0,90 66,8 1,67
2 10,5 1,27 70,3 1,16
3 16,4 1,26 34,0 1,07
4 15,2 1,76 49,2 1,57
5 9,40 1,18 46,9 1,36
6 8,20 1,31 34,0 1,13
7 16,4 1,29 21,1 1,52
8 15,2 1,33 28,1 1,94
9 24,6 1,28 37,5 1,47
10 14,1 1,69 31,6 2,10
11 12,9 1,67 42,2 1,10
12 23,4 1,73 22,3 1,21
13 14,1 1,45 38,7 1,83
14 22,3 1,31 36,3 1,09
15 21,1 1,22 30,5 0,87
Gemiddelde 15,3 1,38 39,3 1,41
Toelichting:
De bijbehorende grafieken kunnen gevonden worden in de bijlagen.
Hierbij correspondeert het nummer van de mier met het nummer van de grafiek.
27
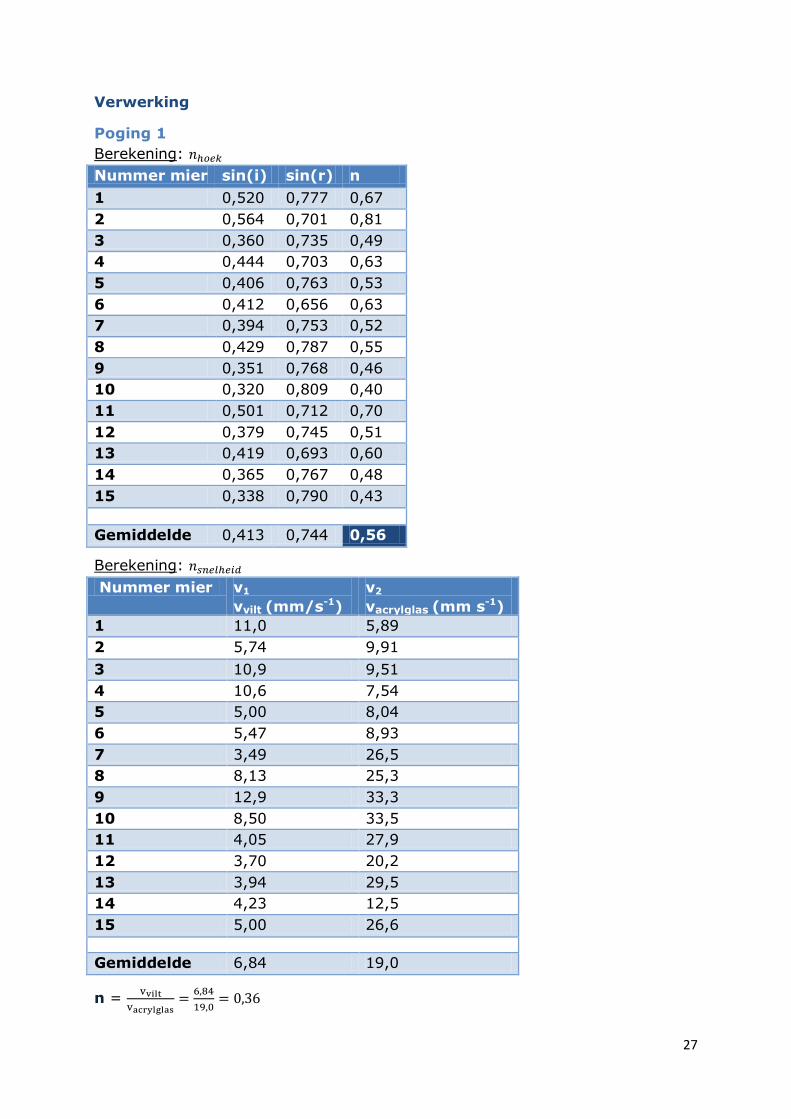
Verwerking
Poging 1
Berekening:
Nummer mier sin(i) sin(r) n
1 0,520 0,777 0,67
2 0,564 0,701 0,81
3 0,360 0,735 0,49
4 0,444 0,703 0,63
5 0,406 0,763 0,53
6 0,412 0,656 0,63
7 0,394 0,753 0,52
8 0,429 0,787 0,55
9 0,351 0,768 0,46
10 0,320 0,809 0,40
11 0,501 0,712 0,70
12 0,379 0,745 0,51
13 0,419 0,693 0,60
14 0,365 0,767 0,48
15 0,338 0,790 0,43
Gemiddelde 0,413 0,744 0,56
Berekening:
Nummer mier v1
vvilt (mm/s-1)
v2
vacrylglas (mm s-1)
1 11,0 5,89
2 5,74 9,91
3 10,9 9,51
4 10,6 7,54
5 5,00 8,04
6 5,47 8,93
7 3,49 26,5
8 8,13 25,3
9 12,9 33,3
10 8,50 33,5
11 4,05 27,9
12 3,70 20,2
13 3,94 29,5
14 4,23 12,5
15 5,00 26,6
Gemiddelde 6,84 19,0
n =
28
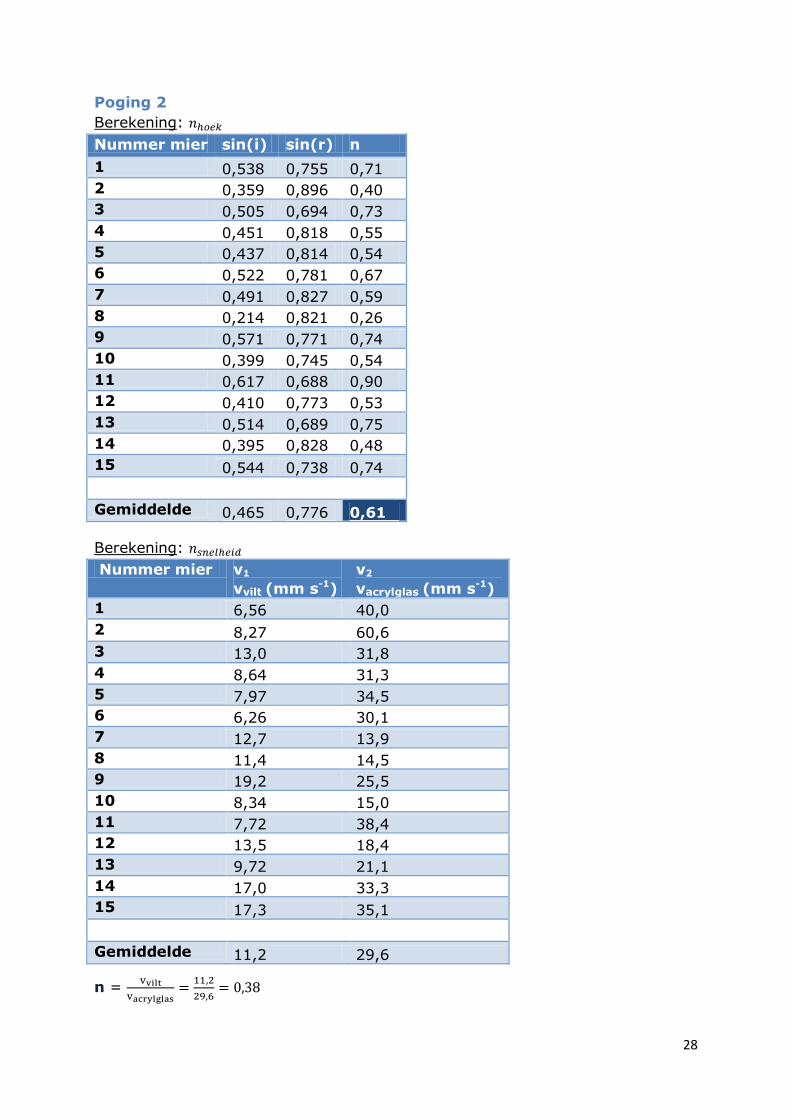
Poging 2
Berekening:
Nummer mier sin(i) sin(r) n
1 0,538 0,755 0,71
2 0,359 0,896 0,40
3 0,505 0,694 0,73
4 0,451 0,818 0,55
5 0,437 0,814 0,54
6 0,522 0,781 0,67
7 0,491 0,827 0,59
8 0,214 0,821 0,26
9 0,571 0,771 0,74
10 0,399 0,745 0,54
11 0,617 0,688 0,90
12 0,410 0,773 0,53
13 0,514 0,689 0,75
14 0,395 0,828 0,48
15 0,544 0,738 0,74
Gemiddelde 0,465 0,776 0,61
Berekening:
Nummer mier v1
vvilt (mm s-1)
v2
vacrylglas (mm s-1)
1 6,56 40,0
2 8,27 60,6
3 13,0 31,8
4 8,64 31,3
5 7,97 34,5
6 6,26 30,1
7 12,7 13,9
8 11,4 14,5
9 19,2 25,5
10 8,34 15,0
11 7,72 38,4
12 13,5 18,4
13 9,72 21,1
14 17,0 33,3
15 17,3 35,1
Gemiddelde 11,2 29,6
n =

29
Evaluatie
Hypothesetoets
Natuurlijk is het zelden zo dat de onderzoeksresultaten precies overeenkomen
met de theorie, doordat toevallige meetfouten deze beïnvloeden. Gelukkig is het
mogelijk om onze hypothese aan te nemen/te verwerpen dankzij de stellingen
van de statistiek.
Volgens de theorie zou gelijk moeten zijn aan , oftewel:
Echter blijkt uit ons experiment dat voor beide pogingen geldt:
. Betekent dit dan dat we onze hypothese moeten verwerpen?
De statistiek veronderstelt dat de gevonden waardes ‘normaal verdeeld’ zijn. Dit
houdt in dat de gevonden waardes symmetrisch verdeeld zijn rond een centrale
waarde (het gemiddelde, bij ons dus de theoretische waarde) in een soort
‘klokvorm’. Afwijkingen van deze centrale waarde worden steeds
onwaarschijnlijker naarmate de afwijking groter is. De kans op bepaalde waardes
kan op verschillende manieren berekend worden . In het kort komt het erop neer
dat de oppervlakte van de normaalkromme precies gelijk aan 1 is (100%), en
dat de rekenmachine vervolgens de integraal van de afwijking neemt om de
bijbehorende verwachtingswaarde te berekenen.
De vraag is nu of onze hypothese aangenomen mag worden of verworpen dient
te worden. We toetsen daarom onze ‘nulhypothese’ aan de ‘alternatieve
hypothese’:
Om te beslissen of onze verwachtingswaarde significant genoeg is, stellen we de
grens op 5% (deze waarde kiest men vaak in de statistiek). Dit betekent dat de
verwachtingswaarde groter moet zijn dan 0,025 (de helft van 0,05; de afwijking
kan namelijk te veel of te weinig zijn) om aan te nemen.
In onze situatie is de centrale waarde 0 en de afwijking waarvan we de
verwachtingswaarde moeten berekenen het interval (A):

30
Hoe groot is nu de kans dat de afwijking groter of gelijk aan 0,2 is?
Bij de berekening van de overschrijdingskans moet men gebruik maken van
,
dit volgt uit de -wet:
Logisch, want hoe groter de steekproef hoe dichter je de populatie benadert en
dus hoe dichter je bij het werkelijke gemiddelde zit. Bij beide pogingen bestond
de steekproef uit 15 mieren, dus in beide gevallen.
De standaardafwijking, ook wel standaarddeviatie genoemd, is een maat voor de
spreiding van uitkomsten rondom een gemiddelde. Hoe hoger deze waarde is,
des te meer verschillen de waardes van elkaar.
De standaardafwijking is als volgt te berekenen:
Om één standaardafwijking te berekenen uit twee standaardafwijkingen, moet de
volgende formule gehanteerd worden:
Formule standaardafwijking
Waarbij geldt: X = waarnemingsgetal n = aantal waarnemingsgetallen
= gemiddelde van de waarneminggetallen
Formule uit
Waarbij geldt: = standaardafwijking uit = standaardafwijking van X = standaardafwijking van Y
Wortel-n-wet:
Hoe groter de steekproef, hoe kleiner de
standaardafwijking.
31
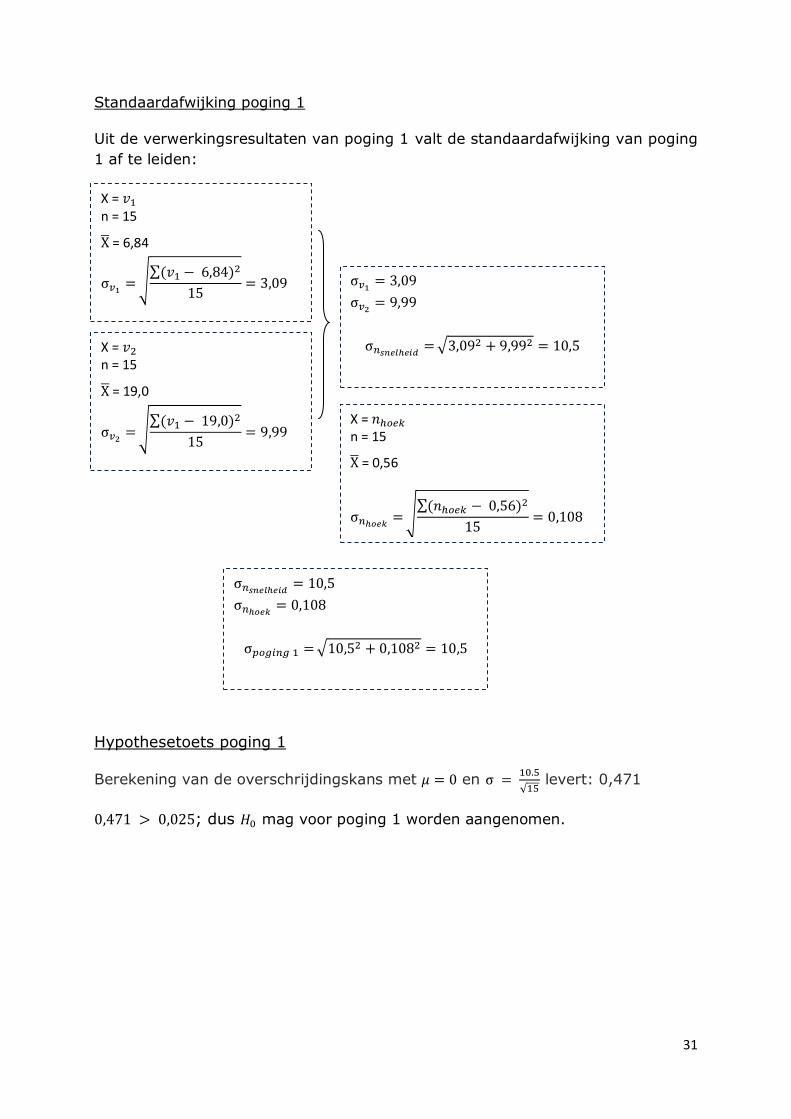
Standaardafwijking poging 1
Uit de verwerkingsresultaten van poging 1 valt de standaardafwijking van poging
1 af te leiden:
Hypothesetoets poging 1
Berekening van de overschrijdingskans met en
levert: 0,471
; dus mag voor poging 1 worden aangenomen.
X = n = 15
= 0,56
X = n = 15
= 6,84
X = n = 15
= 19,0
32
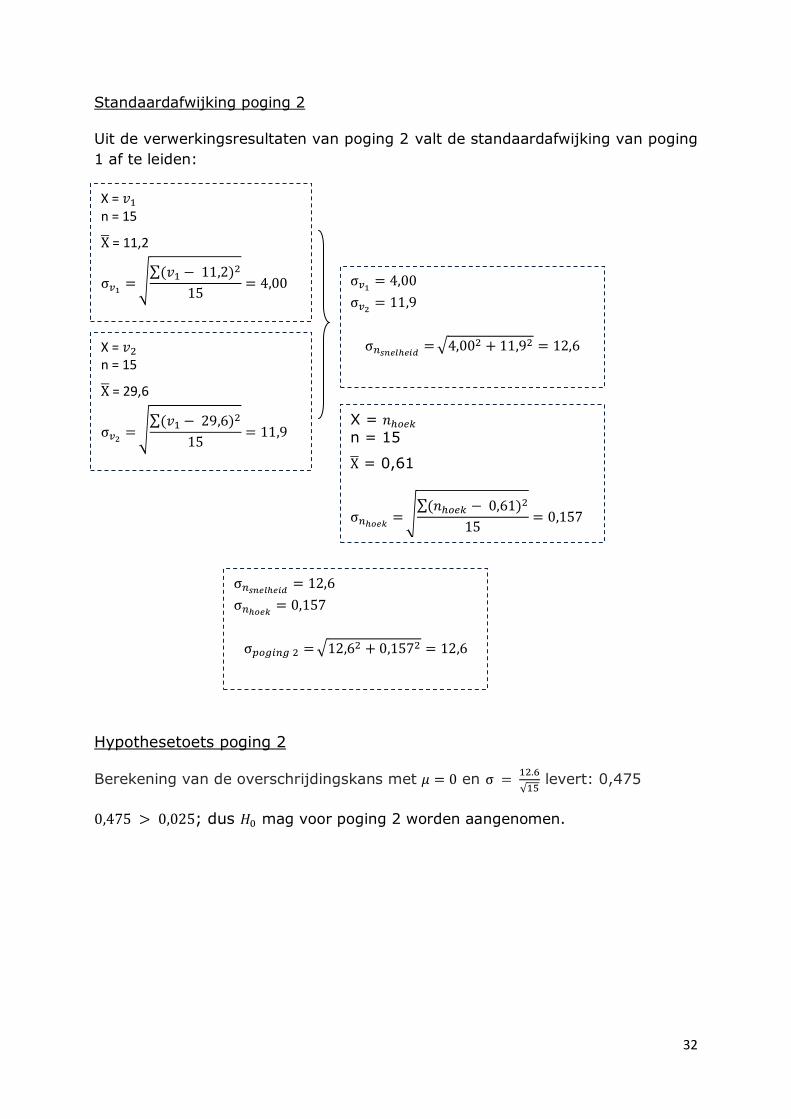
Standaardafwijking poging 2
Uit de verwerkingsresultaten van poging 2 valt de standaardafwijking van poging
1 af te leiden:
Hypothesetoets poging 2
Berekening van de overschrijdingskans met en
levert: 0,475
; dus mag voor poging 2 worden aangenomen.
X =
n = 15
= 0,61
X = n = 15
= 11,2
X = n = 15
= 29,6

33
Correlatie en regressie
Om te onderzoeken of er een sterk lineair verband bestaat tussen de
‘bewegingspunten’ (de plek waar de mier zich bevond op een willekeurig tijdstip)
van de mieren op zowel medium I als medium II, kan de correlatiecoëfficiënt als
maat gebruikt worden. Afhankelijk van of de looppadlijn stijgt of daalt, is de
optimale waarde van de correlatiecoëfficiënt respectievelijk 1 of -1. Deze
waardes geven dan namelijk aan dat er een heel sterk lineair verband is tussen
de ‘bewegingspunten’, en dus alle punten op één lijn liggen. In het meest
ongunstige geval is de correlatiecoëfficiënt gelijk aan 0. Dit betekent dat er dan
geen enkel lineair verband bestaat tussen de ‘bewegingspunten’.
Uit onze meetresultaten valt af te leiden dat er geen enkele correlatiecoëfficiënt
lager dan 0,79 is, in de meeste gevallen is er zelfs een correlatiecoëfficiënt van
bijna 1. We kunnen dus stellen dat er zeker een lineair verband bestaat tussen
de ‘bewegingspunten’.
Dit lineaire verband kan vervolgens zichtbaar gemaakt worden door GeoGebra
een regressielijn te laten tekenen. Deze regressielijn is de ‘best passende’ lijn bij
de verzameling meetgegevens en is gemaakt met behulp van de zogeheten
‘kleine kwadraten methode’. Bij deze methode moeten de sommen van de
kwadraten van het verschil tussen de Y-waarde van elk meetpunt en de Y-
waarde van bijbehorend punt op de ‘best passende’ lijn zo klein mogelijk zijn. De
regressielijn die zo ontstaat, is de beste schatting voor het verband tussen de
‘bewegingspunten’.

34
Conclusie Onze hypothese luidt:
Uit de meetresultaten volgt dat voor beide pogingen geldt:
Bij de evaluatie hebben wij berekend dat statistisch gezien onze hypothese
aangenomen mag worden. Dit betekent dus dat het wel mogelijk is om met Fermats principe van de kortste tijd de refractie van het looppad van miersoort
Lasius niger te voorspellen.
Als = , dan is het mogelijk om met Fermats principe van de kortste
tijd de refractie van het looppad van miersoort Lasius niger te voorspellen.
= 0,6
= 0,4

35
Discussie
Twijfelachtige meetresultaten
Uit de dataset met meetresultaten van de snelheid en de bijbehorende standaarddeviatie blijkt dat deze meetresultaten redelijk ver uit elkaar liggen.
Echter geven de meetresultaten geen aanleiding om te twijfelen aan de theorie.
Wel geven ze aanleiding om te twijfelen aan onze aannames.
Foutieve aannames?
De aanname dat er een volstaand feromonenspoor gevormd wordt dat alle
mieren zullen volgen, blijkt onjuist te zijn. Waarschijnlijk zijn er gedurende de beeldopnames feromonen verdampt, aangezien de mieren nooit de optimale
route lopen. Dat kan komen doordat in tegenstelling tot het onderzoek Fermat’s
Principle of Least Time Predicts Refraction of Ant Trails at Substrate Borders er niet zoveel mieren in de opstelling liepen, laat staan bijna in een rij achter
elkaar.
We hebben van dhr. Dondergoor (software-ingenieur) een zogenaamde
‘heatmap’ toegestuurd gekregen. De heatmap laat zien op welke plaatsen de
mieren regelmatig hebben gelopen. Hoe roder de plek, hoe vaker mieren op die
plaats gelopen hebben. We kunnen uit de theorie afleiden dat op de rode plekken een feromonenspoor gevormd is. Dit is dus duidelijk niet optimaal gebeurd:
Uit de heatmap is af te leiden dat er meerdere paden vaak bewandeld zijn. Dit
kan meerdere oorzaken hebben:
- De camera is tijdens de opname van het experiment verschoven,
waardoor het pad ook verschoven is. - Het vilt was niet vlak. Het buisje van de nestuitgang zat vastgeplakt aan
het vilt en hierdoor werd het vilt iets omhooggetrokken.
- Er zat een relatief groot hoogteverschil tussen het vilt en het acrylglas. Dit verklaart de rechte lijn langs de grenslijn (blauwe lijn in de heatmap).
Heatmap experiment

36
Verder hebben wij aangenomen dat de snelheden op beide oppervlaktes constant
zijn. Gezien de relatief grote standaardafwijking tussen de meetresultaten bleek deze verre van juist te zijn. In tegenstelling tot de mieren die in het onderzoek
Fermat’s Principle of Least Time Predicts Refraction of Ant Trails at Substrate
Borders gebruikt zijn, kunnen we ervan uitgaan dat onze mieren niet puur liepen om voedsel te vergaren. De experts van antforum.nl hebben met een kritische
blik naar onze beelden gekeken. Zij concludeerden dat de mieren ook op zoek
waren naar nieuwe nestruimte en op verkenning gingen. Hierdoor liepen de
mieren soms langzamer dan gebruikelijk is bij het vergaren van voedsel.
Daarnaast maakt de miersoort die in het onderzoek Fermat’s Principle of Least
Time Predicts Refraction of Ant Trails at Substrate Borders gebruikt is, de Wasmannia auropunctata meer gebruik van feromonen dan de miersoort die wij
gebruikt hebben in ons onderzoek, de Lasius niger. De aanname dat onze
mierenkolonie zich hetzelfde gedraagt als de mierenkolonie uit het onderzoek blijkt dus ook onjuist te zijn.
Ook is het vloeibare voedsel uitgelopen, waardoor het voedselpunt verschoof. Dit
heeft er waarschijnlijk toe geleid dat er telkens nieuwe paden gevormd werden, wat kan verklaren waarom de hoeken van inval en hoeken van refractie
onderling verschillen. Het was misschien beter geweest om het vloeibare voedsel
in een recipiënt te stoppen, zodat het vloeibare voedsel duidelijk op één punt bleef.
Het vilt bleek tijdens het onderzoek ook niet helemaal goed vast te zitten op de bodem, aangezien er soms mieren waren die onder het vilt kropen. Mogelijk
heeft dit ook invloed gehad op het feromonenpad.
Gemotiveerde mieren?
Een gebrek aan intrinsieke motivatie kan mogelijk een reden zijn waarom er niet
zoveel mieren in de opstelling rond lopen, of op sommige momenten helemaal
geen. De aanname dat het jaargetijde geen invloed heeft op de activiteit van de mieren kan daarom onjuist zijn. De mieren hadden eigenlijk in een soort
winterslaap moeten zitten, wat ook impliceert dat ze al voldoende voedsel gehad
hadden. We hebben geprobeerd ze toch te motiveren door een dag van te voren
de toegang tot voedsel/water af te sluiten, om de mieren op deze manier te motiveren op zoek te gaan naar voedsel/water tijdens het experiment. Door het
warmtematje hebben we tevens geprobeerd de invloed van de winterse
temperaturen uit te schakelen. Ook moet gezegd worden dat niet alle mieren die in de opstelling rondliepen op zoek waren naar voedsel/water, maar dat sommige
ook op verkenning waren op zoek naar een mogelijk betere plek voor het nest.
Opstelling
Onze opstelling (15 x 24 cm) is beduidend kleiner dan de opzet in het onderzoek
Fermat’s Principle of Least Time Predicts Refraction of Ant Trails at Substrate
Borders (50 x 50 cm). Een groter oppervlak had mogelijk nog mooiere brekingshoeken op kunnen leveren, hoewel dat waarschijnlijk niet het geval was
geweest aangezien onze mierenkolonie een stuk kleiner was en er ook niet
zoveel mieren in de opstelling rondliepen.

37
Literatuurlijst
[1] BENZALDEHYDE [Illustratie]. (2013, 06 september). Geraadpleegd op http://infokimiawan13o1b-6.blogspot.nl/2013/09/bahan-bahan-
kimia_6.html
[2] Bertone, G. (2014). Achter de schermen van het heelal. Amsterdam,
Nederland: Prometheus.
[3] BioNinja. (z.j.). Pheromones. Geraadpleegd op
http://www.vce.bioninja.com.au/aos-2-detecting-and- respond/coordination--regulation/pheromones.html
[4] Dusenbery, D. B. (2009). Living at Micro Scale (Harvard Universtiy Press, Cambridge). Geraadpleegd op
http://www.hup.harvard.edu/catalog.php?isbn=9780674060210
[5] Hildebrand, J. G. (1995). Analysis of chemical signals by nervous systems
(Proc. Natl. Acad. Sci. U.S.A. 92 (1): 67–74). Geraadpleegd op
http://www.pnas.org/content/92/1/67.abstract
[6] Karlson, P., & Lüscher, M. (1959). Pheromones: a new term for a class of
biologically active substances (Nature 183). Geraadpleegd op
http://www.nature.com/nature/journal/v183/n4653/abs/183055a0.html
[7] Kleerebezem, M., & Quadri, L. E. (2001). Peptide pheromone-dependent
regulation of antimicrobial peptide production in Gram-positive bacteria: a case of multicellular behavior. Geraadpleegd op
http://www.sciencedirect.com/science/article/pii/S0196978101004934
[8] Moglich, M., Maschwitz, U., & Holldobler, B. (1974). Tandem calling: A new kind of signal in ant communication (Science (Wash.)). Geraadpleegd op
http://www.ncbi.nlm.nih.gov/pubmed/4469698
[9] Molenaar, L. (2003). Marcel Minnaert, astrofysicus 1893-1970.
Amsterdam, Nederland: Balans.
[10] Nordlund, D. A., & Lewis, W. J. (1976). Terminology of chemical releasing
stimuli in intraspecific and interspecific interactions (J. Chem. Ecol., Vol 2,
No. 2, pp. 211-220.). Geraadpleegd op
http://link.springer.com/article/10.1007%2FBF00987744?LI=true
[11] Oettler J, Schmid V.S., Zankl N., Rey O., Dress A., Heinze J. (2013)
Fermat’s Principle of Least Time Predicts Refraction of Ant Trails at Substrate Borders. PLoS ONE 8(3): e59739.
doi:10.1371/journal.pone.0059739

38
[12] Rozendaal, J., & Toledano, Y. (2010). Fermat: het werk van een
fascinerende wiskundige. Geraadpleegd op
http://books.google.nl/books?id=OSjpAQAAQBAJ&dq=principe+van+fermat+voorbeelden&hl=nl&source=gbs_navlinks_s
[13] Smith, A. D. M., Schouwstra, M., Smith, K., & Boer, B. de. (2010). The Evolution of Language. Geraadpleegd op
https://books.google.nl/books?id=xmn6EPIa8VoC&pg=PA130&lpg=PA130
&dq=Signal:+any+act+or+structure+which+alters+the+behaviour+of+ot
her+organisms,+which+evolved+because+of+that+effect,+and+which+is +effective+because+the+receiver%27s+response+has+also+evolved.+%
28&source=bl&ots=HA_MvYPxFD&sig=nCxknbhNsAArPdJuaQCs8zY79PQ&h
l=nl&sa=X&ei=uUWpVIGYM6y17gab54DoDg&ved=0CCEQ6AEwAA#v=one page&q=Signal%3A%20any%20act%20or%20structure%20which%20alte
rs%20the%20behaviour%20of%20other%20organisms%2C%20which%2
0evolved%20because%20of%20that%20effect%2C%20and%20which%20 is%20effective%20because%20the%20receiver%27s%20response%20ha
s%20also%20evolved.%20%28&f=false
[14] Wheeler, W. M. (1960). Ants : their structure, development and behavior. Geraadpleegd op
http://books.google.nl/books?id=KjHRuG5nfs4C&printsec=frontcover&hl=
nl&source=gbs_ge_summary_r&cad=0#v=onepage&q&f=false
[15] Wyatt, TD (2003). Pheromones and Animal Behaviour (ISBN 0 521
48526 6). Geraadpleegd op http://books.google.nl/books?hl=nl&lr=&id=aty3hz6rdlgC&oi=fnd&pg=PP1
5&dq=pheromones+and+animal+behaviour+T.D.+Wyatt&ots=nZTg4gZjN
U&sig=7uPDheWj42VUwuGHqfQJpD9zCD8#v=onepage&q=pheromones%
20and%20animal%20behaviour%20T.D.%20Wyatt&f=false
[16] Zwiebel, L. (2012). Ants have an exceptionally high-def sense of smell.
Geraadpleegd op http://news.vanderbilt.edu/2012/09/ants-have-an- exceptionally-high-def-sense-of-smell/
39
Bijlagen
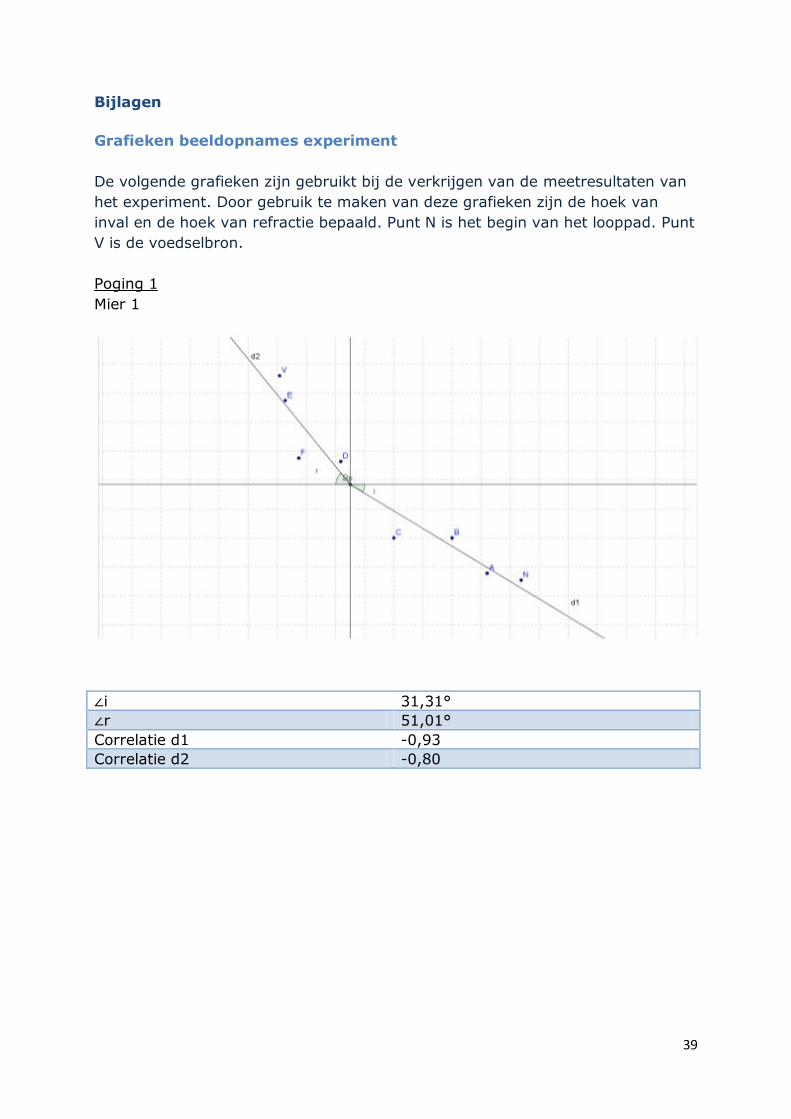
Grafieken beeldopnames experiment
De volgende grafieken zijn gebruikt bij de verkrijgen van de meetresultaten van
het experiment. Door gebruik te maken van deze grafieken zijn de hoek van
inval en de hoek van refractie bepaald. Punt N is het begin van het looppad. Punt
V is de voedselbron.
Poging 1
Mier 1
∠i 31,31°
∠r 51,01°
Correlatie d1 -0,93
Correlatie d2 -0,80
40
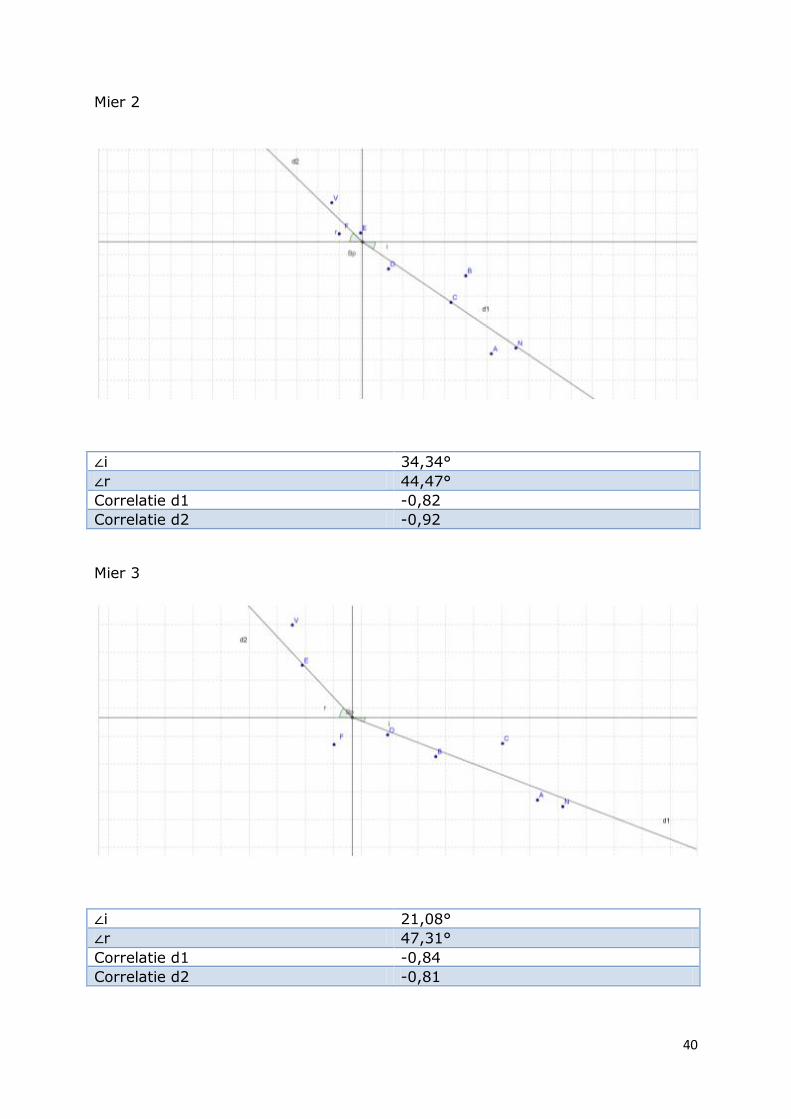
Mier 2
∠i 34,34°
∠r 44,47°
Correlatie d1 -0,82
Correlatie d2 -0,92
Mier 3
∠i 21,08°
∠r 47,31°
Correlatie d1 -0,84
Correlatie d2 -0,81
41
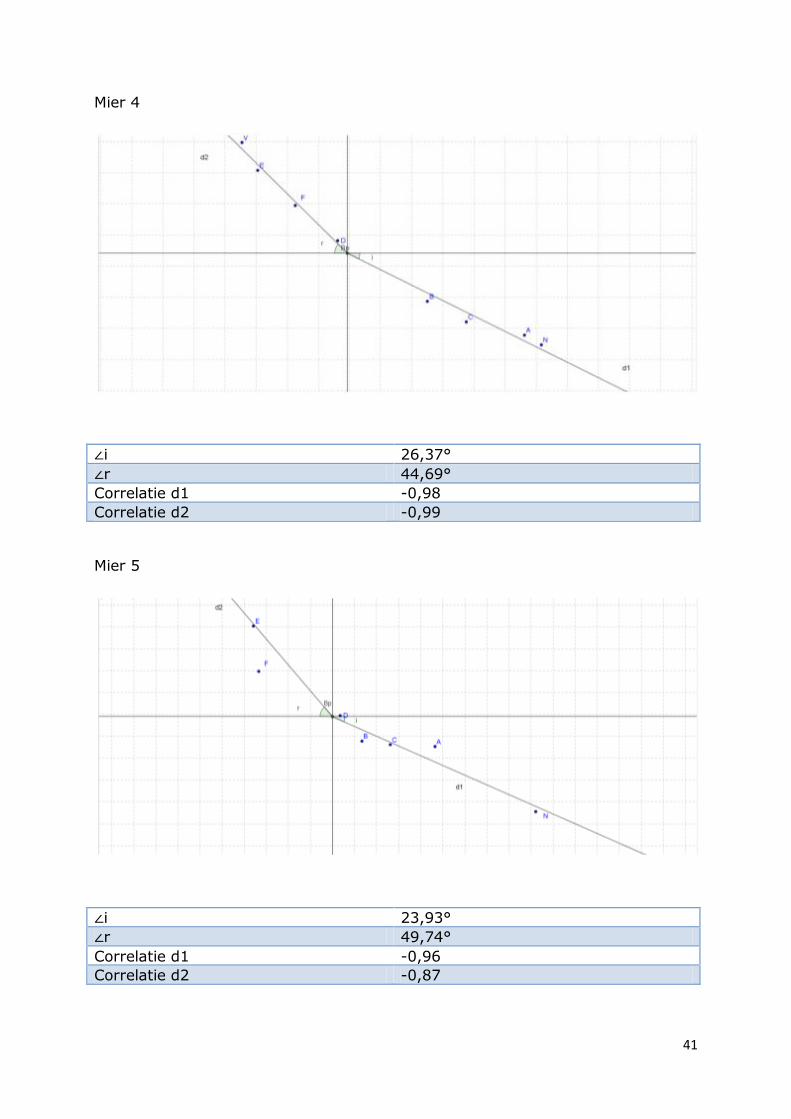
Mier 4
∠i 26,37°
∠r 44,69°
Correlatie d1 -0,98
Correlatie d2 -0,99
Mier 5
∠i 23,93°
∠r 49,74°
Correlatie d1 -0,96
Correlatie d2 -0,87
42
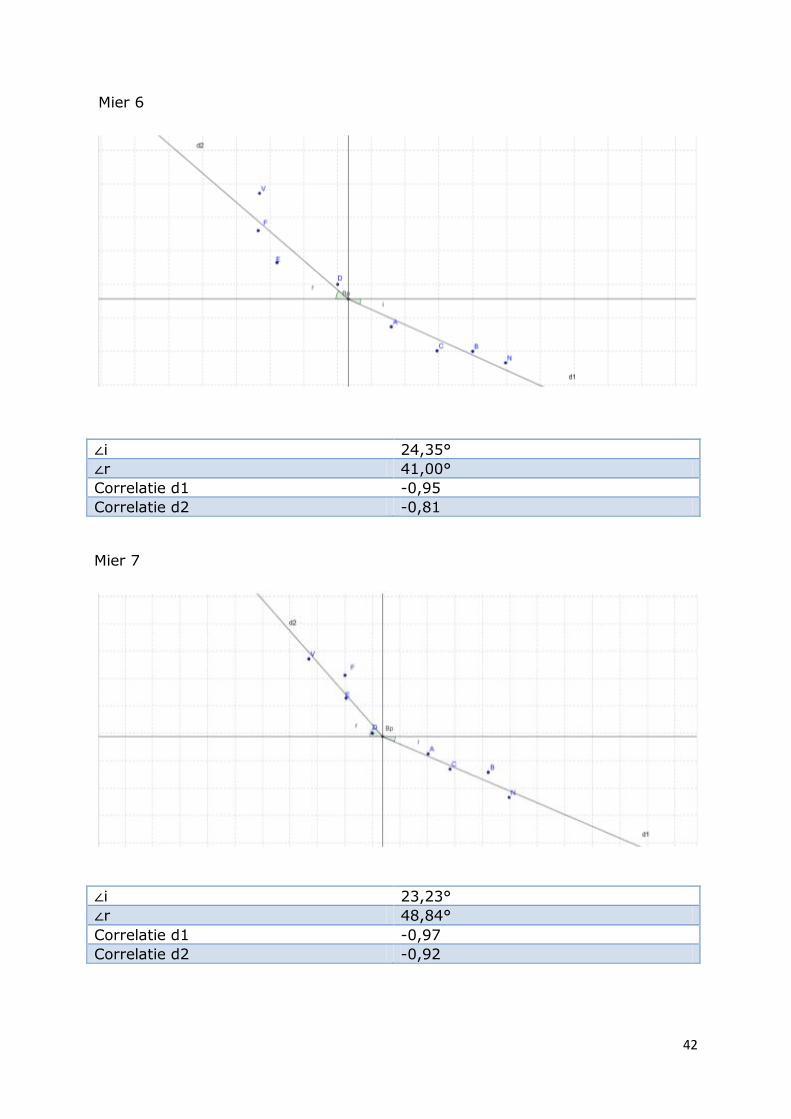
Mier 6
∠i 24,35°
∠r 41,00°
Correlatie d1 -0,95
Correlatie d2 -0,81
Mier 7
∠i 23,23°
∠r 48,84°
Correlatie d1 -0,97
Correlatie d2 -0,92
43
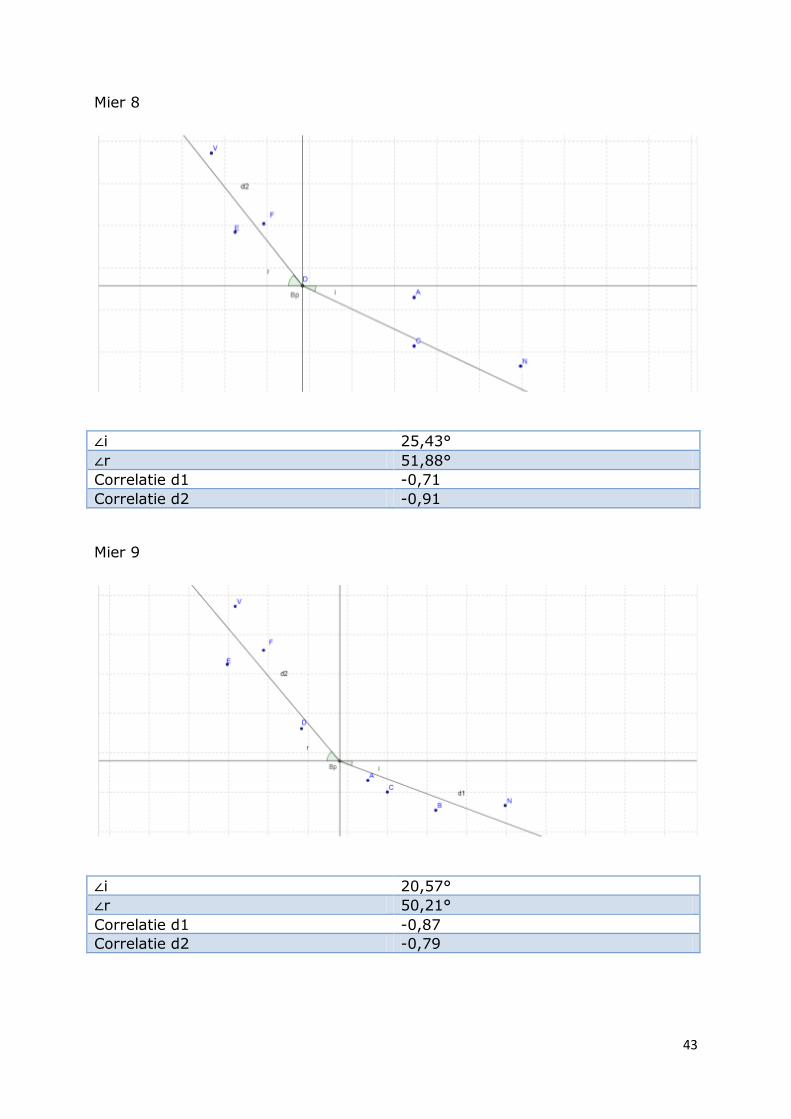
Mier 8
∠i 25,43°
∠r 51,88°
Correlatie d1 -0,71
Correlatie d2 -0,91
Mier 9
∠i 20,57°
∠r 50,21°
Correlatie d1 -0,87
Correlatie d2 -0,79
44
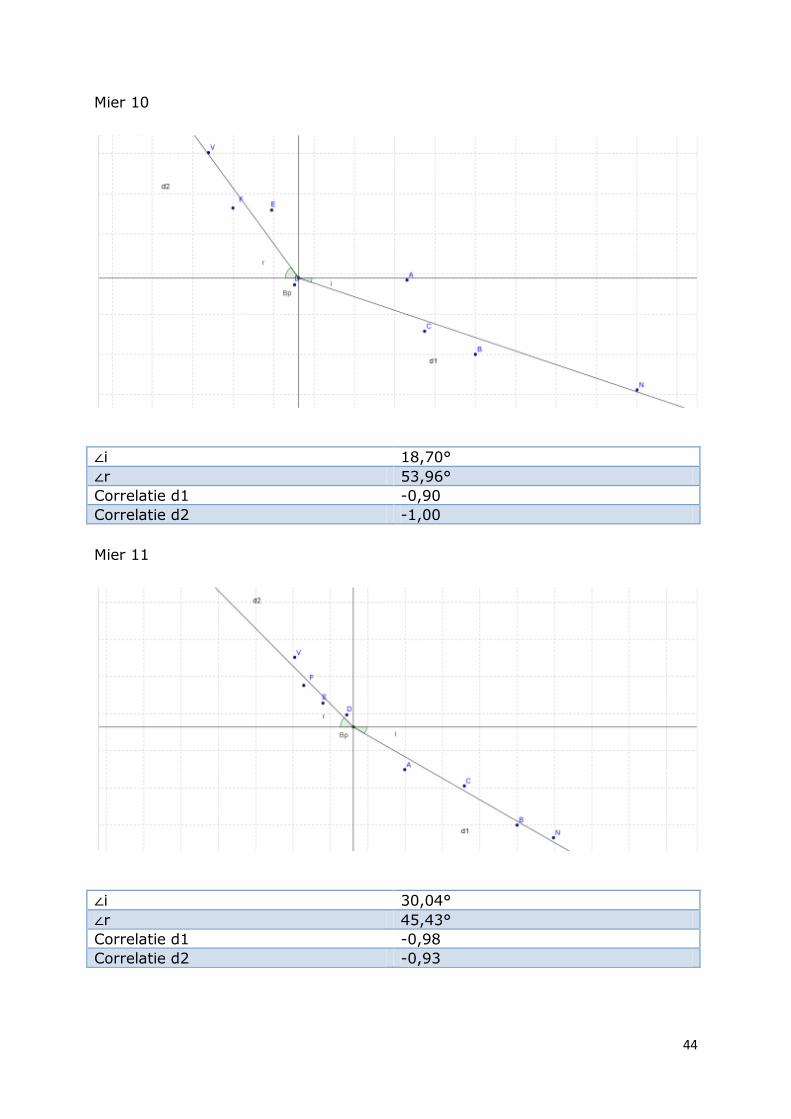
Mier 10
∠i 18,70°
∠r 53,96°
Correlatie d1 -0,90
Correlatie d2 -1,00
Mier 11
∠i 30,04°
∠r 45,43°
Correlatie d1 -0,98
Correlatie d2 -0,93
45
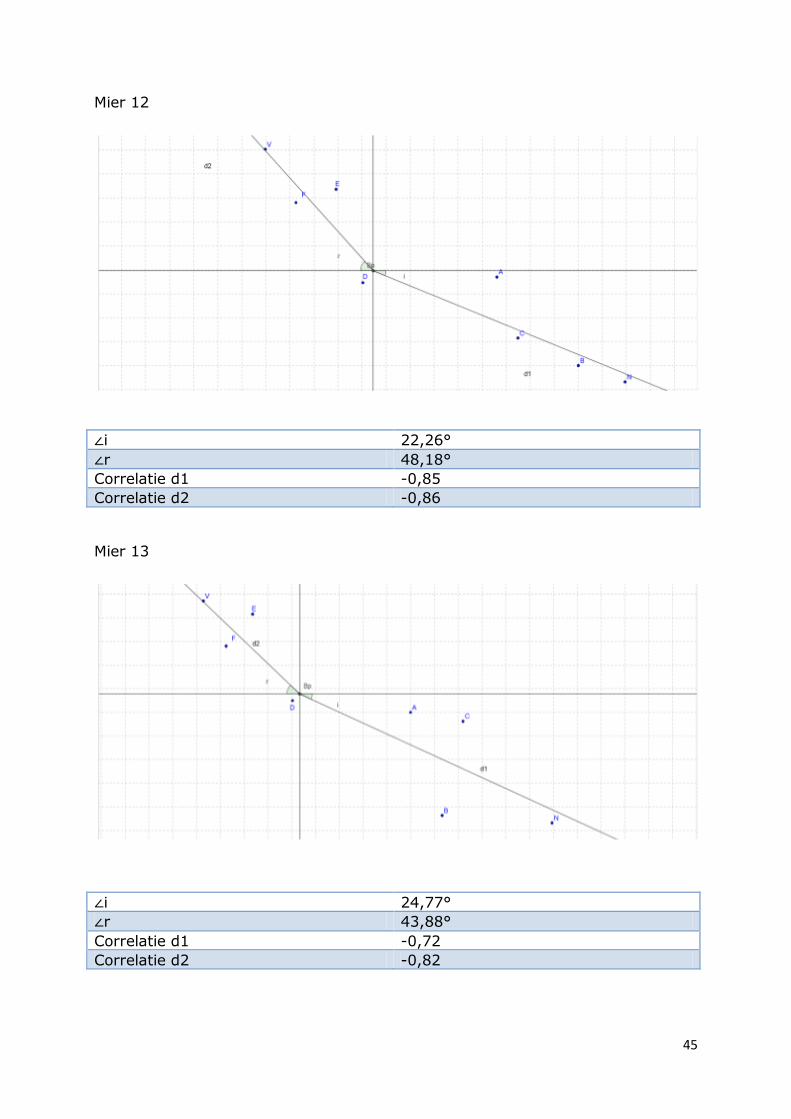
Mier 12
∠i 22,26°
∠r 48,18°
Correlatie d1 -0,85
Correlatie d2 -0,86
Mier 13
∠i 24,77°
∠r 43,88°
Correlatie d1 -0,72
Correlatie d2 -0,82
46
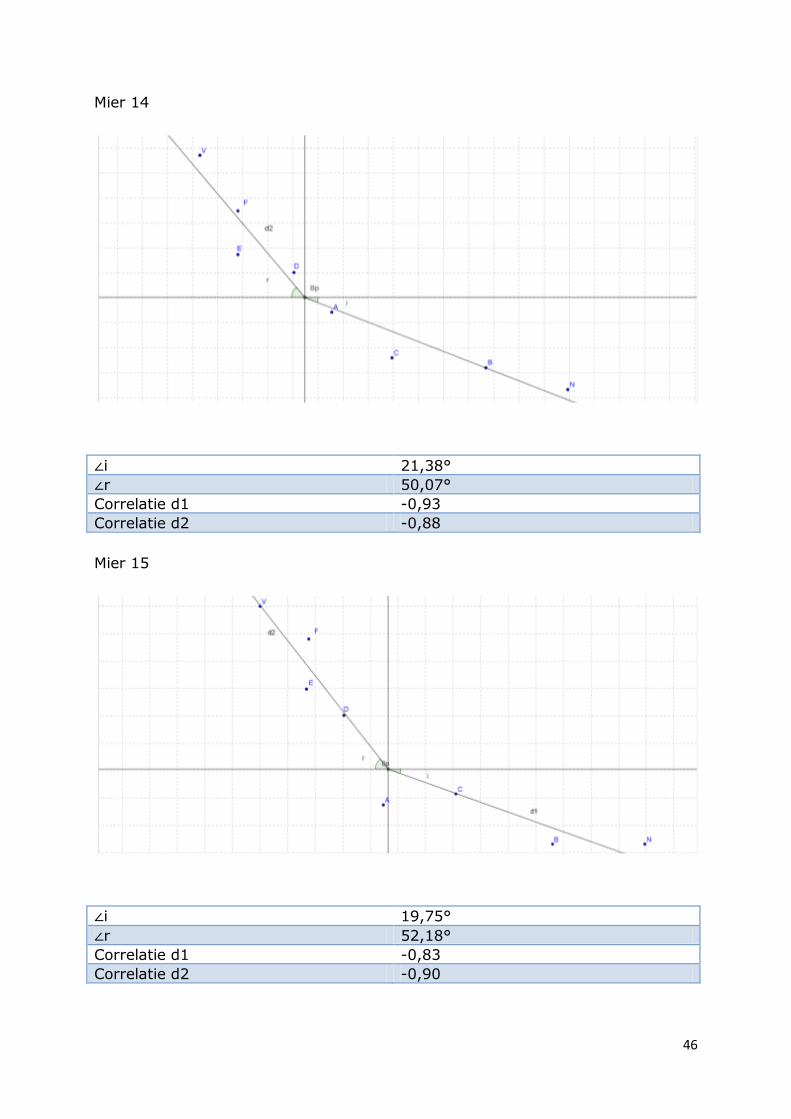
Mier 14
∠i 21,38°
∠r 50,07°
Correlatie d1 -0,93
Correlatie d2 -0,88
Mier 15
∠i 19,75°
∠r 52,18°
Correlatie d1 -0,83
Correlatie d2 -0,90
47
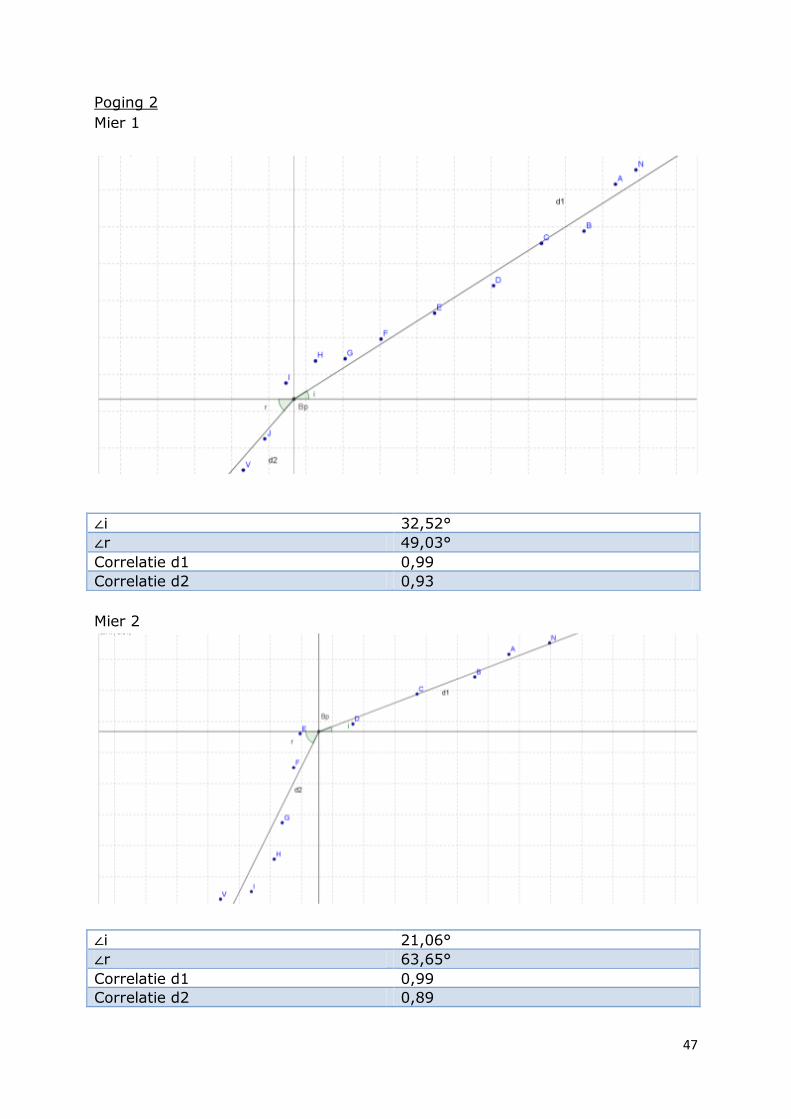
Poging 2
Mier 1
∠i 32,52°
∠r 49,03°
Correlatie d1 0,99
Correlatie d2 0,93
Mier 2
∠i 21,06°
∠r 63,65°
Correlatie d1 0,99
Correlatie d2 0,89
48
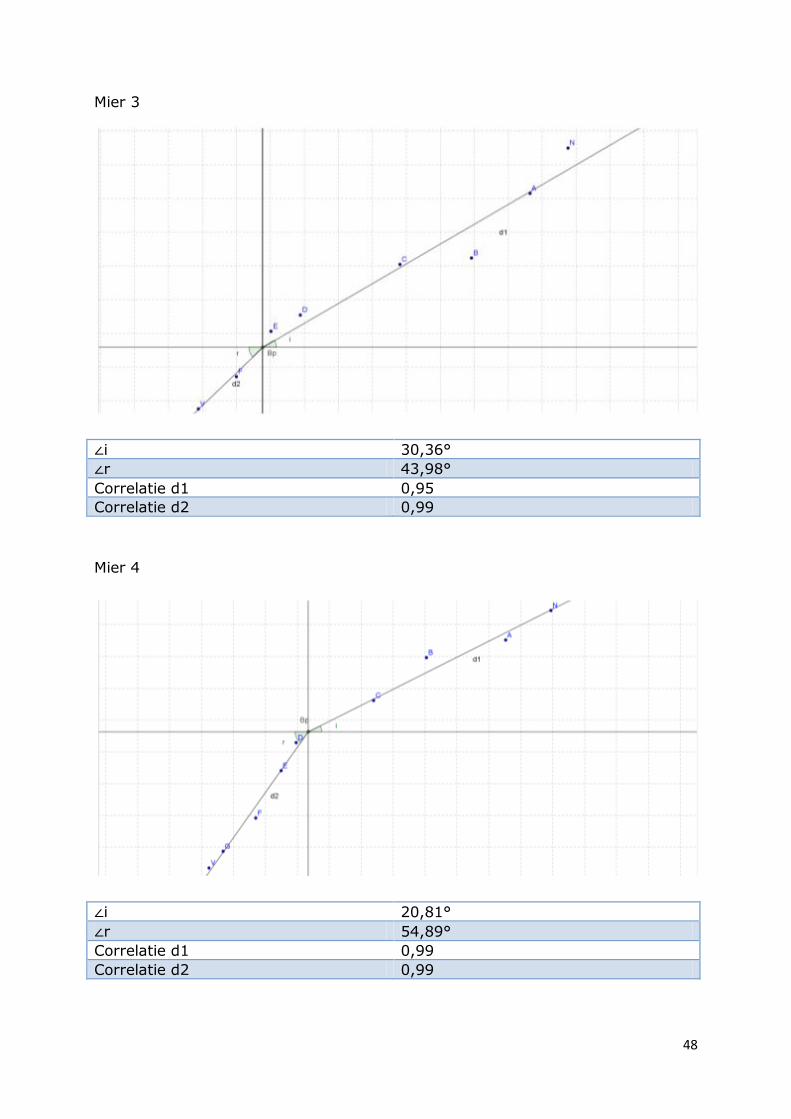
Mier 3
∠i 30,36°
∠r 43,98°
Correlatie d1 0,95
Correlatie d2 0,99
Mier 4
∠i 20,81°
∠r 54,89°
Correlatie d1 0,99
Correlatie d2 0,99
49
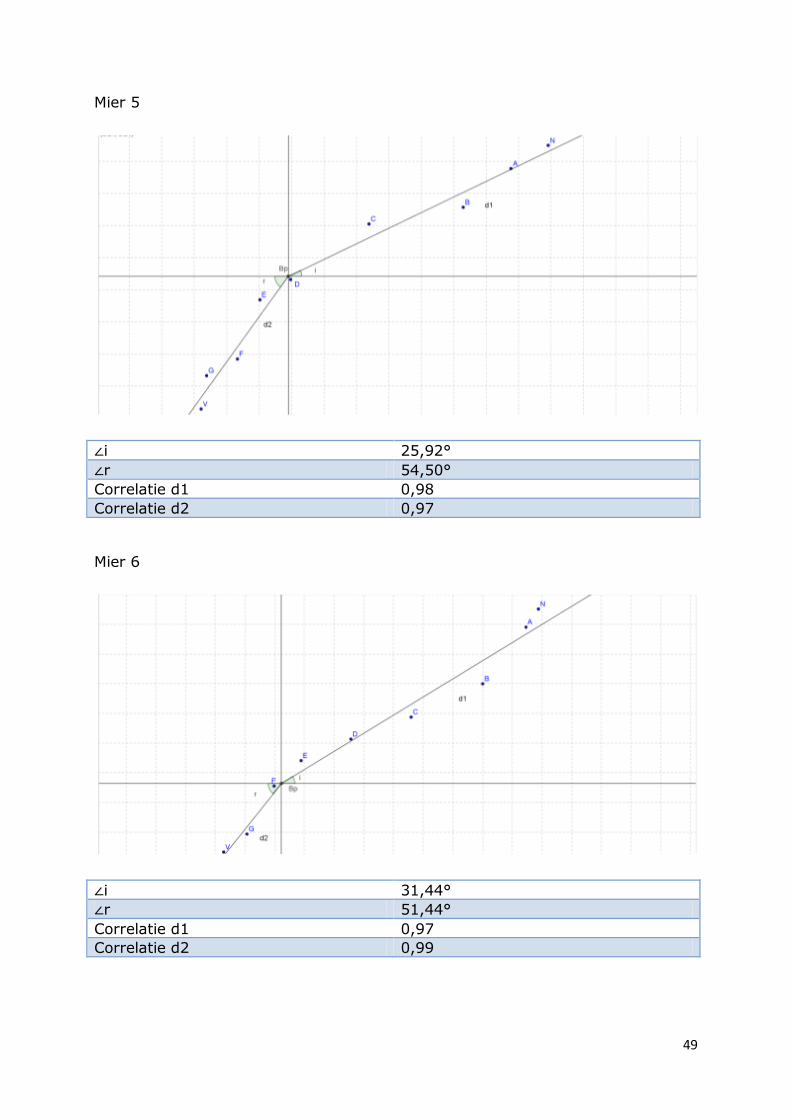
Mier 5
∠i 25,92°
∠r 54,50°
Correlatie d1 0,98
Correlatie d2 0,97
Mier 6
∠i 31,44°
∠r 51,44°
Correlatie d1 0,97
Correlatie d2 0,99
50
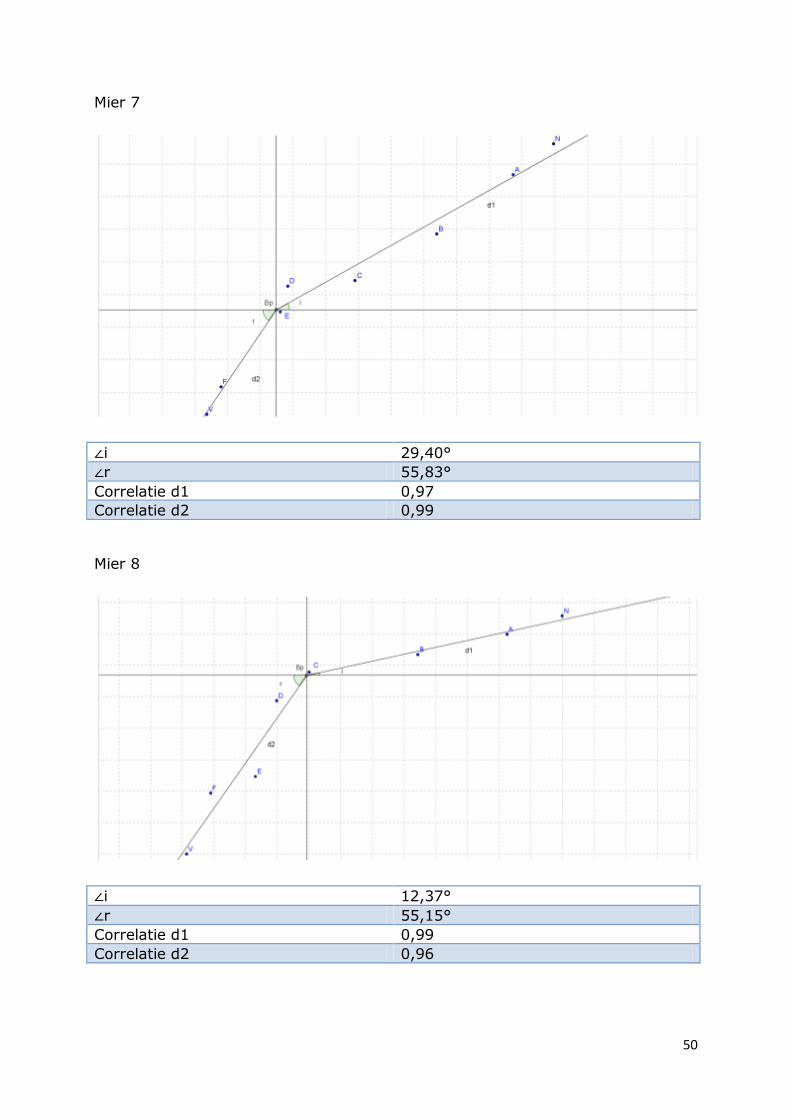
Mier 7
∠i 29,40°
∠r 55,83°
Correlatie d1 0,97
Correlatie d2 0,99
Mier 8
∠i 12,37°
∠r 55,15°
Correlatie d1 0,99
Correlatie d2 0,96
51
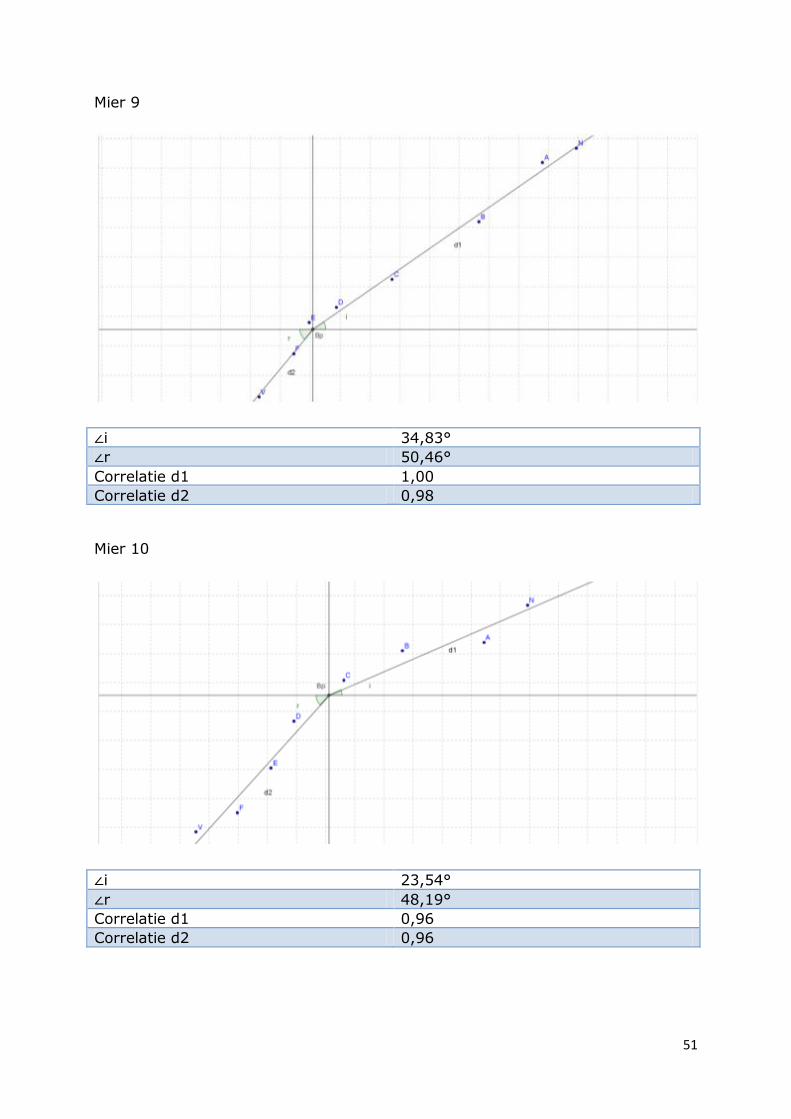
Mier 9
∠i 34,83°
∠r 50,46°
Correlatie d1 1,00
Correlatie d2 0,98
Mier 10
∠i 23,54°
∠r 48,19°
Correlatie d1 0,96
Correlatie d2 0,96
52
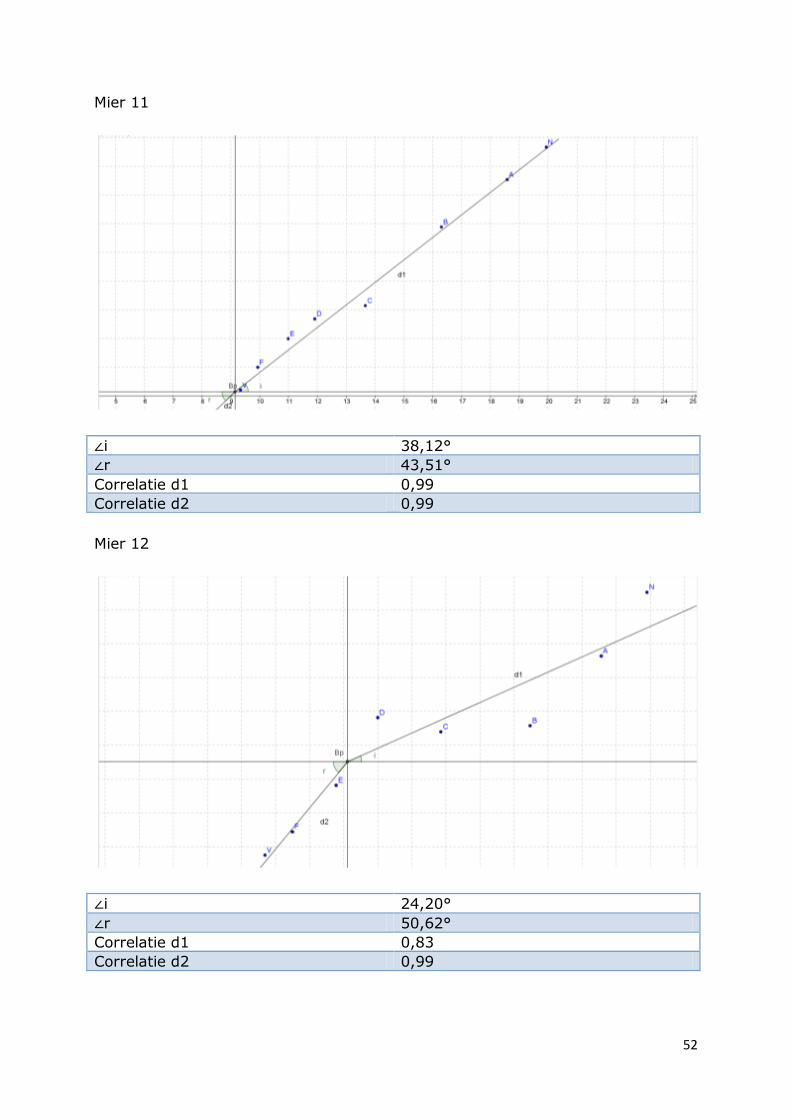
Mier 11
∠i 38,12°
∠r 43,51°
Correlatie d1 0,99
Correlatie d2 0,99
Mier 12
∠i 24,20°
∠r 50,62°
Correlatie d1 0,83
Correlatie d2 0,99
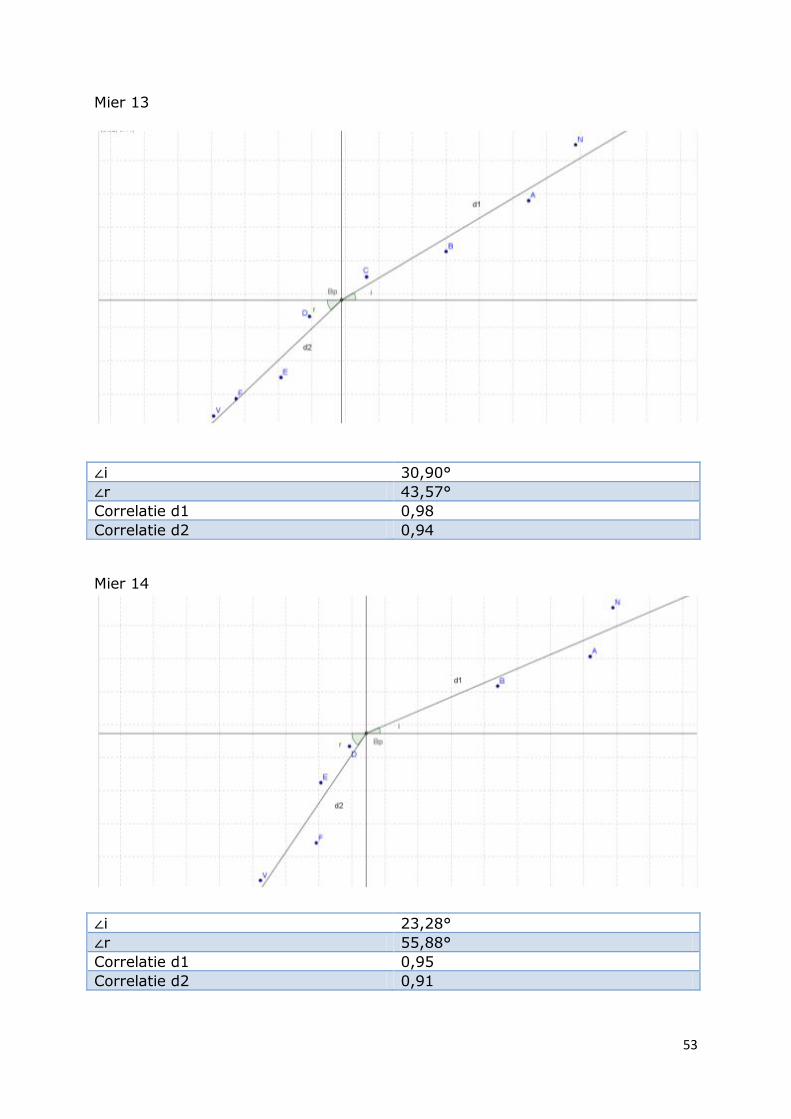
53
Mier 13
∠i 30,90°
∠r 43,57°
Correlatie d1 0,98
Correlatie d2 0,94
Mier 14
∠i 23,28°
∠r 55,88°
Correlatie d1 0,95
Correlatie d2 0,91
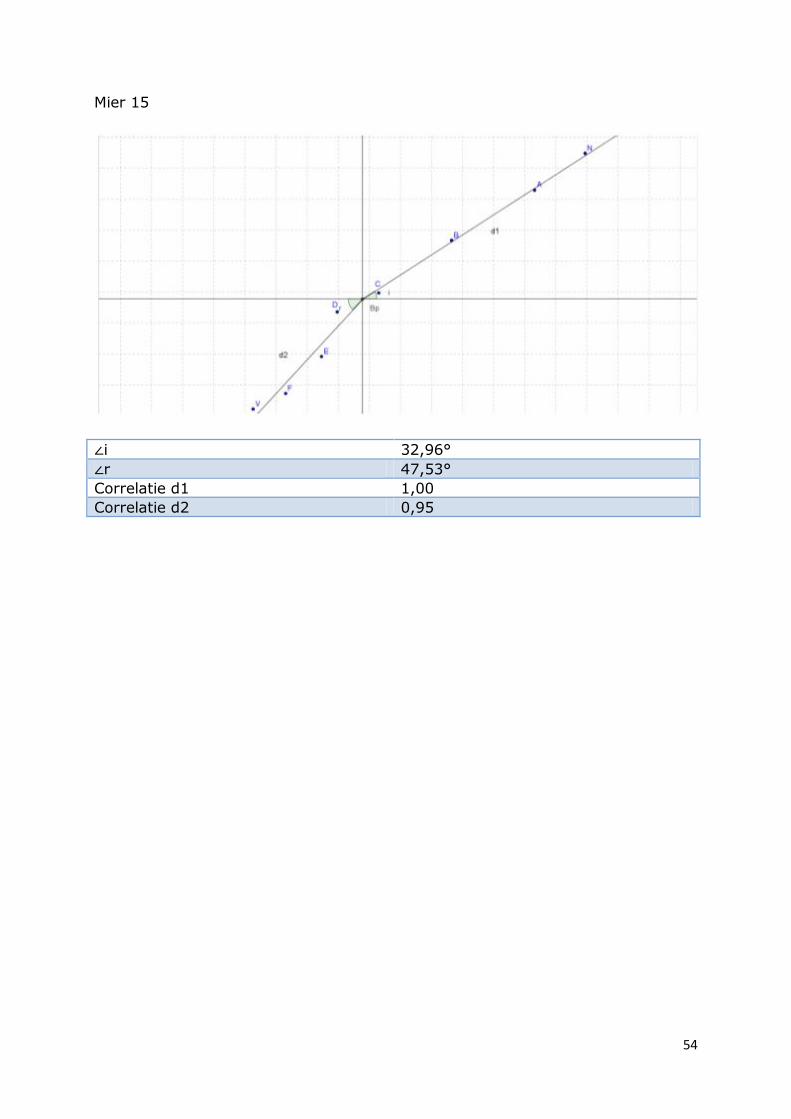
54
Mier 15
∠i 32,96°
∠r 47,53°
Correlatie d1 1,00
Correlatie d2 0,95


















